
Novel Insights into the Mechanisms of CIN85 SH3 Domains Binding to Cbl Proteins: Solution-Based...
17
Novel Insights into the Mechanisms of CIN85 SH3 Domains Binding to Cbl Proteins: Solution-Based Investigations and In Vivo Implications Abdessamad Ababou 1 , Mark Pfuhl 2 and John E. Ladbury 1 ⁎ 1 Department of Biochemistry and Molecular Biology, University College London, Gower Street, London WC1E 6BT, UK 2 Department of Biochemistry, University of Leicester, University Road, Leicester LE1 7RH, UK Received 10 October 2008; received in revised form 20 February 2009; accepted 24 February 2009 Available online 4 March 2009 CIN85 is a multifunctional protein that plays key roles in endocytic down-regulation of receptor tyrosine kinases, apoptosis, cell adhesion, and cytoskeleton rearrangement. Its three SH3 domains (CIN85A, CIN85B, and CIN85C) allow it to recruit multiple binding partners. To understand the manifold interactions of CIN85, we present a detailed high-resolution solution structural study of CIN85A and CIN85B binding to proline–arginine peptides derived from the cognate ligands Cbl and Cbl-b. We report the structure of CIN85B and provide evidence that both CIN85A and CIN85B, in isolation or when linked, form heterodimeric complexes with the peptides. We report unusual curved chemical shift changes for several residues of CIN85A when titrated with Cbl-b peptide, indicating the existence of more than one complex form. Here we demonstrate that CIN85A and CIN85B use different mechanisms for peptide binding. © 2009 Elsevier Ltd. All rights reserved. Edited by M. F. Summers Keywords: ubiquitination; NMR spectroscopy; isothermal titration calorimetry; chemical shifts Introduction The 85-kDa human Cbl-interacting protein CIN85, also known as SETA, Ruk, or SH3KBP1, is an important adaptor protein. 1 It is well known to play a key role in down-regulation of receptor tyrosine kinases and to facilitate endocytosis by interacting with endophilin-associated Cbl pro- teins. 2,3 However, CIN85 has also been implicated in other cellular processes such as apoptosis, via association with the p85 subunit of PI3K and AIP1/Alix, 4,5 as well as adhesion and cytoskeletal rearrangement, via interaction with p130Cas, cor- tactin, and ASAP1. 6–8 CIN85 is involved in additional important physiological processes such as regulation of cellular stress response via interaction with MEKK4, 9 cross-linking of F-actin into bundles, 10 and several others. 1,11–13 In addi- tion, CIN85 is implicated in infectious diseases via interaction with infected cell protein 0 from herpes simplex virus, 14 development of glomerulosclerosis in a diseased mouse model, 15 and breast cancer cell invasion via interaction with AMAP1. 16 CIN85 interacts with binding partners through its three SH3 domains (via atypical proline-rich motifs based on the sequence PXXXPR, where X is any amino acid), a proline-rich region, and a serine-rich region 10,17 (Fig. 1a). The importance of the CIN85 SH3 domains in clustering Cbl proteins has also been reported. 17 Although there are several iso- forms of CIN85, little is known about their roles in promoting or rescuing activity or in competing with full-length functions 5,18 (Fig. 1a). Nonetheless, the full-length protein and the isoform lacking the first SH3 domain show distinct functions. 19 CIN85 lacking the first two SH3 domains competes with the intact protein for binding to the p85α subunit of PI3K. 5 *Corresponding author. Department of Biochemistry and Molecular Biology, University of Texas M. D. Anderson Cancer Center, Unit 1000, 1515 Holcombe Boulevard, Houston, TX 77030, USA. E-mail address: [email protected]. Present address: A. Ababou, Department of Chemistry & Biological Sciences, University of Huddersfield, Queensgate, Huddersfield, HD1 3DH, UK. Abbreviations used: ITC, isothermal titration calorimetry; CS, chemical shift; NOESY, nuclear Overhauser enhancement spectroscopy; HSQC, heteronuclear single quantum coherence; 1D, one-dimensional. doi:10.1016/j.jmb.2009.02.061 J. Mol. Biol. (2009) 387, 1120–1136 Available online at www.sciencedirect.com 0022-2836/$ - see front matter © 2009 Elsevier Ltd. All rights reserved.
Transcript of Novel Insights into the Mechanisms of CIN85 SH3 Domains Binding to Cbl Proteins: Solution-Based...

doi:10.1016/j.jmb.2009.02.061 J. Mol. Biol. (2009) 387, 1120–1136
Available online at www.sciencedirect.com
Novel Insights into the Mechanisms of CIN85 SH3Domains Binding to Cbl Proteins: Solution-BasedInvestigations and In Vivo Implications
Abdessamad Ababou1, Mark Pfuhl2 and John E. Ladbury1⁎
1Department of Biochemistryand Molecular Biology,University College London,Gower Street, London WC1E6BT, UK2Department of Biochemistry,University of Leicester,University Road, Leicester LE17RH, UKReceived 10 October 2008;received in revised form20 February 2009;accepted 24 February 2009Available online4 March 2009
*Corresponding author. DepartmenMolecular Biology, University of TeCancer Center, Unit 1000, 1515 HolcHouston, TX 77030, USA. E-mail [email protected] address: A. Ababou, Dep
& Biological Sciences, University ofQueensgate, Huddersfield, HD1 3DAbbreviations used: ITC, isotherm
calorimetry; CS, chemical shift; NOESenhancement spectroscopy; HSQC, hquantum coherence; 1D, one-dimens
0022-2836/$ - see front matter © 2009 E
CIN85 is a multifunctional protein that plays key roles in endocyticdown-regulation of receptor tyrosine kinases, apoptosis, cell adhesion,and cytoskeleton rearrangement. Its three SH3 domains (CIN85A,CIN85B, and CIN85C) allow it to recruit multiple binding partners. Tounderstand the manifold interactions of CIN85, we present a detailedhigh-resolution solution structural study of CIN85A and CIN85B bindingto proline–arginine peptides derived from the cognate ligands Cbl andCbl-b. We report the structure of CIN85B and provide evidence that bothCIN85A and CIN85B, in isolation or when linked, form heterodimericcomplexes with the peptides. We report unusual curved chemical shiftchanges for several residues of CIN85A when titrated with Cbl-b peptide,indicating the existence of more than one complex form. Here wedemonstrate that CIN85A and CIN85B use different mechanisms forpeptide binding.
© 2009 Elsevier Ltd. All rights reserved.
Keywords: ubiquitination; NMR spectroscopy; isothermal titration calorimetry;chemical shifts
Edited by M. F. SummersIntroduction
The 85-kDa human Cbl-interacting proteinCIN85, also known as SETA, Ruk, or SH3KBP1,is an important adaptor protein.1 It is well knownto play a key role in down-regulation of receptortyrosine kinases and to facilitate endocytosis byinteracting with endophilin-associated Cbl pro-teins.2,3 However, CIN85 has also been implicatedin other cellular processes such as apoptosis, viaassociation with the p85 subunit of PI3K andAIP1/Alix,4,5 as well as adhesion and cytoskeletal
t of Biochemistry andxas M. D. Andersonombe Boulevard,dress:
artment of ChemistryHuddersfield,H, UK.al titrationY, nuclearOverhausereteronuclear singleional.
lsevier Ltd. All rights reserve
rearrangement, via interaction with p130Cas, cor-tactin, and ASAP1.6–8 CIN85 is involved inadditional important physiological processes suchas regulation of cellular stress response viainteraction with MEKK4,9 cross-linking of F-actininto bundles,10 and several others.1,11–13 In addi-tion, CIN85 is implicated in infectious diseases viainteraction with infected cell protein 0 from herpessimplex virus,14 development of glomerulosclerosisin a diseased mouse model,15 and breast cancercell invasion via interaction with AMAP1.16
CIN85 interacts with binding partners through itsthree SH3 domains (via atypical proline-rich motifsbased on the sequence PXXXPR, where X is anyamino acid), a proline-rich region, and a serine-richregion10,17 (Fig. 1a). The importance of the CIN85SH3 domains in clustering Cbl proteins has alsobeen reported.17 Although there are several iso-forms of CIN85, little is known about their roles inpromoting or rescuing activity or in competingwith full-length functions5,18 (Fig. 1a). Nonetheless,the full-length protein and the isoform lacking thefirst SH3 domain show distinct functions.19 CIN85lacking the first two SH3 domains competes withthe intact protein for binding to the p85α subunit ofPI3K.5
d.

Fig. 1. Domain organization and isoforms of CIN85. (a) Domain organization of CIN85 and its natural isoforms. (b)Ribbon representation of the structural model of CIN85AB fragment showing the four prolines located in the linker. (c)Sequence alignment of the SH3 domains CIN85A and CIN85B. Arrows highlight binding site residues. Blue, conserved;red, nonconserved.
1121Binding Mechanisms of CIN85 SH3 Domains
The SH3 domain is one of the best-characterizedinteracting modules involved in protein–proteinassociations.20,21 It binds primarily to proline-richsequences that generally adopt a PPII helix conforma-tion. Binding of peptides containing proline-richsequences to SH3 domains can occur in twoopposing orientations. The stoichiometry of bindingis 1:1, with notable exception of the tandem SH3domains of p47phox, where a single peptide bindsto both SH3 domains.22,23 Recent studies havereported novel binding features of CIN85 SH3domains. In one of these studies that explored themolecular mechanism of the interaction of CIN85with the p85α subunit of PI3K, it was proposed thatthe proline-rich motif from the p85α monomersimultaneously binds to both N-terminal SH3domain A (referred to as CIN85A) and N-terminalSH3 domain B (referred to as CIN85B).24 Taking intoaccount the long linker between CIN85A andCIN85B (Fig. 1b), such complex formation isplausible. Recently, it was reported that binding ofCIN85A to a peptide from Cbl-b results in hetero-trimer complex formation between two SH3domains and one peptide.25 Since the CIN85 SH3domains are selective for proline-rich motifs of typePXXXPR, it is possible that CIN85B may also form aheterotrimeric complex of the form described forCIN85A.
Taking into account the large number of interact-ing proteins with CIN85 via its SH3 domains, theexistence of several of its isoforms with deletion ofone to all SH3 domains, and the recent new bindingmode revealed by its SH3 domains, further detailedstudies of the CIN85 protein—particularly its SH3domains—are important. Hence, to explore thestructural implications and to understand the func-tional role of complex formation between CIN85SH3 domains and proline-rich peptides from c-Cbland Cbl-b proteins (referred to as Cbl and Cbl-b,respectively), we have initiated a detailed investiga-tion of high-resolution structures and interactions insolution of CIN85A and CIN85B, in isolation and inthe context of a CIN85A–linker–CIN85B fragment(referred to as CIN85AB). We report the solutionstructure of CIN85B. Isothermal titration calorime-try (ITC) and NMR spectroscopic experimentsdemonstrate that both domains, in isolation or aspart of CIN85AB, form heterodimeric complexeswith both peptides. An unusual curved chemicalshift(s) (CS) change for several residues in CIN85Awas observed upon titration with Cbl-b. This revealsthe existence of more than one complex withCIN85A in isolated form or in the CIN85AB form.Here we report that the mechanisms of binding ofCIN85A and CIN85B to both peptides are differentand independent of CIN85AB. This suggests that the

1122 Binding Mechanisms of CIN85 SH3 Domains
SH3 domains of CIN85 have evolved to incorporatedifferent modes of recognition for cognate ligands,providing a level of specificity to achieve its multi-functional capability.
Results
Solution structure of CIN85B and the absence ofintramolecular interaction with CIN85A withinthe CIN85AB fragment
The structure of CIN85A was recently reportedin complex with Cbl-b.25 It was shown that there
Fig. 2. Solution structure of CIN85B. (a) Superposition of thand labeling of the best structure. (c) Overlaid 1H–15N HSQCIN85AB (blue) fragments. (d) Mapping of the amino acidsresonances in CIN85AB.
are no substantial structural changes between theapo form and the complex form of CIN85A.26 Thestructure calculation of CIN85B reported here isbased on excellent-quality three-dimensional 15N-and 13C-resolved heteronuclear nuclear Overhau-ser enhancement spectroscopy (NOESY) spectra ofa monomeric protein supported by a rotationalcorrelation time of 6.92 ns at a concentration of∼0.7 mM. The family of structures shown in Fig.2a indicates a highly defined structure, supportedby a root mean square deviation (rmsd) of 0.27 Åfor backbone atoms (see Table S1 in Supplemen-tary Materials). The only regions of disorder arethe N- and C-termini, which extend beyond the
e 27 best structures. (b) Secondary structure representationC spectra of isolated CIN85A (black), CIN85B (red), andof CIN85A with small to substantial CS changes in their

Table 1. ITC binding parameters of CIN85A, CIN85B,and CIN85AB with Cbl and Cbl-b peptides
Protein–peptide n Kd (μM) ΔHobs (kJ/mol)
CIN85A–Cbl 0.84±0.05 31.31±5.78 −14.29±2.05CIN85B–Cbl 0.92±0.02 4.93±0.55 −36.59±1.13CIN85AB–Cbl 1.80±0.26 27.09±4.24 −48.17±5.70CIN85A–Cbl-b 0.89±0.17 9.87±3.80 −7.43±1.09CIN85B–Cbl-b 0.80±0.11 7.06±1.68 −38.22±7.16CIN85AB–Cbl-b⁎ 2.12±0.05 12.65±1.30 −30.67±0.93CIN85A–Cbl-b(F) 0.83±0.12 25.90±5.87 −12.76±3.00CIN85B–Cbl-b(F)⁎ 0.94±0.02 6.22±0.64 −31.56±2.17CIN85AB–Cbl-b(F) 1.97±0.21 33.07±8.65 −38.03±7.76CIN85A–Cbl-b(E) 0.81±0.08 8.46±2.01 −11.11±5.85CIN85B–Cbl-b(E)⁎ 0.86±0.03 6.14±0.87 −29.65±1.19CIN85AB–Cbl-b(E) 2.04±0.04 21.66±3.43 −34.18±2.23
All titrations were carried out two or three times to estimate theerrors as standard deviations, except for titrations with (⁎), where theerrors were estimated as confidence intervals at the 68.3% level.28
Fig. 3. ITC measurements of the proteins with Cblpeptide. (a) ITC data titration with the best-fitting line forCIN85A. (b) ITC data titration with the best-fitting line forCIN85B. (c) ITC data titration with the best-fitting line forCIN85AB. The insets show the raw data of each titration.
1123Binding Mechanisms of CIN85 SH3 Domains
SH3 domain. The solution structure of CIN85Bmaintains a typical SH3 fold consisting of five β-strands organized in an antiparallel β-barrelstructure, with a short 310 helix between the lasttwo β-strands, and two characteristic loops,namely, the RT loop (between β1 and β2) andthe n-Src loop (between β2 and β3), as shown inFig. 2b. Sequence alignment of CIN85A andCIN85B shows about 47% identity. The residuesin the binding pocket are highly conserved, butsome notable differences are apparent (Fig. 1c,blue and red arrows). Structural alignment ofCIN85B with the crystal structure of CIN85A incomplex with Cbl-b shows significant differencesin the RT and n-Src loop regions and an overallrmsd of 3 Å.25
Backbone assignment of polypeptides provides adirect assessment of the existence of any intramo-lecular interactions between CIN85A and CIN85Bwithin CIN85AB. Overlay of the 1H–15N hetero-nuclear single quantum coherence (HSQC) spectraof the isolated domains with that of CIN85ABshows a reasonable match (Fig. 2c). CIN85A hasabout 20 (small to substantial) unmatched reso-nances. Far better matching is observed betweenCIN85B and CIN85AB. Unmatched resonances ofCIN85A were mapped over the whole domainsurface (Fig. 2d). This suggests that no specificintermolecular or intramolecular interaction exists.Thus, these data rule out the possibility of aninteraction of the SH3 domains with the PXXP motif(PEKPLHEVP), which is present in the linker region(see Fig. 1b). Such CS changes may be directlyrelated to the removal of the C-terminal His-tag27and/or the presence of the linker. To control for this,we expressed and purified a new construct withoutthe His-tag (CIN85A_new). The overlay of the1H–15N HSQC spectra of the new construct withCIN85AB revealed a very good match, with almostall its resonances within CIN85AB (see Fig. S1 inSupplementary Materials). This demonstrates thatthe presence of the His-tag in the construct ofCIN85A can affect the CS of a small number
resonances of CIN85A within the CIN85AB poly-peptide. These observations, along with additionalevidence provided below, suggest that CIN85A andCIN85B do not interact within the CIN85ABfragment.

1124 Binding Mechanisms of CIN85 SH3 Domains
CIN85A and CIN85B form 1:1 complexes withCbl and Cbl-b peptides
ITC analysis is summarized in Table 1, and useexample of typical ITC data for the interaction of thedomains with Cbl peptide is shown in Fig. 3. Clearly,the interaction stoichiometry n shows that CIN85A29
and CIN85B form heterodimeric complexes withboth peptides.CIN85A and CIN85B titrations with peptides were
monitored with 1H–15N HSQC spectra. Most of theresidues in the binding site are in a fast-exchangeregime. The combined backbone 1H and 15N CSchanges between apo and saturated complexes areplotted against the domain sequences in Fig. 4a. Forboth domains, there are three major groups ofresidues with substantial CS changes above the⟨Δδtot⟩ threshold. These include residues in the RTloop (8–19 and 108–117 for CIN85A and CIN85B,respectively), at the n-Src loop and β3 (32–39 and128–138, respectively), and near β4 and the 310 helix(48–53 and 145–154, respectively). Upon binding ofthe peptides, several residues of CIN85B showhighly perturbed CS. CIN85A, on the other hand,shows low CS perturbations, except for Asp16 at theRT loop and Asn51 at the 310 helix. These residuesform hydrogen bonds with the peptides.25 Residueswith CS changes above a ⟨Δδtot⟩+1σ threshold aremapped on the domain structures in Fig. 4b.CIN85A and CIN85B binding to Cbl results in
linear CS changes in amide and side-chain signals(Fig. 5). For CIN85B binding to Cbl-b, similar CSchanges were observed. Surprisingly, CIN85A bind-ing to Cbl-b reveals highly curved and slightlycurved CS changes for Phe8, His14, Gly35, Trp36,Asp50, and Asn51. As an illustration of such strikingcurved CS changes, we show the titrations of theTrp36 side chain in CIN85A (Fig. 5). Interestingly inCIN85B, while the Trp135 side-chain titration withCbl peptide shows linear CS changes, we observed adouble signal set at intermediate concentrations(0.17–0.57 Eq) (Fig. 5).To verify further the stoichiometry of the com-
plexes formed by CIN85A and CIN85B with bothpeptides in solution, we have determined therotational correlation times (τc) of the apo andcomplex forms of these domains using NMR relaxa-tion (Table 2). The τc values of the apo domains (∼6–7 ns) increase in the complexes (∼8.8–9.0 ns). Notethat the CIN85A construct is 10 residues shorter thanthe CIN85B construct, which may account for thelower τc values. Taking into account the anisotropicnature of the complexes due to the disorderedtermini of both the domains and/or the peptides,30
τc values provide strong additional evidence that theCIN85A and CIN85B complexes are monomericspecies binding with a 1:1 stoichiometry.
CIN85A and CIN85B form independentheterodimer complexes in the context of CIN85AB
We have shown that isolated SH3 domains formheterodimeric complexes with Cbl and Cbl-b. These
results prompted us to investigate the complexformation of these domains in the context ofCIN85AB. ITC analysis of CIN85AB binding toboth peptides is summarized in Table 1. Thestoichiometry of these complexes is 1:2, stronglysuggesting that CIN85A and CIN85B behave asisolated domains binding one peptide each.CIN85AB titrations with Cbl and Cbl-b, as moni-
tored with 1H–15N HSQC spectra, show CS changessimilar to those of the isolated domains (see examplein Fig. 5), including the curved CS changes observedfor the CIN85A complex with Cbl-b. Overlaying ofthe 1H–15N HSQC spectra of the proteins in complexwith Cbl and Cbl-b peptides shows a very goodmatch between signals of CIN85B and CIN85AB.Again CIN85A shows similar residues with (small tosubstantial) unmatched signals, as observedwith theapo form (Fig. 2; see also Fig. S2a and b inSupplementary Materials). This confirms that sev-eral residues in CIN85A have different CS inCIN85AB, mainly due to effect of the His-tag27 (seeFig. S1 in Supplementary Materials) and/or theprolines of the linker in terms of backbone rigidity,but are unlikely to arise from any intramolecularinteraction as previously reported for an SH3domain with a proline-rich linker.31 In addition, tofurther verify the stoichiometry of the complexesformed by CIN85A and CIN85B with both peptideswithin CIN85AB, we have determined the rotationalcorrelation times (τc) of the apo and complex formsof these domains in CIN85AB (Table 2). The τc valuesof the apo domains in CIN85AB (∼6.8–7.3 ns)increase in the complexes (∼8.4–8.8 ns) and areindicative of the independent behavior of thesedomains within CIN85AB. These observations con-firm that CIN85A and CIN85B form heterodimercomplexes with both peptides in the context ofCIN85AB and behave independently within the apoand complex forms of CIN85AB. Interestingly,residues that showed slightly curved CS changesfor CIN85A in complex with Cbl-b have highlycurvedCS changes inCIN85AB in complexwithCbl-b (see examples in Fig. 6).
CIN85A and CIN85B bind differently to Cbl andCbl-b peptides
The binding affinities between the two peptidesare different for CIN85A, but are similar for CIN85B.CIN85B has a more favorable change in enthalpy,ΔHobs, for binding than CIN85A, suggesting theexistence of clear differences in peptide recognitionbetween these domains. These differences are intri-guing because the peptides have similar binding coremotifs PXXXPR, and both domains have similarbinding sites with conserved residues (Fig. 1c). Inaddition, our NMR titrations reveal the unusualcurved CS changes in several residues of CIN85A incomplex with Cbl-b, but not with Cbl, while no suchcurved changes are observed for CIN85B with eitherpeptide (Fig. 5). Comparison of Cbl and Cbl-b showsthat Ala903 and Arg909 in Cbl-b are replaced byGlu821 and Phe827 in Cbl. To rationalize the

Fig. 4. Combined 1H and 15NCS perturbations of the proteinswith Cbl andwild-type andmutant Cbl-b peptides. In allpanels, the average CS perturbation of all amino acids ⟨Δδtot⟩ is indicated by the first blue horizontal line; the ⟨Δδtot⟩+1σthreshold is shown as the second blue horizontal line. (a)Wild-type Cbl andCbl-b peptides. (b)Mapping of the amino acidsat and above the ⟨Δδtot⟩+1σ threshold onCIN85A andCIN85B structures for titrationwith thewild-type Cbl-b peptide. (c)Mutants of Cbl and Cbl-b peptides.
1125Binding Mechanisms of CIN85 SH3 Domains

Fig. 4 (legend on previous page)
1126 Binding Mechanisms of CIN85 SH3 Domains
difference in the binding affinities of CIN85A forthese peptides in structural terms, we mutated theseresidues in the Cbl-b peptide (since this complex hasbeen characterized25). We will refer to the peptidemutants of Cbl-b as Cbl-b(F) and Cbl-b(E) formutants Arg909-Phe and Ala903-Glu, respectively.CIN85B binds with similar affinity and thermo-
dynamic parameters to both Cbl-b(F) and Cbl-b(E)(Table 1). Surprisingly, CIN85A shows comparablebinding affinities between Cbl and Cbl-b(F), as wellas between Cbl-b and Cbl-b(E). NMR titrationsreveal unexpected effects of these mutations uponbinding to CIN85A and CIN85B. Titrations withCbl-b(F) and Cbl-b(E) do not change the bindingbehavior of CIN85B (Figs. 4c and 5). Indeed, theTrp135 side-chain titration with Cbl-b(F) resulted inlinear CS changes, with a doublet signal observedonly at intermediate concentrations (Fig. 5). This isdue to the presence of Phe827 in Cbl, since mutationof Arg909 to Phe in Cbl-b resulted in similar be-havior, as observed with Cbl. SH3 domains bind toproline-rich peptides using this conserved Trp,which makes a hydrogen bond with the peptidebackbone. Although it has been proposed that thisTrp could dictate the peptide binding orientationthrough a different orientation of its side chain,32
such a small change in side-chain conformation isunlikely to affect the Trp135 side-chain signal.Instead, we suggest that the Phe residue in Cbl orCbl-b(F) exchanges slowly between two transientconformers detected only at intermediate concentra-
tions. Indeed, at intermediate concentrations, thebinding rates may be affected, hence indicating thepresence of other conformational states of the inter-acting residues.33
Titration of CIN85A with Cbl-b(E) produced thesame unusual curved CS changes for residues as inCbl-b titration; however, Cbl-b(F) abrogates thiseffect (Fig. 5). These results show that Arg909(underlined in PXXXPR) has a critical role in givingrise to the observed unusual curved CS changes inCIN85A signals for Cbl-b binding. This is intriguingbecause our mutants do not affect Arg911, whichwas shown to be an important residue for bindingwith CIN85,17,34 or Arg904, which has been reportedto be crucial for peptide-induced dimerization.25
Instead, our mutants have kept the pseudo-symme-try of the binding core motif of Cbl-b as RPPXXXPR.
Discussion
CIN85A formsmore thanoneheterodimer complexwith Cbl-b, but not with Cbl
We have provided evidence that in solution,CIN85A forms a heterodimeric complex with bothCbl and Cbl-b. Interestingly, the NMR titration ofCIN85A with Cbl-b shows unusual curved CSchanges for several residues (Fig. 5). Nonlinear CSchanges have been previously reported;35–37 how-ever, to our knowledge, curved CS changes have

Fig. 5. Spectral perturbations ofNε (NE) of conserved Trp residuesfrom the SH3 domains of CIN85.Trp36 (W36NE) and Trp135(W135NE) are found in CIN85Aand CIN85B respectively. Top tworows: Left - W36 binding to Cbl andCbl-b (titration performed up to 2.8and 2.5 equivalents respectively).Right - W135 on binding to Cbl andCbl-b (1.6 and 2.4 eq. respectively).Middle row: Trp36 (blue arrow)and Trp135 (black arrow) ofCIN85AB on binding to Cbl andCbl-b (4.5 and 4.3 eq. respectively).Bottom two rows: Left - W36 onbinding to Cbl-b(F) and Cbl-b(E)(up to 3.1 and 2.8 eq. respectively).Right - W135 on binding to Cbl-b(F)and Cbl-b(E) (both titrations up to1.7 Eq.).
1127Binding Mechanisms of CIN85 SH3 Domains

Table 2. Rotational correlation time (ns) for the apo and complex forms of separate CIN85A CIN85B, and withinCIN85AB [i.e., CIN85AB(A) or CIN85AB(B)]
State form CIN85A CIN85B CIN85AB(A) CIN85AB(B)
Apo 6.25±0.27 6.92±0.20 6.81±0.76 7. 33±0.87Complex with Cbl 8.75±0.24 9.10±0.22 8.50±0.47 8.79±0.56Complex with Cbl-b 8.55±0.21 8.95±0.24 8.31±0.39 8.58±0.54
1128 Binding Mechanisms of CIN85 SH3 Domains
only been reported in proteins for the temperaturedependence of the amide signals.38 The CS changecurvature was suggested to originate from alter-native protein folding states.38 SH3 domains bindproline-rich peptides without any obvious change intheir folding states. Consequently, since linear CSchanges are the hallmarks of the formation of onehighly populated complex, to explain the origin ofour curved CS changes, we have to consider theexistence of more than one complex population insolution (Fig. 7a). A one-dimensional (1D) lineshapessimulation in the fast-exchange regime between two
Fig. 6. Comparison of spectral changes in His14 and Glyfragment (left) and within the CIN85AB fragment (right).
states shows that lineshapes will shift only in onedirection toward the final complex (data not shown).Inspection of the 1H 1D spectra of Trp36 side-chaintitration reveals that lineshapes shift in one directionup to∼0.5 Eq and then shift in the opposite direction(Fig. 7b). Curved CS changes are only likely to beobserved when the CS in these states is largelydifferent. The most likely way to produce such CSdifferences will be through change in hydrogenbonding.39,40 Hydrogen bonds are affected by theelectronic structure of the residues involved in theirformation and by proximal amino acids.41 The X-ray
35 residues during the binding of Cbl-b in the CIN85A

Fig. 7. Existence of at least two complex forms of CIN85Awith Cbl-b. (a) Schematic three-state model for the curved CS of Trp36 side chain. Linear CS changes (red and blue)represent the CS changes for each heterodimer complex formation in slow-exchange regime. (b) Experimental Trp36 side-chain 1H lineshapes shift change direction at∼0.45 Eq. (c)overleaf CIN85A in complex with Cbl-b, which binds in either C-terminal-to-N-terminal orientation (Complex A left) or N-terminal-to-C-terminal orientation (complex B right).The complexes were taken from the X-ray structure (Protein Data Bank code 2BZ8) and energy minimized. (d) Simulation of lineshapes shift change for the three-state model inslow-exchange regime (we used kex=7 s−1) and fast-exchange regime (we used kex=3000 s−1). (e) Data fit plot of Trp36 side-chain 1H CS change using model 1 (competitionbetween two heterodimers). (f) Data fit plot of Trp36 side-chain 1H CS change using model 2 (competition between a heterotrimer and a heterodimer) (g) Rotational correlationtime of CIN85A in the absence of Cbl-b and at different concentrations of Cbl-b.
1129Binding
Mechanism
sof
CIN85
SH3Dom
ains
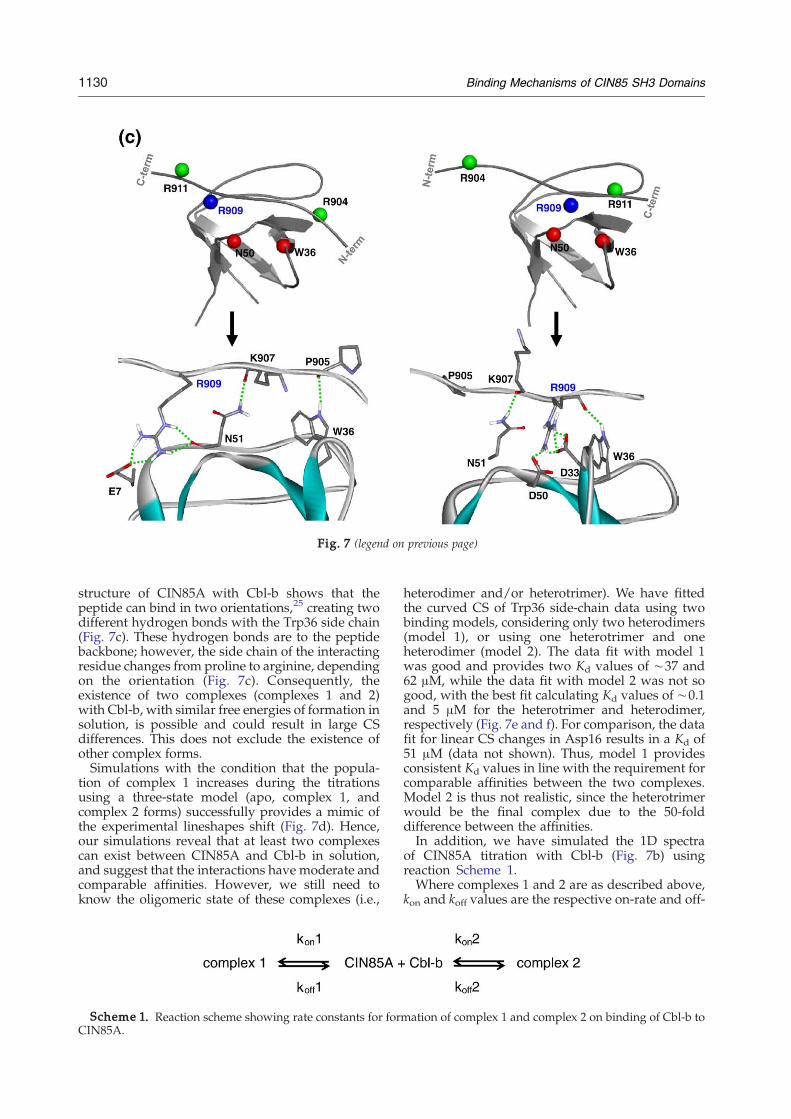
Fig. 7 (legend on previous page)
1130 Binding Mechanisms of CIN85 SH3 Domains
structure of CIN85A with Cbl-b shows that thepeptide can bind in two orientations,25 creating twodifferent hydrogen bonds with the Trp36 side chain(Fig. 7c). These hydrogen bonds are to the peptidebackbone; however, the side chain of the interactingresidue changes from proline to arginine, dependingon the orientation (Fig. 7c). Consequently, theexistence of two complexes (complexes 1 and 2)with Cbl-b, with similar free energies of formation insolution, is possible and could result in large CSdifferences. This does not exclude the existence ofother complex forms.Simulations with the condition that the popula-
tion of complex 1 increases during the titrationsusing a three-state model (apo, complex 1, andcomplex 2 forms) successfully provides a mimic ofthe experimental lineshapes shift (Fig. 7d). Hence,our simulations reveal that at least two complexescan exist between CIN85A and Cbl-b in solution,and suggest that the interactions have moderate andcomparable affinities. However, we still need toknow the oligomeric state of these complexes (i.e.,
Scheme 1. Reaction scheme showing rate constants for forCIN85A.
heterodimer and/or heterotrimer). We have fittedthe curved CS of Trp36 side-chain data using twobinding models, considering only two heterodimers(model 1), or using one heterotrimer and oneheterodimer (model 2). The data fit with model 1was good and provides two Kd values of ∼37 and62 μM, while the data fit with model 2 was not sogood, with the best fit calculating Kd values of ∼0.1and 5 μM for the heterotrimer and heterodimer,respectively (Fig. 7e and f). For comparison, the datafit for linear CS changes in Asp16 results in a Kd of51 μM (data not shown). Thus, model 1 providesconsistent Kd values in line with the requirement forcomparable affinities between the two complexes.Model 2 is thus not realistic, since the heterotrimerwould be the final complex due to the 50-folddifference between the affinities.In addition, we have simulated the 1D spectra
of CIN85A titration with Cbl-b (Fig. 7b) usingreaction Scheme 1.Where complexes 1 and 2 are as described above,
kon and koff values are the respective on-rate and off-
mation of complex 1 and complex 2 on binding of Cbl-b to

Fig. 8. Simulation of lineshapesof the HE1 resonance of Trp36during the titration of CIN85Awith Cbl-b. (a) Superposition ofseven steps of the titration takenat Cbl-b/CIN85A concentrationratios of 0.0, 0.2, 0.5, 0.7, 1.3, 1.8,and 2.5. The progress of the titra-tion is color coded, with the firststep in red, the last step in blue, andthe intervening steps appearing inan appropriate mixture of red andblue. The peaks of the free protein(S), the intermediate (I), and thefinal (F) complex are marked by ablack triangular symbol withappropriate labels. The individualpoints of the experimental spec-trum are shown as nonconnectedsquare symbols, and the simulatedspectra are shown as continuous
lines of the same color. Because of the strong overlap of the lineshapes, three key points of the titration are shownseparately in (b) at ratios of 0.0, (c) 0.2, and (d) 2.5. In (b)–(d), points from the experimental spectra are shown asunconnected blue squares, while the simulated spectra are shown as red continuous lines.
1131Binding Mechanisms of CIN85 SH3 Domains
rate constants for each complex. Figure 8 showsthat the lineshapes can be well described bythis mechanism scheme. The values for the rateconstants obtained from the simulated lineshapeswere 4.0×108 M−1s−1 and 117 s−1 for kon1 and koff1,respectively, and 3.4×108 M−1s−1 and 310 s−1 forkon2 and koff2, respectively. Interestingly, while thekon values were nearly similar for the complexes, thekoff values were different by a factor of 2.65.Furthermore, the calculatedKd values are in a similarrange of 0.3 and 0.9 μM for complexes 1 and 2,respectively, in line with those obtained from thefitting data of model 1 (37 and 62 μM). A detailedkinetic analysis of the interactions of domains ofCin85 will be presented elsewhere.Experimentally, the τc values for the apo and
saturated complexes of CIN85A with Cbl-b wereclearly consistent with monomeric CIN85A species(Table 2); however, to clarify the oligomeric state ofthe complex in solution, we investigated the com-plex formation at low and intermediate concentra-tions. We determined τc of CIN85A in the presenceof Cbl-b at ∼0.12, 0.25, and 0.55 Eq (Fig. 7g). The τcvalues are consistent with monomeric species ofCIN85A, since as Cbl-b concentration increases, itseems that there is only an averaging of τc betweenthe apo form and the heterodimer form. Since ourcalculated theoretical τc (τc
th) of the heterotrimer is∼14.2 ns (for the apo and heterodimer forms, the τc
th
values were 6.2 and 8.3 ns, respectively),42 if itexisted in solution over this Cbl-b concentrationrange, values higher than those reported here wouldbe observed (Fig. 7g). This clearly shows that duringthe course of complex formation, only the hetero-dimeric complexes exist.The binding of CIN85A to Cbl-b(F) results in the
formation of one heterodimeric complex population,similar to the binding of Cbl (Fig. 5). Thus, it is
intriguing that the presence of Arg909 seems to playa major role in the formation of another complexdespite the fact that its side chain does not appear toform hydrogen bonds or salt bridges with CIN85Aresidues in the crystal structure.25 Hence, for the firsttime, we report the importance of the Arg909residue in complex formation between CIN85Aand Cbl-b. At present, it is difficult to provideexperimental evidence to interpret this observation;however, an explanation is possible if we assumethat Arg909 forms hydrogen bonds with CIN85Aresidues in solution. Indeed, Arg909 side chain mayform hydrogen bonds with Asn51 and Glu7 (crystalstructure complex A) or Asp33 and Asp50 (complexB), depending on the peptide orientation, asrevealed by our energy-minimized structures25
(Fig. 7c). This shows the potential of Arg909 tocreate at least two different complex states usingdifferent hydrogen bonding networks. Thus, the Phemutation will abolish these hydrogen bonds, result-ing in one highly populated complex (Fig. 5).
CIN85A and CIN85B bind peptides with differentbinding mechanisms
CIN85A and CIN85B have a sequence similarity of∼47% and similar binding sites. Although CIN85Aand CIN85B form heterodimeric complexes withCbl and Cbl-b in solution, their binding mechanismsare strikingly dissimilar. First, we observe differ-ences in binding affinities and ΔHobs for theinteraction of the domains with peptides sharingsimilar binding core motifs and pseudo-symmetries(Table 1). Second, there is a different pattern in CSperturbations of the domains upon binding (Fig. 4).CIN85A shows low CS perturbations in comparisonto CIN85B, except for Asp16 and Asn51, and nosubstantial difference between the peptides, except

1132 Binding Mechanisms of CIN85 SH3 Domains
for small changes at the acidic pocket (Fig. 4).Change in the extent of CS perturbations is wellcorrelated to the binding affinity.43 However, thesmall increase in CS perturbations in the acidicpocket observed with CIN85A could not account forthe affinity differences on complex formationbetween these peptides. CIN85B, on the otherhand, shows higher CS perturbations upon bindingand a substantial decrease in the CS perturbationsfor Cbl-b binding at several residues (Fig. 4). Thisdecrease could be correlated to the very smallreduction in the binding affinity of Cbl-b. Themapping of residues above the ⟨Δδtot⟩+1σ thresh-old shows that CIN85B has a far higher number ofresidues interacting with the peptide than CIN85A(Fig. 4). This suggests that CIN85A has a ‘pre-formed’ binding site, while in CIN85B, there issubstantial side-chain rearrangement within thebinding site and more distal residues. Comparisonof the structures of the apo N-terminal SH3 domainof CMS (CMSA) to those of CIN85A in complex withCbl-b shows no conformational changes in the n-Srcor RT loops upon binding26 (CMS and CIN85 havesimilar domain compositions with 54% similarity inamino acid sequence and similar proposed func-tions1). Furthermore, structural overlap of theCIN85A monomers of the heterotrimer complexwith Cbl-b shows a very low backbone rmsd of0.11 Å and, importantly, the side-chain conforma-tions of all residues in the binding site areidentical.25 This is in line with the observed CSperturbation pattern for CIN85A. Third, severalresidues of CIN85A involved in the binding of Cbl-bshow curved CS changes, while CIN85B residues donot. We have provided evidence that at least twoheterodimer complexes of CIN85A form in solution(Fig. 7). A plausible way for this to happen is whenbinding takes the form of rigid-body docking,especially when the ligand has certain molecularsymmetry. Fourth, the difference between τc of apoCIN85A and that with either Cbl or Cbl-b complexes(Δτc) is slightly higher by ∼0.3 ns compared to Δτcfor CIN85B (Table 2). Only binding with looser-fitting peptides at the binding site would give rise tosuch an increase in τc. The X-ray structure ofCIN85A in complex with Cbl-b shows that mono-mer B has far less contact with Cbl-b than monomerA, and the electron density for three residues ateither termini of Cbl-b is absent.25 In contrast,CIN85B in complex with both peptides showsresonances matching those of CIN85AB (see Fig.S1a and b in Supplementary Materials). This isconsistent with binding via an induced fit of CIN85Bin isolation or as part of CIN85AB. These observa-tions suggest that CIN85A binds to the peptideswith a rigid-body mechanism while CIN85B uses aninduced-fit mechanism.
Behavior of CIN85 SH3 domains in solution andin vivo implications
A large and increasing number of binding partnershave been identified for CIN85, emphasizing the
multifunctional role of this protein. The majority ofthese interactions involve its SH3 domains.1,7,13,16Recent X-ray structures of both CIN85A and CMSAhave revealed heterotrimeric complex formationwith Cbl-b.25,26 Consequently, it was suggestedthat these heterotrimer complexes are important inthe clustering of Cbl proteins in vivo.25,26 We showthat in solution, CIN85A and CIN85B form 1:1complexes with Cbl and Cbl-b peptides. Further-more, we demonstrate that CIN85A is able to format least two complexes in solution with Cbl-bthrough a rigid-body binding mechanism. Hence,it is possible that the reported crystal structures ofthe heterotrimer complex represent a ‘trapping’ oftwo CIN85A molecules binding to one Cbl-b. Thisview is supported by the observation of bothheterodimeric and heterotrimeric complex struc-tures in two different crystals of the CMSA, withthe CD2 peptide containing the PXXXPR motif.26
Another complex between the SH3 domain fromcortactin and AMAP1 peptide containing a PXXXPRmotif was reported to be a heterotrimer in crystals.44
Our results, as well as previously reported data,17
clearly suggest that the observed crystal heterotrimercomplexes are not physiologically relevant. First, Cblproteins are known to homodimerize at their UBA/LZ domain.45 Our Cbl-b peptide motif is locatedvery close to the UBA/LZ domain, so that it isunlikely that heterotrimer formation is able to takeplace due to steric constraints. Second, in vivo CIN85exists as a dimer and/or a tetramer. This is likely toadd additional molecular constraints to the forma-tion of a possible trimeric complex with the intactCbl-b protein, particularlywhile the latter is engagedin other protein complexes.46 Third, in vivo experi-ments have often not mimicked physiological con-ditions. For example, deletion mutants of CIN85containing only the three SH3 domains are shown tointeract efficiently with proteins such as MEKK4,p85α, and Cbl; however, in the context of the full-length protein, the interaction is less efficient.9,19,24 Inaddition, in vivo experiments often fail to reproducethe effect of the C-terminal phosphorylation of Cbl-bon CIN85 binding.47
Finally, an interesting interaction mechanismbetween CIN85 and p85α has been suggested toinvolve CIN85A and CIN85B binding to the sameproline-rich motif PXXXPR of p85α.24 Although thepeptide sequence is different from Cbl-b and Cblpeptides, our results in solution show clearly thatCIN85AB binds to these peptides similarly as theisolated domains. Although the linker between thedomains in CIN85AB is long, the presence of fourprolines (Fig. 1b) near CIN85A may exist to preventCIN85A and CIN85B from binding the same motifin vivo.
Conclusion
We have provided evidence that, in solution, thefirst two SH3 domains of CIN85 form heterodimerswith Cbl and Cbl-b peptides in either isolated

1133Binding Mechanisms of CIN85 SH3 Domains
(CIN85A and CIN85B) or linked forms (CIN85AB).We have shown that these domains bind to thepeptides using different binding mechanisms,namely, a rigid-body mechanism for CIN85A andan induced fit for CIN85B. For the first time, wehave reported for an SH3 domain unusual curvedCS changes during the binding of Cbl-b peptide toCIN85A, revealing the existence of at least twoheterodimer complexes. We have correlated ourresults with the in vivo implications for CIN85functions. In summary, the manifold function ofCIN85 is dominated by discrete interactions of itsSH3 domains. They exhibit clear differences in theirmechanism of recognition of Cbl proteins, whichhave been reported to have different functions (seereview by Schmidt and Dikic46) and thereforeprovide the potential for specific recognition andindependent function of the domains.
Materials and Methods
Protein preparation
All protein constructs were cloned into the pET21bvector and verified by DNA sequencing. These wereoverexpressed in Escherichia coli strain BL21(DE3) at 37 °C.The proteins were purified using similar protocols, asdescribed previously.43 CIN85AB, however, was ex-pressed in inclusion bodies, so the cell pellet was washedwith 50 mM Tris buffer (pH 7.0), 70 mM NaCl, 5 mM β-mercaptoethanol, and 0.1% Triton X-100. The pellet wasthen dissolved and homogenized in the same buffer with8 M urea without Triton X-100. Cell debris was then spun,and the supernatant was collected for protein refolding.The latter was achieved by successive dialysis in the urea-containing buffer at 4 M urea, 2 M urea, and, finally, 0 Murea. CIN85AB protein was then purified using the sameprotocol as the other soluble polypeptides. Isotope-enriched samples of all proteins were obtained by growingthe bacteria in M9 minimal medium supplemented with15N-ammonium chloride (Cambridge Isotope Labora-tories, Andover, MA) and/or uniformly 13C-enrichedglucose as the unique sources of nitrogen and carbon,respectively.
Isothermal titration calorimetry
Titrations were carried out on a VP-ITC instrument(MicroCal, Inc., Northampton, MA). The peptides were c-Cbl (or Cbl), SQVPERPPKPFPRRINSY, and Cbl-b, SQA-PARPPKPRPRRTAY. The C-terminal tyrosines were intro-duced for concentration determination, and bold lettersrepresent residues that were mutated, as described in thetext. CIN85A, CIN85B, and CIN85AB (20–50 μM) weretitrated with the peptides (500–1000 μM) in 50 mM Tris(pH 7.5) and 50mMNaCl at 25 °C. The titrations consistedof a preliminary 2-μl injection (neglected in data analysis),followed by 24 subsequent 12-μl injections. Four minuteswere left between each injection. Binding data werecorrected by subtraction of the mean value of the heat ofdilution data. Corrected binding data were processedusing the Origin ITC analysis software package suppliedby MicroCal, Inc. The similarity of the affinities of the twoindependent binding events on binding of peptides toCIN85AB meant that deconvolution of the data for the
individual interactions was not possible at the experi-mental temperature.
NMR spectroscopy
NMR data were collected with 0.5–0.8 mM proteinsamples containing 20 mM phosphate buffer (pH 6.3),100 mM NaCl, 2 mM dithiothreitol, 1 mM ethylenediami-netetraacetic acid, and 0.01% NaN3. Each sample wasprepared in 95% (vol/vol) H2O/5% D2O. NMR spectrawere acquired at 25 °C on either a 600-MHz BrukerAVANCEDRX spectrometer, a 500-MHzVarianUnity Plusspectrometer, or a 600-MHz Varian Inova spectrometer.The 1H, 15N, and 13C assignmentsweremade as previouslyreported.43 Relaxation analysis was performed bymeasur-ing 15N R1, R2, and
1H–15N heteronuclear NOE experi-ments on 15N-labeled CIN85A, CIN85B, and CIN85ABprotein samples at 0.35–0.65 mM protein concentrations at25 °C and analyzed as previously reported.43 For CIN85Aand CIN85B, R1 was measured with delays of 51, 202, 302,403 (twice), 503, 603, 805, 1006, 1408, and 1810 ms, and R2was measured with delays of 16, 31, 47, 62 (twice), 94, 125,156, 187, 219, and 250 ms. For CIN85AB, R1 was measuredwith delays of 16, 48, 96, 192, 288 (twice), 384, 512, 704, 960,and 1200ms, andR2 wasmeasuredwith delays of 5.1, 10.3,15.4, 20.5 (twice), 30.8, 41.0, 61.5, 82.0, 112.8, and 143.5 ms.Heteronuclear NOE was measured with a proton satura-tion period of 3 s. We collected total data points of 18–20steps for each NMR titration of the proteins with thepeptides. To calculate the total average change in backboneamide 1H and 15N CS for each resonance, the followingequation was applied: Δδtot = {[0.17Δδ(15N)]2 + [Δδ(1H)]2}1/2, where Δδ(15N) and Δδ(1H) are the CS changes.The change in total CS was then calculated for eachidentified residue, with the exception of those whoseresonances had broadened past detectability in the 1H–15NHSQC NMR spectra. The average total CS ⟨Δδtot⟩ andstandard deviation 1σ associated with each data set werealso calculated.The NMR simulations of exchanging lineshapes were
performed using WINDNMR software (by Dr. Hans J.Reich); however, we used programs written in Mathe-matica (Wolfram Research) for simulations in slow-exchange regime for comparison. The equations adoptedfor the two-state model calculations were previouslydescribed.48 For the three-state model, the Kubo–Sackmatrix approach was used to simulate the lineshapes.49
We performed extensive tests to show that the largedifferences in CS between different complex states dictatethe extent of curvature of the CS changes observedduring the titration in our NMR simulations. The data fitof the curved CS changes was performed for model 1(competition between two heterodimer complexes) usingthe equation: δ=∑i=1,2δi,min+ (δi,max−δi,min)[((L+Kdi+P)− ((L+Kdi+P)
2−4LP))1/2]/2P, where L and P are the totalconcentrations of the ligand and protein, respectively, andKdi is the dissociation constant for each heterodimer. Formodel 2 (competition between heterotrimer and hetero-dimer complexes), we have used Mathematica to solve thesystem of equations: K1=LP
2/T, K2=LP/D, Ptot=P+2⁎T+D, and Ltot=L+T+D, where Ptot and Ltot are the totalconcentrations of the protein and the ligand. L, P, D, and Tare the free concentrations of the ligand, protein, dimer,and trimer, respectively. We then implemented a fittingroutine where we substituted the solutions for T and Dinto the equation: δ=(T/Ptot)δT+((Ptot−T−D)/Ptot)δP+(D/Ptot)δD, where δT, δP, and δD are the CS of the trimer,free protein, and dimer, respectively.

1134 Binding Mechanisms of CIN85 SH3 Domains
Lineshape analysis was achieved using Bloch equationsfor a three-state exchanging system according to Scheme 1(see Discussion) and prepared as a system of coupledequations as described previously50 and solved inMathematica using the DSolve function. A series ofseven free induction decays for the titration was calculatedfor an acquisition time of 1.0 s and a dwell time of 100 μs,followed by Fourier transformation for a comparison ofsimulated with experimental spectra.51 CS for the freeprotein (41.5 Hz) and complex 1 (52.2 Hz), as well as theintrinsic linewidths for these species (30 and 40.0 s−1,respectively), were fixed as obtained from the first and laststeps of the titration, while the CS of complex 2, the fourkinetic constants, and the intrinsic linewidth of complex 2were optimized to best reproduce the experimental data.Simulated and recorded spectra are displayed super-imposed as suggested previously.51,52
Structure calculation
Weperformed simulated annealingmolecular dynamicsusing ARIA protocol (version 1.2),53 as previouslyreported.43 A total of 315 unambiguous proton–protondistances from NOESY spectra were used as a starting setof distance constraints together with 15 hydrogen bondconstraints for protons in residues having appropriate β-sheet-specific interstrand NOEs and 70 ϕ and ψ dihedralangle constraints from CS analysis by TALOS.54 Dihedralrestraints were only added for residues that had asubstantial number of NOE distances and were thusassumed to be in awell-defined conformation. UnassignedNOEs (totaling 2275) from three-dimensional 15N and 13CNOESY-HSQC experiments were introduced as ambig-uous distance constraints. After eight cycles of ARIA, 1323of these were unambiguously assigned. After the finaliteration, structures with no violation of NOE constraintsabove 0.5 Å were selected. A total of 27 structures fulfilledthis criterion.
Acknowledgements
We thank Ivan Dikic for providing the proteinconstructs. The authors are grateful to DidierPhilippe and Bhaumik Dave for the gift of TEV-cleaved 15N-labeled CIN85A. This work wassupported by a Wellcome Trust Senior ResearchFellowship (J.E.L. and A.A.) and BBSRC grant BB/D018994/1 (M.P.).
Supplementary Data
Supplementary data associated with this articlecan be found, in the online version, at doi:10.1016/j.jmb.2009.02.061
References
1. Dikic, I. (2002). CIN85/CMS family of adaptormolecules. FEBS Lett. 529, 110–115.
2. Petrelli, A., Gilestro, G. F., Lanzardo, S., Comoglio, P. M.,Migone, N. & Giordano, S. (2002). The endophilin–
CIN85–Cbl complex mediates ligand-dependentdownregulation of c-Met. Nature, 416, 187–190.
3. Soubeyran, P., Kowanetz, K., Szymkiewicz, I., Lang-don, W. Y. & Dikic, I. (2002). Cbl–CIN85–endophilincomplex mediates ligand-induced downregulation ofEGF receptors. Nature, 416, 183–187.
4. Chen, B., Borinstein, S. C., Gillis, J., Sykes, V. W. &Bogler, O. (2000). The glioma-associated proteinSETA interacts with AIP1/Alix and ALG-2 andmodulates apoptosis in astrocytes. J. Biol. Chem. 275,19275–19281.
5. Gout, I., Middleton, G., Adu, J., Ninkina, N. N.,Drobot, L. B., Filonenko, V. et al. (2000). Negativeregulation of PI 3-kinase by Ruk, a novel adaptorprotein. EMBO J. 19, 4015–4025.
6. Dustin, M. L., Olszowy, M. W., Holdorf, A. D., Li,J., Bromley, S., Desai, N. et al. (1998). A noveladaptor protein orchestrates receptor patterningand cytoskeletal polarity in T-cell contacts. Cell, 94,667–677.
7. Kowanetz, K., Husnjak, K., Holler, D., Kowanetz, M.,Soubeyran, P., Hirsch, D. et al. (2004). CIN85 associateswith multiple effectors controlling intracellular traf-ficking of epidermal growth factor receptors.Mol. Biol.Cell, 15, 3155–3166.
8. Lynch, D. K., Winata, S. C., Lyons, R. J., Hughes, W. E.,Lehrbach, G. M., Wasinger, V. et al. (2003). A cortactin–CD2-associated protein (CD2AP) complex provides anovel link between epidermal growth factor receptorendocytosis and the actin cytoskeleton. J. Biol. Chem.278, 21805–21813.
9. Aissouni, Y., Zapart, G., Iovanna, J. L., Dikic, I. &Soubeyran, P. (2005). CIN85 regulates the ability ofMEKK4 to activate the p38 MAP kinase pathway.Biochem. Biophys. Res. Commun. 338, 808–814.
10. Gaidos, G., Soni, S., Oswald, D. J., Toselli, P. A. &Kirsch, K. H. (2007). Structure and function analysis ofthe CMS/CIN85 protein family identifies actin-bund-ling properties and heterotypic-complex formation. J.Cell Sci. 120, 2366–2377.
11. Schmidt, M. H., Chen, B., Randazzo, L. M. & Bogler,O. (2003). SETA/CIN85/Ruk and its binding partnerAIP1 associate with diverse cytoskeletal elements,including FAKs, and modulate cell adhesion. J. CellSci. 116, 2845–2855.
12. Watanabe, S., Take, H., Takeda, K., Yu, Z. X., Iwata, N.& Kajigaya, S. (2000). Characterization of the CIN85adaptor protein and identification of componentsinvolved in CIN85 complexes. Biochem. Biophys. Res.Commun. 278, 167–174.
13. Le Bras, S., Moon, C., Foucault, I., Breittmayer, J. P. &Deckert, M. (2007). Abl-SH3 binding protein 2, 3BP2,interacts with CIN85 and HIP-55. FEBS Lett. 581,967–974.
14. Liang, Y., Kurakin, A. & Roizman, B. (2005). Herpessimplex virus 1 infected cell protein 0 forms a complexwith CIN85 and Cbl and mediates the degradation ofEGF receptor from cell surfaces. Proc. Natl Acad. Sci.USA, 102, 5838–5843.
15. Tossidou, I., Kardinal, C., Peters, I., Kriz, W., Shaw, A.,Dikic, I. et al. (2007). CD2AP/CIN85 balance deter-mines receptor tyrosine kinase signaling response inpodocytes. J. Biol. Chem. 282, 7457–7464.
16. Nam, J. M., Onodera, Y., Mazaki, Y., Miyoshi, H.,Hashimoto, S. & Sabe, H. (2007). CIN85, a Cbl-interacting protein, is a component of AMAP1-mediated breast cancer invasion machinery. EMBO J.26, 647–656.
17. Kowanetz, K., Szymkiewicz, I., Haglund, K.,

1135Binding Mechanisms of CIN85 SH3 Domains
Kowanetz, M., Husnjak, K., Taylor, J. D. et al. (2003).Identification of a novel proline–arginine motifinvolved in CIN85-dependent clustering of Cbl anddown-regulation of epidermal growth factor recep-tors. J. Biol. Chem. 278, 39735–39746.
18. Buchman, V. L., Luke, C., Borthwick, E. B., Gout, I. &Ninkina, N. (2002). Organization of the mouse Ruklocus and expression of isoforms in mouse tissues.Gene, 295, 13–17.
19. Tibaldi, E. V. & Reinherz, E. L. (2003). CD2BP3,CIN85 and the structurally related adaptor proteinCMS bind to the same CD2 cytoplasmic segment, butelicit divergent functional activities. Int. Immunol. 15,313–329.
20. Ball, L. J., Kuhne, R., Schneider-Mergener, J. &Oschkinat, H. (2005). Recognition of proline-richmotifs by protein–protein-interaction domains.Angew. Chem. Int. Ed. Engl. 44, 2852–2869.
21. Mayer, B. J. (2001). SH3 domains: complexity inmoderation. J. Cell Sci. 114, 1253–1263.
22. Groemping, Y., Lapouge, K., Smerdon, S. J. &Rittinger, K. (2003). Molecular basis of phosphoryla-tion-induced activation of the NADPH oxidase. Cell,113, 343–355.
23. Ogura, K., Nobuhisa, I., Yuzawa, S., Takeya, R.,Torikai, S., Saikawa, K. et al. (2006). NMR solutionstructure of the tandem Src homology 3 domains ofp47phox complexed with a p22phox-derived proline-rich peptide. J. Biol. Chem. 281, 3660–3668.
24. Borthwick, E. B., Korobko, I. V., Luke, C., Drel, V. R.,Fedyshyn, Y. Y., Ninkina, N. et al. (2004). Multipledomains of Ruk/CIN85/SETA/CD2BP3 are involvedin interaction with p85alpha regulatory subunit of PI3-kinase. J. Mol. Biol. 343, 1135–1146.
25. Jozic, D., Cardenes, N., Deribe, Y. L., Moncalian, G.,Hoeller, D., Groemping, Y. et al. (2005). Cbl promotesclustering of endocytic adaptor proteins. Nat. Struct.Mol. Biol. 12, 972–979.
26. Moncalian, G., Cardenes, N., Deribe, Y. L., Spinola-Amilibia, M., Dikic, I. & Bravo, J. (2006). Atypicalpolyproline recognition by the CMS N-terminal Srchomology 3 domain. J. Biol. Chem. 281, 38845–38853.
27. Song, J. & Markley, J. L. (2007). Cautionary tail: thepresence of an N-terminal tag on dynein light-chainRoadblock/LC7 affects its interaction with a func-tional partner. Protein Pept. Lett. 14, 265–268.
28. Ababou, A., Shenvi, R. A. & Desjarlais, J. R. (2001).Long range calcium binding effect and conformationalchange in the N-domain of calmodulin. Biochemistry,40, 12719–12726.
29. Ababou, A., Pfuhl, M. & Ladbury, J. E. (2008). Thebinding stoichiometry of CIN85 SH3 domain A andCbl-b. Nat. Struct. Mol. Biol. 15, 890–891.
30. Ghose, R., Shekhtman, A., Goger, M. J., Ji, H. &Cowburn, D. (2001). A novel, specific interactioninvolving the Csk SH3 domain and its natural ligand.Nat. Struct. Biol. 8, 998–1004.
31. Sarkar, P., Reichman, C., Saleh, T., Birge, R. B. &Kalodimos, C. G. (2007). Proline cis–trans isomeriza-tion controls autoinhibition of a signaling protein.Mol.Cell, 25, 413–426.
32. Fernandez-Ballester, G., Blanes-Mira, C. & Serrano, L.(2004). The tryptophan switch: changing ligand-bind-ing specificity from type I to type II in SH3 domains. J.Mol. Biol. 335, 619–629.
33. Tolkatchev, D., Xu, P. & Ni, F. (2003). Probing thekinetic landscape of transient peptide–protein inter-actions by use of peptide (15)N NMR relaxationdispersion spectroscopy: binding of an antithrombin
peptide to human prothrombin. J. Am. Chem. Soc. 125,12432–12442.
34. Kurakin, A. V., Wu, S. & Bredesen, D. E. (2003).Atypical recognition consensus of CIN85/SETA/RukSH3 domains revealed by target-assisted iterativescreening. J. Biol. Chem. 278, 34102–34109.
35. Marsden, B. J., Hodges, R. S. & Sykes, B. D. (1988).1H-NMR studies of synthetic peptide analogues ofcalcium-binding site III of rabbit skeletal troponin C:effect on the lanthanum affinity of the interchangeof aspartic acid and asparagine residues at themetal ion coordinating positions. Biochemistry, 27,4198–4206.
36. Sugase, K., Dyson, H. J. & Wright, P. E. (2007).Mechanism of coupled folding and binding of anintrinsically disordered protein.Nature, 447, 1021–1025.
37. Velyvis, A., Yang, Y. R., Schachman, H. K. & Kay, L. E.(2007). A solution NMR study showing that active siteligands and nucleotides directly perturb the allostericequilibrium in aspartate transcarbamoylase. Proc. NatlAcad. Sci. USA, 104, 8815–8820.
38. Baxter, N. J., Hosszu, L. L., Waltho, J. P. & Williamson,M. P. (1998). Characterisation of low free-energyexcited states of folded proteins. J. Mol. Biol. 284,1625–1639.
39. Asakura, T., Taoka, K., Demura, M. & Williamson,M.P. (1995). The relationship between amide protonchemical shifts and secondary structure in proteins. J.Biomol. NMR, 6, 227–236.
40. Grzesiek, S., Cordier, F., Javarine, V. & Barfield, M.(2004). Insights into biomolecular hydrogen bondsfrom hydrogen bond scalar couplings. Prog. Nucl.Magn. Reson. Spectrosc. 45, 275–300.
41. Ababou, A., van der Vaart, A., Gogonea, V. & Merz,K.M., Jr (2007). Interaction energy decomposition inprotein–protein association: a quantum mechanicalstudy of barnase–barstar complex. Biophys. Chem. 125,221–236.
42. de la Torre, J. G., Huertas, M. L. & Carrasco, B.(2000). HYDRONMR: prediction of NMR relaxationof globular proteins from atomic-level structuresand hydrodynamic calculations. J. Magn. Reson. 147,138–146.
43. Ababou, A., Gautel, M. & Pfuhl, M. (2007). Dissectingthe N-terminal myosin binding site of humancardiac myosin-binding protein C. Structure andmyosin binding of domain C2. J. Biol. Chem. 282,9204–9215.
44. Hashimoto, S., Hirose, M., Hashimoto, A., Morishige,M., Yamada, A., Hosaka, H. et al. (2006). TargetingAMAP1 and cortactin binding bearing an atypical srchomology 3/proline interface for prevention of breastcancer invasion and metastasis. Proc. Natl Acad. Sci.USA, 103, 7036–7041.
45. Bartkiewicz, M., Houghton, A. & Baron, R. (1999).Leucine zipper-mediated homodimerization of theadaptor protein c-Cbl. A role in c-Cbl's tyrosinephosphorylation and its association with epidermalgrowth factor receptor. J. Biol. Chem. 274, 30887–30895.
46. Schmidt, M. H. & Dikic, I. (2005). The Cbl inter-actome and its functions. Nat. Rev. Mol. Cell Biol. 6,907–919.
47. Szymkiewicz, I., Kowanetz, K., Soubeyran, P.,Dinarina, A., Lipkowitz, S. & Dikic, I. (2002).CIN85 participates in Cbl-b-mediated down-regula-tion of receptor tyrosine kinases. J. Biol. Chem. 277,39666–39672.
48. Sandstrom, J. (1982). Dynamic NMR Spectroscopy.Academic Press, New York.

1136 Binding Mechanisms of CIN85 SH3 Domains
49. Johnson, C. S. & Moreland, C. G. (1973). Thecalculation of NMR spectra for many-site exchangeproblems. J. Chem. Ed. 50, 477–483.
50. Cavanagh, J., Fairbrother, W. I., Palmer, A. G.,Skelton, B. I. & Rance, M. (2006). Protein NMRSpectroscopy: Principles and Practice. Academic Press,San Diego, CA.
51. Günther, U. L. & Schaffhausen, B. (2002). NMRKIN:simulating line shapes from two-dimensional spectraof proteins upon ligand binding. J. Biomol. NMR, 22,201–209.
52. Mittag, T., Schaffhausen, B. & Günther, U. L. (2004).Tracing kinetic intermediates during ligand binding. J.Am. Chem. Soc. 126, 9017–9023.
53. Linge, J. P., O'Donoghue, S. I. & Nilges, M. (2001).Automated assignment of ambiguous nuclear Over-hauser effects with ARIA. Methods Enzymol. 339,71–90.
54. Cornilescu, G., Delaglio, F. & Bax, A. (1999). Proteinbackbone angle restraints from searching a databasefor chemical shift and sequence homology. J. Biomol.NMR, 13, 289–302.




















