
NONVIRAL GENE TRANSFER INTO PRIMARY CULTURES OF HUMAN AND PORCINE MESOTHELIAL CELLS
6
In Vitro Cell. Dev. Biol.~Anima137:402-~07, July/August 2001 2001 Societyfor In VitroBiology 1071-2690/01 $10.00+0.00 NONVIRAL GENE TRANSFER INTO PRIMARY CULTURES OF HUMAN AND PORCINE MESOTHELIAL CELLS J. OHAN, 1 M. A. GILBERT, G. LESECHE, Y. PANIS, E MIDOUX, AND L. DROUET Laboratoire de Chirurgie Vasculaire et Thoracique (J. 0., G. L.), INSERM 353 (M. A. G.), INSERM 450 et Service d'Hdmatologie de l'h6pital lariboisi&e (L. D.), Service de Chirurgie C~m~rale de l'hOpital lariboisi~re (Y. P.), Paris, France, and Centre de Biophysique Moldculaire, Orldans, France (P. M.) (Received 14 December 2000; accepted 31 March 2001) SUMMARY Due to their abundance and accessibility, mesothelial cells may be suitable tools for recombinant reagent expression by gene transfer. Genetically modified porcine mesothelial cells (PMCs) may have the potential for the treatment of vascular diseases in humans. We studied the effect of various transfection reagents on the primary culture of PMCs and human mesothelial cells (HMCs). The cells were transfected with a plasmid encoding a reporter gene (luciferase or green fluorescent protein [GFP]) under the control of the cytomegalovirus promoter. Transfection was achieved using cationic lipids (DOSPER and DOTAP) or calcium phosphate/deoxyribonucleic acid coprecipitation or Fugene 6. Results showed that Fugene 6 was the most efficient and reproducible transfection reagent with both PMCs and HMCs. With Fugene 6, luciferase activity in PMCs (1.5 • 108 relative light units [RLU]/10 ~' cells) was at least 2.5-fold higher than with the other transfection reagents, and it was 100-fold higher than in HMCs. Howevel, the proportion of transfected ceils expressing GFP was only 1%. These preliminary findings open up new avenues for developing experimental studies on the use of genetically modified PMCs. Key words: gene therapy; gene transfer; nonviral vector; green fluorescent protein; luciferase; mesothelial cells. INTRODUCTION Seeding with endothelial cells (ECs) has been suggested as a means of improving the hemostatic performance of vascular pros- theses. ECs can be modified genetically to increase their antico- agulant and fibrinolytic properties and, therefore, their potential for preventing thrombosis within the vascular prostheses. How- ever, genetically modified ECs have a low rate of surface endoth- elialization, a feature associated with a weaker adherence to the prostheses after exposure to in vivo hemodynamic forces (Thomas et al., 1995; Sackman et al., 1995; Sugawara, 1997). In addition, seeding requires a large number of ECs. It has been suggested that microvascular ECs isolated from human capillaries may be a good substitute for ECs and may be available in abundance from the omentum, a tissue rich in capillaries (Les~che and Ohan, 1995; Pasic et al., 1995; Bordenave et al., 1999). Experience has taught, however, that the human omentum yields few microvas- cular ECs, but is a plentiful source of mesothelial cells. Human mesothelial cells (HMCs) have fibrinolytic properties and grow in monolayers similar to the endothelium, two desirable character- istics in candidates for seeding the vascular prosthesis with the goal of reducing thrombosis (van Hinsbergh et al., 1990; Pronk et al., 1993; Ivarsson et al., 1998). In addition, because of their abundance, location, and accessibility, HMCs can be modified ge- netically so that they deliver recombinant proteins directly into To whom correspondence should be addressed at Laboratoire de Chi- rurgie Vasculaire et Thoracique, H6pital BEAUJON, 100 Boulevard du G6n- 6ral Leclerc, 92100 Clichy, France. E-mail: [email protected] the systemic circulation. This ex vivo approach has been tested in small animals including rats and mice (Nagy et al., 1994, 1995a, 1995b; Jackman et al., 1998). The main studies of vascular prosthesis seeding in large animals have been conducted on dogs, a species with differences from humans, including a higher plate- let count and different reactivities of several coagulation and fi- brinolytic factors and of their inhibitors. In contrast, pigs share similar physiological characteristics with humans (Dodds, 1992; Schwartz et al., 1994). To assist in creating a large animal model of vascular disease for investigating gene therapy, we developed a technique for isolating porcine mesothelial cells (PMCs), and dem- onstrated that these cells exhibit phenotypic and functional char- acteristics similar to those of HMCs (Ohan et al., 1999). In the present study, we compared various transfection reagents for trans- ferring a reporter gene into the cultures of PMCs and HMCs. We found that Fugene 6 was the best transfection reagent, and that PMCs were more efficiently transfected than HMCs. These results are a first step in the development of an ex-vivo gene therapy strategy based on genetically modified mesothelial cells from the porcine omentum. MATERIALS AND METHODS Animals. Pieces of porcine omentum were removed surgically from 20 adult (6-mo-old) anesthetized large, white pigs. The animals were killed, and the tissues were placed in phosphate buffer saline (PBS) containing 1 mM Ca ++ and 0.5 mM Mg ++ (PBS+) at 4 ~ C. The mesothelial ceils were isolated on the arrival of the specimens at the laboratory after a 2-h journey. Cell isolation. HMCs were isolated from small pieces of omentum removed 402
Transcript of NONVIRAL GENE TRANSFER INTO PRIMARY CULTURES OF HUMAN AND PORCINE MESOTHELIAL CELLS

In Vitro Cell. Dev. Biol.~Anima137:402-~07, July/August 2001 �9 2001 Society for In Vitro Biology 1071-2690/01 $10.00+0.00
NONVIRAL GENE TRANSFER INTO PRIMARY CULTURES OF HUMAN AND PORCINE MESOTHELIAL CELLS
J. OHAN, 1 M. A. GILBERT, G. LESECHE, Y. PANIS, E MIDOUX, AND L. DROUET
Laboratoire de Chirurgie Vasculaire et Thoracique (J. 0., G. L.), INSERM 353 (M. A. G.), INSERM 450 et Service d'Hdmatologie de l'h6pital lariboisi&e (L. D.), Service de Chirurgie C~m~rale de l'hOpital lariboisi~re (Y. P.), Paris, France, and
Centre de Biophysique Moldculaire, Orldans, France (P. M.)
(Received 14 December 2000; accepted 31 March 2001)
SUMMARY
Due to their abundance and accessibility, mesothelial cells may be suitable tools for recombinant reagent expression by gene transfer. Genetically modified porcine mesothelial cells (PMCs) may have the potential for the treatment of vascular diseases in humans. We studied the effect of various transfection reagents on the primary culture of PMCs and human mesothelial cells (HMCs). The cells were transfected with a plasmid encoding a reporter gene (luciferase or green fluorescent protein [GFP]) under the control of the cytomegalovirus promoter. Transfection was achieved using cationic lipids (DOSPER and DOTAP) or calcium phosphate/deoxyribonucleic acid coprecipitation or Fugene 6. Results showed that Fugene 6 was the most efficient and reproducible transfection reagent with both PMCs and HMCs. With Fugene 6, luciferase activity in PMCs (1.5 • 108 relative light units [RLU]/10 ~' cells) was at least 2.5-fold higher than with the other transfection reagents, and it was 100-fold higher than in HMCs. Howevel, the proportion of transfected ceils expressing GFP was only 1%. These preliminary findings open up new avenues for developing experimental studies on
the use of genetically modified PMCs.
Key words: gene therapy; gene transfer; nonviral vector; green fluorescent protein; luciferase; mesothelial cells.
INTRODUCTION
Seeding with endothelial cells (ECs) has been suggested as a means of improving the hemostatic performance of vascular pros- theses. ECs can be modified genetically to increase their antico- agulant and fibrinolytic properties and, therefore, their potential for preventing thrombosis within the vascular prostheses. How- ever, genetically modified ECs have a low rate of surface endoth- elialization, a feature associated with a weaker adherence to the prostheses after exposure to in vivo hemodynamic forces (Thomas et al., 1995; Sackman et al., 1995; Sugawara, 1997). In addition, seeding requires a large number of ECs. It has been suggested that microvascular ECs isolated from human capillaries may be a good substitute for ECs and may be available in abundance from the omentum, a tissue rich in capillaries (Les~che and Ohan, 1995; Pasic et al., 1995; Bordenave et al., 1999). Experience has taught, however, that the human omentum yields few microvas- cular ECs, but is a plentiful source of mesothelial cells. Human mesothelial cells (HMCs) have fibrinolytic properties and grow in monolayers similar to the endothelium, two desirable character- istics in candidates for seeding the vascular prosthesis with the goal of reducing thrombosis (van Hinsbergh et al., 1990; Pronk et al., 1993; Ivarsson et al., 1998). In addition, because of their abundance, location, and accessibility, HMCs can be modified ge- netically so that they deliver recombinant proteins directly into
To whom correspondence should be addressed at Laboratoire de Chi- rurgie Vasculaire et Thoracique, H6pital BEAUJON, 100 Boulevard du G6n- 6ral Leclerc, 92100 Clichy, France. E-mail: [email protected]
the systemic circulation. This ex vivo approach has been tested in small animals including rats and mice (Nagy et al., 1994, 1995a, 1995b; Jackman et al., 1998). The main studies of vascular prosthesis seeding in large animals have been conducted on dogs, a species with differences from humans, including a higher plate- let count and different reactivities of several coagulation and fi- brinolytic factors and of their inhibitors. In contrast, pigs share similar physiological characteristics with humans (Dodds, 1992; Schwartz et al., 1994). To assist in creating a large animal model of vascular disease for investigating gene therapy, we developed a technique for isolating porcine mesothelial cells (PMCs), and dem- onstrated that these cells exhibit phenotypic and functional char- acteristics similar to those of HMCs (Ohan et al., 1999). In the present study, we compared various transfection reagents for trans- ferring a reporter gene into the cultures of PMCs and HMCs. We found that Fugene 6 was the best transfection reagent, and that PMCs were more efficiently transfected than HMCs. These results are a first step in the development of an ex-vivo gene therapy strategy based on genetically modified mesothelial cells from the porcine omentum.
MATERIALS AND METHODS
Animals. Pieces of porcine omentum were removed surgically from 20 adult (6-mo-old) anesthetized large, white pigs. The animals were killed, and the tissues were placed in phosphate buffer saline (PBS) containing 1 mM Ca ++ and 0.5 mM Mg ++ (PBS+) at 4 ~ C. The mesothelial ceils were isolated on the arrival of the specimens at the laboratory after a 2-h journey.
Cell isolation. HMCs were isolated from small pieces of omentum removed
4 0 2

PMC AND HMC FOR GENE TRANSFER 403
from patients undergoing abdominal surgery for noninfectious conditions. The sampling protocol was approved by the appropriate ethics committee, and written informed consent was obtained from each patient prior to the pro- cedure. Cells were isolated using our adaptation (Dodds, 1992) of the method of Kern et al. (1983). Briefly, omentum was rinsed in Medium 199 (M199) (GIBCO-BRL, Eragny, France), minced, and incubated under gentle shaking for 30 min at 37 ~ C in 0.25% collagenase clostridiopeptidase A (GIBCO-BRL) diluted in PBS (0.1 M sodium phosphate buffer, pH = 7.4, in 0.15 M NaC1, 1:10 v/v). The cell suspension was filtered and centrifuged at 240 • g for 5 min. The cells were resuspended in 5 nil of PBS, layered on to diluted Ficoll* (10:2 v/v; Sigma, Saint Quentin Fallauier, France), and centrifuged at 128 x g for 3 min. The upper 3 ml of the supernatant was discarded, and the remaining supernatant was centrifuged at 240 • g for 5 lnin. After suspension of the pellet in growtfi medium, the cells were seeded on Falcon 25-cm ~ flasks (Becton Dickinson, Le Pont de Claix, France) pre- viously coated with human fibronectin (GIBCO-BRL). Cell viability was de- termined by trypan blue dye exclusion immediately after the isolation pro- cedure.
PMCs were isolated from adult pig omentum as previously reported (Ohan et al., 1999). Briefly, the isolation procedure was as described above, with the following modifications: the enzymatic digestion period was 45 rain, and the cell pellet suspended in PBS was layered on native Ficoll.
Cell culture. HMCs and PMCs seeded on 25-cm 2 flasks coated with hu- man fibronectin (0.5 mg/cm ~) were cultured in M199 containing 20% fetal calf serum (FCS) (Boehringer Mannheim, Meylan, France), 2 m_M L-gluta- mine, 2.5 mg/ml fi_mgizone, and 50 txg/ml gentamicin (GIBCO-BRL). Con- fluent cells were washed twice with PBS, harvested with 0.05% trypsin- 0.02% ethylenediamine-tetraacetic acid (EDTA) (GIBCO-BRL), and seeded at a split ratio of 1/2. The cells we~'e cultured at 37 ~ C in a humidified atnlospherc containing 5% C02 and 95% air. All the experiments were performed with primary cells at passage 2.
Cell characterization. HMCs and PMCs from primary confluent cultures were examined under a light microscope and photographed using light mi- croscopy (Axiplan Zeiss, Le Pecq, France).
Ultrastructural study was done with confluent primary PMCs and HMCs grown in 35-mm fibronectin-coated Petri dishes (9 cm 2) (Becton Dickinson) which were fixed for 1 h at 4 ~ C with 2.5% glutaraldehyde in 0.1 M cac- odylate buffer (pH 7.4), postfixed for 1 h at room temperature in 1% os- mium tetroxide, dehydrated with increasing concentrations of ethanol, and subjected to two-stage embedding in LX-112 resin (LADD Research In- dustries, Burlington, VT). Ultrathin sections (700 nm) were stained with uranyl acetate and lead citrate. All sections were examined under an elec- tron microscope (EM109, Zeiss).
For immunofluorescent staining, cells were fixed for 10 rain in 3% para- formaldehyde and permeabilized with 0.1% Triton-X 100 (Sigma) for 10 min at room temperature. Activated sites were quenched with bovine serum albumin fraction V (Sigma) (0.1% in PBS+) for 20 min. The cells were washed with PBS and incubated for 1 h at room temperature with specific monoclonal antibodies to vimentin (clone: V9-1/25), human cyto- keratin 8 (clone: 35bHl1-1/25), human cytokeratin 18 (MNF6-1/25), HBME1 (1/50) (Dako SA, Trappe, France), and ME1 (1/50, Interchim, As- nitres, France) or the nuclear transcription factors WT1 (a generous gift from Dr. Rougier, INSERM U64 HSpital Tenon, Paris, France). Finally, the (:ells were incubated with fluorescein isothiocyanate-conjugated goat anti- mouse IgG antibody (1:50 dilution) (Silenus, Melbourne, Australia). After extensive washings, the cells were incubated for 10 min with 1,4-diaza bicyclo(2,2,2)actane triethylenedimnine (Dako) to preserve the fluores- cence, mounted on glass slides using Glycergel | (Dako), and examined and photographed under a conventional fluorescence microscope or a confocal laser scanning microscope (Leica, TCS4D, Heidelberg, Germany).
Plasmid vectors, pCMVluc (pUT650 6.2 kb) containing the firefly lucif- erase gene under the control of the human cytomegalovirus promoter was from Cayla (Toulouse, France). The green fluorescent protein expression vector (pEGFP) was purchased from Clontech Laboratories (Paris, France) and contained the promoter of the human cytomegalovirus.
Transfections. HMCs and PMCs were seeded on 12-well cuhnre plates Nunc Primaria culture plates (Becton Dickinson, Le Pont de Claix, France) at a density of 1.5 • 10 a cells/well, and gown overnight in medium con- taining 20% FCS at 37 ~ C. The cells were grown to 70% comquent monolay- ers, which were washed twice with PBS+, then once with M199. The trans- fections were performed as described below.
For transfection with N-(1-(2,3-dioleoyloxy)propyl)N,N,N-trimethylammon-
ium methyl sulfate (DOTAP), the deoxyribonucleic acid (DNA) pCMVluc cationic lipid complexes (lipoplexes) were prepared in sterile 1-ml polysty- rene tubes, according to the manufacturer's instructions, pCMVluc (5 Ixg) was diluted in 50 txl of serum-free M199 medium. In a separate sterile reaction tube, 10-40 ixg of DOTAP (Boehringer Mannheim) was mixed with 100 txl of serum-free M199 medium. The plasmid solution was transfen'ed to the DOTAP solution and mixed by gentle pipetting without vortexing. The solu- tion was kept for 15 min at room temperature; 850 Ix1 of M199 medium containing 1% FCS was then added, and the mixture was added to the cells. After incubation for 6 h, the cells were washed with M199 medium and incubated for 24 h in M199 medium containing 20% FCS at 37 ~ C, in a humidified atmosphere composed of 5% CO2 and 95% air.
For transfection with 1,3-dioleoyloxyl-2,6-carboxyl spermidyl priopylamide (DOSPER), the Iipoplexes were prepared as described above, with a few modifications, according to the manufacturer's instructions, pCMVluc (2.5 p.g) was diluted in 50 I*1 of serum-free M199 medium. In a separate sterile reaction tube, 3~30 txg of DOSPER (Boehringer Mannheim) was mixed with 100 t*1 of serum-free M199 medium. The solution was kept for 15 rain at room temperature, before being made up to 1 nil with M199 medium con- taining 1% FCS. This mixture was added to the cells. After incubation for 3 h, the cells were washed with serum-free M199 medium and incubated for either 6 or 24 h in M199 medium containing 20% FCS at 37 ~ C, in a humidified atmosphere composed of 5% CO2 and 95% air.
For transfection with Fugene 6, 6 td of the reagent (Boehringer Mannheim) was added to 100 txl serum-free M199 and incubated for 5 min at room temperature to eliminate the ethanol, according to the manufacturer's instruc- tions. The solution thus obtained was added to 2 txg pCMVluc. After mixing by gentle pipetting without vortexing, 900 t*l M199 medium containing 20% FCS was added. This mixture was added to the cells, which were incubated for 24 h at 37 ~ C in a humidified atmosphere containing 5% CO2 and 95% air.
Transfection with calcium phosphate/DNA coprecipitation was done by transfecting the HMCs and PMCs by coprecipitation of pCMVluc with cal- cium phosphate. Calcium phosphate (2.5 nrM) was coprecipitated with var- ious amounts of pCMVluc (0.2, 0.4, 0.8, 1, 2, 4, 8 txg) in 100 I*l M199. Then, 900 txl M199 medium containing 1% FCS was added, and this mix- ture was added to the ceils. After incubation for 3 h, the cells were washed with serum-free M199 medium and incubated for 24 h in M199 medium containing 20% FCS at 37 ~ C, in a humidified atmosphere composed of 5% CO: and 95% air.
In the control experiment, cells were incubated with pCMVluc without any transfection reagents. Experiments were perforuled in triplicate wells and repeated with different PMCs and HMCs from different omentum specimens.
Luciferase assay. Luciferase gene expression was measured using the lu- minescence assay" (Luciferase Assays System Freezer Pack, Proem, MD) (De Wet et al., 1987; Brasier et al., 1989). Twenty-four hours after transfection, the medium was discarded, and the cells were washed three times with PBS. Four hundred microliters of homogenization buffer (1.5 g of Tris phosphate in 400 ml of sterile water adjusted to pH 7.8 with H3P04 and containing 8 mM MgClu, 1 mM dithiothreitol, 1 mM EDTA, 1% Triton-X 100, and 13% glycerol) was added to each well. After incubation for 10 min at room tem- perature, the solution was vigorously vortexed. The lysates were recovered and centrifuged at 2000 rpm for 7 rain at 4 ~ C. The superuatants were stored at -80 ~ C until use (no more than 7 d). Ninety-five microliters of 2 mM adenosine triphosphate in homogenization buffer without Triton-X 100 was added to 65 I*1 of cell lysate, and the sample thus obtained was introduced into a counting chamber. Upon addition of 150 I*l luciferin (165 mM in water), the luminescence was recorded for 4 s using a luminometer (Lumal LB9501; Berthold Wild Bearch, Germany). Measurements were done in du- plicate. The data were expressed as the mean -+ standard en'or of the mean (SEM) RLU/106 living cells.
Flow-cytometry analysis of transfected cells. Transfection with EGFP was perforuled as described above for DOTAl?, DOSPER, and Fugene 6 by using the plasmid formulation which has given the best results in transfection with the luciferase plasmid. The expression of EGFP was measured after 24 h. Cells were harvested by wypsin/EDTA, resuspended in PBS, and analyzed using a cell flow cytometer (FACSCAN; Becton Dickinson, Grenoble, France) with a 488-nm excitation wavelength and a 520-nm emission wavelength.
Statistics. Data are expressed as means _+ SEMs from four to eight exper- iments. Student's t-test was used for statistical comparisons.
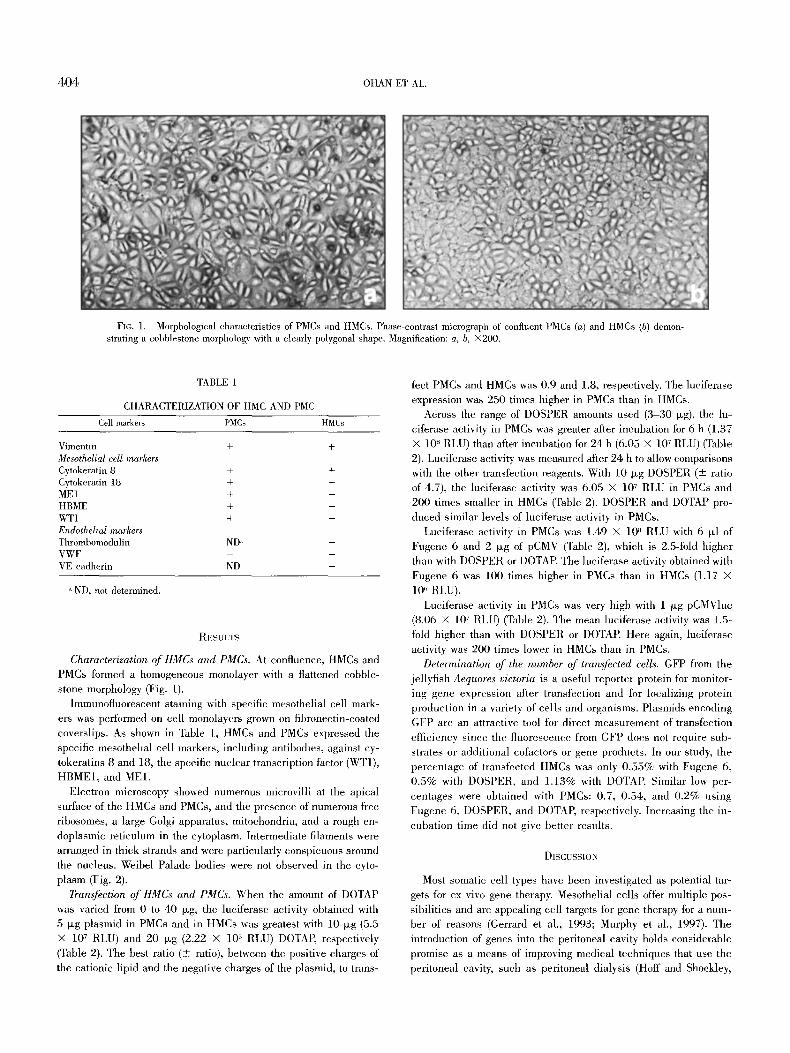
4 0 4 OHAN ET AL.
FIG. 1. Morphological characteristics of PMCs and HMCs. Phase-contrast micrograph of confluent PMCs (a) and HMCs (b) demon- strating a cobblestone morphology with a clearly polygonal shape. Magnification: a, b, •
TABLE 1
CHARACTERIZATION OF HMC AND PMC
Cell markers PMCs HMCs
Vimentin + + Mesothelial cell markers Cytokeratin 8 + + Cytokeratin 18 + + ME1 + +
HBME + + WT1 + + Endothelial markers Thrombomodulin ND" + VWF - - VE cadherin ND -
ND, not determined.
RESUH'S
Characterization of HMCs and PMCs. At confluence, HMCs and PMCs formed a homogeneous monolayer with a flattened cobble- stone morphology (Fig. 1).
Iminunofluorescent staining with specific mesothelial cell mark- ers was performed on cell monolayers grown on fibronectin-coated coverslips. As shown in Table 1, HMCs and PMCs expressed the specific mesothelial cell markers, including antibodies, against cy- tokeratins 8 and 18, the specific nuclear transcription factor (WT1), HBME1, and ME1.
Electron microscopy showed numerous microvilli at the apical surface of the HMCs and PMCs, and the presence of numerous free ribosomes, a large Golgi apparatus, mitochondria, and a rough en- doplasmic reticulum in the cytoplasm. Intermediate filaments were arranged in thick strands and were particularly conspicuous around the nucleus. Weibel Palade bodies were not observed in the cyto- plasm (Fig. 2).
Transfection of HMCs and PMCs. When the amount of DOTAP was varied from 0 to 40 p~g, the luciferase activity obtained with 5 p~g plasmid in PMCs and in HMCs was greatest with 10 ~g (5.5 • 107 RLU) and 20 p~g (2.22 • 1@ RLU) DOTAP, respectively (Table 2). The best ratio (_+ ratio), between the positive charges of the cationic lipid and the negative charges of the plasmid, to trans-
fect PMCs and HMCs was 0.9 and 1.8, respectively. The luciferase expression was 250 times higher in PMCs than in HMCs.
Across the range of DOSPER amounts used (3-30 p~g), the lu- ciferase activity in PMCs was greater after incubation for 6 h (1.37 • 10 ~' RLU) than after incubation for 24 h (6.05 • 107 RLU) (Table 2). Luciferase activity was measured after 24 h to allow comparisons with the other transfection reagents. With 10 ~g DOSPER (+- ratio of 4.7), the luciferase activity was 6.05 x 107 RLU in PMCs and 200 times smaller in HMCs (Table 2). DOSPER and DOTAP pro- duced similar levels of luciferase activity in PMCs.
Luciferase activity in PMCs was 1.49 • 10 ~ RLU with 6 ~1 of Fugene 6 and 2 p~g of pCMV (Table 2), which is 2.5-fold higher than with DOSPER or DOTAP. The luciferase activity obtained with Fugene 6 was 100 times higher in PMCs than in HMCs (1.17 • 10" RLU).
Luciferase activity in PMCs was very high with 1 tLg pCMVluc (8.06 x 107 RLU) (Table 2). The mean luciferase activity was 1.5- fold higher than with DOSPER or DOTAP. Here again, luciferase activity was 200 times lower in HMCs than in PMCs.
Determination of the number of transfected cells. GFP from the jellyfish Aequores victoria is a useful reporter protein for monitor- ing gene expression alter transfection and for localizing protein production in a variety of cells and organisms. Plasmids encoding GFP are an attractive tool for direct measurement of transfection efficiency since the fluorescence from GFP does not require sub- strates or additional cofactors or gene products. In our study, the percentage of transfected HMCs was only 0.55% with Fugene 6, 0.5% with DOSPER, and 1.13% with DOTAP. Similar low per- centages were obtained with PMCs: 0.7, 0.54, and 0.2% using Fugene 6, DOSPER, and DOTAP, respectively. Increasing the in- cubation time did not give better results.
DISCUSSION
Most somatic cell types have been investigated as potential tar- gets for ex vivo gene therapy. Mesothelial cells offer muhiple pos- sibilities and are appealing cell targets for gene therapy for a num- ber of reasons (Gerrard et al., 1993; Murphy et al., 1997). The introduction of genes into the peritoneal cavity holds considerable promise as a means of improving medical techniques that use the peritoneal cavity, such as peritoneal dialysis (Hoff and Shockley,

PMC AND HMC FOR GENE TRANSFER 4 0 5
FIG. 2. Electron photomicrograph of confluent cultures of PMCs and HMCs. Electron photomicrographs of confluent cultures of PMCs (A) and HMCs (B). Numerous microvilli are present at the cell surface (mi) (magnification: • for HMCs; and • for PMCs). Adjacent cells produce tortuous, tight intercellular junctions (tj), near which microvilli are abundant (magnification: • for HMCs). The cytoplasm of PMCs contains numerous mitochondria (mr) and abundant rough endoplasmic reticulum (re) (magnification: • 1550 tor PMCs).
TABLE 2
TRANSFECTION EFFICIENCY WITH HMC AND PMC
PMCs (RLU • 107)" HMCs (RLU • 107y PMCs/HMCs
Control 0.0087 • 0.0043 0.0011 • 0.0002 8 DOTAP 5.50 + 1.61 0.0222 • 0.0131 250 DOSPER 6.05 • 1.66 0.0291 • 0.0075 207 Phosphate Ca ++ 8.06 -+ 2.80 0.0395 • 0.0001 204 Fugene 6 14.90 • 4.14 0.117 • 0.0045 127
Luciferase activity is expressed as RLUs per 106 live cells.
1999). We developed a porcine model to assist in designing an ex vivo strategy for gene therapy in humans. We have accumulated experience with our porcine model of vascular diseases, and we have access to material appropriate for working with this model.
Furthermore, pigs are an accepted mammalian animal model for studying gene transfer (Dodds, 1992). Although several porcine cell types have been used in earlier studies for gene transfer with viral
vectors (adenovirus or retrovirus) or nonviral vectors (liposomes), PMCs have not yet been evaluated for gene transfer (Benhamou et al., 1997; Ghazizadeh et al., 1998; Shiraishi et al., 1998; Rio et al., 1999; Blanton et ah, 2000). Cationic l iposome-mediated trans- fection is effective, reproducible, and fairly simple, and as a result has been widely used to introduce foreign genes into a variety of primal~r and cultured cells (Tokeshita et al., 1994). We have inves- tigated the transfection of primal T PMCs and of HMCs with different transfection reagents, including cationic liposomes (DOSPER and DOTAP), calcium phosphate precipitation, and Fugene 6. We have used a reporter gene encoding luciferase and one encoding GFP gene in order to estimate the number of transfected ceils.
Our resul ts ind ica te that i r respect ive of the t ransfect ion re-

4 0 6 OHAN ET AL.
agent used, the transfeetion efficiency was always higher in PMCs than in HMCs. Fugene 6 is a very efficient transfection reagent for both PMCs and HMCs. The difference in the efficacy of gene transfer between these two cell types was only lO0-fold with Fugene 6 compared with the other reagents tested (200- fold). This suggests that the HMCs may be more sensitive than the PMCs to Fugene 6. The important differences between Fu- gene 6 and the other reagents found in our study may be ascrib- able to the milder eytotoxicity of Fugene 6 and to the fact that this transfeetion reagent can be used in the presence of serum. Debs and eoworkers reported a modified transfection protocol for cell l ines that allows DNA/liposome transfeetion in the presence of serum (Brunette et al., 1992). Furthermore, Fugene 6 provided greater transfer efficiency with a smaller amount of DNA (2 lag as opposed to 5 lag with DOTAP). The concentration of DNA could be a limiting feature of the efficiency of gene transfer.
With both PMCs and HMCs, ealeium phosphate/DNA precipi- tation provided more efficient gene transfer than DOTAP or DOS- PER. It has been shown that the protein kinase C activator ean enhance calcium phosphate-mediated transfeetion, a process that also involves phagoeytosis and endocytosis (Graham and Van Der Eb, 1973; Reston et al., 1991). As the peritoneum is a semiper- meable membrane, this may explain the marked difference in transfer efficiency between calcium phosphate/DNA and DOSPER or DOTAR However, the changes in mesothelial cell morphology observed in this study suggest that the calcium phosphate/DNA method may have eytotoxic effects responsible for cell vaeuoliza- tion and detachment (data not shown). No cytotoxie effects are mentioned in a report by Nagy et al. (1995a) of a study of stron- tium/DNA-precipitation gene transfer into rat peritoneal mesothe- lial cells.
Only about 1% of mesothelial cells from both species were trans- feeted. The number of transfected cells was very low (0.5-1.2%) with the transfeetion reagents tested. This limited number of cells expressing GFP contrasts with the fairly high level of luciferase expression (1 X 10 ~ RLU/IO" PMCs), suggesting that either the tew transfected cells expressed the transgene at a high level or many transfected cells expressed the transgene at a low level.
Our results suggest that the gene transfer into PMCs, especially with Fugene 6, led to high expression of a transgene. Our primat3J results concerning the infection of PMCs and HMCs with the first- generation adenovirus type 5 showed that lueiferase activity mea- sured with adenovirus on PMCs was similar to that obtained with Fugene 6, and that HMCs are more sensitive to adenovirus than to Fugene 6 (results not shown). These first results suggest that we could test Fugene 6 to transfeet PMCs without the disadvantage of adenovirus, This method may be useful for studying genetically modified mesothelial cells of potential interest for achieving the synthesis of therapeutic molecules easily released into the systemic circulation, with the goal of improving the ability of mesothelial cells to protect vascular prostheses from thrombosis. Human and rabbit peritoneal mesothelial cells have been successfully cultured and reimplanted into their donor (Hoff et al., 1998; Zoldhelyi et al., 2000). Additional studies on PMCs as a tool for testing inducible promoters or delivering therapeutic proteins would he of interest. Furthermore, the in vivo behavior of PMCs and their differentiation potential after seeding on to the vascular prosthesis need to be investigated. Quantification of a recombinant protein secreted by PMCs previously transfeeted using Fugene 6 would be of great in-
terest for evaluating the transfer efficiency and determining whether genetically modified PMCs deserve to be investigated in an exper- iinental study in pigs.
REFERENCES
Benhamou, R Y.; Morisoot, C.; Prevost, R; Rolland, E.; Halimi, S.; Chroboe- zek, J. Standardization of procedure for efficient ex vivo gene transfer into porcine panei~atie islets with cationic liposonms. Transplantation 3:1798-1803; 1997.
Blanton Jr., J. R.; Bidwell, C. A.; Sanders, D. A.; Sharkey, C. M.; MeFarland, D. C.; Gerrard, D. E., Grant, A. L. Plasmid transfeetion and retroviral transduction of porcine muscle cells for cell-mediated gene transter. J. Anim. Sci. 4:909-918; 2000.
Bordenave, L.; Remy-Zolghadri, M.; Fernandez, R; Bareille, R.; Midy, D. Clinical performance of vascular grafts lined with endothelial cells. Endothelium 6:267-275; 1999.
Brasier, A. R.; Tate, J. E.; Habener, J. F. Optimized use of the firefly lueiferase assay as a reporter gene in mammalian cell lines. Bioteehniques 7:1116-1123; 1989.
Brunette, E.; Stribling, R.; Debs, R. Lipofeetion does not require the removal of serum. Nucleic-Acids Res. 20:1151-1158; 1992.
De Wet, J. R.; Wood, K. V.; DeLuca, M.; Helinsky, D. R.; Subra Mauri, S. Firefly lueiferase gene structure and expression in mammalian cells. Mol. Cell. Biol. 7:725-737; 1987.
Dodds, W. J. The pig model for biomedical research. FASEB J. 41:247-256; 1992.
Gerrard, A. J.; Hudson, D. L.; Browlnee, G. G.; Watt, F. M. Toward gene therapy for haemophilia B using primary human keratinoeytes. Nat. Genet. 3:180-183; 1993.
Ghazizadeh, S.; Harrington, R.; Garfield, J.; Taiehman, L. B. Retrovirus-me- diated transduetion of porcine keratinocytes in organ culture. J. In- vest. Dermatol. 111:492-496; 1998.
Graham, F. L.; Van Der Eb, A. J. A new technique for the assay of infectivity of human adenovirus DNA. Virology 52:456467; 1973.
Hoff, C. M.; Cusick, J. L.; Masse, E. M.; Jackman, R. W.; Nagy, J. A.; Shock- ley, T. R. Modulation of transgene expression in mesothelial cells by activation of an inducible promnter. Nephrol. Dial. Transplantation 13(6):1420-1429; 1998.
Hoff, C. M.; Shoekley, T. R. The potential of gene therapy in the peritoneal cavity. Perit. Dial. Int. 19:$202-$207; 1999.
Ivarsson, M. L.; Holmdahl, L.; Falk, R; Molne, J.; Risberg, B. Characteriza- tion and fibrinolytic properties of mesothelial cells isolated from peri- toneal lavage. Seand. J. Clin. Lab. Invest. 58:195-203; 1998.
Jaekman, R. W.; Stapleton, T. 1).; Masse, E. M.; Harvey, V. S.; Meyers, M. S.; Shoekley, T. R.; Nagy, J. A. Enhancement of the functional rep- ertoire of rat parietal peritoneal mesolhelium in vivo: directed ex- pression of the anticoagulant and antiinflammatory molecule throm- homodulin. Hum. Gen. Ther. 9:1069-1081; 1998.
Jankowski, R. J.; Wagner, W. R. Directions in eardiovaseular tissue engi- neering. Clin. Plast. Surg. 26:605-616; 1999.
Kern, R A.; Knedler, A.; Eckel, It. H. Isolation and culture of mierovascular endothelium from human adipose tissue. J. Clin. lnvest. 71:1822-1829; 1983.
Les~ehe, G.; Ohan, J. Am61ioration de la pem6abilit6 des proth~ses en Da- cron implant6es en position veineuse grace h l'ensemeneement h haute densit6 de eellules endoth61iales. Ann. Chin Vase. 9(3):325- 326; 1995.
Murphy, J. E.; Rheinwald, J. G. Intraperitoneal injection of genetically mod- ified human mesothelial cells for systemic gene therapy. Hum. Gene Ther. 8:1867-1879; 1997.
Nagy, J. A.; Shoekley, T. R.; Jackman, R. W. Mesothelial cell gene therapy. J. Cell. Bioehem. 18A (Suppl.):244; 1994.
Nagy, J. A.; Shoekley, T. R.; Masse, E. M.; Harvey, V. S.; Holt; C. M.; Jack- man, R. W. Systemic delivery" of recombinant protein by genetically modified mesothelial cells reseeded on the parietal peritoneal surface. Gene Ther. 2:402-410; 1995a.
Nagy, J. A.; Shockley, T. R.; Masse, E. M.; Harvey, V. S.; Jaekman, R. W. Mesothelial cell mediated gene therapy: feasibility of an ex vivo strat- egy. Gene Ther. 2:393-40i; 1995b.

PMC AND HMC FOR GENE TRANSFER 407
Ohan, J.; Gilbert, M. A.; Brouland, J. P.; Rougier, J. P.; Trugnan, G.; Vassef, M.; Les~che, G.; Drouet, L. Phenotypic and functional characteristics of porcine peritoneal mesothelial cells. In Vitro Cell. Dev. Biol. 35A:625-634; 1999.
Pasic, M.; Muller-Glauser, W.; Obermatt, B.; Lachat, M.; Seifert, B.; Turina, M. Seeding with omental cells prevents late neointimat hyperplasia in small-diameter Dacron grafts. Circulation 92:2605-2616; 1995.
Pronk, A.; Leguit, P.; Hoynch van Papendrecht, A. A.; Hagelen, E.; van- Vroonhoven, T. J.; Verbrugh, H. A. A cobblestone cell isolated from the human omentum. The mesothelial cells: isolation, identification and growth characteristics. In Vitro. Cell. Dev. Biol. 29A:127-134; 1993.
Rando, T. A.; Black, H. M. Primary mouse myoblast purification character- istic and transplantation for cell mediated gene therapy. J. Cell Biol. 125:1275-1287; 1994.
Reston, J. T.; GouM-Forgerite, S.; Mannino, R. J. Potentiation of DNA me- diated gene transfer in NIH3T3 cells by activators of protein kinase C. Biochim. Biophys. Acta 1088:270-276; 1991.
Rio, M. D.; Lardner, F.; Meana, A.; Segovia, J.; Alvarez, A.; Jorcano, J. Non viral transfer of gene to pig primary keratynocytes induction of an- giogenesis by composite grafts of modified kcratynocytes overexpres- sion VEGF driven by a keratin promoter. Gene Ther. 10:1734-1741; 1999.
Sackman, J. E.; Freeman, M. B.; Peterseon, M. G.; Allebban, Z.; Niemeyer, G. P.; Lothrop Jr., C. D. Synthetic vascular grafts seeded with genet- ically modified endothelium in the dog: evaluation of the effect of seeding technique and retroviral vector on cell persistence in vivo. Cell Transplantation 2:219-235; 1995.
Schwartz, R. S.; Edward, S. D. W.; Bailey Kent, R.; Camrud, A. R.; Jorgensen, A. M. Differential neointimal response to coronary artery injury in pigs and dogs: implications for restenosis models. Arterioscl. Thromb. 14:395-400, 1994.
Shiraishi, M.; Niroyasu, S.; Nagabama, H.; Taira, K.; Nosato, E.; Towori, H.; Obuchi, Y.; Muto, Y. Adenoviral mediated gene transfer to the porcine liver in vivo. Transplantation Proc. 7:2914-2916; 1998.
Sugawara, Y. Adenovirus-mediated transfer of tissue type plasminogen acti- vator gene to human endothelial cells. Surgery 122:91-100; 1997.
Thomas, S. H.; Theodor, H.; Welling, B. S.; Rajabrata, S.; Messina, L. M.; Stanley, J. C. Effect of retroviral-mediated tissue plasminogen acti- vator gone transfer and expression on adherence and proliferation of canine endothelial cell seeded onto expanded polytetrafluoroethylene. J. Vasc. Surg. 22:795~803; 1995.
Tokeshita, S.; Losolvlo, D. W.; Keamey, M.; Rossow, S. T.; Isner, J. M. Time course of reconrbinant protein secretion after liposome-mediated gene transfer in a rabbit arterial organ culture model. Lab. Invest. 71:387-391; 1994.
van Hinsbergh, V. W.; Kooistra, T.; Scheffer, M. A.; van HajoBockel, J.; van Muijen, G. N. Characterization and fibrinolytic properties of human omental tissue mesothelial ceils: comparison with endothelial cells. Blood 75:1490-1497; 1990.
Zoldhelyi, P.; McNatt, J.; Shelat, H. S.; Yamamoto, Y.; Chen, Z. Q.; Willerson, J. T. Thromboresistance of balloon-injured porcine carotid arteries after local gene transfer of human tissue factor pathway inhibitor. Circulation 101:289-295; 2000.




















