
Microplastics Derived From Disposable Greenhouse Plastic ...
Upload
independentCategory
view
2download
0
NmDef02, a novel antimicrobial gene isolated fromNicotiana megalosiphon confers high-level pathogenresistance under greenhouse and field conditionsRoxana Portieles1, Camilo Ayra1, Ernesto Gonzalez1, Araiz Gallo1, Raisa Rodriguez1, Osmany Chacon2,Yunior Lopez1, Mayra Rodriguez1, Juan Castillo3, Merardo Pujol1, Gil Enriquez1, Carlos Borroto1,Luis Trujillo1, Bart P.H.J. Thomma4 and Orlando Borras-Hidalgo1,*
1Center for Genetic Engineering and Biotechnology, Havana, Cuba2Tobacco Research Institute, San Antonio de los Banos, Havana, Cuba3INCA, San Jose de las Lajas, Havana, Cuba4Laboratory of Phytopathology, Wageningen University, Wageningen, The Netherlands
Received 16 July 2009;
revised 10 December 2009;
accepted 12 December 2009.
*Correspondence (fax +537 33 1779;
email [email protected])
Keywords: plant defensin, recombi-
nant proteins, transgenic plants, dis-
ease control.
SummaryPlant defensins are small cysteine-rich peptides that inhibit the growth of a broad
range of microbes. In this article, we describe NmDef02, a novel cDNA encoding a
putative defensin isolated from Nicotiana megalosiphon upon inoculation with the
tobacco blue mould pathogen Peronospora hyoscyami f.sp. tabacina. NmDef02 was
heterologously expressed in the yeast Pichia pastoris, and the purified recombinant
protein was found to display antimicrobial activity in vitro against important plant
pathogens. Constitutive expression of NmDef02 gene in transgenic tobacco and
potato plants enhanced resistance against various plant microbial pathogens, includ-
ing the oomycete Phytophthora infestans, causal agent of the economically impor-
tant potato late blight disease, under greenhouse and field conditions.
Introduction
Upon pathogen challenge, plants activate a defence-
related metabolism producing several metabolites to
defend themselves against these pathogens, including
antimicrobial peptides such as defensins (Bruce and Pick-
ett, 2007). Plant defensins are a family of small (typically
45–54 amino acids), highly stable, basic cysteine-rich pep-
tides that occur ubiquitous throughout the plant kingdom
(Thomma et al., 2002). These peptides play an important
role as part of the plant’s natural defence system by inhib-
iting growth of a broad range of infectious microbes (Bro-
ekaert et al., 1995; Thomma et al., 2002, 2003).
Members of the defensin family are quite diverse with
respect to their amino acid composition, as only the eight
structure-stabilizing cysteines and a glycine residue (posi-
tion 34; numbering relative to type defensin member
Rs-AFP2; Terras et al., 1992) appear to be conserved
among almost all of the family members, which reflects
the diverse biological activities displayed by different plant
defensins (Broekaert et al., 1995; Thomma et al., 2002,
2003; Lay and Anderson, 2005). Furthermore, an aromatic
residue (position 11) that is always followed by a glycine
(position 13), two residues further along the polypeptide
chain, as well as a serine (position 8) and a glutamic acid
(position 29) residue are often found to be conserved (Lay
and Anderson, 2005).
Plant defensins have been isolated from almost all plant
tissues including seeds (Terras et al., 1992; Broekaert
et al., 1995), leaves (Terras et al., 1995), tubers (Moreno
et al., 1994), fruit (Meyer et al., 1996) and floral tissues
(Thomma and Broekaert, 1998; Park et al., 2002; Lay
et al., 2003). While some defensins exhibit constitutive
expression, others are induced by various abiotic and bio-
tic stress factors (Penninckx et al., 1996; Thomma and
Broekaert, 1998; Do et al., 2004). Many defensins have
been proven to display antimicrobial activity (Thevissen
et al., 2007), while others lack apparent antifungal or anti-
bacterial activity (Osborn et al., 1995; Liu et al., 2006).
The alfalfa (alfAFP; Gao et al., 2000) defensin isolated
from seeds of Medicago sativa was found to display
strong activity against the agronomically important fungal
ª 2010 CIGB678 Journal compilation ª 2010 Blackwell Publishing Ltd
Plant Biotechnology Journal (2010) 8, pp. 678–690 doi: 10.1111/j.1467-7652.2010.00501.x

pathogen Verticillium dahliae (Fradin and Thomma, 2006).
This defensin also inhibits the growth of other fungal plant
pathogens such as Alternaria solani and Fusarium culmo-
rum. However, the same defensin lacks significant in vitro
antimicrobial activity against the oomycete Phytophthora
infestans, an economically important pathogen that causes
potato late blight (Gao et al., 2000).
The mode of action of two different plant defensins
from radish (Raphanus sativus) and dahlia (Dahlia merckii),
Rs-AFP2 and Dm-AMP1, respectively, has been studied in
detail (Thevissen et al., 2000, 2004). Both peptides were
found to bind distinct sphingolipids in fungal outer plasma
membranes and, consequently, showed a differential activ-
ity against fungal and yeasts species, including the human
pathogen Candida albicans (Thevissen et al., 2004). The
antimicrobial activity in vitro of plant defensins suggests
an important role in the plant defence response, which is
further evidenced by enhanced disease resistance pheno-
types observed in different plant species heterologously
overexpressing plant defensin genes (Terras et al., 1995;
Gao et al., 2000; Zhu et al., 2007; Sels et al., 2008). Con-
stitutive expression of the radish defensin RsAFP2 was
found to enhance pathogen resistance of various plant
species, such as tobacco plants to the fungal leaf patho-
gen Alternaria longipes (Terras et al., 1995) and tomato to
A. solani (Parashina et al., 2000). Transgenic canola (Bras-
sica napus) constitutively expressing a pea defensin
showed enhanced resistance against blackleg disease
caused by the fungus Leptosphaeria maculans (Wang
et al., 1999). Furthermore, transgenic potato expressing
an alfalfa defensin exhibited robust resistance against
V. dahliae under field conditions (Gao et al., 2000). These
studies indicate that engineering disease resistance in
crops with a range of plant defensins has the potential to
provide protection against various fungal diseases.
In this study, we report on NmDef02, a novel defensin
gene isolated from the tobacco species Nicotiana megalo-
siphon, a non-cultivated wild tobacco species that is gen-
erally used as a parent in genetic tobacco breeding
programs because of its high resistance towards several
important pathogens (Borras-Hidalgo et al., 2006; Collazo
et al., 2006; Chacon et al., 2009). For example, N. mega-
losiphon has been shown to be highly resistant to Pero-
nospora hyoscyami f.sp. tabacina, an obligate biotrophic
oomycete for which species from the genus Nicotiana are
the only hosts and that causes one of the most important
foliar diseases of tobacco (Borras-Hidalgo et al., 2009). We
report on the activity of NmDef02 against several plant
pathogens in vitro. Furthermore, we show that the expres-
sion of this defensin gene in transgenic tobacco and
potato plants provides high-level resistance levels against
several pathogens under greenhouse and field conditions.
Results
In a suppression subtractive hybridization (SSH) library
prepared from N. megalosiphon leaves inoculated with
P. hyoscyami f.sp. tabacina (Borras-Hidalgo et al., 2006,
2009), a 219 bp clone with homology to plant defensin
genes was identified and named NmDef02. Based on the
deduced amino acid sequence, NmDef02 is a 73 amino
acids polypeptide including a putative 27 amino acids
(81 bp) signal peptide (MDRVALVSLCFVYLVLFVAQEIVV-
TEA) (Figure 1a). An amino acid sequence alignment of
plant defensins collected from public databases and pub-
lished reports (Lay and Anderson, 2005; De-Paula et al.,
Figure 1 (a) The DNA coding sequence for the NmDef02 mature peptide (bold letters) and its 27-amino acid signal peptide are shown below the
amino acid sequence. Underlined and in italics are the start and stop codon (*), respectively. (b) Comparison of the putative mature sequence of
NmDef02 and those of other 74 plant defensins retrieved from the SUCEST database and Psd1 (http://compbio.dfci.harvard.edu/tgi/). The
sequence alignment was performed with CLUSTAL X software (Thompson et al., 1997). The signal peptide sequence has been omitted and spaces
have been introduced to maximize the alignment. A conserved glycine and the eight cysteine residues that are invariant among members of the
plant defensin family are included in the consensus sequence dashed in gray and highlighted with capital letters and with asterisks. The serine resi-
due, not conserved in NmDef02, is dashed in black and with lowercase letters. Also, with lowercase letters are indicated the putative aromatic res-
idue (denoted as ‘a’), glycine and a glutamic acid that are conserved in most of the aligned sequences. All the sequences are coded by their
accession number followed by the abbreviated name of plant species from which they were isolated in bold. Classifiers are as follows: At: Arabid-
opsis thaliana, Bv: Beta vulgaris, Bn: Brassica napus, Bo: Brassica oleracea, Ca: Capsicum annuum, Cc: Capsicum chivense, Cp: Citrus paradisi, Dm:
Dahlia merckii, Ew: Eutrema wasabi, Gm: Glycine max, Hs: Heuchera sanguinea, Hv: Hordeum vulgare, Ms: Medicago sativa, Na: Nicotiana alata,
Np: Nicotiana paniculada, Oe: Olea europaea, Os: Oryza sativa, Pi: Petunia integrifolia, Ph: Petunia hybrida, Ps: Pisum sativum, Pp: Prunus persica,
Ppy: Pyrus pyrifolia, Rs: Raphanus sativa, Sa: Sinapis alba, Si: Setaria italica, Sb: Sorghum bicolor, So: Saccharum officinarum, Sol: Spinacia olera-
cea, Sp: Solanum pimpinellifolium, St: Solanum tuberosum, Ta: Triticum aestivum, Vf: Vicia faba, Vu: Vigna unguiculata, Vr: Vigna radiata, Vv: Vitis
vinifera, Zm: Zea mays. (c). Phylogenetic tree of NmDef02 with other representatives members of plant defensins. The phylogenetic tree was gen-
erated using the neighbour joining method (Saitou and Nei, 1987) (MEGA 4.0 (http://www.megasoftware.net)) using the same 74 defensins
sequences from the alignment represented above. NmDef02 is dashed in gray. Numbers represent bootstrap values and protein classifiers are
identical to those used in the multiple alignment.
ª 2010 CIGBJournal compilation ª 2010 Blackwell Publishing Ltd, Plant Biotechnology Journal, 8, 678–690
NmDef02, a novel antimicrobial protein 679

2008) is presented in Figure 1b. As expected, based on
previously published data (Thomma et al., 2002; Lay and
Anderson, 2005), limited sequence conservation is
observed among the 74 polypeptides (Figure 1b). Strictly
conserved are the eight cysteine residues thought to be
involved in disulphide bridge formation, the aromatic resi-
due at position 11 and glycines at position 13 and 34
(numbering relative to Rs-AFP2). Interestingly, the serine
(S) at position 8 that is conserved in many plant defensins
(Lay and Anderson, 2005) is not conserved in NmDef02.
(a)
(b)
ª 2010 CIGBJournal compilation ª 2010 Blackwell Publishing Ltd, Plant Biotechnology Journal, 8, 678–690
Roxana Portieles et al.680
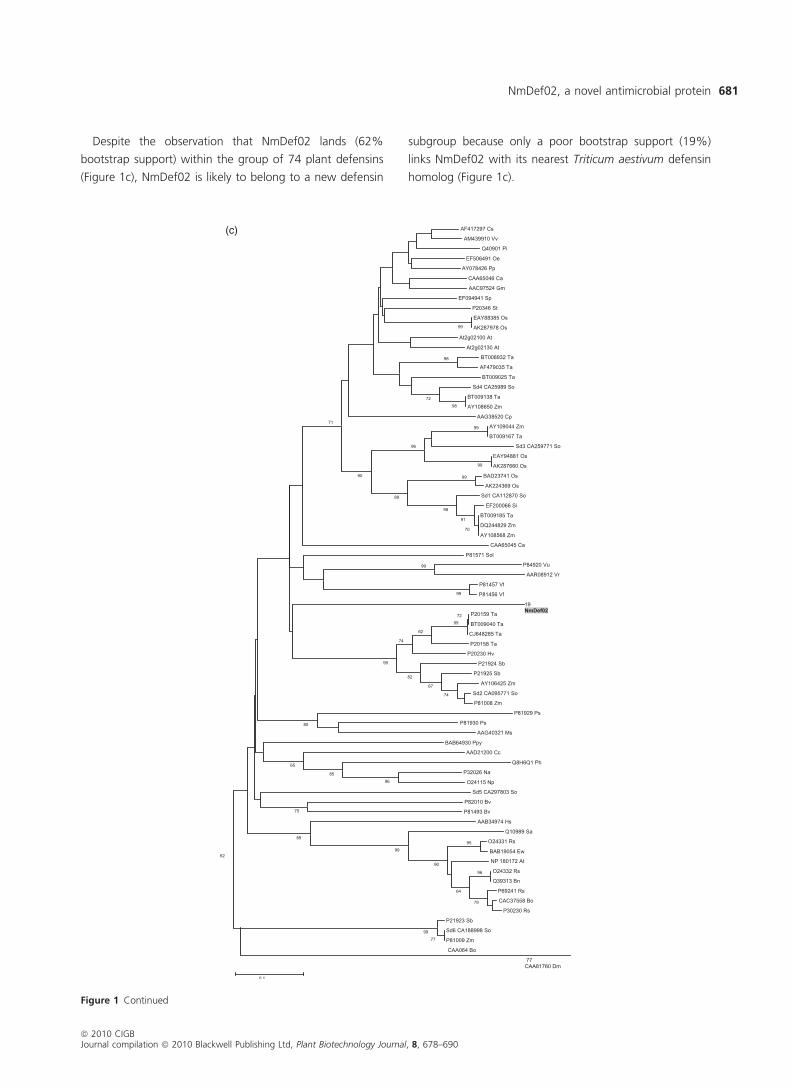
Despite the observation that NmDef02 lands (62%
bootstrap support) within the group of 74 plant defensins
(Figure 1c), NmDef02 is likely to belong to a new defensin
subgroup because only a poor bootstrap support (19%)
links NmDef02 with its nearest Triticum aestivum defensin
homolog (Figure 1c).
(c)
Figure 1 Continued
681
ª 2010 CIGBJournal compilation ª 2010 Blackwell Publishing Ltd, Plant Biotechnology Journal, 8, 678–690
NmDef02, a novel antimicrobial protein
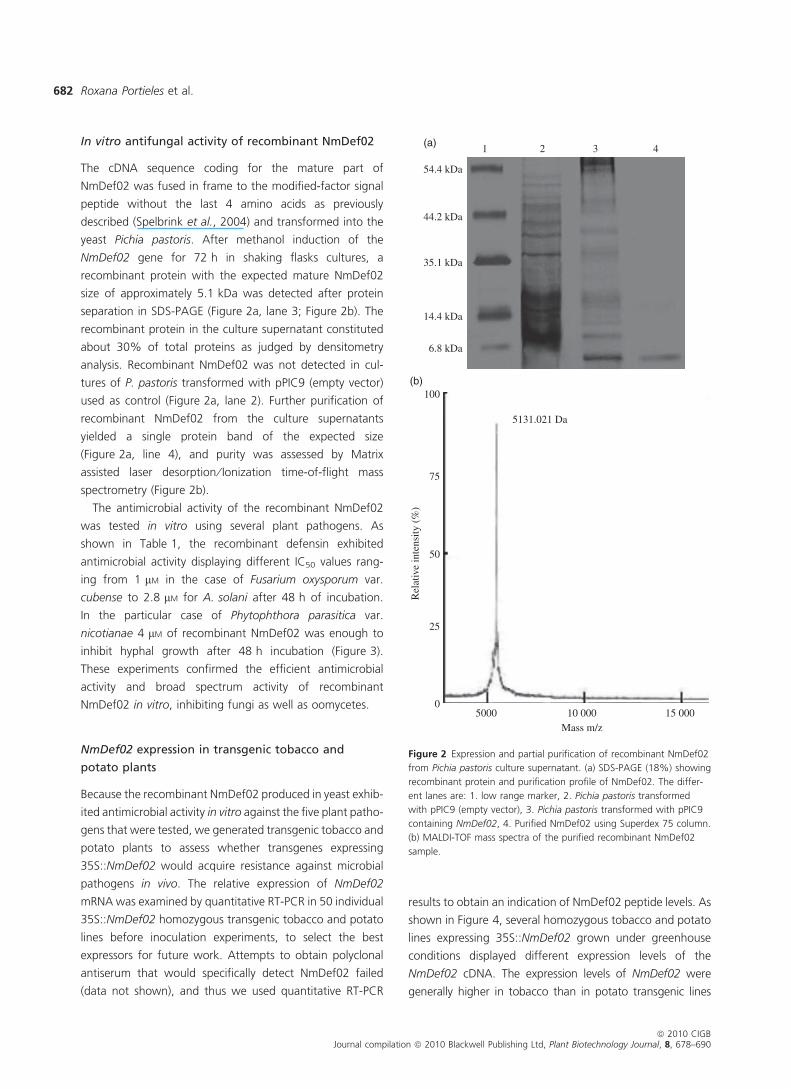
In vitro antifungal activity of recombinant NmDef02
The cDNA sequence coding for the mature part of
NmDef02 was fused in frame to the modified-factor signal
peptide without the last 4 amino acids as previously
described (Spelbrink et al., 2004) and transformed into the
yeast Pichia pastoris. After methanol induction of the
NmDef02 gene for 72 h in shaking flasks cultures, a
recombinant protein with the expected mature NmDef02
size of approximately 5.1 kDa was detected after protein
separation in SDS-PAGE (Figure 2a, lane 3; Figure 2b). The
recombinant protein in the culture supernatant constituted
about 30% of total proteins as judged by densitometry
analysis. Recombinant NmDef02 was not detected in cul-
tures of P. pastoris transformed with pPIC9 (empty vector)
used as control (Figure 2a, lane 2). Further purification of
recombinant NmDef02 from the culture supernatants
yielded a single protein band of the expected size
(Figure 2a, line 4), and purity was assessed by Matrix
assisted laser desorption ⁄ Ionization time-of-flight mass
spectrometry (Figure 2b).
The antimicrobial activity of the recombinant NmDef02
was tested in vitro using several plant pathogens. As
shown in Table 1, the recombinant defensin exhibited
antimicrobial activity displaying different IC50 values rang-
ing from 1 lM in the case of Fusarium oxysporum var.
cubense to 2.8 lM for A. solani after 48 h of incubation.
In the particular case of Phytophthora parasitica var.
nicotianae 4 lM of recombinant NmDef02 was enough to
inhibit hyphal growth after 48 h incubation (Figure 3).
These experiments confirmed the efficient antimicrobial
activity and broad spectrum activity of recombinant
NmDef02 in vitro, inhibiting fungi as well as oomycetes.
NmDef02 expression in transgenic tobacco and
potato plants
Because the recombinant NmDef02 produced in yeast exhib-
ited antimicrobial activity in vitro against the five plant patho-
gens that were tested, we generated transgenic tobacco and
potato plants to assess whether transgenes expressing
35S::NmDef02 would acquire resistance against microbial
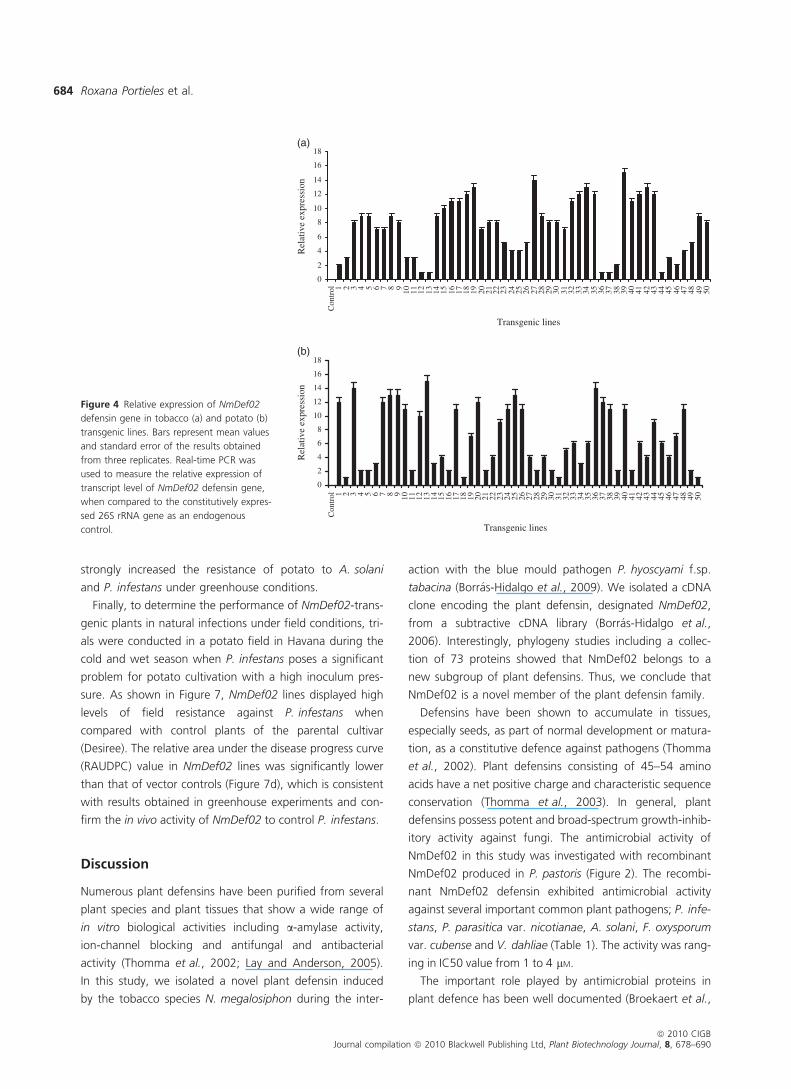
pathogens in vivo. The relative expression of NmDef02
mRNA was examined by quantitative RT-PCR in 50 individual
35S::NmDef02 homozygous transgenic tobacco and potato
lines before inoculation experiments, to select the best
expressors for future work. Attempts to obtain polyclonal
antiserum that would specifically detect NmDef02 failed
(data not shown), and thus we used quantitative RT-PCR
results to obtain an indication of NmDef02 peptide levels. As
shown in Figure 4, several homozygous tobacco and potato
lines expressing 35S::NmDef02 grown under greenhouse
conditions displayed different expression levels of the
NmDef02 cDNA. The expression levels of NmDef02 were
generally higher in tobacco than in potato transgenic lines
1 2 3 4
54.4 kDa
44.2 kDa
35.1 kDa
14.4 kDa
6.8 kDa
100
75
50
25
0
Rel
ativ
e in
tens
ity (
%)
5131.021 Da
5000 10 000 15 000Mass m/z
(a)
(b)
Figure 2 Expression and partial purification of recombinant NmDef02
from Pichia pastoris culture supernatant. (a) SDS-PAGE (18%) showing
recombinant protein and purification profile of NmDef02. The differ-
ent lanes are: 1. low range marker, 2. Pichia pastoris transformed
with pPIC9 (empty vector), 3. Pichia pastoris transformed with pPIC9
containing NmDef02, 4. Purified NmDef02 using Superdex 75 column.
(b) MALDI-TOF mass spectra of the purified recombinant NmDef02
sample.
682
ª 2010 CIGBJournal compilation ª 2010 Blackwell Publishing Ltd, Plant Biotechnology Journal, 8, 678–690
Roxana Portieles et al.

(Figure 4). Twenty transgenic lines of each species with the
highest relative NmDef02 expression were selected for disease
resistance assays under greenhouse and field conditions.
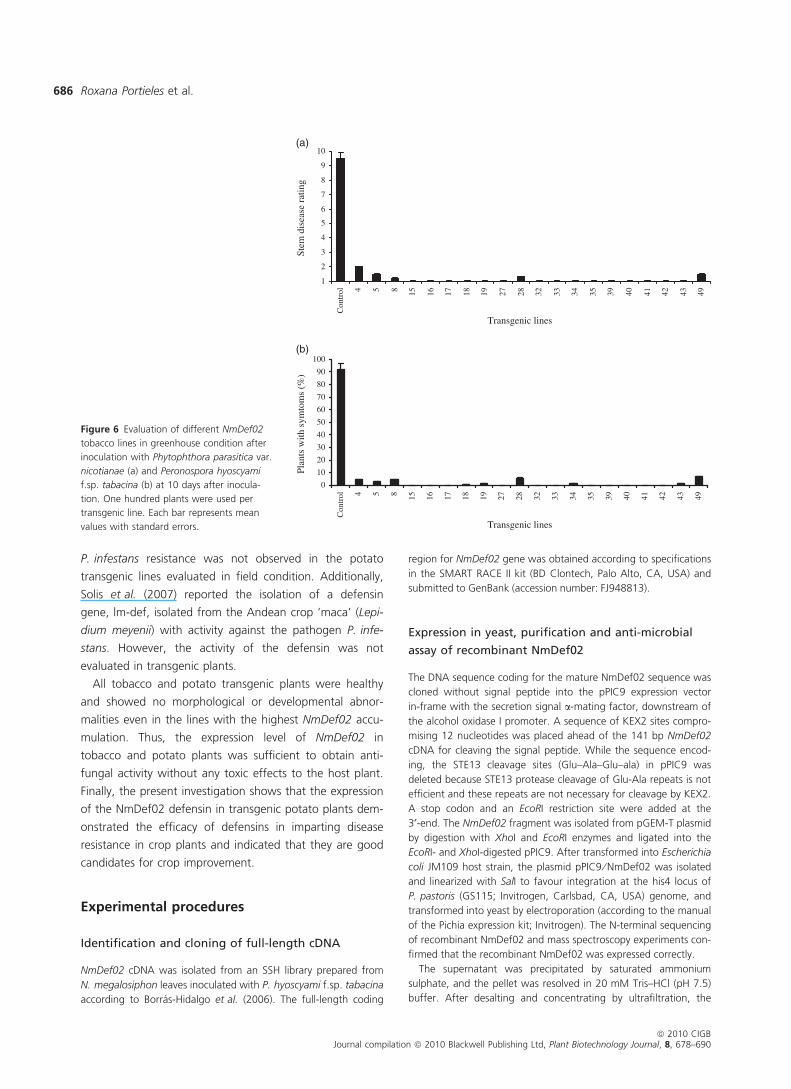
Enhanced disease resistance of transgenic tobacco
plants under greenhouse conditions
Twenty transgenic 35S::NmDef02 tobacco lines and vector
control plants were inoculated with the oomycete patho-
gens P. parasitica var. nicotianae and P. hyoscyami f.sp. ta-
bacina, respectively. At 5 days after inoculation, slight
disease symptoms produced by P. parasitica var. nicotianae
appeared on the vector control plants whereas no symp-
toms were detected at this stage on the assayed transgenic
lines. However, 10 days after infection, severe disease
symptoms such as leaf wilting and stem rot were observed
in all of the vector control plants, and all of them died
5 days later (Figure 5a). However, only few (6.7%) of the
35S::NmDef02 transgenic lines revealed mild disease symp-
toms while the rest remained healthy (Figures 5a and 6a).
Tobacco transgenic lines inoculated with P. hyoscyami f.sp.
tabacina showed a similar behaviour as upon inoculation
with P. parasitica var. nicotianae. The transgenic tobacco
lines remained without disease symptoms at 10 days after
infection, while the vector control plants died during the
same period (Figures 5b and 6b). Interestingly, we found a
high correlation between the relative expression of
NmDef02 as determined by quantitative RT-PCR and resis-
tance against P. parasitica var. nicotianae and P. hyoscyami
f.sp. tabacina in the various transgenic lines (Figure 6). Lines
4, 5, 8, 28 and 49 with a relatively low NmDef02 expression
(Figure 4a) developed symptoms as a consequence of path-
ogen infection (Figure 6), while non-inoculated control
plants displayed no resistance.
Enhanced disease resistance of transgenic potato
plants
Potato is a very important agronomical crop worldwide that
suffers annual yield losses caused by several pathogens. To
study the response of 35S::NmDef02-transgenic potato
plants to A. solani and P. infestans under greenhouse
conditions, potato plants of transgenic 35S::NmDef02 lines
and vector control plants were inoculated with these patho-
gens. At 10 days post-inoculation, the control plants were
readily infected and disease symptoms were clearly
observed. As a consequence of disease progression, the
vector plants died while the transgenic lines remained
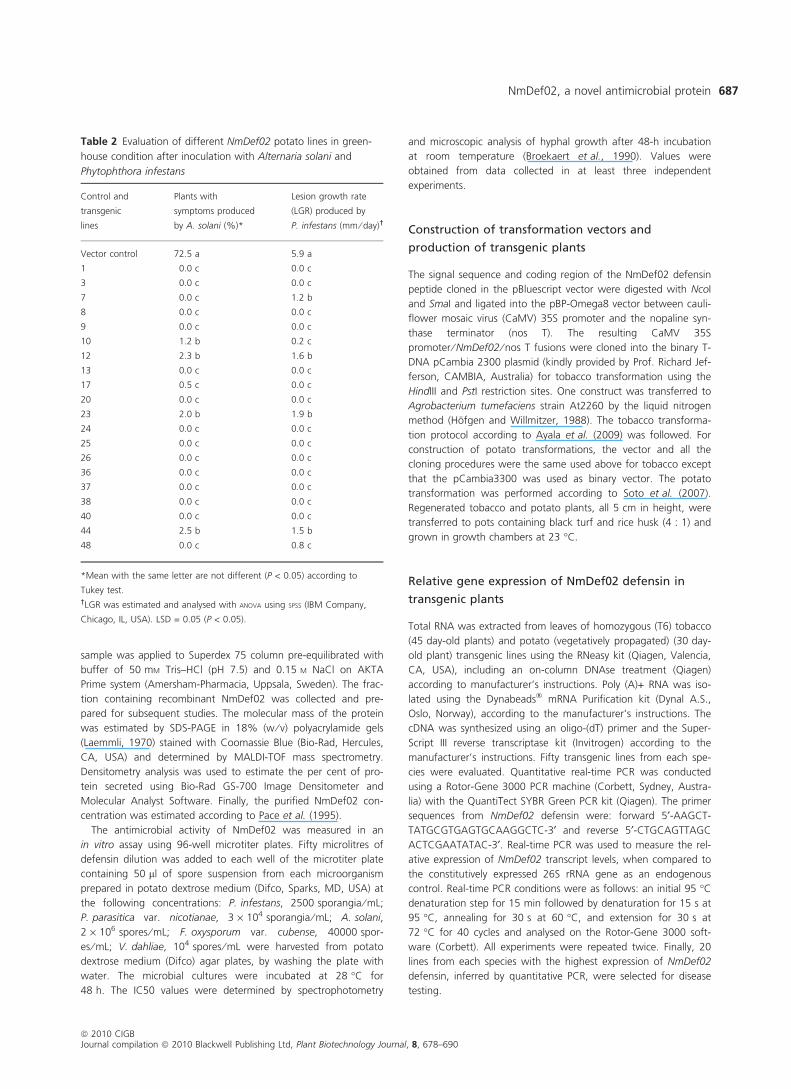
healthy with no obvious disease symptoms. As depicted in
Table 2, non-transformed lines used as controls displayed a
high degree (72.5%) of plants developing symptoms when
compared to the transgenic lines. In a separate experiment,
transgenic plants were also challenged with P. infestans,
causal agent of early blight, using the method described by
Vleeshouwers et al. (1999). Leaves of control plants
displayed typical spreading lesions, with water-soaked areas
and extensive rotting developed at later time points. In
contrast, all plants derived from NmDef02 lines displayed
resistance to P. infestans infection. At 10 days post-inocula-
tion, the lesion growth rate (LGR) found in potato transgenic
lines averaged from 0 to 1.9 mm ⁄ day, respectively. In
contrast to the NmDef02 transformed lines, inoculation of
non-transformed plants resulted in LGR of 5.9 mm ⁄ day
(Table 2).
Similar to tobacco transgenic lines, the level of disease
resistance correlated with the relative level of NmDef02
expression as determined by quantitative RT-PCR also in
potato transgenic lines. Potato transgenic lines 1, 3, 8, 9,
13, 20, 24, 25, 26, 36, 37 and 38 accumulated the high-
est transcript levels and displayed the highest degree of
resistance to both diseases (Table 2). According to these
results and similar to tobacco, the expression of NmDef02
(a) (b)
Figure 3 NmDef02 inhibits microbial growth in vitro. Purified
NmDef02 was evaluated for in vitro antimicrobial activity. Pre-germi-
nated zoospores of Phytophthora parasitica var. nicotianae
(3 · 104 zoospores ⁄ mL) were grown for 48 h at 23 �C in culture
medium in the absence (a) or presence (b) of 4 lM of NmDef02.
Table 1 Antimicrobial activity of NmDef02 on five plant
pathogens
Plant pathogens IC50* (lM)
Phytophthora infestans 1.4
Phytophthora parasitica var. nicotianae 4
Alternaria solani 2.8
Fusarium oxysporum var. cubense 1
Verticillium dahliae 2
*Protein molar concentrations (lM) required for 50% growth inhibition
(IC50) after 48 h of incubation were determined from the dose–response
curves (per cent growth inhibition vs. protein concentration). Five replicates
(N = 10) per experiments were performed.
683
ª 2010 CIGBJournal compilation ª 2010 Blackwell Publishing Ltd, Plant Biotechnology Journal, 8, 678–690
NmDef02, a novel antimicrobial protein
strongly increased the resistance of potato to A. solani
and P. infestans under greenhouse conditions.
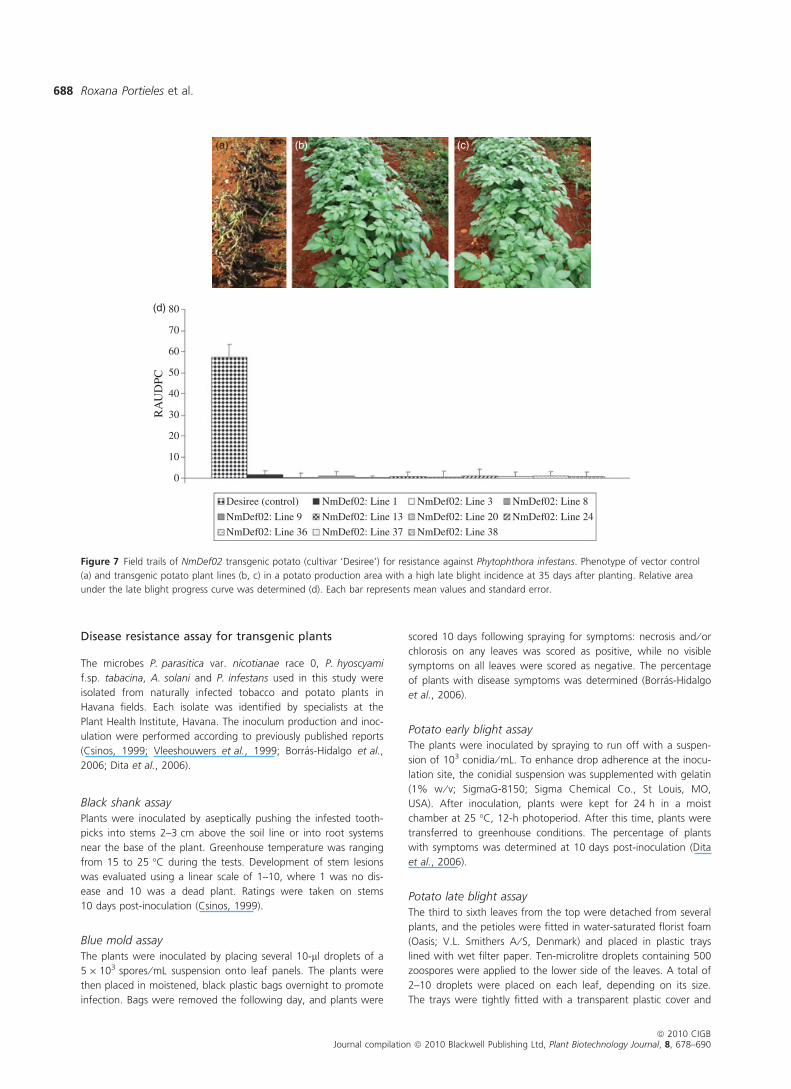
Finally, to determine the performance of NmDef02-trans-
genic plants in natural infections under field conditions, tri-
als were conducted in a potato field in Havana during the
cold and wet season when P. infestans poses a significant
problem for potato cultivation with a high inoculum pres-
sure. As shown in Figure 7, NmDef02 lines displayed high
levels of field resistance against P. infestans when
compared with control plants of the parental cultivar
(Desiree). The relative area under the disease progress curve
(RAUDPC) value in NmDef02 lines was significantly lower
than that of vector controls (Figure 7d), which is consistent
with results obtained in greenhouse experiments and con-
firm the in vivo activity of NmDef02 to control P. infestans.
Discussion
Numerous plant defensins have been purified from several
plant species and plant tissues that show a wide range of
in vitro biological activities including a-amylase activity,
ion-channel blocking and antifungal and antibacterial
activity (Thomma et al., 2002; Lay and Anderson, 2005).
In this study, we isolated a novel plant defensin induced
by the tobacco species N. megalosiphon during the inter-
action with the blue mould pathogen P. hyoscyami f.sp.
tabacina (Borras-Hidalgo et al., 2009). We isolated a cDNA
clone encoding the plant defensin, designated NmDef02,
from a subtractive cDNA library (Borras-Hidalgo et al.,
2006). Interestingly, phylogeny studies including a collec-
tion of 73 proteins showed that NmDef02 belongs to a
new subgroup of plant defensins. Thus, we conclude that
NmDef02 is a novel member of the plant defensin family.
Defensins have been shown to accumulate in tissues,
especially seeds, as part of normal development or matura-
tion, as a constitutive defence against pathogens (Thomma
et al., 2002). Plant defensins consisting of 45–54 amino
acids have a net positive charge and characteristic sequence
conservation (Thomma et al., 2003). In general, plant
defensins possess potent and broad-spectrum growth-inhib-
itory activity against fungi. The antimicrobial activity of
NmDef02 in this study was investigated with recombinant
NmDef02 produced in P. pastoris (Figure 2). The recombi-
nant NmDef02 defensin exhibited antimicrobial activity
against several important common plant pathogens; P. infe-
stans, P. parasitica var. nicotianae, A. solani, F. oxysporum
var. cubense and V. dahliae (Table 1). The activity was rang-
ing in IC50 value from 1 to 4 lM.
The important role played by antimicrobial proteins in
plant defence has been well documented (Broekaert et al.,
(a)
0
2
4
6
8
10
12
14
16
18
Con
trol 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50
Transgenic lines
Rel
ativ
e ex
pres
sion
(b)
0
2
4
6
8
10
12
14
16
18
Con
trol 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50
Transgenic lines
Rel
ativ
e ex
pres
sion
Figure 4 Relative expression of NmDef02
defensin gene in tobacco (a) and potato (b)
transgenic lines. Bars represent mean values
and standard error of the results obtained
from three replicates. Real-time PCR was
used to measure the relative expression of
transcript level of NmDef02 defensin gene,
when compared to the constitutively expres-
sed 26S rRNA gene as an endogenous
control.
684
ª 2010 CIGBJournal compilation ª 2010 Blackwell Publishing Ltd, Plant Biotechnology Journal, 8, 678–690
Roxana Portieles et al.

1995; Francois et al., 2002). Transgenic plants have the
potential to provide a generic resistance against different
pathogens and are likely to reduce dependence on chemi-
cal pesticides. The independent transgenic plants showed
a range of NmDef02 expression levels, which might be
because of position effects, the effects of transgene copy
number and ⁄ or silencing processes. The relative expression
level of NmDef02 in transgenic lines ranged between 1
and 15 (Figure 4). Strong evidences supporting the func-
tion of NmDef02 in plant defence derives from the
enhanced resistance of transgenic tobacco (Figure 5) and
potato plants (Figure 7) constitutively expressing NmDef02.
Analysis of the disease resistance of tobacco (Figure 6)
and potato (Table 2) transgenic lines revealed enhanced
resistance against P. parasitica var. nicotianae, P. hyoscy-
ami f.sp. tabacina, A. solani and P. infestans, respectively,
compared to wild type or transformants containing vector
only. There was a significant correlation between the lev-
els of NmDef02 and the degree of disease resistance
observed in the transgenic lines. We interestingly showed
for the first time a high level of potato resistance to
P. infestans using a plant defensin under greenhouse and
field conditions (Figure 7). Gao et al. (2000) used a plant
defensin to increase potato resistance to V. dahliae, but
(a)
_____________________ __________
(b)
_____________________ ___________ ____________________ NmDef02 transgenic tobacco
NmDef02 transgenic tobacco
Vector control
Vector control
NmDef02 transgenic tobacco
Figure 5 Greenhouse evaluation of NmDe-
f02 transgenic tobacco for resistance against
Phytophthora parasitica var. nicotianae (a)
and Peronospora hyoscyami f.sp. tabacina
(b). Phenotype of vector control and patho-
gen-infected transgenic tobacco plant in the
greenhouse at 10 days after inoculation.
685
ª 2010 CIGBJournal compilation ª 2010 Blackwell Publishing Ltd, Plant Biotechnology Journal, 8, 678–690
NmDef02, a novel antimicrobial protein
P. infestans resistance was not observed in the potato
transgenic lines evaluated in field condition. Additionally,
Solis et al. (2007) reported the isolation of a defensin
gene, lm-def, isolated from the Andean crop ‘maca’ (Lepi-
dium meyenii) with activity against the pathogen P. infe-
stans. However, the activity of the defensin was not
evaluated in transgenic plants.
All tobacco and potato transgenic plants were healthy
and showed no morphological or developmental abnor-
malities even in the lines with the highest NmDef02 accu-
mulation. Thus, the expression level of NmDef02 in
tobacco and potato plants was sufficient to obtain anti-
fungal activity without any toxic effects to the host plant.
Finally, the present investigation shows that the expression
of the NmDef02 defensin in transgenic potato plants dem-
onstrated the efficacy of defensins in imparting disease
resistance in crop plants and indicated that they are good
candidates for crop improvement.
Experimental procedures
Identification and cloning of full-length cDNA
NmDef02 cDNA was isolated from an SSH library prepared from
N. megalosiphon leaves inoculated with P. hyoscyami f.sp. tabacina
according to Borras-Hidalgo et al. (2006). The full-length coding
region for NmDef02 gene was obtained according to specifications
in the SMART RACE II kit (BD Clontech, Palo Alto, CA, USA) and
submitted to GenBank (accession number: FJ948813).
Expression in yeast, purification and anti-microbial
assay of recombinant NmDef02
The DNA sequence coding for the mature NmDef02 sequence was
cloned without signal peptide into the pPIC9 expression vector
in-frame with the secretion signal a-mating factor, downstream of
the alcohol oxidase I promoter. A sequence of KEX2 sites compro-
mising 12 nucleotides was placed ahead of the 141 bp NmDef02
cDNA for cleaving the signal peptide. While the sequence encod-
ing, the STE13 cleavage sites (Glu–Ala–Glu–ala) in pPIC9 was
deleted because STE13 protease cleavage of Glu-Ala repeats is not
efficient and these repeats are not necessary for cleavage by KEX2.
A stop codon and an EcoRI restriction site were added at the
3¢-end. The NmDef02 fragment was isolated from pGEM-T plasmid
by digestion with XhoI and EcoRI enzymes and ligated into the
EcoRI- and XhoI-digested pPIC9. After transformed into Escherichia
coli JM109 host strain, the plasmid pPIC9 ⁄ NmDef02 was isolated
and linearized with SalI to favour integration at the his4 locus of
P. pastoris (GS115; Invitrogen, Carlsbad, CA, USA) genome, and
transformed into yeast by electroporation (according to the manual
of the Pichia expression kit; Invitrogen). The N-terminal sequencing
of recombinant NmDef02 and mass spectroscopy experiments con-
firmed that the recombinant NmDef02 was expressed correctly.
The supernatant was precipitated by saturated ammonium
sulphate, and the pellet was resolved in 20 mM Tris–HCl (pH 7.5)
buffer. After desalting and concentrating by ultrafiltration, the
(a)
1
2
3
4
5
6
7
8
9
10
Con
trol 4 5 8 15 16 17 18 19 27 28 32 33 34 35 39 40 41 42 43 49
Transgenic lines
Stem
dis
ease
rat
ing
(b)
0
10
20
30
40
50
60
70
80
90
100
Con
trol 4 5 8 15 16 17 18 19 27 28 32 33 34 35 39 40 41 42 43 49
Transgenic lines
Plan
ts w
ith s
ymto
ms
(%)
Figure 6 Evaluation of different NmDef02
tobacco lines in greenhouse condition after
inoculation with Phytophthora parasitica var.
nicotianae (a) and Peronospora hyoscyami
f.sp. tabacina (b) at 10 days after inocula-
tion. One hundred plants were used per
transgenic line. Each bar represents mean
values with standard errors.
686
ª 2010 CIGBJournal compilation ª 2010 Blackwell Publishing Ltd, Plant Biotechnology Journal, 8, 678–690
Roxana Portieles et al.
sample was applied to Superdex 75 column pre-equilibrated with
buffer of 50 mM Tris–HCl (pH 7.5) and 0.15 M NaCl on AKTA
Prime system (Amersham-Pharmacia, Uppsala, Sweden). The frac-
tion containing recombinant NmDef02 was collected and pre-
pared for subsequent studies. The molecular mass of the protein
was estimated by SDS-PAGE in 18% (w ⁄ v) polyacrylamide gels
(Laemmli, 1970) stained with Coomassie Blue (Bio-Rad, Hercules,
CA, USA) and determined by MALDI-TOF mass spectrometry.
Densitometry analysis was used to estimate the per cent of pro-
tein secreted using Bio-Rad GS-700 Image Densitometer and
Molecular Analyst Software. Finally, the purified NmDef02 con-
centration was estimated according to Pace et al. (1995).
The antimicrobial activity of NmDef02 was measured in an
in vitro assay using 96-well microtiter plates. Fifty microlitres of
defensin dilution was added to each well of the microtiter plate
containing 50 ll of spore suspension from each microorganism
prepared in potato dextrose medium (Difco, Sparks, MD, USA) at
the following concentrations: P. infestans, 2500 sporangia ⁄ mL;
P. parasitica var. nicotianae, 3 · 104 sporangia ⁄ mL; A. solani,
2 · 106 spores ⁄ mL; F. oxysporum var. cubense, 40000 spor-
es ⁄ mL; V. dahliae, 104 spores ⁄ mL were harvested from potato
dextrose medium (Difco) agar plates, by washing the plate with
water. The microbial cultures were incubated at 28 �C for
48 h. The IC50 values were determined by spectrophotometry
and microscopic analysis of hyphal growth after 48-h incubation
at room temperature (Broekaert et al., 1990). Values were
obtained from data collected in at least three independent
experiments.
Construction of transformation vectors and
production of transgenic plants
The signal sequence and coding region of the NmDef02 defensin
peptide cloned in the pBluescript vector were digested with NcoI
and SmaI and ligated into the pBP-Omega8 vector between cauli-
flower mosaic virus (CaMV) 35S promoter and the nopaline syn-
thase terminator (nos T). The resulting CaMV 35S
promoter ⁄ NmDef02 ⁄ nos T fusions were cloned into the binary T-
DNA pCambia 2300 plasmid (kindly provided by Prof. Richard Jef-
ferson, CAMBIA, Australia) for tobacco transformation using the
HindIII and PstI restriction sites. One construct was transferred to
Agrobacterium tumefaciens strain At2260 by the liquid nitrogen
method (Hofgen and Willmitzer, 1988). The tobacco transforma-
tion protocol according to Ayala et al. (2009) was followed. For
construction of potato transformations, the vector and all the
cloning procedures were the same used above for tobacco except
that the pCambia3300 was used as binary vector. The potato
transformation was performed according to Soto et al. (2007).
Regenerated tobacco and potato plants, all 5 cm in height, were
transferred to pots containing black turf and rice husk (4 : 1) and
grown in growth chambers at 23 �C.
Relative gene expression of NmDef02 defensin in
transgenic plants
Total RNA was extracted from leaves of homozygous (T6) tobacco
(45 day-old plants) and potato (vegetatively propagated) (30 day-
old plant) transgenic lines using the RNeasy kit (Qiagen, Valencia,
CA, USA), including an on-column DNAse treatment (Qiagen)
according to manufacturer’s instructions. Poly (A)+ RNA was iso-
lated using the Dynabeads� mRNA Purification kit (Dynal A.S.,
Oslo, Norway), according to the manufacturer’s instructions. The
cDNA was synthesized using an oligo-(dT) primer and the Super-
Script III reverse transcriptase kit (Invitrogen) according to the
manufacturer’s instructions. Fifty transgenic lines from each spe-
cies were evaluated. Quantitative real-time PCR was conducted
using a Rotor-Gene 3000 PCR machine (Corbett, Sydney, Austra-
lia) with the QuantiTect SYBR Green PCR kit (Qiagen). The primer
sequences from NmDef02 defensin were: forward 5¢-AAGCT-
TATGCGTGAGTGCAAGGCTC-3¢ and reverse 5¢-CTGCAGTTAGC
ACTCGAATATAC-3¢. Real-time PCR was used to measure the rel-
ative expression of NmDef02 transcript levels, when compared to
the constitutively expressed 26S rRNA gene as an endogenous
control. Real-time PCR conditions were as follows: an initial 95 �Cdenaturation step for 15 min followed by denaturation for 15 s at
95 �C, annealing for 30 s at 60 �C, and extension for 30 s at
72 �C for 40 cycles and analysed on the Rotor-Gene 3000 soft-
ware (Corbett). All experiments were repeated twice. Finally, 20
lines from each species with the highest expression of NmDef02
defensin, inferred by quantitative PCR, were selected for disease
testing.
Table 2 Evaluation of different NmDef02 potato lines in green-
house condition after inoculation with Alternaria solani and
Phytophthora infestans
Control and
transgenic
lines
Plants with
symptoms produced
by A. solani (%)*
Lesion growth rate
(LGR) produced by
P. infestans (mm ⁄ day)†
Vector control 72.5 a 5.9 a
1 0.0 c 0.0 c
3 0.0 c 0.0 c
7 0.0 c 1.2 b
8 0.0 c 0.0 c
9 0.0 c 0.0 c
10 1.2 b 0.2 c
12 2.3 b 1.6 b
13 0.0 c 0.0 c
17 0.5 c 0.0 c
20 0.0 c 0.0 c
23 2.0 b 1.9 b
24 0.0 c 0.0 c
25 0.0 c 0.0 c
26 0.0 c 0.0 c
36 0.0 c 0.0 c
37 0.0 c 0.0 c
38 0.0 c 0.0 c
40 0.0 c 0.0 c
44 2.5 b 1.5 b
48 0.0 c 0.8 c
*Mean with the same letter are not different (P < 0.05) according to
Tukey test.†LGR was estimated and analysed with ANOVA using SPSS (IBM Company,
Chicago, IL, USA). LSD = 0.05 (P < 0.05).
687
ª 2010 CIGBJournal compilation ª 2010 Blackwell Publishing Ltd, Plant Biotechnology Journal, 8, 678–690
NmDef02, a novel antimicrobial protein
Disease resistance assay for transgenic plants
The microbes P. parasitica var. nicotianae race 0, P. hyoscyami
f.sp. tabacina, A. solani and P. infestans used in this study were
isolated from naturally infected tobacco and potato plants in
Havana fields. Each isolate was identified by specialists at the
Plant Health Institute, Havana. The inoculum production and inoc-
ulation were performed according to previously published reports
(Csinos, 1999; Vleeshouwers et al., 1999; Borras-Hidalgo et al.,
2006; Dita et al., 2006).
Black shank assay
Plants were inoculated by aseptically pushing the infested tooth-
picks into stems 2–3 cm above the soil line or into root systems
near the base of the plant. Greenhouse temperature was ranging
from 15 to 25 �C during the tests. Development of stem lesions
was evaluated using a linear scale of 1–10, where 1 was no dis-
ease and 10 was a dead plant. Ratings were taken on stems
10 days post-inoculation (Csinos, 1999).
Blue mold assay
The plants were inoculated by placing several 10-ll droplets of a
5 · 103 spores ⁄ mL suspension onto leaf panels. The plants were
then placed in moistened, black plastic bags overnight to promote
infection. Bags were removed the following day, and plants were
scored 10 days following spraying for symptoms: necrosis and ⁄ or
chlorosis on any leaves was scored as positive, while no visible
symptoms on all leaves were scored as negative. The percentage
of plants with disease symptoms was determined (Borras-Hidalgo
et al., 2006).
Potato early blight assay
The plants were inoculated by spraying to run off with a suspen-
sion of 103 conidia ⁄ mL. To enhance drop adherence at the inocu-
lation site, the conidial suspension was supplemented with gelatin
(1% w ⁄ v; SigmaG-8150; Sigma Chemical Co., St Louis, MO,
USA). After inoculation, plants were kept for 24 h in a moist
chamber at 25 �C, 12-h photoperiod. After this time, plants were
transferred to greenhouse conditions. The percentage of plants
with symptoms was determined at 10 days post-inoculation (Dita
et al., 2006).
Potato late blight assay
The third to sixth leaves from the top were detached from several
plants, and the petioles were fitted in water-saturated florist foam
(Oasis; V.L. Smithers A ⁄ S, Denmark) and placed in plastic trays
lined with wet filter paper. Ten-microlitre droplets containing 500
zoospores were applied to the lower side of the leaves. A total of
2–10 droplets were placed on each leaf, depending on its size.
The trays were tightly fitted with a transparent plastic cover and
(d) 80
70
60
50
40
30RA
UD
PC
20
10
0
Desiree (control)
NmDef02: Line 9
NmDef02: Line 1
NmDef02: Line 13
NmDef02: Line 37
NmDef02: Line 3
NmDef02: Line 20
NmDef02: Line 38
NmDef02: Line 8
NmDef02: Line 24
NmDef02: Line 36
(a) (b) (c)
Figure 7 Field trails of NmDef02 transgenic potato (cultivar ‘Desiree’) for resistance against Phytophthora infestans. Phenotype of vector control
(a) and transgenic potato plant lines (b, c) in a potato production area with a high late blight incidence at 35 days after planting. Relative area
under the late blight progress curve was determined (d). Each bar represents mean values and standard error.
688
ª 2010 CIGBJournal compilation ª 2010 Blackwell Publishing Ltd, Plant Biotechnology Journal, 8, 678–690
Roxana Portieles et al.

placed under fluorescent light in a regulated growth chamber
(15 �C for a 16-hr photoperiod). Inoculation spots were examined
for disease symptoms and necrosis daily for 10 days. The largest
length and width (perpendicular to the length) of each lesion
were measured, and the ellipse area (A = 1 ⁄ 4 · p · length ·width) was calculated. Analysis of LGR was estimated by linear
regression over time (Vleeshouwers et al., 1999).
One hundred plants of each of the homozygous transgenic
tobacco (T6) and potato lines (vegetatively propagated) were used
in the experiments under greenhouse conditions. Subsequently,
field evaluation of potato expressing NmDef02 was made. To
determine the performance of NmDef02 under field conditions,
trials were conducted in a potato production area in Havana with
a high inoculum pressure where P. infestans is a significant prob-
lem for potato production each year. During cold and wet season
of 2008, a total of 10 transgenic NmDef02 lines and transgenic
control lines (cultivar ‘Desiree’ transformed with the vector) were
evaluated by planting 100 tubers of each transgenic homozygous
line in the potato production area with a high late blight inci-
dence. The tubers were planted with a random design to look at
positional effects in the field. The RAUDPC was calculated accord-
ing to Kirk et al. (2001). Data were analysed by analysis of vari-
ance or general linear model procedures of SAS (SAS Institute,
Cary, NC, USA). Significant difference among means was deter-
mined by Fisher’s least significant difference mean separation at
P = 0.05.
Acknowledgements
We are grateful to the Cuban State Council for providing
support of this research. The authors would like to thanks
Dr Muluneh Tamiru Oil for comments and critical reading
of the manuscript. The authors thank to anonymous
reviewers and editor for useful suggestion.
References
Ayala, M., Gavilondo, J., Rodrıguez, M., Fuentes, A., Enrıquez, G.,
Perez, L., Cremata, J. and Pujol, M. (2009) Production of
Plantibodies in Nicotiana plants. in: Methods in Molecular
Biology. Recombinant Proteins from Plants (Faye, L. and
Gomord, V., eds), pp. 103–134. New York: Humana Press.
Borras-Hidalgo, O., Thomma, B., Collazo, C., Chacon, O.,
Borroto, C.J., Ayra, C., Portieles, R., Lopez, Y. and Pujol, M.
(2006) EIL2 transcription factor and glutathione synthetase are
required for defense of tobacco against tobacco blue mold.
Mol. Plant Microbe Interact. 19, 399–406.
Borras-Hidalgo, O., Thomma, B., Silva, Y., Chacon, O. and Pujol,
M. (2010) Tobacco blue mould disease caused by Peronospora
hyoscyami f. sp. tabacina. Mol. Plant Pathol. 11, 13–18.
Broekaert, W.F., Terras, F.R.G., Cammue, B.P.A. and
Vanderleyden, J. (1990) An automated quantitative assay for
fungal growth inhibition. FEMS Microbiol. Lett. 69, 55–60.
Broekaert, W.F., Terras, F.R., Cammue, B.P. and Osborn, R.W.
(1995) Plant defensins: novel antimicrobial peptides as
components of the host defense system. Plant Physiol. 108,
1353–1358.
Bruce, T.J. and Pickett, J.A. (2007) Plant defence signalling
induced by biotic attacks. Curr. Opin. Plant Biol. 10, 387–392.
Chacon, O., Hernandez, I., Portieles, R., Lopez, Y., Pujol, M. and
Borras-Hidalgo, O. (2009) Identification of defense-related
genes in tobacco responding to black shank disease. Plant Sci.
177, 175–180.
Collazo, C., Ramos, P.L., Chacon, O., Borroto, C.J., Lopez, Y.,
Pujol, M., Thomma, B.P.H.J., Hein, I. and Borras-Hidalgo, O.
(2006) Phenotypical and molecular characterization of the
tomato mottle taino virus – Nicotiana megalosiphon interaction.
Physiol. Mol. Plant Pathol. 67, 231–236.
Csinos, A.S. (1999) Stem and root resistance to tobacco black
shank. Plant Dis. 83, 777–780.
De-Paula, V.S., Rasear, G., Medeiros, L., Miyamoto, C.A.,
Almeida, M.S., Kurtenbach, E., Almeida, F.C.L. and Valente,
A.P. (2008) Evolutionary relationship between defensins in the
Poaceae family strengthened by the characterization of new
sugarcane defensins. Plant Mol. Biol., 68, 321–335.
Dita, M.A., Brommonschenkel, S.H., Matzuoka, K. and Mizubuti, E.
(2006) Components of resistance to early blight in four potato
cultivars: effect of leaf position. J. Phytopathol. 153, 1–6.
Do, H.M., Lee, S.C., Jung, H.W., Sohn, K.H. and Hwang, B.K.
(2004) Differential expression and in situ localization of a
pepper defensin (CADEF1) gene in response to pathogen
infection, abiotic elicitors and environmental stresses in
Capsicum annuum. Plant Sci. 166, 1297–1305.
Fradin, E.F. and Thomma, B.P.H.J. (2006) Physiology and
molecular aspects of Verticillium wilt diseases caused by
V. dahliae and V. albo-atrum. Mol. Plant Pathol. 7, 71–86.
Francois, I.E., De Bolle, M.F., Dwyer, G., Goderis, I.J., Woutors, P.F.
and Verhaert, P.D. (2002) Transgenic expression in Arabidopsis
of a polyprotein construct leading to production of two different
antimicrobial proteins. Plant Physiol. 128, 1346–1358.
Gao, A.G., Hakimi, S.M., Mittanck, C.A., Wu, Y., Woerner, B.M.,
Stark, D.M., Shah, D.M., Liang, J. and Rommens, C.M. (2000)
Fungal pathogen protection in potato by expression of a plant
defensin peptide. Nat. Biotechnol. 18, 1307–1310.
Hofgen, R. and Willmitzer, L. (1988) Storage of competent cells
for Agrobacterium transformation. Nucleic Acids Res. 16, 9877.
Kirk, W.W., Felcher, K.J., Douches, D.S., Coombs, J.M., Stein,
J.M., Baker, K.M. and Hammerschmidt, R. (2001) Effect of host
plant resistance and reduced rates and frequencies of fungicide
application to control potato late blight. Plant Dis. 85, 1113–
1118.
Laemmli, U.K. (1970) Cleavage of structural proteins during the
assembly of the head of bacteriophage T4. Nature, 227, 681–
685.
Lay, F.T., Schirra, H.J., Scanlon, M.J, Anderson, M.A. and Craik,
D.J. (2003) The three-dimensional solution structure of NaD1, a
new floral defensin from Nicotiana alata and its application to
homology model of the crop defensin alfAFP. J. Mol. Bio., 325,
175–188.
Lay, F.T. and Anderson, M.A. (2005) Defensins – components of
the innate immune system in plants. Curr. Protein Pept. Sci. 6,
85–101.
Liu, Y.J., Cheng, C.S., Lai, S.M., Hsu, M.P., Chen, C.S. and Lyu,
P.C. (2006) Solution structure of the plant defensin VrD1 from
mung bean and its possible role in insecticidal activity against
bruchids. Proteins, 63, 777–786.
689
ª 2010 CIGBJournal compilation ª 2010 Blackwell Publishing Ltd, Plant Biotechnology Journal, 8, 678–690
NmDef02, a novel antimicrobial protein

Meyer, B., Houlne, G., Pozueta-Romero, J., Schantz, M.L. and
Schantz, R. (1996) Fruit-specific expression of a defensin-type
gene family in bell pepper. Plant Physiol. 112, 615–622.
Moreno, M., Segura, A. and Garcia-Olmedo, F. (1994)
Pseudothionin-St1, a potato peptide active against potato
pathogens. Eur. J. Biochem. 223, 135–139.
Osborn, R.W., De Samblanx, G.W., Thevissen, K., Goderis, I.,
Torrekens, S., Van Leuven, F., Attenborough, S., Rees, S.B. and
Broekaert, W.F. (1995) Isolation and characterisation of plant
defensins from seeds of Asteraceae, Fabaceae,
Hippocastanaceae and Saxifragaceae. FEBS Lett. 368, 257–262.
Pace, C.N., Vajdos, F., Fee, L., Grimsley, G. and Gray, T. (1995)
How to measure and predict the molar absorption coefficient
of a protein. Protein Sci. 4, 2411–2423.
Parashina, E.V., Serdobinskii, L.A., Kalle, E.G., Lavorova, N.V.,
Avetisov, V.A., Lunin, V.G. and Naroditskii, B.S. (2000) Genetic
engineering of oilseed rape and tomato plants expressing a
radish defensin gene. Russ. J. Plant Physiol. 47, 417–423.
Park, H.C., Kang, Y.H., Chun, H.J., Koo, J.C., Cheong, Y.H., Kim,
C.Y., Kim, M.C., Chung, W.S., Kim, J.C. and Yoo, J.H. (2002)
Characterization of a stamen specific cDNA encoding a novel
plant defensin in Chinese cabbage. Plant Mol. Biol. 50, 59–69.
Penninckx, I.A., Eggermont, K., Terras, F.R., Thomma, B.P., De
Samblanx, G.W., Buchala, A., Metraux, J.P., Manners, J.M. and
Broekaert, W.F. (1996) Pathogen-induced systemic activation of
a plant defensin gene in Arabidopsis follows a salicylic acid-
independent pathway. Plant Cell, 8, 2309–2323.
Saitou, N. and Nei, M. (1987) The neighbor-joining method:
a new method for reconstructing phylogenetic trees. Mol. Biol.
Evol. 4, 406–425.
Sels, J., Mathys, J., De Coninck, B.M.A., Cammue, B.P.A. and
De Bolle, M.F.C. (2008) Plant pathogenesis-related (PR)
proteins: a focus on PR peptides. Plant Physiol. Biochem. 46,
941–950.
Solis, J., Medrano, G. and Ghislain, M. (2007) Inhibitory effect of
a defensin gene from the Andean crop maca (Lepidium
meyenii) against Phytophthora infestans. J. Plant Physiol. 164,
1071–1082.
Soto, N., Enrıquez, G.A., Ferreira, A., Corrada, M., Fuentes, A.,
Tiel, K. and Pujol, M. (2007) Efficient transformation of potato
stems segments from cultivar Desiree, using phosphinothricin as
selection marker. Biotecnol. Apl. 24, 139–144.
Spelbrink, R.G., Dilmac, N., Allen, A., Smith, T.J., Shah, D.M. and
Hockerman, G.H. (2004) Differential antifungal and calcium
channel-blocking activity among structurally related plant
defensins. Plant Physiol. 135, 2055–2067.
Terras, F.R., Schoofs, H.M., De Bolle, M.F., Van Leuven, F., Rees,
S.B., Vanderleyden, J., Cammue, B.P. and Broekaert, W.F.
(1992) Analysis of two novel classes of plant antifungal proteins
from radish (Raphanus sativus L.) seeds. J. Biol. Chem. 267,
15301–15309.
Terras, F.R., Eggermont, K., Kovaleva, V., Raikhel, N.V., Osborn,
R.W., Kester, A., Rees, S.B., Vanderleyden, J., Cammue, B.P.
and Broekaert, W.F. (1995) Small cysteine-rich antifungal
proteins from radish: their role in host defense. Plant Cell, 7,
573–588.
Thevissen, K., Cammue, B.P.A., Lemaire, K., Winderickx, J.,
Dickson, R.C., Lester, R.L., Ferket, K.K., Van Even, F., Parret,
A.H. and Broekaert, W.F. (2000) A gene encoding a sphingolipid
biosynthesis enzyme determines the sensitivity of Saccharomyces
cerevisiae to an antifungal plant defensin from dahlia (Dahlia
merckii). Proc. Natl Acad. Sci. USA, 97, 9531–9536.
Thevissen, K., Warnecke, D.C., Francois, I.E.J.A., Leipelt, M.,
Heinz, E., Ott, C., Zahringer, U., Thomma, B.P.H.J., Ferket, K.K.
and Cammue, B.P.A. (2004) Defensins from insects and plants
interact with fungal glucosylceramides. J. Biol. Chem. 279,
3900–3905.
Thevissen, K., Kristensen, H.H., Thomma, B.P.H.J., Cammue,
B.P.A. and Francois, I.E.J.A. (2007) Therapeutic potential of
antifungal plant and insect defensins. Drug Discov. Today, 12,
966–971.
Thomma, B.P.H.J. and Broekaert, W.F. (1998) Tissue-specific
expression of plant defensin genes PDF2.1 and PDF2.2 in
Arabidopsis thaliana. Plant Physiol. Biochem. 36, 533–537.
Thomma, B.P.H.J., Cammue, B.P.A. and Thevissen, K. (2002) Plant
defensins. Planta, 216, 193–202.
Thomma, B.P.H.J., Cammue, B.P.A. and Thevissen, K. (2003)
Mode of action of plant defensins suggests therapeutic
potential. Curr. Drug Targets Infect. Disord. 3, 1–8.
Thompson, J.D., Gibson, T.J., Plewniak, F., Jeanmougin, F. and
Higgins, D.G. (1997) The CLUSTAL X windows interface:
flexible strategies for multiple sequence alignment aided by
quality analysis tools. Nucleic Acids Res. 25, 4876–4882.
Vleeshouwers, V.G.A.A., van Dooijeweert, W., Keizer, L.C.P.,
Sijpkes, L., Govers, F. and Colon, L.T. (1999) A laboratory assay
for Phytophthora infestans resistance in various Solanum
species reflects the field situation. Eur. J. Plant Pathol. 105,
241–250.
Wang, Y.P., Nowak, G., Culley, D., Hadwiger, L.A. and Fristensky,
B. (1999) Constitutive expression of pea defense gene DRR206
confers resistance to blackleg (Leptosphaeria maculans) disease
in transgenic canola (Brassica napus). Mol. Plant Microbe
Interact. 12, 410–418.
Zhu, Y.J., Agbayani, R. and Moore, P.H. (2007) Ectopic expression
of Dahlia merckii defensin DmAMP1 improves papaya resistance
to Phytophthora palmivora by reducing pathogen vigor. Planta,
226, 87–97.
690
ª 2010 CIGBJournal compilation ª 2010 Blackwell Publishing Ltd, Plant Biotechnology Journal, 8, 678–690
Roxana Portieles et al.
Copyright © 2022 FDOKUMEN




















