New metallothionein assay in Scrobicularia plana: Heating effect and correlation with other...
8
New metallothionein assay in Scrobicularia plana: Heating effect and correlation with other biomarkers Antonio Romero-Ruiz a, 1 , Jose ´ Alhama a, * , 1 , Julia ´ n Blasco b , Jose ´ Luis Go ´ mez-Ariza c , Juan Lo ´ pez-Barea a a Department of Biochemistry and Molecular Biology, University of Co ´rdoba, Co ´rdoba, Spain b Andalusia Institute of Marine Sciences-CSIC, Rı ´o San Pedro Campus, Puerto Real, Ca ´diz, Spain c Department of Chemistry and Materials Sciences, University of Huelva, Huelva, Spain Quantification of total MTs by RP-HPLC-FD in non-heated bivalve extracts allows metal contamination assessment with high sensitivity and specificity. article info Article history: Received 11 December 2007 Received in revised form 22 February 2008 Accepted 29 February 2008 Keywords: Bivalve molluscs Metals Reversed-phase high performance liquid chromatography Fluorescence detection Don ˜ ana National Park Guadalquivir Estuary Aznalco ´ llar pyrite mine abstract Metallothionein (MT) and other biomarker levels were measured in Scrobicularia plana clams to assess pollution of the Guadalquivir Estuary possibly affected by metals released from Aznalco ´ llar pyrite mine in 1998. After optimizing reagent concentrations for monobromobimane derivatization, MT levels were quantified by reversed-phase high-performance liquid chromatography coupled to fluorescence de- tection (RP-HPLC-FD) in heated or unheated digestive gland extracts and compared to those obtained by differential pulse polarography (DPP). MT content assayed by RP-HPLC-FD in unheated samples was higher than that obtained by DPP and correlated better with metals and anti-oxidant activities. MT assay by RP-HPLC-FD in unheated extracts would be preferable for assessing metal pollution, due to its greater sensitivity and specificity. In addition to MT induction, glyoxalase II inhibition was well correlated with metal contents. Our results suggest that metals at the estuary do not originate from Aznalco ´ llar spill, but from those carried along by the river and deposited at its concave bank. Ó 2008 Elsevier Ltd. All rights reserved. 1. Introduction Transition metals convert O 2 into reactive oxygen species (ROS) that attack many biomolecules, such as fatty acids that generate malondialdehyde (MDA), a major oxidized by-product. Aerobic organisms have several defence lines from oxidative stress: primary anti-oxidant enzymes, such as catalase, detoxify ROS, while ancillary anti-oxidant enzymes, like 6-phosphogluc- onate dehydrogenase (6-PGDH), recycle NADP þ to its reduced form, which is consumed by glutathione reductase (GRase) (Sies, 1986). In addition, glyoxalases (Glyox) I and II use GSH to de- toxify a-oxoaldehydes, derived from cellular oxidative processes (Regoli et al., 1997) and glutathione-S-transferase (GST) conju- gates GSH to electrophilic compounds. Some GSTs also detoxify reactive products derived from lipid peroxidation (Jensson et al., 1986). Don ˜ana National Park (DNP, SW Spain) is located north of Guadalquivir Estuary and was declared by UNESCO a ‘‘World Re- serve of the Biosphere’’. It has been an area of concern since the tailings spill from Aznalco ´llar pyrite mine (April 1998), which released large amounts of acid water and mud full of toxic metals (Fe, Zn, Pb, As, Cu, Cd, Ag, Ni, Mn, Cr) via Guadiamar stream, close to DNP (Achterberg et al., 1999; Grimalt et al., 1999). Some bi- ological effects of this spill have been studied by our group in Algerian mice (Mus spretus) and clams (Scrobicularia plana). Mice living near Guadiamar stream showed greater anti-oxidant and detoxifying activities than controls (Ruiz-Laguna et al., 2001), and Abbreviations: Bh, Bonanza harbour; BT, ‘‘Brazo de la Torre’’; DNP, Don ˜ana Na- tional Park; DTT, dithiothreitol; DPP, differential pulse polarography; EDTA, ethyl- enediaminetetraacetic acid; Glyox, glyoxalase; GRase, glutathione reductase; GSH, reduced glutathione; GSSG, oxidized glutathione; GST, glutathione-S-transferase; MDA, malondialdehyde; MT, metallothionein; mBBr, monobromobimane; PAGE, polyacrylamide gel electrophoresis; 6PGDH, 6-phosphogluconate dehydrogenase; PMSF, phenylmethylsulphonylfluoride; ROS, reactive oxygen species; RP-HPLC-FD, reversed-phase high-performance liquid chromatography coupled to fluorescence detection; SDS, sodium dodecyl sulphate; SRs, ‘‘San Rafael’’ salt works; TFA, tri- fluoroacetic acid; ww, wet weight. * Corresponding author. S. Ochoa Building, Rabanales Campus, Highway A4, Km 396a, 14071 Co ´ rdoba, Spain. Tel.: þ34 957 218082; Fax: þ34 957 218688. E-mail address: [email protected] (J. Alhama). 1 J.A. and A.R.-R. contributed equally to this paper and should be considered as first authors. Environmental Pollution 156 (2008) 1340–1347 Contents lists available at ScienceDirect Environmental Pollution journal homepage: www.elsevier.com/locate/envpol 0269-7491/$ – see front matter Ó 2008 Elsevier Ltd. All rights reserved. doi:10.1016/j.envpol.2008.02.022
Transcript of New metallothionein assay in Scrobicularia plana: Heating effect and correlation with other...

lable at ScienceDirect
Environmental Pollution 156 (2008) 1340–1347
lable at ScienceDirect
Contents lists avaiContents lists avaiEnvironmental Pollution
journal homepage: www.elsevier .com/locate/envpol
Environmental Pollution
journal homepage: www.elsevier .com/locate/envpol
New metallothionein assay in Scrobicularia plana: Heating effect andcorrelation with other biomarkers
Antonio Romero-Ruiz a,1, Jose Alhama a,*,1, Julian Blasco b, Jose Luis Gomez-Ariza c,Juan Lopez-Barea a
a Department of Biochemistry and Molecular Biology, University of Cordoba, Cordoba, Spainb Andalusia Institute of Marine Sciences-CSIC, Rıo San Pedro Campus, Puerto Real, Cadiz, Spainc Department of Chemistry and Materials Sciences, University of Huelva, Huelva, Spain
Quantification of total MTs by RP-HPLC-FD in non-heated bivalve extr
acts allows metal contamination assessment with high sensitivityand specificity.a r t i c l e i n f o
Article history:Received 11 December 2007Received in revised form 22 February 2008Accepted 29 February 2008
Keywords:Bivalve molluscsMetalsReversed-phase high performance liquidchromatographyFluorescence detectionDonana National ParkGuadalquivir EstuaryAznalcollar pyrite mine
Abbreviations: Bh, Bonanza harbour; BT, ‘‘Brazo dtional Park; DTT, dithiothreitol; DPP, differential pulsenediaminetetraacetic acid; Glyox, glyoxalase; GRasereduced glutathione; GSSG, oxidized glutathione; GSMDA, malondialdehyde; MT, metallothionein; mBBrpolyacrylamide gel electrophoresis; 6PGDH, 6-phospPMSF, phenylmethylsulphonylfluoride; ROS, reactivereversed-phase high-performance liquid chromatogrdetection; SDS, sodium dodecyl sulphate; SRs, ‘‘Sanfluoroacetic acid; ww, wet weight.
* Corresponding author. S. Ochoa Building, Rabana396a, 14071 Cordoba, Spain. Tel.: þ34 957 218082; Fa
E-mail address: [email protected] (J. Alhama).1 J.A. and A.R.-R. contributed equally to this paper
first authors.
0269-7491/$ – see front matter � 2008 Elsevier Ltd.doi:10.1016/j.envpol.2008.02.022
a b s t r a c t
Metallothionein (MT) and other biomarker levels were measured in Scrobicularia plana clams to assesspollution of the Guadalquivir Estuary possibly affected by metals released from Aznalcollar pyrite mine in1998. After optimizing reagent concentrations for monobromobimane derivatization, MT levels werequantified by reversed-phase high-performance liquid chromatography coupled to fluorescence de-tection (RP-HPLC-FD) in heated or unheated digestive gland extracts and compared to those obtained bydifferential pulse polarography (DPP). MT content assayed by RP-HPLC-FD in unheated samples washigher than that obtained by DPP and correlated better with metals and anti-oxidant activities. MT assayby RP-HPLC-FD in unheated extracts would be preferable for assessing metal pollution, due to its greatersensitivity and specificity. In addition to MT induction, glyoxalase II inhibition was well correlated withmetal contents. Our results suggest that metals at the estuary do not originate from Aznalcollar spill, butfrom those carried along by the river and deposited at its concave bank.
� 2008 Elsevier Ltd. All rights reserved.
1. Introduction
Transition metals convert O2 into reactive oxygen species(ROS) that attack many biomolecules, such as fatty acids thatgenerate malondialdehyde (MDA), a major oxidized by-product.Aerobic organisms have several defence lines from oxidative
e la Torre’’; DNP, Donana Na-e polarography; EDTA, ethyl-, glutathione reductase; GSH,T, glutathione-S-transferase;, monobromobimane; PAGE,hogluconate dehydrogenase;oxygen species; RP-HPLC-FD,aphy coupled to fluorescenceRafael’’ salt works; TFA, tri-
les Campus, Highway A4, Kmx: þ34 957 218688.
and should be considered as
All rights reserved.
stress: primary anti-oxidant enzymes, such as catalase, detoxifyROS, while ancillary anti-oxidant enzymes, like 6-phosphogluc-onate dehydrogenase (6-PGDH), recycle NADPþ to its reducedform, which is consumed by glutathione reductase (GRase) (Sies,1986). In addition, glyoxalases (Glyox) I and II use GSH to de-toxify a-oxoaldehydes, derived from cellular oxidative processes(Regoli et al., 1997) and glutathione-S-transferase (GST) conju-gates GSH to electrophilic compounds. Some GSTs also detoxifyreactive products derived from lipid peroxidation (Jensson et al.,1986).
Donana National Park (DNP, SW Spain) is located north ofGuadalquivir Estuary and was declared by UNESCO a ‘‘World Re-serve of the Biosphere’’. It has been an area of concern since thetailings spill from Aznalcollar pyrite mine (April 1998), whichreleased large amounts of acid water and mud full of toxic metals(Fe, Zn, Pb, As, Cu, Cd, Ag, Ni, Mn, Cr) via Guadiamar stream, closeto DNP (Achterberg et al., 1999; Grimalt et al., 1999). Some bi-ological effects of this spill have been studied by our group inAlgerian mice (Mus spretus) and clams (Scrobicularia plana). Miceliving near Guadiamar stream showed greater anti-oxidant anddetoxifying activities than controls (Ruiz-Laguna et al., 2001), and

SRs
Guadiamar
stream
Guadalquivir
river
Sanlúcar de
Barrameda
San Rafael
salt works
Doñana
National Park
Doñana
National Park
Guadalquivir
river
Guadiamar
stream
Sanlúcar de
Barrameda
Brazo de
la Torre
BTSan Rafael
salt works
SRs
Across
Bonanza
harbour
Bh
Spain
Fig. 1. Map of Donana National Park (SW Spain) and sampling sites at GuadalquivirEstuary. S. plana specimens were collected at three sites: ‘‘Brazo de la Torre’’ (BT), ‘‘SanRafael’’ salt works (SRs), and across Bonanza harbour (Bh).
A. Romero-Ruiz et al. / Environmental Pollution 156 (2008) 1340–1347 1341
clams transplanted to the Guadalquivir Estuary displayed highmetal uptake and oxidative stress, as revealed by higher levels ofanti-oxidant, MT and GSH-related enzymes, MDA and over-oxidized glutathione status (Romero-Ruiz et al., 2003). The sed-entary lifestyle, filter-feeding habits and metal-concentrationability of bivalves make them excellent bioindicators of metalexposure/effect in marine systems (Cosson, 2000; Hamza-Chaffaiet al., 2000; Isani et al., 2000; Romero-Ruiz et al., 2003; Amiardet al., 2006).
Metallothioneins (MTs) are low Mr (w7 kDa) proteins with highCys content and no aromatic or His residues. In accordance withtheir conservative character and ubiquity, they play crucial bi-ological roles, including: (i) homeostasis of essential metals (Zn, Cu)(Kagi,1991); (ii) detoxification of trace metals (Ag, Cd, Hg, Ni, Pb, Co);and (iii) protection from oxidative injury (Kagi,1991; Viarengo et al.,2000; Kondoh et al., 2001; Haq et al., 2003; Erikson et al., 2004; Minet al., 2005). The inducible expression of MT genes by metals hasresulted in their use as metal-pollution biomarkers (Cosson, 2000;Hamza-Chaffai et al., 2000; Dabrio et al., 2002; Haq et al., 2003;Amiard et al., 2006). Since an integrated view of environmentalstatus requires the use of several biomarkers (Amiard et al., 2006),anti-oxidant defences and oxidative damage to biomolecules areused as sensitive biomarkers of metal pollutants, known to generateoxidative stress (Cajaraville et al., 2000; Rodrıguez-Ortega et al.,2002; Romero-Ruiz et al., 2003; Funes et al., 2006).
A method for total MT assay by RP-HPLC coupled to fluorescencedetection (RP-HPLC-FD) has been described in Chamaelea gallinaclams by heating digestive gland extracts at 95 �C for 10 min(Alhama et al., 2006). MTs thiols are labelled with the fluorogenicreagent monobromobimane (mBBr) in the presence of EDTA, DTTand SDS. Heat-denaturation is also used in the MT assay by differ-ential pulse polarography (DPP), the most popular MT-assay inbivalves (Thompson and Cosson, 1984; Cosson, 2000; Erk et al.,2002; Amiard et al., 2006). A comparative study of purificationprotocols has shown that the MT level diminishes after heating (Erket al., 2002). Although there is a good correlation among differentMT assays, concentrations diverge and are expressed in differentunits (Isani et al., 2000; Amiard et al., 2006). Thus, to compare MTcontents in bivalve tissues with published levels, isolation andquantification procedures should be considered (Isani et al., 2000;Dabrio et al., 2002). The highest MT levels and metal responses arefound in digestive glands compared to gills and other tissues(Cosson, 2000; Geffard et al., 2005; Amiard et al., 2006).
Here, in S. plana, we quantified the contents of MT and of bio-markers responsive to oxidative stress to assess the pollution statusof the Guadalquivir Estuary (SW Spain). MT was assessed by RP-HPLC-FD in heat-treated and unheated extracts, and comparedwith those obtained by DPP. The relationships between MT content,metal levels, anti-oxidant enzymatic defences, and lipid oxidativedamage were also studied.
2. Materials and methods
2.1. Materials, animals and sampling areas
Brilliant Blue G, D-lactoylglutathione, 5,50-dithio-bis-2-nitrobenzoic acid, DTT,reduced (GSH) and oxidized (GSSG) glutathione, leupeptin, methylglyoxal, rabbitliver MT-I, SDS (95%), PMSF, 6-P-gluconate, NADPþ, Tricine, and Mr markers werefrom Sigma (Spain). Monobromobimane (mBBr) was from Fluka (Spain). H2O2, 1-Cl-2,4-dinitrobenzene, sodium dodecyl sulphate (SDS, >99%), ethylenediaminetetra-acetic acid (EDTA), trifluoroacetic acid (TFA), thiobarbituric acid and Tris base werefrom Merck (Germany), and protein dye binding reagent from BioRad (Hercules, CA,USA). Other chemicals were reagent quality.
Clams were sampled on May, June and October 2003 at three sites on the DNPside of Guadalquivir Estuary (Fig. 1) and sent at 4 �C to the laboratory, where shellswere washed and opened. For MT assay, digestive glands were excised, pooled andfrozen at �80 �C, ground in a mortar with liquid N2 and kept at �80 �C until extractpreparation. For metal analysis, the whole soft tissues from 20 clams were frozen at�80 �C, ground in a mortar with liquid N2, and kept at �80 �C until used.
2.2. Metallothionein quantification by high-performance liquid chromatography
For extract preparation, ground digestive glands were disrupted in a T25Ultraturrax (Janke & Kunkel, Germany) in 0.1 M Tris–HCl buffer (pH 9.5) with 1 mMDTT, 50 mM PMSF and 6 mM leupeptin at a ratio of 3 mL/g, and spun 20 min at35 000 � g. Extracts were measured directly or after heating at 95 �C, 10 min, fol-lowed by centrifugation at 14 000 � g, 15 min. For reduction and denaturation, allsamples were treated for 20 min at 70 �C with DTT, EDTA and SDS. For derivatization,mBBr was added and the mixture incubated for 15 min at room temperature awayfrom light (Alhama et al., 2006). Final concentrations were DTT (12 mM), EDTA(2 mM), SDS (3%) and mBBr (12 mM) in 100 mL total volume.
Derivatized proteins in heated/unheated extracts (20 mL) were separated ina Supelcosil LC-18 column (0.46 � 25 cm, 5 mm particle, Supelco, PA, USA), as inC. gallina (Alhama et al., 2006). The HPLC system included a 7725i injection valve(Rheodyne, CA, USA), a 126 solvent module (Beckman, CA, USA) and a FP-2020 PlusFluorescence Detector (Jasco, MD, USA) coupled on-line. Fluorescence of mBBr-labelled molecules was followed with excitation at 382 nm and emission at 470 nm(gain ¼ 1). Peak areas were analyzed with the Beckman 32Karat Software (v. 7.0).
2.3. Metallothionein quantification by differential pulse polarography
Ground digestive glands were disrupted as indicated above in 0.1 M Tris–HClbuffer (pH 8.1) containing 1 mM DTT at a ratio of 3 mL/g, and spun for 1 h at50 000 � g. The supernatants were diluted 10-fold with 0.9% NaCl, heated at 95 �Cfor 5 min and spun for 15 min at 10 000 � g.
DPP assay for MTs was made in a Methrom 746 VA Trace analyzer, in the heat-stable fraction by anodic stripped voltammetry in the Static Mercury Drop Electrodemode (Olafson, 1987) using rabbit liver MT-I as a reference for the calibration curve.
2.4. Electrophoresis
Two types of samples were analyzed: mBBr-labelled cell-free extracts (directlyor heat-denatured) and samples after mBBr-labelling and purification by RP-HPLC-FD as described in Section 2.2. To this end, 40 mL of each type of extracts and 20 mL ofrabbit liver MT-I were separated in 0.2 ml fractions; those containing fluorescentmBBr-MT were pooled, dried and freed from residual TFA as described (Alhamaet al., 2006). The pellets were resuspended in 10 mL of 0.1 M Tris–HCl buffer (pH 9.5)containing 12 mM DTT, 2 mM EDTA and 3% SDS. For mBBr derivatization, Mr markerswere dissolved in 86.6 mL deionized water, heated for 20 min at 70 �C, and labelledas shown in Section 2.2.
Then, 10 mL of sample and 10 mL of 2� sample buffer (90 mM Tris–HCl (pH 8.45),8% SDS, 24% glycerol, 0.01% Brilliant Blue G and 10% b-mercaptoethanol) were mixed.Samples were denatured at 95 �C, 5 min, loaded, and separated in 13.5% Tricine SDS-PAGE gels (7 � 8 cm) at 30 mA/gel in a Mini-PROTEAN 3 cell, using as buffers 0.1 MTris, 0.1 M Tricine (pH 8.2) and 0.1% SDS (cathode), and 0.2 M Tris–HCl (pH 8.9)(anode) (Schagger and von Jagow, 1987). Gels were washed with 50% methanol,30 min; fluorescent protein bands were documented in a Gel Doc EQ system (Bio-Rad), and re-stained with Coomassie Blue R250.

100
80
60
40
20
0
100
80
60
40
20
00 2 4 6 8 10 12 0 1 2 3 4 5 6
SDS (%)
6 9 12 15 18 21mBBr (mM)DTT (mM)
Pe
rc
en
ta
ge
(%
)
Pe
rc
en
ta
ge
(%
)A B C
Fig. 2. Effect of DTT, SDS and mBBr on MT labelling. Purified rabbit liver MT-I (�) and clam digestive gland extracts (B) in 0.1 M Tris–HCl (pH 9.5), were heated at 70 �C, derivatizedwith mBBr and analyzed by RP-HPLC-FD. Values are shown as percentage of maximum.
A. Romero-Ruiz et al. / Environmental Pollution 156 (2008) 1340–13471342
2.5. Biochemical assays
Tissues were disrupted in a T25 Ultraturrax in 10 mM Tris HCl (pH 7.5) con-taining 1 mM EDTA, 1 mM PMSF and 1 mM GSH at a ratio of 4 mL/g. For MDA assay,300 mL of homogenate was set apart and the rest ultracentrifuged at 105 000 � g, 1 h(Beckman L8-80). The supernatant (2 mL) was dialyzed for 12 h (Sigma tubing,12 kDa cut) against 350 volumes of disruption buffer and stored at �80 �C untilanalysis.
Enzymatic activities were assayed spectrophotometrically as reported byRomero-Ruiz et al. (2003). The level of thiobarbituric acid-reactive substances wasassayed to assess MDA level, a known biomarker of lipid oxidative damage (Brownand Kelly, 1996). Protein concentration was determined by the dye-binding methodin unheated supernatants (Bradford, 1976).
16
20A
2.6. Metal analyses
Samples (0.1 g fresh weight) were mixed with 3.0 mL of concentrated H2NO3 inclosed Teflon vessels and digested with microwave assistance. Extracts were dilutedwith Milli-Q water to 50 mL in a volumetric flask and stored at 4 �C. Metals wereanalyzed in a 4500 inductively coupled plasma-mass spectroscopy instrument(Hewlett-Packard, Palo Alto, CA, USA) as reported (Romero-Ruiz et al., 2003). A blankand a reference material (NIST SRM 2977, ‘‘mussel tissue’’) were included in eachsample set to assess results quality.
4
6
12
RF
U
0
2.7. Statistical analysis
The results show the mean � SD of 3 independent derivatizations (RP-HPLC-FD)or 3 independent assays (DPP, metal analysis, and other biomarkers). The statisticalsignificance of differences in MT values in clams from SRs and Bh compared to BTclams was assayed by the Dunnett’s t-test and shown as not significant (ns),*p < 0.05 or **p < 0.01. Previously, normality of data and homogeneity of variancewere checked. Correlation coefficients were calculated by the Pearson parametriclineal correlation method. InStat software, version 2.01/93 (Graphpad, San Diego,CA, USA) was used in both cases.
20
4
6
12
16
RF
U
05
Time (min)
10 15 20 25 300
B
Fig. 3. MT separation of a digestive gland clam extract by RP-HPLC-FD using SDS of95% (A) or >99% (B) purity for derivatization. Arrows show MT peaks.
3. Results and discussion
3.1. Analytical optimization
Fig. 2 shows the effect of DTT, SDS and mBBr concentrations onMT labelling, from rabbit liver or S. plana. With rabbit MT-I, theoptimal DTT concentration was w2 mM (Fig. 2A) and labellingdecreased at higher levels, probably due to mBBr titration by excessdithiol, preventing mBBr binding to MTs thiol groups. Optimal la-belling of clam extracts required higher DTT, w12 mM, suggestingthe presence in the extracts of high oxidant levels. Optimal SDS andmBBr levels were 3% and 12 mM for both MTs (Figs. 2B/C), in-dicating similar reagent needs for both proteins. Using 12 mM DTT,3% SDS and 12 mM mBBr, an excellent linearity (r ¼ 0.997) wasobtained when the area of HPLC-eluted mBBr-MT was plotted vstotal protein content in clam extracts, from 8 to 50 mg. Linearity of
clam extract (protein (mg) ¼ 7.53 � 10�6 � peak area �1.64) wascalculated using 7 points for the regression.
The importance of SDS purity in MT derivatization with mBBrshould be noted. Fig. 3 compares the elution profiles of mBBr-MT-Iusing 3% SDS of different purity, w95% (A) and >99% (B). Accordingto the specifications, the first contains lauryl (66%), myristyl (28%)and cetyl sulphate (6%). With 95% SDS, mBBr-MT-I eluted in 3peaks, whose retention times and relative peak areas changed withSDS content in the derivatization mixture (results not shown), andcannot be taken as being different isoforms. In contrast, with >99%SDS, mBBr-MT-I eluted in a sharp well-defined peak, whose re-tention time and peak area were unaffected by detergent content.

1 2 3 4 5 6 7 8 1 2 3 4 5 6 7 8Mr(kDa)
66
4536
2924
14.26.5
20
A B
Fig. 4. Effects of RP-HPLC purification and heating in SDS-PAGE analysis. Fluorescence of mBBr-labelled (A) and absorbance of Coomassie-stained proteins (B) were assessed in thesame gel. Lane 1, 30 mg of mBBr-labelled standards. Lanes 2 and 7, 1.3 mg of rabbit mBBr-MT-I. Lane 3, 30 mg of mBBr-labelled protein (heated extract). Lane 4, 7 mg of mBBr-labelledMT purified from heated extract. Lane 5, 30 mg of mBBr-labelled protein (unheated extract). Lane 6, 12 mg of mBBr-labelled MT purified from unheated extract. Lane 8, 30 mg ofunlabelled standards.
A. Romero-Ruiz et al. / Environmental Pollution 156 (2008) 1340–1347 1343
Fig. 4 shows the electrophoretic analysis of mBBr-labelled cell-free extracts (directly or heat-denatured), without or with RP-HPLC-FD purification, using rabbit mBBr-MT-I as reference. Gelswere monitored by fluorescence (mBBr-labelled protein thiols, A)and by Coomassie Blue staining (total proteins, B). In heated ex-tracts after RP-HPLC (lanes 4) a sharp and highly fluorescent bandwas visible, with Mr slightly higher than rabbit liver MT-I, whilea more intense band appeared in unheated purified extracts (lanes6), suggesting that part of MT was removed by heating. A sharp MTband was also seen in heated (lanes 3) and unheated extracts (lanes5) not purified by RP-HPLC. The smear observed in heated and,specially, unheated extracts correspond to many additional pro-teins not well resolved. The high purification effect of RP-HPLC isshown by comparing cell-free extracts and purified fractions, whilethe loss of total protein and MT promoted by heat is shown bycomparing heated and unheated samples.
3.2. Field validation
To validate the mBBr-MT assay, S. plana clams from 3 Gua-dalquivir Estuary sites were compared to assess their status mea-suring MT levels by RP-HPLC-FD in digestive gland extracts heated
BT SRs Bh BTSamp
0
40
80
120
160
200
MT
co
nten
t (m
g/g
p
ro
tein
)
May J
*
** **
**
*
**
Fig. 5. MT content of clams from different estuary sites. Analysis was carried out by DPP (wgland extracts. S. plana specimens were collected at three sites: ‘‘Brazo de la Torre’’ (BT), ‘‘Santhe differences is compared with BT, used as reference.
at 95 �C for 10 min or unheated (Fig. 5). While both MT patternswere similar, its content was much higher in unheated extracts,suggesting that heating removes part of MT initially present in theextracts. Since lower MT levels were always obtained at BT, this sitewas used as a reference for statistical analysis. Significant differ-ences were obtained in all samplings, the highest MT level beingfound at Bh in May, at SRs in June, while a similar MT contentexisted at SRs and Bh in October, both of them much higher than inheated extracts. MT levels were also measured by DPP in May andJune samples. MT levels detected by this well-established method(Olafson, 1987) were much lower than those obtained by RP-HPLC-FD, even in heated extracts, since MTs were also determined by DPPin the heat-stable fraction after 95 �C treatment for 5 min. DPPrevealed similar MT levels at BT and Bh in May and June, bothsignificantly lower than in SRs, in contrast to RP-HPLC-FD results.
The monitoring (Fig. 5) and electrophoretic analysis (Fig. 4) re-sults showed that part of MT disappeared during heat-treatment. Ithas been reported that MT co-precipitates during heating, leadingto its underestimation (Geret et al., 1998; Erk et al., 2002). The RP-HPLC-FD assay allows a direct assessment of total MT in extractswithout heat treatment, since mBBr-labelled MT is well separatedby RP-HPLC from other Cys-containing proteins (Fig. 4). This is
SRs Bh BT SRs Bhling site
une October**
**
**
**
**
**
*
hite bars) and RP-HPLC-FD in heated (grey bars) and unheated (black bars) digestiveRafael’’ salt works (SRs), and across Bonanza harbour (Bh). The statistical significance of

0
120
0
40
80
0.0
1.0**
** ** ****
** **
**
*
0.0
1.0
2.0
**** ** **
0
10
0
80
0
40
** ** ****
****
** ***
*
Sampling date
MDA (mmol/mg)
Catalase (U/mg)
Glyox I (mU/mg)
GST (U/mg)GRase (mmol/mg)
Glyox II (mU/mg)
6PGDH (mU/mg)
May June October May June October
Fig. 6. Biomarkers analyzed in the digestive gland of S. plana clams from ‘‘Brazo de la Torre’’ (BT, white bars), ‘‘San Rafael’’ salt works (SRs, grey bars) and Bonanza harbour (Bh, blackbars). The statistical significance of the differences is compared with BT, used as a reference.
A. Romero-Ruiz et al. / Environmental Pollution 156 (2008) 1340–13471344
a further advantage that simplifies the assay, and is quite importantfor using MT as biomarker in monitoring studies. Prior to RP-HPLC-FD, only a derivatization with mBBr is required after EDTA, SDS andDTT treatment at 70 �C.
Table 1Correlation coefficients of MT levels, determined by RP-HPLC with fluorescent detectionbiomarkers in Scrobicularia plana
MT level Catalase 6PGDH GRase
RP-HPLC (NH) 0.705* �0.826** 0.367RP-HPLC (H) 0.495 �0.620 �0.200DPP 0.238 �0.467 �0.220
Coefficients were calculated by the parametric lineal correlation (Pearson) method. Stati
The MT levels detected by RP-HPLC-FD in S. plana, 62–117 mgg�1protein (0.9–1.6 mg g�1ww) in heated and 89–199 mgg�1protein (1.3–3.9 mg g�1ww) in unheated extracts, are similar tothose previously reported by DPP in the digestive gland of other
in non-heated (NH) and heated (H) clam extracts or by DPP, and other biochemical
Glyox I Glyox II GST MDA
0.377 �0.798** 0.788* 0.480�0.037 �0.436 0.470 0.647
0.176 �0.238 0.389 0.120
stical significance: *p < 0.05; **p < 0.01.
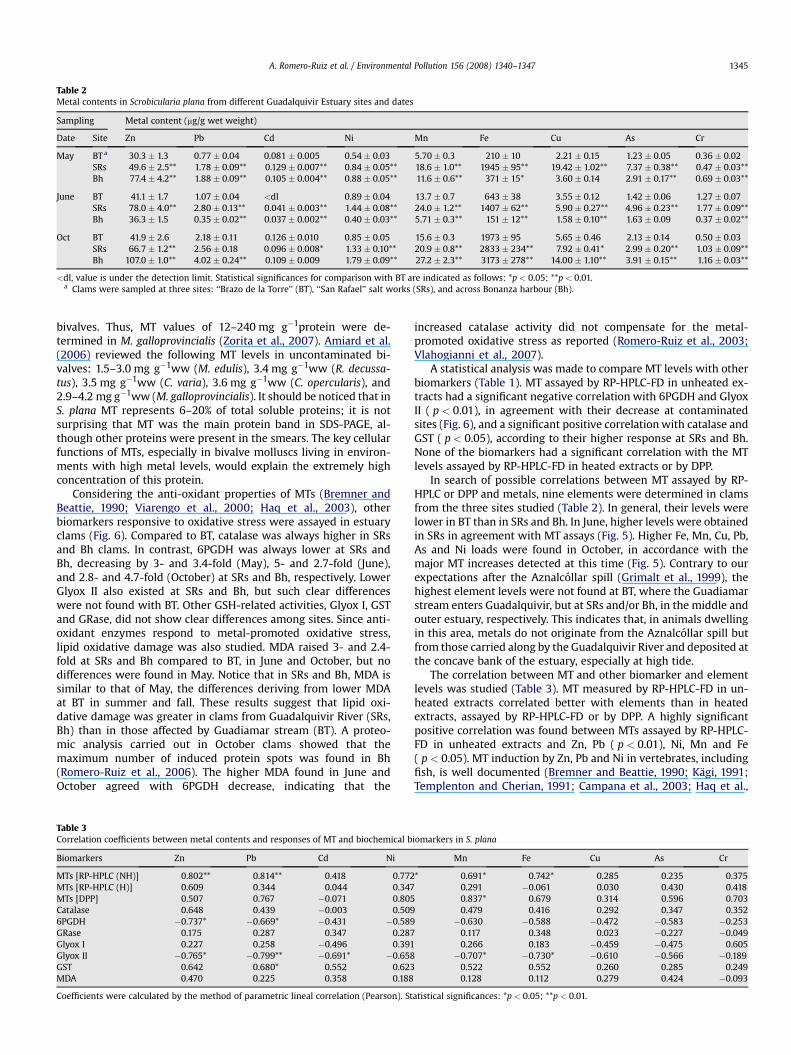
Table 2Metal contents in Scrobicularia plana from different Guadalquivir Estuary sites and dates
Sampling Metal content (mg/g wet weight)
Date Site Zn Pb Cd Ni Mn Fe Cu As Cr
May BT a 30.3 � 1.3 0.77� 0.04 0.081 � 0.005 0.54 � 0.03 5.70� 0.3 210 � 10 2.21 � 0.15 1.23 � 0.05 0.36 � 0.02SRs 49.6 � 2.5** 1.78 � 0.09** 0.129 � 0.007** 0.84 � 0.05** 18.6 � 1.0** 1945 � 95** 19.42 � 1.02** 7.37 � 0.38** 0.47 � 0.03**Bh 77.4 � 4.2** 1.88 � 0.09** 0.105 � 0.004** 0.88 � 0.05** 11.6 � 0.6** 371 � 15* 3.60 � 0.14 2.91 � 0.17** 0.69 � 0.03**
June BT 41.1 � 1.7 1.07� 0.04 <dl 0.89 � 0.04 13.7 � 0.7 643 � 38 3.55 � 0.12 1.42 � 0.06 1.27� 0.07SRs 78.0 � 4.0** 2.80 � 0.13** 0.041 � 0.003** 1.44 � 0.08** 24.0 � 1.2** 1407 � 62** 5.90 � 0.27** 4.96 � 0.23** 1.77 � 0.09**Bh 36.3 � 1.5 0.35� 0.02** 0.037 � 0.002** 0.40 � 0.03** 5.71 � 0.3** 151 � 12** 1.58 � 0.10** 1.63 � 0.09 0.37 � 0.02**
Oct BT 41.9 � 2.6 2.18 � 0.11 0.126 � 0.010 0.85 � 0.05 15.6 � 0.3 1973 � 95 5.65 � 0.46 2.13 � 0.14 0.50 � 0.03SRs 66.7 � 1.2** 2.56 � 0.18 0.096 � 0.008* 1.33� 0.10** 20.9 � 0.8** 2833 � 234** 7.92 � 0.41* 2.99 � 0.20** 1.03 � 0.09**Bh 107.0 � 1.0** 4.02 � 0.24** 0.109 � 0.009 1.79 � 0.09** 27.2 � 2.3** 3173 � 278** 14.00 � 1.10** 3.91 � 0.15** 1.16 � 0.03**
<dl, value is under the detection limit. Statistical significances for comparison with BT are indicated as follows: *p < 0.05; **p < 0.01.a Clams were sampled at three sites: ‘‘Brazo de la Torre’’ (BT), ‘‘San Rafael’’ salt works (SRs), and across Bonanza harbour (Bh).
A. Romero-Ruiz et al. / Environmental Pollution 156 (2008) 1340–1347 1345
bivalves. Thus, MT values of 12–240 mg g�1protein were de-termined in M. galloprovincialis (Zorita et al., 2007). Amiard et al.(2006) reviewed the following MT levels in uncontaminated bi-valves: 1.5–3.0 mg g�1ww (M. edulis), 3.4 mg g�1ww (R. decussa-tus), 3.5 mg g�1ww (C. varia), 3.6 mg g�1ww (C. opercularis), and2.9–4.2 mg g�1ww (M. galloprovincialis). It should be noticed that inS. plana MT represents 6–20% of total soluble proteins; it is notsurprising that MT was the main protein band in SDS-PAGE, al-though other proteins were present in the smears. The key cellularfunctions of MTs, especially in bivalve molluscs living in environ-ments with high metal levels, would explain the extremely highconcentration of this protein.
Considering the anti-oxidant properties of MTs (Bremner andBeattie, 1990; Viarengo et al., 2000; Haq et al., 2003), otherbiomarkers responsive to oxidative stress were assayed in estuaryclams (Fig. 6). Compared to BT, catalase was always higher in SRsand Bh clams. In contrast, 6PGDH was always lower at SRs andBh, decreasing by 3- and 3.4-fold (May), 5- and 2.7-fold (June),and 2.8- and 4.7-fold (October) at SRs and Bh, respectively. LowerGlyox II also existed at SRs and Bh, but such clear differenceswere not found with BT. Other GSH-related activities, Glyox I, GSTand GRase, did not show clear differences among sites. Since anti-oxidant enzymes respond to metal-promoted oxidative stress,lipid oxidative damage was also studied. MDA raised 3- and 2.4-fold at SRs and Bh compared to BT, in June and October, but nodifferences were found in May. Notice that in SRs and Bh, MDA issimilar to that of May, the differences deriving from lower MDAat BT in summer and fall. These results suggest that lipid oxi-dative damage was greater in clams from Guadalquivir River (SRs,Bh) than in those affected by Guadiamar stream (BT). A proteo-mic analysis carried out in October clams showed that themaximum number of induced protein spots was found in Bh(Romero-Ruiz et al., 2006). The higher MDA found in June andOctober agreed with 6PGDH decrease, indicating that the
Table 3Correlation coefficients between metal contents and responses of MT and biochemical b
Biomarkers Zn Pb Cd Ni
MTs [RP-HPLC (NH)] 0.802** 0.814** 0.418 0.772MTs [RP-HPLC (H)] 0.609 0.344 0.044 0.347MTs [DPP] 0.507 0.767 �0.071 0.805Catalase 0.648 0.439 �0.003 0.5096PGDH �0.737* �0.669* �0.431 �0.589GRase 0.175 0.287 0.347 0.287Glyox I 0.227 0.258 �0.496 0.391Glyox II �0.765* �0.799** �0.691* �0.658GST 0.642 0.680* 0.552 0.623MDA 0.470 0.225 0.358 0.188
Coefficients were calculated by the method of parametric lineal correlation (Pearson). St
increased catalase activity did not compensate for the metal-promoted oxidative stress as reported (Romero-Ruiz et al., 2003;Vlahogianni et al., 2007).
A statistical analysis was made to compare MT levels with otherbiomarkers (Table 1). MT assayed by RP-HPLC-FD in unheated ex-tracts had a significant negative correlation with 6PGDH and GlyoxII ( p < 0.01), in agreement with their decrease at contaminatedsites (Fig. 6), and a significant positive correlation with catalase andGST ( p < 0.05), according to their higher response at SRs and Bh.None of the biomarkers had a significant correlation with the MTlevels assayed by RP-HPLC-FD in heated extracts or by DPP.
In search of possible correlations between MT assayed by RP-HPLC or DPP and metals, nine elements were determined in clamsfrom the three sites studied (Table 2). In general, their levels werelower in BT than in SRs and Bh. In June, higher levels were obtainedin SRs in agreement with MT assays (Fig. 5). Higher Fe, Mn, Cu, Pb,As and Ni loads were found in October, in accordance with themajor MT increases detected at this time (Fig. 5). Contrary to ourexpectations after the Aznalcollar spill (Grimalt et al., 1999), thehighest element levels were not found at BT, where the Guadiamarstream enters Guadalquivir, but at SRs and/or Bh, in the middle andouter estuary, respectively. This indicates that, in animals dwellingin this area, metals do not originate from the Aznalcollar spill butfrom those carried along by the Guadalquivir River and deposited atthe concave bank of the estuary, especially at high tide.
The correlation between MT and other biomarker and elementlevels was studied (Table 3). MT measured by RP-HPLC-FD in un-heated extracts correlated better with elements than in heatedextracts, assayed by RP-HPLC-FD or by DPP. A highly significantpositive correlation was found between MTs assayed by RP-HPLC-FD in unheated extracts and Zn, Pb ( p < 0.01), Ni, Mn and Fe( p < 0.05). MT induction by Zn, Pb and Ni in vertebrates, includingfish, is well documented (Bremner and Beattie, 1990; Kagi, 1991;Templenton and Cherian, 1991; Campana et al., 2003; Haq et al.,
iomarkers in S. plana
Mn Fe Cu As Cr
* 0.691* 0.742* 0.285 0.235 0.3750.291 �0.061 0.030 0.430 0.4180.837* 0.679 0.314 0.596 0.7030.479 0.416 0.292 0.347 0.352�0.630 �0.588 �0.472 �0.583 �0.253
0.117 0.348 0.023 �0.227 �0.0490.266 0.183 �0.459 �0.475 0.605�0.707* �0.730* �0.610 �0.566 �0.189
0.522 0.552 0.260 0.285 0.2490.128 0.112 0.279 0.424 �0.093
atistical significances: *p < 0.05; **p < 0.01.

A. Romero-Ruiz et al. / Environmental Pollution 156 (2008) 1340–13471346
2003; Cheung et al., 2004; Chan and Chan, 2008). In contrast, MTinduction by Zn, Pb and Ni in molluscs presents many contradic-tions and inconsistencies since these metals are shown to be effi-cient MT inducers in some papers, while no induction has beenreported in others (Cosson, 2000; Irato et al., 2003; Amiard et al.,2006; Bourdineaud et al., 2006; Marie et al., 2006; Geffard et al.,2007; Martın-Dıaz et al., 2007). Possible explanations for this var-iability have been discussed extensively by Amiard et al. (2006).Correlation with Fe and Mn can be explained by the anti-oxidantrole of MTs (Bremner and Beattie, 1990; Cajaraville et al., 2000;Viarengo et al., 2000; Kondoh et al., 2001). Induced MT levelswould protect from the oxidative stress prompted by Mn (Eriksonet al., 2004) or by Fe that generates hydroxyl radical by Fentonreaction (Sies, 1986; Min et al., 2005). Although Cd is an excellentMT inducer (Bremner and Beattie, 1990; Cajaraville et al., 2000;Isani et al., 2000; Viarengo et al., 2000; Amiard et al., 2006), MT wasnot significantly correlated with Cd in S. plana, probably due to thelow Cd levels detected.
A significant negative correlation was obtained between GlyoxII and Pb ( p < 0.01), Zn, Cd, Mn and Fe ( p < 0.05), suggestingthat the decrease in this GSH-related enzyme is a good biomarkerof metal contamination. The negative correlation of 6PGDH withZn and Pb ( p < 0.05) agrees with Pb inhibition of G6PDH and6PGDH described in haemolysed erythrocytes (Lachant et al.,1984). The positive correlation of GST and Pb ( p < 0.05) is inaccordance with significant increase of this activity found in theearthworm Lampito mauritii exposed to low levels of Pb (Maityet al., 2008).
In addition to higher metal levels, clams living at the end ofthe Guadalquivir Estuary would be exposed to higher salinity inthe following order: Bh > SRs > BT. Conflicting salinity effectshave been reported in MT and several other biomarkers. En-hanced MT synthesis has been claimed in organisms living atlow salinity because of higher free metal concentrations, due tolower complexation by chloride (Martın-Dıaz et al., 2005; Mon-serrat et al., 2007). In contrast, no salinity effect on MT levelswas found in C. gigas larvae (Damiens et al., 2006), and oppositeeffects existed in M. balthica and M. edulis at different ArticEstuaries (Amiard-Triquet et al., 1998). Even if low salinity en-hances MT levels, the differences found in S. plana between Bhand BT sites would be underestimated; so, we think that MTlevels at the Guadalquivir Estuary mainly reflect changes inmetal content. Diverging salinity effects have been found inother biomarkers. Salinity promotes high P-glycoprotein, catalase,and glutathione reductase levels (Luedeking and Koehler, 2004;Khessiba et al., 2005; Lima et al., 2007), but low glucose-6-phosphate dehydrogenase, 6-phosphogluconate dehydrogenase,lactate dehydrogenase, glutathione peroxidase and acetylcholin-esterase activities (Carrion et al., 1978; Menezes et al., 2006;Lima et al., 2007; Rank et al., 2007). The higher catalase andMDA levels of animals living at the end of the GuadalquivirEstuary agree with the higher content of several metals andsalinity found at these sites. The decrease of 6-phosphogluconatedehydrogenase and glyoxalase II activities at these sites might beassigned to the negative effect of salinity on the two pentosephosphate pathway enzymes (Carrion et al., 1978) and anotherwell-established anti-oxidative enzyme, glutathione peroxidase(Lima et al., 2007).
Our results have clearly established the important MT lossesoccurring in heated extracts, although MTs are usually consideredas being heat-resistant proteins (Kagi, 1991; Cosson, 2000; Hamza-Chaffai et al., 2000). In contrast to DPP and other methods based onthiol group detection, that rely on heating pre-purification steps, inthe RP-HPLC-FD method developed by us (Alhama et al., 2006), MTsare separated from other Cys-containing proteins by HPLC asshown by electrophoretic fluorimetric analysis, obtaining a unique
intense fluorescent band after chromatography of non-heated clamextracts.
4. Conclusion
In conclusion, quantification of total MTs by RP-HPLC-FD in non-heated bivalve extracts allows assessment of metal contaminationwith higher sensitivity and specificity than other well-establishedassays (Thompson and Cosson, 1984; Cosson, 2000; Erk et al., 2002;Amiard et al., 2006), since MT content is significantly reduced ifsamples are heated. We propose this improved and simplifiedprocedure as an analytical tool for using MT as a biomarker inmonitoring programmes, since it has been validated both with re-spect to metal contents and to the response of well-establishedbiomarkers. Finally, metal and biomarker levels indicate that themetals detected at the left bank of the Guadalquivir Estuary do notoriginate from the Aznalcollar spill, but from those carried along bythe river and deposited at its concave bank.
Acknowledgements
This work was funded by grant CMT2006-08960-C02 from theSpanish Ministry of Education and Science, a grant from the Donana2005 Program, and grants CVI-151 and FQM-141 from the Anda-lusian Research Plan. A.R.-R. had a predoctoral fellowship from theSpanish Ministry of Education and Science.
References
Achterberg, E.P., Braungardt, C., Morley, N.H., Elbaz-Poulichet, F., Leblanc, M., 1999.Impact of Los Frailes mine spill on riverine, estuarine and coastal waters insouthern Spain. Water Res. 33, 3387–3394.
Alhama, J., Romero-Ruiz, A., Lopez-Barea, J., 2006. Metallothionein quantification inclams by reversed-phase high-performance liquid chromatography coupled tofluorescence detection after monobromobimane derivatization. J. Chromatogr.A 1107, 52–58.
Amiard-Triquet, C., Rainglet, F., Larroux, C., Regoli, F., Hummel, H., 1998. Metal-lothioneins in Artic bivalves. Ecotoxicol. Environ. Safety 41, 96–102.
Amiard, J.-C., Amiard-Triquet, C., Barka, S., Pellerin, J., Rainbow, P.S., 2006. Metal-lothioneins in aquatic invertebrates: their role in metal detoxification and theiruse as biomarkers. Aquat. Toxicol. 76, 160–202.
Bourdineaud, J.-P., Baudrimont, M., Gonzalez, P., Moreau, J.-L., 2006. Challenging themodel for induction of metallothionein gene expression. Biochimie 88, 1787–1792.
Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of micro-gram quantities of protein utilizing the principle of protein-dye binding. Anal.Biochem. 72, 248–254.
Bremner, I., Beattie, J.H., 1990. Metallothionein and the trace minerals. Annu. Rev.Nutr. 10, 63–83.
Brown, R.K., Kelly, F.J., 1996. Peroxides and other products. In: Punchard, N.A.,Kelly, F.J. (Eds.), Free Radicals: A Practical Approach. IRL Press, Oxford, UK, pp.119–131.
Cajaraville, M.P., Bebianno, M.J., Blasco, J., Porte, C., Sarasquete, C., Viarengo, A.,2000. The use of biomarkers to assess the impact of pollution in coastal envi-ronments of the Iberian Peninsula: a practical approach. Sci. Total Environ. 247,295–311.
Campana, O., Sarasquete, C., Blasco, J., 2003. Effect of lead on ALA-D activity, met-allothionein levels, and lipid peroxidation in blood, kidney, and liver of thetoadfish Halobatrachus didactylus. Ecotoxicol. Environ. Safety 55, 116–125.
Carrion, A., Silva, M., Cienfuegos, E., Ruiz-Amil, M., 1978. Influence of temperatureand salinity on glucose-6-phosphate dehydrogenase and 6-phosphogluconatedehydrogenase in mussel (Mytilus edulis L.). Rev. Esp. Fisiol. 34 (3), 241–245.
Chan, W.W.L., Chan, K.M., 2008. Cloning and characterization of a tilapia (Oreo-chromis aureus) metallothionein gene promoter in Hepa-T1 cells following theadministration of various heavy metal ions. Aquat. Toxicol. 86, 59–75.
Cheung, A.P.L., Lam, T.H.-J., Chan, K.M., 2004. Regulation of Tilapia metallothioneingene expression by heavy metal ions. Mar. Environ. Res. 58, 389–394.
Cosson, R.P., 2000. Bivalve metallothionein as a biomarker of aquatic ecosystempollution by trace metals: limits and perspectives. Cell. Mol. Biol. 46, 295–309.
Dabrio, M., Rodrıguez, A.R., Bordin, G., Bebianno, M.J., De Ley, M., Sestakova, I.,Vasak, M., Nordberg, M., 2002. Recent developments in quantification methodsfor metallothionein. J. Inorg. Biochem. 88, 123–134.
Damiens, G., Mouneyrac, C., Quiniou, F., His, E., Gnassia-Barelli, M., Romeo, M., 2006.Metal bioaccumulation and metallothionein concentrations in larvae of Cras-sostrea gigas. Environ. Pollut. 140, 492–499.
Erikson, K.M., Dobson, A.W., Dorman, D.C., Aschner, M., 2004. Manganese exposureand induced oxidative stress in the rat brain. Sci. Total Environ. 334-335, 409–416.

A. Romero-Ruiz et al. / Environmental Pollution 156 (2008) 1340–1347 1347
Erk, M., Ivankovic, D., Raspor, B., Pavicic, J., 2002. Evaluation of different purificationprocedures for the electrochemical quantification of mussel metallothioneins.Talanta 57, 1211–1218.
Funes, V., Alhama, J., Navas, J.I., Lopez-Barea, J., Peinado, J., 2006. Ecotoxicologicaleffects of metal pollution in two mollusc species from the Spanish South At-lantic littoral. Environ. Pollut. 139, 214–223.
Geffard, A., Amiard-Triquet, C., Amiard, J.C., 2005. Do seasonal changes affectmetallothionein induction by metals in mussels, Mytilus edulis? Ecotoxicol.Environ. Safety 61, 209–220.
Geffard, A., Geffard, O., Amiard, J.C., His, E., Amiard-Triquet, C., 2007. Bio-accumulation of metals in sediment elutriates and their effects on growth,condition index, and metallothionein contents in oyster larvae. Arch. Environ.Contam. Toxicol. 53, 57–65.
Geret, F., Rainglet, F., Cosson, R.P., 1998. Comparison between isolation protocolscommonly used for the purification of mollusc metallothioneins. Mar. Environ.Res. 46, 545–550.
Grimalt, J.O., Ferrer, M., Macpherson, E., 1999. The mine tailing accident in Aznal-collar. Sci. Total Environ. 242, 3–11.
Hamza-Chaffai, A., Amiard, J.C., Pellerin, J., Joux, L., Berthet, B., 2000. The potentialuse of metallothionein in the clam Ruditapes decussatus as a biomarker of in situmetal exposure. Comp. Biochem. Physiol. C 127, 185–197.
Haq, F., Mahoney, M., Koropatnick, J., 2003. Signaling events for metallothioneininduction. Mutat. Res. 533, 211–226.
Irato, P., Santovito, G., Cassini, A., Piccinni, E., Albergoni, V., 2003. Metal accumu-lation and binding protein induction in Mytilus galloprovincialis, Scapharca in-aequivalvis, and Tapes philippinarum from the lagoon of Venice. Arch. Environ.Contam. Toxicol. 44, 476–484.
Isani, G., Andreani, G., Kindt, M., Carpene, E., 2000. Metallothioneins (MTs) in ma-rine molluscs. Cell. Mol. Biol. 46, 311–330.
Jensson, H., Guthenberg, C., Alin, P., Mannervik, B., 1986. Rat glutathione transferase8-8, an enzyme efficiently detoxifying 4-hydroxyalk-2-enals. FEBS Lett. 203,207–209.
Kagi, J.H.R., 1991. Overview of metallothionein. Methods Enzymol. 205, 613–626.Khessiba, A., Romeo, M., Aıssa, P., 2005. Effects of some environmental parameters
on catalase activity in the mussel (Mytilus galloprovincialis) exposed to lindane.Environ. Pollut. 133, 275–281.
Kondoh, M., Inoue, Y., Atagi, S., Futakawa, N., Higashimoto, M., Sato, M., 2001.Specific induction of metallothionein synthesis by mitochondrial oxidativestress. Life Sci. 69, 2137–2146.
Lachant, N.E., Tomoda, A., Tanaka, K.R.,1984. Inhibition of pentose phosphate shunt bylead: potential mechanism for hemolysis in lead poisoning. Blood 63, 518–524.
Lima, I., Moreira, S.M., Rendon-Von Osten, J., Soares, A.M.V.M., Guilhermino, L.,2007. Biochemical responses of the marine mussel Mytilus galloprovincialis topetrochemical environmental contamination along the North-western coast ofPortugal. Chemosphere 66, 1230–1242.
Luedeking, A., Koehler, A., 2004. Regulation of expression of multixenobiotic re-sistance (MXR) genes by environmental factors in the blue mussel Mytilusedulis. Aquat. Toxicol. 69, 1–10.
Maity, S., Roy, S., Chaudhury, S., Bhattacharya, S., 2008. Antioxidant responses of theearthworm Lampito mauritii exposed to Pb and Zn contaminated soil. Environ.Pollut. 151, 1–7.
Marie, V., Gonzalez, P., Baudrimont, M., Gourdineaud, J.-P., Boudou, A., 2006. Met-allothionein response to cadmium and zinc exposures compared in twofreshwater bivalves, Dreissena polymorpha and Corbicula fluminea. Biometals 19,399–407.
Martın-Dıaz, M.L., Blasco, J., Gonzalez de Canales, M., Sales, D., DelValls, A., 2005.Bioaccumulation and toxicity of dissolved heavy metals from the GuadalquivirEstuary after the Aznalcollar mining spill using Ruditapes philippinarum. Arch.Environ. Contam. Toxicol. 48, 233–241.
Martın-Dıaz, M.L., Blasco, J., Sales, D., DelValls, T.A., 2007. Biomarkers study for sed-iment quality assessment in Spanish ports using the crab Carcinus maenas and theclam Ruditapes philippinarum. Arch. Environ. Contam. Toxicol. 53, 66–76.
Menezes, S., Soares, A.M.V.M., Guilhermino, L., Peck, M.R., 2006. Biomarker re-sponses of the estuarine brown shrimp Crangon crangon L. to non-toxicstressors: temperature, salinity and handling stress effects. J. Exp. Mar. Biol.Ecol. 335, 114–122.
Min, K.S., Morishita, F., Tetsuchikawahara, N., Onosaka, S., 2005. Induction of hepaticand renal metallothionein synthesis by ferric nitrilotriacetate in mice: the roleof MT as an antioxidant. Toxicol. Appl. Pharmacol. 204, 9–17.
Monserrat, J.M., Martınez, P.E., Geracitano, L.A., Lund Amado, L., Martinez GasparMartins, C., Lopes Leaes Pinho, G., Soares Chaves, I., Ferreira-Cravo, M., Ventura-Lima, J., Bianchini, A., 2007. Pollution biomarkers in estuarine animals: criticalreview and new perspectives. Comp. Biochem. Physiol. C 146, 221–234.
Olafson, R.W., 1987. Electrochemical quantification and characterization of metal-lothioneins. Experientia Suppl. 52, 329–333.
Rank, J., Lehtonen, K.K., Strand, J., Laursen, M., 2007. DNA damage, acetylcholines-terase activity and lysosomal stability in native and transplanted mussels(Mytilus edulis) in areas close to coastal chemical dumping sites in Denmark.Aquat. Toxicol. 84, 50–61.
Regoli, F., Principato, G.B., Bertoli, E., Nigro, M., Orlando, E., 1997. Biochemicalcharacterization of the antioxidant system in the scallop Adamussium colbecki,a sentinel organism for monitoring the Antarctic environment. Polar Biol. 17,251–258.
Rodrıguez-Ortega, M.J., Alhama, J., Funes, V., Romero-Ruiz, A., Rodrıguez-Ariza, A.,Lopez-Barea, J., 2002. Biochemical biomarkers of pollution in the clamChamaelea gallina from south-Spanish littoral. Environ. Toxicol. Chem. 21, 542–549.
Romero-Ruiz, A., Amezcua, O., Rodrıguez-Ortega, M.J., Munoz, J.L., Alhama, J., Ro-drıguez-Ariza, A., Gomez-Ariza, J.L., Lopez-Barea, J., 2003. Oxidative stressbiomarkers in bivalves transplanted to the Guadalquivir estuary after Aznal-collar spill. Environ. Toxicol. Chem. 22, 92–100.
Romero-Ruiz, A., Carrascal, M., Alhama, J., Gomez-Ariza, J.L., Abian, J., Lopez-Barea, J., 2006. Utility of proteomics to assess pollutant response of clams from theDonana bank of Guadalquivir Estuary (SW Spain). Proteomics 6, S245–S255.
Ruiz-Laguna, J., Garcıa-Alfonso, C., Peinado, J., Moreno, S., Ieradi, L.A., Cristaldi, M.,Lopez-Barea, J., 2001. Biochemical biomarkers of pollution in Algerian mouse(Mus spretus) to assess the effects of the Aznalcollar disaster on Donana Park(Spain). Biomarkers 6, 146–160.
Schagger, H., von Jagow, G., 1987. Tricine-sodium dodecyl sulfate-polyacrylamidegel electrophoresis for the separation of proteins in the range from 1 to100 kDa. Anal. Biochem. 166, 368–379.
Sies, H., 1986. Biochemistry of oxidative stress. Angew. Chem. Int. Ed. Engl. 25,1058–1071.
Templenton, D.M., Cherian, M.G., 1991. Toxicological significance of metallothionein.Methods Enzymol. 205, 11–24.
Thompson, J.A.J., Cosson, R.P., 1984. An improved electrochemical method for thequantification of metallothioneins in marine organisms. Mar. Environ. Res. 11,137–152.
Vlahogianni, T., Dassenakis, M., Scoullos, M.J., Valavanidis, A., 2007. Integrated use ofbiomarkers (superoxide dismutase, catalase and lipid peroxidation) in musselsMytilus galloprovincialis for assessing heavy metals’ pollution in coastal areasfrom the Saronikos Gulf of Greece. Mar. Pollut. Bull. 54, 1361–1371.
Viarengo, A., Burlando, B., Ceratto, N., Panfoli, I., 2000. Antioxidant role of metal-lothioneins: a comparative overview. Cell. Mol. Biol. 46, 407–417.
Zorita, I., Apraiz, I., Ortiz-Zarragoitia, M., Orbea, A., Cancio, I., Soto, M., Marigomez, I.,Cajaraville, M.P., 2007. Assessment of biological effects of environmental pol-lution along the NW Mediterranean Sea using mussels as sentinel organisms.Environ. Pollut. 148, 236–250.