
Neurotrophin p75 Receptor (p75NTR) Promotes Endothelial Cell Apoptosis and Inhibits Angiogenesis:...
26
ISSN: 1524-4571 Copyright © 2008 American Heart Association. All rights reserved. Print ISSN: 0009-7330. Online TX 72514 Circulation Research is published by the American Heart Association. 7272 Greenville Avenue, Dallas, DOI: 10.1161/CIRCRESAHA.108.177386 published online Jun 19, 2008; Circ. Res. Madeddu and Costanza Emanueli Aurelie S. Leroyer, Chantal M. Boulanger, Gaia Spinetti, Sung Ok Yoon, Paolo Oikawa, Graciela B. Sala-Newby, Marco Meloni, Brunella Cristofaro, Gallia Graiani, Andrea Caporali, Elisabetta Pani, Anton J.G. Horrevoets, Nicolle Kraenkel, Atsuhiko Neovascularization in Ischemic Limb Muscles Inhibits Angiogenesis. Implications for Diabetes-Induced Impaired ) Promotes Endothelial Cell Apoptosis and NTR Neurotrophin p75 Receptor (p75 http://circres.ahajournals.org/cgi/content/full/CIRCRESAHA.108.177386/DC1 Data Supplement (unedited) at: http://circres.ahajournals.org located on the World Wide Web at: The online version of this article, along with updated information and services, is http://www.lww.com/reprints Reprints: Information about reprints can be found online at [email protected] 410-528-8550. E-mail: Fax: Kluwer Health, 351 West Camden Street, Baltimore, MD 21202-2436. Phone: 410-528-4050. Permissions: Permissions & Rights Desk, Lippincott Williams & Wilkins, a division of Wolters http://circres.ahajournals.org/subscriptions/ Subscriptions: Information about subscribing to Circulation Research is online at by on May 22, 2011 circres.ahajournals.org Downloaded from
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Neurotrophin p75 Receptor (p75NTR) Promotes Endothelial Cell Apoptosis and Inhibits Angiogenesis:...

ISSN: 1524-4571 Copyright © 2008 American Heart Association. All rights reserved. Print ISSN: 0009-7330. Online
TX 72514Circulation Research is published by the American Heart Association. 7272 Greenville Avenue, Dallas,
DOI: 10.1161/CIRCRESAHA.108.177386 published online Jun 19, 2008; Circ. Res.
Madeddu and Costanza Emanueli Aurelie S. Leroyer, Chantal M. Boulanger, Gaia Spinetti, Sung Ok Yoon, Paolo
Oikawa, Graciela B. Sala-Newby, Marco Meloni, Brunella Cristofaro, Gallia Graiani, Andrea Caporali, Elisabetta Pani, Anton J.G. Horrevoets, Nicolle Kraenkel, Atsuhiko
Neovascularization in Ischemic Limb MusclesInhibits Angiogenesis. Implications for Diabetes-Induced Impaired
) Promotes Endothelial Cell Apoptosis andNTRNeurotrophin p75 Receptor (p75
http://circres.ahajournals.org/cgi/content/full/CIRCRESAHA.108.177386/DC1Data Supplement (unedited) at:
http://circres.ahajournals.org
located on the World Wide Web at: The online version of this article, along with updated information and services, is
http://www.lww.com/reprintsReprints: Information about reprints can be found online at
[email protected]. E-mail:
Fax:Kluwer Health, 351 West Camden Street, Baltimore, MD 21202-2436. Phone: 410-528-4050. Permissions: Permissions & Rights Desk, Lippincott Williams & Wilkins, a division of Wolters
http://circres.ahajournals.org/subscriptions/Subscriptions: Information about subscribing to Circulation Research is online at
by on May 22, 2011 circres.ahajournals.orgDownloaded from

Neurotrophin p75 Receptor (p75NTR) Promotes EndothelialCell Apoptosis and Inhibits Angiogenesis
Implications for Diabetes-Induced Impaired Neovascularization inIschemic Limb Muscles
Andrea Caporali,* Elisabetta Pani,* Anton J.G. Horrevoets, Nicolle Kraenkel, Atsuhiko Oikawa,Graciela B. Sala-Newby, Marco Meloni, Brunella Cristofaro, Gallia Graiani, Aurelie S. Leroyer,
Chantal M. Boulanger, Gaia Spinetti, Sung Ok Yoon, Paolo Madeddu, Costanza Emanueli
Abstract—Diabetes impairs endothelial function and reparative neovascularization. The p75 receptor of neurotrophins(p75NTR), which is scarcely present in healthy endothelial cells (ECs), becomes strongly expressed by capillary ECs afterinduction of peripheral ischemia in type-1 diabetic mice. Here, we show that gene transfer-induced p75NTR expressionimpairs the survival, proliferation, migration, and adhesion capacities of cultured ECs and endothelial progenitor cells(EPCs) and inhibits angiogenesis in vitro. Moreover, intramuscular p75NTR gene delivery impairs neovascularization andblood flow recovery in a mouse model of limb ischemia. These disturbed functions are associated with suppression ofsignaling mechanisms implicated in EC survival and angiogenesis. In fact, p75NTR depresses the VEGF-A/Akt/eNOS/NOpathway and additionally reduces the mRNA levels of ITGB1 [beta (1) integrin], BIRC5 (survivin), PTTG1 (securin)and VEZF1. Diabetic mice, which typically show impaired postischemic muscular neovascularization and bloodperfusion recovery, have these defects corrected by intramuscular gene transfer of a dominant negative mutant form ofp75NTR. Collectively, our data newly demonstrate the antiangiogenic action of p75NTR and open new avenues for thetherapeutic use of p75NTR inhibition to combat diabetes-induced microvascular liabilities. (Circ Res. 2008;103:0-0.)
Key Words: neurotrophins � p75NTR� VEGF-A � reparative neovascularization � limb ischemia
Diabetes impairs endothelial function and induces apopto-sis of endothelial cells (ECs) and endothelial progenitor
cells (EPCs). As a result, postischemic reparative angiogen-esis and vasculogenesis are both impaired in diabetic sub-jects.1–12 The consequences are harmful: in the leg, severemicroangiopathy aggravates atherosclerosis-induced muscu-lar ischemia, thus contributing to gangrene and cutaneousulcers with a severely impaired healing potential, whichmakes amputation of the diabetic foot an all-too-frequentnecessity.13,14 A remedy to prevent and treat diabetic micro-vascular complications is urgently needed.
Neurotrophins (NTs) have been extensively studied fortheir actions on the nervous system. However, it isbecoming increasingly evident that the expression andfunction of NTs is also important in the cardiovascularsystem. Both ECs and EPCs express tropomyosin kinasereceptors (trk), which are tyrosine-kinases binding NTswith high affinity. The NT nerve growth factor (NGF) and
brain-derived neurotrophic factor (BDNF), acting on trkAand trkB, respectively, promote EC survival and angiogen-esis.3,15–19 Moreover, NGF, via trkA, is an autocrinesurvival factor for cardiomyocytes.20 NTs have anotherreceptor of 75 kDa MW (p75NTR). The p75NTR contains adeath domain and belongs to the tumor necrosis factor �-�(TNF-�) receptor superfamily, but it does not bind TNF-�.In neural cells, p75NTR mediates apoptosis and cell cyclearrest.21–23 It was proposed that p75NTR has a low affinityfor mature NTs, as it preferentially binds proforms ofNTs.24 –26 However, the mechanisms underpinning p75NTR
activation and downstream molecular signaling have notbeen fully elucidated. Notably, no investigation of the roleof p75NTR on EC and EPC survival and function or onangiogenesis has been attempted to date.
We previously reported that p75NTR is scarcely representedin capillary ECs of healthy murine limb muscles, but thatexpression strikingly increases after induction of type-1
Original received April 11, 2008; revision received June 2, 2008; accepted June 11, 2008.From the Experimental Cardiovascular Medicine (A.C., E.P., N.K., A.O., M.M., B.C., P.M., C.E.) and Vascular Biology Divisions (G.B.S.-N.), Bristol
Heart Institute, University of Bristol, UK; IRCCS Multimedica Hospital (E.P., G.S.), Milan, Italy; the Department of Molecular Cell Biology andImmunology (A.J.G.H.), VU University Medical Center Amsterdam, Netherlands; Pathology (G.G.), University of Parma, Italy; INSERM CardiovascularResearch Center (A.S.L., C.N.B.), Hopital Lariboisiere, Paris, France; and the Center for Molecular Neurobiology (S.O.Y.), Ohio State University,Columbus.
*These authors contributed equally to this study.Correspondence to Costanza Emanueli, BSc, PhD, FAHA, Bristol Heart Institute, University of Bristol, Bristol Royal Infirmary, level 7, Upper Maudlin
Road BS2 8HW Bristol, UK. E-mail [email protected]© 2008 American Heart Association, Inc.
Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/CIRCRESAHA.108.177386
1 by on May 22, 2011 circres.ahajournals.orgDownloaded from

diabetes and hindlimb ischemia, 2 increasingly prevalentconditions that synergistically activate EC apoptosis.3,10,27
This study provides novel evidence that p75NTR impairssurvival and functions of cultured ECs and EPCs and ham-pers reparative neovascularization in ischemic limb muscles.We attribute the antiangiogenic effect of p75NTR to inhibitionof the vascular endothelial growth factor A (VEGF-A)/Aktkinase axis. Finally, we show that p75NTR inhibition benefitspostischemic healing in diabetes.
Materials and MethodsA detailed description of material and methods used is provided asonline supplement (available online at http://circres.ahajournals.org).The experiments involving mice were performed in accordancewith the Guide for the Care and Use of Laboratory Animalsprepared by the Institute of Laboratory Animal Resources and withprior approval of the British Home Office and the University ofBristol.
Cell CulturesHuman umbilical vein ECs (HUVECs) and human microvascularECs (MVECs) were grown in EGM-2 medium. EPCs were enrichedfrom peripheral blood mononuclear cells (PB-MNC) and cultured onfibronectin in the EPC medium EGM-2MV.
Adenoviruses (Ad) and In Vitro Gene TransferWe prepared Ad.p75NTR, containing the complete coding sequence ofhuman p75NTR with or without a V5 tag. The Ad.DN-p75NTR, consist-ing of extracellular and transmembrane domains of rat p75NTR linkedto a kinase-dead form the human EGF (cytoplasmic domain), hasbeen previously shown to inhibit apoptotic responses mediated byligand-mediated activation of the endogenous p75NTR receptor.28
Ad.Null and Ad.GFP served as controls. Ad.VEGF-A and Ad.Myr-Akt(expressing a constitutively active mutant form of Akt) have beenpreviously described.12,27 Cells were infected with Ads at 10 to 500mol/L.O.I.
Western Blot AnalysesWestern blot analyses for: mouse p75NTR, phosphorylated and totaleNOS, phosphorylated and total Akt, VEGF-A, phospho-Rb,phospho-FAK, cleaved-caspase-3, and GADPH (loading control)were performed.
FACS Analysis of HUVECs for p75NTR,Annexin-V, and Propidium IodideSurface antigen expression by HUVECs was analyzed by flowcytometry using a combination of a phycoerythrin-conjugated mouseantihuman p75NTR antibody and FITC-conjugated Annexin-V.Annexin-V binds to phosphatidyl serine exposed on the extracellularmembrane of apoptotic cells. Cells were also stained with propidiumiodide (PI).
In Vitro Apoptosis AssaysCaspase-3 activity was measured with a luminescent cell deathdetection kit. TUNEL assay was performed according to the manu-facturer’s instructions.
Analysis of Endothelial Apoptotic MicroparticlesEndothelial apoptotic microparticles (EMPs) were collected from theHUVEC-conditioned medium by centrifugation. EMPs were num-bered by flow cytometry analysis as AnnexinV-binding micropar-ticles. Analyses were performed using an antihuman p75NTR fluores-cent antibody and its isotype control. EMPs were analyzed in thepresence of Flowcount calibrator beads. EMP gate was defined asevents with a 0.1 to 1 �m diameter and examined on a fluorescence/forward light scatter plot.
Cell Cycle AnalysisTransduced HUVECs were synchronized by serum deprivationbefore being released in complete medium, and harvested at differenttime points. After fixation and RNase treatment, cells were stainedwith PI. DNA content was analyzed by flow cytometric analysis.
BrdU IncorporationTransduced HUVECs and EPCs were incubated with BrdU for 24hours and 72 hours, respectively. BrdU incorporation was measuredusing a BrdU immunofluorescence assay kit (HUVECs) or by FACS(EPCs).
Migration AssayMigration of transduced HUVECs and EPCs toward stromal cell–derived factor-1 (SDF-1) or 0.1% BSA (negative control indicativeof spontaneous migration) was analyzed by using fibronectin-coated24-well plate transwell migration inserts with a polycarbonatemembrane of 8-�m (HUVECs) or 5-�m (EPCs) pore size.
Adhesion AssayAdhesion of transduced HUVECs and EPCs to fibronectin wasevaluated.
Matrigel AssayTransduced HUVECs were seeded in 24-well plates coated withMatrigel supplemented with growth factors. Tube formation wasquantified in randomly captured microscopic fields by counting thenumber of intersection points and by measuring the % of areacovered by connected tubular structures.
NO Generation AssayNO generation by transduced HUVECs was assessed in a FACSCalibur flow cytometer by using DAF-2DA in the presence orabsence of the eNOS preferential inhibitor L-N5-(1-Iminoethyl)ornithine (L-NIO).
Gene Transfer to Murine MusclesAdenoviruses were delivered to the normoperfused or ischemic leftadductor muscles of anesthetized mice.
Illumina Beadarray Gene Expression ProfilingAnalyses are described in supplementary methods. Microarray ex-pression data are available at the NCBI Gene Expression Omnibus(GEO) (http://www.ncbi.nlm.nih.gov/project/geo) under accessionnumber GSE9910.
GEArrayHuman GEArray kit, composed of 113 genes connected to the p53signaling pathway and 2 housekeeping genes (actin and GADPH),was used to analyze the RNA of transduced HUVECs. The resultswere validated by real-time quantitative PCR.
Real-Time Quantitative RT-PCRQuantitative RT-PCR for mouse and human VEGF-A, ITGB1 andVEZF-1; human BIRC5, PTTG1, and p75NTR; and 18s rRNA wasperformed in a LightCycler.
Limb Ischemia and Diabetes ModelsLeft limb ischemia was induced in anesthetized CD1 male mice.Type-1 diabetes was induced by streptozotocin.
Clinical Outcome From Ischemia andBF MeasurementsThe number of autoamputated or necrotic fingers in the ischemic footwas calculated. Superficial BF of the ischemic and contralateral footwas analyzed by using the Lisca color laser Doppler. IntramuscularBF was measured by Oxylite/Oxyflow.
2 Circulation Research July 18, 2008
by on May 22, 2011 circres.ahajournals.orgDownloaded from
Histology and ImmunohistochemistryCapillary density was determined on muscular sections stained withH&E or for isolectin B4 (to identify ECs) and DAPI (to identify thenuclei). Cell apoptosis was recognized by TUNEL and proliferationby staining for the proliferation cell nuclear antigen (PCNA) or themini-chromosome maintenance protein 2 (MCM2). Transgene ex-pression by capillary ECs was determined by costaining sections forV5 and with isolectinB4. Endogenous mouse p75NTR expression byECs was identified by costaining for mouse p75NTR and withisolectinB4.
FACS Analyses of Bone Marrow CellsBM-MNC cells were obtained from mouse legs. Surface antigenexpression of MNC was analyzed by flow cytometry using combi-nations of fluorescent antibodies for Sca-1, lineage markers, andmouse p75NTR plus fluorescent Annexin-V.
Statistical AnalysisResults are expressed as the mean�SEM. Statistical analysis wasperformed via a t test when 2 groups were analyzed, or via an
ANOVA, followed by a Bonferroni post hoc test for multiplecomparisons. A probability value of �0.05 was taken as statisticallysignificant. The statistical analysis used for Illumina data aredescribed apart.
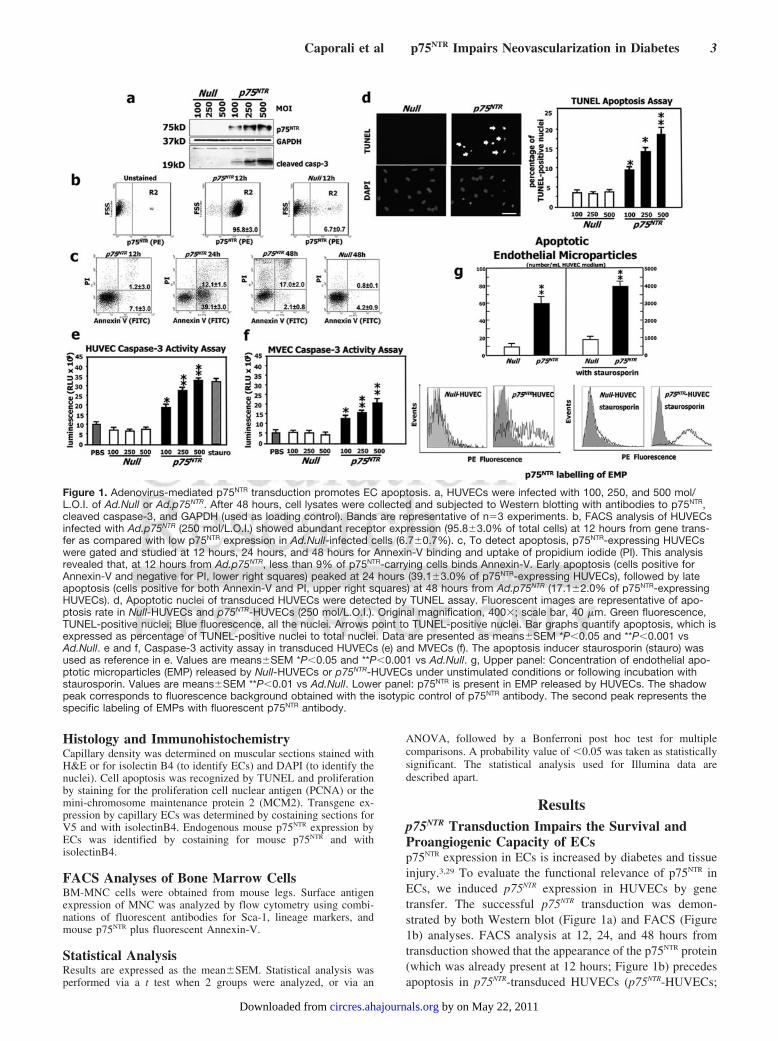
Resultsp75NTR Transduction Impairs the Survival andProangiogenic Capacity of ECsp75NTR expression in ECs is increased by diabetes and tissueinjury.3,29 To evaluate the functional relevance of p75NTR inECs, we induced p75NTR expression in HUVECs by genetransfer. The successful p75NTR transduction was demon-strated by both Western blot (Figure 1a) and FACS (Figure1b) analyses. FACS analysis at 12, 24, and 48 hours fromtransduction showed that the appearance of the p75NTR protein(which was already present at 12 hours; Figure 1b) precedesapoptosis in p75NTR-transduced HUVECs (p75NTR-HUVECs;
Figure 1. Adenovirus-mediated p75NTR transduction promotes EC apoptosis. a, HUVECs were infected with 100, 250, and 500 mol/L.O.I. of Ad.Null or Ad.p75NTR. After 48 hours, cell lysates were collected and subjected to Western blotting with antibodies to p75NTR,cleaved caspase-3, and GAPDH (used as loading control). Bands are representative of n�3 experiments. b, FACS analysis of HUVECsinfected with Ad.p75NTR (250 mol/L.O.I.) showed abundant receptor expression (95.8�3.0% of total cells) at 12 hours from gene trans-fer as compared with low p75NTR expression in Ad.Null-infected cells (6.7�0.7%). c, To detect apoptosis, p75NTR-expressing HUVECswere gated and studied at 12 hours, 24 hours, and 48 hours for Annexin-V binding and uptake of propidium iodide (PI). This analysisrevealed that, at 12 hours from Ad.p75NTR, less than 9% of p75NTR-carrying cells binds Annexin-V. Early apoptosis (cells positive forAnnexin-V and negative for PI, lower right squares) peaked at 24 hours (39.1�3.0% of p75NTR-expressing HUVECs), followed by lateapoptosis (cells positive for both Annexin-V and PI, upper right squares) at 48 hours from Ad.p75NTR (17.1�2.0% of p75NTR-expressingHUVECs). d, Apoptotic nuclei of transduced HUVECs were detected by TUNEL assay. Fluorescent images are representative of apo-ptosis rate in Null-HUVECs and p75NTR-HUVECs (250 mol/L.O.I.). Original magnification, 400�; scale bar, 40 �m. Green fluorescence,TUNEL-positive nuclei; Blue fluorescence, all the nuclei. Arrows point to TUNEL-positive nuclei. Bar graphs quantify apoptosis, which isexpressed as percentage of TUNEL-positive nuclei to total nuclei. Data are presented as means�SEM *P�0.05 and **P�0.001 vsAd.Null. e and f, Caspase-3 activity assay in transduced HUVECs (e) and MVECs (f). The apoptosis inducer staurosporin (stauro) wasused as reference in e. Values are means�SEM *P�0.05 and **P�0.001 vs Ad.Null. g, Upper panel: Concentration of endothelial apo-ptotic microparticles (EMP) released by Null-HUVECs or p75NTR-HUVECs under unstimulated conditions or following incubation withstaurosporin. Values are means�SEM **P�0.01 vs Ad.Null. Lower panel: p75NTR is present in EMP released by HUVECs. The shadowpeak corresponds to fluorescence background obtained with the isotypic control of p75NTR antibody. The second peak represents thespecific labeling of EMPs with fluorescent p75NTR antibody.
Caporali et al p75NTR Impairs Neovascularization in Diabetes 3
by on May 22, 2011 circres.ahajournals.orgDownloaded from

Figure 1c). In fact, early apoptosis (cells positive forAnnexin-V binding and negative for PI) peaked at 24 hours,followed, at 48 hours, by late apoptosis (Annexin-V�PI�
cells). We confirmed the proapoptotic effect of p75NTR inHUVECs by TUNEL apoptosis assay (Figure 1d), Westernblot for the apoptosis marker cleaved (activated) caspase-3(Figure 1a), and caspase-3 activity assay (Figure 1e). Addi-tionally, we measured the release of EMP30 from p75NTR- andNull-transduced HUVECs (Null-HUVECs) with or withoutexposure to the apoptosis inducer staurosporin. Under both
circumstances, p75NTR enhanced EMP release and p75NTR waspresent in EMP (Figure 1g). Taken together, these resultsprovide evidence that p75NTR promotes HUVEC apoptosis,via caspase-3 activation. The proapoptotic effect of p75NTR
was also present in MVECs (Figure 1f).In neural cells, p75NTR promotes cell cycle arrest.21 Because
EC replication is intrinsic to the angiogenesis process, westudied the impact of p75NTR on HUVEC cycle progression.HUVECs were synchronized at G1/S boundary by serumstarvation and the progression of cell cycle on release from
Figure 2. p75NTR inhibits EC cell cycle progression andimpairs EC ability to proliferate, migrate, adhere, andform capillary-like tubular structures in vitro. a, HUVECswere infected with either Ad.Null or Ad.p75NTR (250 mol/L.O.I.). Cells were synchronized by overnight serum star-vation. After release of cell cycle arrest by restoring nor-mal culture medium, cell cycle progression wasassessed by flow cytometric analysis, at the indicatedtime points. The percentage of cells in G1, S, and G2phases is indicated in figures. b, Western blot analysis ofphosphorylated tumor suppressor retinoblastoma protein(p-Rb) (marker of cell cycle progression) on HUVECextracts. GAPDH was used as loading control. c, Prolif-eration (measured by BrdU incorporation) of HUVECsinfected with Ad.Null or Ad.p75NTR (250 mol/L.O.I.) oruninfected (PBS). **P�0.01 vs Ad.Null. d, Migrationtoward SDF-1 (100 ng/mL) or PBS (vehicle) of p75NTR-HUVECs and Null-HUVECs. In bar graph, values aremeans�SEM #P�0.05 vs Ad.Null combined with PBS,*P�0.05 vs Ad.Null combined with SDF-1. Upper panelsshow representative microscopic fields (original magnifi-cation 100�; scale bar: 100 �m) e, Adhesion tofibronectin of p75NTR-HUVECs and Null-HUVECs. Celladhesion was quantified by counting adherent cells fol-
lowing washing of nonadherent cells and DAPI staining of nuclei of adherent cells. In bar graph, values are means�SEM **P�0.01 vsAd.Null. Upper panels show representative photomicrographs (magnification 200�, scale bar: 100 �m) f, Time-course (up to 24 hoursfrom cell seeding) of tubular structure formation in matrigel. Twelve hours before being seeded on Matrigel-coated dishes, HUVECswere infected with Ad.Null or Ad.p75NTR. In bar graphs, the quantification of EC tube network formation at 24 hours from seeding isexpressed as the number of intersecting points of tubular structures for microscopic field (left) as well as the % of microscopic fieldarea covered by connected tubular structures (right). Values are means�SEM **P�0.01 versus Ad.Null. Upper panels show representa-tive photomicrographs (original magnification 100�; scale bar: 100 �m).
4 Circulation Research July 18, 2008
by on May 22, 2011 circres.ahajournals.orgDownloaded from
the G1-block was analyzed by flow cytometry. As shown inFigure 2a, the control Null-HUVECs entered the cell cycleand progressed through S-phase and G2-mol/L. p75NTR-HUVECs were delayed in cycling, as shown by the reducedpercentage of cells in S-phase (P�0.01 versus Null-HUVECsat 21 hours). Correspondingly, the level of phosphorylatedtumor suppressor retinoblastoma protein (Rb), a marker ofcell cycle progression,31 was less in p75NTR-HUVECs (Figure2b). The antiproliferative effect of p75NTR on ECs wasconfirmed by using a BrdU incorporation assay (Figure 2c).
EC migration is essential for neovessel formation. Wetested the effect of p75NTR on EC migratory capacity toward
SDF-1, which is a classic chemotactic stimulus for cells of theendothelial lineage.32 At both 12 hours and 48 hours fromgene transfer, the migratory capacity of p75NTR-HUVECs toSDF-1 was severely impaired compared to Null-HUVECs(Figure 2d for 48 hours; 12-hour data not shown).
Cell-matrix adhesion is also fundamental for vessel growthand maintenance. At both 12 hours and 48 hours frominfection, p75NTR-HUVECs showed reduced adhesion capac-ity to fibronectin (Figure 2e for 48 hours; 12-hour data notshown).
Finally, we tested the angiogenic potential of p75NTR-HUVECs in a matrigel assay, in which ECs stimulated by
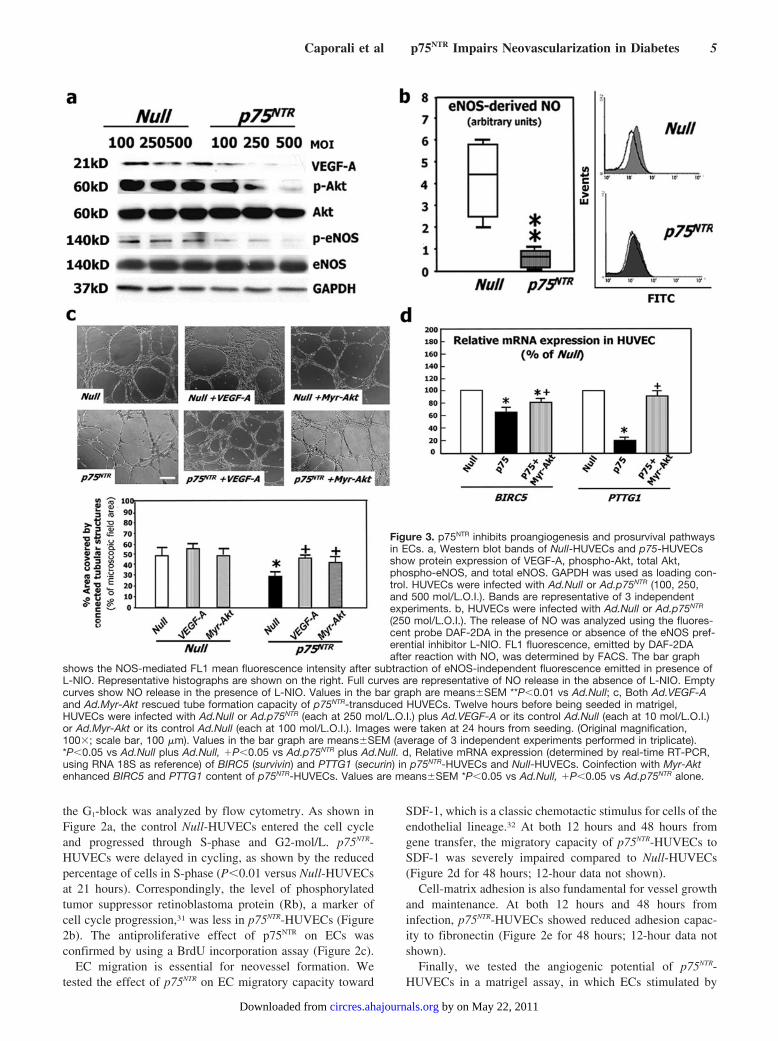
Figure 3. p75NTR inhibits proangiogenesis and prosurvival pathwaysin ECs. a, Western blot bands of Null-HUVECs and p75-HUVECsshow protein expression of VEGF-A, phospho-Akt, total Akt,phospho-eNOS, and total eNOS. GAPDH was used as loading con-trol. HUVECs were infected with Ad.Null or Ad.p75NTR (100, 250,and 500 mol/L.O.I.). Bands are representative of 3 independentexperiments. b, HUVECs were infected with Ad.Null or Ad.p75NTR
(250 mol/L.O.I.). The release of NO was analyzed using the fluores-cent probe DAF-2DA in the presence or absence of the eNOS pref-erential inhibitor L-NIO. FL1 fluorescence, emitted by DAF-2DAafter reaction with NO, was determined by FACS. The bar graph
shows the NOS-mediated FL1 mean fluorescence intensity after subtraction of eNOS-independent fluorescence emitted in presence ofL-NIO. Representative histographs are shown on the right. Full curves are representative of NO release in the absence of L-NIO. Emptycurves show NO release in the presence of L-NIO. Values in the bar graph are means�SEM **P�0.01 vs Ad.Null; c, Both Ad.VEGF-Aand Ad.Myr-Akt rescued tube formation capacity of p75NTR-transduced HUVECs. Twelve hours before being seeded in matrigel,HUVECs were infected with Ad.Null or Ad.p75NTR (each at 250 mol/L.O.I.) plus Ad.VEGF-A or its control Ad.Null (each at 10 mol/L.O.I.)or Ad.Myr-Akt or its control Ad.Null (each at 100 mol/L.O.I.). Images were taken at 24 hours from seeding. (Original magnification,100�; scale bar, 100 �m). Values in the bar graph are means�SEM (average of 3 independent experiments performed in triplicate).*P�0.05 vs Ad.Null plus Ad.Null, �P�0.05 vs Ad.p75NTR plus Ad.Null. d, Relative mRNA expression (determined by real-time RT-PCR,using RNA 18S as reference) of BIRC5 (survivin) and PTTG1 (securin) in p75NTR-HUVECs and Null-HUVECs. Coinfection with Myr-Aktenhanced BIRC5 and PTTG1 content of p75NTR-HUVECs. Values are means�SEM *P�0.05 vs Ad.Null, �P�0.05 vs Ad.p75NTR alone.
Caporali et al p75NTR Impairs Neovascularization in Diabetes 5
by on May 22, 2011 circres.ahajournals.orgDownloaded from
growth factors give rise to vessel-like tubular structures invitro, thus mimicking angiogenesis. In this assay, which wasinitiated at 12-hour postgene transfer, the antiangiogeniceffect of p75NTR was evident (Figure 2f).
p75NTR Induces a Proapoptosis andAntiangiogenesis Signaling Pathway in ECsp75NTR-HUVECs showed reduced levels of VEGF-A andSer473-phospho-Akt (Figure 3a). Phospho-Akt positivelycontrols EC survival and angiogenesis by several downstreammediators, including endothelial nitric oxide synthase(eNOS).33 Correspondingly, p75NTR transduction impairedSer1177-phosphorylation of eNOS (Figure 3a) and eNOS-derived NO production (Figure 3b). VEGF-A gene transferimproved Ser473-phospho-Akt level in p75NTR-HUVECs,whereas Myr-Akt did not rescue VEGF-A expression (datanot shown), thus confirming that VEGF-A lies upstream ofAkt phosphorylation.34 Phospho-eNOS content was increasedby either VEGF-A or a Myr-Akt gene transfer in p75NTR-HUVECs. Focal adhesion kinase (FAK), upstream of Akt,controls EC migration, proliferation, and survival.35 Phospho(active)-FAK was less in p75NTR-HUVECs and, as expected,
Myr-Akt did not restore phospho-FAK to normal levels. Bycontrast, VEGF-A increased phospho-FAK in p75NTR-HUVECs (data not shown). To investigate the role of de-pressed VEGF-A and phospho-Akt in the antiangiogenicaction of p75NTR, we performed a rescue experiment in thematrigel assay model. The capacity of p75NTR-HUVECs toform tubular structures was restored by either VEGF-A orMyr-Akt (Figure 3c), thus providing evidence that the antian-giogenic action of p75NTR is caused by inhibition of theVEGF-A/Akt pathway.
To further understand the p75NTR-induced molecularprogram contributing to EC apoptosis and antiangiogen-esis, we performed a GeArray for apoptosis and cellcycle-related genes on RNA extracted from p75NTR-HUVECs and Null-HUVECs. Results (not shown) werevalidated by real-time RT-PCR. As shown in Figure 3d,p75NTR reduced the mRNA expression of BIRC5 (survivin)and PTTG1 (pituitary tumor-derived transforming gene,also named securin), which both mediate EC survival andangiogenesis and are transcriptionally controlled byphospho-Akt.36 Accordingly, Myr-Akt increased BIRC5and PTTG1 in p75NTR-HUVECs (Figure 3d).
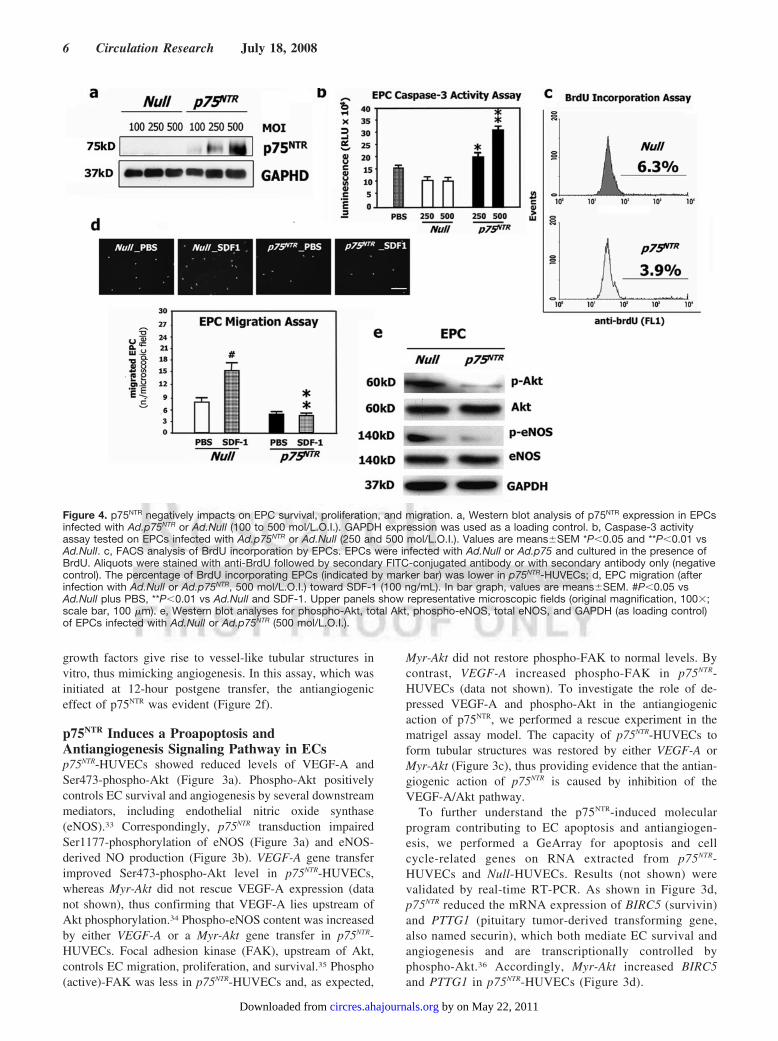
Figure 4. p75NTR negatively impacts on EPC survival, proliferation, and migration. a, Western blot analysis of p75NTR expression in EPCsinfected with Ad.p75NTR or Ad.Null (100 to 500 mol/L.O.I.). GAPDH expression was used as a loading control. b, Caspase-3 activityassay tested on EPCs infected with Ad.p75NTR or Ad.Null (250 and 500 mol/L.O.I.). Values are means�SEM *P�0.05 and **P�0.01 vsAd.Null. c, FACS analysis of BrdU incorporation by EPCs. EPCs were infected with Ad.Null or Ad.p75 and cultured in the presence ofBrdU. Aliquots were stained with anti-BrdU followed by secondary FITC-conjugated antibody or with secondary antibody only (negativecontrol). The percentage of BrdU incorporating EPCs (indicated by marker bar) was lower in p75NTR-HUVECs; d, EPC migration (afterinfection with Ad.Null or Ad.p75NTR, 500 mol/L.O.I.) toward SDF-1 (100 ng/mL). In bar graph, values are means�SEM. #P�0.05 vsAd.Null plus PBS, **P�0.01 vs Ad.Null and SDF-1. Upper panels show representative microscopic fields (original magnification, 100�;scale bar, 100 �m). e, Western blot analyses for phospho-Akt, total Akt, phospho-eNOS, total eNOS, and GAPDH (as loading control)of EPCs infected with Ad.Null or Ad.p75NTR (500 mol/L.O.I.).
6 Circulation Research July 18, 2008
by on May 22, 2011 circres.ahajournals.orgDownloaded from

p75NTR Transduction Impairs EPC Survivaland FunctionsThe growth of new blood vessels needs the participation ofboth angiogenesis, developed by preexisting vessels, andvasculogenesis, which involves EPCs. To study the impact ofp75NTR in EPCs derived from the human blood, we performedp75NTR gene transfer (Figure 4a). We found that p75NTR
negatively impacted on EPC survival (caspase-3 activityassay, Figure 4b), proliferation (FACS analysis of BrdUincorporation, Figure 4c), migration toward SDF-1 (Figure4d), and adhesion (data not shown). Moreover, p75NTR trans-duction reduced the levels of both phospho-Akt and phospho-eNOS in EPCs (Figure 4e).
p75NTR Transduction Impairs PostischemicNeovascularization and Blood Flow Recoveryin a Mouse Model of Limb IschemiaTo test the impact of p75NTR on blood vessel growth in vivo,Ad.p75NTR (V5-tagged) or Ad.Null was injected into theischemic adductor of normoglycemic mice. We verifiedsuccessful gene transfer by Western blot analysis for p75NTR
(supplemental Figure Ia). In addition, by immunohistochem-istry for V5 (red fluorescence), we observed localization ofthe transgene in capillary ECs (recognized by FITC-isolectinstaining -green fluorescence-) of the ischemic muscles (sup-plemental Figure Ib). Limb blood flow (BF) was measured bycolor laser Doppler at 7 and 14 days postischemia (n�12mice per group). Reduce BF to the ischemic foot wasobserved in the p75NTR group. In Figure 5a, the color laserDoppler images are representative of the BF to the ischemicfoot at 14 days postischemia, and the graph shows results atthe same time (data at 7 days are not shown). Correspond-ingly, capillary density was reduced in p75NTR-transduced
muscles (Figure 5b). In additional mice euthanized at 5 dayspostischemia, which, in comparison to 14 days, is a time pointmore informative on ischemia-induced EC proliferation andEC apoptosis responses, we observed increased EC apoptosis(Figure 5c) and reduced EC proliferation (Figure 5d) inp75NTR-transduced muscles.
Expressional Changes Induced by p75NTR
Transduction or Diabetes in Limb MusclesWe have used full genome expression profiling using Illu-mina bead-arrays interrogating 46 600 murine genome tran-scripts to study the global impact of p75NTR transduction onthe gene expression program of normoperfused mouse mus-cle. The overall changes in the transcriptome were analyzedat the levels of specific functional pathways using Panther-annotated protein group analysis.37 This analysis identified, inaddition to a number of inflammatory or muscle-specificpathways, a specific, statistically significant reduction ingene-sets involved in endothelial survival and angiogenesisprocesses, ie, VEGF-signaling and integrin-signaling (Figure6a). Analysis of the corresponding genes in more detailsidentified 3 pivotal genes (VEGF-A, ITGB1-[beta (1)integrin]-, VEZF1) that were suppressed by p75NTR, relative toa number of well-established endothelial-specific genes (datanot shown). By real-time RT-PCR, we confirmed that p75NTR
alters the expression of VEGF-A, ITGB1, and VEZF1 inmurine muscles (Figure 6b). To further confirm that thesemolecular changes are relative to the effect of p75NTR on ECs,we analyzed VEGF-A, ITGB1 and VEZF1 mRNA levels inp75- HUVECs and found all of them reduced in comparisonsto Null-HUVECs (Figure 6c). VEGF-A transfer correctedITGB1, but not VEZF1 deficit in p75NTR-HUVECs (Figure6c). Notably, muscular expression of VEGF-A, ITGB1, and
Figure 5. Effects of p75NTR transduction of ischemic limb muscles. a, Recovery of the bloodflow (BF) to the ischemic foot (expressed as the ratio between the BF in the ischemic foot tothe BF in the contralateral foot) at 14 days after ischemia induction and intramuscular deliv-ery of Ad.p75NTR or Ad.Null. Values are means�SEM. *P�0.05 vs Ad.Null. Representativelaser Doppler images taken at 14 days postischemia induction are also shown. Squares include the ischemic feet. Color from blue tobrown indicates progressive increases in BF. b, Capillary density of ischemic adductor muscles of mice described in a. Values aremeans�SEM. **P�0.01 vs Ad.Null. Representative microphotographs show muscle sections stained with isolectin B4 (red fluores-cence) to recognize ECs and counterstained with DAPI (blue fluorescence) to recognize nuclei. (Original magnification, 400�; scale bar,500 �m). Apoptosis (c; revealed by TUNEL assay) and proliferation (d; revealed by immunhistochemistry for the proliferating cell anti-gen [PCNA]) of ECs in limb muscles at 5 days postischemia induction and gene transfer. Values are means�SEM **P�0.01 vs Ad.Null.
Caporali et al p75NTR Impairs Neovascularization in Diabetes 7
by on May 22, 2011 circres.ahajournals.orgDownloaded from
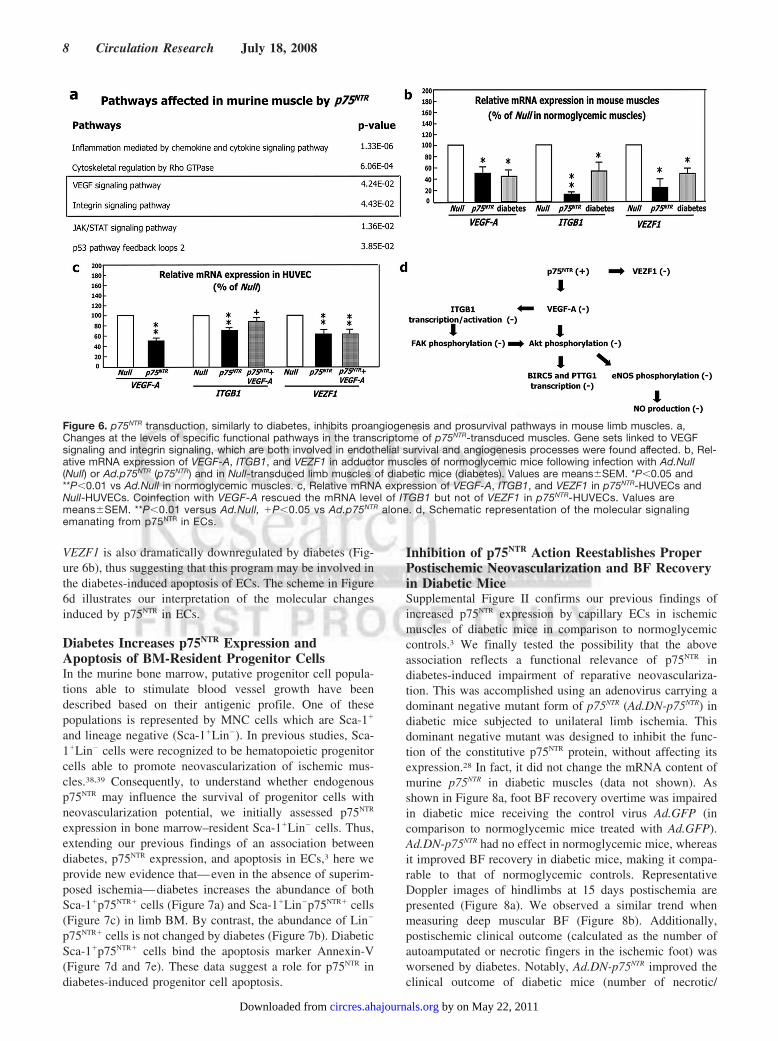
VEZF1 is also dramatically downregulated by diabetes (Fig-ure 6b), thus suggesting that this program may be involved inthe diabetes-induced apoptosis of ECs. The scheme in Figure6d illustrates our interpretation of the molecular changesinduced by p75NTR in ECs.
Diabetes Increases p75NTR Expression andApoptosis of BM-Resident Progenitor CellsIn the murine bone marrow, putative progenitor cell popula-tions able to stimulate blood vessel growth have beendescribed based on their antigenic profile. One of thesepopulations is represented by MNC cells which are Sca-1�
and lineage negative (Sca-1�Lin�). In previous studies, Sca-1�Lin� cells were recognized to be hematopoietic progenitorcells able to promote neovascularization of ischemic mus-cles.38,39 Consequently, to understand whether endogenousp75NTR may influence the survival of progenitor cells withneovascularization potential, we initially assessed p75NTR
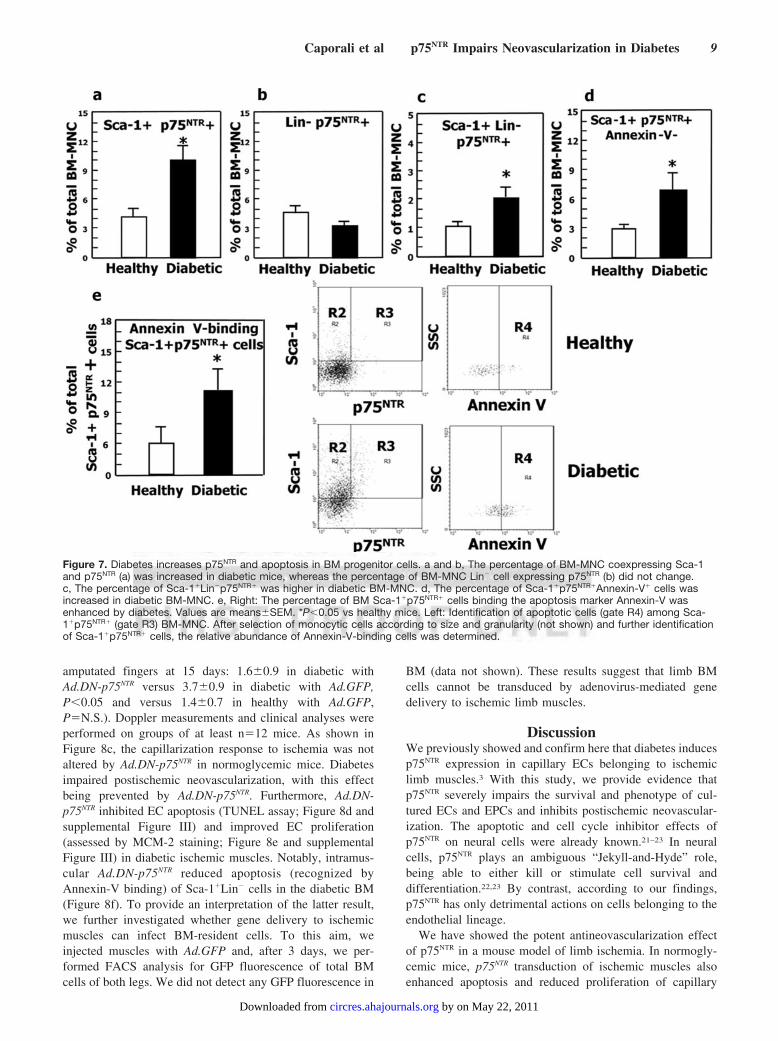
expression in bone marrow–resident Sca-1�Lin� cells. Thus,extending our previous findings of an association betweendiabetes, p75NTR expression, and apoptosis in ECs,3 here weprovide new evidence that—even in the absence of superim-posed ischemia—diabetes increases the abundance of bothSca-1�p75NTR� cells (Figure 7a) and Sca-1�Lin�p75NTR� cells(Figure 7c) in limb BM. By contrast, the abundance of Lin�
p75NTR� cells is not changed by diabetes (Figure 7b). DiabeticSca-1�p75NTR� cells bind the apoptosis marker Annexin-V(Figure 7d and 7e). These data suggest a role for p75NTR indiabetes-induced progenitor cell apoptosis.
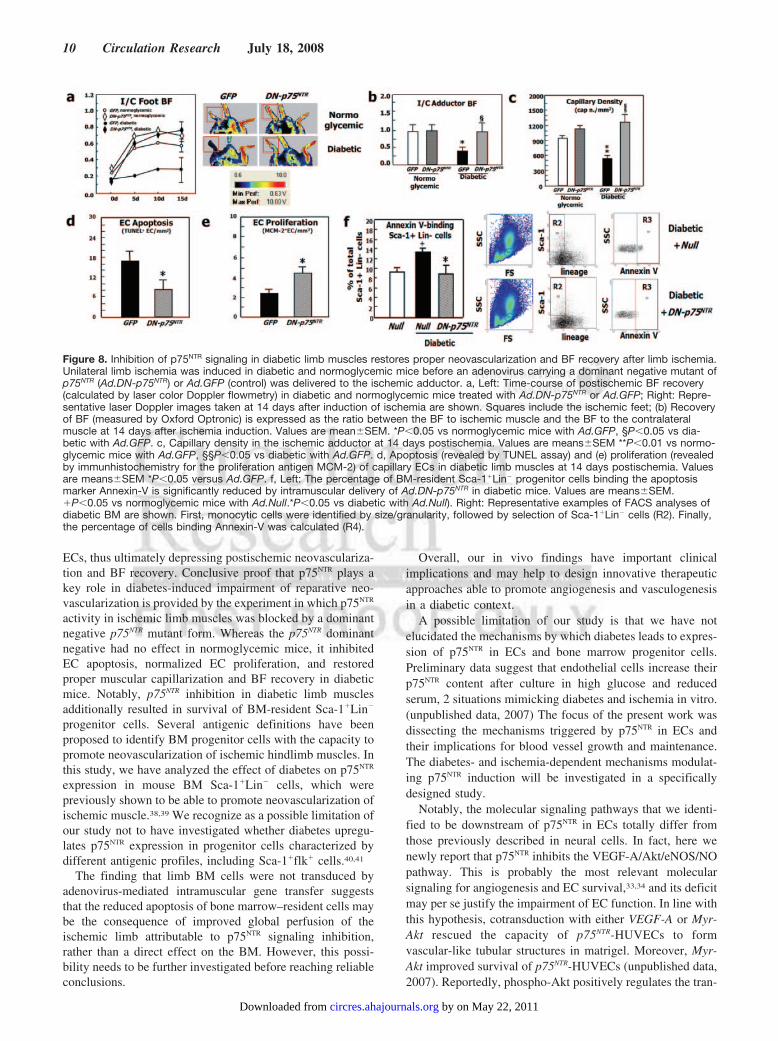
Inhibition of p75NTR Action Reestablishes ProperPostischemic Neovascularization and BF Recoveryin Diabetic MiceSupplemental Figure II confirms our previous findings ofincreased p75NTR expression by capillary ECs in ischemicmuscles of diabetic mice in comparison to normoglycemiccontrols.3 We finally tested the possibility that the aboveassociation reflects a functional relevance of p75NTR indiabetes-induced impairment of reparative neovasculariza-tion. This was accomplished using an adenovirus carrying adominant negative mutant form of p75NTR (Ad.DN-p75NTR) indiabetic mice subjected to unilateral limb ischemia. Thisdominant negative mutant was designed to inhibit the func-tion of the constitutive p75NTR protein, without affecting itsexpression.28 In fact, it did not change the mRNA content ofmurine p75NTR in diabetic muscles (data not shown). Asshown in Figure 8a, foot BF recovery overtime was impairedin diabetic mice receiving the control virus Ad.GFP (incomparison to normoglycemic mice treated with Ad.GFP).Ad.DN-p75NTR had no effect in normoglycemic mice, whereasit improved BF recovery in diabetic mice, making it compa-rable to that of normoglycemic controls. RepresentativeDoppler images of hindlimbs at 15 days postischemia arepresented (Figure 8a). We observed a similar trend whenmeasuring deep muscular BF (Figure 8b). Additionally,postischemic clinical outcome (calculated as the number ofautoamputated or necrotic fingers in the ischemic foot) wasworsened by diabetes. Notably, Ad.DN-p75NTR improved theclinical outcome of diabetic mice (number of necrotic/
Figure 6. p75NTR transduction, similarly to diabetes, inhibits proangiogenesis and prosurvival pathways in mouse limb muscles. a,Changes at the levels of specific functional pathways in the transcriptome of p75NTR-transduced muscles. Gene sets linked to VEGFsignaling and integrin signaling, which are both involved in endothelial survival and angiogenesis processes were found affected. b, Rel-ative mRNA expression of VEGF-A, ITGB1, and VEZF1 in adductor muscles of normoglycemic mice following infection with Ad.Null(Null) or Ad.p75NTR (p75NTR) and in Null-transduced limb muscles of diabetic mice (diabetes). Values are means�SEM. *P�0.05 and**P�0.01 vs Ad.Null in normoglycemic muscles. c, Relative mRNA expression of VEGF-A, ITGB1, and VEZF1 in p75NTR-HUVECs andNull-HUVECs. Coinfection with VEGF-A rescued the mRNA level of ITGB1 but not of VEZF1 in p75NTR-HUVECs. Values aremeans�SEM. **P�0.01 versus Ad.Null, �P�0.05 vs Ad.p75NTR alone. d, Schematic representation of the molecular signalingemanating from p75NTR in ECs.
8 Circulation Research July 18, 2008
by on May 22, 2011 circres.ahajournals.orgDownloaded from
amputated fingers at 15 days: 1.6�0.9 in diabetic withAd.DN-p75NTR versus 3.7�0.9 in diabetic with Ad.GFP,P�0.05 and versus 1.4�0.7 in healthy with Ad.GFP,P�N.S.). Doppler measurements and clinical analyses wereperformed on groups of at least n�12 mice. As shown inFigure 8c, the capillarization response to ischemia was notaltered by Ad.DN-p75NTR in normoglycemic mice. Diabetesimpaired postischemic neovascularization, with this effectbeing prevented by Ad.DN-p75NTR. Furthermore, Ad.DN-p75NTR inhibited EC apoptosis (TUNEL assay; Figure 8d andsupplemental Figure III) and improved EC proliferation(assessed by MCM-2 staining; Figure 8e and supplementalFigure III) in diabetic ischemic muscles. Notably, intramus-cular Ad.DN-p75NTR reduced apoptosis (recognized byAnnexin-V binding) of Sca-1�Lin� cells in the diabetic BM(Figure 8f). To provide an interpretation of the latter result,we further investigated whether gene delivery to ischemicmuscles can infect BM-resident cells. To this aim, weinjected muscles with Ad.GFP and, after 3 days, we per-formed FACS analysis for GFP fluorescence of total BMcells of both legs. We did not detect any GFP fluorescence in
BM (data not shown). These results suggest that limb BMcells cannot be transduced by adenovirus-mediated genedelivery to ischemic limb muscles.
DiscussionWe previously showed and confirm here that diabetes inducesp75NTR expression in capillary ECs belonging to ischemiclimb muscles.3 With this study, we provide evidence thatp75NTR severely impairs the survival and phenotype of cul-tured ECs and EPCs and inhibits postischemic neovascular-ization. The apoptotic and cell cycle inhibitor effects ofp75NTR on neural cells were already known.21–23 In neuralcells, p75NTR plays an ambiguous “Jekyll-and-Hyde” role,being able to either kill or stimulate cell survival anddifferentiation.22,23 By contrast, according to our findings,p75NTR has only detrimental actions on cells belonging to theendothelial lineage.
We have showed the potent antineovascularization effectof p75NTR in a mouse model of limb ischemia. In normogly-cemic mice, p75NTR transduction of ischemic muscles alsoenhanced apoptosis and reduced proliferation of capillary
Figure 7. Diabetes increases p75NTR and apoptosis in BM progenitor cells. a and b, The percentage of BM-MNC coexpressing Sca-1and p75NTR (a) was increased in diabetic mice, whereas the percentage of BM-MNC Lin� cell expressing p75NTR (b) did not change.c, The percentage of Sca-1�Lin�p75NTR� was higher in diabetic BM-MNC. d, The percentage of Sca-1�p75NTR�Annexin-V� cells wasincreased in diabetic BM-MNC. e, Right: The percentage of BM Sca-1�p75NTR� cells binding the apoptosis marker Annexin-V wasenhanced by diabetes. Values are means�SEM. *P�0.05 vs healthy mice. Left: Identification of apoptotic cells (gate R4) among Sca-1�p75NTR� (gate R3) BM-MNC. After selection of monocytic cells according to size and granularity (not shown) and further identificationof Sca-1�p75NTR� cells, the relative abundance of Annexin-V-binding cells was determined.
Caporali et al p75NTR Impairs Neovascularization in Diabetes 9
by on May 22, 2011 circres.ahajournals.orgDownloaded from
ECs, thus ultimately depressing postischemic neovasculariza-tion and BF recovery. Conclusive proof that p75NTR plays akey role in diabetes-induced impairment of reparative neo-vascularization is provided by the experiment in which p75NTR
activity in ischemic limb muscles was blocked by a dominantnegative p75NTR mutant form. Whereas the p75NTR dominantnegative had no effect in normoglycemic mice, it inhibitedEC apoptosis, normalized EC proliferation, and restoredproper muscular capillarization and BF recovery in diabeticmice. Notably, p75NTR inhibition in diabetic limb musclesadditionally resulted in survival of BM-resident Sca-1�Lin�
progenitor cells. Several antigenic definitions have beenproposed to identify BM progenitor cells with the capacity topromote neovascularization of ischemic hindlimb muscles. Inthis study, we have analyzed the effect of diabetes on p75NTR
expression in mouse BM Sca-1�Lin� cells, which werepreviously shown to be able to promote neovascularization ofischemic muscle.38,39 We recognize as a possible limitation ofour study not to have investigated whether diabetes upregu-lates p75NTR expression in progenitor cells characterized bydifferent antigenic profiles, including Sca-1�flk� cells.40,41
The finding that limb BM cells were not transduced byadenovirus-mediated intramuscular gene transfer suggeststhat the reduced apoptosis of bone marrow–resident cells maybe the consequence of improved global perfusion of theischemic limb attributable to p75NTR signaling inhibition,rather than a direct effect on the BM. However, this possi-bility needs to be further investigated before reaching reliableconclusions.
Overall, our in vivo findings have important clinicalimplications and may help to design innovative therapeuticapproaches able to promote angiogenesis and vasculogenesisin a diabetic context.
A possible limitation of our study is that we have notelucidated the mechanisms by which diabetes leads to expres-sion of p75NTR in ECs and bone marrow progenitor cells.Preliminary data suggest that endothelial cells increase theirp75NTR content after culture in high glucose and reducedserum, 2 situations mimicking diabetes and ischemia in vitro.(unpublished data, 2007) The focus of the present work wasdissecting the mechanisms triggered by p75NTR in ECs andtheir implications for blood vessel growth and maintenance.The diabetes- and ischemia-dependent mechanisms modulat-ing p75NTR induction will be investigated in a specificallydesigned study.
Notably, the molecular signaling pathways that we identi-fied to be downstream of p75NTR in ECs totally differ fromthose previously described in neural cells. In fact, here wenewly report that p75NTR inhibits the VEGF-A/Akt/eNOS/NOpathway. This is probably the most relevant molecularsignaling for angiogenesis and EC survival,33,34 and its deficitmay per se justify the impairment of EC function. In line withthis hypothesis, cotransduction with either VEGF-A or Myr-Akt rescued the capacity of p75NTR-HUVECs to formvascular-like tubular structures in matrigel. Moreover, Myr-Akt improved survival of p75NTR-HUVECs (unpublished data,2007). Reportedly, phospho-Akt positively regulates the tran-
Figure 8. Inhibition of p75NTR signaling in diabetic limb muscles restores proper neovascularization and BF recovery after limb ischemia.Unilateral limb ischemia was induced in diabetic and normoglycemic mice before an adenovirus carrying a dominant negative mutant ofp75NTR (Ad.DN-p75NTR) or Ad.GFP (control) was delivered to the ischemic adductor. a, Left: Time-course of postischemic BF recovery(calculated by laser color Doppler flowmetry) in diabetic and normoglycemic mice treated with Ad.DN-p75NTR or Ad.GFP; Right: Repre-sentative laser Doppler images taken at 14 days after induction of ischemia are shown. Squares include the ischemic feet; (b) Recoveryof BF (measured by Oxford Optronic) is expressed as the ratio between the BF to ischemic muscle and the BF to the contralateralmuscle at 14 days after ischemia induction. Values are mean�SEM. *P�0.05 vs normoglycemic mice with Ad.GFP, §P�0.05 vs dia-betic with Ad.GFP. c, Capillary density in the ischemic adductor at 14 days postischemia. Values are means�SEM **P�0.01 vs normo-glycemic mice with Ad.GFP, §§P�0.05 vs diabetic with Ad.GFP. d, Apoptosis (revealed by TUNEL assay) and (e) proliferation (revealedby immunhistochemistry for the proliferation antigen MCM-2) of capillary ECs in diabetic limb muscles at 14 days postischemia. Valuesare means�SEM *P�0.05 versus Ad.GFP. f, Left: The percentage of BM-resident Sca-1�Lin� progenitor cells binding the apoptosismarker Annexin-V is significantly reduced by intramuscular delivery of Ad.DN-p75NTR in diabetic mice. Values are means�SEM.�P�0.05 vs normoglycemic mice with Ad.Null.*P�0.05 vs diabetic with Ad.Null). Right: Representative examples of FACS analyses ofdiabetic BM are shown. First, monocytic cells were identified by size/granularity, followed by selection of Sca-1�Lin� cells (R2). Finally,the percentage of cells binding Annexin-V was calculated (R4).
10 Circulation Research July 18, 2008
by on May 22, 2011 circres.ahajournals.orgDownloaded from

scription of BIRC536 and PTTG1,42 which both resulted beingdownregulated by p75NTR. Notably, Myr-Akt totally or par-tially rescued BIRC5 and PTTG1 mRNA expression levels inp75NTR-HUVECs. BIRC5, which was named survivin for itsprosurvival action, was recently discovered to be a positivemediator of angiogenesis.36,43,44 Interestingly, BIRC5 alsomodulates cell cycle entry by interaction with Cdk4 andactivation of Cdk2/Cyclin E complex leading to Rb phos-phorylation.45 This may be reconciled with the impaired cellcycling capacity and reduced phospho-Rb levels observed inp75NTR-HUVECs. PTTG1 was also reported to positivelymodulate angiogenesis.42,46
Illumina technology-based analysis of p75NTR- and Null-transduced murine muscles followed by real-time RT-PCR ofthe same muscles and of p75- and Null-HUVECs identifiedthat p75NTR downregulates the mRNA expression of ITGB1[beta(1) integrin] and of the endothelial specific VEZF1transcription factor in ECs, in vitro and in vivo. Notably, wealso found downregulated VEGF-A, ITGB1, and VEZF-1 indiabetic muscles, which confirms previous reports ofVEGF-A reduction as the cause of diabetes-induced impairedneovascularization in skeletal muscles and myocardi-um.1,4,47,48 Reduction in ITGB1 content may be explainedwith the p75NTR-induced decrease in VEGF-A. In fact, inmicrovascular ECs, VEGF-A upregulates the mRNA expres-sion of alpha(6)beta(1) integrin. Interestingly, VEGF-A alsomodulates the activity of alpha(6)beta(1) and alpha(9)beta(1)integrins, which in turn mediate VEGF-A driven angiogene-sis.49,50 In line, VEGF-A rescued ITGB1 level in p75NTR-HUVECs, once again suggesting that VEGF-A downregula-tion plays an important role in orchestrating the molecularchanges triggered by p75NTR in ECs. The only outsider to thisVEGF-A–based molecular network seems to be VEZF1,whose impaired mRNA content was not corrected byVEGF-A transfer in p75NTR-HUVECs. VEZF1 is a relativelyscarcely explored transcription factor that was implicated inin utero vasculogenesis, postnatal angiogenesis, and ECsurvival.51–53 Our expressional data discount the control ofVEZF1 by VEGF-A. It is also improbable that VEZF1 liesupstream of VEGF-A, because the antiangiogenic effectcaused by knocking down VEZF1 expression was reportedlynot corrected in the presence of VEGF-A53. Further studiesare necessary to define the role of decreased VEZF1 indiabetes-induced microvascular complications.
Our interpretation of the coordinated changes induced byp75NTR in the molecular program of ECs is summarized inFigure 6d. Future studies are necessary to elucidate themolecular mechanisms by which p75NTR modulates the ex-pressional level of VEGF-A in HUVECs and murine limbmuscles. In addition, considering the importance of VEGF-Afor the survival of neurons,54 we speculate that VEGF-Adownregulation might be also involved in the well establishedproapoptotic effect of p75NTR on neural cells.55 This hypoth-esis should be validated by future studies.
We finally conclude that p75NTR overexpression is suffi-cient and necessary for altering angiogenesis in diabetes andthus becomes an obligatory target for therapy.
AcknowledgmentsWe thank Dr Mark Bond (Bristol Heart Institute) for importantsuggestions and gift of reagents, Dr Andrew Herman (Molecular andCellular Medicine, University of Bristol) for help in setting theconditions for FACS analyses, Mauro Siragusa (Bristol Heart Insti-tute) for help in preparing fluorescent images, and Dr Elinor Griffiths(Bristol Heart Institute) for editing the manuscript.
Sources of FundingThis study was supported by the British Heart Foundation (BHF)(RJ4769, RJ4430, and SM6266 to C.E.) and by the Europeancommunity FP6 through the European Vascular Genomic Networkof Excellence (EVGN) (to A.J.G.H., C.M.B., P.M., and C.E.).
DisclosuresNone.
References1. Yoon YS, Uchida S, Masuo O, Cejna M, Park JS, Gwon HC, Kirchmair
R, Bahlman F, Walter D, Curry C, Hanley A, Isner JM, Losordo DW.Progressive attenuation of myocardial vascular endothelial growth factorexpression is a seminal event in diabetic cardiomyopathy: restoration ofmicrovascular homeostasis and recovery of cardiac function in diabeticcardiomyopathy after replenishment of local vascular endothelial growthfactor. Circulation. 2005;111:2073–2085.
2. Tepper OM, Galiano RD, Capla JM, Kalka C, Gagne PJ, Jacobowitz GR,Levine JP, Gurtner GC. Human endothelial progenitor cells from type IIdiabetics exhibit impaired proliferation, adhesion, and incorporation intovascular structures. Circulation. 2002;106:2781–2786.
3. Salis MB, Graiani G, Desortes E, Caldwell RB, Madeddu P, Emanueli C.Nerve growth factor supplementation reverses the impairment,induced by Type 1 diabetes, of hindlimb post-ischaemic recovery inmice. Diabetologia. 2004;47:1055–1063.
4. Rivard A, Silver M, Chen D, Kearney M, Magner M, Annex B, Peters K,Isner JM. Rescue of diabetes-related impairment of angiogenesis byintramuscular gene therapy with adeno-VEGF. Am J Pathol. 1999;154:355–363.
5. Loomans CJ, de Koning EJ, Staal FJ, Rookmaaker MB, Verseyden C, deBoer HC, Verhaar MC, Braam B, Rabelink TJ, van Zonneveld AJ.Endothelial progenitor cell dysfunction: a novel concept in the patho-genesis of vascular complications of type 1 diabetes. Diabetes. 2004;53:195–199.
6. Krankel N, Adams V, Linke A, Gielen S, Erbs S, Lenk K, Schuler G,Hambrecht R. Hyperglycemia reduces survival and impairs function ofcirculating blood-derived progenitor cells. Arterioscler Thromb VascBiol. 2005;25:698–703.
7. Ingram DA, Lien IZ, Mead LE, Estes M, Prater DN, Derr-Yellin E,Dimeglio L, Haneline LS. In Vitro Hyperglycemia or a Diabetic Intra-uterine Environment Reduces Neonatal Endothelial Colony Forming CellNumbers and Function. Diabetes. 2008;57:724–731.
8. Gallagher KA, Liu ZJ, Xiao M, Chen H, Goldstein LJ, Buerk DG, NedeauA, Thom SR, Velazquez OC. Diabetic impairments in NO-mediatedendothelial progenitor cell mobilization and homing are reversed byhyperoxia and SDF-1 alpha. J Clin Invest. 2007;117:1249–1259.
9. Gadau S, Emanueli C, Van Linthout S, Graiani G, Todaro M, Meloni M,Campesi I, Invernici G, Spillmann F, Ward K, Madeddu P. Benfotiamineaccelerates the healing of ischaemic diabetic limbs in mice throughprotein kinase B/Akt-mediated potentiation of angiogenesis and inhi-bition of apoptosis. Diabetologia. 2006;49:405–420.
10. Emanueli C, Monopoli A, Kraenkel N, Meloni M, Gadau S, Campesi I,Ongini E, Madeddu P. Nitropravastatin stimulates reparative neovascu-larisation and improves recovery from limb Ischaemia in type-1 diabeticmice. Br J Pharmacol. 2007;150:873–882.
11. Emanueli C, Graiani G, Salis MB, Gadau S, Desortes E, Madeddu P.Prophylactic gene therapy with human tissue kallikrein ameliorates limbischemia recovery in type 1 diabetic mice. Diabetes. 2004;53:1096–1103.
12. Emanueli C, Caporali A, Krankel N, Cristofaro B, Van Linthout S,Madeddu P. Type-2 diabetic Lepr(db/db) mice show a defective micro-vascular phenotype under basal conditions and an impaired response toangiogenesis gene therapy in the setting of limb ischemia. Front Biosci.2007;12:2003–2012.
13. Brem H, Tomic-Canic M. Cellular and molecular basis of wound healingin diabetes. J Clin Invest. 2007;117:1219–1222.
Caporali et al p75NTR Impairs Neovascularization in Diabetes 11
by on May 22, 2011 circres.ahajournals.orgDownloaded from

14. Jeffcoate WJ, Harding KG. Diabetic foot ulcers. Lancet. 2003;361:1545–1551.
15. Emanueli C, Salis MB, Pinna A, Graiani G, Manni L, Madeddu P. Nervegrowth factor promotes angiogenesis and arteriogenesis in ischemichindlimbs. Circulation. 2002;106:2257–2262.
16. Cantarella G, Lempereur L, Presta M, Ribatti D, Lombardo G, LazaroviciP, Zappala G, Pafumi C, Bernardini R. Nerve growth factor-endothelialcell interaction leads to angiogenesis in vitro and in vivo. Faseb J.2002;16:1307–1309.
17. Kim H, Li Q, Hempstead BL, Madri JA. Paracrine and autocrinefunctions of brain-derived neurotrophic factor (BDNF) and nerve growthfactor (NGF) in brain-derived endothelial cells. J Biol Chem. 2004;279:33538–33546.
18. Kermani P, Rafii D, Jin DK, Whitlock P, Schaffer W, Chiang A, VincentL, Friedrich M, Shido K, Hackett NR, Crystal RG, Rafii S, HempsteadBL. Neurotrophins promote revascularization by local recruitment ofTrkB� endothelial cells and systemic mobilization of hematopoieticprogenitors. J Clin Invest. 2005;115:653–663.
19. Turrini P, Gaetano C, Antonelli A, Capogrossi MC, Aloe L. Nerve growthfactor induces angiogenic activity in a mouse model of hindlimb ische-mia. Neurosci Lett. 2002;323:109–112.
20. Caporali A, Sala-Newby GB, Meloni M, Graiani G, Pani E, Cristofaro B,Newby AC, Madeddu P, Emanueli C. Identification of the prosurvivalactivity of nerve growth factor on cardiac myocytes. Cell Death Differ.2008;15:299–311.
21. Lopez-Sanchez N, Frade JM. Control of the cell cycle by neurotrophins:lessons from the p75 neurotrophin receptor. Histol Histopathol. 2002;17:1227–1237.
22. Ibanez CF. Jekyll-Hyde neurotrophins: the story of proNGF. TrendsNeurosci. 2002;25:284–286.
23. Reichardt LF. Neurotrophin-regulated signalling pathways. Philos TransR Soc Lond B Biol Sci. 2006;361:1545–1564.
24. Lee R, Kermani P, Teng KK, Hempstead BL. Regulation of cell survivalby secreted proneurotrophins. Science. 2001;294:1945–1948.
25. Nykjaer A, Lee R, Teng KK, Jansen P, Madsen P, Nielsen MS, JacobsenC, Kliemannel M, Schwarz E, Willnow TE, Hempstead BL, Petersen CM.Sortilin is essential for proNGF-induced neuronal cell death. Nature.2004;427:843–848.
26. Kenchappa RS, Zampieri N, Chao MV, Barker PA, Teng HK, HempsteadBL, Carter BD. Ligand-dependent cleavage of the P75 neurotrophinreceptor is necessary for NRIF nuclear translocation and apoptosis insympathetic neurons. Neuron. 2006;50:219–232.
27. Emanueli C, Salis MB, Van Linthout S, Meloni M, Desortes E, SilvestreJS, Clergue M, Figueroa CD, Gadau S, Condorelli G, Madeddu P.Akt/protein kinase B and endothelial nitric oxide synthase mediatemuscular neovascularization induced by tissue kallikrein gene transfer.Circulation. 2004;110:1638–1644.
28. Harrington AW, Kim JY, Yoon SO. Activation of Rac GTPase by p75 isnecessary for c-jun N-terminal kinase-mediated apoptosis. J Neurosci.2002;22:156–166.
29. Graiani G, Emanueli C, Desortes E, Van Linthout S, Pinna A, FigueroaCD, Manni L, Madeddu P. Nerve growth factor promotes reparativeangiogenesis and inhibits endothelial apoptosis in cutaneous wounds ofType 1 diabetic mice. Diabetologia. 2004;47:1047–1054.
30. Boulanger CM, Amabile N, Tedgui A. Circulating microparticles: a potentialprognostic marker for atherosclerotic vascular disease. Hypertension. 2006;48:180–186.
31. Knudsen ES, Wang JY. Differential regulation of retinoblastoma proteinfunction by specific Cdk phosphorylation sites. J Biol Chem. 1996;271:8313–8320.
32. Urbich C, Aicher A, Heeschen C, Dernbach E, Hofmann WK, ZeiherAM, Dimmeler S. Soluble factors released by endothelial progenitor cellspromote migration of endothelial cells and cardiac resident progenitorcells. J Mol Cell Cardiol. 2005;39:733–742.
33. Papapetropoulos A, Garcia-Cardena G, Madri JA, Sessa WC. Nitric oxideproduction contributes to the angiogenic properties of vascular endothe-lial growth factor in human endothelial cells. J Clin Invest. 1997;100:3131–3139.
34. Ackah E, Yu J, Zoellner S, Iwakiri Y, Skurk C, Shibata R, Ouchi N,Easton RM, Galasso G, Birnbaum MJ, Walsh K, Sessa WC. Akt1/proteinkinase Balpha is critical for ischemic and VEGF-mediated angiogenesis.J Clin Invest. 2005;115:2119–2127.
35. Angelucci A, Bologna M. Targeting vascular cell migration as a strategyfor blocking angiogenesis: the central role of focal adhesion proteintyrosine kinase family. Curr Pharm Des. 2007;13:2129–2145.
36. Papapetropoulos A, Fulton D, Mahboubi K, Kalb RG, O’Connor DS, LiF, Altieri DC, Sessa WC. Angiopoietin-1 inhibits endothelial cell apo-ptosis via the Akt/survivin pathway. J Biol Chem. 2000;275:9102–9105.
37. Hochberg Y, Benjamini Y. More powerful procedures for multiple sig-nificance testing. Stat Med. 1990;9:811–818.
38. Chavakis E, Aicher A, Heeschen C, Sasaki K, Kaiser R, El Makhfi N,Urbich C, Peters T, Scharffetter-Kochanek K, Zeiher AM, Chavakis T,Dimmeler S. Role of beta2-integrins for homing and neovascularizationcapacity of endothelial progenitor cells. J Exp Med. 2005;201:63–72.
39. Sbaa E, Dewever J, Martinive P, Bouzin C, Frerart F, Balligand JL, DessyC, Feron O. Caveolin plays a central role in endothelial progenitor cellmobilization and homing in SDF-1-driven postischemic vasculogenesis.Circ Res. 2006;98:1219–1227.
40. Iwakura A, Luedemann C, Shastry S, Hanley A, Kearney M, Aikawa R,Isner JM, Asahara T, Losordo DW. Estrogen-mediated, endothelial nitricoxide synthase-dependent mobilization of bone marrow-derived endothe-lial progenitor cells contributes to reendothelialization after arterialinjury. Circulation. 2003;108:3115–3121.
41. Qin G, Ii M, Silver M, Wecker A, Bord E, Ma H, Gavin M, GoukassianDA, Yoon YS, Papayannopoulou T, Asahara T, Kearney M, Thorne T,Curry C, Eaton L, Heyd L, Dinesh D, Kishore R, Zhu Y, Losordo DW.Functional disruption of alpha4 integrin mobilizes bone marrow-derivedendothelial progenitors and augments ischemic neovascularization. J ExpMed. 2006;203:153–163.
42. Thompson AD III, Kakar SS. Insulin and IGF-1 regulate the expressionof the pituitary tumor transforming gene (PTTG) in breast tumor cells.FEBS Lett. 2005;579:3195–3200.
43. O’Connor DS, Schechner JS, Adida C, Mesri M, Rothermel AL, Li F, NathAK, Pober JS, Altieri DC. Control of apoptosis during angiogenesis bysurvivin expression in endothelial cells. Am J Pathol. 2000;156:393–398.
44. Zwerts F, Lupu F, De Vriese A, Pollefeyt S, Moons L, Altura RA, JiangY, Maxwell PH, Hill P, Oh H, Rieker C, Collen D, Conway SJ, ConwayEM. Lack of endothelial cell survivin causes embryonic defects in angio-genesis, cardiogenesis, and neural tube closure. Blood. 2007;109:4742–4752.
45. Suzuki A, Hayashida M, Ito T, Kawano H, Nakano T, Miura M, AkahaneK, Shiraki K. Survivin initiates cell cycle entry by the competitiveinteraction with Cdk4/p16(INK4a) and Cdk2/cyclin E complex acti-vation. Oncogene. 2000;19:3225–3234.
46. Heaney AP, Horwitz GA, Wang Z, Singson R, Melmed S. Earlyinvolvement of estrogen-induced pituitary tumor transforming gene andfibroblast growth factor expression in prolactinoma pathogenesis. NatMed. 1999;5:1317–1321.
47. Boodhwani M, Sodha NR, Mieno S, Xu SH, Feng J, Ramlawi B,Clements RT, Sellke FW. Functional, cellular, and molecular character-ization of the angiogenic response to chronic myocardial ischemia indiabetes. Circulation. 2007;116:I31–37.
48. Kivela R, Silvennoinen M, Touvra AM, Lehti TM, Kainulainen H, VihkoV. Effects of experimental type 1 diabetes and exercise training onangiogenic gene expression and capillarization in skeletal muscle. FasebJ. 2006;20:1570–1572.
49. Lee TH, Seng S, Li H, Kennel SJ, Avraham HK, Avraham S. Integrinregulation by vascular endothelial growth factor in human brain micro-vascular endothelial cells: role of alpha6beta1 integrin in angiogenesis.J Biol Chem. 2006;281:40450–40460.
50. Vlahakis NE, Young BA, Atakilit A, Hawkridge AE, Issaka RB,Boudreau N, Sheppard D. Integrin alpha9beta1 directly binds to vascularendothelial growth factor (VEGF)-A and contributes to VEGF-A-inducedangiogenesis. J Biol Chem. 2007;282:15187–15196.
51. Xiong JW, Leahy A, Lee HH, Stuhlmann H. Vezf1: A Zn finger tran-scription factor restricted to endothelial cells and their precursors. DevBiol. 1999;206:123–141.
52. Kuhnert F, Campagnolo L, Xiong JW, Lemons D, Fitch MJ, Zou Z, KiossesWB, Gardner H, Stuhlmann H. Dosage-dependent requirement for mouseVezf1 in vascular system development. Dev Biol. 2005;283:140–156.
53. Miyashita H, Kanemura M, Yamazaki T, Abe M, Sato Y. Vascularendothelial zinc finger 1 is involved in the regulation of angiogenesis:possible contribution of stathmin/OP18 as a downstream target gene.Arterioscler Thromb Vasc Biol. 2004;24:878–884.
54. Greenberg DA, Jin K. From angiogenesis to neuropathology. Nature.2005;438:954–959.
55. Dechant G, Barde YA. The neurotrophin receptor p75(NTR): novelfunctions and implications for diseases of the nervous system. NatNeurosci. 2002;5:1131–1136.
12 Circulation Research July 18, 2008
by on May 22, 2011 circres.ahajournals.orgDownloaded from

CIRCRESAHA/2008/177386/R2 page 1
Caporali et al, p75NTR impairs neovascularization in diabetes
EXPANDED METHODS Cell cultures HUVEC and MVEC were purchased from Cambrex (Belgium) and grown in EGM-2 medium (EBM-2 supplemented with EGM-2 SingleQuots, Cambrex) containing 5% FBS (Cambrex). HUVEC and MVEC were used between the second and the fifth passage in culture. Human endothelial progenitor cells (EPC) were enriched from peripheral blood of healthy volunteers. Peripheral blood mononuclear cells (PB-MNC) were separated by density-gradient centrifugation (Histopaque 1077, Sigma), washed in phosphate buffered saline (PBS) and plated on fibronectin (10μg/ml, Sigma)-coated tissue culture plates in the EPC medium EGM-2MV (EBM-2 MV supplemented by EGM-2 MV SingleQuots, Cambrex) containing 10% FBS at a density of 1x106 MNC per cm2. At day 4, non-adherent cells were removed and the adherent EPC were used for experiments. Adenoviruses (Ad) In order to prepare Ad. p75NTR, the complete coding sequence of human p75NTR was excised using EcoRI from a Bluescript plasmid (generous gift from Prof. Dario Marchetti, Louisiana State University, USA) and cloned in the adenovirus shuttle vector pDC515 (Microbix Biosystems, Canada) downstream of the mouse cytomegalovirus promoter.
Replication-deficient adenoviruses were generated by site-specific FLP-mediated recombination of the cotransfected shuttle and genomic plasmids in 293 cells. Viral stocks were amplified, CsCl banded, and titrated as previously described1. To add a C-terminal V5 tag, the complete coding sequence of p75NTR was amplified using KOD proofreading DNA polymerase (Novagen) to remove the stop codon. A perfect clone was excised and subcloned in a pDC515 shuttle vector containing the coding sequence of the V5 tag ready for adenovirus generation. Adenoviral vectors carrying human p75NTR (Ad. p75NTR) with or without a V5-tag were prepared. Ad-rad66 (empty vector, also called Ad.Null) was used as negative control. Ad.DN-p75NTR was provided by the co-author Dr. Yoon, who previously reported its propierties.2 Briefly, the mutant p75NTR contains the extracellular
and transmembrane domains of rat p75NTR, but its cytoplasmic domain was replaced with that of the human EGF that was rendered kinase-dead. Accordingly, this mutant inhibits the signaling capacity of endogenous p75NTR by binding with neurotrophin ligands and it does not influence expression of endogenous p75NTR. The capacity of the dominant negative p75NTR to be expressed on the cell surface and bind neurotrophins as efficiently as the full-length p75NTR has been shown2. It has also been shown that this dominant negative p75NTR specifically inhibits apoptotic responses evoked by neurotrophins in rat
by on May 22, 2011 circres.ahajournals.orgDownloaded from

CIRCRESAHA/2008/177386/R2 page 2
oligodendrocytes which express the endogenous receptor2. Ad.VEGF-A and Ad.Myr-Akt expressing a constitutively active mutant form of Akt were previously used by us4, 5. in vitro gene transfer In order to infect HUVEC, MVEC, or human EPC, Ad.VEGF-A and its control Ad.Null was used at 10 M.O.I. Ad.Myr-Akt and its control Ad.Null was used at 100 M.O.I. The other Ads were used at 100 to 500 M.O.I.
Western-Blot analyses Proteins were extracted from cultured cells, muscles, or BM by using ice-cold buffer A (50mM Hepes, 150mM NaCl, 1mM EDTA, 1mM EGTA, 25mM NaF, 5mM NaPPi, 1%Triton, 1% NP40, 1 mM Na3VO4, 0,25% sodium deoxycholate, 0.5mM Na-orthovanadate, 1 mM benzamidine, 0.1mM phenylmethylsulfonyl fluoride) or ice-cold RIPA buffer. Protein concentration was determined using the Bio-Rad Protein Assay Reagent (Bio-Rad). Detection of proteins by Western blot analysis was done following separation of whole cell extracts (50μg) on SDS-polyacrylamide gels. Proteins were transferred to polyvinylidene difluoride membranes (PVDF, Amersham-Pharmacia) and probed with the following antibodies: anti mouse p75NTR (clone ME20.4, Upstate Biotechnology, 1:1000), Ser 1177-phospho-eNOS (Cell Signaling, 1:1000), eNOS (Santa Cruz Biotechnology, 1:500), VEGF-A (Santa Cruz Biotechnology, 1:2000), Ser473-phospho-Akt (Cell Signaling, 1:1000), Akt (Cell Signaling, 1:1000), phospho-Rb (Upstate Biotechnology, 1:500), p-FAK (pY397, Transduction Laboratories, 1:1000), cleaved-caspase-3 (Cell Signaling, 1:1000), V5 (Novus Biological, 1:1000). GADPH (R&D System, 1:2000) was used as loading control. For detection, secondary antibody goat anti rabbit or anti mouse conjugated to horseradish peroxidase (Amersham Pharmacia, 1:5000) were used, followed by chemiluminescence reaction (ECL, Amersham Pharmacia). FACS analysis of HUVEC for p75NTR, Annexin-V, and PI Surface antigen expression by HUVEC was analyzed by flow cytometry using combinations of a phycoerythrin (PE)-conjugated mouse anti-human p75NTR antibody and FITC-conjugated Annexin-V (both from BD Bioscience Pharmigen (BD). Cells were also stained with PI. Unstained and single stained controls were performed to define positivity. Data were obtained on a FACS Calibur flow cytometer (BD) and analyzed using WinMDI (Purdue University, West Lafayette, USA). Cell numbers are expressed as percent of HUVEC. The experiment was performed in triplicate and repeated 3 times. Caspase-Glo assay HUVEC (7x103/well) and EPC (3x104/well) were seeded in flat bottomed 96-well plates and infected with Ad.Null or Ad.p75NTR (each at 100, 250, and 500 M.O.I.). Caspase-3
by on May 22, 2011 circres.ahajournals.orgDownloaded from

CIRCRESAHA/2008/177386/R2 page 3
activity was measured by using a luminescent cell death detection kit (Caspase-Glo Assay, Promega), according to the instruction of the manufacturer. The apoptosis inducer staurosporin (1μM, for 4h) was used as a positive control of apoptotic death. Each experiment was performed in triplicate and repeated at least 3 times.
TUNEL apoptosis assay of HUVEC and EPC HUVEC or EPC were seeded (1.5x104/well) on gelatine and fibronectin-coated Labtek™ slides (Nalge Nunc Int.) and infected with Ad.Null or Ad.p75NTR overnight. After 48h, cells were fixed with 4% paraformaldehyde. TUNEL assay was performed according to the manufacturer’s instructions (Calbiochem). Nuclei were counterstained with DAPI. At least 500 cells (in eight randomly captured microscopic fields at 400x magnification) were scored in a blinded fashion to calculate the percentage of TUNEL-positive nuclei. Data are expressed as the average percentage of apoptotic nuclei. The experiment was performed in triplicate and repeated 3 times. Flow cytometry analysis of apoptotic microparticles Endothelial microparticles (EMP) were collected from the conditioned culture medium of HUVEC by two centrifugation steps (1500 x g for 15 min, followed by 20500 x g for 5 min) and stored and at -80°C. Then, EMP were numbered by flow cytometry analysis as AnnexinV-binding microparticles, as described6. Analyses were performed using PE-conjugated mouse anti-human p75NTR antibody and its corresponding isotype control provided by BD. EMP (20μL) were incubated with 20μL of antibody for 30min in the dark. EMP were then analysed on a Beckman-Coulter EPICS XL flow cytometer in the presence of Flowcount calibrator beads. EMP gate was defined as events with a 0.1-1μm diameter and examined on a fluorescence/forward light scatter plot. The experiment was repeated 3 times. Cell cycle analysis HUVEC were infected with Ad.p75NTR or Ad.Null (150 M.O.I.) and then synchronized by serum deprivation (0.1% FBS) for 16h. Next, cells were washed twice with PBS, released in complete medium, and harvested at different time points (indicated in Fig. 2a). After fixation in 70% ethanol and RNase A (100μg/ml, Sigma) treatment for 1h at 37°C, cells were stained with PI (50μg/ml, Sigma) on ice in the dark for 30min. DNA content was analyzed by flow cytometric analysis (Becton Dickinson FACScan, San Jose California) and quantification was performed by using the software Modfit LT for MAC version 2.0 (from Verity Software house, Topsham, Maine, USA). The experiment was repeated 3 times. BrdU incorporation
by on May 22, 2011 circres.ahajournals.orgDownloaded from

CIRCRESAHA/2008/177386/R2 page 4
HUVEC (5x103/well) were seeded in a 96-well plate and infected overnight (in EBM2 supplemented with 0.1% FBS) with Ad.Null or Ad.p75NTR (250 M.O.I.). Then, the medium was replaced with complete EGM2 added with BrdU (10μM) for 24h. BrdU incorporation by HUVEC was measured by using a BrdU immunofluorescence assay kit from Roche, according to the manufacturer's instructions. Briefly, HUVEC were fixed and made permeable with FixDenat solution for 1h, then incubated with monoclonal anti-BrdU peroxidase-conjugated antibody (anti-BrdU-POD) for 90min. Bound anti-BrdU-POD was detected by a substrate reaction, then quantified by an ELISA plate reader. Each experiment was performed using 6 wells per group and repeated 3 times.
EPC were plated in a 6-well plate and infected with Ad.Null or Ad. p75NTR (500 M.O.I.) in EBM2 with 0.1% FBS overnight. Then, the medium was changed in complete EGM2 containing BrdU (10μM) for 72h and the incorporation of BrdU by EPC was measured by FACS. Briefly, EPC were trypsinized, washed with PBS, and fixed with 70% ethanol on ice for 30min. Then, DNA was denaturated in 2N HCl with 0.5% Triton X-100 for 30min at room temperature and neutralized in 0.1M Na2B4O7, pH 8.5. Cells were finally incubated with monoclonal anti-BrdU antibody (Dako, 1:100) for 1h at room temperature, centrifuged, washed, stained with FITC-conjugated secondary for 30min and read at a FACS Calibur (Becton Dickinson). Evaluation was performed blind. Data were analyzed using CellQuest software (Becton Dickinson). Each experiment was performed in triplicates and repeated 3 times.
Migration assay The migratory function of p75NTR or Null-transduced HUVEC and EPC was evaluated (at 12h and 48h from gene transfer) by using fibronectin-coated (10μg/ml, Sigma) 24-well-plate transwell migration inserts (Corning, Artington, UK) with a polycarbonate membrane of 8-μm (HUVEC) or 5-μm (EPC) pore size. Briefly, cells were trypsinized and resuspended in EBM-2 containing 0.1% bovine serum albumin (BSA, Sigma). Cells (0.5x105) were placed in the upper chamber, while EBM-2 containing 0.1% BSA and human recombinant SDF-1 (100ng/mL) was placed in the lower chamber. To control for spontaneous migration (negative control), EBM-2 with 0.1% BSA was used in the lower chamber. Cells were incubated in a humidified incubator at 37°C for 5h (HUVEC) or overnight (EPC). Non-migrated cells were removed by gently wiping the upper surface of the membrane with a cotton swab. Membranes were fixed in methanol, excised with a scalpel and mounted on microscope slides with mounting medium containing DAPI (Vectashield, Vector Technologies). Level of migration was assessed by counting the number of cells present on the lower surface of the membrane in 8 randomly captured fluorescence microscopic view fields (magnification 100X). Evaluation was performed blind. Each experiment was performed in triplicate and repeated 3 times.
by on May 22, 2011 circres.ahajournals.orgDownloaded from

CIRCRESAHA/2008/177386/R2 page 5
Adhesion assay Ninety-six-well tissue culture plates were coated for 1h with fibronectin (10μg/ml in PBS) and then blocked for 1h at 37°C with 1% BSA in PBS and washed 3 times with PBS before cell seeding. At 12h and 48h from infection with Ad.p75NTR or Ad.Null (250 M.O.I. for HUVEC, 500 M.O.I. for EPC), transduced HUVEC (15x103/well) or EPC (20x103/well) were seeded in EGM2 with 10% FCS and incubated at 37°C. One hour later, non-adherent cells were discarded by washing 3 times and adherent cells were fixed (10 % formalin in PBS, pH 7.4 for 10min at room temperature) and stained with DAPI (2.5μg/ml, Sigma,) for 10min in the dark. Plates were then rinsed extensively with water and cell adhesion was quantified by counting adherent cells (magnification 200x). Evaluation was performed blind. Each experiment was performed in triplicate and repeated 3 times. Matrigel assay HUVEC (105 cell/well) were infected with Ad.p75NTR or Ad.Null (250 M.O.I.) and, after 12h or 24h, seeded in 24-well plates coated with 300 μL of Matrigel supplemented with growth factors (Becton Dickinson). For rescue experiments with VEGF-A and Myr-Akt, cells were co-infected with Ad.p75NTR or Ad.Null (250 M.O.I.) plus either 10 M.O.I. of Ad.VEGF-A or its control Ad.Null or 100 M.O.I. of Ad.Myr-Akt or its control Ad.Null. Plates were incubated at 37°C and photographed at different times from seeding. Capillary-like structure formation was examined under an inverted phase photomicroscope. Tube formation was quantified by counting the number of intersection
points in randomly captured microscopic fields and/or by calculating the % microscopic field area covered by connected tubular structures (magnification 100x). Evaluation was performed blind. Each experiment was performed in triplicate and repeated 3 times. NO generation assay NO generation was assessed in p75NTR-HUVEC and Null-HUVEC. Briefly, transduced HUVEC were detached by trypsinization and divided into four groups with n=4 replicates containing 2x105 cells. Samples were pre-incubated in sample buffer (200 mM Tris-HCl, 10mM CaCl2, 10mM MgCl2, 1.33mM NaCl, 65mM KCl, 5mM D-glucose, 0.1% BSA and 50μM L-arginine (all Sigma). To the p75NTRL-NIO and NullL-NIO groups, the NOS inhibitor L-N5-(1-Iminoethyl)ornithine (L-NIO, 100μM, Merck, UK) was added for 10min whereas the other two groups remained without inhibitor. DAF-2DA (5μM; Invitrogen, UK) was added to all samples and incubated in the dark for 3h at 37°C. Cells were washed with sample buffer and immediately FL1 fluorescence was determined in a FACS Calibur flow cytometer (BD). FL1 mean fluorescence intensity was calculated and unspecific fluorescence remaining after NOS inhibition by L-NIO was subtracted from
by on May 22, 2011 circres.ahajournals.orgDownloaded from

CIRCRESAHA/2008/177386/R2 page 6
uninhibited FL1 fluorescence intensity to obtain NOS specific DAF fluorescence values reflecting NO generation. Gene transfer to murine muscles Anaesthetized mice received 3 adenovirus injections (for a total of 109p.f.u. virus in 20μL) into 3 equidistant sites of the normoperfused or ischemic left adductor muscles. When desired, ischemia was induced (vide infra) immediately before gene transfer. Illumina beadarray gene expression profiling RNA samples from Null and p75NTR infected murine muscle were amplified and biotinylated using the Illumina TotalPrep RNA amplification Kit (Ambion, Austin, TX). Eight samples that passed quality control were randomly allocated to Sentrix Mouse-6 Expression bead chip arrays (Illumina, San Diego, CA), interrogating 46,600 transcripts from the mouse genome. Samples were hybridized to the beadchip arrays, followed by scanning and feature extraction, all performed at ServiceXS (Leiden, The Netherlands). Array data were extracted using Illumina BeadStudio software. Normalization and statistical analysis of the bead summary data from the arrays was carried out using the limma package 5 in R/Bioconductor (www.bioconductor.org). Bead summary intensities were normalized using quantile normalization and differential expression between the treatments of interest was assessed using a moderated t-test 7. This test is similar to a standard t-test for each probe except that the standard errors are moderated across genes to ensure more stable inference for each gene using Baysian statistics. Genes were considered significant if the P values, adjusted for multiple testing by using Benjamini and Hochberg's method 8 were <0.05. The false discovery rate was thereby controlled to be <5%. Differential modulation of specific pathways was assessed using the Panther-annotated protein group expression statistical analysis 9. Micro-array expression data are available at the NCBI Gene Expression Omnibus (GEO) (http://www.ncbi.nlm.nih.gov/project/geo) under accession number GSE9910. GEArray Human GEArray kit (SuperArray Inc., Bethesda, MD, USA), composed of 113 genes connected to the p53 signaling pathway and two housekeeping genes (actin and GADPH), was used to characterize gene expression profile of HUVEC cells infected with Ad.p75NTR or Ad.Nul (250 M.O.I.). Total RNA was isolated from cells by RNeasy isolation kit (Qiagen, Crawley, UK). cRNA probes for array analysis were synthesized following the manufacturer's directions. Differential gene expression patterns were detected by autoradiography. The experiment was performed in triplicate and data were confirmed by real-time quantitative PCR.
by on May 22, 2011 circres.ahajournals.orgDownloaded from

CIRCRESAHA/2008/177386/R2 page 7
Real-time quantitative RT-PCR
Total RNA was isolated from HUVEC and murine muscles (RNeasy, Qiagen, UK) and RNA quality confirmed using the RNA Nano LabChip in a bioanalyzer (Agilent). RNA was reverse transcribed (Sensiscript reverse transcriptase, Qiagen) and quantitative PCR was performed in a LightCycler (Roche, Burgess Hill, UK) using the following primers: 18s rRNA (forward: 5’- TAGAGGGACAAGTGGCGTTC -3’, reverse: 5’- TGTACAAAGGGCAGGGACTT-3’), murine VEGF-A (forward: GGAGATCCTTCGAGGAGCACTT, reverse: GGCGATTTAGCAGCAGATATAAGAA), human VEGF-A (forward: CTACCTCCACCATGCCAAGTG, reverse: GATGATTCTGCCCTCCTCCTT), human BIRC5 (forward: TCCGGTTGCGCTTTCCT, reverse: TCTTCTTATTGTTGGTTTCCTT), human PTTG1 (forward: CGGCTGTTAAGACCTGCAATA , reverse: TTCAGCCCATCCTTAGCAACC), mouse ITGB1 (forward: CGTGGTTGCCGGAATTGTTC, reverse: ACCAGCTTTACGTCCATAGTTTG), human ITGB1 (forward: AAGCTTTTAATGATAATTCATGACAG, reverse: GCCACTTTGCTTTTTGGATG), murine VEZF1 (forward: CAGCAGCCGAACTTCGTTG, reverse: CAGGCTTACTGGGTTTCTTGAC), human VEZF1 (forward: GACAGCAGCCGAACTTCGTT, reverse: TGCCCGAGGAAGATGTAGTGA), human p75NTR (forward: GTGGGCCTTGTGGCCTAC, reverse: TGTGGCCTACATAGCCTTC). The analyses were performed on at least 4 samples per time and repeated three times. Limb ischemia and diabetes models Left limb ischemia was induced in anaesthetized CD1 male mice, as described5, 10-12. Type-1 diabetes was induced by streptozotocin, as also described13. Diabetic mice did not received insulin and were studied between 1 and 3 months from diabetes onset. In all experiments, normoglycemic controls were age-and gender-matched. Clinical outcome from ischemia and BF measurements Following limb ischemia induction, we evaluated the toe and foot necrosis event, as described 10, 11, 14. Superficial BF of the ischemic and contralateral foot was analyzed at different time-points from ischemia induction by color laser Doppler (Lisca Perimed, Sweden) and the ratio between BF in the ischemic foot and BF in controlateral foot was calculated and used as an index of % BF recovery5,10-12,14. Intra-muscular BF was measured by Oxylite/Oxyflow (Oxford Optronic, Oxford, UK) by inserting a probe in the adductor muscle of anaesthetized mice4. The number of auto-amputated or necrotic fingers in the ischemic foot was calculated, as previously explained10, 11. A minimum of n=12 mice per group were studied.
by on May 22, 2011 circres.ahajournals.orgDownloaded from

CIRCRESAHA/2008/177386/R2 page 8
Capillary density Capillary density was determined on H&E-stained transverse sections of adductor muscles (observed at 400x magnification) by the aid of an ocular graticule, as previously described5,10-12,14 and in fluorescent sections stained for isolectin B4 (red fluorescence) (Molecular Porbes) to identify EC and with DAPI (blue fluorescence) (Sigma Aldrich) to identify nuclei. A minimum of n=6 muscles per each group were used for these analyses. Apoptosis and proliferation in muscles Apoptosis was recognized by TUNEL assay, as previously described5, 10-12,14. Cell proliferation was recognized by IHC for either the proliferation cell nuclear antigen (PCNA) or the mini-chromosome maintenance protein 2 (MCM2), a marker of cycling cells, as also previously published4, 15. Additional sections were triple stained for either TUNEL, isolectin B4, and DAPI or PCNA, isolectin B4, and DAPI. A minimum of n=6 muscles per each group were used for these analyses. Other immunohistochemical analyses of muscular sections Following the delivery of V5-tagged Ad.p75NTR to adductor muscles, transgene expression by capillary EC was evaluated by staining paraffin-embedded adductor sections with a FITC-conjugated primary antibody recognizing V5 (Novus Biological, 1:200) and EC by Alexa 568-conjugated isolectinB4 (Molecular Probe, 1:200). In a separate experiment, expression of endogenous mouse p75NTR by EC was analyzed by co-staining adductor sections with a FITC-conjugated primary antibody recognizing mouse p75NTR (Calbiochem, 1:200) and Alexa 568-conjugated isolectinB4.
FACS analyses of bone marrow cells BM-MNC were obtained from the left leg of freshly killed CD1 mice, as described previously16. Briefly, tibiae and femora were flushed with DMEM (Cambrex) containing 5% FCS (Biochrom, Berlin, Germany). Cells were separated and passed through a 70μm cell strainer (BD, Oxford, UK). Single cell suspensions were centrifuged on Histopaque 1083 (Sigma) to obtain MNC. Surface antigen expression of MNC was analyzed by flow cytometry using combinations of PE-labeled anti-Sca-1 (BD), FITC-labelled anti-lineage markers-Alexa 488 (Caltag), FITC-labeled PE-labeled or biotynilated Annexin-V (BD), and Cy5 labeled-p75NTR (Advance Targeting System, CA, USA) together with PE- or FITC-labelled anti-rabbit secondary antibody (Sigma Aldrich) (all monoclonal). Unstained and staining controls for single colors and secondary antibodies were performed to define positivity. Data were obtained on a FACS Calibur flow cytometer (BD) and analyzed using WinMDI (Purdue University, West Lafayette, USA). Cell numbers are expressed as percent of a sub-population as specified. A minimum of n=6 mice per group were used for these analyses.
by on May 22, 2011 circres.ahajournals.orgDownloaded from

CIRCRESAHA/2008/177386/R2 page 9
FACS for GFP fluorescence of BM cells Anaesthetized normoglycemic and diabetic mice received 3 injections of Ad.GFP (for a total of 109p.f.u. in 20μL) into 3 equidistant sites of the ischemic left adductor muscles. After 3d, BM-MNC were obtained from both legs of freshly killed CD1 mice. The presence of green fluorescent BM-MNC was determined by FACS Calibur flow cytometer (BD). A minimum of n=5 mice per group were used for these analyses.
REFERENCES
1. Sala-Newby GB, Freeman NV, Curto MA, Newby AC. Metabolic and functional consequences of cytosolic 5'-nucleotidase-IA overexpression in neonatal rat cardiomyocytes. Am J Physiol Heart Circ Physiol. 2003;285:H991-998.
2. Harrington AW, Kim JY, Yoon SO. Activation of Rac GTPase by p75 is necessary for c-jun N-terminal kinase-mediated apoptosis. J Neurosci. 2002;22:156-166.
3. He TC, Zhou S, da Costa LT, Yu J, Kinzler KW, Vogelstein B. A simplified system for generating recombinant adenoviruses. Proc Natl Acad Sci U S A. 1998;95:2509-2514.
4. Emanueli C, Salis MB, Van Linthout S, Meloni M, Desortes E, Silvestre JS, Clergue M, Figueroa CD, Gadau S, Condorelli G, Madeddu P. Akt/protein kinase B and endothelial nitric oxide synthase mediate muscular neovascularization induced by tissue kallikrein gene transfer. Circulation. 2004;110:1638-1644.
5. Emanueli C, Caporali A, Krankel N, Cristofaro B, Van Linthout S, Madeddu P. Type-2 diabetic Lepr(db/db) mice show a defective microvascular phenotype under basal conditions and an impaired response to angiogenesis gene therapy in the setting of limb ischemia. Front Biosci. 2007;12:2003-2012.
6. Leroyer AS, Isobe H, Leseche G, Castier Y, Wassef M, Mallat Z, Binder BR, Tedgui A, Boulanger CM. Cellular origins and thrombogenic activity of microparticles isolated from human atherosclerotic plaques. J Am Coll Cardiol. 2007;49:772-777.
7. Baldi P, Long AD. A Bayesian framework for the analysis of microarray expression data: regularized t -test and statistical inferences of gene changes. Bioinformatics. 2001;17:509-519.
8. Thomas PD, Kejariwal A, Guo N, Mi H, Campbell MJ, Muruganujan A, Lazareva-Ulitsky B. Applications for protein sequence-function evolution data: mRNA/protein expression analysis and coding SNP scoring tools. Nucleic Acids Res. 2006;34:W645-650.
by on May 22, 2011 circres.ahajournals.orgDownloaded from

CIRCRESAHA/2008/177386/R2 page 10
9. Benjamini Y H, Y. . Controlling the false discovery rate: a practical and powerful approach to multiple testing. Journal of the Royal Statistical Society. 1995;57:289-300.
10. Gadau S, Emanueli C, Van Linthout S, Graiani G, Todaro M, Meloni M, Campesi I, Invernici G, Spillmann F, Ward K, Madeddu P. Benfotiamine accelerates the healing of ischaemic diabetic limbs in mice through protein kinase B/Akt-mediated potentiation of angiogenesis and inhibition of apoptosis. Diabetologia. 2006;49:405-420.
11. Invernici G, Emanueli C, Madeddu P, Cristini S, Gadau S, Benetti A, Ciusani E, Stassi G, Siragusa M, Nicosia R, Peschle C, Fascio U, Colombo A, Rizzuti T, Parati E, Alessandri G. Human fetal aorta contains vascular progenitor cells capable of inducing vasculogenesis, angiogenesis, and myogenesis in vitro and in a murine model of peripheral ischemia. Am J Pathol. 2007;170:1879-1892.
12. Salis MB, Graiani G, Desortes E, Caldwell RB, Madeddu P, Emanueli C. Nerve growth factor supplementation reverses the impairment, induced by Type 1 diabetes, of hindlimb post-ischaemic recovery in mice. Diabetologia. 2004;47:1055-1063.
13. Emanueli C, Salis MB, Pinna A, Stacca T, Milia AF, Spano A, Chao J, Chao L, Sciola L, Madeddu P. Prevention of diabetes-induced microangiopathy by human tissue kallikrein gene transfer. Circulation. 2002;106:993-999.
14. Emanueli C, Monopoli A, Kraenkel N, Meloni M, Gadau S, Campesi I, Ongini E, Madeddu P. Nitropravastatin stimulates reparative neovascularisation and improves recovery from limb Ischaemia in type-1 diabetic mice. Br J Pharmacol. 2007;150:873-882.
15. Graiani G, Lagrasta C, Migliaccio E, Spillmann F, Meloni M, Madeddu P, Quaini F, Padura IM, Lanfrancone L, Pelicci P, Emanueli C. Genetic deletion of the p66Shc adaptor protein protects from angiotensin II-induced myocardial damage. Hypertension. 2005;46:433-440.
16. Madeddu P, Kraenkel N, Barcelos LS, Siragusa M, Campagnolo P, Oikawa A, Caporali A, Herman A, Azzolino O, Barberis L, Perino A, Damilano F, Emanueli C, Hirsch E. Phosphoinositide 3-kinase gamma gene knockout impairs postischemic neovascularization and endothelial progenitor cell functions. Arterioscler Thromb Vasc Biol. 2008;28:68-76.
by on May 22, 2011 circres.ahajournals.orgDownloaded from
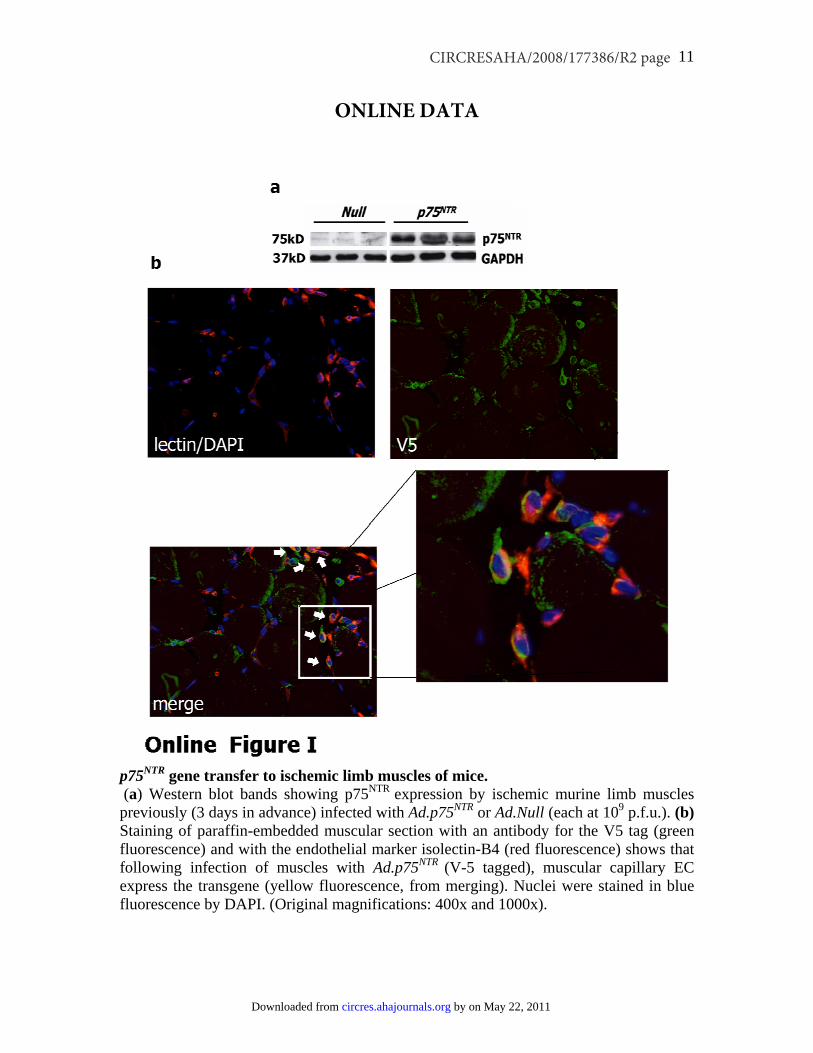
CIRCRESAHA/2008/177386/R2 page 11
ONLINE DATA
p75NTR gene transfer to ischemic limb muscles of mice. (a) Western blot bands showing p75NTR expression by ischemic murine limb muscles previously (3 days in advance) infected with Ad.p75NTR or Ad.Null (each at 109 p.f.u.). (b) Staining of paraffin-embedded muscular section with an antibody for the V5 tag (green fluorescence) and with the endothelial marker isolectin-B4 (red fluorescence) shows that following infection of muscles with Ad.p75NTR (V-5 tagged), muscular capillary EC express the transgene (yellow fluorescence, from merging). Nuclei were stained in blue fluorescence by DAPI. (Original magnifications: 400x and 1000x).
by on May 22, 2011 circres.ahajournals.orgDownloaded from
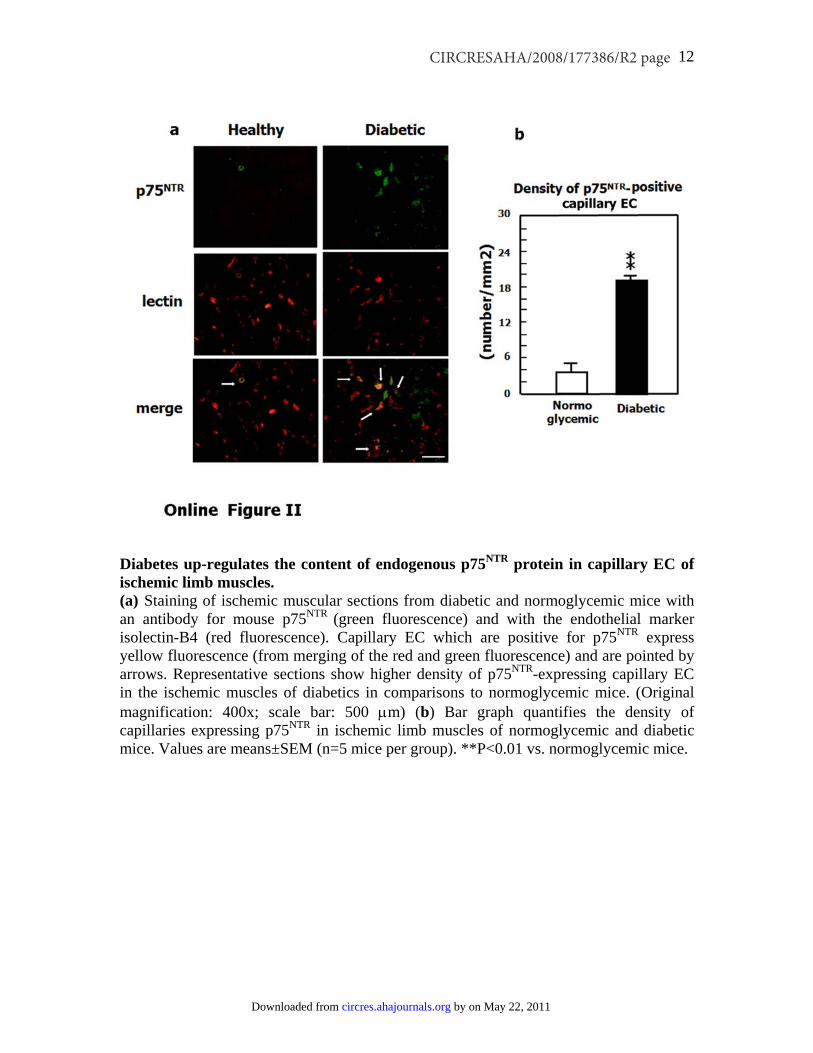
CIRCRESAHA/2008/177386/R2 page 12
Diabetes up-regulates the content of endogenous p75NTR protein in capillary EC of ischemic limb muscles. (a) Staining of ischemic muscular sections from diabetic and normoglycemic mice with an antibody for mouse p75NTR (green fluorescence) and with the endothelial marker isolectin-B4 (red fluorescence). Capillary EC which are positive for p75NTR express yellow fluorescence (from merging of the red and green fluorescence) and are pointed by arrows. Representative sections show higher density of p75NTR-expressing capillary EC in the ischemic muscles of diabetics in comparisons to normoglycemic mice. (Original magnification: 400x; scale bar: 500 μm) (b) Bar graph quantifies the density of capillaries expressing p75NTR in ischemic limb muscles of normoglycemic and diabetic mice. Values are means±SEM (n=5 mice per group). **P<0.01 vs. normoglycemic mice.
by on May 22, 2011 circres.ahajournals.orgDownloaded from
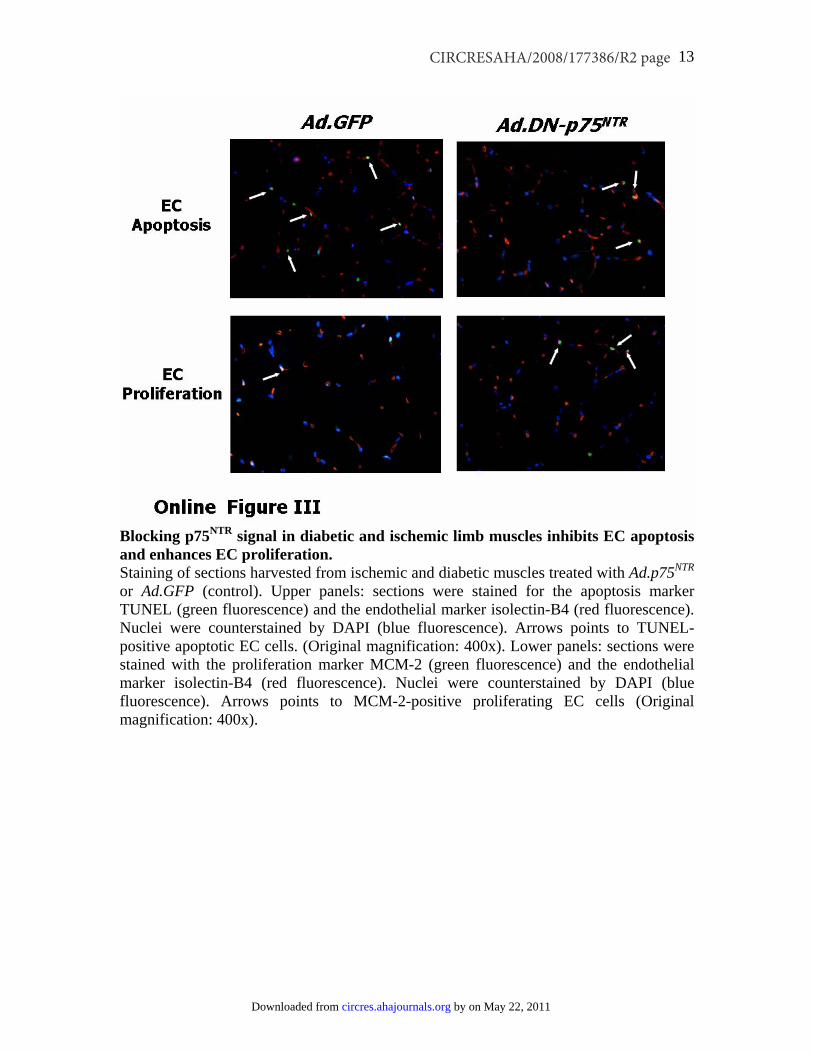
CIRCRESAHA/2008/177386/R2 page 13
Blocking p75NTR signal in diabetic and ischemic limb muscles inhibits EC apoptosis and enhances EC proliferation. Staining of sections harvested from ischemic and diabetic muscles treated with Ad.p75NTR or Ad.GFP (control). Upper panels: sections were stained for the apoptosis marker TUNEL (green fluorescence) and the endothelial marker isolectin-B4 (red fluorescence). Nuclei were counterstained by DAPI (blue fluorescence). Arrows points to TUNEL-positive apoptotic EC cells. (Original magnification: 400x). Lower panels: sections were stained with the proliferation marker MCM-2 (green fluorescence) and the endothelial marker isolectin-B4 (red fluorescence). Nuclei were counterstained by DAPI (blue fluorescence). Arrows points to MCM-2-positive proliferating EC cells (Original magnification: 400x).
by on May 22, 2011 circres.ahajournals.orgDownloaded from




















