Knock‐out mouse for Canavan disease: a model for gene transfer to the central nervous system
Upload
independentCategory
view
0download
0
mltetaccmadnwntbttsaaorrvtl
tn
pmti
aa
Experimental Cell Research 251, 213–223 (1999)Article ID excr.1999.4569, available online at http://www.idealibrary.com on
Nestin Is Expressed during Development and in Myotendinous andNeuromuscular Junctions in Wild Type and Desmin Knock-Out Mice
L. Carlsson,*,† Z. Li,‡ D. Paulin,‡ and L-E. Thornell*,†,1
*Department of Integrative Medical Biology, Section for Anatomy, and †Department of Musculoskeletal Research, National Institute of
Working Life, Umeå University, SE-901 87 Umeå, Sweden; and ‡Universite Paris VII, 2 Place Jussieu, 75005 Paris, FranceDw4fiotpndstl
taeosfnwtnmtm
obdtmstwm
rssap
Desmin, the main component of intermediate fila-ents (IFs) in mature skeletal muscle, forms an inter-
inking scaffold around myofibrils with connections tohe sarcolemma and the nuclear membrane. Desmin isnriched in neuromuscular and myotendinous junc-ions. Mice lacking the desmin gene develop normallynd reproduce. However, postnatally they develop aardiomyopathy and a dystrophy in highly used mus-les. We have investigated whether and how neuro-uscular and myotendinous junctions are affected
nd whether nestin compensates for the lack ofesmin in the knock-out (K/O) mice. We show thateither neuromuscular nor myotendinous junctionsere markedly affected in the desmin K/O mice. Ineuromuscular junctions nestin was present betweenhe postjunctional folds and the subneural nuclei andetween the nucleus and the myofibrillar cytoskele-on. In myotendinous junctions nestin was present be-ween myofibrils at the Z-disc level and in longitudinaltrands close to and at the junction. Nestin expressiont these specialized sites, as well as during myogenesisnd myofibrillogenesis, is independent of the presencef desmin. In desmin K/O mice nestin was also found inegenerating myofibers. The presence of nestin at neu-omuscular and myotendinous junctions might pro-ide enough strength for preservation and organiza-ion of the junctional areas, although desmin isacking. © 1999 Academic Press
Key Words: desmin knock-out mice; cytoskeleton; in-ermediate filaments; immunocytochemistry; immu-oelectron microscopy.
INTRODUCTION
The skeletal muscle fiber cytoskeleton is mainly com-osed of intermediate filaments (IFs), which surroundyofibrils at the Z-disc level and links them laterally to
he sarcolemma [for reviews see Refs. 1 and 2]. Desmins the major IF protein in adult skeletal muscle fibers.
1 To whom correspondence and reprint requests should beddressed. Fax: 146 90 7865480. E-mail: lars-eric.thornell@
onatomy.umu.se.
213
esmin is also present at neuromuscular junctions,here they are related to the postsynaptic foldings [3,], and in myotendinous junctions, at the sites of myo-bril-membrane attachment [5, 6]. In fact the stainingf desmin at the neuromuscular junction is so stronghat it has been used as a marker to distinguish end-lates in human muscle biopsies [4]. In the myotendi-ous junctions desmin intermediate filaments are co-istributed with the actin filaments and have beenuggested to act in parallel with these as separate forceransmitters across the cell membrane to the extracel-ular matrix of the tendon [5].
Vimentin, the IF protein typical of mesenchymalissues [7], is expressed in myotubes and in primarynd secondary generation myofibers during myogen-sis [8, 9]. In mice and human skeletal muscle devel-pment, vimentin becomes downregulated during latertages of myogenic differentiation, when myotubes areormed [8, 9]. Nestin, a protein originally identified ineuroepithelial stem cells [10], is another IF protein,hich is transiently expressed with desmin and vimen-
in during myogenesis [11–14]. Nestin becomes post-atally downregulated in rat [11] and is absent inature human skeletal muscles [15]. In human skele-
al muscle both nestin and vimentin are present inuscle fibers in the early stages of regeneration [15].In mice lacking desmin, due to molecular disruption
f the desmin gene, no defects in myogenesis or myofi-rillogenesis were seen; however, postnatally theesmin K/O mice showed a phenotype with defects inhe heart, in weight-bearing or highly used skeletaluscles, and in smooth muscle cells [16–19]. In the
oleus muscle, an important weight-bearing muscle inhe calf, muscle fiber degeneration occurred in parallelith muscle fiber regeneration and fibrosis as in auscular dystrophy [18].In the present study we have investigated the neu-
omuscular and the myotendinous junctions in limbkeletal muscles of wild type and desmin K/O mice. Weought to discover if these specialized regions wereffected by the lack of desmin. Furthermore, as theresence of desmin is not necessary for the alignment
f myofibrils during myogenesis and myofibrillogen-0014-4827/99 $30.00Copyright © 1999 by Academic Press
All rights of reproduction in any form reserved.

etndbmm
co
fieaa
o
ansoswm
214 CARLSSON ET AL.
sis, we wanted to know if nestin might be involved inhe alignment of myofibrils. Vimentin has been shownot to be upregulated and thus cannot compensate foresmin [17, 18]. How it is with nestin has so far noteen thoroughly investigated, though in one study it isentioned that nestin was not observed in skeletaluscles of adult desmin K/O mice [17].We here report that nestin is present in neuromus-
ular and myotendinous junctions in adult limb muscle
FIG. 1. Serial transverse (A–F) sections of skeletal muscles in and E) and wild-type newborn mice (C and F), peroxidase-stained wiestin is present in all myofibers, whereas strong staining is prearcolemma (short arrows) (A–C). No staining of desmin was seen inbserved in myofibers of wild-type mice (E and F). Note also the enrictrong staining of nestin was present (F). Serial longitudinal sectionith antinestin (G), antidesmin (H), and antimyomesin (I). Note theyotendinous junctions (arrows) (G–I). Bar, 25 mm.
f both wild-type and desmin K/O mice and in muscle d
bers during skeletal muscle development and differ-ntiation in both desmin K/O and wild-type mice. Indult desmin K/O mice nestin was present in regener-ting fibers.
MATERIAL AND METHODS
Material. Wild-type and desmin K/O mice in at least five of eachf the following age groups were examined: fetuses at 16- and 18-
min K/O mouse at g.d. 16 (A and D), and in wild-type at g.d. 18 (Bntibodies against nestin (A–C) and desmin (D–F). Weak staining oft in secondary myotubes (arrows) and at localized areas at the
scles of desmin K/O mice (D), whereas strong staining of desmin wasd staining of desmin (short arrows) in the same areas as in C, wheref muscle fibers from a wild-type mice at g.d. 18, peroxidase-stainedfferent staining patterns and the enriched amount of nestin in the
desth asen
muhes odi
ays of gestation, newborn, 5 days, 11 days, 14 days, 4 weeks, 10

wfiDdmt
amfcm5phbPssR(s(v(wddcc(lJnmwfi
btddvebI(UAusFiwo
tp1ra2iPfai
am(ttow
(1vS(pCctloZ
215NESTIN IN DESMIN KNOCK-OUT MICE
eeks, and 20 weeks of age. The phenotype of the mice were identi-ed by polymerase chain reaction (PCR) and Southern blot using tailNA [for details on the creation of the desmin K/O mice, see [16]. Theiaphragm muscle and the triceps surae muscle, including the soleususcle, were excised and processed for histochemistry, immunocy-
ochemistry, and conventional and immunogold electron microscopy.Tissue processing and sectioning. Specimens for histochemistry
nd immunocytochemistry were allowed to relax for a few minutes,ounted in Tissue Tek OCT Compound (Miles Inc., Elkhart, IL), and
rozen in propane chilled with liquid nitrogen. Frozen samples wereut at 225°C using a Reichert Jung cryostat (Leica, Nussloch, Ger-any) in 8-mm-thick sections for histochemical staining and 2- to
-mm-thick sections for immunocytochemistry. Other muscle sam-les were fixed for 2 h either in freshly prepared 2% paraformalde-yde (PFA) with 0.01% glutaraldehyde (GA) in 0.1 M phosphate-uffered saline (PBS) buffer or in 2.5% GA in 0.1 M PBS buffer. TheFA-fixed samples were cut into 1 mm cubes, infiltrated in 2.3 Mucrose, and frozen in liquid nitrogen. The specimens were cut intoemithin (0.2 mm) and ultrathin (70–100 nm) sections, using aeichert Ultracut microtome equipped with a FCS cryo attachment
Leica, Nussloch, Germany). The semithin sections were collected onlides and stained with antibodies for indirect immunofluorescencesee below), whereas the ultrathin sections were transferred to form-ar-coated carbon grids and stained for immunoelectron microscopysee below). GA-fixed material for conventional electron microscopyere postfixed for 1 h at 4°C in 2% osmium tetroxide, acetoneehydrated, and embedded in polybed 812 resin (Analytical Stan-ards, Kungsbacka, Sweden). Semithin (1 mm) plastic sections wereut and stained with toluidine blue. After identification of neuromus-ular and myotendinous junctions in the light microscope, ultrathin70-nm-thick) sections were cut and stained with uranyl acetate andead citrate on a LKB 2168 ultrastainer. Sections were observed in aeol 1200 EX-II electron microscope (Jeol, Tokyo, Japan). At least 40euromuscular junctions of diaphragm and soleus muscles and 20yotendinous junctions of the soleus muscle from desmin K/O andild-type mice were examined by electron microscopy. The musclebers examined showed a seemingly normal myofibrillar pattern.Antibodies and ligands. A polyclonal antibody Nestin 130 has
een produced in rabbits with a rat nestin polypeptide that con-ained the carboxyterminal tail region, but not the conserved rodomain [20]. This antibody has previously been shown to specificallyetect nestin by Western blots, without cross-reactivity to eitherimentin or desmin [11]. Nestin 130 has been used to study nestinxpression in mice, rat, and human muscles [11, 15, 14]. The anti-ody was kindly provided by T. Sejersen and U. Lendahl (Karolinskanstitute, Stockholm, Sweden). The monoclonal antibody Rat-401Developmental Studies Hybridoma Bank, Department of Biology,niversity of Iowa, Iowa City, IA) against rat nestin was also tested.monoclonal antibody Desmin D33 (Dako, Glostrup, Denmark) was
sed to stain desmin and a monoclonal antibody B4 against myome-in [21] was used as a marker for the myofibrillar M-band [22].luorescein (FITC)-conjugated a-bungarotoxin (Sigma) was used to
dentify neuromuscular junctions. All antibodies for light microscopyere diluted in 0.01 M PBS containing 0.1% BSA and used at theirptimal dilution.Histochemical and immunocytochemical stainings. Serial sec-
ions were stained to demonstrate myofibrillar adenosine triphos-hate (ATPase) activity after preincubation at pH 4.3, 4.6, 9.4, and0.3 [23] and with nicotinamide adenine dinucleotide tetrazoliumeductase (NADH-TR) to evaluate mitochondrial oxidative enzymectivity. For immunocytochemistry, sections were acetone fixed at20°C for 10 min and air dried. After treatment in 1% Triton X-100
n PBS for 15 min, the sections were washed three times in 0.01 MBS. Sections were immersed in 5% normal serum (Dako), obtained
rom the same species as the host animal for the secondary antibody,nd incubated in the primary antibody for 60 min at 37°C. Visual-
zation of antibodies was performed with either indirect peroxidase- anti-peroxidase (PAP) technique [24] or with indirect fluorescenceethod, using either FITC- or Cy 3-conjugated secondary antibodies
Dako, Glostrup, Denmark and Jackson Immuno Research Labora-ories, PA). Control sections were treated as described except thathe primary antibody was omitted or replaced by nonimmune serumr an unrelated primary antibody. Normal litter mates (Des1/1)ere used as positive controls. All sections were photographed with
FIG. 2. Longitudinal semithin cryosections of soleus musclesA–C). 5-day-old control mouse muscle stained with antinestin (A).1-day-old desmin K/O mouse muscle double-stained with antinestinisualized by Cy 3 (B) and antimyomesin visualized by FITC (C).taining of nestin is present at the sarcolemma, around a nucleus
n), and in myofibrils in a striated transverse, partly punctatedattern at the Z-disc level and in some longtudinal strands (A and B).apillaries (short arrows) (B). The two myofibers in the upper leftorner in B is shown in (C). The transverse striations correspond tohe myofibrillar M-bands, whereas the green staining at the sarco-emma is an unspecific reaction. The orange color is bleeding throughf the Cy 3 staining, confirming the localization of nestin at the-disc level. Bars, 10 mm.
Zeiss Axiophot microscope (Carl Zeiss, Obercochen, Germany)

wacmmc
216 CARLSSON ET AL.
FIG. 3. Serial transverse sections of soleus muscles in 10-week-old wild-type (A, C, and E) and desmin K/O mice (B, D, and F) stainedith ATPase preincubated at pH 4.6 (A and B), NADH-TR (C and D), and antinestin (E and F). In the control mouse, the muscle fibers showedcheckerboard pattern of fibertypes (A and C) and capillaries, some running in the plane of the section (short arrows) and others being
ross-sectioned (arrows), were stained with antinestin (E). In the desmin K/O mouse the soleus muscle showed a myopathic pattern withainly slow type I (l) fibers and large variations in fiber size (B). The pattern of mitochondrial enzyme activity in the desmin K/O fibers isore irregular (D). Some myofibers were stained with antinestin (short arrows), in others staining was present at the sarcolemma (arrows);
apillaries (thin arrows) (F). Bar, 50 mm.

e4iSsl3Daadwudb
c(iBrwaedmac
ttRe
E
lfsstwomwmws1wdKwNdn
jdaitcbpption. Bar, 20 mm.
217NESTIN IN DESMIN KNOCK-OUT MICE
quipped with epifluorescence using either an ordinary camera with00ASA Ektachrome film or a 3CCD camera (Dage-MTI, Inc., Mich-gan City), connected to Adobe Photoshop (Adobe Systems, USA).emithin cryosections were stained for immunofluorescence as de-cribed, except excluding the acetone fixation and air-drying. Toocalize acetylcholine receptors (AChRs), an incubation for 15 min at7°C with FITC-conjugated a-bungarotoxin (Sigma) was performed.ouble labeling [25] was then achieved by a second incubation withnti-nestin, followed by a Cy 3-conjugated goat-anti rabbit secondaryntibody. To reveal neuromuscular junctions in wild-type mice, aouble labeling with either a-bungarotoxin or antidesmin togetherith antinestin followed by appropriate secondary antibodies wassed. To determine the localization of nestin within myofibers aouble labeling with antinestin and antimyomesin (B4), visualizedy Cy 3 and FITC, respectively, was performed.Immunogold labeling. Sections on grids were washed in PBS
ontaining 0.5% BSA and 0.15% glycine, immersed in 5% goat serumDako, Glostrup, Denmark) to block nonspecific staining, and thenncubated with primary antibodies at room temperature for 60 min.ound antibodies were visualized by a gold labeled (5 nm) goat-anti-abbit secondary antibody (British Bio Cell, Cardiff, UK). Sectionsere postfixed in 2.5% GA and counterstained with 2% uranyl ox-late and 4% uranyl acetate (Merck, Darmstadt, Germany). Aftermbedding the sections in 1.15% methyl cellulose (Fluka) and airrying, the sections were photographed in a Jeol 1200 EX-II electronicroscope (Jeol, Tokyo, Japan). In control sections the primary
ntibody was replaced by the antibody dilution buffer (0.01 M PBSontaining 0.5% BSA and 0.15% glycine).
RESULTS
Our present study is based on results obtained withhe polyclonal antibody rabbit-anti-nestin 130. Sec-ions stained with the monoclonal antibody nestin-401 showed no reaction in mature muscle fibers inither wild-type or desmin K/O mice.
xpression of Nestin During Developmentof Skeletal Myofibers
In cross-sectioned muscles, antibodies against nestinightly stained the primary myotubes (larger diameter) inetuses of both desmin K/O and wild-type mice, whereasecondary myotubes (smaller diameter) were moretrongly stained (Figs. 1A and 1B). The secondary myo-ubes were few at gestation day (g.d.) 16 (Fig. 1A), butere more abundant at g.d. 18 (Fig. 1B). In cross-sectionsf muscles in newborn wild-type and desmin knock-outice, the myofibers showed a more mature appearance,ith reduced intercellular space (Figs. 1C and 1F). Theyofibers at this stage of development stained lightlyith antibodies against nestin; however, some showed
trong staining in localized areas at the sarcolemma (Fig.C). For comparison adjacent sections to those stainedith antinestin were stained with antibodies againstesmin. No staining of desmin was seen in the desmin/O mice (Fig. 1D), whereas strong staining of desminas observed in the wild-type mice (Figs. 1E and 1F).ote also the enriched subsarcolemmal staining ofesmin in the same areas as those with high amounts of
FIG. 4. A longitudinal semithin cryosection of a myotendinousunction in a soleus muscle of a 4-week-old wild-type mouseouble-stained with antinestin visualized by FITC (A) andntidesmin visualized by Cy 3 (B) and FITC and Cy 3 super-mposed (C). Nestin is present in between the myofibrils athe level of the Z-discs and in longitudinal strands (arrows)lose to and at the myotendinous junction. This interpretation isased on the colocalization of nestin with desmin, the stainingattern of which is well known (37, 32, 38). Note that nestin isresent only in the myofibrils closest to the myotendinous junc-
estin in Fig. 1C (Fig. 1F).

cwlamsaa
P
b1nttcZpacaaM
N
desv(
n(mdi(aswKdfi
N
amllT
jsattistr100 nm.
218 CARLSSON ET AL.
In longitudinally sectioned muscles mainly a diffuseytoplasmic staining was seen at g.d.16 (not shown),hereas at g.d.18 a transverse striated pattern at the
evel of the Z-discs was apparent (Fig. 1G). Enrichedmount of nestin was seen at the sarcolemma and inyotendinous junctions (Fig. 1G). Adjacent sections
tained with antibodies against desmin and myomesinlso showed transverse staining patterns (Figs. 1Hnd 1I).
ostnatal Distribution of Nestin in Mice
Nestin was detected in some of the muscle fibers inoth the wild-type and the desmin K/O mice at 5 and1 days (Figs. 2A and 2B). The presence of nestin didot correlate to a particular muscle or muscle fiberype. In longitudinal sections we observed a striatedransverse staining pattern, which at higher magnifi-ation was seen as dots in between the myofibrils at the-disc level. Longitudinal strands of staining wereresent as well, especially where irregularities in thelignment of the myofibrils were seen, and at the sar-olemma (Fig. 2A). Double labeling with antinestinnd antimyomesin confirmed that nestin was localizedt the Z-discs, whereas myomesin stained the-bands, as expected [21, 22] (Figs. 2B and 2C).
estin in Adult Mice
Muscles in general in the desmin K/O mice do notiffer markedly from those in the wild-type mice. How-ver, highly used muscles, such as the soleus muscle,how distinct differences with respect to fiber types,ariations in fiber size, and mitochondrial organizationfor comparison, see Figs. 3A and 3B; 3C and 3D).
In both controls and desmin K/O mice capillaries anderves were stained with the antibody against nestinFigs. 3E and 3F). In muscle fibers of adult wild-type
ice, staining of nestin was only seen in the myoten-inous and neuromuscular junctions. These special-zed regions were also stained in the desmin K/O micesee further below). In the desmin K/O mice nestin waslso seen in some fibers of varying size (Fig. 3F). Thistaining was most prominent in the soleus muscle,hich is one of the most affected muscles in the desmin/O mice. Typical features of this muscle are signs ofegeneration and regeneration of muscle fibers andbrosis [17, 18] (Figs. 3B, 3D, and 3F).
estin in Myotendinous and Neuromuscular Junctions
At the myotendinous junctions, the muscle fibers of allge groups in both wild-type (Fig. 4A) and desmin K/Oice (Fig. 5A) were strongly stained with antinestin. In
ongitudinal sections the labeling was seen at the sarco-emma and in parts of the myofibrils facing the tendon.
FIG. 5. Longitudinal semithin section of a myotendinousunction in a soleus muscle of a 10-week-old desmin K/O mousetained with antinestin and FITC. Staining of nestin is seens longitudinal strands (arrows) at the sarcolemma facing theendon and as dots in between the myofibrils (A). Immunoelec-ron microscopy of ultrathin frozen sections of soleus musclesn 10-week-old (B) and 5-month-old (C) desmin K/O micetained with antinestin. Gold particles are seen in betweenwo myofibrils at the Z-disc level (B) and in the subsarcolemmalegion. Sarcolemma (small arrows) (C). Bars: A, 20 mm, B and C,
he staining was seen as strands and as dots in between
tbwwlbmprmsldtam
ndjtcwugt(ttbicp
(m ch
219NESTIN IN DESMIN KNOCK-OUT MICE
he myofibrils at the Z-disc level. Nestin was observed toe colocalized with desmin in sections double labeledith antinestin and antidesmin in skeletal muscles ofild-type mice (Figs. 4A–4C). Some variations in the
evel of nestin in different myotendons were observed inoth wild-type and desmin K/O mice. Immunoelectronicroscopy of longitudinally sectioned muscles showed
resence of nestin in between myofibrils at the Z-discegion (Fig. 5B) and at the sarcolemma (Fig. 5C). Trans-ission electron microscopy of myotendinous junctions
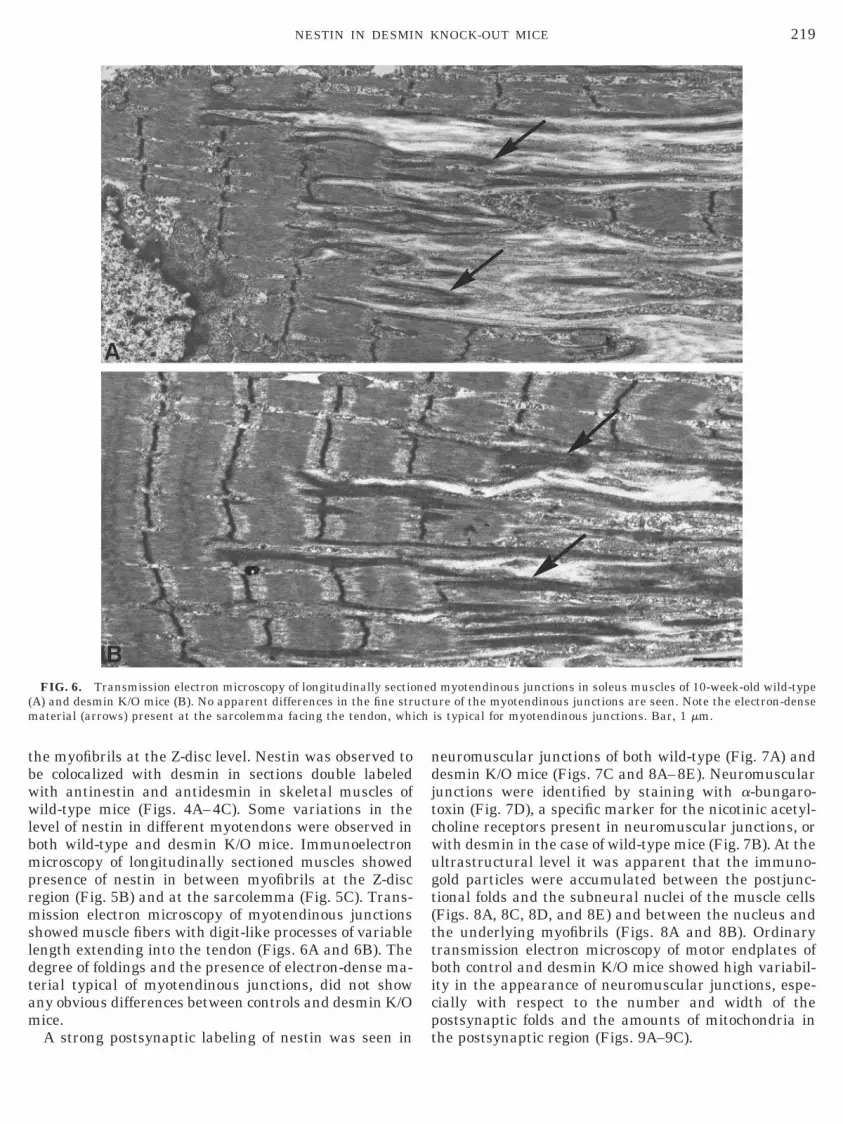
howed muscle fibers with digit-like processes of variableength extending into the tendon (Figs. 6A and 6B). Theegree of foldings and the presence of electron-dense ma-erial typical of myotendinous junctions, did not showny obvious differences between controls and desmin K/Oice.
FIG. 6. Transmission electron microscopy of longitudinally sectioA) and desmin K/O mice (B). No apparent differences in the fine straterial (arrows) present at the sarcolemma facing the tendon, whi
A strong postsynaptic labeling of nestin was seen in t
euromuscular junctions of both wild-type (Fig. 7A) andesmin K/O mice (Figs. 7C and 8A–8E). Neuromuscularunctions were identified by staining with a-bungaro-oxin (Fig. 7D), a specific marker for the nicotinic acetyl-holine receptors present in neuromuscular junctions, orith desmin in the case of wild-type mice (Fig. 7B). At theltrastructural level it was apparent that the immuno-old particles were accumulated between the postjunc-ional folds and the subneural nuclei of the muscle cellsFigs. 8A, 8C, 8D, and 8E) and between the nucleus andhe underlying myofibrils (Figs. 8A and 8B). Ordinaryransmission electron microscopy of motor endplates ofoth control and desmin K/O mice showed high variabil-ty in the appearance of neuromuscular junctions, espe-ially with respect to the number and width of theostsynaptic folds and the amounts of mitochondria in
myotendinous junctions in soleus muscles of 10-week-old wild-typere of the myotendinous junctions are seen. Note the electron-dense
is typical for myotendinous junctions. Bar, 1 mm.
neductu
he postsynaptic region (Figs. 9A–9C).

cmiio1t1mal
lognmutpjaori
fosipktit3cemsimllanahomNwacl
iejseippgaam
itabAcw
ttwb
ltsw(rCWt
220 CARLSSON ET AL.
DISCUSSION
Nestin is here shown to be selectively present in spe-ialized regions of the muscle fibers in adult mice; inotor endplates and in myotendinous junctions. Nestin
s present in these areas irrespective of whether desmins expressed or not. The results in the present study werebtained with the polyclonal antibody rabbit-anti-nestin30, which previously has been characterized and showno selectively identify nestin in rat, human, and mice [11,5, 14]. On the other hand, no staining was seen with theonoclonal antibody nestin R-401. This explains the neg-
tive results reported in another study, in which theatter antibody was used [17].
In the neuromuscular junctions, nestin was accumu-ated in the postsynaptic region from the lower portionsf the junctional folds to the subneural nuclei. This sug-ests that nestin might participate in anchoring the sub-eural nuclei to the endplate region. Such a cytoskeletonight be related to a mechanism, which selectively reg-lates these nuclei to provide mRNA for the synthesis ofhe unique proteins localized at the endplates [26]. It hasreviously been suggested that IFs in neuromuscular
unctions form a podoskeleton, which participates in thessembly and maintenance of the postsynaptic folds andf the subneural organelles [27]. In freeze-etched frog andat motor endplates a meshwork of cytoskeletal filaments
FIG. 7. Serial transverse sections (A and B) of the neuromuscu-ar junction area in a 4-week-old wild-type mouse stained with an-inestin and FITC (A) and with antidesmin and Cy3 (B). Transverseection (C and D) of a 4-week-old desmin K/O mouse double-stainedith antinestin and Cy 3 (C) and a-bungarotoxin coupled to FITC
D). Neuromuscular junctions are marked with arrows. Strong fluo-escence of nestin is present at the neuromuscular junctions (A and), where it is colocalized with desmin (B) and a-bungarotoxin (D).eak staining of nestin is seen in some myofibers (*) at the junc-
ional area (C). Bar, 25 mm.
ncluding actin filaments, IFs, and microtubules were r
ound in the postsynaptic processes [28, 29]. The IFs werebserved in parallel to the synaptic folds in the core of theynaptic protrusions [28]. To our knowledge no furthernformation is available on the organization of IFs and IFroteins at the neuromuscular junctions. In contrast thenowledge on dystrophin and associated proteins andheir association to the sarcolemma and the proteinsnvolved in the clustering of the acetylcholine receptors athe motor endplates is very extensive [for reviews, see 27,0, 31]. In skeletal muscles it has in fact been very diffi-ult to visualize IFs defined as filaments having a diam-ter of approximately 10 nm [32]. In the present study aeshwork of filamentous strands were seen in the cryo-
ections as well as in the ordinary plastic sections. Sim-larly with conventional electron microscopy of human
yopathies it was not possible to identify IF, althougharge foci of desmin-positive areas were observed withight microscopy [33, 34]. This might suggest that desminnd other IF proteins in skeletal muscle might be orga-ized in another form and/or to be sensitive to fixationnd other preparatory steps for electron microscopy. Weave recently reported observations of filamentous linksf unknown components between myofibrils and betweenyofibrils and the sarcolemma in desmin K/O mice [18].estin has been reported to be able to form filamentsith desmin and vimentin but not with itself [35]. Thisrgues in favor of nestin polymerizing with additionalytoskeletal proteins, as both desmin and vimentin areacking in skeletal muscles of adult desmin K/O mice [18].
The presence of endplates with well preserved fold-ngs in desmin K/O mice suggests that desmin is notssential for the organization of the neuromuscularunctions, although some functional impairment ofkeletal muscles in the desmin K/O mice still are un-xplained. A tentative suggestion is that nestin bytself or in addition to other cytoskeletal proteinsresent at the endplates gives enough strength forreservation and organization of the postsynaptic re-ion. The myofibrillar presence of nestin in limitedreas of the junction might also be of importance as andditional link between the subneural domain and theyofibrillar cytoskeleton.Further investigations are obviously needed to clar-
fy the role of the IF proteins at neuromuscular junc-ions and in particular their relationship to the largerray of proteins associated with the plasma mem-rane and the basal lamina [for reviews, see 27, 30, 31].double knock-out of desmin and nestin would help to
larify the role of nestin in neuromuscular junctions, asell as in myotendinous junctions (see below).At myotendinous junctions the contraction force is
ransmitted from the muscle fibers to the tendon andhe myofibrils anchor to the sarcolemma in a similaray as in a fascia adherens [2]. Defects in the linketween the exterior and the muscle cytoskeleton give
ise to muscle fiber degeneration [36]. In our study on
wacc
mpt
d(sAr wn
221NESTIN IN DESMIN KNOCK-OUT MICE
ild-type mice, desmin was observed in highermounts in myotendinous junctions and showed colo-alization with nestin. According to previous immuno-
FIG. 8. Immunoelectron microscopy of ultrathin frozen sectionsesmin K/O mouse stained with antinestin and 5-nm gold particles (nt), postjunctional folds (pf), and a subneural nucleus (n) (A). Highereen in between the junctional folds and the subneural nucleus (C) a
nerve terminal (nt) with postsynaptic folds (pf). Gold particles (shegion close to the sarcolemma (D). The area to the right in D is sho
ytochemical studies desmin is also enriched in frog f
yotendinous junctions and localized in the core of therocesses but not immediately subjacent to the junc-ional membrane [5]. As no major ultrastructural dif-
neuromuscular junctions in the soleus muscle from a 5-month-old). Overview of a neuromuscular junction showing a nerve terminalgnification of the two boxed areas in A (B and C). Gold particles arein the area between the nucleus and the underlying myofibrils (B).arrows) are seen in the muscle part of the folds outside the densein higher magnification (E). Bars: A, 500 nm; B–E, 100 nm.
ofA–Emandort
erences of the myotendinous junctions were here ob-

sctOmjmose
ndhaiftpwmaobpaa[p
mw(utP
jTaamount of postsynaptic folds (arrows). Bar, 500 nm.
222 CARLSSON ET AL.
erved between wild-type and desmin K/O mice, weonclude that desmin is not of crucial importance forhe stability of the myotendinous junctions in mice.ur results showing expression of nestin in the outer-ost part of the myofibers facing the myotendinous
unctions and at the sarcolemma facing the tendonight indicate that nestin by itself or together with
ther cytoskeletal proteins are enough to provide atrong link between the muscle fiber and the tendonven if desmin is lacking.We report here the same presence and localization of
estin during development in both wild-type andesmin-deficient mice. This might suggest that nestinas some function during myofibrillar differentiation,lthough it is not an obligatory component. Previouslyt has been shown that nestin was unnecessary forusion and terminal differentiation in myoblasts of cer-ain cell lines in vitro [12]. That expression of nestin isart of the developmental program for muscle fibersas confirmed by our observation that regeneratingyofibers in adult skeletal muscles of desmin K/O mice
lso expressed nestin. This is also in accordance withbservations on nestin expression in regenerating fi-ers in degenerative human muscle disorders [15]. In arevious ultrastructural study of the soleus muscle indult desmin K/O mice, we observed aberrant regener-tion with myotubes having disorganized myofibrils18]. Thus nestin, although present, is not sufficient forroper myofibrillar assembly during regeneration.
We thank Dr. Maureen Price, Houston, for many helpful com-ents and linguistic improvements on the manuscript. This workas supported by grants from the Swedish Medical Research Council
12X-03934), the Magnus Bergvall Foundation and the Medical Fac-lty of Umeå University to L-E. Thornell, and grants from Associa-ion Francaise contre les Myopathies and Fondation de France to D.aulin.
REFERENCES
1. Price, M. G. (1991). Striated muscle endosarcomeric and exo-sarcomeric lattices. In “Advances in Structural Biology,” pp.175–207.
2. Small, J. V., Furst, D. O., and Thornell, L.-E. (1992). Thecytoskeletal lattice of muscle cells. Eur. J. Biochem. 208, 559–572.
3. Sealock, R., Murnane, A. A., Paulin, D., and Froehner, S. C.(1989). Immunochemical identification of desmin in Torpedopostsynaptic membranes and at the rat neuromuscular junc-tion. Synapse 3, 315–324.
4. Askanas, V., Bornemann, A., and Engel, W. K. (1990). Immu-nocytochemical localization of desmin at human neuromuscularjunctions. Neurology 40, 949–953.
5. Tidball, J. G. (1992). Desmin at myotendinous junctions. Exp.Cell Res. 199, 206–212.
6. Cary, R. B., and Klymkowsky, M. W. (1995). Disruption ofintermediate filament organization leads to structural defectsat the intersomite junction in Xenopus myotomal muscle. De-
FIG. 9. Transmission electron microscopy of neuromuscularunctions in muscle fibers of 5-month-old desmin K/O mice (A–C).he endplate regions are highly variable with respect to the numbernd size of mitochondria (m) in the nerve terminals (nt) and the
velopment 121, 1041–1052.

1
1
1
1
1
1
1
1
1
1
2
2
2
2
2
2
2
2
2
2
3
3
3
3
3
3
3
3
3
RR
223NESTIN IN DESMIN KNOCK-OUT MICE
7. Franke, W. W., Schmid, E., Osborne, M., and Weber, K. (1978).Different intermediate-sized filaments distinguished by immu-nofluorescence microscopy. Proc. Natl. Acad. Sci. USA 10,5034–5038.
8. Furst, D. O., Osborn, M., and Weber, K. (1989). Myogenesis inthe mouse embryo: Differential onset of expression of myogenicproteins and the involvement of titin in myofibril assembly.J. Cell Biol. 109, 517–527.
9. Barbet, J. P., Thornell, L.-E., and Butler-Browne, G. S. (1991).Immunocytochemical characterisation of two generations of fi-bers during the development of the human quadriceps muscle.Mech. Dev. 35, 3–11.
0. Hockfield, S., and McKay, R. D. (1985). Identification of majorcell classes in the developing mammalian nervous system.J. Neurosci. 5, 3310–3328.
1. Sejersen, T., and Lendahl, U. (1993). Transient expression ofthe intermediate filament nestin during skeletal muscle devel-opment. J. Cell Sci. 106, 1291–1300.
2. Kachinsky, A. M., Dominov, J. A., and Miller, J. B. (1994).Myogenesis and the intermediate filament protein, nestin. Dev.Biol. 165, 216–228.
3. Sjoberg, G., Jiang, W. Q., Ringertz, N. R., Lendahl, U., andSejersen, T. (1994). Colocalization of nestin and vimentin/desmin in skeletal muscle cells demonstrated by three-dimen-sional fluorescence digital imaging microscopy. Exp. Cell Res.214, 447–458.
4. Wroblewski, J., Engstrom, M., Edwall-Arvidsson, C., Sjoberg,G., Sejersen, T., and Lendahl, U. (1997). Distribution of nestinin the developing mouse limb bud in vivo and in micro-masscultures of cells isolated from limb buds. Differentiation 61,151–159.
5. Sjoberg, G., Edstrom, L., Lendahl, U., and Sejersen, T. (1994).Myofibers from Duchenne/Becker muscular dystrophy and my-ositis express the intermediate filament nestin. J. Neuropathol.Exp. Neurol. 53, 416–423.
6. Li, Z., Colucci-Guyon, E., Pincon-Raymond, M., Mericskay, M.,Pournin, S., Paulin, D., and Babinet, C. (1996). Cardiovascularlesions and skeletal myopathy in mice lacking desmin. Dev.Biol. 175, 362–366.
7. Milner, D. J., Weitzer, G., Tran, D., Bradley, A., and Capet-anaki, Y. (1996). Disruption of muscle architecture and myo-cardial degeneration in mice lacking desmin. J. Cell Biol. 134,1255–1270.
8. Li, Z., Mericskay, M., Agbulut, O., Butler-Browne, G., Carlsson,L., Thornell, L.-E., Babinet, C., and Paulin, D. (1997). Desmin isessential for the tensile strength and integrity of myofibrils butnot for myogenic commitment, differentiation, and fusion ofskeletal muscle. J. Cell Biol. 139, 129–144.
9. Thornell, L.-E., Carlsson, L., Li, Z., Mericskay, M., and Paulin,D. (1997). Null mutation in the desmin gene gives rise to acardiomyopathy. J. Mol. Cell Cardiol. 29, 2107–2124.
0. Dahlstrand, J., Collins, V. P., and Lendahl, U. (1992). Expres-sion of the class VI intermediate filament nestin in humancentral nervous system tumors. Cancer Res. 52, 5334–5341.
1. Grove, B. K., Kurer, V., Lehner, C., Doetschman, T. C., Perri-ard, J. C., and Eppenberger, H. M. (1984). A new 185.000-dalton skeletal muscle protein detected by monoclonal antibod-ies. J. Cell Biol. 98, 518–524.
2. Obermann, W. M., Gautel, M., Weber, K., and Furst, D. O.(1997). Molecular structure of the sarcomeric M band: Mapping
of titin and myosin binding domains in myomesin and theevised version received February 26, 1999
identification of a potential regulatory phosphorylation site inmyomesin. Embo J. 16, 211–220.
3. Dubowitz, V. (1985). In “Muscle Biology. A Practical Approach,”2nd ed., pp. 29–40, Bailliere Tindall, London.
4. Sternberger, L. A. (1979). The unlabeled antibody peroxidase-antiperoxidase (PAP) method. In “Immunohistochemistry,” pp.104–169, Wiley Medical, New York.
5. Beesley, J. E. (1993). Multiple immunolabelling techniques. In“Immunocytochemistry,” pp. 103–125, Oxford Univ. Press, NewYork.
6. Traub, P. (1995). Intermediate filaments and gene regulation.Physiol. Chem. Phys. Med. NMR 27, 377–400.
7. Carteaud, J., and Changeux, J.-P. (1992). Post-transcriptionalcompartmentalzation of acetylcholine receptor biosynthesis inthe subneural domain of muscle and electrocyte junctions. Eur.J. Neurosci. 5, 191–202.
8. Hirokawa N., and Heuser, J. E. (1982). Internal and externaldifferentiations of the postsynaptic membrane at the neuro-muscular junction. J. Neurocytol. 11, 487–510.
9. Yorifuji, H., and Hirokawa, N. (1989). Cytoskeletal architectureof neuromuscular junction: Localization of vinculin. J. ElectronMicrosc. Tech. 12, 160–171.
0. Ohlendieck, K. (1996). Towards an understanding of the dys-trophin-glycoprotein complex: Linkage between the extracellu-lar matrix and the membrane cytoskeleton in muscle fibers.Eur. J. Cell Biol. 69, 1–10.
1. Berthier, C., and Blaineau, S. (1997). Supramolecular organi-zation of the subsarcolemmal cytoskeleton of adult skeletalmuscle fibers. Biol. Cell. 89, 413–434.
2. Thornell, L.-E., Ericsson, A., and Edstom, L. (1983). Interme-diate filaments in human myopathies. In “Cell and MuscleMotility” (R. B. Dowben, and J. W. Shay, Eds.), pp. 85–135,Plenum, New York.
3. Cameron, C. H., Mirakhur, M., and Allen, I. V. (1995). Desminmyopathy with cardiomyopathy. Acta Neuropathol. 89, 560–566.
4. Nakano, S., Engel, A. G., Waclawik, A. J., Emslie-Smith, A. M.,and Busis, N. A. (1996). Myofibrillar myopathy with abnormalfoci of desmin positivity. I. Light and electron microscopy anal-ysis of 10 cases. J. Neuropathol. Exp. Neurol. 55, 549–562.
5. Sjoberg, G., Meng, J. J., Sejersen, T., and Wallace, I. (1997).Nestin is an obligate heterodimer in intermediate filament for-mation. In “Nestin: Filament Formation and Expression inMyogenesis and Muscle Disorders,” Thesis, Karolinska Insti-tute, Stockholm, Sweden.
6. Martin, P. T., Kaufman, S. J., Kramer, R. H., and Sanes, J. R.(1996). Synaptic integrins in developing, adult, and mutantmuscle: Selective association of alpha1, alpha7A, and alpha7Bintegrins with the neuromuscular junction. Dev. Biol. 174, 125–139.
7. Thornell, L.-E., Edstrom, L., Eriksson, A., Henriksson, K. G.,and Angqvist, K. A. (1980). The distribution of intermediatefilament protein (skeletin) in normal and diseased human skel-etal muscle—An immunohistochemical and electron- micro-scopic study. J. Neurol. Sci. 47, 153–170.
8. Thornell, L.-E., Eriksson, A., Johansson, B., Kjorell, U., Franke,W. W., Virtanen, I., and Lehto, V. P. (1985). Intermediatefilament and associated proteins in heart Purkinje fibers: Amembrane-myofibril anchored cytoskeletal system. Ann. N.Y.
Acad. Sci. 455, 213–240.eceived February 18, 1999
Copyright © 2022 FDOKUMEN