
Expression of R120G-aB-Crystallin Causes Aberrant Desmin and aB-Crystallin Aggregation and...
22
Lorenz, Timothy Hewett and Jeffrey Robbins Xuejun Wang, Hanna Osinska, Raisa Klevitsky, A. Martin Gerdes, Michelle Nieman, John Aggregation and Cardiomyopathy in Mice B-Crystallin α B-Crystallin Causes Aberrant Desmin and α - Expression of R120G Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2001 American Heart Association, Inc. All rights reserved. is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231 Circulation Research doi: 10.1161/hh1301.092688 2001;89:84-91; originally published online June 21, 2001; Circ Res. http://circres.ahajournals.org/content/89/1/84 World Wide Web at: The online version of this article, along with updated information and services, is located on the http://circres.ahajournals.org/content/suppl/2001/06/12/hh1301.092688.DC1.html Data Supplement (unedited) at: http://circres.ahajournals.org//subscriptions/ is online at: Circulation Research Information about subscribing to Subscriptions: http://www.lww.com/reprints Information about reprints can be found online at: Reprints: document. Permissions and Rights Question and Answer about this process is available in the located, click Request Permissions in the middle column of the Web page under Services. Further information Editorial Office. Once the online version of the published article for which permission is being requested is can be obtained via RightsLink, a service of the Copyright Clearance Center, not the Circulation Research in Requests for permissions to reproduce figures, tables, or portions of articles originally published Permissions: by guest on August 6, 2014 http://circres.ahajournals.org/ Downloaded from by guest on August 6, 2014 http://circres.ahajournals.org/ Downloaded from by guest on August 6, 2014 http://circres.ahajournals.org/ Downloaded from by guest on August 6, 2014 http://circres.ahajournals.org/ Downloaded from by guest on August 6, 2014 http://circres.ahajournals.org/ Downloaded from by guest on August 6, 2014 http://circres.ahajournals.org/ Downloaded from by guest on August 6, 2014 http://circres.ahajournals.org/ Downloaded from by guest on August 6, 2014 http://circres.ahajournals.org/ Downloaded from by guest on August 6, 2014 http://circres.ahajournals.org/ Downloaded from by guest on August 6, 2014 http://circres.ahajournals.org/ Downloaded from by guest on August 6, 2014 http://circres.ahajournals.org/ Downloaded from by guest on August 6, 2014 http://circres.ahajournals.org/ Downloaded from by guest on August 6, 2014 http://circres.ahajournals.org/ Downloaded from by guest on August 6, 2014 http://circres.ahajournals.org/ Downloaded from by guest on August 6, 2014 http://circres.ahajournals.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Expression of R120G-aB-Crystallin Causes Aberrant Desmin and aB-Crystallin Aggregation and...

Lorenz, Timothy Hewett and Jeffrey RobbinsXuejun Wang, Hanna Osinska, Raisa Klevitsky, A. Martin Gerdes, Michelle Nieman, John
Aggregation and Cardiomyopathy in MiceB-CrystallinαB-Crystallin Causes Aberrant Desmin and α−Expression of R120G
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2001 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/hh1301.0926882001;89:84-91; originally published online June 21, 2001;Circ Res.
http://circres.ahajournals.org/content/89/1/84World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org/content/suppl/2001/06/12/hh1301.092688.DC1.htmlData Supplement (unedited) at:
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on August 6, 2014http://circres.ahajournals.org/Downloaded from by guest on August 6, 2014http://circres.ahajournals.org/Downloaded from by guest on August 6, 2014http://circres.ahajournals.org/Downloaded from by guest on August 6, 2014http://circres.ahajournals.org/Downloaded from by guest on August 6, 2014http://circres.ahajournals.org/Downloaded from by guest on August 6, 2014http://circres.ahajournals.org/Downloaded from by guest on August 6, 2014http://circres.ahajournals.org/Downloaded from by guest on August 6, 2014http://circres.ahajournals.org/Downloaded from by guest on August 6, 2014http://circres.ahajournals.org/Downloaded from by guest on August 6, 2014http://circres.ahajournals.org/Downloaded from by guest on August 6, 2014http://circres.ahajournals.org/Downloaded from by guest on August 6, 2014http://circres.ahajournals.org/Downloaded from by guest on August 6, 2014http://circres.ahajournals.org/Downloaded from by guest on August 6, 2014http://circres.ahajournals.org/Downloaded from by guest on August 6, 2014http://circres.ahajournals.org/Downloaded from

Expression of R120G–aB-Crystallin Causes AberrantDesmin andaB-Crystallin Aggregation and
Cardiomyopathy in MiceXuejun Wang, Hanna Osinska, Raisa Klevitsky, A. Martin Gerdes, Michelle Nieman, John Lorenz,
Timothy Hewett, Jeffrey Robbins
Abstract—Upregulation ofaB-crystallin (CryAB), a small heat shock protein, is associated with a variety of diseases,including the desmin-related myopathies. CryAB, which binds to both desmin and cytoplasmic actin, may participateas a chaperone in intermediate filament formation and maintenance, but the physiological consequences of CryABupregulation are unknown. A mutation in CryAB, R120G, has been linked to a familial desminopathy. However, it isunclear whether the mutation is directly causative. We created multiple transgenic mouse lines that overexpressed eithermurine wild-type CryAB or the R120G mutation in cardiomyocytes. Overexpression of wild-type CryAB was relativelybenign, with no increases in mortality and no induction of desmin-related cardiomyopathy even in a line in whichCryAB mRNA expression was increased'104-fold and the protein level increased by 11-fold. In contrast, linesexpressing the R120G mutation were compromised, with a high-expressing line exhibiting 100% mortality by earlyadulthood. Modest expression levels resulted in a phenotype that was strikingly similar to that observed for thedesmin-related cardiomyopathies. The desmin filaments in the cardiomyocytes were overtly affected, myofibrilalignment was significantly impaired, and a hypertrophic response occurred at both the molecular and cellular levels.The data show that the R120G mutation causes a desminopathy, is dominant negative, and results in cardiachypertrophy.(Circ Res. 2001;89:84-91.)
Key Words: transgenicn heart diseasen mousen cardiacn genetics
The small heat shock–related proteinaB-crystallin(CryAB) was originally discovered and classified as a
lens protein.1 CryAB is also found in nonlenticular tissuesand is abundant in cardiac and skeletal muscle.2,3 CryABbinds both desmin and cytoplasmic actin and possessesmolecular chaperone function in vitro.4–6 When a cell issubjected to stress, CryAB transits from the cytosol onto thecytoskeleton.7 Phosphorylation by mitogen-activated proteinkinase, p38, and other kinases may regulate this translocationand presumably its chaperone function.8,9 The upregulation ofthe gene and subsequent accumulation of CryAB occurs in anumber of cardiac disorders including familial hypertrophiccardiomyopathy and desminopathy,10–12 as well as degener-ative neural pathologies such as Alexander and Alzheimerdiseases.2,13 However, the pathophysiological significance, ifany, of CryAB protein upregulation in muscle remainsobscure.
A missense mutation (R120G) of CryAB has recently beenlinked to familial desmin-related myopathy (DRM), a diseasethat is characterized by intrasarcoplasmic accumulation of
desmin.14 Restrictive, hypertrophic, and dilated cardiomyopa-thies have all been observed in the desminopathies and oftenresult in death.12,15 Overexpression of R120G-CryAB in amuscle cell line caused formation of electron-dense aggregatescontaining CryAB in the center and desmin at the periphery.14
However, there is no direct in vivo evidence, outside of linkageanalysis, proving that the missense mutation of CryAB causesDRM. Furthermore, if the mutation is directly causative, itremains to be explored how it leads to disease presentation.
To approach these issues, we generated multiple stabletransgenic (TG) mouse lines that express different levels ofeither the wild-type (WT) or mutant CryAB proteins specif-ically in the heart. Whereas overexpression of WT CryABprotein was benign, expression of even very modest levels ofR120G-CryAB protein led to aberrant desmin and CryABaggregation, disruption of the desmin network, perturbationof myofibril alignment, and compromised muscle function.
Materials and MethodsAn expanded Materials and Methods section is available online athttp://www.circresaha.org.
Original received February 6, 2001; revision received May 8, 2001; accepted May 8, 2001.From the Division of Molecular Cardiovascular Biology (X.W., H.O., R.K., T.H., J.R.), Children’s Hospital Research Foundation, Cincinnati, Ohio;
South Dakota Health Research Foundation–Cardiovascular Research Institute (A.M.G.), Sioux Falls, SD; and Department of Molecular and CellularPhysiology (M.N., J.L.), University of Cincinnati Medical Center, Cincinnati, Ohio.
Correspondence to Jeffrey Robbins, Division of Molecular Cardiovascular Biology, Children’s Hospital Research Foundation, 3333 Burnet Ave,Cincinnati, OH 45229. E-mail [email protected]
© 2001 American Heart Association, Inc.
Circulation Researchis available at http://www.circresaha.org
84 by guest on August 6, 2014http://circres.ahajournals.org/Downloaded from

ResultsTG Mouse LinesBoth human genetic and in vitro biochemical data indicatethat the missense mutation (R120G) of CryAB acts in adominant-negative fashion.14,16 Therefore, we chose to use aTG approach in an attempt to show direct causality of themutation in causing cardiovascular disease and to create ananimal model suitable for longitudinal analyses of the patho-genic processes. To control for the possibility that alterationsin the overall stoichiometries of either the CryAB transcriptor protein pools might lead to a phenotype, WT murineCryAB was also used to generate TG lines in parallel with theR120G-CryAB construct (Figure 1 online, available in thedata supplement at http://www.circresaha.org). Three TGlines (lines 11, 13, and 41) were made using WT-CryABcDNA, whereas three TG lines (lines 25, 134, and 708)
expressed the R120G-CryAB transgene. Germline transmis-sion was confirmed, and normal mendelian ratios wereobserved, indicating that no embryonic lethality occurredwith either construct with those particular lines. GenomicSouthern blotting (Figure 1A) showed that the transgene copynumbers of the WT-CryAB lines 11, 13, and 41 were 22, 120,and 60, respectively. Those of R120G lines 708, 25, and 134were 1, 1, and 3, respectively. CryAB transcript and proteinlevels determined (Figures 1B and 1C). Consistent with thecopy numbers, all of the WT-CryAB TG lines showed higherCryAB mRNA and protein levels as compared with themutant lines (Figures 1B through 1D). Increases in solubleCryAB protein levels in the WT-CryAB TG lines accountedfor essentially all of the increase in protein. In contrast, theR120G lines show significant increases in the insoluble, andpresumably aggregated, fraction (Figure 1D). We have nowcarried the WT-CryAB lines for'16 months. To date, thereis no increase in mortality for the WT-CryAB TG miceFigure 1. TG DNA, mRNA, and protein levels. A, Genomic
Southern blots show that transgene copies in WT-CryAB mice(upper band) are substantially higher than in R120G-CryAB ani-mals. An EcoRI-SalI fragment (1.7 kb) derived from thea-myosin heavy chain promoter region was used as a probe,and the endogenous gene (lower band) allowed internal lanecontrols for copy number determinations. B, Transcript levels ofCryAB in different lines. All RNA samples were derived from6-week-old hearts. Total CryAB mRNA levels showed good butnot exact copy number dependency. Compared with the endog-enous CryAB mRNA level in NTG hearts, CryAB mRNA levels inlines 13, 41, 11, 708, 25, and 134 were increased by 106-, 50-,22-, 3.5-, 4.0-, and 9.0-fold, respectively. Histogram representsdegree of overexpression relative to endogenous transcript; val-ues obtained from 1 to 5 animals were averaged. C, Westernblot analyses of CryAB in the insoluble and soluble fractions.Desmin, actin, and CryAB levels in the insoluble fraction weredetermined simultaneously by immunoblotting. GAPDH andCryAB levels in the soluble fraction were also determined simul-taneously by immunoblotting. Actin and GAPDH were used asloading controls. D, Quantification of CryAB protein levels. Eachof the R120G-CryAB TG lines (708, 25, and 134) had increasedCryAB in the insoluble fraction relative to the comparableWT-CryAB TG lines. All data were normalized to the actin signal(for the insoluble fraction) or GAPDH signal (for the soluble frac-tion) in the same lane before being plotted. The experiment wasrepeated twice; error bars indicate SD. Compared with the en-dogenous CryAB protein level of NTG hearts, CryAB proteinexpression was increased in lines 13, 41, 11, 708, 25, and 134by 10-, 9-, 8-, 5-, 5.2-, and 7-fold, respectively.
Figure 2. Kaplan-Meier curve. Lines 11 (WT-CryAB) and 134were analyzed. A majority of R120G-CryAB TG mice died ofcongestive heart failure between 25 and 28 weeks of age, andall died by 32 weeks. No increases in mortality rate relative toNTG controls were detected in WT-CryAB TG mice.
Figure 3. Trichrome-stained myocardial sections. Shown areparaffin-embedded sections prepared from 10% buffered forma-lin perfusion–fixed hearts from 12-week-old mice. Lines 11 (WT-CryAB) and 708 and 134 (R120G-CryAB) were analyzed. Eosino-philic aggregates (arrows) are present in essentially every line134 myocyte. Similar but substantially smaller aggregates werefound in TG hearts of line 708, an R120G line with lower levelsof transgene expression. No aggregates were observed inWT-CryAB TG hearts. Bar520 mm.
Wang et al An aB-Crystallin Mutation Causes Desminopathy 85
by guest on August 6, 2014http://circres.ahajournals.org/Downloaded from
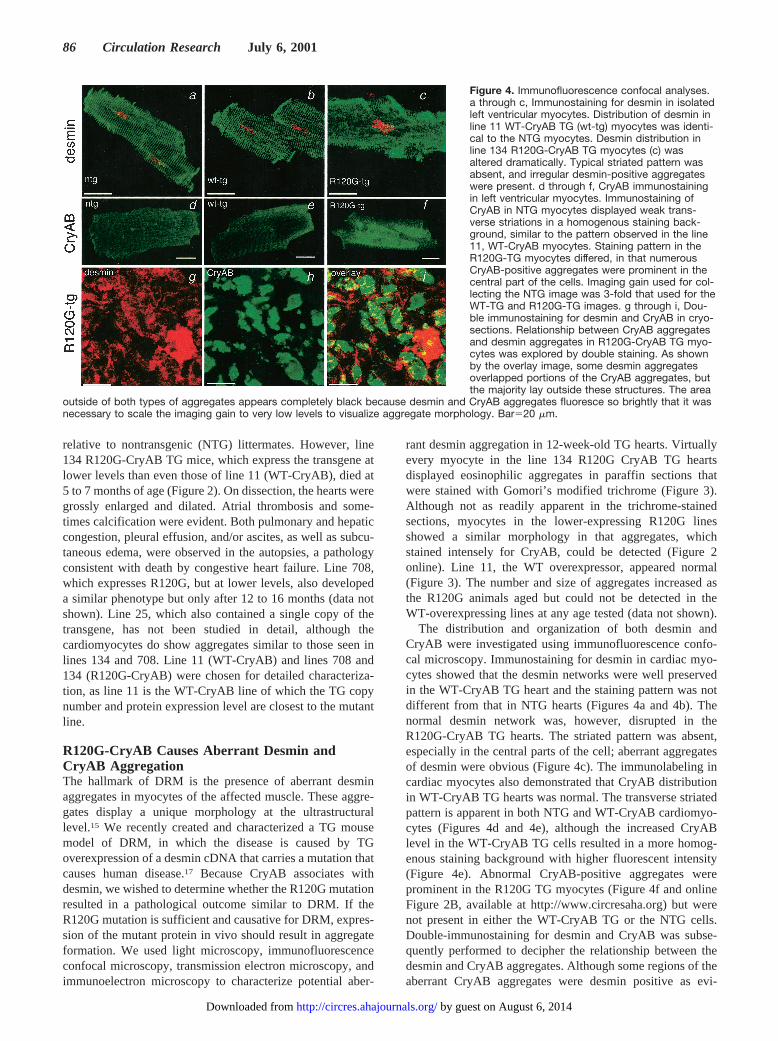
relative to nontransgenic (NTG) littermates. However, line134 R120G-CryAB TG mice, which express the transgene atlower levels than even those of line 11 (WT-CryAB), died at5 to 7 months of age (Figure 2). On dissection, the hearts weregrossly enlarged and dilated. Atrial thrombosis and some-times calcification were evident. Both pulmonary and hepaticcongestion, pleural effusion, and/or ascites, as well as subcu-taneous edema, were observed in the autopsies, a pathologyconsistent with death by congestive heart failure. Line 708,which expresses R120G, but at lower levels, also developeda similar phenotype but only after 12 to 16 months (data notshown). Line 25, which also contained a single copy of thetransgene, has not been studied in detail, although thecardiomyocytes do show aggregates similar to those seen inlines 134 and 708. Line 11 (WT-CryAB) and lines 708 and134 (R120G-CryAB) were chosen for detailed characteriza-tion, as line 11 is the WT-CryAB line of which the TG copynumber and protein expression level are closest to the mutantline.
R120G-CryAB Causes Aberrant Desmin andCryAB AggregationThe hallmark of DRM is the presence of aberrant desminaggregates in myocytes of the affected muscle. These aggre-gates display a unique morphology at the ultrastructurallevel.15 We recently created and characterized a TG mousemodel of DRM, in which the disease is caused by TGoverexpression of a desmin cDNA that carries a mutation thatcauses human disease.17 Because CryAB associates withdesmin, we wished to determine whether the R120G mutationresulted in a pathological outcome similar to DRM. If theR120G mutation is sufficient and causative for DRM, expres-sion of the mutant protein in vivo should result in aggregateformation. We used light microscopy, immunofluorescenceconfocal microscopy, transmission electron microscopy, andimmunoelectron microscopy to characterize potential aber-
rant desmin aggregation in 12-week-old TG hearts. Virtuallyevery myocyte in the line 134 R120G CryAB TG heartsdisplayed eosinophilic aggregates in paraffin sections thatwere stained with Gomori’s modified trichrome (Figure 3).Although not as readily apparent in the trichrome-stainedsections, myocytes in the lower-expressing R120G linesshowed a similar morphology in that aggregates, whichstained intensely for CryAB, could be detected (Figure 2online). Line 11, the WT overexpressor, appeared normal(Figure 3). The number and size of aggregates increased asthe R120G animals aged but could not be detected in theWT-overexpressing lines at any age tested (data not shown).
The distribution and organization of both desmin andCryAB were investigated using immunofluorescence confo-cal microscopy. Immunostaining for desmin in cardiac myo-cytes showed that the desmin networks were well preservedin the WT-CryAB TG heart and the staining pattern was notdifferent from that in NTG hearts (Figures 4a and 4b). Thenormal desmin network was, however, disrupted in theR120G-CryAB TG hearts. The striated pattern was absent,especially in the central parts of the cell; aberrant aggregatesof desmin were obvious (Figure 4c). The immunolabeling incardiac myocytes also demonstrated that CryAB distributionin WT-CryAB TG hearts was normal. The transverse striatedpattern is apparent in both NTG and WT-CryAB cardiomyo-cytes (Figures 4d and 4e), although the increased CryABlevel in the WT-CryAB TG cells resulted in a more homog-enous staining background with higher fluorescent intensity(Figure 4e). Abnormal CryAB-positive aggregates wereprominent in the R120G TG myocytes (Figure 4f and onlineFigure 2B, available at http://www.circresaha.org) but werenot present in either the WT-CryAB TG or the NTG cells.Double-immunostaining for desmin and CryAB was subse-quently performed to decipher the relationship between thedesmin and CryAB aggregates. Although some regions of theaberrant CryAB aggregates were desmin positive as evi-
Figure 4. Immunofluorescence confocal analyses.a through c, Immunostaining for desmin in isolatedleft ventricular myocytes. Distribution of desmin inline 11 WT-CryAB TG (wt-tg) myocytes was identi-cal to the NTG myocytes. Desmin distribution inline 134 R120G-CryAB TG myocytes (c) wasaltered dramatically. Typical striated pattern wasabsent, and irregular desmin-positive aggregateswere present. d through f, CryAB immunostainingin left ventricular myocytes. Immunostaining ofCryAB in NTG myocytes displayed weak trans-verse striations in a homogenous staining back-ground, similar to the pattern observed in the line11, WT-CryAB myocytes. Staining pattern in theR120G-TG myocytes differed, in that numerousCryAB-positive aggregates were prominent in thecentral part of the cells. Imaging gain used for col-lecting the NTG image was 3-fold that used for theWT-TG and R120G-TG images. g through i, Dou-ble immunostaining for desmin and CryAB in cryo-sections. Relationship between CryAB aggregatesand desmin aggregates in R120G-CryAB TG myo-cytes was explored by double staining. As shownby the overlay image, some desmin aggregatesoverlapped portions of the CryAB aggregates, butthe majority lay outside these structures. The area
outside of both types of aggregates appears completely black because desmin and CryAB aggregates fluoresce so brightly that it wasnecessary to scale the imaging gain to very low levels to visualize aggregate morphology. Bar520 mm.
86 Circulation Research July 6, 2001
by guest on August 6, 2014http://circres.ahajournals.org/Downloaded from

denced by the yellow color (overlay), the desmin aggregates(red) were, for the most part, distributed outside of the CryAB(green) aggregates (Figure 4i).
As noted above, the hallmark of DRM is the appearance ofelectron-dense, granular aggregates in the cytoplasm of theaffected cells.18 These structures were abundant in the R120Gcardiomyocytes. Two populations of electron-dense aggre-gates, named type I and II, were present in the intermyofi-brillar space (Figure 5). Type I aggregates (Figure 5c,asterisk) had a relatively low electron density and occupied alarge portion of the central part of the cardiomyocyte. Theywere substantially larger and more regular in shape than typeII. They had clear boundaries but were not enclosed in amembrane. In comparison with type II aggregates, theycontained finer granules. Type II aggregates (Figure 5c,arrow) were relatively small but more numerous. They wereirregular in shape and surrounded by numerous fine fila-ments. Morphologically, type II aggregates resembled thedesmin aggregates observed in TG animals that expressed themutant desmin protein.17 Some of the type II aggregatesappear to be associated with the nuclear envelope and otherswith the Z-band (Figure 5c). No direct physical associationbetween the two aggregate types was apparent (Figure 5c),although occasionally we noted that type II aggregates weretrapped in type I aggregates (data not shown). Immunoelec-tron microscopy analyses confirmed that type I aggregateswere CryAB positive (Figure 6c) with only a few scatteredgrains resulting from immunogold labeling with the desminantibody (Figure 7b). Type II aggregates were CryAB anddesmin positive (Figures 6 and 7). A number of desmin-positive filaments outside of the type II aggregates were alsoobserved, but no filamentous structure was associated withtype I aggregates (Figure 7b). Ultrastructural analyses alsoshowed that the alignment of adjacent myofibrils at theZ-band was perturbed in the R120G-CryAB TG hearts(Figure 5c). Generally in these animals, Z-band thickness wasincreased and its normal uniformity lost (Figures 5 through7). The pathologies were essentially identical between the leftand right ventricles.
Expression of the CryAB Mutation CausesCardiac HypertrophyCommon outcomes of DRM are skeletal muscle atrophy andcardiac hypertrophy and, as expression of the mutant proteinwas restricted to the heart, we wished to determine whetherthe R120G mice recapitulated this aspect of the humandisease. At the molecular level, activation of fetal geneticprograms is common during the early stages of hypertrophy,with upregulation of atrial natriuretic factor andb-myosinand downregulation ofa-myosin, phospholamban, and thesarcoplasm reticulum calcium pump. Transcript levels weremeasured at 1, 3, and 6 months of age as the R120Gpathology developed (Figure 3 online, available at http://www.circresaha.org). The relative amounts of these tran-scripts as well as those of desmin and total CryAB showed apattern consistent with the morphological disarray apparent inthe confocal and transmission electron microscopy analyses.That is, expression of R120G-CryAB but not overexpressionof WT-CryAB triggered a hypertrophic response at the
molecular level in the heart. By 3 months, hypertrophy wasapparent at the gross level as well, with significant increasesin the ventricular weight/tibial length ratios (Figure 4Aonline). Hypertrophy continued to develop and was evenmore pronounced at 6 months.
To determine whether the cardiomyocytes themselves werelarger, the cell volume, profile area, and length, as well as the
Figure 5. Electron microscopy. Sections were obtained from3-month-old NTG (a), WT-CryAB TG (b), and R120G-CryAB TG(c) left ventricles. In both panels a and b, sarcomeres are orga-nized normally with clearly distinguishable I- and A-bands, andM-lines as well as regularly aligned Z-bands. Mitochondria aretypically arranged along myofibrils. The sarcoplasmic reticulumis closely associated with sarcomeres. No major differencesbetween NTG and WT-CryAB TG samples are apparent,although there are subtle alterations in the space where mito-chondria are localized (panel a vs panel b). In R120G-CryAB TGhearts (c), two populations of abnormal aggregates are present.Type I aggregates (*) are relatively large, regularly shaped, andof low electron density. Type II aggregates (arrows) are smallerand irregularly shaped, display relatively high electron density,and are associated with Z-bands. Z-band morphology, particu-larly near the aggregates, is perturbed. Bar51.0 mm.
Wang et al An aB-Crystallin Mutation Causes Desminopathy 87
by guest on August 6, 2014http://circres.ahajournals.org/Downloaded from

transverse sectional area (TSA), were measured at 3 and 6months (Figure 4 online). In both the left and right ventricles,cardiomyocyte size progressively increased (P,0.01) in theR120G-CryAB TG mice as compared with the NTG andWT-CryAB TG controls. At 3 months, the increase in cellsize was due to increases in the TSA. By 6 months, both thecell length and TSA were larger, indicating that concentrichypertrophy at the cellular level occurs in both ventricles at 3months and ventricular chamber dilatation occurs later on asthe heart begins to fail. This is consistent with the clinicalprogression observed in many cardiovascular diseases, inwhich a compensatory hypertrophy is observed early, butlater on decompensated heart failure presents. No abnormal-ities were observed in the cohort that overexpressed the WTprotein.
R120G-CryAB Leads to Cardiac DysfunctionCardiac function in patients with DRM is often significantlycompromised. As the disease progresses, the heart dilates,systolic function becomes compromised, and heart failureoccurs. Considering the effects of R120G-CryAB expressionon the cellular structure and organization, and the resultanthypertrophy, we wished to measure the impact of R120G-CryAB or WT-CryAB TG expression on cardiac function at3 months of age. In the early stages of compensatoryhypertrophy, contractile function is often maintained or evenincreased, whereas deficits in relaxation begin to occur.19 Todivorce the system from endogenousb-adrenergic stimula-tion and determine whether any functional deficits presentedat this stage, an isolated work-performing heart preparation20
was used (Figure 8A). Contractile function, as measuredusing the left ventricular pressure waveform in the isolatedwork-performing heart, showed that1dP/dt was higher in theR120G-CryAB TG hearts than in the NTG and WT-CryABTG controls (P,0.005). Relaxation, as measured by the firstderivative of left ventricular pressure (2dP/dt), was signifi-cantly lower than in either control cohort (P,0.005).
By 6 months, the animals’ presentation was consistent withsevere heart failure and, rather than the working heart, in vivohemodynamics17,21 were used so that the neurohumoral axis,which helps to maintain cardiac function, could be taken intoaccount. In vivo hemodynamics showed that baseline abso-lute values of both dP/dtmax and dP/dtmin were significantlydecreased relative to the NTG and WT-CryAB TG controls.Even when stimulated via catecholamines, the hearts wereunable to maintain normal contractility and an overt responseto b-agonist stimulation via dobutamine infusion was sub-stantially blunted (Figures 8B and 8C). Determination of tau(Figure 8D), the monoexponential time constant of relax-ation, showed that the deficits in relaxation were relativelyload independent. The data show that cardiac function issignificantly compromised.
DiscussionCardiac and skeletal muscles contain the highest CryABlevels among the nonlenticular tissues. Upregulation ofCryAB occurs in a number of cardiac disorders, includingfamilial hypertrophic cardiomyopathy and DRM, but thefunctional consequences are unknown. Linkage of R120G-CryAB to familial DRM suggests that normal CryAB func-tion, which presumably involves chaperone activity, is crucialfor desmin filament formation and/or function. Interestingly,the mutant CryAB protein appears to be relatively resistant todegradation as compared with normal CryAB. As the TGmice age, total CryAB protein in the mutant CryAB heartsprogressively increases, whereas CryAB protein levels stayrelatively constant in the WT-CryAB TG heart despite muchhigher transcript levels (data not shown). The relationshipbetween CryAB, desmin, and DRM pathogenesis remainsobscure. The data presented in this study show that upregu-lation of normal CryAB is not, by itself, detrimental to theheart. However, in the intact animal, the R120G mutationresults in both abnormal CryAB aggregation and aberrantdesmin aggregation. The sequelae accurately recapitulate the
Figure 6. Anti-CryAB immunogold label-ing. CryAB distribution was analyzed atthe ultrastructural level. Left ventricularsections were obtained from NTG (a),WT-CryAB TG (b), and R120G-CryAB TG(c and d) 12-week-old mice. CryAB dis-tribution in the WT-CryAB TG and NTGhearts is similar, although there is someincreased labeling in the intermyofibrillarspace (a and b). Both the type I (*) and II(arrow) aggregates in R120G-CryAB TGhearts (c and d) are CryAB positive, butstaining in type I aggregates was twicethe intensity found in type II aggregates.Bar50.5 mm.
88 Circulation Research July 6, 2001
by guest on August 6, 2014http://circres.ahajournals.org/Downloaded from

progression of cardiovascular disease as the heart first at-tempts to compensate for the structural insult by hypertro-phying but eventually transits into a decompensated, dilatedstate with heart failure as the final outcome.
No discernible phenotype presented in the WT-CryAB TGmice, supporting the conclusion that simple alterations inCryAB stoichiometry are not responsible for the phenotype.Oligomerization is required for CryAB to exert its molecularchaperone function,16,22 and this is consistent with theR120G-missense mutation of CryAB being dominant nega-tive. The genetics of the R120G mutation indicate an auto-somal dominant inherited pattern in the human disease,14 andin vitro analyses of R120G-CryAB polymers, or a mixture ofR120G:WT-CryAB protein, result in altered morphology andcompromised molecular chaperone function.16,22 The TGmice offer possible insights into the functional deficits thatresult. The type I and II aggregates in the R120G mice areintriguing. The two types of aggregates are distinguished bothby their morphology and by their protein composition. TypeI aggregates are CryAB positive and contain only traces ofdesmin, whereas type II aggregates are both desmin and
CryAB positive and are similar to the characteristic desminaggregates found in DRM hearts carrying desmin mutations.It is well established that WT CryAB binds to desmin anddesmin filaments, especially when cells are stressed.4,23,24Wehypothesize that the relative paucity of desmin in the R120G-CryAB–loaded type I aggregates is due to the inability of themutant CryAB to productively interact with desmin. The dataalso indicate that desmin aggregate formation in these mice isnot due to a physical or biochemical interaction betweendesmin and the R120G mutant CryAB. Indeed, formation ofthe aberrant desmin aggregates may be caused by a loss offunction of the mutant CryAB.
We observed significant differences in copy number andexpression levels for the WT versus R120G transgenes(Figure 1). The explanation probably lies in the embryoniclethality of embryos containing higher copy numbers of theR120G transgene. In fact, an R120G-CryAB founder with'30 copies of the transgene died at 8 weeks and so could notbe bred. Another founder, who was mosaic for the transgene,produced TG pups who all died from congestive heart failurebefore 4 weeks. When analyzed, these mice had 40 copies ofthe R120G transgene, and the histology (at 3 weeks) showedtremendous aggregate accumulation in the cardiomyocytes(data not shown).
Previously we produced a mouse model of DRM bycardiac-specific expression of a transgene containing adesmin mutation that causes human disease.17 Those mice didnot exhibit the severe morbidity and mortality that present inthe R120G animals. Similarly, desmin-null mice also exhibita less severe pattern of morbidity and mortality.25–27 Thus,both loss-of-function and dominant-negative alleles ofdesmin result in disease, but the pathology is markedly lesssevere than that resulting from the R120G-CryAB mutation.R120G TG mice develop cardiac hypertrophy that is concen-tric at an early stage (3 months) but leads to dilation andfailure by 5 to 7 months. All mice from line 134, which haveonly three copies of the transgene, die during this period.Unfortunately, there is a lack of human data in terms ofmutant/WT protein expression with which we can comparethe mouse data. The earlier adulthood high mortality of line134 relative to lines 708 and 25 is almost certainly due to thehigher dosage. The higher level of mutant protein expressiongives rise to a more pronounced phenotype at an earlier time.We have noted that line 708 TG mice also tend to dieprematurely, but this is apparent only in the older adultpopulation, and statistically significant data have not yet beenaccumulated, although there are alterations at the cardiomyo-cyte level (Figure 2 online, available at http://www.circresa-ha.org). The desmin-null mice also die prematurely, but asubstantial percentage of the null cohort survives for up to ayear.25,26 The desmin mutation–induced DRM TG cohortslive for at least 18 months.17 These observations suggest thatthe cardiac dysfunction caused by R120G-CryAB must bedue to more than just a loss of desmin function and implyeither additional roles for CryAB that can impact on thegeneral cytoskeletal architecture or other, as-yet-undefinedtargets for CryAB interaction(s).
In the young adults (3 months), systolic function is actuallyincreased as measured using the isolated working heart
Figure 7. Antidesmin immunogold labeling. Distribution ofdesmin was analyzed at the ultrastructural level. Left ventricularsections were obtained from NTG (a) or R120G-CryAB TG (b)12-week-old mice. Immunogold particles in the NTG heart werein the intermyofibrillar space, mainly at the Z-line level and inter-calated disks. In R120G-CryAB TG hearts (b), most label waslocalized in type II aggregates (*), with type I aggregates beingonly slightly positive. No desmin labeling was evident in areasimmediately surrounding type I aggregates, whereas a numberof desmin-positive filaments surrounded the type II aggregates.Bar50.5 mm.
Wang et al An aB-Crystallin Mutation Causes Desminopathy 89
by guest on August 6, 2014http://circres.ahajournals.org/Downloaded from

preparation (Figure 8A). Although this may appear to besomewhat surprising, at this early stage, the R120G TG miceshow concentric hypertrophy (Figure 4 online). The hyper-trophic myocardium displayed an increased1dP/dt in re-sponse to changes in preload, and we have observed similar1/2dP/dt profiles in the early stage of concentric hypertro-phy induced by pressure overload. Occurrence of hypertrophyat this stage might be due to functional deficit in diastole.Recently, we have evaluated in vivo left ventricular functionon 3-month-old mice. Similarly, baseline1dP/dtmax of theR120G TG mice was unaffected (X. Wang and J. Robbins,unpublished observations, 2001). Histological changes at 3months include primarily concentric hypertrophy with mini-mal indications of the more advanced changes that accom-pany failure. We observed the more severe histologicalchanges and increased mortality at 5 to 7 months and beyondand believe the observed increases in1dP/dt at 3 months ofage are consistent with the early stages of a compensatoryhypertrophy. The primary performance effect of the mutationis decreased diastolic function, which most likely leads toinitial compensation at 3 months and ultimate failure at 7months and beyond.
In cardiomyocytes, desmin filaments link adjacent myofi-brils to one another, to the cell membrane, and to the nuclearenvelope.26 As other intermediate filaments do in other cells,the desmin filaments play an important role in maintaining
the structural integrity of myocytes,25 and thus it is notsurprising that alterations in a molecular chaperone, whichfunctions in their transport, can have severe consequences.Previously, Vicart et al14 showed, via transfection of musclecell cultures with the R120G mutant, that characteristicaggregates developed. Our data significantly extend theseobservations, showing that stable expression of a mutatedchaperone in the heart can lead directly first to a compensatedhypertrophy and eventually to heart failure. By combining acombination of biochemical and whole-animal approaches, itshould be possible to define the role(s) of CryAB, both innormal cardiomyocyte function and in cardiovascular disease.
AcknowledgmentsThis work was supported by NIH Grants HL56370, HL41496,HL56620, HL52318, HL60546, and HL56620 (to J.R.) andHL62459 (to A.M.G.), and by the American Heart Association, OhioValley Affiliate (to X.W.).
References1. Piatigorsky J. Lens crystallins and their gene families.Cell. 1984;38:
620–621.2. Iwaki T, Kume-Iwaki A, Liem RK, Goldman JE.a B-Crystallin is
expressed in non-lenticular tissues and accumulates in Alexander’sdisease brain.Cell. 1989;57:71–78.
3. Longoni S, Lattonen S, Bullock G, Chiesi M. Cardiaca-crystallin, II:intracellular localization.Mol Cell Biochem. 1990;97:121–128.
4. Bennardini F, Wrzosek A, Chiesi M.a B-crystallin in cardiac tissue:association with actin and desmin filaments.Circ Res. 1992;71:288–294.
Figure 8. Functional analyses. A, Isolated work-performing heart preparations were used to examine left ventricular function at 3months. No differences were detected between NTG and WT-CryAB TG mice (n56). B and C, Closed chest intact mouse model anddobutamine infusion protocols have been described in detail.21 D, Tau, the monoexponential time constant of relaxation, is relativelyload-independent and was determined from primary data. Each experimental set (n55) consisted of 23-week-old animals. Cardiovascu-lar function was measured at baseline (no dobutamine) and under increasing b-adrenergic stimulation. The three cohorts were all sub-jected to the same regimen, which included a series of 3-minute infusions of increasing concentrations of dobutamine and measure-ment of cardiovascular indices in the last 30 seconds of each period to obtain peak response. Pressure signals from both the COBEand Millar transducers were recorded using a MacLab 4/s data acquisition system (AD Instruments). The software directly determinesarterial systolic and diastolic pressure, mean arterial pressure, heart rate, left ventricular systolic pressure, developed pressure, andboth positive (dP/dtmax) and negative (dP/dtmin) dP/dt. Data were analyzed using a mixed, two-factor ANOVA with repeated measures onthe second factor. At all points, there were no statistically significant differences between the NTG and WT-CryAB cohorts. When nec-essary, post hoc comparisons were performed by single degree-of-freedom contrasts. Compared with NTG and WT-CryAB animals,both dP/dtmax and dP/dtmin differed significantly at the indicated points. *P#0.05, #P#0.005.
90 Circulation Research July 6, 2001
by guest on August 6, 2014http://circres.ahajournals.org/Downloaded from

5. Nicholl ID, Quinlan RA. Chaperone activity ofa-crystallins modulatesintermediate filament assembly.EMBO J. 1994;13:945–953.
6. Wang K, Spector A.a-Crystallin stabilizes actin filaments and preventscytochalasin-induced depolymerization in a phosphorylation-dependentmanner.Eur J Biochem. 1996;242:56–66.
7. Barbato R, Menabo R, Dainese P, Carafoli E, Schiaffino S, Di Lisa F.Binding of cytosolic proteins to myofibrils in ischemic rat hearts.CircRes. 1996;78:821–828.
8. Ito H, Okamoto K, Nakayama H, Isobe T, Kato K. Phosphorylation ofaB-crystallin in response to various types of stress.J Biol Chem. 1997;272:29934–29941.
9. Hoover HE, Thuerauf DJ, Martindale JJ, Glembotski CC.a B-Crystallingene induction and phosphorylation by MKK6-activated p38: a potentialrole for a B-crystallin as a target of the p38 branch of the cardiac stressresponse.J Biol Chem. 2000;275:23825–23833.
10. Xiao X, Benjamin IJ. Stress-response proteins in cardiovascular disease.Am J Hum Genet. 1999;64:685–690.
11. Hwang DM, Dempsey AA, Wang RX, Rezvani M, Barrans JD, Dai KS,Wang HY, Ma H, Cukerman E, Liu YQ, Gu JR, Zhang JH, Tsui SK,Waye MM, Fung KP, Lee CY, Liew CC. A genome-based resource formolecular cardiovascular medicine: toward a compendium of cardiovas-cular genes.Circulation. 1997;96:4146–4203.
12. Arbustini E, Morbini P, Grasso M, Fasani R, Verga L, Bellini O, DalBello B, Campana C, Piccolo G, Febo O, Opasich C, Gavazzi A, FerransVJ. Restrictive cardiomyopathy, atrioventricular block and mild to sub-clinical myopathy in patients with desmin-immunoreactive materialdeposits.J Am Coll Cardiol. 1998;31:645–653.
13. Shinohara H, Inaguma Y, Goto S, Inagaki T, Kato K.a B crystallin andHSP28 are enhanced in the cerebral cortex of patients with Alzheimer’sdisease.J Neurol Sci. 1993;119:203–208.
14. Vicart P, Caron A, Guicheney P, Li Z, Prevost MC, Faure A, Chateau D,Chapon F, Tome F, Dupret JM, Paulin D, Fardeau M. A missensemutation in theaB-crystallin chaperone gene causes a desmin-relatedmyopathy.Nat Genet. 1998;20:92–95.
15. Abraham SC, DeNofrio D, Loh E, Minda JM, Tomaszewski JE, PietraGG, Reynolds C. Desmin myopathy involving cardiac, skeletal, andvascular smooth muscle: report of a case with immunoelectronmicroscopy.Hum Pathol. 1998;29:876–882.
16. Bova MP, Yaron O, Huang Q, Ding L, Haley DA, Stewart PL, HorwitzJ. Mutation R120G inaB-crystallin, which is linked to a desmin-related
myopathy, results in an irregular structure and defective chaperone-likefunction.Proc Natl Acad Sci U S A. 1999;96:6137–6142.
17. Wang X, Osinska H, Dorn GW 2nd, Nieman M, Lorenz JN, Gerdes AM,Witt S, Kimball T, Gulick J, Robbins J. A mouse model of desmin-relatedcardiomyopathy.Circulation. 2001;103:2402–2407.
18. Baeta AM, Figarella-Branger D, Bille-Turc F, Lepidi H, Pellissier JF.Familial desmin myopathies and cytoplasmic body myopathies.ActaNeuropathol (Berl). 1996;92:499–510.
19. Hunter JJ, Tanaka N, Rockman HA, Ross J Jr, Chien KR. Ventricularexpression of a MLC-2v-ras fusion gene induces cardiac hypertrophy andselective diastolic dysfunction in transgenic mice.J Biol Chem. 1995;270:23173–23178.
20. Gulick J, Hewett TE, Klevitsky R, Buck SH, Moss RL, Robbins J.Transgenic remodeling of the regulatory myosin light chains in themammalian heart.Circ Res. 1997;80:655–664.
21. Lorenz JN, Robbins J. Measurement of intraventricular pressure andcardiac performance in the intact closed-chest anesthetized mouse.Am JPhysiol. 1997;272:H1137–H1146.
22. Perng MD, Muchowski PJ, van Den IP, Wu GJ, Hutcheson AM, Clark JI,Quinlan RA. The cardiomyopathy and lens cataract mutation inaB-crystallin alters its protein structure, chaperone activity, and inter-action with intermediate filaments in vitro.J Biol Chem. 1999;274:33235–33243.
23. Golenhofen N, Ness W, Koob R, Htun P, Schaper W, Drenckhahn D.Ischemia-induced phosphorylation and translocation of stress proteinaB-crystallin to Z lines of myocardium.Am J Physiol. 1998;274:H1457–H1464.
24. Head MW, Hurwitz L, Kegel K, Goldman JE.aB-Crystallin regulatesintermediate filament organization in situ.Neuroreport. 2000;11:361–365.
25. Li Z, Mericskay M, Agbulut O, Butler-Browne G, Carlsson L, ThornellLE, Babinet C, Paulin D. Desmin is essential for the tensile strength andintegrity of myofibrils but not for myogenic commitment, differentiation,and fusion of skeletal muscle.J Cell Biol. 1997;139:129–144.
26. Milner DJ, Weitzer G, Tran D, Bradley A, Capetanaki Y. Disruption ofmuscle architecture and myocardial degeneration in mice lacking desmin.J Cell Biol. 1996;134:1255–1270.
27. Milner DJ, Taffet GE, Wang X, Pham T, Tamura T, Hartley C, GerdesAM, Capetanaki Y. The absence of desmin leads to cardiomyocytehypertrophy and cardiac dilation with compromised systolic function.JMol Cell Cardiol. 1999;31:2063–2076.
Wang et al An aB-Crystallin Mutation Causes Desminopathy 91
by guest on August 6, 2014http://circres.ahajournals.org/Downloaded from

Online Supplemental MaterialExpression of R120G-α-B-Crystallin Causes Aberrant Desmin and α-B-Crystallin
Aggregation and Cardiomyopathy in Mice
Xuejun Wang,1 Hanna Osinska,1 Raisa Klevitsky,1 A. Martin Gerdes,2 Michelle Nieman,3
John Lorenz,3 Timothy Hewett1 and Jeffrey Robbins1
Running Title: An -B-crystallin mutation causes desminopathy
1Division of Molecular Cardiovascular Biology, Children’s Hospital Research
Foundation, 3333 Burnet Avenue, Cincinnati, Ohio, USA
2South Dakota Health Research Foundation-Cardiovascular Research Institute, Sioux
Falls, South Dakota, USA
3Department of Molecular and Cellular Physiology, University of Cincinnati Medical
Center, Cincinnati, Ohio; USA
Address correspondence to: Jeffrey Robbins, Division of Molecular Cardiovascular
Biology, The Children’s Hospital Research Foundation, 3333 Burnet Avenue, Cincinnati,
OH 45229, USA. Phone: (513) 636-8098; Fax: (513) 636-3852; E-mail:
Subject Codes: (16) myocardial cardiomyopathy disease; (130) animal models of human
disease; (146) genetically altered mice; (15) hypertrophy; (110) congestive heart failure

MS #2383/R1
2
Supplemental Methods
Transgenic (TG) Mouse Lines
FVB/N mice were used for all studies. Animals were housed in a temperature controlled,
germ-free barrier facility that was AAALAC-approved. All experiments were approved
by the Children’s Hospital Research Foundation Animal Care Review Board. The full-
length CryAB cDNA was cloned using RT-PCR and multiple isolates were sequenced
(Genbank accession #M73741). Although 3 amino acid changes are present between the
human and mouse αB-crystallin proteins, the Arg120 residue and flanking sequences are
well conserved (Figure 1, Online Supplement). The R120G-CryAB was made from the
WT-CryAB cDNA using PCR-based mutagenesis. Subsequently the cDNA’s were placed
in the α-myosin heavy chain (MyHC) promoter1 and used to generate TG mice. Genomic
Southern blots in combination with PCR were employed for genotyping, using a pair of
primers corresponding to the junction between exons 1 and 2. The primer sequences are:
GAGTC TGACCTCTTCTCAACAGCC and CTGGCGTTCTTCGTGCTTGCCGTG.
Copy number was determined using genomic Southern blots.
Transcript and Protein Analyses
Total ventricular RNA was isolated separately from at least 3 hearts of each line and the
integrity of the TG RNA confirmed by Northern analysis and sequencing. RNA levels
were quantitated using dot blots hybridized to transcript-specific oligonucleotides. The
total CryAB protein levels were determined using SDS-PAGE followed by western blots
with a polyclonal antibody against CryAB (Stressgen, Victoria, BC Canada). Protein
extracts were prepared from cytoskeletal (detergent insoluble) and soluble fractions of
myocardial homogenates. Approximately 20 mg of apical ventricular myocardium that
had been snap-frozen in liquid nitrogen and stored at -80°C was homogenized on ice in
PBS, pH 7.4, with 1% Triton X-100, 5 mM EDTA, 1 mM PMSF and 1 tablet/10 ml of

MS #2383/R1
3
protease inhibitor cocktail (Roche, Indianapolis, IN). The homogenates were centrifuged
at 14,000 ×g at 4°C and the supernatant (soluble fraction) separated from the pellet. The
pellet was then resuspended in SDS-PAGE loading buffer, boiled, centrifuged, and the
resultant supernatant was kept as the insoluble fraction. The ECF western blotting
protocol (ECF, Amersham Life Sciences, Arlington Heights, IL) was utilized for
quantification of CryAB protein and an enhanced chemiluminescent protocol (F.
Hoffmann-La Roche Ltd, Basal, Switzerland) used for film exposure.
Morphological Analyses
At least 3 tissue samples from each area of the heart were picked randomly for extensive
ultrastructural analysis. Immuno-electron microscopy (EM) was employed to detect the
localization of desmin and CryAB protein at the ultrastructural level. For immuno-EM,
hearts were perfused with 1% glutaraldehyde/3% paraformaldehyde in cardioplegic
buffer (5% dextrose, 30 mM KCl in PBS), treated in the same fixative in cacodylate
buffer, incubated in 0.1M glycine/PBS, dehydrated in N, N-dimethyl formamide (DMF)
and embedded in LR-White resin. Ultrathin sections were picked up on nickel grids,
dried and etched with a saturated solution of sodium m-periodate and 0.1N HCl. The
immunolabeling procedure consisted of a PBS rinse, a 1.5 hour incubation in 1% BSA,
0.1% cold water fish skin gelatin and 1% Tween20 in PBS, an overnight incubation in
rabbit anti-desmin (Biomeda, Foster City, CA) (1:25) or anti-CryAB (Stressgen) (1:200)
antibody diluted with 1% BSA/PBS, a rinse in PBS, a 1.5 hour incubation in the blocking
solution and then a 2 hour incubation with goat anti-rabbit IgG tagged with colloidal gold
(10 nm particle size, 1:20) (Aurion/Electron Microscopy Sciences, Fort Washington, PA).
After a PBS rinse, the samples were postfixed in 2% glutaraldehyde and rinsed in water.
They were then counter-stained with uranium/lead and viewed in a Zeiss Omega 912
electron microscope at 100 kV. Indirect immunofluorescence staining was carried out on
both fixed isolated myocytes and myocardial cryo-sections.2 Hearts were excised, fixed

MS #2383/R1
4
by coronary perfusion with 4% paraformaldehyde, saturated with 30% sucrose solution
and embedded in Tissue-Tek O.C.T. (Sakura Finetek U.S.A, Torrance, CA).
Cryosections or mounted isolated myocytes were air-dried, incubated with 0.1M glycine
in PBS (pH7.2) for 30 minutes, treated with 1% Triton X-100 for 1 hour, and blocked
with 0.5% BSA/10% goat serum in PBS. The specimens were then incubated with mouse
monoclonal antibodies to desmin (Sigma, St. Louis, MO) and/or rabbit polyclonal
antibodies to CryAB (Stressgen) overnight at 4°C, and subsequently with TRITC
conjugated anti-mouse IgG and FITC conjugated anti-rabbit IgG antibodies (Sigma, St.
Louis, MO) for 1 hour before being examined using confocal microscopy.
Cardiac Functional Analyses
Cardiac function was evaluated at 3 and 6 months of age using isolated working
performing heart preparations and in vivo hemodynamics as described previously.3,4

MS #2383/R1
5
Supplemental Figure Legends
Figure 1. A schematic diagram of α-B-crystallin (CryAB) domains and the transgenic
constructs. The CryAB molecule contains 3 regions: a crystallin homologous region (1-
59), the heat shock protein 20 homologous region (HSP, 60-162), and a low complexity
sequence (LCS, 162-175), which is located at the carboxyl terminus. The human and
mouse CryAB amino acid sequences at and around the mutation are shown and the amino
acid numbers indicated. The missense mutation (R120G) is located in the HSP domain.
Constructs containing either the normal α-B-crystallin (WT-CryAB) or the missense
mutation (R120G-CryAB) were linked to the α-myosin heavy chain (α-MyHC)
promoter: the 3 vertical boxes in the promoter represent noncoding exons in the 5’-
untranslated region. PolyA splicing is provided by the human growth hormone sequences
downstream of the inserted cDNA (hGH).
Figure 2. Immunofluorescence confocal analyses of NTG (A) and line 708 (B)
cryosections. One-month animals were used and the presence of CryAB aggregates
detected by anti-CryAB staining and confocal analysis. Even at 1 month in the less
affected line, aggregates can be detected by this methodology. Numerous CryAB positive
aggregates were prominent in the central part of the cells. The imaging gains used for
collecting the images were identical. Similar aggregates could also be detected in line 25
animals (data not shown). Bar = 20 µm.
Figure 3. Temporal changes in hypertrophy responsive gene expression. Total RNA was
isolated from ventricular myocardium of TG (lines 11 and 134) or nontransgenic (NTG)
mice at 1, 3, and 6 months (m) of age. Three mice were used for each group at each time
point. The transcript levels of GAPDH, total CryAB, α-MyHC, β-MyHC, desmin, atrial
natriuretic factor (ANF), sarcoplasmic reticulum calcium ATPase-2a (SERCA), and
phospholamban (PLB) were quantified using RNA dot blots and transcript specific

MS #2383/R1
6
oligonucleotides probes.5 GAPDH levels were used to correct for variations in loading.
The NTG value at each time point was arbitrarily set as 1 (100%), transcript levels in the
WT-CryAB TG and R120G-CryAB TG groups were normalized to the corresponding
NTG control. CryAB expression in the R120G-CryAB line134 mice was always lower
than that of the line 11 WT-CryAB TG controls, and was substantially down-regulated at
6 months. Expression levels of β-MyHC, desmin, and ANF were progressively up-
regulated and those of α-MyHC, SERCA, and PLB were down-regulated, a pattern that is
consistent with a developing heart failure phenotype.6-8 It should be noted that GAPDH
expression is slightly increased in the older R120G mice, probably as a result of the
pathology that is present or because of alterations in general metabolism. Although it
serves as useful internal loading control, its use in normalization might accentuate
differences in the myosin and total CryAB transcript levels in the 6-month R120G
groups. This may be due to one or more of the following: (1) down regulation of the
endogenous transcript as it has been recently reported that CryAB is down regulated in
human heart failure. (2) modest down regulation of the myosin promoter that drives
transgene expression. In other circumstances, during some forms of hypertrophy, we have
noticed a 10-20% down-regulation of TG expression. (3) potential up-regulation of
GAPDH as above. (4) total CryAB mRNA levels in the NTG control to which they are
compared were slightly increased at 6 months. The error bars are one standard deviation.
After normalization to the corresponding GAPDH signal, the mean intensity value of the
NTG group is set at 1 at each time point. The intensity signal of each individual dot is
then normalized to the mean of the NTG at the corresponding time point. The resultant -
fold number of each of the 3 animals from the same group at the same time was used to
derive the mean and standard deviation, as represented by the error bars. **P< 0.05, *P<
0.01.

MS #2383/R1
7
Figure 4. Temporal changes in cardiac hypertrophy. (A) Changes in ventricle
weight/tibia length ratios. For each group for each time point, 7-9 mice were examined.
(B) Changes in the size of left (LV) and right (RV) ventricular myocytes. Myocytes were
isolated from NTG and R120G CryAB (line 134) LV and RV using 3 or 6 month old
animals. For each group, 6 animals were used for each time point. The myocyte size (CV)
was directly measured from 3000-5000 cells using a Z2 Coulter Sizer (Beckman Coulter,
Inc., Miami, FL). Consistent with the gravimetric data, both LV and RV myocyte size
was significantly increased at 3 months and continued to increase up to failure at 6
months in the R120G-CryAB TG group. No differences were detected between the NTG
and WT-CryAB TG groups (data not shown). (C) Temporal changes in LV myocyte
length (CL) and transverse section area (TSA). Myocyte lengths and profile areas were
measured from photographed images with NIH image software (v1.54), using 50 isolated
myocytes for each sample. TSA was calculated as CV/CL. No differences were detected
between the NTG and WT-CryAB TG groups (data not shown). In the R120G-CryAB
TG group, the TSA of the LV myocytes was significantly increased at both 3 and 6
months while cell length was increased only at 6 months. The error bars represent the
standard deviation. Compared to NTG or WT-CryAB TG, *P< 0.05, **P< 0.01.

MS #2383/R1
8
References
1. Gulick J, Hewett TE, Klevitsky R, Buck SH, Moss RL, Robbins J. Transgenic
remodeling of the regulatory myosin light chains in the mammalian heart. Circ
Res. 1997; 80:655-664.
2. Wang X, Li F, Campbell SE, Gerdes AM. Chronic pressure overload cardiac
hypertrophy and failure in guinea pigs: II. Cytoskeletal remodeling. J Mol Cell
Cardiol. 1999; 31:319-331.
3. Tardiff JC, Hewett TE, Palmer BM, Olsson C, Factor SM, Moore RL, Robbins J,
Leinwand LA. Cardiac troponin T mutations result in allele-specific phenotypes
in a mouse model for hypertrophic cardiomyopathy. J Clin Invest. 1999; 104:469-
481.
4. Lorenz JN, Robbins J. Measurement of intraventricular pressure and cardiac
performance in the intact closed-chest anesthetized mouse. Am J Physiol. 1997;
272:H1137-1146.
5. Jones WK, Grupp IL, Doetschman T, Grupp G, Osinska H, Hewett TE, Boivin G,
Gulick J, Ng WA, Robbins J. Ablation of the murine alpha myosin heavy chain
gene leads to dosage effects and functional deficits in the heart. J Clin Invest.
1996; 98:1906-1917.
6. Boluyt MO, O'Neill L, Meredith AL, Bing OH, Brooks WW, Conrad CH, Crow
MT, Lakatta EG. Alterations in cardiac gene expression during the transition
from stable hypertrophy to heart failure. Marked upregulation of genes encoding
extracellular matrix components. Circ Res. 1994; 75:23-32.

MS #2383/R1
9
7. Vikstrom KL, Bohlmeyer T, Factor SM, Leinwand LA. Hypertrophy, pathology,
and molecular markers of cardiac pathogenesis. Circ Res. 1998; 82:773-778.
8. Just H. Pathophysiological targets for beta-blocker therapy in congestive heart
failure. Eur Heart J. 1996; 17 Suppl B:2-7.


Supplemental Online Figure 2

0
2.5
5
7.5
10
β-M
yHC
1 3 6Age (months)
R120GWt-TGNTG
1m
3m6m
0
0.5
1
1.5
α-M
yHC
1 3 6Age (months)
R120GWt-TGNTG
1m3m6m
GA
PD
HNTG Wt-TG R120G
1m
3m
6m
0
0.4
0.8
1.2
SE
RC
A
1 3 6Age (months)
R120GWt-TGNTG
1m3m6m
0
1
2
3
Des
min
1 3 6Age (months)
R120GWt-TGNTG
1m
3m6m
0
2.5
5
7.5
10
AN
F
1 3 6Age (months)
R120GWt-TGNTG
1m
3m6m
0
0.5
1
1.5
PLB
1 3 6Age (months)
R120GWt-TGNTG
1m3m6m
05
1015202530
Cry
AB
1 3 6Age (months)
R120GWt-TGNTG
1m
3m6m
Supplemental Online Figure 3
*
*
*
*
*
*
*
*
**
***
*
* *

0
3
6
9
12
Ven
tric
le/T
ibia
(m
g/m
m)
0 1 2 3 4 5 6 7Age (months)
R120G-CryAB TGWT-CryAB TGNTG
**
**
0
20000
40000
60000
80000
Myo
cyte
Siz
e (µ
m3 )
3 6 3 6Age (months)
R120G-CRYABNTG
**
**
** **
LV
RV
0
100
200
300
400
500
3 60
30
60
90
120
150
180
Cel
l Len
gth
(µm
)
3 6Age (months)
*
**
**
A
B
C
Supplemental Online Figure 4
TS
A µ
m2












![^_`]ab - DPU – Direitos Humanos](https://static.fdokumen.com/doc/165x107/6334d004d2b728420307a0c7/ab-dpu-direitos-humanos.jpg)






