
An Excitatory Loop with Astrocytes Contributes to Drive Neurons to Seizure Threshold
Upload
independentCategory
view
3download
0
&p.1:Abstract Using the Monte Carlo method, spontaneousfast excitatory postsynaptic currents (mEPSCs) at a hip-pocampal synapse were simulated by releasing150–20,000 glutamate molecules from a point sourcecentred 15 nm above a rectangular grid of 14 × 14 α-amino-3-hydroxy-methyl-isoxazole (AMPA) receptorsand assuming the channel kinetics to be as reported byJonas et al. [J Physiol (Lond) 472:615; 1993]. The rela-tionship between the amplitudes of mEPSCs and theirtime constants of decay is positive, but not pronouncedin physiological conditions (except when the number ofmolecules released is very high). It increases as desensi-tization is reduced and becomes highly pronouncedwhen it is eliminated. mEPSCs are prolonged with re-peated opening of AMPA channels due to enhancementof two concentration-dependent processes: (1) binding ofglutamate molecules by AMPA receptors, and (2) occu-pancy of both activatable bound states. In contrast, thetime constant of decay of the patch currents evoked by ashort glutamate pulse is independent of glutamate con-centration and current amplitude in control conditions,and only moderately concentration dependent in the ab-sence of desensitization. The fast application protocolthus fails to reproduce synaptic currents reliably whenthere is repeated binding of glutamate molecules toAMPA receptors. During an mEPSC, the occupancy ofdesensitized states increases rapidly and it strongly de-pends on the number of glutamate molecules released.Desensitization reaches its maximum after an mEPSCdecays to very low levels, and recovers very slowly(from tens to hundreds of milliseconds), and in a concen-
tration-dependent manner. In conclusion, under physio-logical conditions the desensitization of AMPA receptorsplays a major role in shaping the time course of mEPSCsby minimizing the repeated opening of AMPA channels.
&kwd:Key words Glutamate · AMPA receptor channels ·Desensitization · Buffering · Monte Carlo method&bdy:
Introduction
The mechanisms determining the amplitude and the timecourse of unitary synaptic currents, especially in the cen-tral nervous system (CNS), are not well understood [5, 9,16]. The time course of decay of the fast [i.e. α-amino-3-hydroxy-methyl-isoxazole (AMPA) receptor activated]excitatory postsynaptic currents (mEPSCs) has been at-tributed to the kinetics of postsynaptic channel closure[12, 22], or to some combination of prolonged presence ofneurotransmitter in the cleft [2, 6, 23] and channel desen-sitization [19, 24, 25]. Two comprehensive approacheshave been traditionally taken to describe the diffusion ofneurotransmitter molecules in the cleft coupled to the ki-netic models of channel opening and closing: (1) a numer-ical solution of partial differential equations [27] – in thismethod the channel kinetics are driven by the bulk con-centration of the transmitter in the synaptic cleft; and (2) aMonte Carlo method, in which each transmitter moleculeis followed as it diffuses randomly within the synapticcleft and interacts with the postsynaptic receptors [3, 26]– this allows us to explore the stochastic properties ofboth neurotransmitter diffusion and channel kinetics.
Following the recent measurements of the fast excita-tory currents at the mossy fibre CA3 synapse [7, 17, 20],a kinetic scheme for the ligand-gated channel responsi-ble for the fast excitatory currents was published [17]. Inthe present study we used this kinetic scheme [17] andMonte Carlo methods to investigate the factors affectingthe time course of the spontaneous mEPSCs in the hip-pocampus. Our simulations show that the time constantof decay of mEPSCs (rd), which is amplitude dependent
M. I. GlavinovicDepartment of Physiology, McGill University,3655 Drummond Street, Montreal, PQ, H3G IY6, Canada
M. I. Glavinovic (✉)Department of Anaesthesia Research, McGill University,3655 Drummond Street, Montreal, PQ H3G 1Y6, Canada
H. R. RabieDepartment of Chemical Engineering, McGill University,3655 Drummond Street, Montreal, PQ H3G 1Y6, Canada&/fn-block:
Pflügers Arch – Eur J Physiol (1998) 435:193–202 © Springer-Verlag 1998
O R I G I N A L A RT I C L E
&roles:Mladen I. Glavinovic · Hamid R. Rabie
Monte Carlo simulation of spontaneous miniature excitatory postsynapticcurrents in rat hippocampal synapse in the presence and absenceof desensitization
&misc:Received: 17 April 1997 / Received: after revision: 11 August 1997 / Accepted: 1 September 1997

only when the number of molecules released is high inphysiological conditions, becomes markedly amplitudedependent for any number of molecules when desensiti-zation is reduced or abolished, because the buffering ofglutamate in the synaptic cleft, and the occupancy of thebound activable states by the AMPA receptors are bothincreased and concentration dependent.
Materials and methods
Theory
Synaptic transmission in the excitatory synapse in the hippocam-pus was simulated using a Monte Carlo method [3, 26]. Thismethod was used instead of numerically evaluating the partial dif-ferential equations that describe the macroscopic quantities. It en-ables one to study the fluctuations due to the stochastic nature ofthe diffusion of the individual transmitter molecules, their interac-tions with the receptor molecules, and the stochastic nature of thegating of the associated ion channels. At each discrete time step:(1) every transmitter molecule is associated with a position (x,y,z),and is either flagged as free or bound – free molecules move ran-domly in all three dimensions or interact with receptors; (2) everyreceptor has a fixed position and is considered to be in one of thestates of a given kinetic scheme – every receptor has a finite prob-ability of changing to another state according to the kineticscheme. The changes of the receptor states were assumed to beMarkovian (i.e. they were assumed to depend only on their presentposition and not on their previous history).
Computer simulations
All simulations in this paper have been done using MATLAB(MathWorks, Natick Mass., USA) on either a 486 or a PentiumIBM computer. Since the rise time of mEPSCs is of the order of≈100 µs, we chose the time step in our simulation to be 1µs (seebelow; [26]). The distance travelled by a transmitter molecule (ineach of three dimensions) was chosen randomly from a Gaussiandistribution with mean of 0 and a standard deviation σ given by
σ = √2Dδt (1)
[13, 26], where the δt is the length of the time step and D is thediffusion coefficient of the transmitter molecule. The diffusion co-efficient of glutamine (D = 7.6 × 10–6 cm2/s measured in water at25°C; [18]) was taken to be the diffusion coefficient of glutamatein the synapse. The Q10 for diffusion was assumed to be 1.3 [13].The random numbers were obtained from a random number gener-ator. The diffusion in a restricted space was simulated by assum-ing that the transmitter molecules collide elastically with the“walls” of the space (i.e. presynaptic and postsynaptic membranes;[26]). Finally all molecules that reach the edge of the model syn-apse and diffuse away into the “infinite” space (or return from itinto the synaptic cleft) were also followed throughout simulation.
According to the kinetic scheme of channel gating (Fig. 1) areceptor can be unbound (C1), in a single- or in a double-boundstate (C2 and C3 respectively; these are also called activatablestates), open (O4), or in one of the three desensitized states (C5, C6and C7). Following previous reports [3, 26], we associated witheach state of the receptor a surface area and a probability of bind-ing, given that a transmitter molecule “hits” this receptor surface.The inverse of the receptor surface area is taken to be the densityof the receptor molecules (σr) at the postsynaptic membrane. Theprobability (Pb) that a transmitter molecule, after hitting the recep-tor surface, will bind in a given time step (δt) is related to the mac-roscopic rate constant by
Pb = [(σρκ)/Na]√(πδt)/ D (2)
where Na is Avogadro’s number and κ is the appropriate rate con-stant of binding (in M–1 s–1). For steps in the kinetic scheme thatdid not involve binding of the transmitter, the probability (p) that areceptor will move to a new state in a given time step (δt) is relat-ed to the macroscopic rate constant by
p = 1 –e –k δ t (3)
where k is the appropriate rate constant (in s–1). The time step(chosen to be 1µs in all our calculations) was such that the proba-bility of a receptor changing state twice within one time step was< 1 in 100. It has been shown previously that the results do not be-come more accurate by reducing the time step further [26].
Equation 3 also applies to the situation when a transmitter mole-cule unbinds from a receptor. We have chosen to move a moleculethe mean length of the random jump (0.67σ) perpendicularly awayfrom the receptor surface at unbinding: (1) to ensure that the proba-bility of a given receptor making a transition to a bound state doesnot depend on the receptor’s previous history; and (2) because suchphysical separation between the neurotransmitter and the receptor ac-curately reproduces the macroscopic unbinding rate constant [3]. Inall our calculations we used the rate constants of the kinetic schemeof Jonas et al. [17] without restrictions, as these values have beenfound to be more adequate than those with restrictions (Fig. 1). Therate constants used in the kinetic scheme, however, were adjusted forthe difference in temperature (37°C in all our calculations) and as-suming a Q10 of 4.0 for all rate constants as done previously [26].
We modelled a typical synapse as one with 196 receptors in thepostsynaptic membrane, 14 nm apart in a rectangular array of14 × 14, and the synaptic cleft as a 15-nm-thick sheet [26]. Thischoice was made for a “typical” synapse because, assuming a par-ticle density of 3000 particles per µm2 (from CA1 pyramidal cells,[11]), one obtains 600 particles within 0.2µm2. On the other hand,the lower limit of the number of available fast excitatory channelsin a mossy fibre CA3 synapse has been estimated from the averageamplitude of quantal events as 20–100 [17].
A simulation of voltage-clamp experiments on an outside-outmembrane patch was done by constructing a 100-nm “box” aroundthe receptor grid. We fixed the concentration of glutamate at a givenlevel while keeping the size of the box constant. In order to maintainthe concentration of glutamate, any binding or unbinding of gluta-mate molecules was not associated with removal or addition of glu-tamate molecules from or to the “box”. The glutamate moleculeswere distributed randomly within the box at the beginning of thesimulation. To simulate abrupt changes in the glutamate concentra-tion (pulses), we distributed additional molecules at random posi-tions in the box or removed a certain number of free molecules.
Results
Amplitude dependence of kinetics of synaptic currentsin the presence and absence of desensitization
To simulate the release of a single vesicle of neurotrans-mitter, N molecules were simultaneously released intothe cleft at a point source on the presynaptic membrane,centred above the postsynaptic receptor grid. The timecourse of the number of channels in the “open” state(Fig. 1, state O4) simulates mEPSC that might be record-ed under conditions of perfect voltage clamp and no den-dritic filtering [15]. Figure 2A illustrates such mEPSCssimulated at 37°C. N ranged from 600 to 10,000 as in-dicated (traces are averages of five and ten simulationruns with different random seeds; two upper and threelower traces, respectively). Note that the decay times ofmEPSCs (τd) became clearly longer as N increases.
194

Several studies have shown that mEPSCs are pro-longed following application of compounds such as an-iracetam or cyclothiazide that reduce or abolish desensi-tization [1, 10, 14, 25]. Furthermore our studies havesuggested that the amplitude dependence of τd increasesfollowing such a treatment [1, 10]. Since it may be ar-gued that this is due to the presence of different types ofAMPA channels at different synapses (with different sin-gle-channel conductance, mean open time and sensitivityto desensitization) we simulated the synaptic currents inthe presence and in the absence of desensitization to de-termine what the amplitude dependence of τd is (in thepresence and in absence of desensitization) at a synapsebeing mediated by the same type of AMPA channels.The simulations focused on establishing whether the pu-
195
Fig. 1 Kinetic scheme for gating of AMPA channels. The con-stants taken from Jonas et al. [17] assume no restrictions.They are: K+1 = 3.67 × 107 M–1 s–1, k-1 = 3.41 × 104 s—1, K+2 =22.7 × 107 M–1 s–1, k-2 = 2.61 × 104 s–1, K+3 = 1.02 × 107 M–1 s–1,k-3 = 366 s–1 for glutamate binding; α = 3.39 × 104 s–1, β =7200 s–1 for channel opening; and α1 = 2.31 × 104 s–1, β1 =314 s–1, α2 = 1376 s–1, β2 = 5.82 s–1, α3 = 142 s–1, β3 = 32 s–1,α4 = 134.4 s–1, β4 = 1523 s–1 for the desensitization pathway&/fig.c:
Fig. 2A, B Decay of simulated miniature excitatory postsynapticcurrents (mEPSCs) is concentration, and thus amplitude depen-dent. Two families of simulated mEPSCs are shown, one calculat-ed assuming physiological rate constants for desensitization (A;control), and the other calculated assuming that all desensitizedstates have been eliminated (B; no desensitization). The number ofglutamate molecules released (N) was as indicated. Traces are av-erages of 5 and 10 simulations runs with different random seedsfor the 2 upperand 3 lower tracesrespectively. The time course ofthe postsynaptic current is expressed as the fraction of the totalnumber of channels in the open state at any point in time. The de-cay phase of mEPSCs (τd) increases as N increases and elimina-tion of desensitization prolongs it further&/fig.c:
Fig. 3A The amplitude dependence of decay times (rd) of mE-PSCs increases as desensitization is reduced and becomes verypronounced when it is eliminated (note the semilog scale). B Theamplitude dependence of τd in the absence of desensitization, butwith the density of AMPA receptors reduced to a half or increasedto twice the control value. C The relationship between the numberof molecules released (N) and mEPSC amplitude (A); D N versusτd. [SquaresPhysiological rates of desensitization, upright trian-gles desensitization reduced by decreasing α1 to one-tenth of thecontrol value, circles all desensitization states eliminated, down-ward trianglesand diamonds, dashed lineall desensitization stateseliminated and the diffusion constant reduced to a half and a quar-ter of control value (as indicated), crosses, dotted lineclosure ratereduced to one-third of the control value]&/fig.c:
tative amplitude dependence of τd is due to the concen-tration dependence of channel gating, buffering of gluta-mate molecules by AMPA receptors or some combina-tion of these. Figure 2B illustrates a family of mEPSCsequivalent to that shown in Fig. 2A but with all desensi-tized states abolished. mEPSCs are clearly prolongedfollowing abolition of desensitization. To gain an insightinto whether other factors, and especially the bufferingof glutamate molecules by AMPA receptors, contributeto shaping the time course of mEPSCs, we made addi-tional simulations that were in all respects equivalent,but permitted only a single binding of a glutamate mole-cule to AMPA receptors. Following the unbinding, theglutamate molecule was taken out of the system. Finally,we also determined the amplitude dependence of τd inthe absence of desensitization, but with either half ordouble the density of AMPA receptors. Figure 3 summa-rizes these findings showing the amplitude dependenceof τd (Fig. 3A), and the relationships between the ampli-tude of the mEPSC or τd and N (Fig. 3C and D respec-

tively; the maximal occupancy of the open state was tak-en to be the amplitude).
The relationship between τd and the amplitude of themEPSC was positive when desensitization rates were attheir physiological level, but was not very pronounced, ir-respective of whether a single or a multiple binding of glu-tamate molecules was assumed, except for large values ofN. The relationships involving the single binding are notshown to avoid crowding, and were essentially the same asthose for multiple binding. When desensitization was re-duced (by reducing the rate from the single to the desensi-tized state), and especially when it was eliminated, τd be-came markedly amplitude dependent (Fig. 3A, B). A re-duction of desensitization, by either a reduction of the ratefrom the double-bound state to a corresponding desensi-tized state or alternatively by an increase of the rate lead-ing in the opposite direction, was in all respects similar tothe one presented (not shown). The difference in durationof τd for a case of multiple and single binding becameclear in the absence of desensitization (Fig. 3D). However,even complete elimination of desensitization altered rela-tionship between the amplitude of the mEPSC and N onlymodestly (Fig. 3C); in all cases the buffering of glutamatemolecules had very little effect on the relationship (notshown). Slower diffusion (i.e. lower diffusion constant)leads to a markedly longer τd. With slower closing of theopen channel, the τd versus amplitude relationship wasshifted upwards but the slope was not increased (Fig. 3A).Finally, reducing the density of receptors did not lead toless, but rather to more,pronounced amplitude dependenceof τd (note that to enable a comparison the amplitude val-ues were multiplied by the number of AMPA receptors;Fig. 3B). These findings argue that buffering of glutamatemolecules plays only a minor role in altering either the am-plitudes or the time courses of mEPSCs in the absence ofdesensitization, but plays an important role when desensiti-zation is abolished (especially if the diffusion of glutamatein the cleft is slower than that in solution). These findings,however, also prompt several questions. According to oursimulations, the number of glutamate molecules releasedwas typically greater, and in some cases much greater, thanthe number of AMPA receptors postsynaptically. Howcould so few AMPA receptors buffer so many glutamatemolecules? This is discussed in the following section.
Buffering of glutamate in the synaptic cleft– effect of desensitization
Buffering was calculated as the difference in glutamateconcentration in the synaptic cleft between a case whenAMPA receptors were present and a case when theywere completely absent. In all simulations, the concen-tration of glutamate in the cleft was calculated as an av-erage concentration in a volume centred above the recep-tor grid. We used a volume that covers the postsynapticgrid and overlaps the edges of the grid by 5 nm in everydirection (15 × 192 × 192 nm3; [26]). Figure 4A showsthe dependence of the cleft glutamate concentration on
N. The buffering of glutamate molecules by AMPA re-ceptors was especially important for large values of N(buffering was thus concentration dependent), and wasclearly greater throughout when desensitization wasabolished. Change of the gating mechanism clearly canalter the buffering of glutamate in the synaptic cleft (theelimination of desensitization of AMPA receptors in-creasesthe buffering). Note also that the glutamate con-centration increase due to buffering becomes relativelymore important with time (Fig. 4B).
Occupancy of an unbound and activatable single-and double-bound states – effect of desensitization
The occupancy of the activatable double-bound state(state C3; FCdb) and the transition probability from thedouble-bound to the open state (Tdb,o) determine the fre-quency of opening of AMPA channels. Similar reasoningapplies to the occupancy of the activatable single-boundstate (state C2; FCsb), the transition probability from thesingle-bound to the activatable double-bound state
196
Fig. 4A, B The cleft concentration of glutamate is altered signifi-cantly because of buffering by AMPA receptors. A The glutamatecleft concentration in the absence of AMPA receptors (filled cir-cles), in their presence and with physiological rates of desensitiza-tion (filled downward triangles) or with all desensitization stateseliminated (filled upright triangles). The difference between physio-logical or zero desensitization and no AMPA receptors (open up-right or downward trianglesrespectively). In all cases, the concen-tration was calculated as an average over an interval of from 0.95 to1.0 ms from the start of the release. B Time course of glutamateconcentration in the synaptic cleft (semilog scale). Three families ofthree traces each are shown. The number of molecules released isgiven to the right of the trace. In all cases the upper tracerepresentsthe simulation in the absence of desensitization, the middle tracethephysiological case and the lower trace simulation in absence ofAMPA receptors. The cleft concentration is clearly greater in the ab-sence of desensitization, with the effect increasing with time&/fig.c:

(Tsb,db; Tsb,db was, however, strongly dependent on theglutamate concentration in the synaptic cleft) and thefrequency of transition from the activatable single-boundto the activatable double-bound state. Figure 5A givesthe occupancy of the activatable single-bound and dou-ble-bound states as well as their sum. The occupancy ra-tio is given in Fig. 5B (zero desensitization over control).The ratio, which was always greater than one (straightline), was especially high for low values of N. These ra-tios are expected to be greater than unity throughout thetime interval examined, since over the whole interval theoccupancy of the activatable double-bound state wasgreater in the absence of desensitization (Fig. 5C).
To better understand how changes of the glutamateconcentration in the synaptic cleft and the occupancy ofthe activatable bound states affect the transition probabili-ty and the frequency of opening of the AMPA channels,
we studied a case of physiological and one of zero desen-sitization. We set N to 1250 and assessed the results 1 msafter the start of release. The transitional probability fromthe activatable double-bound to the open state was, in thecontrol case, Tdb,o = α/(α + α2 + k–2), and was equal to0.55; for zero desensitization this gives α/(α + k–2) and0.57. Since the frequency of transitions to the open statefrom the activatable double-bound state was equal toFCdb × Tdb,o, and since FCdb was 1.69% for physiologicaldesensitization and 5.81% for zero desensitization, the fre-quency of transition increases 3.5 times. A similar line ofargument applies to a transition from a single-bound to anactivatable double-bound state. Tsb,db= K+2/(α1 + K+2 + k–1),and was equal to 0.057 for a control case. For a zero de-sensitization case this gives Tsb,db= K+2/(K+2 + k–1), whichwas equal to 0.163. Since the occupancy of the activatablesingle-bound state was 0.82% and 8.1% for the controland the zero desensitization cases, respectively, the fre-quency of transition increases 28.3 times. Thus, the in-crease was so pronounced because of: (1) a much greaterFCsb, and (2) a greater Tsb,db (this occurs mainly becauseof a greater K+2; K+2 was concentration dependent and theglutamate concentration in the cleft increases) and to amuch lesser extent because α1 was set to zero). The rela-tionship between Tsb,dband the concentration of glutamateis strongly nonlinear (not shown). As a result, a very smallfraction of all transitions occurs between a single-boundand an activatable double-bound state at low concentra-tions, but essentially all occur at high concentrations (this
197
Fig. 5A, B The occupancy of AMPA receptors in a single- or adouble-bound activatable state increases when desensitization iseliminated. A (Circles Occupancy of the activatable single-boundstate, upright trianglesthe activatable double-bound state, squarestheir sum.) In all cases open symbolsindicate a physiological rateof desensitization and filled symbolsthe case when all desensitizedstates are eliminated. All estimates are averages over an interval offrom 0.95 to 1.0 ms from the start of the release. B Occupancy ra-tios (zero/physiological desensitization). Filled circles, upright tri-anglesand squaresare the symbols for the activatable, single- ordouble-bound states, and their sum, respectively. Note that the ra-tios are always greater than one (straight line), and are especiallypronounced for low numbers of molecules released. C The occu-pancy of the activatable double-bound state is greater in the ab-sence of desensitization throughout the time interval examined.Both traces are the averages of 10 runs&/fig.c:
Fig. 6A Occupancy of the unbound state is also affected by thepresence or absence of desensitization. The number of moleculesreleased is as indicated. Occupancy was estimated 1 ms fromthe start of release. B The ratio of the occupancies (FCUnb, NoDes/FCUnb, Control) has a maximum when approximately 2500 mole-cules are released, and is approximately four (i.e. four times asmany AMPA channels are in an unbound state in the absence ofdesensitization)&/fig.c:
change in the level of transitions occurs within a very nar-row range of concentrations). When desensitization wasabolished the sensitivity of Tsb,db to glutamate concentra-tion increased, i.e. there was a shift to the left. This in-crease is another reason why a lower desensitization leadsto a higher occurrence of reopening of AMPA channelsand to a prolongation of mEPSCs but only to marginallygreater amplitudes of mEPSCs.
Finally the occupancy of the unbound state (state C1)also changed when desensitization was abolished. Figure6A shows the occupancy of the unbound state 1 ms afterthe release of the transmitter, as a function of N. Notethat irrespective of N more AMPA receptors were in theunbound state in the absence of desensitization than inits presence. However, their ratio (i.e. the ratio of the oc-cupancy in the absence and in the presence of desensiti-zation; Fig. 6B) was about 1 at low values of N. Itreached to a maximum of 4 at N ≈ 2500, and decreasedto 1 with further increases in N. A higher level of occu-pancy of the unbound state results in a greater occupancyof the activatable single-bound state as well as of the act-ivatable double-bound and the open states. It also con-
tributes to the higher concentration of glutamate in thesynaptic cleft (i.e. to greater buffering).
Concentration dependence of desensitizationof synaptic currents
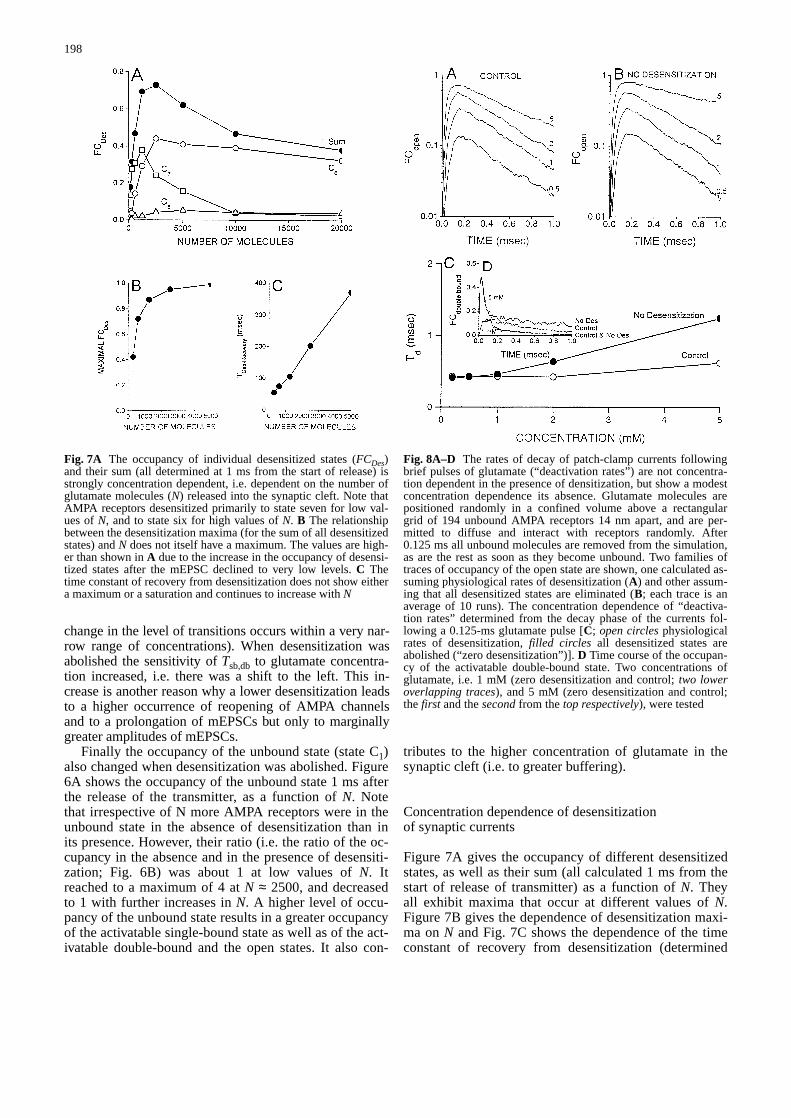
Figure 7A gives the occupancy of different desensitizedstates, as well as their sum (all calculated 1 ms from thestart of release of transmitter) as a function of N. Theyall exhibit maxima that occur at different values of N.Figure 7B gives the dependence of desensitization maxi-ma on N and Fig. 7C shows the dependence of the timeconstant of recovery from desensitization (determined
198
Fig. 7A The occupancy of individual desensitized states (FCDes)and their sum (all determined at 1 ms from the start of release) isstrongly concentration dependent, i.e. dependent on the number ofglutamate molecules (N) released into the synaptic cleft. Note thatAMPA receptors desensitized primarily to state seven for low val-ues of N, and to state six for high values of N. B The relationshipbetween the desensitization maxima (for the sum of all desensitizedstates) and N does not itself have a maximum. The values are high-er than shown in A due to the increase in the occupancy of desensi-tized states after the mEPSC declined to very low levels. C Thetime constant of recovery from desensitization does not show eithera maximum or a saturation and continues to increase with N&/fig.c:
Fig. 8A–D The rates of decay of patch-clamp currents followingbrief pulses of glutamate (“deactivation rates”) are not concentra-tion dependent in the presence of densitization, but show a modestconcentration dependence its absence. Glutamate molecules arepositioned randomly in a confined volume above a rectangulargrid of 194 unbound AMPA receptors 14 nm apart, and are per-mitted to diffuse and interact with receptors randomly. After0.125 ms all unbound molecules are removed from the simulation,as are the rest as soon as they become unbound. Two families oftraces of occupancy of the open state are shown, one calculated as-suming physiological rates of desensitization (A) and other assum-ing that all desensitized states are eliminated (B; each trace is anaverage of 10 runs). The concentration dependence of “deactiva-tion rates” determined from the decay phase of the currents fol-lowing a 0.125-ms glutamate pulse [C; open circlesphysiologicalrates of desensitization, filled circles all desensitized states areabolished (“zero desensitization”)]. D Time course of the occupan-cy of the activatable double-bound state. Two concentrations ofglutamate, i.e. 1 mM (zero desensitization and control; two loweroverlapping traces), and 5 mM (zero desensitization and control;the first and the secondfrom the top respectively), were tested&/fig.c:

from the decay phases of the desensitization occupancytraces) on N. Both the time constant of desensitizationrecovery and the desensitization maxima strongly de-pend on N, but their relationships differ greatly. Whilethe former continues to increase with N, the latter showssaturation with its values approaching the theoreticallimit (i.e. almost all AMPA receptors become desensi-tized following a single mEPSC) even for comparativelylow values of N.
Simulations of macroscopic patch-clamp currents
We simulated macroscopic patch-clamp currents to de-termine how currents, produced with a glutamate pulsewith a well-defined time course and without the contri-bution of buffering, depend on N or on the abolition of
desensitization. Figure 8 gives two families of openchannel traces resulting from 0.125-ms pulses of gluta-mate (concentration in mM as indicated; semilog scale)followed by 0.875 ms of recovery. Figure 8A showsphysiological rates of desensitization, while Fig. 8Bshows zero desensitization. The duration of the gluta-mate pulse was chosen to be this short because the ratesin the kinetic scheme were high (due to the high temper-ature assumed in the present simulation – see Materialsand methods). Deactivation rates were essentially con-centration independent for physiological rates of desensi-tization, whereas they were moderately concentration de-pendent in the absence of desensitization (Fig. 8C). Thisoccurred as a result of the concentration dependence ofthe occupancy of the open state and a high transitionprobability between the activatable double-bound and theopen states. Both factors favour a repeated and a concen-tration-dependent opening of AMPA channels, and, as aresult, a concentration-dependent occupancy of not onlythe open but also of the activatable double-bound state.This is illustrated in Fig. 8D in which the time course ofthe occupancy of the activatable double-bound state isshown for two different concentrations of glutamate:1 mM (zero desensitization and control; two lower over-lapping traces), and 5 mM (zero desensitization and con-trol; the first and the second from the top, respectively).
Figure 9 gives two families of traces, one for the occu-pancy of the open state (A) and the other for the occupan-cy of all desensitized states (C; concentrations in mM wereas indicated). The time constants of desensitization (esti-mated from fits of a single exponential and a constant tothe traces shown in Fig. 9A) did not show a clear concen-
199
Fig. 9A–D The rates of decay of patch-clamp currents duringpulses of glutamate with intermediate duration (“desensitizationrates”) are not concentration dependent. Glutamate molecules arepositioned randomly in a confined volume above a rectangular gridof 194 unbound AMPA receptors 14 nm apart, and are permitted todiffuse and interact with receptors randomly. A The family of trac-es of occupancy of the open state shown was calculated assumingphysiological rates of desensitization. B The concentration depen-dence of “desensitization rates” determined from the decay phaseof the currents during 2-ms glutamate pulses. C Correspondingfamily of traces of occupancy of combined desensitization states(glutamate concentrations were as indicated; 10, 5 and 2 runs wereaveraged when 0.5–2 mM, 5 mM and 10–20 mM were used, re-spectively). D The concentration dependence of occupancy of com-bined desensitization states (determined at the end of glutamatepulse) shows a broad but a clear maximum at ≈0.5 mM

tration dependence (Fig. 9B). The concentration depen-dence of the occupancy of all desensitized states (deter-mined 1 ms after the beginning of simulation) was obvi-ous, however, peaking at approximately 0.5 mM (Fig. 9D).
Discussion
Amplitude and time course of miniature currentsin the presence and absence of desensitization
It is still controversial whether a positive (or any) relation-ship exists between the amplitudes and the kinetics of thefast spontaneous miniature currents. Several reports haveargued against such a positive relationship [4, 14]. In con-trast, our recent results show that such a positive relation-ship exists, as demonstrated by control recordings, andthat it increases dramatically following a reduction of de-sensitization by aniracetam and cyclothiazide [1, 10]. Thisstudy was undertaken to clarify whether the kinetics ofmEPSCs is amplitude dependent at synapse(s) with thesame type of AMPA receptors. The mEPSCs were simu-lated using Monte Carlo methods.
The amplitude of mEPSCs is not greatly affected by areduction or a complete abolition of desensitization, irre-spective of N values or whether single or repeated bind-ing of glutamate molecules is assumed. This is in excel-lent agreement with the experimental findings that neithermeans nor CV values of the amplitude change signifi-cantly following the addition of aniracetam or cyclothi-azide, which profoundly affects their time course [1, 10,14]. The relationship between the decay time of mEPSCs(τd) and its amplitude is positive but not pronounced (ex-cept for large values of N) when the rates of desensitiza-tion are at physiological levels. However, it becomesmarkedly so when desensitization is abolished becausethe prolongation of τd is amplitude dependent. The lowerthe diffusion constant (i.e. the slower the diffusion) thegreater the amplitude dependence of τd. On the otherhand, the rate of closure of open AMPA receptor chan-nels does not increase the amplitude dependence of τd.When only a single binding of glutamate molecules toAMPA receptors is permitted to occur τd values are short-er. However, when the rates of desensitization are atphysiological levels, repeated binding of glutamate mole-cules has little effect on τd. Its effect becomes very pro-nounced when desensitization is abolished. These find-ings all suggest that in the absence of desensitization,mEPSCs are prolonged because of the repeated openingof AMPA channels, itself caused by repeated binding ofglutamate molecules to AMPA receptors. A surprisingfinding is that in the absence of desensitization, a greaterdensity of AMPA receptor channels is not associated witha greater, but with a lower, amplitude dependence of τd.Greater amplitude of mEPSCs and the insensitivity of τdto higher density of AMPA receptor channels explainthese findings, which are in contrast to changes observedat the neuromuscular junction. This is clearly in contrastto changes observed at the neuromuscular junction.
Buffering of glutamate molecules by AMPA receptorsand occupancy of activatable bound states duringan mEPSC
Initially, the cleft glutamate concentration decreases rap-idly in the presence or absence of desensitization and, asexpected during this time interval, the desensitization hasa minimal or no effect on the rate of decrease. However,at approximately 0.2 ms, when the rate of decrease slowsconsiderably and the desensitization has an important ef-fect on the time course of mEPSCs, the glutamate con-centration is already significantly reduced. Thus, thesefindings do not support the interesting recent suggestion[8] that the glutamate concentration has to be high andessentially constant for the duration of an mEPSC if de-sensitization is to have a major impact on the time courseof mEPSCs. As discussed later, concomitant changes ofthe occupancy of the single- and the activatable double-bound state, as well as changes of the transition proba-bility from a single- to the double-bound activatable stateappear to be responsible for these findings.
The increase in the cleft glutamate concentration dueto the buffering (assessed from the concentration differ-ence in the presence and absence of AMPA receptors)depends on the number of glutamate molecules released– buffering is thus concentration dependent. A relativeincrease (i.e. the concentration increasedue to the buff-ering normalized by the glutamate concentration in theabsence of any AMPA receptors) assessed 1 ms after thestart of release of glutamate molecules was greater forlow values of N, and was especially pronounced in theabsence of desensitization: it ranged from 15 to 115%and from 46 to 228% for control and zero desensitiza-tion, respectively. Buffering of glutamate by AMPA re-ceptors clearly and strongly depends on the mechanismof gating of AMPA channels. Note, however, that theglutamate is not likely to be buffered by AMPA receptorsonly. Uptake transporters may also be an important fac-tor in affecting the kinetics of synaptic currents [2, 21].If their contribution is highly significant, they may maskthe differences in the buffering of glutamate by AMPAreceptors.
Prolongation of mEPSCs caused by the frequent re-opening of AMPA channels is also due to a greater occu-pancy of the activatable double-bound state. This mayoccur either because of the greater occupancy of the act-ivatable single-bound state and/or the greater transitionprobability from the single- to the double-bound activa-table state. Both changes occur as N increases and/or asdesensitization is reduced or eliminated; neither is sur-prising. The two rates κ+1 and κ+2, leading to the single-and the double-bound activatable states respectively, areconcentration dependent. Furthermore, with desensitiza-tion reduced or eliminated, the occupancy of all nonde-sensitized states (i.e. the activatable bound states, the un-bound state and the open state) increases markedly andin a concentration-dependent manner. Almost all AMPAreceptors become desensitized at physiological rates ofdesensitization, even for modest values of N.
200

It is beyond the scope of this paper to characterize indetail why altering the density of AMPA receptor chan-nels does not affect τd. However, during the 1-ms timeinterval starting from the beginning of glutamate release,the glutamate concentration in the cleft was (except atthe very beginning) higher in the case of a greater densi-ty of AMPA receptor channels, but not four times higher(not shown). As a result, the repeated binding of gluta-mate molecules per individual AMPA receptor (and therepeated opening of AMPA channels) is less rather thanmore likely to occur.
Occupancy of desensitized states duringand after a single mEPSC
The occupancy of all desensitized states (during and af-ter a single mEPSC) was found to be concentration de-pendent. Generally, and not surprisingly, state seven isoccupied the most at low concentrations of glutamate. Asthe glutamate concentration increases, the occupancy ofstates five and six becomes more important. Maximal de-sensitization (defined as the maximal occupancy of alldesensitized states) is strongly dependent on N, but evenfor a relatively low N a very high level of desensitizationof AMPA channels is reached. The time course of desen-sitization recovery appears to be monoexponential andslow (with a duration that increases as N increases) andis similar to the time course of the paired pulse depres-sion reported recently [14, 25]. Progressively greater oc-cupancy of states six and five is responsible for this pro-longation (their conversion to nondesensitized states andultimately to the unbound state is slower than that for de-sensitized state seven).
Deactivation and desensitization of patch currents
Earlier experimental studies have shown that, for a pro-longed application of glutamate pulses, the time constantof decrease of the macroscopic patch currents (a standardmeasure of desensitization) is essentially concentrationindependent [7]. Our simulations of patch currents dur-ing 2-ms glutamate pulses (2 ms is found to be longenough because the kinetics of gating of AMPA channelsis fast due to the high temperature assumed in the presentsimulation) show a similar concentration independence.This is not unexpected since the kinetic model of AMPAchannel gating is as given by Jonas et al. [17], but itserves as additional evidence that our simulations pro-vide an adequate means for assessing the synaptic andpatch currents. Furthermore, the time course of the de-crease of the current flow through the open channels ofthe simulated macroscopic patch currents following ashort (0.125 ms) glutamate pulse (a standard measure ofdeactivation) was found to be essentially concentrationindependent when the rates of desensitization are atphysiological levels. A modest concentration dependenceis observed when desensitization is abolished.
The agreement between the simulated synaptic cur-rents and the response to short (0.125 ms) glutamatepulses is good when: (1) the synaptic currents resultfrom the release of low number of molecules, and (2) de-sensitization is at a physiological level. However, when-ever the repeated opening of AMPA channels due to therepeated binding of glutamate molecules becomes im-portant, either because of the greater buffering of gluta-mate molecules in the synaptic cleft, or an elevated occu-pancy of the activatable bound states or both (as occursduring reduced or no desensitization), the fast applica-tion protocol becomes inadequate for reproducing thesynaptic currents. Therefore, any conclusion about thesynaptic currents drawn from patch currents should betreated with great caution. Thus, this simulation alsoshows that the time course of deactivation is not insensi-tive to the level of desensitization, emphasizing the needfor caution in using a fast application protocol. Neverthe-less, note that the conclusions reached in our simulationsof the synaptic currents depend on the kinetic schemeused, and might not hold for other combinations of rateconstants or receptor state diagrams.
Finally, good agreement between the theoretical cal-culations of how a reduction or abolition of desensitiza-tion affects the amplitude of mEPSCs, τd and their rela-tionship and the actual experimental findings suggeststhat the experimental findings are due to the events atsynapses with the same type of AMPA channels. It isthus not necessary to postulate the existence of AMPAchannels with different single-channel conductance,mean open time or sensitivity to desensitization at diffe-rent synapses. This is an important point since recent ev-idence indicates that the functional properties of somaticand dendritic glutamate receptors in the CA3 mossy fibresynapses are similar [20]. We did not explore the effectof changes in desensitization on rise times since through-out this study we assumed that release occurs from an in-stantaneous point source. The release from a vesiclethrough a fusion pore is not likely to have an importanteffect on τd but is expected to be a very important factorin determining the duration of τr. In conclusion, thisstudy helps to reconcile the fact that in control record-ings the time constant of decay of mEPSCs is essentiallythe same as that of the deactivation, and the growing evi-dence that desensitization plays a major role in shapingthe time course of mEPSCs.
&p.2:Acknowledgements Drs. K. Krnjevic, M. Mackey and L. Glassread the manuscript and made valuable comments. Supported bythe Medical Research Council of Canada (M.I.G.).
References
1. Atassi B, Godfraind J-M, Glavinovi´c MI (1996) Kinetics ofspontaneous miniature excitatory postsynaptic currents inpresence and in absence of cyclothiazide. Soc Neurosci Abstr26: 1283p
2. Barbour B, Keller BU, Llano I, Marty A (1994) Prolongedpresence of glutamate during excitatory synaptic transmissionto cerebellar Purkinje cells. Neuron 12:1331–1343
201

3. Bartol TM Jr, Land BR, Salpeter EE, Salpeter MM (1991) Mon-te Carlo simulation of miniature endplate current generation inthe vertebrate neuromuscular junction. Biophys J 59:1290–1307
4. Bekkers JM, Stevens CF (1996) Cable properties of culturedhippocampal neurons determined from sucrose-evoked minia-ture EPSCs. J Neurophysiol 75: 1250–1255
5. Bekkers JM, Richerson GB, Stevens CF (1990) Origin of vari-ability in quantal size in cultured hippocampal neurons andhippocampal slices. Proc Natl Acad Sci USA 87:5359–5362
6. Clements JD, Lester RAJ, Tong G, Jahr CE, Westbrook GL(1992) The time course of glutamate in the synaptic cleft. Sci-ence 258:1498–1501
7. Colquhoun D, Jonas P, Sakmann B (1992) Action of briefpulses of glutamate on AMPA/kainate receptors in patchesfrom different neurones of rat hippocampal slices. J Physiol(Lond) 458:261–287
8. Edmonds B, Gibb AJ, Colquhoun D (1995) Mechanisms ofactivation of glutamate receptors and the time course of excita-tory synaptic currents. Annu Rev Physiol 57:495–519
9. Edwards FA, Konnerth A, Sakmann B (1990) Quantal synaptictransmission in the central nervous system; a patch clampstudy of IPSCs in rat hippocampal slices. (Appendix by C.Busch). J Physiol (Lond) 430:231–249
10. Ghamari M, Glavinovi´c MI (1995) Quantitative analysis ofspontaneous excitatory quantal events in hippocampal pyrami-dal neurones. Can J Physiol Pharmacol 73:9p
11. Harris KM, Landis DMD (1986) Membrane structure at syn-aptic junctions in area CA1 of the rat hippocampus. Neurosci-ence 19:857–872
12. Hestrin S, Sah P, Nicoll R (1990) Mechanisms generating thetime course of dual component excitatory synaptic currents re-corded in hippocampal slices. Neuron 5:247–253
13. Hille B (1992) Ionic channels of excitable membranes. Sinau-er, Sutherland, Mass.
14. Issacson JS, Walmsley B (1996) Amplitude and time course ofspontaneous and evoked excitatory postsynaptic currents inbushy cells of the anteroventral cochlear nucleus. J Neuro-physiol 76:1566–1571
15. Johnston D, Brown TH (1983) Interpretation of voltage-clampmeasurements in hippocampal neurons. J Neurophysiol 50:464–486
16. Jonas P, Spruston N (1994) Mechanisms shaping glutamate-mediated excitatory postsynaptic currents in the CNS. CurrOpin Neurobiol 4:366–372
17. Jonas P, Major G, Sakmann B (1993) Quantal components ofunitary EPSCs at the mossy fibre synapse on CA3 pyramidalcells of rat hippocampus. J Physiol (Lond) 472:615–663
18. Longsworth LG (1953) Diffusion measurements at 25°C, ofaqueous solutions of amino acids, peptides and sugars. J AmChem Soc 75:5705–5709
19. Raman IM, Trussell LO (1992) The kinetics of the response toglutamate and kainate in neurons of the avian cochlear nucle-us. Neuron 9:173–186
20. Spruston N, Jonas P, Sakmann B (1995) Dendritic glutamatereceptor channels in rat hippocampal CA3 and CA1 pyramidalneurons. J Physiol (Lond) 482:325–352
21. Takahashi M, Sarantis M, Attwell D (1996) Postsynaptic glu-tamate uptake in rat cerebellar Purkinje cells. J Physiol (Lond)497:523–530
22. Tang C-M, Shi Q-Y, Katchman A, Lynch G (1991) Modulationof the time course of fast EPSCs and glutamate channel kinet-ics by Aniracetam. Science 254:288–290
23. Tong G, Jahr CE (1994) Multivesicular release from excitatorysynapses of cultured hippocampal neurons. Neuron 12:51–59
24. Trussell LO, Fischbach GD (1989) Glutamate receptor desen-sitization and its role in synaptic transmission. Neuron 3:209–218
25. Trussell LO, Zhang S, Raman IM (1993) Desensitization ofAMPA receptors upon multiquantal neurotransmitter release.Neuron 10: 1185–1196
26. Wahl LM, Pouzat C, Stratford KJ (1996) Monte Carlo simula-tion of fast excitatory synaptic transmission at a hippocampalsynapse. J Neurophysiol 75:597–608
27. Wathey JC, Nass, MM, Lester HA (1979) Numerical recon-struction of the quantal event at nicotinic synapses. Biophys J27:145–164
&/fig.c:
202
Copyright © 2022 FDOKUMEN




















