
Molecular and phylogenetic characterization of cytochromes c from Haemonchus contortus and...
9
Molecular and phylogenetic characterization of cytochromes c from Haemonchus contortus and Trichostrongylus vitrinus (Nematoda: Trichostrongylida) ☆ B.E. Campbell a, ⁎, A.J. Nisbet b , J. Mulvenna c , A. Loukas c , R.B. Gasser a, ⁎ a Department of Veterinary Science, The University of Melbourne, Werribee, Victoria 3030, Australia b Parasitology Division, Moredun Research Institute, Penicuik, EH26 OPZ, UK c Queensland Institute of Medical Research, Herston, Queensland 4006, Australia abstract article info Article history: Received 1 May 2008 Received in revised form 11 June 2008 Accepted 16 July 2008 Available online 30 July 2008 Received by M. Di Giulio Keywords: Cytochrome c Nematodes Parasites Haemonchus contortus Trichostrongylus vitrinus Livestock Although cytochrome c genes (cyt c) and proteins (CYT C) have been relatively well studied in mammals, very little is known about them in parasitic helminths. In the present study, we investigated this group of molecules in Haemonchus contortus (barber's pole worm) and Trichostrongylus vitrinus (black scour worm), two parasitic nematodes of small ruminants. The cyt c gene (512 bp) of H. contortus had one intron and encoded a transcript of 345 nucleotides, whilst that of T. vitrinus (792 bp) had two introns and encoded a transcript of 360 nucleotides. The transcription of cyt c in T. vitrinus was substantially greater in adult males compared with females, although no such gender-enrichment was evident in adults of H. contortus. These findings were supported at the protein level by immunoblot analyses. The inferred proteins (designated Hc- CYT C and Tv-CYT C, respectively) shared nucleotide and amino acid identities of 78% and 85%, respectively. The alignment of these and other CYT C sequences from nematodes, flatworms, insects and mammals identified conserved motifs associated with CYT C oxidase- and reductase- as well as haem-binding. One residue (histidine-26) was conserved for mammals, whereas this residue was absent from all nematodes; the functional significance of this difference is not yet known. Both phylogenetic analysis and protein modelling revealed that CYT C proteins of nematodes are structurally distinct from those of mammals and other organisms, suggesting their potential as targets for parasite intervention. © 2008 Elsevier B.V. All rights reserved. 1. Introduction Cytochrome c (CYT C) proteins are involved in key biological processes, including respiration and apoptosis. These proteins are associated with the mitochondrial membrane in both prokaryotes and eukaryotes (Virbasius and Scarpulla, 1988; Jiang and Wang, 2004). CYT C is involved in electron transport during respiration, interacting with other proteins via oxidation (aerobic) and reduction (anaerobic) of the iron atoms within the haem-binding part of the molecule (Takamiya et al., 1996a). In mammals and insects, CYT C proteins also play a key role in programmed cell death (apoptosis) (Chinnaiyan, 1999; Adrain and Martin, 2001; Ho and Zacksenhaus, 2004; Arama et al., 2006). CYT C proteins described from a range of taxa possess well- conserved functional domains (Vanfleteren et al., 1990; Allen et al., 2005), and multiple CYT C genes and isoforms have been identified. Some of these isoforms are expressed in somatic tissue, whilst testis-specific isoforms have been described from mammals and insects (Kim and Sabourin, 1986; Virbasius and Scarpulla, 1988; Narisawa et al., 2002; Schlecht et al., 2004; Arama et al., 2006). Although tissue-specific CYT C proteins exist, there is evidence from Drosophila melanogaster (vinegar fly) that the different proteins can be the same functionally (Arama et al., 2006). To date, two cyt c genes (cyc-2.1 and cyc-2.2) have been characterized in the free-living nematode Caenorhabditis elegans, although, in contrast to mammals, neither CYT C has been found to be testis-specific or involved in apoptosis (Chinnaiyan, 1999; WormBase: www.wormbase.org). In C. elegans, ‘loss-of-function’ phenotypes following gene silencing (RNA interference, RNAi) indicate that these proteins are essential for reproduction and development (www.wormbase.org). In contrast, very little is known about CYT Cs in parasitic nematodes. In Ascaris suum, the common roundworm of pigs, two CYT Cs have been identified, namely type 1, which is expressed in third-stage larvae (L3s) and adults, and type 2, which is specific to adults (Takamiya et al., 1996a,b). Kinetic studies have revealed species- specific differences between the CYT C proteins of A. suum and those of cattle in their interactions with CYT C oxidase (Takamiya et al., 1996b). Also, Nisbet and Gasser (2004) demonstrated by microarray Gene 424 (2008) 121–129 Abbreviations: CYT C or cyt c, cytochrome c; Hc, Haemonchus contortus; Tv, Tri- chostrongylus vitrinus; EST, expressed sequence tag; L3, third-stage larva; PCR, polymerase chain reaction; RT-PCR, reverse transcription polymerase chain reaction; DTT, dithiothreitol; RNAi, double-stranded RNA interference. ☆ The nucleotide sequence data reported in this paper are available in the EMBL and GenBank databases under accession nos. EU679404, EU679405, AJ616633 and FJ147586. ⁎ Corresponding authors. Tel.: +61 3 9731 2000; fax: +61 3 9731 2366. E-mail addresses: [email protected] (B.E. Campbell), [email protected] (R.B. Gasser). 0378-1119/$ – see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.gene.2008.07.028 Contents lists available at ScienceDirect Gene journal homepage: www.elsevier.com/locate/gene
Transcript of Molecular and phylogenetic characterization of cytochromes c from Haemonchus contortus and...

Gene 424 (2008) 121–129
Contents lists available at ScienceDirect
Gene
j ourna l homepage: www.e lsev ie r.com/ locate /gene
Molecular and phylogenetic characterization of cytochromes c from Haemonchuscontortus and Trichostrongylus vitrinus (Nematoda: Trichostrongylida)☆
B.E. Campbell a,⁎, A.J. Nisbet b, J. Mulvenna c, A. Loukas c, R.B. Gasser a,⁎a Department of Veterinary Science, The University of Melbourne, Werribee, Victoria 3030, Australiab Parasitology Division, Moredun Research Institute, Penicuik, EH26 OPZ, UKc Queensland Institute of Medical Research, Herston, Queensland 4006, Australia
Abbreviations: CYT C or cyt c, cytochrome c; Hc, Hchostrongylus vitrinus; EST, expressed sequence tagpolymerase chain reaction; RT-PCR, reverse transcriptioDTT, dithiothreitol; RNAi, double-stranded RNA interfer☆ The nucleotide sequence data reported in this paperGenBank databases under accession nos. EU679404, EU67⁎ Corresponding authors. Tel.: +61 3 9731 2000; fax:
E-mail addresses: [email protected] (B.E. Campb(R.B. Gasser).
0378-1119/$ – see front matter © 2008 Elsevier B.V. Alldoi:10.1016/j.gene.2008.07.028
a b s t r a c t
a r t i c l e i n f oArticle history:
Although cytochrome c gen Received 1 May 2008Received in revised form 11 June 2008Accepted 16 July 2008Available online 30 July 2008Received by M. Di Giulio
Keywords:Cytochrome cNematodesParasitesHaemonchus contortusTrichostrongylus vitrinusLivestock
es (cyt c) and proteins (CYT C) have been relatively well studied in mammals, verylittle is known about them in parasitic helminths. In the present study, we investigated this group ofmolecules in Haemonchus contortus (barber's pole worm) and Trichostrongylus vitrinus (black scour worm),two parasitic nematodes of small ruminants. The cyt c gene (512 bp) of H. contortus had one intron andencoded a transcript of 345 nucleotides, whilst that of T. vitrinus (792 bp) had two introns and encoded atranscript of 360 nucleotides. The transcription of cyt c in T. vitrinus was substantially greater in adult malescompared with females, although no such gender-enrichment was evident in adults of H. contortus. Thesefindings were supported at the protein level by immunoblot analyses. The inferred proteins (designated Hc-CYT C and Tv-CYT C, respectively) shared nucleotide and amino acid identities of 78% and 85%, respectively.The alignment of these and other CYT C sequences from nematodes, flatworms, insects and mammalsidentified conserved motifs associated with CYT C oxidase- and reductase- as well as haem-binding. Oneresidue (histidine-26) was conserved for mammals, whereas this residue was absent from all nematodes; thefunctional significance of this difference is not yet known. Both phylogenetic analysis and protein modellingrevealed that CYT C proteins of nematodes are structurally distinct from those of mammals and otherorganisms, suggesting their potential as targets for parasite intervention.
© 2008 Elsevier B.V. All rights reserved.
1. Introduction
Cytochrome c (CYT C) proteins are involved in key biologicalprocesses, including respiration and apoptosis. These proteins areassociated with the mitochondrial membrane in both prokaryotes andeukaryotes (Virbasius and Scarpulla,1988; Jiang andWang, 2004). CYTC is involved in electron transport during respiration, interacting withother proteins via oxidation (aerobic) and reduction (anaerobic) of theiron atomswithin the haem-binding part of themolecule (Takamiya etal., 1996a). In mammals and insects, CYT C proteins also play a key rolein programmed cell death (apoptosis) (Chinnaiyan, 1999; Adrain andMartin, 2001; Ho and Zacksenhaus, 2004; Arama et al., 2006).
CYT C proteins described from a range of taxa possess well-conserved functional domains (Vanfleteren et al., 1990; Allen et al.,
aemonchus contortus; Tv, Tri-; L3, third-stage larva; PCR,n polymerase chain reaction;ence.are available in the EMBL and9405, AJ616633 and FJ147586.
+61 3 9731 2366.ell), [email protected]
rights reserved.
2005), and multiple CYT C genes and isoforms have been identified.Some of these isoforms are expressed in somatic tissue, whilsttestis-specific isoforms have been described from mammals andinsects (Kim and Sabourin, 1986; Virbasius and Scarpulla, 1988;Narisawa et al., 2002; Schlecht et al., 2004; Arama et al., 2006).Although tissue-specific CYT C proteins exist, there is evidence fromDrosophila melanogaster (vinegar fly) that the different proteins canbe the same functionally (Arama et al., 2006). To date, two cyt cgenes (cyc-2.1 and cyc-2.2) have been characterized in the free-livingnematode Caenorhabditis elegans, although, in contrast to mammals,neither CYT C has been found to be testis-specific or involved inapoptosis (Chinnaiyan, 1999; WormBase: www.wormbase.org). In C.elegans, ‘loss-of-function’ phenotypes following gene silencing (RNAinterference, RNAi) indicate that these proteins are essential forreproduction and development (www.wormbase.org). In contrast,very little is known about CYT Cs in parasitic nematodes.
In Ascaris suum, the common roundworm of pigs, two CYT Cs havebeen identified, namely type 1, which is expressed in third-stagelarvae (L3s) and adults, and type 2, which is specific to adults(Takamiya et al., 1996a,b). Kinetic studies have revealed species-specific differences between the CYT C proteins of A. suum and thoseof cattle in their interactions with CYT C oxidase (Takamiya et al.,1996b). Also, Nisbet and Gasser (2004) demonstrated by microarray

122 B.E. Campbell et al. / Gene 424 (2008) 121–129
analysis that the transcription of a cyt c gene, linked to the expressedsequence tag (EST) TVm07_A08 (accession no. AJ616633), was 4.5-foldgreater in adult males than females of the trichostrongylid nematodeTrichostrongylus vitrinus, an economically important parasite of smallruminants. Extending this work, the aims of the present study were to(i) isolate and characterize corresponding full-length cyt c genes fromT. vitrinus and the related, blood-feeding trichostrongylid Haemonchuscontortus, (ii) to investigate the transcription of these genes and theirproducts in adult male and female nematodes and (iii) compare theproteins phylogenetically and structurally with those available incurrent databases from a range of organisms. Also, comparisons weremade to CYT Cs from the free-living nematode C. elegans to assess therole(s) of these proteins in nematode survival, development andreproduction, thus adding to the paucity of information presentlyavailable on these molecules in nematodes.
2. Materials and methods
2.1. Parasite material
Merino lambs (8–12 weeks of age), maintained under helminth-free conditions, were inoculated intra-ruminally with 10,000 or40,000 infective third-stage larvae (L3) of H. contortus or T. vitrinus(Animal Ethics Approval Number 03235; The University of Mel-bourne). The patency of the infection (∼21–25 days) was establishedbased on the detection of strongylid eggs in the faeces using theMcMaster flotation method (MAFF, 1977). For the collection of adultworms, infected lambs were euthanized with an overdose ofpentobarbitone sodium (Lethobarb, Virbac Pty. Ltd.), administeredintravenously 30 days after inoculation. Adult wormswere immediatelycollected (using fine forceps) from the abomasum (H. contortus) or fromthe first 4 m of the small intestine (T. vitrinus), washed extensively inphosphate-buffered saline (PBS; pH 7.4), male and female worms wereseparated, washed again and then snap frozen in liquid nitrogen forsubsequent storage at −70 °C.
2.2. Isolation, purification, treatment and storage of nucleic acids
Total genomic DNAwas extracted from ∼100 mg of worms using asmall-scale sodium dodecyl sulphate (SDS)/proteinase K extractionprocedure (Gasser et al., 1993), followed by purification over a mini-column (Wizard™ DNA Clean-Up, Promega). Total RNA was isolatedfrom each sex ofH. contortus and T. vitrinus (following homogenizationin liquid nitrogen using a mortar and pestle) employing the TriPure®
isolation reagent, according to the manufacturer's protocol (RocheMolecular Biochemicals). Amounts of nucleic acid were measuredspectrophotometrically (NanoDrop ND-1000, NanoDrop Technolo-gies). DNA and RNA samples were both stored at −70 °C.
2.3. Cloning and sequencing
The polymerase chain reaction (PCR)-based amplification of cyt cfrom first-strand cDNA (1 μl) or genomic DNA (100 ng) wasperformed using the Phusion High-fidelity DNA polymerase enzyme(Finnzymes). Reactions consisted of 1× HF reaction buffer, 2 pmol ofeach primer, 0.2 mM of dNTPs and 1 U of the DNA polymerase in avolume of 50 μl. Based on the T. vitrinus EST TVm07_A08 (accessionno. AJ616633; Nisbet and Gasser, 2004), the primers CYT C—F(forward; 5′-ATGGGTAAAGGATCTGATTCAGGAAG-3′ and CYT C—R(reverse; 5′-AACTTTCACGGTTCTTCGGTAGAATT-3′), each with a melt-ing temperature (Tm) of 56 °C, were designed. Cycling conditions usedwere: initial denaturation of 98 °C/30 s; then 30 cycles of 98 °C/10 s,55 °C/30 s, 72 °C/2 min, with a final extension of 72 °C/10 min, beforebeing held at 4 °C. The EST TVm07_A08, cloned in the plasmid vectorpGEM®-T Easy (Promega), was used as a positive control. Productswere resolved in agarose gels (2%), stained for 10 min in ethidium
bromide (10 μ/ml), detected on a transilluminator and photographeddigitally.
Phusion High-fidelity DNA polymerase produces blunt-endedproducts, which require 3′ A-tailing prior to ligation into pGEM®-TEasy. In brief, this reaction was performed using Taq DNA polymerase(Promega). The reaction consisted of 5 μl of amplicon, 1 μl of 10× PCRbuffer, 0.6 μl of 25 mM MgCl2, 0.2 μl of 0.2 mM dATP, 1 μl of Taq DNApolymerase (5 U/μl) and 2.2 μl of H2O. The reaction was incubated at70 °C for 30 min, the products then precipitated with isopropanol andresuspended in 5 μl of H2O. The 3′ A-tailed product was ligated intopGEM®-T Easy, and Escherichia coli (strain JM109; Promega) (108 colonyforming units/μg) transformed with recombinant plasmids using theheat shockmethod (Hanahan et al., 1991) and grown overnight at 37 °Con Luria Bertani (LB) agar plates containing 10 mg/ml ampicillin.Colonies were screened by PCR using Go-Taq Flexi DNA polymerase(Promega), according to the manufacturer's instructions using theprimers CYT C—F and CYT C—R and 4 mM MgCl2 (final concentration),employing the following thermocycling conditions: 95 °C/2min, followedby30cyclesof 95 °C/30 s; 55 °C/30 s; 72 °C/1min, followedby72 °C/5min.Single colonieswere isolated and suspended in 20 μl of H20, and 2 μl usedas a template in the PCR to screen for inserts. Bacterial cultures weregrownovernight from the remainder of the suspension inoculated into LBbroth (containing 10 mg/ml ampicillin) at 37 °C, shaking. Plasmid DNAwas isolated from recombinant clones and purified from the overnightcultures using Wizard® Miniprep columns (Promega). Inserts weresequenced inbothdirectionsusingprimers T7andSP6 (topGEM®-TEasy,Promega), employingBigDyeTerminator v.3.1 chemistry in an automatedABI-PRISM sequencer (Applied Biosystems).
2.4. Bioinformatic analyses
The cDNA sequences were subjected to BLASTx analysis usingprograms in the National Center for Biotechnology Information(www.ncbi.nlm.nih.gov/BLAST) and the parasite genome BLASTserver from the European Bioinformatics Institute (www.ebi.ac.uk/parasites/parasite-genome) to confirm that they coded for CYT Cproteins. The manual alignment of the cDNAs and correspondinggenomic DNAs for individual species was used to determine intron/exon boundaries (AG–GT rule, Breathnach and Chambon, 1981). Thesequences were then conceptually translated using the translateoption in the Molecular Toolkit (www.vivo.colostate.edu/molkit/index). The protein sequences were then aligned with published CYTC sequences (Vanfleteren et al., 1990; Allen et al., 2005) to identify theCYT C oxidase- and reductase- as well as haem-binding domains. Thesesequences were also analysed using ProSite to verify the presence of thecharacteristic motifs within each protein (www.expasy.ch/tools/scnpsit1.html). Information on the C. elegans homologues and interac-tion datawere also retrieved fromWormBase (www.wormbase.org) andWormNet (www.functionalnet.org/wormnet/). These databases pro-vide information about gene ontology, expression, inferred interac-tions and/or function(s) in C. elegans. The genetic interaction data arebased on naïve Bayesian analysis of data from a range of eukaryoticorganisms (Lee et al., 2008). The cDNA sequences predicted to encodeCYT Cs of nematodes were subjected to BLASTx and BLASTn analysesagainst the sheep ESTand genomicDNAdatabases (www.ncbi.nlm.nih.gov/projects/genome/guide/sheep/) to ensure that they were notderived from host tissue.
Full-length (CYT C) protein sequences representing a number ofnematodes, insects and trematodes and mammals from existingdatabases (www.ncbi.nlm.nih.gov, www.ebi.ac.uk; see Table 1) werealigned using CLUSTALX (ftp://ftp.ebi.ac.uk/pub/software/clustalw2;Thompson et al., 1997), adjusted manually and saved as a nxs file.The phylogenetic analysis of these data were performed using twoalgorithms, employing the CYT C of the yeast Pichia stipitis as theoutgroup. Bayesian inference (BI) was carried out using the softwarepackage MrBayes 3.1.2 (mrbayes.csit.fsu.edu/index.php); probabilities

Table 1Selected full-length cytochrome c amino acid sequences (and relevant information from BLASTx analysis) used for the analysis of phylogenetic relationship to cytochrome c fromHaemonchus contortus (Hc-CYT C) and Trichostrongylus vitrinus (Tv-CYT C)
Group Species Accession no. Length % amino acid identityto Hc-CYT C
% amino acid identityto Tv-CYT C
Nematode Haemonchus contortus EU679404 114 85Trichostrongylus vitrinus AJ616633 119 85Ancylostoma caninum BG438360 112 72 83Ancylostoma ceylanicum CB17591 112 72 83Ascaris suum D73391 112 68 78Ascaris suum P92504 109 59 61Brugia malayi EDP34055 108 72 80Caenorhabditis briggsae CBG017 112 69 79Caenorhabditis briggsae CBG085 129 71 77Caenorhabditis elegans E04A4.7 111 71 81Caenorhabditis elegans ZC116.2 123 71 75Necator americanus BU086533.1 111 72 82Nippostrongylus brasiliensis EB184981 109 74 82Onchocerca volvulus BE132516 108 73 80Pratylenchus penetrans BQ627045 109 72 80Strongyloides stercoralis BF015012 111 72 81Teladorsagia circumcincta CB036274 119 78 92Teladorsagia circumcincta CB042978.1 112 73 81Toxocara canis CD571677 113 70 76
Platyhelminth Schistosoma japonicum AAP06143.1 108 52 55Schistosoma mansoni ABG21809.1 108 50 51
Protozoa Leishmania braziliensis XP001563738.1 114 48 50Leishmania braziliensis XP001563739.1 113 47 50Leishmania infantum XP001464620.1 113 48 52Leishmania major XP001682212.1 113 48 50Plasmodium falciparum XP001348211.1 115 50 56Trypanosoma brucei XP847342.1 114 47 49Trypanosoma cruzi XP809186.1 114 50 51
Arthropods Drosophila melanogaster AAF53553 (testis-specific) 105 53 56Drosophila melanogaster AAF53554 108 54 62
Mammal Bos taurus NP001039526.1 105 55 60Bos taurus NP001071431.1 (testis-specific) 105 56 55Capra hircus ABA06540 105 55 61Equus caballus XP001500681.1 (testis-specific) 105 58 60Homo sapiens AAP36314.1 106 53 54Monodelphis domestica XP001377302.1 (testis-specific) 104 54 53Mus musculus NP031834.1 105 55 61Mus musculus NP034119.1 (testis-specific) 105 57 60Oryctolagus cuniculus P00008 105 55 60Ovis aries P62896 105 55 60Rattus norvegicus NP036972.1 (testis-specific) 105 57 60Rattus norvegicus XP001075173.1 105 55 61Sus scrofa P62895 105 55 60
Yeast Pichia stipitis XP001384941.1 110 50 46
123B.E. Campbell et al. / Gene 424 (2008) 121–129
were calculated using200,000 generations (ngen=200,000burnin=20),employing four simultaneous tree-building chains (nchains=4), savingevery 100th tree (samplefreq=100). Theheuristic neighbour-joining (NJ)analysis was performed employing PAUP⁎ v4.0b10 (Swofford, 1999)using the absolute difference model (2000 replications) with thealignment gaps treated as missing data. Using the program MODELLER(Fiser and Sali, 2003), the three-dimensional structures of the CYT Csfrom H. contortus and T. vitrinus were predicted based on comparativemodelling to rice ferricytochrome c (PDB ID 1CCR; Ochi et al. 1983) andbovine heart CYT C (PDB ID 2B4Z; Mirkin et al. unpublished data; seewww.rcsb.org/pdb/), respectively. The covalent geometry of the mod-elled structures was in agreement with the template structures, with91.5% (H. contortus) and 92.1% (T. vitrinus) of the residues occupying themost favoured region of the Ramachandran plot. The quality of thestereochemical structures of the models was determined using PRO-CHECK-NMR (Laskowski et al., 1996).
2.5. First-strand synthesis of cDNA and reverse-transcriptionPCR (RT-PCR) of cyt c
First-strand cDNA synthesis was performed using the SuperScriptReverse Transcriptase II kit (Invitrogen). In brief, 1 μg of total RNA
and 0.5 μg of oligo d(T) n=12–18 primer (0.5 μg/μl), in a totalreaction volume of 12 μl, were incubated at 70 °C for 10 min andthen chilled on ice. Subsequently, 4 μl of first-strand cDNA buffer(250 mM Tris–Cl, pH 8.3, 375 mM KCl, 15 mM MgCl2), 2 μl of 0.1 Mdithiothreitol (DTT) and 1 μl of 10 mM dNTPs were added. Thereactions were then incubated at 42 °C for 2 min, and 200 U (1 μl)of SuperScript II added. The contents were mixed and incubated at25 °C for 10 min, then at 42 °C for 50 min, and the reactionsterminated by inactivating the enzyme at 70 °C for 15 min. Usingthe first-strand cDNA as a template, RT-PCR, utilizing Go-Taq FlexiDNA polymerase (Promega), was performed to assess the levels oftranscription of cyt c in adult males and females of bothH. contortus andT. vitrinus. The conditions and primers used were as described insubsection2.3. A187bp fragmentof the large subunitof nuclear ribosomalRNA (28S) genewas used as a reference control in the PCR employing theprimers 28S—F (forward; 5′-GCATAAGCTCTCGCGTTACC-3′) and 28S—R(reverse; 5′-GAGAGGGACAGCAGGTTCAG-3′). The T. vitrinus ESTTVm07_A08 in pGEM®-T Easy was used as a positive control inthe PCR. The experiment was repeated three times (with threereplicates per gene) on three separate days. Products were resolvedin agarose gels (2%), documented and sequenced as described insubsection 2.3.

Fig. 1. Diagrammatic representations of the genomic organization of cytochrome cgenes from Haemonchus contortus, Trichostrongylus vitrinus and Caenorhabditis elegans.The organization of the C. elegans gene was determined from the published nucleotidesequences from WormBase (www.wormbase.org). Black boxes represent exonsencoding protein. The lines joining the exons indicate introns.
124 B.E. Campbell et al. / Gene 424 (2008) 121–129
2.6. Subcloning, expression of recombinant proteins and immunoblotanalysis
The cDNAs amplified by PCR using primers CYT C—F and CYT C—Rand Phusion High-fidelity DNA polymerase were cloned blunt-endedinto the SmaI site of the glutathione S-transferase (GST) fusion proteinexpression vector pGEX-3TEX (GE Healthcare Life Sciences). To ensurethat the fragments were cloned in the correct frame, each putativepositive clone was sequenced with the primers pGEX 5′ (forward; 5′-GGGCTGGCAAGCCACGTTTGGTG-3′) and pGEX 3′ (reverse; 5′-CCG-GGAGCTGCATGTGTCAGAGG-3′) (GE Healthcare Life Sciences). The E.coli BL21 strain (Promega) was transformed (Hanahan et al., 1991) andgrown on LB plates containing 10 mg/ml ampicillin. Induced cloneswere screened for the expression of fusion protein. In brief, singlecolonies were inoculated into 5 ml of LB broth (containing 10 mg/mlampicillin) and grown at 37 °C, shaking, for 4 h. Following the removalof 1 ml aliquots from un-induced samples, protein expression wasinduced by the addition of 0.2 mM isopropyl-β-D-thiogalactopyrano-side (IPTG). Cellswere grown for a further 2 h at 37 °C, shaking. Aliquotsof 1 ml were centrifuged (10,000 g) at room temperature (22 °C) for2 min and the cell pellet, including the un-induced controls,resuspended in 200 μl of sodium dodecyl sulphate (SDS) sample buffercontaining 5% β-mercaptoethanol. Samples were heated at 95 °C for5minand then centrifuged. Individual supernatants (5 μl aliquots)wereseparated in 10% SDS polyacrylamide gels (SDS-PAGE), usingmid-rangemolecular weight protein markers (Promega) for size estimations.Proteins were transferred (20 V for 30 min) to polyvinylidene fluoride(PVDF) membrane in a semi-dry blotting cell (Bio-Rad). Following theprotein transfer, the lanes containing the markers were removed andstained with Ponceau S stain (Sigma-Aldrich) for 1 h at roomtemperature and then destained with 50% methanol. Lanes containingthe proteins to be detected were incubated overnight at 4 °C inphosphate-buffered saline (PBS) containing 0.1% Tween 20 (PBST) and5% skimmilk powder.Membraneswere incubatedwith affinity purifiedrabbit anti-GST IgG antibody (1:1000) for 1 h and then affinity purifiedgoat anti-rabbit (IgG antibody) horseradish peroxidase (HRP) conjugate(1: 5000; both antibodies from Sigma-Aldrich) for another hour, withfour PBST washes between each antibody step (1×15 min; 3×5 min).The HRP reaction was detected using the SuperSignal West PicoChemiluminescent substrate (Pierce Biotechnology), used according tothe instructions. Chemiluminescencewas detected by exposure of eachmembrane to autoradiographic film (Agfa Ultra UV-G) for 10–60 s.Aliquots of the expressed protein and un-induced controls were alsosubjected to the detection of CYT C by chemiluminescence (seesubsection 2.8).
2.7. Preparation of protein extracts and SDS-PAGE analysis
In order to conserve the haem bound to haem-binding proteins,such that CYT C could be detected, the protein preparations wereneither heated nor exposed to reducing agent (β-mercaptoethanol).Aliquots (100 mg) of adult males or females of each H. contortus andT. vitrinus were suspended in 50 μl of SDS sample buffer per 50 mgof worms in 1.5 ml microcentrifuge tubes. Worms were homo-genised using two freeze (liquid nitrogen)/thaw cycles on ice andvortexing with glass beads. Samples were centrifuged at 12,000 g for2 min at 4 °C. Haemolytic serum was prepared from sheep (host)blood. For electrophoresis and subsequent haem-binding proteindetection, a 10 μl aliquot of this serum was mixed with 10 μl of2×SDS sample buffer. Protein amounts were measured spectro-photometrically (NanoDrop ND-1000, NanoDrop Technologies) usingthe Bradford assay (in triplicate) (Bradford, 1976) and bovine serumalbumin (BSA) as the standard. Proteins (50 μg of each preparation),including 5 μl of mid-range molecular weight protein markers wereresolved by SDS-PAGE (15% gels) for ∼2 h at 100 V, transferred toPVDF membranes (subsection 2.6), washed in PBS for 10 min and
then used for the detection of haem-binding proteins (subsection2.8).
2.8. Biochemical detection of haem-binding proteins
Two methods, both detecting the haem-binding capacity of themolecule, were used to detect the presence of CYT C in protein extractsfrom adult nematodes and expressed fusion protein. These methodsfollowed Feissner et al. (2003), one being based on colorimetric andthe other on chemiluminescent detection. For the colorimetric assay,6.3 mM 3, 3′, 5, 5′-tetramethylbenzidine (TMBZ) was dissolved inmethanol immediately before staining. Three parts (30 ml) of TMBZstain were added to seven parts (70 ml) of 0.25 mM sodium acetate(pH 5.0) to prepare the stain. Membranes containing the wormproteins were incubated in the stain for 30 min at room temperaturein the dark. Subsequently, hydrogen peroxide (30%) was added to afinal concentration of 0.3%, and the membrane incubated for a further30 min. Staining of each membrane was documented by digitalphotography and each membrane air-dried. Chemiluminescentdetection of CYT C was achieved using the SuperSignal West PicoChemiluminescent substrate (Thermo Scientific), by incubating themembrane in the substrate for 5 min at room temperature. Thissubstrate detects the haem bound to CYT C. Membranes were exposedto autoradiographic film (Agfa Ultra UV-G) for 5–60 s.
3. Results and discussion
3.1. Characterisation and classification of CYT C from H. contortus andT. vitrinus
The full-length genes coding for CYT C isolated from adult males ofH. contortus and T. vitrinus were 512 and 792 nucleotides in length,respectively. The H. contortus gene, encoding a transcript of 345nucleotides, had one intron, whereas the T. vitrinus gene had twointrons and encoded a transcript of 360 nucleotides (Fig. 1). Theconceptually translated proteins (designated Hc-CYT C and Tv-CYT C)were 114 and 119 amino acids in length for H. contortus and T. vitrinus,respectively. Hc-CYT C and Tv-CYT C shared 78% and 85% identity atthe nucleotide and amino acid levels, respectively. Comparisons of Hc-CYT C and Tv-CYT C were made (via BLASTx, National Center forBiotechnology Information www.ncbi.nlm.nih.gov) with related pro-teins from selected organisms, such as other nematodes (including A.suum, Teladorsagia circumcincta, C. elegans and C. briggsae), platyhel-minths, arthropods and mammals (see Table 1). Hc-CYT C and Tv-CYTC had amino acid identities of 59–83% to related molecules from othernematodes, both showing the highest homology to proteins inferredfrom the genes cyc-2.1 (E04A4.7) and/or cyc-2.2 (ZC116.2) of C. elegans

125B.E. Campbell et al. / Gene 424 (2008) 121–129
(cf. www.wormbase.org/). Lower identities were recorded to CYT Csfrom a number of platyhelminths (50–55%), protozoa (47–56%),mammals (55–61%), arthropods (53–62%) and yeasts (46–50%)(Table 1).
In spite of conservation in the functional domains among the CYT Cproteins from the different species (Table 1), there was considerablesequence divergence among taxa in other regions. Based on thephylogenetic analysis of amino acid sequence data for a range ofmetazoans (Table 1), CYT C proteins grouped broadly into those ofnematodes, platyhelminths (blood flukes of the genus Schistosoma),protozoa (species of Plasmodium, Leishmania and Trypanosoma),insects and mammals (nodal support of 0.95–1.0 for BI; see Fig. 2).The CYT Cs of nematodes clustered (nodal support of 0.99) to theexclusion of CYT Cs from all other metazoan groups. Within the‘nematode group’, proteins Hc-CYT C and Tv-CYT C clustered with amolecule (accession no. CB036274) from another trichostrongylidnematode (T. circumcincta). The analysis also revealed a grouping ofthe three trichostrongylid CYT Cs with CYC-2.2 from both C. elegans(ZC116.2) and C. briggsae (Fig. 2; cf. Table 1). Interestingly, selected CYTCs from filarial nematodes (Onchocerca volvulus and Brugia malayi)grouped (nodal support of 0.96) separately from those of othernematodes representing the orders Strongylida, Ascaridida andRhabditida. This unique grouping might reflect the biology and lifecycles of filarial nematodes, which involve arthropod intermediatehosts, rather than being monoxenous (direct) (as is the case for manynematodes of the other orders) (see Bowman, 1995).
Fig. 2. Phylogenetic tree constructed following analysis by Bayesian inference of amino acidother metazoan species (cf. Table 1) for which full-length peptide sequences were availabA cytochrome c sequence from the yeast Pichia stipitis was used as an outgroup. Nodal supp
Within the nematode group of CYT Cs, protein CYC-2.1 (E04A4.7)from C. elegans clustered with molecules from selected strongylids(Ancylostoma spp., Necator americanus, Nippostrongylus brasiliensisand T. circumcincta), an ascaridoid (A. suum) and a plant parasiticnematode (Pratylenchus penetrans). Teladorsagia circumcincta, A. suum,C. elegans and C. briggsae each had two CYT C proteins (see Fig. 2) thatlikely represent distinct isoforms. Given that two types of CYT C (i.e.,testis-specific and somatic) occur in some species of mammals (Kimand Sabourin, 1986; Virbasius and Scarpulla, 1988; Narisawa et al.2002) and insects (Drosophila melanogaster; Arama et al., 2006),separate phylogenetic analyses were conducted to establish whetherany of the CYT C proteins from nematodes grouped together witheither of these types from selected mammals or insects. The analysesrevealed clearly that all nematode CYT Cs clustered (with nodalsupport of 0.99) to the exclusion of both testis-specific and somaticCYT Cs from other animals (data not shown). Overall, there wasconcordance between the trees constructed using the BI and NJmethods, with minor variation in the position of some branches (notshown).
Although there is presently a paucity of information on CYT Cs ofparasitic nematodes, considerable data are available for the free-living nematode C. elegans. Tv-CYT C was most similar in sequence(81%) to that encoded by the gene cyc-2.1 of C. elegans (E04A4.7),whereas the Hc-CYT C was equally similar (71%) to proteins encodedby both cyc-2.1 (E04A4.7) and cyc-2.2 (ZC116.2). Available data(www.wormbase.org) also indicated that the proteins CYC-2.1 and
sequences of cytochromes c from Haemonchus contortus, Trichostrongylus vitrinus andle from databases (www.ncbi.nlm.nih.gov, www.ebi.ac.uk/parasites/parasite-genome).ort values ≥0.80 are indicated above each branch.

Fig. 3. The detection of haem-binding proteins in adult male (M) and female (F) Haemonchus contortus (Hc) and Trichostrongylus vitrinus (Tv). Cytochrome c (CYT C), indicated by anarrow, was expressed equally in adult male and female H. contortus, whereas expression of cytochrome c in T. vitrinus was substantially higher in males than females (panel A). Thecytochromes cwere expressed as glutathione S-transferase (GST) fusion proteins to ensure that the corresponding genes were not pseudogenes (panel B). The sizes and numbers ofhaem-binding proteins in haemolytic sheep serum (HSS) were also assessed to ensure that the proteins detected in worm preparations were not from the host (panel C). None of theproteins detected in sheep blood were the same size as the proteins detected in H. contortus, indicating that those detected in the nematode were not host proteins. The sizes of theprotein standards (kDa) used for the analysis are indicated to the left of each panel.
Fig. 4. Analysis of transcription of cytochromes c in adult male (M) and female (F)Haemonchus contortus and Trichostrongylus vitrinus by reverse-transcription PCR,including a positive control PCR using primers for the amplification of a region(187 bp) from the large subunit of nuclear ribosomal RNA (28S). The transcription of Tv-cyt c (360 bp)was greater in adultmales comparedwith females. The transcription ofHc-cyt cwas shown to be similar between adult males and females of H. contortus (345 bp).
126 B.E. Campbell et al. / Gene 424 (2008) 121–129
CYC-2.2 from C. elegans are involved in the molecular functions ofelectron transport and are linked to the biological processes ofembryonic development, larval development, positive regulation ofgrowth rate and positive regulation of multicellular organism growth.Gene knock-down using RNAi is known to result in ‘loss-of-function’phenotypes, including embryonic lethality (Emb), slow growth (Slo)and larval arrest (Lva) for cyc-2.1 and slow growth (Slo) for cyc-2.2(www.wormbase.org/). Further investigation of cyc-2.1 and cyc-2.2 inWormNet (www.functionalnet.org/wormnet/), which contains 82% ofthe genes of C. elegans (Lee et al., 2008), identified a number of genespredicted to interact with these two CYT Cs. Twelve and twenty-onegenes were predicted to interact with cyc-2.1 and cyc-2.2, respec-tively; amongst these are genes, such as those encoding initiationfactors iff-1 and iff-2 and some sperm proteins, which are enriched inthe germline and are required for its development (Hanazawa et al.,2004; www.wormbase.org). Moreover, gene ontology (GO) dataindicate that these genes and their interaction partners are requiredfor key biological processes, including reproduction, gametogenesis,translational initiation, locomotory behaviour, growth, embryonicdevelopment, larval development and for the development of thegenitalia in C. elegans (hermaphrodite). Although there are presentlyno data for the spatial or temporal transcription or expressionpatterns for these genes, genetic interaction predictions suggest thatthese molecules may be expressed in gonadal tissues and/or areassociated with the production of sperm. Hc-CYT C shows equalamino acid identity to CYC-2.1 and CYC-2.2 (71%, Table 1), suggestingthat it may be expressed in somatic tissue, including that of thegonads, and may have a dual role in somatic and testicular tissues.The Tv-CYT C shows the highest homology to the CYC-2.2 of C. eleganswhich interacts with genes including ssp-2 and sss-10 (both represent-ing a yet uncharacterised family of [class P] sperm-specific proteins),suggesting that the Tv-CYT C might be involved in functions such asmale gametogenesis. Localizing the CYT C proteins to specific tissueswould allow these hypotheses to be tested.
3.2. Transcription and expression in H. contortus and T. vitrinus
As cyt c pseudogenes have been described for mammals (∼30 inrats and ∼49 in humans) (Scarpulla, 1985; Evans and Scarpulla, 1988;Zhang and Gerstein, 2003), it was important to show that the cyt cgenes cloned encoded proteins. In the present study, Hc-CYT C and Tv-CYT C were expressed in E. coli as GST fusion proteins and detectedusing a specific anti-GST antibody (Fig. 3, panel A). Immunoblotanalysis detected fusion proteins of the expected sizes, namely
38.6 kDa for H. contortus and 38.9 kDa for T. vitrinus. These findingsconfirmed that the genes isolated were not pseudogenes. Theexpressed proteins did not bind to haem, using the chemilumine-scence approach, which was likely due to having been expressed in aprokaryotic system, which would neither allow correct folding of theproteins nor post-translational modification(s).
Two approaches were used independently to verify that the haem-binding proteins detected in adult H. contortus originated from theworm rather than the host. Haemolytic sheep serumwas tested for thepresence of haem-binding proteins, including CYT C, haemoglobin andcytochrome P450s (Fig. 3, panel B). Based on the predicted sizes ofsheep CYT C (11.7 kDa; Swiss Prot, ca.expasy.org/uniprot, accession no.P62896), cytochrome P450s (e.g., ∼58 kDa; accession no. CAB40563;www.ncbi.nlm.nih.gov) and haemoglobin (∼64 kDa, e.g., accessionnos. P68240 and P02075; www.ncbi.nlm.nih.gov), there was noevidence that Hc-CYT C or Tv-CYT C were derived from host bloodor tissue; the failure of an anti-sheep antibody to detect proteins inworm extracts confirmed this finding (not shown).
Following the confirmation that the cyt c genes were indeednematode protein-coding genes, the transcription of the cyt c genes inadult H. contortus and T. vitrinus was investigated. Analysis by RT-PCRrevealed no clear difference in the level of transcription between adultmales and females of H. contortus, suggesting that CYT C may not betestis-specific. The cyt c transcript levels in adult H. contortus reflectedthe protein levels detected by colorimetric and chemiluminescenceassays (Figs. 3 and 4). Given that blood-feeding nematodes, such as H.
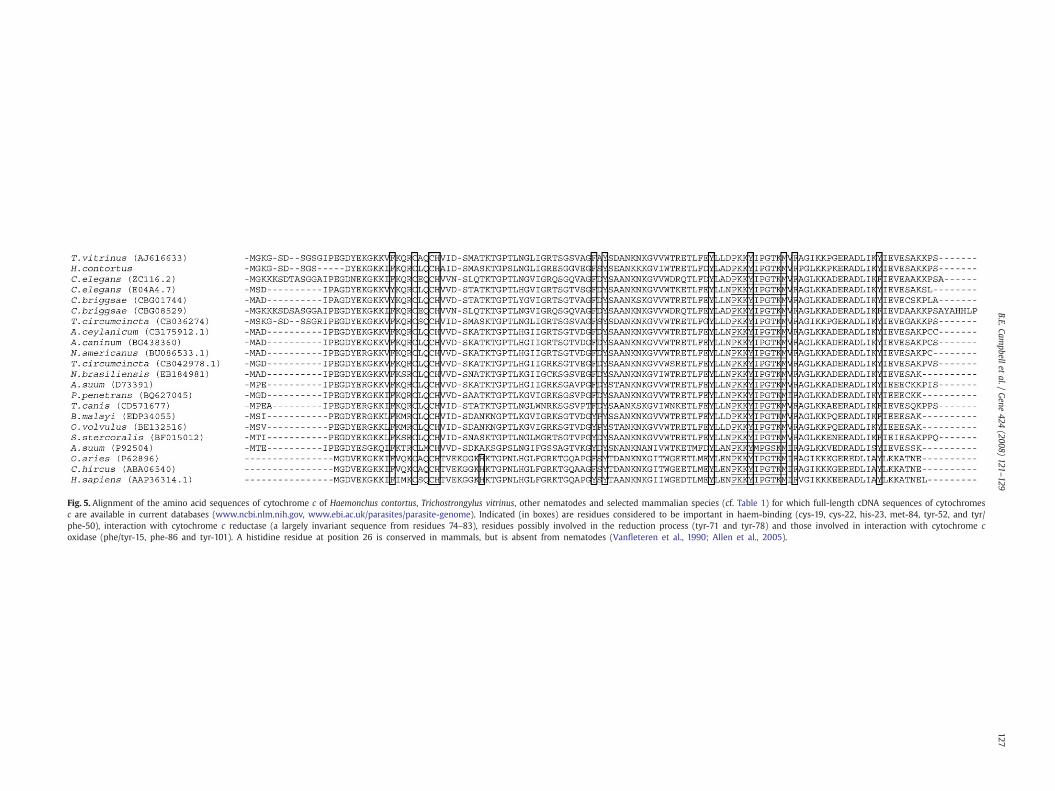
Fig. 5. Alignment of the amino acid sequences of cytochrome c of Haemonchus contortus, Trichostrongylus vitrinus, other nematodes and selected mammalian species (cf. Table 1) for which full-length cDNA sequences of cytochromesc are available in current databases (www.ncbi.nlm.nih.gov, www.ebi.ac.uk/parasites/parasite-genome). Indicated (in boxes) are residues considered to be important in haem-binding (cys-19, cys-22, his-23, met-84, tyr-52, and tyr/phe-50), interaction with cytochrome c reductase (a largely invariant sequence from residues 74–83), residues possibly involved in the reduction process (tyr-71 and tyr-78) and those involved in interaction with cytochrome coxidase (phe/tyr-15, phe-86 and tyr-101). A histidine residue at position 26 is conserved in mammals, but is absent from nematodes (Vanfleteren et al., 1990; Allen et al., 2005).
127B.E.Cam
pbelletal./
Gene
424(2008)
121–129

Fig. 6. Three-dimensional ribbon structures of cytochrome c proteins from thetrichostrongylid nematodes Haemonchus contortus and Trichostrongylus vitrinus,modelled using the program MODELLER (Fiser and Sali, 2003). PROCHECK-NMR(Laskowski et al., 1996) was used to confirm the quality of the stereochemical structurespredicted.
128 B.E. Campbell et al. / Gene 424 (2008) 121–129
contortus, live as adults in a microaerobic environment in the host, itwas not unexpected that CYT C expressionwas similar between malesand females. Takamiya et al. (1996a) suggested that, of the two CYT Csin A. suum, type 2 CYT C (adult-specific) has evolved to assist theworms to adapt to the anaerobic environment in the gut of the host.The second CYT C was transcribed mostly in third-stage larvae (L3s) ofA. suum (see Takamiya et al., 1996b).
In contrast to Hc-cyt c, the transcription of Tv-cyt c was greater inadult males comparedwith females, suggesting a key role for Tv-CYT Cin biological processes in males (Fig. 4). The differential transcriptionbetween males and females is consistent with previous evidence ofa 4.5-fold higher transcription in males of the mRNA linked to ESTTVm07_A08 (Nisbet and Gasser, 2004) as well as the abundance ofCYT C in adult males of T. vitrinus and no detectable protein infemales (see Fig. 4). Given that the chemiluminescence techniquecan detect picomolar amounts of protein (Feissner et al., 2003), it ishighly unlikely that the assay would not detect the presence of CYTC. It is possible that increased CYT C activity in male T. vitrinusrelates to function/s similar to that/those in mice, in which it isbelieved that the testicular CYT C rapidly catalyses the reduction ofreactive oxygen species preventing DNA damage, and has a role inthe elimination of sperm with damaged DNA (Liu et al., 2006).Hence, the expression of Tv-CYT C may be up-regulated in males toachieve this function with or without the necessity for a testis-specific CYT C.
Although phylogenetic, genetic and immunological analyses haverevealed that the testis-specific and somatic CYT C proteins aredistinctly different in structure and function (Kim and Sabourin, 1986;Virbasius and Scarpulla, 1988; Hake and Hecht, 1993; Arama et al.,2006), there is evidence from D. melanogaster (see Arama et al., 2006)that these proteins show some functional conservation. Caspaseactivation, required for apoptosis during spermatid differentiation incyt-c–d (testis-specific CYT C) null mutants, could be restored by thetransformation and subsequent expression of either the testis-specificor somatic cyt c genes. The differences in expression of the differentCYT Cs between somatic and testicular tissues in the vinegar fly wereconsidered to be the result of a differential regulation of transcriptionin these tissues, in which cyt c–d was predominantly expressed inmale germ cells (Kim and Sabourin, 1986; Virbasius and Scarpulla,1988; Vanfleteren et al., 1990; Takamiya et al., 1996a,b; Arama et al.,2006). Multiple, distinct cyt c transcripts have been detected in mice.Proximal initiation of transcription has been shown to produce foursmaller cyt c transcripts in somatic cells, germ cells and early meioticcells (Hake and Hecht, 1993); distal initiation of transcriptionproduced a larger transcript, which was detected exclusively inpost-meiotic cells (Hake and Hecht, 1993). Interestingly, the distancebetween the two transcription initiation sites was shown to be N1 kb(Hake and Hecht, 1993). Testis-specific and somatic CYT Cs have alsobeen identified in rats, rabbits, cattle and insects, indicating that morethan one gene encoding CYT C may be required to provide thefunctional and/or regulatory specificity that a single gene could not(Kim and Sabourin, 1986; Virbasius and Scarpulla, 1988; Narisawa etal., 2002; Arama et al., 2006).
3.3. Comparisons of CYT Cs of nematodes with those of other organisms
The alignment of the amino acid sequences of CYT Cs fromnematodes compared with those of mammals indicated conservedelements and some distinct differences (Fig. 5). This alignmentshowed a high degree of conservation, including haem-bindingresidues (cys-19, cys-22, his-23, tyr/phe-50, tyr-52 and met-84)(Vanfleteren et al., 1990; Allen et al., 2005). A region of ten aminoacids (residues 74–83) towards the C-terminus of the protein, thoughtto be involved in interaction with CYT C reductase (Vanfleteren et al.,1990; Allen et al., 2005), was also conserved across both nematodesand mammals. It was also conserved across a wider range of taxa,
including birds, reptiles, amphibians, fish, arthropods, yeast and fungi(data not shown). The exception to this was a CYT C from A. suum(accession no. P92504), which had a methionine residue at position78, whilst all other sequences had an isoleucine. Residues tyr-71 andmet/tyr-78, invariant across most taxa studied, are also thought to beinvolved in the reduction reaction (Vanfleteren et al., 1990). Functionalinteraction with CYT C oxidase, via phe/tyr-15, tyr-101 and phe-86,was also conserved. Interestingly, all CYT C sequences from nematodeslacked a histidine at position 26, which is present and conserved in theCYT Cs of mammals (Vanfleteren et al., 1990). This alteration is notthought to cause a conformational change in the protein (Vanfleterenet al., 1990), but its functional significance is presently unknown and isthus worthy of study.
Amino acid alignments reveal useful information about levels ofsequence conservation and divergence. The most informativecomparisons between CYT C proteins from different taxa, however,are made using three-dimensional models. Such models of the Hc-CYT C and Tv-CYT C were developed (see Fig. 6) based oncomparisons with CYT Cs from cattle (PDB ID 2b4z; Mirkin et al.,unpublished) and rice (PDB ID 1CCR; Ochi et al., 1983). Hc-CYT C,modelled using a rice ferri-CYT C, was significantly different from thestructure predicted for the Tv-CYT C and bovine CYT C (Fig. 6),suggesting functional differences among the molecules. The inferredprotein Tv-CYT C resembled the bovine protein, but had a longer N-terminus prior to the first alpha-helix and differed in electrostaticpotential across the protein (data not shown). This informationsuggests that the structure of Tv-CYT C is similar to CYT C of sheep(Swiss Prot; ca.expasy.org/uniprot, primary accession no. P62896).Kinetic studies of interactions between CYT C oxidase and CYT Csfrom A. suum and those of cattle have demonstrated species-specificdifferences in activities (Takamiya et al., 1996a,b).
Differences in the kinetic properties and binding specificitiesbetween nematode and mammalian CYT C might allow theidentification of drug targets, enabling the design of compoundsspecifically against nematodes. Targeting specific CYT Cs may have a“two-fold effect” on nematode reproduction. Firstly, a reduction insperm production and the production of defective sperm wouldreduce fertilization. Secondly, sperm has been shown to initateoocyte meiotic maturation and ovulation (Harris et al., 2006).Therefore, a reduction in sperm production would also reducefecundity in the female. Although there is presently no evidence ofthe involvement of CYT Cs of nematodes in apoptosis, current dataand evidence for C. elegans (available in WormBase) suggest thatthese molecules are involved in gametogenesis in parasitic nema-todes and adult survival in environments of low oxygen saturationinside a host (Takamiya et al., 1996a). The disruption of theseprocesses would thus have serious consequences for nematodesurvival, development and/or reproduction.

129B.E. Campbell et al. / Gene 424 (2008) 121–129
Acknowledgments
Thanks to Youssef Abs EL-Osta for confirming the sequence of ESTTVm07_A08 and Ian Beveridge for comments on the manuscript.Funding from the Australian Research Council (ARC) (LP0667795),Genetic Technologies Limited (GTG) and Meat and Livestock Australia(MLA) is gratefully acknowledged.
References
Adrain, C., Martin, S.J., 2001. The mitochondrial apoptosome: a killer unleashed by thecytochrome seas. Trends Biochem. Sci. 26, 390–397.
Allen, J.W.A., Leach, N., Ferguson, S.J., 2005. The histidine of the c-type cytochromeCXXCH haem-binding motif is essential for haem attachment by the Escherichia colicytochrome c maturation (Ccm apparatus). Biochem. J. 389, 587–592.
Arama, E., Bader, M., Srivastava, M., Bergmann, A., Steller, H., 2006. The two Drosophilacytochrome C proteins can function in both respiration and caspase activation.EMBO J. 25, 232–243.
Bowman D.D., 1995. Georgis' Parasitology for Veterinarians 6th Edition. W.B. SaundersCompany. pp. 160, 162, 163, 205.
Breathnach, R., Chambon, P., 1981. Organization and expression of eukaryotic split genescoding for proteins. Annu. Rev. Biochem. 50, 349–383.
Bradford,M.,1976.A rapid and sensitivemethod for thequantitationofmicrogramquantitiesof protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254.
Chinnaiyan, A.M., 1999. The apoptosome: heart and soul of the cell death machine.Neoplasia. 1, 5–15.
Evans, M.J., Scarpulla, R.C., 1988. The human somatic cytochrome c gene: two classes ofprocessed pseudogenes demarcate a period of rapid molecular evolution(accelerated evolution/respiratory chain/primate lineage). Proc. Natl. Acad. Sci.USA 85, 9625–9629.
Feissner, R., Xiang, Y., Kranz, R.G., 2003. Chemiluminescent-based methods to detectsubpicomole levels of c-type cytochromes. Anal. Biochem. 315, 90–94.
Fiser, A., Sali, A., 2003. Modeller: generation and refinement of homology-based proteinstructure models. Methods Enzymol. 374, 461–491.
Gasser, R.B., Chilton, N.B., Hoste, H., Beveridge, I., 1993. Rapid sequencing of rDNA fromsingle worms and eggs of parasitic helminths. Nucleic Acids Res. 21, 2525–2526.
Hake, L.E., Hecht, N.B., 1993. Utilization of an alternative transcription initiation site ofsomatic cytochrome c in the mouse produces a testis-specific cytochrome c mRNA.J. Biol. Chem. 268, 4788–4797.
Hanahan, D., Jessee, J., Bloom, F.R., 1991. Plasmid transformation of Escherichia coli andother bacteria. Methods Enzymol. 204, 63–113.
Hanazawa, M., Kawasaki, I., Kunitomo, H., Gengyo-Ando, K., Bennett, K.L., Mitani, S., Iino,Y., 2004. The Caenorhabditis elegans eukaryotic initiation factor 5A homologue, IFF-1, is required for germ cell proliferation, gametogenesis and localization of the P-granule component PGL-1. Mech. Dev. 121, 213–224.
Harris, J.E., Govindan, J.A., Yamamoto, I., Schwartz, J., Kaverina, I., Greenstein, D., 2006.Major sperm protein signalling promotes oocyte microtubule reorganization priorto fertilization in Caenorhabditis elegans. Dev. Biol. 299, 105–121.
Ho, A.T., Zacksenhaus, E., 2004. Splitting the apoptosome. Cell Cycle 3, 446–448.Lee, I., Lehner, B., Crombie, C., Wang, W., Fraser, A.G., Marcotte, E.M., 2008. A single
network comprising the majority of genes accurately predicts the phenotypiceffects of gene perturbation in C. elegans. Nat. Genet. 40, 181–188.
Jiang, X., Wang, X., 2004. Cytochrome C-mediated apoptosis. Annu. Rev. Biochem. 73,87–106.
Kim, I.C., Sabourin, C.L., 1986. Antigenic and size differences between somatic andtesticular cytochromes c. Biochem. Biophys. Res. Commun. 141, 131–136.
Laskowski, R.A., Rullmannn, J.A., MacArthur, M.W., Kaptein, R., Thornton, J.M., 1996.AQUA and PROCHECK-NMR: programs for checking the quality of protein structuressolved by NMR. J. Biomol. NMR. 8, 477–486.
Liu, Z., Lin, Y., Ye, S., Liu, Q-y.,Meng, Z., Zhang, C-m., Xiz, Y.,Margoliash, E., Rao, Z., Liu, X-J.,2006. Remarkably high activities of testicular cytochrome c in destroying reactiveoxygen species and in triggering apoptosis. Proc Natl Acad Sci USA 103, 8965–8970.
MAFF, 1977. Manual of veterinary parasitological laboratory techniques. TechnicalBulletin 18. Her Majesty's Stationary Office, London, UK.
Narisawa, S., Hecht, N.B., Goldberg, E., Boatright, K.M., Reed, J.C., Millán, J.L., 2002. Testis-specific cytochrome c-null mice produce functional sperm but undergo earlytesticular atrophy. Mol. Cell Biol. 22, 5554–5562.
Nisbet, A.J., Gasser, R.B., 2004. Profilingof gender-specific gene expression forTrichostrongylusvitrinus (Nematoda: Strongylida) by microarray analysis of expressed sequence taglibraries constructed by suppressive-subtractive hybridisation. Int. J. Parasitol. 34,633–643.
Ochi, H., Hata, Y., Tanaka, N., Kakudo, M., Sakurai, T., Aihara, S., Morita, Y., 1983. Structureof rice ferricytochrome c at 2.0 A resolution. J. Mol. Biol. 166, 407–418.
Schlecht, U., Demougin, P., Koch, R., Hermida, L., Widerkehr, C., Descombes, P., Pineau, C.,Jégou, B., Primig, M., 2004. Expression profiling of mammalian male meiosis andgametogenesis identifies novel candidate genes for roles in the regulation offertility. Mol. Biol. Cell 15, 1031–1043.
Scarpulla, R.C., 1985. Association of a truncated cytochrome c processed pseudogenewith a similarly truncated member from a long interspersed repeat family of rat.Nucleic Acids Res. 13, 755–763.
Swofford D.L., 1999. PAUP⁎4.0b10. Sunderland: Sinauer Associates.Takamiya, S., Hirawake, H., Kuramochi, T., Aoki, T., Kojima, S., Kita, K., 1996a. cDNA
cloning for mitochondrial cytochrome c and its adult-specific isoform from Ascarissuum. Mol. Biochem. Parasitol. 76, 293–297.
Takamiya, S., Yu, Y., Cavaleante, M.E., Murayama, K., Taka, H., Tateno, S., Takeuchi, T.,Aoki, T., 1996b. Molecular and functional properties of cytochrome c from adultAscaris suum muscle. Mol. Biochem. Parasitol. 79, 61–70.
Thompson, J.D., Gibson, T.J., Plewniak, F., Jeanmougin, F., Higgins, D.G., 1997. TheCLUSTAL_X windows interface: flexible strategies for multiple sequence alignmentaided by quality analysis tools. Nucleic Acids Res. 25, 4876–4882.
Vanfleteren, J.R., Evers, E.A.I.M., Van deWerken, G., Van Beeumen, J.J., 1990. The primarystructure of cytochrome c from the nematode Caenorhabditis elegans. Biochem. J.271, 613–620.
Virbasius, J.V., Scarpulla, R.C., 1988. Structure and expression of rodent genes encodingthe testis-specific cytochrome c — differences in gene structure and evolutionbetween somatic and testicular variants. J. Biol. Chem. 263, 6791–6796.
Zhang, Z., Gerstein, M., 2003. The human genome has 49 cytochrome c pseudogenes,including a relic of a primordial gene that still functions in mouse. Gene 312,61–72.




















