
Phylogenetic Analysis of Morphological Data from Extinct and ...
36
455 Syst. Biol. 48(3):455–490, 1999 The Position of Cetacea Within Mammalia: Phylogenetic Analysis of Morphological Data from Extinct and Extant Taxa MAUREEN A. O’LEARY 1,3 AND JONATHAN H. GEISLER 2 1 Department of Anatomical Sciences, State University of New York, Stony Brook, New York 11794-8081, USA; E-mail: [email protected] 2 Department of Vertebrate Paleontology, American Museum of Natural History, New York, New York 10024-5192, USA; E-mail: [email protected] Abstract.— Knowledge of the phylogenetic position of the order Cetacea (whales, dolphins, and porpoises) within Mammalia is of central importance to evolutionary biologists studying the trans- formations of biological form and function that accompanied the shift from fully terrestrial to fully aquatic life in this clade. Phylogenies based on molecular data and those based on morphological data both place cetaceans among ungulates but are incongruent in other respects. Morphologists ar- gue that cetaceans are most closely related to mesonychians, an extinct group of terrestrial ungulates. They have disagreed, however, as to whether Perissodactyla (odd-toed ungulates) or Artiodactyla (even-toed ungulates) is the extant clade most closely related to Cetacea, and have long main- tained that each of these orders is monophyletic. The great majority of molecule-based phylogenies show, by contrast, not only that artiodactyls are the closest extant relatives of Cetacea, but also that Artiodactyla is paraphyletic unless cetaceans are nested within it, often as the sister group of hip- popotamids. We tested morphological evidence for several hypotheses concerning the sister taxon relationships of Cetacea in a maximum parsimony analysis of 123 morphological characters from 10 extant and 30 extinct taxa. We advocate treating certain multistate characters as ordered because such a procedure incorporates information about hierarchical morphological transformation. In all most-parsimonious trees, whether multistate characters are ordered or unordered, Artiodactyla is the extant sister taxon of Cetacea. With certain multistate characters ordered, the extinct clade Mesonychia (Mesonychidae + Hapalodectidae) is the sister taxon of Cetacea, and Artiodactyla is monophyletic. When all fossils are removed from the analysis, Artiodactyla is paraphyletic with Cetacea nested inside, indicating that inclusion of mesonychians and other extinct stem taxa in a phylogenetic analysis of the ungulate clade is integral to the recovery of artiodactyl monophyly. Phylogenies derived from molecular data alone may risk recovering inconsistent branches because of an inability to sample extinct clades, which by a conservative estimate, amount to 89% of the ingroup. Addition of data from recently described astragali attributed to cetaceans does not over- turn artiodactyl monophyly. [Artiodactyla; astragalus; Cetacea; fossils; homoplasy; Mesonychia; morphology; phylogeny.] The importance of fossils in phylogeny reconstruction is most keenly appreciated when a clade is of great antiquity and has suffered numerous extinctions. Gauthier et al. (1988), Donoghue et al. (1989), and No- vacek (1992a, 1994) have emphasized that under such circumstances, the importance of fossils in phylogeny reconstruction can outweigh the drawback of their incomplete- ness. These authors have shown that fossils often capture the primitive morphotype of a clade, because many fossil taxa have had less time to evolve homoplasies than have extant members of the same clades. Because fossils often have combinations of primi- tive and derived features not found in ex- tant taxa, they can be critical for untangling problems of long-branch attraction, where homoplasies masquerade as homologies. Depending on the sampling of characters and taxa, convergent similarities can have the potential to draw taxa together phylo- genetically, as in the celebrated example of the paraphyletic clade “Haematothermia” that links birds and mammals but excludes crocodiles (Gauthier et al., 1988; Eernisse and Kluge, 1993). Mammalian systematists in particular continue to spar (Catze is, 1993; Graur, 1993a, 1993b; Novacek, 1993) over the sig- ni cance of the often strongly differing tree topologies that emerge from phylogenetic analyses of data sets partitioned between molecular and morphological data or be- tween extinct and extant taxa. The problem is exacerbated by different types of missing data: phylogenies based on molecular data cannot sample extinct taxa, and those based 3 Address correspondence to Dr. Maureen A. O’Leary, Department of Anatomical Sciences, HSC T- 8 (040), SUNY at Stony Brook, Stony Brook, New York 11794–8081, USA. Downloaded from https://academic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Phylogenetic Analysis of Morphological Data from Extinct and ...

455
Syst. Biol. 48(3):455–490, 1999
The Position of Cetacea Within Mammalia: Phylogenetic Analysis ofMorphological Data from Extinct and Extant Taxa
MAUREEN A. O’LEARY1,3 AND JONATHAN H. GEISLER2
1Department of Anatomical Sciences, State University of New York, Stony Brook, New York 11794-8081 ,USA; E-mail: [email protected]
2Department of Vertebrate Paleontology, American Museum of Natural History, New York, New York10024-5192 , USA; E-mail: [email protected]
Abstract.— Knowledge of the phylogenetic position of the order Cetacea (whales, dolphins, andporpoises) within Mammalia is of central importance to evolutionary biologists studying the trans-formations of biological form and function that accompanied the shift from fully terrestrial to fullyaquatic life in this clade. Phylogenies based on molecular data and those based on morphologicaldata both place cetaceans among ungulates but are incongruent in other respects. Morphologists ar-gue that cetaceans aremost closely related to mesonychians, an extinctgroup of terrestrial ungulates.They have disagreed, however, as to whether Perissodactyla (odd-toed ungulates) or Artiodactyla(even-toed ungulates) is the extant clade most closely related to Cetacea, and have long main-tained that each of these orders is monophyletic. The great majority of molecule-based phylogeniesshow, by contrast, not only that artiodactyls are the closest extant relatives of Cetacea, but also thatArtiodactyla is paraphyletic unless cetaceans are nested within it, often as the sister group of hip-popotamids. We tested morphological evidence for several hypotheses concerning the sister taxonrelationships of Cetacea in a maximum parsimony analysis of 123 morphological characters from10 extant and 30 extinct taxa. We advocate treating certain multistate characters as ordered becausesuch a procedure incorporates information about hierarchical morphological transformation. Inall most-parsimonious trees, whether multistate characters are ordered or unordered, Artiodactylais the extant sister taxon of Cetacea. With certain multistate characters ordered, the extinct cladeMesonychia (Mesonychidae + Hapalodectidae) is the sister taxon of Cetacea, and Artiodactyla ismonophyletic. When all fossils are removed from the analysis, Artiodactyla is paraphyletic withCetacea nested inside, indicating that inclusion of mesonychians and other extinct stem taxa in aphylogenetic analysis of the ungulate clade is integral to the recovery of artiodactyl monophyly.Phylogenies derived from molecular data alone may risk recovering inconsistent branches becauseof an inability to sample extinct clades, which by a conservative estimate, amount to 89% of theingroup. Addition of data from recently described astragali attributed to cetaceans does not over-turn artiodactyl monophyly. [Artiodactyla; astragalus; Cetacea; fossils; homoplasy; Mesonychia;morphology; phylogeny.]
The importance of fossils in phylogenyreconstruction is most keenly appreciatedwhen a clade is of great antiquity and hassuffered numerous extinctions. Gauthier etal. (1988), Donoghue et al. (1989), and No-vacek (1992a, 1994) have emphasized thatunder such circumstances, the importanceof fossils in phylogeny reconstruction canoutweigh the drawback of their incomplete-ness. These authors have shown that fossilsoften capture the primitive morphotype ofa clade, because many fossil taxa have hadless time to evolve homoplasies than haveextant members of the same clades. Becausefossils often have combinations of primi-tive and derived features not found in ex-tant taxa, they can be critical for untangling
problems of long-branch attraction, wherehomoplasies masquerade as homologies.Depending on the sampling of charactersand taxa, convergent similarities can havethe potential to draw taxa together phylo-genetically, as in the celebrated example ofthe paraphyletic clade “Haematothermia”that links birds and mammals but excludescrocodiles (Gauthier et al., 1988; Eernisseand Kluge, 1993).
Mammalian systematists in particularcontinue to spar (Catze�is, 1993; Graur,1993a, 1993b; Novacek, 1993) over the sig-ni�cance of the often strongly differing treetopologies that emerge from phylogeneticanalyses of data sets partitioned betweenmolecular and morphological data or be-tween extinct and extant taxa. The problemis exacerbated by different types of missingdata: phylogenies based on molecular datacannot sample extinct taxa, and those based
3Address correspondence to Dr. Maureen A.O’Leary, Department of Anatomical Sciences, HSC T-8 (040), SUNY at Stony Brook, Stony Brook, New York11794–8081, USA.
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

456 SYSTEMATIC BIOLOGY VOL. 48
on morphological data and drawing on fos-sil taxa cannot sample all morphologicalcharacters, even from the most exquisitelypreserved fossils (Gauthier et al., 1988; No-vacek, 1994; Simmons and Geisler, 1998).
The part of the mammalian tree relatingto the origin of Cetacea (whales, dolphins,and porpoises) has been the subject of par-ticular controversy of late because of dif-fering results from phylogenetic analysesbased on molecular data and those basedon morphological data. Both types of analy-ses support the hypothesis that cetaceans arenested within ungulates (Novacek, 1992b),but there is virtually no agreement as to thesister taxon of Cetacea or whether certainungulate clades close to Cetacea are mono-phyletic. Without knowledge of the sistertaxon of Cetacea, we cannot begin to under-stand the drastic anatomical and physiolog-ical transformations that occurred as terres-trial mammals returned to aquatic life andevolved into some of the most specializedknown vertebrates.
Initial phylogenetic analyses of mor-phological evidence bearing on the ques-tion of cetacean origins nothwithstanding(Prothero et al., 1988; Thewissen, 1994), acomprehensive phylogenetic analysis test-ing morphological evidence for the mono-phyly of various modern ungulate orderswith respect to Cetacea has been conspic-uously absent from the paleontological lit-erature until recently (Geisler and O’Leary,1997; Geisler and Luo, 1998). This is a seriousshortcoming, because detailed morphologi-cal analyses are an integral part of any to-tal evidence analysis (Kluge, 1989; and ref-erences therein) combining molecular andmorphological data (also referred to as si-multaneous analysis; Nixon and Carpen-ter, 1996). Lack of a comprehensive phylo-genetic analysis of morphological data haspersisted despite the suggestion by somemolecular biologists thatbecause of the “nu-merous extinctions along the stem lineageof cetaceans, most major insights into thetransformation from a terrestrial ungulate toa fully aquatic cetacean will come from fos-sil taxa” (Gatesy et al., 1996:960) . Here weattempt to correct this shortcoming.
BACKGROUND ON CETACEAN ORIGINS
Morphological ContributionsMorphologists at one time viewed under-
standing the phylogenetic relationships ofcetaceans to other orders of mammals as anintractable problem because the anatomy ofcetaceans is so transformed relative to thatof other mammals (e.g., Simpson, 1945).Within the last three decades, however, at-tributable in no small part to discoveriesof new fossils, paleontologists have devel-oped the hypothesis that an extinct order ofcarnivorous, hoofed mammals, the Mesony-chia, is most closely related to Cetacea (VanValen, 1966, 1968, 1969, 1978; McKenna,1975; Prothero et al., 1988; Thewissen, 1994;Zhou et al., 1995; McKenna and Bell, 1997;Geisler and Luo, 1998; O’Leary, 1998a). VanValen (1966) initially based this hypothe-sis on a variety of dental and cranial simi-larities. Mesonychians (Mesonychidae andHapalodectidae [but not Andrewsarchus; seeVan Valen, 1978; McKenna and Bell, 1997;O’Leary, 1998a]) , a group that some paleon-tologists argue is paraphyletic (Thewissen,1994; Geisler and Luo, 1998), are knownfrom the Early Tertiary of the Holarctic. Be-cause they had hoofs, mesonychians havegenerally been classi�ed among ungulates(Van Valen, 1966), but they differ fromvirtually all other ungulates in possessinglaterally compressed, homodont lower den-titions that are strongly suggestive of a car-nivorous diet (Szalay and Gould, 1966; Sza-lay, 1969a, 1969b; Zhou et al., 1992; O’Learyand Rose, 1995a, 1995b). McKenna (1975)classi�ed Cetacea and Acreodi (Mesonychi-dae) together in the mirorder Cete withinthe grandorder Ungulata, thereby formal-izing morphological arguments advancedby Van Valen (1966; see also McKenna andBell, 1997). Paleontologists have not arguedthat mesonychians are nested among any ofthe following ungulate clades that have ex-tant members: Artiodactyla, the even-toedhoofed mammals (ruminants, pigs, hip-pos, and camels); Perissodactyla, the odd-toed hoofed mammals (horses, rhinos andtapirs); or among the paenungulates: Pro-boscidea (elephants), Hyracoidea (hyraxes),and Sirenia (dugongs and manatees).
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

1999 O’LEARY AND GEISLER—PHYLOGENY OF CETACEA 457
Initially, interpretations of morpholog-ical evidence for the extant clade mostclosely related to Cetacea tended to con-verge on Perissodactyla or were simply am-biguous. Among more recent treatments,Novacek (1982) placed cetaceans in anunresolved polytomy with Artiodactyla,Perissodactyla, and tentatively, Tubuliden-tata (aardvarks), with paenungulates asthe next most closely related clade. Aseries of subsequent parsimony analysesof the mammalian orders tackled moreprecisely the morphological evidence forcetacean–ungulate relationships. On thebasis of cranial traits, Novacek (1986)found cetaceans to be related to ungu-lates in the following manner: Artiodactyla(Cetacea (Perissodactyla (Hyracoids (Sire-nians + Proboscideans))))–a result broadlyconsistent with information from patternsof variation in the stapedial artery (Wible,1987). Novacek and Wyss (1986) and No-vacek (1989) obtained similar results basedon morphological information from severalanatomical systems but introduced the pos-sibility that Perissodactyla might be moreclosely related to Cetacea than is Artio-dactyla. In a study including a variety ofextinct “condylarths,” Prothero et al. (1988)argued that Cetacea was the sister group ofAndrewsarchus and that, of the extant ungu-late orders, Perissodactyla was most closelyrelated to Cetacea. Their tree, however, maynot be the most–parsimonious explanationof their data because these authors did notperform a parsimony analysis on their en-tire data set. Thus, phylogenetic analyses ofmorphological data initially tended to favora close relationship between paenungulates ,perissodactyls, and Cetacea but were unableto demonstrate this relationship with greatcertainty.
In many of the studies described above,monophyly of Artiodactyla was assumedand not tested. The assumption that Artio-dactyla is monophyletic originates primar-ily from the observation that artiodactylsshare an ankle joint morphology known asthe double-pulleyed astragalus (Schaeffer,1947; Vaughan, 1986) (Fig. 1), a unique con-dition. This skeletal feature is one of the fewcharacters that can be directly veri�ed in
both extant and extinct forms, and its pres-encehas been the primary criterion for mem-bership in the order, particularly for fos-sil taxa. The signi�cance of the artiodactylankle is that it restricts the distal hindlimbto parasagittal motion and is thought to beless likely to dislocate as the animal en-gages in high–speed quadrupedal runningacross a terrestrial substrate (Schaeffer, 1947;O’Leary and Rose, 1995b). Despite the broadradiation of this group, the morphology ofthis joint has remained remarkably constantsince its �rst known appearance (Schaeffer,1947) at the base of the Eocene ( ~ 55 millionyears ago). Figure 1 shows the astragali ofvarious mammals associated with the basalungulate radiation. Relatively unspecializedEarly Tertiary mammals like Chriacus havean astragalus with a relatively �at proxi-mal end (trochlea) and a convex distal end(head). The astragalus of Phenacodus, a morederived ungulate, has a convex head buta grooved trochlea. Artiodactyls, perisso-dactyls, and some mesonychians also havea relatively grooved trochlea. These taxadiffer, however, in the structure of the as-tragalar head, speci�cally in its articularfacet for the navicular, and each morphol-ogy is thought to be a convergent special-ization for cursorial locomotion (O’Learyand Rose, 1995b). Both perissodactyls andmesonychians exhibit a saddle-shaped head(O’Leary and Rose, 1995b) but differ in thatmesonychians have a distinct facet for thecuboid. Artiodactyls have a deeply groovedhead with a more substantial cuboid ar-ticulation (e.g., Bunophorus [Fig. 1]; Schaef-fer, 1947). Despite variations in body sizeand locomotor capabilities among artio-dactyls, the morphology of the astragalusremains a diagnostic feature of the order(Schaeffer, 1947; Vaughan, 1986). Stem taxaoutside of Artiodactyla exhibiting transi-tional morphologies leading to the primitiveartiodactyl morphotype remain virtuallyunknown (Rose, 1987, 1996). A completeastragalus associated with other diagnos-tic material of a very primitive cetaceanhas never been described (Milinkovitch andThewissen, 1997). For the archaic cetacean,Ambulocetus, the taxon for which we havethe most detailed knowledge of skeletal
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

458 SYSTEMATIC BIOLOGY VOL. 48
FIGURE 1. Right astragali of Early Tertiary mammals associated with the basal ungulate radiation: the proxi-mal end of the astragalus (trochlea); the articular facet for the distal tibia (character 109), towards the top of eachdrawing; the distal end of the astragalus (head); the articular facet for the navicular; and sometimes the cuboid(characters 104 and 106), towards the bottom of the page. Chriacus (AMNH-VP 92832) , an oxyclaenid arctocyonid(morphology resembles that of Arctocyon coded in this analysis); Phenacodus (AMNH-VP 15287), a stem taxonto extant perissodactyls; Dissacus (AMNH-VP 3359) and Pachyaena (AMNH 16154), mesonychians; Bunophorus(AMNH-VP 92847), an artiodactyl (morphology resembles that of Diacodexis coded in this analysis, representativeof the artiodactyl double-pulleyed astragalus); Heptodon (AMNH-VP 95866), a perissodactyl; and Ambulocetus(H-GSP 18507) , an archaic cetacean (redrawn from Thewissen et al., 1996). The head and part of the trochlea ofthe Ambulocetus astragalus are not preserved in this specimen. Scale bar = 10 mm.
anatomy, only the proximal astragalus isknown (Fig. 1). It has a grooved trochleathat distinguishes it from generalized mam-mals such as Chriacus and Arctocyon, butthe more diagnostic head is not preserved.Two partial astragali have recently been at-tributed to thearchaic cetaceans Ambulocetusand Pakicetus on the basis of their groovedtrochleae, large size, and faunal associations(Thewissen et al., 1998). The authors inter-pret these bones as possessing certain de-rived similarities that might link cetaceansto artiodactyls but not to mesonychians but
qualify their argument by stating that these“cetacean” astragali do not have trochleatedheads as in artiodactyls. Because these fos-sils are fragmentary and are not associatedwith diagnostic cetacean material, we con-tinue to consider the morphology of thecetacean astragalus to be relatively poorlyestablished.
Three other relatively recent fossil discov-eries also suggest the possibility of a closelink between Cetacea and Artiodactyla. Thehind foot of the archaic cetacean Basilosaurus(Gingerich et al., 1990) exhibits a paraxonic
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

1999 O’LEARY AND GEISLER—PHYLOGENY OF CETACEA 459
condition (i.e., the axis of symmetry passesbetween the third and fourth digits), a fea-ture consistent with a close phylogeneticrelationship between cetaceans and bothMesonychia and Artiodactyla. The mor-phology of the incus of another archaiccetacean, Pakicetus (Thewissen and Hus-sain, 1993), has been argued to support aclose relationship between Artiodactyla andCetacea, as have features of the skeleton of athird archaic cetacean Ambulocetus (Thewis-sen et al., 1994, 1996), such as hoofs and aparaxonic foot, which reinforce a link withMesonychidae as well.
Thewissen (1994) conducted the �rst par-simony analysis of morphological data bear-ing on the question of cetacean origins andidenti�ed several dental synapomorphiessupporting a sister-taxon relationship be-tween cetaceans and certain mesonychids.He found Mesonychia (Mesonychidae + Ha-palodectidae) and Mesonychidae to be pa-raphyletic with respect to Cetacea and, incontrast to Thewissen and Hussain (1993),argued that the extant sister taxon to Cetaceawas Perissodactyla, a hypothesis champi-oned by Prothero (1993) as well. Thewis-sen’s (1994) analysis did not directly testmonophyly of Artiodactyla because it in-cluded only one artiodactyl, Diacodexis.However, when Geisler and O’Leary (1997)and Geisler and Luo (1998) included sev-eral artiodactyls in parsimony analyses ofmorphological data, support emerged for aclade that included Artiodactyla, Cetacea,and Mesonychia to the exclusion of Perisso-dactyla, and for monophyly of Artiodactyla,Perissodactyla, Cetacea, and Mesonychi-dae. New dentitions of archaic cetaceans ledO’Leary (1998a) to argue that Mesonychi-dae and Mesonychia are each monophyleticclades (in contrast to Prothero et al., 1988;Thewissen, 1994), the former being nestedwithin the latter. Our work here representsa combination and expansion of these mostrecent phylogenetic analyses.
Molecular Contributions
Like morphological analyses, molecularanalyses of cetacean origins have varied intheir taxon and character sampling, meth-ods of phylogenetic reconstruction, and
ultimately in their conclusions. Some ini-tial parsimony analyses of amino acid se-quences (Goodman et al., 1985; Miyamotoand Goodman, 1986) or of a combinationof amino acid sequences, immunodiffusion,and morphology (Shoshani, 1986) gener-ally revealed no greater resolution thanthat Cetacea formed a clade with Artio-dactyla and Perissodactyla. Evidence sup-porting a close relationship between Peris-sodactyla and Cetacea to the exclusion ofArtiodactyla emerged from parsimony anal-yses of amino acid sequences of eye lens pro-teins (De Jong, 1985; McKenna, 1992), andfrom analysis of a combination of nucleargene sequences and amino acid sequences(Stanhope et al., 1993). This last result hasbeen among the minority of results basedon molecular data. The molecule-based hy-pothesis that Artiodactyla is the closest liv-ing relative of Cetacea was �rst put forth onthe basis of serological precipitin tests (Boy-den and Gemeroy, 1950). Maximum parsi-mony analyses of amino acid sequences of aand b hemoglobin chains (Shoshani et al.,1985; Czelusniak et al., 1990b), pancreaticribonucleases (Beintema et al., 1986, 1988),and a combination of eight different typesof amino acid sequences (Czelusniak et al.,1990a) also showed that Cetacea groupedmore closely with artiodactyls than withperissodactyls.
Several molecular analyses, however, be-gan to recover phylogenies with Cetaceanested within Artiodactyla, a highly con-troversial result from a morphological per-spective, given the reknowned monophylyof Artiodactyla. Czelusniak et al. (1990b)recovered a clade of cetaceans and peco-rans (deer, sheep, giraffe, and their closerelatives) to the exclusion of other artio-dactyls in maximum parsimony analyses ofamino acid sequences. They quali�ed theirresult as likely to be “phylogenetically in-correct” (Czelusniak et al., 1990b:614) be-cause the new topology differed from estab-lished ideas of relationships and was basedonly on a small subset of all molecular ev-idence. Using a combination of amino acidand nucleotide sequences for 18 artiodactyls(including Hippopotamus , camels, suids, andpecorans, a fair representation of taxonomic
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

460 SYSTEMATIC BIOLOGY VOL. 48
variation within Artiodactyla), Czelusniaket al. (1990a) found some evidence for artio-dactyl paraphyly, but equally parsimonioustrees also supported artiodactyl monophyly.Their cautious interpretations of trees sup-porting artiodactyl paraphyly were not fol-lowed by other researchers examining nu-cleotide sequence data.
Parsimony analyses of nucleotide se-quence data of the mitochondrial cy-tochrome b gene (Arnason et al., 1991)and of the 12S and 16S ribosomal genes(Milinkovitch et al., 1993) recovered a sister-taxon relationship between Artiodactylaand Cetacea to the exclusion of Perisso-dactyla, based on relatively small samplesof artiodactyls (1 to 3 taxa). Increasing theartiodactyl sample to 11 taxa for mitochon-drial cytochrome b gene sequences, Irwinet al. (1991) recovered a paraphyletic Ar-tiodactyla with Cetacea nested inside asthe sister taxon of camels. A sister taxonrelationship between Cetacea and rumi-nants resulted from parsimony analyses ofcombined nucleotide sequences of both cy-tochrome b and cytochrome c oxidase sub-unit II (Honeycutt et al., 1995) and receivedmixed support from nuclear gene sequences(Stanhope et al., 1996). Graur and Higgins(1994) analyzed 11 nuclear-encoded proteinsequences and �ve mitochondrial DNA se-quences in four taxa, using maximum like-lihood (with the assumption of constantrates of substitution [Hasegawa and Adachi,1996]), neighbor joining, and maximum par-simony, and found that Cetacea was the sis-ter group of the cow to the exclusion ofpigs and camels. They advocated revisionsto mammalian ordinal-level taxa to formal-ize their result, despite its genesis from rel-atively poor taxonomic sampling.
Others criticized the four-taxon method ofGraur and Higgins (1994), recognizing thatit risks recovering “robust, but false relation-ships” (Philippe and Douzery, 1994:149; seealso Adachi and Hasegawa, 1996) becauseof such confounding factors as long-branchattraction. Authors of other nucleotide se-quence analyses that had recovered artio-dactyl paraphyly (Queralt et al., 1995; Smithet al., 1996) quali�ed their results as tenta-tive, sensitive to such variables as outgroup
choice and ingroup sampling, and possiblyre�ective of the unique evolutionary historyof a particular gene but not of the clades inquestion. Hasegawa and Adachi (1996) reex-amined the data sets of both Graur and Hig-gins (1994) and Irwin and Arnason (1994),combined with hemoglobin sequences, us-ing a maximum-likelihood analysis witha rate heterogenous model for nucleotidesubstitution. They found greatly reducedstatistical support for artiodactyl paraphylybut still achieved that result. This suggestedto them that artiodactyl monophyly is a vi-able alternative hypothesis that could notbe dismissed until more genes had beenexamined.
None of the above nucleotide sequenceanalyses included hippopotamids in thesample of artiodactyls, and none explicitlytested the effect of this group of artiodactylson tree topology. Once hippopotamids wereincluded with 11 other artiodactyls in a par-simony analysis of nucleotide and aminoacid sequences of the cytochrome b gene,hippopotamids formed a clade with Cetaceato the exclusion of other artiodactyls (Ir-win and Arnason, 1994). Gatesy et al. (1996)found a similar result from parsimony anal-ysis of milk-protein gene sequences, con-cluding that the next outgroup to the hippo-cetacean clade was ruminant artiodactyls,then pigs and peccaries, and �nally camels.Increasing the number of genes and thenumber of taxa examined (Gatesy, 1997) pro-duced similar results, and Montgelard et al.(1997), using cytochrome b and 12s rRNAsequences, also found support for an An-codonta (Hexaprotodon + Hippopotamus) +Cetacea clade to the exclusion of other ar-tiodactyls. By contrast, the hippo-cetaceanclade had not been recovered in an earlierparsimony analysis of amino acid sequences(Czelusniak et al., 1990a).
Finally, Shimamura et al. (1997) found ev-idence for artiodactyl paraphyly on the ba-sis of retroposons: nucleotide sequences thathave been inserted into a genome at par-ticular loci (Li, 1997). As implemented byShimamura et al. (1997), the homologystatement for retroposons is their presenceor absence in a particular position in thegenome. Theresults of theseauthors demon-
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

1999 O’LEARY AND GEISLER—PHYLOGENY OF CETACEA 461
strated a close relationship between Hip-popotamus, ruminants, and cetaceans, to theexclusion of the camel pig and several out-group taxa. On the basis of these charac-ters alone, Shimamura et al. (1997:669) as-serted “we believe that recent moleculardata will lead to the reinterpretation by pa-leontologists of many fossil records of Artio-dactyla to match our conclusions.” Further-more, Milinkovitch and Thewissen (1997)claimed that retroposons are “noise-free,”arguing that it is improbable that retropo-sitional elements inserted themselves inde-pendently into orthologous positions in thegenomes of different taxa. No phylogeneticdata, however, are known a priori to be freeof homoplasy (Wiley et al., 1991).
Thus, molecular papers arguing for ar-tiodactyl paraphyly are numerous. Differ-ent molecular analyses have found peris-sodactyls, hippopotamids, ruminants, andcamels each to be most closely relatedto cetaceans. A hippopotamid–ruminant–cetacean grouping describes the most com-mon �nding of many of these studies,notably those with the densest characterand taxon sampling. These unconventionaltopologies recovered from molecular analy-ses have implications not only for cetaceanphylogeny but also for the monophyly ofclades within Artiodactyla. A previous phy-logenetic analysis of morphological charac-ters divided Artiodactyla into two clades,each with extant and extinct members (Gen-try and Hooker, 1988): Selenodontia, con-sisting of various ruminating artiodactyls(including Tragulidae, Cervidae, Giraf�dae,Antilocapridae, and Tylopoda and fossil rel-atives); and Bunodontia (including Suidae,Tayassuidae, and Hippopotamidae and fos-sil relatives). Many of the molecular topolo-gies, however, support paraphyly of theclade including Suidae, Tayassuidae, andHippopotamidae, challenging the hypoth-esis that these taxa are more closely re-lated to each other than any is to eithercamels or ruminants (Matthew, 1929; Pick-ford, 1983; Gentry and Hooker, 1988). Ananalysis of cytochrome b and 12s rRNAsequences, combined with morphologicaldata for extant taxa only (Montgelard et al.,1998), and aimed at testing the monophyly
of the clade that includes Suidae, Tayas-suidae, and Hippopotamidae, reinforced thenotion that con�icting signals are present inmorphological and molecular data sets. Thisanalysis did not include cetaceans, therebymaking it dif�cult to interpret the results inthe context of many other molecular analy-ses. At the same time, monophyly of rumi-nants and camels, to the exclusion of pigs,peccaries, and hippos, has been argued onthe basis of suites of morphological charac-ters (Webb and Taylor, 1980; Vaughan, 1986;Gentry and Hooker, 1988; Langer, 1988)but is disrupted in many of the molecularphylogenies.
The hypothesis that Artiodactyla isparaphyletic with respect to Cetacea con-tradicts traditional morphological ideas ofartiodactyl monophyly but merits explicittesting in the wake of the numerous molec-ular studies supporting artiodactyl para-phyly. Furthermore, the morphological hy-pothesis of artiodactyl monophyly has beenformulated without knowledge of the mor-phology of the astragalus of a primitivecetacean—the skeletal character argued tobe among the most important for determin-ing membership within Artiodactyla (Luck-ett and Hong, 1998). Because it is para-phyly, not polyphyly, of Artiodactyla thatis in question, lack of evidence about thecetacean astragalus invites the query: onwhat basis do morphologists know thatwhales are not highly derived artiodactyls?We therefore investigated the morpholog-ical evidence for the following four ques-tions: (1) Is Artiodactyla monophyletic withrespect toCetacea; (2) what is thesister taxonof Cetacea; (3) does exclusion of fossil taxafrom the phylogenetic analysis result in apruned version of the tree based on extanttaxa alone or a different tree; and (4) whatimpact does the morphology of the astra-galus of a primitive cetacean have on treetopology?
MATERIALS AND METHODS
The Signi�cance of a Morphological DataPartition When Most of the Ingroup Is Extinct
We subscribe fully to the notion that a to-tal evidence analysis (i.e., one based on com-
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

462 SYSTEMATIC BIOLOGY VOL. 48
bined molecular and morphological data) isan important step in the overall investiga-tion of the position of Cetacea within Mam-malia and that it is preferable to a taxonomiccongruence approach for reasons outlinedby Kluge (1989). It also has been argued inthe overall discussion of the signi�cance oftotal evidence that system-based data par-titions (i.e., molecules, osteology, behavior,or various subsets of these partitions) arenot at present derived from an understand-ing of biological processes but are insteadbased on intuition (Kluge and Wolf, 1993).Total evidence analyses combining molec-ular data for a few extant taxa with mor-phological data for extant taxa and a largenumber of extinct taxa, however, face anoperational obstacle imposed by extinction.With > 99% of all organisms extinct (No-vacek and Wheeler, 1992), can such total ev-idence analyses be considered robust whenthere are no molecular, soft tissue, or be-havioral data for the vast majority of life?Because of the potential importance of nu-merous extinct taxa for reconstructing thephylogeny of Cetacea, taxa that are knownonly from morphology and not from molec-ular biology, we believe it is also importantto investigate and present the morpholog-ical (predominantly osteological) signal in-dependently. In such a data partition, extincttaxa, which can far outnumber extant taxa inan ingroup, can be maximally in�uential ontree topology.
The vast majority of ingroup taxa rel-evant to cetacean phylogeny are extinct.Molecular and morphological contributionsto the problem of the phylogenetic posi-tion of Cetacea are congruent in nestingCetacea within ungulates, or the grandorderUngulata (McKenna and Bell, 1997). Ungu-lata (sensu McKenna and Bell, 1997) is di-verse and includes several extinct cladesbesides Artiodactyla, Cete (which includesCetacea), and Perissodactyla (Table 1). Inthis study (see below) �ve orders comprisethe ingroup: Artiodactyla, Perissodactyla,Procreodi, Cete, and “Condylarthra.” Most(89%) of the genera in these orders are ex-tinct (McKenna and Bell, 1997). Maximally,a phylogenetic study based on extant taxaalone can access no more than 11% of genera
in the ingroup. At present, larger molecule-based phylogenies (Gatesy, 1997) have sam-pled only 1% of ingroup genera. Even thepresent study, which includes 37 ingrouptaxa, almost triple that of Gatesy (1997),uses only 3% of relevant genera. Since theaim of this analysis is to recover deep splitsbetween mammalian orders such as Artio-dactyla, Perissodactyla, and Cetacea, sam-pling exhaustively within the ingroup maynot be essential if the most primitive mem-bers of a clade can be sampled. It is not,however, likely that extant mammals areprimitive members of the clades in ques-tion, because both molecular and morpho-logical estimates indicate that the splitsbetween mammalian orders are ancient(Novacek, 1992b; Kumar and Hedges, 1998),and their extant members have had muchtime to evolve homoplasies.
TABLE 1. Estimate (derived from McKenna andBell, 1997) of the extinction in the ingroup in this analy-sis and other recent molecule-based phylogenetic anal-yses of the position of Cetacea within Mammalia (e.g.,Gatesy, 1997, and references therein). Shown are thenumber of extinct and extant genera from �ve ordersthat make up the ingroup. This estimate of the num-ber of ingroup taxa is conservative because the in-group may include all genera within Ungulata (seeMcKennaand Bell, 1997) (i.e., Tubulidenatata, Arctosty-lopida, Litopterna, Notoungulata, Astrapotheria, Xe-nungulata, Pyrotheria, and Urantotheria) and becauseit is highly unlikely that the fossil record has preservedall ingroup taxa.
Taxon Extinct genera Extant genera
Artiodactyla 589 (86%) 94 (14%)Perissodactyla 236 (97.5%) 6 (2.5%)Cete 243 (86%) 39 (14%)“Condylarthra” 59 (100%) 0Procreodi 29 (100%) 0
Total 1156 (89%) 139 (11%)
The problem of looking at only a few taxa,particularly only a few extant mammaliantaxa, is thatbecause mammalian orders haveancient splits (long terminal branches) sep-arated by short internodes (Novacek et al.,1998), sampling of the extant taxa alone risksreinforcing an inconsistent branching pat-tern attributable to long-branch attraction(Felsenstein, 1978). Adding fossils to a data
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

1999 O’LEARY AND GEISLER—PHYLOGENY OF CETACEA 463
matrix has been argued to be one way to ex-pose and overturn a long-branch attractionproblem (Gauthier et al., 1988). The studyof Gauthier et al., 1988, and others of seedplants (Doyle and Donoghue, 1987) and eu-therians (Novacek, 1992a), all show that par-simony analysis of a matrix of morpholog-ical data for extant taxa alone results in atree that is incongruent with results of a par-simony analysis of a more complete matrix(extant + extinct taxa) of the same charac-ters. Interestingly, none of these empiricalanalyses has shown that the tree based onextant taxa alone was simply a pruned ver-sion of the tree based on the more completematrix. The assumption that the tree result-ing from more taxa is the most robust isimplicit in this methodology and is logical,because there is no justi�cation for exclud-ing ingroup taxa. Each study performed ex-periments in which fossil taxa were deletedand parsimony analyses run to investigatechanges to the tree topology (Donoghueet al., 1989). Drawing on independent evi-dence from simulations where the true treeis known, Huelsenbeck (1991) recognizedthat such deletion/restoration experimentsare one of the few means of testing whetherfossils have a signi�cant effect on tree topol-ogy. Inclusion of fossil taxa is particularlyimportant when studying the relationshipsof taxa separated by large stretches of timebecause fossils aid in the identi�cation of ho-moplasies that might otherwise go unrecog-nized (Doyle and Donoghue, 1987; Huelsen-beck, 1991). Essentially, fossils may break uplong branches.
Fossil taxa are particularly likely tobreak up long branches in the radiationof cetaceans and other ungulates. Morpho-logical and molecular evidence argues thatthe splits among Cetacea, Artiodactyla, andPerissodactyla are > 50 million years old.A fossil-based estimate for the divergenceof Cetacea from Artiodactyla is early Pale-ocene or very Late Cretaceous ( ~ 65 millionyears ago; Gingerich and Uhen, 1998); forPerissodactyla and Artiodactyla, Late Cre-taceous (Novacek, 1992b; but see also No-vacek et al., 1998). A molecule-based esti-mate for the divergence of Cetacea fromArtiodactyla is ~ 58 million years ago, and
that for Cetacea + Artiodactyla from Peris-sodactyla is ~ 83 million years ago (Kumarand Hedges, 1998). Thus, by all estimates ofdivergence, the split under consideration isancientand hence vulnerable to long-branchattraction problems. Mesonychians, whichrange from early Paleocene through lateEocene (McKenna and Bell, 1997), are fossilsthat meet two phylogenetically signi�cantcriteria (Huelsenbeck, 1991): They are rela-tively complete, and they have a time of ap-pearance relatively close to that of Cetacea.Many of the diverse array of exinct artio-dactyls, perissodactyls, and “condylarths”known ful�ll similar criteria.
Other studies have reinforced the im-portance of taxonomic sampling. Wheeler(1992) demonstrated on the basis of com-puter simulations that accuracy of clado-grams is more affected by number of taxaincluded than by model of evolution, num-ber of characters (speci�cally, length of nu-cleotide sequences), or rate of evolution. Al-though Kim (1996) argued that adding taxadoes not always increase phylogenetic accu-racy, Graybeal (1998) recognized that manyof the taxa added in his study did not breakup long branches. She found that phylo-genetic accuracy improved as the numberof taxa increased, even if simultaneouslythe number of characters decreased. Impor-tantly, the taxa added speci�cally broke uplong branches, and she emphasized that it isthe addition of such taxa that improves phy-logenetic accuracy, a result consistent withpaleontological studies.
We follow the methods of Doyle andDonoghue (1987) and Gauthier et al. (1988)and use deletion/restoration experimentswith fossil taxa to examine the effect of ex-tinction on tree topology. Indeed, investiga-tion of the morphological (again, primarilyosteological) signal alone is one of the fewways to maximize the empirical effect of ex-tinct clades on tree topology. If tree topologychanges on addition of morphological datafrom fossil taxa, one hypothesis explainingthis result is that it would also happen ifmolecular data were accessible for all extinctingroup taxa. Obviously, we cannot test thishypothesis empirically because of missingdata. However, the operational reality faced
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

464 SYSTEMATIC BIOLOGY VOL. 48
by total evidence analyses is that extinctioncreates a data partition that can in�uence re-sults and obscure phylogenetic signals. Be-cause osteology can be studied across ex-tinct and extant organisms, its signal alonedeserves consideration, for its potential tooffer an important perspective on relation-ships not apparent from neontological data.
Data Collection
Scoring all extant and extinct ungulategenera, particularly Artiodactyla, Perisso-dactyla, Cetacea, Mesonychia, and archaicungulates, was an unrealistic point of de-parture because, as noted above, some ofthese taxa are highly diverse. From amongthe clades mentioned above we chose 37genera to form the ingroup (Appendix 1),primarily following principles outlined byHillis (1998:5 [methods 3 and 4]), and as-sumed that each of these genera is mono-phyletic on the basis of the morphologi-cal similarity of the species within it. Theingroup consists primarily of fossil gen-era known from relatively complete speci-mens, chosen both because we believe theyare representative of forms close to thebasal morphotype of various extant ungu-late clades and because they are represen-tative of the diversity of ungulate cladesargued to be closest to Cetacea (Gentryand Hooker, 1988; Prothero et al., 1988;McKenna and Bell, 1997). We did not sampleextensively within paenungulates becausemolecular and morphological phylogeniesdescribed above do not argue for a closerelationship between Artiodactyla, Cetacea,and paenungulates . We did, however, in-clude several taxa that have been arguedon the basis of cladistic analysis (Thewis-sen and Domning, 1992) to be among themost primitive members of the paenungu-late clade (i.e., Meniscotherium, Phenacodus;see Appendix 1, “archaic ungulates”). Otherpaenungulates are assumed to be nestedamong the taxa sampled.
The artiodactyl sample includes theoldest member of this order, Diacodexis(Rose, 1982); an entelodont, Archaeotherium;an anthracothere, “Elomeryx”; an oreodon-toid, Agriochoerus; a camelid, Poebroth-erium; and several extant forms: Sus (pig),
Hippopotamus, Hexaprotodon (= Choeropsis,pygmy hippotamus), Ovis (sheep), Trag-ulus (chevrotain), and Camelus (camel)(McKenna and Bell, 1997). The taxonomyof anthracotheres is much in need of re-vision, and we place “Elomeryx” in quotesbecause we have also used specimens at-tributed to Bothriodon (Ancodus) to scorethe morphology of “Elomeryx”; these taxaappear to be very similar, are from simi-lar deposits, and may be synonyms. Oursample of perissodactyls includes Equus(horse), an extinct equid, Hyracotherium,and an extinct tapiroid, Heptodon. All rel-atively complete genera of mesonychiansare included in our analysis, as well asa number of extinct cetaceans—Pakicetus,Ambulocetus , and Remingtonocetus, amongothers—and two extant cetaceans: odon-tocete Tursiops (bottle-nosed dolphin) anda mysticete Balaenoptera (rorqual whale).Several relevant taxa, collectively describedas “archaic ungulates” (see below andAppendix 1) or “condylarths,” were alsoincluded.
One genus for which the assumptionof monophyly remains controversial is Pa-kicetus. We follow Thewissen and Hussain(1998) as to which specimens constitutePakicetus. These authors caution that someof these identi�cations remain poorly sub-stantiated. Where possible, original speci-mens were examined, but character cod-ing was supplemented from the literaturewhen original specimens were unavailable(Appendix 1).
Ordinal-level relationships of mammalsare incompletely resolved (Novacek, 1992b),making outgroup choice ambiguous ina study of this kind examining varia-tion across more than four orders. InNovacek’s (1992b) tree, the node be-low our ingroup (Artiodactyla, Perisso-dactyla, and Cetacea) is “Condylarthra,”a paraphyletic assemblage of primitive,herbivorous-omnivorous, placental mam-mals loosely allied with ungulates (Car-roll, 1988). Although “Condylarthra” is notspeci�cally designated as an outgroup inour analysis, our tree contains a num-ber of “condylarths”: Arctocyon, Hyposodus,Meniscotherium, Phenacodus, triisodontines,
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

1999 O’LEARY AND GEISLER—PHYLOGENY OF CETACEA 465
and mesonychians (Carroll, 1988), which ef-fectively polarize characters within Artio-dactyla, Perissodactyla, and Cetacea. Thesister taxon of “Condylarthra” is an unre-solved polytomy consisting of archontans,glires, carnivorans, and insectivorans (No-vacek, 1992b). We chose one outgroup fromthis assemblage, the insectivoran Leptictis,and two others at still lower nodes on themammalian tree: the primitive eutherianAsioryctes and the extant marsupial Didel-phis. We decided against using Tubuliden-tata as an outgroup, despite the hypothe-sized proximity of this order to our ingroup(Novacek, 1992b), because the highly de-rived dentition of tubulidentates would beuseless for polarizing dental characters inthe ingroup.
Our matrix heavily emphasizes osteolog-ical characters. We scored 123 morphologi-cal characters; 33 basicranial, 9 other cranial,45 dental, 29 postcranial, and 7 soft morpho-logical characters (Appendix 2). The amountof missing data for a given taxon rangedfrom 2% to 75% (average 29%; Appendix 3).Most (88, or 72%) of the characters were bi-nary, and 35 (28%) were multistate. Multi-state characters were treated in twoways: (1)all unordered, or (2) 24 of the 35 multistatecharacters ordered (20% of the total num-ber of characters, 68% of multistate charac-ters); these characters are speci�ed in Ap-pendices 2 and 3. Ordering was sequentialas follows: 0 « 1 « 2 « 3. Multistate char-acters were not ordered if we did not hy-pothesize a hierarchical transformation forthat character.
Treatment of multistate characters as or-dered or unordered constitutes an assump-tion about evolutionary process (Hauserand Presch, 1991; Wilkinson, 1992; Barrieland Tassy, 1993; Slowinski, 1993) that mustbe faced a priori in cladistic analysis. Un-ordered multistate characters assume thatone character state can transform directlyinto any other character state without pass-ing through an intermediate character state.Ordered multistate characters assume thattransformations do pass through such an in-termediate stage. Wilkinson (1992) arguedthat ordering certain characters is a logi-cal extension of Hennig’s auxiliary principle
and “explains the similarity between a sub-set of the character states in terms of synapo-morphy” (Wilkinson, 1992:380). Slowinski(1993) demonstrated that neither orderingnor unordering necessarily increases taxo-nomic congruence, one measure of phylo-genetic accuracy. He concluded that eitherway of treating multistate characters is valid(Slowinski, 1993:163) and proposed order-ing as favorable if a transformation seriesdescribes morphoclinal variation. The char-acters treated as ordered herecapture hierar-chical morphological change, and we arguethat treating these characters as unorderedwould constitute a loss of information im-portant for reconstructing this phylogeny.
Certain characters discussed as importantfor substantiating the monophyly of Artio-dactyla were not included in our study if wecould not score them in a consistent fashion(e.g., the relative expansion of the pars fa-cialis of the lacrimal bone). We excluded onesoft morphological character, relative elon-gation of the blastocyst (Thewissen, 1994;Geisler and Luo, 1998), because it has notbeen described in suf�cient detail for manyof the extant taxa in question. The hypoconeof artiodactyls has been described as non-homologous to the hypocones of various ofthe ungulates (Prothero et al., 1988). Thiscusp is thought to develop from the meta-conule in Artiodactyla and from the lingualcingulum in many other ungulates (Hunterand Jernvall, 1995). Instead of assuming apriori that the hypocones are nonhomolo-gous, we scored both the metaconule andthe hypocone on the basis of position onthe tooth: the hypocone being the cusp inthe distolingual corner of the upper molar,and the metaconule being the cusp posi-tioned more labially or anterolabially on adiagonal line between the metacone and theprotocone.
Outgroup taxa do not necessarily havestate 0 for every character (Appendix 3) fortworeasons: (1) we use several genera ratherthan a hypothetical ancestor as an outgroup,and there is morphological variation amongthe outgroups; and (2) certain ordered char-acters (e.g., character 46) branch in two di-rections from the hypothesized primitivestate, which is most easily coded by desig-
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

466 SYSTEMATIC BIOLOGY VOL. 48
nating the primitive state as 1 and the de-rived states as 0 and 2.
Character Independence
An assumption of parsimony analysis isthat the characters used in a phylogeneticanalysis (homology statements) are inde-pendent. As discussed by Kluge and Wolf(1993) independence in phylogenetic analy-sis must be addressed in two forms: (1) logi-cal independence of characters, and (2) con-current origin of the characters. The mostegregious violation of the assumption of in-dependence is to score redundant charac-ters, that is, those that fail the test of logi-cal independence. Farris (1983:20) describesan example of two characters that are notlogically independent as scoring “tarsal seg-ments and twice that number” separately.We strive not to include duplications of thisnature in our data matrix.
The second criterion for independence,concurrent origin, is much more dif�cultto verify (Kluge and Wolf, 1993), par-ticularly for fossil taxa. Rejection of thehypothesis that two synapomorphiesevolved independently requires knowledgeof tokogenetic relationships that “even ourbest-supported phylogenetic propositionsdo not provide” (Kluge and Wolf, 1993:192).Numerous biological processes (e.g., ge-netic, selective, developmental) may con-tribute to the non-independence of differentcharacters. However, as Kluge and Wolf(1993:192) contend, “the actual processesresponsible for nonindependent evolutioncannot be read from even the most detailedpatterns of organism relationship.” We in-terpret this to mean that similar distributionof character states in two or more charac-ters is not necessarily indicative of the non-independence of those characters. Hence,to dismiss covarying characters withoutexquisite knowledge of tokogenetic rela-tionships risks an a priori dismissal of phy-logenetic signal (character congruence).
We, therefore, include all logically inde-pendent characters in our analysis, eventwo characters that appear to covary ex-actly (i.e., characters 81 and 82). Exact co-variance does not seem suf�cient reason tocombine two characters in the matrix. These
characters cannot be scored for all taxa inthe matrix, and thus it is quite possible thatthe exact covariance may disappear as ourknowledge of the fossil record improves.Secondly, we are studying taxa whose ori-gin is separated by millions of years, mak-ing it very likely that we have not discov-ered, and have not scored, all relevant forms.Although in certain cases characters mayappear to have been acquired at the samenode (i.e., covary exactly), this may be a by-product of extinction, fossilization, and thescale of the analysis, not evidence of the non-independence of the synapomorphies. Thispaper contains references to basicranial,cranial, dental, postcranial, and soft mor-phological data partitions. These partitionsare, however, simply constructs to organizeinformation and are not meant at present toconvey any knowledge of biologically basedcharacter-relatedness.
Phylogenetic Analyses
The matrix was compiled in MacClade3.07 (Maddison and Maddison, 1992) andanalyzed by using either the heuristicor branch-and-bound search algorithms inPAUP 3.1.1 (Swofford, 1993), depending onthe size of the character-taxon matrix ina particular run. Once trees were foundin PAUP, character distributions were ana-lyzed in MacClade. Because of the large sizeof the data set, a branch-and-bound analysisin PAUP was possible only in runs that ex-cluded all fossils (runs 3 and 4; see below).All other searches were heuristic with thefollowing settings: addition sequence ran-dom; number of replications = 1,000; tree-bisection and reconnection (TBR) branch-swapping performed; MULPARS option ineffect; steepest descent option not in ef-fect; branches having maximum length 0collapsed to yield polytomies; trees un-rooted; uninformative characters excluded;and multistate taxa interpreted as polymor-phism. Trees were rooted with Didelphis asoutgroup. All tree lengths reported are cal-culated in PAUP 3.1.1 (Swofford, 1993).
The character-taxon matrix was subject tomaximum parsimony analysis under the fol-lowing parameters: run 1, certain multistatecharacters ordered (Appendices 2 and 3);
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

1999 O’LEARY AND GEISLER—PHYLOGENY OF CETACEA 467
run 2, all multistate characters unordered;run 3, extant taxa only, and certain multistatecharacters ordered; run 4, extant taxa only,and all multistate characters unordered; run5, certain multistate characters ordered, andevidence from Thewissen et al. (1998) in-cluded; run 6, all multistate characters un-ordered, and evidence from Thewissen et al.(1998) included. Runs 3 and 4 with extanttaxa only were constructed to mimic taxonsampling available in molecular analyses,thereby testing the effect of fossils on treetopology.
On the basis of available data, we testedthe hypothesis that the morphology of thecetacean astragalus is central to establishingartiodactyl monophyly (Milinkovitch andThewissen, 1997; Luckett and Hong, 1998)by adding in runs 5 and 6 new data onwhat are described as astragali of primitivecetaceans (Thewissen et al., 1998). The char-acters described by Thewissen et al. (1998)are found on two specimens, neither ofwhich is associated with diagnostic cetaceanmaterial. For this reason, we treat these datawith particular caution. These specimenspotentially provide data on three charactersthat are missing data in our initial matrix: thenavicular facet (character 104), the susten-tacular facet (character 105), and the lateralprocess (character 107). (The cetacean condi-tions for the astragalar canal [character 103]and the proximal astragalus [character 109],are known for Ambulocetus [Thewissen et al.,1996].) On the basis of the new bones, wecoded character 104 as a new state 3, �at(in Pakicetus); character 105 as state 0, nar-row (in Pakicetus); and character 107 as state1, absent (in Ambulocetus). The bone de-scribed by Thewissen et al. (1998) appearsto introduce new variability to character 105not orignally re�ected in our coding regime.Where our matrix emphasizes width of thischaracter, Thewissen et al. (1998) emphasizelength. This character is very likely in needof more detailed description in the future toaccommodate the variation in the ingroup.
We calculated decay values (Bremer, 1988)for nodes in the consensus trees from runs1 and 2 by conducting heuristic searches(with 100 replications, using TBR branch-swapping and random addition) in PAUP
3.1.1 (Swofford, 1993) and using constrainttrees to �nd trees that do not contain aparticular clade in question. The differ-ence in length between these trees and ourmost-parsimonious tree equals the decayindex for each node. We emphasize thatthe decay values are estimates because allsearches are heuristic. Estimated bootstrapvalues (Felsenstein, 1985) were calculated inPAUP * 4.0b1 (Swofford, 1998) for the un-ordered and ordered trees by using heuris-tic searches (1,000 replications) with sim-ple addition and TBR branch-swapping,and including groups compatible with the50% majority rule consensus. Optimiza-tion of characters was performed by usingboth ACCTRAN and DELTRAN options inPAUP 3.1.1. Finally, to compare the topologyfound in one of the more densely sampledmolecule-based analyses (Gatesy, 1997) withthe morphology-based results here, we useda backbone constraint tree in PAUP * 4.061to search for the most-parsimonious mor-phological tree under this molecular topo-logical constraint. This exercise provided anestimate of whether or not forcing the topol-ogy generated on the basis of molecular dataalone substantially increased the length ofthe treebased on morphology alone. This ex-periment was performed on the matrix withordered characters only.
RESULTS
Run 1, in which 20% of the total num-ber of characters (69% of multistate char-acters) were ordered (Fig. 2, Appendices 2and 3), produced two most-parsimonioustrees of 536 steps each, the strict consen-sus of which supported monophyly of eachthe following clades: Artiodactyla, Peris-sodactyla, Mesonychia, Mesonychidae, andCetacea. The sister taxon of Cetacea isMesonychia (Hapalodectidae and Mesony-chidae), and the sister group to the cetacean-mesonychyian clade is the triisodontinearctocyonid, Andrewsarchus. Among cladeswith extant members, Artiodactyla is moreclosely related to the mesonychian-cetaceanclade than is Perissodactyla. Within Artio-dactyla, Sus, the hippopotamids, the en-telodont, and the anthracothere are more
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

468 SYSTEMATIC BIOLOGY VOL. 48
FIGURE 2. Strict consensus of two most-parsimonious trees of 536 steps each from run 1 (CI = 0.384, RI = 0.696,HI = 0.698, RC = 0.267), 20% of total characters ordered (68% of multistate characters) (Appendices 2 and 3).Artiodactyla (A), Cetacea (C), Mesonychidae (D), Mesonychia (M), and Perissodactyla (P) are each monophyletic.Table 2 describes synapomorphies found with ACCTRAN and DELTRAN optimization at nodes marked withletters. Extant taxa = bold, extinct taxa = y. Numbers above branches are estimated decay values; numbers belowbranches are estimated bootstrap support.
closely related to each other than any is tothe ruminant clade, which consists of Ovis,Tragulus, Camelus, and Poebrotherium. Menis-cotherium, Phenacodus, and Hyopsodus form
a monophyletic clade with Perissodactyla.The ingroup overall is not monophyletic;the “condylarth” Arctocyon falls outside ofa clade that includes the insectivoran Lep-
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

1999 O’LEARY AND GEISLER—PHYLOGENY OF CETACEA 469
tictis. The data are not strong enough tounite the two triisodontine arctocyonids An-drewsarchus and Eoconodon as monophyletic,or even to draw Eoconodon into the ingroup,where it is hypothesized to belong based oncurrent taxonomy (McKenna and Bell, 1997).Estimated decay indices indicate that themonophyly of Artiodactyla is more stronglysupported than either the sister taxon rela-tionship between Mesonychia and Cetaceaor the clade Artiodactyla + (Andrewsarchus+ (Mesonychia + Cetacea))).
Run 2 (all characters unordered; Fig. 3)resulted in 36 most-parsimonious trees of505 steps each, the strict consensus of whichsupports monophyly of each of the follow-ing clades: Artiodactyla, Perissodactyla, andCetacea. The sister taxon of Cetacea is nowthe mesonychian Hapalodectes, the polaritywithin Mesonychidae has reversed in com-parison with Figure 2, and both Mesony-chia and Mesonychidae are paraphyleticwith respect to Cetacea. Two results occurin all most-parsimonious trees, as in run1: Poebrotherium and Camelus form a cladewith the ruminants (Tragulus and Ovis), andHyopsodus, Phenacodus, and Meniscotheriumform a clade with Perissodactyla (Fig. 2).The tree topology within Artiodactyla is rel-atively similar to that found in run 1, exceptthat the oreodontoid Agriochoerus joins theruminant clade.
Removing all fossils from the character-taxon matrix and treating certain charac-ters as ordered (run 3; Fig. 4a-c) recoversthree most-parsimonious trees of 173 stepseach, the strict consensus of which is a pa-raphyletic Artiodactyla with internal rela-tionships of the ingroup poorly resolved.The three most-parsimonious trees (Fig. 4)support a cetacean sister group relationshipwith Ovis or with Ovis + (Camelus + Tragu-lus). Running extant taxa only with all multi-state characters unordered (run 4; Fig. 4d-e)resulted in two most-parsimonious trees of150 steps each, again with a paraphyletic Ar-tiodactyla and Cetacea nested inside as thesister taxon of either hippopotamids or hip-popotamids + Sus.
The backbone constraint tree (Fig. 5b),which forced the most-parsimonious solu-tion for the morphological matrix congruent
with one topology generated on the basisof molecules (Gatesy, 1997), was 551 steps,15 steps longer than the most-parsimonioustree that was based on our morphologi-cal matrix with certain multistate charac-ters ordered (Fig. 2). Under this constraint,mesonychians are still the sister taxon ofCetacea, and this entire clade falls inside Ar-tiodactyla.
Optimizations
Synapomorphies for the tree found inrun 1 (certain characters ordered) calcu-lated with both ACCTRAN and DELTRANoptimization algorithms (Table 2) are de-scribed below. Such synapomorphies de-scribe the strongest support for particularnodes because they persist whether paral-lelism or reversals are favored. The cladeincluding Perissodactyla + (Artiodactyla +(Andrewsarchus + (Mesonychia + Cetacea)))is supported by four such synapomorphies(Table 2; Fig. 2: node a), which come frombasicranial, dental, and postcranial parti-tions of the data matrix. Artiodactyla +(Andrewsarchus + (Mesonychia + Cetacea))is supported by 12 synapomorphies (Fig. 2:node b) from basicranial, cranial, postcra-nial, and soft-morphological partitons ofthe data matrix. Mesonychia + Cetacea,however, is supported by only two synapo-morphies (Fig. 2: node d). This is duein large part to the fragmentary na-ture of Andrewsarchus, the outgroup tothis clade, which makes polarity cal-culations ambiguous. Cetacea is one ofthe more strongly supported nodes with15 synapomorphies from basicranial, cra-nial, and dental data (Fig. 2: node e).Mesonychia (Hapalodectes + Mesonychidae)(Fig. 2: node f), by contrast, is supportedby three dental synapomorphies, andMesonychidae by four synapomorphies,from basicranial, cranial, and dental datapartitions (Fig. 2: node g). Artiodactyla(Fig. 2: node j) is supported by 8 synapomor-phies from basicranial, dental, and postcra-nial partitions of the data matrix. Peris-sodactyla, and clades that include stemtaxa leading to it, are well-supportednodes with several synapomorphies each(Table 2).
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

470 SYSTEMATIC BIOLOGY VOL. 48
FIGURE 3. Strict consensus of 36 most-parsimonious trees, 505 steps each (CI = 0.406, RI = 0.694, HI = 0.679,RC = 0.282), all characters unordered (run 2). Artiodactyla (A), Cetacea (C), and Perissodactyla (P) are eachmonophyletic; Mesonychidae (“D”), and Mesonychia (“M”) are paraphyletic. Numbers above branches repre-sent estimated decay values, numbers below branches represent estimated bootstrap support, branches withoutbootstrap values are nodes recovered in < 5% of bootstrap replicates . Extant taxa are in bold; extinct taxa = y.Unequivocal synapomorphies supporting artiodactyl monophyly in all 36 trees are the following: 27. Alisphenoidcanal, present (1) ® absent (0); 32. Post-temporal canal, present (0) ® absent (1); 86. dp4 , resembles M1 (0) ®six-cusped (1); 92. Entepicondyle, wide (0) ® narrow (1); 93. Entepicondylar foramen, present (0) ® absent (1); 96.Proximal radius, two fossae (1) ® three fossae (2); 105. Sustentaculum, narrow (0) ® wide (1); 107. Lateral processof astragalus, present (0) ® absent (1); 109. Proximal astragalus, �at (0) ® deeply grooved (2). The unequivocalsynapomorphies supporting Hapalodectes + Cetacea are these: 37. Lacrimal tubercle, present (1) ® absent (0); 60.M3 , absent (3) ® equal to M2 in size (1); 83. Reentrant grooves, distal (2) ® proximal (0); and 95. Olecranonprocess, deep (1) ® shallow (0).
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

1999 O’LEARY AND GEISLER—PHYLOGENY OF CETACEA 471
FIGURE 4. Results of parsimony analyses with all extinct taxa removed from the data matrix (Appendix 3).(a-c) The three most-parsimonious trees of 173 steps each (CI = 0.618, RI = 0.590, HI = 0.422, RC = 0.365), based onthe matrix with 20% of multistate characters ordered (Appendices 2 and 3) (run 3); Cetacea is the sister taxon ofOvis in (a) and (b) or Ovis + (Camelus + Tragulus) in (c). Synapomorphies supporting the sister taxon relationshipbetween Cetacea and other taxa found with both ACCTRAN and DELTRAN algorithms are as follows: Tree a-1.Subarcuate fossa, present (0) ® absent (1); 15. Articulation of ectotympanic to squamosal, broad (0) ® absent(3); and 16. Exoccipital-ectotympanic contact, present (1) ® absent (0); tree b-1 and 15; tree c-26. Foramen ovale,medial, posterior wall formed by petrosal (2) ® anterior, posterior wall formed by alisphenoid (0); 94. Lengthof olecranon process, long (1) ® short (0); 113. Second metatarsal, unreduced (0) ® highly reduced (2); and 114.Fifth metatarsal, unreduced (0) ® highly reduced (2). (d and e) The two most-parsimonious trees of 150 stepseach (run 4) (CI = 0.660, RI = 0.617, HI = 0.387, RC = 0.407), based on the matrix with all multistate charactersunordered. Synapomorphies supporting Cetacea as the sister taxon of Sus + (Hippopotamus + Hexaprotodon) (treed) are these: 7. Mastoid, exposed (0) ® not exposed (1); 23. Postglenoid foramen, present, enclosed by squamosal(0) ® absent (2). The following support Cetacea as the sister taxon of Hippopotamus + Hexaprotodon (tree e): 1.Subarcuate fossa: present (0) ® absent (1); 15. Articulation of ectotympanic to squamosal: broad (0) ® absent (3);121. Hair: abundant (0) ® absent (1); and 122. Sebaceous glands: present (0) ® absent (1).
Because removal of extinct taxa from thedata matrix changed the topology of thetree (compare Figs. 2 and 4a-c, or Figs.3 and 4d-e), we examined which of thecharacters speci�cally supporting the sis-ter taxon relationship with Cetacea wereacting as homologies in the cladograms ofextant taxa alone (Fig. 4) but were homo-plasies when the fossils were included. (Figs.2 and 3; Table 2). These characters, listedin the caption to Figure 4, came from thecranial, postcranial, or soft-morphologicaldata partitions.
Addition of Astragalar Data from Thewissenet al. (1998)
When the data from Thewissen et al.(1998) are incorporated into the matrix (runs5 and 6), cetaceans are scored as having onecharacter that was shown above (Table 2) tobe a synapomorphy for Artiodactyla: char-acter 107, lateral process of the astragalusabsent. Adding these characters to the ma-trix with certain multistate characters or-dered (run 5) results in 2 trees of 539 stepseach with a topology no different from thatof Figure 2. Adding these characters to thematrix with all multistate characters un-ordered results in a concensus tree no dif-ferent from that found in Figure 3; that is,the new data do not change the toplogiesof the trees.
DISCUSSION
Results of these parsimony analyses showthat on the basis of 123 morphologicalcharacters (116 osteological and 7 soft mor-
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

472 SYSTEMATIC BIOLOGY VOL. 48
FIGURE 5. Comparison of backbone constraint treeto molecule-based tree, both addressing the questionof artiodactyl monophyly. (a) pruned tree from Gatesy(1997) used as the constraint; (b) strict consensus of ninemost-parsimonious trees of 551 steps each resultingfrom a heuristic search using the tree from (a) as a back-bone constraint and the matrix with certain multistatecharacters ordered. Extant taxa are in bold. Cetacea(C), Mesonychidae (D), Mesonychia (M), and Perisso-dactyla (P) are each monophyletic, “A” = Artiodactylahere is monophyletic only if Cetacea are included.
phological) from 10 extant and 30 extincttaxa, Artiodactyla is more closely relatedto Cetacea than is Perissodactyla. This con-clusion is supported when certain multi-state characters (20% of the total data set)are treated as ordered, when all multistatecharacters are treated as unordered, andwhen we exclude fossils from the analy-sis. We believe that results of runs withcertain multistate characters ordered betterre�ect hierarchical morphoclinal transfor-mations and relationships between differentcharacter states (Wilkinson, 1992; Slowin-ski, 1993), that are important to recon-structing phylogeny. With certain charac-ters ordered, the sister taxon of Cetacea isa monophyletic Mesonychia, as has been ar-gued on the basis of dental characters alone(O’Leary, 1998a). The triisodontine arctocy-onid Andrewsarchus is the stem taxon to thiscetacean-mesonychian clade, and a mono-phyletic Artiodactyla is the sister taxon tothat clade (Fig. 2). This tree also supportstraditional groupings within Artiodactyla(Gentry and Hooker, 1988): a ruminant-camel clade and a hippopotamid-suid clade,each with extinct relatives. That Cetacea ismore closely related to Artiodactyla thanto Perissodactyla corroborates the conclu-sions of virtually all molecule-based phylo-genetic analyses of this question noted inthe introduction, as well as several exclu-sively morphological descriptions or anal-yses (Gingerich et al., 1990; Novacek, 1992b;Thewissen and Hussain, 1993; Geisler andO’Leary, 1997; Geisler and Luo, 1998), butis incongruent with conclusions of severalother morphological analyses (Novacek andWyss, 1986; Wible, 1987; Prothero et al., 1988;Novacek, 1989; Prothero, 1993; Thewissen,1994).
When all multistate characters are treatedas unordered, the strict consensus of36 most-parsimonious trees also has amonophyletic Artiodactyla, but Mesony-chia, Mesonychidae, and triisodontinearctocyonids are paraphyletic. Other nodesat which this tree is congruent with thatfrom the run with certain multistatecharacters ordered are these: monophylyof Perissodactyla, the order of stem taxaleading to Perissodactyla (Meniscotherium,Phenacodus, Hyopsodus), a clade including
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

1999 O’LEARY AND GEISLER—PHYLOGENY OF CETACEA 473
TABLE 2. Character optimizations for particular nodes in the two most-parsimonious trees from run 2 (certainmultistate characters ordered, Fig. 2). Character state transformations were calculated by using both ACCTRANand DELTRAN algorithms in PAUP 3.1.1. Complete character state descriptions are given in Appendix 2. Indicatedare transformations found with (A) ACCTRAN only, (D) DELTRAN only, or (A/D) ACCTRAN and DELTRAN(in bold).
Node Character Transformation
a A 2. Tegmen tympani (0) Unin�ated ® (1) in�atedA/D 11. Ectotympanic (0) Ring ! (1) bullaA/D 60. M3 equal to M2 (2) Smaller than M2 ! (1) equalA 95. Olecranon fossa (0) Shallow ® (1) deepA/D 97. Distal radius (0) Single, concave fossa ! (1) scaphoid
and lunate fossaeA/D 115. Distal phalanges (0) Curved ! (1) straight
b A/D 16. Exoccipital-ectotympanic (0) Absent ! (1) presentcontactA/D 17. Sigmoid process (0) Absent ! (1) presentA 20. Squamosal part of external (0) Absent ® (1) deep grooveauditory meatusA/D 37. Lacrimal tubercle (1) Present ! (0) absentA 56. M1-M2 hypocone (1) Present ® (0) absentA 58. Paraconule on M2 (1) Present ® (0) absentA 59. Metaconule on M2 (1) Present ® (0) absentA/D 92. Entepicondyle (0) Wide ! (1) narrowA 93. Entepicondylar foramen (0) Present ® (1) absentA/D 99. Manus (0) Mesaxonic ! (1) paraxonicA/D 104. Navicular facet (0) Convex ! (1) saddle-shapedA/D 108. Pes (0) Mesaxonic ! (1) paraxonic.A/D 112. First metatarsal (1) Reduced ! (2) highly reducedA/D 118. Lumen (0) Unilocular ! (1) plurilocularA/D 119. Stomach lumen (0) Absent ! (1) presentA/D 120. Cavernous tissue of penis (0) Abundant ! (1) sparseA/D 123. Lung bronchi (0) Two ! (1) three
c D 20. Squamosal part of external (0) Absent ® (1) deep grooveauditory meatusA 22. Internal carotid foramen (0) Absent ® (1) presentA/D 23. Post-glenoid foramen (0) Enclosed by squamosal ! (1) medial to
petrosal/squamosal sutureA 25. Position of foramen for ramus (0) In petrosal/squamosal suture ® (1) anterolateralsuperior of stapedial artery to epitympanic recessA 26. Foramen ovale (1) Medial ® (0) anteriorA 30. Foramen rotundum (0) Absent ® (1) presentA 31. Mastoid foramen (0) Present ® (1) absentA 33. Preglenoid process (0) Absent ® (1) presentA 52. M2 metacone (0) Subequal to paracone ® (1) half the size of
paraconeD 56. M1-M2 hypocone (1) Present ® (0) absentA/D 57. Trigon basin (0) Broad ! (1) narrowA 71. M2 paraconid (0) Present ® (1) absentA 73. M3 paraconid (0) Present ® (1) absentA 78. Protoconid (0) Subequal to height of talonid ® (1) twice
height of talonidA 83. Reentrant grooves (1) Absent ® (0) proximalA 94. Olecranon process (0) Short ® (1) longA 95. Olecranon fossa (1) Deep ® (0) shallowA 98. Centrale (1) Absent ® (0) presentA 116. Distal phalanges (0) Compressed ® (1) broadA 121. Hair (0) Abundant ® (1) absentA 122. Sebaceous glands (0) Present ® (1) absent
(continued on next page)
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

474 SYSTEMATIC BIOLOGY VOL. 48
TABLE 2. (Continued)
Node Character Transformation
d D 2. Tegmen tymapani (0) Unin�ated ® (1) in�atedD 31. Mastoid foramen (0) Present ® (1) absentA 45. Embrasure pits (0) Absent ® (1) presentD 52. M2 metacone (0) Sub-equal to paracone ® (1) half size of
paraconeA 54. Lingual cingulum on M2 (0) Present ® (1) absentA/D 57. Trigon basin (1) Narrow ! (2) very narrowD 73. M3 paraconid (0) Present ® (1) absentD 83. Reentrant grooves (1) Absent ® (0) proximalA/D 84. Talonid basins (0) Basined ! (1) reducedA/D 85. M3 hypoconulid (0) Long ! (2) absentD 94. Olecranon process (0) Short ® (1) longD 116. Distal phalanges (0) Compressed ® (1) broad
e A 3. Anterior process of petrosal (0) Absent ® (1) presentA 4. Tensor tympani fossa (1) Circular pit, no groove ® (2) circular pit,
deep grooveD 4. Tensor tympani fossa (0) Elongate fossa ® (2) circular pit, deep grooveA 5. Sulcus for internal caroid (0) Present ® (1) absentarteryA/D 6. Proximal stapedial artery (0) Present ! (1) absentsulcusA/D 7. Mastoid process external (0) Present ! (1) absentexposureD 9. Facial nerve sulcus (0) Absent ® (2) anterior wall = mastoid processA 9. Facial nerve sulcus (3) Anterior wall formed by meatal tube ® (2)
anterior wall formed by mastoid processA/D 12. Pachyosteosclerotic (0) Absent ! (1) presentinvolucrumA/D 15. Articulation of ectotympanic (0) Broad ! (1) circular facetbulla to squamosalA/D 24. Foramen for ramus superior (0) Present ! (1) absentof stapedial arteryD 26. Foramen ovale (1) Medial to glenoid fossa, posterior wall =
alisphenoid ® (0) anterior to glenoid fossa,posterior wall formed by alisphenoid
A/D 27. Alisphenoid canal (1) Present ! (0) absentA 33. Preglenoid process (1) Present ® (0) absentA 35. Postorbital bar (0) Absent ® (1) present, almost completeA 44. Premaxillae (0) Short ® (1) elongateA/D 48. P4 protocone (0) Present ! (1) absentA/D 49. P4 paracone (0) Equal to height of M1 paracone ! (1) twice height
of M1 paraconeA 52. M2 metacone (1) Half the size of the paracone ® (2) highly
reducedA/D 67. Lingual cingulid (0) Absent ! (1) presentA/D 68. M1 paraconid (0) Present ! (1) absentA/D 70. M2 paraconid (0) Present ! (1) absentA 72. M3 paraconid (0) Present ® (1) absentA/D 74. M1 metaconid (0) Present ! (1) absentA/D 75. M2 metaconid (0) Present ! (1) absentA 76. M3 metaconid (0) Present ® (1) absentA/D 87. Elongate shearing facets (0) Absent ! (1) presentA 91. Scapular spine (0) Acromion overhangs glenoid ® (2) acromion
does not overhang glenoidA 96. Proximal radius (1) Two fossae ® (0) one fossa
(continued on next page)
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

1999 O’LEARY AND GEISLER—PHYLOGENY OF CETACEA 475
TABLE 2. (Continued)
Node Character Transformation
f A 14. Median furrow of tympanic bulla (1) Median notch ® (0) absentA 19. Ectotympanic part of meatal tube (1) Short ® (2) longD 30. Foramen rotundum (0) Absent ® (1) presentD 33. Preglenoid process (0) Absent ® (1) presentA 41. Mandibular condyle height (1) Even with dentition ® (0) below dentitionA/D 50. P4 metacone (0) Absent ! (1) presentA/D 53. M1 parastyle (0) Absent ! (2) strongD 54. Lingual cingulum (0) Absent ® (1) presentA/D 69. M1 paracristid position (0) Lingual ! (1) anteriorD 71. M2 paracristid position (0) Lingual ® (1) anteriorA 90. Number of sacrals (2) Four ® (1) two or threeA 93. Entepicondylar foramen (1) Absent ® (0) present
g D 4. Tensor tympani fossa (0) Elongate fossa ® (1) circular pitA 9. Facial nerve sulcus (3) Anterior wall formed by meatal tube ® (1)
anterior wall formed by squamosalA 22. Internal carotid foramen (1) Present ® (0) absentA/D 23. Post-glenoid foramen (1) Present ! (2) absentD 25. Foramen for ramus superior of (0) In petrosal/squamosal suture ® (1)stapedial artery anterolateral to epitympanic recessD 26. Foramen ovale (1) Medial to glenoid fossa, posterior wall formed by
alisphenoid ® (0) anterior to glenoid fossaA/D 29. Posterior opening (0) Separated from foramen ovale ! (1) in aof alisphenoid canal recess with the foramen ovaleA 37. Lacrimal tubercle (0) Absent ® (1) presentD 41. Height of mandibular condyle (1) Even with dentition ® (0) below dentitionA 45. Embrasure pits (1) Present ® (0) absentA/D 60. M3 size (1) Equal to M2 ! (2) smallA/D 83. Reentrant grooves (0) Proximal ! (2) distalA/D 95. Olecranon fossa (0) Shallow ! (1) deep
h D 2. Tegmen tympani (0) Unin�ated ® (1) in�atedA 14. Median furrow of tympanic bulla (1) Median notch ® (2) anteroposterior furrowA 41. Condyle (1) Even with dentition ® (2) superior to dentitionD 58. Paraconule on M2 (0) Absent ® (1) presentD 59. Metaconule on M2 (0) Absent ® (1) presentA/D 65. P3 metaconid (0) Absent ! (1) presentA/D 66. P4 metaconid (0) Absent ! (1) presentA/D 77. M1-M2 hypoconulid (0) Absent ! (1) presentD 78. Protoconid (1) Twice height of talonid ! (0) subequal to
height of talonidD 90. Number of sacral vertebrae (1) Two or three ® (2) fourD 95. Olecranon fossa (0) Shallow ® (1) deepA/D 106. Astragalar-cuboid contact (1) Present ! (0) absent
i A 4. Fossa for tensor tympani (0) Elongate fossa ® (1) circular pitD 5. Transpromontorial sulcus for (0) Present ® (1) absentinternal carotid arteryD 6. Proximal stapedial artery (0) Present ® (1) absentD 53. M1 parastyle (0) Absent ® (1) weakA/D 79. Protoloph (0) Absent ! (1) presentA 90. Number of sacral vertebrae (2) Four ® (3) �ve or sixA 91. Scapular spine (0) Large acromion ® (1) small acromionA/D 92. Entepicondyle (0) Wide ! (1) narrowA/D 93. Entepicondylar foramen (0) Present ! (1) absentA/D 103. Astragalar canal (0) Present ! (1) absentA/D 104. Navicular facet of (0) Convex ! (1) saddle-shapedastragalus
(continued on next page)
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

476 SYSTEMATIC BIOLOGY VOL. 48
TABLE 2. (Continued)
Node Character Transformation
A/D 109. Proximal astragalus (1) Grooved ! (2) deeply groovedA/D 112. First metatarsal (1) Reduced ! (2) highly reducedA/D 114. Fifth metatarsal (1) Reduced ! (2) highly reduced
j A 2. Tegmen tympani (1) In�ated ® (0) unin�atedD 4. Tensor tympani fossa (0) Elongate fossa ® (1) circular pitA/D 27. Alisphenoid canal (1) Present ! (0) absentA/D 32. Post-temporal canal (0) Present ! (1) absentA/D 36. Frontal and maxillary (0) Absent ! (1) presentcontactD 78. Protoconid (1) Twice height of talonid ® (0) subequal to
height of talonidA/D 86. dp4 (0) Resembles M1 ! (1) six-cuspedA 91. Acromion (0) Large ® (1) smallD 93. Entepicondylar foramen (0) Present ® (1) absentD 95. Olecranon fossa (0) Shallow ® (1) deepA/D 96. Proximal radius (1) Two fossae ! (2) three fossaeD 98. Centrale (0) Present ® (1) absentA/D 101. Third trochanter (0) Present ! (1) highly reducedA/D 104. Navicular facet of (1) Saddle-shaped ! (2) trochleatedastragalusA/D 105. Sustentacular width (0) Narrow ! (1) wideA 106. Astragalar-cuboid contact (1) Small ® (2) largeA/D 107. Lateral process of (0) Present ! (1) absentastragalusA 109. Proximal astragalus (0) Flat ® (2) deeply grooved
camels and ruminants, and one includinghippopotamids, Sus, and fossil relatives.
Milinkovitch and Thewissen (1997) ar-gued that determination of the sister groupof Cetacea hinges strongly on the recov-ery of an astragalus of a primitive cetacean(e.g., Pakicetus, Ambulocetus). Two fragmen-tary astragali argued to be cetaceans on thebasis of faunal data have recently been de-scribed (Thewissen et al., 1998). These bonesare not, however, associated with diagnos-tic cetacean material, and we treat their im-pact on the phylogeny of Cetacea with somecaution. The new fossils are extremely frag-mentary, and we have concluded that thesebones introduce new and possibly autapo-morphic character states not previously rec-ognized. Addition of these new fossils doesnot change the tree topology or the num-ber of trees found whether multistate char-acters are treated as ordered or unordered.These bones have been described as exhibit-ing more similarities to artiodactyls thanto mesonychians (Thewissen et al., 1998);nonetheless, the invarianceof our resultmay
be related to the fact that artiodactyl mono-phyly is supported by basicranial, den-tal, and postcranial synapomorphies, notsimply synapomorphies based on anklemorphology.
As found in other phylogenetic analy-ses (Gauthier et al., 1988), removing fossilsfrom an analysis does not simply resultin a pruned version of the tree generatedfrom the matrix with extinct and extant taxa,but instead results in a tree with a differenttopology. When all extinct taxa are removedfrom the character-taxon matrix, mimick-ing the taxon sampling of many molecu-lar analyses, we consistently recover artio-dactyl paraphyly with Cetacea as the sistertaxon of Ovis, a clade including Ovis +(Camelus+ Tragulus), hippopotamids, or hip-popotamids + Sus. Characters supportingthese sister taxon relationships with Cetaceacome from a variety of anatomical systemsand are interpreted as homoplasies in themore complete matrix that includes extantand extinct taxa. Interestingly, artiodactylparaphyly is also the result found in most
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

1999 O’LEARY AND GEISLER—PHYLOGENY OF CETACEA 477
molecule-based analyses of the phyloge-netic position of cetaceans, studies that can-not sample extinct taxa. The topology sup-ported in the molecule-based analysis ofGatesy (1997) would increase the length ofthe most-parsimonious tree based on ourmatrix by 15 steps, a substantial number ina tree based on only 123 characters.
Given a conservative estimate that 89% ofthe ingroup genera relevant to the phyloge-netic position of Cetacea are extinct, we pro-pose that analysis of a morphology-based,largely osteological, phylogenetic signal isparticularly important, because extinct taxacan be maximally effective in such an analy-sis. We cannot test the effect of most molec-ular data on tree topology, because they arenot available for mesonychians, triisodon-tine arctocyonids, other “condylarths,” lep-ticitids, basal artiodactyls, or basal perisso-dactyls. These extinct taxa may have evenmore information on branching sequencesand character transformation than do extanttaxa, because by virtue of their antiquity theextinct taxa have had less time to evolvehomoplasies.
The stem taxa that we have addedhere, mesonychians—“condylarths,” tri-isodontines, and basal artiodactyls andperissodactyls—introduce so much data oncharacter transformation that they over-turn nodes supported in most-parsimonioustrees that are based on extant taxa alone(Figs. 2–4). These stem taxa simultaneouslyintroduce substantial homoplasy: The con-sistency index (CI) for the trees based on ex-tant taxa alone (Fig. 4a-c) is 0.618, whereasthat for the more complete matrix (extant+ extinct) taxa (Fig. 2) is 0.391. Adding in-group taxa to a phylogenetic problem, as wehave done, typically increases homoplasy(Sanderson and Donoghue, 1989), some-thing that may affect the bootstrap and de-cay values of the �nal tree. Strict compar-isons between decay and bootstrap valuesfound in this study and those in molecule-based studies are not a means of assess-ing relative robustness of the different phy-logenetic hypotheses they support becausethe trees are derived from different char-acters and taxa. Values presented here forthe decay and bootstrap are lower than
those found in a molecule-based analysisof Gatesy (1997), which supports a sistertaxon relationship between hippopotamidsand cetaceans, not artiodactyl monophyly. Ithas been shown elsewhere that addition ofan extra ingroup taxon with homoplasies toa parsimony analysis can reduce the stabil-ity of the ingroup (i.e., make it decay faster)(Novacek, 1991). However, if that taxon ispart of the ingroup, it simultaneously con-tributes to an increase in understanding ofcharacter transformation and relationships.Given such effects, Novacek (1991:347) ar-gued that “cladistic results arenotuniformlyamenable to evaluation by simple measuresof the stability of their components.” Suchmeasures include the bootstrap and decayindices. Novacek (1991) argued further thataddition of stem taxa may elucidate charac-ter transformation at the price of destabiliz-ing certain nodes.
In an important effort to synthesize in-formation on differing molecular and mor-phological hypotheses of cetacean origins,Gatesy et al. (1996) proposed three hy-potheses of relationship incorporating ex-tinct and extant taxa: (1) cetaceans andmesonychians are derived artiodactyls; (2)cetaceans are derived artiodactyls and thesister group of hippopotamids, and mesony-chians are a separate clade outside of Ar-tiodactyla; and (3) Artiodactyla is mono-phyletic and mesonychians are the sistertaxon of cetaceans. On the basis of all themorphological evidence presented here, nei-ther mesonychians nor cetaceans are de-rived artiodactyls (hypotheses 1 and 2),making hypothesis 3 the best explanationof the morphological data. Hypothesis 3renders as homoplasies the six morpho-logical and behavioral synapomorphies ofhippopotamids and cetaceans proposed byGatesy et al. (1996). These include absenceof hair, aquatic habitat, and underwatervocalization.
These results suggest future avenues ofinvestigation likely to impact the phylo-genetic position of Cetacea. The triisodon-tine arctocyonid Andrewsarchus does nothave the close relationship to Cetacea pro-posed by Prothero et al. (1988), but insteadfalls outside of a mesonychian-cetacean
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

478 SYSTEMATIC BIOLOGY VOL. 48
clade largely on the basis of dental evi-dence, as argued by O’Leary (1998a) . An-drewsarchus forms part of a paraphyleticgrouping with Eoconodon, but expandedinvestigation of the triisodontine arctocy-onids, a relatively primitive and poorlyknown group, is necessary before the pa-raphyly of this group should be recognizedby taxonomic changes. In particular, knowl-edge of such features as the morphologyof the distal phalanges of Hapalodectes andEoconodon will be important for establish-ing their positions within ungulates. The re-sult that the insectivoran Leptictis, an out-group taxon, falls closer to our ingroup thandoes the primitive “condylarth” Arctocyonalso warrants further testing and may bean artifact of including only one insectivo-ran. It is not clear that the hypocone in allmembers of Artiodactyla is derived fromthe metaconule, because both the hypoconeand the metaconule are present togetherin the clade comprising Archaeotherium,“Elomeryx,” and Sus (Appendix 3). Phylo-genetic analyses focusing on the relation-ships within Cetacea (Uhen, 1996) do notsupport the positions of Basilosaurus andDorudon found in this analysis but insteadplace Dorudon closer to extant cetaceans. Be-cause the aim of this study is to resolve morebasal nodes in the ungulate tree, we suspectthat the addition of more characters or morebasilosaurid cetaceans to this matrix mayresult in the topology supported in Uhen(1996).
Results based on the data set with cer-tain multistate characters ordered (Fig. 2)require that cetaceans diverged by themiddle Paleocene (Torrejonian), the timeof appearance of the oldest mesonychian,Dissacus, a result also supported by otherphylogenetic analyses (Geisler and Luo,1998; O’Leary, 1998a). This tree topologyspeci�es the existence of a cetacean ghostlineage ~ 10 million years long that re-mains to be discovered, something alsosuggested by the large number of synapo-morphies uniting cetaceans (Table 2; Fig. 2:node e). This estimate of the time of ori-gin of Cetacea is greater than likelihood-based estimates (Gingerich and Uhen,1998), which place the origin of Cetacea
in the Eocene, much closer to the �rstappearance of archaeocetes in the fossilrecord.
As emphasized by the count of extinctand extant taxa relevant to the question ofthe position of Cetacea within Mammalia(Table 1), this analysis only begins to sam-ple the diverse array of taxa likely to impactthe position of Cetacea within Mammalia.Besides numerous extinct artiodactyls andperissodactyls, 12 other mesonychian gen-era and �ve other triisodontine genera,known from fragmentary remains, are likelyto affect the relationships presented here(McKenna and Bell, 1997). Furthermore,located either within the ungulate clade oras stem taxa to it are 65 other “condy-larth” genera (McKenna and Bell, 1997),many of which are also poorly knownand have not been scored here. Other crit-ical taxa include the pakicetid Nalacetus,which has been argued to have the mostprimitive dental morphology of any knowncetacean (O’Leary, 1998a; Thewissen andHussain, 1998); Chriacus (Rose, 1996), whichexhibits modi�cations of the postcranialskeleton that suggest a link between arctocy-onid “condylarths” and basal artiodactyls;Microclaenodon, a triisodontine arctocyonid,which is the �rst outgroup to Mesonychiaon the basis of dental evidence (O’Leary,1998a), but which is known from little othermaterial; and Sinonyx (Zhou et al., 1995), amesonychian from the Paleocene of Asia,which is well-preserved postcranially butremains undescribed. Discovery and de-scription of material of these key fossils willbe crucial for testing the stability of the treetopologies presented here.
ACKNOWLEDGMENTS
We acknowledge the generosity of many colleagueswho provided helpful discussion and access to speci-mens, including undescribed fossil material: P. D. Gin-gerich, R. E. Heinrich, E. Heizmann, L. T. Holbrook,S. G. Lucas, W. P. Luckett, D. Lunde, Z. Luo, R. D. E.MacPhee, P. J. Makovicky, M. C. McKenna, M. J. No-vacek, K. D. Rose, A. E. Sanders, N. B. Simmons, andJ. G. M. Thewissen. We are particularly grateful to J. G.M. Thewissen for unpublished data on Ambulocetus, toM. D. Uhen for assisting with the coding of Dorudon,to L. B. Nash for preparing Figure 1, and to C. Heezywho provided data on the postorbital bars of several
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

1999 O’LEARY AND GEISLER—PHYLOGENY OF CETACEA 479
ungulates. M. J. Novacek, C. F. Ross, and D. M. Sabatinigenerously allowed us to conduct hours of phyloge-netic analysis on their computers. R. J. Asher, J. Gatesy,G. J. P. Naylor, M. J. Novacek, C. F. Ross, N. B. Simmons,J. B. Slowinski, and W. C. Wheeler read and greatly im-proved earlier versions of the manuscript.
REFERENCES
ADACHI, J., AND M. HASEGAWA. 1996. Instabil-ity of quartet analyses of molecular sequencedata by the maximum likelihood method: TheCetacea/Artiodactyla relationships. Mol. Phylo-genet. Evol. 6:72–76.
ARNASON, U., A. GULLBERG, AND B.WIDEGREN. 1991. Thecomplete nucleotide sequence of the mitochondrialDNA of the �n whale, Balaenoptera physalus. J. Mol.Evol. 33:556–568.
BARNES, L. G. 1984. Whales, dolphins and porpoises:Origin and evolution of the Cetacea. Pages 139–154in Mammals. Notes for a short course, Studies in Ge-ology 8(1-4) (P. D. Gingerich and C. E. Badgely, eds.).Department of Geological Sciences, Univ. Tennessee,Knoxville.
BARRIEL, V., AND P. TASSY. 1993. Characters, observa-tions and steps: Comment on Lipscomb’s “Parsi-mony, homology and the analysis of multistate char-acters.” Cladistics 9:223–232.
BEINTEMA, J. J., W. M. FITCH, AND A. CARSANA.1986. Molecular evolution of pancreatic-type ri-bonucleases. Mol. Biol. Evol. 3:262–275.
BEINTEMA, J. J., C. SCHULLER, M. IRIE, AND A. CARSANA.1988. Molecular evolution of theribonuclease super-family. Prog. Biophys. Mol. Biol. 51:165–192.
BOYDEN, A., AND D. GEMEROY. 1950. The relative po-sition of Cetacea among the orders of Mammalia asindicated by precipitin tests. Zoologica 35:145–151.
BREMER, J. L. 1904. On the lung of the opossum. Am.J. Anat. 3:67–73.
BREMER, K. 1988. The limits of amino acid sequencedata in angiosperm phylogenetic reconstruction.Evolution 42:795–803.
CARROLL, R. L. 1988. Vertebrate paleontology andevolution. W. H. Freeman, New York.
CATZEFLIS, F. 1993. Mammalian phylogeny: Morphol-ogy and molecules. Trends Ecol. Evol. 8:340–341.
CHOW, M. 1959. A new arctocyonid from the upperEocene of Lushih, Honan. Vertebr. Palasiat. 3:133–138.
CIFELLI, R. L. 1982. Thepetrosal structure of Hyopsoduswith respect to that of some other ungulates, and itsphylogenetic implications . J. Paleontol. 56:795–805.
CZELUSNIAK, J., M. GOODMAN, B. F. KOOP, D. O. TAGLE,J. SHOSHANI, G. BRAUNITZER, T. K. KLEINSCHIMDT , W.W. DE JONG, AND G. MATSUDA. 1990a. Perspectivesfrom amino acid and nucleotide sequences on cladis-tic relationships among higher taxa of Eutheria.Pages 545–572 in Current mammalogy, volume 2 (H.H. Genoways, ed.). Plenum, New York.
CZELUSNIAK, J., M. GOODMAN, N. D. MONCRIEF, AND S.M. KEHOE. 1990b. Maximum parsimony approachto construction of evolutionary trees from aligned
homologous sequences. Methods Enzymol. 183:601–615.
DAUDT,W. 1898. Beitragezur Kenntnis des Urogenita-lapparates der Cetaceen. Jena. Z. Naturwiss. 32:231–312.
DE JONG, W. W. 1985. Superordinal af�nities of Ro-dentia studied by sequence analysis of eye lensproteins. Pages 211–226 in Evolutionary relation-ships among rodents: A multidisciplinary analysis(W. P. Luckett and J.-L. Hartenberger, eds.). Plenum,New York.
DONOGHUE, M. J., J. A. DOYLE, J. GAUTHIER, A. G. KLUGE,AND T. ROWE. 1989. The importance of fossils inphylogeny reconstruction. Annu. Rev. Ecol. Syst.20:431–460.
DOYLE, J. A., AND M. J. DONOGHUE. 1987. The impor-tance of fossils in elucidating seed plant phylogenyand macroevolution. Rev. Palaeobot. Palynol. 50:63–95.
EERNISSE, D. J., AND KLUGE, A. G. 1993. Taxonomiccongruence versus total evidence and amniote phy-logeny inferred from fossils, molecules and morphol-ogy. Mol. Biol. Evol. 10:1170–1195.
FARRIS, J. S. 1983. The logical basis of phylogeneticanalysis. Pages 1–36 in Advances in Cladistics, Vol.2 (N.I. Platnick and V.A. Funk, eds.). Columbia Uni-versity Press.
FELSENSTEIN , J. 1978. Cases in which parsimony orcompatibility methods will be positively misleading.Syst. Zool. 27:401–410.
FELSENSTEIN , J. 1985. Con�dence limits on phyloge-nies: An approach using the bootstrap. Evolution39:783–791.
FRAAS, E. 1904. Neue Zeuglodonten aus dem unterenMitteleocan vom Mokattam bei Cairo. Geol. Palaeon-tol. Abh. 6:199–220.
GASKIN, D. E. 1978. Form and function in the diges-tive tract and associated organs in Cetacea, with aconsideration of metabolic rates and speci�c energybudgets. Oceanogr. Mar. Biol. Ann. Rev. 16:313–345.
GATESY, J. 1997. More DNA support for a Cetacea/Hippopotamidae clade: The blood-clotting proteingene gamma-�brinogen. Mol. Biol. Evol. 14:537–543.
GATESY, J., C. HAYASHI, M. A. CRONIN, AND P.ARCTANDER. 1996. Evidence from milk casein genesthat cetaceans are close relatives of hippopotamid ar-tiodactyls. Mol. Biol. Evol. 13:954–963.
GAUTHIER, J., A. G. KLUGE, AND T. ROWE. 1988. Am-niote phylogeny and the importance of fossils.Cladistics 4:105–209.
GAZIN, C. L. 1965. A study of the early Tertiary condy-larthran mammal Meniscotherium. Smithson. Misc.Collect. 149:1–98.
GAZIN, C. L. 1968. A study of the Eocene condy-larthran mammal Hyopsodus. Smithson. Misc. Col-lect. 153:1–90.
GEISLER, J. H., AND Z. LUO. 1996. The petrosal and in-ner ear of Herpetocetus sp. (Mammalia, Cetacea) andtheir implications for the phylogeny and hearing ofarchaic myticetes. J. Paleont. 70:1045–1066.
GEISLER, J. H., AND Z. LUO. 1998. Relationships ofCetacea to terrestrial ungulates and the evolution
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

480 SYSTEMATIC BIOLOGY VOL. 48
of cranial vasculature in Cete. Pages 163–212 inThe emergence of whales (J. G. M. Thewissen, ed.).Plenum, New York.
GEISLER, J. H., AND M. A. O’LEARY. 1997. A phylogenyof Cetacea, Artiodactyla, Perissodactyla, and archiacungulates: The morphological evidence. J. Vertebr.Paleontol. 17-(Suppl. to No. 3):48A.
GEISLER, J. H., A. E. SANDERS, AND Z. LUO. 1996. Anew protocetid cetacean from the Eocene ofSouth Carolina, U.S.A.: Phylogenetic and bio-geographic implications . Paleontol. Soc. Spec.Publ. 8:139.
GENTRY, A. W., AND J. J. HOOKER. 1988. The phy-logeny of the Artiodactyla. Pages 235–272 in Thephylogeny and classi�cation of the tetrapods, vol-ume 2. Mammals Syst. Assoc. Spec. Vol. 35B(M. J. Benton, ed.). Clarendon Press, Oxford,England.
GINGERICH, P. D., M. ARIF, AND W. C. CLYDE. 1995. Newarchaeocetes (Mammalia, Cetacea) from the middleEocene Domanda Formation of the Sulaiman Range;Punjab (Pakistan). Contrib. Mus. Paleontol. Univ.Mich. 29:291–230.
GINGERICH, P. D., AND D. E. RUSSELL. 1981. Pakicetus in-achus, a new archaeocete (Mammalia, Cetacea) fromthe early-middle Eocene Kuldana Formation of Ko-hat (Pakistan). Contrib. Mus. Paleontol. Univ. Mich.25:235–246.
GINGERICH, P. D., AND D. E. RUSSELL . 1990. Dentitionof the early Eocene Pakicetus (Mammalia, Cetacea).Contrib. Mus. Palentol. Univ. Mich. 28:1–20.
GINGERICH, P. D., B. H. SMITH, AND E. L. SIMONS.1990. Hind limbs of Eocene Basilosaurus: Evidenceof feet in whales. Science 249:154–157.
GINGERICH, P. D., AND M. D. UHEN. 1998. Likelihoodestimation of the time of divergence of Cetacea andArtiodactyla. Paleontol. Electron. 2:1–47.
GOODMAN, M., J. CZELUSNIAK, AND J. E. BEEBER.1985. Phylogeny of primates and other eutherian or-ders: A cladistic analysis using amino acid and nu-cleotide sequence data. Cladistics 1:171–185.
GRAUR, D. 1993a. Molecular phylogeny and thehigher classi�cation of eutherian mammals. TrendsEcol. Evol. 8:141–147.
GRAUR, D. 1993b. Reply from D. Graur. Trends Ecol.Evol. 8:341–342.
GRAUR, D., AND D. G. HIGGINS. 1994. Molecular evi-dence for the inclusion of cetaceans within the orderArtiodactyla. Mol. Biol. Evol. 11:357–364.
GRAYBEAL , A. 1998. Is it better to add taxa or charac-ters to a dif�cult phylogenetic problem? Syst. Biol.47:9–17.
GUTHRIE, D. A. 1967. The mammalian fauna ofthe Lysite Member, Wind River Formation (earlyEocene), of Wyoming. Mem. South. Calif. Acad. Sci.5:1–53.
HASEGAWA, M., AND J. ADACHI. 1996. Phylogenetic po-sition of cetaceans relative to artiodactyls: Reanalysisof mitochondrial and nuclear sequences. Mol. Biol.Evol. 13:710–717.
HAUSER, D. L., AND W. PRESCH. 1991. The effect ofordered characters on phylogenetic reconstruction.Cladistics 7:243–265.
HILLIS, D. M. 1998. Taxonomic sampling, phyloge-netic accuracy, and investigator bias. Syst. Biol.47:3–8.
HONEYCUTT, R. L., M. A. NEDBAL , R. M. ADKINS, AND L.L. JANECEK. 1995. Mammalian mitochondrial DNAevolution: A comparison of the cytochrome b and cy-tochrome c oxidase II genes. J. Mol. Evol. 40:260–272.
HUELSENBECK , J. P. 1991. When are fossils better thanextant taxa in phylogenetic analysis? Syst. Biol.40:458–469.
HUNTER, J. P., AND J. JERNVALL. 1995. The hypocone as akey innovation in mammalian evolution. Proc. Natl.Acad. Sci. USA. 92:10718–10722.
IRWIN, D. M., AND U. ARNASON. 1994. Cytochrome bgene of marine mammals: Phylogeny and evolution.J. Mammal. Evol. 2:37–55.
IRWIN, D. M., T. D. KOCHER, AND A. C. WILSON.1991. Evolution of the cytochrome b gene of mam-mals. J. Mol. Evol. 32:128–144.
KELLOGG, A. R. 1936. A review of the Archaeoceti.Carnegie Inst. Washington Publ. 482:1–366.
KIELAN-JAWOROWSKA, Z. 1977. Evolution of the the-rian mammals in the Late Cretaceous of Asia. PartII. Postcranial skeleton in Kennalestes and Asioryctes.Palaeontol. Pol. 37:65–83.
KIELAN-JAWOROWSKA, Z. 1981. Evolution of the the-rian mammals in theLate Cretaceous of Asia. Part. IV.Skull structure in Kennalestes and Asioryctes. Palaeon-tol. Pol. 42:25–78.
KIM, J. 1996. General inconsistency conditions formaximum parsimony: Effects of branch lengths andincreasing numbers of taxa. Syst. Biol. 45:363–374.
KITTS, D. B. 1956. American Hyracotherium (Perisso-dactyla, Equidae). Bull. Am. Mus. Nat. Hist. 110:1–60.
KLUGE, A. G. 1989. A concern for evidence and aphylogenetic hypothesis of relationships among Ep-icrates (Boidae, Serpentes). Syst. Zool. 38:7–25.
KLUGE, A. G., AND A. J. WOLF. 1993. Cladistics: What’sin a word? Cladistics 9:183–199.
KUMAR, S., AND B. HEDGES. 1998. A moleculartimescale for vertebrate evolution. Nature 392:917–920.
KUMAR, K., AND A. SAHNI. 1986. Remingtonocetus haru-diensis, new combination, a middle Eocene archaeo-cete (Mammalia, Cetacea) from Western Kutch, India.J. Vertebr. Paleontol. 6:326–349.
LANGER, P. 1988. The mammalian herbivore stomach:Comparative anatomy, function and evolution. Fis-cher, Stuttgart.
LI, W.-H. 1997. Molecular evolution. Sinauer, Sunder-land, Massachusetts.
LUCKETT, W. P., AND N. HONG. 1998. Phylogenetic re-lationships between the orders Artiodactyla andCetacea: A combined assessment of morphologicaland molecular evidence. J. Mammal. Evol. 5:127–182.
LUO, Z., AND K. MARSH. 1996. Petrosal (periotic) andinner ear of a Pliocene kogiine whale (Kogiinae,Odontoceti): Implications on realionships and hear-ing evolution of toothed whales. J. Vertebr. Paleontol.16:328–248.
MACFADDEN, B. J. 1992. Fossil horses. CambridgeUniv. Press, New York.
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

1999 O’LEARY AND GEISLER—PHYLOGENY OF CETACEA 481
MACLEOD, N., AND K. D. ROSE. 1993. Inferring loco-moter behavior in Paleogene mammals via eigen-shape analysis. Am. J. Sci. 293-A:300–355.
MACPHEE, R. D. E. 1981. Auditory regions of pri-mates and eutherian insectivores: Morphology, on-togeny, and charcater analysis. Contrib. Primatol.18:1–282.
MACPHEE, R. D. E. 1994. Morphology, adaptations,and relationships of Plesiorycteropus, and a diagno-sis of a new order of eutherian mammals. Bull. Am.Mus. Nat. Hist. 220:1–214.
MADDISON , W. P., AND D. R. MADDISON . 1992. Mac-Clade: Analysis of phylogeny and character evolu-tion. Version 3. Sinauer, Sunderland, Massachusetts.
MATTHEW, W. D. 1897. A revision of the Puerco Fauna.Bull. Am. Mus. Nat. Hist. 9:59–110.
MATTHEW, W. D. 1929. Reclassi�cation of the artio-dactyl families. Bull. Geol. Soc. Am. 40:403–408.
MATTHEW, W. D. 1937. Paleocene faunas of the SanJuan Basin, NewMexico. Trans. Am. Philos.Soc. 30:1–510.
MCCRADY, E., JR. 1940. The development and fate ofthe urogenital sinus in the opossum, Didelphy virgini-ana. J. Morphol. 66:131–154.
MCKENNA, M. C. 1975. Toward a phylogenetic classi-�cation of the Mammalia. Pages 21–46 in Phylogenyof the primates (W. P. Luckett and F. S. Szalay, eds.).Plenum, New York.
MCKENNA, M. C. 1992. The alpha crystallin A chain ofthe eye lens and mammalian phylogeny. Ann. Zool.Fenn. 28:349–360.
MCKENNA, M. C., AND S. K. BELL. 1997. Classi�cationof mammals above the species level. Columbia Univ.Press, New York.
MILINKOVITCH , M.C.,G. ORTI, AND A. MEYER. 1993. Re-vised phylogeny of whales suggested by mitochon-drial ribosomalDNA sequences. Nature 361:346–348.
MILINKOVITCH , M. C., AND J. G. M. THEWISSEN. 1997.Even-toed �ngerprints on whale ancestry. Nature388:622–624.
MIYAMOTO, M. M., AND M. GOODMAN. 1986. Biomolec-ular systematics of eutherian mammals: Phyloge-netic patterns and classi�cation. Syst. Zool. 35:230–240.
MONTGELARD, C., F. M. CATZEFLIS, AND E. DOUZERY.1997. Phylogenetic relationships of artiodactyls andcetaceans as deduced from the comparison of cy-tochrome b and 12S RNA mitochondrial sequences.Mol. Biol. Evol. 14:550–559.
MONTGELARD, C., S. DUCROQ, AND E. DOUZERY. 1998.What is a suiforme(Artiodactyla)? Mol. Phylog. Evol.9:528–532.
MULLER, O. 1898. Untersuchungen uber die Ve-randerungen, welche die Respirationsorgane derSaugetiere durch die anpassung an das Leben imWasser erlitten haben. Jena. Z. Naturwiss. 32:95–230.
NIXON, K. C., AND J. M. CARPENTER. 1996. On simulta-neous analysis. Cladistics 12:221–241.
NOVACEK, M. J. 1977. Aspects of the problem of vari-ation, origin and evolution of the eutherian bulla.Mammal Rev. 7:131–149.
NOVACEK, M. J. 1980. Cranioskeletal features in tupai-ids and selected Eutheria as phylogenetic evidence.
Pages 35–93 in Comparative biology and evolution-ary relationships of the tree shrews (W. P. Luckett,ed.). Plenum, New York.
NOVACEK, M. J. 1982. Information for molecular stud-ies from anatomical and fossil evidence on higher eu-therian phylogeny. Pages 3–41 in Macromolecular se-quences in systematic and evolutionary biology (M.Goodman, ed.). Plenum, New York.
NOVACEK, M. J. 1986. The skull of leptictid insectivo-rans and the higher-level classi�cation of eutherianmammals. Bull. Am. Mus. Nat. Hist. 183:1–111.
NOVACEK, M. J. 1989. Higher mammal phylogeny: Themorphological/molecular synthesis. Pages 421–435in The hierarchy of life, Nobel Symposium 70 (R.Fernholm, K. Bremer, and H. Jornvall, eds.). ElsevierScience, Amsterdam.
NOVACEK, M. J. 1991. “All tree histograms” and theevaluation of cladistic evidence: Some ambiguities.Cladistics 7:345–349.
NOVACEK, M. J. 1992a. Fossils, topologies, missingdata, and the higher-level phylogeny of eutherianmammals. Syst. Biol. 41:58–73.
NOVACEK, M. J. 1992b. Mammalian phylogeny: Shak-ing the tree. Nature 356:121–125.
NOVACEK, M. J. 1993. Mammalian phylogeny: Mor-phology and molecules. Trends Ecol. Evol. 8:339–340.
NOVACEK, M. J. 1994. Morphological and molecularinroads to phylogeny. Pages 85–130 in Interpretingthe hierarchy of nature (L. Grande and O. Rieppel,eds.). Academic Press, San Diego.
NOVACEK, M. J., K. GAO, M. A. NORRELL, AND
G. ROUGIER. 1998. Ghost lineages, phylogeny, andranges of selected vertebrate lineages across the K/Tboundary. J. Vertebr. Paleontol. 18(Suppl. to No.3):67A.
NOVACEK, M. J., AND Q. D. WHEELER. 1992. Extinct taxa:Accounting for 99.999...% of the earth’s biota. Pages1–16 in Extinction and phylogeny (M. J. Novacekand Q. D. Wheeler, eds.). Columbia Univ. Press, NewYork.
NOVACEK, M. J., AND A. R. WYSS. 1986. Higher-levelrelationships of the eutherian orders: Morphologicalevidence. Cladistics 2:257–287.
O’LEARY, M. A. 1998a. Phylogenetic and morphomet-ric reassessment of the dental evidence for a mesony-chian and cetacean clade. Pages 133–161 in The emer-gence of whales (J. G. M. Thewissen, ed.). Plenum,New York.
O’LEARY, M. A. 1998b. Morphology of the humerusof Hapalodectes (Mammalia, Mesonychia). Am. Mus.Novit. 3242:1–6.
O’LEARY, M. A., AND K. D. ROSE. 1995a. New mesony-chian dentitions from the Paleocene and Eocene ofthe Bighorn Basin, Wyoming. Ann. Carnegie Mus.64:147–172.
O’LEARY, M. A., AND K. D. ROSE. 1995b. Postcra-nial skeleton of the early Eocene mesonychidPachyaena (Mammalia: Mesonychia). J. Vertebr. Pa-leontol. 15:401–430.
PETERSON, O. A. 1931. New mesonychids from theUinta. Ann. Carnegie Mus. 20:333–339.
PHILIPPE, H., AND E. DOUZERY. 1994. The pitfalls ofmolecular phylogeny based on four species, as illus-
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

482 SYSTEMATIC BIOLOGY VOL. 48
trated by the Cetacea/Artiodactyla relationships. J.Mammal. Evol. 2:133–152.
PICKFORD, M. 1983. On the orgins of Hippopotami-dae together with descriptions of two new species,a new genus and a new subfamily from the Mioceneof Kenya. Geobios 16:193–217.
PROTHERO, D. R. 1993. Ungulate phylogeny: Molec-ular vs. morphological evidence. Pages 173–181 inMammal phylogeny, volume 2. Placentals (F. S. Sza-lay and M. J. Novacek, eds.). Springer-Verlag, NewYork.
PROTHERO, D. R., E. M. MANNING, AND M.FISCHER. 1988. The phylogeny of the ungulates.Pages 201–234 in The phylogeny and classi�cationof the tetrapods, volume 2. Mammals. Syst. Assoc.Spec. Vol. 35B (M. J. Benton, ed.). Clarendon, Oxford,England.
QUERALT, R., R. ADROER, R. OLIVA, R. J. WINKFEIN, J. D.RETIEF, AND G. H. DIXON. 1995. Evolution of pro-tamine P1 genes in mammals. J. Mol. Evol. 40:601–607.
RADINSKY, L. B. 1965. Evolution of the tapiroid skele-ton from Heptodon to Tapirus. Bull. Mus. Comp. Zool.34:1–106.
ROSE, K. D. 1982. Skeleton of Diacodexis, oldest knownartiodactyl. Science 216:621–623.
ROSE, K. D. 1985. Comparative osteology of NorthAmerican dichobunid artiodactyls. J. Paleontol.59:1203–1226.
ROSE, K. D. 1987. Climbing adaptations in the earlyEocene mammal Chriacus and the origin of Artio-dactyla. Science 236:314–316.
ROSE, K. 1990. Postcranial skeletal remains and adap-tations in early Eocene mammals from the WillwoodFormation, Bighorn Basin, Wyoming. Geol. Soc. Am.Spec. Pap. 243:107–133.
ROSE, K. D. 1996. On the origin of Artiodactyla. Proc.Natl. Acad. Sci. USA 93:1705–1709.
ROSE, K. D., AND M. A. O’LEARY. 1995. The manus ofPachyaena gigantea (Mammalia: Mesonychia). J. Ver-tebr. Paleontol. 15:855–859.
RUSSELL , D. E. 1964. Les mammiferes Paleocenesd’Europe. Mem. Mus. Natl. Hist. Nat. Ser. C (Paris)13:1–324.
RUSSELL , D. E., J. G. M. THEWISSEN, AND D. SIGOGNEAU-RUSSELL . 1983. A new dichobunid artiodactyl(Mammalia) from the Eocene of North-West Pak-istan, Part II. Cranial osteology. Proc. K. Ned. Akad.Wet., Ser. B 86:285–299.
SANDERSON , M. J., AND M. J. DONOGHUE. 1989. Pat-terns of variation in levels of homoplasy. Evolution43:1781–1795.
SCHAEFFER, B. 1947. The origin of a mammalian ordi-nal character. Evolution 2:164–175.
SCOTT, W. B. 1888. On some new and little known cre-odonts. J. Acad. Nat. Sci. Phila. 9:155–185.
SCOTT, W. B. 1894. The structure and relationships ofAncodus. J. Acad. Nat. Sci. Phila. 9:461–497.
SHIMAMURA, M., H. YASUA, K. OHSHMIA, H. ABE, H.KATO, T. KISHIRO, M. GOTOS, I. MUNECHIKAI, AND
N. OKADA. 1997. Molecular evidence from retro-posons that whales form a clade within even-toedungulates. Nature 388:666–670.
SHOSHANI, J. 1986. Mammalian phylogeny: Compar-ison of morphological and molecular results. Mol.Biol. Evol. 3:222–242.
SHOSHANI, J., M. GOODMAN, J. CZELUSNIAK, AND G.BRAUNITZER. 1985. A phylogeny of Rodentia andother eutherian orders: Parsimony analysis utilizingamino acid sequences of alpha and beta hemoglobinchains. Pages 191–210 in Evolutionary relationshipsamong rodents: A multidisciplinary approach (W.P. Luckett and J.-L. Hartenberger, eds.). Plenum,New York.
SIMMONS, N. B., AND J. H. GEISLER. 1998. Phylo-genetic relationships of Icaronycteris, Archaeonyc-teris, Hassianycteris, and Palaeochiropteryx to ex-tant bat lineages, with comments on the evo-lution of echolocation and foraging strategiesin Microchiroptera. Bull. Am. Mus. Nat. Hist.235:1–182.
SIMPSON, G. G. 1945. The principles of classi�cationand a classi�cation of mammals. Bull. Am. Mus. Nat.Hist. 85:1–350.
SISSON, S., AND J. D. GROSSMAN . 1953. The anatomy ofthe domestic animals. W. B. Saunders, Philadelphia.
SLIJPER, E. J. 1936. Die Cetaceen Vergleichend-Anatomische und Systematisch. Capita Zool. 7:1–590.
SLOWINSKI, J. B. 1993. “Unordered” versus “ordered”characters. Syst. Biol. 42:155–165.
SMITH, M. R., M. S. SHIVJI, V. G. WADDELL , AND M.J. STANHOPE. 1996. Phylogenetic evidence from theIRBP gene for the paraphyly of toothed whales, withmixed support for Cetacea as a suborder of Artio-dactyla. Mol. Biol. Evol. 13:918–922.
SMUTS, M.M.S. AND A. J. BEZUIDENHOUT . 1987. Anatomyof the dromedary. Clarendon Press, Oxford.
SOKOLOV, V. E. 1982. Mammal skin. Univ. CaliforniaPress, Berkeley.
STANHOPE, M. J., W. J. BAILEY, J. CZELUSNIAK, M.GOODMAN, J.-S. SI, J. NICKERSON, J. G. SGOUROS, G. A.M. SINGER, AND T. K. KLEINSCHMIDT . 1993. A molec-ular view of primate supraordinal relationships fromboth nucleotide and amino acid sequences. Pages251–292 in Primates and their relatives in phyloge-netic perspective (R. D. E. MacPhee, ed.). Plenum,New York.
STANHOPE, M. J., M. R. SMITH, V. G. WADDELL, C. A.PORTER, M. S. SHIVJI, AND M. GOODMAN. 1996. Mam-malian evolution and the interphotoreceptor retinoidbinding protein (IRPB) gene: Convincing evidencefor several superordinal clades. J. Mol. Evol. 43:83–92.
SWOFFORD, D. L. 1993. PAUP: Phylogenetic analysisusing parsimony, version 3.1.1. Illinois Natural His-tory Survey, Champaign.
SWOFFORD, D. L. 1998. PAUP* 4.0b1: PhylogeneticAnalysis Using Parsimony ( * and Other Methods).Sinauer Associates, Sunderland, Massachusetts.
SZALAY, F. S. 1969a. The Hapalodectinae and a phy-logeny of the Mesonychidae (Mammalia, Condy-larthra). Am. Mus. Novit. 2361:1–26.
SZALAY, F. S. 1969b. Origin and evolution of functionof the mesonychid condylarth feeding mechanism.Evolution 23:703–720.
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

1999 O’LEARY AND GEISLER—PHYLOGENY OF CETACEA 483
SZALAY, F. S., AND S. J. GOULD. 1966. Asiatic Mesony-chidae (Mammalia, Condylarthra). Bull. Am. Mus.Nat. Hist. 132:128–173.
THEWISSEN, J. G. M. 1990. Evolution of Paleoceneand Eocene Phenacodontidae (Mammalia, Condy-larthra). Univ. Mich. Pap. Paleontol. 29:1–107.
THEWISSEN, J. G. M. 1994. Phylogenetic aspects ofcetacean origins: A morphological perspective. J.Mammal. Evol. 2:157–184.
THEWISSEN, J. G. M., AND D. P. DOMNING. 1992. The roleof phenacodontids in the origin of the modern ordersof ungulate mammals. J. Vertebr. Paleontol. 12:494–504.
THEWISSEN, J. G. M., AND S. T. HUSSAIN. 1990. Postcra-nial osteology of the most primitive artiodactyl Di-acodexis pakistanensis (Dichobunidae). Anat. Histol.Embryol. 19:37–48.
THEWISSEN, J. G. M., AND S. T. HUSSAIN. 1993. Origin ofunderwater hearing in whales. Nature 361:444–445.
THEWISSEN, J. G. M., AND S. T. HUSSAIN. 1998. Sys-tematic review of the Pakicetidae, early and middleEocene Cetacea (Mammalia) from Pakistan and In-dia. Ann. Carnegie. Mus. 34:220–238.
THEWISSEN, J. G. M., S. T. HUSSAIN, AND M. ARIF. 1994.Fossil evidence for the origin of aquatic locomotionin archaeocete whales. Science 263:210–212.
THEWISSEN, J. G. M., S. I. MADAR, AND S. T.HUSSAIN. 1996. Ambulocetus natans, an Eocenecetacean (Mammalia) from Pakistan. Cour.Forschungsinst. Senckenb. 191:1–86.
THEWISSEN, J. G. M., S. I. MADAR, AND S. T.HUSSAIN. 1998. Whale ankles and evolutionary re-lationships. Nature. 395:452.
THEWISSEN, J. G. M., D. E. RUSSELL, P. D. GINGERICH,AND S. T. HUSSAIN. 1983. A new dichobunid artio-dactyl (Mammalia) from the Eocene of North-WestPakistan, dentition and classi�cation. Proc. K. Ned.Akad. Wet. Ser. B 86:153–180.
TING, S., AND C. LI. 1987. The skull of Hapalodectes(Acreodi, Mammalia), with notes on some ChinesePaleocene mesonychids. Vertebr. Palasiat. 25:161–186.
UHEN, M. D. 1996. Dorudon atrox (Mammalia, Cet-acea): Form, function, and phylogenetic relationshipsof an archaeocete from the late middle Eocene ofEgypt. Ph.D. thesis, Univ. Michigan, Ann Arbor.
UHEN, M. D. 1998. Middle to late Eocenebasilosaurines and dorudontines. Pages 29–61 inThe emergence of whales (J. G. M. Thewissen, ed.).Plenum, New York.
VAN VALEN, L. 1966. The Deltatheridia, a new orderof mammals. Bull. Am. Mus. Nat. Hist. 132:1–126.
VAN VALEN, L. 1968. Monophyly or diphyly in the ori-gin of whales. Evolution 23:37–41.
VAN VALEN, L. 1969. The multiple origins of the pla-cental carnivores. Evolution 23:118–130.
VAN VALEN, L. 1978. Thebeginning of theage of mam-mals. Evol. Theory 4:45–80.
VAUGHAN, T. A. 1986. Mammalogy. Harcourt, Brace,Jovanovich, Fort Worth, Texas.
WEBB, S. B., AND B. E. TAYLOR. 1980. Phylogeny ofhornless ruminants and a description of the craniumof Archaeomeryx. Bull. Am. Mus. Nat. Hist. 167:1–157.
WEST, R. M. 1981. Geology and paleontology of theBridger Formation, Southern Green River Basin,Southwestern Wyoming. Part 5. Harpagolestes macro-cephalus and comments on structure, function anddiversity of middle Eocene to early Oligocene largemesonychids. Contrib. Biol. Geol. Milwauk. Publ.Mus. 43:1–17.
WHEELER, W. C. 1992. Extinction, sampling andmolecular phylogenetics. Pages 205–215 in Extinctionand phylogeny (M. J. Novacek and Q. D. Wheeler,eds.). Columbia Univ. Press, New York.
WIBLE, J. R. 1987. The eutherian stapedial artery:Character analysis and implications for superordinalrelationships. Zool. J. Linn. Soc. 91:107–135.
WIBLE, J. R. 1990. Petrosals of Late Cretaceous marsu-pials from North America, and a cladistic analysis ofthe petrosal in therian mammals. J. Vertebr. Paleon-tol. 10:183–205.
WILEY, E. O., D. SIEGEL-CAUSEY, D. R. BROOKS, AND V.A.FUNK. 1991. The Compleat Cladist. Univ. KansasMus. Nat. Hist. Spec. Publ. 19:1–158.
WILKINSON, M. 1992. Ordered versus unordered char-acters. Cladistics 8:375–385.
WILLIAMSON , T. E., AND S. G. LUCAS. 1992. Menis-cotherium (Mammalia, “Condylarthra”) from the Pa-leocene/Eocene of western North America. N. M.Mus. Nat. Hist. Sci., Bull. 1:1–75.
WORTMAN, J. L. 1901. Studies of Eocene Mammalia inthe Marsh Collection, Peabody Museum. Am. J. Sci.11:1–90.
ZHOU, X., AND P. D. GINGERICH. 1991. New species ofHapalodectes (Mammalia, Mesonychia) from the earlyWasatchian, early Eocene of northwestern Wyoming.Contrib. Mus. Paleontol. Univ. Mich. 28:215–220.
ZHOU, X., W. J. SANDERS, AND P. D. GINGERICH.1992. Functional and behavioral implications ofverterbral structure in Pachyaena ossifraga (Mam-malia, Mesonychia). Contrib. Mus. Paleontol. Univ.Mich. 28:289–319.
ZHOU, X., R. ZHAI, P. D. GINGERICH, AND L. CHEN.1995. Skull of a new mesonychid (Mammalia,Mesonychia) from the late Paleocene of China. J. Ver-tebr. Paleontol. 15:387–400.
Received 29 April 1998; accepted 14 August 1998Associate Editor: G. Naylor
APPENDIX 1.SPECIMENS AND LITERATURE REFERENCES USED
TO COMPILE THE CHARACTER-TAXON MATRIX
FOR PARSIMONY ANALYSIS
Data on the deciduous fourth premolar throughoutmatrix are from Luckett and Hong (1998). AMNH-M= Department of Mammalogy, AMNH-VP, Depart-ment of Vertebrate Paleontology, American Museumof Natural History, New York; ChM PV = CharlestonMuseum, Charleston, South Carolina; GSP-UM = Ge-ological Survey of Pakistan/University of Michigan,Ann Arbor; H-GSP = Howard University/GeologicalSurvey of Pakistan, Washington, D.C.; IVPP = Instituteof Vertebrate Paleontology and Paleoanthropology, Bei-
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

484 SYSTEMATIC BIOLOGY VOL. 48
jing, China; MCZ = Museum of Comparative Zoology,Harvard University, Cambridge, Massachusetts;NMMNH = New Mexico Museum of Natural His-tory, Albuquerque; USGS = Johns Hopkins Univer-sity/United States Geological Survey (now at theSmithsonian Institution, Washington, D.C.); USNM =National Museum of Natural History, Smithsonian In-stitution, Washington, D.C.; YPM = Yale Peabody Mu-seum, New Haven, Connecticut; YPM-PU, PrincetonUniversity collection (now at Yale Peabody Museum).
Archaic Ungulates
Andrewsarchus.—AMNH-VP 20135; Chow (1959).Arctocyon.—AMNH-VP 55900 (cast) , 55901 (cast),
55902; Russell (1964).Eoconodon.—AMNH-VP 764, 774, 3177, 3181, 3187,
3280, 4052, 16329, 16341; Matthew (1897, 1937).Hyopsodus.—AMNH-VP 1, 39, 1717, 10977, 10979,
11330, 11349, 11350, 11363, 11393, 11415, 11899; Gazin(1968); Thewissen and Domning (1992).
Meniscotherium.—AMNH-VP 2560, 4414, 4426, 4434,4447, 48083, 48126, 48129, 48555; Gazin (1965); Cifelli(1982); Williamson and Lucas (1992).
Phenacodus.—AMNH-VP, 2961, 4370, 4378, 4403,15262, 15266, 15268, 15271, 15275, 15279, 15284, 15286,117195; Thewissen (1990); O’Leary and Rose (1995b).
Artiodactyla
Agriochoerus.—AMNH-VP 685, 1349, 1355, 7407,7409, 9808, 9811, 38843, 38932, 95324, 95332, 99275.
Archaeotherium.—AMNH-VP 39127, 39455.Camelus.—AMNH-M 2911, 14109, 35463, 35563,
69405, 80227, 90433; Smuts and Bezuidenhout (1987);Langer (1988 [based on condition in Llama]).
Diacodexis.—AMNH-VP 16141; USGS 2352; Rose(1982, 1985); Russell et al. (1983); Thewissen et al. (1983);Thewissen and Hussain (1990); Thewissen (1994).
“Elomeryx”-(includes specimens attributed to Bothri-odon).—AMNH-VP 579, 582, 583, 1242, 1245, 1259, 1483,10041, 12461, 39015, 101668; Scott (1894).
Hexaprotodon.—AMNH-M 2423, 54265, 81899, 89626,146848, 146849, 148452, 185383, 202423; Vaughan (1986);Langer (1988); Sokolov (1982).
Hippopotamus.—AMNH-M 15898, 24282, 24284,24285, 24289, 53773, 54248, 80813, 81856, 130247, 176118;Vaughan (1986); Langer (1988); Sokolov (1982).
Ovis.—AMNH-M 6231, 6239, 10074, 14515, 15584,35520, 53598, 88702, 100012, 100072, 146547; Sisson andGrossman (1953); Langer (1988); Sokolov (1982).
Poebrotherium.—AMNH-VP 8955, 39085, 42240,42248, 42249, 42261, 42272, 42276, 42281, 42284, 42292,47003, 47008, 47052, 47103, 47182, 47284, 47317, 47324,47333, 47077, 47907, 63701, 63704, 63712, 63713, 97103.
Sus.—AMNH-M 20871, 69422, 69442, 100260,235190, 235192, 236144, 238325, 238331; Sisson andGrossman (1953); Langer (1988); Sokolov (1982).
Tragulus.—AMNH-M 10101, 14137, 14139, 34252,32645, 53602, 53609, 60759, 90101, 90193, 102091, 102176,106552, 240913; Langer (1988); Sokolov (1982).
Cetacea
Ambulocetus.—H-GSP 18507; Thewissen et al. (1996);Thewissen pers. comm. (pelvis data).
Balaenoptera.—AMNH-M 28274, 84870, 148407,219212, 219220; Daudt (1898); Muller (1898); Sokolov(1982); Gaskin (1978).
Basilosaurus.—AMNH-VP 14381, 129577 (cast); GSP-UM 97507; Kellogg (1936).
Cross Whale.—ChM PV 5401; Geisler et al. (1996).Dorudon.—Uhen (1996, 1998).Pakicetus.—GSP-UM 084 (cast); H-GSP 96231, 96386,
96431, 18410, 18570, 96505; Gingerich and Russell(1981, 1990); Thewissen and Hussain (1993); Thewissen(1994); Thewissen and Hussain (1998).
Protocetus.—Fraas (1904).Remingtonocetus.—GSP-UM 3009, 3054, 3057; Kumar
and Sahni (1986); Gingerich et al. (1995).Tursiops.—AMNH-M 120920, 184930, 212554; Daudt
(1898 [based on condition in other odontocetes]);Muller (1898 [based on condition in other odontocetes]);Langer (1988).
Mesonychidae
Ankalagon.—AMNH-VP 776, 777, 2454; NMMNH16309.
Dissacus.—AMNH-VP 3356, 3357, 3359, 3360, 3361,15996, 39276, 55925 (cast); IVPP 4266 (cast); YPM-PU16159; O’Leary and Rose (1995a, 1995b).
Harpagolestes.—AMNH-VP 1878, 1892, 1945, 2308,26267, 26300, 26301; USNM 14708, 14915; Wortman(1901); Peterson (1931); West (1981); Zhou et al. (1995).
Mongolestes.—AMNH-VP 26064, 26065.Mongolonyx.—AMNH-VP 26661, 26662.Mesonyx.—AMNH-VP 1716, 5021, 11552, 12641,
12643 (cast), 19204, 122122; Scott (1888); O’Leary andRose (1995b).
Pachyaena.—AMNH-VP 72, 75, 1522, 2823, 2959,4262, 15224, 15228, 15728, 15730, 16154; USGS 7185;YPM 50000; YPM-PU 14708; O’Leary and Rose (1995a,1995b); Rose and O’Leary (1995).
Sinonyx.—VPP V10760 (skull and lower dentition);Zhou et al. (1995).
Synoplotherium.—AMNH-VP 19203,122122; Wortman (1901).
Hapalodectidae
Hapalodectes.—AMNH-VP 78, 12781, 14748 (cast),128561, 17558, 20172, 80802 (cast); IVPP 5254 (cast);USGS 275 (cast), 9628, 10293 (cast); Guthrie (1967); Sza-lay (1969a); Ting and Li (1987); Zhou and Gingerich(1991); O’Leary (1998b).
Perissodactyla
Equus.—AMNH-M 1244C, KMM 99; AMNH-VP 113(VP), LA 118; Sisson and Grossman (1953); Vaughan(1986); Langer (1988).
Heptodon.—AMNH-VP 294, 4855, 4858, 14865, 14884,16861, 141881; Radinsky (1965).
Hyracotherium.—AMNH-VP 55267, 55268, 55269,96274, 96277, 96284, 96298, 129209; Kitts (1956); Mac-Fadden (1992); Rose (1990); Thewissen and Domning(1992).
Outgroups
Asioryctes.—Kielan-Jaworowska (1977, 1981).
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

1999 O’LEARY AND GEISLER—PHYLOGENY OF CETACEA 485
Didelphis.—AMNH-M 7310, 14335, 70089, 180358,217701, 219212, 219220, 240517, 242660; Bremer (1904);McCrady (1940); Vaughan (1986); Langer (1988).
Leptictis.—AMNH-VP 1413a, 5346, 38920, 39444,90256, 96766; MCZ 19678; USNM 336367; Novacek(1980, 1986).
APPENDIX 2. CHARACTERS AND CHARACTER
STATES FOR PARSIMONY ANALYSIS
Multistate characters treated as ordered are speci-�ed below. The character-taxon matrix is presented inAppendix 3.
Basicranial Characters
1. Subarcuate fossa.—Present (0); absent (1) (Novacek,1986).
2. Shape of tegmen tympani (ordered).—Unin�ated,forms thin lamina lateral to facial nerve canal (0);in�ated, forms barrel-shaped ossi�cation lateral tothe facial nerve canal (1); hyperin�ated, transversewidth greater than or equal to width of promonto-rium (2) (modi�ed from Cifelli, 1982; Geisler andLuo, 1998).
3. Anterior process of petrosal.—Absent (0); present, an-terior edge of tegmen tympani extending far ante-rior to edge of pars cochlearis (1) (Luo and Marsh,1996; Geisler and Luo, 1998).
4. Fossa for tensor tympani muscle.—Shallow, anteropos-teriorly elongate fossa (0); circular pit, no groove (1);circular pit with deep tubular anterior groove (2);long narrow groove between tegmen tympani andpromontorium (3) (Luo and Marsh, 1996; Geislerand Luo, 1998).
5. Transpromontorial sulcus for internal carotid artery.—Present, forms anteroposterior groove on promon-torium, medial to fenestrae rotunda and ovalis (0);absent (1) (Cifelli, 1982; Thewissen and Domning,1992).
6. Sulcuson promontoriumforproximal stapedial artery.—Present, forms a groove that branches from thetranspromontorial sulcus anteromedial to the fen-estra rotunda and extends to the medial edge of thefenestra vestibuli (0); absent (1) (Cifelli, 1982; Wible,1987; Thewissen and Domning, 1992).
7. Mastoid process of petrosal.—Exposed externally onposterior face of braincase as a triangle betweenlambdoidal crest of the squamosal dorsolaterally,the exoccipital ventrally, and the supraoccipital me-dially (0); not exposed posteriorly, lambdoidal crestof squamosal in continuous contact with exoccipitaland supraoccipital (1).
8. Stylomastoid foramen.—Complete, ectotympaniccontacts tympanohyal laterally and petrosal me-dially, in some cases ectotympanic separated frompetrosal by a narrow (< 1 mm) �ssure (0); incom-plete, ectotympanic does not contact the petrosaleither anterior or posterior to the fenestra rotunda,medial side of foramen open (1) (modi�ed fromGeisler and Luo, 1998).
9. Facial nerve sulcus distal to stylomastoid foramen.—Absent (0); present, anterior wall of sulcus formedby squamosal (1); present, anterior wall formed by
mastoid process of petrosal (2); anterior wall formedby meatal tube of ectotympanic (3) (modi�ed fromGeisler and Luo, 1996, 1998). Some taxa coded aspolymorphic for this character are not truly poly-morphic, and the character state is simply ambigu-ous, but certain states can be eliminated from thelist of possibilities.
10. Articulation of pars cochlearis with basisphenoid/basioc-cipital.—Present (0); absent (1) (Thewissen andDomning, 1992).
11. Ectotympanic .—Simple ring, no bulla formation (0);medial edge expanded into bulla (1) (derived fromNovacek, 1977; MacPhee, 1981; Geisler and Luo,1998).
12. Pachyosteosclerotic involucrum of bulla.—Absent (0);present (1) (Thewissen, 1994).
13. Lateral furrow of tympanic bulla.—Absent (0); present,forms a groove on the lateral surface of the ecto-tympanic bulla anterior to the base of the sigmoidprocess or meatal tube (1) (Geisler and Luo, 1998).
14. Median furrow of tympanic bulla for tympanohyal(ordered).—Absent (0); median notch on posteriorrim of bulla (1); prominent anteroposteriorly ori-ented furrow splits the ventral surface of the bullainto medial and lateral halves (2) (Geisler and Luo,1998).
15. Articulation of ectotympanic bulla to squamosal(ordered).—Broad articulation with medial base ofpostglenoid process (0); circular facet on short en-toglenoid process (1); contact with a crest of the en-toglenoid process (2); contact absent (3) (Geisler andLuo, 1998).
16. Contact between exoccipital and ectotympanic bulla.—Absent (0); present (1) (Geisler and Luo, 1998).
17. Sigmoid process (= anterior crus of tympanic ring).—Absent (0); present, forms transverse plate thatprojects laterally from the anterior crus of the ecto-tympanic ring and forms the anterior wall of the ex-ternal auditory meatus (1) (modi�ed from Thewis-sen, 1994; Geisler and Luo, 1998).
18. Sigmoid process shape.—Thin and transverse plate(0); broad and �aring, base of the sigmoid processforms dorsoventral ridge on lateral surface of ecto-tympanic bulla (1) (Geisler and Luo, 1998).
19. Ectotympanic part of the meatal tube (ordered).—Absent (0); present but short, length of tube < 30%maximum width of bulla (1); present and long,length > 60% maximum width of bulla (2) (Geislerand Luo, 1998).
20. Squamosal portion of external auditory meatus.—Absent or shallow, depth < 25% of transverse length(0); deep groove, depth > 35% of transverse length(1).
21. Basioccipital crests (falcate processes).—Absent (0);present, forming ventrolaterally �aring basioccipi-tal processes (1) (derived from Barnes, 1984; modi-�ed from Thewissen, 1994).
22. Internal carotid foramen.—Absent or con�uent withthe piriform fenestra (0); present at basisphe-noid/basioccipital suture with lateral wall of fora-men formed by both these bones and thus sepa-rated from the piriform fenestra (1) (Geisler andLuo, 1998).
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

486 SYSTEMATIC BIOLOGY VOL. 48
23. Postglenoid foramen (ordered).—Present, enclosedby squamosal, immediately posterior to post-glenoid process (0); present, medial to pet-rosal/squamosal suture (if bulla present, a sec-ondary ventral opening between bulla andsquamosal may form) (1); absent (2) (Novacek,1986; Geisler and Luo, 1998).
24. Foramen for ramus superior of stapedial artery.—Present (0); absent (1) (modi�ed from Novacek,1986; Thewissen and Domning, 1992).
25. Position of foramen for ramus superior of stapedialartery.—In petrosal/squamosal suture on dorsolat-eral edge of epitympanic recesss (0); anterolateral toepitympanic recess (1) (modi�ed from Geisler andLuo, 1998). Cannotbescored for taxa that lack a fora-men for the ramus superior of the stapedial artery.
26. Foramen ovale.—Anterior to glenoid fossa, posteriorwall formed by alisphenoid (0); medial to glenoidfossa, posterior wall formed by alisphenoid (1); me-dial to glenoid fossa, posterior wall formed by pet-rosal (2); posterior to glenoid fossa (3).
27. Alisphenoid canal (alar canal).—Absent (0); present(1) (Novacek, 1986; Thewissen and Domning, 1992).
28. Anterior opening of alisphenoid canal.—Within sphe-norbital �ssure, cannot be distinguished from sphe-norbital �ssure in lateral view (0); posterior to sphe-norbital �ssure (1) (Novacek, 1982; Thewissen andDomning, 1992).
29. Posterior opening of alisphenoid canal.—Well-separated from foramen ovale (0); within a deepgroove or recess with the foramen ovale (1) (de-rived from Zhou et al., 1995).
30. Foramen rotundum.—Absent, maxillary division oftrigeminal nerve exits skull through sphenorbital�ssure (0); present (1) (Novacek, 1986; Thewissenand Domning, 1992).
31. Mastoid foramen.—Present, skull in posterior view(0); absent (1) (de�nition of structure followsMacPhee, 1994).
32. Post-temporal canal (for arteria diploetica magna,also called pericranial foramen).—Present, occursat petrosal/squamosal suture with skull in pos-terior view, the canal continuing within the pet-rosal/squamosal suture (0); absent (1) (Wible, 1990;MacPhee, 1994).
33. Preglenoid process.—Absent (0); present, forms trans-verse, ventrally projecting ridge at anterior edge ofglenoid fossa (1) (modi�ed from Thewissen, 1994).
Cranial Characters
34. Supraorbital process.—Absent, region over orbit doesnot project laterally from sagittal plane (0); present,laterally elongate and tabular (1) (derived fromBarnes, 1984).
35. Postorbital bar (ordered).—Absent (0); present andalmost complete (1); present and complete (2).
36. Contact of frontal and maxilla in orbit.—Absent (0);present (1).
37. Lacrimal tubercle.—Absent (0); present, situated onanterior edge of orbit adjacent to the lacrimal fora-men (1) (Novacek, 1986).
38. Palatine �ssure.—Small (0); large (1).39. Posterior margin of external nares (ordered).—
Anterior to or over the canines, or at anterior edge ofpremaxilla if canine not present (0); between P1 andP2 (1); posterior to P2 (2) (Geisler and Luo, 1998).
40. Mandibular foramen.—Small, maximum height ofopening # 25% the height of the mandible at M3 (0);large, continuous with a large posterior fossa, max-imum height $ 50% the height of the mandible atM3 (1) (modi�ed from Thewissen, 1994; Geisler andLuo, 1998).
41. Height of mandibular condyle relative to dentition(ordered).—Below level of the dentition (0); evenwith superior aspect of dentition (1); substantiallysuperior to dentition (2).
42. Angle of dentary.—Distal end at same level as ven-tral edge of dentary below molars (0); forms dis-tinct �ange that projects posteroventrally well be-low ventral edge of dentary (1) (modi�ed from Gen-try and Hooker, 1988).
Dental Characters
43. Upper incisors.—Present (0); absent (1).44. Premaxillae.—Short with incisors arranged in trans-
verse arc (0); elongate, incisors aligned longitu-dinally with intervening diastemata (1) (modi�edfrom Prothero et al., 1988; Thewissen, 1994).
45. Embrasure pits on palate.—Absent (0); present (1)(modi�ed from Thewissen, 1994).
46. P1 (ordered).—Absent (0); present, one-rooted (1);present, two-rooted (2) (Zhou et al., 1995).
47. P3 roots.—Three (0); two (1) (Zhou et al., 1995).48. P4 protocone.—Present (0); absent (1) (Thewissen,
1994).49. P4 paracone.—Equal or subequal to height of para-
cone of M1 (0); greater than twice the height of M1paracone (1) (Thewissen, 1994).
50. P4 metacone.—Absent (0); present (1) (Thewissen,1994).
51. Stylar shelf.—Present (0); absent (1).52. M2 metacone (ordered).—Distinct cusp, sub-equal to
paracone (0); distinct cusp approximately half thesize of the paracone (1); highly reduced, indistinctfrom paracone (2) (Zhou et al., 1995).
53. M1 parastyle (ordered).—Absent (0); weak (1); mod-erate to strong (2) (O’Leary, 1998a) .
54. Lingual cingulum on M2 .—Present (0); absent (1)(O’Leary, 1998a).
55. Ectocingula.—Present (0); absent (1) (O’Leary,1998a).
56. M1-M2 hypocone.—Absent (0); present (1) (Thewis-sen and Domning, 1992; O’Leary, 1998a).
57. Trigon basin (ordered).—Broad (0); somewhat nar-row (1); very narrow (2) (modi�ed from Thewissen,1994; O’Leary, 1998).
58. Paraconule on M2.—Absent (0); present (1) (O’Leary,1998a).
59. Metaconule on M2.—Absent (0); present (1) (O’Leary,1998a).
60. M3 size (ordered).—Present, larger than M2
(0); present, approximately equal to M2
(1); present, small, maximum mesiodis-tal length than < 60% the length of M2
(2); absent (3) (modi�ed from Zhou et al.,1995).
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

1999 O’LEARY AND GEISLER—PHYLOGENY OF CETACEA 487
61. Lower canines.—Approximately twice as large as in-cisors or larger (0); subequal in size to incisors, wellseparated from postcanine teeth (1).
62. Lower incisors.—Small and vertical (0); tusk-like (1).63. Lower canine cross-section shape.—Oval (0); triangu-
lar (1) (Gentry and Hooker, 1988).64. P1.—Present (0); absent (1) (Zhou et al., 1995).65. P3 metaconid.—Absent (0); present (1) (Thewissen
and Domning, 1992).66. P4 metaconid.—Absent (0); present (1) (Thewissen
and Domning, 1992).67. Lingual cingulid on lower molars.—Poorly de�ned or
absent (0); continuous from mesial to distal extreme(1) (O’Leary, 1998a).
68. M1 paraconid.—Present (0); absent (1) (derived fromO’Leary, 1998a) .
69. M1 paraconid/paracristid position.—Cusp lingual orparacristid winding lingually (or both) (0); cuspanterior or paracristid (or both) straight mesiodis-tally, sometimes poorly developed (1) (derived fromO’Leary, 1998a) .
70. M2 paraconid.—Present (0); absent (1) (derived fromO’Leary, 1998a) .
71. M2 paraconid/paracristid position.—Cusp lingual orparacristid (or both) winding lingually (0); cusp an-terior or paracristid (or both) straight mesiodistally,sometimes poorly developed (1) (O’Leary, 1998a) .
72. M3 paraconid.—Present (0); absent (1) (O’Leary,1998a).
73. M3 paraconid/paracristid position.—Cusp lingual orparacristid winding lingually (or both) (0); cusp an-terior or paracristid (or both) straight mesiodistally,sometimes poorly developed (1) (O’Leary, 1998a) .
74. M1 metaconid.—Present, forms distinct cusp (0); ab-sent or occasionally present as swelling on lingualside of protoconid (1) (Thewissen, 1994; Zhou et al.,1995).
75. M2 metaconid.—Present, forms distinct cusp (0); ab-sent or occasionally present as swelling on lingualside of protoconid (1) (modi�ed from Zhou et al.,1995).
76. M3 metaconid.—Present, forms distinct cusp (0); ab-sent or occasionally present as swelling on lingualside of protoconid (1) (modi�ed from Zhou et al.,1995).
77. M1-M2 hypoconulid.—Absent (0); present (1) (The-wisen, 1994).
78. Molar protoconid.—Subequal to height of talonid(0); closer to twice height of talonid or greater (1)(O’Leary, 1998a).
79. Protoloph.—Absent (0); present (1).80. Metaloph.—Absent (0); present (1).81. Metalophid formation on anterior aspect of lower teeth
(between protoconid and metaconid).—Absent (0);present (1).
82. Hypolophid formation on posterior aspect of lower teeth(between entoconid and hypoconid).—Absent (0);present (1) (Thewissen and Domning, 1992).
83. Reentrant grooves.—Proximal (0); absent (1); distal(2) (Thewissen, 1994; O’Leary, 1998a).
84. Molar talonid.—Basined, or slightly basined,hypoconid and entoconid present (0); reduced (1)(modi�ed from Thewissen, 1994).
85. M3 hypoconulid.—Long, protrudes as separate dis-tal lobe (0); short, does not protrude substantiallybeyond rest of talonid (1); absent (2) (Thewissen,1994).
86. dp4 .—Resembles M1 (0); six-cusped, additionalneomorphic cusp on paracristid (1) (Gentry andHooker, 1988; Luckett and Hong, 1998).
87. Elongate shearing facets on lower molars extending be-low the gum line.—Absent (0); present (1).
Postcranial Characters
88. Cervical vertebrae.—Long, length of centrum greaterthan or equal to the centra of the anterior thoracics(0); short, length shorter than centra of anterior tho-racics (1); very long (2) (derived from Gingerich etal., 1995).
89. Articulation between sacral vertebrae and ilium ofpelvis (ordered).—Broad area of articulation be-tween pelvis and S1 and sometimes S2 (0); narrowarticulation of pelvis with end of transverse processof S1 (1); articulation absent (2) (Geisler and Luo,1998).
90. Number of sacral vertebrae (ordered).—One (0); twoor three (1); four (2); �ve or six (3); cannot be scoredfor taxa that lack an articulation between the verte-bral column and the ilium (Thewissen and Domn-ing, 1992; Gingerich et al., 1995).
91. Scapular spine.—Bears large acromion pro-cess that overhangs glenoid fossa (0); scapu-lar spine with acromion process small or ab-sent, does not encroach upon glenoid fossa (1);acromion process large, directed anteriorly anddoes not encroach upon the glenoid fossa (2)(O’Leary and Rose, 1995b; Geisler and Luo,1998).
92. Entepicondyle of humerus.—Wide, $ 50% width of ul-nar and radial articular facets (0); narrow, # 25%width of ulnar and radial articular facets (1) (de-rived from O’Leary and Rose, 1995b; Geisler andLuo, 1998).
93. Entepicondylar foramen.—Present (0); absent (1)(Thewissen and Domning, 1992).
94. Length of olecranon process.—Short, < 10% total ul-nar length (0); long, > 20% total ulnar length (1)(derived from O’Leary and Rose, 1995b).
95. Olecranon fossa.—Shallow (0); deep, sometimes per-forate (1) (modi�ed from O’Leary and Rose, 1995b).
96. Proximal end of radius.—Single fossa for trochlea andcapitulum of humerus (0); two fossae, one for the ca-pitulum and one for the medial edge of trochlea (1);three fossae, same as (1) but with additional fossafor the lateral lip of the humeral articular surface (2)(O’Leary and Rose, 1995b; Geisler and Luo, 1998).
97. Distal articular surface of radius.—Single concavefossa (0); split into scaphoid and lunate fossae (1);convex to �at, distinct facets (2) (derived fromO’Leary and Rose, 1995b).
98. Centrale.—Present (0); (1) absent (Thewissen, 1994).99. Manus.—Mesaxonic, axis of symmetry passes along
center of digit 3 (0); paraxonic, axis of symmetrypasses between digits three and four (1).
100. Height of greater trochanter of femur (ordered).—Inferior to head of femur (0); approximately equal to
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

488 SYSTEMATIC BIOLOGY VOL. 48
head of femur (1); above head of femur (2) (derivedfrom O’Leary and Rose, 1995b).
101. Third trochanter of femur (ordered).—Present (0);highly reduced (1); absent (2) (derived fromO’Learyand Rose, 1995b).
102. Patellar articular surface on femur.—Wide (0); narrow(1).
103. Astragalar canal.—Present (0); absent (1) (derivedfrom Shoshani, 1986).
104. Navicular facet of astragalus (ordered) (see Fig. 1).—Convex (0); saddle-shaped (1); trochleated (2) (mod-i�ed from Thewissen and Domning, 1992; O’Learyand Rose, 1995b).
105. Sustentacular width.—Narrow, # 50% width of theastragalus (0); wide, $ 70% width of the astragalus(1) (Geisler and Luo, 1998).
106. Contact of distal astragalus with cuboid (ordered)(see Fig. 1).—Absent (0); present but small, contact# 30% of the width of the distal articular surface (1);present and large, contact $ 40% of the articular sur-face (2) (O’Leary and Rose, 1995b; Geisler and Luo,1998).
107. Lateral process of astragalus.—Present (0); absent (1)(Schaeffer, 1956).
108. Pes (ordered).—Mesaxonic, axis of symmetry of footpasses along center of digit three (0); paraxonic, axislies between digits three and four (1); mesaxonic,axis passes along center of digit four (2) (derivedfrom Gingerich et al., 1990; Thewissen, 1994; ourinterpretation of this character differs from that ofthese previous authors).
109. Proximal astragalus (ordered).—Nearly �at toslightly concave (0); grooved, depth of trochlea< 25% width of trochlea (1); deeply grooved, depthof trochlea > 30% width of trochlea (2) (derivedfrom Schaeffer, 1947; O’Leary and Rose, 1995b).
110. Cuboid and navicular.—Unfused (0); fused (1) (Webband Taylor, 1980).
111. Proximal fusion of third and fourth metatarsals.—Absent (0); present (1).
112. First metatarsal (ordered).—Unreduced, length >50% length of third metatarsal (0); reduced, length< 50% length of third metatarsal (1); highly reducedin form of nodule or small splint or absent (2).
113. Second metatarsal (ordered).—Unreduced, length$ 50% length of third metatarsal (0); reduced, length# 50% length of third metatarsal (1); highly re-duced in form of nodule, small splint or completelyabsent (2).
114. Fifth metatarsal (ordered).—Unreduced, length$ 50% length of third metatarsal (0); reduced,length # 50% length of third metatarsal (1); highlyreduced in form of nodule or small splint orabsent (2).
115. Ventral border of distal phalanges.—Curved inferiorly(0); straight (1) (derived from MacLeod and Rose,1993).
116. Distal phalanges in dorsal view.—Phalanx com-pressed transversely (0); broad transversely, eachphalanx bilaterally symmetricalwith central antero-posterior axis (1); broad transversely, each phalanxasymmetrical (2) (derived from MacLeod and Rose,1993).
Soft Tissue Characters
117. Stomach epithelium.—Nonglandular (strati�ed squa-mous epithelium) (0); composite (nonglandularand glandular mucosa) (1); discoglandular (smallpatches of glandular epithelium surrounded bynonglandular mucosa) (2); glandular (mucosa withglands) (3) (derived from Langer, 1988).
118. Lumen.—Unilocular (stomach with simple cham-ber only) (0); plurilocular (lumen of stomach sub-divided by folds into two or more chambers) (1)(derived from Langer, 1988).
119. Diverticulation of stomach lumen (blind tubes or sacsbranching from the main gastric lumen).—Absent(0); present (1) (derived from Langer, 1988).
120. Cavernous tissue of penis.—Abundant (0); sparse (1)(derived from Slijper, 1936; Thewissen, 1994).
121. Hair.—Abundant to common on body (0); almostcompletely absent (1) (Gatesy, 1997).
122. Sebaceous glands.—Present (0); absent (1) (Gatesy,1997).
123. Primary bronchi of lung.—Two (0); three (1) (Thewis-sen, 1994).
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022

1999 O’LEARY AND GEISLER—PHYLOGENY OF CETACEA 489
APPENDIX 3. Data matrix of 123 characters for 40 taxa. ? = missing data; n = inapplicable character; characterstates described in Appendix 2. Polymorphic characters as follows: states 0 and 1 = 5; 0 and 2 = 6; 0 and 3 = 7;1 and 2 = 8; 2 and 3 = 9; earliest appearance of the genus (McKenna and Bell, 1997) (E. = early; M. = middle; L.= late; Cret. = Cretaceous; Pal. = Paleocene; Eoc. = Eocene; Mioc. = Miocene; Pleist. = Pleistocene), and percentmissing data listed at end of matrix. Characters in bold are treated as ordered in certain analyses, extant taxa inbold. The matrix is available at http://www.herbaria. harvard.edu/treebase/ under the accession number S387.
(continued on next page)
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022
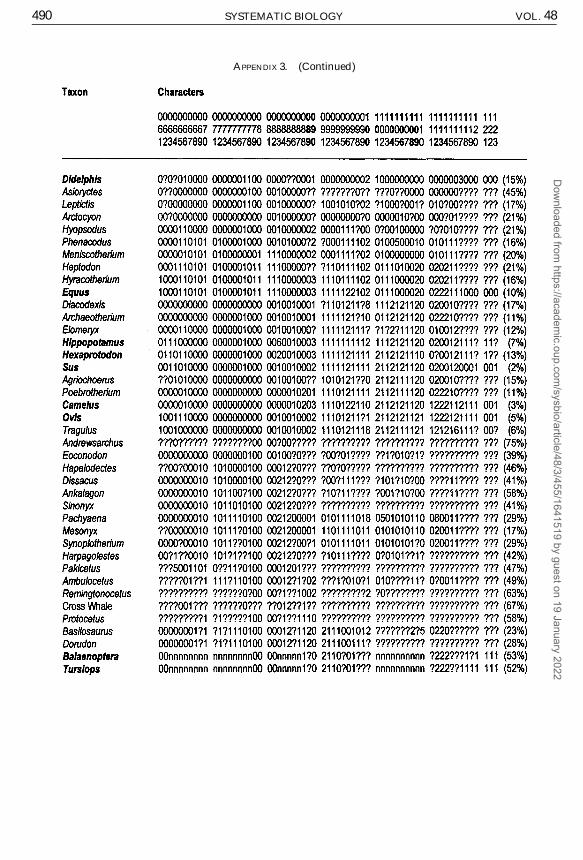
490 SYSTEMATIC BIOLOGY VOL. 48
APPENDIX 3. (Continued)
Dow
nloaded from https://academ
ic.oup.com/sysbio/article/48/3/455/1641519 by guest on 19 January 2022




















