
Phylogenetic and morphological assessment of five new species of Thozetella from an Australian...
14
1074 Mycologia, 96(5), 2004, pp. 1074–1087. q 2004 by The Mycological Society of America, Lawrence, KS 66044-8897 Phylogenetic and morphological assessment of five new species of Thozetella from an Australian rainforest Barbara Paulus 1 Paul Gadek School of Tropical Biology, James Cook University, Cairns, QLD 4870, Australia Kevin Hyde Centre for Research in Fungal Diversity, Department of Ecology and Biodiversity, University of Hong Kong, Pokfulam Road, Hong Kong, SAR, People’s Republic of China Abstract: During an investigation of saprobic micro- fungi in leaf litter from an Australian rainforest, five new species of Thozetella, namely T. acerosa, T. boon- jiensis, T. falcata, T. gigantea and T. queenslandica, were identified and these are described and illustrat- ed here. The morphology of specimens derived from cultures grown under different conditions and from natural substrata was compared. DNA sequence data of ITS regions within nuclear rDNA confirmed the morphological species concept and indicated that Thozetella species are anamorphs of the ascomycete genus Chaetosphaeria. Key words: anamorphic fungi, Chaetosphaeria, Chaetosphaeriacae, key, microfungi, morphology, taxonomy, tropical fungi INTRODUCTION Thozetella O. Kuntze (1891) is a nomen novum for Thozetia Berk. et F. Mueller, which was antedated by Thozetia F. Mueller ex Bentham (Asclepiadaceae). Pi- rozynski and Hodges (1973) revised the genus and accepted four species. Since then, a further five spe- cies were added by Nag Raj (1976), Sutton and Cole (1983), Castan ˜eda Ruiz (1984), Castan ˜eda Ruiz and Arnold (1985) and Mercado Sierra et al (1997). The genus Thozetella is characterized by phialidic conidi- ogenesis, aseptate, hyaline, curved conidia with an unbranched setula on each pole, microawns and spo- rodochial, synnematal or effuse conidiomata (Piro- zynski and Hodges 1973). Microawns are distinctive, sterile elements, which are produced from conidio- phores and found in the spore mass. Their function Accepted for publication January 14, 2004. 1 Corresponding author. E-mail: [email protected] is unknown but they might aid spore dispersal. The generic definition outlined above make the inclusion of T. ciliata (Mercado-Sierra et al 1997) doubtful, be- cause this species lacks microawns and produces se- tae. Species are primarily saprobes of decaying plant material, including terrestrial litter, bark and palm fronds (Pirozynski and Hodges 1973, Yanna et al 2002), submerged wood (Sivichai et al 2002), and T. tocklaiensis was observed as a nonpathogenic root en- dophyte (Waipara et al 1996). While revising the genus, Pirozynski and Hodges (1973) questioned the species concept in Thozetella because knowledge of variation with growth condi- tions and age was limited. Interpretation of species concepts in the genus is complicated by the fact that some species are known only from cultures and oth- ers only from fruiting bodies grown on natural sub- strata (Sutton and Cole 1983). Consequently, Sutton and Cole (1983) showed that characters of T. effusa can vary greatly depending on the substratum on which the fungus has grown. During an investigation of the diversity of saprobic microfungi on leaf litter in an Australian rainforest (Paulus et al 2003), we isolated a number of speci- mens belonging in Thozetella. They could be assigned to five morphological groups based on microawn length and/or morphology. Two of these groups were known only from cultures, while the remaining three were available both as fruiting bodies from nat- ural substrata and the cultures derived from them. Three of the morphological groups were differenti- ated mainly by the length of their microawns. To test whether they were indeed separate species rather than different stages of development of the same spe- cies or artifacts of cultural conditions, sequence anal- ysis from internal transcribed spacer regions (ITS) of the nuclear ribosomal repeat (nrDNA) was undertak- en. Sequence data also were used to clarify the place- ment of the genus among ascomycete lineages. METHODS Morphology of specimens from natural substrata and from cul- ture.—Specimens of Thozetella were isolated after they spor- ulated on decaying leaves incubated in moist chambers (Booth 1971). In addition, spore suspensions were spread on 2% tap water agar containing 50 mg L 21 streptomycin and after 3–4 d small colonies were subcultured (Booth
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Phylogenetic and morphological assessment of five new species of Thozetella from an Australian...

1074
Mycologia, 96(5), 2004, pp. 1074–1087.q 2004 by The Mycological Society of America, Lawrence, KS 66044-8897
Phylogenetic and morphological assessment of five new species of Thozetellafrom an Australian rainforest
Barbara Paulus1
Paul GadekSchool of Tropical Biology, James Cook University,Cairns, QLD 4870, Australia
Kevin HydeCentre for Research in Fungal Diversity, Department ofEcology and Biodiversity, University of Hong Kong,Pokfulam Road, Hong Kong, SAR, People’s Republicof China
Abstract: During an investigation of saprobic micro-fungi in leaf litter from an Australian rainforest, fivenew species of Thozetella, namely T. acerosa, T. boon-jiensis, T. falcata, T. gigantea and T. queenslandica,were identified and these are described and illustrat-ed here. The morphology of specimens derived fromcultures grown under different conditions and fromnatural substrata was compared. DNA sequence dataof ITS regions within nuclear rDNA confirmed themorphological species concept and indicated thatThozetella species are anamorphs of the ascomycetegenus Chaetosphaeria.
Key words: anamorphic fungi, Chaetosphaeria,Chaetosphaeriacae, key, microfungi, morphology,taxonomy, tropical fungi
INTRODUCTION
Thozetella O. Kuntze (1891) is a nomen novum forThozetia Berk. et F. Mueller, which was antedated byThozetia F. Mueller ex Bentham (Asclepiadaceae). Pi-rozynski and Hodges (1973) revised the genus andaccepted four species. Since then, a further five spe-cies were added by Nag Raj (1976), Sutton and Cole(1983), Castaneda Ruiz (1984), Castaneda Ruiz andArnold (1985) and Mercado Sierra et al (1997). Thegenus Thozetella is characterized by phialidic conidi-ogenesis, aseptate, hyaline, curved conidia with anunbranched setula on each pole, microawns and spo-rodochial, synnematal or effuse conidiomata (Piro-zynski and Hodges 1973). Microawns are distinctive,sterile elements, which are produced from conidio-phores and found in the spore mass. Their function
Accepted for publication January 14, 2004.1 Corresponding author. E-mail: [email protected]
is unknown but they might aid spore dispersal. Thegeneric definition outlined above make the inclusionof T. ciliata (Mercado-Sierra et al 1997) doubtful, be-cause this species lacks microawns and produces se-tae. Species are primarily saprobes of decaying plantmaterial, including terrestrial litter, bark and palmfronds (Pirozynski and Hodges 1973, Yanna et al2002), submerged wood (Sivichai et al 2002), and T.tocklaiensis was observed as a nonpathogenic root en-dophyte (Waipara et al 1996).
While revising the genus, Pirozynski and Hodges(1973) questioned the species concept in Thozetellabecause knowledge of variation with growth condi-tions and age was limited. Interpretation of speciesconcepts in the genus is complicated by the fact thatsome species are known only from cultures and oth-ers only from fruiting bodies grown on natural sub-strata (Sutton and Cole 1983). Consequently, Suttonand Cole (1983) showed that characters of T. effusacan vary greatly depending on the substratum onwhich the fungus has grown.
During an investigation of the diversity of saprobicmicrofungi on leaf litter in an Australian rainforest(Paulus et al 2003), we isolated a number of speci-mens belonging in Thozetella. They could be assignedto five morphological groups based on microawnlength and/or morphology. Two of these groupswere known only from cultures, while the remainingthree were available both as fruiting bodies from nat-ural substrata and the cultures derived from them.Three of the morphological groups were differenti-ated mainly by the length of their microawns. To testwhether they were indeed separate species ratherthan different stages of development of the same spe-cies or artifacts of cultural conditions, sequence anal-ysis from internal transcribed spacer regions (ITS) ofthe nuclear ribosomal repeat (nrDNA) was undertak-en. Sequence data also were used to clarify the place-ment of the genus among ascomycete lineages.
METHODS
Morphology of specimens from natural substrata and from cul-ture.—Specimens of Thozetella were isolated after they spor-ulated on decaying leaves incubated in moist chambers(Booth 1971). In addition, spore suspensions were spreadon 2% tap water agar containing 50 mg L21 streptomycinand after 3–4 d small colonies were subcultured (Booth
1075PAULUS ET AL: FIVE NEW SPECIES OF THOZETELLA
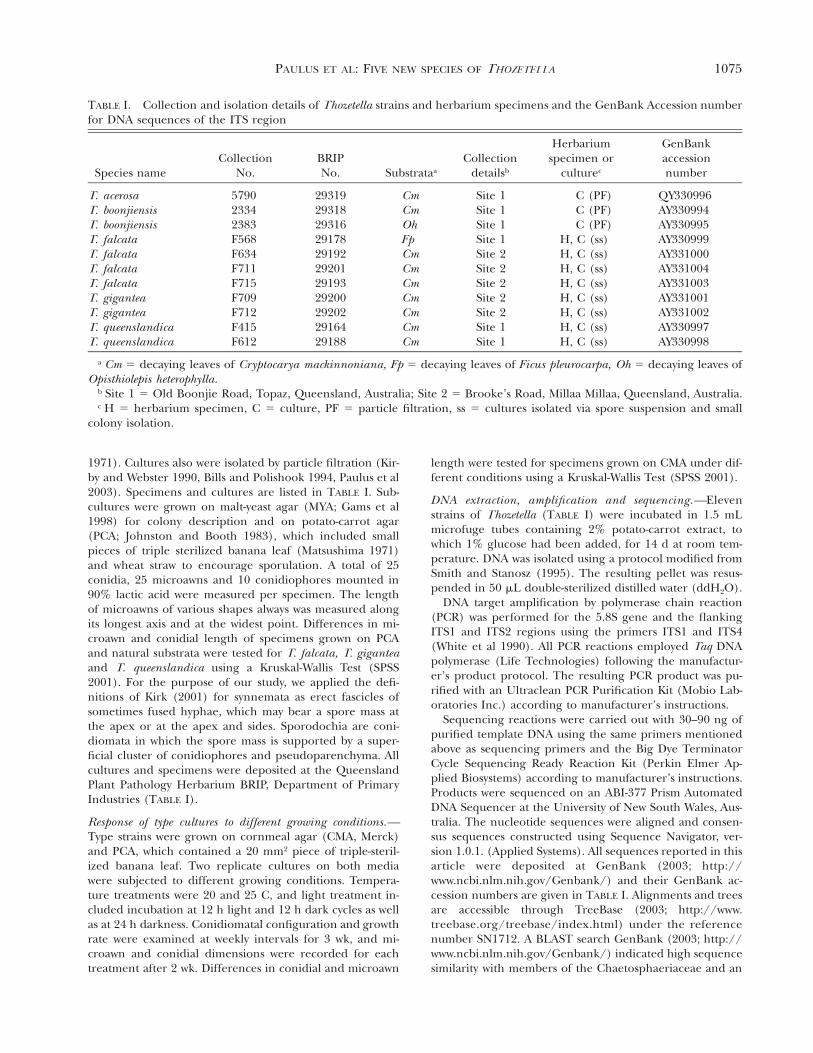
TABLE I. Collection and isolation details of Thozetella strains and herbarium specimens and the GenBank Accession numberfor DNA sequences of the ITS region
Species nameCollection
No.BRIPNo. Substrataa
Collectiondetailsb
Herbariumspecimen or
culturec
GenBankaccessionnumber
T. acerosaT. boonjiensisT. boonjiensisT. falcataT. falcata
579023342383F568F634
2931929318293162917829192
CmCmOhFpCm
Site 1Site 1Site 1Site 1Site 2
C (PF)C (PF)C (PF)
H, C (ss)H, C (ss)
QY330996AY330994AY330995AY330999AY331000
T. falcataT. falcataT. giganteaT. giganteaT. queenslandicaT. queenslandica
F711F715F709F712F415F612
292012919329200292022916429188
CmCmCmCmCmCm
Site 2Site 2Site 2Site 2Site 1Site 1
H, C (ss)H, C (ss)H, C (ss)H, C (ss)H, C (ss)H, C (ss)
AY331004AY331003AY331001AY331002AY330997AY330998
a Cm 5 decaying leaves of Cryptocarya mackinnoniana, Fp 5 decaying leaves of Ficus pleurocarpa, Oh 5 decaying leaves ofOpisthiolepis heterophylla.
b Site 1 5 Old Boonjie Road, Topaz, Queensland, Australia; Site 2 5 Brooke’s Road, Millaa Millaa, Queensland, Australia.c H 5 herbarium specimen, C 5 culture, PF 5 particle filtration, ss 5 cultures isolated via spore suspension and small
colony isolation.
1971). Cultures also were isolated by particle filtration (Kir-by and Webster 1990, Bills and Polishook 1994, Paulus et al2003). Specimens and cultures are listed in TABLE I. Sub-cultures were grown on malt-yeast agar (MYA; Gams et al1998) for colony description and on potato-carrot agar(PCA; Johnston and Booth 1983), which included smallpieces of triple sterilized banana leaf (Matsushima 1971)and wheat straw to encourage sporulation. A total of 25conidia, 25 microawns and 10 conidiophores mounted in90% lactic acid were measured per specimen. The lengthof microawns of various shapes always was measured alongits longest axis and at the widest point. Differences in mi-croawn and conidial length of specimens grown on PCAand natural substrata were tested for T. falcata, T. giganteaand T. queenslandica using a Kruskal-Wallis Test (SPSS2001). For the purpose of our study, we applied the defi-nitions of Kirk (2001) for synnemata as erect fascicles ofsometimes fused hyphae, which may bear a spore mass atthe apex or at the apex and sides. Sporodochia are coni-diomata in which the spore mass is supported by a super-ficial cluster of conidiophores and pseudoparenchyma. Allcultures and specimens were deposited at the QueenslandPlant Pathology Herbarium BRIP, Department of PrimaryIndustries (TABLE I).
Response of type cultures to different growing conditions.—Type strains were grown on cornmeal agar (CMA, Merck)and PCA, which contained a 20 mm2 piece of triple-steril-ized banana leaf. Two replicate cultures on both mediawere subjected to different growing conditions. Tempera-ture treatments were 20 and 25 C, and light treatment in-cluded incubation at 12 h light and 12 h dark cycles as wellas at 24 h darkness. Conidiomatal configuration and growthrate were examined at weekly intervals for 3 wk, and mi-croawn and conidial dimensions were recorded for eachtreatment after 2 wk. Differences in conidial and microawn
length were tested for specimens grown on CMA under dif-ferent conditions using a Kruskal-Wallis Test (SPSS 2001).
DNA extraction, amplification and sequencing.—Elevenstrains of Thozetella (TABLE I) were incubated in 1.5 mLmicrofuge tubes containing 2% potato-carrot extract, towhich 1% glucose had been added, for 14 d at room tem-perature. DNA was isolated using a protocol modified fromSmith and Stanosz (1995). The resulting pellet was resus-pended in 50 mL double-sterilized distilled water (ddH2O).
DNA target amplification by polymerase chain reaction(PCR) was performed for the 5.8S gene and the flankingITS1 and ITS2 regions using the primers ITS1 and ITS4(White et al 1990). All PCR reactions employed Taq DNApolymerase (Life Technologies) following the manufactur-er’s product protocol. The resulting PCR product was pu-rified with an Ultraclean PCR Purification Kit (Mobio Lab-oratories Inc.) according to manufacturer’s instructions.
Sequencing reactions were carried out with 30–90 ng ofpurified template DNA using the same primers mentionedabove as sequencing primers and the Big Dye TerminatorCycle Sequencing Ready Reaction Kit (Perkin Elmer Ap-plied Biosystems) according to manufacturer’s instructions.Products were sequenced on an ABI-377 Prism AutomatedDNA Sequencer at the University of New South Wales, Aus-tralia. The nucleotide sequences were aligned and consen-sus sequences constructed using Sequence Navigator, ver-sion 1.0.1. (Applied Systems). All sequences reported in thisarticle were deposited at GenBank (2003; http://www.ncbi.nlm.nih.gov/Genbank/) and their GenBank ac-cession numbers are given in TABLE I. Alignments and treesare accessible through TreeBase (2003; http://www.treebase.org/treebase/index.html) under the referencenumber SN1712. A BLAST search GenBank (2003; http://www.ncbi.nlm.nih.gov/Genbank/) indicated high sequencesimilarity with members of the Chaetosphaeriaceae and an

1076 MYCOLOGIA
unidentified leaf litter ascomycete (GenBank Accessionnumber AF502902).
DNA sequence analysis.—Two datasets were generated. Thefirst aligned ITS rDNA sequences from two representativeThozetella species, T. gigantea and T. queenslandica, with theChaetosphaeria dataset of Reblova and Winka (2000) avail-able from TreeBase (2003; http://www.treebase.org/treebase/index.html) under the reference number M1287.As a starting point, we accepted the alignment and out-groups proposed by Reblova and Winka (2000) but weremore conservative in the analysis by treating gaps as missingdata as opposed to fifth characters. The second dataset ofthe 11 available Thozetella sequences was used to examinethe phylogeny of Thozetella. We chose to undertake a sep-arate analysis because the alignment with the dataset of Re-blova and Winka (2000) was ambiguous in places and char-acters had to be excluded. Based on the results of the firststudy, we selected Codinaeopsis gonytrichoides (Shearer etJ.L. Crane) Morgan-Jones and Striatosphaeria codinaeaphoraSamuels et E. Mull. outgroups. Indels initially were exclud-ed from analysis, but to test the sensitivity of analysis tovariable alignment, indels were scored as presence/ab-sence. We were unable to use DNA of previously describedThozetella species for this study, which would have providedfurther information.
For both datasets, parsimony analysis was performed inPAUP* 4.0b10 (Swofford 2002). Gaps were treated as miss-ing data. The computing scheme used for parsimony anal-ysis was: heuristic search option with stepwise addition ran-dom, 1000 replications, tree bisection-reconnection branchswapping (TBR), MulTrees option in effect. A strict consen-sus tree was constructed from all most parsimonious treesand support for clades was estimated with a bootstrappingprocedure (Felsenstein 1985) using 1000 replications. Pair-wise genetic distances were calculated based on ‘‘uncor-rected p’’ (transitions plus transversions/total number ofnucleotides) as performed in PAUP*.
RESULTS
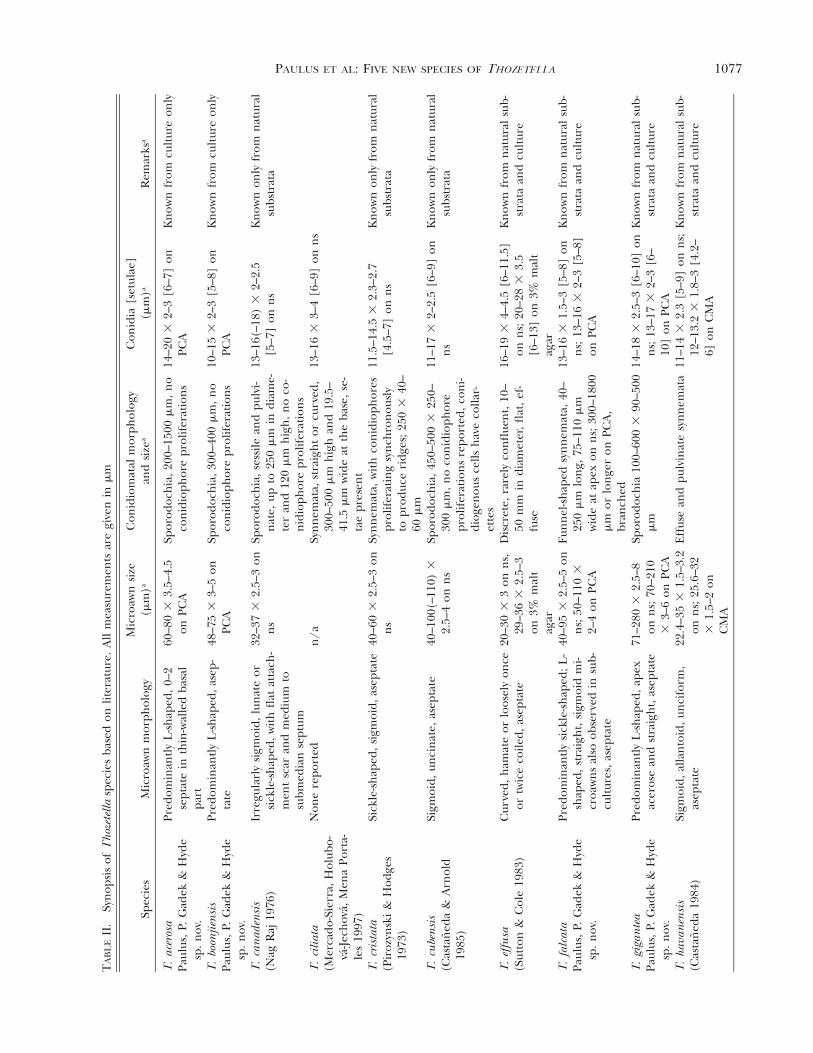
Taxonomy.—Microawn, conidioma and conidium siz-es and their morphology from fruiting bodies de-rived from natural substrata and from cultures fornew and previously described Thozetella species weresummarized (TABLE II).
Thozetella Kuntze, Revis. gen. pl. (Leipzig) 2:873(1891)
Type species: Thozetella nivea (Berk.) Kuntze, Re-vis. gen. pl. (Leipzig) 2:873 (1891)
Thozetella acerosa Paulus, P.Gadek et K.D. Hyde,sp.nov. FIG. 1AAd fungos conidiales, hyphomycetes pertinens. Coni-
diomatae sporodochiae vel effusae, albae, 200–1500 mm.Conidiophorae macronematae, brunnae, 2.5–4 mm latae.Cellulae conidiogenae pallidae brunneae, 12–20 3 2–4 mm.Microaristae angulatae, 0–2 septatae ad basi, apice acerosae
et basi leviter obtusae, hyalinae, 60–80 3 5–4.5 mm in PCA.Conidiae lunatae, unicellulariae, hyalinae, 14–20 3 2–3 mmin PCA, laeves et utrinque solo appendice, filiformi prae-dita, 6–7 mm longa.
Etymology. ‘‘acerasa’’ referring to the needle-likeapices of the microawns.
Colonies cream and light brown, flat, woolly to sub-felty, margin incised and indistinct. Conidiomata spo-rodochial or effuse, superficial, sessile, forming aconvex or flat hymenium, topped by a moist sporemass, 200–1500 mm diam. Conidiophores macrone-matous, brown, irregularly cylindrical, branched, aris-ing from a basal plate, 2.5–4 mm wide. Conidiogen-ous cells monophialidic, integrated, determinate, ter-minal, light brown, irregularly cylindrical, with no orminute collarette, periclinal wall thickened, 12–20 32–4 mm. Microawns produced from conidiophores,predominantly L-shaped, basal part thin-walled andhyaline, with 0–2 septa depending on growing con-ditions (TABLE III), apical part acerose and slightlyundulating, smooth, thick-walled with the upper por-tion becoming solid, refractive, 60–80 3 3.5–4.5 mmin PCA. Conidia lunate, aseptate, finely guttulate, hy-aline and smooth, 14–20 3 2–3 mm in PCA, with onefiliform setula at each end, 6–7 mm long.
Specimens examined. AUSTRALIA, QUEENSLAND: Ath-erton Tablelands, Topaz, Old Boonjie Road, on decayingleaves of Cryptocarya mackinnoniana, 8 Sep 2001, B. Paulusand C. Pearce, BP5970, HOLOTY PE: BRIP 29319.
Commentary. Thozetella acerosa resembles T. niveain microawn shape. Our isolate differs in havingshorter and narrower conidia and longer microawnsthan reported for T. nivea. In addition, our speci-mens have 0–2 septa at the basal part of the mi-croawns at 25 C while microawns of T. nivea are asep-tate. Two other species described below, namely T.boonjiensis and T. gigantea, also have L-shaped mi-croawns and form sporodochia. Mature specimens ofT. gigantea are distinguished easily because they havemuch longer microawns. Mean values of microawnand conidial length, as well as the presence or ab-sence of septation in microawns, clearly differentiatebetween T. acerosa and T. boonjiensis.
Thozetella boonjiensis Paulus, P.Gadek et K.D. Hyde,sp. nov. FIG. 1B–EAd fungos conidiales, hyphomycetes pertinens. Coloniae
cremeae et fuscae Conidiomatae sporodochiae vel effusae,120–250 3 300–400 mm. Conidiophorae macronematae,brunnae, ramosae, 1.5–3 mm latae. Cellulae conidiogenaepallidae brunneae, 15–20 3 1.5–2.5 mm. Microaristae an-gulatae, aseptatae, apice acerosae, refractivae et basi leviterobtusae, hyalinae, 48–75 3 3–5 mm in PCA. Conidiae lun-atae, unicellulariae, hyalinae, 10–15 3 2–3 mm in PCA, lae-ves et utrinque solo appendice, filiformi praedita, 5–8 mmlonga.
1077PAULUS ET AL: FIVE NEW SPECIES OF THOZETELLA
TA
BL
EII
.Sy
nop
sis
ofT
hoze
tella
spec
ies
base
don
liter
atur
e.A
llm
easu
rem
ents
are
give
nin
mm
Spec
ies
Mic
roaw
nm
orph
olog
yM
icro
awn
size
(mm
)a
Con
idio
mat
alm
orph
olog
yan
dsi
zea
Con
idia
[set
ulae
](m
m)a
Rem
arks
a
T.ac
eros
aPa
ulus
,P.
Gad
ek&
Hyd
esp
.n
ov.
Pred
omin
antl
yL
-sh
aped
,0–2
sept
ate
inth
in-w
alle
dba
sal
part
60–8
03
3.5–
4.5
onPC
ASp
orod
och
ia,2
00–1
500
mm
,n
oco
nid
ioph
ore
prol
ifer
atio
ns
14–2
03
2–3
[6–7
]on
PCA
Kn
own
from
cult
ure
only
T.bo
onjie
nsi
sPa
ulus
,P.
Gad
ek&
Hyd
esp
.n
ov.
Pred
omin
antl
yL
-sh
aped
,ase
p-ta
te48
–75
33–
5on
PCA
Spor
odoc
hia
,300
–400
mm
,n
oco
nid
ioph
ore
prol
ifer
atio
ns
10–1
53
2–3
[5–8
]on
PCA
Kn
own
from
cult
ure
only
T.ca
nad
ensi
s(N
agR
aj19
76)
Irre
gula
rly
sigm
oid,
lun
ate
orsi
ckle
-sh
aped
,wit
hfl
atat
tach
-m
ent
scar
and
med
ium
tosu
bmed
ian
sept
um
32–3
73
2.5–
3on
ns
Spor
odoc
hia
,ses
sile
and
pulv
i-n
ate,
upto
250
mm
indi
ame-
ter
and
120
mm
hig
h,
no
co-
nid
ioph
ore
prol
ifer
atio
ns
13–1
6(–1
8)3
2–2.
5[5
–7]
onn
sK
now
non
lyfr
omn
atur
alsu
bstr
ata
T.ci
liata
(Mer
cado
-Sie
rra,
Hol
ubo-
va-Je
chov
a,M
ena
Port
a-le
s19
97)
Non
ere
port
edn
/aSy
nn
emat
a,st
raig
ht
orcu
rved
,30
0–50
0m
mh
igh
and
19.5
–41
.5m
mw
ide
atth
eba
se,
se-
tae
pres
ent
13–1
63
3–4
[6–9
]on
ns
T.cr
ista
ta(P
iroz
ynsk
i&
Hod
ges
1973
)
Sick
le-s
hap
ed,s
igm
oid,
asep
tate
40–6
03
2.5–
3on
ns
Syn
nem
ata,
wit
hco
nid
ioph
ores
prol
ifer
atin
gsy
nch
ron
ousl
yto
prod
uce
ridg
es;
250
340
–60
mm
11.5
–14.
53
2.3–
2.7
[4.5
–7]
onn
sK
now
non
lyfr
omn
atur
alsu
bstr
ata
T.cu
ben
sis
(Cas
tan
eda
&A
rnol
d19
85)
Sigm
oid,
unci
nat
e,as
epta
te40
–100
(–11
0)3
2.5–
4on
ns
Spor
odoc
hia
,450
–500
325
0–30
0m
m,
no
con
idio
phor
epr
olif
erat
ion
sre
port
ed,
con
i-di
ogen
ous
cells
hav
eco
llar-
ette
s
11–1
73
2–2.
5[6
–9]
onn
sK
now
non
lyfr
omn
atur
alsu
bstr
ata
T.ef
fusa
(Sut
ton
&C
ole
1983
)C
urve
d,h
amat
eor
loos
ely
once
ortw
ice
coile
d,as
epta
te20
–30
33
onn
s,29
–36
32.
5–3
on3%
mal
tag
ar
Dis
cret
e,ra
rely
con
flue
nt,
10–
50m
min
diam
eter
,fl
at,
ef-
fuse
16–1
93
4–4.
5[6
–11.
5]on
ns;
20–2
83
3.5
[6–1
3]on
3%m
alt
agar
Kn
own
from
nat
ural
sub-
stra
taan
dcu
ltur
e
T.fa
lcat
aPa
ulus
,P.
Gad
ek&
Hyd
esp
.n
ov.
Pred
omin
antl
ysi
ckle
-sh
aped
;L-
shap
ed,
stra
igh
t,si
gmoi
dm
i-cr
oaw
ns
also
obse
rved
insu
b-cu
ltur
es,a
sept
ate
40–9
53
2.5–
5on
ns;
50–1
103
2–4
onPC
A
Fun
nel
-sh
aped
syn
nem
ata,
40–
250
mm
lon
g,75
–110
mm
wid
eat
apex
onn
s;30
0–18
00m
mor
lon
ger
onPC
A,
bran
ched
13–1
63
1.5–
3[5
–8]
onn
s;13
–16
32–
3[5
–8]
onPC
A
Kn
own
from
nat
ural
sub-
stra
taan
dcu
ltur
e
T.gi
gan
tea
Paul
us,
P.G
adek
&H
yde
sp.
nov
.
Pred
omin
antl
yL
-sh
aped
,ape
xac
eros
ean
dst
raig
ht,
asep
tate
71–2
803
2.5–
8on
ns;
70–2
103
3–6
onPC
A
Spor
odoc
hia
100–
600
390
–500
mm
14–1
83
2.5–
3[6
–10]
onn
s;13
–17
32–
3[6
–10
]on
PCA
Kn
own
from
nat
ural
sub-
stra
taan
dcu
ltur
e
T.ha
van
ensi
s(C
asta
ned
a19
84)
Sigm
oid,
alla
nto
id,u
nci
form
,as
epta
te22
.4–3
53
1.5–
3.2
onn
s;25
.6–3
23
1.5–
2on
CM
A
Eff
use
and
pulv
inat
esy
nn
emat
a11
–14
32.
3[5
–9]
onn
s;12
–13.
23
1.8–
3[4
.2–
6]on
CM
A
Kn
own
from
nat
ural
sub-
stra
taan
dcu
ltur
e

1078 MYCOLOGIAT
AB
LE
II.
Con
tin
ued
Spec
ies
Mic
roaw
nm
orph
olog
yM
icro
awn
size
(mm
)a
Con
idio
mat
alm
orph
olog
yan
dsi
zea
Con
idia
[set
ulae
](m
m)a
Rem
arks
a
T.n
ivea
(Kun
tze
1891
)Pr
edom
inan
tly
L-s
hap
ed,
undu
late
,ace
rose
apex
,ase
ptat
e50
–70
33–
4Sp
orod
och
ium
,200
–250
mm
indi
amet
er(1
7.5–
)21(
–24)
33–
3.8
[6–9
]K
now
non
lyfr
omn
atur
alsu
bstr
ata
T.qu
een
slan
dica
Paul
us,
P.G
adek
&H
yde
sp.
nov
.
Pred
omin
antl
yh
amat
e,as
epta
te24
–33
32–
3.5
onn
s;21
–34
32–
4on
PCA
Spor
odoc
hia
,up
to20
0–25
03
50–1
25m
mw
ith
con
idio
phor
espr
olif
erat
ing
syn
chro
nou
sly
inso
me
spec
imen
sto
prod
uce
ridg
es
10–1
23
1.5–
2.5
[3–6
]on
ns;
9–12
31.
5–2.
5[3
–6]
onPC
A
Kn
own
from
nat
ural
sub-
stra
taan
dcu
ltur
e
T.ra
dica
ta(P
iroz
ynsk
i&
Hod
ges
1973
)
Cur
ved,
sigm
oid
orL
-sh
aped
,as
epta
te30
–60
33–
4.5
incu
ltur
eFu
nn
elsh
aped
syn
nem
ata
(up
to75
0m
mlo
ng)
11–1
33
2.5–
3[3
]bin
cult
ure
Kn
own
from
cult
ure
only
T.to
ckla
ien
sis
(Pir
ozyn
ski
&H
odge
s19
73)
L-s
hap
edan
dot
her
con
figu
ra-
tion
s,as
epta
te18
–38(
–44)
31.
5–3
onm
alta
gar
On
nat
ural
subs
trat
umsp
orod
och
ia(1
000
mm
wid
e3
300
mm
hig
h),
syn
nem
atal
incu
ltur
e(1
–1.5
mm
lon
g)
9–13
(–18
)3
1.5–
3[3
–8(–
13)]
onm
alt
agar
Kn
own
from
nat
ural
sub-
stra
taan
dcu
ltur
e
an
s5
nat
ural
subs
trat
a.b
Acc
ordi
ng
toPi
rozy
nsk
ian
dH
odge
s(1
973)
,or
igin
alde
scri
ptio
n13
–17
32.
5–3
mm
.
Etymology. ‘‘boonjiensis’’ referring to the place ofcollection near the Old Boonjie Road in northernQueensland.
Colonies cream and pale brown, subfelty, slimy, re-verse pale brown. Conidiomata sessile sporodochia,300–400 mm diam, or flat effuse development of co-nidiophores. Conidiophores macronematous, brown,cylindrical, branched, arising from a basal plate ofcells, 1.5–3 mm wide. Conidiogenous cells mono-phialidic, integrated, determinate, terminal, lightbrown, irregularly cylindrical, lacking an apical col-larette, no or slight periclinal wall thickening, 15–203 1.5–2.5 mm. Microawns produced from conidio-phores, predominantly L-shaped, aseptate, smooth,apical part acerose, straight or slightly undulating,thick-walled, refractive, basal part thin-walled, hya-line, aseptate, 48–75 3 3–5 mm on PCA. Conidia lu-nate, continuous, finely guttulate or eguttulate, hya-line, smooth, 10–15 3 2–3 mm on PCA, supplied witha single, 5–8 mm long, filiform setula at each end.
Specimens examined. AUSTRALIA, QUEENSLAND, Ath-erton Tablelands, Topaz, Old Boonjie Road, on decayingleaves of C. mackinnoniana, 11 Mar 2001, B. Paulus and C.Pearce, BP2334, HOLOTY PE: BRIP 29318. AUSTRALIA,QUEENSLAND: Atherton Tablelands, Topaz, Old BoonjieRoad, on decaying leaves of Opisthiolepis heterophylla, 11Mar 2001, B. Paulus and C. Pearce, BP2383, BRIP 29316.
Commentary. Thozetella boonjiensis resembles T.tocklaiensis in microawn shape and conidial dimen-sions. Our isolates differ in having longer microawns(48–75 3 3–5 mm rather than 18–38[–44] 3 1.5–5mm) and in producing sporodochia or effuse coni-diomata rather than the synnemata reported for cul-tures of T. tocklaiensis. Two other species describedhere, namely T. acerosa and T. gigantea, also have L-shaped microawns and form sporodochia. Maturespecimens of T. gigantea are easily distinguished be-cause they have much longer microawns. Young spec-imens of T. gigantea may show an overlap in mi-croawn dimensions with T. boonjiensis but can be dif-ferentiated because they have longer conidia. Meanvalues of microawn and conidial length, as well as thepresence or absence of septation in microawns clearlydifferentiate between T. acerosa and T. boonjiensis.
Thozetella falcata Paulus, P. Gadek et K.D. Hyde, sp.nov. FIG. 1F–LAd fungos conidiales, hyphomycetes pertinens. Coni-
diomatae synnematae, brunneae, 40–250 3 12–35 mm adbasim et 75–110 mm ad apicem in foliis dejectis, 300–18003 12–80 mm ad basim et 120–300 mm ad apicem in PCA.Conidiophorae macronematae, brunnae, 1.5–3 mm latae.Cellulae conidiogenae pallidae brunneae, 6–19 3 2–3 mm.Microaristae hamatae, angulatae, sigmoidae vel rectae, api-ce crassitunicatae, refractivae et basi leviter obtusae, hyali-
1079PAULUS ET AL: FIVE NEW SPECIES OF THOZETELLA
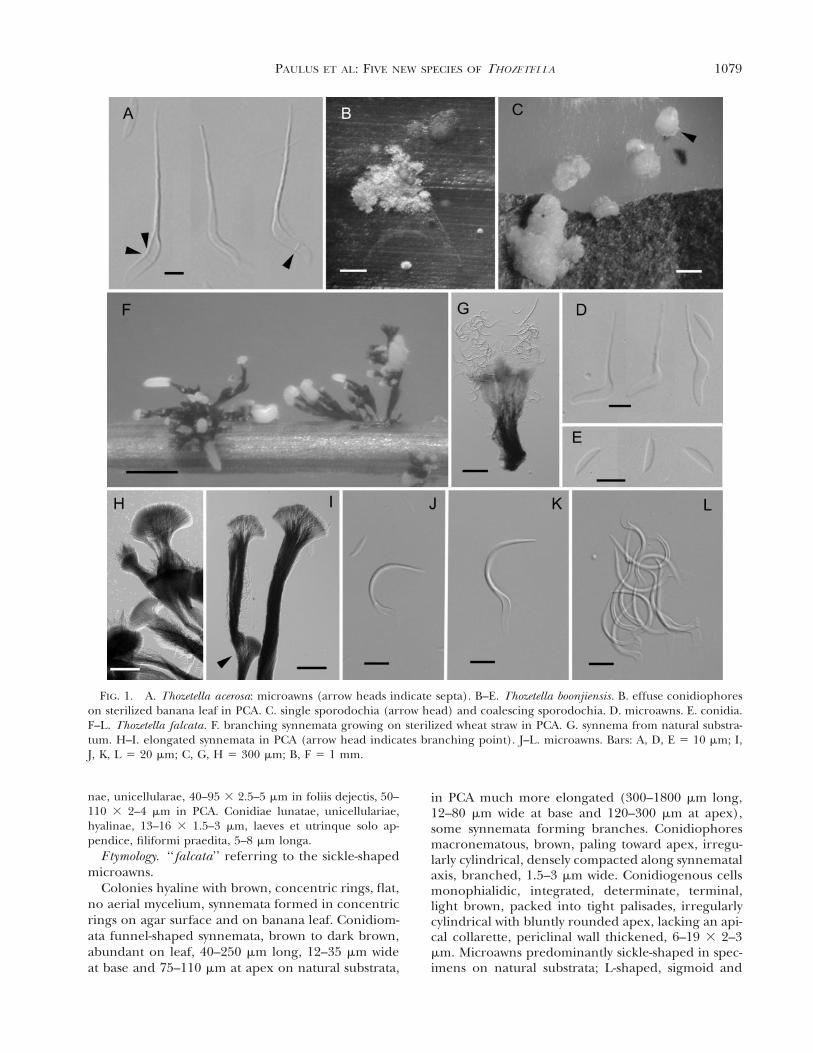
FIG. 1. A. Thozetella acerosa: microawns (arrow heads indicate septa). B–E. Thozetella boonjiensis. B. effuse conidiophoreson sterilized banana leaf in PCA. C. single sporodochia (arrow head) and coalescing sporodochia. D. microawns. E. conidia.F–L. Thozetella falcata. F. branching synnemata growing on sterilized wheat straw in PCA. G. synnema from natural substra-tum. H–I. elongated synnemata in PCA (arrow head indicates branching point). J–L. microawns. Bars: A, D, E 5 10 mm; I,J, K, L 5 20 mm; C, G, H 5 300 mm; B, F 5 1 mm.
nae, unicellularae, 40–95 3 2.5–5 mm in foliis dejectis, 50–110 3 2–4 mm in PCA. Conidiae lunatae, unicellulariae,hyalinae, 13–16 3 1.5–3 mm, laeves et utrinque solo ap-pendice, filiformi praedita, 5–8 mm longa.
Etymology. ‘‘falcata’’ referring to the sickle-shapedmicroawns.
Colonies hyaline with brown, concentric rings, flat,no aerial mycelium, synnemata formed in concentricrings on agar surface and on banana leaf. Conidiom-ata funnel-shaped synnemata, brown to dark brown,abundant on leaf, 40–250 mm long, 12–35 mm wideat base and 75–110 mm at apex on natural substrata,
in PCA much more elongated (300–1800 mm long,12–80 mm wide at base and 120–300 mm at apex),some synnemata forming branches. Conidiophoresmacronematous, brown, paling toward apex, irregu-larly cylindrical, densely compacted along synnematalaxis, branched, 1.5–3 mm wide. Conidiogenous cellsmonophialidic, integrated, determinate, terminal,light brown, packed into tight palisades, irregularlycylindrical with bluntly rounded apex, lacking an api-cal collarette, periclinal wall thickened, 6–19 3 2–3mm. Microawns predominantly sickle-shaped in spec-imens on natural substrata; L-shaped, sigmoid and

1080 MYCOLOGIA
TABLE III. Percentage of microawns in Thozetella acerosathat formed septa under different growing conditions
Media Temperature Light
No. of septa
0 1 2 3
CMA 20 C
25 C
light/darkdarklight/darkdark
95403525
5505565
555
555
PCA 20 C
25 C
light/darkdarklight/darkdark
100904540
104550
105 5
FIG. 2. A–I. Thozetella queenslandica. A. sporodochium from natural substratum. B. older sporodochium from naturalsubstratum. Arrow heads indicate ridges formed by conidiophore proliferation. C. conidiophores. D. sporodochia on abaxialsurface of Cryptocarya mackinnoniana leaf. E–H. microawns. I. conidia. J–M. Thozetella gigantea. J. part of sporodochium,spore mass and microawns of subcultures on PCA. K, L. microawns. M. microawn and conidia. Bars: A, B 5 100 mm; H–I 525 mm. C, E–F, J–M 5 10 mm; D 5 1 mm.
straight microawns also were observed in subculturesin addition to sickle-shaped microawns, 40–95 3 2.5–5 mm on natural substrata, 50–110 3 2–4 mm onPCA. Conidia lunate, aseptate, finely guttulate oreguttulate, hyaline, smooth, 13–16 3 1.5–3 mm onnatural substrata, 13–16 3 2–3 mm on PCA, providedwith a single, filiform setula at either pole, 5–8 mmlong.
Specimens examined. AUSTRALIA, QUEENSLAND: Ath-erton Tablelands, Millaa Millaa, Brooke’s Road, on decayingleaves of C. mackinnoniana, 28 May 2002, B. Paulus andI.G. Steer, BP F715, HOLOTY PE: BRIP29193. AUSTRALIA,QUEENSLAND: Atherton Tablelands, Topaz, Old BoonjieRoad, on decaying leaves of Ficus pleurocarpa, 23 Mar 2002,B. Paulus and I.G. Steer, BP F568, BRIP 29178. AUSTRA-LIA, QUEENSLAND, Atherton Tablelands, Millaa Millaa,
Brooke’s Road, on decaying leaves of C. mackinnoniana, 28May 2002, B. Paulus and I.G. Steer, BP F634, BRIP 29192.AUSTRALIA, QUEENSLAND: Atherton Tablelands, MillaaMillaa, Brooke’s Road, on decaying leaves of C. mackinnon-iana, 21 Jun 2002, B. Paulus and I.G. Steer, BP F711, BRIP29201.
Commentary. Thozetella falcata, T. cristata, T. radi-cata and T. tocklaiensis all produce synnemata. Thoz-etella falcata is also similar to T. cristata and T. radi-cata in microawn shape and in conidial dimensions.However, T. falcata differs from these two species inhaving considerably longer microawns on naturalsubstrata and in culture. In addition, T. falcata differsfrom T. radicata in having branched and longer syn-nemata. Conidiomata of T. falcata are also differentfrom those of T. cristata, in which conidiophores pro-liferate simultaneously to form visible ridges. In ad-dition, Thozetella cristata forms consistently cylindri-cal synnemata while those in T. falcata are funnel-shaped. The length and morphology of synnemata inT. tocklaiensis are similar to those of T. falcata, how-ever, these two fungal taxa differ in microawn mor-phology and size.
Thozetella gigantea Paulus, P.Gadek et K.D. Hyde, sp.nov. FIG. 2J–MAd fungos conidiales, hyphomycetes pertinens. Coni-
diomatae sporodochiae, 100–600 3 90–500 mm. Conidi-ophorae macronematae, brunnae, 2.5–4 mm latae. Cellulaeconidiogenae pallidae brunneae, 12–20 3 2–4 mm. Microar-
1081PAULUS ET AL: FIVE NEW SPECIES OF THOZETELLA
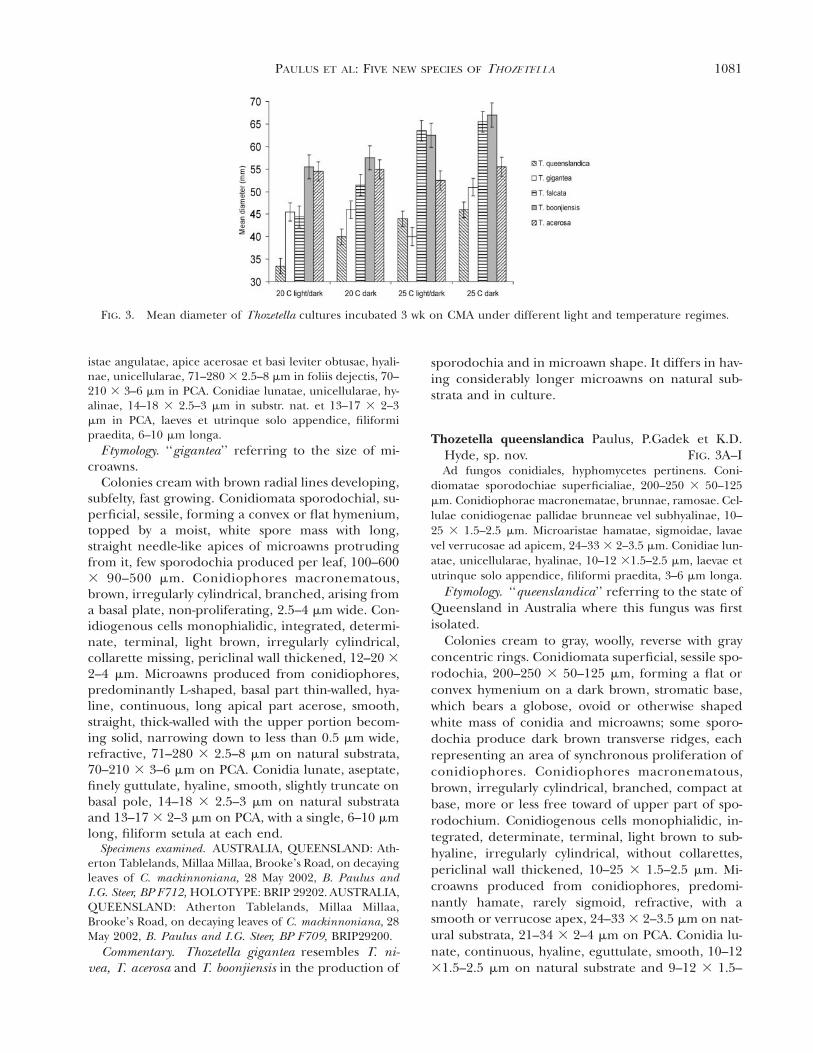
FIG. 3. Mean diameter of Thozetella cultures incubated 3 wk on CMA under different light and temperature regimes.
istae angulatae, apice acerosae et basi leviter obtusae, hyali-nae, unicellularae, 71–280 3 2.5–8 mm in foliis dejectis, 70–210 3 3–6 mm in PCA. Conidiae lunatae, unicellularae, hy-alinae, 14–18 3 2.5–3 mm in substr. nat. et 13–17 3 2–3mm in PCA, laeves et utrinque solo appendice, filiformipraedita, 6–10 mm longa.
Etymology. ‘‘gigantea’’ referring to the size of mi-croawns.
Colonies cream with brown radial lines developing,subfelty, fast growing. Conidiomata sporodochial, su-perficial, sessile, forming a convex or flat hymenium,topped by a moist, white spore mass with long,straight needle-like apices of microawns protrudingfrom it, few sporodochia produced per leaf, 100–6003 90–500 mm. Conidiophores macronematous,brown, irregularly cylindrical, branched, arising froma basal plate, non-proliferating, 2.5–4 mm wide. Con-idiogenous cells monophialidic, integrated, determi-nate, terminal, light brown, irregularly cylindrical,collarette missing, periclinal wall thickened, 12–20 32–4 mm. Microawns produced from conidiophores,predominantly L-shaped, basal part thin-walled, hya-line, continuous, long apical part acerose, smooth,straight, thick-walled with the upper portion becom-ing solid, narrowing down to less than 0.5 mm wide,refractive, 71–280 3 2.5–8 mm on natural substrata,70–210 3 3–6 mm on PCA. Conidia lunate, aseptate,finely guttulate, hyaline, smooth, slightly truncate onbasal pole, 14–18 3 2.5–3 mm on natural substrataand 13–17 3 2–3 mm on PCA, with a single, 6–10 mmlong, filiform setula at each end.
Specimens examined. AUSTRALIA, QUEENSLAND: Ath-erton Tablelands, Millaa Millaa, Brooke’s Road, on decayingleaves of C. mackinnoniana, 28 May 2002, B. Paulus andI.G. Steer, BP F712, HOLOTY PE: BRIP 29202. AUSTRALIA,QUEENSLAND: Atherton Tablelands, Millaa Millaa,Brooke’s Road, on decaying leaves of C. mackinnoniana, 28May 2002, B. Paulus and I.G. Steer, BP F709, BRIP29200.
Commentary. Thozetella gigantea resembles T. ni-vea, T. acerosa and T. boonjiensis in the production of
sporodochia and in microawn shape. It differs in hav-ing considerably longer microawns on natural sub-strata and in culture.
Thozetella queenslandica Paulus, P.Gadek et K.D.Hyde, sp. nov. FIG. 3A–IAd fungos conidiales, hyphomycetes pertinens. Coni-
diomatae sporodochiae superficialiae, 200–250 3 50–125mm. Conidiophorae macronematae, brunnae, ramosae. Cel-lulae conidiogenae pallidae brunneae vel subhyalinae, 10–25 3 1.5–2.5 mm. Microaristae hamatae, sigmoidae, lavaevel verrucosae ad apicem, 24–33 3 2–3.5 mm. Conidiae lun-atae, unicellularae, hyalinae, 10–12 31.5–2.5 mm, laevae etutrinque solo appendice, filiformi praedita, 3–6 mm longa.
Etymology. ‘‘queenslandica’’ referring to the state ofQueensland in Australia where this fungus was firstisolated.
Colonies cream to gray, woolly, reverse with grayconcentric rings. Conidiomata superficial, sessile spo-rodochia, 200–250 3 50–125 mm, forming a flat orconvex hymenium on a dark brown, stromatic base,which bears a globose, ovoid or otherwise shapedwhite mass of conidia and microawns; some sporo-dochia produce dark brown transverse ridges, eachrepresenting an area of synchronous proliferation ofconidiophores. Conidiophores macronematous,brown, irregularly cylindrical, branched, compact atbase, more or less free toward of upper part of spo-rodochium. Conidiogenous cells monophialidic, in-tegrated, determinate, terminal, light brown to sub-hyaline, irregularly cylindrical, without collarettes,periclinal wall thickened, 10–25 3 1.5–2.5 mm. Mi-croawns produced from conidiophores, predomi-nantly hamate, rarely sigmoid, refractive, with asmooth or verrucose apex, 24–33 3 2–3.5 mm on nat-ural substrata, 21–34 3 2–4 mm on PCA. Conidia lu-nate, continuous, hyaline, eguttulate, smooth, 10–1231.5–2.5 mm on natural substrate and 9–12 3 1.5–

1082 MYCOLOGIA
TABLE IV. Sporulation of type cultures after 2 wk incubation on two media and under different conditions
20 C
Light/dark Dark
25 C
Light/dark Dark
CMA T. acerosaT. boonjiensisT. falcataT. giganteaT. queenslandica
sp & eff 1—sy 11sp (1)sp (1)
sp 11sp (1)sy 11—sp 1
sp 11sp & eff 1sy 11sp 1sp 1
sp 111—sy 11sp 1—
PCA T. acerosaT. boonjiensisT. falcataT. giganteaT. queenslandica
sp & eff 11—sy 11—sp 1
sp & eff 11—sy 1—sp 1
sp & eff 11sp & eff, sy 1sy 11(sp (1)sp 11
sp 11—sy 11——
sp 5 sporodochia.sy 5 synnemata.eff 5 effuse layer of conidiophores.— none, (1) 5 few, 1 5 some, 11 5 many, 111 5 abundant sporulating structures.
2.5 mm on PCA, provided with a single filiform setulaat each end, 3–6 mm long.
Specimens examined. AUSTRALIA, QUEENSLAND: Ath-erton Tablelands, Topaz, Old Boonjie Road, on decayingleaves of C. mackinnoniana, 28 May 2002, B. Paulus andI.G. Steer, BP F612, HOLOTY PE: BRIP 29188. AUSTRALIA,QUEENSLAND: Atherton Tablelands, Topaz, Old BoonjieRoad, on decaying leaves of C. mackinnoniana, 23 Mar2002, B. Paulus and I.G. Steer, BP F415, BRIP 29164.
Commentary. Thozetella queenslandica can be dif-ferentiated from other known species of Thozetella onthe basis of its distinctive, short hamate microawnsand the production of sporodochia, which show pro-liferation ridges. The microawns of T. queenslandicaoverlap in length with those of T. effusa and T. hav-anensis, but these species differ from T. queenslandicain both microawn and conidiomatal morphology.
Growth and sporulation of cultures under different grow-ing conditions.—Although some variation of mi-croawn and conidial lengths were observed in speci-mens grown on PCA and on natural substrata, thesewere not found to be significant (df 5 1; P 5 0.524and df 5 1; P 5 0.727, respectively). Similarly, nosignificant differences in microawn and conidiallengths were noted for cultures grown on differentmedia (df 5 1; P 5 0.457 and df 5 1; P 5 0.583,respectively). Microawn length also did not differ sig-nificantly for cultures grown under different lightand temperature treatments (df 5 3; P 5 0.968). Incontrast, conidial length differed significantly underdifferent cultures conditions (df 5 3; P 5 0.002).Examination of raw data (not shown) revealed thatconidia of T. gigantea were markedly longer at 25 Ccompared to 20 C (18.3 and 15.2 mm respectively).The mean diameter of type strains incubated 3 wk onCMA at different temperature and light regimes is
given in FIG. 3. All cultures grew well on CMA andPCA, and generally similar patterns were observed onboth media. One group, which included T. queen-slandica, T. falcata and T. boonjiensis, appeared togrow faster at 25 C. In contrast, T. gigantea and T.acerosa appeared to favor 20 C with the slower growthrate observed at 25 C with light/dark cycles.
A summary of sporulation patterns after 2 wk isprovided in TABLE IV. Thozetella acerosa and T. falcatacommenced sporulation in the first week and contin-ued to sporulate profusely on both media and underall conditions. Scant sporulation from the secondweek onward was observed for T. gigantea and T.boonjiensis. Thozetella queenslandica sporulated wellunder a relatively wide range of conditions but didnot form fruiting structures at 25 C in the dark oneither medium (TABLE IV). With some exceptions,conidiomata of all species appeared to be consistentin their basic configuration over the 3 wk observationperiod. Exceptions included the development of ef-fuse conidiophores in addition to sporodochia in T.acerosa and T. boonjiensis under some conditions.Thozetella boonjiensis also developed cream to lightbrown, submerged synnemata at 25 C on PCA whenexposed to light/dark cycles. At 25 C synnemata ofT. falcata were submerged in the agar.
KEY TO THE DESCRIBED SPECIES OF THOZETELLA
1. microawns present . . . . . . . . . . . . . . . . . . . . . . . . . 21. microawns absent . . . . . . . . . . . . . . . . . . . . . T. ciliata
2. microawns predominantly L-shaped . . . . . . . . . 32. microawns not L-shaped or variously shaped2 . . 6
3. microawns 0–2 septate, 60–80 3 3.5–4.5 mm (mean72 3 3.9 mm) . . . . . . . . . . . . . . . . . . . . . . . T. acerosa
3. microawns aseptate . . . . . . . . . . . . . . . . . . . . . . . . . 44. microawns very long, 70–280 mm 3 2.5–8 mm

1083PAULUS ET AL: FIVE NEW SPECIES OF THOZETELLA
(mean 153.8 3 5 mm on natural substrata, 132.43 4.2 mm on PCA) . . . . . . . . . . . . . . . T. gigantea
4. microawns shorter than 75 mm . . . . . . . . . . . . 55. conidia (17.5–)21(–24) 3 3–3.8 mm, microawns 50–
70 3 3–4 mm, microawn apex undulating or genic-ulate . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . T. nivea
5. conidia 10–15 3 2–3 mm, microawns 48–75 3 3–5mm (mean 60.1 3 3.6 mm), microawn apex straightor slightly undulating . . . . . . . . . . . . . . T. boonjiensis6. microawns predominantly sickle-shaped, unci-
nate, hamate or otherwise strongly curved . . . 76. microawns predominantly straight, sigmoid or
any other shape . . . . . . . . . . . . . . . . . . . . . . . 117. conidiomata predominantly synnematous . . . . . . . . 87. conidiomata predominantly sporodochial . . . . . . 10
8. synnemata cylindrical, proliferating conidio-phores form ridges, microawns 40–60 3 2.5–3mm, conidia 11.5–14.5 3 2.3–2.7 mm . . T. cristata
8. synnemata funnel-shaped, conidiophores non-proliferating . . . . . . . . . . . . . . . . . . . . . . . . . . . 9
9. microawns 40–95 3 2.5–5 mm (mean 59.9 3 3.6 mmon natural substratum, 77.2 3 3.3 mm in PCA), co-nidia 13–16 3 1.5–3 mm, synnemata branching . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . T. falcata
9. microawns 30–60 3 3–4.5 mm, conidia 11–13 3 2–2.5 mm . . . . . . . . . . . . . . . . . . . . . . . . . . . . T. radicata10. microawns 21–34 3 2–4 mm (mean 27.9 3 3 mm
on natural substrata, 26.9 3 2.9 mm on PCA),proliferation ridges on sporodochia, conidia 9–12 3 1.5–2.5 mm . . . . . . . . . . . . . T. queenslandica
10. microawns 40–110 3 2.5–4 mm, conidia 11–173 2–2.5 mm, sporodochia without proliferationridges . . . . . . . . . . . . . . . . . . . . . . . . . T. cubensis
11. conidiomata effuse, never sporodochial or synne-matous, microawns 20–30 3 3 mm, conidia 16–19 34–4.5 mm . . . . . . . . . . . . . . . . . . . . . . . . . . . . T. effusa
11. conidiomata sporodochial or synnematous . . . . . . 1212. conidiomata sporodochial, microawns 32–37 3
2.5–3 mm, conidia 13–16(–18) 3 2–2.5 mm . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . T. canadensis
12. conidiomata synnematous . . . . . . . . . . . . . . . . 1313. microawns variously shaped2, bulbous base, acerose
apex, straight, undulate, uncinate or bent 18–38(–44) 3 1.5–4 mm, conidia 9–13(–18) 3 1.5–3 mm. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . T. tocklaiensis
13. microawns with 6 uniform width, sigmoid, allan-toid, uncinate, verruculose, 22.4–35 3 1.5–3.2 mm,conidia 11–14 3 2.3 mm, conidiomata synnematousor effuse . . . . . . . . . . . . . . . . . . . . . . . . T. havanensis
ITS DNA sequence analyses.—The aligned dataset ofThozetella and Chaetosphaeria ITS sequences con-tained 529 positions. Of the 503 characters includedin analysis, 284 were variable and 219 were constant.Among the variable sites, 209 were parsimony infor-
2 The original descriptions of T. tocklaiensis shows microawns ofvarious shapes, whereas Pirozynski and Hodges (1973) show themicroawns of T. tocklaiensis to be L-shaped. We have accepted theoriginal description for this key.
mative. The heuristic search identified two equallyparsimonious trees, each 872 steps long, which dif-fered only in their placement of Chaetosphaeria dila-bens Reblova et W. Gams. The strict consensus treeshowed 11 clades, which were supported by bootstrapvalues higher than 55% (FIG. 4). The two represen-tative taxa of Thozetella grouped next to Striatosphaer-ia codinaeaphora and Codinaeopsis gonytrichoides withthis clade having a bootstrap support of less than55% (FIG. 4). A branch supporting the majority ofChaetosphaeria species has high bootstrap support(92%). The genetic distances of all Chaetosphaeriaspecies excluding outgroups and excluding Thozetellawere 0.0–25.5%; the genetic distances of Thozetellaand Chaetosphaeria species were 11.0–31.8% (detailsnot shown).
The aligned dataset of specimens of Thozetella plustwo outgroups contained 494 positions excluding in-del scores. Of the 467 characters included in theanalyses, 133 were variable and 334 were constant.Among the variable sites, 66 were parsimony infor-mative. Intraspecific genetic distances were low (0–0.7%), whereas interspecific genetic differences were2.3–8.5%. The lowest interspecific genetic distancewas observed between T. acerosa and T. boonjiensis(2.3 and 2.4%). The next closest species was T. gi-gantea, which differed from these two species be-tween 4.1 and 4.4%. The greatest genetic distancewas observed between T. gigantea and T. queenslan-dica (8.2 and 8.4%), while T. falcata differed from allother species between 4.9 and 6.4%. When indelswere excluded from analysis, three most parsimoni-ous trees were obtained, each 189 steps long and dif-fering only in the arrangement of terminal brancheswithin the clade representing Thozetella falcata. Thestrict consensus tree of three most parsimonious treesshows two clades and five subclades (FIG. 5). Thoze-tella boonjiensis and T. acerosa are sister clades andgroup next to T. gigantea. This tree also shows T.falcata and T. queenslandica to be sister clades. Boot-strap support is generally moderate to high (82–100%) but two branches have bootstrap values ofonly 56 and 59% (FIG. 5). One of these branches (viz.the branch supporting T. queenslandica and T. fal-cate) collapsed to form a polytomy when the datawere re-analyzed with indel scores included (notshown).
DISCUSSION
Relationship of Thozetella and Chaetosphaeria.—Analysis of ITS sequence data associated the ana-morph genus Thozetella with the ascomycete genusChaetosphaeria (FIG. 4). Reblova and Winka (2000)discussed the phylogeny of Chaetosphaeria and used

1084 MYCOLOGIA
FIG. 4. Phylogenetic relationships among species of Chaetosphaeria and its anamorphs based on ITS rDNA sequences.The phylogram, which is rooted by the outgroup method, depicts the strict consensus tree of five most parsimonious trees.Significant bootstrap values (.55%) are given above branches.
two separate genomic regions in their analysis. OurITS rDNA analysis was conservative and, hence, theresulting strict consensus tree was less resolved thanthat of Reblova and Winka (2000). Subgroups 1A and1B inferred by Reblova and Winka (2000) clusteredtogether, and the two Thozetella sequences insertedin this clade (FIG. 4). In addition to two teleomorphgenera, Chaetosphaeria pulviscula and Striatosphaeriacodinaeaphora, subgroup 1A includes the anamorphsDictyochaeta, Menispora and Codinaeopsis, which arecharacterized by producing oblong, fusiform, oftencurved conidia with terminal setulae (Reblova andWinka 2000). The genus Thozetella shares these mor-phological characteristics and, therefore, morpholog-ical data are consistent with the results of ITS se-quence analysis, despite the low bootstrap support
values obtained for those clades. The teleomorphs ofindividual Thozetella species remain unknown be-cause cultures did not produce ascomata and nucle-otide divergence among available Thozetella andChaetosphaeria sequences consistently was greaterthan 11%.
Phylogenetic analysis of Thozetella species.—This studyapplied both phylogenetic and morphological spe-cies recognition (Taylor et al 2000) and both werecongruent. For example, sequence analysis of ITSrDNA supported recognition of the morphologicalspecies as five distinct clades (FIG. 5). The strict con-sensus tree of the Thozetella sequences suggests thatthey are a strongly supported monophyletic group(100%, FIG. 5). Two main sister clades are resolved

1085PAULUS ET AL: FIVE NEW SPECIES OF THOZETELLA
FIG. 5. Phylogenetic relationships among species of Thozetella based on ITS rDNA sequences. The phylogram, which isrooted by the outgroup method, depicts the strict consensus tree of three most parsimonious trees. Significant bootstrapvalues (.55%) are given above the branches.
with the clade grouping T. acerosa, T. boonjiensis andT. gigantea showing a moderately high bootstrap val-ue (82%, FIG. 5). These species share the same basicmicroawn shape and predominant conidiomatal con-figuration but differ in microawn length (TABLE II).In addition to their morphological similarity, theyalso have the smallest interspecific genetic differenc-es observed in our study.
Despite its relatively low bootstrap value, the cladesupporting T. acerosa and T. boonjiensis remained sta-ble when indel scores were included (not shown).The other clade, which includes T. queenslandica andT. falcata, was weakly supported (bootstrap value59%, FIG. 5) and the branch supporting this cladecollapsed when indel scores were included in analy-sis. Because insertions and deletions are thought tobe the primary source of ITS variation in fungi (Hib-bett 1992), the close relationship between T. queen-slandica and T. falcata cannot be inferred with asmuch confidence as that of the species in the firstclade. This uncertainty is mirrored in the morphol-ogy of these species and the moderately high geneticdistance (6.2 and 6.4%). Although T. queenslandicaand T. falcata share a similar microawn shape (i.e.,
curved), their basic conidiomatal configurations dif-fer (TABLE II). Including ITS sequences of other spe-cies and testing the concordance of more than onegene genealogy might help to resolve the phylogenyof Thozetella in the future (Taylor et al 2000).
Species delimitation based on morphology.—Re-exami-nation of morphological characters following ITS se-quence analysis suggested that basic microawn mor-phology, such as L-shaped versus curved, microawnlength and basic conidiomatal configuration, such assporodochia versus synnemata, are key characters inspecies delimitation. Conidium and conidiophoremorphology and dimensions, which are usually im-portant characters in differentiating anamorphic fun-gi species, can play only a secondary role in Thozetelladue to their similarity among some species (TABLE
II). With one exception, each new species in our col-lection differs from the most similar, previously de-scribed taxa in at least one key criterion and usuallyby one or more secondary characteristics (TABLE II,Key). Thozetella boonjiensis does not differ from T.nivea by a key taxonomic character but by three oth-er characters, namely conidial length, conidial widthand finer microawn morphology (TABLE II, Key).

1086 MYCOLOGIA
Comparison of fruiting structures from cultures andfrom leaves.—One of the criticisms raised by Suttonand Cole (1983) in delimiting Thozetella species wasthat new species had been described from few spec-imens and in a number of instances were known onlyeither from culture or from natural substrata. Suttonand Cole (1983) reported that there was a consider-able increase in conidial size and a moderate increasein microawn length, as well as greater variation ofconidiomata morphology in cultures of T. effusa, ascompared to those developing on natural substrata.In contrast, Agnihothrudu (1958) and CastanedaRuiz (1984) observed minimal size variation in co-nidia and microawns of T. tocklaiensis and T. hava-nensis from natural substrata and from cultures.
In our study, cultures were available for all five spe-cies and for three of these specimens also could bestudied from natural substrata (TABLE I). A compar-ison of microawn and conidial dimensions revealedsome variation among specimens derived in vitro andfrom natural substrata, but these differences were notstatistically significant. Similarly, microawn length ap-peared to decrease in cultures of all species with in-creasing temperature and with dark treatment (datanot shown), but no statistical significance was detect-ed. In contrast, conidial length differed significantlyamong cultures grown under different conditions.This difference could be attributed to a marked in-crease in conidial length in cultures of T. giganteawhen grown at 25 C rather than at 20 C. Overall,microawn and conidial dimensions did not differ sig-nificantly on CMA and PCA. However, temperaturepreferences as observed by different growth rates(FIG. 3) and peaks in sporulation (TABLE IV) mightprovide additional taxonomic information.
The identification of many anamorphic genera(e.g., Cylindrocladium, Cylindrocladiella and Fusari-um) has been based almost solely on cultural char-acteristics under standard conditions (e.g., Joffe1986, Crous et al 1992, Crous and Wingfield 1994).The same approach has been suggested for Thozetella(Sutton and Cole 1983) and may be particularly use-ful if conidiomatal configuration is to be utilized asa taxonomic character. Specimens of the same Thoz-etella species have been observed to differ in the typeof conidiomata formed even when derived from thesame substratum (Castaneda Ruiz pers comm), butconidiomatal configuration remained basically stablein cultures of the five species investigated (TABLE IV).Although long-term stability was not evaluated, werecommend that Thozetella species be characterizedroutinely on CMA that includes a sterilized bananaleaf, after incubation at 20 C for 2 wk. This recom-mendation is based on the slow development of con-idiomata in some of the species and differs from oth-
er anamorphs where assessment usually is undertak-en after 7 d (e.g., Fisher et al 1982, Samuels andBrayford 1990, Crous et al 1992).
ACKNOWLEDGMENTS
We gratefully acknowledge these institutions for financingthis project: the Centre for Research of Fungal Diversity atthe Department of Ecology and Biodiversity; The Universityof Hong Kong; the Cooperative Research Centre for Rain-forest Ecology and Management; and the School of Tropi-cal Biology, James Cook University. Kylie McKenzie (TrinityAnglican School, Cairns) contributed to the assessment ofcultures under different growing conditions as part of a stu-dent research scheme. B.P. thanks N. Tucker for access toone of the collection sites, C. Pearce and I. Steer for theircompany on collection trips, H. Leung for providing sup-port with photography, M. Harrington for advice on phy-logenetic analysis and useful comments on the manuscriptand L. Jones and M. Pye for assistance with technical as-pects. We especially thank R. Castaneda Ruiz for sharing histaxonomic knowledge and experience. Two anonymous re-viewers are thanked for their valuable comments.
LITERATURE CITED
Agnihothrudu V. 1958. Notes on fungi from north-east In-dia. I. A new genus of Tuberculariaceae. Mycologia 50:570–579.
Bills GF, Polishook JD. 1994. Abundance and diversity ofmicrofungi in leaf litter of a lowland rain forest in Cos-ta Rica. Mycologia 86:187–198.
Booth C. 1971. Methods in microbiology. Vol. 4. London,UK: Academic Press. 795 p.
Castaneda Ruiz RF. 1984. Nuevos taxones de Deuteromy-cotina: Arnoldiella robusta gen. et sp.nov., Roigiella lig-nicola gen. et sp. nov., Sporidesmium pseudolmediaesp.nov., y Thozetella havanensis sp. nov. Revista del Jar-din Botanique Nacional, Universidad de Havana 5:57–87.
Castaneda Ruiz RF, Arnold GRW. 1985. Deuteromycotinade Cuba. I. Hyphomycetes. Revista del Jardin Bota-nique Nacional, Universidad de Havana 6:47–67.
Crous PW, Phillips AJW, Wingfield MJ. 1992. Effects of cul-tural conditions on vesicle and conidium morphologyin species of Cylindrocladium and Cylindrocladiella. My-cologia 84:497–504.
Crous PW, Wingfield MJ. 1994. A monograph of Cylindro-cladium, including anamorphs of Calonectria. Mycotax-on 51:341–435.
Felsenstein J. 1985. Confidence limits on phylogenies: anapproach using the bootstrap. Evolution 39:783–791.
Fisher NL, Burgess LW, Toussoun TA, Nelson PE. 1982. Car-nation leaves as a substrate and for preserving culturesof Fusarium species. Phytopathology 72:151–153.
Gams W, Hoekstra ES, Aptroot A. 1998. CBS Course of My-cology. 4th ed. Delft, The Netherlands: Centraalbureauvoor Schimmelcultures. 165 p.
Hibbett DS. 1992. Ribosomal RNA and fungal systematics.

1087PAULUS ET AL: FIVE NEW SPECIES OF THOZETELLA
Transactions of the Mycological Society of Japan 33:533–556.
Joffe AZ. 1986. Taxonomic problems of the genus Fusarium.In: Joffe, AZ, ed. Fusarium species: their biology andtoxicology. New York, USA: J. Wiley and Sons. 588 p.
Johnston A, Booth C. 1983. Plant pathologist’s pocketbook.2nd ed. Slough, England: Commonwealth AgriculturalBureau. 439 p.
Kirby JJH, Webster J. 1990. A particle plating method foranalysis of fungal community composition and struc-ture. Mycological Research 94:621–626.
Kirk PM. 2001. Ainsworth & Bisby’s dictionary of the fungi.9th ed. Wallingford, UK: C.A.B. International. 672 p.
Kuntze O. 1891. Revisio generum plantarum. A. Felix, Leip-zig, Germany. 1011 p.
Matsushima T. 1971. Microfungi of the Solomon Island andPapua New Guinea. Kobe, Japan: Matsushima FungusCollection. 78 p.
Mercado-Sierra A, Holubova-Jechova V, Mena Portales J.1997. Monografie XXIII. Hifomycetes demaciaceos deCuba. Enteroblasticos. Torino, Cuba: Museo RegionaleDi Scienze Naturali. p 352–354.
Nag Raj TR. 1976. Miscellaneous microfungi. I. CanadianJournal of Botany 54:1370–1376.
Paulus B, Gadek P, Hyde KD. 2003. Estimation of microfun-gal diversity in tropical rain forest leaf litter using par-ticle filtration: the effects of leaf storage and surfacetreatment. Mycological Research 107:748–756.
Pirozynski KA, Hodges CS. 1973. New Hyphomycetes fromSouth Carolina. Canadian Journal of Botany 51:157–173.
Reblova M, Winka K. 2000. Phylogeny of Chaetosphaeria andits anamorphs based on morphological and moleculardata. Mycologia 92:939–954.
Samuels GJ, Brayford D. 1990. Variation in Nectria radicicolaand its anamorph Cylindrocarpon destruens. Mycologi-cal Research 94:433–442.
Sivichai S, Jones EBG, Hywel-Jones N. 2002. Fungal coloni-sation of wood in a freshwater stream at Tad Ta Phu,Khao Yai National Park, Thailand. Fungal Diversity 10:113–129.
Smith DR, Stanosz GR. 1995. Confirmation of two distinctpopulations of Sphaeriopsis sapinae in the North Cen-tral United States using RAPDs. Phytopathology 85:699–704.
SPSS. 2001. Chicago, Illinois, USA: Lead Technologies Inc.Sutton B, Cole GT. 1983. Thozetella (hyphomycetes): an ex-
ercise in diversity. Transactions of the British Mycolog-ical Society 81:97–107.
Swofford DL. 2002. PAUP*. Phylogenetic analysis using par-simony (*and other methods). Sunderland, Massachu-setts, USA: Sinauer Associates.
Taylor JW, Jacobson DJ, Kroken S, Kasuga T, Geiser DM,Hibbett DS, Fisher MC. 2000. Phylogenetic species rec-ognition and species concepts in fungi. Fungal Genet-ics and Biology 31:21–32.
Waipara NW, Di Menna ME, Cole ALJ, Skipp RA. 1996.Characterisation of Thozetella tocklaiensis isolated fromthe roots of three grass species in Waikato pastures,New Zealand. New Zealand Journal of Botany 34:517–522.
White TJ, Bruns TD, Lee S, Taylor J. 1990. Amplificationand direct sequencing of fungal ribosomal DNA genesfor phylogenetics. In: Innis, MA, Gelfand, DH, Sninsky,J J and White, TJ, eds. PCR Protocols. San Diego, USA:Academic Press. p 315–322.
Yanna, Ho WH, Hyde KD. 2002. Fungal succession onfronds of Phoenix hanceana in Hong Kong. Fungal Di-versity 10:185–211.




















