
Micropaleontologic record of Quaternary paleoenvironments in the Central Albemarle Embayment, North...
23
Micropaleontologic record of Quaternary paleoenvironments in the Central Albemarle Embayment, North Carolina, U.S.A. Stephen J. Culver a, ⁎, Kathleen M. Farrell b , David J. Mallinson a , Debra A. Willard c , Benjamin P. Horton d , Stanley R. Riggs a , E. Robert Thieler e , John F. Wehmiller f , Peter Parham a , Scott W. Snyder a , Caroline Hillier g a Department of Geological Sciences, Graham Rm 101, East Carolina University, Greenville, NC 27858, USA b North Carolina Geological Survey, Raleigh Field Office and Core Repository, MSC 1620, Raleigh, NC 27699, USA c U.S. Geological Survey, 926A National Center, Reston, VA 20192, USA d Sea Level Research Laboratory, Department of Earth and Environmental Science, University of Pennsylvania, Philadelphia, PA 19104, USA e U.S. Geological Survey, Coastal and Marine Geology Program, 384 Woods Hole Road, Woods Hole, MA 02543, USA f Department of Geological Sciences, University of Delaware, Newark, DE 19716, USA g Argus Ecology Ltd., Annfield Plain, Durham DH97XN, UK abstract article info Article history: Received 14 September 2010 Received in revised form 8 March 2011 Accepted 11 March 2011 Available online 17 March 2011 Keywords: Foraminifera Diatoms Pollen Paleoenvironments Pleistocene Holocene To understand the temporal and spatial variation of eustatic sea-level fluctuations, glacio–hydro–isostacy, tectonics, subsidence, geologic environments and sedimentation patterns for the Quaternary of a passive continental margin, a nearly complete stratigraphic record that is fully integrated with a three dimensional chronostratigraphic framework, and paleoenvironmental information are necessary. The Albemarle Embayment, a Cenozoic regional depositional basin in eastern North Carolina located on the southeast Atlantic coast of the USA, is an ideal setting to unravel these dynamic, interrelated processes. Micropaleontological data, coupled with sedimentologic, chronostratigraphic and seismic data provide the bases for detailed interpretations of paleoenvironmental evolution and paleoclimates in the 90 m thick Quaternary record of the Albemarle Embayment. The data presented here come from a transect of cores drilled through a barrier island complex in the central Albemarle Embayment. This area sits in a ramp-like setting between late Pleistocene incised valleys. The data document the episodic infilling of the Albemarle Embayment throughout the Quaternary as a series of transgressive–regressive (T–R) cycles, characterized by inner shelf, midshelf, and shoreface assemblages, that overlie remnants of fluvial to estuarine valley-fill. Barrier island and marginal marine deposits have a low preservation potential. Inner to mid-shelf deposits of the early Pleistocene are overlain by similar middle Pleistocene shelf sediments in the south of the study area but entirely by inner shelf deposits in the north. Late Pleistocene marine sediments are of inner shelf origin and Holocene deposits are marginal marine in nature. Pleistocene marine sediments are incised, particularly in the northern half of the embayment by lowstand paleovalleys, partly filled by fluvial/floodplain deposits and in some cases, overlain by remnants of transgressive estuarine sediments. The shallowing through time of Quaternary sediments reflects the eastward progradational geometry of the continental shelf. The preservation potential of marginal marine deposits (barrier island, shoreface, backbarrier deposits) is not high, except in topographic lows associated with late Pleistocene paleovalleys and inlets because the current interglacial highstand has not yet reached its highest level. Given the documented increase in rate of relative sea-level rise in this region, shallow marine conditions are likely to return to the central Albemarle Embayment in the near future. © 2011 Elsevier B.V. All rights reserved. 1. Introduction The stratigraphic record in the Albemarle Embayment (Fig. 1A) is one of the thickest (ca. 90 m) and most complete Quaternary sections on the North American Atlantic margin (Riggs et al., 1992; Mallinson et al., 2005, 2010a). Foraminiferal, diatom and pollen data are valuable tools for reconstructing paleoenvironmental change in the late Pliocene to Holocene sedimentary record preserved in the Albemarle Embayment (Culver et al., 2006, 2007, 2008). A three-dimensional framework is provided by extensive seismic data that define the geometry of late Cenozoic to recent strata in the central and eastern part of the basin currently occupied by the Albemarle–Pamlico estuarine system (APES) and the Outer Banks (OBX) barrier islands (Mallinson et al., 2005, 2010a). A number of chronostratigraphic Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 227–249 ⁎ Corresponding author. Tel.: + 1 252 328 6360; fax: +1 252 328 4391. E-mail address: [email protected] (S.J. Culver). 0031-0182/$ – see front matter © 2011 Elsevier B.V. All rights reserved. doi:10.1016/j.palaeo.2011.03.004 Contents lists available at ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo
Transcript of Micropaleontologic record of Quaternary paleoenvironments in the Central Albemarle Embayment, North...

Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 227–249
Contents lists available at ScienceDirect
Palaeogeography, Palaeoclimatology, Palaeoecology
j ourna l homepage: www.e lsev ie r.com/ locate /pa laeo
Micropaleontologic record of Quaternary paleoenvironments in the CentralAlbemarle Embayment, North Carolina, U.S.A.
Stephen J. Culver a,⁎, Kathleen M. Farrell b, David J. Mallinson a, Debra A. Willard c, Benjamin P. Horton d,Stanley R. Riggs a, E. Robert Thieler e, John F. Wehmiller f, Peter Parham a, Scott W. Snyder a, Caroline Hillier g
a Department of Geological Sciences, Graham Rm 101, East Carolina University, Greenville, NC 27858, USAb North Carolina Geological Survey, Raleigh Field Office and Core Repository, MSC 1620, Raleigh, NC 27699, USAc U.S. Geological Survey, 926A National Center, Reston, VA 20192, USAd Sea Level Research Laboratory, Department of Earth and Environmental Science, University of Pennsylvania, Philadelphia, PA 19104, USAe U.S. Geological Survey, Coastal and Marine Geology Program, 384 Woods Hole Road, Woods Hole, MA 02543, USAf Department of Geological Sciences, University of Delaware, Newark, DE 19716, USAg Argus Ecology Ltd., Annfield Plain, Durham DH97XN, UK
⁎ Corresponding author. Tel.: +1 252 328 6360; fax:E-mail address: [email protected] (S.J. Culver).
0031-0182/$ – see front matter © 2011 Elsevier B.V. Aldoi:10.1016/j.palaeo.2011.03.004
a b s t r a c t
a r t i c l e i n f oArticle history:Received 14 September 2010Received in revised form 8 March 2011Accepted 11 March 2011Available online 17 March 2011
Keywords:ForaminiferaDiatomsPollenPaleoenvironmentsPleistoceneHolocene
To understand the temporal and spatial variation of eustatic sea-level fluctuations, glacio–hydro–isostacy,tectonics, subsidence, geologic environments and sedimentation patterns for the Quaternary of a passivecontinental margin, a nearly complete stratigraphic record that is fully integrated with a three dimensionalchronostratigraphic framework, and paleoenvironmental information are necessary. The AlbemarleEmbayment, a Cenozoic regional depositional basin in eastern North Carolina located on the southeastAtlantic coast of the USA, is an ideal setting to unravel these dynamic, interrelated processes.Micropaleontological data, coupled with sedimentologic, chronostratigraphic and seismic data provide thebases for detailed interpretations of paleoenvironmental evolution and paleoclimates in the 90 m thickQuaternary record of the Albemarle Embayment. The data presented here come from a transect of coresdrilled through a barrier island complex in the central Albemarle Embayment. This area sits in a ramp-likesetting between late Pleistocene incised valleys.The data document the episodic infilling of the Albemarle Embayment throughout the Quaternary as a seriesof transgressive–regressive (T–R) cycles, characterized by inner shelf, midshelf, and shoreface assemblages,that overlie remnants of fluvial to estuarine valley-fill. Barrier island andmarginal marine deposits have a lowpreservation potential. Inner to mid-shelf deposits of the early Pleistocene are overlain by similar middlePleistocene shelf sediments in the south of the study area but entirely by inner shelf deposits in the north. LatePleistocene marine sediments are of inner shelf origin and Holocene deposits are marginal marine in nature.Pleistocene marine sediments are incised, particularly in the northern half of the embayment by lowstandpaleovalleys, partly filled by fluvial/floodplain deposits and in some cases, overlain by remnants oftransgressive estuarine sediments. The shallowing through time of Quaternary sediments reflects theeastward progradational geometry of the continental shelf.The preservation potential of marginal marine deposits (barrier island, shoreface, backbarrier deposits) is nothigh, except in topographic lows associated with late Pleistocene paleovalleys and inlets because the currentinterglacial highstand has not yet reached its highest level. Given the documented increase in rate of relativesea-level rise in this region, shallow marine conditions are likely to return to the central AlbemarleEmbayment in the near future.
+1 252 328 4391.
l rights reserved.
© 2011 Elsevier B.V. All rights reserved.
1. Introduction
The stratigraphic record in the Albemarle Embayment (Fig. 1A) isone of the thickest (ca. 90 m) and most complete Quaternary sectionson the North American Atlantic margin (Riggs et al., 1992; Mallinson
et al., 2005, 2010a). Foraminiferal, diatom and pollen data are valuabletools for reconstructing paleoenvironmental change in the latePliocene to Holocene sedimentary record preserved in the AlbemarleEmbayment (Culver et al., 2006, 2007, 2008). A three-dimensionalframework is provided by extensive seismic data that define thegeometry of late Cenozoic to recent strata in the central and easternpart of the basin currently occupied by the Albemarle–Pamlicoestuarine system (APES) and the Outer Banks (OBX) barrier islands(Mallinson et al., 2005, 2010a). A number of chronostratigraphic
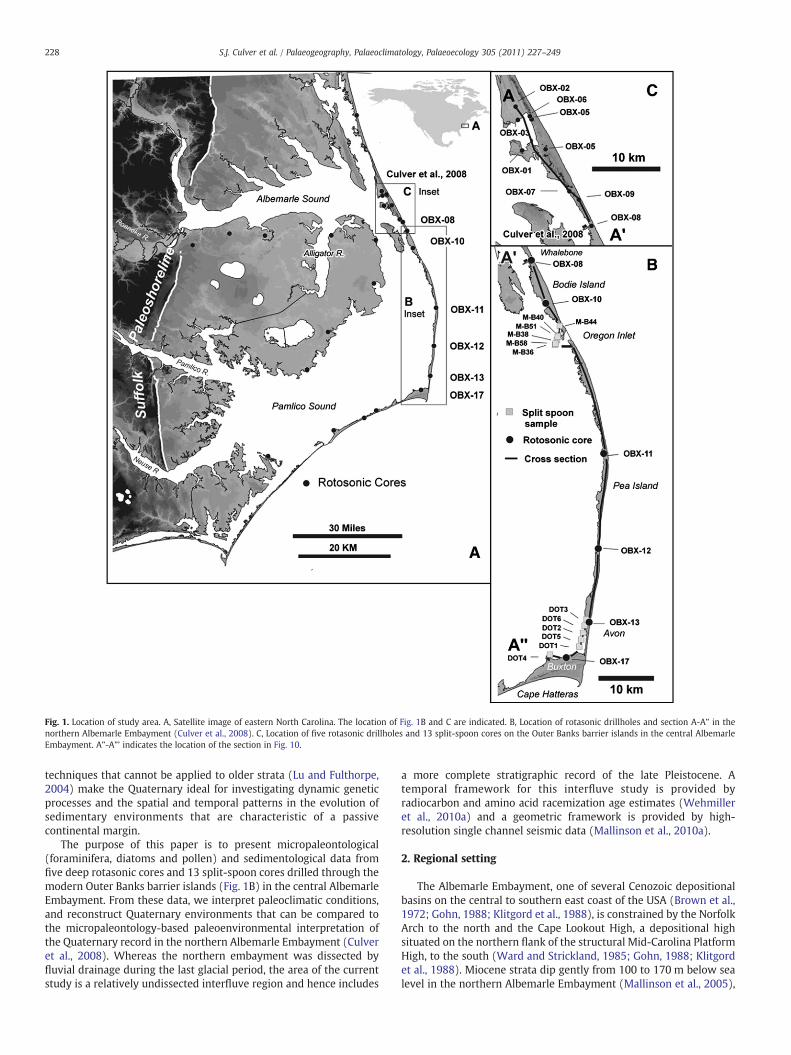
Fig. 1. Location of study area. A, Satellite image of eastern North Carolina. The location of Fig. 1B and C are indicated. B, Location of rotasonic drillholes and section A-A” in thenorthern Albemarle Embayment (Culver et al., 2008). C, Location of five rotasonic drillholes and 13 split-spoon cores on the Outer Banks barrier islands in the central AlbemarleEmbayment. A”-A”’ indicates the location of the section in Fig. 10.
228 S.J. Culver et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 227–249
techniques that cannot be applied to older strata (Lu and Fulthorpe,2004) make the Quaternary ideal for investigating dynamic geneticprocesses and the spatial and temporal patterns in the evolution ofsedimentary environments that are characteristic of a passivecontinental margin.
The purpose of this paper is to present micropaleontological(foraminifera, diatoms and pollen) and sedimentological data fromfive deep rotasonic cores and 13 split-spoon cores drilled through themodern Outer Banks barrier islands (Fig. 1B) in the central AlbemarleEmbayment. From these data, we interpret paleoclimatic conditions,and reconstruct Quaternary environments that can be compared tothe micropaleontology-based paleoenvironmental interpretation ofthe Quaternary record in the northern Albemarle Embayment (Culveret al., 2008). Whereas the northern embayment was dissected byfluvial drainage during the last glacial period, the area of the currentstudy is a relatively undissected interfluve region and hence includes
a more complete stratigraphic record of the late Pleistocene. Atemporal framework for this interfluve study is provided byradiocarbon and amino acid racemization age estimates (Wehmilleret al., 2010a) and a geometric framework is provided by high-resolution single channel seismic data (Mallinson et al., 2010a).
2. Regional setting
The Albemarle Embayment, one of several Cenozoic depositionalbasins on the central to southern east coast of the USA (Brown et al.,1972; Gohn, 1988; Klitgord et al., 1988), is constrained by the NorfolkArch to the north and the Cape Lookout High, a depositional highsituated on the northern flank of the structural Mid-Carolina PlatformHigh, to the south (Ward and Strickland, 1985; Gohn, 1988; Klitgordet al., 1988). Miocene strata dip gently from 100 to 170 m below sealevel in the northern Albemarle Embayment (Mallinson et al., 2005),

229S.J. Culver et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 227–249
and the overlying Pliocene unit is composed of at least seven,southward prograding seismic sequences. The overlying 90 m thickQuaternary section consists of up to 18 seismic sequences (Riggs et al.,1992) of mainly inner to mid-shelf sand and muddy sand. LatestPleistocene to Holocene sediments fill the paleo-Roanoke River valleythat is incised into the subhorizontal Pleistocene units (Sager andRiggs, 1998; Mallinson et al., 2005; Parham et al., 2007; Culver et al.,2008). The northern embayment is dissected by a large fluvialdrainage system incised during the last glacial maximum. In contrast,the present study area is relatively undissected and includes a morecomplete stratigraphic record for the later Pleistocene.
The late Pliocene to Pleistocene record is predominantly ofsubhorizontal, open marine deposits with minor brackish marineand non-marine units (Mallinson et al., 2005; Culver et al., 2008)infilling remnants of incised valleys beneath marine transgressivesurfaces. Pleistocene deposits were deeply incised during the lastglacial lowstand by the paleo-Roanoke, paleo-Neuse and paleo-Tardrainages (Riggs et al., 1992; Mallinson et al., 2005; Culver et al., 2007,2008). Subsequent valley-fill is composed of latest Pleistocene fluvialgravel and estuarine silt and mud overlain (depending on location) byearly to mid Holocene shallow marine and bay deposits and lateHolocene barrier island and/or estuarine sediments (Culver et al.,2007, 2008). In interfluve regions underlying the Outer Banks,Pleistocene deposits extend to within ca. 8 m of the surface (Riggset al., 1992; Mallinson et al., 2005). Along the mainland coast ofPamlico Sound, Pleistocene deposits occur near to the modern landsurface, overlain only by modern soil or various paralic deposits(Hoffman et al., 2007).
3. Materials and methods
Five near-continuous rotasonic cores (OBX-10, 11, 12, 13 and 17)were taken on the central Outer Banks (Fig. 1B; Table 1) to the southof cores OBX-01 to 09 (Mallinson et al., 2005; Culver et al., 2008). Coresites were selected based on geophysical data (Mallinson et al., 2005;2010a) to penetrate the entire Quaternary stratigraphic section in thecentral Albemarle Embayment in a transect extending from immedi-ately south of the paleo-Roanoke river valley southwards to CapeHatteras. Cores were logged (using a process-based method indepen-dent from composition and modified after Folk, 1980) and sampledfor grain-size analysis, radiocarbon and amino-acid racemization ageestimation, and microfossil study. Microfossil samples were approx-imately 3 cm-thick quarter-rounds of core taken above and belowcontacts and at varying intervals within thicker units.
Table 1Depth, elevation and location data for rotasonic and split-spoon cores. Depth is from land s
Core No. Latitude(N)
Longitude(W)
OBX-10 35.83517714 75.57228508OBX-11 35.59157226 75.46847005OBX-12 35.43891288 75.48587282OBX-13 35.32088928 75.50892826OBX-17 35.2652778 75.556355M-B33 35.756639 75.563528M-B36 35.767861 75.557111M-B38 35.775472 75.553194M-B40 35.783361 75.550389M-B44 35.797917 75.546556M-B51 35.780722 75.551306M-B58 35.76975 75.556DOT1 35.282222 75.53DOT2 35.304167 75.521667DOT3 35.325833 75.513889DOT4 35.270833 75.588056DOT5 35.293333 75.525833DOT6 35.315556 75.518056
Six discontinuous split-spoon cores (DOT 1–6) that providedbagged samples (10 cm of core at 1.5 m intervals) were taken by theNorth Carolina Department of Transportation. These cores provideadditional sedimentological, foraminiferal and temporal information,albeit within an incomplete stratigraphic record, between Avon (OBX-13) and Buxton (OBX-17) (Jomp et al., 2004; Fig. 1B; Table 1). Sevensimilar NC DOT cores (M-B33, 36, 38, 40, 44, 51, 58) providestratigraphic information for the considerable gap between OBX-10and 11 (Fig. 1B; Table 1).
3.1. Foraminifera
One hundred and twenty samples from five cores wereprocessed (using the protocol of Culver et al., 2008) and foraminif-era were concentrated using a sodium polytungstate floatingmethod (Munsterman and Kerstholt, 1996). Where possible, 100to 300 foraminifera were picked at random from aliquots or theentire N63 μ portion of each sample. Thirty-six samples were barrenof foraminifera.
Foraminifera were identified by comparison with illustrations inthe published literature (e.g., Culver and Buzas, 1980, and includedreferences) and identifications were confirmed by comparison withtype material in the Smithsonian Institution, Washington, D.C. andThe Natural History Museum, London.
The benthic foraminiferal data (available from S.J. Culver) wereanalyzed using cluster analysis (Mello and Buzas, 1968). The analysis,run in SYSTAT and utilizing Ward's linkage method and Euclidiandistances, included normalized abundance data of all taxa (64)constituting 2% or more of the assemblage in any one sample (toensure comparability with Culver et al., 2008). Eighty-four sampleswere clustered into four groups. The species composition of eachgroup was examined by calculating occurrence (O; the number oftimes a species occurred in a cluster group), constancy (C; thepercentage of samples in which the species occurred in each clustergroup), and biofacies fidelity (BF; a measure of species faithfulness toa cluster group). Fidelity and constancy are expressed in tens roundedto the nearest whole number (Hazel, 1977).
3.2. Diatoms
Fourteen samples for diatom analysis were prepared from coresOBX-10, 11 and 13 using the same sampling protocol as Culver et al.(2008) and followed the methodology of Zong and Horton (1999).Samples from OBX-11 proved to be barren of diatoms.
urface.
Core depth(m)
Elevation(NAVD 88) (m)
Elevation(msl) (m)
68.6 2.568.6 122.9 1.6119.8 1.3357.9 0.6946.4 −0.646.2 −0.275.8 −3.546.4 −0.544.4 0.7677.4 −1.4675.6 −1.750 −1.151.4 −1.251 1.947.8 1.450.5 −0.750.6 −0.7

230 S.J. Culver et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 227–249
Valves were identified by reference to Hendey (1964) and Van derWerff and Huls (1958–1966) and their ecological classification wasbased on Hudstedt (1953, 1957), Denys (1991–1992), Vos and deWolf (1993); polyhalobous, fully marine; mesohalobous, brackishwater; oligohalobous–halophilous, brackish–fresh; oligohalobousindifferent, fresh–brackish; halophobous, fresh water. The diatomdata for each core (available from B.P. Horton) were run separatelythrough a CONISS cluster analysis (Grimm, 1992) to determinesample groupings within cores OBX-10.
3.3. Pollen
Following the sampling protocol of Culver et al. (2008), pollen wasisolated from 40 samples taken from cores OBX-10 to 13 usingstandard palynological techniques (Traverse, 1988; Willard et al.,2003); at least 300 pollen grains per sample were counted todetermine percent abundance and pollen concentration. Data arelodged in the North American Pollen Database (NAPD); http://www.ngdc.noaa.gov/paleo/pollen.html. Pollen data, compared to a databaseof pollen from marine and estuarine surface samples collected inMississippi Sound (Edwards and Willard, 2001), Chesapeake Bay(Willard et al., 2003), the western Atlantic Ocean (Litwin and Andrie,1992; Willard et al., 2005), and the eastern Canada Shelf (Mudie,1980, 1982) using the modern analogue technique and squared chorddistance (SCD) measure (Overpeck et al., 1985), were considered tobe analogous to modern samples when they had SCD values ≤0.15.Samples in OBX-10 were run through a CONISS cluster analysis(Grimm, 1992) to define pollen zones.
3.4. Amino acid racemization age estimates
Seventy-eight mollusc samples (Mercenaria, Chione,Mulinia, Ensis,and Spisula) from OBX-10, 11, 12, 13, and 17 (Table 2) were analyzedfor amino acid racemization (AAR) using gas chromatographic (GC)methods outlined in Wehmiller and Miller (2000) or reverse phaseliquid chromatographic (RPLC) methods described in Kaufman andManley (1998). The latter method is particularly well suited for theanalysis of small Mulinia valves, which are common in several of thecores. Although Mercenaria specimens were the first priority foranalysis (Wehmiller et al., 2010a), specimens of this taxon were notfound throughout all the cores, so Mulinia results were used to fillsome significant data gaps in selected cores.
Three aminozones at Stetson Pit, NC were originally observed byYork et al. (1989) and later defined as AZ2, AZ3, and AZ4, in order ofincreasing stratigraphic age. These zones were based on D-alloisoleu-cine/L-isoleucine values in Mercenaria and Mulinia specimens andwere interpreted as late, middle, and early Pleistocene (~100 ka,~325–500 ka, and ~750–1200 ka, respectively). Subsequent modifi-cations of these age estimates (Riggs et al., 1992; Wehmiller et al.,2010a) and the addition of new aminozones are the consequence ofboth new results and refined kinetic modeling. Eight statisticallydistinct Mercenaria-based aminozones, defined on the basis of valineand leucine D/L values, are now recognized: AZ2, AZ2+, AZ3−, AZ3+,AZ3.5, AZ4−, AZ4, AZ4+ (Table 3). The younger and older ageestimates for aminozones (Table 3) are derived from a parabolic and anon-linear kinetic model, respectively, both of which have beenevaluated using stratigraphic, theoretical, and independent calibra-tion using Sr-isotope analysis (Wehmiller et al., 2010a,b). Althoughthe parabolic model has some value in certain applications, it is notconsidered a useful model for the AAR results from the OBX samplesdiscussed here. Consequently, all further discussion of AAR ageestimates includes only the non-linear model ages and theiruncertainties, as shown in Table 3.
Clusters of AG (the mean of D/L aspartic and glutamic acid D/Lvalues) measured inMulinia are defined as Groups A, B, and C, broadlyequivalent to AZ2, AZ3, and AZ4, respectively. In several cases each of
the Mulinia groups can be divided into smaller units: A1, A2, B1, B2,B3, and C1 and C2, in increasing stratigraphic order, respectively(Wehmiller et al., 2010a). In most cases whole Mulinia valves weresampled for analysis, but only rarely was more than 40% of a singleMercenaria valve available, and in many cases multiple Mercenariafragments at a single depth may have represented a single valve. Eachtaxon has advantages for this study. The robust Mercenaria valves arepotentially more resistant to diagenetic alteration of interior shelllayers used for analysis, but the shells themselves have greaterpotential to survive age-mixing processes. The thin-shelled Muliniamay bemore vulnerable to alteration, but their fragile nature suggeststhat they are more likely to represent the age of their confining unit.Age estimates for sedimentary units are based on the AAR results incombinationwith taphonomic and lithostratigraphic information, andwith several examples of older material being reworked into youngerbeing recognized.
3.5. Radiocarbon age estimates
Eleven radiocarbon age estimates were obtained for wood, plantfragments (stems, leaves, bark), and mollusks (Table 4). Inorganiccarbon sampleswere treatedwith phosphoric acid and organic carbonsamples were washed in distilled water and pretreated with a seriesof acid–base–acid leaches to remove inorganic carbon and mobilehumic/fluvic phases. Radiocarbon ages of samples (analyzed at theNational Ocean Sciences Accelerator Mass Spectrometry [NOSAMS]facility) were calibrated using the Marine 04 dataset (Hughen et al.,2004) for shells (with no local reservoir correction) and the IntCal04dataset (Reimer et al., 2004) for plant material.
4. Results and paleoenvironmental interpretations
After describing the general patterns of foraminifera in the fiverotasonic cores, followed by discussion of the results of the clusteranalysis, paleoenvironmental change through time for each core isdescribed and illustrated below from north to south (OBX-10, OBX-11,OBX-12, OBX-13, OBX-17).
4.1. Foraminiferal patterns
Figs. 2, 5, 6, 7, 8 show that the uppermost portions (ca. 5 m) of allOBX cores lack foraminifera. Below the barren sections, foraminiferalabundance is variable (e.g., OBX-11; Fig. 5). Species richness (numberof species in a sample) is a reflection of number of specimens pickedbut it is also generally highest when the calculated number ofspecimens per unit weight of sediment is highest (e.g., OBX-17;Fig. 8). Planktonic foraminifera are generally rare and scattered(Figs. 2, 5–8) but reach 10% of foraminiferal assemblages in sampleswhere specimen abundance and species richness are high (e.g., OBX-17; Fig. 8).
Cluster analysis of benthic foraminiferal data defined two majorgroups of samples containing several nested subgroups (Fig. 9). Fourgroups (A to D) were determined to be paleoenvironmentallymeaningful; the group membership of each sample containingforaminifera is indicated on the core logs (Figs. 2, 5–8). A north–south geologic section showing correlations between cores based onmicrofossils (foraminifera, diatoms, pollen), lithology, seismic dataand gamma logs, and paleoenvironmental interpretations is shown inFig. 10. A second cluster analysis was run that, in addition to OBX-10to 13 foraminiferal data, also included data from the southernmostcore (OBX-08) from the Culver et al. (2008) study. This aidedcorrelations to the northern Albemarle Embayment set of cores(Fig. 10).
Occurrence, constancy and biofacies fidelity were calculated todetermine the species responsible for the clustering of samples incores OBX-10, 11, 12, 13 and 17 (Table 5). Thirteen taxa are unique to

Table 2Amino acid racemization data for the OBX cores. VL is the average of valine and leucine D/L values. Aminozones (AZ2, AZ2+, etc.) are based on VL values inMercenaria. Groups (A, A1,A2, etc.) are based on AG values inMulinia (Wehmiller et al., 2010a). In a few cases, aminozones or groups are assigned based on results for taxa other thanMercenaria orMulinia. Ageestimates for aminozones are given in Table 3.
UDAMS Locality ID Sampled interval, feet Interval top, meters Sample ID Sample type SubsampleID A-Zone(Merc VL)
A-Group(Mul AG)
07530 OBX-10 29.7 9.05 JW2003-026-002 Mercenaria 2004024 H07530 OBX-10 65.6 20 JW2003-031-1 Mercenaria 2004025 AZ2+07530 OBX-10 65.6 20 JW2003-031-2 Mercenaria 2004026 AZ2+07530 OBX-10 65.75 20.05 JW2003-032 Mercenaria 2004027 AZ2+07530 OBX-10 66 20.12 JW2003-033 Mercenaria 2005498 AZ2+07530 OBX-10 73.95–74.1 22.55 JW2003-038 Mercenaria 2004028 AZ2+07530 OBX-10 122.5–122.7 37.35 JW2003-053-002 Mercenaria 2005499 AZ3+07530 OBX-10 127.8–128.4 38.96 JW2003-055 Mercenaria 2005500 AZ3+07531 OBX-11 28.5–29.1 8.69 JW2003-118-1 Mercenaria 2004019 H07531 OBX-11 28.5–29.1 8.69 JW2003-118-2 Mercenaria 2004020 H07531 OBX-11 32.0–32.9 9.76 JW2003-119-1 Mercenaria 2004010 H07531 OBX-11 32.0–32.9 9.76 JW2003-119-6 Mercenaria 2004011 H07531 OBX-11 73.8 22.5 JW2003-131-001 Mercenaria 2003169 M AZ3-07531 OBX-11 73.8 22.5 JW2003-131-002 Mercenaria 2003170 M AZ3-07531 OBX-11 79.65–79.85 24.28 JW2003-134-1 Mulinia 2005295 A2? b
07531 OBX-11 79.65–79.85 24.28 JW2003-134-2 Mulinia 2005296 A2? b
07531 OBX-11 84.5–84.75 25.76 JW2003-137-3 Ensis 2004031 A2? a b c
07531 OBX-11 84.5–84.75 25.76 JW2003-137-4 Ensis 2004032 A2? a b c
07531 OBX-11 88.7–89.2 27.05 JW2003-142-001 Mulinia 2007346 A207531 OBX-11 88.7–89.2 27.05 JW2003-142-002 Mulinia 2007347 A207531 OBX-11 88.7–89.2 27.05 JW2003-142-003 Mulinia 2007348 A207531 OBX-11 88.7–89.2 27.05 JW2003-142-004 Mulinia 2007349 A207531 OBX-11 90.4–90.7 27.56 JW2003-143-001 Mercenaria 2003171 AZ3−07531 OBX-11 103.2–103.4 31.46 JW2003-146-001 Mulinia 2007350 B107531 OBX-11 103.2–103.4 31.46 JW2003-146-002 Mulinia 2007351 B107531 OBX-11 103.2–103.4 31.46 JW2003-146-003 Mulinia 2007352 B107531 OBX-11 168.3–169.15 51.31 JW2003-150-003 Mercenaria? 2006038 AZ4−?07531 OBX-11 168.3–169.15 51.52 JW2003-150 Ensis 2007353 C c
07531 OBX-11 177.7–178.3 54.17 JW2003-152-001 Mercenaria 2005342 AZ4+07531 OBX-11 177.7–178.3 54.17 JW2003-152-002 Mercenaria 2003149 AZ4+07531 OBX-11 177.7–178.3 54.17 JW2003-152 Mercenaria 2007354 AZ4+07532 OBX-12 24.2 7.38 JW2003-073-001 Chione 2003168 M H07532 OBX-12 58.8–59.2 17.93 JW2003-084-003 Mercenaria 2006040 AZ207532 OBX-12 67.15–67.5 20.47 JW2003-089-001 Mulinia 2006041 A07532 OBX-12 67.15–67.5 20.47 JW2003-089-002 Mulinia 2006042 A07533 OBX-13 38.8 11.83 JW2003-103-001 Mercenaria 2006102 AZ2+07533 OBX-13 38.8 11.83 JW2003-103-002 Mulinia 2005340 A07533 OBX-13 53.2 16.22 JW2003-105-001 Mercenaria 2006103 AZ2+07533 OBX-13 53.2 16.22 JW2003-105-002 Mulinia 2006104 A07533 OBX-13 53.85 16.42 JW2003-106 Mercenaria 2006105 AZ2+07533 OBX-13 54.1 16.49 JW2003-107 Mercenaria 2004070 AZ2+07533 OBX-13 62.4 19.02 JW2003-113-001 Mercenaria 2003145 AZ2+07533 OBX-13 62.4 19.02 JW2003-113-002 Mercenaria 2003146 AZ2+07533 OBX-13 62.4 19.02 JW2003-113-003 Mercenaria 2003147 AZ2+07547 OBX-17 50.1 15.27 JW2004-027-001 Mulinia 2007356 A107547 OBX-17 50.1 15.27 JW2004-027-002 Mulinia 2007357 A107547 OBX-17 50.1 15.27 JW2004-027-003 Mulinia 2008079 A107547 OBX-17 50.1 15.27 JW2004-027-004 Mulinia 2008080 A107547 OBX-17 50.1 15.27 JW2004-027-005 Spisula 2008081 A1 a
07547 OBX-17 50.1 15.27 JW2004-027-006 Mercenaria 2008082 AZ207547 OBX-17 61.3 18.68 JW2004-028-001 Mulinia 2008083 A107547 OBX-17 61.3 18.68 JW2004-028-002 Mulinia 2008084 A107547 OBX-17 61.3 18.68 JW2004-028-003 Mulinia 2008085 A107547 OBX-17 61.3 18.68 JW2004-028 S Spisula 2008086 A1 c
07547 OBX-17 61.3 18.68 JW2004-028 M Mercenaria 2008087 AZ207547 OBX-17 67 20.42 JW2004-029 Spisula A1 c
07547 OBX-17 67.7 20.64 JW2004-030 Mulinia 2008088 A107547 OBX-17 69.6 21.22 JW2004-031-001 Mercenaria 2005327 AZ207547 OBX-17 69.6 21.22 JW2004-031-002 Mercenaria 2005328 AZ207547 OBX-17 69.6 21.22 JW2004-031-003 Mercenaria 2005343 AZ207547 OBX-17 70 21.34 JW2004-032-001 Mercenaria 2004207 AZ207547 OBX-17 70 21.34 JW2004-032-002 Mercenaria 2006106 AZ207547 OBX-17 70 21.34 JW2004-032-003 Mercenaria 2006107 AZ207547 OBX-17 79.2 24.15 JW2004-038-001 Ensis 2007358 A2 c
07547 OBX-17 79.2 24.15 JW2004-038-002 Mulinia 2007359 A207547 OBX-17 79.2 24.15 JW2004-038-003 Mulinia 2007360 A207547 OBX-17 79.2 24.15 JW2004-038-004 Mulinia 2007361 A207547 OBX-17 81.55 24.86 JW2004-039-001 Mulinia 2008089 A207547 OBX-17 81.55 24.86 JW2004-039-002 Mulinia 2008090 A207547 OBX-17 81.55 24.86 JW2004-039-003 Mulinia 2008091 A207547 OBX-17 81.55 24.86 JW2004-039-004 Mulinia 2008092 A2
(continued on next page)
231S.J. Culver et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 227–249

Table 2 (continued)
UDAMS Locality ID Sampled interval, feet Interval top, meters Sample ID Sample type SubsampleID A-Zone(Merc VL)
A-Group(Mul AG)
07547 OBX-17 105.05 32.03 JW2004-050-001 Mulinia 2007366 B107547 OBX-17 105.05 32.03 JW2004-050-002 Mulinia 2007367 B107547 OBX-17 105.05 32.03 JW2004-050-003 Mulinia 2007368 B107547 OBX-17 105.05 32.03 JW2004-050-004 Mulinia 2007369 B107547 OBX-17 136.4 41.58 JW2004-054-001 Mulinia 2007370 C107547 OBX-17 136.4 41.58 JW2004-054-002 Mulinia 2007371 C107547 OBX-17 136.4 41.58 JW2004-054-003 Mulinia 2007372 C107547 OBX-17 164.8 50.23 JW2004-059-002 Mulinia 2007377 C207547 OBX-17 164.8 50.23 JW2004-059-003 Mulinia 2007378 C2
a) Analytical results were evaluated in context of sedimentology and stratigraphy to determine if samples were reworked.b) Data obtained by GC. In all probability, these are A2 but the data were obtained by a different method than that used for the A-group designations.c) A-group designations are formally based onMulinia. Ensis and Spisula have racemization rates that are quite similar toMulinia, hence we can substitute Ensis or Spisula forMuliniawhen necessary.
232 S.J. Culver et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 227–249
a group (BF=10). Most have low occurrences (occur in few samples)except for Hopkinsina atlantica and Stainforthia complanata, whichoccur in approximately half of the samples of group C (Table 5). Fewspecies have both high BF and C values; therefore, those taxa in groupswith BF and C values of 4 or more were arbitrarily defined as“characteristic” of that group (Table 5). Elphidium excavatum occurs inall samples in this study (C=10 in every group) and Ammoniaparkinsoniana is almost as ubiquitous (C ranging from 7 to 10). Theirlow BF scores mean that these species are not included as“characteristic” species for any group. Cluster group A has relativelylow species richness (it contains 14 taxa) (Table 5). In contrast, clustergroups B, C and D contain at least three times as many taxawith 42, 55and 50 taxa respectively. Cluster group B has high dominance (onlyone characteristic species, in addition to the generally ubiquitousElphidium excavatum and Ammonia parkinsoniana), whereas clustergroups C and D have 17 and 14 characteristic species respectively and,thus, exhibit less dominance. The number of species comprisinggroups C and D (55 and 50) is considerably higher than the number ofspecies comprising any of the five cluster groups (4, 35, 31, 34, 35, and1 taxa, respectively) in the northern Albemarle Embayment (Culveret al., 2008).
4.2. Paleoenvironmental interpretations of foraminiferal assemblages
Paleoenvironmental interpretations of the foraminiferal assem-blages comprising cluster groups A to D are based on moderndistributional data (e.g., Grossman and Benson, 1967; Schnitker, 1971;Ellison and Nichols, 1976; Poag et al., 1980; Scott and Medioli, 1980;
Table 3Summary of Mercenaria leucine age estimates for AAR zones derived from alternativekinetic models, and corresponding Mulinia subgroups (modified from Wehmiller et al.,2010a). Age ranges quoted for each aminozone or subgroup are derived from theextrapolated range of D/L values for calibration samples.
Age range, ka (rounded to nearest 10 ka)
Aminozone Parabolicd Non-lineard Mulinia subgroupc
AZ2 80 kaa 80 kaa A1AZ2+ 125 kab 125 kab A2AZ3- 130–160 170–220 B1/B2AZ3+ 200–240 340–430 B2AZ3.5 280–340 620–770 B2/B3AZ4– 320–380 770–980 B3/C1AZ4 390–480 1130–1450 C1AZ4+ 450–540 1440–1840 C2
d Parabolic and Non-Linear Ages are based on Fig. 13 in Wehmiller et al., 2010a.c Corresponding Mulinia subgroups based on intergeneric relations (see Wehmiller
et al., 2010a).a AZ2 assigned an age of 80 ka based on local calibration.b AZ2+ appears to be early MIS5 based on AZ2 calibration and local stratigraphic
relations.
Workman, 1981; Culver and Snedden, 1996; Culver et al., 1996; Wooet al., 1997; Abbene et al., 2006; Culver et al., 2006; Vance et al., 2006;Horton and Culver, 2008; Kemp et al., 2009) and are quite similar to theinterpretations documented in OBX cores in the northern AlbemarleEmbayment (Culver et al., 2008).
The foraminiferal assemblages in the seven samples comprisingcluster group A are dominated by Elphidium excavatum and Ammoniaparkinsoniana (Table 5). These seven samples do not, however, forman ecologically coherent group. OBX-10 5.94 m (Fig. 2) contains a fewspecimens of typical salt-marsh species, Haplophragmoides wilberti,Trochammina inflata and Tiphotrocha comprimata (Culver and Horton,2005; Horton and Culver, 2008; Kemp et al., 2009a) and this sample isinterpreted as representing a back-barrier estuarine environmentadjacent to a marsh. Three additional samples, two in OBX-11 and oneOBX-13 (Figs. 5 and 7), are also interpreted as estuarine. A secondsample from OBX-13 is considered to be reworked. Two deep samplesin OBX-10 (at 41.18 and 56.24 m; Fig. 2) occur adjacent to typicalopen shelf assemblages and OBX-10 56.24 m contains a few speci-mens of planktonic foraminifera.
Cluster group B comprises 31 samples that tend to occur in cleansands in the shallower portions of all OBX cores (Figs. 2, 5–8). Thesesamples are strongly dominated by Elphidium excavatum and are oflow to moderate species richness. Planktonic foraminifera are rare.One species, Elphidium subarcticum, is characteristic of this assem-blage (Table 5). This species can occur in high salinity estuaries (e.g.,Abbene et al., 2006) but it is more typically an inner shelf species(Culver and Snedden, 1996). Other relatively common taxa in group B(e.g., Buccella frigida, Elphidium mexicanum, Hanzawaia strattoni,Nonionella atlantica, Quinqueloculina seminula and Textularia cf.T. gramen) are also typical on the modern inner shelf off NorthCarolina (Schnitker, 1971; Workman, 1981; Stanton, 2008). Thisshallow inner shelf assemblage is comparable to an assemblagerecorded in cores OBX-1 to 9 in the northern Albemarle Embayment(group D in Culver et al., 2008).
The 19 samples of group C contain diverse assemblages (20 to 30species) comparable to those of cluster analysis groups Bs and Bm inthe northern Albemarle Embayment (Culver et al., 2008). They arerestricted to fine sands and muds occurring in the lower parts of thethree deep cores (OBX-10, 11 and 17); foraminifera of this assemblageare generally small (b100 μ) and abundant (Figs. 2, 5, 8). Elphidiumexcavatum occurs in all samples and Ammonia parkinsoniana in 16 of19, but 17 additional taxa are characteristic of group C (Table 5).Several of these (e.g., Buliminella elegantissima, Cassidulina reniforme,Fursenkoina fusiformis and Stainforthia complanata) are species thatcommonly occur in reduced oxygen environments (Poag, 1985;Snyder et al., 1988; Sen Gupta and Machain-Castillo, 1993).Stainforthia complanata, Fursenkoina fusiformis, Hopkinsina atlanticaand Bolivina lowmani are generally found today at middle shelf depths(Culver and Buzas, 1980; Poag et al., 1980; Culver and Snedden, 1996).The presence of planktonic foraminifera (Figs. 2, 5, 8), sometimes

Table4
AMS
14Cag
eestimates
forOBX
corescalib
ratedus
ingCA
LIB5
.0(Stuiver
andRe
imer,1
993)
.Med
ianprob
ability
ageestimates
areplottedon
thefigu
res.NAVD88
is~15
cmab
ovemea
nsealeve
linthestud
yarea
.
SampleID
Lab
Num
ber
Material
Dep
th(m
)in
Core
Dep
th(m
)NAVD88
Age
(C-14yr
BP)
Error
(yr)
δC-13
2-σAge
Rang
eRe
lative
Prob
ability
Med
ianProb
ability
Age
(yr)
Calib
ration
Data
OBX
-10-24
.2OS-41
166
Crassostreavirginica
7.38
−4.88
2840
40−
1.68
2457
2716
126
03Hug
henet
al.,20
04OBX
-10-89
.5OS-41
178
Plan
tfrag
men
ts27
.28
−24
.78
N48
,000
−26
.29
––
––
−OBX
-12-58
-59.2
OS-63
816
Mercena
riamercena
ria
17.86
−16
.25
43,200
480
0.41
––
––
–
OBX
-12-24
.2OS-41
112
Chione
7.38
−5.77
3080
350.86
2753
2947
128
47Hug
henet
al.,20
04OBX
-13-43
OS-41
111
Ana
dara
13.11
−11
.78
4910
40−
0.31
5044
5327
152
41Hug
henet
al.,20
04OBX
-13-44
.8-A
1OS-44
292
Ano
mia
simplex
13.66
−12
.33
4870
35−
0.25
5045
5285
151
83Hug
henet
al.,20
04OBX
-13-44
.8-O
1OS-44
293
Crassostreavirginica
13.66
−12
.33
4880
351.87
5052
5292
151
99Hug
henet
al.,20
04OBX
-13-44
.8-O
2OS-44
294
Crassostreavirginica
13.66
−12
.33
5810
30−
2.53
6169
6292
162
34Hug
henet
al.,20
04OBX
-13-47
.0-48.7
OS-44
218
Woo
d14
.58
−13
.25
8950
45−
24.63
9916
10,093
0.56
10,063
Reim
eret
al.,20
0410
,113
10,222
0.44
OBX
-17-50
.1OS-67
832
Mulinia
lateralis
15.27
−14
.58
31,700
230
1.54
––
––
–
OBX
-17-67
.0OS-45
327
Spisulasolid
issima(?)
20.42
−19
.73
28,700
160
0.7
––
––
–
BUX-D
OT1
-36.6-38
.6Be
ta-199
577
Peat
11to
12−
12.1
to−13
.111
,600
40−
26.5
13,320
13,584
113
,432
Reim
eret
al.,20
04
233S.J. Culver et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 227–249
more than 10% (Fig. 8), in this group indicates inner to middle shelfenvironments (Grimsdale and Morkhoven, 1955; Murray, 1976). Insummary, assemblage C accumulated in relatively low energyconditions, with, at times, reduced oxygen, at inner to mid-shelfdepths.
The samples comprising cluster groupD are from clean sands, oftenwithmollusc shell fragments, at variable depths in OBX-11, 12, 13 and17 (Figs. 5–8). Elphidium excavatum and Ammonia parkinsonianadominatemoderately diverse assemblages (Table 5) but 14 additionalspecies are considered to be characteristic of group D (Table 5). All ofthese are typical inner shelf taxa (e.g., Schnitker, 1971; Workman,1981; Culver and Snedden, 1996; Stanton, 2008). Five of the 14characteristic taxa are species of Quinqueloculina, which can occur atbarrier island inlets (e.g., Abbene et al., 2006; Vance et al., 2006), butwhich are also indicative of normal salinity shelf conditions (Culverand Buzas, 1980). Planktonic foraminifera, indicating open inner shelfwaters (Grimsdale andMorkhoven, 1955; Murray, 1976), can occur ingroupD (Figs. 3–6) but never as abundantly as in group C. In summary,like groupB, the assemblages comprising groupD are typical of normalsalinity, shallow shelf environments that characterize the inner shelfoff North Carolina today (Schnitker, 1971). Group D of this study iscomparable to group D in the northern Albemarle Embayment OBXcores (Culver et al., 2008).
4.3. OBX-10
4.3.1. Age estimatesA Crassostrea virginica valve from 7.38 m core depth has a 2-sigma
age range of 2457 to 2716 cal yr B.P. (Table 4). Plant fragments from27.28 m core depth are radiocarbon dead (Table 4; Fig. 2).Mercenariasamples from 20 to 22.55 m core depth (Fig. 2) belong to aminozoneAZ2+, with an age estimate of 125 ka (Tables 2, 3; Wehmiller et al.,2010a). Two deeper samples from ~38 m (Fig. 2) representaminozone AZ3+, 300 to 430 ka (Tables 2, 3; Wehmiller et al.,2010a). Geophysical data indicate that the Pliocene is approximately10 m below the base of this core (Fig. 10; Mallinson et al., 2010a).Although the three lowest pollen samples in the core (57.6 to 60.5 m)contain small amounts of Pterocarya pollen (Fig. 3), indicative of aPliocene age, the geophysical data indicate likely reworking of thepollen.
4.3.2. Paleoenvironmental interpretationsThe basal approximately 10 m of OBX-10 is barren of foraminifera
and this generally sandy section is interpreted as fluvial in origin. From62 to 55 m alternating sand and mud layers contain foraminifera(assemblages C andA) anddiatoms (Figs. 2, 4) indicative of open/innerto mid shelf environments. Pollen assemblages (pollen zone 5) in thiscore segment (Fig. 3) have no modern analogs, but the relatively highabundance of Fagus (beech) pollen indicates cooler than modernconditions. The overlying sand (up to 50.5 m) contains lowerabundance inner shelf foraminiferal assemblage B (Fig. 2).
From 50.5 to 46 m, a burrowed mud contains foraminiferalassemblage C (Fig. 2) and is dominated by Raphoneis amphiceros, apolyhalobous taxon (Fig. 4). Pollen assemblages (pollen zone 4) inthis mud (Figs. 2, 3) have greater abundance of Pinus pollen than inpollen zone 5, but Fagus pollen is still abundant. We interpret thismud to have been deposited in low energy, reduced oxygenconditions at inner to mid-shelf depths under cooler than moderntemperatures.
A laminated mud, barren of foraminifera, overlies the burrowedmud from 46 to 43 m. The greater abundance of Picea (spruce), Abies(fir), and Tsuga (hemlock) and decreased abundance of Fagus pollenin this zone indicates further cooling. The lower of two samples in thelaminated mud contains abundant Cyclotella striata (a mesohalobousdiatom species) and the oligohalobous-indifferent (i.e., fresh–brack-ish water) Navicula pusilla (Figs. 2, 4). The laminated mud is

234 S.J. Culver et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 227–249

Fig. 2. Lithologic log of core OBX-10 indicating sample depths, number of benthic foraminiferal species per sample, number of specimens picked per sample, percent planktonics,calculated number of specimens in 100 g dry weight of sediment, percent polyhalobous (P), mesohalobous (M), and oligohalobous (O) diatoms, pollen zones 1 to 5. b, barren offoraminifera. Horizontal fine dashed lines indicate level of AAR and median probability 14C age estimates. Shading indicates the Holocene section.
235S.J. Culver et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 227–249
interpreted as being deposited in a low energy estuarine setting undercolder than modern conditions.
From 43 to 33.5 m, sands are characterized by shallow inner shelfforaminiferal assemblage B. Up-core increase in numbers of forami-nifera per unit weight of core and the presence of planktonicspecimens where benthic foraminifera are most abundant (Fig. 2)indicates up-core increased depth of water and/or distance fromshore.
The overlying sandy section from 33.5 to 26 m is barren offoraminifera and is interpreted as fluvial in origin. Near the top ofthis section, two samples from a radiocarbon dead freshwater peat arecharacterized by dominant Pinus pollen, abundant fern spores, andcommon Piceapollen (pollen zone 3; Fig. 3). There is nomodern analogbut the abundance of Picea and Abies pollen in the lower sample issuggestive of colder than modern conditions. The sand immediatelyoverlying the freshwater peat contains Thalassinoides-type burrowsand is interpreted as estuarine in origin. Pinus and Quercus pollendominate assemblages in the two samples from this sand (pollen zone2; Figs. 2, 3). Fagus pollen is common, and Picea and Abies pollen arerare, indicating warmer conditions than in the underlying peat.
The overlying sand and shelly gravel from 26 to 20 m contains theshallow inner shelf foraminiferal assemblage B. Consistent with anearshore setting, planktonic specimens are absent.
From 20 to 12 m, a coarsening upwards sequence of sand isgenerally barren of foraminifera except for two samples (Fig. 2) thatcontain the shallow inner shelf assemblage B. Five pollen sampleswithin this section (pollen zone 1; Figs. 2, 3) are dominated by Pinuspollen (50–60%). Picea and Quercus are subdominant, Fagus is absentand herbaceous pollen and fern spores are common. There are nomodern analogues to these assemblages but the high abundance ofPicea suggests cooler conditions than those of modern North Carolina.
The abundance of herbaceous pollen and fern spores is consistentwith proximity to the shoreline.
From12 to9 m,burrowed sand, barrenof foraminifera, is interpretedas a fluvial/floodplain unit. The Pleistocene/Holocene boundary at ca.9 m core depth, above a transgressive lag, is overlain by a coarsening-upwards sequence. The upper part, composed of barrier island sand, isbarren of foraminifera but the finer grained lower part contains theshallow inner shelf assemblage B and assemblage A (with salt-marshforaminifera); a back-barrier- estuarine setting is indicated.
4.4. OBX-11
4.4.1. Age estimatesAmino acid racemization age estimates from both Mercenaria and
Mulinia between 22.5 and 54.2 m depth provide age control for coreOBX-11. Inconsistent results between the two taxa between 22.4 and27.05 m depth are suggestive of age mixing of the older Mercenariainto the unit that is ~120 ka in age based on theMulinia results (Tables2, 3). Below 27.05 m, to 31.46 m (or deeper), internally consistentMercenaria AZ3− and Mulinia Group B (B1) results suggest an age of170 to 220 ka (Tables 2, 3). Aminozone 4+ (450 to 1840 ka) isencountered at a depth of 54.17 m, and this result is supported byresults for Ensis samples at 51.52 m (Tables 2, 3; Fig. 5). Although onespecimen of Pterocarya pollen, indicative of the Pliocene, is present inthe sample from 59 m core depth, it is considered to be reworked.Seismic data indicates that the Pliocene occurs at ca. 20 m below thebase of OBX-11 (Fig. 10; Mallinson et al., 2010a).
4.4.2. Paleoenvironmental interpretationsA basal mud (up to 66.5 m core depth) contains foraminiferal
assemblage C, indicating a reduced oxygen, inner to mid-shelf, low

Fig. 3. Percent abundance of palynomorphs of major plant taxa, core OBX-10. Asterisks indicate presence of palynomorphs. Pollen concentration is provided in number of grains pergram dry sediment. Pollen zonation is based on a combination of visual inspection and the dendrogram based on total sum of squares (Euclidean distance with no transformation)calculated using CONISS (Grimm, 1992).
236 S.J. Culver et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 227–249
energy environment (Fig. 5). A pollen assemblage from 68.1 m isdominated by Pinus (ca. 80%) with subdominant Picea and lowabundances of other tree and herb taxa, suggesting cooler thanmodern conditions.
Approximately 1 m of fluvial sand and generally leached (twosamples contain foraminiferal assemblage B) inner shelf sand (Fig. 5)overlies the basal mud up to 54.5 m. Although still dominant, Pinuspollen is less abundant than in the basal mud, and Quercus pollen issubdominant. Other common taxa include Picea, Abies, Carya andherbaceous (7–19%) pollen. The lower abundance of Picea pollensuggests warmer conditions than those of the basal mud.
Sands from 54.5 to 50.5 m contain the moderately diverse innershelf foraminiferal assemblage D. A deepening upwards shelfsequence is consistent with the up-core increasing number offoraminifera within a standard volume of sediment (Fig. 5).
A fining-upwards sequence of apparently fluvial sands between50.5 and 41 m core depth is barren of foraminifera. Flaser-beddedmuddy sand (from 41 m to 39 m) are interpreted as estuarine. Asample from near the top of this muddy sand contains a fewforaminifera of shallow inner shelf, low diversity assemblage Bprobably as a result of displacement downcore by macrofaunalburrowing from the foraminifera-rich overlying marine unit.
From39 to24 maveryfine tofinemuddysandcontainshighlydiverseforaminiferal assemblages (belonging to assemblage C) indicating lowenergy, reducedoxygen, inner tomid-shelf environments. Extremelyhighforaminiferal concentrations in these samples are consistent with thisinterpretation (Fig. 5).
Higher energy, shallow inner shelf conditions are indicated bycoarser sand containing the moderate diversity, inner shelf forami-niferal assemblage D from 24 to 13 m (Fig. 5). This marine sand isoverlain by 2 m of fluvial/floodplain sand (13–11 m).
The Pleistocene/Holocene boundary at 11 m core depth is overlainby 1.5 m of sand with Thalassinoides-type burrows and the lowdiversity, estuarine foraminiferal assemblage A (Fig. 5). From 9.5 to1.5 m, coarse tomedium sands of themodern barrier island are barrenof foraminifera except for one occurrence of the shallow inner shelfforaminiferal assemblage B (Fig. 5). The thin peat and overlying sandat the top of the core (Fig. 5) are fill deposits.
4.5. OBX-12
4.5.1. Age estimatesRadiocarbon and AAR dating of a Chione valve from 7.38 m core
depth (Fig. 6) indicate a late Holocene age (2753 to 2947 cal yr BP,Table 2). A Mercenaria valve from 17.93 m core depth was alsosubjected to paired analyses. AAR indicates AZ2 and an age estimate of~80 ka (Tables 2, 3); radiocarbon returned a finite (interpreted asminimum) age of 43,200 C-14 yr BP (Table 4; Fig. 6). A singleMulinia-based age estimate (80 to 125 ka) from 20.47 m is consistent with theMercenaria-based AAR age estimate from 17.93 m (Tables 2, 3; Fig. 6).
4.5.2. Paleoenvironmental interpretationsCoarse sandwith shell gravel andmoderately diverse foraminiferal
assemblage D containing a few planktonic foraminifera indicate innershelf deposition for the bottom 11 m of the core (Fig. 6). Mediumsands from 12.5 m core depth to the Pleistocene/Holocene boundaryat ca. 9 m contain the lower abundance and lower diversity inner shelfforaminiferal assemblage B (Fig. 6). Similar sand and foraminiferalassemblages occur immediately above the boundary up to ca. 6 m.Three pollen samples from these medium sands (two above and onebelow the Holocene/Pleistocene boundary) were dominated by Pinuspollen with varying abundances of Picea, Carya and Quercus pollen.

Fig. 4. Percent abundance of diatoms, core OBX-10. O−I=oligohalobous-indifferent. Dendrogram based on total sum of squares (Euclidean distance with no transformation) calculated using CONISS (Grimm, 1992).
237S.J.Culver
etal./
Palaeogeography,Palaeoclimatology,Palaeoecology
305(2011)
227–249

238 S.J. Culver et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 227–249

Fig. 6. Lithologic log of core OBX-12 indicating sample depths, number of benthic foraminiferal species per sample, number of specimens picked per sample, percent planktonics,calculated number of specimens in 100 g dry weight of sediment, and depth of pollen samples. b, barren of foraminifera. Horizontal fine dashed lines indicate level of AAR andmedian probability 14C age estimates. Shading indicates the Holocene section. See Fig. 2 for key to symbols.
239S.J. Culver et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 227–249
The greater abundance of Picea pollen in the lowermost (Pleistocene)assemblage suggests cooler than modern climate conditions. Thiscontrasts with the upper two Holocene assemblages, which areanalogous to modern assemblages from the pine-oak–hickory foreststhat characterize much of the Atlantic and Gulf coastal plains andindicate the shift to a modern climate regime. The upper 6 m of core iscomposed of modern barrier island sands; two samples from this unitare barren of foraminifera (Fig. 6).
4.6. OBX-13
4.6.1. Age estimatesAlthough AAR age estimates on Mercenaria and Mulinia from 11.83
to 19.02 m core depth indicate a late Pleistocene age (AZ2+ or group A;125 ka) (Tables 2, 3; Fig. 7), Holocene radiocarbon age estimates onshells collected above ~16 m (Fig. 7) indicate that Pleistocene shellsfrom 11.83 m were reworked. A radiocarbon age estimate (Table 4)from Anadara collected at 13.11 m is 5044–5327 cal yr BP. Additionalshells (one Anomia and two Crassostrea virginica) from the same sectionof the core gave very similar 2-sigma age estimates (Table 4; Fig. 7).Wood fragments from 14.56 m core depth, ca. 1.5 m above thePleistocene/Holocene boundary yielded a 2-sigma age range estimateof 9916 to 10,093 cal yr BP. We interpret the Pleistocene shells at 16.22to 16.49 m as part of a lag deposit at the base of the Holocene unit.
4.6.2. Paleoenvironmental interpretationsCoarse shelly gravelly sands comprising the lower ca. 4 m of core
contain the moderate diversity inner shelf foraminiferal assemblage D(Fig. 7). The Pleistocene/Holocene boundary at ca. 16 m is overlain (to13.8 m) by medium-grained fluvial/floodplain sands (based on terres-
Fig. 5. Lithologic log of core OBX-11 indicating sample depths, number of benthic foraminifcalculated number of specimens in 100 g dry weight of sediment, depth of pollen samples. bprobability 14C age estimates. Shading indicates the Holocene section. See Fig. 2 for key to
trial plant fragments with δ13C values of −24.63 o/oo at 14.56 m coredepth). A sample fromthebaseof this terrestrial unit contains a few(17)foraminifera that cluster in assemblage A. These specimens are likelyreworked from the underlying, highly fossiliferous, Pleistocene marinesand. Fine muddy sands from 13.8 to 12.3 m contain oysters,foraminiferal assemblage A, and a mix of polyhalobous, mesohalobous,and marine–brackish epiphytic diatoms. Collectively, these indicatedeposition in an estuarine setting. Pollen assemblages from this samesection are dominated by Pinus pollen with common (10–20%)Quercus,indicating the occurrence of southeastern conifer forests that charac-terize Holocene sediments of the southeastern United States.
Medium sands and gravel lags (12.3 to 2.0 m) contain inner shelfforaminiferal assemblages D and B (Fig. 7). Ground penetrating radar(GPR) data taken across the coring site document shallow (b10 m)reflections dipping towards the south that are interpreted torepresent a spit prograding to the south across an inlet (Mallinsonet al., 2010b). We interpret sands from 7 to 12.3 m to haveaccumulated on the shoreface, and sediments above 7 m as inlet-fillsands; foraminiferal assemblages from modern inlets along the OuterBanks (Abbene et al., 2006; Robinson and McBride, 2006; Smith et al.,2009) are consistent with this interpretation.
The upper 2 m of coarse sand in this core is interpreted as modernbarrier island overwash sand. A sample from this section is barren offoraminifera.
4.7. OBX-17
4.7.1. Age estimatesAmino acid racemization age estimates on Mulinia shells from
50.23 m core depth indicate a maximum age of 1840 ka for the core.
eral species per sample, number of specimens picked per sample, percent planktonics,, barren of foraminifera. Horizontal fine dashed lines indicate level of AAR and mediansymbols.

Fig. 7. Lithologic log of core OBX-13 indicating sample depths, number of benthic foraminiferal species per sample, number of specimens picked per sample, percent planktonics,calculated number of specimens in 100 g dry weight of sediment, percent polyhalobous (P), mesohalobous (M), and oligohalobous (O) diatoms, depth of pollen samples. b, barren offoraminifera. Horizontal fine dashed lines indicate level of median probability 14C and AAR age estimates. Shading indicates the Holocene section. See Fig. 2 for key to symbols.
240 S.J. Culver et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 227–249
Foraminiferal assemblages from 40.93, 42.31, and 43.77 m containthe planktonic taxa Globigerina bulloides, G. falconensis, G. calida,G. quinqueloba, Globigerinoides ruber, Globigerinita glutinata andGloborotalites hexagona, indicating a Quaternary age. No evidencefor Pliocene sediments was recovered. AAR age estimates on multipleMercenaria (aminozone AZ2) and Mulinia (group A1) specimensdemonstrate the stratigraphic reliability of AAR for these genera(Wehmiller et al., 2010a). Age estimates between 15.27 and 21.34 mare ~80 ka (Tables 2, 3; Fig. 8). Radiocarbon dating of shells collectedwithin this interval provide minimum ages of 28,700 14C years(Spisula at 20.42 m) and 31,700 14C years (Mulinia at 15.27 m). Aslightly older age (125 ka) is indicated by Mulinia results (group A2)from 24.15 to 24.86 m. Three more Mulinia age groups (B1, C1, andC2) are recognized at depths of 32.03, 41.58, and 50.23 m, respec-tively, suggesting ages of 170 to 220 ka, 1130 to 1450 ka, and 1440 to1840 ka, respectively (Tables 2, 3; Fig. 8).
4.7.2. Paleoenvironmental interpretationsThe lower 14 m of the core (up to 44 m) is characterized by
medium to coarse sands and the moderate diversity inner shelfforaminiferal assemblage D (Fig. 8). Seismic data (Mallinson et al.,2010a) indicate that these sands comprise prograding clinoforms. Thepresence of planktonic foraminifera and slightly higher diversityforaminiferal assemblages near the base of this section suggests ashallowing upward sequence from the lower shoreface to the uppershoreface (Mallinson et al., 2010a).
From 44 to 22.5 m core depth, a coarsening (shoaling) upwardsequence from muddy sand to medium sand is generally characterizedby reduced oxygen, mid-shelf, low energy, foraminiferal assemblage C(Fig. 8).High concentrationsof foraminifera (nearly 5million specimensper 100 g dry weight of sediment) occur near the base of this section,where assemblages are diverse, and planktonic foraminifera compriseup to 12% of assemblages (Fig. 8). Fluctuations in environmentalconditions are evident from variability in foraminiferal assemblagesthroughout this section. The lower 2 m (44–42 m) are characterized bydominance of Rosalina spp., abundance of Elphidium excavatum, and
common occurrence of Bolivina lowmani, Textularia cf. T. gramen,Hanzawaia strattoni and Quinqueloculina spp. Benthic foraminiferaldiversity and concentration are high, and planktonic foraminiferacomprise a few percent of assemblages, indicating an open mid-shelfenvironment. From 42 to 33 m, planktonic foraminifera are moreabundant (generally N5%), and low oxygen-tolerant benthic species(e.g., Buliminella elegantissima, Fursenkoina fusiformis, Stainforthiacomplanata) are less abundant than lower in the core. Theseassemblages indicate reduced oxygen levels most likely associatedwith deeper mid-shelf conditions throughout most of this interval. Theobservation of at least one interval of decreased foraminiferalconcentrations and planktonic foraminiferal abundance and a shift tobenthic foraminiferal assemblages characteristic of shallower shelf, welloxygenated conditions at 37.5 m (Fig. 8) indicates environmentalvariability within this section. From 33 to 26 m, decreased abundanceof reduced oxygen-tolerant species and foraminiferal concentrationsindicate increasingly well oxygenated conditions, but mid-shelf depthsstill are indicated by the high species richness and high percentages ofplanktonic foraminifera (Fig. 8). From 26 to 22.5 m, a coarse to mediumsand contains fewer specimens, fewer planktonic foraminifera, and thelow diversity inner shelf foraminiferal assemblage B, indicatingsignificant shoaling near the top of the 21.5 m thick shoaling upwardunit (Fig. 8).
Coarse shelly sand beds characterized by the moderately diverse,shallow shelf foraminiferal assemblage D extend from 22.5 m to thePleistocene/Holocene boundary at 14.3 m core depth. The decreasednumber of species, specimens and planktonic foraminifera upwards inthis section suggest shallowing and/or increasing proximity to shorethrough time.
Medium sands from 14.3 to 3.3 m are characterized by the lowdiversity, shallow inner shelf foraminiferal assemblage B. Becausecore OBX-17 was drilled through beach ridges, this section of coreprobably represents the stacked, shoreface portions of one or moreridges. The upper 3.3 m of core is barren of foraminifera andrepresents the eolian portion of a beach ridge. A thin peat at 0.9 mis barren of foraminifera and composed of woody plant fragments,

Fig. 8. Lithologic log of core OBX-17 indicating sample depths, number of benthic foraminiferal species per sample, number of specimens picked per sample, percent planktonics, andcalculated number of specimens in 100 g dry weight of sediment. b, barren of foraminifera. Horizontal fine dashed lines indicate level of AAR and median probability 14C ageestimates. Shading indicates the Holocene section. See Fig. 2 for key to symbols.
241S.J. Culver et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 227–249
seeds and stalks, indicating an inter-ridge, freshwater swampenvironment.
4.8. Oregon Inlet DOT cores
Sedimentological data from seven split-spoon cores, ranging inpenetration from 44 to 77 m and taken across and slightly west of
Oregon Inlet (Fig. 1), provide stratigraphic information betweenOBX-10 and OBX-11 along the northern portion of the transect.Seismic data (Mallinson et al., 2010a) suggest that two DOT cores(M-B38 and M-B51) penetrate Pliocene deposits at ca. 76 m belowmodern sea level. Uppermost Pliocene sediments are characterizedby dense, iron stained, shelly sandy mud with occasional ferricreteaggregates. The character of the overlying unconformity (Q0) is

Fig. 9. Dendrogram resulting from cluster analysis of the 64 benthic foraminiferal taxa that comprised 2% or more of the assemblage in any single sample in cores OBX-10, 11, 12, 13and 17 and showing sample membership of cluster groups A, B, C, and D.
242 S.J. Culver et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 227–249
unknown due to the sampling interval in the boreholes (10 cm longsplit-spoon samples at 1.5 m intervals). However, the unconformityis indicated by a sharp decrease in the split-spoon coring blow-countrecord.
Pliocene deposits are overlain by a fining upward sequence offluvial sand and gravel to estuarine mud between ca. 76 m and 63 mbelow modern sea level. These in turn are overlain by shelly muddysand and sandy mud marine deposits that are interpreted to correlate
with AZ4 along the OBX cross section (Fig. 10). Seismic reflection Q30separates AZ4 from younger deposits.
Within the Oregon Inlet cores, Q30 is locally overlain by 3 to 6 m offluvial sand and gravel that forms valley-fill beneath the Q40 reflector(Fig. 10). In this area, the base of the valley-fill locally coincides withQ30. These deposits are overlain by an upward succession from shellymud to shelly muddy sand that is interpreted to reflect a transitionfrom estuarine to marine conditions, correlates with AZ3, and is

243S.J. Culver et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 227–249
bounded above by Q50 at ca. 20 below present sea level (Fig. 10). Atthe north and south ends of the Oregon Inlet transect (cores M-B44andM-B33 in Fig. 10), Q50 is overlain by shelly muddy sand (betweenca. −20 m and −12 m) that correlates with AZ2 on the OBX crosssection (Fig. 10). Within the central portion of the Oregon Inlettransect, AZ2 deposits are truncated by valley-fill deposits that gradeupward from fluvial sand and gravel to a thin layer of peat (on theflanks) to estuarine mud and are interpreted to be post LGM andHolocene in age (Fig. 10). The upper ca. 8 m of the entire section iscomposed of inlet and barrier-island sand that is interpreted asHolocene in age.
4.9. Avon–Buxton DOT cores
Sedimentological, foraminiferal and chronostratigraphic (a radio-carbon age estimate) data from six split-spoon cores taken betweenAvon and Buxton (Fig. 1) document a Quaternary stratigraphic recordclosely comparable to that in the adjacent, more continuous rotasoniccores OBX-13 and 17 (Fig. 10). Based on foraminiferal data, shellyinner shelf marine sands comprise much of the Pleistocene section(Fig. 10; Jomp et al., 2004). Reduced oxygen inner to mid-shelfassemblages (equivalent to assemblage C in OBX-10 to 13, 17) occurbelow seismic reflection Q30 in DOT2; in DOT1 they occur in the lowerpart of the overlying sequence (bounded by Q30 and Q50; Fig. 10) atthe same depth below modern sea level (ca. 25 to 40 m) as in theadjacent OBX-17.
Seismic reflection Q99, and the immediately overlying organic-rich sediments representing the deposits that accumulated underterrestrial conditions on the last glacial maximum surface during thelatest Pleistocene, can be traced in DOT1 to DOT6 at the sameapproximate depth as in OBX-13 and 17 (Fig. 10). Peat from 11 to12 m core depth in DOT1 yielded a 2-sigma age range of 13,320 to13,584 cal yr BP. Similar material in OBX-13 from 14.58 m core depthgave a 2-sigma age of 9916 to 10,093 cal yr BP. The δ13Cvalues of−26.5 and −24.6 o/oo, respectively, for these two samples (Table 4)indicate terrestrial organic material.
The Holocene record in DOT1 to 6 is composed of fine to mediumshelly sand. In DOT1 and 2, a diverse foraminiferal assemblage ofnormal marine salinity shallow water foraminifera is consistent withdeposition in an inlet environment (Abbene et al., 2006; Robinson andMcBride, 2006; Vance et al., 2006; Smith et al., 2009). The AshWednesday nor'easter (Davis and Dolan, 1993) opened an inletbetween Avon and Buxton in 1962; it was closed artificially thefollowing year.
5. Nature and timing of paleoenvironmental change
Correlation between OBX-10, 11, 12, 13 and 17, and between OBX-10 and OBX-08 to the north (Culver et al., 2008), is based on severallines of evidence: sedimentology, stratigraphy, foraminifera, diatoms,pollen, radiocarbon age estimates, AAR age estimates (Mercenaria andMulinia-based), gamma logs, and seismic data (Fig. 10). Gaps betweenrotasonic cores OBX-10 and 11 and OBX-13 and 17 were partiallyfilled with two sets of split-spoon cores. The resulting generally north-south geologic section along the modern Outer Banks provides adetailed understanding of the Quaternary deposits filling the centralAlbemarle Embayment (Fig. 10).
The seismic stratigraphy of this region has been described byMallinson et al. (2010a,b). The major, high amplitude, regionallycontinuous acoustic reflections (Q0, Q10, Q20?, Q30, Q40?, Q50 andQ99) and the seismic stratigraphic units (SSUs) that they define aresummarized in Table 6. The age of SSUs are constrained, in part, byAAR data (Tables 2, 3; Fig. 10). Reflections Q0, Q30, Q50 and Q99, inparticular, proved useful in determining correlations between OBX-10and 17.
The high amplitude, regionally continuous reflection (Q0) thatmarks the boundary between Pliocene and Pleistocene strata(Mallinson et al., 2010a) dips towards the center of the section fromboth north and south (Fig. 10). Thus, OBX-11 is located in the deepestpart of the Albemarle Embayment along this transect. Q0 waspenetrated in OBX-07 to the north (Culver et al., 2008), but noother OBX cores in the northern or central Albemarle basin proved tobe deep enough to reach the base of the Quaternary section. Two NCDOT split-spoon cores taken in Pamlico Sound at Oregon Inlet areinterpreted as having penetrated into the Pliocene (Fig. 10).
Sediments comprising SSU I-III, between reflections Q0 and Q30(Table 6), comprise the early Pleistocene section (Fig. 10A). Seismicreflection correlations indicate that sediments below Q0 are Pliocenein age based on planktonic foraminifera in OBX-07 at Nags Head to thenorth (Culver et al., 2008) and macrofossils (P. Weaver, personalcommunication) in carbonate sediments below Q0 in OBX-14, arotasonic core on the northernmargin of the Cape Lookout High to thesouth (Fig. 1). AAR data for SSU I-III indicate assignment to AZ4 andAZ4+ and deposition between 1130 to 1840 ka BP (Tables 2, 3).
The majority of the early Pleistocene section is composed of innerto mid-shelf marine deposits (OBX-08, 10, 11, 17; M-B51, 38, 58;Fig. 10B). Pollen data from OBX-10 and 11 indicate air temperaturescooler than those of today (Fig. 10B). Two thick fluvial sequencesoccur towards the north of the cross section (Fig. 10). A much thinnerfluvial section was penetrated in the center of the embayment (OBX-11; Fig. 10). Estuarine deposits were penetrated immediately beneathQ30 in OBX-10 (Fig. 10).
The middle Pleistocene record is bounded by Q30 and Q50(Table 6; Fig.10A); the age of these deposits (SSU IV) is indicated bythe presence of AZ 3+ to AZ3− (deposition between 170 and 430 kaBP; Tables 2, 3). Like the early Pleistocene section, the middlePleistocene is composed mainly of inner to mid-shelf marine deposits(OBX-11, 17; M-B44, 40, 51, 38, 58, 36, 33; DOT3, 6, 2, 5, 1, 4; Fig. 10B).Also like the early Pleistocene, themiddle Pleistocene section containsextensive fluvial deposits that extend into the central AlbemarleEmbayment (OBX-11; Fig. 10). The younger fluvial sequence is cappedby non-marine, probably floodplain mud in OBX-08 and freshwaterpeat and estuarine sand in OBX-10 (Fig. 10B) which contain pollenassemblages indicative of slightly cooler air temperatures than thosein modern North Carolina.
High amplitude seismic reflection Q50 is mapped on Fig. 10 as athin (ca. 3 m) zone composed of a marine transgressive ravinementsurface and overlying marine, shelly lag deposits. The position of Q50in OBX11 and OBX17 as plotted in Fig. 10 represents an interpretationbased on lithologic properties of the cores. The cores were located0.5 km (OBX11) and 1.5 km (OBX17) from the nearest seismic data,hence there is some uncertainty in the placement of the Q50 reflectorin each core. The AAR data for OBX11 and OBX17 suggest that Q50might be ~3 to 4 m deeper than shown in Fig. 10, although burrowingand sediment mixing around the interval of Q50 could be responsiblefor this ambiguity.
Based on the 80 to 125 ka age estimates (AZ2 and AZ2+) for theoverlying deposits (indicating preservation of marine strata fromMarine Isotope Stages 5a and 5e; Mallinson et al., 2008), Q50 (Table 3)likely represents the MIS 6 to 5 transgressive ravinement surface(Mallinson et al., 2010a). Thus, the majority (see below) of the latePleistocene record is bounded by reflections Q50 and Q99 (Fig. 10A).The late Pleistocene section along the transect is shallow inner shelfmarine sand. Pollen assemblages from this section (in OBX-10 and 12)indicate air temperatures slightly cooler than those of modern NorthCarolina (Fig. 10B).
Q99 (Table 6) is a medium to high amplitude semi-continuousreflection that defines the land surface and paleodrainage pattern ofthe last glacial stage in the Albemarle Embayment (Mallinson et al.,2010a). In core it was identified using a combination of criteria. Q99defines an erosional surface at the base of incised valleys that is
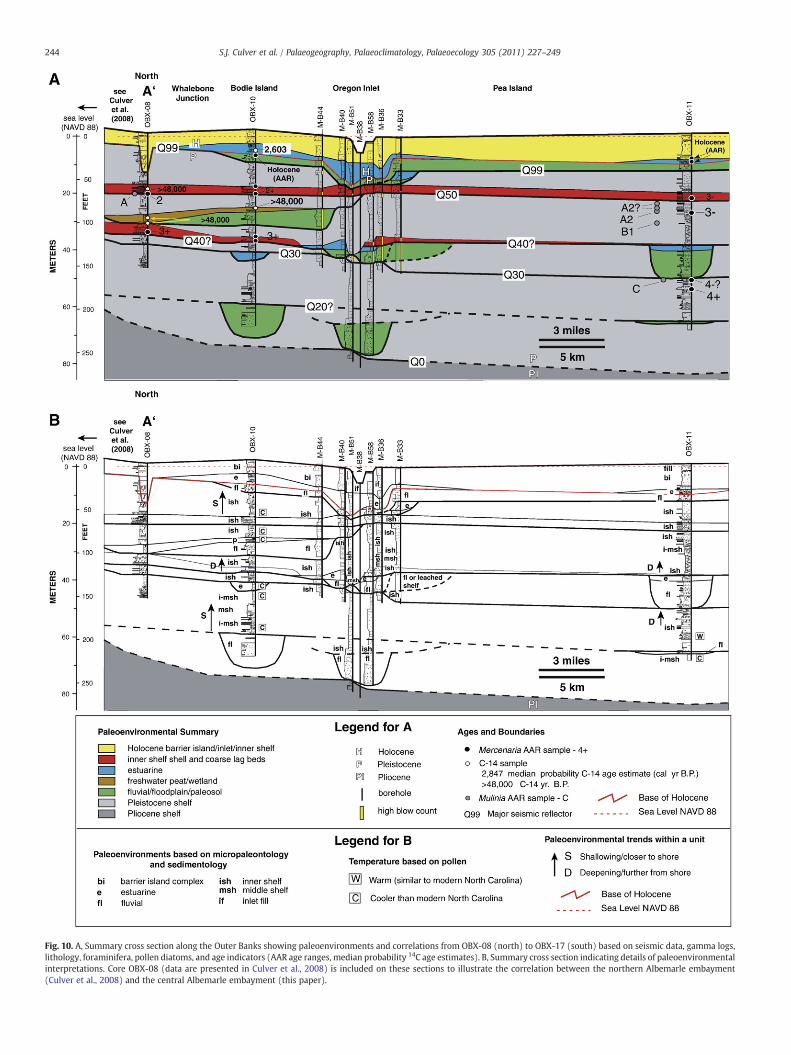
Fig. 10. A, Summary cross section along the Outer Banks showing paleoenvironments and correlations from OBX-08 (north) to OBX-17 (south) based on seismic data, gamma logs,lithology, foraminifera, pollen diatoms, and age indicators (AAR age ranges, median probability 14C age estimates). B, Summary cross section indicating details of paleoenvironmentalinterpretations. Core OBX-08 (data are presented in Culver et al., 2008) is included on these sections to illustrate the correlation between the northern Albemarle embayment(Culver et al., 2008) and the central Albemarle embayment (this paper).
244 S.J. Culver et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 227–249

Fig. 10 (continued).
245S.J. Culver et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 227–249
contiguous with a flooding surface or facies contact that is mappableacross interfluves. In the current study area, Q99 dips to define apaleovalley beneath the modern Oregon Inlet and, possibly, a smallerfluvial valley between Avon and Buxton (Fig. 10). Localities withincised paleovalleys commonly include basal, well-defined fluvial toestuarine deposits. Between valleys, however, Q99 is inferred from acontact that separates an underlying weathering profile or paleosolremnant from overlying relatively unweathered deposits that mayinclude marine shelf to barrier island deposits. For mapping purposes,paleosols are included in the fluvial/floodplain unit. Q99 is overlain byup to 5 m of weathered fluvial/floodplain/paleosol sands (Fig. 10B)that accumulated or weathered in place during the latest Pleistocene(post last glacial maximum) upon the terrestrial landscape.
The Holocene section in several cores commences with estuarinemuddy sands (Fig. 10B) representing the initial flooding of the last glacialdrainage system.When this occurred depends on the depth of individualparts of the drainage system. In the paleo-Roanoke valley in northernAlbemarle Embayment, estuarine deposits commenced accumulating ca.12,000 cal yr BP (Culver et al., 2008). Sea level was at −35.7+/−1.1 mMSLat 11,062–10,576 cal yr BP (Hortonet al., 2009). Estuarine conditionsin the paleo-Neuse/Tar drainage of the southern Albemarle Embaymentwere established ca. 9000 cal yr BP based on our understanding of thedepth of the Pleistocene surface (Mallinson et al., 2010a) and theHolocene sea level curve for North Carolina (Horton et al., 2009). Shallowestuarine deposits in OBX-10 (Figs. 2, 10A) are dated at 2716–2457 cal yrBP and probably represent Pamlico Sound estuarine deposits that
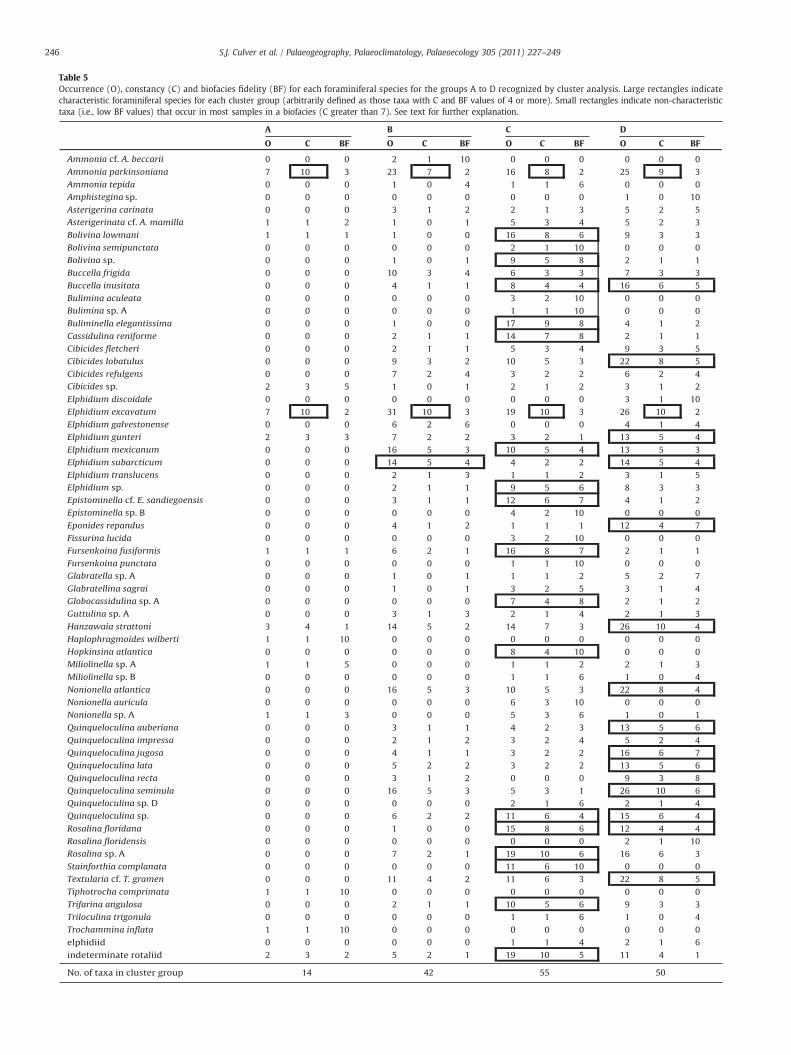
Table 5Occurrence (O), constancy (C) and biofacies fidelity (BF) for each foraminiferal species for the groups A to D recognized by cluster analysis. Large rectangles indicatecharacteristic foraminiferal species for each cluster group (arbitrarily defined as those taxa with C and BF values of 4 or more). Small rectangles indicate non-characteristictaxa (i.e., low BF values) that occur in most samples in a biofacies (C greater than 7). See text for further explanation.
A B C D
O C BF O C BF O C BF O C BF
Ammonia cf. A. beccarii 0 0 0 2 1 10 0 0 0 0 0 0Ammonia parkinsoniana 7 10 3 23 7 2 16 8 2 25 9 3Ammonia tepida 0 0 0 1 0 4 1 1 6 0 0 0Amphistegina sp. 0 0 0 0 0 0 0 0 0 1 0 10Asterigerina carinata 0 0 0 3 1 2 2 1 3 5 2 5Asterigerinata cf. A. mamilla 1 1 2 1 0 1 5 3 4 5 2 3Bolivina lowmani 1 1 1 1 0 0 16 8 6 9 3 3Bolivina semipunctata 0 0 0 0 0 0 2 1 10 0 0 0Bolivina sp. 0 0 0 1 0 1 9 5 8 2 1 1Buccella frigida 0 0 0 10 3 4 6 3 3 7 3 3Buccella inusitata 0 0 0 4 1 1 8 4 4 16 6 5Bulimina aculeata 0 0 0 0 0 0 3 2 10 0 0 0Bulimina sp. A 0 0 0 0 0 0 1 1 10 0 0 0Buliminella elegantissima 0 0 0 1 0 0 17 9 8 4 1 2Cassidulina reniforme 0 0 0 2 1 1 14 7 8 2 1 1Cibicides fletcheri 0 0 0 2 1 1 5 3 4 9 3 5Cibicides lobatulus 0 0 0 9 3 2 10 5 3 22 8 5Cibicides refulgens 0 0 0 7 2 4 3 2 2 6 2 4Cibicides sp. 2 3 5 1 0 1 2 1 2 3 1 2Elphidium discoidale 0 0 0 0 0 0 0 0 0 3 1 10Elphidium excavatum 7 10 2 31 10 3 19 10 3 26 10 2Elphidium galvestonense 0 0 0 6 2 6 0 0 0 4 1 4Elphidium gunteri 2 3 3 7 2 2 3 2 1 13 5 4Elphidium mexicanum 0 0 0 16 5 3 10 5 4 13 5 3Elphidium subarcticum 0 0 0 14 5 4 4 2 2 14 5 4Elphidium translucens 0 0 0 2 1 3 1 1 2 3 1 5Elphidium sp. 0 0 0 2 1 1 9 5 6 8 3 3Epistominella cf. E. sandiegoensis 0 0 0 3 1 1 12 6 7 4 1 2Epistominella sp. B 0 0 0 0 0 0 4 2 10 0 0 0Eponides repandus 0 0 0 4 1 2 1 1 1 12 4 7Fissurina lucida 0 0 0 0 0 0 3 2 10 0 0 0Fursenkoina fusiformis 1 1 1 6 2 1 16 8 7 2 1 1Fursenkoina punctata 0 0 0 0 0 0 1 1 10 0 0 0Glabratella sp. A 0 0 0 1 0 1 1 1 2 5 2 7Glabratellina sagrai 0 0 0 1 0 1 3 2 5 3 1 4Globocassidulina sp. A 0 0 0 0 0 0 7 4 8 2 1 2Guttulina sp. A 0 0 0 3 1 3 2 1 4 2 1 3Hanzawaia strattoni 3 4 1 14 5 2 14 7 3 26 10 4Haplophragmoides wilberti 1 1 10 0 0 0 0 0 0 0 0 0Hopkinsina atlantica 0 0 0 0 0 0 8 4 10 0 0 0Miliolinella sp. A 1 1 5 0 0 0 1 1 2 2 1 3Miliolinella sp. B 0 0 0 0 0 0 1 1 6 1 0 4Nonionella atlantica 0 0 0 16 5 3 10 5 3 22 8 4Nonionella auricula 0 0 0 0 0 0 6 3 10 0 0 0Nonionella sp. A 1 1 3 0 0 0 5 3 6 1 0 1Quinqueloculina auberiana 0 0 0 3 1 1 4 2 3 13 5 6Quinqueloculina impressa 0 0 0 2 1 2 3 2 4 5 2 4Quinqueloculina jugosa 0 0 0 4 1 1 3 2 2 16 6 7Quinqueloculina lata 0 0 0 5 2 2 3 2 2 13 5 6Quinqueloculina recta 0 0 0 3 1 2 0 0 0 9 3 8Quinqueloculina seminula 0 0 0 16 5 3 5 3 1 26 10 6Quinqueloculina sp. D 0 0 0 0 0 0 2 1 6 2 1 4Quinqueloculina sp. 0 0 0 6 2 2 11 6 4 15 6 4Rosalina floridana 0 0 0 1 0 0 15 8 6 12 4 4Rosalina floridensis 0 0 0 0 0 0 0 0 0 2 1 10Rosalina sp. A 0 0 0 7 2 1 19 10 6 16 6 3Stainforthia complanata 0 0 0 0 0 0 11 6 10 0 0 0Textularia cf. T. gramen 0 0 0 11 4 2 11 6 3 22 8 5Tiphotrocha comprimata 1 1 10 0 0 0 0 0 0 0 0 0Trifarina angulosa 0 0 0 2 1 1 10 5 6 9 3 3Triloculina trigonula 0 0 0 0 0 0 1 1 6 1 0 4Trochammina inflata 1 1 10 0 0 0 0 0 0 0 0 0elphidiid 0 0 0 0 0 0 1 1 4 2 1 6
indeterminate rotaliid 2 3 2 5 2 1 19 10 5 11 4 1
No. of taxa in cluster group 14 42 55 50
246 S.J. Culver et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 227–249

Table 6Seismic reflection and seismic stratigraphic unit (SSU) summary (modified fromMallinson et al., 2010a). Age estimates based on AAR data fromWehmiller et al. (2010a)and strontium isotope data from Mallinson et al. (2010a). MIS=Marine Isotope Stage.HST, LST and FSST=highstand, lowstand and falling stage systems tracts, respectively.
Reflection SSU Age Interpretation
Q100 Holocene Modern sediment/water interfaceVI Holocene Estuarine, barrier island and shallow
open shelf sedimentsQ99 Late Pleistocene Last glacial maximum subaerial
unconformityV Late Pleistocene
MIS 5Multiple depositional sequences withextensive estuarine,shelf, and fluvial valley-fill
Q50 Late Pleistocene Transgressive ravinement surfaceassociated withMIS 6/5 sea-level rise
IV Early to middlePleistocene
Multiple eroded depositional sequences.FSST and TST.
MIS 7–13? Multiple incised valleysQ30 Early to middle
PleistoceneSubaerial unconformity
III Early Pleistocene Multiple parasequences. Coastal to shelfdeposits
Q20 Early Pleistocene Subaerial to shallow marineunconformity; defineslowstand shelf
II Early Pleistocene Multiple depositional sequencesexhibitingLST, TST and HST
Q10 Early Pleistocene Unconformity - minimal subaerialexposure
I Early Pleistocene Mid to outer shelfQ0 Pliocene/Pleistocene
boundarySubmarine unconformity
247S.J. Culver et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 227–249
accumulated behind barrier islands thatwere present ca. 3000 to 2500 yrBP based on optically stimulated luminescence dating (OSL; Mallinson etal., 2008). Overlying the estuarine deposits, from OBX-11 to the north, isbarrier island sand (Fig. 10B) that is generally barren of foraminifera(Culver et al., 2006). At OBX-13, a thin estuarine sequence is overlain byinlet-fill sand and, finally, by barren barrier island sand (Figs. 7, 10B).Pollen from the estuarine unit indicates air temperatures similar to thosein this region today.
At OBX-12 and 17, shallow marine sand overlies the post lastglacial maximum terrestrial deposits (Fig. 10B). At OBX-12, this sand,dated at 2947–2753 cal yr BP and containing a pollen assemblagesimilar to those found in modern Chesapeake Bay, represents innershelf shoal sands that accumulated prior to barrier island formation atthis location (Ricardo et al., 2004; Ricardo, 2005). They are overlain bybarrier island sand (Figs. 6, 10B). The entire Holocene section at OBX-17 is composed of beach-ridge sand with a lower shorefacecomponent bearing shallow inner shelf foraminifera, overlain by asubaerial component barren of foraminifera (Figs. 8, 10B). Theformation of the Cape Hatteras beach ridges is likely to havecommenced ca. 3000 to 2500 yr BP (based on OSL age estimates onKitty Hawk beach ridges in the northern Outer Banks; Mallinson et al.,2008) but the oldest OSL age estimate yet acquired near the site ofOBX-17 is ca. 1600 yr BP (McDowell, 2009).
Stacked inner shelf sequences, representing successive transgres-sive–regressive cycles characterize the southern part of the study area.The bases of these units are marked by reflectors interpreted astransgressive surfaces. In some areas,where data density is high, such asthe central and northern part of the study area, thick fluvial/floodplaindeposits fill paleovalleys, andmay be overlain by thin estuarine units orinner tomid-shelf deposits. Thick finer grained units (sandymuds)mayinclude subtle sequence boundaries where microfossils demonstratethat inner to mid-shelf, low energy, perhaps reduced oxygen depositsare directly overlain by estuarine deposits (e.g., OBX-10). Gamma logs
and sedimentary structures help confirm the existence of suchboundaries (Figs. 2, 10).
The progression of paleoenvironments of the central Albemarleembayment demonstrates a shallowing of the depositional basin over aseries of transgressive–regressive cycles throughout Quaternary time.Inner tomid-shelf early Pleistocenedeposits, representing transgressiveto highstand systems tracts (TST to HST) are overlain by similar middlePleistocene deposits in the southern part of the study area but by innershelf deposits in the north. Late Pleistocene marine deposits weredeposited in inner shelf settings. Thin lowstand deposits, commonlywith basal fluvial deposits, are preserved in remnants of incised valleysbeneath each TST–HST cycle. Holocene deposits are marginal marine inorigin and represent shoreface, inlet, barrier island, beach ridge andestuarine environments. This pattern reflects thefilling of theAlbemarleEmbayment through time and the eastward progradational construc-tion of the continental shelf (Mallinson et al., 2010a). Thus, as a corollaryto this basin infilling and progradational depositional pattern, the earlyand/or middle Pleistocene shoreline probably formed the regionallymapped topographic feature known as the Surry Scarp (Winker andHoward, 1977), whereas the late Pleistocene shoreline was located atthe Suffolk Scarp (Fig. 1) during MIS 5a and 5c (Mallinson et al., 2008;Parham, 2009) and possibly MIS 5e as well, although evidence for is thelatter is currently lacking. The modern oceanic shoreline wouldprobably be located in the vicinity of the mainland shoreline of theAlbemarle–Pamlico estuarine system were it not for the presence of agenerally north–south last glacial lowstand interstream divide uponwhich the late Holocene Outer Banks barrier islands formed (Culveret al., 2007) and into which the cores of this study were drilled.
6. Conclusions
Micropaleontological (foraminifera, diatoms and pollen) and sedi-mentological data, placed in a seismic (geometric) framework and achronostratigraphic framework, based on radiocarbon and AAR data,help elucidate the paleonenvironmental evolution of the centralAlbemarle embayment of North Carolina during the Quaternary. Thesedata also provide an increased understanding of the stratigraphicarchitecture of a generally inner to mid-shelf basinal setting thatdeveloped during a period of frequent, high amplitude sea-levelfluctuations.
The Albemarle Embayment was flooded during several transgres-sive–regressive cycles,with incomplete preservationof the stratigraphicrecord. Throughout theQuaternary, the preserved shelf deposits show ashallowing trend as the embayment was gradually filled while thecontinental shelf prograded eastwards. Shelf deposits are incised bylowstand paleovalleys which are partly filled by a succession of fluvial/floodplain to estuarine and evenmarinedeposits. Preservation potentialformarginal marine deposits is poor; those that characterize this regiontodaywill likely be removed, at least in part, as continued rising sea levelfloods the region and returns the Albemarle Embayment to a shallowinner shelf environment. The resulting stratigraphic pattern will be arepeat of that seen several times in the underlying Pleistocenestratigraphic record.
Acknowledgments
We thank D.R. Corbett, I. Abbene, C.W. Hoffman, D. Merritt, B.Landacre, C. Grand Pre, A. Kemp, J. Smith, M. Robertson, J. Roberts, J.Jomp, J. Foley, J. Ricardo, J. Pierson, R. Pruitt and T. Sheehan for their helpand support and T. Cronin, C. Bernhardt, C. Brunner and S. Gallagher forreviews. M.A. Buzas and J. Jett provided access to the CushmanCollection of foraminifera, Smithsonian Institution and J. Whittakerand C. Jones to the foraminiferal collections in The Natural HistoryMuseum, London. This research is part of the North Carolina CoastalGeology Cooperative Program (NCCGC). Funding for the USGS cooper-ative agreement awards 02ERAG0044, O2ERAG0050, and 01CRAG0015

248 S.J. Culver et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 227–249
and for NSF Cooperative agreement award OCE-9807266 is gratefullyacknowledged.
References
Abbene, I.J., Culver, S.J., Corbett, D.R., Buzas, M.A., Tully, L.S., 2006. Distribution offoraminifera in Pamlico Sound, North Carolina over the past century. Journal ofForaminiferal Research 36, 135–151.
Brown, P.M., Miller, J.A., Swain, F.M., 1972. Structural and stratigraphic framework, andspatial distribution of permeability of Atlantic coastal plain, North Carolina to NewYork. U.S. Geological Survey. Professional Paper 796, 1–79.
Culver, S.J., Buzas, M.A., 1980. Distribution of recent benthic foraminifera off the NorthAmerican Atlantic coast. Smithsonian Contributions to the Marine Sciences 13,1–512.
Culver, S.J., Horton, B.P., 2005. Infaunal marsh foraminifera from the Outer Banks, NorthCarolina, USA. Journal of Foraminiferal Research 35, 148–170.
Culver, S.J., Snedden, J.W., 1996. Foraminifera and their implications for the formationof New Jersey shelf sand ridges. Palaios 11, 161–175.
Culver, S.J., Woo, H.J., Oertel, G.E., Buzas, M.A., 1996. Foraminifera of coastal depositionalenvironments, Virginia, U.S.A: distribution and taphonomy. Palaios 11, 459–486.
Culver, S.J., Ames, D.V., Corbett, D.R., Mallinson, D.J., Riggs, S.R., Smith, C.G., Vance, D.J.,2006. Foraminiferal and sedimentary record of late Holocene barrier islandevolution, Pea Island, North Carolina. Journal of Coastal Research 22, 406–416.
Culver, S.J., Grand Pre, C.A., Mallinson, D.J., Riggs, S.R., Corbett, D.R., Foley, J., Hale, M.,Metger, L., Ricardo, J., Rosenberger, J., Smith, C.G., Smith, C.W., Snyder, S.W.,Twamley, D., Farrell, K., Horton, B.P., 2007. Late Holocene barrier island collapse:Outer Banks, North Carolina, USA. The Sedimentary Record 5, 4–8.
Culver, S.J., Farrell, K.M., Mallinson, D.J., Horton, B.P., Willard, D.A., Thieler, E.R., Riggs,S.R., Snyder, S.W., Wehmiller, J.F., Bernhardt, C.E., Hillier, C., 2008. Micropaleon-tologic record of late Pliocene and Quaternary paleoenvironments in the northernAlbemarle Embayment, North Carolina, U.S.A. Palaeogeography, palaeoclimatol-ogy. Palaeoecology 264, 54–77.
Davis, R.E., Dolan, R., 1993. Nor'easters. American Scientist 81, 428–439.Denys, L., 1991–1992. A check-list of the diatoms in the Holocene deposits of the
western Belgian coastal plain with a survey of their apparent ecologicalrequirements. Professional Paper Belgian Geological Survey 246, 1–41.
Edwards, L.E., Willard, D.A., 2001. Dinoflagellate cysts and pollen from sedimentsamples, Mississippi Sound and Gulf of Mexico. In: Gohn, G.S. (Ed.), Stratigraphicframework of the Neogene and Quaternary sediments of the Mississippi coastalzone, Jackson County, Mississippi. U.S. Geological Survey Open-file Report 01–415(CD-ROM).
Ellison, R.L., Nichols, M.M., 1976. Modern and Holocene foraminifera in the ChesapeakeBay region. In: Schafer, C.T., Pelletier, B.R. (Eds.), First International Symposium onBenthonic Foraminifera of Continental Margins: Maritime Sediments, SpecialPublication 1, pp. 131–151.
Folk, R.L., 1980. Petrology of Sedimentary Rock. Hemphill Publishing Company, Austin,Texas.
Gohn, G.S., 1988. Late Mesozoic and early Cenozoic geology of the Atlantic coastal plain:North Carolina to Florida. In: Sheridan, R.E., Grow, J.A. (Eds.), The Geology of NorthAmerica, v.1–2, The Atlantic Continental Margin, U.S. Geological Society of America,pp. 107–130.
Grimm, E.C., 1992. CONISS: a Fortran 77 program for stratigraphically constrainedcluster analysis by the method of incremental sum of squares. Computers andGeosciences 13, 13–35.
Grimsdale, T., Morkhoven, F., 1955. The ratio between pelagic and benthonicforaminifera as a means of estimating the depth of deposition of sedimentaryrocks. IV World Petrology Congress, Proceedings, Section I/D, Report 4, 473–491.
Grossman, S., Benson, R., 1967. Ecology of Rhizopodea and Ostracoda of SouthernPamlico Sound Region, North Carolina. The University of Kansas PaleontologicalContributions 44, 1–90.
Hazel, J.E., 1977. Use of certain multivariate and other techniques in assemblage zonebiostratigraphy: examples utilizing Cambrian, Cretaceous and Tertiary benthicinvertebrates. In: Kauffman, E.G., Hazel, J.E. (Eds.), Concepts and Methods ofBiostratigraphy: Dowden. Hutchinson and Ross, Inc., Stroudsberg, pp. 187–212.
Hendey, N.I., 1964. An introductory account of the smaller algae of British coastalwaters, bacillariophyceae (diatoms). Fishery Investigation Series N, London, HMSO.
Hoffman, C.W., Farrell, K.M., Pierson, J.A., Ward, A.N., Nickerson, J.G., Chapman, J.S.,Brooks, R.W., Ganey, J.G., 2007. Rotasonic cores from northeastern North CarolinaVolume III: cores MLD-01 through MLD-05. North Carolina Geological Survey,Open-File Report 2007–01. .
Horton, B.P., Culver, S.J., 2008. Modern intertidal foraminifera of the Outer Banks, NorthCarolina, USA, and the applicability for sea-level studies. Journal of Coastal Research24, 1110–1125.
Horton, B.P., Peltier, W.R., Culver, S.J., Drummond, R., Engelhart, S.E., Kemp, A.C.,Mallinson, D., Thieler, E.R., Riggs, S.R., Ames, D.V., Thomson, K.H., 2009. Holocenesea-level changes along the North Carolina coastline and their implications forglacial isostatic adjustment models. Quaternary Science Reviews 28, 1725–1736.
Hudstedt, F., 1953. Die systematik der Diatomeen in ihren Beziehungen zur Geologieund Okologie nebst einer Revisions des Halobian-systems. Svensk BotanicalTidskrifter 47, 509–519.
Hudstedt, F., 1957.Diediatomeenflorades Fluss-systemsderWeser imGebietderHansestadtBremen. Abhandlungen der Naturwissenschatflichen Bremen 34, 181–440.
Hughen, K.A., Baillie, M.G.L., Bard, E., Bayliss, A., Beck, J.W., Bertrand, C.J.H., Blackwell,P.G., Buck, C.E., Burr, G.S., Cutler, K.B., Damon, P.E., Edwards, R.L., Fairbanks, R.G.,
Friedrich, M., Guilderson, T.P., Kromer, B., McCormac, F.G., Manning, S.W., BronkRamsey, C., Reimer, P.J., Reimer, R.W., Remmele, S., Southon, J.R., Stuiver, M.,Talamo, S., Taylor, F.W., van der Plicht, J., Weyhenmeyer, C.E., 2004. Marine04Marine radiocarbon age calibration, 25–0 ka BP. Radiocarbon 46, 1059–1086.
Jomp, J., Mallinson, D., Culver, S.J., Riggs, S., 2004. Defining the Quaternary geologicframework of the Buxton Inlet area, North Carolina. Geological Society of America,Abstracts with Programs 36 (5), 497.
Kaufman, D.S., Manley, W.F., 1998. A new procedure for determining DL amino acidratios in fossils using reverse phase liquid chromatography. Quaternary ScienceReviews (Quaternary Geochronology) 17, 987–1000.
Kemp, A.C., Horton, B.P., Culver, S.J., 2009. Distribution of modern salt-marshforaminifera in the Albemarle–Pamlico estuarine system of North Carolina, USA:implications for sea-level research. Marine Micropaleontology 72, 222–238.
Klitgord, K.D., Hutchinson, D.R., Schouten, H., 1988. U.S. Atlantic continental margin;structural and tectonic framework. In: Sheridan, R.E., Grow, J.A. (Eds.), The Geologyof North America, v.1–2, The Atlantic Continental Margin, U.S. Geological Society ofAmerica, pp. 19–55.
Litwin, R.J., Andrie, V.A.S., 1992. Modern palynomorph and weather census data fromthe U.S. Atlantic coast (Continental Margin Program) samples and selected NOAAweather stations. U.S. Geological Survey Open-file Report, pp. 92–263.
Lu, H., Fulthorpe, C.S., 2004. Controls on sequence stratigraphy of a middle Miocene–Holocene, current-swept, passive margin: offshore Canterbury Basin, New Zealand.Geological Society of America Bulletin 116, 1345–1366.
Mallinson, D., Riggs, S.R., Thieler, E.R., Foster, D.S., Culver, S.J., Corbett, D.R., Farrell, K.,Wehmiller, J.F., 2005. Late Neogene evolution of the northeastern North Carolinacoastal system: filling the Albemarle Embayment. Marine Geology 217, 97–117.
Mallinson, D., Burdette, K., Mahan, S., Brook, G., 2008. Optically stimulatedluminescence age controls on late Pleistocene and Holocene lithosomes, NorthCarolina, USA. Quaternary Research 69, 97–109.
Mallinson, D.J., Culver, S.J., Riggs, S.R., Thieler, E.R., Foster, D., Wehmiller, J., Farrell, K.,Pierson, J., 2010a. Regional seismic stratigraphy and controls on the Quaternaryevolution of the Cape Hatteras region of the Atlantic passive margin, USA. MarineGeology 268, 16–33.
Mallinson, D.J., Smith, C.W., Culver, S.J., Riggs, S.R., Ames, D.V., 2010b. Geologicalcharacteristics and spatial distribution of paleo-inlet channels beneath the OuterBanks barrier islands, North Carolina, USA. Estuarine, Coastal and Shelf Science 88,175–189.
McDowell, K., 2009. Holocene geologic development of Central Hatteras Flats andBuxton Beach ridges, Outer Banks, North Carolina. Unpublished Master's thesis,Department of Geological Sciences, East Carolina University, Greenville, NorthCarolina, USA.
Mello, J.T., Buzas, M.A., 1968. An application of cluster analysis as a method ofdetermining biofacies. Journal of Paleontology 42, 747–758.
Mudie, P.J., 1980. Palynology of later Quaternarymarine sediments, eastern Canada. Ph.D.Thesis, Dalhousie University, Halifax, Nova Scotia.
Mudie, P.J., 1982. Pollen distribution in recent marine sediments, eastern Canada.Canadian Journal of Earth Sciences 19, 729–747.
Munsterman, D., Kerstholt, S., 1996. Sodium polytungstate, a new non-toxic alternativeto bromoform in heavy liquid separation. Review of Palaeobotany and Palynology91, 417–422.
Murray, J.W., 1976. A method of determining proximity of marginal seas to an ocean.Marine Geology 22, 103–119.
Overpeck, J.T., Webb III, T., Prentice, I.C., 1985. Quantitative interpretation of fossilpollen spectra: dissimilarity coefficients and the methods of modern analogs.Quaternary Research 23, 87–108.
Parham, P., 2009. The late quaternary stratigraphy and geologic history of NortheasternNorth Carolina and Southeastern Virginia. Unpublished PhD dissertation, Depart-ment of Geological Sciences, East Carolina University, Greenville, North Carolina,USA.
Parham, P., Riggs, S., Culver, S., Mallinson, D., Wehmiller, J.F., 2007. Quaternarydepositional patterns and sea-level fluctuations, northeastern North Carolina.Quaternary Research 67, 83–99.
Poag, C.W., 1985. Benthic foraminifera as indicators of potential petroleum sources.Gulf Coast Section, Society of Economic Paleontologists and Mineralogists, FourthAnnual Research Conference Proceedings 275–284.
Poag, C.W., Knebel, H.J., Todd, R., 1980. Distribution of modern benthic foraminifers onthe New Jersey outer continental shelf. Marine Micropaleontology 5, 43–69.
Reimer, P.J., Baillie, M.G.L., Bard, E., Bayliss, A., Beck, J.W., Bertrand, C.J.H., Blackwell, P.G.,Buck, C.E., Burr, G.S., Cutler, K.B., Damon, P.E., Edwards, R.L., Fairbanks, R.G.,Friedrich, M., Guilderson, T.P., Hogg, A.G., Hughen, K.A., Kromer, B., McCormac, F.G.,Manning, S.W., Ramsey, C.B., Reimer, R.W., Remmele, S., Southon, J.R., Stuiver, M.,Talamo, S., Taylor, F.W., van der Plicht, J., Weyhenmeyer, C.W., 2004. IntCal04Terrestrial radiocarbon age calibration, 26–0 ka BP. Radiocarbon 46, 1029–1058.
Ricardo, J., 2005. Late Holocene paleoenvironmental change of the Salvo–Gull Island–Little Kinnekeet Area, Outer Banks. North Carolina. Unpublished M.S. thesis,Department of Geology, East Carolina University, Greenville, North Carolina, USA.
Ricardo, J., Culver, S.J., Mallinson, D., Riggs, S., 2004. Back-barrier/estuarine stratigraphyand recent morphological evolution of Hatteras Flats, Gull Island vicinity, OuterBanks, North Carolina. Geological Society of America, Abstracts with Programs 36(5), 290.
Riggs, S.R., York, L.L., Wehmiller, J.F., Snyder, S.W., 1992. Depositional patterns resultingfrom high-frequency Quaternary sea-level fluctuations in northeastern NorthCarolina. Quaternary Coasts of the United States: Marine and Lacustrine Systems,SEPM Special Publication 48, 141–153.
Robinson, M.M., McBride, R.A., 2006. Benthic foraminifera from a relict flood tidal deltaalong the Virginia/North Carolina Outer Banks. Micropaleontology 50, 67–80.

249S.J. Culver et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 305 (2011) 227–249
Sager, E.D., Riggs, S.R., 1998. Models for the Holocene valley-fill history of AlbemarleSound, North Carolina. USA. In: Tidalites: Processes and Products. SEPM SpecialPublication 61, 119–127.
Schnitker, D., 1971. Distribution of foraminifera on the North Carolina continental shelf.Tulane Studies in Geology and Paleontology 8, 169–215.
Scott, D.B., Medioli, F.S., 1980. Quantitative studies of marsh foraminiferal distributionsin Nova Scotia: implications for sea level studies. Cushman Foundation forForaminiferal Research, Special Publication 17, 1–58.
Sen Gupta, B.K., Machain-Castillo, M.L., 1993. Benthic foraminifera in oxygen-poorhabitats. Marine Micropaleontology 20, 183–201.
Smith, C.G., Culver, S.J., Mallinson, D.J., Riggs, S.R., Corbett, D.R., 2009. Recognizingformer flood tidal delta deposits in the late Holocene stratigraphic record:examples from the Outer Banks of North Carolina, USA. Stratigraphy 6, 61–78.
Snyder, S.W., Waters, V.J., Moore, T.L., 1988. Benthic foraminifera and paleoecology ofMiocene Pungo River formation sediments in Onslow Bay, North Carolinacontinental shelf. In: Snyder, S.W. (Ed.), Micropaleontology of Miocene sedimentsin the shallow subsurface of Onslow Bay, North Carolina continental shelf.Cushman Foundation for Foraminiferal Research Special Publication 25, pp. 43–96.
Stanton, C.L.T., 2008. Holocene inner continental shelf stratigraphy, micropaleontologyand paleoenvironmental change off the Outer Banks. North Carolina. UnpublishedM.S. thesis, Department of Geology, East Carolina University, Greenville, NorthCarolina, USA.
Stuiver, M., Reimer, P.J., 1993. Extended 14C database and revised CALIB radiocarboncalibration program. Radiocarbon 35, 215–230.
Traverse, A., 1988. Paleopalynology. Unwin-Hyman, Boston.Van der Werff, H., Huls, H., 1958–1966. Diatomeenflora van Nederland. Published
privately, De Haef, The Netherlands. .Vance, D.J., Culver, S.J., Corbett, D.R., Buzas, M.A., 2006. Foraminifera in the Albemarle
estuarine system, North Carolina: distribution and recent environmental change.Journal of Foraminiferal Research 36, 15–33.
Vos, P.C., de Wolf, H., 1993. Methodological aspects of paleoecological diatom researchin coastal areas of the Netherlands. Geologie en Mijnbouw 67, 31–40.
Ward, L.W., Strickland, G.L., 1985. Outline of Tertiary stratigraphy and depositionalhistory of the U.S. Atlantic margin. In: Poag, C.W. (Ed.), Geologic Evolution of theUnited States Atlantic Margin. Van Nostrand Reinhold, New York, pp. 125–188.
Wehmiller, J.F., Miller, G.H., 2000. Aminostratigraphic dating methods in Quaternarygeology. In: Noller, J. S., Sowers, J. M., Lettis, W. R. (Eds.), Quaternary Geochronology,Methods and Applications. American Geophysical Union Reference Shelf 4, 187–222.
Wehmiller, J.F., Thieler, E.R., Miller, D., Pellerito, V., Bakeman Keeney, V.R., Riggs, S.R.,Culver, S., Mallinson, D., Farrell, K., York, L.L., Pierson, J., Parham, P., 2010a.Aminostratigraphy of surface and subsurface Quaternary units, Coastal Plain ofNorth Carolina, USA. Quaternary Geochronology 5, 459–492.
Wehmiller, J.F., Harris, W.B., Farrell, K.M., 2010b. Evaluation of racemization-derivedage estimates using strontium 87/86 values in mollusks from early and middlePleistocene units, North Carolina coastal plain. Geological Society of America,Abstracts with Programs 42 (1), 68.
Willard, D.A., Cronin, T.M., Verardo, S., 2003. Late-Holocene climate and ecosystemhistory from Chesapeake Bay sediment cores, USA. Holocene 13, 201–214.
Willard, D.A., Bernhardt, C.E., Korejwo, D.A., Meyers, S.R., 2005. Impact of millennial-scale Holocene climatic variability on eastern North American terrestrial ecosys-tems: pollen-based climatic reconstruction. Global and Planetary Change 47, 17–35.
Winker, C.D., Howard, J.D., 1977. Correlation of tectonically deformed shorelines on thesouthern Atlantic coastal plain. Geology 5, 123–127.
Woo, J.H., Culver, S.J., Oertel, G.F., 1997. Benthic foraminiferal communities of a barrier–lagoon system, Virginia, U.S.A. Journal of Coastal Research 13, 1192–1200.
Workman, R.R., 1981. Foraminiferal assemblages of the nearshore inner continentalshelf, Nags Head and Wilmington areas. North Carolina. Unpublished M.S. thesis,Department of Geology, East Carolina University, Greenville, North Carolina, USA.
York, L.L., Wehmiller, J.F., Cronin, T.M., Ager, T.A., 1989. Stetson Pit, Dare County, NorthCarolina: an integrated chronologic, faunal, and floral record of subsurface coastalsediments. Palaeogeography, Palaeoclimatology, Palaeoecology 72, 115–132.
Zong, Y., Horton, B.P., 1999. Diatom-based tidal-level transfer functions as an aid inreconstructing Quaternary history of sea-level movements in Britain. Journal ofQuaternary Science 14, 153–167.




















