
Metal accumulation in earthworms inhabiting floodplain soils
9
Metal accumulation in earthworms inhabiting floodplain soils Martina G. Vijver a,b, * , Jos P.M. Vink b , Cornelis J.H. Miermans b , Cornelis A.M. van Gestel a a Institute of Ecological Science, Dept. Animal Ecology, Vrije Universiteit, Amsterdam, The Netherlands b Institute for Inland Water Management and Waste Water Treatment (RIZA), Dept. Chemistry and Ecotoxicology, Lelystad, The Netherlands Received 27 July 2006; received in revised form 16 October 2006; accepted 22 October 2006 Metal levels in earthworms show large variation among sites, among seasons and among epigeic and endogeic species. Abstract The main factors contributing to variation in metal concentrations in earthworms inhabiting floodplain soils were investigated in three flood- plains differing in inundation frequency and vegetation type. Metal concentrations in epigeic earthworms showed larger seasonal variations than endogeic earthworms. Variation in internal levels between sampling intervals were largest in earthworms from floodplain sites frequently inun- dated. High and low frequency flooding did not result in consistent changes in internal metal concentrations. Vegetation types of the floodplains did not affect metal levels in Lumbricus rubellus, except for internal Cd levels, which were positively related to the presence of organic litter. Internal levels of most essential metals were higher in spring. In general, no clear patterns in metal uptake were found and repetition of the sampling campaign will probably yield different results. Ó 2006 Elsevier Ltd. All rights reserved. Keywords: Field results; Soil organisms; Floodplain systems; Metals; Earthworms 1. Introduction It is relatively unknown how metal levels in organisms in- habiting floodplains alter throughout the year. Despite high contamination levels, floodplain soil properties, such as high pH and high organic matter content, cause low soluble metal concentrations (Janssen et al., 1997a; Sauve ´ et al., 2000; Van Leeuwen, 1999). Soluble metal is believed to be an impor- tant source of exposure for most organisms (Van Gestel, 1997), especially for organisms having a water permeable epi- dermis (Vijver et al., 2003a,b). However, elevated metal con- centrations in earthworms were detected in Dutch floodplains, whereas isopods and millipedes (having a less wa- ter-permeable epidermis) showed similar metal levels as in species from unpolluted reference areas (Hobbelen et al., 2004). The explanation of this controversy remained indis- tinct, probably because of the involvement of many interfering factors. In literature, field studies in which internal and external metal concentrations are measured usually have a static char- acter, while floodplain ecosystems by definition are subjected to fluctuating conditions. Inundation, which is by definition a dynamic process, is known to influence metal speciation in soils (Vink and Meeussen, 2007). We tested the hypothesis that this variation in speciation is also reflected in bioaccumu- lation. To investigate dynamics of metal bioaccumulation, the epigeic Lumbricus rubellus, and the endogeic species Allolo- bophora chlorotica and Aporrectodea caliginosa were moni- tored. The epigeic species have their habitat in the top layers of soil and in litter layers, whereas the habitat of endogeic spe- cies is predominantly in deeper soil layers (Bouche ´, 1977). Three different floodplain sites were selected, based on differ- ences in inundation frequency and vegetation types. Internal metal concentrations in earthworms were related to external factors, such as soil properties, metal concentrations, river * Corresponding author. Present address: Institute of Environmental Sci- ences (CML), Department of Environmental Biology, P.O. Box 9518, 2300 RA Leiden, The Netherlands. Tel.: þ31 71 527 1487; fax: þ31 71 527 5587. E-mail address: [email protected] (M.G. Vijver). 0269-7491/$ - see front matter Ó 2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.envpol.2006.10.046 Environmental Pollution 148 (2007) 132e140 www.elsevier.com/locate/envpol
-
Upload
leidenuniv -
Category
Documents
-
view
5 -
download
0
Transcript of Metal accumulation in earthworms inhabiting floodplain soils

Environmental Pollution 148 (2007) 132e140www.elsevier.com/locate/envpol
Metal accumulation in earthworms inhabiting floodplain soils
Martina G. Vijver a,b,*, Jos P.M. Vink b, Cornelis J.H. Miermans b, Cornelis A.M. van Gestel a
a Institute of Ecological Science, Dept. Animal Ecology, Vrije Universiteit, Amsterdam, The Netherlandsb Institute for Inland Water Management and Waste Water Treatment (RIZA), Dept. Chemistry and Ecotoxicology, Lelystad, The Netherlands
Received 27 July 2006; received in revised form 16 October 2006; accepted 22 October 2006
Metal levels in earthworms show large variation among sites, among seasons and among epigeic and endogeic species.
Abstract
The main factors contributing to variation in metal concentrations in earthworms inhabiting floodplain soils were investigated in three flood-plains differing in inundation frequency and vegetation type. Metal concentrations in epigeic earthworms showed larger seasonal variations thanendogeic earthworms. Variation in internal levels between sampling intervals were largest in earthworms from floodplain sites frequently inun-dated. High and low frequency flooding did not result in consistent changes in internal metal concentrations. Vegetation types of the floodplainsdid not affect metal levels in Lumbricus rubellus, except for internal Cd levels, which were positively related to the presence of organic litter.Internal levels of most essential metals were higher in spring. In general, no clear patterns in metal uptake were found and repetition of thesampling campaign will probably yield different results.� 2006 Elsevier Ltd. All rights reserved.
Keywords: Field results; Soil organisms; Floodplain systems; Metals; Earthworms
1. Introduction
It is relatively unknown how metal levels in organisms in-habiting floodplains alter throughout the year. Despite highcontamination levels, floodplain soil properties, such as highpH and high organic matter content, cause low soluble metalconcentrations (Janssen et al., 1997a; Sauve et al., 2000;Van Leeuwen, 1999). Soluble metal is believed to be an impor-tant source of exposure for most organisms (Van Gestel,1997), especially for organisms having a water permeable epi-dermis (Vijver et al., 2003a,b). However, elevated metal con-centrations in earthworms were detected in Dutchfloodplains, whereas isopods and millipedes (having a less wa-ter-permeable epidermis) showed similar metal levels as inspecies from unpolluted reference areas (Hobbelen et al.,
* Corresponding author. Present address: Institute of Environmental Sci-
ences (CML), Department of Environmental Biology, P.O. Box 9518, 2300
RA Leiden, The Netherlands. Tel.: þ31 71 527 1487; fax: þ31 71 527 5587.
E-mail address: [email protected] (M.G. Vijver).
0269-7491/$ - see front matter � 2006 Elsevier Ltd. All rights reserved.
doi:10.1016/j.envpol.2006.10.046
2004). The explanation of this controversy remained indis-tinct, probably because of the involvement of many interferingfactors.
In literature, field studies in which internal and externalmetal concentrations are measured usually have a static char-acter, while floodplain ecosystems by definition are subjectedto fluctuating conditions. Inundation, which is by definitiona dynamic process, is known to influence metal speciation insoils (Vink and Meeussen, 2007). We tested the hypothesisthat this variation in speciation is also reflected in bioaccumu-lation. To investigate dynamics of metal bioaccumulation, theepigeic Lumbricus rubellus, and the endogeic species Allolo-bophora chlorotica and Aporrectodea caliginosa were moni-tored. The epigeic species have their habitat in the top layersof soil and in litter layers, whereas the habitat of endogeic spe-cies is predominantly in deeper soil layers (Bouche, 1977).Three different floodplain sites were selected, based on differ-ences in inundation frequency and vegetation types. Internalmetal concentrations in earthworms were related to externalfactors, such as soil properties, metal concentrations, river

133M.G. Vijver et al. / Environmental Pollution 148 (2007) 132e140
water composition, seasonal changes in vegetation, and inun-dation frequency.
2. Materials and methods
2.1. Research sites and abiotic parameters
Three sites along the rivers Nieuwe Merwede (coded M and P) and Waal
(coded S) were selected. River Waal switches over to the Nieuwe Merwede, so
comparable water characteristics are found. Geographical information on the
sites and vegetation types are summarized in Table 1.
Cattle grazed the sites M, P and S. The sites are located on gradually slop-
ing riverbanks, and are subjected to periodic inundation. Plots from which
earthworms were sampled, measured 10 by 10 meter. Fluctuations in water
level and inundation frequency are shown in Fig. 1.
Inundation patterns in the fresh water estuary floodplain M and P are influ-
enced by sea tides and wind, an example of a typical cycle is given in Fig. 1
(inset). Water levels are monitored on an hourly basis by Rijkswaterstaat and
are available on http://www.waterbase.nl. At all sites we measured inundation
frequency, surface water composition of both rivers, and soil and pore water
properties, metal concentrations in soil, pore water, litter, and organisms.
Soil characteristics and total metal concentrations were determined in Novem-
ber 1999 and August 2000. Pore water was collected using different tech-
niques. At all three sites, pore water was collected by centrifugation, and
filtered over 0.45 mm before chemical analysis. In addition, at site M pore wa-
ter was monitored in situ regularly from November 2000 to December 2001,
using a permeable pore water sampler (Rhizon SMS-MOM, Rhizosphere Re-
search Products, Wageningen, The Netherlands). Organic litter was collected
three times in 2003 and 2004 for metal analyses. Due to logistic reasons, sam-
pling was done at different times.
2.2. Collection of organisms
From November 2000 to February 2004, earthworms of the species Lum-
bricus rubellus were collected from the floodplain sites M and P at three to five
months intervals. Endogenic earthworm species were collected from Novem-
ber 2000 to May 2002, also site S was visited during these years. At all sites
collection was random within the 10 � 10 m plot. At least 5 individuals of
each pedo-ecological group were taken. After taking to the laboratory, the
earthworms were allowed to defaecate on wet filter paper for 48 h. Wet
weights were determined, followed by storage in a freezer at �18 �C until
analyses.
2.3. Chemical analyses
Animals and litter were freeze-dried and dry weights were determined. Ly-
ophilized organisms were digested in concentrated HNO3 solution using
a Mars5 destruction microwave oven and measured by ICP-MS (Perkin Elmer,
SciEx ELAN 6000, Concord, Ontario, Canada). Soil samples were dried,
sieved (<2 mm), and freeze dried before analysis at �50 �C for 18 h at
�0.04 kPa. As, Cd, Cr, Cu, Pb, Zn, Mn, Fe concentrations in aqua regia
(NEN 6465, 1992) were measured by ICP-AES (Spectros, Specto Flame)
and ICP-MS. Certified reference material Dolt-2 (BCR, Brussels, Belgium),
and blancs were treated similar as the samples. No systematic correction
was applied to the body residue analyses, since recovery of the standard addi-
tion was within acceptance limits (90e110%). Measured metal concentrations
of the standard reference were within performance acceptance limits (95e
105%). The organic carbon content of the soils was analysed by wet oxidation
with K2Cr2O7 (Wallinga et al., 1992). Clay content was measured by sedimen-
tation according to Houba et al. (1996).
2.4. Statistics
Internal metal concentrations of organisms were tested for outliers using
the Grubbs test (Funk et al., 1985). The test is used to detect data outliers
from a univariate and normally distributed data set. For each group of animal
species, homogeneity of data was tested using Fmax, followed by analysis of
covariance (ANCOVA) to correct metal accumulation for body weight (de-
pendent ¼ internal concentration and covariable ¼ weight and factor ¼ time).
For each group of animal species, one-way ANOVA tested the significance of
time of measurement (dependent ¼ internal concentration and factor ¼ time).
This test was followed by a post-hoc test (Tukey) to analyse for significance of
the differences that can be attributed to seasonality, specifically ( p < 0.05).
All statistics were performed using the software packages Systat 9.0 and
SPSS 10.0.
3. Results
Sites M, P, and S differed in inundation frequency (Fig. 1).Due to the influence of sea tides, M and P were flooded peri-odically during the year, but were only permanently inundatedduring some winter periods. Flooding of site S was long-last-ing and occurred usually in winter and early spring, and lastedfor up to three months, depending on rainfall.
Water composition did not differ between the rivers Waaland Nieuwe Merwede. Soil characteristics and total metal con-centrations are summarized in Table 2. Pollution levels of thethree floodplain soils are specified as pollution class 4. Thismeans that concentrations of one or more metals exceed theregulatory standards according to the Dutch regulations (4thNational Policy Document). To compare metal levels withthe quality criteria, a correction for clay and organic mattercontent was applied as described by Vegter (1995).
Site P is the most polluted location followed by site M andsite S. Concentrations of Zn, Cd, Cu, Cr and Pb exceed the in-tervention values in most cases at sites M and P, and targetvalues at site S. The metals exceeding these criteria aremarked in Table 2. A summary of the pore water compositionsat the different sites is given in Table 3.
Compared to the control and reference soils, only Cd con-centrations in pore water of site M were elevated (Hobbelenet al., 2004). Pore water concentrations of all other metalswere within boundary range of reference values. Both methodsused for pore water sampling, i.e. rhizon and centrifugation,
Table 1
Characteristics of the floodplain sites selected for determining dynamics of metal concentrations in earthworms
Code Location x-coordinates y-coordinates Height above NAP (cm) Rivers Vegetation type
M Lage Hof 111.314 420.428 80 Nieuwe Merwede Grassland
P Ruitersplaat 111.235 420.000 100 Nieuwe Merwede Mixed vegetation
S Stiftse Waard 155.240 427.360 457 Waal Grassland
x and y coordinates were determined using a GPS-system calibrated at Amersfoort, The Netherlands. NAP ¼ Dutch reference mean sea level. Mixed vegetation
includes reed Phragmites australis, herbs e.g., Urtica dioica, Symphytum officinale and Valeriana officinalis and shrub species, e.g., Salix. The grassland vegetation
mainly consists of grass and some Phragmites australis.
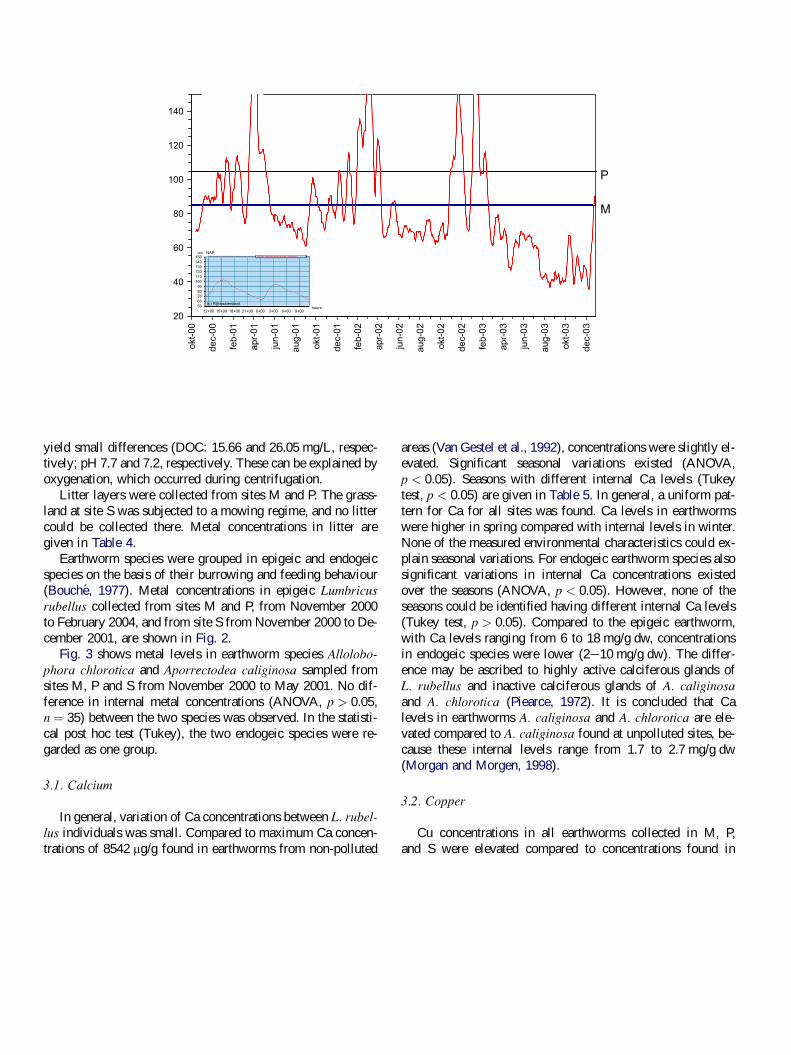
Table 2Soil properties and total metal concentrations at thethree floodplainsites expressed on a dry weight basisSite OM [%]<
2
m
m [%] Ph 0.01 M CaCl
[mg/kg] Cd [mg/kg] Cr [mg/kg] Cu [mg/kg] Pb [mg/kg] Zn[mg/kg] Fe [g/kg] Ca [g/kg]
M 10 5 3245 7.61441658
507
316 561 2308
4141 4249
P 1120 340 7.21951845 527 343
5512444
n.d.n.d.
S 241 1 7.5 7442
1.79 7550 430 116 354 052 43.7
Metal concentrations exceeding the Dutch target values for soil areitalic, concentrations exceeding the intervention values arebold. Testing to these quality stan-dards correction for organic matter and clay content as described byVegter (1995)
.
134
M.G. Vijver et al. / Environmental Pollution 148 (2007)132e
20
40
60
80
100
120
140
okt-00
dec-00
feb-01
apr-01
jun-01
aug-01
okt-01
dec-01
feb-02
apr-02
jun-02
aug-02
okt-02
dec-02
feb-03
apr-03
jun-03
aug-03
okt-03
dec-03
P
M
s yield small differences (DOC: 15.66 and 26.05 mg/L, respec-tively; pH 7.7 and 7.2, respectively. These can be explained byoxygenation, which occurred during centrifugation.Litter layers were collected from sites M and P. The grass-land at site S was subjected to a mowing regime, and no littercould be collected there. Metal concentrations in litter aregiven in Table 4.
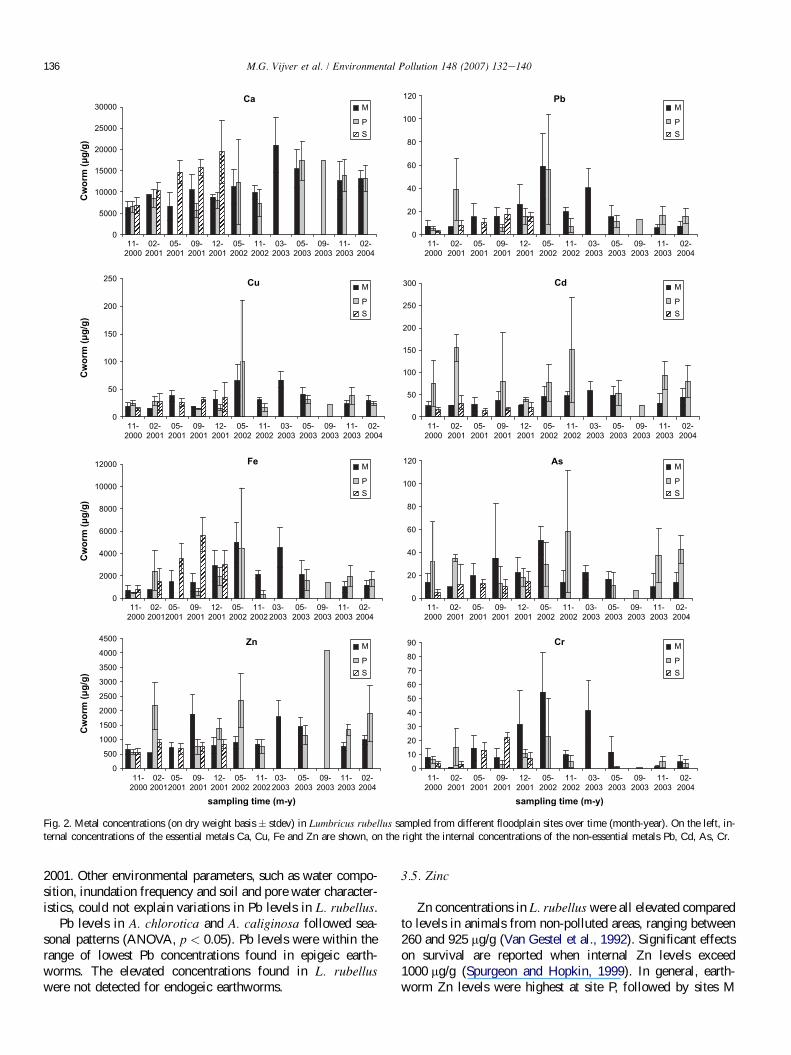
Earthworm species were grouped in epigeic and endogeicspecies on the basis of their burrowing and feeding behaviour(Bouche, 1977). Metal concentrations in epigeic Lumbricusrubellus collected from sites M and P, from November 2000to February 2004, and from site S from November 2000 to De-cember 2001, are shown in Fig. 2.
Fig. 3 shows metal levels in earthworm species Allolobo-phora chlorotica and Aporrectodea caliginosa sampled fromsites M, P and S from November 2000 to May 2001. No dif-ference in internal metal concentrations (ANOVA, p > 0.05,n ¼ 35) between the two species was observed. In the statisti-cal post hoc test (Tukey), the two endogeic species were re-garded as one group.
3.1. Calcium
In general, variation of Ca concentrations between L. rubel-lus individuals was small. Compared to maximum Ca concen-trations of 8542 mg/g found in earthworms from non-polluted
2 areas (Van Gestel et al., 1992), concentrations were slightly el-evated. Significant seasonal variations existed (ANOVA,p < 0.05). Seasons with different internal Ca levels (Tukeytest, p < 0.05) are given in Table 5. In general, a uniform pat-tern for Ca for all sites was found. Ca levels in earthwormswere higher in spring compared with internal levels in winter.None of the measured environmental characteristics could ex-plain seasonal variations. For endogeic earthworm species alsosignificant variations in internal Ca concentrations existedover the seasons (ANOVA, p < 0.05). However, none of theseasons could be identified having different internal Ca levels(Tukey test, p > 0.05). Compared to the epigeic earthworm,with Ca levels ranging from 6 to 18 mg/g dw, concentrationsin endogeic species were lower (2e10 mg/g dw). The differ-ence may be ascribed to highly active calciferous glands ofL. rubellus and inactive calciferous glands of A. caliginosaand A. chlorotica (Piearce, 1972). It is concluded that Calevels in earthworms A. caliginosa and A. chlorotica are ele-vated compared to A. caliginosa found at unpolluted sites, be-cause these internal levels range from 1.7 to 2.7 mg/g dw(Morgan and Morgen, 1998).
3.2. Copper
Cu concentrations in all earthworms collected in M, P,and S were elevated compared to concentrations found in
A
135M.G. Vijver et al. / Environmental Pollution 148 (2007) 132e140
Table 3
Metal concentrations in pore water sampled at floodplain sites M, P and S
Site Sampling
method
Date
(month -yr)
DOC
[mg/L]
pH [�] Ca
[mg/L]
Fe
[mg/L]
Cu
[mg/L]
Zn
[mg/L]
Cd
[mg/L]
Pb
[mg/L]
As
[mg/L]
Cr
[mg/L]
M Rhizon Nov-01 24.4 7.52* 164 582 28.3 482 3.00 2.60 18.6 3.2
M Feb-01 13.7 7.03 142 767 22.6 720 5.67 1.94 15.7 4.2
M May-01 12.5 7.34 160 1144 22.3 580 5.69 1.26 17.5 2.3
M Sep-01 16.5 6.87 187 d.l. 53.6 759 7.48 1.39 17.1 6.1
M Dec-01 11.2 7.07 172 382 27.4 727 5.52 2.11 15.0 5.8
M Centrifugation Aug-00 28.8 7.88 135 d.l. 57.1 168 2.92 1.56 12.6 n.d.
M Feb-01 23.3 7.51 109 d.l. 42.2 308 3.71 1.28 9.81 n.d.
P Aug-00 23.5 7.52 111 d.l. 57.1 225 2.74 1.87 8.82 n.d.
S Feb-01 20.5 7.69 88 d.l. 13.6 144 0.60 0.62 4.28 n.d.
The concentrations that exceed Dutch quality standards are in bold. n.d. ¼ not determined, d.l. ¼ below detection limit, * measured in laboratory instead of in situ.
L. rubellus from non-polluted soil (14.7 � 5.2 mg/g; Irelandand Richards, 1977). Effects on survival of L. rubellus arefound when internal Cu levels exceed 55 mg/g (Langdonet al., 2001). This concentration was exceeded in earthwormssampled in May 2002 (sites M and P), and in March 2003 (siteM). Significant differences (ANOVA, p < 0.05) were foundwithin earthworms collected from the three sites. Large bio-logical variation (Fmax > Fcrit) existed between organismscollected from site M in May 2002, probably because of inter-nal concentrations that are close to effect levels. Analysis ofvariance (see Fmax) could not be performed due to variationof levels within sampling times, which is reflected in largestandard error exceeding the mean. Epigeic earthworms col-lected in February 2001 and May 2002 (site P) also had a bio-logical variation that was larger than the variation between thedifferent groups. Detailed seasonal analysis of internal Cu con-centrations from the sites revealed the same pattern as foundfor Ca. Especially in spring and summer, higher internal con-centrations were found. None of the external characteristicscould explain Cu concentrations in L. rubellus. Even differ-ences in total metal concentrations (a factor of 7 to 8) betweensoil M and P compared to site S, were not reflected in internallevels. No significant variation (ANOVA, p > 0.05) in Culevels was found over the season for endogeic earthworms.
3.3. Iron
Average Fe concentrations in L. rubellus ranged from343 mg/g (P, Nov 2002) to 5664 mg/g (S, sept 2001). Signifi-cant seasonal variations (ANOVA, p < 0.05, post hoc Tukey)were found in earthworms collected from site M and site S.Seasons that were identified to induce altered internal Fe levels
are given in Table 5. In contrast, earthworms sampled at site Pshowed no significant seasonal differences (ANOVA,p ¼ 0.066). The absence of seasonal variations in Fe concen-trations of L. rubellus inhabiting site P may be explained bythe presence of a metal-binding litter layer. Total metal con-centrations in soil could not explain differences, neither couldinundation frequency or water composition. DOC concentra-tions in pore water showed variation over the seasons. DOCconcentrations measured in November 2000 were two times2 higher than in December 2001.
Fe concentrations in endogeic earthworms were compara-ble to those in epigeic earthworms. Internal Fe concentrationsin A. chlorotica and A. caliginosa varied with time (ANOVA,p < 0.05). This pattern is in agreement with the trend found inL. rubellus.
3.4. Lead
Pb concentrations in L. rubellus were mostly within therange of concentrations (24.7 � 6.2 mg/g) found in earth-worms collected from non-polluted areas (Ireland and Ri-chards, 1977). Significant differences over time (ANOVA,p < 0.05) were found within earthworms. Table 5 presentsthe time periods in which changes in internal concentrationswere observed. Internal Pb gave statistical relationship toDOC concentrations measured in pore water. Note that accu-mulation is the net effect of uptake, elimination and storage(Vijver, 2005), and therefore not necessarily reflects the mech-anism of uptake. Pb is known to have a high affinity to DOC(Christensen et al., 1999), and pore water samples that weretaken in November 2000 had DOC concentrations that weretwice as high as samples taken in September and December
Table 4
Metal concentrations (dry weight � stdev, n ¼ 3) in the litter layer of the floodplain sites M and P
Site Time
(month-yr)
As [mg/kg] Cd [mg/kg] Cr [mg/kg] Cu [mg/kg] Pb [mg/kg] Zn [mg/kg] Fe [mg/kg] Ca [mg/kg]
M 05-2003 27.3 � 0.98 4.79 � 0.16 153 � 5.63 110 � 3.50 124 � 3.87 818 � 34.2 26,445 � 991 33,827 �11-2003 38.6 � 0.93 5.66 � 0.19 162 � 6.07 115 � 2.68 143 � 4.36 895 � 8.21 22,045 � 829 43,635 � 2834
02-2004 69.2 � 7.54 9.00 � 0.73 290 � 29.8 189 � 20.2 239 � 27.3 1334 � 129 31,624 � 3222 46,590 � 3556
P 05-2003 37.2 � 4.63 5.74 � 0.64 118 � 12.8 85.6 � 8.01 156 � 18.3 1226 � 133 21,577 � 2313 59,076 � 2978
11-2003 43.8 � 17.4 10.5 � 3.70 169 � 60.7 120 � 41.2 185 � 68.2 1617 � 599 14,837 � 5144 61,572 � 21,599
02-2004 49.4 � 7.18 7.70 � 1.25 156 � 24.9 120 � 19.6 193 � 30.1 1491 � 225 19,810 � 3110 38,149 � 6995
136 M.G. Vijver et al. / Environmental Pollution 148 (2007) 132e140
Zn
0
500
1000
1500
2000
2500
3000
3500
4000
4500
sampling time (m-y) sampling time (m-y)
Cd
0
50
100
150
200
250
300
Pb
0
20
40
60
80
100
120
Cu
0
50
100
150
200
250
Fe
0
2000
4000
6000
8000
10000
12000
Ca
0
5000
10000
15000
20000
25000
30000
11-2000
02-2001
05-2001
09-2001
12-2001
05-2002
11-2002
03-2003
05-2003
09-2003
11-2003
02-2004
11-2000
02-2001
05-2001
09-2001
12-2001
05-2002
11-2002
03-2003
05-2003
09-2003
11-2003
02-2004
11-2000
02-2001
05-2001
09-2001
12-2001
05-2002
11-2002
03-2003
05-2003
09-2003
11-2003
02-2004
11-2000
02-2001
05-2001
09-2001
12-2001
05-2002
11-2002
03-2003
05-2003
09-2003
11-2003
02-2004
11-2000
02-2001
05-2001
09-2001
12-2001
05-2002
11-2002
03-2003
05-2003
09-2003
11-2003
02-2004
11-2000
02-2001
05-2001
09-2001
12-2001
05-2002
11-2002
03-2003
05-2003
09-2003
11-2003
02-2004
11-2000
02-2001
05-2001
09-2001
12-2001
05-2002
11-2002
03-2003
05-2003
09-2003
11-2003
02-2004
11-2000
02-2001
05-2001
09-2001
12-2001
05-2002
11-2002
03-2003
05-2003
09-2003
11-2003
02-2004
Cw
orm
(µ
g/g
)C
wo
rm
(µ
g/g
)C
wo
rm
(µ
g/g
)C
wo
rm
(µ
g/g
)
As
0
20
40
60
80
100
120
Cr
0
10
20
30
40
50
60
70
80
90
M
PS
M
PS
M
PS
M
PS
M
PS
M
PS
M
PS
M
PS
Fig. 2. Metal concentrations (on dry weight basis � stdev) in Lumbricus rubellus sampled from different floodplain sites over time (month-year). On the left, in-
ternal concentrations of the essential metals Ca, Cu, Fe and Zn are shown, on the right the internal concentrations of the non-essential metals Pb, Cd, As, Cr.
2001. Other environmental parameters, such as water compo-sition, inundation frequency and soil and pore water character-istics, could not explain variations in Pb levels in L. rubellus.
Pb levels in A. chlorotica and A. caliginosa followed sea-sonal patterns (ANOVA, p < 0.05). Pb levels were within therange of lowest Pb concentrations found in epigeic earth-worms. The elevated concentrations found in L. rubelluswere not detected for endogeic earthworms.
3.5. Zinc
Zn concentrations in L. rubellus were all elevated comparedto levels in animals from non-polluted areas, ranging between260 and 925 mg/g (Van Gestel et al., 1992). Significant effectson survival are reported when internal Zn levels exceed1000 mg/g (Spurgeon and Hopkin, 1999). In general, earth-worm Zn levels were highest at site P, followed by sites M

137M.G. Vijver et al. / Environmental Pollution 148 (2007) 132e140
Pb
0
10
20
30
40
50Ca
0
2000
4000
6000
8000
10000
12000
14000
nov-00
jan-01
mrt-01
mei-01
jul-01
sep-01
nov-01
jan-02
mrt-02
mei-02
nov-00
jan-01
mrt-01
mei-01
jul-01
sep-01
nov-01
jan-02
mrt-02
mei-02
nov-00
jan-01
mrt-01
mei-01
jul-01
sep-01
nov-01
jan-02
mrt-02
mei-02
nov-00
jan-01
mrt-01
mei-01
jul-01
sep-01
nov-01
jan-02
mrt-02
mei-02
nov-00
jan-01
mrt-01
mei-01
jul-01
sep-01
nov-01
jan-02
mrt-02
mei-02
nov-00
jan-01
mrt-01
mei-01
jul-01
sep-01
nov-01
jan-02
mrt-02
mei-02
nov-00
jan-01
mrt-01
mei-01
jul-01
sep-01
nov-01
jan-02
mrt-02
mei-02
nov-00
jan-01
mrt-01
mei-01
jul-01
sep-01
nov-01
jan-02
mrt-02
mei-02
(u
g/g
)(u
g/g
)
(u
g/g
)(u
g/g
)(u
g/g
)(u
g/g
)
(u
g/g
)(u
g/g
)
Fe
0
2000
4000
6000
8000
Cd
0
50
100
150
Cu
0
20
40
60
80
Cr
0
10
20
30
40
50Zn
0
200
400
600
800
1000
1200
1400
1600
1800
2000
As
05101520253035404550
MPS
MPS
MPS
MPS
MPS
MPS
MPS
MPS
Fig. 3. Metal concentrations (on dry weight basis � stdev) in Allolobophora chlorotica and Aporrectodea caliginosa sampled from different floodplain sites over
time (month-year). On the left, essential metals are shown, on the right the non-essential metals.
and S. Significant differences over time (ANOVA, p < 0.05)were found within earthworms collected from all sites. Resultsare summarized in Table 5. The general trend is that internalZn concentrations in earthworms correlated with pore watermetal concentrations together with pH. Internal levels weresignificantly higher in February, March, May and Septembercompared to other months. Environmental factors other than
pore water concentrations could not explain the variations ob-served in earthworm Zn levels.
3.6. Cadmium
Cd concentrations in L. rubellus ranged from 12.6 mg/g (S,May 2001) to 150 mg/g (P, Feb 2001 and Nov 2002), far
138 M.G. Vijver et al. / Environmental Pollution 148 (2007) 132e140
Table 5
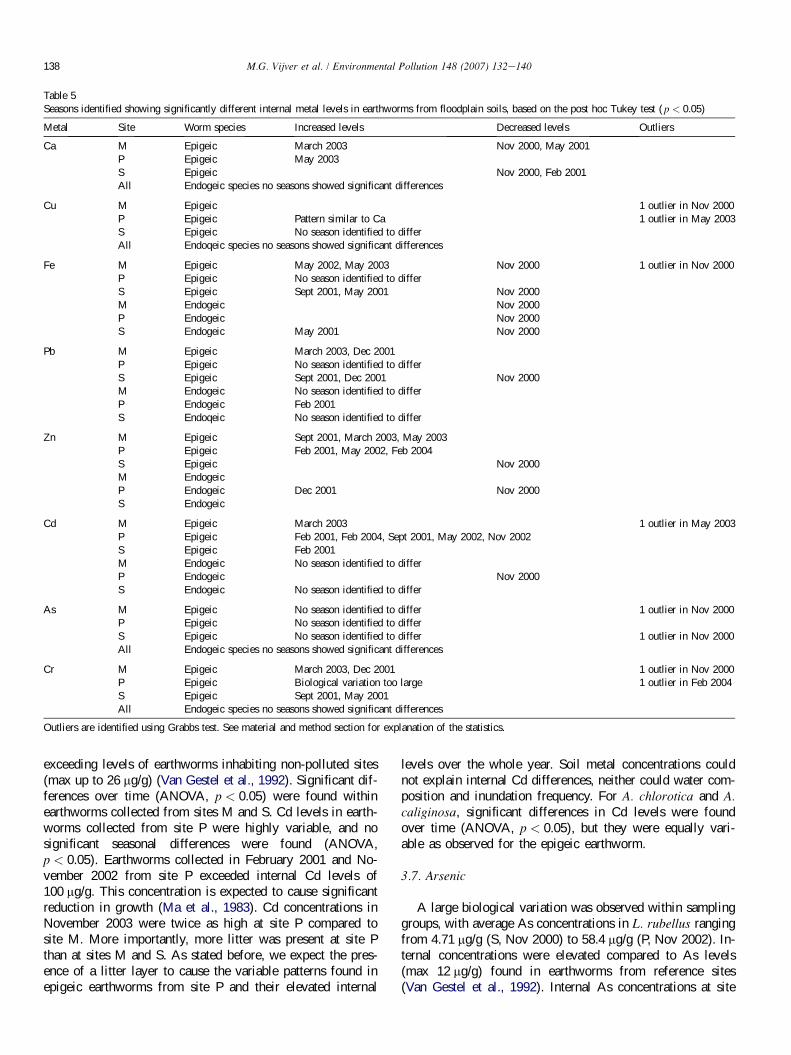
Seasons identified showing significantly different internal metal levels in earthworms from floodplain soils, based on the post hoc Tukey test ( p < 0.05)
Metal Site Worm species Increased levels Decreased levels Outliers
Ca M Epigeic March 2003 Nov 2000, May 2001
P Epigeic May 2003
S Epigeic Nov 2000, Feb 2001
All Endogeic species no seasons showed significant differences
Cu M Epigeic 1 outlier in Nov 2000
P Epigeic Pattern similar to Ca 1 outlier in May 2003
S Epigeic No season identified to differ
All Endoqeic species no seasons showed significant differences
Fe M Epigeic May 2002, May 2003 Nov 2000 1 outlier in Nov 2000
P Epigeic No season identified to differ
S Epigeic Sept 2001, May 2001 Nov 2000
M Endogeic Nov 2000
P Endogeic Nov 2000
S Endogeic May 2001 Nov 2000
Pb M Epigeic March 2003, Dec 2001
P Epigeic No season identified to differ
S Epigeic Sept 2001, Dec 2001 Nov 2000
M Endogeic No season identified to differ
P Endogeic Feb 2001
S Endoqeic No season identified to differ
Zn M Epigeic Sept 2001, March 2003, May 2003
P Epigeic Feb 2001, May 2002, Feb 2004
S Epigeic Nov 2000
M Endogeic
P Endogeic Dec 2001 Nov 2000
S Endogeic
Cd M Epigeic March 2003 1 outlier in May 2003
P Epigeic Feb 2001, Feb 2004, Sept 2001, May 2002, Nov 2002
S Epigeic Feb 2001
M Endogeic No season identified to differ
P Endogeic Nov 2000
S Endogeic No season identified to differ
As M Epigeic No season identified to differ 1 outlier in Nov 2000
P Epigeic No season identified to differ
S Epigeic No season identified to differ 1 outlier in Nov 2000
All Endogeic species no seasons showed significant differences
Cr M Epigeic March 2003, Dec 2001 1 outlier in Nov 2000
P Epigeic Biological variation too large 1 outlier in Feb 2004
S Epigeic Sept 2001, May 2001
All Endogeic species no seasons showed significant differences
Outliers are identified using Grabbs test. See material and method section for explanation of the statistics.
exceeding levels of earthworms inhabiting non-polluted sites(max up to 26 mg/g) (Van Gestel et al., 1992). Significant dif-ferences over time (ANOVA, p < 0.05) were found withinearthworms collected from sites M and S. Cd levels in earth-worms collected from site P were highly variable, and nosignificant seasonal differences were found (ANOVA,p < 0.05). Earthworms collected in February 2001 and No-vember 2002 from site P exceeded internal Cd levels of100 mg/g. This concentration is expected to cause significantreduction in growth (Ma et al., 1983). Cd concentrations inNovember 2003 were twice as high at site P compared tosite M. More importantly, more litter was present at site Pthan at sites M and S. As stated before, we expect the pres-ence of a litter layer to cause the variable patterns found inepigeic earthworms from site P and their elevated internal
levels over the whole year. Soil metal concentrations couldnot explain internal Cd differences, neither could water com-position and inundation frequency. For A. chlorotica and A.caliginosa, significant differences in Cd levels were foundover time (ANOVA, p < 0.05), but they were equally vari-able as observed for the epigeic earthworm.
3.7. Arsenic
A large biological variation was observed within samplinggroups, with average As concentrations in L. rubellus rangingfrom 4.71 mg/g (S, Nov 2000) to 58.4 mg/g (P, Nov 2002). In-ternal concentrations were elevated compared to As levels(max 12 mg/g) found in earthworms from reference sites(Van Gestel et al., 1992). Internal As concentrations at site

139M.G. Vijver et al. / Environmental Pollution 148 (2007) 132e140
M were positively related to body weight (ANCOVA,p ¼ 0.004). In general, accumulation in earthworms was high-est during autumn and winter, and body weight contributed totemporal fluctuations in internal metal concentrations. Earth-worms from sites P and S showed no significant differencesin As concentrations over time. No relationship between inter-nal As levels and environmental parameters was found. In theendogeic earthworm species, a mean As concentration of20 mg/g was found and did not show significant variation intime (ANOVA, p < 0.05).
3.8. Chromium
Cr concentrations in L. rubellus showed large biologicalvariation within the sampling groups. Body concentrationswere elevated compared to levels (max 3 mg/g) found in earth-worms from non-polluted areas (Van Gestel et al., 1992).
Although internal levels were elevated, no toxic effects areexpected for L. rubellus Critical effect concentrations derivedfrom toxicity experiments are much higher, e.g., EC50 level forcocoon production of Eisenia fetida was 890 mg/kg dw for Cr(III) (Lock and Janssen, 2002) Significant differences overtime (ANOVA, p < 0.05) were found for sites M and S. Earth-worms from site P showed a large biological variation (Fmax)and no statistical tests for time-relations were performed. Nospecific environmental parameter could explain the differ-ences. It should be noted that Cr concentrations in porewaterwere slightly higher than detection limit.
Cr concentrations in A. chlorotica and A. caliginosa wereup to 15 mg/g and did not vary over time (ANOVA,p < 0.05). Internal Cr concentrations in endogeic earthwormswere comparable to the lowest Cr concentrations found inL. rubellus.
4. Discussion
Metal uptake by soil-invertebrates varies depending uponthe biology of species and physico-chemical modifying factorspresent in soil. Location-specific differences in Zn, Cd, As,and Cr concentrations in L. rubellus were found, whereasCa, Cu, Fe and Pb levels had a similar pattern over the seasonsand over the three different floodplain sites. For the endogeicearthworm species, location-specific differences in internal Pband Cd concentrations were found, whereas all other internalmetal and As levels were similar over time and over the threesites. Hence, earthworm-specific physiology has an impact oninternal metal levels. A similar sampling campaign was exe-cuted for isopods species Trachelipus rathkii, Philoscia mus-corum, Oniscus asellus within the same time frame and atthe same locations. Details on that study are reported by Vijver(2005). It was concluded that metal concentrations in variousisopod species did not differ. For isopods, neither location-spe-cific accumulation differences nor seasonal differences weredetected, except for internal Cd levels.
Floodplain soils are generally characterized by low metalconcentrations in pore water, and depletion of metals frompore water in the direct surroundings of earthworms has
been reported (Pinheiro et al., 2004). The desorption-limitedpore water pool has strong consequences for chemical avail-ability and subsequent uptake by organisms (Vink, 2002).This effect may be responsible for at least a part of theobserved variation in metal concentrations in organisms. Espe-cially location-specific differences over time may be explainedthis in way.
Redox mechanisms mechanisms for Cr were summarizedby, for example, Kozuh et al. (2000). Alterations of chemicalavailability over the seasons were, however, not reflected bythe observed uptake patterns. The same conclusion was drawnfor As. Internal As concentrations however showed large var-iation within individuals, and no descriptive environmentalcharacteristic could be deduced. Internal levels of essentialmetals mostly were elevated during spring, from February toMay. But also the pattern could not be attributed to any envi-ronmental characteristic. Some authors have suggested thatmetal concentrations in the body burden of earthworms areclosely related to pore water concentrations (e.g., Janssenet al. (1997b) for Eisenia andrei). In our field survey, however,we found no conclusive environmental characteristic that canconfirm this statement. This can be explained by antagonisticmasking effects of a variety of environmental factors (Grime,1989), or the internal storage enabling earthworms (Vijveret al., 2006) to mask the effect of soluble metals.
5. Conclusion
Metal levels in earthworms inhabiting floodplain areasshowed large variations among sites, among pedo-ecologicalclassification, and among seasons. Although four-years sur-veillance data on epigeic earthworms and two-years surveil-lance data on endogeic earthworms were collected, no cleargeneral patterns of metal accumulation could deduced. Thevariations in metal accumulation could not be explained byany individual field parameter. Risk assessments should there-fore be apprehensive to a single stress approach, and be pre-pared for masking effects of environmental characteristics.
Acknowledgements
We thank our colleagues from the IMLA Department(RIZA) for their analytical work. Thomas Schroder for theporewater sampling and analyses. We are grateful to AminaDjorai for analysing the data, and Prof. Dr. Nico van Straalenfor his valuable comments on an earlier draft.
References
Bouche, M.B., 1977. Strategies lombriciennes. In: Lohm, U., Persson, T.
(Eds.), Soil Organisms as Components of Ecosystems, vol. 25. Ecological
Bulletin, Stockholm.
Christensen, J.B., Botma, J.J., Christensen, T.H., 1999. Complexation of Cu
and Pb by DOM in polluted groundwater: a comparison of experimental
data and predictions by computer speciation models (WHAM and MIN-
TEQA2). Water Research 15, 3231e3238.
Funk, W., Dammann, V., Voderheid, C., Oehler, G., 1985. Statistische Meth-
oden in der Wasseranalytik. VCH verlagsgesellsschaft mbH, Weinheim.

140 M.G. Vijver et al. / Environmental Pollution 148 (2007) 132e140
Grime, J.P., 1989. The stress debate; symptom of impending synthesis? Bio-
logical Journal of the Linnean Society 37, 3e17.
Hobbelen, P.H.F., Koolhaas, J.E., Van Gestel, C.A.M., 2004. Risk assessment
of heavy metal pollution for detrivores in floodplain soils in the Biesbosch,
the Netherlands, taking bioavailability into account. Environmental Pollu-
tion 129, 409e419.
Houba, V.J.G., Lexmond, Th.M., Novozamsky, I., Van der Lee, J.J., 1996.
State of the art and future developments in soil analysis for bioavailability
assessment. Science of the Total Environment 178, 21e28.
Ireland, M.P., Richards, K.S., 1977. The occurrence and localisation of heavy
metals and glycogen in the earthworms Lumbricus rubellus and Dendro-
baena rubida from a heavy metal site. Histochemistry 51, 153e166.
Janssen, R.P.T., Peijnenburg, W.J.G.M., Posthuma, L., Van den
Hoop, M.A.G.T., 1997a. Equilibrium partitioning of heavy metals in Dutch
field soils. I. Relationship between metal partition coefficients and soil
characteristics. Environmental Toxicology and Chemistry 16, 2470e2478.
Janssen, R.P.T., Posthuma, J., Baerselman, R., Den Hollander, H.A., Van
Veen, R.P.M., Peijnenburg, W.J.G.M., 1997b. Equilibrium partitioning of
heavy metals in Dutch fields soils. II. Prediction of metal accumulation
in earthworms. Environmental Toxicology and Chemistry 12, 2479e2788.
Kozuh, N., Supar, J., Gorenc, B., 2000. Reduction and oxidation processes of
chromium in soils. Environmental Science and Technology 34, 112e119.
Langdon, C.J., Piearce, T.G., Meharg, A.A., Semple, K.T., 2001. Resistance to
copper toxicity in populations of the earthworms Lumbricus rubellus and
Dendrodrilus rubidus from contaminated mine wastes. Environmental
Toxicology and Chemistry 20, 2336e2341.
Lock, K., Janssen, C.R., 2002. Ecotoxicity of chromium (III) to Eisenia fetida,
Enchytraeus albidus, and Folsomia candida. Ecotoxicology and Environ-
mental Safety 51, 203e205.
Ma, W., Edelman, T., Van Beersum, I., Jans, T., 1983. Uptake of cadmium,
zinc, lead and copper by earthworms near a zinc-smelting complex: influ-
ence of soil pH and organic matter. Bulletin of Environmental Contamina-
tion Toxicology 30, 424e427.
Morgan, J.E., Morgen, A.J., 1998. The distribution and intracellular compart-
mentation of metals in the endogeic earthworm Aporrectodea caliginosa
sampled from an unpolluted and a metal-contaminated site. Environmental
Pollution 99, 167e175.
NEN, 1992. Monstervoorbehandling van slib, slibhoudend water, luchtstof en
grond voor de bepaling van elementen met atomaire-absorptie-spectrome-
trie. In: Ontsluiting met salpeterzuur en zoutzuur, second ed., 6465. Neder-
lands Normalisatie-Instituut, Delft, The Netherlands (In Dutch).
Piearce, T.G., 1972. The calcium relations of selected Lumbricidae. Journal of
Animal Ecology 41, 167e188.
Pinheiro, J.P., Galceran, J., Van Leeuwen, H.P., 2004. Metal speciation dynam-
ics and bioavailability: bulk depletion effects. Environmental Science &
Technology 38, 2397e2405.
Sauve, S., Hendershot, W., Allen, H.E., 2000. Solid-solution partitioning of
metals in contaminated soils: dependence on pH, total metal burden and
organic matter. Environmental Science and Technology 34, 1125e1131.
Spurgeon, D.J., Hopkin, S.P., 1999. Comparisons of metal accumulation and
excretion kinetics in earthworms (Eisenia fetida) exposed to contaminated
field and laboratory soils. Applied Soil Ecology 11, 227e243.
Van Gestel, C.A.M., 1997. Scientific basis for extrapolating results from soil
ecotoxicity tests to field conditions and the use of bioassays. In: Van
Straalen, N.M., Løkke, H. (Eds.), Ecological Risk Assessment of Contam-
inants in Soil. Chapman & Hall, London.
Van Gestel, C.A.M., Dirven-van Breemen, E.M., Kamerman, J.W., 1992.
Beoordeling van gereinigde grond. IV. Toepassing van bioassays met
planten en regenwormen op referentiegronden. RIVM-rapport
216402004. Bilthoven, The Netherlands.
Van Leeuwen, H.P., 1999. Metal speciation dynamics and bioavailability:
inert and labile complexes. Environmental Science & Technology 33,
3743e3748.
Vegter, J.J., 1995. Soil protection in the Netherlands. In: Salomons, W.,
Forstner, U., Mader, P. (Eds.), Heavy Metals-Problems and Solutions.
Springer-Verlag, Berlin-Heidelberg.
Vink, J.P.M., 2002. Measurement of heavy metal speciation over redox
gradients in natural water-sediment interfaces and implications for
uptake by benthic organisms. Environmental Science & Technology 36,
5130e5138.
Vink, J.P.M., Meeussen, J.C.L., 2007. BIOCHEM-ORCHESTRA: a tool
for evaluating chemical speciation and ecotoxicological impacts of
heavy metals in river floodplain systems. Environmental Pollution, in
press.
Vijver, M.G., 2005. The Ins and Outs of Bioaccumulation. Metal Bioaccumu-
lation Kinetics in Soil Invertebrates in Relation to Availability and Physi-
ology. ISBN no 903695701X. PhD-thesis, Vrije Universiteit, Amsterdam,
The Netherlands.
Vijver, M.G., Vink, J.P.M., Miermans, C.J.H., Van Gestel, C.A.M., 2003a. Oral
sealing using glue: a new method to distinguish between intestinal and der-
mal uptake of metals in earthworms. Soil Biology and Biochemistry 35,
125e132.
Vijver, M., Jager, T., Posthuma, L., Peijnenburg, W., 2003b. Metal uptake from
soils and soil-sediment mixtures by larvae of Tenebrio molitor (L.) (Cole-
optera). Ecotoxicology and Environmental Safety 54, 277e289.
Vijver, M.G., Van Gestel, C.A.M., Van Straalen, N.M., Lanno, R.P.,
Peijnenburg, W.J.G.M., 2006. Biological significance of metals partitioned
to subcellular fractions within earthworms (Aporrectodea caliginosa).
Environmental Toxicology and Chemistry 25, 807e814.
Wallinga, I., Kithome, M., Novozamsky, I., Houba, V.J.G., Van der Lee, J.J.,
1992. Spectrophotometric determination of organic carbon in soil.
Communications in Soil Science and Plant Analyses 23, 1935e1944.




















