
Algal–bacterial interactions in metal contaminated floodplain sediments
11
Algalebacterial interactions in metal contaminated floodplain sediments M.E.Y. Boivin a,b , G.D. Greve a,b , J.V. Garcı ´a-Meza c , B. Massieux d , W. Sprenger c , M.H.S. Kraak c, * , A.M. Breure a , M. Rutgers a , W. Admiraal c a National Institute for Public Health and the Environment, P.O. Box 1, 3720 BA Bilthoven, The Netherlands b Department of Animal Ecology, IES, Vrije Universiteit, De Boelelaan 1085, 1081 HV Amsterdam, The Netherlands c Department of Aquatic Ecology and Ecotoxicology, IBED, University of Amsterdam, Kruislaan 320, 1098 SM Amsterdam, The Netherlands d Netherlands Institute of Ecology, Centre for Limnology, Rijkstraatweg 6, 3631 AC Nieuwersluis, The Netherlands Received 20 April 2006; accepted 1 May 2006 Algal and bacterial communities in floodplain sediments are interlinked, but are not affected by metal pollution. Abstract The aim of the present study was to investigate algalebacterial interactions in a gradient of metal contaminated natural sediments. By means of multivariate techniques, we related the genetic structure (denaturing gradient gel electrophoresis, DGGE) and the physiological structure (community-level physiological profiling, CLPP) of the bacterial communities to the species composition of the algal communities and to the abiotic environmental variables, including metal contamination. The results revealed that genetic and physiological structure of the bacterial communities correlated with the species composition of the algal community, but hardly to the level of metal pollution. This must be interpreted as an indication for a strong and species-specific linkage of algal and bacterial species in floodplain sediments. Metals were, however, not proven to affect either the algal or the bacterial communities of the Dutch river floodplains. Ó 2006 Elsevier Ltd. All rights reserved. Keywords: Algae; Bacteria; Community; Sediment; Metal pollution 1. Introduction Microorganisms have the ability to colonise extremely pol- luted habitats, yet they are also known to be sensitive to metal pollution (Babich and Stotzky, 1985). Observations in metal polluted environments have shown that the density and the diversity of bacterial communities decreased and that the surviving bacterial communities had genetic and physiological characteristics differing from reference communities (Tyler, 1974; Ba ˚a ˚th et al., 1998; Rutgers et al., 1998; Gong et al., 2002; Van Beelen et al., 2004). Metal exposed bacterial communities may acquire metal tolerance, either by physiological adaptation, genetic changes or the succession of sensitive species by more tolerant ones (Blanck et al., 1988; Boivin et al., 2002). These processes have been tradi- tionally studied in environments clearly affected by metal pollution. However, at lower levels of metal pollution abiotic and biotic interactions complicate the establishment of a mean- ingful relationship between metal contamination and changes in bacterial communities. Factors such as pH, organic matter and clay content are likely to confound the direct biological effects of low concentrations of metals (Ba ˚a ˚th and Anderson, 2003). Nold and Zwart (1998) pointed to the importance of interspecies interactions among microbial communities, mod- ifying the potential response to a toxic agent. Interactions between heterotrophic bacterial communities and autotrophic algal communities do also occur (Scha ¨fer et al., 2002). Bell et al. (1984) pointed out that long-term selection favours * Corresponding author. Tel.: þ31 20 525 7717. E-mail address: [email protected] (M.H.S. Kraak). 0269-7491/$ - see front matter Ó 2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.envpol.2006.05.003 Environmental Pollution 145 (2007) 884e894 www.elsevier.com/locate/envpol
Transcript of Algal–bacterial interactions in metal contaminated floodplain sediments

Environmental Pollution 145 (2007) 884e894www.elsevier.com/locate/envpol
Algalebacterial interactions in metal contaminated floodplain sediments
M.E.Y. Boivin a,b, G.D. Greve a,b, J.V. Garcıa-Meza c, B. Massieux d, W. Sprenger c,M.H.S. Kraak c,*, A.M. Breure a, M. Rutgers a, W. Admiraal c
a National Institute for Public Health and the Environment, P.O. Box 1, 3720 BA Bilthoven, The Netherlandsb Department of Animal Ecology, IES, Vrije Universiteit, De Boelelaan 1085, 1081 HV Amsterdam, The Netherlands
c Department of Aquatic Ecology and Ecotoxicology, IBED, University of Amsterdam, Kruislaan 320, 1098 SM Amsterdam, The Netherlandsd Netherlands Institute of Ecology, Centre for Limnology, Rijkstraatweg 6, 3631 AC Nieuwersluis, The Netherlands
Received 20 April 2006; accepted 1 May 2006
Algal and bacterial communities in floodplain sediments are interlinked, but are not affected by metal pollution.
Abstract
The aim of the present study was to investigate algalebacterial interactions in a gradient of metal contaminated natural sediments. By meansof multivariate techniques, we related the genetic structure (denaturing gradient gel electrophoresis, DGGE) and the physiological structure(community-level physiological profiling, CLPP) of the bacterial communities to the species composition of the algal communities and tothe abiotic environmental variables, including metal contamination. The results revealed that genetic and physiological structure of the bacterialcommunities correlated with the species composition of the algal community, but hardly to the level of metal pollution. This must be interpretedas an indication for a strong and species-specific linkage of algal and bacterial species in floodplain sediments. Metals were, however, not provento affect either the algal or the bacterial communities of the Dutch river floodplains.� 2006 Elsevier Ltd. All rights reserved.
Keywords: Algae; Bacteria; Community; Sediment; Metal pollution
1. Introduction
Microorganisms have the ability to colonise extremely pol-luted habitats, yet they are also known to be sensitive to metalpollution (Babich and Stotzky, 1985). Observations in metalpolluted environments have shown that the density and thediversity of bacterial communities decreased and that thesurviving bacterial communities had genetic and physiologicalcharacteristics differing from reference communities (Tyler,1974; Baath et al., 1998; Rutgers et al., 1998; Gong et al.,2002; Van Beelen et al., 2004). Metal exposed bacterialcommunities may acquire metal tolerance, either by
* Corresponding author. Tel.: þ31 20 525 7717.
E-mail address: [email protected] (M.H.S. Kraak).
0269-7491/$ - see front matter � 2006 Elsevier Ltd. All rights reserved.
doi:10.1016/j.envpol.2006.05.003
physiological adaptation, genetic changes or the successionof sensitive species by more tolerant ones (Blanck et al.,1988; Boivin et al., 2002). These processes have been tradi-tionally studied in environments clearly affected by metalpollution. However, at lower levels of metal pollution abioticand biotic interactions complicate the establishment of a mean-ingful relationship between metal contamination and changesin bacterial communities. Factors such as pH, organic matterand clay content are likely to confound the direct biologicaleffects of low concentrations of metals (Baath and Anderson,2003). Nold and Zwart (1998) pointed to the importance ofinterspecies interactions among microbial communities, mod-ifying the potential response to a toxic agent. Interactionsbetween heterotrophic bacterial communities and autotrophicalgal communities do also occur (Schafer et al., 2002). Bellet al. (1984) pointed out that long-term selection favours

885M.E.Y. Boivin et al. / Environmental Pollution 145 (2007) 884e894
bacterial communities that are adapted to the spectrum oforganic carbon provided by the algal community. So far,however, there is no information on the role of autotropheheterotroph interactions in the response of bacteria to metals.The aim of the present study was therefore to investigatealgalebacterial interactions in a gradient of metal contami-nated natural sediments. We focused on bacterial communitiesof biofilms colonising sediment surface layers of pollutedfloodplains of the river Rhine in the Netherlands. Althoughthese sediments contain substantial concentrations of metals,mainly as a result of historic pollution of the river, the biolog-ical effects are still under debate. Here, by means of multivar-iate techniques, we related the genetic structure (denaturinggradient gel electrophoresis, DGGE) and the physiologicalstructure (community-level physiological profiling, CLPP) ofthe bacterial communities to the species composition of thealgal communities and to the abiotic environmental variables,including metal contamination.
2. Materials and methods
2.1. Study area
The Afferdensche and Deestsche Waarden (ADW) and the Sliedrechtse
Biesbosch (B) are floodplains subjected to flooding by the river Rhine. The
floodings have resulted in the sedimentation of particulate matter from river
water contaminated with metals and persistent organic compounds especially
at the time when the river Rhine was heavily polluted (ca. 1960e1980).
Deeper layers of severely contaminated sediments are covered by recent de-
posits with only traces of contaminants.
2.2. Sampling location selection
In May 2003, samples were taken at four ADW lakes, giving a total of 100
sediment top layer samples, three from lake Rijswaard (R1, R2 and R3) and one
from lake Deest1 (D1), lake Deest3 (D3) and lake Deest4 (D4). Sampling
locations were chosen based on the total concentrations of Zn, Pb and Cu.
The six selected locations were marked with a plastic flag and were sampled
again in July and September 2003. In September 2003, three samples were
taken at three locations in the Biesbosch (B1, B2 and B3).
2.3. Sampling
At the selected locations the submerged sediment top layer (top 1 cm layer
at 20 cm water depth) was collected. Because of water evaporation during the
sampling-months, the subsequent samples were taken in lower parts of the
lake, causing variations in metal concentrations and sediment properties
between the sampling-months. It was impossible to sample R3 in September,
because it was completely dried.
Each sample was mixed and divided into four portions. The first one was
used to measure pH, organic matter content, clay content and total and calcium
chloride (CaCl2) extractable metal concentrations. The second one was used to
determine the physiological structure of the bacterial community by means of
CLPP analysis (Rutgers et al., 1998, 2006). The third one was used to assess
the genetic structure of the microbial community by DGGE analysis of PCR-
amplified fragments of SSU-rDNA using a primer combination specific for
Bacteria and Eukarya. The fourth one was used for microscopical assessment
of algal diversity and abundance.
2.4. Analysis of abiotic factors
Abiotic parameters measured were water pH, light intensity, conductivity,
oxygen concentration and temperature of the water and sediments.
For the analysis of metal concentrations, sediment samples were dried
overnight at 100 �C and subsequently grounded using a mortar. The calcium
chloride exchangeable fraction (CaCl2) was determined to get an estimate of
the exposure of sediment organisms to metals. Thirty millilitres of CaCl2(0.01 M, pH 7) was added to 3 g dry sediment. Sediment suspensions were
shaken subsequently over night at 150 rpm. The pH of the solution was mea-
sured and further referred to as sediment pH. The supernatant was filtered
(0.45 mm) and acidified to pH 1 to keep extracted metals in solution
(w15 ml HNO3 per 10 ml sample; Suprapur, 65%: Merck, Darmstadt, Ger-
many). Total metal concentrations were determined by adding 0.25 g of dry
sediment to 3 ml milli-Q water, 6 ml HCl (37%; Merck) and 2 ml HNO3
(65%; Merck). Digestion of the sediments was done using a microwave
(CEM, Mars 5), at a pressure of 100 psi and a temperature of 150 �C for
60 min. After cooling down, 50 ml milli-Q water was added to the samples.
All metal analyses were performed on a ICP-MS (HP-4500) using a standard
operational procedure from the National Institute for Public Health and the
Environment in The Netherlands (RIVM; SOP LAC M419).
For the analysis of total organic matter content the sediment samples were
divided over porcelain crucibles that were previously weighed. The samples
were weighed and placed in an oven at 100 �C overnight. The samples were
reweighed and then ashed in a muffle furnace at 450 �C for 6 h. The weight
loss upon ignition (LOI) was considered to represent total organic matter con-
tent. The samples were further ashed in a muffle furnace at 900 �C for 1 h. The
weight loss was considered to represent inorganic matter content.
To determine the clay content of the sediment samples, a ‘‘laser particle
sizer’’ A222 was used (Fritsch Gmbh, Idar Oberstein, Germany) according
to Konert and Van den Berghe (1997). The clay content was corrected for
inorganic matter content.
2.5. Analysis of biota
Algae (chlorophytes, euglenophytes, diatoms and cyanobacteria), were
identified in fixed sediment samples (field samples in 2% formaldehyde).
The identification of diatoms was done using acid-cleaned frustules. The iden-
tifications were done by microscopic observations, using an optical micro-
scope (Olympus BH2, Tokyo, Japan) equipped with contrast-phase yield. To
identify the species of the field samples, few drops of the preserved samples
were placed on a 24 � 24 mm area (the cover-slide size) and observed at
a magnification of 1000�. The identified species were grossly counted. The
presence of species was indicated in two classes: abundant species and infre-
quent species.
To extract the bacteria from the sediment, every sediment sample was
mixed and 1 ml of sediment suspension was added to 4 ml of 10 mM of sterile
Bis-Tris buffer, pH 7. The mixture was sonicated three times for 30 s
(68 watts) and centrifuged (500 � g) for 10 min. The supernatant was trans-
ferred into 2 ml microtubes, frozen in liquid nitrogen, and maintained
at �70 �C until use for CLPP analysis.
Bacterial DNA from the sediment samples was extracted by using a stan-
dard extraction kit (FastDNA� SPIN Kit for Soil, BIO 101, USA). Subse-
quently, PCR was performed with the DNA extracts, to amplify the region
between no. 357 and no. 518 of the 16s rRNA gene (nucleotide positions
according to Escherichia coli). Subsequently, the genetic structure of bacterial
communities was determined by means of DGGE according to Muyzer et al.
(1993). The denaturing agents, urea and formamide, used for the DGGE were
ranging from 35% at the top to 60% at the bottom of the gel. The DGGE gel
was submerged in a buffer containing 40 mM Tris, 40 mM acetic acid, 1 mM
EDTA, pH 7.6 (0.5� TAE) and submitted to 75 V for 16 h. In contrast to or-
dinary agarose gel electrophoresis, the amplicons have the same length, but
migrate differently if they differ in GC content. Total denaturation is prevented
by a so-called GC clamp at one end of the amplicon. The DGGE gel was an-
alysed using the program Bionumerics. The banding patterns were analysed
with a Dice correlation also using the program Bionumerics. A similarity
matrix was constructed from band presence and absence. The dendrogram
obtained was subjected to a redundancy analysis (RDA).
For the determination of the physiological structure of bacterial communi-
ties, CLPPs were generated using 96-well ECO-plates (BIOLOG Inc.,
Hayward, CA, USA), which contain a triplicate set of 31 different carbon

886 M.E.Y. Boivin et al. / Environmental Pollution 145 (2007) 884e894
substrates plus a no-substrate control as well as a mineral medium and the
redox dye tetrazolium. Twelve subsequent threefold dilutions of the bacterial
suspensions (3�1 to 3�12) were made in sterile 0.9% physiological salt, pH 7.
The plates were inoculated with 100 ml of bacterial dilution per well and were
incubated in the dark at 20 �C at a relative humidity of w85%. Colour devel-
opment in each well was measured spectrophotometrically at 590 nm every 8 h
during 7 days. CLPPs were calculated using a regression approach applied to
the average well colour development (AWCD) according to Rutgers et al.
(2006) and Boivin et al. (2005). To assess the overall activity of the bacterial
community in the ECO-plate, the AWCD after 3 days incubation was
calculated.
2.6. Statistics
All abiotic parameters were verified for normality using Kolmogorove
Smirnov test (GraphPad Prism, version 4.01, January 2004). Next, it was
analysed if there were correlations between the different abiotic parameters.
Pearson correlation was used in case of normal distributed data and Spearman
correlation in case of non-normal distributed data. The significance level was
corrected for the number of comparisons made using a Bonferroni correction.
The algal communities were analysed using a redundancy analysis (RDA).
The dominant species as well as the infrequent species were used for the anal-
ysis and therefore different weights were given to the data. The presence of
a dominant species in a sample weighted ten times more than the presence
of an infrequent species. In a direct analysis, such as in a RDA, the number
of variables should be considerably lower than the number of samples. In
this study, the number of variables was higher than the number of samples
and therefore a selection procedure was performed to reduce the number of
variables. Prior to the analysis, each variable was tested in the forward selec-
tion procedure available in the software program Canoco V.4.0 (CPRO-
DLO, Wageningen, The Netherlands; Ter Braak and Smilauer, 1998). The
variables having significant values were included in the analysis (Monte Carlo
permutation test, 9999 permutation, p � 0.05). The variance inflation factors
(VIF) were also investigated. Indeed, high VIF (VIF � 20) indicates multi-
collinearity between variables, which should be avoided in this type of analysis
(Ter Braak, 1986). Since a large part of the observed variation was caused by
the locations sampled, this variable was taken as a covariable.
The results of the DGGE were first analysed in the same manner as the
algal communities, using the abiotic data set as explanatory variables (organic
matter and clay content, pH, metal concentrations etc.). Next, the DGGEs were
analysed using the biotic data set as explanatory variables (the weighed algae
species present at the locations). Finally, both the abiotic and the biotic data
sets together were used as explanatory variables. The results of the CLPPs
were analysed in the same manner as the DGGEs.
3. Results
3.1. Field characteristics
Table 1 presents abiotic variables measured in the Affer-densche and Deestsche Waarden (R1,2,3 and D1,3,4) and the Sli-edrechtse Biesbosch (B1,2,3). The sediment pH was uniformduring the sampling months and between sampling locations,in contrast to the water pH. The location D1 sampled in Julyand September showed a water pH above 9. Similar pH valueswere observed at D3 in July and also at R2 in September. Also,the oxygen concentrations of the water layer were very high onsome occasions, especially in September at R2, D1 and D4.These high pH and oxygen levels result from the active algalmats in these shallow waters. The water conductivity at D1 andD3 also increased during the course of the observation period.Clay and organic matter content were variable both in timeand space, but this was expected because the sampling
locations in the small lakes had to be moved because of thedecreasing water level.
Table 2 presents metal concentrations measured at the dif-ferent locations. The CaCl2 exchangeable metal concentra-tions (mg/kg dry weight) varied from 124 to 1027 (Zn), 2 to50 (Pb), 157 to 1655 (Cu) and 8 to 1190 (Cd). Total metal con-centrations (mg/kg dry weight) varied from 32 to 1043 (Zn), 9to 291 (Pb), 4 to 134 (Cu) and 0.02 to 0.2 (Cd).
The CaCl2 exchangeable fraction of Cu was significantlypositively correlated with clay and organic matter content hav-ing a coefficient correlation of 0.83 and 0.84, respectively. TheCaCl2 exchangeable fraction of Pb, Zn and Cd did not signif-icantly correlate with clay or organic matter content. The totalmetal concentrations correlated better with clay and organicmatter content than the CaCl2 exchangeable metal concentra-tions. Total concentrations of Cu, Pb, Zn and Cd were signif-icantly positively correlated with organic matter content, withcorrelation coefficient of 0.86, 0.78, 0.84 and 0.85, respec-tively. Total concentrations of Cu, Pb, Zn and Cd were alsosignificantly positively correlated with the clay content: coef-ficient correlation 0.80, 0.55, 0.63 and 0.85, respectively.Because sediment pH was uniform, none of the total metalconcentrations or CaCl2 exchangeable fraction of the metalshad a correlation with sediment pH.
3.2. Algal community
Table 3 shows the algal species found at the different loca-tions of the Afferdensche and Deestsche Waarden (R1,2,3 andD1,3,4) and the Sliedrechtse Biesbosch (B1,2,3). R1 was thelocation that harboured the largest number of algal speciescompared with the other locations, and all locations were dom-inated by diatoms. At every location, September was thesampling month counting the lowest number of algal species.
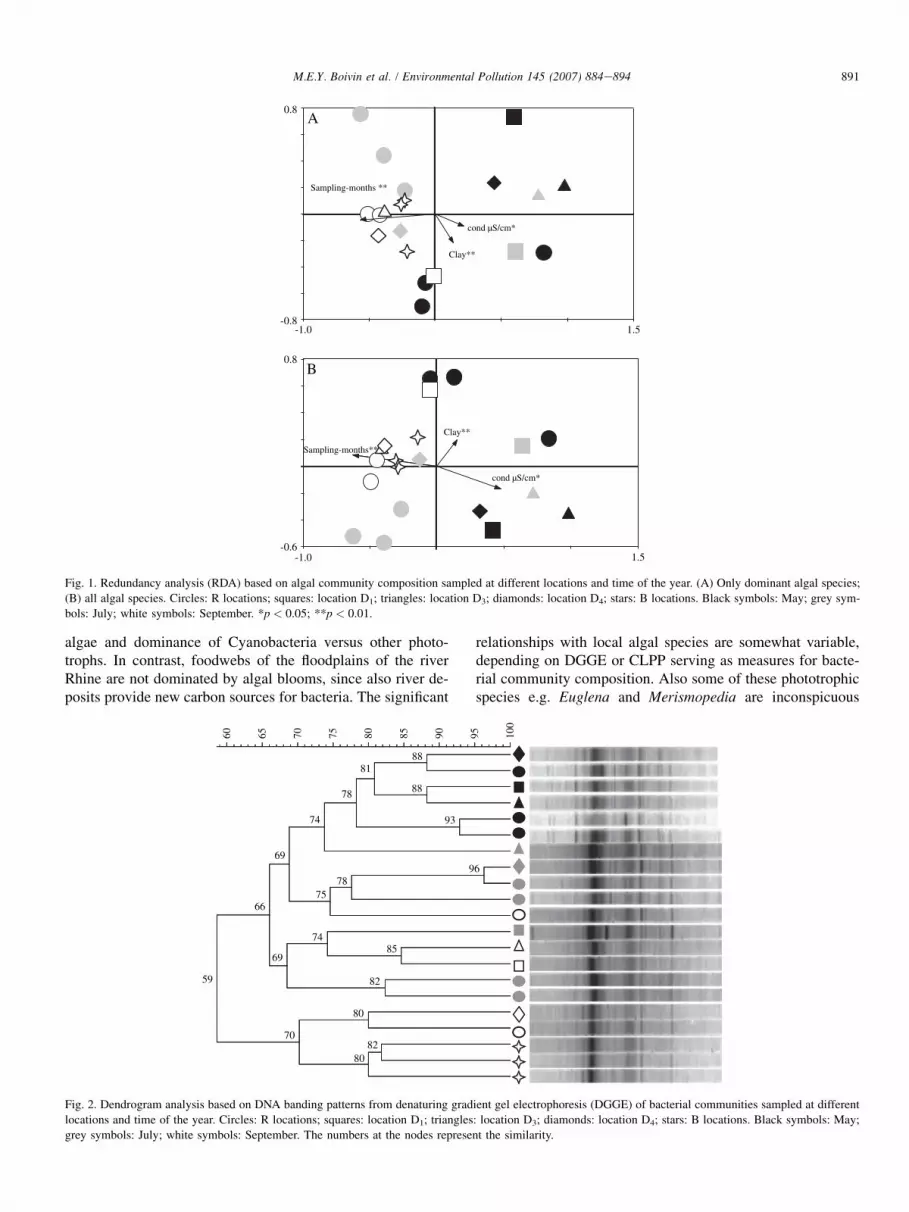
Fig. 1A represents an RDA based on the dominant algalspecies and Fig. 1B represents an RDA based on all algal spe-cies. The algal species composition showed no significantcorrelations with any of the four metals, neither as total con-centrations, nor as extractable concentrations. In contrast,the variability observed in the algal community composition,both for dominant species and all species was significantlycorrelated with the sampling-months, the clay content andthe conductivity of the water.
3.3. Denaturing gradient gel electrophoresis
Bacterial genotypes were investigated with DGGE analysis.Fig. 2 presents the banding patterns and a dendrogram analysisbased on the bacterial DGGE profiles made from sedimentsamples taken at the R, D and B locations. Different clustersmay be observed. In particular, bacterial communities sampledfrom the B locations cluster apart from the other communities.Also, bacterial communities sampled in May cluster indepen-dently from where they were sampled.
Fig. 3 presents an RDA of the DGGE analysis. The DGGEbanding patterns did not show any significant correlation withmetal concentrations. In contrast, as suspected after analysing

887M.E.Y. Boivin et al. / Environmental Pollution 145 (2007) 884e894
Table 1
Physical and chemical parameters measured at the field locations
R1 R2 R3 D1 D3 D4 B1 B2 B3
pH water May 7.1 7.8 7.6 8.6 8.9 8.4 e e e
July 8.5 8.8 8.6 9.8 9.1 7.5 e e e
September NA 9.1 NA 9.3 7.0 7.8 7.8 7.4 7.4
pH sediment May 6.8 6.6 6.7 6.8 6.8 6.8 e e e
July 6.5 6.6 6.7 6.9 6.8 6.8 e e e
September 6.9 6.6 6.8 6.8 7.0 6.7 6.8 7.0 6.8
Water temperature ( �C) May 16 16.4 16 16 16.2 16.1 e e eJuly 24 26 25 28 29 26 e e e
September 13.4 13.6 NA 23.8 17.1 16.1 11 11.5 11.5
Conductivity (mS/cm) May 496 494 499 363 421 510 e e eJuly 525 522 520 403 474 570 e e e
September NA 483 NA 489 570 537 NA NA NA
Oxygen (%) May 66 77 50 71 72 75 e e e
July 95 80 87 191 162 97 e e eSeptember NA 213 NA 324 4 113 83 83 83
Clay (%) May 9.1 6.3 11.5 8.1 26.3 25.3 e e e
July 2.6 2.5 12.4 49.5 33.4 20.5 e e e
September 23.3 10.5 25.3 46.8 32 23.6 25.8 44.7 40.9
Organic matter (%) May 1.8 1.3 1.4 1.3 7.4 9.3 e e e
July 0.6 0.5 1.5 7.1 6.4 5.6 e e e
September 5.5 1.3 NA 8.8 6.8 6.3 4.9 9.4 8.9
NA, not available; e, no samples were taken.
the dendrogram (Fig. 2), changes in genetic composition of thebacterial communities were significantly correlated with thesampling-months (Fig. 3A,D,E). Fig. 3B shows correlationsbetween DGGE pattern shifts and the dominant algal speciesobserved in the field. The DGGE pattern shifts were signifi-cantly correlated with the chlorophyte Oocystis marssonniand the diatoms Navicula cf. fiesta and Cyclotella atomus.Fig. 3C shows correlations between DGGE pattern shifts and
all algal species observed in the field. The DGGE patternsshifts were again significantly correlated with the chlorophyteO. marssonni and the diatom Cyclotella atomus, but now alsowith the euglenophyte Euglena sp. and the diatom Asterionellaformosa. Fig. 3D shows correlations between DGGE patternsshift and the combined set of variables of dominant algalspecies and abiotic parameters. The DGGE patterns shiftswere significantly correlated with the chlorophyte
Table 2
Cu, Zn, Cd and Pb concentrations in the sediment based on total destruction and CaCl2 exchangeable fraction
Total destruction (mg/kg dry wt.) CaCl2 exchangeable (mg/kg dry wt.)
Cu Zn Cd Pb Cu Zn Cd Pb
R1 May 11 68 0.03 16 702 1027 110 50
July 4 32 0.02 9 314 175 13 2
September 46 276 0.12 66 1638 424 17 6
R2 May 10 79 0.04 18 352 201 20 3
July 5 36 0.04 11 157 124 8 2
September 10 75 0.04 16 295 174 21 3
R3 May 8 63 0.03 15 417 265 14 3
July 10 76 0.03 16 377 210 19 4
September NA NA NA NA NA NA NA NA
D1 May 9 57 0.05 13 436 227 69 4
July 51 296 0.13 68 1558 439 71 11
September 48 282 0.15 64 1655 528 13 7
D3 May 48 322 0.10 68 752 698 1190 7
July 52 346 0.10 71 875 792 31 11
September 51 350 0.11 76 738 517 18 5
D4 May 134 1043 0.12 291 1504 476 88 6
July 46 320 0.11 83 821 580 244 6
September 54 361 0.11 86 837 592 22 7
B1 September 80 551 0.14 127 971 667 21 3
B2 September 63 413 0.11 111 1176 632 40 5
B3 September 95 609 0.15 143 1832 846 257 5
NA, not available.

888 M.E.Y. Boivin et al. / Environmental Pollution 145 (2007) 884e894
O. marssonni and the diatoms Navicula cfr. festiva andAulacoseira granulata as well as with sampling-months.Fig. 3E shows correlations between DGGE patterns shift andall other parameters. The DGGE patterns shifts were still sig-nificantly correlated with the chlorophyte O. marssonni andagain with the euglenophyte Euglena sp. and the diatomsA. formosa and A. granulata, but now also with the cyanobac-terium Merismopedia. Some of these correlations can betraced directly in the data set: for example, O. marssonniwas only present in July and at the R sites and Euglena sp.was also found at R sites, but only in May.
3.4. Community-level physiological profiles
The metabolic capacity of the bacterial communities wasinvestigated using CLPP analysis. The activity of the hetero-trophic bacterial communities in the Biolog plate, expressedas average well colour development (AWCD), did not differbetween locations or sampling-seasons and therefore no sig-nificant correlations were found between those parametersand any abiotic or biotic variables. Fig. 4 presents an RDAbased on the variability observed in the different CLPPs.The position of the CLPP in the multivariate space signifi-cantly correlated with total Cd concentration and with theO2 concentration of the water layer above the sediments,the water conductivity and pH (Fig. 4A). Fig. 4B shows cor-relations between CLPP shifts and the dominant algalspecies. The CLPP shifts were correlated with the chloro-phytes Oocystis marssonni and Chlorella sp. and the diatomsAulacoseira granulata and Cyclotella atomus. Fig. 4C showscorrelations between CLPP shifts and all algal species. TheCLPP shifts were again significantly correlated with thechlorophyte O. marssonni and the diatom A. granulata, butnow also with the chlorophyte Scenedesmus quadricaudavar. longispina. Fig. 4D shows correlations between meta-bolic profiles shifts and the combined set of variables ofdominant algal species and abiotic parameters. The CLPPshifts were again significantly correlated with the diatomsA. granulata and C. atomus and the chlorophyte Chlorellasp. and with only one abiotic parameter: the total Cd concen-tration. Fig. 4E shows correlations between CLPP shifts andall other parameters. The CLPP shifts were still significantlycorrelated with the chlorophyte O. marssonni and the diatomA. granulata, but now also with the diatom Gomphonemaaffine. The CLPP shifts were now also significantly relatedto several abiotic parameters: clay content, oxygen leveland sampling months.
The shifts in CLPPs were due to changes in the utilisationof carbon sources. Simple regressions were made between theO2 concentrations of the water and the different carbon sub-strates. The O2 concentration was used since it significantlycorrelated with the CLPP shifts and is generally correlatedwith algal blooms. Bacterial communities extracted fromlocations showing low O2 concentration utilised i-erythritol,L-threonine, a-cyclodextrine and g-hydroxybutyric acid toa greater extent than bacterial communities from locationshaving higher O2 concentration. On the opposite, D-malic
acid, phenylethylamine and Tween-40 were relatively moreutilised by bacterial communities extracted from locationsshowing higher O2 concentration.
4. Discussion
The present study revealed that genetic and physiologicaldiversity of floodplain bacteria was correlated with the diver-sity of algae, but hardly to the level of metal pollution. Thesetwo different groups of potential drivers of bacterial diversitywill be discussed below.
Sample locations were selected to reflect major differencesin metal pollution in the floodplains of the river Rhine. Totalconcentrations of Cd, Cu, Pb and Zn indeed showed a tenfolddifference between locations. Conforming to expectation, VanGriethuysen et al. (2004) observed that the structure of thebenthic invertebrate communities in the same floodplain sedi-ments correlated with metal level. In addition, De Haas et al.(2004) observed that these metal contaminated sediments weretoxic to aquatic insects in laboratory bioassays. The presentstudy does not confirm the observations by Van Griethuysenet al. (2004) and De Haas et al. (2004). The genetic structureof the bacterial community (DGGE) showed no significantcorrelations with any of the four metals, neither as total con-centrations, nor as extractable concentrations. The physiolog-ical structure of the bacterial community (CLPP) correlatedonly with the total Cd content of the sediments, among signif-icant correlations with other abiotic factors like O2, pH andconductivity. Thus the present study revealed that there wasa lack of a robust correlation between bacterial communitycomposition and metal content. Analogously, Hobbelen et al.(2004) observed no effects of metals on soil detrivores suchas isopods and earthworms in soils from floodplains of thelower river Rhine, close to the freshwater tidal range of theriver near our locations in the Biesbosch. Hobbelen et al.(2004) noted that the total metal concentrations in soil depositsexceeded Dutch soil quality criteria, but showed low availabil-ity in CaCl2 exchangeable fraction and consequently lowbioavailability, which may have prevented biological effects.Indeed, the metal concentrations in CaCl2 extracts of soilwere low in the Biesbosch and similar to those of unpollutedsoil (Hobbelen et al., 2004). In contrast, the metal concentra-tions in CaCl2 extracts of the present set of sediments covereda clear gradient and were sometimes high, but meanwhile, weobserved that metal concentrations showed strong correlationswith the contents of clay and organic matter. It is questionableif clay and organic matter bound metals are readily availableto algae and bacteria, even though they are CaCl2 extractable.The inconspicuous effect of metals on bacteria (in this study)and soil fauna (Hobbelen et al., 2004) and the more evidenteffects on invertebrates (De Haas et al., 2004; Van Griethuysenet al., 2004) may thus be caused by variable bioavailabilityrather than to differences in sensitivities of the studiedorganisms.
Changes in CLPP were also found to correlate with oxygenconcentration of the water layer above the sediment and withthe clay content of the sediment. During the sampling period,

889M.E.Y. Boivin et al. / Environmental Pollution 145 (2007) 884e894
Table 3
Algal species observed at the different locations
Algea spp.M J S M J S M J S M J S M J M J S S
Amphora sp.
Ankyra judayi
Asterionella formosa
Aulacoseira cf. ambigua
Aulacoseira granulata
Aulacoseira sp.
Caloneis amphisbaena
Caloneis sp.
Chlorella sp.
Chlorella vulgaris
Chloroccocum sp.
Closterium kuetzingii
Closterium limneticum
Cocconeis placentula
Cosmarium sp.
Cyclotella atomus
Cyclotella bodanica
Cyclotella glomerata
Cyclotella meneghiniana
Cymatopleura solea
Cymbella sp.
Diatoma vulgaris
Euglena sp.
Golenkenia cf. paucispina
Golenkinia radiata
Gomphonema affine
Gyrosigma cf. perkerii
Gyrosigma sp.
Keratococcus caudatus
Kirchneriella irregularis
Kirchneriella obesa
Leptolyngbya tenuis
Merismopedia glauca
Merismopedia sp.
Merispondia sp.
Microcystis sp.
Monoraphidium arcuatum
Monoraphidium contortum
Monoraphidium sp.
D1 D3 D4 R1 R2 R3 B1
Navicula capitata
Navicula capitatoradiata
Navicula cf. festiva
Navicula cryptocephala
Navicula halophila
Navicula phyllepta
Navicula pupula
Navicula trivialis
Navicula veneta
Nitzschia acicularis
Nitzschia capitata

890 M.E.Y. Boivin et al. / Environmental Pollution 145 (2007) 884e894
D1 D3 D4 R1 R2 R3 B1 B2 B3Algea spp.M MS
Nitzschia dissipata
Table 3 (continued)
Nitzschia gracilis
Nitzschia palea
Nitzschia pellucida
Nitzschia plana
Nitzschia pupula
Nitzschia pusilla
Nitzschia recta
Nitzschia sigmoidea
Nitzschia sublinearis
Nitzschia viridula var. rostellata
Oocistys cf. marssonnii
Oocystis pusilla
Oscillatoria sp.
Pediastrum boryanum
Pediastrum duplex
Pediastrum sp.
Phormidium tenue
Pinnularia microstauron
Planktothrix sp.
Pleurosigma angulatum
Pleurosigma sp.
Scenedesmus abundans
Scenedesmus acuminatus
Scenedesmus communis
Scenedesmus protuberans
Scenedesmus quadricauda var. longispina
Scenedesmus subapicatus
Scenesdesmus pecsensis
Staurastrum sp.
Stephanodiscus hantzschii
Stephanodiscus parvus
Tetraedron caudatum
Tetraedron sp.
Tetrastrum elegans
Tetrastrum triangulare
J J MS J MS J MS M J S S S SJ
White squares represent locations lacking the species, grey squares represent locations where the species was present in low numbers, and black squares represent
locations where the species was dominant. Synonyms: Sc. quadricauda is also known as Sc. communis; Sc. subapicatus as Sc. abundans; Navicula pupula as
Sellaphora pupula.
evaporation and eutrophication of the water caused large var-iations in the water pH, oxygen concentration and temperature,parameters all known to influence bacterial and algal commu-nities (Wu and Conrad, 2001; Eiler et al., 2003; Tank et al.,2003; Kirchman et al., 2004). Increased oxygen concentrationsare caused by photosynthetic activity of algal communities,but the abundance of algae at our field locations invites a fur-ther analysis of the potential interactions between bacteria andalgae beyond the effect of increased oxygen.
DGGE pattern shifts correlated with different algal species,Asterionella formosa, Aulacoseira granulata, Cyclotellaatomus, Euglena sp., Merismopedia sp., Navicula cfr. festivaand Oocystis marssonni. In agreement with the differences
in DGGE profiles, the CLPP profiles correlated with A.granulata, Chlorella sp., C. atomus, Gomphonema affine,O. marssonni and Scenedesmus quadricauda var. longispina.This matches well with the findings of Bell et al. (1984) dem-onstrating that the spectrum of organic carbon produced byparticular algae favoured particular bacteria. This was experi-mentally verified by Schafer et al. (2002), who observed thatlaboratory diatom cultures had distinct bacterial ‘satellite’ as-semblages. Besides these two studies, there is not muchevidence in natural waters that the composition of bacterialcommunities is linked to micro-algal species. Van der Guchtet al. (2001) indicated that bacterial diversity differed betweentwo freshwater bodies with markedly different densities of
891M.E.Y. Boivin et al. / Environmental Pollution 145 (2007) 884e894
-1.0 1.5-0.8
0.8
Sampling-months **
Clay**
A
-1.0 1.5-0.6
0.8
Clay**
cond µS/cm*
cond µS/cm*
B
Sampling-months**
Fig. 1. Redundancy analysis (RDA) based on algal community composition sampled at different locations and time of the year. (A) Only dominant algal species;
(B) all algal species. Circles: R locations; squares: location D1; triangles: location D3; diamonds: location D4; stars: B locations. Black symbols: May; grey sym-
bols: July; white symbols: September. *p < 0.05; **p < 0.01.
algae and dominance of Cyanobacteria versus other photo-trophs. In contrast, foodwebs of the floodplains of the riverRhine are not dominated by algal blooms, since also river de-posits provide new carbon sources for bacteria. The significant
relationships with local algal species are somewhat variable,depending on DGGE or CLPP serving as measures for bacte-rial community composition. Also some of these phototrophicspecies e.g. Euglena and Merismopedia are inconspicuous
100
9590858075706560
88
88
81
93
78
74
9678
75
69
8574
82
69
66
80
8280
70
59
Fig. 2. Dendrogram analysis based on DNA banding patterns from denaturing gradient gel electrophoresis (DGGE) of bacterial communities sampled at different
locations and time of the year. Circles: R locations; squares: location D1; triangles: location D3; diamonds: location D4; stars: B locations. Black symbols: May;
grey symbols: July; white symbols: September. The numbers at the nodes represent the similarity.

892 M.E.Y. Boivin et al. / Environmental Pollution 145 (2007) 884e894
-0.8 0.8-0.6
0.8
Sampling-months **
A
-1.0 1.0-0.6
0.8
77
Cyclotella atomus **
Navicula cfr.festiva*
Navicula cfr.festiva*
Oocystis marssonni**
Oocystis marssonni**
Oocystis marssonni**
Oocystis marssonni**
Asterionella formosa**
Aulacoseria granulata*Sampling-months ***
Sampling-month ***
Cyclotella atomus**
Euglena sp**
Euglena sp. *
77
B
-0.6 0.8-0.8
0.8C
-1.0 1.0-0.6
0.8D
-1.0 1.0-0.8
0.8E
Asterionella formosa*
Aulacoseria granulata*
Merispondia sp.*
Fig. 3. Redundancy analysis (RDA) based on DNA banding patterns from denaturing gradient gel electrophoresis (DGGE) of bacterial communities sampled at
different locations and time of the year. (A) Abiotic data set as explanatory variables; (B) dominant algal species as explanatory variables; (C) all algal species as
explanatory variables; (D) as A plus dominant algal species as explanatory variables; (E) as A plus all algal species as explanatory variables. Circles: R locations;
squares: location D1; triangles: location D3; diamonds: location D4; stars: B locations. Black symbols: May; grey symbols: July; white symbols: September.
*p < 0.05; **p < 0.01; ***p < 0.001.
components of the micro-phytobenthos, indicating organicallyrich and porous sands, respectively. The main components ofthe micro-phytobenthos in these floodplain lakes, Nitzschiaand Navicula species have been shown to be stable (Van derGrinten et al., 2004) and thus may not show up in the presentcorrelations despite their potential impact on the biodiversityof the bacteria.
5. Conclusion
We conclude that the significant, but somewhat variable cor-relations between the bacterial community composition andsediment living micro-algal species must be interpreted as an in-dication for a strong and species-specific linkage between bac-terial and algal species. We cannot prove, however, that metals

893M.E.Y. Boivin et al. / Environmental Pollution 145 (2007) 884e894
-0.8 0.8-0.6
1.0
Sampling-months **
Conductivity *
Cyclotella atomus **
Chlorella sp.*
Aulacoseira granulata *
Cyclotella atomus **
Gomphonema affine *
Aulacoseira granulata *
Aulacoseira granulata *
Chlorella sp. *
O2 *
ACd (total destruction) ***
pHwater *
-1.0 1.0
-1.0 1.0
-0.8
1.0
-0.8
1.0
B
-0.6 1.0-1.0
1.0
Scenedesmus quadricauda var. Longispina **
Oocystis marssonni **
Oocystis marssonni *
Oocystis marssonni *
Aulacoseria granulata *
C
-0.8 0.8-0.6
1.0D
Cd.T ***
Sampling-months **
O2 *E Clay **
Fig. 4. Redundancy analysis (RDA) based on community-level physiological profiles (CLPP) of bacterial communities sampled from different locations and time
of the year. (A) Abiotic data set as explanatory variables; (B) dominant algal species as explanatory variables; (C) all algal species as explanatory variables; (D) as
A plus the dominant algal species as explanatory variables; (E) as A and all algal species as explanatory variables. Circles: R locations; squares: location D1;
triangles: location D3; diamonds: location D4; stars: B locations. Black symbols: May; grey symbols: July; white symbols: September. *p < 0.05; **p < 0.01.
affect either the algal community or the bacterial community, orthat the interaction of algae and bacteria is critically affected.
Acknowledgements
We thank the NIOO for allowing us to use their facilities,Paul van der Brink for his statistical advice, and Willem
Roelofs for his help in building a macro to treat the data.We also thank the Dutch Organisation for Scientific Research(NWO), the Stimulation Program on System-Oriented Ecotox-icological Research (SSEO), and the Dutch National Instituteof Public Health and the Environment (RIVM) for funding theproject under number 0123.060/061 (NWO/SSEO) and projectnumber M/860711 (RIVM).

894 M.E.Y. Boivin et al. / Environmental Pollution 145 (2007) 884e894
References
Baath, E., Anderson, T.H., 2003. Comparison of soil fungal/bacterial ratios in
a pH gradient using physiological and PFLA-based techniques. Soil Biol-
ogy and Biochemistry 35, 955e963.
Baath, E., Dıaz Ravina, M., Frostegard, A., Campbell, C.D., 1998. Effect of
metal-rich sludge amendments on the soil microbial community. Applied
and Environmental Microbiology 64, 238e245.
Babich, H., Stotzky, G., 1985. Heavy metal toxicity to microbe-mediated eco-
logic processes: a review and potential application to regulatory policies.
Environmental Research 36, 111e137.
Bell, R.A., Ogden, N., Kramer, J.R., 1984. The biotic ligand model and a cel-
lular approach to class B metal aquatic toxicity. Comparative Biochemistry
and Physiology Part C: Toxicology and Pharmacology 133, 175e188.
Blanck, H., Wangberg, S.A., Molander, S., 1988. Pollution-Induced Commu-
nity Tolerance. A new ecotoxicological tool. In: Cairns, J.J., Pratt, J.R.
(Eds.), Functional testing of Aquatic Biota for Estimating Hazards of
Chemicals, pp. 219e230 (ASTM STP 988).
Boivin, M.E.Y., Breure, A.M., Posthuma, L., Rutgers, M., 2002. Determination
of field effects of contaminantsdsignificance of pollution-induced commu-
nity tolerance. Human and Ecological Risk Assessment 8, 1035e1055.
Boivin, M.E.Y., Massieux, B., Breure, A.M., Van Den Ende, F.P., Greve, G.D.,
Rutgers, M., Admiraal, W., 2005. Effects of copper and temperature on
structural and functional traits of aquatic bacterial communities. Aquatic
Toxicology 71, 345e356.
De Haas, E.M., Paumen, M., Koelmans, A.A., Kraak, M.H.S., 2004. Com-
bined effects of copper and food on the midge Chironomus riparius in
whole sediment bioassays. Environmental Pollution 127, 99e107.
Eiler, A., Farnleitner, A.H., Zechmeister, T.C., Herzig, A., Hurban, C.,
Wesner, W., Krachler, R., Velimirov, B., Kirschner, A.K.T., 2003. Factors
controlling extremely productive heterotrophic bacterial communities in
shallow soda pools. Microbial Ecology 46, 43e54.
Gong, P., Siciliano, S.D., Srivastava, S., Greer, C.W., Sunahara, G.I., 2002.
Assessment of pollution-induced microbial community tolerance to heavy
metals in soil using ammonia-oxidizing bacteria and biolog assay. Human
and Ecological Risk Assessment 8, 1067e1081.
Hobbelen, P.H.F., Koolhaas, J.E., Van Gestel, C.A.M., 2004. Risk assessment
of heavy metal pollution for detritivores in floodplain soils in the Bies-
bosch, the Netherlands, taking bioavailability into account. Environmental
Pollution 129, 409e419.
Kirchman, D.L., Dittel, A.I., Findlay, S.E.G., Fischer, D., 2004. Changes in bac-
terial activity and community structure in response to dissolved organic mat-
ter in the Hudson River, New York. Aquatic Microbial Ecology 35, 243e257.
Konert, M., Van den Berghe, J., 1997. Comparison of laser grain size analysis
with pipette and sieve analysis: a solution for the underestimation of the
clay fraction. Sedimentology 44, 523e535.
Muyzer, G., Dewaal, E.C., Uitterlinden, A.G., 1993. Profiling of complex
microbial-populations by denaturing gradient gel electrophoresis analysis
of polymerase chain reaction-amplified genes coding for 16S ribosomal-
RNA. Applied and Environmental Microbiology 59, 695e700.
Nold, S.C., Zwart, G., 1998. Patterns and governing forces in aquatic micro-
bial communities. Aquatic Ecology 32, 17e35.
Rutgers M., Sweegers, B.M.C., Wind, B.S., van Veen, R.P.M., Folkerts, A.J.,
Posthuma, L., Breure, A.M., 1998. Pollution-induced community tolerance
in terrestrial microbial communities, In: Contaminated Soil ’98. Proceed-
ings of the Sixth International FZK/TNO Conference on Contaminated
Soil, London, UK, pp. 337e343.
Rutgers, M., Breure, A.M., Insam, H., 2006. Substrate utilization in Biolog
plates for analysis of CLPP. In: Bloem, J., Benedetti, A., Hopkin, D.W.
(Eds.), Microbiological Methods for Assessing Soil Quality. CAB,
Wallingford, pp. 212e227.
Schafer, H., Abbas, B., Witte, H., Muyzer, G., 2002. Genetic diversity of ‘‘sat-
ellite’’ bacteria present in cultures of marine diatoms. FEMS Microbiology
Ecology 42, 25e35.
Tank, S.E., Schindler, D.W., Arts, M.T., 2003. Direct and indirect effects of
UV radiation on benthic communities: epilithic food quality and inverte-
brate growth in four montane lakes. Oikos 103, 651e667.
Ter Braak, C.J.F., 1986. Canonical correspondence analysis: a new eigenvector
technique for multivariate direct gradient analysis. Ecology 67,
1167e1179.
Ter Braak, C.J.F., Smilauer, P., 1998. Canoco Reference Manual and User’s
Guide to Canoco for Windows: Software for Canonical Community Ordi-
nation (version 4). Centre for Biometry, Wageningen.
Tyler, G., 1974. Heavy metal pollution and soil enzymatic activity. Plant and
Soil 41, 303e311.
Van Beelen, P., Wouterse, M., Posthuma, L., Rutgers, M., 2004. Location-
specific ecotoxicological risk assessment of metal-polluted soils.
Environmental Toxicology and Chemistry 23, 2769e2779.
Van der Grinten, E., Simis, S.G.H., Barranguet, C., Admiraal, W., 2004. Dom-
inance of diatoms over cyanobacterial species in nitrogen-limited biofilms.
Archiv fur Hydrobiologie 161, 98e111.
Van der Gucht, K., Sabbe, K., De Meester, L., Vloemans, N., Zwart, G.,
Gilles, M., Vyverman, W., 2001. Contrasting bacterioplankton community
composition and seasonal dynamics in two neighbouring hypertrophic
freshwater lakes. Environmental Microbiology 3, 680e690.
Van Griethuysen, C., van Baren, J., Peeters, E.T.H.M., Koelmans, A.A.,
2004. Trace metal availability and effect on benthic community structure
in floodplain lakes. Environmental Toxicology and Chemistry 23,
668e681.
Wu, X.L., Conrad, R., 2001. Functional and structural response of a cellulose-
degrading methanogenic microbial community to multiple aeration stress
at two different temperatures. Environmental Microbiology 3, 355e362.




















