
Metabolite profiling of the benthic diatom Cocconeis scutellum by GC-MS
12
Metabolite profiling of the benthic diatom Cocconeis scutellum by GC-MS Michela Nappo & Strahil Berkov & Carles Codina & Conxita Avila & Patrizia Messina & Valerio Zupo & Jaume Bastida Received: 12 March 2008 / Revised and accepted: 25 July 2008 / Published online: 11 September 2008 # Springer Science + Business Media B.V. 2008 Abstract Cocconeis scutellum is a benthic diatom pro- ducing one or more compounds responsible for the early programmed cell death (apoptosis) of the male gonad and the androgenic gland of the protandric shrimp Hippolyte inermis Leach. The metabolite composition of both the ether and butanol extracts was studied by gas chromatog- raphy-mass spectrometry (GC-MS) in both electron impact (EI) and chemical ionization (CI) mode. The compounds were identified as trimethylsilyl (TMSi) derivatives. The structure of the fatty acids (FA) was confirmed after conversion to their methyl esters. The ether fraction of C. scutellum contained FA (76%), of which 30% were saturated (SFA), 24% monounsaturated (MUFA) and 22% polyunsaturated (PUFA). Mono- (8%) and diglycerides (3%) as well as sterols (5%) and isoprenoid compounds (4%) were also found in this fraction. The butanol extract consisted of FA (45%), carbohydrates (25%), amino acids and N-containing metabolites (10%), fatty alcohols (9%), glycerides (4%) and organic acids (3%). In the literature, many reports deal with the chemistry and the chemical ecology of planktonic diatoms; contrastingly, benthic species are still less studied, especially with respect to their chemical composition, due to their difficult cultiva- tion. Hence, our investigation represents a preliminary approach to clarify on chemical bases the ecological role of Cocconeis on decapods in the marine benthos, as well as a description of the metabolic pattern of this benthic diatom. Keywords Cocconeis scutellum . Hippolyte inermis . Apoptosis . Metabolites . GC-EI/MS . GC-CI/MS Introduction Diatoms are eukaryotic unicellular algae characterized by a siliceous cell wall (Lebeau and Robert 2003) and belonging to the class Bacillariophyceae. They are widely distributed in most aquatic habitats, and they are responsible for more than 50% of marine primary production (Nelson et al. 1995). Furthermore, they play a key role in the global carbon cycle (Pohnert 2005), constituting a rich diet for many herbivores (Newell and Newell 1963; Raymont 1983) both in the marine benthos and in the water column, due to their high concentration of lipids (Sicko-Goad and Andresen 1991). This study is based on previous investigations dealing with the influence of benthic diatoms (i.e., living close to the sea bottom) of the genus Cocconeis on the sex reversal of the shrimp Hippolyte inermis Leach 1915 (Zupo 1994, 2000, 2001). This small decapod lives in shallow waters of the Mediterranean Sea and along the Atlantic coast of Spain (Zariquey 1968), where it forms stable populations in seagrass meadows (Gambi et al. 1992). H. inermis is a protandric crustacean (i.e., individuals undergo J Appl Phycol (2009) 21:295–306 DOI 10.1007/s10811-008-9367-8 M. Nappo : S. Berkov : C. Codina : J. Bastida (*) Departament de Productes Naturals, Biologia Vegetal i Edafologia, Facultat de Farmàcia, Universitat de Barcelona, Av. Joan XXIII s/n, 08028 Barcelona, Catalunya, Espanya e-mail: [email protected] C. Avila Departament de Biologia Animal (Invertebrats), Facultat de Biologia, Universitat de Barcelona, Av. Diagonal 645, 08028 Barcelona, Catalunya, Espanya P. Messina : V. Zupo Stazione Zoologica “A. Dohrn”, Laboratorio di Ecologia del Benthos, Punta S. Pietro, 80077 Ischia, Napoli, Italia
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Metabolite profiling of the benthic diatom Cocconeis scutellum by GC-MS

Metabolite profiling of the benthic diatom Cocconeisscutellum by GC-MS
Michela Nappo & Strahil Berkov & Carles Codina &
Conxita Avila & Patrizia Messina & Valerio Zupo &
Jaume Bastida
Received: 12 March 2008 /Revised and accepted: 25 July 2008 /Published online: 11 September 2008# Springer Science + Business Media B.V. 2008
Abstract Cocconeis scutellum is a benthic diatom pro-ducing one or more compounds responsible for the earlyprogrammed cell death (apoptosis) of the male gonad andthe androgenic gland of the protandric shrimp Hippolyteinermis Leach. The metabolite composition of both theether and butanol extracts was studied by gas chromatog-raphy-mass spectrometry (GC-MS) in both electronimpact (EI) and chemical ionization (CI) mode. Thecompounds were identified as trimethylsilyl (TMSi)derivatives. The structure of the fatty acids (FA) wasconfirmed after conversion to their methyl esters. Theether fraction of C. scutellum contained FA (76%), ofwhich 30% were saturated (SFA), 24% monounsaturated(MUFA) and 22% polyunsaturated (PUFA). Mono- (8%) anddiglycerides (3%) as well as sterols (5%) and isoprenoidcompounds (4%) were also found in this fraction. The butanolextract consisted of FA (45%), carbohydrates (25%), aminoacids and N-containing metabolites (10%), fatty alcohols
(9%), glycerides (4%) and organic acids (3%). In theliterature, many reports deal with the chemistry and thechemical ecology of planktonic diatoms; contrastingly,benthic species are still less studied, especially with respectto their chemical composition, due to their difficult cultiva-tion. Hence, our investigation represents a preliminaryapproach to clarify on chemical bases the ecological role ofCocconeis on decapods in the marine benthos, as well as adescription of the metabolic pattern of this benthic diatom.
Keywords Cocconeis scutellum .Hippolyte inermis .
Apoptosis . Metabolites . GC-EI/MS . GC-CI/MS
Introduction
Diatoms are eukaryotic unicellular algae characterized by asiliceous cell wall (Lebeau and Robert 2003) and belongingto the class Bacillariophyceae. They are widely distributedin most aquatic habitats, and they are responsible for morethan 50% of marine primary production (Nelson et al.1995). Furthermore, they play a key role in the global carboncycle (Pohnert 2005), constituting a rich diet for manyherbivores (Newell and Newell 1963; Raymont 1983) bothin the marine benthos and in the water column, due to theirhigh concentration of lipids (Sicko-Goad and Andresen1991). This study is based on previous investigationsdealing with the influence of benthic diatoms (i.e., livingclose to the sea bottom) of the genus Cocconeis on the sexreversal of the shrimp Hippolyte inermis Leach 1915 (Zupo1994, 2000, 2001). This small decapod lives in shallowwaters of the Mediterranean Sea and along the Atlanticcoast of Spain (Zariquey 1968), where it forms stablepopulations in seagrass meadows (Gambi et al. 1992). H.inermis is a protandric crustacean (i.e., individuals undergo
J Appl Phycol (2009) 21:295–306DOI 10.1007/s10811-008-9367-8
M. Nappo : S. Berkov :C. Codina : J. Bastida (*)Departament de Productes Naturals,Biologia Vegetal i Edafologia, Facultat de Farmàcia,Universitat de Barcelona,Av. Joan XXIII s/n,08028 Barcelona, Catalunya, Espanyae-mail: [email protected]
C. AvilaDepartament de Biologia Animal (Invertebrats),Facultat de Biologia, Universitat de Barcelona,Av. Diagonal 645,08028 Barcelona, Catalunya, Espanya
P. Messina :V. ZupoStazione Zoologica “A. Dohrn”,Laboratorio di Ecologia del Benthos,Punta S. Pietro,80077 Ischia, Napoli, Italia

a male stage before switching to female about a year afterhatching), with a life cycle characterized by 2-yearlyperiods of recruitment: autumn and spring. Autumn-borndecapods experience a male phase before reverting tofemales (α females, according to Zupo 1994) in the nextspring. The sex change naturally occurs in H. inermis about7–12 months after birth, while the female gonad derivesfrom undifferentiated tissues only after the whole disruptionof the testes (Reverberi 1950). Spring recruits, instead,show a different sexual maturation: they immediatelyproduce in the field both male and female individuals.The seasonal difference has been explained by the influenceof various species of benthic diatoms upon which thecrustaceans feed. These diatoms are epiphytes of Posidoniaoceanica leaves and bloom mainly in spring, while they arescarcely represented in the periphyton community inautumn (Buia et al. 2000; Zupo 2001). It was hypothesizedthat dietary diatoms contain one or more compounds able todestroy selectively the male gonad of H. inermis during itsdevelopment, thus producing primary (β) females (Zupo1994, 2000; Zupo et al. 2007). The effect of diatoms wouldbe triggering the cell death in postlarva’s male gonad, aprocess that in absence of the microalgae would happenabout 1 year after birth in mature male individuals (Zupo2000). The action of the diatom apoptotic compound(s) isspecies-specific and extremely selective for the male gonadand the androgenic gland of H. inermis (Zupo and Messina2007). This shrimp is largely adapted to P. oceanica’s lifecycle (d’Udekem d’Acoz 1996), and the effect of thedietary microalgae could be a spring signal for thedevelopment of β females, whose presence is crucial forkeeping a constant sex ratio (Zupo 1994; Buia et al. 2000).Our recent studies indicated that C. scutellum is the specieswith the major effect in the sex reversal of H. inermis(authors, unpublished data). This diatom plays a key functionin the ecology of the crustacean, resulting in a large number offemales (α and β) for the next autumn mating period. Such amechanism of regulation is unknown for planktonic diatoms(i.e., free-floating organisms) which, contrastingly, exercisedeleterious effects towards their crustacean predators (Miraltoet al. 1995, 1999; Ianora et al. 2004). In fact, diatoms likeSkeletonema costatum and Thalassiosira rotula have abor-tive and teratogenic effects on copepods by production ofoxylipins, mainly aldehydes, derived from the oxidation ofpolyunsaturated fatty acids (PUFA) (Fontana et al. 2007).The chemical ecology of planktonic diatoms is welldocumented (Pohnert and Boland 2002; Pohnert 2005;Fontana et al. 2007); in contrast, few studies are reportedabout benthic diatoms and even fewer about their metabolitecomposition (Simental et al. 2001) and effects on otherorganisms (Taylor et al. 2007). One of the reasons for thelimited information in this field is that benthic diatoms aremuch more difficult to sample, quantify and cultivate than
planktonic ones. In particular, Cocconeis is characterized bylow growth rates and strong adhesion to the substratum,which makes the isolation of the diatoms and the manipu-lation of their cultures difficult (Raniello et al. 2007).
In the related literature on diatom chemistry, greatattention is paid to the lipid fractions due to their nutritionalvalue in the marine food web (Newell and Newell 1963;Raymont 1983; Gordon et al. 2006). Sterol fractions aremainly analyzed as TMSi derivatives or without derivati-zation (Gladu et al. 1991; Souchet and Laplante 2007)while fatty acids (FA) are usually identified by GC-MS astheir methyl esters (FAME) (Mjos and Pettersen 2003;Rousch et al. 2003; Dayhuff and Wells 2005; Gordon et al.2006). Various analytical methods, including HPLC-MSand GC-MS, have been reported for analysis of free aminoacids and carbohydrates in algae (Hokputsa et al. 2003;Gordon et al. 2006). Several papers on metabolite profilinghave proved that GC-MS is a reliable, reproducible, rapid,selective and sensitive method requiring a small biomassfor simultaneous identification of plant metabolites fromvarious classes, such as amino acids, phenolic compounds,saccharides, organic acids, FA, sterols, glycerides, etc. afterderivatization (Roessner et al. 2000; Benkeblia et al. 2007;Schliemann et al. 2008).
To our knowledge, no detailed reports on the metaboliteprofiling of benthic diatoms have been published. Hencethe objective of this study was to characterize the metabolicpattern of these diatoms as a basis to clarify their ecologicalrole on selected decapods in the marine benthos.
Materials and methods
Cocconeis scutellum diatoms were isolated and cultivated atthe Stazione Zoologica “A. Dohrn” of Ischia, Naples, Italy.Axenic cultures were cultivated in sterilized Petri dishes(14 cm diameter) containing 100 mL of F2 medium (SigmaAldrich) and incubated in a thermostatic chamber (18°C,12/12 h photoperiod). After 16 days, the Petri dishes wereopened, the culture medium was drained and the disheswere washed twice with distilled water, then freeze-driedand scraped with a metal blade, to collect the producedbiomass (Zupo et al. 2007; Raniello et al. 2007).
Extraction
The extraction of freeze-dried diatoms of C. scutellum (1.5 gdry wt) was carried out in agreement with the protocoldescribed by Gavagnin et al. (2005). The material waspreviously reconstituted in distilled water (50 mL) and thentreated with acetone (3×100 mL) at room temp. in anultrasound bath for 2–3min, in order to increase themembranebreakdown and the release of compounds from the cells. The
296 J Appl Phycol (2009) 21:295–306

suspension was filtered and the filtrate was concentrated byrotary evaporator. The residual water was extracted withdiethyl ether (3×150 mL) by means of a separatory funnel.After separation, the ether phase was treated with anhydrousNa2SO4 and evaporated to dryness under reduced pressure,obtaining 182 mg of dry extract. The aqueous residue wasfurther extracted with n-butanol (100 mL). The n-butanolsoluble part was evaporated until dryness and yielded 74 mg.The extraction with diethyl ether and n-butanol allows for arepartition of the acetone-extracted compounds in apolar andpolar fractions. The remaining water phase was lyophilizedand then weighted to obtain 8 mg of residue.
In order to search for aldehydes, another diatom sample(1.0 g dry wt) was sonicated in distilled water for 1 min andleft on the bench for 30 min. Acetone was added and theresulting suspension was centrifuged three times at 4,000gfor 10 min. The supernatant was exhaustively extractedwith CHCl3. The organic layers were combined, dried overanhydrous Na2SO4 and then evaporated under reducedpressure, affording 91 mg of dry extract.
Derivatization
Ether fraction (ca. 2–3 mg) was silylated with 50 μL of N,O-bis-(trimethylsilyl)trifluoro-acetamide (BSTFA) in 50 μLof pyridine for 2 h at 50°C. Butanol fraction (ca. 2–3 mg)was first methoxymated with 100 μL of methoxyaminehydrochloride (MOA, 20 mg mL−1 in pyridine) for 90 minat 50°C to stabilize the carbonyl moiety and prevent ringformation in sugars, then silylated by addition of 100 μL ofBSTFA and heating at 50°C for 1 h. The mixtures wereevaporated under a stream of N2 and redissolved in 100 μLof chloroform for further GC-MS analysis. In parallel,FAME were obtained from ca. 5 mg of dry ether fractiondissolved in 2 mL of 1.5% solution of HCl in absolutemethanol and heated overnight (12 h) at 60°C. Aftercooling, 2 mL of water containing 5% of NaCl were addedto the reaction mixture and the acid solution was extractedwith n-hexane (2×3 mL). The organic layer was separatedand washed with 3 mL of water containing 5% of Na2CO3
and dried over anhydrous Na2SO4. After evaporation of then-hexane, the residue was dissolved in 100 μL ofchloroform for further GC-MS analysis.
The chloroform extract (ca. 3 mg) was treatedwith (carbetoxyethylidene)-triphenylphosphorane (CET-triphenylphosphorane) in order to converse eventual un-stable aldehydes into ethyl esters by Witting reaction(d’Ippolito et al. 2002).
GC-MS analysis
The data were recorded on a Hewlett Packard 6890+MSD5975 (Hewlett Packard, Palo Alto, CA, USA) operating in
EI and CI mode with a HP-5 MS capillary column (30 m×0.25 mm×0.25 μm). The temperature program was: 100–180°C at 15°C min−1, 1 min at 180°C, 180–300°C at 5°Cmin−1, and 1 min held at 300°C. Injector temperature was280°C. The flow rate of carrier gas (helium in EI mode andmethane in CI mode) was 0.8 mL min−1 and the electronpotential was 70 eV. In order to detect aldehydes, the GC-MS conditions were the same as those reported in theliterature (d’Ippolito et al. 2002).
Identification of metabolites
The metabolites of the ether extract were identified as TMSiderivatives comparing their mass spectra and KovatsIndexes (RI) with those of an on-line available plant-specific database (The Golm Metabolome Database; http://csbdb.mpimp-golm.mpg.de/csbdb/gmd/home/gmd_sm.html) and the NIST 05 database. Additionally, the structureof FA was confirmed after their conversion to FAMEconsidering the characteristic mass spectral fragmentationof each subgroup, molecular ion, retention time and massspectra available in the on-line lipid library (http://www.lipidlibrary.co.uk/ms/ms01/index.htm), NIST 05 databaseand literature data as indicated in Table 1. Peaks in theGC-MS of the butanol extract were identified as TMSi and/or MOA derivatives with the help of the NIST 05 databaseand on-line available plant-specific database (The GolmMetabolome Database) as well as with literature data on thebasis of the match of mass spectra and RI as indicated inTable 1. The measured mass spectra were deconvoluted bythe Automated Mass Spectral Deconvolution and Identifica-tion System (AMDIS), before comparison with the databases.Then, the spectra of individual components were transferredto the NIST Mass Spectral Search Program MS Search 2.0where they were matched against reference compounds of theNIST Mass Spectral Library 2005 and the Golm MetabolomeDatabase. RI of the compounds were recorded with standardn-hydrocarbon calibration mixture (C9-C36) (Restek, Cat no.31614, supplied by Teknokroma, Spain) using AMDIS 3.6software. The search for aldehydes was performed afterderivatization of the chloroform extract with CET-phosphor-ane. The MS data were deconvoluted by AMDIS as above,transferred to the NIST program and compared with thelibrary of derivatized standards which we previously built.
Results
The GC-MS analysis of both the ether and butanol fractionsresulted in the detection of more than 150 metabolites,about 100 of which were identified. The metabolite patternofC. scutellum is presented in Tables 1 and 2. The extraction
J Appl Phycol (2009) 21:295–306 297

Table 1 Metabolites of C. scutellum presented as % of TIC
Compounds Rt Key ions (m/z)* Ether extract Butanol extract
1 L-Alanine a, b 4.00 116, 147, 180 0.142 L-Valine a, b 5.16 144, 218, 246 0.233 Urea a, b 5.33 147, 189 2.344 UC 5.71 0.125 L-Leucine a, b 5.72 158, 232, 260 0.336 Glycerol a, b 5.75 205, 147, 218 0.07 8.627 Phosphoric acid a, b 5.79 299, 314 0.018 L-Isoleucine a, b 5.95 158, 218 0.269 Succinic acid a, b 6.09 147, 247 0.2010 UC 6.14 0.9611 UC 6.38 0.4012 L-Serine a, b 6.59 204, 218, 278 0.0713 L-Threonine a, b 6.86 218, 219, 291 0.0314 Glutaric acid a, b 6.94 147, 216, 158 0.0715 Decanoic acid (10:0) a, b, c 7.40 117, 229, 244** 0.02 0.0416 Aspartic acid a, b 7.65 232, 334 0.0117 Dodecanol a, b, c 8.49 243***, 75, 103 0.0218 Hexadecane a, b, c 8.82 57, 71, 226** 0.0119 Tridecanoic acid ME (13:0) h 9.10 74, 87, 228** tr20 Ornithine a, b 9.11 142, 204, 348** 0.0821 1-Tetradecanol a, b, c 9.20 271***, 75, 103 0.0122 L-Phenylalanine a, b 9.30 218, 192, 294*** 0.2723 Pentonic acid-1,4-lactone b 9.43 217, 147, 231 0.2824 Dodecanoic acid (12:0) a, b, c 9.44 117, 257***, 272** 0.0425 Arabinose 1 a, b, d 9.50 217, 230, 191 0.1526 Arabinose 2 a, b, d 9.68 217, 230, 191 0.5327 UM 9.88 0.4128 Heptadecane a, b, c 10.01 57, 71, 240** 0.0129 1-Pentadecanol a, b, c 10.34 285***, 103, 75 tr30 UM 10.65 3.5631 Glycerol-2-phosphate a, b 10.67 243, 299, 445*** 0.2432 Hydroxycinnamic acid a, b 10.94 179, 192, 310** 0.0633 Glycerol-3-phosphate a, b 11.17 357, 299, 445*** 0.02 0.0934 UC 11.23 0.0735 Octadecane a, b, c 11.33 57, 71, 254** 0.0136 UC 11.44 0.3937 2-Ketogluconic acid a, b 11.50 292, 217, 305 1.7838 Hypoxanthine a, b 11.63 265***, 280** 0.2839 9-Tetradecenoic acid (14:1Δ9) a, b, c 11.78 117, 298***, 283** 0.02 6.9440 UC 11.93 0.4041 Tetradecanoic acid (14:0) a, b, c 12.13 117, 285***, 300** 7.18 6.9442 Galactofuranose a, b, d 12.42 217, 191, 204 0.3443 Adenine a, b 12.54 264***, 279** 0.4644 UM 12.67 0.1245 9,12-Hexadecadienoic acid ME (16:2 Δ9,12) h 12.70 81, 95, 266** tr46 UM 13.00 0.5947 UC 13.01 0.4548 Talose a, b, d 13.19 204, 191, 217 3.4749 Pentadecanoic acid (15:0) a, b, c 13.55 117, 299***, 314** 6.89 0.4450 L-Tyrosine a, b 13.68 218, 280, 382*** 0.3251 1-Hexadecanol a, b, c 13.75 75, 103, 299*** 0.1052 UC 14.22 0.1053 6,9,12,15-Hexadecatetraenoic acid (16:4Δ6,9,12,15) c 14.53 79, 305***, 320** 4.1654 6,9,12-Hexadecatrienoic acid (16:3Δ6,9,12) b, c 14.60 79, 307***, 322** 0.9255 Glucose a, b, d 14.61 204, 191, 217 3.9956 9-Hexadecenoic acid (16:1Δ9) a, b, c 14.96 117, 311***, 326** 21.74 5.64
298 J Appl Phycol (2009) 21:295–306

Table 1 (continued)
Compounds Rt Key ions (m/z)* Ether extract Butanol extract
57 11-Hexadecenoic acid (16:1Δ11) a, c 15.03 117, 311***, 326** 0.3758 Hexadecanoic acid (16:0) a, b, c 15.23 117, 313***, 328** 12.29 16.5959 UC 15.34 79, 91, 108 0.2860 Heneicosane a, b, c 15.92 57, 71, 296** 0.0261 Inositol a, b, d 16.34 305, 217, 191 0.1262 Heptadecanoic acid (17:0) a, b, c 16.67 117, 327***, 342** 0.0163 Octadecanol a, b, c 16.87 327***, 75, 103 0.08 0.1964 Phytol a, b 17.23 143, 123, 353*** 0.74 0.3465 Octadecanoic acid polyene (18:?5) 17.31 79, 91, 117 0.0666 6,9,12-Octadecatrienoic acid (18:3Δ6,9,12) a, b, c 17.47 79, 335***, 350** 0.3467 6,9,12,15-Octadecatetraenoic acid (18:4Δ6,9,12,15) c 17.59 79, 333***, 348** 0.7868 9,12-Octadecadienoic acid (18:2Δ9,12) a, b, c 17.76 79, 337***, 352** 0.3169 cis-11-Octadecenoic acid (18:1Δ11) a, b, c 17.86 117, 339***, 354** 0.69 0.1170 trans-11-Octadecenoic acid (18:1Δ11) a, b, c 17.97 117, 339***, 354** 1.39 1.0671 Octadecanoic acid (18:0) a, b, c 18.26 117, 341***, 356** 0.70 0.7672 UC 18.36 0.1573 UM 18.69 0.2574 Tricosane a, b, c 19.08 57, 71, 324** 0.1375 4,8,12,16-Tetramethylheptadecane-4-olide a, b 19.99 99, 114, 324** 0.0276 2-O-Glycerol-D-galactopyranoside (floridoside) a, b, e 20.16 204, 337, 671*** 11.2277 Arachidonic acid (20:4 Δ5,8,11,14) a, c 20.28 117, 361***, 376** 2.68 0.2078 5,8,11,14,17-Eicosapentaenoic acid (20:5Δ5,8,11,14,17) c 20.51 117, 359***, 374** 12.44 1.3379 Eicosatrienoic acid (20:3) c 20.61 129, 363***, 378** 0.0580 Tetracosane a, b, c 20.66 57, 71, 338** 0.1081 1-Monotetradecanoylglycerol (14:0) a, c 20.86 343, 147, 431** 0.36 0.2282 11-Eicosenoic acid (20:1Δ11) a, b, c 21.07 117, 367***, 382** 0.0483 Eicosanoic acid (20:0) a, b, c 21.36 117, 369***, 384** 1.51 2.8884 Uridine a, b 21.78 217, 259, 445*** 0.4485 Pentacosane a, b, c 22.17 57, 71, 352** 0.1186 2-Monohexadecenoylglycerol (16:1) c 23.03 129, 457***, 472** 1.05 0.0687 UC 23.29 0.5788 2-Monohexadecanoylglycerol (16:0) a, c 23.30 129, 218, 459*** 1.30 0.1289 1-Monohexadecenoylglycerol (16:1) c 23.54 369, 457***, 472** 2.05 0.4090 Hexacosane a, b, c 23.67 57, 71, 366** 0.0591 1-Monohexadecanoylglycerol (16:0) a, b, c 23.80 371, 459***, 474** 2.94 1.1292 Docosanoic acid (22:0) a, b, c 24.30 117, 397***, 412** 0.11 0.2593 Adenosine a, b 24.70 230, 445***, 460** 4.2794 Sucrose a, b, d 25.26 361, 217, 437 0.8795 2-Monooctadecenoylglycerol (18:1) c 25.81 103, 339, 485*** 0.1096 1-Monooctadecenoylglycerol (18:1) a, c 26.28 397, 485***, 500** 0.0897 1-Monooctadecanoylglycerol (18:0) a, b, c 26.51 399, 487*** 0.0398 Tetracosenoic acid (24:1) c 26.76 117, 423***, 438** 0.0799 Squalene a, b 26.95 69, 81, 410** 0.15100 Tetracosanoic acid (24:0) a, b, c 27.08 117, 425***, 440** 0.70 1.33101 UC 27.58 0.61102 UC 28.19 0.33103 Sterol A a, c 28.50 380**, 255 0.27 0.56104 1-Monoeicosanoylglycerol (20:0) a, c 29.12 427, 515*** 0.03105 Triacontane a, b, c 29.18 57, 71, 422** 0.02106 Hexacosanoic acid (26:0) a, b, c 29.71 117, 453***, 468** 0.01 0.06107 β-Tocopherol a 29.94 151, 416** tr108 α-Tocopherol a, b 31.22 502**, 237 2.63 0.82109 Cholesterol a, b, c 31.29 329, 368, 458** 0.07 0.08110 Brassicasterol a, c 31.86 255, 380, 470** 0.10 0.49111 UC 31.96 0.77112 24-Methylenecholesterol f 32.57 314, 271, 398** 3.33 1.26
J Appl Phycol (2009) 21:295–306 299
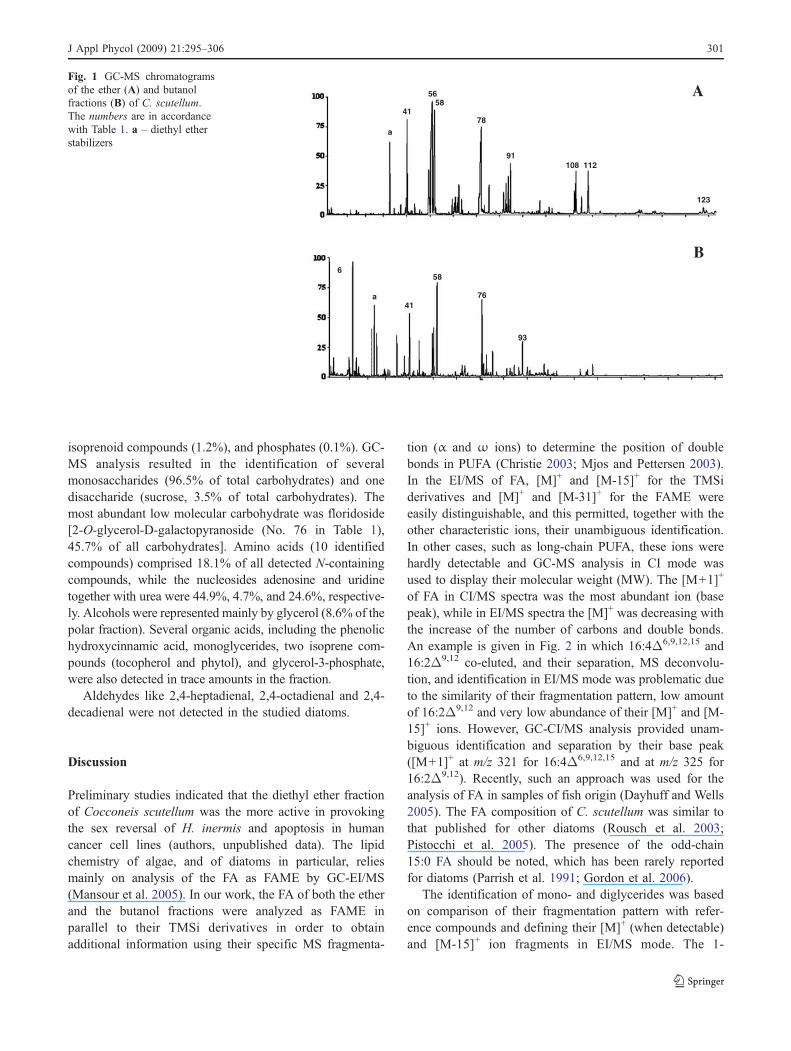
procedure allowed for partitioning the diatom compoundsinto an apolar ether fraction (12% of the total diatombiomass) and a polar butanol fraction (5% of the originalsample). Representative chromatograms of both the ether andbutanol fractions are shown in Fig. 1. All the identifiedcompounds and those still to be characterised (but compris-ing more than 0.2% of the total ion current, TIC) areincluded in the tables. The chemical pattern of the etherfraction consisted of FA (75.8%), glycerides (10.8%), sterols(4.7%), isoprenoid compounds (3.6%) as well as alkanes,fatty alcohols and phosphates (Table 2). It was characterizedby FA from C10 to C26, dominated (expressed as percent oftotal FA) by C16 and C20: 16:0 (16.2%), 16:1Δ9 (29.2%),16:4Δ6,9,12,15 (5.5%) and 20:5Δ5,8,11,14,17 (16.4%). Relative-ly high levels of 14:0 (9.5%) and 15:0 (9.1%) FAwere foundas well. FA pattern was dominated by saturated FA (SFA)(38.9%), followed by monounsaturated FA (MUFA) (32.1%)and PUFA (28.6%). The glyceride pattern was dominated bymonoglycerides with a ratio mono-diglycerides of 2.8:1. Theglycerol-esterified FA were 14:0, 16:0, 16:1, 18:0 and 18:1.The main esterified FA (presented as % of total glycerides) inmonoglycerides are 16:0 (53.3%) and 16:1 (39.0%). Thediglycerides were esterified mainly with 14:0 and 16:1 or16:0 (24.3%) and 16:1 and/or 16:0 (75.7%) FA. Five sterolswere detected in C. scutellum. 24-methylenecholesterol(51.9% of total sterols) was the main compound followedby sterol A (23.0%), brassicasterol (20.0%), cholesterol(3.4%) and campesterol (1.6%).
The profile of the butanol fraction is dominated by FA(44.6%) followed by low molecular carbohydrates (24.5%),amino acids and other N-containing compounds (9.5%),alcohols (8.8%), glycerides (4.4%), organic acids (2.5%),
Table 1 (continued)
Compounds Rt Key ions (m/z)* Ether extract Butanol extract
113 Campesterol a, b, c 32.63 342, 382, 472** 0.07 0.04114 UC 33.93 0.08115 1-Tetradecanoyl-2-hexadecenoylglycerol (14:0; 16:1) c 39.91 129, 357, 595*** 0.23116 1-Tetradecanoyl-2-hexadecanoylglycerol (14:0; 16:0) c 40.16 145, 129, 597*** 0.28117 1-Tetradecanoyl-3-hexadecenoylglycerol (14:0; 16:1) c 40.80 343, 369, 595*** 0.09118 1-Tetradecanoyl-3-hexadecanoylglycerol (14:0; 16:0) c 41.07 343, 371, 597*** 0.09119 1,2-Dihexadecenoylglycerol (16:1; 16:1) c 45.21 129, 383, 621*** 0.22120 1-Hexadecanoyl-2-hexadecenoylglycerol (16:0; 16:1) c 45.62 129, 385, 623*** 1.00121 1,2-Dihexadecanoylglycerol (16:0; 16:0) a, b 45.96 129, 385, 625*** 0.43122 1,3-Dihexadecenoylglycerol (16:1; 16:1) c 46.55 369, 385, 621*** 0.07123 1-Hexadecanoyl-3-hexadecenoylglycerol (16:0; 16:1) c 46.94 371, 369, 623*** 0.39124 1,3-Dihexadecanoylglycerol (16:0; 16:0) a, b, c 47.27 371, 385, 625*** 0.06
* First key ion (m/z) is the base peak** Molecular ion [M] +
*** [M-15]+ ionUC Unidentified compound, UM unidentified monosaccharideIdentification:a NIST Mass spectral library 2005b The Golm Metabolome Databasec Recursive procedure for metabolite assignmentd Medeiros and Simoneit 2007e Nagashima and Fukuda 1981f Souchet and Laplante 2007 (as TMSi-derivative) and Gladu et al. 1991 (as non-derivatized compound)h The Lipid Library
Table 2 Metabolite groups of C. scutellum presented as % of TIC
Compounds Ether extract Butanol extract
Amino acids and N-containingmetabolites
9.51
Organic acids 2.51Phosphates 0.27 0.10Fatty acids 75.82 44.59Saturated 29.47 29.30Monoenes 24.32 13.76Dienes 0.31Polyenes 21.72 1.53Alcohols 0.28 8.81Alkanes 0.47Carbohydrates 24.54Monosaccharides 23.67Disaccharides 0.87Glycerides 10.8 4.35Monoglycerides 7.95 1.93Diglycerides 2.85 2.42Sterols 4.74Isoprenoids 3.55 1.17Total identified compounds(% of TIC)
95.93 95.58
300 J Appl Phycol (2009) 21:295–306
isoprenoid compounds (1.2%), and phosphates (0.1%). GC-MS analysis resulted in the identification of severalmonosaccharides (96.5% of total carbohydrates) and onedisaccharide (sucrose, 3.5% of total carbohydrates). Themost abundant low molecular carbohydrate was floridoside[2-O-glycerol-D-galactopyranoside (No. 76 in Table 1),45.7% of all carbohydrates]. Amino acids (10 identifiedcompounds) comprised 18.1% of all detected N-containingcompounds, while the nucleosides adenosine and uridinetogether with urea were 44.9%, 4.7%, and 24.6%, respective-ly. Alcohols were represented mainly by glycerol (8.6% of thepolar fraction). Several organic acids, including the phenolichydroxycinnamic acid, monoglycerides, two isoprene com-pounds (tocopherol and phytol), and glycerol-3-phosphate,were also detected in trace amounts in the fraction.
Aldehydes like 2,4-heptadienal, 2,4-octadienal and 2,4-decadienal were not detected in the studied diatoms.
Discussion
Preliminary studies indicated that the diethyl ether fractionof Cocconeis scutellum was the more active in provokingthe sex reversal of H. inermis and apoptosis in humancancer cell lines (authors, unpublished data). The lipidchemistry of algae, and of diatoms in particular, reliesmainly on analysis of the FA as FAME by GC-EI/MS(Mansour et al. 2005). In our work, the FA of both the etherand the butanol fractions were analyzed as FAME inparallel to their TMSi derivatives in order to obtainadditional information using their specific MS fragmenta-
tion (α and ω ions) to determine the position of doublebonds in PUFA (Christie 2003; Mjos and Pettersen 2003).In the EI/MS of FA, [M]+ and [M-15]+ for the TMSiderivatives and [M]+ and [M-31]+ for the FAME wereeasily distinguishable, and this permitted, together with theother characteristic ions, their unambiguous identification.In other cases, such as long-chain PUFA, these ions werehardly detectable and GC-MS analysis in CI mode wasused to display their molecular weight (MW). The [M+1]+
of FA in CI/MS spectra was the most abundant ion (basepeak), while in EI/MS spectra the [M]+ was decreasing withthe increase of the number of carbons and double bonds.An example is given in Fig. 2 in which 16:4Δ6,9,12,15 and16:2Δ9,12 co-eluted, and their separation, MS deconvolu-tion, and identification in EI/MS mode was problematic dueto the similarity of their fragmentation pattern, low amountof 16:2Δ9,12 and very low abundance of their [M]+ and [M-15]+ ions. However, GC-CI/MS analysis provided unam-biguous identification and separation by their base peak([M+1]+ at m/z 321 for 16:4Δ6,9,12,15 and at m/z 325 for16:2Δ9,12). Recently, such an approach was used for theanalysis of FA in samples of fish origin (Dayhuff and Wells2005). The FA composition of C. scutellum was similar tothat published for other diatoms (Rousch et al. 2003;Pistocchi et al. 2005). The presence of the odd-chain15:0 FA should be noted, which has been rarely reportedfor diatoms (Parrish et al. 1991; Gordon et al. 2006).
The identification of mono- and diglycerides was basedon comparison of their fragmentation pattern with refer-ence compounds and defining their [M]+ (when detectable)and [M-15]+ ion fragments in EI/MS mode. The 1-
B
a
41
56 58
78
91 108 112
123
6
a 41
58
76
93
AFig. 1 GC-MS chromatogramsof the ether (A) and butanolfractions (B) of C. scutellum.The numbers are in accordancewith Table 1. a – diethyl etherstabilizers
J Appl Phycol (2009) 21:295–306 301

monosubstituted glycerides showed an intensive [M-103]+
(often as base ion) and detectable [M-15]+ and [M]+. The 2-monosubstituted glycerides showed a base peak ion at m/z103 or 129 and characteristic losses [M-15]+, [M-90]+, [M-161]+ and [M-acyl]+. In contrast to EI/MS, the CI/MSspectra of 1- or 2-monosubstituted glycerides displayedsimilar fragmentation pattern with relatively intensive [M+1]+ ion, confirming their MW (Fig. 3). Diglyceridespossessed relatively intensive [M-15]+ ion and characteris-tic fragmentation depending on the position of substitutionas indicated in Fig. 4. The presence of triglycerides was not
confirmed by GC-MS as their boiling point would be abovethe highest temperature used in the elution program.
The structures of sterols were confirmed by comparingboth EI/MS of TMSi derivatives (together with RI) andnon-derivatized compounds with those of reference com-pounds or literature data. Sterols determine the diatomvalue as food, which can affect predators’ growth, asreported in oysters (Patterson et al. 1993). Unlike higherplants, algae and microalgae contain a large diversity ofsterols (Boutry et al. 1979; Veron et al. 1998) whoseprofiles can be sometimes characteristic of a particular
40 60 80 100 120 140 160 180 200 220 240 260 280 300 3200
50
100
4155
67
73
91
105 117129
147161 189 201 213 225 239 252 305 320
[M-15]+ [M]+
m/z 321
m/z 325
m/z 323
A
B
C
D
[M+1]+
[M+29]+
[M+41]+
Fig. 2 GC-CI-MS separation of 16:4Δ6,9,12,15 ([M+1]+ at m/z 321), 16:2Δ9,12 ([M+1]+ at m/z 325) and 16:3Δ6,9,12 ([M+1]+ at m/z 323). TICchromatogram (A), selected ion chromatogram (B), GC-CI-MS of 16:4Δ6,9,12,15 (C) and GC-EI-MS of 16:4Δ6,9,12,15 (D)
302 J Appl Phycol (2009) 21:295–306

class, family, genus or even species, and thus used forchemotaxonomic and phylogenetic purposes (Volkman1986; Patterson 1992). 24-methylenecholesterol (No. 112in Table 1) was the principal sterol like in other diatoms(Gladu et al. 1991; Pistocchi et al. 2005). It showed a [M]+
at m/z 398, a base peak at m/z 314 (characteristic of the24-methylene group), and an intensive ion at m/z 271 (lossof side chain) was identified in accordance with literaturedata (Gladu et al. 1991; Souchet and Laplante 2007). Thiscompound was isolated from plant pollens, like that fromPyrus malus (Hügel 1962), and from the soft coralSinularia sp. (Ahmed et al. 2006), and is a commonconstituent not only of algae, but also of clams, silkworms
and oysters. Sterol A (No. 103 in Table 1) showed equalMS spectra in both FAME fraction and TMSi-derivatizedsample indicating absence of a hydroxyl group, a molecularion atm/z 380 (confirmed by CI) and an intensive ion at m/z 255 which is characteristic for Δ5,22 sterols (Souchet andLaplante 2007). In addition, [M-18]+, characteristic forcompounds with hydroxyl group (at C-3), was notdetected. Its MS was similar to that of an ergost-3,5,22-trien structure.
Several isoprenoid compounds were detected in the etherfraction, including a γ-lactone-4,8,12,16-tetramethylhepta-decane-4-olide (No. 75 in Table 1), previously found inmarine sediments (Rontani and Volkman 2005), α- and β-
[M+1]+
[M-Acyl]+
[M-161]+
[M-90]+
[M-15]+
A-1
[M-103]+
[M-15]+
B-1
A-2
[M+1]+
B-2
Fig. 3 GC-EI-MS (A) and GC-CI-MS (B) of 1-monohexadecenoylglycerol (1) and 2-monohexadecenoylglycerol (2)
J Appl Phycol (2009) 21:295–306 303

tocopherols, found in many microalgae (Brown et al. 1999),phytol, and squalene (precursor of sterols). Relatively lowamounts (<0.5% of total compounds) of phosphates(phosphoric acid, glycerol-2-phosphate, and glycerol-3-phosphate), fatty alcohols (from C12 to C18) as well as n-alkanes (from C16 to C30) were detected in the studied etherfraction. Despite the characteristic fragmentation and RI,the n-alkanes showed detectable [M]+ in EI mode.
The metabolites in the butanol fraction (amino acids, N-containing compounds, organic acids, and saccharides)were identified by matching their mass spectra and RI withthe databases and the literature (Medeiros and Simoneit2007; Nagashima and Fukuda 1981) as indicated in Table 1.The FA composition is dominated by SFA and MUFA,while PUFA were considerably less abundant compared to
the apolar fraction (Table 2). Floridoside (No. 76 inTable 1), found to also be the dominant monosaccharidein other unicellular algae, showed under GC conditionsdistinguishable [M-15]+ at m/z 671, mass fragmentationpattern and RI identical to those reported in the literature(Nagashima and Fukuda 1981) and databases. Galactose,arabinose, glucose, talose and two unidentified carbohy-drates were also found in the butanol fraction, as in otheralgae (Abdel-Fattah and Edrees 1977; Nagashima andFukuda 1981). The high relative amount of N-containingcompounds in the polar fraction (9.5%) was noteworthy.
Aldehydes, responsible for teratogenic and abortiveeffects in planktonic copepods (d’Ippolito et al. 2002),were not detected in the studied diatoms. Therefore, thisclass of compounds is not involved in the sex reversal of H.
O
O
OC14H27 OC14H27
TMSi
O
O
O
O
211
239357
211
343
239
385
371
TMSi
[M-15]+
[M-15]+
A
B
A
OC16H31
OC16H31
B
Fig. 4 MS of 1-O-tetradecanoyl-2-O-hexadecanoylglycerol (A) and 1-O-tetradecanoyl-3-O-hexadecanoylglycerol (B)
304 J Appl Phycol (2009) 21:295–306

inermis, according to in vivo experiments carried out byZupo et al. (2007). However, the participation of otherkinds of oxylipins cannot yet be excluded and furtheranalyses of this topic should be performed. Bio-guidedassays on H. inermis postlarvae with the active etherfraction and its subfractions are currently in progress inorder to identify the compound(s) that are responsible,alone or synergistically, for the sex change in the modelshrimp Hippolyte inermis. Parallel tests with diatomextracts on human cancer cell lines will also allow theestablishing of the presence of a possible apoptotic activityon mammalian cells, in the perspective of eventualpharmacogical applications. In vitro and in vivo studieswith C. scutellum extracts have also been carried out oncommercially important crustaceans in order to evaluatetheir possible use in aquaculture.
Conclusions
This paper represents not only one of the few reports aboutnatural products from benthic diatoms, but the firstchemical report about Cocconeis scutellum. It also addsinformation about apoptotic compounds deriving frombenthic microalgae (Zupo et al. 2007). Moreover, our studydemonstrates the advantage of GC-MS analysis for rapidand simultaneous identification of metabolites from variousgroups. As the method is proven to be highly reproducible,it could be used for studies on metabolite dynamics anddistribution or influence of different abiotic factors on theirsynthesis. Further work on the chemistry of the benthicdiatom Cocconeis scutellum may reveal a chemoecologicalfunctionality and reveal potential apoptotic compound(s)for application in different fields.
Acknowledgements This study was carried out within the PHAR-MAPOX project, supported by the European Community (FP6–2003-NEST-A/STREP 4800). We are grateful to all the participants of thePHARMAPOX research team, Dr. A. Sagi (Ben Gurion University,Beer Sheva, Israel), Dr. S. Zupo (National Institute of Cancer, Genoa,Italy) and their groups, as well as Mr. S. Taboada (CEAB-CSIC,Blanes, Spain) The authors thank Generalitat de Catalunya(2005SGR-00020) for financial support and Dr. A. Marín of theServeis Cientificotècnics, University of Barcelona (Faculty of Phar-macy) for performing the GC-MS analyses.
References
Abdel-Fattah A, Edrees M (1977) Carbohydrates of the brownseaweed Padina pavonia. Phytochemistry 16:939–941doi:10.1016/S0031-9422(00)86699-6
Ahmed AF, Ya-Ting H, Zhi-Hong W, Yang-Chang W, Jyh-Horng S(2006) Polyoxygenated sterols from the formosan soft coral
Sinularia gibberosa. J Nat Prod 69:1275–1279 doi:10.1021/np0601509
Benkeblia N, Shinano T, Osaki M (2007) Metabolite profiling andassessment of metabolome compartmentation of soybean leavesusing non-aqueous fractionation and GC-MS analysis. Metab-olomics 3:297–305 doi:10.1007/s11306-007-0078-y
Boutry JL, Saliot A, Barbier M (1979) The diversity of marine sterolsand the role of algal biomasses; from facts to hypothesis.Experientia 35:1541–1543 doi:10.1007/BF01953179
Brown MR, Mular M, Miller I, Farmer C, Trenerry C (1999) Thevitamin content of microalgae used in aquaculture. J Appl Phycol11:247–255 doi:10.1023/A:1008075903578
Buia MC, Gambi MC, Zupo V (2000) Structure and functioning ofMediterranean Seagrass ecosystem: an overview. Biol MarMediterr 7:167–190
Christie WW (2003) Lipid analysis. The Oily Press, Bridgewater,England. http://www.lipidlibrary.co.uk/ms/ms01/index.htm
Dayhuff LE, Wells M (2005) Identification of fatty acids in fishescollected from the Ohio River using gas chromatography-massspectrometry in chemical ionization and electron impact modes. JChromatogr A 1098:144–149 doi:10.1016/j.chroma.2005.08.049
d’Ippolito G, Iadicicco O, Romano G, Fontana A (2002) Detection ofshort-chain aldehydes in marine organisms: the diatom Thalas-siosira rotula. Tet Lett 43:6137–6140
d’Udekem d’Acoz C (1996) The genus Hippolyte Leach, 1814(Crustacea: Decapoda: Caridea: Hippolytidae) in the EastAtlantic Ocean and the Mediterranean Sea, with a checklist ofall species in the genus. Zool Verh 303:1–133
Fontana A, d’Ippolito G, Cutignano A, Miralto A, Ianora A, Romano G,Cimino G (2007) Chemistry of oxylipin pathway in marinediatoms. Pure Appl Chem 79:481–490
Gambi MC, Lorenti M, Russo GF, Scipione MB, Zupo V (1992)Depth and seasonal distribution of some groups of vagile faunaof the Posidonia oceanica leaf stratum: structural and trophicanalyses. PSZNI Mar Ecol 13:17–39
Gavagnin M, Carbone M, Nappo M, Mollo E, Roussis V, Cimino G(2005) First chemical study of anaspidean Syphonota geo-graphica: structure of degraded sterols aplykurodinone-1 and -2. Tetrahedron 61:617–621
Gladu PK, Patterson GW, Wikfors GH, Chitwood DJ, Lusby WR(1991) Sterols of some diatoms. Phytochemistry 30:2301–2303
Gordon N, Neori A, Shpigel M, Lee J, Harpaz S (2006) Effect ofdiatom diets on growth and survival of the abalone Haliotisdiscus hannai postlarvae. Aquaculture 252:225–233
Hokputsa S, Hu C, Paulsen BS, Harding SE (2003) A physico-chemical comparative study on extracellular carbohydrate poly-mers from five desert algae. Carbohydr Polym 54:27–32
Hügel MF (1962) Étude de quelque constituants du pollen. AnnAbeille 5:97–133
Ianora A, Miralto A, Poulet SA, Carotenuto Y, Buttino I, Romano G,Casotti R, Pohnert G,Wichard T, Colucci-D’Amato L, Terrazzano G,Smetacek V (2004) Aldehyde suppression of copepod recruitment inblooms of a ubiquitous planktonic diatom. Nature 429:403–407
Lebeau T, Robert JM (2003) Diatom cultivation and biotechnologi-cally relevant products. Part I: Cultivation at various scales. ApplMicrobiol Biotechnol 60:612–623
Mansour MP, Frampton DMF, Nichols PD, Volkmann JK, Blackburn SI(2005) Lipid and fatty acid yield of nine stationary-phase micro-algae: Applications and unusual C24-C28 polyunsaturated fattyacids. J Appl Phycol 17:287–300
Medeiros PM, Simoneit BRT (2007) Analysis of sugars in environ-mental samples by gas chromatography-mass spectrometry. JChromatogr A 1141:271–278
Miralto A, Ianora A, Poulet SA (1995) Food type induces differentreproductive responses in the copepod Centropages typicus. JPlankton Res 17:1521–1534
J Appl Phycol (2009) 21:295–306 305

Miralto A, Barone G, Romano G, Poulet SA, Ianora A, Russo GL,Buttino I, Mazzarella G, Laabir M, Cabrini M, Giacobbe MG(1999) The insidious effect of diatoms on copepod reproduction.Nature 402:173–176
Mjos SA, Pettersen J (2003) Determination of trans double bonds inpolyunsaturated fatty acid methyl esters from their electronimpact mass spectra. Eur J Lipid Sci Technol 105:156–164
Nagashima H, Fukuda I (1981) Low molecular weight carbohydratesin Cyanidium caldarium and some related algae. Phytochemistry20:439–442
Nelson DM, Treguer P, Brzezinski MA, Leynaert A, Queguiner B (1995)Production and dissolution of biogenic silica in the ocean. Revisedglobal estimates, comparison with regional data and relationship tobiogenic sedimentation. Glob Biogeochem Cycles 9:359–372
Newell GE, Newell RC (1963) Marine plankton. Hutchinson, LondonParrish CC, de Freitas AS, Bodennec G, Macpherson E, Ackman R
(1991) Lipid composition of the marine diatom Nitzschiapungens. Phytochemistry 30:113–116
Patterson GW (1992) Sterols in algae. In: Patterson GW, Nes WD(eds) Physiology and biochemistry of sterols III. American OilChemists Society, Champaign, pp 118–157
Patterson GW, Tsitsa-Tzardis E, Wikfors GH, Gladu PK, Chitwood DJ,Harrison D (1993) Sterols of Tetraselmis (Prasinophyceae). CompBiochem Physiol B 105:253–256
Pistocchi R, Trigari G, Serrazanetti GP, Taddei P, Monti G,Palamidesi S, Guerrini F, Bottura G, Serratore P, Fabbri M,Pirini M, Ventrella V, Pagliarani A, Boni L, Borgatti AR (2005)Chemical and biochemical parameters of cultured diatoms andbacteria from the Adriatic Sea as possible biomarkers ofmucilage production. Sci Total Environ 353:287–299
Pohnert G (2005) Diatom/copepod interactions in plankton: the indirectchemical defence of unicellular algae. ChemBioChem 6:946–959
Pohnert G, Boland W (2002) The oxylipin chemistry of attraction anddefense in brown algae and diatoms. Nat Prod Rep 19:108–122
Raniello R, Iannicelli MM, NappoM, Avila C, Zupo V (2007) Productionof Cocconeis neothumensis (Bacillariophyceae) biomass in batchcultures and bioreactors for biotechnological applications: light andnutrient requirements. J Appl Phycol 19:383–391
Raymont JEG (1983) Plankton and Productivity in the Oceans. In:Phytoplankton 1. Food Industry, Moscow
Reverberi G (1950) La situazione sessuale di Hippolyte viridis e lecondizioni che la reggono. Boll Zool Ital 4–6:91–94
Roessner U, Wagner C, Kopka J, Trethewey RN, Willmitzer L (2000)Simultaneous analysis of metabolites in potato tuber by gaschromatography-mass spectrometry. Plant J 23:131–142
Rontani JF, Volkman JK (2005) Lipid characterization of coastalhypersaline cyanobacterial mats from the Camargue (France).Org Geochem 36:251–272
Rousch JM, Bingham SE, Sommerfeld MR (2003) Changes in fatty acidprofiles of thermo-intolerant and thermo-tolerant marine diatomsduring temperature stress. J Exp Mar Biol Ecol 295:145–156
Schliemann W, Ammer C, Strack D (2008) Metabolite profiling ofmycorrhizal roots of Medicago truncatula. Phytochemistry69:112–146
Sicko-Goad L, Andresen NA (1991) Effect of growth and light/darkcycles on diatom lipid content and composition. J Phycol27:710–718
Simental JA, Sánchez MP, Correa JG (2001) Biochemical compositionof benthic marine diatoms using as culture media a commonagricultural fertilizer. J Shellfish Res 20:611–617
Souchet N, Laplante S (2007) Seasonal and geographical variations ofsterol composition in snow crab hepatopancreas and pelagic fishviscera from Eastern Quebec. Comp Biochem Physiol B147:378–386
Taylor RL, Caldwell GS, Dunstan HJ, Bentley MG (2007) Short-termimpacts of polyunsaturated aldehyde-producing diatoms on theharpacticoid copepod, Tisbe holothuriae. J Exp Mar Biol Ecol341:60–69
Véron B, Dauguet JC, Billard C (1998) Sterolic biomarkers in marinephytoplankton. II. Free and conjugated sterols of seven speciesused in mariculture. J Phycol 34:273–279
Volkman JK (1986) A review of sterol markers for marine andterrigenous organic matter. Org Geochem 9:83–99
Zariquey R (1968) Crustáceos Decápodos Ibéricos. Invest Pesq 32:1–510Zupo V (1994) Strategies of sexual inversion in Hippolyte inermis
Leach (Crustacea, Decapoda) from a Mediterranean seagrassmeadow. J Exp Mar Biol Ecol 178:131–145
Zupo V (2000) Effect of microalgal food on the sex reversal ofHippolyte inermis (Crustacea: Decapoda). Mar Ecol Prog Ser201:251–259
Zupo V (2001) Influence of diet on sex differentiation of Hippolyteinermis Leach (Decapoda: Natantia) in the field. Hydrobiologia449:131–140
Zupo V, Messina P (2007) How do dietary diatoms cause the sexreversal of the shrimp Hippolyte inermis Leach (Crustacea,Decapoda). Mar Biol 151:907–917
Zupo V, Messina P, Buttino I, Sagi A, Avila C, Nappo M, Bastida J,Codina C, Zupo S (2007) Do benthic and planktonic diatomsproduce equivalent effects in crustaceans? Mar Freshw BehavPhysiol 40:169–181
306 J Appl Phycol (2009) 21:295–306




















