
Brominated Furanones Inhibit Biofilm Formation by Salmonella enterica Serovar Typhimurium
Upload
independentCategory
view
0download
0
Chem.-Biol. Interactions, 49 (1984) 351--368 351 Elsevier Scientific Publishers Ireland Ltd.
METABOLISM OF 1,8-DINITROPYRENE BY S A L M O N E L L A T Y P H I M U R I U M *
D.W. BRYANT a. D.R. McCALLA a'** , P. LULTSCHIK a, M.A. QUILLIAM b and B.E. McCARRY b
aDepartment of Biochemistry and bDepartment of Chemistry, McMaster University, 1200 Main St. West, Hamilton, Ontario L8N 3Z5 (Canada)
(Received September 15th, 1983) (Revision received December 13th, 1983) (Accepted January 3rd, 1984)
SUMMARY
Earlier work has shown that many nitroaromatic and nitroheterocyclic compounds are directly 'activated' to their ultimate mutagenic forms through the action of bacterial nitroreductase enzymes. However, in the case of 1,8-dinitropyrene (DNP) and certain other nitroarenes the pathway of activation is more complex and neither the identity of the ultimate mutagens nor the nature of the DNA adducts formed are known. We now show that Salmonella typhirnurium strains TA98 and TA1538, which are sensitive to DNP and have wild type nitroreductase complements, do metab- olize DNP to 1-amino-8-nitropyrene (ANP) and 1,8-diaminopyrene (DAP) but that these compounds axe much weaker mutagens than DNP. These two strains (TA98 and TA1538) contain two separable components of nitroreductase activity as determined using nitrofurazone as the substrate. The major component , at least, is capable of reducing both 1-nitropyrene (NP) and DNP although the rates are much lower than with nitrofurazone. TA98NR, a mutant of TA98 that is resistant to nitrofurazone and NP but not to DNP, lacked the major nitroreductase but retained two minor com- ponents. In contrast, a mutant (DNP6) which is resistant to DNP (but not to NP) contained a full complement of nitroreductases. When the metabolism of [3H]DNP by crude extracts of TA98 was re-examined, previously un- detected metabolites were found. These were more polar than DAP and
*Some of this material was presented in preliminary form at the CIIT Conference on Nitroarenes (Raleigh, NC, January 1982). **To whom correspondence should be sent. Abbreviations: ANP, 1-amino-8-nitropyrene; DAP, 1,8-diaminopyrene; DMSO, dimethyl- sulfoxide; DNP, 1,8-dinitropyrene; dpm, disintegrations per minute; HPLC, high pressure liquid chromatography; MOPS, morpholino propane sulfonic acid; NF, nitrofurazone; NP, 1-nitropyrene; PAH, polycyclic aromatic hydrocarbon.
0009-2797/84/$03.00 © 1984 Elsevier Scientific Publishers Ireland Ltd. Printed and Published in Ireland

352
ANP and were also seen when TA98NR was used as the source of enzyme. These metaboli tes were not formed when enzymes from TA98DNP6 or TA98NR/DNP6 were used. This work supports the notion that some en- zymic activity other than (or in addition to) nitroreductase is required for the activation of DNP and that the new polar metabolites may be related to this process.
K e y words: 1,8-Dinitropyrene -- S. t y p h i m u r i u m -- Nitroreductase metab- olites - Chromatography -- Mutagenicity
INTRODUCTION
Nitroarenes have recently been shown to be common mutagenic en- vironmental contaminants that are formed in a variety of combust ion processes through the nitration of polycyclic aromatic hydrocarbons (PAHs) [1--5]. Prominent amongst these nitro-PAHs are the nitropyrenes. NP has moderate mutagenic activity in the Salmonella assay, while the' dinitro derivatives are much more po ten t mutagens. NP is reduced by Salmonel la t y p h i m u r i u m to yield 1-aminopyrene, 1-acetylaminopyrene and several minor products [6]. When bacteria are incubated with tritiated NP, labelled adducts are formed in DNA. The same adducts are formed when calf thymus DNA is incubated with NP in the presence of xanthine oxidase (a mam- malian enzyme which can reduce a variety of nitro compounds under hy- poxic conditions) and hypothanthine [7]. The principal adduct has been characterized by Howard et al. [8,9] to be N-(deoxyguanosin)-8-yl-l-amino- pyrene.
Bacterial mutants that are partially deficient in nitroreductase activity are resistant to NP, indicating that reduction of the nitro group is involved in formation of the ultimate mutagen from this compound [10,11]. While the exact nature of the reactive species is not known with certainty it is likely that the hydroxylamine derivatives are involved [12]. With DNP the situation is different since nitroreductase-deficient strains are as sensitive as wild type. Further, a mutant (DNP6) selected by McCoy and Rosenkranz [13] on the basis of resistance to DNP was not cross-resistant to NP. Origin- ally, it was proposed that these observations could be explained by the existence of a family of nitroreductases having different specificities for various nitro compounds. This hypothesis has proven to be incorrect and more recently it has been suggested that the enzyme deficiency in DNP6 may be the lack of a transacetylase [14,15].
In this paper we examine the metabolism of DNP by S. t y p h i m u r i u m TA98 and resistant mutants and demonstrate the following: (1) while the reduction products ANP and DAP are formed, they are only weak muta- gens; (2) the nitroreductase enzymes of Salmonel la can be separated by DEAE cellulose chromatography into 2 components ; (3) the major nitro-

353
reductase component is absent from NP-resistant strains but the nitro- reductase profile of the DNP-resistant mutant is the same as wild type; (4) strains of Salmonella which are sensitive to DNP produce two new DNP metabolites that are not formed by DNP-resistant strains or by partially purified nitroreductase.
MATERIALS AND METHODS
Strains S. typhimurium strains TA98 and TA1538 were obtained from Dr. B.N.
Ames, University of California, Berkeley, CA. Strains TA98NR, TA98NR/1,8- DNP6, and DNP6 (also known as LF6) [13,14] were supplied by Dr. H.S. Rosenkranz, Case Western Reserve University, Cleveland, OH.
Chemicals NAD, NADPH and glucose-6-phosphate dehydrogenase from Leucog-
nostoc mesenteroides were supplied by Boehringer (Montreal). Glucose-6- phosphate, FMN and pronase E (bacterial protease Type XIV) were obtained from Sigma Chemical Co. (St. Louis, MO). NP (Aldrich) was purified by high performance liquid chromatography (HPLC), using a cyano bonded phase column (Altex 5 /zm Ultrasphere, 25 X 0.46 cm) with isopropanol/hexane (5:95) as the mobile phase. A mixture of dinitropyrene isomers was pre- pared by the nitration of pyrene with HNO3 in acetic anhydride (see Ref. 12 for details). DNP (Fig. 1) was isolated from the crude mixture by L.C. Services (Boston, MA). [3H]DNP (6 Ci/mrnol) was prepared by nitration of [3H]pyrene (obtained by the tritiation of 1-bromopyrene (Amersham)) followed by isolation of the 1,8-isomer by normal phase HPLC on the cyano column. Nitrofurazone (5-nitro-2-furaldehyde semicarbazone) was a gift of Norwich Pharmical, Norwich, NY.
9 ~ 2 1 0 'NO 8 9 1
FC)TO1 7 ~ N 02 O 2 N ~ 3 1,8-Dinitropyrene 6 ~ , , ~ , ~ j , 3
7 " - ~ v j " ~ 4 5 4 6 5
NH 2
O2N ~ 1"Amino, 8 -n i t ropyrene O2N ~'~sl O~//~ N" ~ H
NH 2 9 10 1
H2 N 1,8-Diamiopyrene 8 ~ ~ - : 7 H2 6 5 4
Fig. 1. Structures of some of the pyrene derivatives studied.
2-Nit rof luorene
Niridazole
2-Am inoanthracene

354
Synthesis of 1,8-ANP and 1,8-DAP Reduct ion of the 1,8-DNP with H2/PtO2/CH3OH gave the 1,8-diamino
derivative (1,8-DAP, Fig. 1) directly. The electron-impact mass spectrum of this compound gave ions at m/z 232 (100%, M), 204 (9%, M-H2CN) and 116 (20%, M/2). The UV spectrum was pH dependent. At pH 7.6 in phosphate buffer, absorbance maximal were seen at 376 nm (e = 1.2 × 104 M -1 cm -1 ) and 228 nm (e = 1.3 × 104 M -l cm-1).
To obtain ANP (Fig. 1), 1-N-acetylaminopyrene (prepared by reduction of NP and acylation of the resulting AP) was nitrated with fuming nitric acid in acetic anhydride to give two isomeric N-acetylaminonitropyrenes that were separated by reverse phase HPLC. On hydrolysis, each of these products afforded a single red aminonitropyrene. To establish which of these compounds was the desired 1,8-isomer, authentic samples of 1,3- 1,6- and 1,8-DNP were individually reduced to the aminonitropyrenes using xanthine oxidase. The retention times of the 1,8- and 1,6-isomers corres- ponded to those of the main products obtained via nitration of N-acetyl- aminopyrene. The electron-impact mass spectrum of the ANP give ions at m/z 262 (76%, M), 232 (100% M-NO), 231 (29%, M-HNO) and 216 (62%, M-NO2). The UV spectrum (acetonitrile) showed maxima at 490 nm (e = 1.1 × 104 M -~ cm-l) , 389 nm (e = 4.7 × 1 0 3 M -1 c m - ' ) , 3 7 1 n m ( e = 4 . 5 × 10 -3 M -1 cm-l) , 303 nm (e = 1.1 × 104 M - ' cm -1) and 246 nm (e = 2.3 × 10 4 M-I cm-1).
Preparation of crude extracts of S. typhirnurium Six liters of cells were grown overnight in Oxoid nutrient broth at 37°C
to an absorbance at 600 nm of 1.1--1.3. Cells were harvested by centri- fugation and washed in 0.067 M potassium phosphate buffer containing 6% (w/v) glycerol. Cells were resuspended in 30 ml of this buffer and soni- cated (Fisher Dismembrator 300) in ice. The resultant sonicate was centri- fuged (30 000 × g) to remove cell debris and the supernatant brought to 20% saturation with (NH4)2SO4. After further centrifugation (30 000 × g) the supernatant fraction was brought to 65% saturation with (NH4)2SO4 and the centrifugation repeated. The pellet was dissolved in 10 ml potassium phos- phate buffer (0.015 M, pH 7.2) containing 6% glycerol and 10 -6 M FMN. After 14 h dialysis against several changes of this buffer, the volume of the extract was determined and aliquots frozen at - 6 0 ° C . Nitroreductase activity of these extracts appeared to be stable for several months.
DEAE chromatography of crude extracts Crude extracts were chromatographed on a 1.5 × 60 cm column of
DEAE cellulose (Sigma) pre-equilibrated with 0.015 M potassium phosphate buffer (pH 7.2) containing 6% glycerol (w/v) and 0.02% sodium azide (column volume: 106 cm3; Vo: 48.5 ml). After loading a volume of crude extract, the column was washed with 5.5 void volumes of starting buffer

355
followed by a linear gradient from 0 to 0.3 M KC1 in buffer. The flow rate was 0.87 ml/min and 4.4 ml fractions were collected. Each collection tube contained sufficient FMN to give a final concentration of 10 -6 M in 4.5 ml. A final wash of 1.0 M KC1 was applied before regenerating the column.
Assay for nitroreductase activity Aliquots from crude extracts (or DEAE cellulose column fractions) were
assayed for ability to reduce nitrofurazone (NF) using either NADH or NADPH as a cofactor. This assay has been described previously [16,17] and was generally carried out within 24 h of chromatographic separation. Nitro- reductase activity was defined by the rate of reduction of NF as determined by the loss of absorbance at 375 nm over a 5-min period compared with a blank containing nitrofurazone. Nitroreductase activity was expressed as nmoles NF reduced per min per ml or per fraction using the extinction coefficient of nitrofurazone of e37s = 1.585 X 104 M -1 cm -1.
NP and DNP metabolism Metabolism of 1-NP or 1,8-DNP was studied under argon atmosphere.
One milliliter of crude extract (or fraction from DEAE cellulose) was added to a tube containing 12 mg G-6-P; 250 pg NADP or NAD; 1.25 X 10 -4 mol (50 mM) morpholino propane sulfonic acid (MOPS) buffer (pH 7.0); 10 -6 M FMN (to permit maximum nitroreductase activity and to help stabilize the enzyme) and 0.5 units glucoseS-phosphate dehydrogenase in a final volume of 2.5 ml. The mixture was equilibrated in an argon atmosphere (15--20 min) then 1,8-[3H]DNP (2.84 mCi/mmol), or unlabelled 1,8-DNP or 1-NP was added as appropriate. After 4 h or longer at 22°C, protease (6.8 units) was added to each assay to facilitate extraction of metabolites with organic solvents. Incubation continued for a further 3 h. Metabolites were recovered from the assay mixture by repeated extractions with ethyl acetate (3X) and dichloromethane (3X). These extracts were combined, concentrated on a rotary evaporator and the residue taken up in 200 pl dimethylsulfoxide (DMSO).
Fluorescence assay Fluorescent metabolites from reaction mixtures containing either 1-NP
or 1,8-DNP and fractions eluted from DEAE cellulose chromatography or crude cell extracts were assayed essentially as described by Lu et al. [18]. This procedure detects AP (excitation 356 nm, emission 440 n m ) o r DAP (excitation 396 nm, emission 451 nm) in organic extracts of enzyme assay mixtures evaporated to dryness and dissolved in methanol. Fluorescence was measured using a Perkin Elmer MFP44 spectrofluorimeter calibrated using known concentrations of AP and DAP as standards.

356
Reduction o f DNP by xanthine oxidase Reactions were carried out in sealed tubes under argon. DNP (100 #g in
DMSO) was added to 10 ml of 3.7 mM hypoxanthine in a 50 mM potassium phosphate buffer (pH 7.0) and a final concentration of 30% DMSO. This DMSO concentration had little, if any effect on the nitroreductase activity of xanthine oxidase. After equilibrating the mixture under argon (30 min), xanthine oxidase (25 units) was added and incubation carried out for several hours. The reaction was terminated by extraction of the mixture with ethyl acetate (3 × 3 vol.) and dichloromethane (3 X 3 vol.) Organic solvents were removed on a rotary evaporator at 25°C and the residue analysed by reverse phase HPLC. Fractions were collected and concentrated.
HPLC analysis Metabolites of 1,8-DNP were analysed using a reverse-phase HPLC system
(Beckman) connected to a variable wavelength detector (Hitachi model 110) and a fluorescence detector (Varian Fluorichrom equipped with 355 nm excitation filter and 430 nm emission filter). The column (Altex 5 ~m Ultra- sphere ODS, 25 X 0.46 cm) was equilibrated with a 0.02 F potassium phosphate buffer (pH 7.6) at 1.0 ml/min. A linear gradient from 0 to 90% acetonitrile over 30 min was begun at the time of injection. The column was subsequently eluted for a further 15 min with 90% acetonitrile/10% buffer before termination of the program. Fractions recovered for sub- sequent mass spectral analysis were collected in glass tubes and evaporated to dryness with a Savant Speed-Vac concentrator. The residue was then transferred into melting point capillaries, which, after evaporation of solvent, were sealed and submitted for mass spectrometry.
Mass spectrometry Electron impact mass spectra were acquired on a VG Micromass 7070F
mass spectrometer with an electron energy of 70 eV and a source tempera- ture of 200°C. Samples were introduced via the solid probe inlet.
Scintillation counting The HPLC eluant from experiments using [3H]DNP was collected in
1.0 ml fractions and added to 10 ml of a xylene based scintillation fluid (xylene, Triton 114, POPOP and POP) for counting of aqueous samples. Radioactivity was measured in a Beckman LS8000 scintillation counter programmed with a set of quenched standards to calculate the disintegra- tions per minute (dpm) in each fraction.
RESULTS
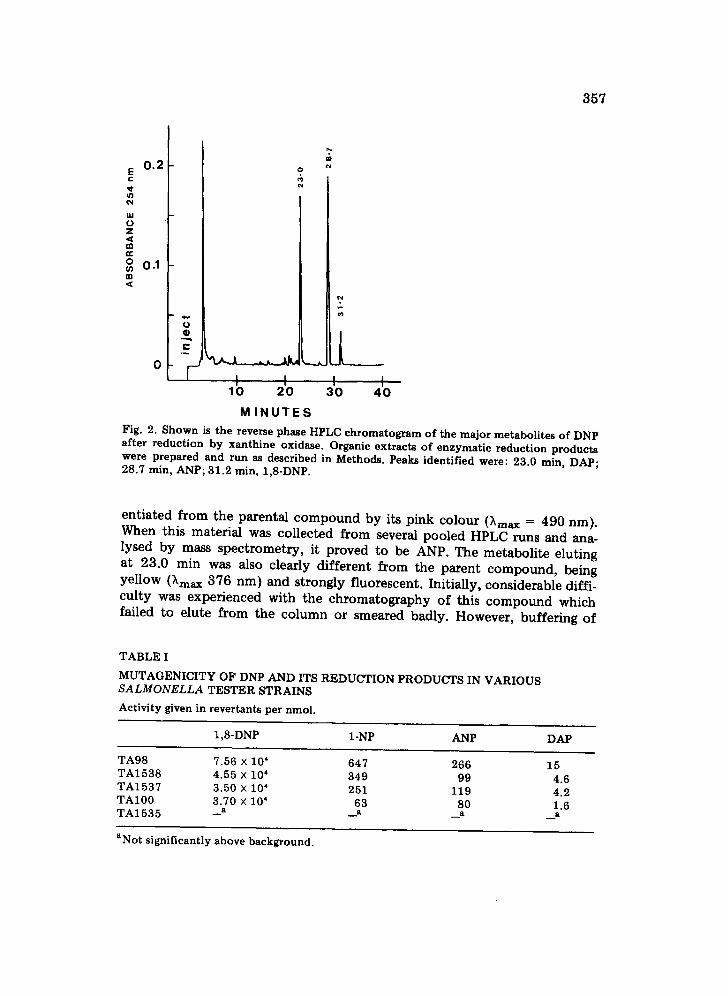
The enzymatic reduction of DNP by xanthine oxidase produced two metabolites that were observed by reverse phase HPLC analysis (Fig. 2, peaks at 23 and 28.7 min). These components are more polar in nature than the 1,8-DNP (31 min). The metabolite eluting at 28.7 min was differ-
357
0.2 E c
~N
UJ 0 Z .< m r r
0.1
0
"c-
r 10 20 30 4 0
M I N U T E S
Fig. 2. Shown is the reverse phase HPLC chromatogram of the major metabolites of DNP after reduction by xanthine oxidase. Organic extracts of enzymatic reduction products were prepared and run as described in Methods. Peaks identified were: 23.0 rain, DAP; 28.7 min, ANP; 31.2 min, 1,8-DNP.
entiated from the parental compound by its pink colour (kmax ---- 490 nm). When this material was collected from several pooled HPLC runs and ana- lysed by mass spectrometry, it proved to be ANP. The metabolite eluting at 23.0 min was also clearly different from the parent compound, being yellow (kma x 376 nm) and strongly fluorescent. Initially, considerable diffi- culty was experienced with the chromatography of this compound which failed to elute from the column or smeared badly. However, buffering of
TABLE I
MUTAGENICITY OF DNP AND ITS REDUCTION PRODUCTS IN VARIOUS SALMONELLA TESTER STRAINS
Activity given in revertants per nmol.
1,8-DNP 1-NP ANP DAP
TA98 7.56 X 10' 647 TA1538 4.55 X 10' 349 TA1537 3.50 x 10' 251 TA100 3.70 X 10' 63 TA1535 _a __.a
aNot significantly above background.
266 15 99 4.6
119 4.2 80 1.6
_a __a

358
the mobile phase to pH 7.6 with phosphate resulted in a sharp peak. This material was found to be DAP by retention time and mass spectrometry.
Neither of these metabolites (ANP or DAP) was particularly genotoxic compared to the parent compound (1,8-DNP) when tested without micro- somal activation in a variety of Salmonella strains (Table I). TA98 was most sensitive to the effects of DNP, followed by TA1538. Strain TA1535 (which preferentially detects base substitutions} was much less sensitive to DNP, though its plasmid-containing derivative TA100 showed nearly the same activity as TA1538. These results are essentially as reported by Mermelstein et al. [10] who at tr ibuted the extra sensitivity of TA100 compared to TA1535 to the enhanced inducibility of error-prone repair functions con- ferred on TA100 by the presence of the plasmid pMK101. The fact that ANP and DAP are more than 250-fold and 500-fold less active, respectively, as mutagens in the Salmonella than the parent compound indicates that their contr ibut ion to the overall mutagenic activity of DNP is minor. These results further indicate that the extremely genotoxic effect of DNP must be due to other metabolites.
Although the addition of microsomal activation nearly abolishes the mutagenicity of 1,8-DNP (results not shown, see also Mermelstein et al. [10] ), it is clear that this direct acting mutagen does require metabolism by endogenous bacterial enzymes before its genotoxici ty is manifest [11-- 13]. This is apparent from Table II which compares the responses of strain TA98 and three mutant strains. DNP6, a strain selected for resistance to DNP, shows substantially reduced sensitivity to DNP but remained as sensi- tive to niridazole and to NP as TA98. In contrast TA98NR, originally selected for niridazole resistance [19], was also resistant to NP but has wild type sensitivity to DNP and a number of other mutagens including 2-nitro- fluorene. 2-aminoanthracene and 2-acetylaminofluorene. The double mutant TA98NR/1,8-DNP6 showed the greatest resistance of all the strains tested (Table II) indicating an additive relationship may be responsible for confer- ring increased resistance to nitroarenes [11,13] . The data of Table II and earlier work by Rosenkranz et al. [11] indicate that the alteration which
T A B L E II
G E N O T O X I C A C T I V I T Y O F S E V E R A L N I T R O A N D AMINO C O M P O U N D S ON R E S I S T A N T S T R A I N S O F S. TYPHIMURIUM
Act iv i ty given in r eve r t an t s per nmol . nd , no t done . 2-AA, 2 - a m i n o a n t h r a c e n e ; AAF, 2 -ace ty l aminof luo rene .
1 ,8-DNP 1-NP 2-NF + $9 Nir idazole 2-AA A A F + $9
T A 9 8 2.05 × 105 647 833 255 644 12.4 T A 9 8 N R 2.03 × 105 116 1267 __a 792 11.3 DNP 6 3.12 × 103 396 3.8 356 82 1.1 T A 9 8 N R / 1 , 8 - D N P , 512 80 nd nd nd 2.2
a N o t s igni f icant ly above background .
TA
BL
E I
II
NIT
RO
RE
DU
CT
AS
E
AC
TIV
ITY
IN
SA
LM
ON
EL
LA
TE
ST
ER
S
TR
AIN
S
Nan
om
ole
s n
itro
fura
zon
e re
du
ced
in
1 r
ain
by
th
e to
tal
cru
de
extr
acts
fro
m
an e
qu
ival
ent
nu
mb
er
of c
ells
; or
by
th
e p
oo
led
fr
acti
on
s re
cov
ered
fro
m D
EA
E c
ellu
lose
ch
rom
ato
gra
ph
y f
oll
ow
ing
ap
pli
cati
on
of
the
cru
de
extr
act
acti
vit
y s
ho
wn
.
Co
fact
or
Cru
de
extr
act
Str
ain
DEAE cellulose fractions b
Component A
Component B
NA
DH
N
AD
PH
N
AD
H
NA
DP
H
To
tal
reco
ver
ed (
A &
B)
NA
DH
N
AD
PH
N
AD
H
NA
DP
H
TA
98
21
7
20
T
A9
8N
R
22
45
D
NP
, 38
7
50
T
A9
8N
R/1
,8-D
NP
6
0 19
18
22
13
00
9
40
1
30
0
96
0
33
38
78
0 11
1 38
17
23
8
40
1
40
0
86
0
15
00
24
23
73
0
97
23
aSee
Fig
. 2.
GO
O
1 ¢£
)

360
makes TA98NR resistant to niridazole and NP is quite separate from the function which confers sensitivity to DNP.
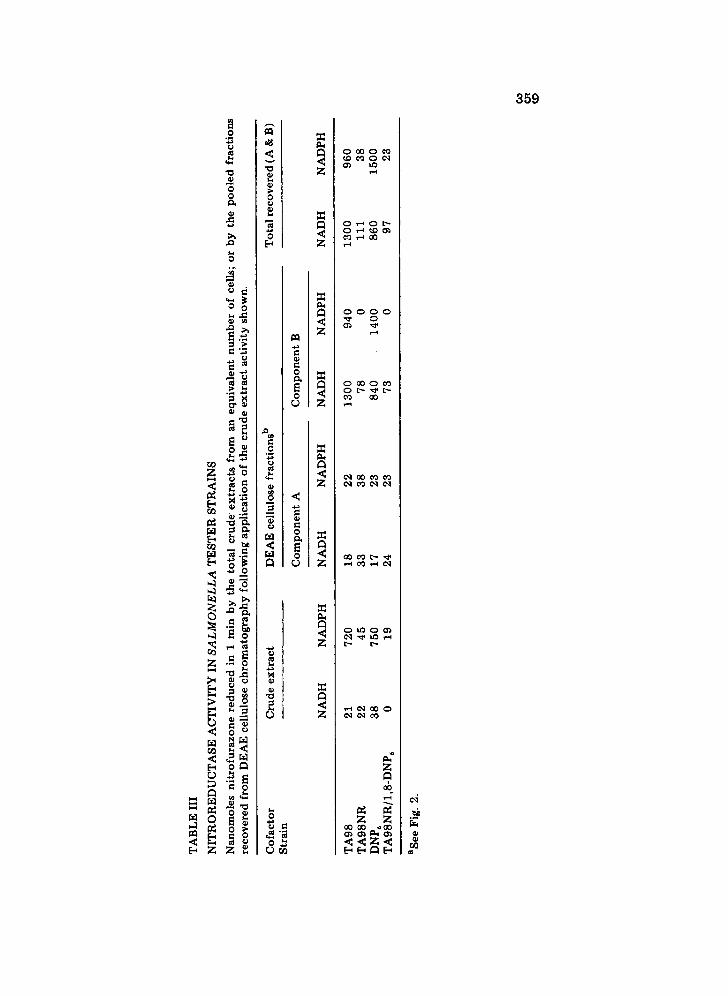
Crude extracts of TA98 and the resistant strains were prepared and assayed for 'classical' nitroreductase activity using nitrofurazone as sub- strate. Table III shows the oxygen-insensitive nitroreductase activity in various strains of Salmonella detected by ability of extracts to catalyse cofactor-dependent reduction of nitrofurazone [16]. Crude extracts of both TA98 and DNP6 showed substantial NADPH-dependent nitroreductase, while TA98NR and the double mutant TA98NR/DNP6 mutan t had sub- stantially reduced activity under similar conditions Activity within crude extracts was low for all strains with NADH as cofactor.
E 1.5 e-,
O oO O4
U,,I
z 1.0 cD
O u) en < 0.5
TA98 NR
L ~X
50 90 125 160
F R A C T I O N
4 0
30
20 .F.
W
10 ~ a w
w z O N
n,"
O n," I-,-
3.0 •
O
E 2 .0 c
1 . 0
Fig. 3. D E A E c h r o m a t o g r a p h y of c rude ex t rac t s o f T A 9 8 ( u p p e r panel ) or T A 9 8 N R ( lower pane l ) resu l ted in the sepa ra t ion of two n i t r o r educ t a se activit ies. The first (Peak A) s h o w n at a 10× e x p a n d e d scale e lu ted at a b o u t 0.07 M KCI, whi le the second (Peak B) was e lu ted by 0 .23 M KC1. F rac t ions were assayed for abi l i ty to r educe n i t r o f u r a z o n e as descr ibed. P r o t e i n is e s t ima ted b y a b s o r b a n c e at 280 m m (closed ovals). Ni t ro r e d u c t i o n was measu red wi th e i the r N A D P H (fil led circles) or NADH ( o p e n triangles). No te the d i f fe rence in scale b e t w e e n the u p p e r and lower panel.

361
Upon DEAE cellulose chromatography, the nitroreductase activity of TA98 separated into two components (Fig. 3). The first (peak A, fraction 90) eluted with 0.07 M KC1 and represents approx. 2% of the total nitroreductase activity recovered. This activity appeared to reduce the NF equally well using either NADPH or NADH as cofactor. The second (peak .B, fraction 125) represented 98% of the recovered activity using NADPH as a cofactor. An interesting feature of this partially purified reduc- tase was its ability to use NADH as a cofactor. This activity was not apparent in crude extracts (Table III). The reason for the lack of NADH dependent reductase activity in crude extracts is unknown. In general, we found that the total reductase activity (NADH and NADPH) recovered after chromato- graphy exceeded that applied to the column by a factor of about 3.
D N P ~
TA98 NR/1,8-DNP 6
(
l O x i
t °
40
3O
e , , . B
E
20 m o
u.I cc u.I
10 z o N < e,,-
o I -
Z 3.0 =
G)
E
2.0
i i i
I
i i
i J i t
: : 1.0 , i
h o , &
90 125
FRACTION
Fig. 4. DEAE chromatography of crude extracts of DNP+ (upper) and TA98NR/I,8- DNP+ (lower panel). Conditions of H a y and symbols are as for Fig. 2.
362
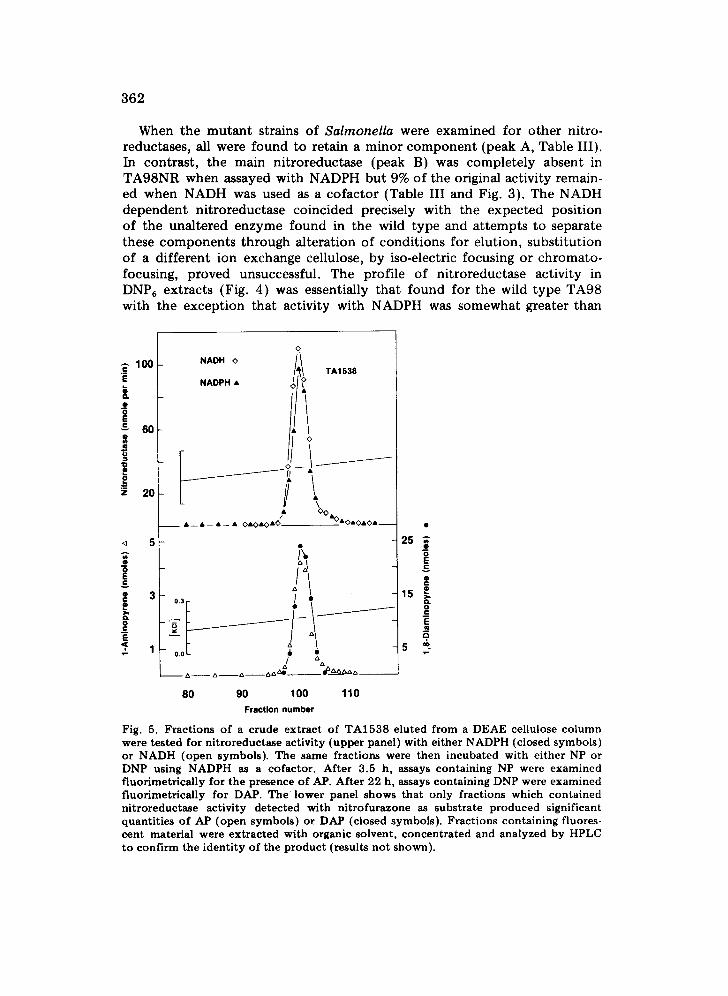
When the mutant strains of Salmonella were examined for other nitro- reductases, all were found to retain a minor component (peak A, Table III). In contrast, the main nitroreductase (peak B) was completely absent in TA98NR when assayed with NADPH but 9% of the original activity remain- ed when NADH was used as a cofactor (Table III and Fig. 3). The NADH dependent nitroreductase coincided precisely with the expected position of the unaltered enzyme found in the wild type and at tempts to separate these components through alteration of conditions for elution, substitution of a different ion exchange cellulose, by iso-electric focusing or chromato- focusing, proved unsuccessful. The profile of nitroreductase activity in DNP6 extracts (Fig. 4) was essentially that found for the wild type TA98 with the exception that activity with NADPH was somewhat greater than
A 1 0 0
E
0
60 S
i 2
z 20
~ 5
3
O _¢ E
o NADH <> /AI
I'~! TA1538 N A D P H • ~ / ~
• -- • _ • - - • (>AO40•O <>AOAOA-
?/ .3 I •
.o ,6
80 90 100 110 Fraction number
25 "~ !
E
s
Fig. 5. Fractions of a crude extract of TA1538 eluted from a DEAE cellulose column were tested for nitroreductase activity (upper panel) with either NADPH (closed symbols) or NADH (open symbols). The same fractions were then incubated with either NP or DNP using NADPH as a cofactor. After 3.5 h, assays containing NP were examined fluorimetrically for the presence of AP. After 22 h, assays containing DNP were examined fluorimetrically for DAP. The lower panel shows that only fractions which contained nitroreductase activity detected with nitrofurazone as substrate produced significant quantities of AP (open symbols) or DAP (closed symbols). Fractions containing fluores- cent material were extracted with organic solvent, concentrated and analyzed by HPLC to confirm the identity of the product (results not shown).

363
with NADH. This difference was observed in several separate analyses of the same enzyme preparation but extracts prepared at different times yielded somewhat variable activities with the two cofactors.
The lower panel included in Fig. 4 shows the nitroreductase profile of the double mutant TA98NR/1,8-DNP6. This is identical to that of TA98NR, showing peak A and the residual NADH-stimulated activity at the position of peak B (see Table III).
To examine the ability of the major classical nitroreductase to catalyse the reduction of nitropyrenes aliquots of column fractions were incubated separately with each of these compounds plus appropriate cofactors. Fluor- escence was measured after 3.5 h (NP) or overnight (DNP) incubation to determine the amounts of AP and DAP formed. The upper panel of Fig. 5 shows the major classical nitroreductase activity of TA1538 obtained with both cofactors while the lower panel indicates production of AP and DAP using the same fractions. These two activities coincide, though it is evident that the rate of formation of these two metabolites is slow relative to the re- duction of nitrofurazone. These results indicate that the classical nitrore- ductase acts upon both NP and DNP.
As was shown in Table I, neither ANP nor DAP are particularly muta- genic. It is of particular interest that the nitroreductase is present in un- diminished amounts in the dinitropyrene resistant strain DNP6. Thus, it would appear that if nitroreduction is involved in the production of the ultimate mutagen from DNP, it is not the rate limiting step even in the nitroreductase-deficient strain.
To determine whether other metabolites of DNP are formed in Salmonella, we examined the metabolism of [3H]-I-8-DNP by crude, unfractionated extracts of strain TA98 and its derivatives. Following a 4-h t reatment under anaerobic conditions, organic-soluble metabolites were extracted as described in the Methods and chromatographed on a C-18 reverse phase column. Eluted fractions were collected and counted. Recoveries of total radioactivity were determined from the total dpm in the organic and aque- ous fraction. For TA98, 90% of the original radioactivity in the assay was recovered. The distribution of radioactivity between organic and aqueous soluble fractions varied somewhat from one preparation to another but recoveries of label in the organic extractable fraction generally remained between 75% and 85%.
Figures 6--8 show the results of the reverse phase HPLC analysis of organic-extractable radioactive metabolites formed by the various Salmonella strains. Figure 6 compares the results where crude extracts of strains TA98NR/1,8-DNP6 and DNP6 were incubated with DNP. The double mutant metabolized little 1,8-DNP, and produced mainly ANP (eluted at 29.7 min). An additional unidentified compound eluting later (less polar) than the original dinitropyrene (33 min) was also observed. Little material of a more polar nature was produced by this extract. By contrast, DNP6, which has a 'normal ' nitroreductase complement bu t is resistant to DNP, metabolized most of the DNP forming some more polar products some of which were

TA
98
TA
98 P
eak
B
2o
s0
° l,l
o
10
10
i i
i
10
20
30
40
10
20
30
40
10
20
30
40
10
20
30
40
MIN
UTE
S
MIN
UTE
S
Fig
. 6.
Sh
ow
n a
re r
ad
ioch
rom
ato
gra
ms
of
me
tab
olit
es
of
[~H
]-I,
8-D
NP
ext
ract
ed
fro
m a
ssay
s usi
ng c
rud
e e
xtra
ct f
rom
eit
he
r th
e d
ou
ble
m
uta
nt
(lef
t)
or t
he
DN
P
resi
stan
t m
uta
nt
(rig
ht).
A
ssay
s w
ere
pre
par
ed
as d
escr
ibed
in
Met
ho
ds
and
met
abo
lite
s se
par
ated
o
n r
ever
se
ph
ase
HP
LC
us
ing
a p
ho
sph
ate
bu
ffer
/ace
ton
itri
le
pro
gra
m
also
des
crib
ed.
Po
siti
on
s o
n t
he
par
ent
com
po
un
d
(DN
P)
and
an
iden
tifi
ed
met
abo
lite
AN
P a
re s
ho
wn
. T
he
figu
re i
s fu
rth
er d
iscu
ssed
in
the
tex
t.
Fig
. 7.
R
ever
se p
has
e H
PL
C
rad
ioch
rom
ato
gra
ms
of
met
abo
lite
s o
f D
NP
p
rod
uce
d
by e
ith
er a
cru
de
extr
act
of
TA
98
(le
ft)
or a
par
tial
ly
pu
rifi
ed
nit
rore
du
ctas
e p
rep
arat
ion
fro
m a
cru
de
extr
act
of T
A9
8 f
ract
ion
ated
on
DE
AE
cel
lulo
se (
righ
t).
Po
siti
on
s of
kn
ow
n m
etab
oli
tes
are
sho
wn
in
dic
atin
g t
he
rad
ioac
tiv
e m
etab
oli
tes
pro
du
ced
by
TA
98
cru
de
extr
act
are
dif
fere
nt
fro
m a
min
on
itro
py
ren
e or
dia
min
op
yre
ne.
N
itro
red
uct
ase
acti
vity
co
nta
ined
in
Pea
k
B (
cf.
Fig
. 2)
was
in
suff
icie
nt
to
gen
erat
e d
etec
tab
le a
mo
un
ts
of
thes
e d
iffe
ren
t m
etab
oli
tes.
N
ote
th
e d
iffe
ren
ces
in s
cale
for
th
e ri
gh
t p
anel
. T
he
tota
l n
itro
red
uct
ase
pre
sen
t in
th
e p
uri
fied
fra
ctio
n w
as o
ne-
ten
th t
hat
of
the
cru
de
extr
act
used
. A
dju
stm
ent
of
acti
viti
es t
o g
ive
equ
al a
mo
un
ts o
f n
itro
red
uct
ase
did
no
t ap
pre
ciab
ly a
lter
th
e re
sult
(n
ot
sho
wn
).
CO
O
~

365
4 0
3 0
=Z = 20 - f :
TA98NR ( c r u d e ex t rac t )
i
1 0 -
10 2 0 3 0 4 0
M I N U T E S
Fig. 8. A reverse phase HPLC r a d i o c h r o m a t o g r a m of me tabo l i t e s of DNP p r o d u c e d by T A 9 8 N R c rude ex t rac t . Cond i t ions were the same as for Fig. 6. Relat ive pos i t ions of k n o w n m e t a b o l i t e s are shown. In add i t i on to a m i n o n i t r o p y r e n e , on ly two new meta- bol ic p r o d u c t s u n i q u e to DNP sensi t ive s t ra ins were observed.
poorly resolved and eluted between 22 and 27 min. The sharp peak at 30 min represents ANP. The right hand side of Fig. 7 shows the effects of a partially purified nitroreductase preparation (Peak B from TA98 in Fig. 1). The scale difference should be noted; <50% of the DNP was metabolized in this assay. As in the case of DNP6, ANP was produced along with other poorly resolved polar metabolites indicating that the metabolism which occurs in the extract of the DNP-resistant mutant is largely nitroreduction (as seen in Fig. 4).
The results for TA98 crude extract (Fig. 7 (left)) are in sharp contrast to those for the partially purified nitroreductase. The crude extract metabo- lized DNP rapidly and almost completely to two new metabolites. The first of these new metabolites eluted at 19.5 min and the second at 21.5 min. These compounds separated clearly from 1,8-DAP when extracts were co-chromatographed with synthetic DAP. Finally, a crude extract of TA98NR (Fig. 8) produced a pattern similar to that found with TA98 and quite different from that of the double mutant or DNP6. Though the relative amounts of the products appear to be different in the case of TA98NR (the extent of metabolism is dependent on the age of the extract), metabolites eluting at 19.5 and 21.5 min are formed only in strains which are sensitive to DNP.
In another series of experiments, [3H]DNP was incubated with intact Salmonella rather than extracts. The patterns of metabolites ob ta ined with the various strains were essentially identical to those obtained with crude extracts. {Data not shown).

366
DISCUSSION
These observations confirm that enzymes present in Salmonella reduce 1,8-DNP. Previous work has shown that the nitroreductases are responsible for the activation of nitroheterocyclic compounds and of NP to the ultimate mutagens [6,8,16,19,20]. In contrast, simple reduction products of DNP generated by nitroreductase are not responsible for the mutagenicity of this compound.
Tatsumi et al. [20] have reported oxygen-insensitive nitroreductase activities in S typhimurium TA100 and in nitrofurazone resistant mutants. These authors found two nitrofurazone reducing activities, one of which was exclusively dependent upon NADPH while the other utilized both NADH and NADPH cofactors. These two components eluted together from DEAE columns (cf, Fig. 3, Peak B) but Peak A was not detected. Depending upon the level of resistance to nitrofurazone, one or both of the nitroreductase activities was absent from the mutant strains. Strain TA98NR, like similar strains selected in our laboratory (unpublished data) represents a multistep mutant having high resistance.
The two new DNP metaboli tes reported here seem to be formed only in strains which are sensitive to DNP. These are likely related to the mutagenic action of DNP, perhaps being stable compounds derived from the ultimate mutagens themselves. We are currently in the process of trying to isolate and characterize these products.
Rosenkranz and coworkers [13,15] have studied the mutational response of several Salmonella strains to a variety of nitroarene mutagens and have suggested that the enzyme missing in DNP6 may be a transacetylase which converts relatively unreactive metabolites of DNP (and some other com- pounds) to highly reactive esters which are the ult imate mutagens. According to this hypothesis, reduction of nitro group(s) of DNP would be required to generate a substrate (perhaps the hydroxylamine) for the transacetylase. The difference between nitroarenes which, like NP, act in the presence of the DNP6 mutat ion and those like DNP which do not, may lie in the ability of the reduced metaboli tes to react directly with DNA. With com- pounds which react directly, nitroreductase activity would be rate-limiting and the NR mutant would be e~xpected to show considerable resistance. In contrast, the reduction products formed from compounds like DNP could be less reactive toward DNA and require further metabolism by the enzyme which is controlled by the DNP6 locus (inactivated in strain DNP6) before they are reactive enough to damage DNA. Since the NR mutants are not measurably resistant to DNP, the nitroreductase step cannot be rate limit- ing -- presumably because the few percent of wild type activity retained by these strains is sufficient to saturate the subsequent enzymic steps. However, until the relevant metabolites have been characterized and the products formed with DNA identified, there remains the possibility that other pro- cesses which do not involve reduction of the nitro groups are involved in the metabolic activation of 1,8-DNP.


368
15 E.C. McCoy, D.D. Marco, E.J. Rosenkranz, M. Anders, H.S. Rosenkranz and R. Mermelstein, 5-Nitroacenapthene: a newly recognized role for the nitro function in mutagenicity, Environ. Mutagen., 5 (1983) 17.
16 D.R. McCalla, P. Olive, Y. Tu and M.L. Fan, Nitrofurazone-reducing enzymes in E. coli and their role in drug activation in vivo, Can. J. Microbiol., 21 (1975) 1484.
17 D.W. Bryant, D.R. McCalla, M. Leeksma and P. Laneuville, Type I nitroreductases of Escherichia coli, Can. J. Microbiol., 27 (1981) 81.
18 C. Lu, D.R. McCalla and B.E. McCarry, The 1-nitropyrene reductase of Salmonella typhimurium, Chem.-Biol. Interact., 43 (1983) 67.
19 W.T. Speck, J.L. Blumer, E.J. Rosenkranz and H.S. Rosenkranz, Effect of genotype on mutagenicity of niridazole in nitroreductase deficient bacteria, Cancer Res., 41 (1981) 2305.
20 K. Tatsumi, T. Doi, H. Yoshimura, H. Koga and T. Horiuchi, Oxygen-insensitive nitrofuran reductases in Salmonella typhimurium TA100, J. Pharmacol. Dyn., 5 (1982) 423.
Copyright © 2022 FDOKUMEN
















![1,8-Di-aza-bicyclo-[5.4.0]undec-7-en-8-ium bromido-(phthalocyaninato)zincate](https://static.fdokumen.com/doc/165x107/633691fc02a8c1a4ec024048/18-di-aza-bicyclo-540undec-7-en-8-ium-bromido-phthalocyaninatozincate.jpg)



