
Chemicals mutagenic in Salmonella typhimurium strain TA1535 but not in TA100
Upload
independentCategory
view
0download
0
© 2002 Blackwell Science Ltd
Salmonella enterica serotype Typhimurium ShdA is anouter membrane fibronectin-binding protein that isexpressed in the intestine
sion is induced in vivo in the murine caecum, a tissuein which a cognate receptor of this outer membraneprotein is expressed.
Introduction
Salmonella enterica subspecies I serotypes are frequentlyisolated from food animals. In contrast, serotypes of S.enterica subspecies II–VII and Salmonella bongori arecommonly isolated from cold-blooded vertebrates. Thepersistence of S. enterica subspecies I serotypes in theintestine of livestock and domestic fowl is a critical factorfor food safety, as faecal contamination is a major sourceof contamination of derived food products (Galbraith,1961). The importance of S. enterica subspecies I sero-types for food safety is illustrated by the fact that this groupaccounts for >99% of human cases of salmonellosis(Aleksic et al., 1996).
One difference between S. enterica subspecies Iserotypes and reptile-associated serotypes of S. entericasubspecies II–VII and S. bongori is the presence of agenetic island located at 55 min on the S. enterica serovarTyphimurium (S. Typhimurium) chromosome. This gen-etic island encodes the shdA gene, which is present in S. enterica subspecies I serotypes but absent fromserotypes of S. enterica subspecies II–VII and S. bongori(Kingsley et al., 2000). Insertional inactivation or deletionof the shdA gene in S. Typhimurium reduces both the bacterial numbers recovered from faeces of mice and the time period for which bacteria are shed from animals(Kingsley et al., 2000; R. A. Kingsley, A. D. Humphries, A.M. Keestra and A. J. Bäumler, submitted). During thecourse of an S. Typhimurium infection in mice, the caecumrepresents the main reservoir for bacterial populationsshed with the faeces (R. A. Kingsley, A. D. Humphries, A.M. Keestra and A. J. Bäumler, submitted). Furthermore, anintact shdA gene is required for colonization of caecal walland caecal contents by S. Typhimurium (R. A. Kingsley, A.D. Humphries, A. M. Keestra and A. J. Bäumler, submit-ted). These data suggest that shdA-mediated colonizationof the caecum is required for persistence of S.Typhimurium in the mouse intestine.
The deduced amino acid sequence of shdA shareshomology with outer membrane-associated proteins ofthe autotransporter family of Gram-negative bacteria,
Molecular Microbiology (2002) 43(4), 895–905
Robert A. Kingsley,1 Renato L. Santos,2
A. Marijke Keestra,1 L. Garry Adams2 and Andreas J. Bäumler1*1Department of Medical Microbiology and Immunology,College of Medicine, Texas A and M University SystemHealth Science Center, Reynolds Medical Building,College Station, TX 77843-1114, USA.2Department of Veterinary Pathobiology, College ofVeterinary Medicine, Texas A and M University, College Station, TX 77843-4467, USA.
Summary
The shdA gene is the only determinant known to be required for persistence of Salmonella entericaserotype Typhimurium (S. Typhimurium) in themurine caecum and for efficient and prolonged shed-ding of the organism with the faeces. To study the biological activity of the ShdA protein, we examinedits expression and binding activity. ShdA was notdetected with anti-ShdA antiserum in S. Typhimuriumstrain ATCC14028 grown in vitro, suggesting that thisprotein is not expressed under standard conditionsof bacterial cultivation in the laboratory. However, inmice infected with S. Typhimurium, an immunofluo-rescence signal detected with anti-ShdA antiserumco-localized with that generated by anti-O4 antiserumin thin sections from the caecum. Expression of thecloned shdA gene from the T7 promoter in vitroresulted in detection of ShdA in the outer membraneof S. Typhimurium and in binding of fibronectin to thebacterial surface. Binding of purified glutathione-S-transferase (GST)–ShdA fusion protein to fibronectinwas dose dependent and could be partially inhibitedby preincubation with antifibronectin antibodies.GST–ShdA bound to connective tissue and the base-ment membrane in thin sections from the murinecaecum in situ. A similar labelling pattern was pro-duced when thin sections of the murine caecum werestained with antifibronectin antiserum. Collectively,these data demonstrate that ShdA is a surface-localized, fibronectin-binding protein whose expres-
Accepted 14 November, 2001. *For correspondence. E-mail [email protected]; Tel. (+1) 979 862 7756; Fax (+1) 979 845 3479.
including AIDA of diffuse adhering Escherichia coli andIcsA (VirG) of Shigella flexneri (Kingsley et al., 2000).However, sequence homology with IcsA and AIDAis limited to the carboxy-terminus of ShdA (residues1560–2036). Based on sequence homology withmembers of the autotransporter family, this conserved C-terminus is predicted to form a pore, through which a pas-senger domain involved in binding host macromoleculesis transported to the bacterial surface. The passengerdomain of AIDA binds epithelial cells through an uniden-tified receptor, whereas the N-terminal domain of IcsAbinds vinculin on the bacterial surface (Benz and Schmidt,1989; Bernardini et al., 1989). The fact that the N-terminal domains of IcsA and AIDA bind host macro-molecules raises the possibility that the ShdA N-terminusmay also have a cognate receptor in the murine caecum.However, sequence homology of the ShdA C-terminuswith IcsA and AIDA does not provide information regard-ing a possible host receptor of ShdA.
Collectively, these considerations raise the question asto whether ShdA is expressed at the surface of bacteriain vivo and binds a host macromolecule that is ex-pressed in the caecum of mice. To address these ques-tions, we investigated the expression and subcellularlocalization of the ShdA protein in S. Typhimurium. Fur-thermore, we identified a host protein that is bound byShdA and investigated the binding of ShdA to the murinecaecum in situ.
Results
Localization of S. Typhimurium in the murine caecum
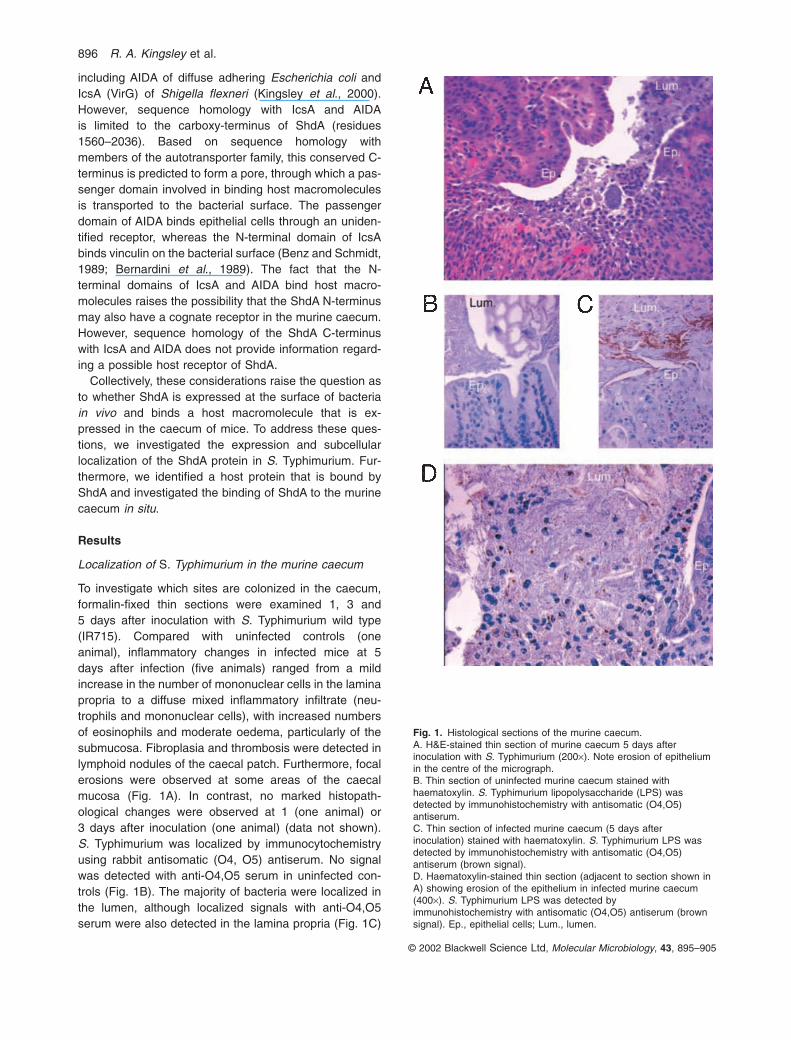
To investigate which sites are colonized in the caecum,formalin-fixed thin sections were examined 1, 3 and5 days after inoculation with S. Typhimurium wild type(IR715). Compared with uninfected controls (one animal), inflammatory changes in infected mice at 5days after infection (five animals) ranged from a mildincrease in the number of mononuclear cells in the laminapropria to a diffuse mixed inflammatory infiltrate (neu-trophils and mononuclear cells), with increased numbersof eosinophils and moderate oedema, particularly of thesubmucosa. Fibroplasia and thrombosis were detected inlymphoid nodules of the caecal patch. Furthermore, focalerosions were observed at some areas of the caecalmucosa (Fig. 1A). In contrast, no marked histopath-ological changes were observed at 1 (one animal) or3 days after inoculation (one animal) (data not shown). S. Typhimurium was localized by immunocytochemistryusing rabbit antisomatic (O4, O5) antiserum. No signalwas detected with anti-O4,O5 serum in uninfected con-trols (Fig. 1B). The majority of bacteria were localized inthe lumen, although localized signals with anti-O4,O5serum were also detected in the lamina propria (Fig. 1C)
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 895–905
896 R. A. Kingsley et al.
Fig. 1. Histological sections of the murine caecum.A. H&E-stained thin section of murine caecum 5 days afterinoculation with S. Typhimurium (200¥). Note erosion of epitheliumin the centre of the micrograph.B. Thin section of uninfected murine caecum stained withhaematoxylin. S. Typhimurium lipopolysaccharide (LPS) wasdetected by immunohistochemistry with antisomatic (O4,O5)antiserum.C. Thin section of infected murine caecum (5 days afterinoculation) stained with haematoxylin. S. Typhimurium LPS wasdetected by immunohistochemistry with antisomatic (O4,O5)antiserum (brown signal).D. Haematoxylin-stained thin section (adjacent to section shown inA) showing erosion of the epithelium in infected murine caecum(400¥). S. Typhimurium LPS was detected byimmunohistochemistry with antisomatic (O4,O5) antiserum (brownsignal). Ep., epithelial cells; Lum., lumen.
Expression of S. enterica ShdA 897
and in focally eroded areas of the epithelium in the caecalmucosa at day 5 after inoculation (Fig. 1D).
GST–ShdA fusion protein binds the extracellular matrixprotein fibronectin
Wound repair of damaged epithelium involves the recruit-ment of activated macrophages and subsequent release ofcytokines that mediate fibroplasia. Fibroblasts presentduring this response synthesize and secrete extracellularmatrix proteins, particularly fibronectin and collagen(Kurkinen et al., 1980; Grinnell et al., 1981; Raghow, 1994;Witte and Barbul, 1997). The presence of S. Typhimuriumin focal erosions of the epithelium (Fig. 1D) and the requirement of shdA for colonization of the caecum (R. A. Kingsley, A. D. Humphries, A. M. Keestra and A. J.Bäumler, submitted) prompted us to examine the possibil-ity that ShdA binds a component of the extracellular matrix.
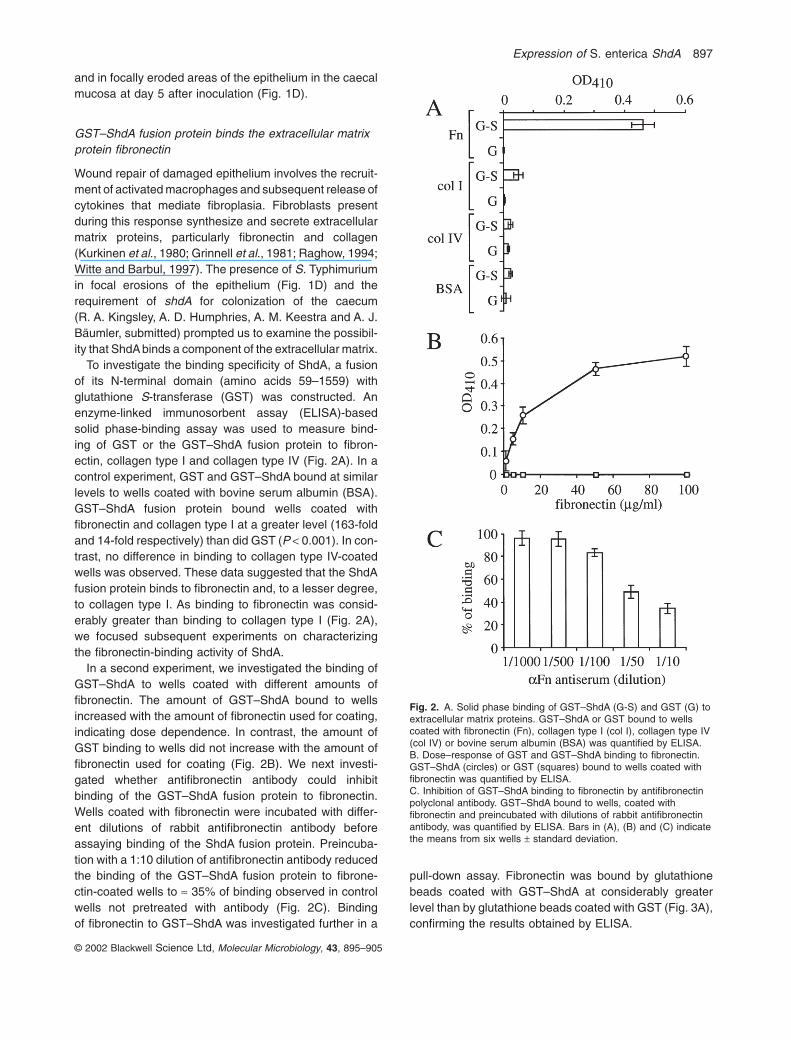
To investigate the binding specificity of ShdA, a fusionof its N-terminal domain (amino acids 59–1559) with glutathione S-transferase (GST) was constructed. Anenzyme-linked immunosorbent assay (ELISA)-basedsolid phase-binding assay was used to measure bind-ing of GST or the GST–ShdA fusion protein to fibron-ectin, collagen type I and collagen type IV (Fig. 2A). In acontrol experiment, GST and GST–ShdA bound at similarlevels to wells coated with bovine serum albumin (BSA).GST–ShdA fusion protein bound wells coated withfibronectin and collagen type I at a greater level (163-foldand 14-fold respectively) than did GST (P < 0.001). In con-trast, no difference in binding to collagen type IV-coatedwells was observed. These data suggested that the ShdAfusion protein binds to fibronectin and, to a lesser degree,to collagen type I. As binding to fibronectin was consid-erably greater than binding to collagen type I (Fig. 2A),we focused subsequent experiments on characterizingthe fibronectin-binding activity of ShdA.
In a second experiment, we investigated the binding ofGST–ShdA to wells coated with different amounts offibronectin. The amount of GST–ShdA bound to wellsincreased with the amount of fibronectin used for coating,indicating dose dependence. In contrast, the amount ofGST binding to wells did not increase with the amount offibronectin used for coating (Fig. 2B). We next investi-gated whether antifibronectin antibody could inhibitbinding of the GST–ShdA fusion protein to fibronectin.Wells coated with fibronectin were incubated with differ-ent dilutions of rabbit antifibronectin antibody beforeassaying binding of the ShdA fusion protein. Preincuba-tion with a 1:10 dilution of antifibronectin antibody reducedthe binding of the GST–ShdA fusion protein to fibrone-ctin-coated wells to ª 35% of binding observed in controlwells not pretreated with antibody (Fig. 2C). Binding of fibronectin to GST–ShdA was investigated further in a
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 895–905
Fig. 2. A. Solid phase binding of GST–ShdA (G-S) and GST (G) toextracellular matrix proteins. GST–ShdA or GST bound to wellscoated with fibronectin (Fn), collagen type I (col I), collagen type IV(col IV) or bovine serum albumin (BSA) was quantified by ELISA.B. Dose–response of GST and GST–ShdA binding to fibronectin.GST–ShdA (circles) or GST (squares) bound to wells coated withfibronectin was quantified by ELISA.C. Inhibition of GST–ShdA binding to fibronectin by antifibronectinpolyclonal antibody. GST–ShdA bound to wells, coated withfibronectin and preincubated with dilutions of rabbit antifibronectinantibody, was quantified by ELISA. Bars in (A), (B) and (C) indicatethe means from six wells ± standard deviation.
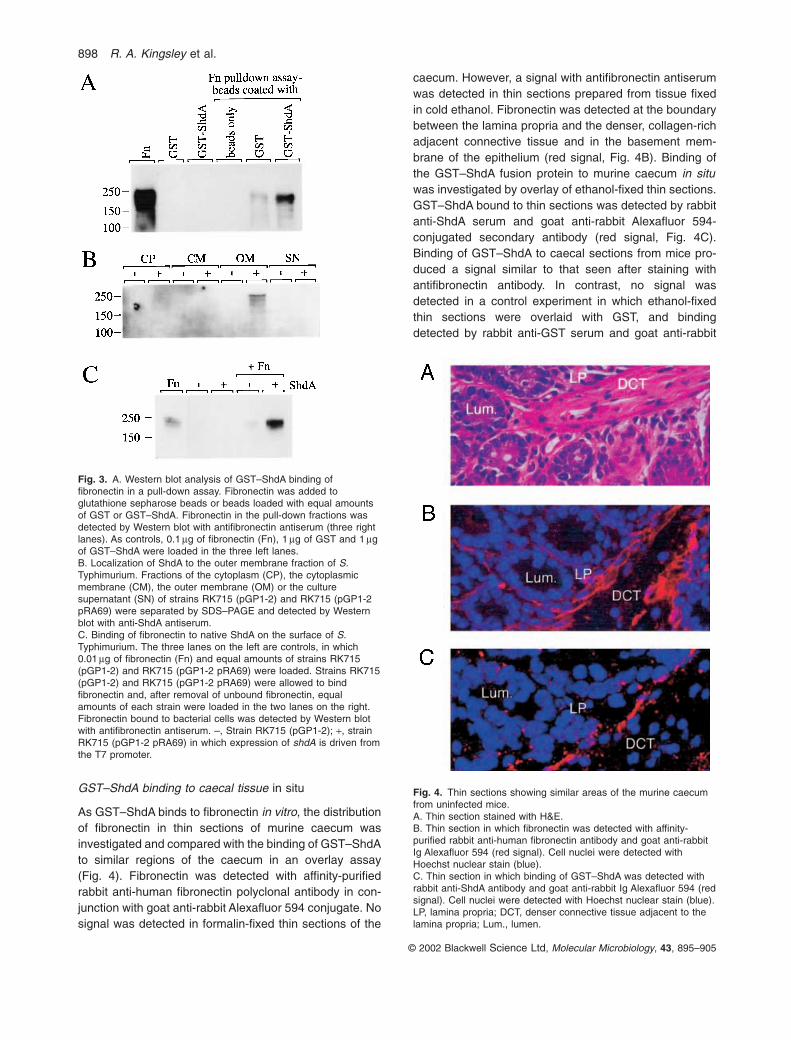
pull-down assay. Fibronectin was bound by glutathionebeads coated with GST–ShdA at considerably greaterlevel than by glutathione beads coated with GST (Fig. 3A),confirming the results obtained by ELISA.
GST–ShdA binding to caecal tissue in situ
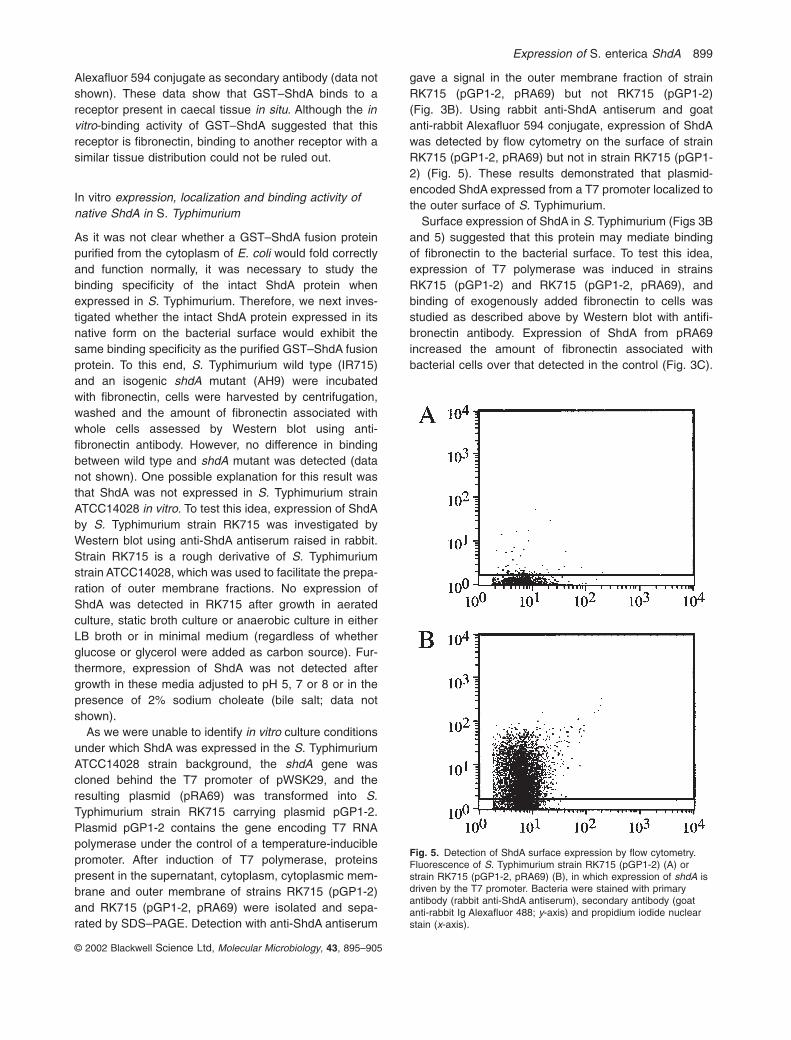
As GST–ShdA binds to fibronectin in vitro, the distributionof fibronectin in thin sections of murine caecum wasinvestigated and compared with the binding of GST–ShdAto similar regions of the caecum in an overlay assay(Fig. 4). Fibronectin was detected with affinity-purifiedrabbit anti-human fibronectin polyclonal antibody in con-junction with goat anti-rabbit Alexafluor 594 conjugate. Nosignal was detected in formalin-fixed thin sections of the
caecum. However, a signal with antifibronectin antiserumwas detected in thin sections prepared from tissue fixedin cold ethanol. Fibronectin was detected at the boundarybetween the lamina propria and the denser, collagen-richadjacent connective tissue and in the basement mem-brane of the epithelium (red signal, Fig. 4B). Binding ofthe GST–ShdA fusion protein to murine caecum in situwas investigated by overlay of ethanol-fixed thin sections.GST–ShdA bound to thin sections was detected by rabbitanti-ShdA serum and goat anti-rabbit Alexafluor 594-conjugated secondary antibody (red signal, Fig. 4C).Binding of GST–ShdA to caecal sections from mice pro-duced a signal similar to that seen after staining withantifibronectin antibody. In contrast, no signal wasdetected in a control experiment in which ethanol-fixedthin sections were overlaid with GST, and bindingdetected by rabbit anti-GST serum and goat anti-rabbit
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 895–905
898 R. A. Kingsley et al.
Fig. 3. A. Western blot analysis of GST–ShdA binding offibronectin in a pull-down assay. Fibronectin was added toglutathione sepharose beads or beads loaded with equal amountsof GST or GST–ShdA. Fibronectin in the pull-down fractions wasdetected by Western blot with antifibronectin antiserum (three rightlanes). As controls, 0.1 mg of fibronectin (Fn), 1 mg of GST and 1 mgof GST–ShdA were loaded in the three left lanes.B. Localization of ShdA to the outer membrane fraction of S.Typhimurium. Fractions of the cytoplasm (CP), the cytoplasmicmembrane (CM), the outer membrane (OM) or the culturesupernatant (SN) of strains RK715 (pGP1-2) and RK715 (pGP1-2pRA69) were separated by SDS–PAGE and detected by Westernblot with anti-ShdA antiserum.C. Binding of fibronectin to native ShdA on the surface of S.Typhimurium. The three lanes on the left are controls, in which 0.01 mg of fibronectin (Fn) and equal amounts of strains RK715(pGP1-2) and RK715 (pGP1-2 pRA69) were loaded. Strains RK715(pGP1-2) and RK715 (pGP1-2 pRA69) were allowed to bindfibronectin and, after removal of unbound fibronectin, equalamounts of each strain were loaded in the two lanes on the right.Fibronectin bound to bacterial cells was detected by Western blotwith antifibronectin antiserum. –, Strain RK715 (pGP1-2); +, strainRK715 (pGP1-2 pRA69) in which expression of shdA is driven fromthe T7 promoter.
Fig. 4. Thin sections showing similar areas of the murine caecumfrom uninfected mice.A. Thin section stained with H&E.B. Thin section in which fibronectin was detected with affinity-purified rabbit anti-human fibronectin antibody and goat anti-rabbitIg Alexafluor 594 (red signal). Cell nuclei were detected withHoechst nuclear stain (blue).C. Thin section in which binding of GST–ShdA was detected withrabbit anti-ShdA antibody and goat anti-rabbit Ig Alexafluor 594 (redsignal). Cell nuclei were detected with Hoechst nuclear stain (blue).LP, lamina propria; DCT, denser connective tissue adjacent to thelamina propria; Lum., lumen.
Expression of S. enterica ShdA 899
Alexafluor 594 conjugate as secondary antibody (data notshown). These data show that GST–ShdA binds to areceptor present in caecal tissue in situ. Although the invitro-binding activity of GST–ShdA suggested that thisreceptor is fibronectin, binding to another receptor with asimilar tissue distribution could not be ruled out.
In vitro expression, localization and binding activity ofnative ShdA in S. Typhimurium
As it was not clear whether a GST–ShdA fusion proteinpurified from the cytoplasm of E. coli would fold correctlyand function normally, it was necessary to study thebinding specificity of the intact ShdA protein whenexpressed in S. Typhimurium. Therefore, we next inves-tigated whether the intact ShdA protein expressed in itsnative form on the bacterial surface would exhibit thesame binding specificity as the purified GST–ShdA fusionprotein. To this end, S. Typhimurium wild type (IR715) and an isogenic shdA mutant (AH9) were incubated with fibronectin, cells were harvested by centrifugation,washed and the amount of fibronectin associated withwhole cells assessed by Western blot using anti-fibronectin antibody. However, no difference in bindingbetween wild type and shdA mutant was detected (datanot shown). One possible explanation for this result wasthat ShdA was not expressed in S. Typhimurium strainATCC14028 in vitro. To test this idea, expression of ShdAby S. Typhimurium strain RK715 was investigated byWestern blot using anti-ShdA antiserum raised in rabbit.Strain RK715 is a rough derivative of S. Typhimuriumstrain ATCC14028, which was used to facilitate the prepa-ration of outer membrane fractions. No expression ofShdA was detected in RK715 after growth in aeratedculture, static broth culture or anaerobic culture in eitherLB broth or in minimal medium (regardless of whetherglucose or glycerol were added as carbon source). Fur-thermore, expression of ShdA was not detected aftergrowth in these media adjusted to pH 5, 7 or 8 or in thepresence of 2% sodium choleate (bile salt; data notshown).
As we were unable to identify in vitro culture conditionsunder which ShdA was expressed in the S. TyphimuriumATCC14028 strain background, the shdA gene wascloned behind the T7 promoter of pWSK29, and theresulting plasmid (pRA69) was transformed into S.Typhimurium strain RK715 carrying plasmid pGP1-2.Plasmid pGP1-2 contains the gene encoding T7 RNApolymerase under the control of a temperature-induciblepromoter. After induction of T7 polymerase, proteinspresent in the supernatant, cytoplasm, cytoplasmic mem-brane and outer membrane of strains RK715 (pGP1-2)and RK715 (pGP1-2, pRA69) were isolated and sepa-rated by SDS–PAGE. Detection with anti-ShdA antiserum
gave a signal in the outer membrane fraction of strainRK715 (pGP1-2, pRA69) but not RK715 (pGP1-2)(Fig. 3B). Using rabbit anti-ShdA antiserum and goat anti-rabbit Alexafluor 594 conjugate, expression of ShdAwas detected by flow cytometry on the surface of strainRK715 (pGP1-2, pRA69) but not in strain RK715 (pGP1-2) (Fig. 5). These results demonstrated that plasmid-encoded ShdA expressed from a T7 promoter localized tothe outer surface of S. Typhimurium.
Surface expression of ShdA in S. Typhimurium (Figs 3Band 5) suggested that this protein may mediate bindingof fibronectin to the bacterial surface. To test this idea,expression of T7 polymerase was induced in strainsRK715 (pGP1-2) and RK715 (pGP1-2, pRA69), andbinding of exogenously added fibronectin to cells wasstudied as described above by Western blot with antifi-bronectin antibody. Expression of ShdA from pRA69increased the amount of fibronectin associated with bacterial cells over that detected in the control (Fig. 3C).
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 895–905
Fig. 5. Detection of ShdA surface expression by flow cytometry.Fluorescence of S. Typhimurium strain RK715 (pGP1-2) (A) orstrain RK715 (pGP1-2, pRA69) (B), in which expression of shdA isdriven by the T7 promoter. Bacteria were stained with primaryantibody (rabbit anti-ShdA antiserum), secondary antibody (goatanti-rabbit Ig Alexafluor 488; y-axis) and propidium iodide nuclearstain (x-axis).
These data demonstrated that ShdA expressed in S.Typhimurium localizes to the outer membrane where itmediates binding to fibronectin.
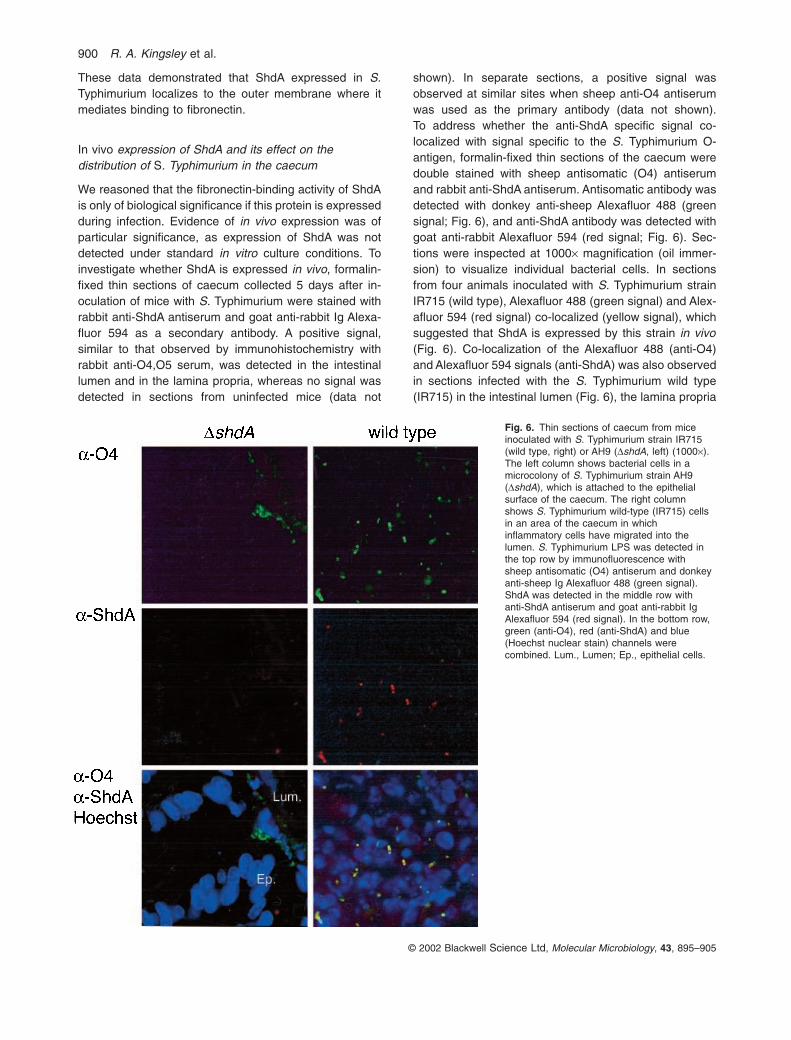
In vivo expression of ShdA and its effect on thedistribution of S. Typhimurium in the caecum
We reasoned that the fibronectin-binding activity of ShdAis only of biological significance if this protein is expressedduring infection. Evidence of in vivo expression was ofparticular significance, as expression of ShdA was notdetected under standard in vitro culture conditions. Toinvestigate whether ShdA is expressed in vivo, formalin-fixed thin sections of caecum collected 5 days after in-oculation of mice with S. Typhimurium were stained withrabbit anti-ShdA antiserum and goat anti-rabbit Ig Alexa-fluor 594 as a secondary antibody. A positive signal,similar to that observed by immunohistochemistry withrabbit anti-O4,O5 serum, was detected in the intestinallumen and in the lamina propria, whereas no signal wasdetected in sections from uninfected mice (data not
shown). In separate sections, a positive signal wasobserved at similar sites when sheep anti-O4 antiserumwas used as the primary antibody (data not shown). To address whether the anti-ShdA specific signal co-localized with signal specific to the S. Typhimurium O-antigen, formalin-fixed thin sections of the caecum weredouble stained with sheep antisomatic (O4) antiserumand rabbit anti-ShdA antiserum. Antisomatic antibody wasdetected with donkey anti-sheep Alexafluor 488 (greensignal; Fig. 6), and anti-ShdA antibody was detected withgoat anti-rabbit Alexafluor 594 (red signal; Fig. 6). Sec-tions were inspected at 1000¥ magnification (oil immer-sion) to visualize individual bacterial cells. In sectionsfrom four animals inoculated with S. Typhimurium strainIR715 (wild type), Alexafluor 488 (green signal) and Alex-afluor 594 (red signal) co-localized (yellow signal), whichsuggested that ShdA is expressed by this strain in vivo(Fig. 6). Co-localization of the Alexafluor 488 (anti-O4)and Alexafluor 594 signals (anti-ShdA) was also observedin sections infected with the S. Typhimurium wild type(IR715) in the intestinal lumen (Fig. 6), the lamina propria
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 895–905
900 R. A. Kingsley et al.
Fig. 6. Thin sections of caecum from miceinoculated with S. Typhimurium strain IR715(wild type, right) or AH9 (DshdA, left) (1000¥).The left column shows bacterial cells in amicrocolony of S. Typhimurium strain AH9(DshdA), which is attached to the epithelialsurface of the caecum. The right columnshows S. Typhimurium wild-type (IR715) cellsin an area of the caecum in whichinflammatory cells have migrated into thelumen. S. Typhimurium LPS was detected inthe top row by immunofluorescence withsheep antisomatic (O4) antiserum and donkeyanti-sheep Ig Alexafluor 488 (green signal).ShdA was detected in the middle row withanti-ShdA antiserum and goat anti-rabbit IgAlexafluor 594 (red signal). In the bottom row,green (anti-O4), red (anti-ShdA) and blue(Hoechst nuclear stain) channels werecombined. Lum., Lumen; Ep., epithelial cells.

Expression of S. enterica ShdA 901
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 895–905
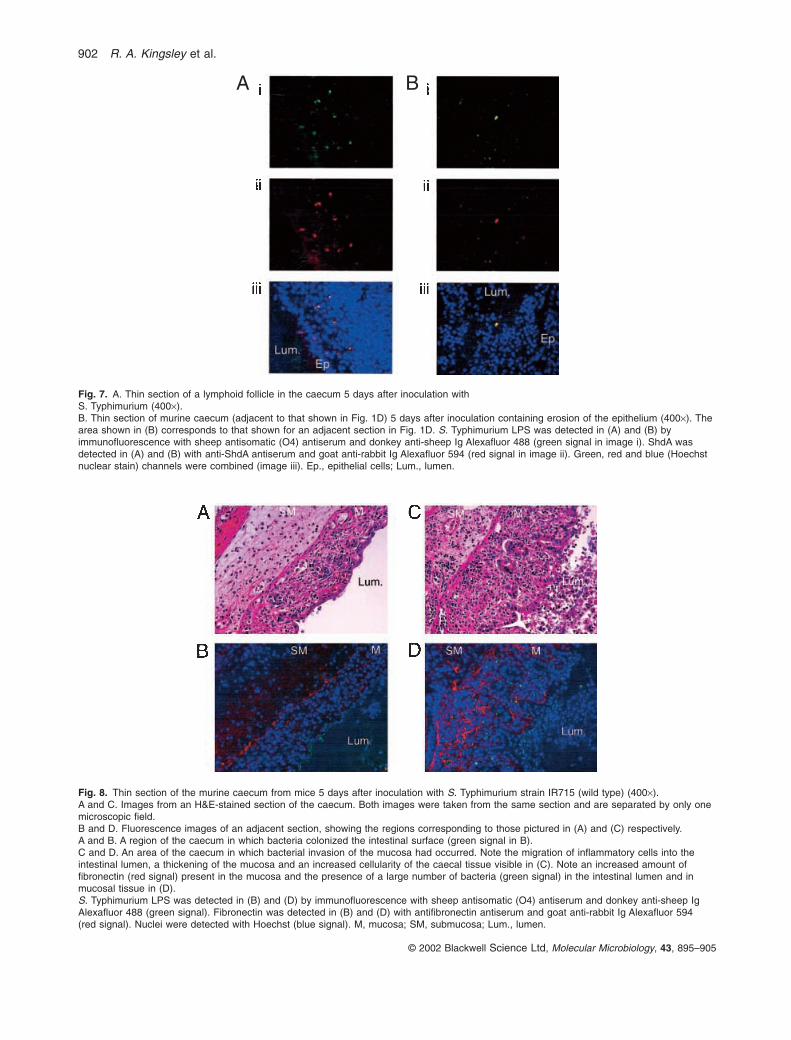
and at the site of focal erosion in sections from animalsinoculated with strain IR715 (Fig. 7A and B). In contrast,no co-localization of Alexafluor 594 signal (anti-ShdA)with Alexafluor 488 (anti-O4) signal was observed in sections from three animals inoculated with the S.Typhimurium shdA deletion mutant, AH9 (Fig. 6).
To investigate whether inactivation of the shdAgene would alter the distribution of S. Typhimurium within the caecum, thin sections from eight mice collected5 days after oral inoculation with strains IR715 (wild type)or AH9 (DshdA) were analysed by immunofluorescencemicroscopy after fixation in cold ethanol and detectionwith antifibronectin and anti-O4 antiserum. Strain IR715could be detected in the lumen (Fig. 8B) of the caecumand within the mucosa (Fig. 8D) in three out of five mice inoculated with this strain. The areas of bacterialinvasion of the caecal mucosa were localized (i.e. theregion shown in Fig. 8D is separated from that shown in Fig. 8B by only one microscopic field). In regions of the caecum in which S. Typhimurium was not detectableor in which it was present only in the lumen, fibronectinwas detectable at the junction of the lamina propria, theunderlying connective tissue of the submucosa and in the basement membrane (Fig. 8B). At sites in which S. Typhimurium was detected in the mucosa, the amountof fibronectin in the mucosa appeared to be elevated(Fig. 8D). The S. Typhimurium shdA mutant (AH9) wasnot detected in the mucosa of 36 thin sections from threeanimals from which the caecum had been collected at5 days after inoculation. In each section, S. Typhimuriumwas either detected exclusively in the lumen of thecaecum, where it was found in low numbers, or notdetected at all (data not shown).
Discussion
Sequence homology suggests that shdA encodes aprotein of the autotransporter family. AIDA and IcsA, theautotransporters that share the highest sequence homol-ogy to ShdA, are outer surface-localized proteins (Benzand Schmidt, 1989; Bernardini et al., 1989). Consistentwith the idea that shdA encodes a surface-localizedprotein, we found that expression of this gene from the T7promoter led to the production of an ª 200 kDa outer mem-brane protein in S. Typhimurium strain ATCC14028, whichwas recognized by anti-ShdA antiserum (Fig. 3B). Fur-thermore, fluorescence labelling of S. Typhimurium cellsexpressing shdA from the T7 promoter demonstrated thatShdA is exposed to the bacterial surface (Fig. 5). The outersurface-associated proteins AIDA and IcsA mediate bac-terial binding to HEp2 cells and vinculin respectively (Benzand Schmidt, 1989; Bernardini et al., 1989). As AIDA andIcsA function as non-fimbrial adhesins, we investigatedwhether ShdA mediates binding to a host protein. Expres-
sion of shdA from the T7 promoter in S. Typhimuriumresulted in binding of fibronectin to the bacterial surface(Fig. 3C). Furthermore, binding of GST–ShdA fusionprotein to fibronectin in a solid phase-binding assay wasdose dependent (Fig. 2B) and could be partially inhibitedby antifibronectin antibody (Fig. 2C).
Fibronectin is a large glycoprotein (220 kDa) present in a multimeric insoluble form in the extracellular matrixand basement membrane (Mosher, 1989). As the shdAgene is required for colonization of the murine caecum(Kingsley et al., 2001), we investigated the distribution of fibronectin in uninfected tissue by fluorescencemicroscopy using anti-human fibronectin antibody.Fibronectin localized to the basement membrane andconnective tissue of the murine caecum (Fig. 4B). The dis-tribution of receptors for ShdA in caecal tissue detectedby overlay with GST–ShdA fusion protein was similar tothat of fibronectin (Fig. 4C). These data demonstrated thata cognate receptor of ShdA is expressed in the murinecaecum, which is likely to be fibronectin. However, aspreincubation with antifibronectin antibodies resulted inonly an approximately twofold inhibition of GST–ShdAbinding to fibronectin in vitro (Fig. 2C), it is not feasible todetect blocking of GST–ShdA binding to thin sections insitu by fluorescence microscopy. Thus, the possibility thatShdA binds to receptors in addition to fibronectin in themurine caecum cannot currently be ruled out. None-theless, this study further supports a role for shdA inintestinal colonization by demonstrating the presence ofa cognate receptor in this tissue.
Microbial surface components, which mediate adher-ence, invasiveness or immune evasion by recognizingand binding adhesive matrix molecules (MSCRAMMs),are a recurring theme in host–pathogen interactions (Joh et al., 1999). For example, Mycobacterium aviumcomplex encodes the fibronectin attachment protein(FAP) (Schorey et al., 1996), which is involved in adher-ence to the human respiratory mucosa (Middleton et al.,2000). Furthermore, serum opacity factor (SOF) of M type2 Streptococcus pyogenes binds fibronectin and isrequired for full virulence in a murine model of infection(Courtney et al., 1999). A second fibronectin-bindingprotein of Streptococcus pyogenes, protein F, has beenimplicated in the invasion of HeLa cells through the inter-action of fibronectin with the b1-integrin receptor (Ozeriet al., 1998). Type I fimbriae of S. Typhimurium and E.coli bind to oligomannoside chains of laminin (Collinsonet al., 1993; Kukkonen et al., 1993). Thin aggregative fim-briae of S. enterica serotype Enteritidis bind fibronectin(Collinson et al., 1993; Kukkonen et al., 1993). Finally, the enteric pathogen Campylobacter jejuni encodes afibronectin-binding protein termed CadF (Konkel et al.,1997), which is required for colonization and persistencein the caecum of poultry (Ziprin et al., 1999). However,
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 895–905
902 R. A. Kingsley et al.
A B
Fig. 7. A. Thin section of a lymphoid follicle in the caecum 5 days after inoculation with S. Typhimurium (400¥).B. Thin section of murine caecum (adjacent to that shown in Fig. 1D) 5 days after inoculation containing erosion of the epithelium (400¥). Thearea shown in (B) corresponds to that shown for an adjacent section in Fig. 1D. S. Typhimurium LPS was detected in (A) and (B) byimmunofluorescence with sheep antisomatic (O4) antiserum and donkey anti-sheep Ig Alexafluor 488 (green signal in image i). ShdA wasdetected in (A) and (B) with anti-ShdA antiserum and goat anti-rabbit Ig Alexafluor 594 (red signal in image ii). Green, red and blue (Hoechstnuclear stain) channels were combined (image iii). Ep., epithelial cells; Lum., lumen.
Fig. 8. Thin section of the murine caecum from mice 5 days after inoculation with S. Typhimurium strain IR715 (wild type) (400¥).A and C. Images from an H&E-stained section of the caecum. Both images were taken from the same section and are separated by only onemicroscopic field.B and D. Fluorescence images of an adjacent section, showing the regions corresponding to those pictured in (A) and (C) respectively.A and B. A region of the caecum in which bacteria colonized the intestinal surface (green signal in B).C and D. An area of the caecum in which bacterial invasion of the mucosa had occurred. Note the migration of inflammatory cells into theintestinal lumen, a thickening of the mucosa and an increased cellularity of the caecal tissue visible in (C). Note an increased amount offibronectin (red signal) present in the mucosa and the presence of a large number of bacteria (green signal) in the intestinal lumen and inmucosal tissue in (D).S. Typhimurium LPS was detected in (B) and (D) by immunofluorescence with sheep antisomatic (O4) antiserum and donkey anti-sheep IgAlexafluor 488 (green signal). Fibronectin was detected in (B) and (D) with antifibronectin antiserum and goat anti-rabbit Ig Alexafluor 594 (red signal). Nuclei were detected with Hoechst (blue signal). M, mucosa; SM, submucosa; Lum., lumen.

Expression of S. enterica ShdA 903
none of these bacterial adhesins exhibits significantsequence homology to ShdA.
Although thin aggregative fimbriae of S. Enteritidis havebeen implicated in fibronectin binding (Collinson et al.,1993; Kukkonen et al., 1993), no differences between S.Typhimurium wild type and an agf mutant were detectedin their ability to bind fibronectin to the bacterial surfaceunder conditions used in this study (R. A. Kingsley and A. J. Bäumler, unpublished results). The reason for thiswas not investigated further but may be related to phase variation of the agf operon, variation in bindingspecificity between S. Typhimurium and S. Enteritidis fimbriae or poor expression of agf-encoded fimbriaeunder the conditions used in this study. Similarly, we foundthat S. Typhimurium wild type and shdA mutant did not differ in their ability to bind fibronectin in vitro, as the ShdA protein was not expressed under a variety of dif-ferent in vitro conditions. However, signals generatedusing antisomatic (O4) and anti-ShdA antibodies in thinsections of infected caecum co-localized (Figs 6 and 7).These data showed that expression of ShdA in S.Typhimurium strain ATCC14028 is induced by anunknown signal both in the lumen of the caecum (Fig. 6)and in the caecal mucosa (Fig. 7) in vivo. The strikingupregulation of ShdA expression in vivo further supporteda role for this fibronectin-binding protein in host–pathogeninteraction.
Experimental procedures
Bacterial strains and culture conditions
Strain IR715 is a nalidixic acid-resistant derivative of S.Typhimurium strain ATCC14028 (Stojiljkovic et al., 1995).AH9, a derivative of IR715 carrying a deletion of shdA, hasbeen described previously (R. A. Kingsley, A. D. Humphries,A. M. Keestra and A. J. Bäumler, submitted). Strain 14028ris a rough derivative of strain ATCC14028 (Fields et al.,1986). Strain RK715 is a spontaneous nalidixic acid-resistant derivative of 14028r. E. coli strain BL21 waspurchased from Invitrogen. Unless otherwise stated, strainswere cultured in standard laboratory media (Miller, 1972) supplemented as appropriate with carbenicillin (100 mg l–1),chloramphenicol (30 mg l–1), kanamycin (60 mg l–1), nalidixicacid (50 mg l–1) or deoxycholate (2%).
Animal experiments
Six- to 8-week-old female BALB/c (ByJ, Jackson Laboratory)mice were used throughout this study (n = 24). Bacteria wereroutinely cultured as static broth cultures in Luria–Bertani(LB) broth before intragastric inoculation with 1 ¥ 109 cfu in0.2 ml of 0.85% saline. In all experiments, the bacterial titreof the inoculum was determined by spreading serial 10-folddilutions on agar plates containing appropriate antibiotics anddetermining colony-forming units (cfu).
Anti-ShdA antiserum was raised in a 6 lb New Zealand
White rabbit. A prebleed (5 ml) was taken, followed by theprimary immunization of a 1:1 mixture of GST–ShdA protein(0.5 mg) and Titre-Max Gold (Sigma) in a total volume of 1ml injected subcutaneously at more than 10 different locations. This procedure was repeated after 14 and 28days. After a further 2 weeks, the rabbit was anaesthetized(ketamine, xylaxine and acepromazine cocktail) and exsan-guinated by cardiac puncture. The serum was prepared bystandard methodology (Harlow, 1988) and preabsorbed with E. coli BL21 pGEX4T/1 (expressing GST) and S.Typhimurium strain AH9 (shdA).
Histopathology, immunocytochemistry andimmunofluorescence microscopy
At necropsy, tissue samples from the caecum were coded for blind examination, fixed in 10% formalin at room tem-perature or in cold ethanol for 24 h and paraffin embeddedby standard procedures. Sections of 0.005 mm weremounted on siliconized glass slides and stained with haema-toxylin and eosin (H&E) for histopathological examination.For immunocytochemistry and immunofluorescence, sec-tions were deparaffinized by incubation at 50∞C for 20 min,three 5 min washes in xylene and subsequent stepwise rehydration through solutions with decreasing alcoholcontent. Sections were incubated with blocking buffer (2%dried non-fat milk and 0.1% Tween 20 in PBS) for 1 h andthen with primary antibody for 1 h. For immunocytochemistry,the primary antibody was rabbit antisomatic (O4, O5) antiserum diluted 1:3000 in blocking buffer. For immuno-fluorescence, the primary antibodies were sheep anti-O4antiserum (SA Scientific), rabbit anti-ShdA antiserum, affinity-purified anti-human fibronectin antiserum or goat anti-GST antiserum (Amersham Pharmacia), each diluted1:500 in blocking buffer. Tissue sections were washed exten-sively in blocking buffer, followed by incubation in secondaryantibody for 1 h. For immunocytochemistry, the secondaryantibody was goat anti-rabbit immunoglobulin (Ig) diluted1:1000 in blocking buffer, and sections were counterstainedwith haematoxylin. For immunofluorescence, the secondaryantibodies were goat anti-rabbit Alexafluor 594 (MolecularProbes) or donkey anti-sheep Alexafluor 488 (MolecularProbes) diluted 1:500 in blocking buffer. Tissue sections wereexamined using an epifluorescence microscope (OlympusVanox AHB53).
For binding of GST–ShdA or GST to thin sections of murine caecum in overlay experiments, sections fixed withcold ethanol were incubated with blocking buffer, followed by overlay with 0.05 mg ml–1 GST–ShdA or GST in PBS.Unbound protein was removed by submersion in block-ing buffer, and the remaining protein was detected with rabbit anti-ShdA or goat anti-GST followed by goat anti-rabbit Ig Alexafluor 594 or donkey anti-goat Ig Alexafluor 594. Sections were examined using an epifluorescence microscope.
Flow cytometry
Cultured bacterial cells were fixed in 1% paraformaldehydein PBS for 15 min at room temperature and washed in PBS.
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 895–905

The cells were blocked with 5% normal goat serum, thenincubated with rabbit anti-ShdA antiserum followed bywashes in PBS and incubation with secondary antibody (goatanti-rabbit Alexafluor 488; Molecular Probes) and 0.04 mMpropidium iodide. The cells were then analysed by flowcytometry (FACSCalibur; Becton Dickinson).
Expression and purification of GST and GST–ShdAfusion protein
A translational fusion of the ShdA N-terminal domain, extend-ing from the predicted signal peptide cleavage site to thebeginning of the C-terminal domain predicted by sequencehomology with IcsA and AIDA (residues 59–1559), with GST was constructed as follows. A region of the shdAgene was polymerase chain reaction (PCR) amplified using the primers 5¢-GTCGACTACGTGCTGGAGCTGCTACC and 5¢-GAATTCGCCGATAATCAGGTATCATATGC-3¢.The PCR product was digested with EcoRI and SalI andligated into pGEXT4/1 (Amersham Pharmacia) digested withEcoRI and SalI. The resulting plasmid (pRA118) was intro-duced into E. coli strain BL21, cells were grown to an OD600
of 0.6, and expression of the fusion protein was induced bythe addition of IPTG to a final concentration of 1 mM. After 3 h, cells were harvested and lysed using a French press at12 000 p.s.i. The GST–ShdA fusion protein was purified byaffinity chromatography from the soluble fraction using glu-tathione sepharose (Amersham Pharmacia) according to themanufacturer’s instructions. Purification of GST was per-formed by the same method using E. coli BL21(pGEXT4/1).
Expression and subcellular localization of ShdA
An ª 9 kb HindIII fragment extending from ª 900 bp upstreamof the shdA ATG start to 1 kb downstream of the shdA stopcodon was cloned into the HindIII site of pWSK29 (Wang andKushner, 1991) to give rise to plasmid pRA69. Sequenceanalysis of pRA69 revealed that shdA was cloned in an ori-entation such that expression could be driven by the T7 pro-moter of pWSK29. Plasmids pRA69 and pGP1-2 (Tabor andRichardson, 1985) were introduced into strain RK715, andexpression of T7 RNA polymerase from pGP1-2 was inducedby a 30 min incubation at 42∞C as described previously(Tabor and Richardson, 1985).
Subcellular fractionation of S. enterica was performed asdescribed previously (Hantke, 1981). Proteins were sepa-rated by electrophoresis on a 4–15% Tris-HCl polyacrylamidegradient gel (Readygel; Bio-Rad) and transferred to Immo-bilon-P membrane (Millipore) using a Trans-Blot SD semi-dryelectrophoretic transfer cell (Bio-Rad) according to the manu-facturer’s instructions. Immobilized protein was detected with rabbit anti-ShdA antibody (1:500 dilution) and goat anti-rabbit Ig alkaline phosphatase conjugate (Bio-Rad). Signalwas generated using Immun-star substrate (Bio-Rad).
Extracellular matrix protein-binding assays
Fibronectin (bovine plasma), collagen type I (calf skin), col-lagen type IV (human placenta) and bovine serum albumin(BSA) used in binding assays were purchased from Sigma.
For the solid phase-binding assay, 96-well microtitre platewells (Maxisorp; Nunc) were coated by the addition of 0.05 ml per well of test protein (fibronectin, collagen I, colla-gen IV, BSA) at the indicated concentration in Tris-HCl,pH 8.5, and allowed to bind overnight at 4∞C. Unboundprotein was removed by washing eight times with d.H2O, and0.2 ml per well of blocking buffer with 0.004% (v/v) antifoamA and 0.1% (w/v) sodium azide were added. Wells werewashed four times with d.H2O followed by the addition of 0.1ml of 0.01mg ml–1 GST or GST–ShdA fusion protein inPBS and incubation for 2 h at room temperature. Wells werewashed eight times with d.H2O and incubated with a 1:1000dilution of goat anti-GST immune serum in blocking buffer(Amersham Pharmacia) for 90 min at room temperature.Wells were washed eight times with d.H2O, and a 1:1000 dilu-tion of secondary antibody (mouse anti-goat Ig alkaline phos-phatase conjugate; Bio-Rad) in blocking buffer was added.Alkaline phosphatase activity in each well was determinedafter the addition of 1 mg ml–1 p-nitrophenyl phosphate(pNPP) by measuring the absorbance at 405 nm. Beforeanalysis of data, the background signal, from wells in whichGST or GST–ShdA were not added, was subtracted from thesignal in test wells. For inhibition of GST–ShdA binding, wellswere coated with fibronectin and then incubated with rabbitanti-human fibronectin antibody (Sigma) diluted 1:1000,1:500, 1:100, 1:50 or 1:10 in PBS. Binding in antibody-treatedwells was expressed as a percentage of binding observed incontrol wells.
For the fibronectin pull-down assays, a 0.2 ml bed-volumeof glutathione sepharose (Amersham Pharmacia) beads wasloaded with 0.05 mg of GST or GST–ShdA in PBS, washedtwice with PBS. The beads were resuspended in 0.05 mg ofbovine plasma fibronectin in 0.2 ml of PBS, incubated at roomtemperature for 90 min and washed twice with PBS. GST orGST–ShdA was eluted with 0.2 ml of 10 mM reduced glu-tathione (Sigma) in 50 mM Tris-HCl, pH 8.0. Eluted proteinwas analysed by SDS–PAGE, transferred to Immobilon-Pmembrane and detected with rabbit anti-human fibronectinimmune serum and goat anti-rabbit Ig alkaline phosphataseconjugate.
For fibronectin binding to S. Typhimurium RK715 (pGP1-2) and RK715 (pGP1-2, pRA69), expression of T7 poly-merase was induced as described above. Approximately1 ¥ 109 cfu were harvested, washed twice in 0.5 ml of PBSand finally resuspended in 0.2 ml of PBS. Fibronectin (0.05 mg) was added and incubated at room temperature for1 h. Unbound fibronectin was removed by washing cells fourtimes in PBS. Cell-associated fibronectin was detected byseparation of whole-cell proteins on 4–15% SDS–PAGE (Bio-Rad), Western transfer and detection with rabbit anti-human fibronectin immune serum.
Acknowledgements
We would like to thank Renée M. Tsolis and Clare V. Smithfor critical comments on this manuscript, and R. Vollmar, S.Hill and S. Ambrus (VTPB, Histology Department, VeterinarySchool, Texas A and M University, USA) for technical assis-tance with histopathology and immunohistochemistry. Thismaterial is based in part upon work supported by the TexasAdvanced Research (Technology) Program under grant
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 895–905
904 R. A. Kingsley et al.

Expression of S. enterica ShdA 905
number 000089-0051-1999. Work in A.B.’s laboratory is sup-ported by Public Health Service grants AI40124 and AI44170.
References
Aleksic, S., Heinzerling, F., and Bockemühl, J. (1996) Humaninfection caused by salmonellae of subspecies II to VI inGermany, 1977–92. Zbl Bakt 283: 391–398.
Benz, I., and Schmidt, M.A. (1989) Cloning and expressionof an adhesin (AIDA-I) involved in diffuse adherence of enteropathogenic Escherichia coli. Infect Immun 57:1506–1511.
Bernardini, M.L., Mounier, J., d’Hauteville, H., Coquis-Rondon, M., and Sansonetti, P.J. (1989) Identification oficsA, a plasmid locus of Shigella flexneri that governs bacterial intra- and intercellular spread through interactionwith F-actin. Proc Natl Acad Sci USA 86: 3867–3871.
Collinson, S.K., Doig, P.S., Doran, J.L., Clouthier, S., Trust,T.J., and Kay, W.W. (1993) Thin, aggregative fimbriaemediate binding of Salmonella enteritidis to fibronectin.J Bacteriol 175: 12–18.
Courtney, H.S., Hasty, D.L., Li, Y., Chiang, H.C., Thacker,J.L., and Dale, J.B. (1999) Serum opacity factor is a majorfibronectin-binding protein and a virulence determinant ofM type 2 Streptococcus pyogenes. Mol Microbiol 32:89–98.
Fields, P.I., Swanson, R.V., Haidaris, C.G., and Heffron, F.(1986) Mutants of Salmonella typhimurium that cannotsurvive within the macrophage are avirulent. Proc NatlAcad Sci USA 83: 5189–5193.
Galbraith, N.S. (1961) Studies of human salmonellosis inrelation to infection in animals. Vet Record 73: 1296–1303.
Grinnell, F., Billingham, R.E., and Burgess, L. (1981) Distrib-ution of fibronectin during wound healing in vivo. J InvestDermatol 76: 181–189.
Hantke, K. (1981) Regulation of the ferric iron transport in Escherichia coli K12: isolation of a constitutive mutant.Mol Gen Genet 182: 288–292.
Harlow, E., and Lane, D. (1988) Antibodies: A LaboratoryManual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Joh, D., Wann, E.R., Kreikemeyer, B., Speziale, P., and Hook,M. (1999) Role of fibronectin-binding MSCRAMMs in bacterial adherence and entry into mammalian cells.Matrix Biol 18: 211–223.
Kingsley, R.A., van Amsterdam, K., Kramer, N., and Baumler,A.J. (2000) The shdA gene is restricted to serotypes of Salmonella enterica subspecies I and contributes to efficient and prolonged fecal shedding. Infect Immun 68:2720–2727.
Konkel, M.E., Garvis, S.G., Tipton, S.L., Anderson, D.E., Jr,
and Cieplak, W., Jr (1997) Identification and molecularcloning of a gene encoding a fibronectin- binding protein(CadF) from Campylobacter jejuni. Mol Microbiol 24:953–963.
Kukkonen, M., Raunio, T., Virkola, R., Lahteenmaki, K.,Makela, P.H., Klemm, P., et al. (1993) Basement mem-brane carbohydrate as a target for bacterial adhesion:binding of type I fimbriae of Salmonella enterica andEscherichia coli to laminin. Mol Microbiol 7: 229–237.
Kurkinen, M., Vaheri, A., Roberts, P.J., and Stenman, S.(1980) Sequential appearance of fibronectin and collagenin experimental granulation tissue. Lab Invest 43: 47–51.
Middleton, A.M., Chadwick, M.V., Nicholson, A.G., Dewar, A.,Groger, R.K., Brown, E.J., and Wilson, R. (2000) The roleof Mycobacterium avium complex fibronectin attachmentprotein in adherence to the human respiratory mucosa.Mol Microbiol 38: 381–391.
Miller, J.H. (1972) Experiments in Molecular Genetics. ColdSpring Harbor, NY: Cold Spring Harbor Laboratory Press.
Mosher, D.F. (1989) Fibronectin. New York: Academic Press.Ozeri, V., Rosenshine, I., Mosher, D.F., Fassler, R., and
Hanski, E. (1998) Roles of integrins and fibronectin in theentry of Streptococcus pyogenes into cells via protein F1. Mol Microbiol 30: 625–637.
Raghow, R. (1994) The role of extracellular matrix in postin-flammatory wound healing and fibrosis. FASEB J 8:823–831.
Schorey, J.S., Holsti, M.A., Ratliff, T.L., Allen, P.M., andBrown, E.J. (1996) Characterization of the fibronectin-attachment protein of Mycobacterium avium reveals afibronectin-binding motif conserved among mycobacteria.Mol Microbiol 21: 321–329.
Stojiljkovic, I., Bäumler, A.J., and Heffron, F. (1995)Ethanolamine utilization in Salmonella typhimurium.nucleotide sequence, protein expression and mutationalanalysis of the cchA cchB eutE eutJ eutG eutH genecluster. J Bacteriol 177: 1357–1366.
Tabor, S., and Richardson, C.C. (1985) A bacteriophage T7RNA polymerase/promoter system for controlled exclusiveexpression of specific genes. Proc Natl Acad Sci USA82: 1074–1078.
Wang, R.F., and Kushner, S.R. (1991) Construction of versatile low-copy-number vectors for cloning, sequencingand gene expression in Escherichia coli. Gene 100:195–199.
Witte, M.B., and Barbul, A. (1997) General principles ofwound healing. Surg Clin North Am 77: 509–528.
Ziprin, R.L., Young, C.R., Stanker, L.H., Hume, M.E., andKonkel, M.E. (1999) The absence of cecal colonization ofchicks by a mutant of Campylobacter jejuni not express-ing bacterial fibronectin-binding protein. Avian Dis 43:586–589.
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 895–905
Copyright © 2022 FDOKUMEN




















