
Hydrogen peroxide and hypochlorous acid influx through the major S. Typhimurium porin OmpD is...
8
1 3 Hydrogen peroxide and hypochlorous acid influx through the major 4 S. Typhimurium porin OmpD is affected by substitution 5 of key residues of the channel Q1 6 7 8 Daniel Aguayo b,d,1 Q2 , Nicolás Pacheco a,1 , Eduardo H. Morales c,1 , Bernardo Collao a , Roberto Luraschi a , 9 Carolina Cabezas a , Paulina Calderon a , Fernando D. González b,d , Fernando Gil a , Iván L. Calderón a , 10 Claudia P. Saavedra a,⇑ 11 a Laboratorio de Microbiología Molecular, Departamento de Ciencias Biológicas, Facultad de Ciencias Biológicas, Universidad Andres Bello, Santiago, Chile 12 b Center for Bioinformatics and Integrative Biology, Facultad de Ciencias Biológicas, Universidad Andres Bello, Santiago, Chile 13 c Laboratorio de Microbiología Molecular, Departamento de Biología, Facultad de Química y Biología, Universidad de Santiago de Chile, Chile Q3 14 d Centro Interdisciplinario de Neurociencia de Valparaíso, Facultad de Ciencias, Universidad de Valparaíso, Valparaíso 2366103, Chile 15 16 17 19 article info 20 Article history: 21 Received 1 October 2014 22 and in revised form 7 January 2015 23 Available online xxxx 24 Keywords: 25 OmpD porin 26 Hydrogen peroxide 27 Hypochlorous acid 28 Homology modeling 29 Molecular simulations Q4 30 31 abstract 32 OmpD is the major Salmonella enterica serovar Typhimurium (S. Typhimurium) porin and mediates 33 hydrogen peroxide (H 2 O 2 ) influx. The results described herein extend this finding to hypochlorous acid 34 (HOCl), another reactive oxygen species that is also part of the oxidative burst generated by the 35 phagosome. S. Typhimurium cells lacking OmpD show decreased HOCl influx, and OmpD-reconstituted 36 proteoliposomes show an increase in the uptake of the toxic compound. To understand this physiologi- 37 cally relevant process, we investigated the role of key OmpD residues in H 2 O 2 and NaOCl transport. Using 38 a theoretical approach, residue K16 was defined as a major contributor to the channel electrostatic 39 properties, and E111 was shown to directly participate in the size-exclusion limit of the channel. 40 Together, we provide theoretical, genetic, and biochemical evidence that OmpD mediates H 2 O 2 and 41 NaOCl uptake, and that key residues of the channel are implicated in this process. 42 Ó 2015 Published by Elsevier Inc. 43 44 45 46 Introduction 47 Q5 Porins are water-filled channels that span the outer membrane 48 (OM) 2 of Gram-negative bacteria and that mediate the uptake of 49 hydrophilic solutes [1,2], nutrients, and toxic compounds from the 50 external environment into the periplasm, and vice versa [3,4]. The 51 Escherichia coli OmpF porin has served as a model to characterize 52 several properties of porins including ionic preference [1], the effect 53 of ionic strength, antibiotic uptake, antibiotic resistance [5], and the 54 contribution of protein residues to the diffusion potential, among 55 others. The availability of the three-dimensional structure of OmpF 56 and other porins at atomic resolution has allowed determining a 57 relation between the sequence, channel structure, and functional 58 properties of porins [5]. In example, OmpF monomers, which form 59 trimers in the outer membrane, exhibit a b-barrel domain 60 constricted by an extracellular loop (L-3) that folds into the channel 61 vestibule [6,7]. In the inner barrel wall, the positively charged 62 residues K16, R42, R82, and R132 are located on the opposite side 63 of the negatively charged residues D113 and E117 at L-3 [8]. The dis- 64 tribution of these residues defines the geometrical and electrostatic 65 properties of the constriction zone, termed ‘‘eyelet’’, and delimits the 66 size, charge, and properties of the molecules to be channeled. Muta- 67 tion of the eyelet residues in E. coli OmpF and OmpC [9,10] and other 68 homologous porins affects antibiotic diffusion and susceptibility to 69 the compounds [11–17]. 70 OmpD is the most abundant OM porin of Salmonella enterica 71 serovar Typhimurium (S. Typhimurium) [18] and mediates the 72 uptake of hydrogen peroxide (H 2 O 2 ) [4,11], a reactive oxygen 73 species (ROS) produced in the oxidative burst by macrophages 74 [19]. Interestingly, the expression of nmpC (encoding the OmpD 75 porin) is down-regulated when S. Typhimurium is exposed to 76 H 2 O 2 , hypochlorous acid (NaOCl) [11], and when residing inside 77 macrophages [4,11,20]. This suggests that the regulation of OmpD http://dx.doi.org/10.1016/j.abb.2015.01.005 0003-9861/Ó 2015 Published by Elsevier Inc. ⇑ Corresponding author at: Laboratorio de Microbiología Molecular, Departa- mento de Ciencias Biológicas, Universidad Andres Bello, República 217, Santiago, Chile. E-mail address: [email protected] (C.P. Saavedra). 1 Authors contributed equally. 2 Abbreviations used: OM, outer membrane; ROS, reactive oxygen species; LB, Luria Bertani; DHR123, dihydrorhodamine123; POPC, palmitoyl oleoyl phosphotidyl choline. Archives of Biochemistry and Biophysics xxx (2015) xxx–xxx Contents lists available at ScienceDirect Archives of Biochemistry and Biophysics journal homepage: www.elsevier.com/locate/yabbi YABBI 6877 No. of Pages 8, Model 5G 16 January 2015 Please cite this article in press as: D. Aguayo et al., Arch. Biochem. Biophys. (2015), http://dx.doi.org/10.1016/j.abb.2015.01.005
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Hydrogen peroxide and hypochlorous acid influx through the major S. Typhimurium porin OmpD is...

1
3
4
5 Q1
6
7
8 Q2
9
10
111213 Q314
151617
1 9
20212223
242526272829 Q430
3 1
45
46
47 Q5
48
49
50
51
52
53
54
55
56
57
Archives of Biochemistry and Biophysics xxx (2015) xxx–xxx
YABBI 6877 No. of Pages 8, Model 5G
16 January 2015
Contents lists available at ScienceDirect
Archives of Biochemistry and Biophysics
journal homepage: www.elsevier .com/ locate/yabbi
Hydrogen peroxide and hypochlorous acid influx through the majorS. Typhimurium porin OmpD is affected by substitutionof key residues of the channel
http://dx.doi.org/10.1016/j.abb.2015.01.0050003-9861/� 2015 Published by Elsevier Inc.
⇑ Corresponding author at: Laboratorio de Microbiología Molecular, Departa-mento de Ciencias Biológicas, Universidad Andres Bello, República 217, Santiago,Chile.
E-mail address: [email protected] (C.P. Saavedra).1 Authors contributed equally.2 Abbreviations used: OM, outer membrane; ROS, reactive oxygen species; LB, Luria
Bertani; DHR123, dihydrorhodamine123; POPC, palmitoyl oleoyl phosphotidylcholine.
Please cite this article in press as: D. Aguayo et al., Arch. Biochem. Biophys. (2015), http://dx.doi.org/10.1016/j.abb.2015.01.005
Daniel Aguayo b,d,1, Nicolás Pacheco a,1, Eduardo H. Morales c,1, Bernardo Collao a, Roberto Luraschi a,Carolina Cabezas a, Paulina Calderon a, Fernando D. González b,d, Fernando Gil a, Iván L. Calderón a,Claudia P. Saavedra a,⇑a Laboratorio de Microbiología Molecular, Departamento de Ciencias Biológicas, Facultad de Ciencias Biológicas, Universidad Andres Bello, Santiago, Chileb Center for Bioinformatics and Integrative Biology, Facultad de Ciencias Biológicas, Universidad Andres Bello, Santiago, Chilec Laboratorio de Microbiología Molecular, Departamento de Biología, Facultad de Química y Biología, Universidad de Santiago de Chile, Chiled Centro Interdisciplinario de Neurociencia de Valparaíso, Facultad de Ciencias, Universidad de Valparaíso, Valparaíso 2366103, Chile
a r t i c l e i n f o a b s t r a c t
3233343536373839404142
Article history:Received 1 October 2014and in revised form 7 January 2015Available online xxxx
Keywords:OmpD porinHydrogen peroxideHypochlorous acidHomology modelingMolecular simulations
43
OmpD is the major Salmonella enterica serovar Typhimurium (S. Typhimurium) porin and mediateshydrogen peroxide (H2O2) influx. The results described herein extend this finding to hypochlorous acid(HOCl), another reactive oxygen species that is also part of the oxidative burst generated by thephagosome. S. Typhimurium cells lacking OmpD show decreased HOCl influx, and OmpD-reconstitutedproteoliposomes show an increase in the uptake of the toxic compound. To understand this physiologi-cally relevant process, we investigated the role of key OmpD residues in H2O2 and NaOCl transport. Usinga theoretical approach, residue K16 was defined as a major contributor to the channel electrostaticproperties, and E111 was shown to directly participate in the size-exclusion limit of the channel.Together, we provide theoretical, genetic, and biochemical evidence that OmpD mediates H2O2 andNaOCl uptake, and that key residues of the channel are implicated in this process.
� 2015 Published by Elsevier Inc.
44
58
59
60
61
62
63
64
65
66
67
68
69
70
Introduction
Porins are water-filled channels that span the outer membrane(OM)2 of Gram-negative bacteria and that mediate the uptake ofhydrophilic solutes [1,2], nutrients, and toxic compounds from theexternal environment into the periplasm, and vice versa [3,4]. TheEscherichia coli OmpF porin has served as a model to characterizeseveral properties of porins including ionic preference [1], the effectof ionic strength, antibiotic uptake, antibiotic resistance [5], and thecontribution of protein residues to the diffusion potential, amongothers. The availability of the three-dimensional structure of OmpFand other porins at atomic resolution has allowed determining arelation between the sequence, channel structure, and functional
71
72
73
74
75
76
77
properties of porins [5]. In example, OmpF monomers, which formtrimers in the outer membrane, exhibit a b-barrel domainconstricted by an extracellular loop (L-3) that folds into the channelvestibule [6,7]. In the inner barrel wall, the positively chargedresidues K16, R42, R82, and R132 are located on the opposite sideof the negatively charged residues D113 and E117 at L-3 [8]. The dis-tribution of these residues defines the geometrical and electrostaticproperties of the constriction zone, termed ‘‘eyelet’’, and delimits thesize, charge, and properties of the molecules to be channeled. Muta-tion of the eyelet residues in E. coli OmpF and OmpC [9,10] and otherhomologous porins affects antibiotic diffusion and susceptibility tothe compounds [11–17].
OmpD is the most abundant OM porin of Salmonella entericaserovar Typhimurium (S. Typhimurium) [18] and mediates theuptake of hydrogen peroxide (H2O2) [4,11], a reactive oxygenspecies (ROS) produced in the oxidative burst by macrophages[19]. Interestingly, the expression of nmpC (encoding the OmpDporin) is down-regulated when S. Typhimurium is exposed toH2O2, hypochlorous acid (NaOCl) [11], and when residing insidemacrophages [4,11,20]. This suggests that the regulation of OmpD

78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
2 D. Aguayo et al. / Archives of Biochemistry and Biophysics xxx (2015) xxx–xxx
YABBI 6877 No. of Pages 8, Model 5G
16 January 2015
is an adaptive response to avoid the harmful conditions encoun-tered during the infective process.
Recent studies have provided biochemical and genetic evidencefor the role of OmpD in H2O2 and NaOCl resistance [4,11]; however,surprisingly no studies have investigated how these toxiccompounds are channeled through this porin. To understand thisphysiologically relevant process, we investigated the role of keyOmpD residues in H2O2 and NaOCl transport. First, we generateda homology model of OmpD by using the structures of E. coli OmpFand OmpC, for which structures have been resolved at atomic res-olution [21,22] and that show high sequence identity with OmpD.Then, this model was used to perform molecular dynamics simula-tion to identify candidate residues important for H2O2 and NaOCltransport. Our predictions where then tested in vivo and in vitroby using cells expressing point mutants of OmpD and using thesesame variants in proteoliposomes, respectively.
135
136
137
138
139
140
141
142
143
Materials and methods
Bacterial strains and growth conditions
Strains used in this study are listed in Table 1. Bacteria weregrown routinely at 37 �C in Luria Bertani (LB) medium withshaking (250 rpm). When required, LB was supplemented withampicillin (Amp, 100 mg/ml). Genetically complemented strainswere grown in culture media supplemented with 1 mM arabinose.
144
145
146
147
148
149
150
151
152
153
154
155
Cloning, overexpression and purification of OmpD
The S. Typhimurium ompD gene was amplified by PCR, clonedinto pBAD-TOPO TA and transformed into E. coli Top 10 (Invitro-gen�). Primer sequences were 50 ATGAAACTTAAGTTAGTGGC 30
(pBAD-ompDF) and 50 GAACTGGTAGTTCAGACCAA 30 (pBA-DompDR). E. coli Top 10 carrying pBAD-ompD were grown in 1Lof LB medium at 37 �C to an OD600 �0.6. Protein overexpressionwas performed for 5 h in the presence of 1 mM arabinose. Mem-brane proteins were obtained as described by Surrey and Jahnig[23]. His-tagged porins were purified by affinity chromatographyusing HisTrap HP columns (Amersham�) according to themanufacturer’s instructions.
156
157
158
159
160
161
162
163
CFU assay
The number of viable microorganisms was estimated by themethod described by Miles et al. [24] and was expressed as cfu/mL. Briefly, bacterial cultures at their exponential phase(OD600 � 0.4) were exposed for 30 min to the toxic compounds.The cultures were then serially diluted, plated into M9 mediumand incubated for 12 h prior to the colony counting process.
Table 1Bacterial strains used in this study.
Strain Genotype
14028s Wild type SalmonE. coli Top 10 F�mcrAD(mrr-hsd
araD139 D(ara-leS. Typhimurium ompD DompD::FRTS. Typhimurium ompD-pBAD DompD::FRT/pBAS. Typhimurium-pBAD DompD::FRT/pBAS. Typhimurium ompD-pBAD ompD K16Q DompD::FRT/pBAS. Typhimurium ompDpBADompD E111C DompD::FRT/pBAE. coli Top 10 pBAD-ompD Top 10/pBAD-omE. coli Top 10 pBAD-ompD K16Q Top 10/pBAD-omE. coli Top 10 pBAD-ompD E111C Top 10/pBAD-om
Please cite this article in press as: D. Aguayo et al., Arch. Biochem. Biophys. (2
In vivo diffusion of H2O2 and NaOCl
The levels of these toxic compounds were determined using thefluorescent probe dihydrorhodamine123 (DHR123) as previouslydescribed [25]. Briefly, an overnight culture was diluted 100-foldinto fresh LB medium and grown at 37 �C with shaking to anOD600 �0.6. A two ml aliquot was centrifuged for 3 min at4500 rpm (centrifuge 5702 Eppendorf�), cells were recovered andresuspended in 1 ml of 50 mM sodium phosphate buffer, pH 7.2.Two hundred ml aliquot of this bacterial suspension was incubatedfor 30 min with 2 mM H2O2 or 5 mM NaOCl, and then vacuum-filtered using polycarbonate filters of 0.025 mm (Millipore). Thepass through (extracellular fraction) was collected and bacteriaretained in the filter (intracellular fraction) were suspended with2 ml of 50 mM sodium phosphate buffer pH 7.2 and disrupted bysonication. Both the extracellular and intracellular fractions wereincubated for 30 min separately with 10 ll of (2.5 lg/ml)DHR123 and fluorescence was measured at 485 nm (excitation)and 430 nm (emission) in a TBS-380 Fluorometer (Turner BioSys-tems). Hydrogen peroxide and NaOCl uptake was determined asthe extracellular/intracellular fluorescence ratio. The backgroundfluorescence from control bacterial suspension (not exposed toH2O2 and NaOCl) was subtracted from all measurements and theresults were normalized by protein concentration.
In vitro diffusion of H2O2 and NaOCl
Proteoliposomes prepared as described by Nikaido and Rosen-berg [26] with modifications by Cubillos et al. [27] were incubatedwith 2 mM H2O2 or 5 mM NaOCl for 5 min and then vacuum-fil-tered to determine fluorescence as described above. Fluorescencewas measured in both the extra-liposomal and intra-liposomalfractions and H2O2 or NaOCl uptake was determined as theextra-/intra-liposomal fluorescence ratio as described above.
PCR conditions for megaprimer synthesis
PCRs were performed using Pfu DNA polymerase (Stratagene�).A 50 ll reaction mixture consisted of 1–10 ng of plasmid DNAcarrying the target DNA template (pBAD-TOPO TA/ompD),1.3 mmoles of MgCl2, 10 nmoles of dNTP, 20 pmol each of forwardand reverse primers (Table 2) and 2.5 units of DNA polymerase.Thermal cycling conditions were: 95 �C for 3 min followed by 30cycles of 95 �C for 30 s, 55 �C for 30 s, and 72 �C for 60 s and a finalstep at 72 �C for 10 min.
Overlapping extension
Five microliters from the first PCR reactions were added into15 ll of a PCR reaction mixture without addition of flanking prim-ers. The reactions were carried out for ten cycles to generate
Source
ella enterica serovar Typhimurium C. SaavedraRMS-mcrBC) U80lacZDM15 DlacX74 recA1u)7697 galUgalKrpsL (StrR) endA1nupG
Invitrogen
C. SaavedraD-ompD This workD This workD-ompDK16Q This workD-ompDE111C This workpD This workpD K16Q This workpD E111C This work
015), http://dx.doi.org/10.1016/j.abb.2015.01.005

164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202 Q6
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
271
272
Table 2Primers used in this study.
Primers Nucleotidic sequence (50–30) Tm (�C)
ompD F GCAAATAATCGGATTCATAA 54ompD R AGCCATTAGGCTGATCAGTA 53ompD K16Q F GGCAATAAACTGGATCTGTACGGTCAGGTTCACGC 78ompD K16Q R GCTGAGCGTGAACCAGACGGTACAGATCCAGTTTA 78ompD E111C F TGGACCGACGTTCTGCCATGCTTTGGTGGCGATACC 70ompD E111C R GGTATCGCCACCAAAGCATGGCAGAACGTCGGTCC 70
D. Aguayo et al. / Archives of Biochemistry and Biophysics xxx (2015) xxx–xxx 3
YABBI 6877 No. of Pages 8, Model 5G
16 January 2015
mutated templates. Then, an equal volume (25 ll) of a PCR reactionmixture containing 20 pmol of primers (Table 2) and 2.5 units ofTaq DNA polymerase was added into the reaction to obtain a50 ll reaction mixture. Amplification of the mutated product wasperformed for 30 cycles as described above. The mutated DNAproducts (50–100 ng) were gel purified and cloned into the pBADTOPO� vector. The vector was transformed into E. coli Top 10.Ten white colonies growing on a LB containing ampicillin(100 lg/ml) and X-gal (30 lg/ml) agar plate were randomly pickedfor sequencing. Nucleotide sequences were analyzed using ABIPRISM� 3100 Genetic Analyzer from Applied Biosystems.
OmpD homology modeling
The amino acid sequence of OmpD from S. Typhimurium (Uni-Prot accession number: D0ZXQ1) was used to find OmpD-homolo-gous proteins with known structure using a Blastp sequence-basedsearch against the Protein Data Bank [28]. The best-alignedsequences were those from OmpK36 (UniProt Q48473) from Kleb-siella pneumoniae, and OmpC (UniProt Q9K597) and OmpF (UniProtP02931) from E. coli. The OmpF structure at 2.4 Å resolution (pdb:2OMF, [8]) was selected as the 3D structural template, based on itsresolution and sequence alignment. Several full atomic three-dimensional models of OmpD trimers and selected amino acid sub-stitutions were generated using the program MODELLER v9.10[29]. The best models obtained according to their DOPE potentialwere further relaxed using the CHARMM 36 force field [30,31]and the NAMD v2.9 program [32] as previously described [33].Briefly, the amino acid’s protonation states were determined usingthe PROPKA3 method [34]. Then, an energy minimization protocolwas performed using the steepest-descent algorithm to removeinitial amino acids clashes, filling vacuum pockets and undesiredatom contacts, followed by a molecular dynamics with the proteinbackbone restricted using an harmonic potential of 0.5 kcal/mol Å2,with the protein loops set to move freely. Molecular dynamics sim-ulations lasted until energetic plateaus were observed.
Molecular dynamics simulations
Molecular dynamics simulations (MD) were carried out startingfrom the best model obtained above. OmpD and OmpD-substitutedtrimers were embedded independently into a palmitoyl oleoylphosphotidyl choline (POPC) lipid bilayer and solvated using a3 nm slab of water from the farthest protein atom. The hydratedsystems were neutralized using Na+ and Cl� counterions at0.150 M. The hydrated systems were first energy-minimized for5000 steps and equilibrated for 1 ns at constant volume and310 K (NVT), with the protein backbone restricted using a har-monic potential of 0.5 kcal/mol Å2. The systems were then equili-brated at constant pressure and temperature (NPT) for 2 ns withonly the protein backbone restrained, and equilibrated, withoutrestraints for a period of 2 ns prior to the production simulation,which lasted 20 ns per system. The MD simulations were carriedout with a time step of 2 fs and a 12-Å spherical cutoff for short-range non-bonded interactions, including a switching function
Please cite this article in press as: D. Aguayo et al., Arch. Biochem. Biophys. (2
from 10 Å for the van der Waals term and shifted electrostatics.For long-range electrostatic interactions, the particle-mesh Ewaldmethod [35] was utilized with grid spacing in the order of 1 Å orless. The production simulations were performed at 310 K in theNPT ensemble with fully anisotropic pressure coupling using theLangevin piston method [36]. The temperature was maintainedconstant using Langevin dynamics with a damping coefficient of1 ps�1. Periodic boundary conditions were applied to all simula-tions. The equations of motion were integrated with the RESPAmultiple time-step algorithm [37], with an effective time step of2, 2 and 4 fs for bonded, short- and long-range non-bonded inter-actions, respectively. The TIP3P model of water was used [38]. Allthe simulations used the CHARMM 36 force field for lipids and pro-teins with CMAP corrections [30,31].
Electrostatic and pore geometrical properties calculations
The electrostatic potential profiles of the systems were calcu-lated using the APBS program [39]. Pore radius profiles were ana-lyzed using the HOLE algorithm [40]. The pore axis was calculatedbased on the pore-slab center position calculated by the HOLEalgorithm and aligned with the z-Cartesian coordinate axis. Molec-ular graphics images were generated using VMD [41]. Electrostaticpotential and channel analyses were performed for each proteinmonomer and over the complete production simulations, whichlasted 20 ns, and averages over this period of time are reportedherein.
Results
Typhimurium OmpD mediates H2O2 and NaOCl uptake
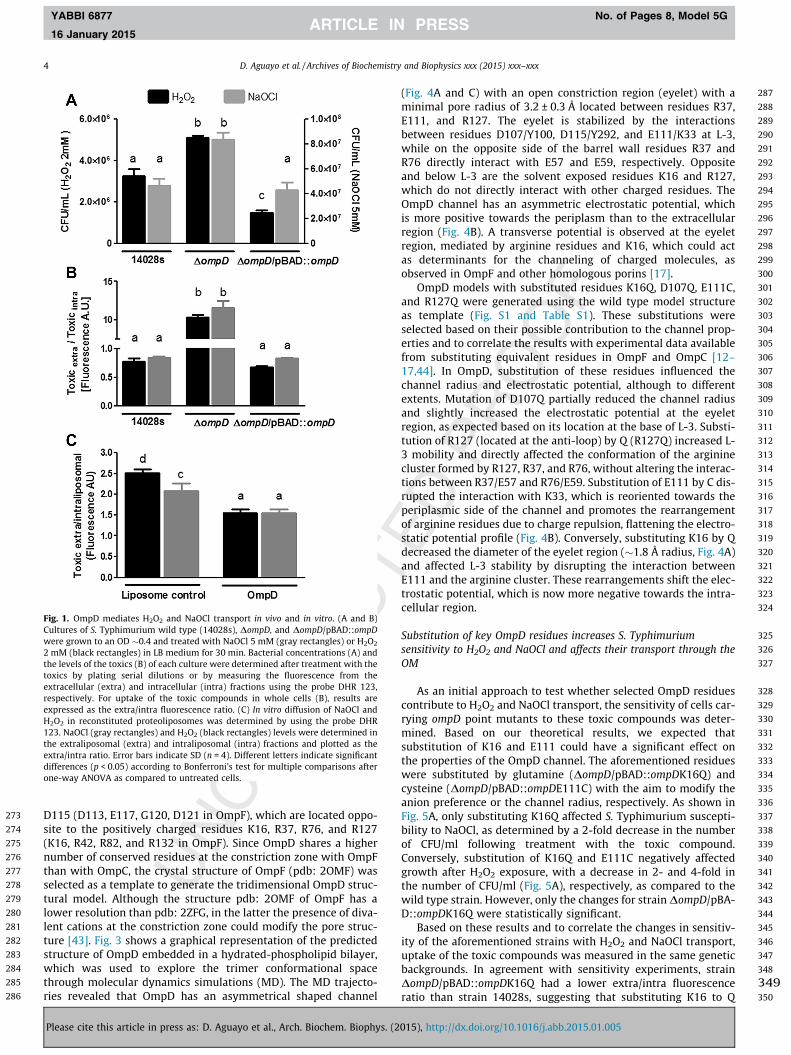
It has been shown that OmpD mediates H2O2 uptake [4], how-ever, its role in NaOCl transport has not been investigated. Toaddress this, we first tested the sensitivity of a DompD strain toH2O2 (control) and NaOCl, and compared it to its isogenic wild typestrain. Supporting a role for OmpD in NaOCl transport, inactivationof ompD increased S. Typhimurium resistance to NaOCl (2-fold), asdetermined by colony forming units (Fig. 1A). A similar result wasobserved when cells were challenged with H2O2 (Fig. 1A), as previ-ously reported [4]. To directly test if NaOCl is transported throughOmpD, whole cells and proteoliposomes with purified OmpD wereused to measure the uptake of the toxic compounds. ompD deletiondecreased H2O2 and NaOCl uptake in whole cells (�15-fold), ascompared to the wild type strain (Fig. 1B), and OmpD-proteolipo-somes incubated with H2O2 or NaOCl had a lower extra-/intra-lipo-somal fluorescence ratio than liposomes alone (Fig. 1C), indicatingincreased transport of both toxicants in the presence of OmpD.Taken together, the genetic and biochemical evidence indicatesthat OmpD mediates H2O2 and NaOCl uptake.
Alignments, molecular simulations and pore properties
To investigate the mechanism of H2O2 and NaOCl transportthrough OmpD, a structural model was first generated. To identifythe best structure to be used as a template, the OmpD sequencewas aligned to that of porins having a reported structure in theprotein data bank. The alignment showed that the closest OmpDhomologues were K. Pneumoniae OmpK36, and E. coli OmpC andOmpF (Fig. 2). OmpD had an overall sequence identity of 64% withthe aforementioned porins. Based on the alignment of the second-ary structural elements [42], the low-conserved regions of OmpDare mainly located in loops, with large insertions facing the extra-cellular regions. The PEFGGD porin-specific motif and inner loop(L-3) residues are conserved, including D107, E111, G113 and
015), http://dx.doi.org/10.1016/j.abb.2015.01.005
273
274
275
276
277
278
279
280
281
282
283
284
285
286
287
288
289
290
291
292
293
294
295
296
297
298
299
300
301
302
303
304
305
306
307
308
309
310
311
312
313
314
315
316
317
318
319
320
321
322
323
324
325
326
327
328
329
330
331
332
333
334
335
336
337
338
339
340
341
342
343
344
345
346
347
348
349350
Fig. 1. OmpD mediates H2O2 and NaOCl transport in vivo and in vitro. (A and B)Cultures of S. Typhimurium wild type (14028s), DompD, and DompD/pBAD::ompDwere grown to an OD �0.4 and treated with NaOCl 5 mM (gray rectangles) or H2O2
2 mM (black rectangles) in LB medium for 30 min. Bacterial concentrations (A) andthe levels of the toxics (B) of each culture were determined after treatment with thetoxics by plating serial dilutions or by measuring the fluorescence from theextracellular (extra) and intracellular (intra) fractions using the probe DHR 123,respectively. For uptake of the toxic compounds in whole cells (B), results areexpressed as the extra/intra fluorescence ratio. (C) In vitro diffusion of NaOCl andH2O2 in reconstituted proteoliposomes was determined by using the probe DHR123. NaOCl (gray rectangles) and H2O2 (black rectangles) levels were determined inthe extraliposomal (extra) and intraliposomal (intra) fractions and plotted as theextra/intra ratio. Error bars indicate SD (n = 4). Different letters indicate significantdifferences (p < 0.05) according to Bonferroni’s test for multiple comparisons afterone-way ANOVA as compared to untreated cells.
4 D. Aguayo et al. / Archives of Biochemistry and Biophysics xxx (2015) xxx–xxx
YABBI 6877 No. of Pages 8, Model 5G
16 January 2015
D115 (D113, E117, G120, D121 in OmpF), which are located oppo-site to the positively charged residues K16, R37, R76, and R127(K16, R42, R82, and R132 in OmpF). Since OmpD shares a highernumber of conserved residues at the constriction zone with OmpFthan with OmpC, the crystal structure of OmpF (pdb: 2OMF) wasselected as a template to generate the tridimensional OmpD struc-tural model. Although the structure pdb: 2OMF of OmpF has alower resolution than pdb: 2ZFG, in the latter the presence of diva-lent cations at the constriction zone could modify the pore struc-ture [43]. Fig. 3 shows a graphical representation of the predictedstructure of OmpD embedded in a hydrated-phospholipid bilayer,which was used to explore the trimer conformational spacethrough molecular dynamics simulations (MD). The MD trajecto-ries revealed that OmpD has an asymmetrical shaped channel
Please cite this article in press as: D. Aguayo et al., Arch. Biochem. Biophys. (2
(Fig. 4A and C) with an open constriction region (eyelet) with aminimal pore radius of 3.2 ± 0.3 Å located between residues R37,E111, and R127. The eyelet is stabilized by the interactionsbetween residues D107/Y100, D115/Y292, and E111/K33 at L-3,while on the opposite side of the barrel wall residues R37 andR76 directly interact with E57 and E59, respectively. Oppositeand below L-3 are the solvent exposed residues K16 and R127,which do not directly interact with other charged residues. TheOmpD channel has an asymmetric electrostatic potential, whichis more positive towards the periplasm than to the extracellularregion (Fig. 4B). A transverse potential is observed at the eyeletregion, mediated by arginine residues and K16, which could actas determinants for the channeling of charged molecules, asobserved in OmpF and other homologous porins [17].
OmpD models with substituted residues K16Q, D107Q, E111C,and R127Q were generated using the wild type model structureas template (Fig. S1 and Table S1). These substitutions wereselected based on their possible contribution to the channel prop-erties and to correlate the results with experimental data availablefrom substituting equivalent residues in OmpF and OmpC [12–17,44]. In OmpD, substitution of these residues influenced thechannel radius and electrostatic potential, although to differentextents. Mutation of D107Q partially reduced the channel radiusand slightly increased the electrostatic potential at the eyeletregion, as expected based on its location at the base of L-3. Substi-tution of R127 (located at the anti-loop) by Q (R127Q) increased L-3 mobility and directly affected the conformation of the argininecluster formed by R127, R37, and R76, without altering the interac-tions between R37/E57 and R76/E59. Substitution of E111 by C dis-rupted the interaction with K33, which is reoriented towards theperiplasmic side of the channel and promotes the rearrangementof arginine residues due to charge repulsion, flattening the electro-static potential profile (Fig. 4B). Conversely, substituting K16 by Qdecreased the diameter of the eyelet region (�1.8 Å radius, Fig. 4A)and affected L-3 stability by disrupting the interaction betweenE111 and the arginine cluster. These rearrangements shift the elec-trostatic potential, which is now more negative towards the intra-cellular region.
Substitution of key OmpD residues increases S. Typhimuriumsensitivity to H2O2 and NaOCl and affects their transport through theOM
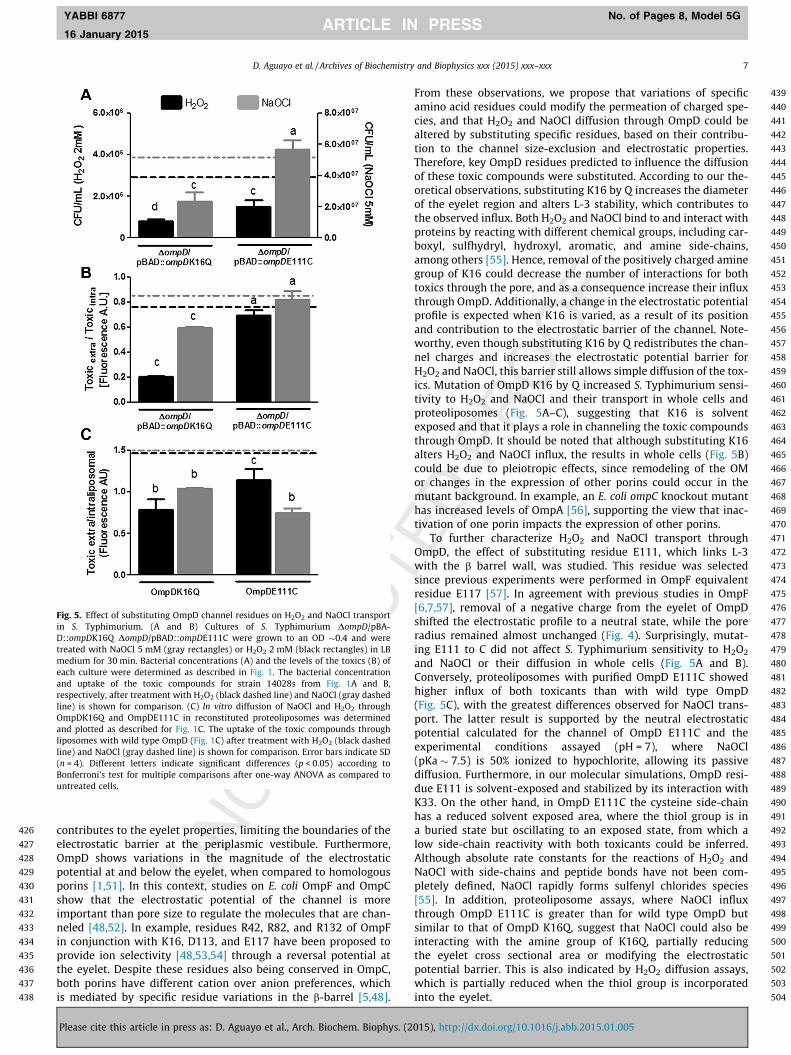
As an initial approach to test whether selected OmpD residuescontribute to H2O2 and NaOCl transport, the sensitivity of cells car-rying ompD point mutants to these toxic compounds was deter-mined. Based on our theoretical results, we expected thatsubstitution of K16 and E111 could have a significant effect onthe properties of the OmpD channel. The aforementioned residueswere substituted by glutamine (DompD/pBAD::ompDK16Q) andcysteine (DompD/pBAD::ompDE111C) with the aim to modify theanion preference or the channel radius, respectively. As shown inFig. 5A, only substituting K16Q affected S. Typhimurium suscepti-bility to NaOCl, as determined by a 2-fold decrease in the numberof CFU/ml following treatment with the toxic compound.Conversely, substitution of K16Q and E111C negatively affectedgrowth after H2O2 exposure, with a decrease in 2- and 4-fold inthe number of CFU/ml (Fig. 5A), respectively, as compared to thewild type strain. However, only the changes for strain DompD/pBA-D::ompDK16Q were statistically significant.
Based on these results and to correlate the changes in sensitiv-ity of the aforementioned strains with H2O2 and NaOCl transport,uptake of the toxic compounds was measured in the same geneticbackgrounds. In agreement with sensitivity experiments, strainDompD/pBAD::ompDK16Q had a lower extra/intra fluorescenceratio than strain 14028s, suggesting that substituting K16 to Q
015), http://dx.doi.org/10.1016/j.abb.2015.01.005

351
352
353
354
355
356
357
358
359
360
361
362
363
364
365
366
367
368
369
370
371
372
373
374
375
376
377
378
379
380
381
382
383
384
385
Fig. 2. Alignment of the mature amino acid sequence of OmpD with the homologous porins OmpF, OmpC, and OmpK36. The structural features of OmpD are indicated abovethe alignment.
D. Aguayo et al. / Archives of Biochemistry and Biophysics xxx (2015) xxx–xxx 5
YABBI 6877 No. of Pages 8, Model 5G
16 January 2015
increases H2O2 and NaOCl uptake through OmpD (Fig. 5B). Surpris-ingly, and despite being more sensitive to H2O2 and NaOCl(Fig. 5A), strain DompD/pBAD::ompDE111C showed similar uptakelevels of the toxic compounds than strain 14028s (Fig. 5B).
Substitution of key OmpD residues affects H2O2 and NaOCl transportthrough proteoliposomes
In vitro functional assays using reconstituted proteoliposomeswere carried out to demonstrate that residues K16Q and E111Care implicated in H2O2 and NaOCl uptake. For this purpose, oxida-tion of DHR123 was measured in proteoliposomes with purifiedOmpD, OmpD K16Q, or OmpD E111C, after incubation with H2O2
or NaOCl. After incubation with both toxics independently, theextra-/intra-liposomal fluorescence ratio was higher in proteolipo-somes with OmpD K16Q and OmpD E111C than with wild typeOmpD (Fig. 5C). This indicates that mutation of residues K16 andE111 increased H2O2 and NaOCl diffusion through OmpD, and thatthese residues are implicated, at least in part, in their transport.
Please cite this article in press as: D. Aguayo et al., Arch. Biochem. Biophys. (2
Discussion
Hydrogen peroxide and NaOCl are reactive oxygen species(ROS) generated by phagocytic cells that damage proteins, lipids,and nucleic acids, among other targets [45,46]. The concentrationof ROS inside the bacterium is tightly controlled by several mech-anisms, including ROS diffusion across membranes, its scavenging,and influx into the cytoplasm [47]. Under physiological conditionsH2O2 is stable and has a molecular radius of about 1.2–1.4 Å, it isnot charged but polar with a dipole moment of 2.26 � 10�18 elec-trostatic units (eu), similar to that of water (1.85 � 10�18 eu) [47].For NaOCl, at pH = 7 it is 50% ionized (pKa � 7.5) to hypochlorite,and therefore it is negatively charged [19]. Since lipid membranesare only permeable to small and non-polar molecules, H2O2 andNaOCl influx relies on the composition of the membrane and onthe presence of protein channels, including porins. Supporting thishypothesis, a recent study and the results presented herein showthat S. Typhimurium lacking OmpD is more resistant than the wildtype strain to H2O2 [4] and NaOCl and incorporates less of the toxic
015), http://dx.doi.org/10.1016/j.abb.2015.01.005

386
387
388
389
390
391
392
393
394
395
396
397
398
399
400
401
402
403
404
405
406
407
408
409
410
411
412
413
414
415
416
417
418
419
420
421
422
423
424
425
Fig. 3. Molecular structure of an OmpD trimer embedded into a lipid bilayer.Snapshot of a MD simulation of the OmpD channel embedded in a lipid bilayer. Toallow visualization of the L-3 inner loop and key OmpD residues, the front of theporin is transparent. Ions, water molecules and some lipids are not shown. The poresurface is depicted as a yellow surface. Residues K16 and E111 are shown in blueand red, respectively. (For interpretation of the references to color in this figurelegend, the reader is referred to the web version of this article.)
6 D. Aguayo et al. / Archives of Biochemistry and Biophysics xxx (2015) xxx–xxx
YABBI 6877 No. of Pages 8, Model 5G
16 January 2015
compounds (Fig. 1A and B). Moreover, proteoliposomes withOmpD show increased H2O2 and NaOCl uptake (Fig. 1C), indicatingthat both ROS are channeled through this porin.
There is growing evidence for the role of porins in ROS uptake[4,11]; however, the underlying transport mechanism(s) is poorlyunderstood and further studies are warranted. To investigate thisprocess, the effect of substituting key OmpD residues on H2O2
Fig. 4. In silico modification of key OmpD residues and their effect on the properties ofOmpD, OmpD K16Q, and E111C were calculated using the HOLE algorithm and Poisson–and the molecular surface in gray. Pink spheres represent the pore axis. Lateral (D) and suare shown. (For interpretation of the references to color in this figure legend, the reade
Please cite this article in press as: D. Aguayo et al., Arch. Biochem. Biophys. (2
and NaOCl uptake was studied. We combined sequence-basedalignment, homology modeling, and molecular dynamics simula-tions to identify key residues that could play different roles inthe channel properties, and therefore influence diffusion of the tox-ics through OmpD.
General diffusion porins have different cation vs. anion prefer-ences, which depends on the spatial arrangement of specificresidues inside the b barrel [48]. In a recent study [49], by usingsequence-based and similarity networks and analysis of the calcu-lated surface properties, we observed that OmpD resembles anintermediate state between the closer homologous OmpF andOmpC from E. coli and OmpK36 from K. pneumoniae. Extendingour theoretical work, and considering that diffusion-driving prop-erties depend on atomic details [50], MD was conducted to obtaina conformational ensemble from which the channel propertiescould be averaged. First, and since the atomic structure of OmpDis not available, a three-dimensional model was generated and fur-ther energy minimized (Fig. 3, see Methods). Analyses of moleculardynamic trajectories of the predicted structure of an OmpD trimer(Fig. 4) shows that conserved residues in OmpD located at theeyelet region (D107, E111, G113 and R37) and the anti-loop (R76and R127) stabilize and control the pore size, that is wide enough(�3 Å) to allow H2O2 and NaOCl influx through the porin (Fig. 4). InOmpD, the region below L-3 is not conserved (Y in OmpD vs W inOmpF [42]). The N-terminal region has variations near residue K16(Figs. 3 and 4), and different experiments have demonstrated thatvariations in this region of the channel alter its charge preference.The negative electrostatic potential observed along the pore axisallows inferring a cation over anion preference; however, the smalldifference with respect to the bulk (�2 kBT/e) suggests that nega-tively charged molecules could easily overcome this electrostaticbarrier. Residues E111, R37, R76, and R127 generate a transversalelectric field in the middle of the channel. Residue K16 also
the channel. The pore radius (A) and electrostatic potential (B) profiles of wild typeBoltzmann equation, respectively. (C) The pore surface of OmpD is depicted in blueperior (E) view of the OmpD channel. Residues K16 at the barrel wall and E111 at L-3r is referred to the web version of this article.)
015), http://dx.doi.org/10.1016/j.abb.2015.01.005
426
427
428
429
430
431
432
433
434
435
436
437
438
439
440
441
442
443
444
445
446
447
448
449
450
451
452
453
454
455
456
457
458
459
460
461
462
463
464
465
466
467
468
469
470
471
472
473
474
475
476
477
478
479
480
481
482
483
484
485
486
487
488
489
490
491
492
493
494
495
496
497
498
499
500
501
502
503
504
Fig. 5. Effect of substituting OmpD channel residues on H2O2 and NaOCl transportin S. Typhimurium. (A and B) Cultures of S. Typhimurium DompD/pBA-D::ompDK16Q DompD/pBAD::ompDE111C were grown to an OD �0.4 and weretreated with NaOCl 5 mM (gray rectangles) or H2O2 2 mM (black rectangles) in LBmedium for 30 min. Bacterial concentrations (A) and the levels of the toxics (B) ofeach culture were determined as described in Fig. 1. The bacterial concentrationand uptake of the toxic compounds for strain 14028s from Fig. 1A and B,respectively, after treatment with H2O2 (black dashed line) and NaOCl (gray dashedline) is shown for comparison. (C) In vitro diffusion of NaOCl and H2O2 throughOmpDK16Q and OmpDE111C in reconstituted proteoliposomes was determinedand plotted as described for Fig. 1C. The uptake of the toxic compounds throughliposomes with wild type OmpD (Fig. 1C) after treatment with H2O2 (black dashedline) and NaOCl (gray dashed line) is shown for comparison. Error bars indicate SD(n = 4). Different letters indicate significant differences (p < 0.05) according toBonferroni’s test for multiple comparisons after one-way ANOVA as compared tountreated cells.
D. Aguayo et al. / Archives of Biochemistry and Biophysics xxx (2015) xxx–xxx 7
YABBI 6877 No. of Pages 8, Model 5G
16 January 2015
contributes to the eyelet properties, limiting the boundaries of theelectrostatic barrier at the periplasmic vestibule. Furthermore,OmpD shows variations in the magnitude of the electrostaticpotential at and below the eyelet, when compared to homologousporins [1,51]. In this context, studies on E. coli OmpF and OmpCshow that the electrostatic potential of the channel is moreimportant than pore size to regulate the molecules that are chan-neled [48,52]. In example, residues R42, R82, and R132 of OmpFin conjunction with K16, D113, and E117 have been proposed toprovide ion selectivity [48,53,54] through a reversal potential atthe eyelet. Despite these residues also being conserved in OmpC,both porins have different cation over anion preferences, whichis mediated by specific residue variations in the b-barrel [5,48].
Please cite this article in press as: D. Aguayo et al., Arch. Biochem. Biophys. (2
From these observations, we propose that variations of specificamino acid residues could modify the permeation of charged spe-cies, and that H2O2 and NaOCl diffusion through OmpD could bealtered by substituting specific residues, based on their contribu-tion to the channel size-exclusion and electrostatic properties.Therefore, key OmpD residues predicted to influence the diffusionof these toxic compounds were substituted. According to our the-oretical observations, substituting K16 by Q increases the diameterof the eyelet region and alters L-3 stability, which contributes tothe observed influx. Both H2O2 and NaOCl bind to and interact withproteins by reacting with different chemical groups, including car-boxyl, sulfhydryl, hydroxyl, aromatic, and amine side-chains,among others [55]. Hence, removal of the positively charged aminegroup of K16 could decrease the number of interactions for bothtoxics through the pore, and as a consequence increase their influxthrough OmpD. Additionally, a change in the electrostatic potentialprofile is expected when K16 is varied, as a result of its positionand contribution to the electrostatic barrier of the channel. Note-worthy, even though substituting K16 by Q redistributes the chan-nel charges and increases the electrostatic potential barrier forH2O2 and NaOCl, this barrier still allows simple diffusion of the tox-ics. Mutation of OmpD K16 by Q increased S. Typhimurium sensi-tivity to H2O2 and NaOCl and their transport in whole cells andproteoliposomes (Fig. 5A–C), suggesting that K16 is solventexposed and that it plays a role in channeling the toxic compoundsthrough OmpD. It should be noted that although substituting K16alters H2O2 and NaOCl influx, the results in whole cells (Fig. 5B)could be due to pleiotropic effects, since remodeling of the OMor changes in the expression of other porins could occur in themutant background. In example, an E. coli ompC knockout mutanthas increased levels of OmpA [56], supporting the view that inac-tivation of one porin impacts the expression of other porins.
To further characterize H2O2 and NaOCl transport throughOmpD, the effect of substituting residue E111, which links L-3with the b barrel wall, was studied. This residue was selectedsince previous experiments were performed in OmpF equivalentresidue E117 [57]. In agreement with previous studies in OmpF[6,7,57], removal of a negative charge from the eyelet of OmpDshifted the electrostatic profile to a neutral state, while the poreradius remained almost unchanged (Fig. 4). Surprisingly, mutat-ing E111 to C did not affect S. Typhimurium sensitivity to H2O2
and NaOCl or their diffusion in whole cells (Fig. 5A and B).Conversely, proteoliposomes with purified OmpD E111C showedhigher influx of both toxicants than with wild type OmpD(Fig. 5C), with the greatest differences observed for NaOCl trans-port. The latter result is supported by the neutral electrostaticpotential calculated for the channel of OmpD E111C and theexperimental conditions assayed (pH = 7), where NaOCl(pKa � 7.5) is 50% ionized to hypochlorite, allowing its passivediffusion. Furthermore, in our molecular simulations, OmpD resi-due E111 is solvent-exposed and stabilized by its interaction withK33. On the other hand, in OmpD E111C the cysteine side-chainhas a reduced solvent exposed area, where the thiol group is ina buried state but oscillating to an exposed state, from which alow side-chain reactivity with both toxicants could be inferred.Although absolute rate constants for the reactions of H2O2 andNaOCl with side-chains and peptide bonds have not been com-pletely defined, NaOCl rapidly forms sulfenyl chlorides species[55]. In addition, proteoliposome assays, where NaOCl influxthrough OmpD E111C is greater than for wild type OmpD butsimilar to that of OmpD K16Q, suggest that NaOCl could also beinteracting with the amine group of K16Q, partially reducingthe eyelet cross sectional area or modifying the electrostaticpotential barrier. This is also indicated by H2O2 diffusion assays,which is partially reduced when the thiol group is incorporatedinto the eyelet.
015), http://dx.doi.org/10.1016/j.abb.2015.01.005

505
506
507
508
509
510
511
512
513
514
515
516
517
518
519
520 Q7
521
522 Q8
523
524
525
526
527528529530531532533534535536537538539540541542543544545546547548549550551552553554
555556557558559560561562563564565566567568569570571572573574575576577578579580581582583584585586587588589590591592593594595596597598599600601602603604605606607608609610611612613614615616617618619620
8 D. Aguayo et al. / Archives of Biochemistry and Biophysics xxx (2015) xxx–xxx
YABBI 6877 No. of Pages 8, Model 5G
16 January 2015
Conclusions
Together, and for the first time, we provide both genetic andbiochemical evidence that OmpD mediates H2O2 and NaOCl uptakeand that key residues of the channel are implicated in this process.Furthermore, our theoretical results suggest that diffusion of bothtoxic compounds through OmpD depends on the overlap of specificdeterminants in the pore radius and electrostatic potential. Newtheoretical and functional assays are currently underway todemonstrate the proposed role of the amino acid side-chain reac-tivity on the diffusion of the toxicants through the porin channel.It is expected that in conjunction with the experimental and theo-retical observations reported herein, the new assays will providefurther insights into the physiological aspects and the role ofOmpD in the response to ROS by the bacterium.
Acknowledgments
This work was supported in part by FONDECYT grants CS#1120384 and DA #11130576 and Millenium Initiative 09-022-F(FG).
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.abb.2015.01.005.
References
[1] H. Nikaido, Microbiol. Mol. Biol. Rev. 67 (2003) 593–656.[2] A. Delcour, Front. Biosci. 8 (2003) d1055–71.[3] C. Santiviago, J. Fuentes, S. Bueno, A. Trombert, A. Hildago, L. Socias, P.
Youderian, G. Mora, Mol. Microbiol. 46 (2002) 687–698.[4] I. Calderón, E. Morales, N. Caro, C. Chahúan, B. Collao, F. Gil, J.M. Villarreal, F.
Ipinza, G. Mora, C. Saavedra, Res. Microbiol. 162 (2011) 214–222.[5] B.K. Ziervogel, B. Roux, Structure 21 (2013) 76–87.[6] W. Achouak, T. Heulin, J.M. Pages, FEMS Microbiol. Lett. 199 (2001) 1–7.[7] A. Alcaraz, E.M. Nestorovich, M. Aguilella-Arzo, V.M. Aguilella, S.M. Bezrukov,
Biophys. J. 87 (2004) 943–957.[8] E. Yamashita, M. Zhalnina, S. Zakharov, O. Sharma, W. Cramer, EMBO J. 27
(2008) 2171–2180.[9] W. Im, B. Roux, J. Mol. Biol. 319 (2002) 1177–1197.
[10] A. Doménech-Sánchez, S. Hernández-Allés, L. Martínez-Martínez, V.J. Benedí, S.Albertí, J. Bacteriol. 181 (1999) 2726–2732.
[11] E. Morales, I. Calderón, B. Collao, F. Gil, S. Porwollik, M. McClelland, C.Saavedra, BMC Microbiol. 12 (2012) 63.
[12] N. Saint, K. Lou, C. Widmer, M. Luckey, T. Schirmer, J. Rosenbusch, J. Biol. Chem.271 (1996) 20676–20680.
[13] D. Jeanteur, T. Schirmer, D. Fourel, V. Simonet, G. Rummel, C. Widmer, J.Rosenbusch, F. Pattus, J.-M. Pagès, Proc. Natl. Acad. Sci. U.S.A. 91 (1994)10675–10679.
[14] V. Simonet, M. Mallea, J.M. Pages, Antimicrob. Agents Chemother. 44 (2000)311–315.
[15] P. Phale, A. Philippsen, C. Widmer, V.P. Phale, J. Rosenbusch, T. Schirmer,Biochemistry 40 (2001) 6319–6325.
[16] J. Bredin, N. Saint, M. Malléa, E. Dé, G. Molle, J.-M. Pagès, V. Simonet, Biochem.J. 363 (2002) 521–528.
621
Please cite this article in press as: D. Aguayo et al., Arch. Biochem. Biophys. (2
[17] M. Masi, J.-M. Pagès, Open Microbiol. J. 7 (2013) 22–33.[18] C. Santiviago, C. Toro, A. Hidalgo, P. Youderian, G. Mora, J. Bacteriol. 185 (2003)
5901–5905.[19] C. Winterbourn, A. Kettle, Antioxid. Redox Signal. 18 (2013) 642–660.[20] S. Eriksson, S. Lucchini, A. Thompson, M. Rhen, J.C.D. Hinton, Mol. Microbiol. 47
(2003) 103–118.[21] S. Cowan, R. Garavito, J. Jansonius, J. Jenkins, R. Karlsson, N. König, E. Pai, R.
Pauptit, P. Rizkallah, J. Rosenbusch, G. Rummel, T. Schirmer, Structure 3 (1995)1041–1050.
[22] A. Baslé, G. Rummel, P. Storici, J. Rosenbusch, T. Schirmer, J. Mol. Biol. 362(2006) 933–942.
[23] T. Surrey, F. Jahnig, Proc. Natl. Acad. Sci. U.S.A. 89 (1992) 7457–7461.[24] A. Miles, S. Misra, J. Irwin, J. Hyg. (Lond.) 38 (1938) 732–749.[25] S. Hempel, G. Buettner, Y. O’Malley, D. Wessels, D. Flaherty, Free Radical Biol.
Med. 27 (1999) 146–159.[26] H. Nikaido, E. Rosenberg, J. Foulds, J. Bacteriol. 153 (1983) 232–240.[27] M. Cubillos, E. Lissi, E. Abuin, Chem. Phys. Lipids 112 (2001) 41–46.[28] F. Bernstein, T. Koetzle, G. Williams, E. Meyer, Eur. J. Biochem. 80 (1977) 319–
324.[29] B. Webb, A. Sali, Methods in Molecular Biology, Springer, New York, NY, 2014.[30] J. Huang, A. MacKerell Jr., J. Comput. Chem. 34 (2013) 2135–2145.[31] A. MacKerell Jr., J. Comput. Chem. 25 (2004) 1584–1604.[32] J. Phillips, R. Braun, W. Wang, J. Gumbart, E. Tajkhorshid, E. Villa, C. Chipot, R.
Skeel, L. Kale, K. Schulten, J. Comput. Chem. 26 (2005) 1781–1802.[33] S. McDonald, D. Aguayo, F.D. Gonzalez-Nilo, J.T. Patton, J. Virol. 83 (2009)
6135–6148.[34] M. Olsson, C. Søndergaard, M. Rostkowski, J. Jensen, J. Chem. Theory Comput. 7
(2011) 525–537.[35] T. Darden, D. York, L. Pedersen, J. Chem. Phys. 98 (1993) 10089.[36] S. Feller, Y. Zhang, R. Pastor, B. Brooks, J. Chem. Phys. 103 (1995) 4613–4621.[37] M. Tuckerman, B. Berne, A. Rossi, J. Chem. Phys. 94 (1991) 1465–1469.[38] W. Jorgensen, J. Chandrasekhar, J. Madura, R. Impey, M. Klein, J. Chem. Phys. 79
(1983) 926.[39] N. Baker, D. Sept, S. Joseph, M. Holst, J. McCammon, Proc. Natl. Acad. Sci. U.S.A.
98 (2001) 10037–10041.[40] O. Smart, J. Neduvelil, X. Wang, B. Wallace, M. Sansom, J. Mol. Graph. 14
(1996). 354–60, 376.[41] W. Humphrey, A. Dalke, K. Schulten, J. Mol. Graph. 14 (1996) 33–38.[42] A. Basle, G. Rummel, P. Storici, J. Rosenbusch, T. Schirmer, J. Mol. Biol. 362
(2006) 933–942.[43] E. García-Giménez, A. Alcaraz, V.M. Aguilella, Biochem. Res. Int. 2012 (2012)
1–12.[44] H. Lou, M. Chen, S. Black, S. Bushell, M. Ceccarelli, T. Mach, K. Beis, A. Low, V.
Bamford, I. Booth, H. Bayley, J.H. Naismith, PLoS ONE 6 (2011) e25825.[45] D. Hassett, M. Cohen, FASEB J. 3 (1989) 2574–2582.[46] J. Imlay, Annu. Rev. Biochem. 77 (2008) 755–776.[47] G. Bienert, J. Schjoerring, T. Jahn, Biochim. Biophys. Acta 1758 (2006) 994–
1003.[48] V. Aguilella, M. Queralt-Martín, M. Aguilella-Arzo, A. Alcaraz, Integr. Biol. 3
(2011) 159.[49] F. Ipinza, B. Collao, D. Monsalva, V.H. Bustamante, R. Luraschi, M. Alegría-
Arcos, D. Almonacid, D. Aguayo, I. Calderón, F. Gil, C.A. Santiviago, E.H. Morales,E. Calva, C. Saavedra, PLoS ONE 9 (2014) e111062.
[50] A. Aksimentiev, K. Schulten, Biophys. J. 88 (2005) 3745–3761.[51] M. Aguilella-Arzo, A. Andrio, V.M. Aguilella, A. Alcaraz, Phys. Chem. Chem.
Phys. 11 (2009) 358.[52] J. Wibbenmeyer, D. Provenzano, C. Landry, K. Klose, A. Delcour, Infect. Immun.
70 (2002) 121–126.[53] H. Schweizer, Expert Opin. Drug Discov. 7 (2012) 633–642.[54] H. Miedema, M. Vrouenraets, J. Wierenga, W. Meijberg, G. Robillard, B.
Eisenberg, Nano Lett. 7 (2007) 2886–2891.[55] O. Panasenko, I.V. Gorudko, A.V. Sokolov, Biochem. (Mosc.) 78 (2014) 1466–
1489.[56] M. De la Cruz, E. Calva, J. Mol. Microbiol. Biotechnol. 18 (2010) 24–36.[57] M. Vrouenraets, J. Wierenga, W. Meijberg, H. Miedema, Biophys. J. 90 (2006)
1202–1211.
015), http://dx.doi.org/10.1016/j.abb.2015.01.005




















