
Mémoire de MAGISTER Miloud BENAISSA - Theses
94
Université d’ORAN Faculté des Sciences Département de Biotechnologie Mémoire de MAGISTER en Biotechnologie Option : Ecosystèmes microbiens complexes Présentée par Miloud BENAISSA Intitulée Devant le Jury : Président : Pr. N-E. KARAM Université d’Oran Examinateurs : Dr. B. DJEGHRI Université d’Annaba Dr. F. DALACHE Université de Mostaganem Rapporteur : Pr. H. ZADI- KARAM Université d’Oran Soutenue le : / / 2010 ELABORATION D’UN MILIEU DE CULTURE A BASE DE LACTOSERUM DOUX POUR LACTOBACILLUS
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Mémoire de MAGISTER Miloud BENAISSA - Theses

Université d’ORAN Faculté des Sciences
Département de Biotechnologie
Mémoire de MAGISTER
en Biotechnologie Option : Ecosystèmes microbiens complexes
Présentée par
Miloud BENAISSA Intitulée
Devant le Jury :
Président : Pr. N-E. KARAM Université d’Oran Examinateurs : Dr. B. DJEGHRI Université d’Annaba
Dr. F. DALACHE Université de Mostaganem Rapporteur : Pr. H. ZADI- KARAM Université d’Oran
Soutenue le : / / 2010
ELABORATION D’UN MILIEU DE CULTURE
A BASE DE LACTOSERUM DOUX POUR LACTOBACILLUS


Remerciements
Je remercie en premier lieu le Créateur des cieux et des terres, notre Grand Dieu Tout Puissant qui par Sa volonté nous a permis d’affronter les difficultés rencontrées et aboutir à la réalisation de ce travail.
Je tiens à exprimer ma profonde reconnaissance à Mme Zadi-Karam H. sans qui ce travail n’aurait sans doute jamais abouti. Merci pour vos précieux conseils, la confiance et le temps que vous m’avez accordé malgré vos nombreuses occupations. J’ai particulièrement apprécié votre rigueur et votre passion dans votre métier de chercheur. Votre gentillesse et vos encouragements ont été une des clés de la construction de ce mémoire. Je ne l’oublierai pas.
Un profond respect et un merci tout spécial à Mr. Karam N. pour son soutien permanent, sa confiance, son esprit scientifique, sa patience. Merci aussi d’avoir accepté de présider le jury de mon travail.
J’adresse également mes remerciements à Mme Djeghri-Hocine B et Mlle Dalache F. pour m’avoir fait l’honneur de juger ce travail.
Je tiens également à remercier les membres de l’équipe des bactéries lactiques qui ont allumé ma petite flamme pour la recherche.
Mes remerciements vont aussi aux membres du Centre Algérien du Contrôle de la Qualité et de l'Emballage (CACQE) d’Oran qui m’ont toujours chaleureusement accueilli. Un grand merci à Mme Reguieg la directrice du laboratoire d’analyse de la qualité EL-FETH, qui m’a permis de réaliser des analyses très utiles à ce travail.
Rien n’aurait été pareil sans la présence indéfectible et inestimable de Amel, Khalida et Amina, je leur dois bien plus qu’un merci. Cette aventure aurait probablement été moins belle sans les moments partagés avec Mustapha, Mohamed, Amaria, Asmaa et Halima.
Enfin, que tous ceux qui m’ont ouvert leur porte et offert leur amitié soient assurés de ma reconnaissance.

Dédicaces
« Comment définir ce que l’on veut infinir,… » Ocean, Jean-Louis Aubert
Je dédie cet événement marquant de ma vie à :
A ma mère, à mon adorable mère, à celle qui est toujours présente et continue de l’être pour faire mon bonheur. Merci pour t’être sacrifiée pour que tes enfants grandissent et prospèrent. Merci de trimer sans relâche au bien-être de tes enfants. Je ne sais ce que serait ma vie sans toi, t’avoir à mes côtés vaut pour moi tout l’or du monde, et toutes les joies de cette vie. Enfin ! Merci tout simplement d’être ma mère.
Que Dieu te protège et te prête bonne santé et longue vie. A mes chers frères et leurs petites familles. Rien n’aurait été pareil sans votre présence indéfectible et inestimable. Je voudrais tous vous remercier pour l’attention que vous m’avez apportée. Merci pour votre soutien, votre aide de tous les instants, votre amour et votre stimulante fierté. Les mots sont faibles pour exprimer la force de mes sentiments et la reconnaissance que je vous porte.
A ma sœur chérie, à l’unique sœur que j’ai au monde. Merci d’être toujours à mes côtés, par ta présence, par ton amour, pour donner du goût et du sens à notre vie de famille.
A mes adorables nièces. Merci de remplir ma vie de joie et de bonheur. A mes petits oursons (mes neveux) qui illuminent mes journées en toutes circonstances.
Enfin, j’espère du fond du cœur que tout ce petit monde, mon monde à moi, trouve ici un mot de reconnaissance, et que chacun se reconnaisse en ce qui le concerne. J’espère aussi que l’effort déployé dans le présent travail répond aux attentes des uns et des autres.

SOMMAIRE Pages
Liste des figures i Liste des tableaux ii 1 – INTRODUCTION 1
1-1 – Lactosérum : 4 1-1-2- Valorisation du lactosérum : 5 1-1-3- Utilisation du lactose : 8 1-1-4- Utilisation des protéines : 8* 1-1-5- Fermentations diverses du lactosérum : 9 1-2- Les lactobacilles : 11 1-2-1-Taxonomie et classification des lactobacilles : 11 1-2-2- Exigences nutritionnelles des lactobacilles : 12 1-3- Métabolisme des lactobacilles : 17 1-3-1- Métabolisme glucidique : 17 1-3-1-1- Systèmes de transport actif : 17 1-3-1-2- Systèmes PEP-PT : 17 1-3-1-3- Métabolisme des sucres : 21 1-3-1-4- Régulation du métabolisme des sucres : 24 1-3-1-5- Régulation du système de transport des sucres : 24 1-3-2- Métabolisme du citrate : 27 1-3-3- Métabolisme azoté : 28
2 - MATERIEL & METHODES 30
2-1- Matériel : 31 2-1-1- Souches bactériennes : 31 2-1-2- Lactosérum : 31 2 -2- Méthodes : 32 2-2-1-Vérification de la pureté et l’appartenance des souches au groupe lactique : 32 2-2-2- Conservation des bactéries : 32 2-2-3- Déprotéinisation du lactosérum : 32 2-2-4- Préparation du jus de tomate : 32 2-2-5- Analyses physicochimiques : 32 2-2-5-1- Mesure de l’acidité : 33 2-2-5-2- Teneur en matière sèche totale : 33 2-2-5-3- Teneur en cendres : 33 2-2-5-4- Détermination de la matière grasse : 34 2-2-5-5-Teneur en glucides : 34 2-2-6- Optimisation du milieu de culture : 35 2-2-7- Cinétique d’apparition d’activité protéolytique : 37 2-2-8 - Culture en fermenteur : 37
3 - RESULTATS 38
3-1- Vérification de la pureté des souches et leur appartenance au groupe lactique 39 3-2- Analyses physicochimiques du lactosérum et du jus de tomate 39 3-3- Optimisation du milieu de culture à base de lactosérum 40 3-3-1- Croissance en milieu LY (Lactosérum + extrait de levure) 41

3-3-2- Croissance en milieu LT (Lactosérum + jus de tomate) 41 3-3-3- Croissance en milieu LYT (lactosérum + extrait de levure + jus de tomate) 43 3.3-4- Croissance des souches de lactobacilles en milieu M2 44 3-3-5- Caractéristiques technologiques des lactobacilles cultivés en milieu M2 46 3-3-5-1- Production d’acide lactique en milieu M2 46 3-3-5-2- Production de diacétyle 48 3-4- Activité protéolytique en milieu M2 49 3-4-1- Recherche de l’activité protéolytique en milieu M2 49 3-4-2- Optimisation de l’activité protéolytique en milieu M2 52 3-4-3- Cinétique de production de protéases en milieu BMK 55 3-5- Cinétique de croissance, d’acidification, de production de diacétyle et de consommation du 57 substrat en fermenteur 3-5-1- Croissance et production d’acide lactique 57 3-5-2- Consommation du substrat 60 3-5-3- Production de diacétyle 61
4 - CONCLUSION 62
5 – REFERENCES BIBLIOGRAPHIQUES 65 ANNEXE 80

Liste des figures
Figure 1 : Voies technologiques permettant l’obtention des principaux types de lactosérums 4 issus de la première transformation du lait Figure 2 : Les produits finis résultants des différents procédés industriels appliqués au lactosérum 7 Figure 3 : Phosphorylations d’Hpr, PEP-dépendante par P~ EI ou ATP- dépendante par Hprk/P, 18 en présence de glucose Figure 4 : Principaux systèmes de transport des substrats à travers la membrane cellulaire 19 Figure 5 : Voies de transport et de métabolisme des glucides par les bactéries lactiques 20 Figure 6 : Principale voies du métabolisme du glucose par les bactéries lactiques 22 Figure 7 : Transport et métabolisme du lactose et du galactose chez les bactéries lactiques 23 Figure 8 : Modèle d’exclusion de l’inducteur chez les bactéries à Gram positif 26 Figure 9 : Modèle du mécanisme d’expulsion de l’inducteur chez les bactéries à Gram positif 27 Figure 10 : Principales étapes du métabolisme du citrate par les bactéries lactiques 28 Figure 11 : Catabolisme des acides aminés par Lc. lactis sp 29 Figure 12 : Analyse physicochimique du lactosérum 39 Figure 13 : Croissance en milieu LY 41 Figure 14 : Croissance en milieu LT 42 Figure 15 : Croissance en milieu LYT de la souche BH14 43 Figure 16 : Croissance des souches BH14 et CHTD27 dans les différents milieux 44 Figure 17: Croissance des lactobacilles en milieu M2 45 Figure 17 (suite): Croissance des lactobacilles en milieu M2 45 Figure 18 : Production d’acide lactique des souches en milieu M2 46 Figure 18 (suite) : Production d’acide lactique des souches en milieu M2 47 Figure 19 : Production de diacétyle par la souche BH14 48 Figure 20 : Activité protéolytique des souches en milieu M2 50 Figure 20 (suite) : Activité protéolytique des souches en milieu M2 50 Figure 21: Activité protéolytique en milieu solide MRSm-lait (1%) 51 Figure 22: Activité protéolytique en milieu solide M2-lait (1%) 51 Figure 23: Activité protéolytique de la souche V7 52 Figure 24: Activité protéolytique de la souche CHM9 52 Figure 25: Activité protéolytique de la souche CHTD27 53 Figure 26 : Activité protéolytique des lactobacilles en milieu MRSm et BMK 54 Figure 27: Activité protéolytique en milieu solide MRSm-lait (1%) 54 Figure 28: Activité protéolytique en milieu solide BMK-lait (1%) 54 Figure 29 : Cinétique de croissance et libération des protéases de la souche BH14 sur milieu MRSm 55 Figure 30: Cinétique de croissance et libération des protéases de la souche BH14 sur milieu BMK 55 Figure 31: Cinétique de croissance et libération des protéases de la souche CHTD27 sur milieu MRSm 56 Figure 32: Cinétique de croissance et libération des protéases de la souche CHTD27 sur milieu BMK 56 Figure 33 : Activité protéolytique dans le surnageant de culture de la souche BH14 sur milieu agar-lait 56 Figure 34 : Croissance, production d’acide lactique et évolution du pH en milieu MRSm de la souche 57 BH14. Figure 35 : Croissance, production d’acide lactique et évolution du pH en milieu BMK de la souche 58 BH14. Figure 36 : Croissance, production d’acide lactique et évolution du pH en milieu MRSm de la souche 58 CHTD27. Figure 37 : Croissance, production d’acide lactique et évolution du pH en milieu BMK de la souche 59 CHTD27.
i

Liste des tableaux
Tableau 1: Composition de différents types de lactosérum 5 Tableau 2 : Les fermentations du lactosérum 10 Tableau 3 : Besoins en acides aminés des lactobacilles 14 Tableau 4 : Compositions physico-chimiques de l’extrait de levure et jus de tomate 16 Tableau 5 : Code et origine des bactéries lactiques 31 Tableau 6 : Enrichissement du lactosérum par l’extrait de levure ou le jus de tomate 36 Tableau 7 : Analyse physicochimique du jus de tomate 40 Tableau 8 : Production de diacétyle 49 Tableau 9 : Concentration du substrat 60 Tableau 10 : Paramètres d’état des cultures 61 Tableau 11 : Production de diacétyle par la souche BH14 61
ii

1 - INTRODUCTION

Introduction
2
L’industrie agroalimentaire doit faire face à un problème de plus en plus crucial ces dernières années. Il s’agit de la production créée par les déchets et les rejets de cette industrie. L’industrie laitière est à l’origine de la production de grandes quantités de lactosérum (neuf fois le tonnage de fromages fabriqués), rejeté dans les cours d’eau et les égouts. Du fait de sa richesse en éléments nutritifs tels que le lactose, les protéines solubles, les vitamines, les matières grasses et les éléments minéraux, le lactosérum constitue un excellent milieu de culture pour les microorganismes, ce qui fait de lui un facteur de pollution redoutable.
Aujourd’hui, la lutte contre la nuisance conduit à interdire cette pratique et le déversement du sérum dans un cours d’eau doit être nécessairement précédé d’une épuration très coûteuse et qui vise à éliminer le lactose et les protéines, les constituants les plus précieux du sérum. Le développement des techniques fines de séparation moléculaire a offert à l’industrie alimentaire, pharmaceutique et autre une large gamme de produits issus du lactosérum et de ses dérivés. En plus, l’ingénierie microbiologique a trouvé dans l’industrie nouvelle du lactosérum un champ d’application privilégié pour produire l’alcool, des protéines et des acides organiques.
Parmi les microorganismes utilisés, les bactéries lactiques, qui possèdent des qualités exceptionnelles et un potentiel métabolique important, peuvent utiliser le lactosérum pour croitre, se multiplier et donner une importante quantité de biomasse et des métabolites souvent indispensables pour l’homme.
Dans notre travail, nous nous sommes intéressés à la valorisation du lactosérum doux comme milieu de culture pour les bactéries lactiques appartenant au genre Lactobacillus. Cette étude comporte les étapes suivantes :
1- Confirmation de l’appartenance des souches utilisées dans cette étude au groupe des bactéries lactiques.
2- Optimisation du lactosérum comme milieu de culture pour les lactobacilles : des expériences ont porté sur l’influence de l’ajout dans ce milieu de l’extrait de levure et/ ou de jus de tomate sur la croissance de deux lactobacilles. Nous avons ensuite examiné la croissance d’une trentaine d’autres souches de Lactobacillus sur le milieu optimisé à base de lactosérum.
3- Etude des caractéristiques technologiques des lactobacilles en milieu optimisé à base de lactosérum: activité protéolytique, production d’acide lactique et de diacétyle.
4- Dans une dernière étape nous avons réalisé une étude cinétique de croissance, de production d’acide lactique, et de consommation du substrat dans le même milieu à base de lactosérum, en fermenteur en mode discontinu et dans des conditions de pH non contrôlé. Parallèlement à cette étude et dans les mêmes conditions de culture,

Introduction
3
nous avons suivi en fonction du temps les productions de protéases et de diacétyle par deux souches de lactobacilles.
Avant de décrire notre travail nous présentons une étude bibliographique qui permettra de définir et de souligner l’importance technologique des lactobacilles et du lactosérum, et de passer en revue les principaux travaux réalisés pour valoriser ce sous-produit.

Analyse bibliographique
4
1-1 – Lactosérum : Appelé autre fois petit lait, le lactosérum est la phase liquide (jaune verdâtre), se séparant du caillé lors de la transformation du lait en fromage ou de la production de caséines. (Roufik, 2001).
Selon le procédé d’obtention, différents types de lactosérums peuvent ainsi être obtenus comme illustré sur la figure 1. Ces lactosérums peuvent être classés en deux principales catégories selon l’acidité du liquide obtenu (Alais, 1984) : è Les lactosérums doux : dont l’acidité varie entre 15 et 22° Dornic (pH=6,5). Ils sont issus
de la production de pâtes pressées et/ou cuites ou molles (Edam, St Paulin, Emmental). è Les lactosérums acides : obtenus lors de la fabrication des pâtes fraîches ou lors de la
production des caséines atteignent 120° Dornic, soit un pH proche de 4,5. LAIT
Figure 1 : Voies technologiques permettant l’obtention des principaux types de
lactosérums issus de la première transformation du lait (Alais, 1984).
La composition dépend du lait d’origine et du procédé de coagulation des caséines. Il contient essentiellement du lactose, des protéines globulaires, une fraction azotée non protéique, de la matière grasse et des minéraux (Fox, 1989).
Pasteurisation- Ecrémage -Standardisation
Crème
LAIT ÉCREMÉ LAIT STANDARDISÉ
Caséinate acide Caséinerie présure
Fromagerie pâtes fraiches
Fromagerie pâtes pressées pâtes molles
Caséine acide
Caséine présure
Fromage frais
Fromages
Ultrafiltration
Protéines
Centrifugation
Sérum acide de caséine
Sérum doux de caséine
Sérum acide de pâtes fraîches
Sérum doux de fromagerie
Sérum de protéine
Perméat d’ultrafiltration
Analyse bibliographique
5
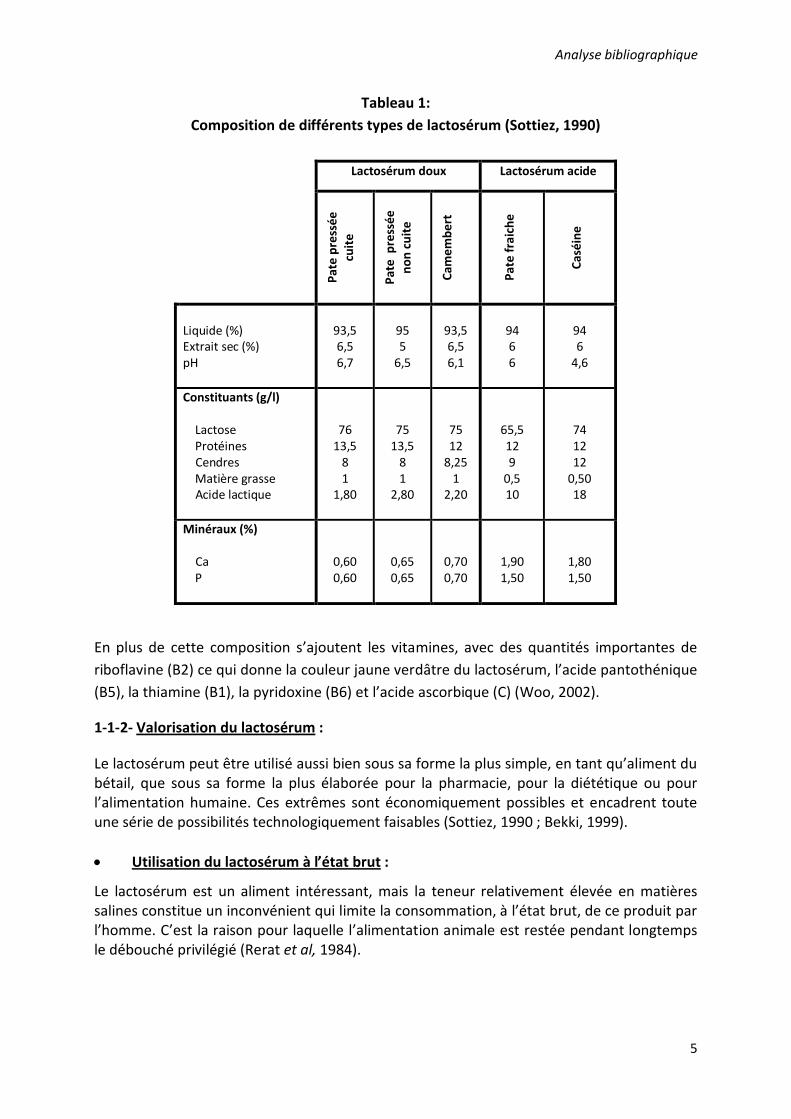
Tableau 1: Composition de différents types de lactosérum (Sottiez, 1990)
Lactosérum doux Lactosérum acide
Pate
pre
ssée
cu
ite
Pate
pre
ssée
no
n cu
ite
Cam
embe
rt
Pate
frai
che
Ca
séin
e
Liquide (%) Extrait sec (%) pH
93,5 6,5 6,7
95 5
6,5
93,5 6,5 6,1
94 6 6
94 6
4,6
Constituants (g/l) Lactose Protéines Cendres Matière grasse Acide lactique
76 13,5
8 1
1,80
75 13,5
8 1
2,80
75 12
8,25 1
2,20
65,5 12 9
0,5 10
74 12 12
0,50 18
Minéraux (%) Ca P
0,60 0,60
0,65 0,65
0,70 0,70
1,90 1,50
1,80 1,50
En plus de cette composition s’ajoutent les vitamines, avec des quantités importantes de riboflavine (B2) ce qui donne la couleur jaune verdâtre du lactosérum, l’acide pantothénique (B5), la thiamine (B1), la pyridoxine (B6) et l’acide ascorbique (C) (Woo, 2002).
1-1-2- Valorisation du lactosérum :
Le lactosérum peut être utilisé aussi bien sous sa forme la plus simple, en tant qu’aliment du bétail, que sous sa forme la plus élaborée pour la pharmacie, pour la diététique ou pour l’alimentation humaine. Ces extrêmes sont économiquement possibles et encadrent toute une série de possibilités technologiquement faisables (Sottiez, 1990 ; Bekki, 1999). • Utilisation du lactosérum à l’état brut :
Le lactosérum est un aliment intéressant, mais la teneur relativement élevée en matières salines constitue un inconvénient qui limite la consommation, à l’état brut, de ce produit par l’homme. C’est la raison pour laquelle l’alimentation animale est restée pendant longtemps le débouché privilégié (Rerat et al, 1984).

Analyse bibliographique
6
Alimentation animale : Le débouché principal des lactosérums est l’alimentation des veaux et de façon plus fluctuante l’alimentation animale dans son ensemble (Adrian, 1971). C’est sur cette utilisation croissante que se sont penchées de nombreuses équipes de recherches spécialisées dans ce domaine, pour améliorer cette alimentation et diminuer les troubles gastro-intestinaux, ainsi :
Ø l’utilisation du lactosérum fermenté avec Lactobacillus acidophilus pour l’alimentation des veaux a montré une meilleure croissance sans aucun désordre gastro-intestinal. Cet essai a également réussi chez les volailles et les porcs. Ø l’enrichissement du lactosérum en azote non protéique par fermentation et neutralisation a donné des résultats satisfaisants pour l’alimentation de bœufs et de vaches laitières. Ø de nombreux travaux ont signalé le développement réussi d’un ensilage de paille avec le lactosérum pour l’alimentation des ruminants.
(Bekki, 1999)
Alimentation humaine : Ø l’incorporation du lactosérum dans la fabrication des laits fermentés : Le lactosérum est incorporé dans la fabrication du yaourt et du leben comme remplaçant de l’eau dans le processus de reconstitution du lait à partir de la poudre du lait. Cette incorporation permet une amélioration notable du point de vue aspect et arôme du yaourt, due essentiellement à l’effet bénéfique exercé par les protéines sériques et le lactose du lactosérum. Ø boisson de lactosérum : Un lactosérum pasteurisé a été additionné à cinq types de jus de fruits différents (orange, raisin, fraise, banane et pomme) en plus de l’ajout de saccharose et d’acide ascorbique. Les résultats ont effectivement montré la bonne qualité des boissons obtenues, en particulier celui du jus d’orange, de pomme et de fraise. Ainsi, on peut avoir, à partir du lait, du fromage et des boissons fruitées à base de lactosérum (Vojnovic et al., 1993).
• Utilisation lactosérum traité : L’un des développements les plus significatifs de l’industrie laitière concerne la transformation du lactosérum en de nombreux dérivés, dont les propriétés nutritionnelles et fonctionnelles intéressent différents secteurs de l’industrie agroalimentaire, en derma-tologie et en cosmétologie. Le diagramme illustré dans la figure 2 résume les différents procédés utilisés dans le traitement du lactosérum et montre les produits finis obtenus.

Analyse bibliographique
7
Figure 2 : Les produits finis résultants des différents procédés industriels appliqués au lactosérum (Dairy processing handbook, 1995).
Lactosérum
Séparation Concentration des
solides totaux
Osmoses inverses
Évaporation
Séchage
¯Poudre de lactosérum ¯Lactosérum concentré ¯Lactosérum concentré sucré
Crème de
sérum
Caséines fines
Fractionnement des solides totaux
Récupération des protéines
Ultrafiltration Chromatographie
Récupération du lactose
Concentré protéique de lactosérum
(CPLs)
¯Lactoperoxidase ¯Lactoferrine ¯ α-lactalbumine ¯ β-lactoglobuline
Lactose
Déminéralisation
Crème de
sérum Caséines
fines
Poudre de lactosérum
partiellement déminéralisée
Poudre de lactosérum
déminéralisée

Analyse bibliographique
8
1-1-3- Utilisation du lactose :
Ø Industries chimiques : l’utilisation du lactose par ces industries a été mise au point dans les années 70, où un procédé original de fabrication d’écumes de polyuréthane a connu une réussite importante. Ces écumes sont largement utilisées dans la construction, l’isolation thermique et phonique et la fabrication des emballages (Ustad et al, 1970).
Ø Industrie pharmaceutique : elle utilise de grandes quantités de lactose raffiné comme charge dans la préparation de nombreux médicaments. En effet le lactose est considéré comme l’un des glucides les mieux adaptés à la préparation des milieux de fermentation destinés au développement des moisissures dans la fabrication des antibiotiques (Harper, 1992).
Ø Industries alimentaires : les industriels diversifient de plus en plus les débouchés du lactose en industrie alimentaire, notamment en charcuterie, confiserie, boulangerie, biscuiterie et pâtisserie ; dans la fabrication des chips et des pommes de terre frites pour favoriser les réactions de brunissement et de caramélisation (Hoppe et Higgins, 1992 ; Sottiez, 1990)
Ø Autres utilisations : le lactose peut être utilisé en aliment pour enfant et destiné à rempla-cer le lait maternel.
1-1-4- Utilisation des protéines :
Les protéines du lactosérum constituent 20% de la totalité des protéines contenues dans le lait (Morr, 1982; de Wit, 1981). Elles sont composées de plusieurs types de protéines dont la β-lactoglobuline et l’α-lactaglobine sont majoritaires. Les immunoglobulines, la sérum albumine bovine (BSA), le glycomacropeptide (GMP) et les protéoses-peptones font partie des constituants protéiques mineurs du lactosérum. Ø en industrie pharmaceutique la lactoferrine et la lactoperoxidase, possédant des
propriétés antibactériennes, sont utilisées pour fabriquer différents produits de désinfection utilisés en oto-rhino-laryngologie, comme les solutions pour bain de bouche, les pastilles pour la gorge et les dentifrices (Bekki, 1999).
Ø la β-lactoglobuline peut remplacer le sérum de veau fœtal dans la culture de cellules d’hybridomes de souris avec des résultats satisfaisants tant en croissance qu’en production d’anticorps, avec un prix de revient beaucoup moins cher (Capiaumont et al, 1994).
Ø l’α-lactalbumine, permettrait de lutter contre la déficience hormonale en sérotonine, puisqu’elle contient dans sa chaîne, en 4 exemplaires, un acide aminé rare, le tryptophane, qui rentre dans la composition de la sérotonine. Les expériences de production par génie génétique de cet acide aminé, aux USA et au Japon, ne semblent pas concluantes (Pearce, 1983).
Ø pour certains types d’alimentation spécifique ou généralement pour tous ceux qui pour une raison quelconque (ablation partielle de l’intestin, maladie immunodéficitaire, syndromes allergiques…) ne peuvent s’alimenter normalement, l’INRA (France) a mis au point, à partir du lactosérum, une solution nutritive composé uniquement de petits fragments protéiques, directement assimilables par l’organisme pour synthétiser les protéines dont il a besoin (Roger, 1979).

Analyse bibliographique
9
Ø les protéines du lactosérum sont utilisées dans les laits infantiles pour augmenter le taux des protéines sériques (plus élevé dans le lait maternel que dans le lait de vache) (Hilpert et al, 1977 ; Hilpert et al, 1987 ; Fukumoto et al, 1994).
Ø elles sont utilisées dans le monde du conditionnement physique et ‘’bodybuilding’’, car ces protéines jouent un grand rôle dans la construction des fibres musculaires qui ont subi des micro-déchirures lors de l’entrainement (FAO, 1995).
Ø les protéines du lactosérum peuvent donner d’excellents gels pouvant être utilisés dans certains desserts comme les glaces, les crèmes et les flans (Kinsella, 1984). Elles sont également utilisées en boulangerie, biscuiterie, charcuterie et salaisonnerie pour l’apport protéique, leur valeur nutritionnelle, la rétention d’eau, leur pouvoir gélifiant et leurs propriétés moussantes (Britten et al, 1994 ; Plock et Kessler, 1993).
1-1-5- Fermentations diverses du lactosérum : Le lactosérum constitue un excellent milieu de culture, notamment pour tous les micro-organismes susceptibles de métaboliser le lactose. Il peut être soumis à de nombreuses fermentations conduisant à des acides, des alcools, des enzymes, des vitamines, des boissons alcoolisées ou non (Veisseyre, 1979). En effet, l’utilisation des micro-organismes a trouvé dans ces industries un champ d’application privilégié pour obtenir toute une gamme de produits de plus en plus élaborés (Hansen, 1980 ; Hiroshi et al, 1993 ; Roukas et al, 1991). Quelques types de fermentations réalisées jusqu’à présent sur le lactosérum sont représentés dans le tableau 2.

Analyse bibliographique
10
Tableau 2 : Les fermentations du lactosérum (Yang et Silva, 1995 modifié).
Produits Organismes Milieu Référence
P.O.U Alcool Bio-éthanol Levure de boulangerie Acide lactique Acide acétique Propionate Polysaccharide Huile Enzymes β-Galactosidase Acétone-butanol Lysine Vitamine B12 Acide citrique Acide L-ascorbique Glycerol Insecticides Gomme de Xanthane
Kluyveromyces fragilis Rhodopseudomonas spheroides Bacillus megaterium Kluyveromyces marxianus Candida pseudotropicalis Lactobacillus casei spp. rhamnosus Lactobacillus acidophilus Lactobacillus delbrueckii ssp. bulgaricus Kluyveromyces fragilis Kluyveromyces marxianus Kluyveromyces marxianus Saccharomyces cerevisiae Lactobacillus delbrueckii ssp lactis Lactobacillus helveticus Streptococcus thermophilus Lactobacillus helveticus Streptococcus lactis Clostridium formicoaceticum Propionobacterium acidipropionici Propionobacterium sp. Lactobacillus delbrueckii ssp. Bulgaricus RR Apiotrichum curvatum Candida curvata Aspergillus niger Candida pseudotropicalis Clostridium acetobutylicum Mutant d’Escherichia coli Propionibacterium sp Propionibacterium shermanii Aspergillus niger Mutant de Candida norvegensis K. fragilis K. marxianus Bacillus thuringiensis Xanthomonas campestris
Lactosérum doux Lactosérum acide Lactosérum + E.L Lactosérum des fromages blanc Lactosérum acide Lactosérum Perméat de lactosérum Perméat de lactosérum acide + E.L Perméat de lactosérum Lactosérum doux Lactosérum Lactosérum + E.L Perméat de lactosérum Lactosérum doux Lactosérum Perméat de lactosérum Lactose Lactosérum + E.L Perméat de lactosérum + E.L Lactosérum Perméat de lactosérum Lactosérum doux + E.L Perméat de lactosérum Perméat de lactosérum doux Perméat de lactosérum Lactosérum doux Perméat de lactosérum hydrolysé + E.L
Reinbold et Takemoto, 1988 Marwson, 1988 Gomez et Castillo, 1983 Sandhu et Waraich, 1983 Djeghri et al., 2007 Gawel et Kosikowski, 1978 Tin et Mawson, 1993 Sooch et Singh , 2002 Champagne et al, 1990 Mehaia et Cheryern, 1986 Chiarini et al., 1992 Boudjemaa, 2008 Amrane et Prigent, 1997 Amrane et Prigent, 1998 Tamura et al., 1993 Yang et al., 1994 Crow, 1988 Briczinski et Roberts, 2002 Ykema et al, 1988 Floetenmeyer et al, 1985 Leuchtenberger et al, 1989 Gomez et Castillo, 1983 Ennis et Maddox, 1985 Sobotkova et al., 1989 Marwaha et al., 1983 Somkuti et bencivengo, 1981 Cayle et al., 1989 Jeng et al., 1989 Rapin et al., 1994 Salama et al., 1983 Maldonado, 1992
P.O.U. : Protéines d’Organisme Unicellulaire

Analyse bibliographique
11
1-2- Les lactobacilles : Les lactobacilles colonisent tous les habitats contenant des glucides fermentescibles, des produits d’hydrolyse des protéines, des vitamines, des facteurs de croissance et une basse tension d’oxygène. On les trouve dans les végétaux, les intestins des animaux et de l’homme ; on peut les isoler également des cavités naturelles de l’organisme. Le genre Lactobacillus appartient à la famille des Lactobacillaceae, il regroupe de nombreuses espèces bactériennes, omniprésentes dans la nature et rarement pathogènes. Il s’agit de bacilles souvent allongés, Gram positif, non sporulés, parfois groupés en paires ou en chainettes, généralement immobiles. Ils sont catalase négatif (certains ont une pseudocata-lase) mais sont benzidine négatif, microaérophiles ou anaérobies. Ils ont un métabolisme fermentaire produisant de l’acide lactique. Certaines espèces sont homolactiques, d’autres hétérolactiques produisant des acides volatils, de l’éthanol et du CO2 en plus de l’acide lactique. Les lactobacilles se montrent généralement plus résistants au stress acide que les lactocoques, possédant une large gamme de température de croissance (2 à 53°C), et un pH optimal de croissance qui se situe selon les espèces entre 4,5 et 6,4 (Ho Thi, 2008 ; Euzéby, 2000 ; de Roissart et luquet, 1994). Ils ont la capacité de limiter ou d’empêcher le développement de microorganismes dangereux ou indésirables (Desmazeaud, 1983 ; de Roissart et Luquet, 1994). 1-2-1-Taxonomie et classification des lactobacilles :
En 1919 Orla-Jensen propose de diviser le genre Lactobacillus en 3 sous-genres : Thermo-bacterium, Streptobacterium et Betabacterium.
Plus récemment les lactobacilles ont été répartis en 3 groupes basés sur les voies fermentatives de différentes sources de carbone (Kandler et Weiss, 1986 ; de Roissart et Luquet, 1994 ; Klein et al., 1998): le groupe I rassemble les lactobacilles homofermentaires stricts, se développant à 45°C mais pas à 15°C; le groupe II les lactobacilles hétérofermentaires facultatifs mésophiles, fermentant les pentoses par la voie hétérofermentaire et les hexoses par la voie homofermentaires : le groupe III les lactobacilles hétérofermentaires stricts. Ces 3 groupes correspondent sensiblement aux divisions d’Orla Jensen (1919)
La classification phylogénique et l’identification au niveau de l’espèce, compte aujourd’hui largement sur des méthodes moléculaires basées sur des régions hautement conservées dans le génome microbien, comme le gène ADN ribosomal 16s (ADNr16s) (Sang et al., 2000). Les groupes I, II et III renferment des ‘’Complexes’’ rassemblant des espèces syntoniques : ainsi le groupe I par exemple comporte deux complexes (Complexe Lb. delbrueckii et complexe Lb. acidophilus) et des espèces non regroupées comme Lb. jensenii, Lb. aviarius, …

Analyse bibliographique
12
1-2-2 Exigences nutritionnelles des lactobacilles : Les lactobacilles se caractérisent par des besoins nutritionnels complexes. Ces exigences ont été déterminées par leur aptitude à croitre ou non dans un milieu chimiquement défini (MCD). Généralement, les MCDs contiennent des sels minéraux, une source de carbone, des vitamines, des acides aminés et des bases azotées (Chervaux et al., 2000; Christensen et steele, 2003 ; Grobben et al., 1998 ; Lauret et al., 1996 ; Ledesma et al., 1977 ; Morishita et al., 1981 ; Morishita et al., 1974). Ø Exigences en vitamines
Toutes les espèces de lactobacilles présentent une exigence absolue en pantothénate de calcium (B5) et en niacine (B3) et des exigences différentes pour les autres vitamines. Les déficiences en cobalamine (B12) peuvent induire une diminution de la synthèse d’ADN et entrainer des changements morphologiques, les cellules devenant filamenteuses. Une telle élongation cellulaire a aussi été rapportée pour Lb. helveticus ssp jugurti lors de déficiences en cobalamine (B12) ou en acide folique (B9). Bien que les résultats obtenus par les différents auteurs sur les besoins en vitamines ne concordent pas toujours, il apparaît clairement que plusieurs vitamines du groupe B sont essentielles au métabolisme de l’ensemble des bactéries lactiques (de Roissart et Luquet, 1994). Ø Exigences en bases azotées
Dans les milieux synthétiques, les lactobacilles exigent la présence d’adénine, de cytosine, de désoxyguanosine, de guanosine, de thymidine et d’uracile (Ledesma et al., 1977). L’acide orotique, qui est un intermédiaire de la synthèse des bases puriques, est un facteur de croissance pour Lb. bulgaricus. Il régule la synthèse des acides ribonucléiques, tandis que la synthèse des acides désoxyribonucléiques n’est pas affectée. En général, chez les lactobacilles, les exigences en bases azotées sont très variables selon les souches ; l’addition de ces composés peut même, chez certaines souches, entrainer des phénomènes d’inhibition de la croissance (Desmazeaud, 1992). Ø Exigences en acides aminés
Les besoins en acides aminés (ou auxotrophies) des lactobacilles sont variables. Les lactobacilles homofermentaires (comme Lb. delbrueckii et Lb. helveticus) ainsi que les hétérofermentaires facultatifs (comme Lb. casei, Lb. sakei et Lb. curvatus) sont les bactéries lactiques les plus exigeantes, avec de 11 à 17 auxotrophies. On note une exception avec l’espèce Lb. plantarum qui est caractérisée par au maximum 7 auxotrophies, proches de celles des lactobacilles hétérofermentaires obligatoires (comme Lb. brevis, Lb. buchneri et Lb. fermentum). Le tableau 3 montre les exigences en acides aminés de quelques souches Lactobacillus.

Analyse bibliographique
13
Ø Influence des cations
• Magnésium C’est le principal cation divalent chez les cellules vivantes. Il est lié aux nucléotides intracellulaires et intervient dans les systèmes de transport intramembranaires. Le magnésium est un activateur des différentes réactions métaboliques : division cellulaire, stabilisation des acides nucléiques ou hydrolyse peptidique. De plus, le magnésium est essentiel pour les phosphokinases impliquées dans la glycolyse. Cet ion serait indispensable pour la croissance de Lb. helveticus et essentiel pour celle de Lb. delbrueckii (Desmazeaud, 1992).
• Manganèse Le manganèse est nécessaire à la structure et le fonctionnement des enzymes, dont l’ARN polymérase, la LDH, la NADH oxydase (Archibald, 1986), la superoxyde dismutase et la catalase formée lors de la croissance sur hématine de Lactobacillus. (Johnston et Delwiche, 1965). Il est nécessaire pour la détoxification des cellules mises en présence de l’oxygène. Dans ce cas notamment chez Lb. plantarum. le manganèse se substituerait au superoxyde dismutase pour éliminer les radicaux O2- superoxyde (Desmazeaud, 1992). Par ailleurs, l’addition de Mn2+ au milieu de culture peut stimuler considérablement la croissance des lactobacilles durant la fermentation (Leroy et al., 2004).
• Fer Le fer n’a pas d’action générale sur la croissance ou sur la production d’acide lactique (Boyaval, 1989). Lb. plantarum n’exige pas le fer et ne l’accumule pas (Archibald et Fridovich, 1981).
• Calcium Il est souvent cité pour son rôle dans la paroi cellulaire en particulier pour l’activité des protéases de paroi. Il stimule la croissance de Lb. casei.

Analyse bibliographique
14
Tableau 3 : Besoins en acides aminés des lactobacilles (selon la nomenclature de Stiles et Holzapfel, 1997)
Acide aminé
Lactobacilles homofermentaires
Lactobacilles hétérofermentaires
obligatoire
Lactobacilles hétérofermentaires
facultatifs
Lb. acidophilus
Lb. delbrueckii
Lb. helveticus
Lb. b
revi
s
NCT
C 81
07
Lb.b
uchn
eri
NCD
O 1
10
Lb. f
erm
entu
m
ATC
C 93
38
Lb. casei
Lb. plantarum
Lb.sakei Lb. curvatus
N
CTC
1899
ATC
C 11
506
NCD
O 2
13
Ssp
bulg
aric
us
CNRZ
397
Ssp
bulg
aric
us
NCF
B 27
72
CRL
1062
CRL
974
ATC
C 15
009
CNRZ
32
ATT
C 74
69
S1
ATT
C 84
14
ATC
C 10
241
23
55
64
LR 1
02
LR 1
26
L-Acide aspartique ND + ND - S - - ND ND ND - ND + S ND - - - + + L-Acide glutamique + + + + S + + ND + + + + + + + - - - + + DL/L Alanine - S + + - S ND - - - - - - - - + + S S L-Arginine S + + + + + + ND + - + + + S - + + + + + L-Asparagine S - + - S/+ - - ND - - - - - - - + - + ND ND L-Cystéine - + + - + - S ND - + - + S S - - - - + + L-Glutamine - - - - S - - ND - - - - - - - + - + ND ND Glycine - S + - S - S ND - - - - - - - + + + + + L-Histidine - + + + + + + ND S - S - - - - + + + + + L-Isoleucine S + - + + + + + + S + + + + S + + + + + L-Leucine + + + + + + + + + + + + + + +/S + + + + + L-Lysine - S - - + - + ND - - - S/- + - - + + + + + L-Méthionine - + + + + + + + S - + S/- + + - + + + + + L-Phénylalanine - + - + + + + + + + + +/- S + - + + + + + L-Proline - + - + + + + + - - - - - - - + + + + + L-Sérine - + - + + + S ND - - - S/- + - - S S S + + L-Thréonine - + - - S/+ + + + - - - S/- + S - + + + + + L-Tryptophane S + + + + + + ND + - + + + + +/S S S S S + L-Tyrosine S + + + + + + ND + - + + + S S + + + + + L-Valine + + + + + + + + + + + + + + + + + + + + Auxotrophie totale 3 15 11 13 16 13 13 14* 8 5 9 10 12 7 4 14 13 15 16 17
+ : essentiel ; - : non essentiel ; ND : non déterminé ; S : effet stimulant ; * : détermination incomplète ; S : effet stimulant

Analyse bibliographique
15
• Autres cations : - Le Potassium joue un rôle important dans la régulation du pH intracellulaire.
Cet ion est exigé pour la croissance de Lb. helveticus et Lb. casei. - Le Sodium exerce un effet sélectif sur les différentes espèces. - Le Cadmium, contaminant du lait à faible concentration (6.10-5mM), peut
inhiber la croissance et la production d’acide chez Lb.delbrueckii et Lb. helveticus (Korkeale et al., 1984).
- Le Césium inhibe la croissance de Lb.arabinosus dés 2,25mM (Boyaval, 1989). - Le Cuivre inhibe la croissance de Lb. caseï et Lb.delbrueckii ssp bulgaricus dans
le lait et la production d’acide lactique pour des concentrations variant selon les espèces, entre 10-2 et 10-4 M (Boyaval, 1989). Le cuivre augmente la production de diacétyle (Kaneko et al., 1987).
- Le Cobalt, partie intégrante de la vitamine B12, n’est pas exigé par Lb. arabinosus ni par Lb. delbrueckii (Boyaval, 1989).
Actuellement de nombreux milieux riches et complexes, comme le milieu MRS (De Man et al., 1960) ou M17 (Terzaghi et Sandine, 1975), sont disponible et conviennent spécialement à la culture des bactéries lactiques. Ils contiennent des sources d’azote complexes tels que la peptone, l’extrait de levure et l’extrait de viande, ainsi que d’autres facteurs de croissance. Cependant leur utilisation est limitée à des usages spécifiques à cause de leurs coûts élevés. Plusieurs chercheurs se sont intéressés à l’optimisation des milieux de culture pour les bactéries lactiques par l’ajout de sources d’azote. Diverses sources d’azote ont été testées : extrait soluble de maïs(Kurbanoglu, 2004), hydrolysat protéiques du lactosérum (Cornelius et al., 1996), extrait de germes de malt (Mc Comas et Gilliland, 2003 ; Watanabe et al., 2004),, hydrolysat de caséines (Vahvaselkä et Linko, 1987 ; Chiarini et al., 1992), cellules de levure (Altaf et al., 2007), substrat végétal (Gardner et al., 2001 ; Nancib et al., 2001 ; Chalampopoulos et al., 2002 ; Djeghri-Hocine et al., 2006 ; Djeghri-Hocine et al., 2007 a), hydrolysats de viscères de poisson (Horn et al., 2007), jaune d’œuf (Djeghri-Hocine et al., 2007 b). La source d’azote la plus efficace semble être l’extrait de levure, mais ce produit est onéreux en production industrielle. Pour réduire les coûts, des tentatives ont été donc faites de remplacer l'extrait de levure par des extraits de bactéries lactiques (Amrane, 2000 ; Gaudreau et al., 2005). La substitution de l’extrait de levure par des autolysats des cellules de levures à été également testé (Barallé et Borzani, 1988 ; Amrane et Prigent, 1993 ; Timbuntam et al., 2006). Gibson et Abdel-Malek (1945) ont mis en évidence un milieu de culture pour la recherche du type fermentaire des souches. Parmi les constituants de ce dernier on trouve le jus de tomate. Le jus de tomate est riche en sources de sucres simples (essentiellement glucose et fructose), en minéraux et en vitamine B. Miller et Puhan (1980) et Babu et al. (1992) ont montré que le jus de tomate stimule la croissance de Lactobacillus acidophilus. Ils ont observé une augmentation de la biomasse bactérienne, un temps de génération plus court et une amélioration de la production d’acide dans le milieu.

Analyse bibliographique
16
Le tableau 4 permet de comparer les compositions physicochimiques de l’extrait de levure et du jus de tomate (Cotte, 2000 ; Ghaly et al., 2003).
Tableau 4 : Compositions physico-chimiques de l’extrait de levure et jus de tomate
(Cotte, 2000 ; Ghaly et al., 2003)
Constituant Extrait de levure Tomate crue Composition (%) Eau Azote total Protéines Glucides Lipides Acides aminés (%) Alanine Acide aminobutyrique Arginine Asparagine Cystéine Acide glutamique Glycine Histidine Isoleucine Leucine Lysine Méthionine Ornithine Phénylalanine Proline Sérine Thréonine Tryptophane Tyrosine Valine Sels minéraux (ppm) Calcium Fer Magnésium Potassium Sodium Phosphore Vitamines (ppm) Thiamine Riboflavine Pyridoxine Niacine Pantothénate de calcium
-
8,8 55 - -
3,4 0,1 2,1 3,8 0,3 7,2 1,6 0,9 2,0 2,9 3,2 0,5 0,3 1,6 1,6 1,9 1,9
- 0,8 2,3
- - - - - -
20-30 50-70 25-35 600 200
93,5 0,75
1 3.6 0,3
- - - - - - - - - - - - - - - - - - - -
2,95-3,95 0,6-0,8 2,5-4
18,7-29,5 15,7-17,6
2,4-2,9
0,6 0,4 0,8 0,5
-

Analyse bibliographique
17
1-3- Métabolisme des lactobacilles :
1-3-1- Métabolisme glucidique :
Les bactéries lactiques transportent et accumulent les glucides soit sous forme de radicaux libres, par transport actif, soit sous forme de dérivés phosphorylés, par des systèmes phosphotransphérase-phosphoénolpyruvate dépendant (PEP-PT). 1-3-1-1- Systèmes de transport actif : En 1961, Peter Mitchell a proposé une théorie pour expliquer comment le transport des électrons peut être couplé à la génération d’ATP. D’après ce concept chimiosmotique, une enzyme membranaire, l’adénosine 5’-triphosphate-phosphohydrolase, activée par le complexe Ca2+ + Mg2+, couple l’hydrolyse de l’ATP à la sortie de protons (2H+) de la cellule. Le flux de protons sortant de la cellule crée une différence dans la concentration de protons et de charge électrique de part et d’autre de la membrane cytoplasmique de la bactérie. Par la suite, en réponse à cette force promotrice, le sucre peut être assimilé grâce à un gradient de concentration (Thompson et Gentry-Weeks, 1994). 1-3-1-2- Systèmes PEP-PT : Les systèmes de translocation de groupe les mieux connus sont les systèmes phosphotransférases, dépendantes du phosphoénolpyruvate (PEP-PT). Trois protéines au moins (EI, HPr, et EII) sont requises pour le transport et la phosphorylation du sucre. Ces protéines forment une chaine de transporteurs qui prélèvent des groupes phosphate riches en énergie sur le phosphoénolpyruvate et les transfèrent au sucre qui doit pénétrer dans la cellule. Les deux premiers transporteurs sont identiques dans toutes les chaines PTS ; il s’agit de l’enzyme EI et d’une petite protéine thermostable HPr (Heat resistant Protein). L’enzyme I, cytoplasmique est capable de s’autophosphoryler aux dépens du PEP. Suite à cette première réaction, EI catalyse le transfert du groupement phosphate acquis vers la protéine HPr. La phosphorylation de cette dernière a lieu sur l’histidine (Kalbitzer et al., 1982 ; Weigel et al., 1982). Chez les bactéries à Gram positif et à faible pourcentage en GC, la protéine HPr peut également être phosphorylée sur la sérine (Figure 3). Cette réaction est catalysée par une enzyme bifonctionnelle, l’HPr kinase/phosphatase (Kravanja et al., 1999). L’enzyme III cytoplasmique est inductible par le sucre, accepte le radical phosphoryl de l’HPr pour former l’EIII~P. L’EIII~P transfère le radical phosphoryle à l’EII qui effectue la reconnaissance, la liaison, la phosphorylation et la translocation du sucre à travers la membrane de la cellule. Les voies de transport et le métabolisme des glucides par les bactéries lactiques sont résumées dans les Figures 4 et 5.

Analyse bibliographique
18
Figure 3 : Phosphorylations d’Hpr, PEP-dépendante par P~ EI ou ATP- dépendante par Hprk/P, en présence de glucose (Deutcher et al., 2002)
P-His-Hpr
P-Ser-Hpr
HPr Kinase/ Phosphatase

Analyse bibliographique
19
S : Substrat assimilable Hpr : Protéine membranaire non spécifique du substrat M : Médiateur de transport EI : Enzyme cytoplasmique non spécifique du substrat ATP : Adénosine triphosphate EII : Enzyme membranaire spécifique du substrat PEP : Phosphoénolpyruvate EIII : Enzyme cytoplasmique spécifique du substrat
Figure 4 : Principaux systèmes de transport des substrats à travers la membrane cellulaire (Thompson et Gentry-Weeks, 1994)

Analyse bibliographique
20
Figure 5 : Voies de transport et de métabolisme des glucides par les bactéries lactiques
(Thompson et Gentry-Weeks, 1994)

Analyse bibliographique
21
1-3-1-3- Métabolisme des sucres : Comme écrit plus haut les bactéries lactiques sont divisées en deux groupes principaux d’espèces, homofermentaire ou hétérofermentaire selon la nature et la concentration des produits terminaux issus de la fermentation des sucres (Figures 6 et 7). • Voie homofermentaire ou EMP (EMBDEN-MEYERHOF-PARNAS) :
Les lactobacilles homofermentaires convertissent presque exclusivement le glucose en acide lactique (90-95%). Une des enzymes clé de la glycolyse, présente dans toutes les espèces homo-fermentaires, est la fructose-1,6 diphosphate aldolase. C’est une enzyme qui transforme le fructose diphosphate en deux trioses phosphates. Les bactéries homolactiques contiennent habituellement de fortes concentrations de fructose diphosphate (FDP). Cette molécule, importante pour la régulation, sert d’activateur de la synthèse des enzymes terminales de la voie glycolique (pyruvate kinase et lactate déshydrogénase). • Voie hétérofermentaire ou PPC (PENTOSE PHOSPHOCETOLASE) :
La fermentation hétérolactique doit sa dénomination au fait qu’en dehors du lactate, elle aboutit à la formation d’éthanol, de CO2 et éventuellement d’acétate. Les bactéries lactiques hétérofermentaires ne contiennent pas de FDP aldolase ni de triose-phosphate isomérase. Elles sont également dépourvues d’un système phosphotransphérase-PEP pour le glucose. Chez ces bactéries, le glucose est accumulé par l’intermédiaire d’un transport actif, puis est phosphorylé par une glucokinase ATP-dépendante. Le glucose-6P est transformé en acide 6P-gluconique puis décarboxylé en 5P avec libération de CO2. Ce pentose-P est métabolisé en triose phosphate et en acétyl-P. Enfin l’acétyl-P est réduit en éthanol, et le triose phosphate est métabolisé en acide lactique par les dernières réactions de la voie EMP (Djidel, 2007).

Analyse bibliographique
22
P : Phosphate NADH : Nicotinamide adénine dinucléotide ADP : Adénosine diphosphate (1) : Lactate déshydrogénase ATP : Adénosine triphosphate (2) : Alcool déshydrogénase NAD+ : Nicotinamide adénine dinucléotide
Figure 6 : Principale voies du métabolisme du glucose par les bactéries lactiques. (Wee et al., 2006)

Analyse bibliographique
23
Figure 7 : Transport et métabolisme du lactose et du galactose chez les bactéries lactiques. (Thompson et Gentry-Weeks, 1994)

Analyse bibliographique
24
1-3-1-4 Régulation du métabolisme des sucres :
La fermentation des sucres s’effectue essentiellement en trois étapes : 1. le transport du sucre à travers la membrane cellulaire 2. le catabolisme intracellulaire du sucre 3. la formation et l’expulsion extracellulaire des métabolites terminaux.
Cette fermentation aboutit principalement à la production d’ATP et d’acide lactique. La dégradation des hexoses est soumise à un mécanisme rapide d’activation-inhibition des enzymes allostériques dont le rôle est important pendant la croissance bactérienne. Ainsi la conversion du phosphoénolpyruvate (PEP) en lactate fait intervenir deux enzymes allostériques : la pyruvate kinase (PK) et la lactate déshydrogénase (LDH) (toutes les deux stimulées par le fructose-1-6-diphoshate) (Crow et Pritchard, 1976 et 1977). Le PEP constitue non seulement une étape de la chaine glycolytique, mais intervient aussi dans le transport des sucres à travers la membrane. Il a été constaté que lorsqu’une bactérie lactique est soumise à une carence en sucre, la teneur en PEP croît notablement (inhibition de la pyruvate kinase), tandis que les intermédiaires métaboliques qui stimulent la PK décroissent (Poolman et al., 1987 ; Otto et al., 1985). Dans ces conditions, la dégradation se ralentit mais produit assez d’ATP pour entretenir la vie cellulaire et la concentration élevée en PEP permet la reprise du transport du sucre dès que celui-ci est à nouveau disponible (Thomas, 1976 ; Thompson et Thomas, 1977). 1-3-1-5- Régulation du système de transport des sucres : De nombreux facteurs déterminent si un sucre donné sera ou non transporté par des bactéries lactiques, et les mécanismes de transport sont soumis à diverses contraintes génétiques ou environnementales. Pour accumuler un sucre donné, une bactérie doit posséder l’information génétique correspondante à la protéine de transport appropriée. • Transport préférentiel des sucres : La protéine HPr peut être présente sous trois formes : HPr phosphorylée, HPr phosphorylée par l’enzyme EI sur l’histidine et HPr phosphorylée par une protéine kinase (HPrK/P) sur la sérine. Cette dernière enzyme joue un rôle clé dans la régulation du transport par le PTS et dans la répression/activation catabolique. En présence d’un sucre rapidement métabolisable comme le glucose, l’activité augmente et conduit à l’augmentation de la concentration en fructose diphosphate (FDP) et en ATP (Mijakovic et al., 2002 ; Neves et al., 1999 ; Thompson et Torchia, 1984) ; l’HPrK/P, stimulée par ces effecteurs, va phosphoryler l’HPr de façon ATP-dépendante sur la sérine (Galinier et al., 1998 ; Galinier et al., 1997 ; Jault et al., 2000 ; Ramstrom et al., 2003 ; Reizer et al., 1998). Lors d’études concernant le transport des sucres pour de cellules au repos de Lactococcus lactis ML3, Thompson et al., (1978) ont montré que dans un mélange de

Analyse bibliographique
25
sucres métabolisables (glucose, galactose et lactose) et d’analogues non métabolisables (2-désoxy-glucose ou 2DG et méthyl-β-D-thiogalactopyranoside ou TMG) la souche transporte de préférence le glucose et son analogue. Ce transport préférentiel a été attribué (1) à la compétition entre les différents enzymes II (EII) pour le donneur commun de phosphoryles, L’HPr-P, (2) à la compétition entre les substrats (lactose et TMG) pour EIIlac commune.
• Exclusion de l’inducteur :
De manière générale, on peut distinguer deux facettes dans la régulation de l’expression des gènes du métabolisme du carbone. Le mécanisme global de la répression/activation catabolique qui permet, d’une part de réprimer l’expression des gènes impliqués dans l’utilisation de sources de carbone non préférentielles en présence de sucre rapidement métabolisable comme le glucose (Luesink et al., 1998) et d’autre part, à l’opposé, de stimuler l’expression des gènes impliqués dans l’utilisation de ces sources préférées de carbone et d’énergie. En présence d’un mélange de glucose, lactose et galactose, les cellules de Lactococcus lactis ML3 co-métabolisent le glucose et le lactose, mais le galactose n’est consommé qu’après épuisement des sucres précédents (Thompson et al., 1978). Liberman et Bleiwiss (1984) ont décrit lors de la croissance en présence de glucose et de lactose dans le milieu de culture, des profils de croissance diauxique, ainsi qu’un métabolisme sélectif du glucose par rapport au lactose pour plusieurs espèces bactériennes. L’utilisation préférentielle du glucose est une conséquence de l’exclusion des molécules non préférentielles de la cellule (Figure 8) (Saier et Crasnier, 1996). Chez Lactococcus lactis la protéine HPr-sérine interagit avec la perméase PTS spécifique du lactose pour inhiber son activité (Ye et al., 1994). On peut citer aussi l’exemple de l’accumulation de galactose dans différents milieux de culture et qui est due à la fermentation incomplète du lactose par Streptococcus thermophilus (Hutkins et al., 1985). Contrairement à l’idée qui a longtemps prévalu, l’incapacité de cette souche à métaboliser le galactose n’est pas dû à l’absence de l’information génétique requise pour la synthèse des voies cataboliques appropriée. Les causes de ce phénotype ne sont pas connues : il serait dû en partie à une répression sévère de la synthèse de ces enzymes exercée par le glucose issu de l’hydrolyse du lactose. Ce phénomène d’inhibition de l’utilisation d’une source de carbone secondaire par la présence d’un sucre métabolisable est appelé répression catabolique (Miwa et al., 2000).

Analyse bibliographique
26
• Expulsion de l’inducteur :
Il s’agit de mécanisme par lequel l’ajout d’un sucre rapidement métabolisable dans le milieu de culture conduit à l’expulsion des autres sucres préalablement accumulés dans le cytoplasme (Luesink et al., 1998 ; Thompson et Saier, 1981; Reizer et Panos, 1980). Chez Lactococcus lactis le méthyle-β-D-thiogalactopyranoside (TMG) est accumulé dans la cellule sous forme phosphorylée (TMG-6P). mais l’ajout d’un sucre rapidement métabolisé provoque une expulsion du TMG sous forme déphosphorylée (Thompson et Saier, 1981). Ainsi en présence de glucose, HPr se trouve majoritairement sous la forme HPr-Ser-P (Deutcher et al., 1986; Deutcher et Saier, 1983). Celle-ci stimule l’activité d’une hexose-phosphate-phosphohydrolase qui va alors déphosphoryler le sucre phosphate pour permettre son expulsion hors la cellule (Reizer et al., 1983 ; Thompson et Chassy, 1983). La figure 9 illustre ce mécanisme d’expulsion de l’inducteur.
Figure 8 : Modèle d’exclusion de l’inducteur chez les bactéries à Gram positif. (Saier et Crasnier, 1996)

Analyse bibliographique
27
Figure 9 : Modèle du mécanisme d’expulsion de l’inducteur chez les bactéries à Gram positif (Saier et Crasnier, 1996).
1-3-2- Métabolisme du citrate :
Le citrate est un précurseur du diacétyle, qui contribue à l’arôme de certains types de laits fermentés, de fromages frais ou affinés, de beurres et de crèmes fraîches. Il conduit également à la formation de CO2, qui participe à la formation des ouvertures dans les fromages. Chez les bactéries lactiques, le transport du citrate à travers la membrane cellulaire est assuré par une citrate perméase. Une fois dans la cellule, le citrate est clivé en acétate et oxaloacétate par une citrate lyase. L’oxaloacétate est décarboxylé en pyruvate qui est transformé en diacétyle par le biais d’une série de réactions intermédiaires. Le pool du pyruvate disponible pour les voies de synthèse du diacétyle dépend de l’activité des enzymes impliquées dans la bioconversion du citrate en pyruvate. Il est à noter que le maintien du gradient de concentration dépend de la vitesse de disparition du citrate dans le milieu intracellulaire, donc de l’activité du citrate lyase (dégradation du citrate en oxaloacétate et acétate) et de l’oxaloacétate décarboxylase (décarboxylation de l’oxaloacétate en pyruvate). L’activité de la citrate lyase est inductible par le citrate et disparaît rapidement après épuisement du citrate dans le milieu. Lindegreen et al. (1990) ont pu démontrer le co-métabolisme du lactate et du citrate chez Lactobacillus plantarum dans des conditions d’anaérobiose. Le citrate est ainsi transformé en acétate et en succinate. Dans ces conditions, il y a aussi induction de la pyruvate formate
P-His-HPr
P-Ser-Hpr
P-Ser-Hpr

Analyse bibliographique
28
lyase sans formation de diacétyle et d’acétoine. L’acide succinique a été retrouvé par Kanenchi et al. (1988) dans le métabolisme de l’acide citrique des lactobacilles. Ces auteurs ont pu constater que l’acide succinique pouvait être formé à partir de l’acide fumarique, mais pas à partir de l’acide pyruvique ni de l’acide α-cétoglutarique. Figure 10 : Principales étapes du métabolisme du citrate par les bactéries lactiques
(Bourel et al., 2001). (1) Citrate perméase, (2) Citrate lyase, (3) Oxaloacétate décarboxylase, (4) Lactate déshydrogénase, (5)
α-acétolactate synthase,(6) α -acétolactate décarboxylase,(7) Décarboxylation non enzymatique, (8) 2,3-butanediol déshydrogénase, (9) Acétate kinase, (10) Phosphotransacétylase (11) Alcool
déshydrogénase.
1-3-3- Métabolisme azoté :
Dans les fromages à maturation lente, les lactobacilles qui représentent à certaines périodes la microflore dominante, sont susceptibles de jouer un rôle dans la dégradation de la caséine et de participer ainsi à la formation de la saveur et de l'arôme du fromage suite à la libération des acides aminés (Thivierge, 2001). Or l'aptitude réelle de ces micro-organismes à la production d'enzymes protéolytiques est encore mal connue (Fira et al., 2001 ; Courtin et al., 2002 ; Kabadjova-Hristova et al., 2006), peu d'études systématiques portant sur un grand nombre de souches ayant été réalisées. Les études génétiques, biochimiques et ultrastructurales ont permis de conclure que les systèmes protéolytiques des lactocoques et des lactobacilles sont remarquablement similaires, en ce qui concerne leurs composants et leurs mode d’action (Kunji et al., 1996).

Analyse bibliographique
29
La dégradation des protéines est un processus comportant plusieurs étapes. Dans un premier temps, les protéases et les endopeptidases séparent la paracaséine composée des caséines αs1, αs2, de la caséine β et de la paracaséine κ en peptides de plus petite taille. On nomme ce processus la protéolyse primaire ou « protéolyse en largeur ». Lors de la protéolyse secondaire, les fragments de protéines continuent à être décomposés jusqu’au niveau des acides aminés. On parle aussi de « protéolyse en profondeur ». La figure 10 présente le catabolisme des acides aminés par Lactococcus lactis sp.
Figure 11 : Catabolisme des acides aminés par Lc. lactis sp. (Grattepanche, 2005)
Dpp : système de transport des di- et tripeptides ; Opp : système de transport des oligopeptides ; aat : système de transport des acides aminés

2 - MATERIEL & METHODES

Matériel et Méthodes
31
2-1- Matériel : 2-1-1 Souches bactériennes : Les souches bactériennes utilisées dans cette étude appartiennent à la collection du Laboratoire de Biologie des Microorganismes et Biotechnologie (LBMB). Nous avons utilisé une trentaine de souches appartenant au genre Lactobacillus. Le tableau 5 montre le code et l’origine des bactéries lactiques utilisées.
Tableau 5 : Code et origine des bactéries lactiques
Code Origine Référence
CHM9,CHM11,CHM12,CHM16, CHM17,CHM18,CHM19,CHM20
Lait cru de chamelles
Mauritanie
Cheikh, 2008
BH14, BH21
Lait cru de chamelles
Illizi
Bounoua, 2005
CHTD25, CHTD27, CHTD29
Lait cru de chamelles
Tindouf
Belkheir , 2004
9(b)
Saumure d’olive
Sig
Hebali, 2006
H3
Hamoum Relizane
Bouziani et Sabaha, 2006
V7, V8, V14, V20, V21, V23
Viande bovine fraiche
Mostaganem
Benbernou, 2008
LVK9, LVK11, LVK12, LVK13, LVK14, LVK15, LVK29, LVK30, LVK31, LVK32
Lait cru de vache
Oran
Karam, 1995
2-1-2- Lactosérum : Le lactosérum provient de l’unité de fabrication des fromages à pâte molle (camembert) située à Maraval (Oran).

Matériel et Méthodes
32
2 -2- Méthodes : 2-2-1-Vérification de la pureté et l’appartenance des souches au groupe lactique : La purification est obtenue par la technique des quadrants sur milieu MRS solide (de Man, Rogosa et Sharpe, 1960). Les colonies isolées obtenues sur milieu solide après incubation à 30°C pendant 24 à 48 h sont soumises à une observation macroscopique afin de préciser leurs tailles, pigmentations, contours, aspects, ... La morphologie des cellules bactériennes (forme, taille et mode d’association) est révélée après coloration par la technique de Gram par observation microscopique à l’immersion. La recherche de la catalase est réalisée sur une partie de la colonie émulsionnée dans une goutte d’eau oxygénée. Une réaction positive est révélée par le dégagement d’oxygène (Stiles et Holzapfel, 1997) Les lactobacilles se présentent sous forme de bâtonnets, Gram positif et catalase négative. 2-2-2- Conservation des bactéries : La conservation de courte durée des bactéries est effectuée à 4°C en gélose inclinée en tubes, avec un repiquage toutes les 4 semaines. Pour la conservation de longue durée, les souches sont conservées à – 20°C dans du lait écrémé à 10% et également dans du milieu MRS supplémenté de 40% glycérol (Herrero et al, 1996 ; Samelis et al, 1994). 2-2-3- Déprotéinisation du lactosérum : Le lactosérum est déprotéiné par un chauffage au bain Marie à 100°c pendant 30min, puis centrifugé deux fois de suite à 5000 tr/min pendant 15 min. Le surnageant obtenu est filtré sur un papier filtre standard (DURIEUX, réf: 66301130). (Derouiche, 2002 modifié). 2-2-4- Préparation du jus de tomate : La tomate est lavée puis broyée. Le broyat obtenu est centrifugé à 5000 tr/min pendant 15 min afin de séparer des débris cellulaires. Le surnageant recueilli est filtré sur un papier filtre standard (DURIEUX, réf: 66301130). 2-2-5- Analyses physicochimiques : Les analyses physicochimiques ont été réalisées au niveau du laboratoire de contrôle de qualité et répression des fraudes (CACQE) d’Oran. Elles permettent de déterminer les paramètres physicochimiques du lactosérum et du jus de tomate selon des techniques standards officielles, décrites dans l’ouvrage de référence AFNOR, édition 1986. Les analyses des sels minéraux pour le jus de tomate ont été réalisées au niveau du laboratoire d’analyse de la qualité (EL-FETH).

Matériel et Méthodes
33
2-2-5-1- Mesure de l’acidité : Elle se fait immédiatement après prélèvement d’un échantillon de 30 ml de la culture bactérienne, réparti dans 3 béchers, à raison de 10ml chacun. On mesure d’abord le pH et on dose ensuite la quantité d’acide lactique par titration avec de la soude N/9 en présence de quelque goutes de phénophtaléine (à 1% dans l’éthanol). Le volume de soude utilisé pour obtenir le virage de la couleur au rose pâle est alors noté et les résultats sont exprimés en degrés Dornic suivant la relation :
Acidité °D = VNaOH . 10 avec VNaOH : volume moyen de la soude utilisé pour titrer 10ml.
2-2-5-2- Teneur en matière sèche totale : Le principe de cette méthode consiste à peser le résidu après dessiccation à l’étuve à plus de 103°C. Une capsule sèche et tarée contenant 5 ml de l’échantillon est pesée puis placée dans l’étuve à 103°C pendant 3h. Après refroidissement dans le dessiccateur la capsule avec le résidu sec est de nouveau pesée. Le taux de la matière sèche est calculé (en g/l ou en %) selon les formules :
M.S (g/l) = ((m2-m0)/5). 1000 M.S (%) = ((m2-m0)/(m1-m0)). 100
avec : m0 : masse de la capsule vide ; m1 : masse de la capsule avec l’échantillon ; m2 : masse de la capsule avec le résidu sec. 2-2-5-3- Teneur en cendres : Le principe de cette méthode réside dans l’incinération de la matière sèche de l’échantillon à 530°C. Dans une capsule calcinée et tarée, on introduit 5 ml de l’échantillon puis on procède à une préincinération en maintenant la capsule au dessus d’une petite flamme de bec Bunsen. Le résidu obtenu est alors incinéré dans le four à moufles réglé à 530°C jusqu’à disparition des particules charbonneuses. Ce résultat est généralement obtenu au bout de 2 à 4h. La capsule est pesée après refroidissement dans un dessiccateur. La teneur (en g/l ou en %) est donnée par les formules :
C (g/l) = ((m2-m0)/5). 1000 C (%) = ((m2-m0)/(m2-m0)). 100
avec: m0: masse de la capsule vide ; m1 : masse de la capsule avec l’échantillon ; m2 : masse de la capsule avec les cendres.

Matériel et Méthodes
34
2-2-5-4- Détermination de la matière grasse : • Dans le lactosérum : Le principe consiste dans un premier temps à faire dissoudre totalement les protéines, puis dans une deuxième étape à mesurer le taux de la matière grasse (après chauffage à 65°C) directement dans le butyromètre. 11 ml de lactosérum sont introduits dans un butyromètre contenant 10ml d’acide sulfurique. A ce mélange, on ajoute 1ml d’alcool isoamylique. On agite délicatement jusqu’à dissolution totale des protéines (aucune particule blanche). Par la suite, le mélange est centrifugé à une vitesse maximale pendant 10min. Après centrifugation, le butyromètre contenant le mélange est placé au bain Marie à 65°C pendant 5min. La matière grasse apparaissant avec une couleur jaunâtre dans la colonne du butyromètre est estimée comme suit :
M.G (g/l) = (B – A). 10 avec : A : valeur de la graduation au dessous de la colonne lipidique. B : valeur de la graduation au dessus de la colonne lipidique. • Dans le jus de tomate : Le principe consiste à traiter l’échantillon avec de l’acide chlorhydrique dilué bouillant pour libérer les fractions lipidiques. Après filtration et séchage de la masse résultante, l’extraction de la matière grasse retenue sur filtre est réalisée au moyen de l’hexane. 100ml de jus de tomate sont mis dans un ballon à col rodé, on y ajoute 50ml d’acide chlorhydrique (4N) et des billes en verre (stabilisateur d’ébullition). Après un chauffage pendant 1h sur une petite flamme, on filtre à chaud, puis on lave le papier filtre avec de l’eau chaude jusqu’à élimination totale de l’acide chlorhydrique (absence de précipité blanc après traitement du liquide de lavage par le nitrate d’argent). Le papier filtre est ensuite séché pendant 1h à 103°C. Après refroidissement, l’extraction de la matière grasse se fait pendant 4h en présence du solvant d’extraction l’hexane. Après élimination de l’hexane, le tout est séché à 103°C pendant 1h. La teneur en matière grasse totale est donnée par la formule : (m2 – m1). 100/m0
avec : m0 : masse en gramme de la prise d’essai m1 : masse en gramme de la fiole vide. m2 : masse en gramme de la fiole et la matière grasse après séchage.
2-2-5-5-Teneur en glucides : Elle est estimée en réalisant un dosage par la méthode de Bertrand, qui repose sur les propriétés réductrices des glucides. Le dosage se déroule en trois étapes : Ø Réduction de la liqueur de Fehling par les glucides réducteurs : (1) ; 2 glucides réducteurs + 2n Cu2+ + 2n OH- à 2 produits d’oxydation + n Cu2O + n électron

Matériel et Méthodes
35
Ø Isolement du cuivre formé : (2); Cu2O + 2 H+ + 2Fe3+ à 2 Cu2+ + H2O + 2 Fe2+ Ø Dosage du cuivre par manganimétrie: Grâce à l’équation bilan (2) on peut écrire ; n (Cu2+) = n (Fe2+) 5 Fe2+ + MnO4
- + 8 H+ à Mn2+ + 4 H2O + 5 Fe3+
On procède de la manière suivante:
- 20ml de l’échantillon à traiter sont mis dans un ballon à col rodé, on y ajoute 50ml d’eau distillée et 20 gouttes de HCl concentré.
- après chauffage au bain Marie bouillant pendant 4h, on filtre puis on neutralise avec NaOH 30% en présence de phénolphtaléine (1%) jusqu’à l’obtention d’une coloration rose.
- on y ajoute 2ml de ferrocyanure de K (10,6%) et 2ml d’acétate de zinc (30%), et on complète avec l’eau distillée jusqu’à 100ml. On laisse reposer à température ambiante pendant 15min puis on filtre.
- à 20ml du filtrat obtenu on ajoute 20ml de liqueur tartrique et 20ml de liqueur cuivrique. Après un chauffage pendant 3 min sur une petite flamme, l’oxyde cuivreux rouge se dépose au fond de l’erlen et la couleur bleue de la liqueur surnageante indique la présence d’un excès de cuivre.
- On filtre la liqueur surnageante avec un entonnoir en verre fritté (N°4), en ayant soin d’entrainer le moins possible le précipité. On dissout entièrement l’oxyde cuivreux par la liqueur ferrique, ce qui donne une liqueur vert limpide qui est filtrée puis titrée par du permanganate de K (0,1N), jusqu’à virage de la couleur au rose. Le résultat est déduit d’une table établie expérimentalement par Bertrand, qui relie la quantité de cuivre isolé à celle des glucides dosés (Annexe). 2-2-6- Optimisation du milieu de culture : Le lactosérum déprotéiné est supplémenté en différentes quantités d’extrait de levure (Difco) pour obtenir le milieu LY, ou en jus de tomate fraiche (pH est ajusté à 5,7 avec NaOH ou HCl 5N) pour obtenir le milieu LT. La supplémentation en extrait de levure et en jus de tomate permet d’obtenir le milieu LYT (tableau 6). Ces milieux sont stérilisés à 120°C pendant 15min. Des cultures jeunes (DO~1) ont été ensemencées à 1% dans 40ml de chaque milieu. Après incubation de 24h à 30°C, la croissance bactérienne est estimée par mesure de la densité optique (DO) à 600nm à l’aide d’un spectrophotomètre et par un dénombrement microbien sur milieu MRS solide incubé à 30°C pendant 48h (les résultats obtenus sont exprimé en U.F.C/ml).

Matériel et Méthodes
36
Tableau 6
Enrichissement du lactosérum par l’extrait de levure ou le jus de tomate Lactosérum
Extrait de levure (% p/v)
0
1
2
3
4
5
6
7
8
9
10
Jus de tomate (% v/v)
0
5
10
15
20
25
30
35
40
45
50
Ce milieu optimisé LYT a été utilisé pour mesurer divers paramètres technologiques en comparaison avec le milieu MRS et le lactosérum (non complémenté).
• Mesure de l’acidité produite : L’acidité est mesurée par pH métrie et par titration. Sous agitation mécanique on ajoute goutte à goutte de la soude (N/9) tout en mesurant le pH. Dès que la valeur du pH atteint celle du pH initial on note le volume de la soude utilisée. Les résultats sont exprimés en degrés Dornic.
Acidité = n . 10 n = volume de la soude utilisé pour atteindre la valeur du pH initial.
1°D = 0,1 g/l d’acide lactique produit
• Recherche du diacétyle : 1 ml du milieu ensemencé est placé dans un tube à hémolyse stérile, la réaction de Voges Proskauer est effectuée en ajoutant 1 ml de réactif α-naphtol à 6% dans l’alcool absolu (VPI) et 1ml de la soude (NaoH) à 16% dans l’eau distillée (VPII). Après agitation des tubes et un délai de quelques minutes à température ambiante, une coloration rose traduit la présence de l’acétoine. Cette substance se transforme en diacétyl sous l’action de la soude qui se combine avec l’ α-naphtol en un complexe rouge.
• Mise en évidence de l’activité protéolytique cellulaire : L’activité protéolytique est recherchée en milieu solide (MRS, LYT) tamponné à pH7 par le tampon phosphate (KH2PO4 / Na2 HPO4 0,1M) additionné de 1% de lait écrémé stérile reconstitué à 10%. Selon la méthode décrite par Van Den Berg et al. (1993), les souches bactériennes à tester sont ensemencées simultanément en touches à la surface du milieu de culture à l’aide de l’inoculateur multipoint. Après incubation à 30°C pendant 48h, l’activité protéolytique est révélée par l’apparition d’un halo clair autour de chaque colonie bactérienne. La dimension de ce halo est mesurée.

Matériel et Méthodes
37
2-2-7- Cinétique d’apparition d’activité protéolytique : Parallèlement à la croissance bactérienne, nous avons suivi l’activité protéolytique de deux souches (BH14 et CHTD27) en fonction du temps afin de déterminer à quel moment de la croissance sont synthétisées les enzymes protéolytique dans chaque milieu de culture. Pour cette étude nous avons utilisé 100ml de milieu (MRS, LYT) contenant 2% de lait écrémé stérile reconstitué à 10%, ce dernier est inoculé à 1% avec une culture bactérienne jeune. Toutes les deux heures, 1ml de la phase claire de la culture est récupérée et centrifugée à 12.000 tr/min pendant 10min à 4°C. Les surnageants recueillis sont testés pour leur activité enzymatique par la méthode des puits : 50μl de chaque surnageant est déposé dans un puits creusé dans une gélose au lait à 2% contenant 0,02% d’azide de sodium (NaN3). L’hydrolyse des caséines du lait se manifeste après 48 h d’incubation à 30°C, par l’apparition d’un halo clair autour des puits contenant l’extrait enzymatique (Chantawanakul et al., 2002). Une courbe d’étalonnage (A = F[C]) est établie en utilisant une gamme de concentration de trypsine (la trypsine 2µg/ml est préparée dans un tampon phosphate stérile 0,1M, pH7) de 0 ; 0,06 ; 0,125 ; 0,25 ; 0,5 ; 1 ; 2 µg/ml. La concentration d’enzyme protéolytique des échantillons est déterminée en rapportant les diamètre des zones de clarification sur la courbe étalon. 2-2-8 - Culture en fermenteur : Nous avons réalisé des cultures en conditions de pH non contrôlé, de deux souches BH14 et CHTD27 dans un fermenteur (OMNI CULTURE VIRTIS) avec une capacité de 2 litres. Un volume de 1 litre de milieu est préparé (pH 5,7), transvasé dans la cuve en verre du fermenteur et l’ensemble est stérilisé par autoclavage à 120°C pendant 15 à 20 min. Après refroidissement, et afin de vérifier la stérilité du milieu, la cuve est placée sur son support, la température est maintenue constante à 30°C, et l’agitation est réglée à 75 tr/min pendant 24 heures. Le fermenteur est ensuite inoculé à l’aide d’un injecteur automatique par une préculture fraiche (DO~1). La taille de l’inoculum représente 1% du volume total. L’évolution de la biomasse, la production d’acide lactique et l’apparition d’acétoïne dans le milieu, sont suivis par prélèvements d’échantillons toutes les deux heures. Le dosage du sucre est réalisé au début et à la fin de fermentation.

3 - RESULTATS

Résultats et Discussion
39
3-1- Vérification de la pureté des souches et leur appartenance au groupe lactique : Après culture sur milieu MRS solide, nous avons observé des petites colonies blanchâtres, de diamètre compris entre 1 et 4 mm. Ces colonies ont un contour régulier ou irrégulier selon les souches. L’examen microscopique a révélé que toutes les bactéries étudiées sont Gram positif. Elles se présentent sous forme de bâtonnets à bouts arrondis ou carrés ou de coccobacilles. Le test de catalase est négatif pour toutes les souches. Tous ces résultats confirment l’appartenance des souches utilisées dans cette étude au groupe lactique et appartenant au genre Lactobacillus. 3-2- Analyses physicochimiques du lactosérum et du jus de tomate :
L’utilisation du lactosérum doux comme milieu de culture pour les lactobacilles, nécessite au préalable une connaissance de sa composition physicochimique. Ceci permettra par la suite de réaliser des essais de supplémentation afin d’obtenir un milieu de culture adéquat. Comme le montre la figure 12, les deux types de lactosérum (brut et déprotéiné) ne présentent pas de grande différence en matière sèche totale. On remarque que le lactose est le constituant le plus important du lactosérum brut, il représente plus de 60% de l’extrait sec. Dans le lactosérum déprotéiné la concentration de ce dernier a diminué de 6,08g/l par rapport au lactosérum brut, la même remarque peut être faite pour les cendres où on note une diminution de 2,29 g/l. Par contre, on remarque une disparition totale de la matière grasse dans le lactosérum déprotéiné. La diminution en lactose est peut être due à son hydrolyse thermique à 100°C, en revanche la matière grasse a probablement sédimenté sous l'effet du champ de pesanteur avec les protéines. L’α-lactoglobine est une des protéines majoritaire dans le lactosérum ; cette dernière est une métalloprotéine (protéine qui a la propriété de fixer spécifiquement des ions) plus particulièrement le calcium (Jacquet-Violleau, 1999), ce qui explique la diminution du taux de cendres.
Figure 12 : Analyse physicochimique du lactosérum
0102030
40506070
8090
Lactosérum brut Lactosérum déprotéiné
pHAcidité (°D)
Matière séche total (g/l)
Lactose (g/l)
Cendres (g/l)
Matière grasse (g/l)

Résultats et Discussion
40
Pour le jus de tomate (tableau 7), on observe qu’il est beaucoup moins riche en matière grasse, possédant un pH acide (3,95), et une teneur moyenne en glucides (25,73g/l) et en les cendres (3,72g/l) par rapport au lactosérum. Le potassium est le principal minéral présent dans les cendres avec plus de 238 mg/l. Selon Cotte (2000), les sucres sont essentiellement représentés par le glucose et le fructose. L’acidité augmente avec la maturité du fruit, elle est due à la présence des acides organiques tels que l’acide malique et citrique. On trouve aussi des vitamines (acide ascorbique, vitamines A, E et B), en quantités notables.
Tableau 7 : Analyse physicochimique du jus de tomate
Constituant (g/l)
Jus de tomate
pH Acidité Matière sèche totale Matières grasses Glucides Cendres ; Calcium (mg/l) Chlore (mg/l) Fer (mg/l) Potassium (mg/l) Magnésium (mg/l) Sodium (mg/l) Phosphore (mg/l) Soufre (mg/l) Zinc (mg/l)
3,95 ND ND
0,044 25,73 3,72
9,31
27,99 0,55
238,19 8,43 4,08
25,11 13,29 0,18
ND : non déterminé. 3-3- Optimisation du milieu de culture à base de lactosérum :
Nous avons démarré cette étude avec deux lactobacilles, Lb. plantarum BH14 et Lb. brevis CHTD27 (Benaissa et Chouiref, 2007 ; Belkheir, 2004) afin de comparer l’effet de l’addition de l’extrait de levure (E.L) et/ou du jus de tomate (J.T) dans le lactosérum déprotéiné sur la biomasse. Nous avons ensuite voulu généraliser l’utilisation de ce milieu optimisé à base de lactosérum en le testant sur la croissance d’une trentaine d’autres souches de Lactobacillus.

Résultats et Discussion
41
3-3-1- Croissance en milieu LY (Lactosérum + extrait de levure) : Les résultats obtenus (figure 13) montrent clairement que la production de la biomasse a été améliorée par rapport au lactosérum non supplémenté. On remarque que la production de la biomasse de CHTD27 est proportionnelle à la quantité d’extrait de levure additionnée, jusqu’à des concentrations supérieures à 8%, puis elle se stabilise. Pour BH14 la biomasse continue d’augmenter jusqu’à 10% en extrait de levure. On note des différences de biomasse entre les deux souches pour une même concentration en E.L ; par exemple pour la souche BH 14 à une concentration de 7% on obtient 1,3. 108
ufc/ml, alors que pour la souche CHTD27 on atteint 2,12. 108 ufc/ml. Ceci est dû probablement à l’effet souche.
Figure 13 : Croissance en milieu LY
L’effet stimulant de E.L sur la croissance bactérienne a été souvent rapportée (Barallé et Borzani, 1988 ; Amrane et Prigent, 1993 ; Champagne et al., 1999 ; Nancib et al., 2005, Djeghri-Hocine et al., 2007). En revanche Boudjemaa (2008) et Ghaly et al. (2003) ont signalé qu’à des concentrations élevées en extrait de levure, la concentration cellulaire diminue sous l’effet d’une forte toxicité de l’extrait de levure. Du point de vue du coût du milieu, l’utilisation d’un milieu de culture avec des telles concentrations en E.L n’est pas rentable. C’est pour cette raison que nous avons substitué l’extrait de levure par le jus de tomate.
3-3-2- Croissance en milieu LT (Lactosérum + jus de tomate)
La figure 14 montre les résultats obtenus. Les mêmes remarques que précédemment peuvent être faites. L’addition du jus de tomate à des concentrations croissantes stimule la biomasse bactérienne proportionnellement, exception faite pour la souche BH14 où nous avons enregistré un maximum de croissance en présence de 35% (v/v) de jus de tomate (milieu LT7). Une diminution de la biomasse est observée pour des quantités supérieures en jus de tomate (milieux LT8, LT9 et LT10).

Résultats et Discussion
42
Ceci pourrait s’expliquer par la réduction de l’activité de la β galactosidase à cause de la forte concentration des acides trouvés dans le jus de tomate. Dans le même ordre d’idées, Inagawa et al (1987) ont montré que l’acide phytique trouvé dans le sirop de maïs à une concentration de 4,4% réduit l’activité de la β-galactosidase in vivo, et par conséquent l’assimilation du lactose par la bactérie.
Figure 14 : Croissance en milieu LT
Des différences légères de biomasse entre les deux souches sont observées pour une même concentration de J.T : ainsi en milieu LT7 on obtient 8,5.107 ufc/ml pour la souche BH14, et 9,3.107 ufc/ml pour la souche CHTD27. Nos observations concordent bien avec les travaux de Babu et al. (1992), qui ont montré que l’addition du jus de tomate dans le lait écrémé stimule la croissance des lactobacilles. Cependant, la quantité de la biomasse produite dans le milieu enrichi avec le J.T est nettement plus faible que celle obtenue avec EL : ceci est probablement dû à une carence en source d’azote dans le J.T. Pour pallier à ces limitations et dans le but d’obtenir une bonne productivité de la biomasse, nous avons réalisé des essais de croissance des bactéries dans un milieu à base de lactosérum contenant les deux suppléments (extrait de levure et jus de tomate).
Le lactosérum est additionné de quantités croissantes de jus de tomate : 1 - LT1 : 5 % de jus de tomate 6 - LT6 : 30 % de jus de tomate 2 - LT2 : 10 % de jus de tomate 7 - LT7 : 35 % de jus de tomate 3 - LT3 : 15 % de jus de tomate 8 - LT8 : 40 % de jus de tomate 4 - LT4 : 20 % de jus de tomate 9 - LT9 : 45 % de jus de tomate 5 - LT5 : 25 % de jus de tomate 10 - LT10 : 50 % de jus de tomate

Résultats et Discussion
43
3-3-3- Croissance en milieu LYT (lactosérum + extrait de levure + jus de tomate) A partir des résultats obtenus précédemment, nous avons retenu le milieu LT6 contenant 30% du jus de tomate, car au-delà de cette concentration le milieu devient trouble, ce qui influence la lecture spectrophotométrique. Par ailleurs, nous nous sommes limités à une concentration maximale de 1% EL, car à cette concentration on a une bonne production de biomasse. Amrane (2000) a travaillé sur la production d’acide lactique par Lactobacillus helveticus cultivée sur lactosérum supplémenté par EL, et il conclue que les concentrations d’extrait de levure supérieures à 20 g/l deviennent toxiques pour le microorganisme. De même, Ghaly et al. (2003) ont signalé qu’à des concentrations élevées en extrait de levure, la concentration cellulaire diminue sous l’effet de la forte toxicité de l’extrait de levure. Bouguettoucha et al. (2007), Kulozik et Wilde (1999), Amrane et Prigent (1997), Aeshlimai (1989) , qui ont travaillé sur une bactérie lactique thermophile libre Lactobacillus helveticus, - ainsi que Norton et al.(1994), Schepers et al.(2006) et Sirisanneyakul et al.(2007) - qui ont travaillé respectivement sur des bactéries lactiques thermophiles immobilisées Lactobacillus lactis IO1, Lactobacillus helveticus - ont jugé que 10 g/l d’extrait de levure est la concentration optimale pour ces souches afin qu’elles puissent donner une bonne croissance et production d’acide lactique. Nous avons donc réalisé des essais de croissance de la souche BH14 en milieu LT6 additionné de 0,5 ou de 1 % d’extrait de levure. Les résultats obtenus sont représentés dans la figure 15.
Figure 15 : Croissance en milieu LYT de la souche BH14
Milieu MRSm : Milieu MRS modifié où la source de carbone est le lactose Milieu LAC : Lactosérum déprotéiné Milieu LT6 : lactosérum + 30 % de jus de tomate Milieu M1 : LT6 + 0,5 % d’extrait de levure Milieu M2 : LT6 + 1 % d’extrait de levure
Les résultats obtenus montrent clairement que la production de la biomasse bactérienne a été améliorée après l’addition d’extrait de levure dans le milieu LT6. L’utilisation du milieu M2 a conduit à des croissances bactériennes proches de celles observées en milieu MRSm.

Résultats et Discussion
44
L’ensemble des résultats précédents nous a mené à retenir le milieu M2 (lactosérum + 30% jus de tomate + 1% d’extrait de levure) pour la suite de l’étude. 3.3-4- Croissance des souches de lactobacilles en milieu M2 : Nous avons commencé cette étude en examinant la croissance des souches BH14 et CHTD27 en milieu M2. La figure 16 regroupe les résultats obtenus pour ces deux souches.
Figure 16 : Croissance des souches BH14 et CHTD27 dans les différents milieux Les remarques suivantes peuvent être faites : - les croissances bactériennes observées en milieu MRSm et M2 pour une même souche sont similaires. - sur milieu LAC les croissances sont nettement plus faibles qu’en milieu MRSm et milieu M2. Ce résultat était prévisible. Au vu de ces résultats, obtenus avec deux souches de lactobacilles, nous avons voulu généraliser l’étude en testant la croissance en milieu M2 de toutes les souches de lactobacilles, toujours en comparaison avec les deux milieux témoins MRSm et LAC.
Les résultats sont montrés dans la figure 17.

Résultats et Discussion
45
Figure 17: Croissance des lactobacilles en milieu M2 (MRSm et LAC sont des milieux témoins)
V7 V8V14 V20 V21 V23
LVK9
LVK11
LVK12
LVK13
LVK14
LVK15
LVK29
LVK30
LVK31
LVK320
1
2
3
4
5
6
7
8
DO
600
nm
Souches
MRSmM2
LAC
Figure 17 (suite): Croissance des lactobacilles en milieu M2 (MRSm et LAC sont des milieux témoins)
Ces résultats conduisent à faire deux remarques:
CHM9
CHM11
CHM12
CHM16
CHM17
CHM18
CHM19
CHM20BH21
BH14 H3
CHTD25
CHTD27
CHTD29 9 b0
1
2
3
4
5
6
7
8
DO
600
nm
Souches
MRSmM2LAC

Résultats et Discussion
46
- la première remarque est que toutes les souches testées croissent dans le milieu M2 et atteignent des densités cellulaires importantes (1.5<DO600nm<8) ce qui indique que le milieu M2 leur est favorable. - la seconde remarque est que la croissance est aussi fonction de la souche considérée : en effet on peut distinguer trois catégories de souches selon leur croissance dans ce milieu :
• celles dont la croissance est nettement plus importante en milieu MRSm qu’en milieu M2 : c’est le cas de 9b, BH14, V7, V8, V14, V20, V21 et V23.
• celles présentant des croissances quasiment identiques dans les deux milieux : c’est le cas des souches BH21, CHM9, CHM11, CHM12, CHM16, CHM19, CHM20, CHTD25, CHTD27, H3, LVK29, LVK30 et LVK32.
• celles dont la croissance est plus importante en milieu M2 qu’en milieu MRSm : c’est le cas de CHTD29, LVK9, LVK11, LVK12, LVK13, LVK14, LVK15et LVK31.
Toutes ces observations indiquent que le milieu M2 peut donc être utilisé pour cultiver ces bactéries lactiques et nous avons voulu examiner si les principales caractéristiques technologiques de ces bactéries étaient exprimées dans ces conditions trophiques. 3-3-5 Caractéristiques technologiques des lactobacilles cultivés en milieu M2 : 3-3-5-1 Production d’acide lactique en milieu M2 : La production d’acide lactique par les bactéries cultivées en milieu M2 a été évaluée par titration. Les résultats obtenus sont représentés dans la figure 18.
Figure 18 : Production d’acide lactique des souches en milieu M2 (MRSm et LAC : Milieux témoins)

Résultats et Discussion
47
Figure 18 (suite) : Production d’acide lactique des souches en milieu M2 (MRSm et LAC : Milieux témoins)
- sur milieu M2, la souche CHTD27 donne une quantité d’acide lactique presque équivalente à celle produite sur milieu MRSm (5 et 5,5 g/l respectivement), mais une différence très significative est observée pour la souche BH14 où la quantité d’acide lactique produite en MRSm est le double de celle produite en milieu M2. - des différences sont également observées pour un même milieu et en fonction des souches ; en milieu M2 une quantité de 6,4 g/l d’acide lactique est produite par la souche BH14, contre 5 g/l pour la souche CHTD27.
Au vu de ces résultats on peut classer les souches en fonction de leur pouvoir acidifiant (Karam et Zadi-Karam, 1994), et on distingue alors :
• Souches fortement acidifiantes (acidité ≥ 7,9 g/l) sur les milieux M2 et MRSm : c’est le cas de BH14, 9b, V7, V8, V14, V20, V21, V23. La production d’acide lactique est liée à la croissance et elle est plus élevée en milieu MRSm ; c’est le cas par exemple de la souche BH14 qui produit 17,5g/l d’acide lactique en milieu MRSm et 9,7 g/l en milieuM2.
• Souches fortement acidifiantes sur un milieu et moyennement acidifiantes sur l’autre ; c’est le cas par exemple de la souche BH21 qui produit 8,5g/l d’acide lactique en milieu MRSm et 7 g/l M2.
• Souches moyennement acidifiantes (4 g/l<acidité<7,9 g/l) sur les milieux M2 et MRSm : c’est le cas de H3, CHTD25, LVK9, LVK11, LVK12, LVK13, LVK14, LVK15, LVK29, LVK30, LVK32 et CHM18. La production d’acide lactique est liée à la croissance et elle est plus élevée en milieu M2 ; c’est le cas par exemple de la souche LVK11 qui produit 4,3g/l d’acide lactique en milieu MRSm et 5,7 g/l en milieu M2.
• Souches moyennement acidifiantes (4 g/l<acidité<7,9 g/l) sur les milieux M2 et MRSm : c’est le cas de CHM9, CHM11, CHM12, CHM16, CHM17 et CHTD27. La production d’acide lactique est non liée à la croissance et elle est plus élevée en

Résultats et Discussion
48
milieu M2 ; c’est le cas par exemple de la souche CHM16 qui produit 4,6g/l d’acide lactique en milieu MRSm et 5,3 g/l M2.
• Souche moyennement acidifiante sur un milieu et faiblement acidifiante sur l’autre : c’est le cas de CHM19, CHM20, LVK31 et CHTD29. La production d’acide lactique est liée à la croissance et elle est plus élevée en milieu M2 : c’est le cas par exemple de la souche CHTD29 qui produit 0,5g/l d’acide lactique en milieu MRSm et 4,8 g/l en milieu M2.
• La production d’acide lactique en milieu LAC est très faible pour toutes les souches, à l’exception des souches V7, V8, V14, V20 et BH14 qui sont moyennement acidifiantes. La valeur la plus élevée est enregistrée pour la souche V8 (5,2 g/l).
Ces résultats concordent avec les observations rapportées par plusieurs auteurs quant à l’effet de l’extrait de levure et aux différences de pouvoir acidifiant des souches. Amrane et Prigent (1998), Kulozik et Wilde (1999) et Sherpers et al. (2002), ont montré que pour une utilisation complète du lactose, il est indispensable de supplémenter le lactosérum en sources azotées. Selon Zhang et al. (2007), l’addition d’extrait de levure au lactosérum déprotéiné a considérablement augmenté non seulement le taux d’acidification par Lactobacillus bulgaricus 11842 mais aussi l’activité de la β-galactosidase. Cette activation entraine la dégradation de lactose et une bonne assimilation du glucose qui se traduit par une bonne croissance. Karam (1995), Benaissa et Chouiref (2007), Dellali (2008) et Benasla et Benchagra (2009) au laboratoire ont montré une différence d’acidification des souches. L’activité acidifiante est spécifique de chaque souche et dépend de sa capacité à dégrader et à assimiler les substances dans le milieu. 3-3-5-2 Production de diacétyle : La production de diacétyle a été recherchée chez toutes les souches sur les trois milieux testés. La figure 19 montre l’exemple de la souche BH14. On peut remarquer que l’intensité de la couleur rouge, qui reflète la quantité de diacétyle, produite sur milieu LAC est plus forte que sur les milieux M2 et MRSm. Ceci est noté pour toutes les souches produisant du diacétyle et peut s’expliquer par la présence de fortes concentrations de citrate dans le lactosérum. Le tableau 8 montre les résultats obtenus pour toutes les souches.
Figure 19 : Production de diacétyle par la souche BH14
T BH14 T BH14 T BH14 (MRSm) (MRSm) (M2) (M2) (LAC) (LAC)

Résultats et Discussion
49
Tableau 8 : Production de diacétyle
+ : réaction positive ; ++ : réaction positive intense, - : réaction négative Nous n’avons pas observé de différence entre les résultats obtenus en milieu MRSm ou en milieu M2. Toutes les souches sont productrices de diacétyle à l’exception de 5 souches (BH21, H3, CHTD25, CHTD27, CHTD29) qui n’en produisent pas quel que soit le milieu. Le résultat concernant la souche CHTD27 a été aussi observé par Benasla et Benchegra en 2009.
3-4-Activité protéolytique en milieu M2 : L’activité protéolytique des lactobacilles est une caractéristique technologique importante, en particulier dans l’affinage des fromages. 3-4-1 Recherche de l’activité protéolytique en milieu M2 : Nous avons recherché et comparé cette activité chez nos souches sur les trois milieux MRSm, LAC et M2, additionnés de 1% de lait écrémé reconstitué 10% (m/v). Les résultats obtenus sont montrés dans la figure 20.
Milieu Souche
MRSm M2 LAC Milieu Souche
MRSm M2 LAC
CHM9 + + ++ V8 + + ++ CHM11 + + ++ V14 + + ++
CHM12 + + ++ V20 ++ ++ ++ CHM16 + + ++ V21 + + ++ CHM17 + + ++ V23 + + ++ CHM18 + + + LVK9 + + ++ CHM19 + + + LVK11 + + ++ CHM20 + + + LVK12 + + ++ BH21 - - - LVK13 + + ++ BH14 + + ++ LVK14 + + ++
H3 - - - LVK15 + + ++ CHTD25 - - - LVK29 + + ++ CHTD27 - - - LVK30 + + ++ CHTD29 - - - LVK31 + + ++
9b + + ++ LVK32 + + ++ V7 + + ++ CHTD27 - - -

Résultats et Discussion
50
Figure 20 : Activité protéolytique des souches en milieu M2 (MRSm et LAC : Milieux témoins)
Figure 20 (suite) : Activité protéolytique des souches en milieu M2 (MRSm et LAC : Milieux témoins)
On peut faire les observations suivantes : - Aucune souche n’exprime d’activité protéolytique en milieu LAC. - Toutes les souches présentent une activité protéolytique en milieu MRSm, qui se manifeste
par un halo clair autour de la colonie, à l’exception la souche CHTD29. Les valeurs des rapports h/c [h : diamètre du halo clair (mm) ; c : diamètre de la colonie (mm)] varient entre 1,75 et 5.
- Les mêmes souches testées sont protéolytiques en milieu MRSm mais ne le sont pas en milieu M2 (Figures 21 et 22).

Résultats et Discussion
51
- Les résultats obtenus montrent que l’activité protéolytique dépend en partie de la composition du milieu de culture. En effet on remarque que 4 souches seulement (CHM18, CHM19, CHM20, CHTD25) présentent une activité protéolytique en milieu M2, et les valeurs du rapport (h/c) se rapprochent de celles enregistrées en MRSm, à l’exception de la souche CHM20 où elle est nettement plus élevée en milieu M2 (5,5 en M2 et 3,7 en MRSm).
Ces observations indiquent qu’il existe dans le milieu MRSm un ou plusieurs éléments qui stimulent cette activité protéolytique. Il a été signalé par de Roissart (1994), que la vitesse globale d’une réaction enzymatique ou d’un enchainement de réactions enzymatiques dépend, d’une part, des concentrations en enzymes, coenzymes et substrats et d’autre part des niveaux de pH, de température et de viscosité du milieu. Ces différents facteurs interviennent dans la régulation du métabolisme. L’étude de l’activité protéolytique ayant été réalisée, pour les deux milieux de culture, dans les mêmes conditions de pH, de température et de concentration en substrat (1%), il nous restait donc à considérer les cofacteurs importants comme le Magnésium (Mg) et le Manganèse (Mn), qui sont présents dans le milieu MRSm et qui jouent un rôle très important dans l’expression de l’activité protéolytique (de Roissart, 1994). Le paramètre ‘’viscosité des milieux’’ est négligeable dans notre cas, car les milieux testés ne sont pas visqueux. Par ailleurs on sait aussi que l’acide citrique stimule la production des protéases (Kojic et al., 1991).
Figure 21: Activité protéolytique en milieu
solide MRSm-lait (1%)
Figure 22: Activité protéolytique en milieu
solide M2-lait (1%)

Résultats et Discussion
52
En vue d’optimiser l’activité protéolytique des lactobacilles en milieu M2, nous avons donc modifié sa composition, en testant l’effet de l’addition du Mg, du Mn et du citrate. Nous avons retenu les mêmes concentrations de ces composés que celles trouvées dans le milieu MRS. 3-4-2 Optimisation de l’activité protéolytique en milieu M2 : Dans un premier temps, nous avons utilisé 3 souches, V7, CHM9 et CHTD27, pour cette étude. Les résultats obtenus sur milieu M2 et milieu M2 supplémenté sont montrés comparativement au milieu MRSm, respectivement dans les figures 23, 24 et 25.
0
2
4
6
Milieu
DO 600nm
Rapport (h/c)
Figure 23: Activité protéolytique de la souche V7
0
1
2
3
Milieu
DO 600nm
Rapport (h/c)
Figure 24: Activité protéolytique de la souche CHM9

Résultats et Discussion
53
01234
5
Milieu
DO 600nm
Rapport (h/c)
Figure 25: Activité protéolytique de la souche CHTD27
Les résultats obtenus confirment que l’activité protéolytique dépend bien de la composition du milieu de culture. En effet on remarque que toutes les souches testées sur le milieu M2 enrichi par le Mg, le Mn et le citrate présentent une activité protéolytique qui se manifeste par un halo clair autour de la colonie ensemencée. On remarque aussi que le milieu M2 additionné de magnésium et de manganèse, donne sensiblement les mêmes valeurs (rapports h/c) que celles obtenues en MRSm. On peut donc retenir le milieu M2 supplémenté suivant comme milieu conduisant à des résultats se rapprochant le plus de ceux observés en milieu MRSm La composition de ce nouveau milieu à base de lactosérum doux, nommé milieu BMK est, pour 1 litre :
Lactosérum 700ml Jus de tomate 300ml Extrait de levure 10g Sulfate de magnésium 0,1g Sulfate de manganèse 0,05g
Dans une deuxième étape, et pour généraliser les résultats précédents, nous avons recherché l’activité protéolytique chez toutes les autres bactéries en milieu BMK, toujours comparativement avec le milieu MRSm.

Résultats et Discussion
54
Les résultats obtenus sont montrés dans les figures 26, 27 et 28: on observe sans équivoque que l’activité protéolytique se manifeste de la même manière dans les deux milieux de culture testés. Les rapports (h/c) sont pratiquement identiques dans les deux milieux de culture.
Figure 26 : Activité protéolytique des lactobacilles en milieu MRSm et BMK.
Figure 27: Activité protéolytique en milieu
solide MRSm-lait (1%)
Figure 28: Activité protéolytique en milieu
solide BMK-lait (1%)

Résultats et Discussion
55
3-4-3- Cinétique de production de protéases en milieu BMK : Les résultats précédents nous ont conduits à examiner la cinétique d’apparition des enzymes protéolytiques dans le milieu BMK, en comparaison avec le milieu MRS, parallèlement à la croissance bactérienne. Cette étude a été réalisée avec les deux bactéries BH14 et CHTD27. Nous avons recherché les enzymes protéolytiques dans les surnageants de centrifugation des milieux de culture, par la méthode des puits comme décrit dans ‘’Matériels et méthodes’’. Les figures 29, 30, 31 et 32 montrent l’évolution de la croissance et de la libération des protéases en fonction du temps. La figure 33 montre un exemple de résultats obtenus en milieu solide agar-lait.
Conc
entr
atio
n de
s pr
otéa
ses
(mg/
ml)
0
1
2
3
4
5
6
7
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30Temps (h)
DO
600
nm
0
0,02
0,04
0,06
0,08
0,1
0,12Protéases DO
Figure 29 : Cinétique de croissance et libération des protéases de la souche BH14 sur milieu MRSm
Conc
entr
atio
n de
s pr
otéa
ses
(mg/
ml)
0
1
2
3
4
5
6
7
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30Temps (h)
DO
600
nm
00,010,020,030,040,050,060,070,080,09
Protéases DO
Figure 30: Cinétique de croissance et libération des protéases de la souche BH14 sur milieu BMK
Conc
entr
atio
n de
s pr
otéa
ses
(µg/
ml)
Conc
entr
atio
n de
s pr
otéa
ses
(µg/
ml)

Résultats et Discussion
56
Conc
entr
atio
n de
s pr
otéa
ses
(mg/
ml)
0123456789
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30Temps (h)
DO
600
nm
0
0,02
0,04
0,06
0,08
0,1
0,12
Protéases DO
Figure 31: Cinétique de croissance et libération des protéases de la souche CHTD27 sur milieu MRSm
Conc
entr
atio
n de
s pr
otés
es (m
g/m
l)
0
1
2
3
4
5
6
7
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30Temps (h)
DO
600
nm
00,010,02
0,03
0,040,050,06
0,070,080,09
Protéases DO
Figure 32: Cinétique de croissance et libération des protéases de la souche CHTD27 sur milieu BMK
Figure 33 : Activité protéolytique dans le surnageant de culture de la souche BH14 sur milieu agar-lait
(T : Témoin)
Conc
entr
atio
n de
s pr
otéa
ses
(µg/
ml)
Conc
entr
atio
n de
s pr
otéa
ses
(µg/
ml)

Résultats et Discussion
57
• On remarque que la production des protéases débute en phase exponentielle de croissance comme cela a été rapporté par Back et al. (1983), Hebert et al. (1996) et Dellali (2008). On constate qu’il faut atteindre une certaine concentration cellulaire pour qu’il y ait production de ces enzymes.
• Le temps d’apparition de l’activité protéolytique varie selon les souches et le milieu de culture. La libération des protéases par la souche BH14 débute au bout de 10h sur milieu MRSm et au bout de 12h sur milieu BMK. La même constatation peut être faite pour la souche CHTD27, les protéases sont produites au bout de 10h sur milieu MRSm et à 14h sur milieu BMK.
• La production des protéases des deux souches sur les deux milieux augmente en fonction du temps, au fur et à mesure que la croissance augmente : cela indique qu’elle est liée à la croissance.
• On observe aussi que l’activité protéolytique continue pendant la phase stationnaire.
L’ensemble de ces résultats indiquent que le milieu BMK peut tout à fait être substitué aux milieux MRS ou MRSm pour cultiver des lactobacilles et rechercher des propriétés d’intérêt technologiques. Ceci nous a encouragés à tester ce milieu en condition de fermenteur.
3-5- Cinétique de croissance, d’acidification, de production de diacétyle et de consommation du substrat en fermenteur :
Pour cette étude nous avons retenu les souches BH14 et CHTD27.
3-5-1 Croissance et production d’acide lactique :
Les figures 34, 35, 36 et 37 montrent les cinétiques de croissance et d’acidification pour les deux souches cultivées en fermenteur en mode discontinu dans les milieux MRS et BMK.
Figure 34 : Croissance, production d’acide lactique et évolution du pH en milieu MRSm de la souche BH14.

Résultats et Discussion
58
Figure 35 : Croissance, production d’acide lactique et évolution du pH en milieu BMK de la souche BH14.
0
1
2
3
4
5
6
02468
1012141618
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30
DO
(600
nm),
Aci
de la
ctiq
ue (g
/l)
Temps (h)
pH
D.O Acidité pH
Figure 36 : Croissance, production d’acide lactique et évolution du pH en milieu MRSm de la souche CHTD27.

Résultats et Discussion
59
0
1
2
3
4
5
6
0
1
2
3
4
5
6
7
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30
DO
(600
nm),
Aci
de la
ctiq
ue (g
/l)
Temps (h)
pH
D.O Acidité pH
Figure 37 : Croissance, production d’acide lactique et évolution du pH en milieu BMK de la souche CHTD27.
On peut faire les remarques suivantes :
- La croissance bactérienne, pour les deux souches et dans les deux milieux, se déroule selon un schéma classique d’une culture discontinue. Pendant la phase exponentielle et simultanément à la croissance bactérienne il ya production d’acide lactique : c’est le cas d’une production associée à la croissance. - Pour la souche CHTD27, la phase exponentielle de croissance débute plus tôt dans le milieu MRSm (après environ 6h) que dans le milieu BMK (après environ 8h), par contre pour la souche BH14, il n’ya pas de différence dans les deux milieux de culture, elle commence après 6h. On remarque aussi qu’après la phase exponentielle, il y a production d’acide lactique même si la croissance cellulaire est pratiquement arrêtée. Ceci s’explique par le fait qu’en fin de fermentation, la croissance s’arrête plus rapidement que la production d’acide lactique, car elle est plus sensible aux phénomènes d’inhibition par l’acide lactique. La production d’acide lactique devient dépendante de la concentration bactéri-enne et l’activité bactérienne existe encore pendant cette période. Les deux phéno-mènes sont alors découplés. Ces observations rejoignent celles faites par Engasser (1993), qui lors de la modélisation de la fermentation lactique par Lactobacillus casei, a montré que le glucose continue à être transformé en acide lactique alors que la croissance bactérienne est pratiquement arrêtée. - Les concentrations finales en acide lactique dans le milieu MRSm, pour les deux souches, sont plus élevées que celles produites dans le milieu BMK. Ceci confirme les résultats précédents pour ces deux souches.
Boudjelel et Nancib (2001) ont montré que l’addition du Tween 80 dans le milieu de culture stimulait la production d’acide lactique car le Tween 80 permet une meilleure excrétion du lactate par la cellule en créant des pores dans la membrane.

Résultats et Discussion
60
Amrane (1990) et White et al. (1976) ont montré qu’un milieu limitant en phosphore augmente considérablement la productivité volumique en acide lactique. Les auteurs expliquent ce résultat de la manière suivante : lorsque le phosphore est limitant, le pool d’ATP doit être rephosphorylé plus vite, ce qui entraine une production plus rapide de l’acide lactique.
3-5-2-Consommation du substrat : Le tableau 9 montre les concentrations du substrat dans les deux milieux au début (0h) et à la fin de la cinétique bactérienne (30h).
Tableau 9 : Concentration du substrat
Pour les deux souches après 30h de croissance sur milieu MRSm la totalité du substrat a été consommée, par contre dans le milieu BMK 32 g/l de substrat reste non consommé et la croissance est pourtant arrêtée. Ce résultat confirme bien que l’inhibition de la croissance est due à la forte acidité du milieu (pH bas) qui résulte de l’accumulation de l’acide lactique. Les travaux de Konings et Otto (1983) et Amrane (2001), ont montré que lorsque le pH du milieu extérieur diminue, par l’accumulation des acides organiques (acide lactique), la bactérie lactique maintient son pH intracellulaire plus alcalin (pH=6,6) que le milieu extracellulaire, car les cellules excrètent rapidement l’acide lactique dissocié dans le milieu extracellulaire. De plus la membrane cellulaire est imperméable pour les protons extracellulaires (molécule de lactate) (Hutkins et Nannen, 1993). La différence de pH entre le cytoplasme et le milieu extracellulaire forme un gradient de pH (∆pH). Pour excréter les protons à l’extérieur, la bactérie consomme l’ATP en activant l’enzyme H+, ATPase (Nannen et Hutkins, 1991), mais en fin de phase exponentielle de croissance, la quantité d’ATP disponible ne suffit plus, le gradient de pH disparaît, les activités hexokinasiques baissent et entrainent l’inhibition de la croissance et de l’activité des enzymes intracellulaires telles que la β-galactosidase.
Le tableau 10 regroupe les paramètres d’état des cultures. Ils ont été déterminés selon les données de la littérature (Deneuville, 1991).
Ces résultats permettent de faire les remarques suivantes :
• La comparaison des taux de croissance et des temps de génération montre que la souche BH14 a pratiquement le même taux de croissance (environ 1,9 h-1) et le même temps de génération (environ 0,36 h) dans les deux milieux. La croissance de la souche CHTD27 est plus rapide en milieu BMK (temps de génération 0,29).
Temps (h)
Concentration du substrat (g/l) BH14 CHTD27
MRS BMK MRS BMK t = 0 20 46,25 20 46,25
t = 30 0 32,4 0 32,7

Résultats et Discussion
61
• La productivité volumique horaire maximale de la biomasse et de l’acide lactique
sont plus élevée en milieu MRSm. Ceci confirme les observations précédentes.
• Le rendement global de transformation du substrat en biomasse (RX/S ) est plus faible pour les deux souches que celui de la transformation du substrat en acide lactique (RP/S ). Cela explique que le substrat est surtout utilisé pour la production d’acide lactique.
Tableau 10 : Paramètres d’état des cultures
μ : taux de croissance exprimés en heure-1. G : temps de génération exprimés en heure. Rs : vitesse maximum de consommation du substrat pendant la phase exponentielle, exprimée en grammes par litre et par heure.
W : pourcentage du substrat qui sert réellement à la fabrication d’acide lactique. Z : pourcentage du substrat
qui sert à la croissance et à la maintenance. Pvhglo(BIO) : productivité volumique horaire globale de la biomasse
exprimée en gramme par litre par heure biomasse sèche. Pvhglo(P) : productivité volumique horaire globale de l’acide lactique produite par fermentation homolactique associé à la croissance exprimés en gramme par litre
par heure. RP/S : Rendement global de transformation du substrat en acide lactique. RX/S : Rendement global de transformation du substrat en biomasse.
3-5-3-Production de diacétyle :
La production de diacétyle par la souche BH14 apparaît en même temps dans les deux milieux testés, au bout de 6h (Tableau 11), alors que le pH du milieu est de 5,5. Ce pH correspond au pH optimum d’activité de la citrate perméase (pH5,5-6 selon Marjo et al., 1991) et donc le citrate du milieu est dégradé activement dans ces conditions.
Tableau 11 : Production de diacétyle par la souche BH14
Temps (h)
Souche
0
2
4
6
8
10
12
14
16
18
20
22
24
26
28
30
BH14 Milieu MRSm - - - + + + + + + + + + + + + +
Milieu BMK - - - +/- + + + + + + + + + + + +
+ : production de diacétyle - : pas de production de diacétyle
Souches Paramètres
BH14 CHTD27 MRS BMK MRS BMK
μ 1,875 1,95 1,9 2,333 G 0,368 0,354 0,363 0,295
Pvhglo(BIO) 0,156 0,135 0,194 0,147 RX/S 0,235 0,292 0,291 0,311
Pvhglo(P) 0,516 0,330 0,505 0,178 RP/S 0,770 0,714 0,777 0,376

4 – CONCLUSION & PERSPECTIVES

Conclusion et Perspectives
63
Les analyses physico-chimiques du lactosérum doux issu de la fabrication du camembert et du jus de tomate fraîche montrent que ces derniers sont riches en matières nutritives et qu’ils présentent des qualités adéquates pour élaborer des milieux de culture de bactéries. Nos essais de culture sur lactosérum déprotéiné ont montré qu’il est trop pauvre en regard des besoins des lactobacilles. Son optimisation par ajout d’extrait de levure et/ou de jus de tomate a conduit à l’élaboration de milieux ‘’peu couteux’’ permettant la croissance de certains lactobacilles de manière similaire à celle obtenue en milieu MRS. Ces milieux ont été obtenus en ajoutant différentes quantités d’extrait de levure et /ou de jus de tomate fraîche au lactosérum déprotéiné, afin d’apporter les ingrédients nécessaires aux lactobacilles (vitamines, acides aminés,…), la source de carbone restant le lactose du lactosérum. Les premiers résultats obtenus montrent que la biomasse produite sur lactosérum additionné d’extrait de levure est supérieure à celle observée en présence de jus de tomate, mais dans les deux cas, la biomasse reste inférieure à celle obtenue en milieu témoin MRSm. Par contre, sur le milieu M2 [lactosérum additionné d’un mélange de jus de tomate et d’extrait de levure (à des concentrations 30% et 1% respectivement)], la biomasse obtenue est comparable à celle observée en milieu MRSm. Ce milieu M2 permet la production de diacétyle tout comme le milieu témoin. Cependant, il n’est pas suffisant pour l’expression de l’activité protéolytique bactérienne. Il était donc nécessaire de le supplémenter avec des cofacteurs, magnésium, manganèse, et du citrate afin de stimuler cette activité. Les résultats obtenus sur ce milieu M2 supplémenté ont montré que ces ajouts conduisent à une nette amélioration de l’activité protéolytique. L’optimisation de ce milieu avec l’utilisation d’une combinaison magnésium et manganèse à des concentrations de 0,1 et 0,05 g/l respectivement nous a permis d’élaborer le milieu BMK qui a conduit à des résultats similaires, voire meilleurs, de ceux obtenus en MRS. La comparaison du pouvoir acidifiant des bactéries en fonction du milieu de culture, à montré que ce dernier varie beaucoup d’un milieu à l’autre selon les bactéries considérées : parfois c’est sur le milieu MRSm qu’il y a une bonne production d’acide, parfois c’est sur le milieu BMK et parfois les résultats d’acidification sont identiques dans les deux milieux. Ceci indique et confirme l’effet souche. L’étude comparative des cinétiques d’acidification, de croissance, de consommation du substrat, de production de diacétyle et de protéase en fonction du temps, réalisée avec les souches BH14 et CHTD27, a montré que la croissance bactérienne se déroule selon un schéma classique d’une culture discontinue, pour les deux souches et dans les deux milieux. Pendant la phase exponentielle, et simultanément à la croissance bactérienne, il ya production d’acide lactique : c’est le cas d’une production associée à la croissance. Pour la souche CHTD27, la phase exponentielle de croissance débute plus tôt dans le milieu MRSm (environ au bout de 6h) que dans le milieu BMK (environ au bout de 8h); par contre pour la souche BH14 elle commence au bout de 6h pour les deux milieux. Pour les deux souches, la concentration finale en acide lactique est plus élevée en milieu MRSm qu’en milieu BMK. La comparaison des taux de croissance et des temps de génération montre que la souche BH14 a pratiquement le même taux de croissance (environ 1,9 h-1) et le même temps de

Conclusion et Perspectives
64
génération (environ 0,36 h-1) dans les deux milieux. La croissance de la souche CHTD27 est néanmoins plus rapide en milieu BMK (temps de génération 0,29 h-1) qu’en milieu MRSm. L’étude des cinétiques de protéolyse a montré que la production des protéases par les deux souches débute en phase exponentielle de croissance et continue pendant la phase stationnaire, elle est donc partiellement liée à la croissance. Le temps d’apparition de l’activité protéolytique varie selon les souches et le milieu de culture. La production de diacétyle par la souche BH14 apparaît en même temps, au bout de 6h. L’ensemble des résultats obtenus au cours de ce travail permet d’envisager plusieurs perspectives :
- Réaliser des essais supplémentaires concernant la substitution de l’extrait de levure par d’autres constituants à faible coût.
- Utiliser ce milieu BMK pour d’autres bactéries lactiques comme les lactocoques et les leuconostocs, en l’optimisant éventuellement.
- Tester le milieu BMK pour des cultures mixtes, en vue de mettre en évidence les interactions interbactériennes (inhibitions, stimulations)
- Tester le milieu en conditions de fermenteur en mode de fermentation en semi-continu et continu.
- S’orienter dans le sens de la production des métabolites en utilisant ce milieu.

5 – REFERENCES BIBLIOGRAPHIQUES

Abraham M.J et Srinivasen R.A. (1978). Utilisation of whey for production of microbial protein and lipid. J. Food Sci. Technol . 16 :11.
Aeschlimani A. (1989). Productions of lactic acid from whey permeate by Lactobacillus helveticus. Thése. Ecole Polytechnique Fédéral de Lausanne,165p. Adrian J. (1971). A propos des poudres de lait et des lactoprotéines levurées. Ind. Alim. Agric., 88 : 1607. AFNOR. (1986). Contrôle de la qualité des produits laitiers 3éme édition. P 107-343. Alais C. (1984). Sciences du lait. Principes des Techniques Laitières, SEPAIC, Paris, 4e edn.. Altaf M.D., Naveena B.J., Reddy G. (2007). Use of inexpensive nitrogen sources and starch for L(+) lactic acid production in anaerobic submerged fermentation. Bioresource Technology. 98: 498-503. Amrane A. (1991). Optimisation de la productivité d’un fermenteur effectuant le convertion en acide lactique de perméat de lactosérum supplémenté. Thése. Université de Renne I. France. Amrane A et Prigent Y. (1993). Influence of media composition on lactic acid production rate from whey by Lactobacillus helveticus. Biotechnology Letters. 15: 239-244. Amrane A. et Prigent Y. (1997). Growth and lactic acid production coupling for Lactobacillus helveticus. Cultivated on supplemented whey: influence of peptidic nitrogen deficiency. Journal of Biotechnology. 55: 1-8. Amrane A. et Prigent Y. (1998). Analysis of growth and production coupling for batch cultures of Lactobacillus helveticus with the help of an unstructured model. Process Bioche-mistry. 34: 1-10. Amrane A. (2000). Effect of inorganic phosphate on lactate production by Lactobacillus helveticus grown on supplemented whey permeate . Journal of chemical technology and biotechnology. 75: 223- 228. Amrane A. (2001). Lactic acid production during the associated and the deceleration growth phases of Lactobacillus helveticus cultivated in various conditions and media. Physiology, metabolism. Lait . 81: 91-103. Archibald F.S. (1986). Manganese: its acquisition by and function in the lactic acid bacteria. CRC Crit. Rev. Microbiol. 13: 63-109. Archibald F.S. et Fridovich I. (1981). Manganese and defences against oxygen toxicity in Lactobacillus plantarum. Journal of Bacteriology. 145: 442-451.

Atlan D., Laloi P., Portalier R. (1989). Isolation and caracterization of aminopeptidase- deficient Lactobacillus bulgaricus mutants. Appl Environ Microbiol. 55 :1717-1723. Babu V., Mital B.K. et Garg S.K. (1992). Effect of tomato juice addition on the growth and activity of Lactobacillus acidophilus. International Journal of Food Microbiology. 17: 67-70. Back Y.J., Yoon K.B., Yoon Y.H et Kim H.U. (1983). Hanguk Ch’uks an Hukhoechi. 25(1): 1-7. Barallé S.B et Borzani W. (1988). Use of yeast autolysate to improve batch lactic fermentation. Arquivos de biologia Y technologia. 31: 273-274. Belkheir K. (2004). Identification des souches de Lactobacillus isolées de lait de chamelle de Tindouf .Etude de leur activité protéolytique .Mémoire d’Ingénieur d’Etat en Biotechnologie. Université d’Oran, Algérie. Benaissa M. et Chouiref K. (2007). Comparaison de l’activité acidifiante de souches de Lactobacillus dans les laits de vache et de chèvres. Mémoire d’ingénieur d’état en Biotechnologie. Université d’Oran. Algérie. Benasla A. et Benchagra K. (2009). Effet des substrats carbonés sur la croissance et les caractéristiques technologiques de Lactobacillus. Mémoire d’ingénieur d’état en Biotechnologie. Université d’Oran. Algérie. Benbernou S. (2008). Effet inhibiteur des bactéries lactiques isolées de viandes bovines sur des germes indésirables. Mémoire de Magister. école nationale vétérinaire. Alger. Bekki R.M.A. (1999). Recherche de levures laitiére en vue de la valorisation du lactosérum. Mémoire de Magistére. Université Djillali Liabes. Sidi-Bel-Abbèse. Algérie. Boudjelal A. et Nancib N. (2001). Production d’acide lactique par Lactobacillus rhamnosus sur milieu à base de jus de dattes. Rev. Energ. Ren : Production et Valorisation-Biomasse. 41-46. Boudjema Khaled. (2008). Essai d’optimisation de la production d’acide lactique sur lactosérum par Streptococcus thermophilus. Mémoire de magister en biologie. Université M’Hamed Bougara. Boumerdès. Algérie. Bougattoucha A., Balannec B., Nacef S. et Amrane A. (2007). A generalized unstructured model for batch cultures of Lactobacillus helveticus. Enzyme and microbial technology. 41: 377-382. Bounoua M.D. (2005). Isolement et pré identification de bactéries lactiques de différents laits de chamelles du sud Algérien. Mémoire d’Ingénieur d’Etat .Université d’Oran, Algérie. Bourel G., Henini S., Krantar K., Oraby M., Divies C., Garmyn D. (2001). Métabolisme sucre-citrate chez Leuconostoc mesenteroides. Lait. 81: 75–82.

Bouziani I et Sebaha F.Z (2006). Isolement et Identification des bactéries lactiques à partir de Blé Fermenté. Etude Technologique et extraction d’ADN plasmidique. Mémoire d’Ingé-nieur d’Etat. Université de Mostaganem, Algérie. Boyaval P. (1989). Lactic acid bacteria and metal ions. Le lait. 69: 87-113. Callebaut A., Voets A.M., Motte J.C. (1990). Anthocyanin production by plant cell cultures on media based on milk whey . Biotechnol. Lett. 12 :215.
Capiaumant J., Legrand C., Dausset B., Parmentelet I., Liden G., Belleville F., Nabet P. (1994).Whey and 3-lactoglobulin: 2 milk by products which can replace fetal calf serum in mousse hybridoma cell culture. Lait. 74: 127-137. Cayle T., Roland J., Mehnert D., Dinwoodie R ., Larson R ., Mathers J., Raines M., Alm W., Ma’ayeb S. Kiang S., Saunders R.(1989). Production of L-ascorbic acid from whey. Page 57 in Biotechnology in food Processing. Harlander S.K. et Labusa T.P. ed. Noyes Publ . Park Ridge . NJ.
Champagne C.P., Goulet J., Lachance R.A. (1990). Production of backer’s yeast in cheese whey ultrafiltrate. Appl. Environ. Microbiol. 56 : 426.
Champane C.P., Gaudreau H., Conway J., Chartier N et Fonchy E. (1999). Evaluation of yeast extracts as growth media supplements for lactococci and lactobacilli by using automated spectrophotometry. Journal of General and Applied Microbiology. 45: 17-21. Chalampopoulos D., Pandiella S.S., Webb C. (2002). Growth studies of potentially probiotic lactic acid bacteria in cereal based substrates. Journal of Applied Microbiology. 92: 851-859. Chantawannakul P.,Oncharoen A.,Klanbut K.,Chukeatirote E.,Lumyong S. (2002). Characterization of protease of Bacillus subtilis strain 38 isolated from traditionally fermented soybean in northern Thailand. Research article Sci Asia. 28: 241-245. Cheik O.A.M. (2008). Résistance et adaptation au sel chez des lactobacilles isolées de lait de chamelle de Mauritanie. Mémoire d’ingénieur d’état en biotechnologie.Université d’Oran. Algérie. Chervaux C., Ehrlich D., Maguin E. (2000). Physiological study of Lactobacillus delbrueckii subsp. Bulgaricus strains in a novel chemically defined medium. Appl Enveron Microbiol. 66: 5306-5311. Chiarini L., Mara L., Tabacchioni S. (1992). Influence of growth supplements on lactic acid production in whey ultrafiltrate by Lactobacillus helveticus. Applied Microbiology and Biotechnology. 36: 461-464. Christensen J., Steele J. (2003). Impaired growth rates in milk of Lactobacillus helveticus Peptidase mutants can be overcome by use of amino acid supplements. J Bacteriol. 185: 3297-3306.

Cotte Fabrice. (2000). Etude de la valeur alimentaire des pulpes de tomates chez les ruminants. Thèse à l’université Claude-Bernard. Lyon I. France. Courtin P., Nardi M,Wegmann U., Joutsjok V., Ogier J.C., Gripon J.C.,Palva A., Henrich B., Monnet V. (2002). Accelerating cheese proteolysis by encheriching Lactoba-cillus lactis proteolytic system with Lactobacillus peptidases. Int Dairy J .12: 447-454. Cornelius C., Erpicum T.H., Jacques P.H., Thonart P.H. (1996). Comparison of fermentation industrial components such as corn steep and yeast extract for lactic acid bacteria production. Medical Fac Landbouww University Gent. 61: 1461-1463. Crow .V.L.et Pritchard G.G.(1976). Purification and properties of pyruvate Kinase from Streptococcus lactis. Biochimica et Biophysica Acte. 438: 90-101. Crow V.L.et Pritchard G.G.(1977). Fructose 1,6 diphosphate-activated L-lactate dehydroge-nase from Streptococcus lactis ; Kinetic properties and factors affecting activation. Journal of Bacteriology.131(1) :82-91. Crow V.L. (1988). Polysaccharide production by Probionibacteria during lactose fermentation. Appl. Environ. Microbiol. Microbiol. 54 : 1892.
Dairy processing handbook . (1995). Ed. Teknotext AB. Publisher, Tetra Pak Processing Systems AB S-221 86 Lind, Sweden. Chap. 6.4, 15. Dallali A. (2008). Caractéristiques technologiques des lactocoques isolés de « ghars » de la région de Biskra. Mémoire d’ingénieur d’état en biotechnologie. Université d’Oran. Algérie. De Man J.C., Rogosa M., Sharpe M.E. (1960). A medium for the cultivation of lactobacilli. Journal of Applied Bacteriology, 23:130-135. De Roissart, H., Luquet, F.M. (1994). Bactéries lactiques Vol. I et II, Edition Lorica. Deneuville F. (1991) Génie fermentaire (Travaux pratiques). Paris, Doin éditeur. Derouiche K. (2002). Production de l’aspartyl- protéase par Penicillium camemberti cultivée sur milieu à base de lactosérum. Essai de purification et propriétés physico-chimique de l’extrait enzymatique. Mémoire de magister. Université Mentouri. Constantine .algérie. Desmazeaud M.J. (1983).L’état des connaissances en matière de nutrition des bactéries lactiques. Lait, 63:267-316. Desmazeaud M.J. (1992). Les bactéries lactiques. In les groupes microbiens d’intérêt laitier, Eds.J.Hermier, J. Lenoir et F. Weber ; CEPIL, Paris. pp 9-60. Deutcher J et Saier M.H. (1983). ATP-dependent protein Kinase-catalyzed phophorylation of a seryl residue in HPr, a phosphate carrier protein of the phosphotransferase system in Streptococcus pyogenes. Procceding of National Academy of Sciences. 80: 6790-6794.

Deutcher J., Pevec B., Beyreuther K., Kiltz H.H. et Hengstentenberg W. (1986). Streptococcal phosphoénolpyruvate-sugar phosphotransferase system ; amino acid sequence and site of ATP-dependent phosphorylation of HPr. Biochemistry.25 : 6543-6551. Deutscher J., Galinier A. et Martin-Verstraete I. (2002). Carbohydrate Uptake and Metabolism. In Bacillus subtilis and its Closets Relatires : from Genes to Cells. Edited by Sonenshein A.L., Hoch J.A., Losick. R. et Washington D.C. : American Society for Micobiology Press.129-150. Djeghri-Hocine B., Boukhemis M., Zidoune N., Amrane A. (2007a). Growth of lactic acid bacteria on oilseed crop pea and chickpea based media. World Journal of Microbiology and Biotechnology. 23: 765-769. Djeghri-Hocine B., Boukhemis M., Zidoune M.N. et Amrane A. (2007b). Evaluation of de-lipidated egg yolk and yeast autolysate as growth supplements for lactic acid bacteria culture. International Journal of Dairy Technology. 60. No 1. Djidel A. (2007). Production d’acide lactique par Lactobacillus casei subsp. rhamnosus sur jus de datte : cinétique et optimisation en cultures discontinues, semi-continues et continus. Ecole National Supérieur d’Agronomie et des Industries Alimentaires. France. Dossonnet V., Monedero V., Zagorec M., Galinier A., Perez-Martinez G. et Deutscher J. (2000). Phosphorylation of HPr by the bifunctional HPr kinase/P-ser-HPr phosphatase from Lactobacillus casei controls catabolite repression and inducer exclusion but not inducer explusion . Journal of Bacteriology. 182: 2528-2590. Ennis B.M et Maddox I.S. (1985). Use of Clostridium acetobutylicum P262 for production of solvents from whey permeate. Biotechnol. Lett. 7: 601.
Engasser J.M. (1993) Modélisation des procédés de fermentation. In Biotechnologie ,4ème édition, coordonnateur : René Scriban. Edition Technique et Documentation : 281-290. Euzéby J. P. (2000). Dictionnaire de bactériologie vétérinaire. http://www.sv.cict.fr/bacterio/bacdico/garde.html FAO. (1995). Le lait et les produits laitiers dans la nutrition humaine. Alimentation et nutrition n° 28. ISBN 92-5-20534-6. Fira Dj., Kojic M., Banina A., Spasojevic I., Strahinic I., Topisirovic L. (2001). Characterization of cell envelope-associated proteinases of thermophilic lactobacilli. J. Appl. Microbiol. 90 : 123-130. Floetenmeyer M.D., Glatz B.A., Hammond E.G. (1985). Continuous culture fermentation of whey permeate to produce microbial oil. J. Dairy Sci. 68: 633.
Fox P.F. (1989). Developments in dairy industry, New York: Elsevier Applied Sciences.

Fukumoto L.R., Li-Chan E., Kwan L., Nakai S. (1994). Isolation of immunoglobulin from cheese whey using ultrafiltration and immobilized metal affinity chromatography. Food Res. Int. 27: 335–348. Galinier A., Haiech J., Kilhoffer M.C., Jaquinod M., Stulke J., Deutscher J. et Martin-Verstraete I. (1997).The Bacillus subtilis crh gene encodes a HPr-like protein involved in carbon catabolite repression. Proceeding of National Academy of sciences USA. 94: 8439-8444. Galinier A., Kravanja M., Engelmann R., Hengstenberg W., Kilhoffer M.C., Deutscher J. et Heiech J. (1998). Now protein kinase and protein phosphatase families mediate signal transduction in bacterial catabolite repression. Proceeding of National Academy of Sciences USA . 95: 1823-1828. Gaudreau H., Champagne C.P., Jelen P. (2005). The use of crude cellular extracts of Lactobacillus delbrueckii ssp. Bulgaricus 11482 to stimulate growth of a probiotic Lactobacillus rhamnosus culture in milk. Enzyme and Microbial Technology. 36: 83-90. Gardner N., Gardner J., Savard T., Abernier P., Calwell G., Champagne C.P. (2001). Selection and characterization of mixed starter cultures for lactic acid fermentation of carrot, cabbage, beet and onion vegetable mixtures. International Journal of Food Microbiology. 64: 261-275. Gawel J et Kosikowski F.V. (1978). Improving alcohol fermentation in concentrated ultrafiltration permeates of cottage cheese whey. J.Food Sci. 43: 1717.
Gibson T. et Abdel-Malek Y. (1945). The formation of carbon dioxide by lactic acid bacteria and Bacillus licheniformis and a cultural method of detecting the process. Journal of Dairy Research. 14 : 35-44. Ghaly A.E., Tango M.S.A et Dams M.A.A. (2003). Enhanced lactic acid production from cheese whey with nutrient supplement addition. Agriculturel Engineering international. The CIGR journal of scientific research and developpement. Gomez A et Castillo F.J. (1983). Production of biomass and ß-D-galactosidase by Condida pseudotro-picalis in continous culture on whey. Biotechnol. Bioeng. 25: 1341.
Grattepenche F. (2005). Etude d’un systéme de préfermentation en continu du lait par une culture mixte immobilisée fonctionnelle. Thése. Université Laval. Quebec .Canada. Grobben G., Chin-joe I., Kitzen A., Boels C., Boer F., Sikkema J., Smith M., de Bont J. (1998). Enhancement of exopolysaccharide production by Lactobacillus delbrueckii subsp. Bulgaricus NCFB 2772 with a simpliefied defined medium. Appl Environ Microbiol. 64 : 1333- 1337. Hansen R. (1980). Carbery milk products in Ireland produces alcohol from whey. Nordeuropaisk Mejeri Tidsskr. 46: 10-13. Harper W.J. (1992). Lactose and lactose derivatives in : Whey and lactose processing/ed. by Zadow J.G., London : Elsevier Applied Science.317-360.

Hebali M. (2006). Bactéries lactiques: isolement à partir de saumure d’olives fermentées et données de la nouvelle taxonomie des bactéries lactiques. Mémoire d’Ingénieur d’Etat. Université d’Oran, Algérie. Hebert E.M., Raya R.R., Giori G.S. (2000). Nutritional requirements and nitrogen dependent regulation of proteinase activity of Lactobacillus helveticus CRL 1062. Appl Enviro Microbiol. 66 : 5316-5321. Hebert E.M., Raya R., Oliver G. et Savoyde Giori G. (1996). Microbiol Aliments Nutr. 14 : 65-72. Herrero M., Mayo B., Gonzalez B., Suarez J.E. (1996). Evaluation of technologically important traits in lactic acid bacteria isolated from spontaneous fermentation. Journal of Applied Bacteriology. 81: 565-570. Hilpert H., Gerber H., Amster J. J., Pahud A., Ballabriga L., Arcalis F., Farriaux E., DePeyer E., Nussle D. (1977). Bovine immunoglobulins: their possible utilization in industrially prepared infant’s milk formula. Page 182 in Food and Immunology. L. Hambraeus, L. A. Hanson and M. Mcfarlane, eds. Almquist and Wiksel Int., Stockholm, Sweden. HO Thi Nguyet Thu. (2008). Etude de la flore lactique du Nem Chua, produit carné fermenté cru traditionnel du sud Vietnam et maitrise du processus de fermentation par ajout de souches lactiques sélectionnées spécifiques du produit. Thèse de l’université Bordeaux 1. Frances. Horn S.J., Aspmo S.I., Eijsink V.G.H. (2007). Evalution of different cod viscera fractions and their seasonal variation used in a growth medium for lactic acid bacteria. Enzyme and Microbial Technology. 40: 1328-1334. Hoppe G. et Higgins J. (1992). Demineralization. In: Whey and lactose processing (JG Zadow, ed), Elsevier,London. Hutkins R., Horward A., Morris A., Larry L. et McKay. (1985). Galactose transport in Streptococcus thermophilus. Applied and Environment Microbiology. 50: 772-776. Hutkins R.W. et Nannen N.L. (1993). pH homostasis in lactic acid bacteria. Journal of dairy science. 76: 2354-2365. Inagawa J., Kiyosawa I. et Nagasawa T. (1987). Effect of phytic acid on the hydrolysis of lactose wih β-galactosidase. Agric. Biol. Chem. 51(11): 3027-3032. Jacquet Violleau Valérie. (1999). Déminéralisation par électrodialyse en présence d’un complexant application au lactosérum. Thése. Institut National Polytechnique de Toulouse. France.

Janicka I., Maliszewska M., Pedziwilk F. (1976). Utilization of lactose and production of corrinoids in selected strains of propionic acid bacteria in cheese whey and casein media. Acte Microbiol. Polonica. 25: 205.
Jault J.M., Fieulaine S., Nessler S., Gonzalo P., Di Pietro A .,Deutscheret J., Galimier A. (2000). The HPr Kinase from Bacillus subtilis is a homo-oligomeric enzyme which exhibits strong positive cooêrativity for nucleotide and fructose 1,6-bisphosphate binding. Journal of Biological Chemistry. 275: 1773-1780. Johnston M.L., Delwiche E.A. (1965). Distribution and characteristics of the catalase of Lactobacillaceae. Journal of Bacteriology. 50: 347-357.
Kabadjova-Hristova P., Bakalova S., Gocheva B., Moncheva P. (2006). Evidence for proteolytic activity of Lactobacilli isolated from kefir grains. Biotechnol. & Biotechnol. 20:2
Kalbitzer H.R., Hengstenberg W., Rosch P., Muss P., Bernsmann P., Engelmann R., Dorschug M., Deutscher J.(1982). HPr proteins of different microorganisms studied by hydrogen-1 high-resolution nuclear magnetic resonance: similarities of structures and mechanisms. Biochemistry. 21 : 2879-2885. Kandler O., Weiss N., (1986). Regular non-sporing Gram-positive rods. Bergey’s Manual of Systematic Bacteriology. P.H.A. Senath, Mair, N.S. and M.E. sharp Edition Williams and Wilkins. Baltimore, 2:1209-1234. Kaneko T., Susuki H., Tkahashi T. (1987). The effects of metal ions on diacétyle production by Streptococcus lactis ssp. Diacetylactis 3022. Agricultural and Biological Chemistry. 51: 2315-2320. Kanenchi C., Seki M. et Komagata K. (1988). Appl. Env. Microbiol. 54: 3053. Karam, N-E. (1995) Constitution d'un souchier de bactéries lactiques à intérêt biotechno-logique: Etude biochimique et moléculaire. Thèse dEtat, Université d'Oran. Algérie. Karam, N-E. et Karam H. (1994). Alimentation, génétique et santé de l’enfant, Eds J.F. Desjeux et M.Touhami, L’harmattan, Paris.pp 257-264. Kinsella J.E. (1984). Milk proteins: physicochemical and functional properties. CRC Crit Rev Food Sei Nutr.21: 197-262. Klein G., Pack A., Bonaparte C., and Realer G. (1998).Taxonomy and physiology of probiotic lactic acid bacteria. Int.J.Food Microbiol, 41: 103-125. Kojic M., Fira D., Banina A. et Topisirovi C.L. (1991). Characterization of the cell wall-Bound Proteinase of Lactobacillus casei HN14. Applied and Environmental Microbiology. 57: 1753-1757.

Koning W.N et Otto. (1983). Mécanismes de transport des nutriments dans les bactéries lactiques : les bactéries lactiques T1, Aspects fondamentaux et technologiques. Ed Loria, Lavoisier, 604p. Korkeala H., Soback S., Hirin J. (1984). Effet of cadium on the growth of Lactobacillus lactis, Lactobacillus helveticus and Streptococcus thermophilus in milk. Journal of dairy Research. 51: 591-596. Kravanja M ., Engelmann R., Dossonnet V., Bluggel M., Meyer H.E., Frank R., Galinier A., Dentscher J., Schnell N. et Hengstenberg W. (1999). The hpr K gene of Enterococcus faecalis . encodes a novel bifunctional enzyme ; the HPr Kirase /phosphatase. Moleculer Micobiology. 31:59-66. Kulozik N. et Wilde J. (1999). Rapid lactic acid production at high cell concentrations in whey ultrafiltrate by Lactobacillus helveticus. Enzume and Microbiol Technology. 24: 297-302. Kunji E.R., Mireau I., Hagting A., Poolman B., Konings W.N. (1996). The proteolytic system of lactic acid bacteria. Antonie van Leeuwenhoek. 70 : 187-221. Kurbanoglu E.B. (2004). Enhancement of lactic acid production with ram horn peptone by Lactobacillus casei. World Journal of Microbiology and Biotechnology. 20: 37-42. Lauret R., Morel-Deville F, Berthier F., Champomier-Verges M., Postma P., Ehrlich D., Zagorec M. (1996). Carbohydrate uilization in Lactobacillus sake. Appl Environ Microbiol. 62: 1922-1927. Ledezma O.V., De Ruiz Holgado A.P., Oliver G., De Giori G.S., Raibaud P., Galpin J.V. (1977). A synthetic medium for comparative nutritional studies of lactobacilli. J Appl Bacteriol. 42: 123-133. Lenq W., Speckman R.A., Crang R.E., Steinberg M.P. (1989). Enhanced conversion of lactose to glycerol by Kluyveromyces fragilis utilizing whey permeate as a substrate. Appl. Environ. Microbiol. 55: 573.
Leroy F., De Vuyst L. (2004). Lactic acid bacteria as functional starter cultures for the food fermentation industry. Food Science & Technology. 15: 67-78. Leuchtenberger A., Friese E., Ruttloff H. (1989). Synthesis of variable enzyme spectrum by immobilized mycelium of A. niger. Zentralbl. Mikrobiol.144:139.
Liberman E.S et Bleiweis A.S. (1984). Role of the phosphoenolpyruvate dependent glucose phosphotransferase system of Streptococcus mutans GS5 in the regulation of lactose uptake. Infection and Immunity. 43: 536-542. Lindgreen S.E., Axelsson L.T. et McFeeters R.F. (1990). Anaerobic L-lactate degradation by Lactobacillus plantarum. FEMS Microbiol. Letters. 66: 209-214.

Luesink E.J., Kuiper O.P. et De Vos W.M. (1998) . Regulation of the carbohydrate metabo-lism in Lactococcus Lactis and other lactic acid bacteria. Le Lait. 78: 69-76. Lund B., Norddahl B., Ahring B. (1992). Production of lactic acid from whey using hydrolysed whey protein as nitrogen source. Biotechnol. Lett. 14: 851.
Mahaia M.A et Cheryan M. (1986). Lactic acid from acid whey permeate in a membrane recycle bioreactor. Enzyme Microbiol. Technol.8: 289.
Maldonado A. (1992). Effet of medium composition on xanthan gum production from whey permeate using Xanthomonas campestris NRRL .B-1459.M.S.Thesis. The Ohio State Univ. Colombus.
Marjo J., Starrenburg C., Hugenholtz J. (1991). Citrate fermentation by Lactoccoccus and Leuconostc ssp. American Society for Microbiology. 57: 3535-3540. Marwaha S.S., Kennedy J.F., Sethi R.P. (1983). Vitamine B12 production from whey and simulation of optimal cultural conditions. Proc. Biochem. (Dec): 24.
Mawson A.J. (1988). Yeast biomass production from acid whey permeate. Biotechnol. Lett. 10: 503.
Mc Comas K.A.J et Gilliland S.E. (2003). Growth of probiotic and traditional yogurt cultures in milk supplemented with whey protein hydrolysate. Journal of Food Science. 68: 2090-2095. Mijakovic I., Poncet S., Galimier A., Monedero V., Fieulaine S ., Janin J., Nessler S., Marquez J.A, Scheffzek K., Hasenbein S., Hengstenberg W. et Deutscher j. (2002). Pyrophosphate-producing protein dephosphorylation by HPr Kinase/phospharylase : A relic of early life? Proceeding of National Academy of Sciences USA. Miller B. et Puhan Z. (1980). Möglichkeiten zur Verkürzung der Fermentation bei Acidophilus-Milch. Sehweig. Milchw. Forschung. 9: 49-56. Miwa Y., Nakata A., Ogiwara A., Yamamoto M. et Fujita Y. (2000). Evalution and charaterization of catabolite-responsive elements (cre) of Bacillus subtilis. Nucleic Acids Research. 28: 1206-1210. Montel M.C., Labadie J. (1986). Specific nutritional requirements of lactobacilli ssp. From meat. Zentrablatt fur Bakteriologie Mikrobiologie und Higiene. 183: 23-27. Morishita T., Deguchi Y., Yajima M., Sakurai T., Yura T. (1981). Multiple nutritional requirements of lactobacilli: genetic lesions affecting amino acid biosynthesis pathways. J Bacteriol. 148: 64-71. Morishita T., Fukuda T., Shirota M., Yura T. (1974). Genetic basis of nutritional requirements in Lactobacillus casei. J Bacteriol. 120: 1078-1084.

Morr, C. V. (1982). Functional Properties of Milk Proteins and their Use as Food Ingredients. In : Dairy Chemistry-1 , Ed., P. F. Fox, NY, 375-398. Nancid A., Nancid N., Meziane-Cherif D, Boubendir A., Fick M. et Boudrant J. (2005). Join effect of nitrogen sources and B vitamin supplementation of date juice on lactic acid production by Lactobacillus casei subsp. rhamnosus. Bioresource Technology. 96: 63-67. Nancib N., Nancib A., Boudjelal A., Benslimane C., Blanchard F., Boudrant J. (2001). The effect of supplementation by different nitrogen sources on the production of lactic acid from date juice by Lactobacillus casei subsp. rhamnosus. Bioresource Technology. 78: 149-153. Nannen N.L et Hutkins R.W. (1991). Proton-translocating Adenosine Triphosphatase Activity in lactic acid bacteria. Journal of dairy science. 74: 745-751. Neves A.R., Ramos A., Nunes M.C., Kleerebezem M., Hugenholtz J.,. de Vos W.M Almeida j. et Santos H. (1999). In vivo magnetic resonance studies of glycolytic kinetics in Lactococcus lactis. Biotechnology and Bioengineering. 64:200-212. Norton S., Lacroix C. et Vuillemard J.C. (1994). Reduction of yeast extracts supplementa-tion in lactic acid fermentation of whey permeat by immobilized cell technology. Journal of Dairy Science. 77: 2494-2508. Orla –Jensen, S. (1919). The lactic acid bacteria; Host and Son, Copenhagen. Otto R .,Vije T., Ten Brink B., Klont B. et Konings W.N. (1985). Energy metabolism in Streptococcus cremoris during lactose starvation. Archives of Microbiology. 141: 348-352. Pearce R.J. (1983). Thermal separation of /3-lactoglobulin and a-Iactalbumin in bovine cheddar cheese whey. Aust J Dairy Technol. 38 : 144-149. Poolman B., Smid E.J., Velkamp H. et Knings W.N. (1987). Bioenergetic consequences of lactose starvation for continuously cultured Streptococcus cremoris. Journal of Bacteriology. 169: 1460-1468. Ramstrom H., Samglier S., Leize-Wagner E., Philippe C., Van Dorsselaer A. et Haiech J. (2003). Properties and regulation of the bifunctional enzyme HPr.kinese/phosphatase in Bacillus subtilis. Journal of Biological Chemistry. 278 : 1174-1185. Rapin J.D. Marison I.W., Reilly P.J. (1994). Glycerol production by yeast fermentation of whey permeate. Enzyme Microbiol. Technol. 16. 143.
Reinbold R.S. et Takemoto J. (1988). Use of Swiss cheese whey permeate by kluyveromyces fragilis and mixed culture of Rhodopseudomonas sphaeroides and Bacillus megaterium. J. Dairy Sci. 71:1799.
Reizer J., Hoischen C., Titgemeyer F., Rivolta C., Rabus R., Stulke J., Karameta D., Saier M.H. et Hillen W. (1998). A novel protein kinase that controls carbon catabolite repression in bacteria. Molecular Microbiology. 27: 1157-1169.

Reizer J., Navotny M.J., Panos.C. et Saier M.H. (1983). Mechamism of inducer explusion in Streptococcus pyogenes : a two-step process activated by ATP. Journal of Bacteriology. 156: 354-361. Reizer J. et Penos C.(1980). Regulation of beta-galaotoside phosphate accumulation in Streptococcus pyogenes by an expulsion mechanism. Proceeding of National Academy of Sciences USA. 77: 5497-5501. Rerat A., Lacrois M., Simoes-Munes C. Vaugelade P., Vaissade P. (1984). Absorption intestinale comparée d'un mélange d'hydrolysats ménagés de protéines laitières et d'un mélange d'acides aminés libres de même composition chez le porc éveillé. Bull. Acad. Natl. Med. 168 : 385-391. Roger L. (1979). Contributions à la recherche d'une meilleure utilisation en alimentation humaine des composants glucidiques et protéiques du lactosérum grâce à l'emploi des techniques à membranes. Thèse de Doctorat de 3e cycle, Rennes (ENSAR).France. Roufik S. (2001). Les concentrés protéiques du lactosérum: étude comparative de la composition globale, séparation et caractérisation des fractions protéiques. Mémoire présenté à la Faculté des études supérieures de l'Université Laval. Canada. Saier M.H et Crasnier M.(1996). Inducer exlusion and the regulation of sugar transport. Research in Microbiology. 147: 482-489. Salama H.S., Foda M.S., El-Sharaby A., Selim M.H. (1983). A novel approach for whey recycling in production of bacterial insecticides. Entomophaga. 28: 151.
Samelis J., Maurogenakis F., Metaxopoulos J. (1994). Characterization of lactic acid bacteria isolated from naturally fermented creak dry Somali. Int. J. food. Microbiol,23: 179-196. Sandhu D.K. et Waraich M.K. (1983). Conversion of cheese whey to single-cell protein. Biotechnol. Bioeng. 25: 797.
Sang Y.,Kato N.,Lau C.X.,Matsumiya Y.,Kato H.,Watanabo K.(2000). Rapid identification of 11 human intestinal Lactobacillus species by multiplex PCR assays using group and species specific primers derived from 16S-23S rRNA intergenic spacer region and its flanking 23s rRNA. FEMS Microbiol .187:167-173. Schepers A.W., Thibault J. et Lacroix C. (2006). Continuous lactic acid productions in whey permeate/yeast extract medium with immobilized Lactobacillus helveticus in a two stage process: model and experiments. Enzyme Microb. Technol. 38: 324-337. Schwartz R.D. (1987). Biopolymers from whey page 56 in Bull. Int. Dairy Fed. No. 212. Int. Dairy fed. Brussels. Belgium.
Sirisaneeyakul S., Luangpipat T., Vanichsriratana W., Scrinophakun T., Chen H.H. et Chisti Y. (2007). Optimization of lactic acid production by immobilized Lactococcus lactis Io1. Journal of Industry Microbiology and Biotechnology. 34: 381-391.

Sobotkovà L., Sikyta B., Smékal F. (1989). Production of lysine by mutants of Escherichia coli K12 in medium with lactose. Acta. Biorechnol. 9: 173.
Somkuti G.A. et Bencivengo M.M. (1981). Citric acid fermentation in whey permeate. Dev. Ind. Microbiol. 22: 557.
Sooch B. S.et Singh R. S. (2002). Characterization of cheese whey and its suitability for ethanol production by alginate entrapped yeast cells. Journal of food science and technology. 39: 21-25. Sottiez P. (1990). Produits dérivés des fabrications fromagères in : lait et produits laitées ; vache, brebis, chèvre. Edition Lavoisier, Paris.633p. Stiles M.E., Holzapfel W.H. (1997).Lactic acid bacteria of foods and their current taxonomy.Int.J.Food.Microbiol.36: 1-29. Tamura Y., Mizota T., Shimamura et Tomita M. (1993). Lactulose and its application to food and pharmaceutical industries. Page 43 in Bull. Int. Dairy Fed . No. 289. Int. Dairy Fed. Brussels Belgium.
Terzaghi B.E. et Sandine W.E. (1975). Improved medium for lactic streptococci and their bacteriophage. Applied Microbiology. 29: 807-813. Thivierge C. M., Bernier J.F., Dubreuil P., Lapierre H. (2002). The effect of jugular or abomasal infusion of amino acids on milk yield in lactating cows fed a protein deficient diet. Reprod. Nutr. Dev. 42: 1–13. Thomas T.D. (1976). Regulation of lactose metabolism in group N Streptocci. Applied and Environment Microbiology. 32: 474-478. Thompson J.T. et Thomas D. (1977). Phosphoenol pyruvate and 2-phosphoglycerate : endogenous energy source(s) for sugar accumulation by starved cells of Streptococcus lactis. Journal of Bacteriology. 130: 583-595. Thompson J., Turner K.W. et Thomas T.D. (1978). Catabolite inhibition and sequential metabolism of sugars by Streptococcus lactis. Journal of Bacteriology. 133: 1163-1174. Thompson J et Saier M.H. (1981). Régulation of methyl-β-D-thiogalatoside-6-phosphate accumulation in Streptococcus lactis by exclusion and expulsion mechemisms. Journal of Becteriology. 146: 885-894. Thompson J. et Chessy B.M. (1983). Intracellular hexose-6-phosphohydrolase from Streptococcus lactis : purification, properties, and fonction. Journal of Bacteriology. 156: 70-80.

Thompson J. et Torchia D.A. (1984). Use of 31 P nuclear magnetic resonance spectroscopy and 14 C fluorography in studies of glycolysis and regulation of pyruvate kinase in Streptococcus lactis. Journal of Bacteriology. 158: 791-800. Thompson J. et Gentry-Weeks C.R (1994). Métabolisme des sucres par les bactéries lactiques. Bactéries lactiques, Ed. De Roisserd H et al.lolica-Uriage.1 : 229-239. Tin C.S.F. et Mawson A.J. (1993). Ethanol production from whey in a membrane recycle bioreactor. Proc. Biochem. 28. 217.
Vahvaselkä M.I. et Linko P. (1987). Lactic acid fermentation in milk ultrafiltrate by Lactobacillus helveticus. In: proceedings of the 4th European Congress on Biotechnology, Vol.3.pp.317-320. Neijssel O.M., van der Meer R.R. and LUYBEN K Ch A.M, eds. Amsterdam: Elsevier. Van Den Berg, D. J. C., Smits, A., Pot, B., Ledeboer, A. M., Kersters, K., Verbakel, J. M. A., et Verrips, C. T. (1993). Isolement, screening and identification of lactic acid bacteria from traditional food fermentation process and culture collections. Food biotechnology. 7: 189-205. Vojnovic V., Ritz M., Vahcic N. (1993). Sensory evolution of whey-based fruit beverages. Nahrung. 37: 285-304. Watanabe M., Kaburagi T. Seto Y., Morita F., Suzuki Y. Nakajima H. (2004). Whey protein hydrolyzate for growth promotion of lactic acid bacteria. Japan Kokai Tokkyo Koho. pp 11. Coden: JKXXAF JP 2004057047. Wee.Y.J., Kim J.N. et Ryu H.W. (2006). Biotechnological production of lactic acid and its Recent applications. Food Technology and Biotechnology. 442: 163-172. Weigel N., Powers D.A. et Roseman S. (1982). Sugar transport by the becterial phospho-transferese system. Primary structure and active site of a general phosphocarrier plotein (HPr) from Salmonella typhimurium . Journal of biological Chemistry. 257: 14499-14509. White G.E., Cooney C.L., Sinskey A.J et Miller S.A. (1976). Continuous culture studies on the growth and physiology of Streptococcus mutans. J. Dent. Res. 55 (2): 239-243. Woo A. (2002). La grande diversité du lactosérum. Agriculture et agroalimentaire. Canada. p 3-13. Yang S.T., Zhu H., Silva E.M. (1993). Production of value-added products from agricultural and food rocessing by products. Page 47 in biotechnol. Ogy in the 21st Century. C. Ayyanna, ed tate McGraw-Hill Publ. Co. Lid., New Delhi. India.
Yang S.T. et Silva E.M. (1995). Novel Products and New Technologies for use of a familiar carbohydrate, milk lactose. Journal of Dairy Science. 78: 2541-2562.

Ye J.J., Reizer J., Cui X. et Saier M .H.(1994). Inhibition of the phosphoenolpyruvate : lactose phosphtransfease system and activation of cytoplasmic sugar-phophate phosphatase in Lactococcus lactis by ATP-dependent metabolite-activated phosphorylation of serine 46 in the phosphocarrier protein HPr. Journal of biological chemistry. 269: 11837-11844. Zhang Z.Y., Jin B. et Kelly J.M. (2007). Production of lactic acid from renewable materials by Rhizopus fungi. Biochemical engineering Journal. 35: 251-263.

ANNEXE

1 -Milieu MRS (De Man, Rogosa et Sharpe, 1960) : utilisé pour la culture des souches
Peptone 10g Extrait de viande 10g Extrait de levure 5g Glucose 20g Acétate de sodium 5g Citrate d’ammonium 1g Sulfate de magnésium 0.1g Sulfate de manganèse 0,05g Phosphate dipotassique 2g Tween 80 1ml Eau distillée qsp 1000ml Agar 15g pH= 6,5 Stérilisation à 120°C pendant 20 minutes.
2 - Lait écrémé : utilisé pour la conservation des souches et l’étude de l’activité acidifiante des bactéries
Lait écrémé 10g Extrait de levure 0,5g Eau distillée 100ml Stérilisation à 120°c pendant 10 minutes.
3 - Tampon phosphate sodium:
KH2PO4 1M 1.36% (m/v) Na2HPO4 1M 3.58% (m/v)
4 - Réactifs utilisés pour la méthode de Bertrand :
• Liqueur Tartrique : Tartrate double Na,K 200 g Soude 150g Eau distillé qsp 1000ml
• Liqueur cuivrique : Sulfate de cuivre 40g Eau distillé qsp 1000ml
• Liqueur ferrique : Sulfate ferrique 50g Acide sulfurique 200g Eau distillé qsp 1000ml
• Ferrocyanure de K4 106 g/l • Acétate de zinc 30%

5 - Dosage des sucres par la technique de G. Bertrand :
Table de conversion
KMnO4 N /10 en ml
GLUCOSE en mg
SUCRE INTERVERTI en mg
7,0 22,2 22,0 ,1 22,5 22,4 ,2 23,0 22,7 ,3 23,2 23,0 ,4 23,5 23,4 ,5 24,0 23,7 ,6 24,2 24,1 ,7 24,5 24,4 ,8 25,0 24,7 ,9 25,2 25,1
8,0 25,5 25,5 ,1 26,0 25,8 ,2 26,2 26,1 ,3 26,6 26,5 ,4 27,0 26,8 ,5 27,2 27,1 ,6 27,6 27,5 ,7 28,0 27,8 ,8 28,2 28,1 ,9 28,6 28,5
9,0 29,0 28,8 ,1 29,2 29,1 ,2 29,6 29,5 ,3 30,0 29,8 ,4 30,2 30,1 ,5 30,6 30,5 ,6 31,0 30,8 ,7 31,3 31,1 ,8 31,6 31,5 ,9 32,0 31,8
10,0 32,3 32,2 ,1 32,7 32,5 ,2 33,0 32,9 ,3 33,4 33,3 ,4 33,7 33,6 ,5 34,0 33,9 ,6 34,4 34,3 ,7 34,7 34,6 ,8 35,1 35,0 ,9 35,5 35,3
11,0 35,8 35,6 ,1 36,1 36,0 ,2 36,5 36,4 ,3 36,8 36,7 ,4 37,1 37,0 ,5 37,5 37,4 ,6 37,8 37,7 ,7 38,2 38,1 ,8 38,5 38,4 ,9 38,8 38,7
12,0 39,2 39,1 ,1 39,6 39,4 ,2 39,9 39,7 ,3 40,3 40,2 ,4 40,6 40,5 ,5 41,0 40,8 ,6 41,3 41,2
,7 41,7 41,5
KMnO4 N /10 en ml
GLUCOSE en mg
SUCRE INTERVERTI en mg
12,8 42,0 42,0 ,9 42,4 42,3
13,0 42,7 42,6 ,1 43,1 43,0 ,2 43,4 43,3 ,3 43,8 43,7 ,4 44,1 44,1 ,5 44,5 44,4 ,6 45,0 44,7 ,7 45,2 45,2 ,8 45,6 45,5 ,9 46,0 45,9
14,0 46,3 46,3 ,1 46,7 46,6 ,2 47,0 47,0 ,3 47,4 47,3 ,4 47,7 47,6 ,5 48,1 48,0 ,6 48,5 48,4 ,7 48,8 48,8 ,8 49,1 49,1 ,9 49,5 49,5
15,0 49,8 49,8 ,1 50,2 50,2 ,2 50,5 50,5 ,3 51,0 51,0 ,4 51,3 51,3 ,5 51,6 51,6 ,6 52,0 52,1 ,7 52,4 52,4 ,8 52,7 52,7 ,9 53,1 53,1
16,0 53,5 53,5 ,1 54,0 53,9 ,2 54,2 54,2 ,3 54,6 54,6 ,4 55,0 55,0 ,5 55,3 55,3 ,6 55,7 55,7 ,7 56,1 56,1 ,8 56,4 56,4 ,9 56,8 56,9
17,0 57,1 57,2 ,1 57,5 57,5 ,2 57,8 57,9 ,3 58,2 58,3 ,4 58,6 58,7 ,5 59,0 59,0 ,6 59,3 59,4 ,7 59,7 59,8 ,8 60,0 60,1 ,9 60,4 60,4
18,0 60,8 61,0 ,1 61,2 61,3 ,2 61,6 61,6 ,3 62,0 62,1 ,4 62,3 62,4 ,5 62,8 62,9

KMnO4 N /10 en ml
GLUCOSE en mg
SUCRE INTERVERTI en mg
18,6 63,1 63,2 ,7 63,4 63,5 ,8 63,8 64,0 ,9 64,2 64,4
19,0 64,5 64,8 ,1 65,0 65,1 ,2 65,2 65,4 ,3 65,7 65,9 ,4 66,0 66,2 ,5 66,4 66,5 ,6 66,8 67,1 ,7 67,1 67,4 ,8 67,5 67,8 ,9 68,0 68,2
20,0 68,3 68,7 ,1 68,7 69,0 ,2 69,0 69,3 ,3 69,4 69,7 ,4 69,8 70,1 ,5 70,2 70,5 ,6 70,5 71,0 ,7 71,0 71,3 ,8 71,3 71,6 ,9 71,7 72,1
21,0 72,1 72,4 ,1 72,5 72,9 ,2 72,8 73,2 ,3 73,3 73,6 ,4 73,6 74,1 ,5 74,0 74,4 ,6 74,5 74,9 ,7 74,8 75,2 ,8 75,3 75,6 ,9 75,6 76,0
22,0 76,0 76,4 ,1 76,4 76,8 ,2 76,8 77,2 ,3 77,2 77,6 ,4 77,6 78,0 ,5 78,0 78,3 ,6 78,4 78,8 ,7 78,7 79,2 ,8 79,1 79,5 ,9 79,5 80,0
23,0 80,0 80,3 ,1 80,3 80,8 ,2 80,6 81,1 ,3 81,1 81,5 ,4 81,5 82,0 ,5 81,8 82,3 ,6 82,3 82,8 ,7 82,6 83,2 ,8 83,0 83,5 ,9 83,5 84,0
24,0 84,0 84,4 ,1 84,3 84,8 ,2 84,7 85,2 ,3 85,1 85,6 ,4 85,5 86,0 ,6 90,3 90,9 ,7 90,6 91,2 ,8 91,0 91,6
,9 91,5 92,1 26,0 92,0 92,5
,1 92,3 93,0 ,2 92,7 93,3 ,3 93,1 93,8 ,4 93,5 94,1 ,5 94,0 94,5 ,6 94,3 95,0 ,7 94,7 95,3 ,8 95,1 95,8 ,9 95,5 96,2
27,0 96,0 96,6 ,1 96,3 97,0 ,2 96,6 97,3 ,3 97,1 97,9 ,4 97,5 98,2 ,5 98,0 98,6 ,6 98,3 99,1 ,7 98,7 99,4 ,8 99,1 99,9 ,9 99,5 100,0
28,0 100,0

Le taux du sucre interverti est calculé (en %) selon la formule : [%]= Masse du sucre interverti (Tableau) . volume de la prise d’essai . 100 Volume de échantillon à dosé Masse de la prise d’essai 1000 6 - Fermenteur utilisé lors de la conduite des cinétiques
7 - Courbe étalon pour le dosage de la fraction enzymatique dans les surnageants de cultures (courbe établie à l’aide de trypsine).

Résumé Nous avons cherché à utiliser le lactosérum doux pour élaborer un milieu de culture pour les lactobacilles. Les bactéries ont été cultivées sur lactosérum déprotéiné seul ou supplémenté en différentes quantités d’extrait de levure et/ou de jus de tomate fraîche. Les résultats obtenus montrent que la production de biomasse produite sur lactosérum additionné d’extrait de levure est supérieure à celle observée en présence de jus de tomate. Dans les deux cas, la biomasse reste inférieure à celle obtenue en milieu témoin MRS. Par contre, sur le milieu dénommé milieu BMK (lactosérum additionné d’un mélange de jus de tomate et d’extrait de levure), la biomasse obtenue est comparable à celle observée en milieu MRS. Nous avons ensuite examiné quelques caractéristiques de ces bactéries lactiques (croissance, consommation du substrat, acidification, production de diacétyle, activité protéolytique, …) sur ce milieu BMK, toujours en comparaison avec le milieu témoin. Les résultats obtenus avec le milieu BMK sont tout à fait comparables à ceux obtenus avec le milieu témoin MRS. Mots-clés : Lactobacillus, lactosérum doux, extrait de levure, jus de tomate fraîche, milieu BMK, croissance bactérienne, acidification, activité protéolytique.
Summary We sought to use the sweet whey to develop an culture medium for lactobacilli. The bacteria were grown on deproteinized whey alone or supplemented with different amounts of yeast extract and / or juice fresh tomato. The results show that the production of biomass production on whey supplemented with yeast extract is higher than that observed in the presence of tomato juice. In both cases, biomass is still less than that obtained in control medium MRS. But, on a medium we set up and called medium BMK (whey supplemented with a mixture of tomato juice and yeast extract), the obtained biomass is comparable to that observed in MRS medium. We examined some characteristics of these lactic acid bacteria (growth, substrate consumption, acidification, diacetyl production, proteolytic activity ...) on the medium BMK, in comparison with the control medium. The results with the medium BMK are quite comparable to those obtained with the control medium MRS. Keywords: Lactobacillus, sweet whey, yeast extract, fresh tomato juice, BMK medium, bacterial growth, acidification, proteolytic activity.
ملخص.lactobacilles سعینا الستخدام مصل اللبن الحلو لتطویر وسط غدائي لبكتریا حمض اللبن
قمنا بزرع مجموعة من البكتیریا في وسط بھ فقط مصل اللبن منزوع البروتینات أو مستكمل مع كمیات مختلفة من خالصة الخمیرة وأیضا مع عصیر الطماطم الطازجة. أظھرت النتائج أن الكتلة الحیویة المنتجة في مصل اللبن مع إضافة خالصة الخمیرة ھي أعلى من
تلك التي لوحظت في وجود عصیر الطماطم. في كلتا الحالتین، الكتلة الحیویة ال تزال اقل من تلك التي تم الحصول علیھا في الوسط الشاھد MRS. على العكس، في الوسط المسمى BMK (مصل اللبن زائد مزیج من عصیر الطماطم و خالصة الخمیرة)، فإن إنتاج
.MRS الكتلة الحیویة ھو مماثل للوسط بعد ذلك قمنا بدراسة بعض الخصائص لبكتیریا حمض اللبن (النمو، استھالك مصدر الكربون، الحموضة، إنتاج diacetyle و النشاط
البروتیني...)على الوسط BMK دائما بالمقارنة مع الوسط الشاھد. النتائج المتحصل علیھا في الوسط BMK ھي مماثلة لتلك التي تم الحصول علیھا
.MRS في الوسط الشاھد
كلمات مفتاحیھ: Lactobacillus، مصل اللبن الحلو، خالصة الخمیرة، عصیر الطماطم الطازجة، وسط BMK، النمو البكتیري، الحموضة، النشاط البروتینى.




















