
1981DavisonPhD.pdf - Enlighten: Theses
492
https://theses.gla.ac.uk/ Theses Digitisation: https://www.gla.ac.uk/myglasgow/research/enlighten/theses/digitisation/ This is a digitised version of the original print thesis. Copyright and moral rights for this work are retained by the author A copy can be downloaded for personal non-commercial research or study, without prior permission or charge This work cannot be reproduced or quoted extensively from without first obtaining permission in writing from the author The content must not be changed in any way or sold commercially in any format or medium without the formal permission of the author When referring to this work, full bibliographic details including the author, title, awarding institution and date of the thesis must be given Enlighten: Theses https://theses.gla.ac.uk/ [email protected]
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of 1981DavisonPhD.pdf - Enlighten: Theses

https://theses.gla.ac.uk/
Theses Digitisation:
https://www.gla.ac.uk/myglasgow/research/enlighten/theses/digitisation/
This is a digitised version of the original print thesis.
Copyright and moral rights for this work are retained by the author
A copy can be downloaded for personal non-commercial research or study,
without prior permission or charge
This work cannot be reproduced or quoted extensively from without first
obtaining permission in writing from the author
The content must not be changed in any way or sold commercially in any
format or medium without the formal permission of the author
When referring to this work, full bibliographic details including the author,
title, awarding institution and date of the thesis must be given
Enlighten: Theses
https://theses.gla.ac.uk/

DETAILED STRUCTURAL ASPECTS OP THE
HERPES SIMPLEX VIRUS GENOME
Andrew John D a v iso n
A T h e s is p r e s e n te d f o r th e D egree o f
D o cto r o f P h ilo s o p h y
IN
The P a c u lty o f S c ie n c e
a t th e U n iv e r s i t y o f G lasgow
I n s t i t u t e o f V ir o lo g y S ep tem b er, 1^81Church S t r e e t G lasgow G il 5JH S c o t la n d

ProQuest Number: 10778000
All rights reserved
INFORMATION TO ALL USERS The quality of this reproduction is dependent upon the quality of the copy submitted.
In the unlikely event that the author did not send a com p le te manuscript and there are missing pages, these will be noted. Also, if material had to be removed,
a note will indicate the deletion.
uestProQuest 10778000
Published by ProQuest LLC(2018). Copyright of the Dissertation is held by the Author.
All rights reserved.This work is protected against unauthorized copying under Title 17, United States C ode
Microform Edition © ProQuest LLC.
ProQuest LLC.789 East Eisenhower Parkway
P.O. Box 1346 Ann Arbor, Ml 48106- 1346

'f ixers
% 1

Rage
TABLE OP CONTENTS
AC KNOWLEDGEMENTS
SUMMARY
ABBREVIATIONS
PART A: INTRODUCTION 1 -3 7
THE HERPESVIRUSES 1
C l a s s i f i c a t i o n and p a t h o g e n ic i t y 1
S tr u c tu r e o f th e v ir io n 4
S tr u c tu r e o f th e HSY genome 6
S tr u c tu r e s o f o th e r h e r p e s v ir u s genom es 10
THE LYTIC CYCLE OP HERPES SIMPLEX VIRUS 14
I n t e r a c t io n o f v ir u s w ith th e c e l l 14
DNA s y n t h e s is 19
RNA s y n t h e s is 24
P r o te in s y n t h e s is and m o d if ic a t io n 29
Enzyme a c t i v i t i e s in d u ced by HSV 34
PART B; MATERIALS AND METHODS 3 8 -7 0
MATERIALS 38
V ir u s e s 38
C e l l s 38
B a c te r ia 38
T is s u e c u ltu r e m edia and s o lu t i o n s 38
R a d io ch em ica ls 39
C h em ica ls 40
DNA 41
Enzymes 41
O ther m a te r ia ls 42
Commonly used s o lu t io n s and b u f fe r s 42

METHODS 4 3
Growth o f c e l l s 43
P r o d u c t io n o f v ir u s s to c it s 43
V ir u s t i t r a t i o n 43
P la q u e p u r i f i c a t i o n o f HSV 44
G en era tio n o f iiS V -l/ilS V -2 i n t e r t y p i c 44recom b in an t v i r u s e s , .
P r e p a r a tio n o f ilSV DI\1 A 45
P r e p a r a t io n o f PRV DNA 47■>
P r e p a r a t io n o f b a c te r io p h a g e lam bda DNA 48
E s t im a t io n o f DNA c o n c e n tr a t io n s 49
P u r i f i c a t io n o f r e s t r i c t i o n e n d o n u c le a se s 49
R e s t r ic t io n e n d o n u c lea se d ig e s t io n 51
A garose g e l e l e c t r o p h o r e s i s 51
P o ly a c r y la m id e g e l e l e c t r o p h o r e s i s 52
G en era l l a b e l l i n g o f DNA w ith *^P 52
I s o l a t i o n o f r e s t r i c t i o n fr a g m e n ts 54
T r a n s fe r o f DNA fra g m en ts to n i t r o c e l l u l o s e 55
B lo t h y b r id is a t io n 55
A u torad iograp hy 57
R e s t r ic t io n fragm en t n o m en cla tu re 57
C a lc u la t io n o f r e l a t i v e m o la r i t i e s 58o f r e s t r i c t i o n fra g m en ts
R a t io n a le f o r mapping r e s t r i c t i o n s i t e s 58
C on tam in ation o f r e s t r i c t i o n fra g m en ts 59i s o l a t e d from g e l s
G en era tio n o f b a c t e r ia l c lo n e s b e a r in g 60recom b in an t p lasim lds w ith riSV DNA i n s e r t s
32L a b e l l in g o f DNA te r m in i w ith P 64
D e t e r m i n a t i o n o f t h e n u c l e o t i d e b7s e q u e n c e o f DuA

PART C: RESULTS AND DISCUSSION
SECTION 1 : MAPPING OP RESTRICTION SITES ON IISV-1 ANN HSV-2 DNA
R e s u lt s
I n tr o d u e t io n
M o lecu la r w e ig h t c a l i b r a t i o n
R e la t iv e m o la r i t i e s o f r e s t r i c t i o n fra g m en ts
IISV-1 Kpnl p h y s ic a l map: r e c le a v a g e d a ta
HSV-1 Kpnl p h y s ic a l map: d o u b le d i g e s t d a ta
HSV-1 Kpnl p h y s ic a l map: h y b r id is a t io n d a ta
IISV-1 Kpnl p h y s ic a l map: r e l a t i v e o r d e r o f fra g m en ts n and p
HSV-l Kpnl p h y s ic a l map
P h y s ic a l maps f o r o th e r r e s t r i c t i o n e n d o n u c le a se s
D is c u s s io n
SECTION 2 : HOMOLOGY BETWEEN HERPESVIRUS GENOMES
R e s u lt s
I n tr o d u c t io n
D is t r ib u t io n o f hom ologous se q u e n c es betw een HSV-1 and HSV—2 DNA
C o l in e a r i t y o f th e genomes o f HSV-1 and HSV-2
N o n -c o lin e a r hom ology betw een th e genom es o f IISV-1 and IISV-2
Homology betw een th e HSV genome and the genom es o f BHV-1 and PRV
Homology betw een th e HSV and CMV genom es
S tu d ie s o f re c o m b in a tio n betw een th e HSV-1 and ilSV-2 genom es

P is c u s s io n
f e a t u r e s o f n u c le ic a c id h y b r id is a t io n
C o l in e a r i t y o f th e genom es o f HSV-1 and HSV-2
C onserved r e g io n s in th e genom es o f HSV-1, HSV-2, EHV-1 and PRV
Homology betw een th e genom es o f HSV and HCMV
D is t r ib u t io n o f r e g io n s hom ologous to ]JSV DNA in th e genom es o f EHV- 1 and PRV
N o n -c o l in e a r hom ology
G e n e tic r e la t e d n e s s o f h e r p e s v ir u s e s
Homology and r e co m b in a tio n i n HSV
D oes o n ly one HSV genome arrangem ent p a r t i c ip a t e in . reco m b in a tio n ?
SECTION 3 : NUCLEOTIDE SEQUENCES OP THE JOINT BETWEEN THE- L AND S SEGMENTS OP THE GENOMES OP. HSV-1 AJp HSV-2
R e s u lt s
I n tr o d u c t io n
S s t I r e s t r i c t i o n maps o f th e HSV-1 and HSV-2 j o i n t r e g io n s
O ther r e s t r i c t i o n maps o f th e HSV-1 and IISV-2 j o i n t r e g io n s
S iz e v a r i a b i l i t y i n th e j o i n t r e g io n
Homology betw een th e j o in t r e g io n s o f o f HSV-1 and HSV-2 DNA
1P in e r e s t r i c t i o n maps o f th e j o in t r e g io n s o f HSV-1 and HSV-2
L o c a t io n o f th e v a r ia b le r e g io n w ith in th e HSV-1 a seq u en ce
D e f i n i t i o n o f th e genom ic te r m in i and l o c a t i o n o f th e a seq u en ce
Tandem r e i t e r a t i o n s i n th e j o i n t r e g io n
C om parison o f n u c le o t id e seq u en c es
E x p r e ss io n o f th e IISV-1 j o in t r e g io n
gage
92
92
94
95
98
99
100
101 103
103
107
107
107
107
109
112114
115
115
116
118
120
121

Page
D is c u s s io n 124
F e a tu r e s o f r e i t e r a t i o n s 124
C oding p o t e n t i a l a t th e 3 ' end o f 125Vmw IE 175 mRNA
R o le o f th e a seq u en ce in c o d in g f o r 126p o ly p e p t id e
Homology betw een th e j o in t r e g io n s o f 127th e ilSV-1 and HSV-2 genom es
R o le o f th e a seq u en ce i n m a tu r a tio n o f 127genome le n g t h DNA
R ole o f th e a seq u en ce i n segm ent in v e r s io n 128
SECTION 4 : INVERSION OP THE TWO SEGMENTS 130OP-THE HSV GENOME . . .
R e s u l t s 130
I n tr o d u c t io n 130
S tr u c tu r e s o f th e j o i n t s and te r m in i 131o f B x l(2 8 -1 )
G ross s t r u c t u r e s o f E x l ( 2 8 - l ) s u b c lo n e s 133
S t r u c tu r e s o f th e j o i n t s and te r m in i o f 134B x l( 2 8 -1 ) su b c lo n e s
S t r u c tu r e s o f th e j o i n t s and te r m in i 138p f o t h e r reco m b in a n ts
E x p r e s s io n o f IE 1 1 0 /1 1 8 by B x l(2 8 -1 ) 140s u b c lo n e s
S t r u c tu r e s o f s u b c lo n e s o f B x l(2 8 -1 ) 142s u b c lo n e s
S tr u c tu r e o f th e genome o f RE4 142
D is c u s s io n 147
P ix e d and f r e e reco m b in a n ts 147
M odels f o r segm ent in v e r s io n 149
M echanism s f o r genome c i r c u l a r i s a t i o n 151and m a tu ra tio n
E x p r e s s io n o f B x l(2 8 -1 ) s u b c lo n e s 154d e le t e d in TR^/IR^
Genome s tr u c tu r e o f RE4 155
REFERENCES

AC KN Q \ i LEDGem eN TS
I sh o u ld l iK e to thank P ro f e s s o r John H. Subak-Sharpe
f o r £ )r o v is io n o f l a c i l i t i e s in th e I n s t i t u t e o f V ir o lo g y and
f o r h i s o v e r a l l s u p e r v is io n o f and en co u ra g in g i n t e r e s t in
t h i s work. I am g r e a t ly in d e b te d to Dr N e i l M. W ilk ie f o r
h i s g en ero u s a d v ic e and g u id a n ce th ro u g h o u t t h i s work and
f o r v a lu a b le c r i t i c i s m o f th e m a n u sc r ip t . I sh o u ld a l s o
l i k e to th an k Dr H .3 . lviarsden and Dr F .J . R ixon f o r t h e ir
c o l la b o r a t io n , Dr J.M . W halley f o r m aking a v a i la b le EHV-1
r e s t r i c t i o n en d o n u c lea se maps p r io r to p u b l i c a t io n , Mrs
M. Murphy f o r i s o l a t i n g s u b c lo n e s o f B x l ( 2 8 - 1 ) , and th e
many members o f th e I n s t i t u t e who p ro v id ed much a d v ic e and
s t im u la t in g d i s c u s s io n . The s e r v i c e s p ro v id e d by th e
Y/ashroom, S to r e , and M edia and C y to lo g y D epartm ents were
much a p p r e c ia te d .
D u rin g t h i s work th e a u th o r was em ployed by th e
M ed ica l R esea rch C o u n c il and, e x c e p t where o th e r w is e
s p e c i f i e d , a l l th e r e s u l t s r e p o r te d were o b ta in e d by th e
a u th o r ’ s own e f f o r t s .

JU. M.ull'
Xuj. o LiiCo-i-o i c o 'C i i ue 3 Lne a o r i v a t i o n o i r o 3 i r i c Lion
e n d o n u c l e a s e maps l o r n3V-1 and nSV—2 HnA; a s t u d y o l t h e
h o m o lo g o u s r e g i o n s oe tw een t h e genomes o f IiSV-1, HSV-2, EHV-1
and PRV, and Hie e f f e c t o f ho o o l o g y upon r e c o m b i n a t i o n
b e tw e e n HSV-1 ana . .3V-2; n u c l e o t i d e s e q u e n c e s 01 t n e L-S
j o i n t r e g i o n s i n HSvr- l and 113V-2 Hi.A; and an a n a l y s i s o f two
HSV—l/H SV -2 i n t e r t.ypic r e c o m b i n a n t s w h ich f a i l t o i n v e r t
n o r m a l l y i n one o r oo th s e g m e n ts o f t h e genome.
R e s t r i c t i o n e n d o n u c l e a s e maps were a l r e a d y a v a i l a b l e
o f HSV-1 HNA f o r A bal , i l i n d l l l , He oh I , Bgl 11 and H p a l , a n a o f
HSV-2 HHA f o r t n c s e enzymes and Kpnl ( W i l k i e , 1 9 76 ; S h a re
and Summers, 197'/; hors .e e t a l . , 1977 ; C o r t i n i and W i l k i e ,
1 9 7 8 ) . wap3 o f HSV-1 HHA f o r Kpnl , BamHI, n h o l and P v u I I ,
an a o f HSV-2 HHA f o r BamHI, were d e t e r m i n e d u s i n g t h rt
t e c h n i q u e s o f s i m u l t a n e o u s d i g e s t i o n w i t h two e n d o n u c l e a s e s ,
r e c l e a v a g e o f i s o l a t e d r e s t r i c t i o n f r a g m e n t s , and b l o t
h y b r i d i s a t i o n . P r e v i o u s l y r e p o r t e d s i z e h e t e r o g e n e i t y a t
t h e L t e r m i n u s and L-S j o i n t o f t h e two genomes was c o n f i r m e d .
The r e s u l t s s u g g e s t e d t h a t nSV-2 HHA i s 1 -2 x 1 0 ° l a r g e r i n
m o l e c u l a r w e i g h t i n t h e S’ seg m e n t t h a n HSV-1 HHA.
HSV-1 an a nSV-2 hha s n a r e a p p r o x i m a t e l y 50^ hom ology ,
ana PhV HHA p o s s e s s e s n o t g r e a t e r t h a n 8f* homology w i t h4
iiSV-1 HHA ( K i e f f e t a l . , 1972 ; B ronson e t a l . , 1972 ; Rand32ana B e n - P o r a t , I 9 6 0 ) , u y o r i d i s a t i o n o f P - l a b e l l e d
r e c o m b i n a n t p l a s m i d s c o n t a i n i n g HSV-1 o r HSV-2 DNA i n s e r t s t o
b l o t s t r i p s o f r e s t r i c t i o n e n d o n u c l e a s e d i g e s t s o f HSV-1 o r
iiSV-2 Hi.A snowed tuali t n e two genomes a r e c o l i n e a r , w i t h i n
ti ie r e s o l u t i o n a t t a i n e d i n t h e s e e x p e r i m e n t s . H y b r i d i s a t i o n
o f ^‘‘P - l a b e l i ou !3V-1 o r nSV-2 HHA wo s i m i l a r b l o t s t r i p s
a l i o we u s e v e n r e ;io*is 01 t h e genome to be i d e n t i f i e d w h ich

a r c more hom ologous t n a n n e i , . n b o u r in g r e g i o n s . HHV—1 and
PuV jJLA a l s o showea j r j - j o r i iomoiogj bo t h e s e r e g i o n s o f
t h e nSV genome t n a n to o 'boo rs , b u t no s i g n i f i c a n t h o r o l o g y
b e tw e e n * S V i>iJA an a riOnV ±fuA was u w b e c t e u . Homologous r e g i o n s
p r o b a b l y r e f l e c t p.*eater c o n s e r v a t i o n o f t h e s t r u c t u r e s o f
p o l y p e p t i d e s encoaod by tnem . f i v e good c a n d i d a t e s f o r
c o n s e r v e d uSV-1 p o l y p e p t i a e s a r e t h e m a j o r HHA-binning; p r o t e i n ,
t h e m a j o r c a p s i u p r o t e i n , t h e DUA p o l y m e r a s e , and two
i m m e d i a t e - e a r l y p o l y p e p t i d e s , Vmw i h 175 and V|i;v/ I n 1 3 6 ' ( 1 4 3 ) .
f i y b r i d i s a t i o n o f c l o n e d nSV DU A f r a g m e n t s to b l o t s t r i p s o f
nnV-1 o r PhV r e s t r i c t i o n e n d o n u c l e a s e d i g e s t s shovvea t h a t
hom ologous r e g i o n s i n t h e L segm ent o f t h e HHV-1 genome a r e
o o l i n e a r w i t n t h e n s eg m en t o f t n e HSV-1 genome i n biie
a r r a n g e m e n t . Homologous r e g i o n s be tw een HSV an a PuV jjuA
were shown n o t to oe a r r a n g e d i n a s i m p l e c o l i n e a r f a s h i o n .
The ^cnome s t r u c t u r e s were a n a l y s e d o f more ' than a
h u n d r e d L S V -l /aS V -2 i n t e r t y p i c r e c o m b i n a n t s p ro d u c e d by
m a r k e r r e s c u e o f HSV-1 r a h w i t h HSV-2 r e s t r i c t i o n f r a g m e n t s
s p a n n i n g t h e L-o j o i n t . The r e c o m b i n a n t s p o s s e s s e u c r o s s o v e r s
p r e f e r e n t i a l l y i n Homologous regio**s. At l e a s t two o f t h e
genome a r r a n g e m e n t s (P and I g ) r e c o m b i n e d , a r e s u l t w h ich
d i s p r o v e s t h e e a r l i e r p r o p o s i t i o n t h a t o n l y one o f t h e s e
two a r r a n g e m e n t s i s a b l e to t a k e p a r t i n t h e g e n e r a t i o n o f
v i a b l e r e c o m b i n a n t s •
HSV hi.A i s b e r m i n a i l y r e d u n d a n t , p o s s e s s i n g a d i r e c t l y
r e p e a t e d s e q u e n c e o f 250 -300 b a s e p a i r s ( t h e a s e q u e n c e ) a t
t h e t e r m i n i which i s a l s o p r e s e n o i n i n v e r s e o r i e n t a t i o n a t
t h e L—b j o i n t ( S u e l d r i c k and j e r t h e l o t , 1 9 7 4 ; G ra f s ^ ro m e t a l . ,
1 j 74 a n a 1J7 3; , /aguer and Summers, 1 9 7 b ) . P i n e s t r u c t u r e
r e s t r i c t i o n enaon e l e a s e «ups o f t h e L-S j o i n t r e g i o n o f HSV-1
m u iSV-2 HHA v.er d c t e r m i n u a , an a n u c l e o t i d e s e q u e n c e s were
a o r i v e a u s l u c ivn .sa i i-rn,u >.cn b s • H e l a b i v e bo b n j s e

s e q u e n c e s ' tne mor.ani a r e i n b o t h c a s e s l o c a l e d c l o s e
t o a s n o r t diroc i repeat o. 1 7 -2 1 Ouse j j a . t r s a t tne o -a and
a - c j u n c t i o n s . Tne . .oV'-l n - o j o i n t r e g i o n c o n t a i n . ; irhrce
s e p a r a t e t u n u e dircc t r e i t e r a t i o n s o i s n o r t s e q u e n c e s
( 1 2 , 16 and 17 e a s e p a i r s ) , and t n e s i z e h e t e r o g e n e i t y o i
v i r i o n LLA i n t n e a a . n c s e q u e n c e s i s a l m o s t c e r t a i n l y
due to v a r i a b l e copy n um bers o f t h e s e r e p e a t e d u n i t s .
A s e c o n d t y p e o f s i x e h e t e r o g e n e i t y was f o u n d t o be due to
i n s e r t i o n o f an a d d i t i o n a l a s e q u e n c e i m m e d i a t e l y a d j a c e n t t o
t h e a s e q u e n c e a t t h e L-S j o i n t . The r e g i o n c o d i n g f o r t n e
3> t e r m i n u s o f HSV-1 VuW I d 175 mAhA was i d e n t i f i e d
a p p r o x i m a t e l y SOU b a s e p a i r s f ro m t h e a s e q u e n c e , b u t i t i s
l i h e l y t h a t t n e a s e q u e n c e and i m m e d i a t e l y a d j a c e n t r e g i o n s
do n o t code f o r p o l y p e p t i d e .
The iISV-i /i iSV-2 i n t e r t y p i c r e c o m b i n a n t B x l ( 2 8 - 1 ) p o s s e s s e s
a m a j o r i t y o f v i r i o n i)l\IA m o l e c u l e s w i t h t h e L s eg m en t i n one
o r i e n t a t i o n ( . f ix ed i n h ) , w h e r e a s t n e S segm ent i n v e r t s
n o r m a l l y ( P r e s t o n e t a l . , 1 9 7 8 ) . An a n a l y s i s o f B x l ( 2 8 - 1 )
and tw e n ty s u b c l o n e s t h e r e o f , some o f w h ic h a r e f i x e d and some
o f w hich i n v e r t n o r m a l l y i n L, showed t h a t f i x e d genomes a r e
t o t a l l y h e t e r o l o g o u s f o r fib^ and 1 t h , and t h a t genoir.es w h ich
i n v e r t n o r m a l l y p o s s e s s a d d i t i o n a l c r o s s o v e r s w h ich g e n e r a t e
r e g i o n s o f t y p e - s p e c i f i c homology b e tw een TR^ and IRg .
Homology be tw een a s e q u e n c e s i s s u f f i c i e n t t o a l l o w n o rm a l
i n v e r s i o n o f L, s u g g e s t i n g t h a t s e g m e n t i n v e r s i o n o c c u r s by a
s i t e - s p e c i f i c e v e n t i n o r n e a r t h e a s e q u e n c e . A l e s s
e x p e n s i v e a n a l y s i s u f n_>i, a r e c o m b i n a n t w hich i s f i x e d i n
L arid S, s u g g e s t s w a t a s i m i l a r r e q u i r e m e n t a p p l i e s t o t h e
3 s e g m e n t . As p r e d i c t e d , f i x e d b ; : l ( 2 8 - l ) s u b c l o n e s were a b l e
to g i v e r i s e l,o n o r m a l l y i n v e r t i n g p r o g e n y , but t h e r e v e r s e
c o n v e r s i o n was a s u o o s o r v e d . The a n a l y s i s was e x t e n d e d to

i n c l u d e a number oT o t n o r r e c o m b i n a n t s which were i h - i t i a l l y
t h o u g h t t o be h e t e r o l o g o u s i o r l i t J IK o r TRg/IRg, an a y e t
i n v e r t e d n o r m a l l y , n a c n o f t n e s e was fo u n d t o p o s s e s s
t y p e - s p e c i f i c homology b e tw een r e p e a t r e g i o n s by v i r t u e o f
p r e v i o u s l y u n d e t e c t e d c r o s s o v e r s . I n v e s t i g a t i o n om t n e
im m e d i a t e e a r l y p o l y p e p t i d e s i n d u c e d by 1 x 1 ( 2 8 - 1 ) s u b c l o n e s ,
i n c l u d i n g two w hich lac ' . : p a r t o f IKg o r TRp, showed t h a t
e x p r e s s i o n o f o n l y one o f t h e two r e p e a t s i s n e c e s s a r y f o r
g r o w th o f HSV i n v i t r o .

ABBiimV lATIOiMS
A u b r e v ia t io n s f o r m edia and s o lu t i o n s a r e g iv e n in th e
M a te r ia ls and th o se f o r th e genome r e g io n s o f HSV are
g iv e n on page 7 and in P ig u re A l . l . A l l tem p era tu res a re
g iv e n in d e g r e e s c e n t ig r a d e .
A a d e n in e -c o n ta in in g m o ie ty
ACV a c y c lo v ir
ATP a d e n o s in e tr ip h o s p h a te
111K baby ham ster k id n ey c e l l s
H..V b o v in e m a m m ilit is v ir u s
bp b ase p a ir s
C c y t o s in e - c o n t a in in g m o ie ty
00V ch a n n e l c a t f i s h v ir u s
cPhA com plem entary DNA
CiviV (human) c y to m e g a lo v ir u s
c . p . e . c y to p a th ic e f f e c t
c*p .m . c o u n ts p er m inute
dATP d eo x y a d en o sin e t r ip h o s p h a te
dCTP d e o x y c y t id in e tr ip h o s p h a te
dGTP d co x y g u a n o sin e t r ip h o s p h a te
PMSO d im e th y l .su lp h o x id e
Phase d e o x y r ib o n u c le a se
dPyK d eo x y p y r im id in e k in a se
dTTP th ym id in e t r ip h o s p h a te
LBV L p s te in -P a r r v ir u s
KDTA e th y ie n e d ia m in e t e t r a - a c e t i c a c id
BHY- 1 oquid h e r p e s v ir u s 1
LllV-2 oquid h e r p e s v ir u s 2
G g u a n in e -c o n ta in in g m o ie ty
GC c o n t e n t a o l e s p e r c e n t deoxyguano s in e p lu su eoxycy t i d i n e m o i e t i e s

HCkiV human c y to m e g a lo v ir u s
hr hours
HSV n erp es s im p lex v ir u s
HSV-1 n erp es s im p le x v ir u s ty p e 1
HSV-2 h erp es s im p lex v ir u s ty p e 2
HVA n e r p e s v ir u s a t e l e s
HVP h e r p e s v ir u s p a p io
HVS h e r p e s v ir u s s a im ir i
HVT h e r p e s v ir u s o f tu r k e y s
IK im m ediate e a r ly
kb k i lo b a s e s
kbp k i lo b a s e p a ir s
LHV Lucke h e r p e s v ir u s
MCMV m urine c y to m e g a lo v ir u s
MCP m ajor c a p s id p r o t e in
MJ)J3P m ajor H NA-binding p r o t e in
MTV h a r e k ’ s d i s e a s e v ir u s
m in m in u tes
m .o . i . m u l t i p l i c i t y o f i n f e c t i o n
mKNA m essen ger RHA
m .wt. m o le cu la r w e ig h t
n n u c le o t id e s
N u n s p e c if ie d n u c le o t id e (A, 0 , G or T)
H?40 h o n id e t P4-8
02 ounces
PAA p h o sp h o n o a ce tic a c id
p .f # u . p la q u e -fo r m in g u n i t s
p . i . p o s t i n f e c t i o n
PiV p se u d o r a b ie s v ir u s
Pa p u rin e
Py p y r im id in e
RNase r ib o n u c le a s e

r .p .m .
rRNA
SGMV
SI)S
SV40
syn
syn +
T
T■Lm
tRNA
t s
U
UV
v o l / v o l
v z v '
W't
w t / v o l
X
r e v o lu t io n s p er m inute
r ib o so m a l RHA
sim ia n c y to m e g a lo v ir u s
sodium d o d e c y l s u lp h a te
sim ia n v ir u s 40
s y n c y t ia l
n o n - s y n c y t ia l
th y m id in e -c o n ta in in g m o ie ty
m e lt in g tem p era tu re
t r a n s f e r RIM; A
t e m p e r a t u r e - s e n s i t iv e
u n i t s , or u r a c i l m o ie ty
u l t r a v i o l e t
volu m e/volu m e
v a r i c e l l a - z o s t e r v ir u s
w eig h t
w e ig h t/v o lu m e
u n s p e c i f ie d n u c le o t id e (A, C, G o r T)
Arrangem ent o f d a ta
T a b le s and th en F ig u r e s have been in c lu d e d a f t e r
each P a r t or S e c t io n , and a d d i t io n a l c o p ie s o f key F ig u r e s
have been in c lu d e d a t th e i n s id e back cover*

i' A j-i i ‘ ii
1 i. U! H O j j U C T I O 14

Knowledge o f h e r p e s v ir u s e s h as exx)anded c o n s id e r a b ly
d u r in g th e p a s t iew y e a r s , and th e r e fo r e th e f o l lo w in g x^&ges
s t r e s s h erx^ esvirus genome s tr u c tu r e and th e e x p r e s s io n o f th e
HSV genome in th e l y t i c a l l y in f e c t e d c e l l . N e c e s s a r i ly ,
o th e r a s i> e c ts o f eq u a l i n t e r e s t and im p o rta n ce a re d e a l t
w ith o n ly b r i e f l y .
THE HEHPESVIKUSES
C l a s s i f i c a t i o n and p a th o g e n ic i ty
h e r p e s v ir u s e s are c h a r a c t e r is e d by v ir io n m orpnology, •
genome s i z e and s i t e o f v ir u s r e p l i c a t i o n . P a r t i c l e s w ith
h e r p e s v ir u s m orphology have been o b serv ed in a w ide v a r ie t y
o f o rg a n ism s: fu n g i (Kazama and S c h o r n s te in , 1 9 7 2 j , f i s h
( e . g . ch a n n e l c a t f i s h v ir u s , CCV; Yi/olf and D a r lin g to n , 1971) >
am phib ia ( e . g . Lucke h e r p e s v ir u s , LEV; S ta c k p o le , 1 9 6 9 ) , b ir d s
( e . g . h a rem 's d i s e a s e v ir u s , MDV; C h u r c h il l and B ig g s , 1 9 6 7 ) ,
m a rsu p ia ls ( e . g . Parma w a lla b y v ir u s ; F in n ie e t a l . , 1 9 7 6 )
and a la r g e number o f mammals. S e v e r a l h e r p e s v ir u s e s have been
i s o l a t e d from non-human p r im a te s ( e . g . h e r p e s v ir u s s a im ir i ,
iiVE, and herx^esvirus a t e l e s , HVA; Barahona e t a l . , 1974;
h e r p e s v ir u s p a p io , HVP; Falk egt a l . , 1 9 7 6 ) , and f i v e from
humans (herxjes s im p lex v ir u s ty p e s 1 and 2 , HSV-1 and HSV-2;
human c y to m e g a lo v ir u s , ilGr.iV; E p s te in -B a r r v i r u s , EBV;
v a r i c e l l a - z o s t e r v ir u s , VEV).
The number o i members o f th e fa m ily H e r p e s v ir id a e h a s
in c r e a s e d aiJuce d u rin g r e c e n t y e a r s and w ith i t th e la b o u r
in a s s ig n in g an in t e r n a t io n a l n o m en cla tu re (Hoizman e t a l . ,
1978; M athews, 1 9 7 9 ) . Three s u b f a m il ie s have been su g g e s te d
wnich a re d is t in g u is h e d by h o s t r a n g e , f e a t u r e s o f th e
r e p l ic a t - iv e c y c l e , and, to some e x t e n t , genome o r g a n is a t io n .

The a lp h a h e r p e s v ir in a e p o s s e s s a v a r ia b le h o s t ran ge and a
r e l a t i v e l y sh o r t r e p l i c a t i v e c y c l e , and in c lu d e iiSV -±, HSV-2,
eq u id herx>esv irus 1 (n h V - i; , su id n e r p e s v ir u s 1 or p se u d o r a b ie s
v ir u s (P hV ), aru oov id h e r p e s v ir u s 2 o r b ov in e mammi .JL t i s
v ir u s (BivjV ). A number o f o th e r c y to m e g a lo v ir u s e s a s w e l l a s
HCi/iV c o n s t i t u t e the b e ta h e r p e s v ir in a e . w hich have a narrow
h o s t ran ge and a r e l a t i v e l y lo n g r e p r o d u c t iv e c y c l e , and
u s u a l ly grow bes u in f i b r o b l a s t s . EBV, 1IVP, HVS and HVA are
cou n ted among th e g a m a a h e r p e sv ir in a e , w hich h ave a narrow
h o s t ran ge and r e p l i c a t e in ly m p h o b la s to id c e l l s . These
v ir u s e s a re s p e c i f i c f o r e i t h e r B - o r T -ly m p h o c y te s .
B i o l o g i c a l a s p e c t s o f th e human h e r p e s v ir u s e s have
been s tu d ie d in some d e t a i l . HSV-1 and HSV-2 a re r e la t e d
g e n e t i c a l l y and a n t i g e n i c a l l y , b u t form two d i s t i n c t s e r o ty p e s
(Plummer, 1964; P a u ls and D ow dle, 1 9 6 7 ) . HSV-1 i s r e s p o n s ib le
fo r v e s ic u l a r J c s io n s o f th e l i p s and mouth and f o r o c u la r
h e r p e t ic k e r a t i t i s . More s e v e r e symptoms, and a ls o
g e n e r a l is e d f a t a l i n f e c t i o n , have been d e s c r ib e d . HSV-1 i s
m a in ta in ed in a l a t e n t form in th e t r ig e m in a l , and som etim es
o th e r , sym x>athetic g a n g l ia (B a r in g er and Sw oveland , 1973;
L on sd a le e t a l . , 1 9 7 9 ) . The v ir u s may be r e a c t iv a t e d
p e r i o d i c a l l y le a d in g to r e c u r r e n t sym ptom s. The m echanism s
by w h ich la t e n c y i s e s t a b l i s h e d and m a in ta in e d , and by w hich
r e a c t iv a t io n and r e c u r r e n c e tak e p la c e , are n o t known.
V iru s la t e n c y i s a f e a tu r e o f s e v e r a l , i f n o t a l l , h e r p e s v ir u s e s
anu t h e i r p a th o g e n e s is .
iiSV-2 i s s e x u a l ly t r a n s m it te d , l e s i o n s o c c u r r in g i n
the g e n i t a l r e g io n . There i s an a s s o c ia t io n betw een HSV-2
ana c e r v i c a l carcinom a in women (N aib e t a l . , 1 9 6 6 ; Uawls
e t a l . , 1 9 6 c i). ru..iour t is s u e has been shown, in a t l e a s t
so lie c a s e s , to o p p ress JiSV-2 s p e c i f i c a n t ig e n s and to c o n ta in

RhA com plem entary oo HbV-2 l)HA (R oyston and A u r e lia n , 1970;
P r en k e l e t a l . , 1972; E g lin e t a l . , 1 9 8 1 ) . Both HSV-1 and
HSV-2 c a u se m o rp h o lo g ic a l tr a n sfo r m a tio n o f ro d en t c e l l s in
v i t r o , e i t h e r a s u e b i l ib a te d v ir u s or a s HHA fra g m en ts
(B u ff and Rapp, 1971; D arai and Munk, 1973;
MaeNab, 1 9 7 4 ; Camacho and S p ea r , 1 978;
R eyes e t a l . , 1979; J a r iw a lla e t a l . , 1 9 80; G allow ay and
M cD ougall, 1981; I . Cameron, p e r so n a l co m m u n ica tio n ).
Tumours can be in d u ced in e x p e r im e n ta l a n im a ls by in o c u la t io n
w ith in v i t r o tran sform ed c e l l s , b u t n o t by i n f e c t i o n w ith
v ir u s p a r t i c l e s . The HSV-1 tr a n sfo r m in g r e g io n has been
mapped a t 0 .3 1 - 0 .4 2 f r a c t i o n a l genome u n i t s , and t h a t o f HSV-2
a t e i t h e r 0 . 43- 0 .3 8 or 0 . 58- 0 . 6 2 , d ep en d in g on th e tr a n sfo r m a tio n
a s s a y em p loyed . The n a tu r e o f tr a n s fo r m a tio n by HSV a t th e
m o le c u la r l e v e l rem ain s o b sc u r e .
1ICMV i n f e c t i o n i s u s u a l ly m ild , a lth o u g h i t may be
s e v e r e in n e o n a te s , and t h i3 v ir u s h a s been a s s o c ia t e d w ith
K a p o s i’ s sarcom a (G ira ld o e t a l . , 1 9 7 5 ) . VZV c a u s e s c h ick e n
p o x , u s u a l ly in c h ild h o o d , and, in a d u lth o o d , a p a in f u l l o c a l
v e s ic u l a r c o n d it io n known a s s h i n g l e s . VZV h a s been r e p o r te d
to tra n sfo rm c e l l s in v i t r o (G elb e t a l . , 1 9 8 0 ) . EBV i s
B -ly m p h o cy te s p e c i f i c and c a u s e s i n f e c t i o u s m o n o n u c le o s is
and i s in v o lv e d in th e a e t io lo g y o f two tum ours: B u r k i t t f s
lymphoma, p erh ap s prom oted by endem ic m a la r ia , and
n a so p h a ry n g e a l ca rc in o m a , in a s s o c i a t io n w ith a g e n e t ic
p r e d is p o s i t io n (Old e t a l . , 1966; H enle e t a l . , 1968 and 1 9 6 9 ) .
PRV i s a p a r a s i t e o f sw ine and i s u s u a l ly in a p p a r e n t ,
but g iv e s r i s e to th e a la rm in g symptoms o f ,fmad i t c h ” and
then d e a th in c a t t l e . EHV-1 c a u s e s p reg n a n t m ares to a b o r t .
Those two v i r u s e s have n o t y e t been im p lic a te d in
tu m o u r tg e n e s is in v iv o , in c o n t r a s t w ith s e v e r a l o th e r n o n -
human h e r p e s v ir u s e s . i.DV c a u se s a h ig h ly c o n ta g io u s

n eu ro lym p h om atosis in c i i ic x e n s , now c o n t r o l le d by im m u n isa tion
w ith th e r e la t e d h e r p e s v ir u s o f tu r k e y s (HVT; Marek, 190'7;
P u rch ase e t a l . , 1 9 7 1 ) . LHV p rod u ces a r e n a l adenocarcinom a
in f r o g s , th e tumour and v ir u s grow th d e m o n str a tin g opposed
s e a s o n a l d ep en d en ce (L ucke, 1934; P a w c e tt , 1 9 5 6 ) . H erp esv ir u s
s i l v i l a g u s c a u s e s a ly m p h o p r o l i f e r a t iv e d i s e a s e in c o t to n
t a i l r a b b i t s (H in z e , 1 9 7 1 ) . The T -lym phocyte s p e c i f i c Hew
World s im ia n h e r p e s v ir u s e s , HVS and HVA, in v a r ia b ly produce
m a lig n a n t lymphomas in c e r t a in h e te r o lo g o u s p r im a te h o s t s
(M elendez e t a l . , 1969 and 1 9 7 2 ) .
S tr u c tu r e o f th e v ir io n
The h e r p e s v ir io n co m p r ise s fo u r m o r p h o lo g ic a lly d i s t i n c t
s t r u c t u r e s : th e c o r e , c a p s id , tegum ent and e n v e lo p e . The co r e
c o n t a in s th e d o u b le -s tr a n d e d DNA genome (E p s te in , 1962a;
B en -P o ra t and K aplan, 1962) t o r o i d a l l y arran ged around a
c e n t r a l p r o te in a c e o u s m a tr ix (C h a i, 1971; F u rlo n g e t a l . >
1972; H a z e r ia n , 1974; H eine and C o t t le r -F o x , 1 9 7 5 ) .
P leom orp h ic fo rm s, p erh ap s r e p r e s e n t in g d i f f e r e n t d ev e lo p m e n ta l
s t a g e s , h ave a l s o been o b se rv ed ( N i i and Y asuda, 1975; Okada
e t a l* , 1 9 7 9 ) . The ic o s a h e d r a l c a p s id w h ich surrou n d s th e
co r e i s a p p r o x im a te ly 100 nm in d ia m e te r , and c o n s i s t s o f
162 cap so m eres i n 5 :3 :2 a k ia l symmetry (W ildy e t a l . , I 9 6 0 ) .
The ca p so m eres a re h o llo w e lo n g a te d p r ism s , and in te r c a p s o m e r ic
f i b r i l s h ave been o b serv ed ( W ildy e t a l . , 1 960; Vernon e t a l . ,
1 9 7 4 ) . A f ib r o u s la y e r known a s th e tegum ent su rrou n d s th e
c a p s id (Hoizman and F u r lo n g , 1 9 7 4 ) , th e w id th o f w hich i s
d eterm in ed a t l e a s t- in p a r t by th e v ir u s (McCombs e t a l . , 1 9 7 1 ) .
The f r a g i l e v ir io n en v e lo p e c o n s i s t s o f a t r i la m e l l a r
membrane w ith s p ik e s p r o j e c t in g from th e o u te r s u r fa c e
( Wildy e t a i . , 19 oO)•

H e r p e s v ir io n s c o n ta in 1 8 -3 3 p o ly p e p t id e s o f m .w t.
1 1 ,0 0 0 -2 9 0 ,0 0 0 (S p ear and Roizm an, 1972; H eine e t a l . , 1974;
Perdue e t a l • , 1974; S t e v e ly , 1975; S tm a d and A u r e lia n ,
1976; Kim e t a l . , 1976a and b; D o ly n iu k e t a l . , 1976; D ixon
and P a rb er , 1 9 8 0 ) . About h a l f o f th e s t r u c t u r a l p r o t e in s o f
HSV-1 have been a s s ig n e d r e l a t i v e p o s i t i o n s in th e v ir io n
on th e b a s i s o f ex p er im en ts in v o lv in g s e l e c t i v e rem oval o f
p o ly p e p t id e s by d e te r g e n t tr e a tm e n t (Roizman and F u rlo n g ,
1 9 7 4 ) , d i f f e r e n t i a l c h e m ic a l tr e a tm e n t (O lsh ev sk y and B eck er ,
1972; Roizman and F u r lo n g , 1974) and v i r u s n e u t r a l i s a t i o n
(P o w e ll e t a l . , 1974; Cohen e t a l . , 1 9 7 8 ) .
The e n v e lo p e c o n ta in s m ost o r a l l o f th e v ir u s -c o d e d
s t r u c t u r a l g ly c o p r o t e in s (S p ea r and Roizm an, 1 9 7 2 ) , c e l l
l i p i d s (A sher e t a l • , 1 9 6 9 ) , an ATPase p ro b a b ly o f c e l l u l a r
o r ig in (E p s te in and H o lt , 1 9 6 3 ) , and sp erm id in e (G ibson and
Roizm an, 1 9 7 1 ) . In c o n t r a s t to o th e r v i r i o n p r o t e in s ,
e l e c t r o p h o r e t ic p r o f i l e s o f c a p s id p r o t e in s from v a r io u s
h e r p e s v ir u s e s a r e s im i la r . Cohen e t a l . (1980b) i d e n t i f i e d
sev en m ajor c a p s id p o ly p e p t id e s in HSV-1, r a n g in g in m .w t.
from 1 2 ,0 0 0 to 1 5 5 ,0 0 0 , and Zweig e t a l . (1 9 7 9 ) showed t h a t
th e m ajor c a p s id p r o t e in ( 1 5 5 , 0 0 0 ) p a r t i c ip a t e s in d is u lp h id e
b on d in g in th e i n t e g r a l v i r i o n s t r u c t u r e . G ibson and
Roizman (1972 and 1974) p r e se n te d e v id e n c e f o r th e p r o c e s s in g
o f a p h o sp h o r y la te d c a p s id p r o te in to a l e s s t i g h t l y bound
form d u r in g v ir u s m a tu r a tio n . A c y c l i c n u c le o t id e -
in d ep e n d en t p r o t e in k in a s e i s ca p a b le o f p h o sp h o r y la t in g
ab ou t sev en s t r u c t u r a l p o ly p e p t id e s , in c lu d in g a c a p s id
p r o t e in (R u b en ste in e t aJL., 1972; le m a s te r and Roizm an, 1 9 8 0 ) .
The s ig n i f i c a n c e in v ir u s m a tu ra tio n and i n f e c t i v i t y o f th e s e
o b s e r v a t io n s i s n o t known. P o ly p e p t id e s w hich have n o t been
lo c a t e d in th e c a p s id o r en v e lo p e a re th o u g h t to r e s id e in
th e tegu m en t. Only one p r o te in h a s been s u g g e s te d a s a

s t r u c t u r a l com ponent o f th e c o r e , in a d d it io n to th e p o ly a m in e ,
sperm in e (G ib son and Roizm an, 1 9 7 1 ) .
S tr u c tu r e o f th e HSV genome
DNA e x tr a c te d from HSV-1 v i r i o n s i s a l i n e a r d u p lex
w ith a m .w t. o f a p p ro x im a te ly 100 x 1 0 5 a s d eterm in ed by
s e d im e n ta t io n , e le c t r o n m ic r o s c o p ic , and r e s t r i c t i o n
e n d o n u c le a se a n a ly s e s (B eck er e t a l . , 1 968; K ie f f e t a l • ,
1971; W ilk ie , 1973; G rafstrom e t a l . , 1 9 7 4 ; Wadsworth e t a l . ,
1975; C lem en ts e t a l . , 1976; W ilk ie , 1 9 76; Skare and Summers,
1 9 7 7 ) . I t h a s a d eo x y g u a n o sin e p lu s d e o x y c y t id in e (GC) c o n te n t o f
67 ft ( K ie f f e t a l . , 1 9 7 1 ) and, u n l ik e c e l l DNA, d o e s n o t have
a s c a r c i t y o f th e n u c le o t id e d o u b le t CpG (Subak-Sharpe e t a l . ,
1 9 6 6 ) . B oth s in g l e and d o u b le stra n d ed DNA a re i n f e c t i o u s
in c e l l m o n o la y ers (Lando and R y h in er , 1 9 6 9 ; Graham e t a l . ,
1973; S h e ld r ic k e t a l . , 1973; W ilk ie e t a l . , 1974; P a rb er ,
1976; Stow and W ilk ie , 1 9 7 6 ) .
The l a b i l i t y o f HSV-1 DNA in a l k a l i h a s been a t t r ib u t e d
to r ib o n u c le o t id e s c o v a le n t ly bound t o th e DNA, d e te c te d by
th e in c o r p o r a t io n o f r a d io a c t iv e u r id in e i n t o m ature DNA
( I l ir s c h and Vonka, 1974; M u ller e t a l . , 1 9 7 9 a ) . T h is
c o n c lu s io n i s in su p p o rt o f th e work o f G ordin e t a l . (1 9 7 3 )
who r e p o r te d th a t formamide d en aturation ^ r e s u l t e d in i n t a c t
s in g l e s t r a n d s . On th e o th e r hand, Hyman e t a l . (1 9 7 7 )
found t h a t th e m a jo r ity o f a l k a l i - l a b i l e s i t e s c o u ld be
r e p a ir e d by DNA p o ly m era se and DNA l i g a s e , and E cker and
Hyman ( 1 9 8 1 ) "claim ed th a t s in g l e s tr a n d s w ere fragm en ted even
u s in g g e l e l e c t r o p h o r e s i s sy stem s w hich l e a v e r ib o s e m o ie t i e s
i n t a c t . T hese a u th o r s co n c lu d ed th a t m ost o r a l l a l k a l i -
s e n s i t i v e s i t e s a re due to n ic k s o r gaps in th e s in g l e
s tr a n d s . T h is c o n c lu s io n , how ever, dep en d s on th e assu m p tio n
th a t th e DNA was n o t damaged upon r e l e a s e from v i r i o n s .

P ren k e l and Roizman (1 9 7 2 a )co n c lu d ed from r e n a tu r a t io n
s t u d ie s t h a t IISV-1 DNA c o n ta in s u n iq u e ly s i t u a t e d n ic k s on
o n ly one s tr a n d o f th e d u p le x , r a th e r l i k e b a c te r io p h a g e
15 . W ilk ie (1 9 7 3 ) co n c lu d ed from s im i la r e x p er im e n ts t h a t
in t e r r u p t io n s a re lo c a t e d on both s tr a n d s w ith e q u a l d i s t r i b u t i o n ,
and m oreover showed by com parison w ith b a c te r io p h a g e T5 DNA
th a t th ey a re random ly s i t u a t e d on both s tr a n d s (W ilk ie e t
a l . . 1 9 7 4 ) . The work o f E cker and Hyman (1 9 8 1 ) , w ith th e
c a v e a t m en tion ed a b o v e , i s in su p p o rt o f t h i s v ie w .
B en -P o ra t e t a l . (1 9 7 9 ) r e p o r te d th a t in t e r r u p t io n s in PRV
DUA a r e lo c a t e d a t random in th e s in g le s t r a n d s .
HSV-1 DNA i s n o t c i r c u l a r l y perm uted (H ir sc h e t a l . ,
1 9 7 5 ) . R e n a tu r a tio n o f HSV-1 DNA w hich h a s been t r e a te d w ith
p r o c e s s iv e e x o n u c le a s e s , such a s 3 ‘- e x o n u c le a s e I I I or
lambda 5 1- e x o n u c le a s e , r e s u l t s in th e fo rm a tio n o f d ou b le
s tra n d ed c i r c l e s ( S h e ld r ic k and B e r t h e lo t , 1 9 7 4 ; G rafstrom
e t a l . , 1974 and 1975; Wadsworth e t a l . , 1976.; Hyman e t a l . ,
1976; K udler and Hyman, 1 9 7 9 ) . T h is i s in t e r p r e t e d a s th e
p r e se n c e o f a d i r e c t r e p e t i t i o n a t th e two genome te r m in i ,
th e s i z e o f w hich f o r v a r io u s s t r a i n s o f HSV-1 h a s been
e s t im a te d to be 4 0 0 -7 5 0 0 bp, a lth o u g h th e more r e l i a b l e
e s t im a t e s p la c e i t betw een 400 and 1600 bp .
Upon ex a m in a tio n o f a n n ea led i n t a c t s i n g l e s tr a n d s o£
HSV-1 DNA in th e e le c t r o n m ic r o sc o p e , S h e ld r ic k and B e r th e lo t
(1 9 7 4 ) found s t r u c t u r e s c o n s i s t e n t w ith th e f o l lo w in g m od el.
The genome c o n s i s t s o f two c o v a le n t ly jo in e d segm en ts
(L and S ) , each o f w hich co m p r ise s a u n iq u e r e g io n (Uj, and
Ucj) bounded by in v e r te d r e p e t i t i o n s (TRp, and IRj,, TRg and IR s)*
a s sum m arised in P ig u re A l . l . They s u g g e s te d t h a t th e two
segm en ts m ight in v e r t by in tr a m o le c u la r r e c o m b in a tio n ,
th e re b y g e n e r a t in g fo u r d i s t i n c t genome a rra n g em en ts .
A lth ough th e s e w orkers supposed th a t TR^/IR^ and

- 8 -
are i d e n t i c a l , f u r th e r e le c t r o n m ic r o s c o p ic , d e n a tu r a t io n ,
and r e s t r i c t i o n en d o n u c lea se a n a ly s e s showed th a t th e two
s e t s o f r e p e a t s a re d i f f e r e n t (Wadsworth e t a l • , 1975;
Hayward e t a l . , 1 9 7 5 b ;D e liu s and C lem en ts , 1976; C lem ents
e t a l . , 1976; ’W ilk ie and C o r t in i , 1 9 7 6 ) . T hese r e s u l t s a ls o
con firm ed t h a t v ir io n DNA c o n ta in s fo u r eq u im o la r genome
arran gem en ts r e s u l t i n g from th e a b i l i t y o f L and S to in v e r t
r e l a t i v e to each o t h e r . The e s ta b lis h m e n t o f r e s t r i c t i o n
en d o n u c lea se maps showed t h a t h a l f - and q u a r te r -m o la r
fra g m en ts map a t th e te r m in i o f L and S and t h e i r m utual
j o in t , a s p r e d ic t e d by th e model and i l l u s t r a t e d in F ig u re ■
A i . l (W ilk ie , 1 976; W ilk ie e t a l . , 1 9 7 7 a ;S k a re and Summers,
1 9 7 7 ) .
The te r m in a l r e p e t i t i o n o f HSV-1 DNA, o r a se q u e n c e ,
i s a l s o p r e s e n t in in v e r s e o r ie n t a t io n a t th e j o i n t betw een
L and S ( f ig u r e A l . l ) . Wagner and Summers (1 9 7 8 ) o b ta in e d
d e t a i l e d r e s t r i c t i o n maps o f t h i s r e g io n , and e s t im a te d th e
s i z e o f th e a seq u en ce in HSV-1 s t r a in KOS to be 265 bp .
Two ty p e s o f s i z e h e t e r o g e n e i t y a re a s s o c ia t e d w ith th e
te r m in i and j o in t r e g io n . The f i r s t c o r r e sp o n d s to i n s e r t i o n s
o f a p p r o x im a te ly 300 bp, th o u g h t to c o n ta in th e a se q u e n c e ,
a t th e L term in u s and j o in t (W ilk ie , 1976; Wagner and Summers,
1978; L ocker and F r e n k e l, 1 9 7 9 b ) . T h is h a s a l s o been
o b serv ed f o r th e HSV-2 genome (W ilk ie e t g l« » 1 9 7 7 a ) . The
secon d ty p e o f h e te r o g e n e ity c o n s i s t s o f v a r ia b le i n s e r t i o n s
o f 1 0 -5 0 bp w ith in th e a seq u en ce and th e a d ja c e n t c
(TK^/IKg) seq u en ce ( ’Wagner and Summers, 1 9 7 8 ) .
R en a tu r a tio n o f e x o n u c le a s e - t r e a t e d HSV—1 DNA r e s u l t s ,
in some in s t a n c e s , in a sm a ll lo o p a t one te r m in u s , in t e r p r e te d
a s th e p r e se n c e o f p a r t o f th e w hole o f th e a seq u en ce in
in v e r s e o r ie n t a t io n a few hundred bp away from th e term in u s
(W adsworth‘e t a i . , 1976; Hyman e t a l . , 1 9 7 6 ) . W ilk ie and

C o r t in i (1 9 7 6 ) s u g g e s te d from experim ents- in v o lv in g en d -
l a b e l l i n g o f IiSV-1 DNA th a t t h i s s t r u c t u r e i s p r e s e n t a t
th e L te r m in u s . H i l l e r e t a l . (1 9 8 0 ) have o b se rv ed a w ide
v a r ie t y o f fo ld b a c k and s te m -lo o p s t r u c t u r e s in r e a n n e a le d
s in g l e s tr a n d s , some o f w hich co rresp o n d to r e c o g n is e d
r e p e a t s w ith in th e genom e, and o th e r s o f w h ich a re o f unknown
s i g n i f i c a n c e .
K udler and Hyman (1 9 7 9 ) o b serv ed t h a t p a r t i a l l y
d e p r o t e in is e d DNA i s more r e s i s t a n t to 3 1- e x o n u c le a s e than
to 5 *- e x o n u c le a s e , and su g g e s te d t h a t t h i s was due to a
p r o t e in bound a t th e 3 1 t e r m in i . In d ee d , fo u r s p e c i f i c
p r o t e in s rem ain n o n -c o v a le n t ly a s s o c ia t e d w ith HSV-l DNA
f o l lo w in g d e te r g e n t tr ea tm e n t (Hyman, 1 9 8 0 ) . Wu e t a l* (1 9 7 9 )
a t ta c h e d d in itr o p h e n y l (DNP) grou p s to th e bound p r o t e in s
and v i s u a l i s e d them in th e e le c t r o n m icro sco p e by tr e a tm e n t
w ith an anti-D N P im m u n og lob u lin . They found p r e f e r e n t i a l
b in d in g c l o s e to th e two te r m in i and a t two p o s i t i o n s
c o r r e sp o n d in g to th e a seq u en ce a t th e j o i n t , and p rop osed
th a t a p r o t e in s p e c i f i c a l l y i n t e r a c t s w ith th e a s e q u e n c e .
The s ig n i f i c a n c e o f t h i s o b s e r v a t io n h a s y e t to be e lu c id a te d *
HSV-2 DHA h as a 2 ft h ig h e r GC c o n te n t th an HSV-l DNA
(G oodheart e t a l . , 1968; K ie f f e t a l . , 1971; H a ll ib u r to n ,
1 9 7 2 ) . A p p rox im ately 50 o f HSV-l and HSV-2 DNA se q u e n c e s
a re hom ologous w ith 6 5 -8 5 ^ m atch in g o f n u c le o t id e s ( K ie f f
e t a l . , 1972; F ren k e l e t a l . , 1973; Sugino and K in gsb u ry , 1 9 7 6 ) .
The genom es o f th e two s e r o ty p e s a re s t r u c t u r a l l y v e r y
s im i la r , but p o s s e s s d i s t i n c t i v e arran gem en ts o f r e s t r i c t i o n
s i t e s (Hayward e t a l • , 1975a and b; Skare e t* a l * , 1 9 7 5 ;
C o r t in i and W ilk ie , 1 9 7 8 ) . These d i f f e r e n c e s have been
e x p lo i t e d a s a d ia g n o s t ic t o o l f o r i d e n t i f i c a t i o n o f HSV-l
and HSV-2 i s o l a t e s and f o r s tu d y in g e p id e m io lo g ic a l a s p e c t s
o f o v e r t and l a t e n t i n f e c t i o n s in man (linnem an e t a l . , 1978;

Buchman e t a l . , 1978 and 1980; L o n sd a le e t a l • , 1979 and 1980;
L o n sd a le , 1 9 7 9 ) . R e s t r ic t io n fra g m en ts r e p r e s e n t in g th e
m a jo r ity o r e n t i r e t y o f th e IISV-1 and HSV-2 genom es have
r e c e n t ly been c lo n e d in t o b a c t e r ia l p la s m id s , b o th in Glasgow
and e lse w h e r e (P o s t e t a l . , 1980; G allow ay and Sw ain , 1980;
G old in . e t a l . , 1 9 8 1 ) .
S tr u c tu r e s o f o th e r h e r p e s v ir u s genom es
A lth ough a number o f genome s t r u c t u r e s have been
d em o n stra ted among th e h e r p e s v ir u s e s , in each c a s e a l i n e a r
d u p le x , t h e s e may be r e p r e s e n te d i n c i r c u l a r form in o rd e r to
show th e fo u r b a s ic o r g a n is a t io n s found among th o s e so f a r
a n a ly se d (F ig u r e A 1 .2 ) . GC c o n te n ts o f h e r p e s v ir u s DNAs are
g iv e n in F ig u re A 1 .3 .
Two subgroups are. in c lu d e d in group I b e c a u s e , a lth o u g h
th ey a re d i s t i n c t , i t i s p o s s ib l e t h a t th e y r e f l e c t a b a s ic
s i m i l a r i t y . The f i r s t i s common to H SV -l, HSV-2, BMV and
UCMV. BkV DNA p o s s e s s e s 1 4 # hom ology w ith HSV-l DNA (S te r z
e t a l . , 1 9 7 4 ) and has a ra.wt. o f a p p ro x im a te ly 90 x 1 0 ^,
p o s s e s s in g s m a lle r in v e r te d r e p e a t s th an HSV-l DNA (Buchman
and Roizm an, 1 9 7 8 a ) . The BIvlV genome h as a te r m in a l r e p e t i t i o n
o f a p p r o x im a te ly 1000 bp (Buchman and Roizm an, 1 9 7 8 b )• The
genome o f HCiuV has a m.wt*. o f 150 x 10^ (D em archi e t a l . , 1978
G eelen e t a l . , .1978; S t in s k i e t a l . , 1979; W e s ts tr a te e t a l . ,
1 9 8 0 ) , a lth o u g h i t was e a r l i e r r e p o r te d t h a t HCMV DNA h a s a
m .w t. o f 100 x 10^ (S arov and Friedm ann, 1 976; K i lp a tr ic k
e t a l . , 1 9 7 6 ; Huang e t a l . , 1 9 7 3 ) . K i lp a tr ic k and Huang
(1 9 7 7 ) d e t e c te d both s i z e c l a s s e s , and Lakeman and Osborn
(1 9 7 9 ) a fu r th e r c l a s s o f 130 x 1 0 ^. i t i s c l e a r th a t th e
s i z e c l a s s o f 150 x 1 0 6 r e p r e s e n ts th e f u l l l e n g th genome,
b oth from d e te r m in a t io n s o f i t s i n f e c t i v i t y and from
r e s t r i c t i o n maps (G eelen e t a l . , 1978; Lakeman and O sborn,
1979; W e s ts tr a te e t a l . , 1 9 8 0 ) . The s m a lle r s i z e s o f DNA

- 11 -
m o le c u le s a re thought to r e s u l t from d e f e c t i v e v i r i o n s .
The HCMV genome i s s t r u c t u r a l ly s im i la r to th a t o f HSV-l
(W e s ts tr a te e t a l . , 1 9 8 0 ) . and Us a r e la r g e r than in H SV -l,
and th e r e g io n o f l e a s t hom ology betw een two c l o s e l y r e la t e d
s t r a i n s i s in rfR g/IR g (com p arison o f th e r e s u l t s o f P r i t c h e t t ,
1980 w ith th e r e s t r i c t i o n maps o f W e s ts tr a te e t a l . , 1 9 8 0 ) .
Jean e t a l . (1 9 7 8 ) have p o s tu la te d from i n d i r e c t e v id e n c e t h a t
HCMV PH A i s t e r m in a l ly red u n d a n t. R e s t r i c t io n p r o f i l e s o f
s im ia n CMV (SCMV) i s o l a t e s d i f f e r s i g n i f i c a n t l y from th o se
o f HCMV ( iC ilp a tr ic k e t a l . , 1 9 7 6 ) , and th e r e h as been a t
l e a s t one c a s e o f a c y to m e g a lo v ir u s i s o l a t e d from a human
p a t ie n t who d ied from c l i n i c a l e n ce p h a lo p a th y w h ich bore
a lm o st t o t a l n u c le ic a c id hom ology w ith SCMV s t r a i n s but none
w ith HCMV (Huang e t a l . , 1 9 7 8 ) . Mosmann and Hudson (1973)
r e p o r te d th a t th e DNA o f m urine CMV (MCMV) h a s a m .w t. o f6132 x 10 . T aking in t o a cc o u n t t h e i r u n d e r e s t im a te o f th e
s i z e o f th e HSV-l genom e, t h i s i s e q u iv a le n t t o 150 x 10 .
However, genome s t r u c t u r e s o f MCMV, SCMV and th e c y to m eg a lo
v ir u s iilIV-2 ( Wharton e t a l * , 1981) have n o t been p u b lis h e d .
N e v e r t h e le s s , i t i s c l e a r t h a t th e HSV ty p e o f genome
o r g a n is a t io n i s found in r e p r e s e n t a t iv e s o f b o th a lp h a -
and b e t a - n e r p e s v ir in a e .
The secon d subgroup o f group I c o n t a in s th e PRV and
BIIV-1 genom es, w hich a re s im i la r in s i z e to t h a t o f HSV-l
but la c k in v e r te d r e p e a t s f la n k in g th e 1 seg m en t, and in
w hich o n ly S in v e r t s ( S t e v e ly , 1977; P o w e ll , 1979; B en -P orat
e t a l . , 1979; Soehner e t a l . , 1965; W halley e t a l . , in p r e s s ) .
There i s 110 d i r e c t e v id e n c e t h a t e i t h e r PRV o r EHV-1 DNA i s
t e r m in a l ly red u n d a n t, a s i t h as n o t proved p o s s ib l e to
c i r c u l a r i s e e i t h e r a f t e r e x o n u c le a se I I I tr e a tm e n t (Ben—P o r a t
e t a l . , 1979; P . S h e ld r ic k , p e r s o n a l co m m u n ica tio n ).
Group I I c o n ta in s EBV and i t s s im ia n c o u n te r p a r t , HVP.

The EBV genome has a m .w t. o f a p p r o x im a te ly 110 x lO *’3
( P r i t c h e t t e t a l . , 1975; Given and K ie f f , 1978 and 1 9 7 9 ) .
The te r m in i a re redundant and o f v a r ia b le s i z e , c o n ta in in g
up to 12 d i r e c t r e p e a t s o f a 500 bp seq u en ce (G iven and K ie f f ,
1 978; G iven e t a l . , 1979; Kimtner and Sugden, 1 9 7 9 ) , and , a s
w ith H SV -l, th e l i n e a r genome can be c i r c u l a r i s e d v i t r o by
a n n e a lin g f o l lo w in g e x o n u c le a se tr e a tm e n t (K in tn e r and Sugden,
1 9 7 9 ) . C ir c u la r DBA produced by r e p r e s e n t a t iv e s o f group I
has so f a r been d e te c te d o n ly in t h i s f a s h io n or in r e p l i c a t in g
IMA d u r in g l y t i c i n f e c t i o n , b ut E B V -transform ed c e l l s have
been shown to c o n ta in EBV genom es a s e p iso m a l c o v a le n t ly
c lo s e d c i r c l e s (L in d a h l e t a l • , 1 9 7 6 ) . The two u n iq u e
segm en ts o f th e l i n e a r genome do n o t in v e r t (G iven and K ie f f ,
1979; Hayward eti a l . , 1980) and a re se p a r a te d by a p p ro x im a te ly
10 d i r e c t r e p e a t s o f 3 kbp (IK ), w hich a r e G C -rich and n o t
hom ologous to th e te r m in a l r e p e a t s (Sugden , 1 977; Rymo and
Porsblum , 1978; Given and K ie f f , 1979; Hayward e t a l . , 1 9 8 0 ) .
G iven and K ie f f (1 9 7 9 ) d e te c te d in t e r n a l hom ology w ith in IR,
and co n c lu d ed th a t th e 3 kbp seq u en ce i t s e l f c o n ta in s a
r e p e a te d u n i t . They a l s o d e te c te d hom ology betw een IR and
two l o c a t io n s in L. The v a r io u s s t r a in s o f EBV d i f f e r in
t h a t some la c k se q u e n c es p o s s e s s e d by o th e r s ( P r i t c h e t t e t a l . ,
1975; Sugden e t a l . , 1 9 76; G iven and K ie f f , 1978; Rymo and
Porsblum , 1978; Raab-Traub e t a l , 1978 and 1980; Hayward e t
a l . , 1 9 8 0 ; Bomkamm e t a l . , 1980; H e l le r et. a l . , 1983a)..
Some s t r a i n s a re u n a b le to tra n sfo rm c e l l s , but i t rem a in s to
be shown p r e c i s e l y w h ich r e g io n s o f th e genome a r e e s s e n t i a l
fo r tr a n s fo r io a t io n .
HVP e x h i b i t s a p p ro x im a te ly 4 0 DNA hom ology w ith EBV,
a s do th e gam m aherpesvirinae o f o th e r Old World p r im a te s
such a s th e oran gu tan (h e r p e s v ir u s p o n g o ), th e ch im panzee
(h e r p e s v ir u s pan) and th e g o r i l l a (h e r p e s v ir u s g o r i l l a )

- 13 -
(G erber e t a l . , 1976; F a lk e t a l . , 1976; R abin e t g l . , 1978;
Heubauer e t a l . , 1979; Lee e t a l . , 1 9 8 0 ) . R e s t r i c t io n mapping
o f IIVP DNA and h y b r id is a t io n , u s in g in one c a s e c lo n e d EBV DNA
fr a g m e n ts , have shown t h a t th e two genom es a re s t r u c t u r a l ly
i d e n t i c a l and c o l in e a r (Lee e t a l . , 1981; H e l le r e t a l . , 1981b;
H e l le r and K ie f f , 1 9 8 1 ) .
The genom es o f HVS and HVA, w h ich a r e in group I H ,
b oth c o n s i s t o f a s in g l e u n iq u e r e g io n o f 3 5 # GC bounded on
b oth s id e s by m u lt ip le d i r e c t r e p e a t s o f a seq u en ce o f
a i> p rox im ately 1500 bp c o n ta in in g 7 0 $ GC ( F le c k e n s te in and
W olf, 1 9 7 4 ; F le c k e n s te in e t a l . , 1975 and 1978; BornXamm
e t a l . , 1 9 7 6 ) . As w ith EBV, H V S-transform ed c e l l s c o n ta in
e p iso m a l c lo s e d c i r c u la r HVS genom es, a lth o u g h such m o le c u le s
have a t l e a s t i n one c a s e been shown to co m p rise rea rra n g em en ts
o f th e HVS genome (Werner e t a l . , 1 9 7 7 ) . I a th e l i n e a r
genom e, th e te r m in a l r e p e a t s a re a p p r o x im a te ly c o n s ta n t in
num ber, a c c o u n t in g f o r 3 0 $ o f th e genome s i z e , but can be
d i s t r i b u t e d a sy m m e tr ic a lly a t th e two t e r m in i . The HVS and
HVA genom es have a m .w t. o f 90 x 1 0 6 , and d e f e c t i v e genom es
com posed e n t i r e l y o f r e i t e r a t i o n s o f th e te r m in a l r e p e a t
have been c h a r a c t e r i s e d .
CCV i s th e o n ly i d e n t i f i e d member o f group IV .
C housterm an e t a l . (1 9 7 9 ) p r e se n te d r e s t r i c t i o n maps o f CCV
DNA, show ing t h a t th e genome c o n s i s t s o f a u n iq u e r e g io n o f
62 x 1 0 in m .w t ., bounded on each s id e by a d i r e c t r e p e a t
o f 12 x 1 0 ^ . Ho m inor bands were d e t e c t e d and , u n l ik e IIVS
and ilVA, th e te r m in i were n o t h ig h ly r e i t e r a t e d .
The LEA o f two o th e r h e r p e s v ir u s e s has to some e x t e n t
been c h a r a c t e r i s e d , but d e te r m in a t io n o f t h e ir genome
s t r u c t u r e s lias been hampered by d i f f i c u l t i e s in c l o n a l l y

- 14 -
p u r i f y in g th e s e v ir u s e s from p o s s ib l e d e f e c t i v e v i r u s .
VZV DNA h as a m .w t. o f 8 0 -1 0 0 x 10 6 and r e s t r i c t i o n p r o f i l e s
o f s e v e r a l c l i n i c a l i s o l a t e s a re v er y s im i la r (O akes e t a l . ,
1 9 7 2 ; l i t i s e t a l . , 1977; Rapp e t a l . , 1 9 7 7 ; R ich a rd s e t a l . ,
1 979; Dumas e t a l . , 1 9 8 0 ) . Rapp e t a l . (1 9 7 7 ) f a i l e d to
d e t e c t r e p e a te d seq u e n c es by k i n e t i c h y b r id i s a t io n , but
subm olar r e s t r i c t i o n fra g m en ts have been d e t e c t e d .
IviDV DNA lias a m .w t. s im i la r to t h a t o f H SV -l, and subm olar
r e s t r i c t i o n fra g m en ts have been d e te c te d (Lee e t a l . , 1971;
B achenheim er e t a l . , 1 9 7 2 ; H ir a i e t a l . , 1 9 7 9 ) .
THE LYTIC CYCLE OP HERPES SIMPLEX VIRUS
I n t e r a c t io n o f v ir u s w ith th e c e l l
A fte r e n tr y o f HSV in t o th e p e r m is s iv e c e l l , th e DNA
e n t e r s th e n u c le u s , where i t i s t r a n s c r ib e d and th e mRNA
tr a n sp o r te d to th e c y to p la sm . M ost s y n t h e s is e d p o ly p e p t id e s
a re tr a n sp o r te d in t o th e n u c le u s where DNA r e p l i c a t i o n and
v ir u s a ssem b ly ta k e p la c e . In th e c o u r se o f i n f e c t i o n , HSV
i n h i b i t s h o s t c e l l m acrom olecu lar s y n t h e s i s . V iru s may
spread by r e l e a s e o f i n f e c t i o u s p a r t i c l e s from th e c e l l o r
by c e l l f u s i o n .
H erpes s im p lex v i r f o n s w i l l a d so rb to th e c e l l s u r fa c e
a t 4 ° , but p e n e t r a t io n r e q u ir e s a h ig h e r tem p era tu re
(Fam ham and Newton, 1959; Holmes and W atson, 1 9 63; Huang
and Wagner, 1 9 0 4 ) . The r o u te by w hich v i r i o n s e n te r th e
c e l l has r a is e d c o n s id e r a b le f u r o r e . I t was co n c lu d ed fronr
s e v e r a l e a r l i e r e le c t r o n m ic r o sc o p ic s t u d ie s t h a t th e v ir u s
i s ta k en up by p in o c y t o s i s in t o v a c u o le s (H olm es and W atson,
1963; ilummeler e t a l . , 1969; D a le s and S i lv e r b e r g , 1 9 6 9 ) .
On th e o th e r hand, morgan e t a l . (1 9 6 8 ) i n s i s t e d t h a t v ir u s
e n tr y o c c u r s by d i r e c t f u s io n w ith th e c e l l membrane.

- 13 -
S u b seq u en t work i s c o n s i s t e n t w ith th e p r e se n c e o f s p e c i f i c
v ir u s r e c e p t o r s on th e c e l l s u r fa c e (H ochberg and B eck er , 1968
B lom berg, 1979; Vahlne e t a l . , 1 9 7 9 ) . M oreover, e v id e n c e
t h a t v i r io n g ly c o p r o t e in s fu n c t io n in a d s o r p t io n and
p e n e t r a t io n makes d i r e c t f u s io n th e more t e n a b le H y p o th e s is
a t p r e s e n t (Cohen e t a l . , 1973; Sarm iento e t a l . * 1 9 7 9 ) .
The v i r i o n e n v e lo p e i s m ost p ro b a b ly r e q u ir e d f o r i n f e c t i v i t y
(S m ith , 1 9 64; S p r in g and Roizm an, 1 968; S t e in e t a l . , 1 9 7 0 ;
R u b en ste in e t a l . , 1 9 7 2 ) . A f te r e n tr y th e v i r io n s t r u c t u r e
i s degraded in th e cy to p la sm and th e DNA tr a n s p o r te d r a p id ly
by an unknown m echanism in t o th e n u c le u s (H ochberg and B eck er ,
1968; Hummeler e t a l . , 1 9 6 9 ) . Knipe e t a l . , (1 9 8 1 ) have
r e c e n t ly d e s c r ib e d a t s m utant w hich com plem ents o th e r t s
m u tan ts p o o r ly , and w hich e x p r e s s e s no p o ly p e p t id e s a t th e
n o n p e r m iss iv e te m p er a tu r e . The l e s i o n may map in a gen e
o o d in g a t l a t e t im e s i n i n f e c t i o n f o r a s t r u c t u r a l p r o t e in ,
and th e a u th o r s p ro p o se t h a t th e d e f e c t i s in u n c o a t in g th e
v i r u s .
I n f e c t io n o f c e l l s i n t i s s u e c u l t u r e w it h HSV r e s u l t s
i n c e s s a t i o n o f m it o s i s (S to k e r and N ew ton, 1 9 5 9 ; W ildy e t
a l . , 1 9 6 1 ) , and i n i t i a t i o n o f th e i n f e c t i o u s c y c l e i s
in d e p en d e n t o f th e c e l l u l a r phase o f DNA s y n t h e s is (Cohen
e t a l . , 1 9 7 1 ) . Soon a f t e r i n f e c t i o n , HSV c a u s e s an i n h i b i t i o n
o f c e l l DNA s y n t h e s is w hich i s d ep en d en t on p r o t e in s y n t h e s i s
(Roizman and R oane, 1 9 6 4 ) . The f a c t t h a t some HSV-2 t s
m u tan ts e i t h e r f a i l to i n h i b i t , o r s t im u la t e , h o s t c e l l DNA
s y n t h e s is a t th e n o n p e r m iss iv e tem p era tu re s u g g e s t s th a t
v ir u s f u n c t io n s are in v o lv e d in i n h i b i t i o n o f h o s t DNA
s y n t h e s i s (H a llib u r to n and Tim bury, 1973; Y am anishi e t a l . ,
1 9 7 9 ) . I t i s n o t known w h eth er th e i n h i b i t i o n i s a r e s u l t o f
i n h i b i t i o n o f c e l l RNA and p r o t e in s y n t h e s is (Roizman e t a l . ,
1 9 6 3 ) , r a th e r than a d i r e c t e f f e c t on DNA s y n t h e s i s .

- 16 -
I n h ib i t io n o f p r o te in s y n t h e s is in v o lv e s d is a g g r e g a t io n
o f h o s t p o ly r iu o so m e s and t h e i r r e a sse m b ly ( S y d is k is and
Roizman, 1 9 6 6 ) . The t o t a l p r o c e in s y n t h e s i s in g c a p a c it y
d e c r e a s e s th ro u g h o u t i n f e c t i o n , a f a c t w h ich S i l v e r s t e i n and
S n g e lh a rd t (1 9 7 9 ) in t e r p r e t e d a s th e a ccu m u la tio n o f non
f u n c t io n a l p o ly r ib o so m e s , Fenw ick and W alker (1 9 7 3 ) have
shown t h a t UV i r r a d ia t io n o f HSV-2 d o es n o t a f f e c t i n h i b i t i o n
o f h o s t c e l l m acrom olecu lar s y n t h e s i s , and co n c lu d ed t h a t
i n h i b i t i o n i s due to a s t r u c t u r a l com ponent o f th e v i r u s .
The same a u th o r s (1 9 7 9 ) have n o ted th e s p e c i f i c p h o sp h o r y la t io n
o f a h o s t r ib o so m u l p r o te in upon i n f e c t i o n , but co n c lu d ed
t h a t t h i s i s n o t th e ca u se o f i n h i b i t i o n o f h o s t p r o t e in
s y n t h e s i s . The HSV-2 fu n c t io n r e s p o n s ib le f o r i n h i b i t i o n h as
been mapped a t 0 .5 6 - 0 .5 7 f r a c t i o n a l genome u n i t s (Fenw ick
e t a l . , 1 9 79; H. 3 . M arsden, p e r s o n a l co m m u n ica tio n ).
IISV-1 i n f e c t i o n le a d s to th e e x p r e s s io n o f v i r u s -
s p e c i f i e d RNA, and c e l l RNA s y n t h e s i s i s r a p id ly su p p resse d
(Hay e t a l . , 1966; F la n a g a n , 1 9 6 7 ) . The p a r t i c u l a r l y ra p id
su p p r e s s io n o f rRNA s y n t h e s i s may be a t t r ib u t e d to two
in h ib i t o r y s t e p s (Wagner and Roizm an, 1 9 6 9 b ). F i r s t l y ,
th e r e i s a d e c r e a s e in th e r a t e o f s y n t h e s i s o f 45 S p r e c u r s o r
RNA, p erh a p s m ed iated by a v ir u s - in d u c e d i n h i b i t o r o f RNA
p o ly m era se I (S a sa k i e t a l . , 1 9 7 4 ) . S e c o n d ly , p r o c e s s in g o f
p r e c u r so r rRNA in t o m ature 18 S and 28 S rRNA i s i n h i b i t e d .
Talley-J3row n and G i l l e t t e (1 9 7 9 ) have n o te d ch a n g es in th e
p a t t e r n s o f sm a ll c y to p la s m ic RNA s y n t h e s i s e a r ly in i n f e c t i o n
l eroderm a pigm entosum c e l l s , ' in c lu d in g th e ap p earan ce o f
a n o v e l 380 n u c le o t id e s p e c i e s .
The a c t i v i t y o f KiJA p o ly m era se I I d e c l in e s on i n f e c t i o n ,
a f e a t u r e w hich P r e s to n and IJev/ton (1 9 7 6 ) a t t r ib u t e d n o t o n ly
to i n h i b i t i o n o f c e l l p r o te in s y n t h e s is but a l s o t o - a v i r u s -
in d u ced p o ly p e p t id e w hich »ay i n t e r a c t e i t h e r w ith th e

- 17 -
enzyme o r w ith th e ch rom atin com p lex . U s in g a c e l l l i n e
vh ich e x p r e s s e s a h ig h l e v e l o f g lo b in mRNA, N is h io k a and
S i l v e r s t e i n (1977 and 1 9 78) showed th a t HSV-l i n f e c t i o n
ca u ses d e g r a d a t io n o f p r e - e x i s t i n g c e l l mRNA. U n lik e
d i s s o c i a t i o n o f p o ly r ib o so m e s , t h i s i s d ep en d en t upon genome
e x x jress lo n . N e v e r t h e le s s , a low l e v e l o f c e l l mRNA i s
s y n th e s is e d even l a t e in i n f e c t i o n ( S t r in g e r e t a l . , 1 9 7 7 ) .
E n g lis and Newton (1 9 8 1 ) rexported t h a t HSV-l mRNA i s
t r a n s la t e d i n v i t r o w ith a lo w e r e f f i c i e n c y th an c e l l mRNA,
and s u g g e s te d t h a t , i f t h i s a p p l ie s in v i v o , i n h i b i t i o n o f
.lo s t c e l l RNA s y n t h e s i s would be in s tr u m e n ta l in e f f i c i e n t
v ir u s g ro w th .
C e l l chrom osom al b rea k a g es w hich o c c u r e a r ly in HSV
i n f e c t i o n a re th o u g h t to be due to a v ir u s - in d u c e d enzym e,
s in c e v ir u s DNA i s n o t i ) h y s ic a l ly a s s o c ia t e d w ith th e
b reakages (Waublce e t a l . , 1 9 6 8 ) . A lth ou gh chrom osom al
breakage seem s n o t to be cau sed by in t e g r a t io n o f v ir u s DNA
in to c e l l DNA, i t has been r e p o r te d th a t a b o u t 200 HSV genome
e q u iv a le n ts p er c e l l a re c o v a le n t ly a s s o c ia t e d w ith n o s t DNA
l a t e in i n f e c t i o n ( B ie g e le i s e n and Rush, 1976; B ie g e l e i s e n
3t a l . , 1 9 77; Y anagi e t a l . , 1 9 7 9 ) . The s ig n i f i c a n c e o f
th e se o b s e r v a t io n s in t h e . i n f e c t i o u s c y c le rem ain s to be s e e n .
The pathway by w hich v ir u s i s a ssem b led i n th e n u c le u s
i s p o o r ly u n d e r s to o d . E le c tr o n m ic r o sc o p ic s t u d ie s o f
n o rm a lly in f e c t e d c e l l s show c o n d e n sa t io n and m a rg in a tio n .o f
h o s t ch rom atin fo llo w e d by th e ap p earan ce o f c a p s id s o f n o ,
low o r h ig h e l e c t r o n - d e n s i t y , a s w e l l a s more b iz a r r e g r a n u la r ,
c r y s t a l l i n e , f i la m e n to u s and tu b u la r s t r u c t u r e s (Morgan e t a l . ,
1959; N i i e t a l . , 196 8 a ; Miyamoto and Morgan, 1971; Sm ith and de
H arven, 1 9 7 3 ) . o th e r p o t e n t i a l l y u s e f u l a t te m p ts to u n r a v e l
what i s u n d o u b ted ly a com plex p r o c e s s have in c lu d e d th e u se o f

~ 18 ~
m e ta b o lic i n h i b i t o r s (N ii e t a l . , 1 1 68b;Friedm ann e t a l . , 1975;
M arcian o-C ab ra l e t a l . , 19 / 7 ) , and s t u d i e s o f HSV-2 t s m utants
(C ab ra l and S c h a f fe r , 1976; A tk in son e t a l . , 1 9 7 0 ) . U n lik e
H SV -l, HSV-2 p ro d u ces tu b u la r s t r u c t u r e s in th e n u c le i o f
in f e c t e d c e l l s (Murphy e t a l • , 1 9 6 7 ) . T h is l a t e e v e n t seem s
n o t to r e q u ir e e n t ir e genome e x p r e s s io n , and k i n e t i c s t u d ie s
u s in g i n h i b i t o r s s u g g e s t t h a t su ch s t r u c t u r e s are in te r m e d ia te s
in v ir u s a ssem b ly r a th e r th an r e s u l t s o f a b er ra n t c a p s id
fo r m a tio n (Iw asak a e t a l . , 1979; Oda e t a l . , 1 9 7 9 ) .
H exagon al and p e n ta g o n a l in t r a n u c le a r c r y s t a l s o f c a p s id s have
f r e q u e n t ly been o b se rv ed (Morgan e t a l • , 1 9 5 8 ; M eln ick e t a l . ,
I 9 6 0 ) . I t i s n o t known how DNA i s e n c a p s id a te d in t o v i r i o n s ,
b u t P i g n a t t i and C a ssa i (1 9 8 0 ) i s o l a t e d n u c le o p r o te in co m p lex es
w hich in c lu d e d n o n -n u c leo so m a l DNA w ith c a p s id - l i k e s t r u c t u r e s
a t one te r m in u s . I t i s n o t c l e a r , h ow ever, w h eth er th e s e
r e p r e s e n t tr u e in te r m e d ia te s in th e e n c a p s id a t io n p r o c e s s ,
o r breakdown p r o d u c ts .
lidV i n f e c t i o n i s accom panied by e x t e n s iv e r e d u p l ic a t io n
o f th e n u c le a r membrane, a t w hich c a p s id s a c q u ir e e n v e lo p e s
a s th e y p a s s in t o th e p e r in u c le a r c i s t e m a (F a lk e e t a l • , 1959;
E p s te in , 1 9 6 2 b ;D a r lin g to n and M oss, 1968; N i i , 1 9 7 1 ) .
A tk in so n e t a l . (1 9 7 8 ) r e p o r te d t h a t en velop m en t may o ccu r
by cle novo s y n t h e s is o f membrane in th e n u c le u s . Envelopm ent
a t c y to p la s m ic membranes may o ccu r f o l lo w in g d i s i n t e g r a t i o n
o f th e n u c le u s . M ature v ir u s p a r t i c l e s l e a v e th e c e l l by
r e v e r s e p h a g o c y to s is (D a r lin g to n and M oss, 1 9 6 8 ) , o r v i a
c y to p la s m ic d u c ts (S ch w artz and Roizm an, 1 9 6 9 ) , o r by b oth
m echanism s ( N i i , 1 9 7 1 ) . O b se r v a tio n s o f v ir u s e g r e s s a re
d ep en d en t on c e l l ty p e (S c h le h o fe r e t a l . , 1 9 7 9 ) .
P r o d u c t iv e i n f e c t i o n k i l l s th e h o s t c e l l , and c e l l u l a r
com ponents le a k in t o th e medium (Y/agner and Roizm an, 1969b;
McCormick, 1 9 7 8 ) . V iru s sp rea d s from c e l l to c e l l v i a th e

- 19 -
iiedium, and a l s o by d i r e c t i n t e r c e l l u l a r .p a ssa g e (S to k e r , 1 9 5 8 ) .
V iId -ty p e v ir u s u s u a l ly c a u se s rou n d in g up o f in f e c t e d c e l l s ,
out some m utant s t r a in s ( s.vn) ca u se e x t e n s iv e c e l l f u s io n
and th e fo r m a tio n o f s y n c y t ia (Gray e t a l . , 1958; Iloggan and
Roizman, 1 9 5 9 ) . The r e c r u itm e n t o f c e l l s in t o s y n c y t ia seem s
la r g e ly d ep en d en t upon th e r e c r u i t in g r a th e r than th e r e c r u i t e d
c e l l (Lee and S p ea r , 1980; B z ik and P e r so n , 1 9 8 1 ) . C e l l
fu s io n i s d ep en d en t on v ir u s e x p r e s s io n , s in c e s e v e r a l
in h ib i t o r s o r UV i r r a d ia t io n a b o l i s h s y n c y t ia fo r m a tio n
(O’Donovan and Roizm an, 1961; K ou sou las e t a l . , 1 9 7 8 ) .
I n h ib it io n - by 2 -d e o x y -D -g lu c o se o r tu n ica m y c in s t r o n g ly
in d ic a t e s t h a t v ir u s g ly c o p r o t e in s a re in v o lv e d in c e l l f u s io n
(G a lla h e r e t a l . , 1973; K e l le r , 1 9 76; K now les and P e r so n , 1976;
P iz e r e t a l . , 1 9 8 0 ) . In d eed , u s in g reco m b in a n ts betw een a
v ir u s d e f e c t i v e i n g ly c o p r o t e in C (gC) and one w ith a t s l e s i o n
in g ly c o p r o t e in B (g B ), M a n serv ig i e t a l . , (1 9 7 7 ) co n c lu d ed
th a t gB i s a f u s io n prom oter and gC a f u s io n s u p p r e s s o r .
However, m u lt ip le v i r a l and c e l l u l a r f a c t o r s a r e in v o lv e d in
c e l l f u s io n (Yamamoto e t a l . , 1975; Yamamoto and K abuta, 1977;
Huyechan e t a l . , 1979; Lee and S p ea r , 1 9 8 0 ) .
DBA s y n t h e s is
kuch o f th e more c o n c lu s iv e work on h e r p e s v ir u s DNA
s y n t h e s is has been done w ith PRV, so th e f o l lo w in g d i s c u s s io n
in c lu d e s b oth HSV and PRV.
R ice e t a l . (1 9 7 6 ) r e p o r te d t h a t , soon a f t e r i n f e c t i o n ,
in p u t DNA becom es s p e c i f i c a l l y a s s o c ia t e d w ith a c e l l n u c le a r
p r o t e in . In c o n t r a s t , th ey d id n o t f in d nev/1 y s y n t h e s is e d DNA
in such a com p lex , and Du A s y t h e s i s d o es n o t in v o lv e th e
norm al c e l l n u c leo so m a l a p p a ra tu s ( k o u t t e t e t a l • , 1 9 79;
L ein bach and Summers, 1 9 8 0 ) . Jacob and Roizman (1 9 7 7 )
r ep o r te d th a t onl^ a sm a ll t r a c t io n o f p a r e n ta l DNA e n t e r s

- 20 -
th e r e p l i c a t i v e p o o l , w hereas th e m a jo r ity o f p a r e n ta l and
progeny PRV DNA can r e p l i c a t e (Jean and B en -P o r a t, 1976;
B en -P o ra t e t a l . , 1 9 7 6 a ) • D e n s i t y - s h i f t ex p er im e n ts have
shown th a t HSV and PRV DNA a re r e p l i c a t e d s e m ic o n s e r v a t iv e ly
(K aplan and B en -P o r a t, 1964; K o lb er , 1975; Khan e t a l . , 1 9 7 8 ) .
M ost s i n g l e s tra n d b reak ages in p a r e n ta l DNA a re r e p a ir e d p r io r
to DNA r e p l i c a t io n (B en -P o ra t e t a l . , 1 9 7 & d ),b u t n a s c e n t DNA
c o n t a in s s in g l e s tra n d ed r e g io n s and c o v a le n t ly l in k e d RNA
(B isw a l e t a l . , 1974; Murray and B is w a l, 1 9 7 4 ; H ir sc h e t a l . ,
1976; Sh lom ai e t a l . , 1976; M u ller e t a l . , 1 9 7 9 a ) . P u ls e -c h a s e
e x p e r im e n ts in d ic a t e th a t new ly s y n t h e s is e d DNA se d im e n ts
a s la r g e h e te r o g e n e o u s m o le c u le s , and i s r a p id ly p r o c e s s e d
to a form sed irtien tin g w ith th e p r o p e r t ie s o f u n i t - l e n g t h
m o le c u le s ( h ir s c h e t a l . , 1976; Sh lom ai e t a l . , 1 9 7 6 ; Jacob
and Roizm an, 1 9 7 7 ; B en -P o ra t and T ok azew sk i, 1 9 7 7 ) . A s in g le
round o f PRV DNA r e p l i c a t io n ta k e s 25 o r 45 m in, d ep en d in g
on w hether r e p l i c a t io n i s u n i - or b i - d i r e c t i o n a l (B en -P o ra t-
e t a l . , 1 9 7 7 ) . Most sy ste m s f o r in v i t r o DNA s y n t h e s i s a l lo w
c o m p le t io n o f n a sc e n t DNA but n o t r e i n i t i a t i o n ( B e l l , 1 974;
B is w a l and Murray, 1974; Shlom ai and B eck er , 1975; B o ld en e t
a l . , 1975; K o lb er , 1975; B eck er and A sh er, 1 9 7 5 ; Sh lom ai e t
a l . , 1977; J o n g en ee l and B achenheim er, 1 9 8 0 ) . P r o m is in g
a tte m p ts have been made to e x t r a c t th e p h y s io lo g ic a l
r e p l i c a t i o n com plex from in f e c t e d c e l l n u c le i ( P ig n a t t i e t
a l . , 1979 ; K nopf, 1 9 7 9 ) .
T e m p e r a tu r e -s e n s it iv e m u tan ts in ab o u t h a l f o f th e
i d e n t i f i e d com p lem en ta tion grou p s o f HSV and PRV a re d e f i c i e n t
i n DNA s y n t h e s is to a g r e a te r or l e s s e r e x t e n t , and t h e r e fo r e
i t i s l i k e l y th a t .many gene p ro d u cts a re in v o lv e d i n t h i s
p r o c e s s , in c lu d in g th e v ir u s -c o d e d DNA p o ly m era se (H a ll ib u r to n
and Tim bury, 1973 and 1 9 7 6 ; P r in g le e t a l . , 1973; S c h a f fe r
e t a l . . 1973; iSsparza e t a l . , 1974; D en yesh -M eln ick e t g l . » 1 9 7 4 ;

P u r ifo y and B e n y e s n -^ e ln ic k , 1975; Aron e t a l . » 1975; C ourtney
e t a l . , 1 9 76; Bone e t a l . , 1 9 7 8 ) .
e l e c t r o n m ic r o sc o p ic ex a m in a tio n o f r e p l i c a t in g DNA
h a s r e v e a le d l i n e a r m o le c u le s s m a lle r th a n , e q u a l in s i z e t o ,
o r la r g e r than v ir io n DNA, c i r c l e s o f u n i t s i z e o r s m a lle r ,
and la r g e ta n g le d m o le c u le s . Such m o le c u le s may c o n ta in
s i n g l e s tr a n d ed r e g io n s , f o r k s , e y e s , lo o p s o r c o l la p s e d
r e g io n s (S h lom ai e t a l . , 1976; Jean and B e n -P o r a t, 1976;
B en -P o r a t e t a l . , 1 976b ;H ixon , 1977; Jean e t a l . , 1977;
H ir sc h e t a l . , 1977; Priedraann e t a l . , 1 9 77; Jacob and Roizm an,
1 9 7 7 ) . The f o l lo w in g r e p l i c a t io n m odel h a s been d e r iv e d from
th e s e and o th e r d a ta . O b se r v a tio n s th a t HSV-l DNA i s
t e r m in a l ly red u n d a n t, th a t te r m in a l r e s t r i c t i o n fra g m en ts
o f IiSV-1 DNA a re red u ced in abundance in in f e c t e d c e l l n u c le i
(J a co b e t a l • , 1 9 7 9 ) , th a t a p r o p o r t io n o f in p u t PRV DNA
a c q u ir e s s in g l e s tra n d ed ends (Jean and B e n -P o r a t, 1976) and
a p r o p o r t io n becom es c i r c u l a r , and t h a t PRV DNA i n th e f i r s t
round o f r e p l i c a t io n i s c i r c u la r (B en -P o ra t and V each, 1 9 8 0 ) ,
a re in su p p o rt o f th e h y p o th e s is t h a t l i n e a r in p u t DNA
c i r c u l a r i s e s a l t e r i n f e c t i o n , p ro b a b ly in v o lv in g th e a c t io n
o f an e x o n u c le a s e . H e a d - t o - t a i l co n ca tem ers a re th en form ed
by an unknown m echanism , but w hich may be a r o l l i n g c i r c l e
(Jean and B en -P o r a t, 1976; Jean e t a l • , 1 9 77; B en -P o ra t and
T ok azew sk i, 1977; B en -P o ra t and R ixon , 1 979; Jacob e t a l . , 1 9 7 9 ) .
The q u e s t io n o f how u n i t - l e n g t h DNA m o le c u le s r e s u l t from
c o n ca to m c rs , w ith fo u r eq u im olar arran gem en ts o f 1 and S,
i s c o n s id e r e d in th e D is c u s s io n . M odels f o r DNA s y n t h e s i s
in v o lv in g in d ep en d en t r e p l i c a t io n o f L and S ( Skare and Summers,
1 9 7 7 ) o r r e p l i c a t i o n o f l i n e a r m o le c u le s from a te r m in a l
h a ir p in lo o p (R ixon , 197'/; Jacob and Roizm an, 1 977) f a i l e d to
f in d su b seq u en t su p p o r t . B en -P orat and Kaplan (1 9 6 3 ) showed

- 22 -
th a t a lth o u g h a m in o r ity o f m ature PRV DNA i s e neap s i d a te d ,
i t i s s e l e c t e d a t random from th e p o o l . S im i la r ly a m in o r ity
o f HSV-l DNA i s e n c a p s id a te d ( R u s s e l l e t a l . , 1 9 6 4 ) , and
P r en k e l and Roizman (1 9 7 2 a ) co n c lu d ed t h a t m o le c u le s w ith
fe w er n ic k s a re p r e f e r e n t i a l l y p ack aged , a lth o u g h i t i s e q u a lly
l i k e l y t h a t n ic k s a r e r e p a ir e d d u r in g p a c k a g in g .
Two ap p ro a ch es have in d ic a te d t h a t HSV-l and PRV DNA
c o n ta in an o r ig in o f r e p l i c a t io n in th e r e g io n o f th e S
term in u s , and p erh ap s a n o th e r i n U- . Sh lom ai e t a l . (1 9 7 6 )
and Priodmann e t aJ_. ( 1 9 7 7 ) d e sc r ib e d two o r ig in s o f
r e p l i c a t io n from e le c t r o n m ic r o sc o p ic s t u d ie s o f r e p l i c a t in g
iiSV-1 DMA, one c o r r e sp o n d in g to TR and IR0 , and th e o th e r to
a p o s i t i o n 40# from one te r m in u s . H ir sc h e t a l . (1 9 7 7 )
d e te c te d g r e a te r in c o r j jo r a t io n o f i s o t o p e in a p u lse d l a b e l
in r e s t r i c t i o n fra g m en ts c o n ta in in g th e j o i n t r e g io n .
Jean e t a l . (1 9 7 7 ) o b ta in e d r e s u l t s s im i la r to th o s e
d e s c r ib e d above from t h e i r e le c t r o n m ic r o sc o p ic a n a ly s i s
o f r e p l i c a t in g PRV HR A, a lth o u g h 1 0 # o f m o le c u le s w ith4
r e p l i c a t io n e y e s a p p a r e n t ly e x h ib it e d m u lt ip le r e p l i c a t io n
s i t e s . H en-P orat and Veuch (1 9 8 0 ) co n c lu d ed from b lo t
h y b r id is a t io n o f PRV nil A th o u g h t to have been i s o t o p i c a l l y
l a b e l l e d d u r in g th e f i r s t round o f r e p l i c a t i o n t h a t th e main
o r ig in o f r e p l i c a t io n i s a t o r n ea r th e S te r m in u s , from
w hich DliA s y n t h e s is p ro ce ed s u n i d i r e c t i o n a l l y . They su g g e s te d
th a t th e o th e r o r ig in 40# from one term in u s i s u sed r e l a t i v e l y
in f r e q u e n t ly d u r in g th e f i r s t round o f r e p l i c a t i o n .
As a secon u ap p roach , s t r u c t u r a l a n a ly s i s o f d e f e c t iv e
v ir u s genomes h as p ro v id ed some e v id e n c e f o r th e l o c a t io n s
o f o r ig in s o f J)AA r e p l i c a t i o n . High m u l t i p l i c i t y p a ssa g e
o f HSV or PRV r e s u l t s in a c y c l i c p a t te r n o f p a r t i c l e to '
i x . f e c t i v i t y r a t i o end o f te n tn e ap p earan ce o f DNA o f n o v e l

B en -P o ra t e t a l . , 1974; hu rray e t a l * , 1 9 75; Stegmari e t a l . ,
1 9 7 8 ) . DNA from th e d e f e c t i v e i n t e r f e r i n g p a r t i c l e s i s
a p p ro x im a te ly th e same s i z e a s , hut h as a lo w e r k i n e t i c
c o m p le x ity th a n , sta n d a rd DNA (V/agner e t a l . , 1974; Bookout
e t a l . , 1 9 7 9 ) . P a r t i a l d e n a tu r a t io n m apping has shown th a t
d e f e c t i v e DNA c o n s i s t s o f m u lt ip le h e a d - t o - t a i l r e i t e r a t i o n s
o f a b a s ic u n i t ( liu b e n s te in and K aplan, 1 9 7 5 ; P ren k e l e t a l . t
1 9 7 5 ) .
Two ty p e s o f rej^eat u n i t have been d e s c r ib e d from
r e s t r i c t i o n e n d o n u c le a se a n a ly s i s o f HSV-l d e f e c t i v e DNA
(K aerner e t a l • , 1979; P r en k e l e t a l . , 1 9 8 0 ) . That o f c l a s s I
c o n t a in s axjp rox im ately 8 .5 kbp o f DNA c o m p r is in g TR and an
a d ja c e n t p o r t io n o f . (P re n k e l e t . a l . , 1 9 7 5 , 1976 and 1980;
lo c k e r and P r e n x e l, 1978 and 1979a;Graham e t a l . , 1978;
V lazny and P r e n k e l, 1 9 8 1 ) . Y/hereas one end o f th e d e f e c t iv e
genome i s u n iq u e and p ro b a b ly p o s s e s s e s an a se q u e n c e , th e
o th e r term in u s i s th o u g h t to be random in n a tu r e . Such a
s t r u c t u r e s u g g e s t s t h a t m a tu ra tio n and p a ck a g in g o f DNA
o ccu r by a s p e c i f i c r e c o g n it io n o f th e S term in u s and an
ap p rox im ate le n g th m easurem ent• T h is i s u n l ik e ly to be th e
c a s e f o r m a tu r a tio n o f sta n d a rd IISV-1 DNA b eca u se both
te r m in i a re u n iq u e . I t i s n o t y e t c l e a r w h eth er th e a
seq u en ce i s p r e s e n t in ea ch r e p e a t u n i t o f c l a s s I d e f e c t i v e
genom es. S iz e h e t e r o g e n e i ty o f th e r e p e a t u n i t in d i f f e r e n t
d e f e c t i v e genom es o f t n i s c l a s s i s due to v a r ia t io n in th e
le n g tn o f se q u e n c e s in c lu d e d t h e r e in . The k i n e t i c s o f
d e f e c t i v e DNA s y n t a c s i s , i t s s e n s i t i v i t y to th e v ir u s DNA■> •»
p o lym erase i n h i b i t o r p h o s if io n o a c e t ic a c id (PAA), and th e
f a c t t h a t c o i n i c c l i o n v /itn se p a r a te d r e jje a t u n i t s generates

- R4 -
f u l l le n g th d e f e c t i v e genom es, argue t h a t d e f e c t i v e DNA
c o n ta in s th e IISV-1 r e p l i c a t io n o r ig in w hich i s f u n c t io n a l
in th e r e p l i c a t i o n o f d e f e c t i v e DNA (P r e n k e l e t a l . , 1980;
V lazny and P r e n k e l, 1 9 8 1 ) . The o b s e r v a t io n t h a t s in g l e
d e f e c t i v e DNA m o le c u le s c o n ta in o n ly one 3 i s e o f r e p e a t u n it
makes i t t e n a b le th a t th ey a re s y n t h e s is e d by a r o l l i n g c i r c l e
m echanism from a c i r c u l a r r e p e a t u n i t . In su p p o r t o f t h i s
h y p o th e s is , B ecker e t a l . (1 9 7 8 ) o b se rv ed c i r c u l a r and
c i r c u l a r - l i n e a r m o le c u le s o f v a r io u s s i z e s by e le c t r o n
m icro sco p y o f r e j iL ic a t in g d e f e c t i v e DNA.
The r e p e a t u n i t o f c l a s s I I d e f e c t i v e DNA c o n s i s t s o f
th e S te r m in a l r e g io n l in k e d to a r e g io n o f m apping a t
0 .3 4 - 0 .4 0 f r a c t i o n a l genome u n i t s . The g r e a t e r s e n s i t i v i t y
o f t h i s c l a s s than c l a s s I to PAA su p p o r ts th e p r e se n c e o f a
s e p a r a te r e p l i c a t io n o r ig in in U^. In d ee d , i t h as been n o ted
th a t a t te m p ts to c lo n e in t o p la sm id s fra g m en ts from t h i s
r e g io n o f have c o n s i s t e n t l y le d to th e d e l e t io n o f ab ou t
100 bp a t a s p e c i f i c l o c a t io n , p erh ap s i n d ic a t in g th e
p r e se n c e o f an in v e r te d r e p e a t in v ir u s DNA w hich p r o v e s
u n s ta b le in p la sm id s (P o s t e t a l . , 1 9 80; B. M atz, p e r s o n a l
co m m u n ica tio n ).
RNA s y n t h e s is
iiSV-1 RNA s y n t h e s is i s a s s e n s i t i v e a s c e l l RNA s y n t h e s i s
to i n h i b i t i o n by *>c-amani t i n , an i n h i b i t o r o f RNA p o ly m era se I I
(A lv/ine e t a l . , 1974; B en-Z eev e t a l . , 1 9 7 6 ) , and a l l s t a g e s
o f v i r a l t r a n s c r ip t io n a re e q u a lly s e n s i t i v e (C o sta n zo e t a l • ,
1 9 7 7 ) . m oreover, u3V~l RNA s y n t h e s is i s r e s i s t a n t to th e
urug in c e l l s p o s s e s s in g an cx’-a m a n it in r e s i s t a n t RNA
p o lym erase I I (C ostan zo e t a l . , 1 9 7 7 ; B en-Z eev and B eck er ,
1 9 7 7 ) . Lowe (1 8 7 8 ) was u n ab le to d e t e c t RNA p o ly m era se
a c t i v i t i e s in th e in x ecteu . c e l l o th e r than th o s e n o rm a lly

- 25 -
p r e s e n t in the u n in fe c te d c e l l . These o b s e r v a t io n s s t r o n g ly
in d ic a t e t h a t c e l l RNA p o lym erase I I i s r e s p o n s ib le f o r
IiSV-1 RNA s y n t h e s i s a t a l l s ta g e s o f i n f e c t i o n , a lth o u g h
m o d if ic a t io n o f th e t r a n s c r ip t io n a l s p e c i f i c i t y o f t h i s
enzyme d u r in g i n f e c t i o n i s n o t r u le d o u t .
HSV-l mRNAs a re p o ly a d e n y la te d a f t e r t r a n s c r ip t io n
(B aclienheim er and Roizm an, 1 9 7 2 ) . I t h a s been r e p o r te d th a t
a s i g n i f i c a n t p r o p o r t io n o f mRNA i s n o t p o ly a d e n y la te d
( S i l v e r s t e i n e t a l . , 1 9 7 3 ; S tr in g e r et; a l . , 1 9 7 7 ) , but
S i l v c r s t e i n e t a l . (1 9 7 6 ) showed t h a t m ost o r a l l v ir u s
mRNAs a re p o ly a d e n y la te d , i n g e n e r a l p o s s e s s in g 3 0 , 50 o r
155 a d e n y lic a c id r e s id u e s a t th e 3 1 te r m in u s . The
c o m p a r tm e n ta lisa t io n o f s h o r te r p o ly a d e n y lic a c id t r a c t s in
th e n u c le u s , and th e lo n g e r in th e c y to p la sm , was in t e r p r e t e d
by th e l a t t e r a u th o r s a s two s e p a r a te p r o c e s s in g s t e p s in
p o ly a d e n y la t io n . HSV-l mRNAs a l s o p o s s e s s a 7 -m e th y lg u a n o s in e
5 ' - t r ip h o s p h a t e cap and i n t e r n a l l y m eth y la ted n u c le o t id e s
(B a r tk o sk i and Roizm an, 1976; Moss e t a l . , 1977)* Jacquem ont
and H uppert (1 9 7 7 ) showed t h a t HSV-l r e p l i c a t i o n i s r e v e r s ib ly
b lo ck ed by 5 f - S - i s o b u t y la d e n o s in e , an i n h i b i t o r o f m e th y la t io n ,
e s p e c i a l l y o f th e c a p . Assum ing t h a t t h i s i s th e s i g n i f i c a n t
in h ib i t o r y a c t io n o f t h e ‘d ru g , t h i s r e s u l t s u g g e s t s t h a t
m e th y la t io n i s e s s e n t i a l f o r t r a n s la t io n o f mRNA. C e s s a t io n
o f m e th y la t io n o f in t e r n a l n u c le o t id e s a t l a t e t im e s may be
due to e x p r e s s io n o f an e a r ly gene (B a r tk o sk i and Roizm an, 1 9 7 8 ) .
HSV t r a n s c r ip t io n i s a com plex p r o c e s s under t ig h t
tem p ora l c o n t r o l . H arly work u s in g l iq u id h y b r id is a t io n showed
t h a t m ost o f th e H3V-1 genome i s e x p r e sse d a t e a r ly (b e fo r e
th e o n s e t o f DNA s y n t h e s i s ) and l a t e t im e s (P r e n k e l and
Roizm an, 1972b;W agner, 1972; P ren k e l e t a l . , 1973; Swanstrom
anu Wagner, 1974; S tr in g e r e t a l . , 1 9 7 7 ) . B a r ly mRNA

c o m p r ise s a s u b s e t o f l a t e mRNA, and ea ch group c o n s i s t s o f
s c a r c e and abundant t r a n s c r ip t s , Wagner e t a l . (1 9 7 2 ) p rop osed
a v e r y e a r ly c l a s s w hich form s a s u b s e t o f e a r ly s p e c i e s .
The e v id e n c e f o r th r e e m ajor c l a s s e s o f t r a n s c r ip t s was
su p p o rted by ex p e r im e n ts in v o lv in g th e u se o f m e ta b o lic
i n h i b i t o r s . I n i t i a l l y i t was r e p o r te d t h a t mRNA made i n th e
a b se n c e o f de novo p r o t e in s y n t h e s i s was s im i la r to t h a t made
a t e a r ly t im e s (P r e n k e l e t a l . , 1973; Swanstrom e t a l . , 1 9 7 5 ) ,
but i t soon became c l e a r th a t im m ediate e a r ly (IE ) mRNA
produced in th e p r e se n c e o f c y c lo h e x im id e form s a more
r e s t r i c t e d c l a s s (Kozak and Roizm an, 1 9 7 4 ; Roizman e t a l . , 1 9 7 4 ) .
I n h ib i t io n o f DNA s y n t h e s is c a u s e s th e p r o d u c t io n o f mRNA
v e r y s im i la r to t h a t made a t e a r ly t im e s (Wagner e t a l . , 1972;
Murray e t a l . , 1974; Swanstrom and Wagner, 1974; Swanstrom
e t a l . , 1 9 7 5 ) .
H y b r id is a t io n o f IE mRNA to n i t r o c e l l u l o s e s t r i p s
c o n t a in in g r e s t r i c t i o n fra g m en ts o f HSV-l DNA a llo w e d IE g e n e s
to be mapped on th e genome (C lem en ts e t a l . , 1977; Jon es
e t a l . , 1 9 7 7 ) . The d i r e c t i o n s o f t r a n s c r ip t io n o f t h e s e
n o n -c o n t ig u o u s g en es were e s t a b l i s h e d by h y b r id is a t io n u s in g
s h o r t cDNA s y n t h e s is e d u s in g p o ly a d e n y la te d mRNA (C lem en ts
e t a l . , 1979; Anderson e t ' a l . , 1 9 8 0 a ;J . B. C lem en ts , p e r s o n a l
co m m u n ica tio n ). I s o l a t io n o f mRNAs from m ethylraercui*ic
h y d ro x id e a g a r o se g e l s fo llo w e d by t h e ir t r a n s l a t io n i n v i t r o
h as a llo w e d th e c o r r e la t io n o f mRNA s i z e , genome l o c a t io n ,
and p o ly p e p t id e s i z e (W atson e t a l . , 1979; T a lley -B ro w n and
M i l l e t t e , 1979; Anderson e t a l . , 1 9 8 0 a ) . C lem en ts e t a l . (1 9 7 9 )
p rop osed a m odel f o r t r a n s c r ip t io n o f IE g e n e s u t i l i s i n g a
s in g l e prom oter in THcj/IRg, but- su b seq u en t work h as shown
th a t each IE gene has i t s own prom oter (E a sto n and C lem en ts ,
1980; h i l l e t t e and iCLaiber, 1580; Mackem and Roizm an, 1980;

- 27 -
H. J . R ixon , p e r s o n a l co m m u n ica tio n ). D ata c o n c e r n in g th e
HSV-l ID g e n es are summarised in P ig u re A 1 .4 . A lth ough
P r en k e l e t a l . (1 973) reported , th a t th e t r a n s c r ip t io n a l
program o f HSV-2 i s s i g n i f i c a n t l y d i f f e r e n t from t h a t o f
H SV -l, H aston and C lem en ts (1 9 8 0 ) have c l e a r l y shown by
b lo t h y b r id is a t io n , mRNA i s o l a t i o n and in v i t r o t r a n s la t io n
t h a t th e two v ir u s e s sh a re v er y s im i la r IE t r a n s c r ip t io n p a t t e r n s .
H arly mRNA form s a more com plex c l a s s mapping th ro u g h o u t
th e HSV-l genome (C lem en ts e t a l . , 1977; S tr in g e r e t a l . , 1 9 7 8 ) .
Some 16 s p e c ie s have been mapped, m ost o f w hich se e m in g ly
have in d iv id u a l p rom oters (H o lla n d e t a l . , 1 979; M i l l e t t e and
K la ib e r , 1 9 8 0 ) . Anderson e t a l . (1 9 7 9 ) lo c a t e d on th e genome
a p p r o x im a te ly 40 l a t e mRNAs r a n g in g i n s i z e from 1 .5 to
g r e a t e r th a n 8 kb . C o sta e t a l . (1 9 8 1 ) have mapped th e
m ajor l a t e mRNA, w hich i s p ro b a b ly n o t s p l i c e d and c o d e s fo r
th e m ajor c a p s id p r o te in o f m .w t. 1 5 5 ,0 0 0 . I t i 3 o b v io u s
t h a t d i s s e c t i n g tn e c o m p le x it ie s o f th e s e t r a n s c r ip t io n a l
p a t t e r n s w i l l occupy i n v e s t i g a t o r s f o r n o t a few y e a r s .
R egard in g th e tem p ora l c o n t r o l o f th e t r a n s c r ip t io n a l
program o f H SV -l, Y/ard and S te v e n s (1 9 7 5 ) showed th a t when
t r a n s c r ip t io n was b lo ck ed by a c tin o m y c in D, e a r l i e r mRNAs
ten d ed to be more s h o r t - l iv e d th an l a t e r o n e s , s u g g e s t in g
a c o n t r o l mechanism o p e r a t in g a t t h i s l e v e l . A seco n d ty p e
o f c o n t r o l , a c t in g a t th e l e v e l o f p r o c e s s in g o f i n i t i a l
t r a n s c r ip t s in th e n u c le u s and t r a n s lo c a t io n o f m ature mRNAs
to th e c y to p la sm , i s l i k e l y to be o p e r a t iv e e s p e c i a l l y in
l a t e r s t a g e s o f i n f e c t i o n . T r a n s c r ip ts i s o l a t e d from n u c le i
a t l a t e t im e s a re la r g e r than c y to p la s m ic mRNAs, and some
n u c le a r s p e c ie s are u n d e r -r e p r e se n te d in th e cy to p la sm
(iVagner ana Roizman, 1969a and b; Wagner, 1972; Kozak and

- 28 -
Roizman, 1974-; C lem ents e t a l . , 1977; J o n es and Roizm an, 1979;
Jacquem ont e t a l . , 1 9 8 0 ) . However, s im i la r c la im s th a t m ost
o r a l l o f th e genome i s t r a n s c r ib e d in th e n u c le u s under IE
c o n d i t io n s , and t h a t m ost t r a n s c r ip t s a ccu m u la te in th e n u c le u s
so th a t o n ly c e r t a in s p e c ie s e n te r th e cy to p la sm (Kozak and
Roizm an, 1974; J o n es and Roizm an, 1 9 79) have been c o u n tered
by o th e r s who showed t h a t n u c le a r and c y to p la s m ic IE RNAs a re
o f s im i la r c o m p le x it ie s (C lem en ts e t a l . , 1 9 77; E aston and
C lem en ts, 1 9 8 0 ; A nderson e t a l . , 1 9 8 0 a ) . l i q u i d h y b r id is a t io n
s t u d ie s showed t h a t a s i g n i f i c a n t p r o p o r t io n o f th e genome i s
sy m m e tr ic a lly t r a n s c r ib e d a t l a t e t im e s (B en -Z eev and Becker",
1975; Jacquem ont and Roizm an, 1975a and b; Kozak and Roizm an,
1975; M iov ic and P iz e r , 1 9 8 0 ) .
The m ost im p o rta n t l e v e l o f c o n t r o l i s th o u g h t to be
t h a t o f t r a n s c r ip t io n a l i n i t i a t i o n . The HSV-l m u tan ts
tsK , tsD and t s T have in d ep en d en t l e s i o n s i n th e gene c o d in g
f o r IE mRNA 3 , w hich i s t r a n s la t e d in t o th e IE p o ly p e p t id e
Vmw IE 175 (IC P4) (P r e s to n e t a l . , 1978; Stow and W ilk ie ,
1978; P r e s to n , in p r e s s ) . A lth ou gh t s K com plem ents tsD and
t s T in some t e s t s (C rom bie, 1 9 7 5 ) , i t i s now o v erw h elm in g ly
l i k e l y t h a t th e th r e e l e s i o n s a r e i n one g e n e . The m u ta tio n
in tsK i s due to a s in g le , b ase p a ir change r e s u l t i n g in th e
rep la cem en t o f an a la n in e by a v a l in e r e s id u e (M -J. M urchie,
p e r s o n a l co m m u n ica tio n ). T hese m u tan ts p rod u ce a t non
p e r m is s iv e tem p era tu re mRNA and p o ly p e p t id e p r o f i l e s more
or l e s s s im i la r to th e IE p a t te r n s (M arsden e t a l . , 1976;
Watson and C lem en ts , 1 9 7 7 , 1978 and 1980; D ixon and S c h a f fe r ,
1 9 8 0 ) . The p r o p e r t ie s o f t h e s e m u tan ts show t h a t a t l e a s t
one IE f u n c t io n ( i . e . Vmw IE 175) i s c o n t in u o u s ly r e q u ir e d
f o r th e t r a n s i t i o n from IE to e a r ly t r a n s c r ip t io n a l p a t t e r n s .
P r e s to n (1 9 7 9 ) showed th a t t h i s fu n c t io n i s d i r e c t l y in v o lv e d
in th e a c t i v a t i o n a t th e t r a n s c r ip t io n a l l e v e l o f an e a r ly

- 29 -
gen e (d eo x y p y r im id in e k in a s e , dPyK). I n h i b i t i o n o f DNA
s y n t h e s is le a d s to an e a r ly t r a n s c r ip t io n a l p a t te r n and nuch
red u ced p r o d u c t io n o f l a t e p o ly p e p t id e s ( lio n e s s and Roizm an,
1974; H oness and W atson, 1 9 7 7 a ;C lem en ts e t a l . , 1977;
Watson and C lem en ts , 1 9 7 7 ; H olland e t a l . , 1 9 8 0 ) . A lth ough
W olf and Roizman (1 9 7 8 ) in t e r p r e t e d t h i s a s a req u irem en t
f o r both p a r e n ta l and p rogeny DNA te m p la te s f o r a norm al
l e v e l o f l a t e p o ly p e p t id e s y n t h e s i s , i t seem s more l i k e l y
th a t v ir u s DNA s y n t h e s is i s r e q u ir e d f o r th e t r a n s i t i o n from
e a r ly to l a t e p a t te r n s o f t r a n s c r ip t io n .
P r o te in s y n t h e s i s and m o d if ic a t io n
A p p rox im ately 50 HSV-induced p o ly p e p t id e s have been
d e t e c t e d i n in f e c t e d c e l l s by S D S -p o ly a cry la m id e g e l
e l e c t r o p h o r e s i s (Honess- and Roizm an, 1 973; M arsden e t a l . ,
1 9 7 6 ) . T w o-d im en sion a l g e l e l e c t r o p h o r e s i s h a s a llo w e d
250 p o ly p e p t id e s to be d i s t in g u i s h e d , a lth o u g h u n d o u b ted ly
many o f t h e s e r e s u l t from p o s t - t r a n s l a t i o n a l m o d if ic a t io n o f
p rim ary t r a n s l a t io n p r o d u c ts (H aarr and M arsden, 1 9 8 1 ) .
V iru s p r o t e in s a r e s y n t h e s is e d in th e cy to p la sm and
m o st, w ith th e e x c e p t io n o f g ly c o s y la t e d p o ly p e p t id e s , a re
th en tr a n sp o r te d to th e n u c le u s ( S y d is k is and Roizm an, 1968;
S p ear and Roizm an, 1968 and 1 9 7 0 ) . I n i t i a l s t u d ie s o f th e
k i n e t i c s o f s y n t h e s is o f v ir u s - in d u c e d p o ly p e p t id e s
d em o n stra ted tem p ora l c o n t r o l o f p r o t e in s y n t h e s i s , and c o n t r o l
o f p r o t e in abundance (H oness and Roizm an, 1 9 7 3 ) . E nzym atic
f u n c t io n s ten d to be produced e a r ly i n i n f e c t i o n , and m ost
s t r u c t u r a l p r o t e in s l a t e r ( R u s s e l l e t a l . , 1 9 6 4 )* The
su b seq u en t u se o f m e ta b o lic i n h i b i t o r s su g g e s te d th e e x i s t e n c e
o f th r e e k i n e t i c c l a s s e s o f p o ly p e p t id e s , a , and y% w hich
co rresp o n d in g e n e r a l to th e IE , e a r ly and l a t e mRNA s p e c ie s
(H on ess and Roizman, 1974 and 1 9 7 5 a ) . oi p o ly p e p t id e s are

- 30 -
s y n t h e s is e d im m ed ia te ly a l t e r r e l e a s e o f a c y c lo h e x im id e b lo ck
a t l e a s t one o f t h i s c l a s s i s r e q u ir e d f o r th e p r o d u c t io n o f
/5 p o ly p e p t id e s , f t f u n c t io n s a re r e q u ir e d f o r th e i n h i b i t i o n
o f <k p o ly p e p t id e s y n t h e s i s and th e s t im u la t io n o f y
p o ly p e p t id e s , w h ich in tu rn i n h i b i t s y n t h e s i s .
The t h r e e f o ld c l a s s i f i c a t i o n o f v ir u s - in d u c e d p o ly p e p t id e s
i s a s i m p l i s t i c m odel, s in c e P e r e ir a e t a l . (1 9 7 7 ) i d e n t i f i e d
two c l a s s e s o f j$ p o ly p e p t id e (^q and y#^), ea ch o f w h ich i s
p ro b a b ly d ep en d en t upon an c* f u n c t io n f o r e x p r e s s io n
(P r e s to n , 1 9 7 9 ) . S tu d ie s o f p o ly p e p t id e s y n t h e s i s in th e
p r e s e n c e o f an i n h i b i t o r o f DNA s y n t h e s i s have le d to th e
i d e n t i f i c a t i o n o f a t l e a s t two y c l a s s e s (H on ess and W atson,
1 9 7 7 a ;P ed erso n e t a l . , 1 9 8 1 ) . The la t t e r " a u t h o r s d e f in e d
s i x c l a s s e s o f p o ly p e p t id e on th e b a s i s o f t h e i r e x p r e s s io n
in th e p r e se n c e o f an i n h i b i t o r o f DNA s y n t h e s i s , and M arsden
e t a l . (1 9 7 6 ) d e s c r ib e d n in e c l a s s e s o f d ep en d en ce r e l a t i o n s h i p
b etw een p o ly p e p t id e s y n t h e s is and th e DNA p h en otyp e o f HSV-l
t s m u ta n ts . The p r o p e r t ie s o f a dom inant l e t h a l e a r ly
m utant i s o l a t e d by J o fr e e t a l . (1 9 8 1 ) a r e in su p p o r t o f th e
e x i s t e n c e o f a s l e a s t two fi and y c l a s s e s . P o s t e t a l . (1 9 8 1 )
e s t a b l i s h e d a c e l l l i n e c o n ta in in g a f t gen e IdPyK) under
oc (Vmw IK 1 7 5 ) t r a n s c r ip t io n a l c o n t r o l and fou n d t h a t dPyK
a c t i v i t y was in d u c e d by i n f e c t i o n w ith dPyK v i r u s . T h is*
e f f e c t was shown n o t to be due to a v ir u s oC f u n c t io n , so
th e y in t e r p r e t e d i t a s th e s t im u la t io n o f t r a n s c r ip t io n from
th e cx prom oter by a v i r io n s t r u c t u r a l p o ly p e p t id e , t h a t i s ,
a pre-cx f u n c t io n . I t i s c l e a r t h a t a more o r l e s s com plex
s e t o f c a sc a d e c o n t r o l s i s o p e r a t iv e in th e tem p o ra l e x p r e s s io n
o f HSV-induced p o ly p e p t id e s , p ro b a b ly a c t in g a t th e
t r a n s c r ip t io n a l l e v e l .
T reatm ent o f 1-JSV-infected c e l l s w ith p r o te a s e

- 31 -
i n h i b i t o r s f a i l e d to show ra p id p o s t - t r a n s l a t i o n a l c le a v a g e
o f v ir u s - in d u c e d p o ly p e p t id e s (H oness and Roizm an, 1973;
P e r e ir a e t a l . , 1 9 7 7 ) . In c o n t r a s t , ex p o su re o f v i r u s -
in f e c t e d c e l l s to th e i n h i b i t o r f o r lo n g e r p e r io d s o f tim e
showed a l t e r e d p r o c e s s in g o f s e v e r a l p o ly p e p t id e s (I).
M acDonald, M. Suh and U .S . M arsden, u n p u b lish e d d a t a ) .
A p p rox im ately 20 p h o sp h o r y la ted p o ly p e p t id e s have been
i d e n t i f i e d i n H S V -in fected c e l l s (M arsden e t a l . , 1978;
Bookout and L evy, 1980; W ilcox e t a l . , 1 9 8 0 ) . The
p h o sp h o r y la t io n pathway may d i f f e r betw een p o ly p e p t id e s and
th e l e v e l o f p h o sp h o r y la t io n may p la y a p a r t in th e b in d in g .
a f f i n i t y o f c e r t a in p h o sp h o p r o te in s f o r DNA (F enw ick e t a l . ,
1 9 80; W ilcox et, a l . , 1 9 8 0 ) . The p r o c e s s in g o f th e
p h o sp h o r y la te d c* p o ly p e p t id e Vmw IE 175 (IGP4) h a s been s tu d ie d
in some d e t a i l . T h is p o ly p e p t id e e x i s t s i n th r e e form s -
d i f f e r i n g in ap p aren t m o le c u la r w e ig h t , and i t i s th o u g h t
th a t ICP4a i 3 p a s s iv e ly tr a n sp o r te d in t o th e n u c le u s , where
i t i s p r o c e s se d in t o ICP4b th en ICP4c ( P e r e ir a e t a l . , 1977;
Fenw ick e t a l . , 1 9 7 8 ) . The n a tu r e o f t h e s e p r o c e s s in g e v e n ts
has n o t been d e term in ed , a lth o u g h a m utant w ith a l e s i o n in
th e ICP4 gen e h as been s tu d ie d w hich a ccu m u la tes ICP4a and
f a i l s to tr a n s p o r t i t in t o th e n u c le u s (C ou rtn ey and P o w e ll ,
1975; C ourtney e t a l . , 1976; C ab ral e t a l . , 1 9 8 0 ) . The
p h o sp h a te m o ie ty i s s t a b ly a s s o c ia t e d w ith ICP4b but c y c l e s
on and o f f ICPAa and ICP4c (W ilcox e t a l . , 1 9 8 0 ) . Bookout
and Levy (1 9 8 0 ) have r e c e n t ly d e s c r ib e d fo u r form s o f ICP4,
o n ly two o f w hich a re found in th e c y to p la sm .
A number o f nSV -induced p o ly p e p t id e s a re g ly c o s y la t e d ,
jjrob ab ly by h o s t c e l l enzym es (K e lle r e t a l . , 1 9 7 0 ) .
G ly c o s y la t io n o f p o ly p e p t id e s o c c u r s in s i t u , soon a f t e r
p o ly p e p t id e s y n t h e s is and t h e ir b in d in g to c e l l membranes

- 32 -
(S p ear and Roizm an, 1 9 7 0 ) . F iv e m ajor g ly c o p r o t e in s (gA, gB,
gC, gl) and gB) have been i d e n t i f i e d in th e v i r io n e n v e lo p e
and in f e c t e d c e l l membrane (S p ear e t a l . , 1970; S p ea r , 1976;
M arsden e t a l . , 1976; Baucke and S p ea r , 1 9 7 9 ) . gA i s r a r e in
v i r i o n s , and gB f u n c t io n s i n v ir u s p e n e t r a t io n and c e l l
f u s io n (M a n serv ig i and S p ea r , 1 9 77; S arm ien to e t a l . , 1 9 7 9 ) .
T hese two g ly c o p r o t e in s a re th o u g h t to o r ig in a t e from th e
same p r e c u r so r p o ly p e p t id e (E b e r le and C ou rtn ey , 1 9 8 0 b ) .
gC h a s an in h ib i t o r y e f f e c t upon c e l l f u s io n , and i s n o t
e s s e n t i a l f o r v ir io n i n f e c t i v i t y (M a n serv ig i and S p ea r , 1977;• .
Ruyechan e t a l . , 1 9 7 9 ) . The l e a s t s tu d ie d o f th e g ly c o p r o t e in s ,
gE, i s e s s e n t i a l f o r v i r io n i n f e c t i v i t y and a c t s a s an Fc
r e c e p to r f o r y ' -g lo b u l in , a f u n c t io n w hich may i n h i b i t th e
h o s t immume c y t o l y t i c r e sp o n se m ed ia ted by v i r a l g ly c o p r o t e in s
ex p o sed a t th e in f e c t e d c e l l s u r fa c e (Baucke and S p ea r , 1979;
P ara e t a l . , 1 9 80; M a ch tig er e t a l . # 1 9 8 0 ) .
G ly c o s y la t io n o f p r e c u r so r p o ly p e p t id e s i s a f t o r y
e v e n t o c c u r r in g in two s ta g e s * r a p id a d d it io n o f an
o lig o m a n n o sy l co r e fo llo w e d by s lo w e r a d d it io n o f fu c o s e and
s i a l i c a c id r e s id u e s (H oness and Roizm an, 1 9 7 5 b ;E isen b e rg
e t a l . , 1 9 7 9 ; Cohen e t a l . , 1 9 8 0 a ) . A ttem p ts have been made
to i d e n t i f y p r e c u r s o r form s by im m u n o lo g ica l a n a l y s i s , th e
u se o f d ru gs w hich b lo c k th e g ly c o s y la t io n pathw ay, tr e a tm e n t
o f p r o c e s se d p o ly p e p t id e s w ith g l y c o s id a s e s , and by p u ls e - c h a s ebetry
e x p e r im e n ts . C o n c lu s io n s h a v e ^ c r i t ic i s e d on th e grou n d s t h a t
breakdown p r o d u c ts and tr u e p r e c u r s o r s a r e d i f f i c u l t to
d i s t i n g u i s h . N e v e r t h e le s s , tw o -d im e n s io n a l e l e c t r o p h o r e t i c
a n a l y s i s s t r o n g ly i n d i c a t e s th a t 16 s p e c ie s a r e in v o lv e d in
th e fo r m a tio n o f gB, 1 5 -2 0 in gC, and 11 i n gD (Cohen e t a l . ,
1980a; Haarr and M arsden, 1 9 8 1 ) .
IISV-1 g ly c o p r o t e in s a re a l s o s u lp h a te d , and a su lp h a te d

- 33 -
breakdown p ro d u ct o f gl) i s e x c r e te d from th e i n f e c t e d c e l l
(R .G . Hope and H .S . M arsden, p e r s o n a l co m m u n ica tio n ).
The m ajor g ly c o p r o t e in o f P R V -in fec te d c e l l s i s su lp h a te d
su b seq u en t to g ly c o s y la t io n and th en e x c r e te d (E r ick so n and
K aplan, 1973; K aplan and B en -P o ra t, 1 9 76; P e n n in g to n and
McCrea, 1 9 7 7 ) /
Much o f th e p r e s e n t u n d e r sta n d in g o f H S V -sp e c if ie d
f u n c t io n s , and th e l o c a t io n o f e s s e n t i a l g e n e s i n th e genom e,
h as come from a n a ly s e s o f m u ta n ts . In t h i s r e s p e c t th e
commonest and m ost g e n e r a l ly u s e f u l ty p e i s th e t s m utant,
w hich was f i r s t d e s c r ib e d i n HSV by Subak-Sharpe (1 9 6 9 ) .
S in c e th en a la r g e number o f t s m u tan ts have been i s o l a t e d .
23 com p lem en ta tion grou p s in HSV-l and 20 i n HSV-2 w ere
i d e n t i f i e d in a c o l la b o r a t iv e s tu d y by S c h a f fe r e t a l . ( 1 9 7 8 ) ,
and th e r e c e n t u se o f s p e c i f i c m u ta g e n e s is o f DNA fra g m en ts
h as added a t l e a s t th r e e more to HSV-l (Chu e t a l . , 1979;
C onley e t a l . , 1 9 8 1 ) . L in e a r g e n e t ic l in k a g e maps f o r t s ,
p la q u e m orphology and drug r e s i s t a n c e m arkers have been
deduced (Brown e t a l . , 1973; S c h a f fe r e t a l . , 1 9 74;
Subak-Sharpe e t a l . , 1974; B en y esh -M eln ick e t a l . , 1974;
Timbury and C a ld er , 1976; Brown and Jam ieson , 1 9 7 7 ) .
R ecom b in ation in HSV was f i r s t n o ted by W ildy (1 9 5 5 ) ,
and in t e r t y p i c com p lem en ta tion and r ec o m b in a tio n by Timbury
and Subak-Sharpe ( 1 9 7 3 ) . R ep eated rounds o f m atin g and
r ec o m b in a tio n o c c u r d u rin g i n f e c t i o n , and reco m b in a n ts seem
to be produced by th e fo rm a tio n o f p a r t ia l h e t e r o z y g o te s
(Brown and R i t c h ie , 1975; R it c h ie e t a l . , 1 9 7 7 ) . E x c e l le n t
u se h as been made o f in t r a t y p ic (m arker r e s c u e ) and in t e r t y p ic
r ec o m b in a tio n to p h y s i c a l ly map IiSV m u ta tio n s and f u n c t io n s
on th e genome (W ilx ie e t a l . , 1977b; Marsden e t a l . , 1978;
Morse e t a l . , 1978; P r e s to n e t a l . , 1978; Stow and W ilk ie ,

1978; Stow e t a l . , 1978; H a ll ib u r to n , 1980; P a r r is e t a l . ,
1980; C hartrand e t a l . , 1981; S a n d r i-G o ld in e t a l . , 1 9 8 1 ) .
HSV-l/HSV-2 i n t e r t y p i c reco m b in a n ts have p roved e s p e c i a l l y
p o w erfu l in a s s ig n in g s p e c i f i c p o ly p e p t id e s and b io c h e m ic a l
f u n c t io n s to r e g io n s o f th e genom e, s in c e r e s t r i c t i o n
e n d o n u c lea se mapping a l lo w s c r o s s o v e r s to be a c c u r a t e ly
d e f in e d , and th e p o ly p e p t id e p r o f i l e s o f th e two s e r o ty p e s
are s u f f i c i e n t l y d i f f e r e n t to d i s t i n g u i s h th e o r ig in o f
many o f th e p o ly p e p t id e s in d u ced by r e c o m b in a n ts . A summary
o f th e mapping p o s i t i o n s o f s e v e r a l p o ly p e p t id e s i s shown
in F ig u r e A1.5*
Enzyme a c t i v i t i e s in d u ced by HSV
S e v e r a l en zym atic a c t i v i t i e s a re in d u ced i n H S V -in fected
c e l l s . T hese in c lu d e p y r im id in e d e o x y n u c le o s id e k in a s e (dPyK)
(K it and Dubbs, 1 9 6 3 a ) , BNA p o lym erase (K e ir and G old, 1963)>
a l k a l in e d e o x y r ib o n u c le a se (M orrison and K e ir , 1 9 6 8 ) ,
r ib o n u c le o t id e r e d u c ta se (Ponce de Leon e t a l . , 1977;
H uszar and B a c c h e t t i , 1 9 8 1 ) , a d e n y lic a c id sd e o x y th y m id in e
5 1-p h o s p h o tr a n s fe r a s e (Jam ieson e t a l . , 1976a; F a lk e e t a l . ,
1 9 8 1 ) , d e o x y r ib o p y r im id in e t r ip h o s p h a ta s e (W ohlrahb and
F ra n ck e , 1 9 8 0 ) , and a p r o t e in k in a s e (B lu e and S to b b s , 1 9 8 1 ) .
The in d u c t io n o f d e o x y c y t id in e d eam inase (Chan, 1977;
B. B u t ia , p e r s o n a l com m unication) and d e o x y c y t id y la t e
d eam in ase (H o lto n and K e ir , 1974; l a n g e l i e r e t a l « , 1 9 7 8 )
i s d i s p u t e d . A BNA g y r a se has been so u g h t but n o t y e t
d em on stra ted ( Francke and M a rg o lin , 1 9 8 1 ) .
The i s o l a t i o n o f an HSV-2 t s m utant w h ich a l s o h a s a
n o n - le t h a l m u ta tio n in th e s t r u c t u r a l gen e f o r th e a lk a l in e
d e o x y r ib o n u c le a se has d em on strated th a t t h i s enzyme i s a t
l e a s t in p a r t coded by th e v ir u s genome (F rancke e t a l . , 1 9 7 8 ) .
The p u r i f i e d enzyme h as a ra.wt. o f 9 0 ,0 0 0 , but i t s f u n c t io n

- 35 -
in v iv o i s a m a tter o f s p e c u la t io n ( S t r o b e l - F id le r and F rancke,
1 9 8 0 J . Moss, e t a l . (1 9 7 9 ) mapped th e n u c le a s e gen e to
0 .1 2 - 0 .2 1 f r a c t i o n a l genome u n i t s .
The v ir u s -c o d e d BNA p o lym erase h a s a m .w t.
o f 1 5 0 ,0 0 0 and seem s to be a c t i v e i n v i t r o a s a monomer
(P o w e ll and P u r ifo y , 1 9 7 7 ) . T h is enzyme i s r e a d i ly
d is t in g u is h e d from th e c e l l BNA p o ly m era se by im m u n o lo g ica l
(K e ir e t a l . , 1 9 6 6 ) , g e n e t i c ' (Aron e t a l . , 1 9 7 5 ) and i n h i b i t o r
s t u d i e s . The HSV BNA p o ly m era se i s v e r y s e n s i t i v e to
i n h i b i t i o n by p h o sp h o n o a c e t ic a c id (PAA; Mao and R obishaw ,
1 9 7 5 ; Hay and Sub ak -S h arp e, 1 9 76) and by a c y c lo g u a n o s in e
( a c y c l o v i r , ACV) in th e t r ip h o s p h a te form (Furman e t a l . ,
1 9 7 9 ; Barby e t a l . , 1 9 8 0 ) . M utant v ir u s e s r e s i s t a n t to PAA
p o s s e s s a P A A -r e s is ta n t . BNA p o ly m era se ( J o fr e e t a l . , 1977;
P u r ifo y and P o w e ll, 1 9 7 7 ) , s u g g e s t in g t h a t a s in g l e lo c u s i s
in v o lv e d i n P A A - s e n s i t iv i t y . A C V - s e n s it iv i t y , on th e o th e r
hand, i s a s s o c ia t e d w ith two d i s t i n c t l o c i c o r r e sp o n d in g to
th e BNA p o lym erase and dPyK (Crumpacker e t a l . , 1 9 8 0 ) . T h is
d i f f e r e n c e i s due to d i r e c t i n t e r a c t i o n o f PAA w ith th e
p o ly m e r a se , w hereas ACV i s f i r s t p h o sp h o r y la te d by dPyK, th e n c e
to th e tr ip h o s p h a te w hich i n h i b i t s th e p o ly m era se ( E lio n e t a l . >
1 9 7 7 ) . I t i s p o s s ib l e t h a t two c i s t r o n s a r e in v o lv e d i n
BNA p o ly m era se e x p r e s s io n , s in c e t s m u tan ts from two
co m p lem en ta tio n g ro u p s, w hich a re c l o s e l y a s s o c ia t e d i n th e
r e g io n o f 0 .4 f r a c t i o n a l genome u n i t s (C hartrand e t a l . ,
I 9 6 0 ) , both e x p r e s s a t s BNA p o lym erase (P u r ifo y and P o w e ll , 1 9 8 1 ) .
The v ir u s -c o d e d dPyK, a s i t s name s u g g e s t s , i s a b le to
p h o sp h o r y la te both th y m id in e and d e o x y c y t id in e , and seem s a l s o
to e x h ib i t a n u c le o s id e p h o sp h o tr a n s fe r a s e a c t i v i t y (Jam ieson
e t a l . , 1974 and 1 9 7 6 a ;J a m ieso n and Subak -S h arp e, 1 9 7 4 ) .
S tro n g e v id e n c e th a t t h i s enzyme i s v ir u s -c o d e d com es from

- 56 -
im m u n o lo g ic a l, b io c h e m ic a l and g e n e t ic s t u d ie s (K lem perer
e t a l . , 1967; Buchan and W atson, 1 9 6 9 ; Buchan e t a l . , 1970;
T h o u le s s , 1 972; Aron e t a l . , 1 9 7 3 ) . The a c t i v e p o ly p e p t id e
in d u ced i n v iv o o r produced in v i t r o by t r a n s la t io n o f mRNA
h as an ap p aren t m .w t. o f 4 0 ,0 0 0 (H oness and W atson, 1974;
Summers e t a l . , 1975; T h o u le ss and W ildy, 1 9 75; P r e s to n , 1977;
Cremer e t a l . , 1977 and 1 9 7 8 ) . T w o -d im en sio n a l g e l
e le c t r o p h o r e s i s o f H SV-induced p o ly p e p t id e s showed th a t t h i s
p o ly p e p t id e may e x i s t i n th r e e d i f f e r e n t l y ch arged form s
(H aarr and M arsden, 1 9 8 1 ) . P r e s to n and McGeoch (1 9 8 1 )
i d e n t i f i e d a s l i g h t l y s m a lle r p o ly p e p t id e e x p r e s se d in v i t r o
and in v i v o , c o n c lu d in g t h a t t h i s was th e r e s u l t o f f a l s e
i n i t i a t i o n o f t r a n s l a t io n from a s i n g l e mRNA s p e c i e s .
The r e s u l t s o f ex p er im e n ts u s in g mixed i n f e c t i o n s o f dPyK
and dPyK v ir u s e s s u g g e s t t h a t th e a c t i v e enzyme i s
p ro b a b ly a rau ltim er (Jam ieson and S u b ak -S h arp e, 1 9 7 8 ) .
A number o f n u c le o s id e a n a lo g u e s i n h i b i t v i r u s grow th b ecau se
t h e y a r e p h o sp h o r y la ted by dPyK, and th e n c e b ind to th e
BNA p o ly m era se o r become l e t h a i l y in c o r p o r a te d in t o n ew ly
s y n t h e s is e d v ir u s BNA. Among t h e s e a re ACV, b ro m o d eo x y cy tid in e ,
b rom od eoxyu rid in e and 1 -^ - a r a b in o s y l th ym in e (K it and Bubbs,
1 9 6 3 ^ ;C ooper, 1973; S c h ild k r a u t e t a l . , 1 9 75; M il le r e t a l . ,
1 9 7 7 ; E lio n e t a l . , 1 9 7 7 ; M u ller e t a l . , 1 9 7 9 b ) . Such
i n h i b i t o r s have been u sed to s e l e c t v ia b le dPyK~ m u ta n ts .
A lth o u g h dPyK a c t i v i t y i s n o t r e q u ir e d f o r v ir u s r e p l i c a t io n
in a c t i v e l y d iv id in g c e l l s , i t i s an e s s e n t i a l f u n c t io n f o r
grow th in r e s t i n g c e l l s , f o r f u l l n e u r o v ir u le n c e , and f o r
norm al e s ta b lis h m e n t o f la t e n c y (Jam ieson e t a l . , 1974;
F ie ld and B arby, 1978 and 1980; Barby e t a l . , 1 9 8 1 ) .
The c o n s tr u c t io n o f b a c t e r ia l p la sm id s c o n ta in in g th e

- 37 -
HSV-1 dPyK gen e (HSV-1 Bamil l £ mapping a t a p p ro x im a te ly 0 .3
f r a c t i o n a l genome u n i t s ) has opened th e way to u n d ersta n d in g
th e m o le c u la r g e n e t i c s o f t h i s f u n c t io n (W ilk ie e t a l . , 1979a;
B n q u ist e t a l . , 1979; C o lb ere-G a ra p in e t a l . , 1 9 7 9 ) . The
DMA seq u en ce o f th e gene has been d eterm in ed (M cKhight, 1 9 80;
Wagner e t a l . , 1981; P. McGeoch, p e r s o n a l co m m u n ica tio n ), and
v ia b le dPyK"" d e l e t io n m u tan ts have been c o n s tr u c te d in v i t r o
(W ilk ie e V a l . , 1980; S m ile y , 1 9 8 0 ) . The dPyK i s a ^
p o ly p e p t id e w h ich r e q u ir e s f o r i t s p r o d u c t io n i n th e in f e c t e d
c .e l l an oL f u n c t io n w hich a c t s a t th e t r a n s c r ip t io n a l l e v e l
(G a r f in k le and M cAuslan, 1974; Leung e t a l * , 1 9 8 0 ) . The
gene i s t r a n s c r ib e d from i t s own prom oter le f tw a r d s w ith
r e s p e c t to th e P o r ie n t a t io n o f th e genom e, and th e mRNA i s
n o t s p l i c e d (W ilk ie e t a l . , 1980; S m iley e t a l . , 1 9 8 0 ) .
The HSV-1 dPyK gen e has been u sed a s a s e p a r a te e n t i t y
from th e genome from w hich i t was d e r iv e d . C e l l
l i n e s la c k in g th e c e l l th y m id in e k in a s e have been b io c h e m ic a lly
tra n sfo rm ed u s in g in a c t iv a t e d HSV or DNA fra g m e n ts (Munyon
e t a l • , 1971; Jam ieson e t a l . , 1 9 7 8 b ;M a itla n d and M cD ougall,
19 77; W ig ler e t a l . , 1977)* U sin g a c lo n e d DNA fragm en t
c o n ta in in g th e dPyK g e n e , P e l l i c e r e t a l . (1 9 7 8 ) have r e p o r te d
t h a t th e v ir u s DNA i s in t e g r a t e d i n t o th e c e l l DNA, a lth o u ^ i
n o t .at a s p e c i f i c s i t e . O ther exogen ou s DNA seq u e n c es have
been s t a b ly in tr o d u c e d in t o c e l l s by em p loy in g th e dPyK gene
a s a s e l e c t a b l e m arker, i n some e x p e r im e n ts o o v a le n t ly l in k e d
to th e ex o g en o u s DNA (M antei e t a l . , 1979; W ig ler e t a l . , 1979)
A p p rox im ately 20 H S V -l-in d u ced DNA b in d in g p r o t e in s
have been i d e n t i f i e d , in c lu d in g the. oi p o ly p e p t id e s , th e
DNA p o ly m er a se , and p ro b a b ly th e a lk a l in e d e o x y r ib o n u c le a se
and dPyK ( B a y l i s s e t a l . , 1975; P o w e ll and P u r ifo y , 1976;
B ookout and L evy , 1980; Becker e t a l . , 1 9 8 0 ; Hay and Hay, 1 9 8 0 )*

Figure Al.lUPPER PANEL S tr u c tu r a l arrangem ent o f th e HSV-1
genom e. The genome co m p r ises two c o v a le n t ly l in k e d segm ents
(L, S ) , each o f w hich c o n s i s t s o f a u n iq u e seq u en ce (UL, Ug)
bounded by in v e r te d r e p e t i t i o n s (TH^, IR^, TRg, IR § ). '
and Us a re sh o r te n e d in t h i s diagram to em p h asise th e
l o c a t i o n s and o r i e n t a t io n s (arrow ed) o f th e a s e q u e n c e s , or
t e r m in a l r e p e t i t i o n s ( t r ) a t th e genome te r m in i and L-S
j o i n t . A lso shown a re th e b and c seq u e n c e s ( TRj, = b + a ,
I R1 * y TRg» c + a , IRg** c f+ a f ) . S i z e s a r e g iv e n in kbp.
A S in g le DNA m o le c u le c o n ta in s each segm ent i n e i t h e r
o r ie n t a t io n w ith e q u a l p r o b a b i l i t y , a s shown by th e la r g e r
a rro w s, a l lo w in g fo u r p erm u ta tio n s o f seq u en ce arran gem en t.
The genome s t r u c t u r e o f HSV-2 i s e s s e n t i a l l y i d e n t i c a l .
LOWER PANEL G en era tio n o f subm olar r e s t r i c t i o n
e n d o n u c le a se fra g m en ts from HSV DNA. A h y p o th e t ic a l
e n d o n u c le a se c le a v e s th e DNA in th r e e l o c a t i o n s . As a
co n seq u en c e o f in v e r s io n o f th e two seg m en ts, fo u r q u a r te r -
m olar fra g m en ts sp a n n in g th e L-S ju n c t io n (a -t -e , c + e ,
a-t-b , c + b ) , fo u r h a lf -m o la r fra g m en ts from th e te r m in i
(a , b, c , e ) , and one m olar fragm en t (d ) a re g e n e r a te d .
E x te n s io n o f t h i s argum ent shows t h a t i f one s e t o f
r e p e a t s a l s o c o n t a in s a r e s t r i c t i o n s i t e , h a lf -m o la r but
n o t q u a r te r -m o la r fra g m en ts would be g e n e r a te d . I f both
s e t s o f r e p e a t s a l s o c o n ta in e d a r e s t r i c t i o n s i t e , o n ly
m olar fr a g m en ts w ould r e s u l t .
In t h i s F ig u re th e fo u r arran gem en ts have been
term ed P , 1^, I g , Igq, in accord an ce w ith th e n om en cla tu re
p ro p o sed by Roizman e t a l . (1 9 7 9 ) . R e s t r ic t io n maps o f
HSV BNA a re c o n v e n t io n a l ly d isp la y e d in th e P arran gem en t.
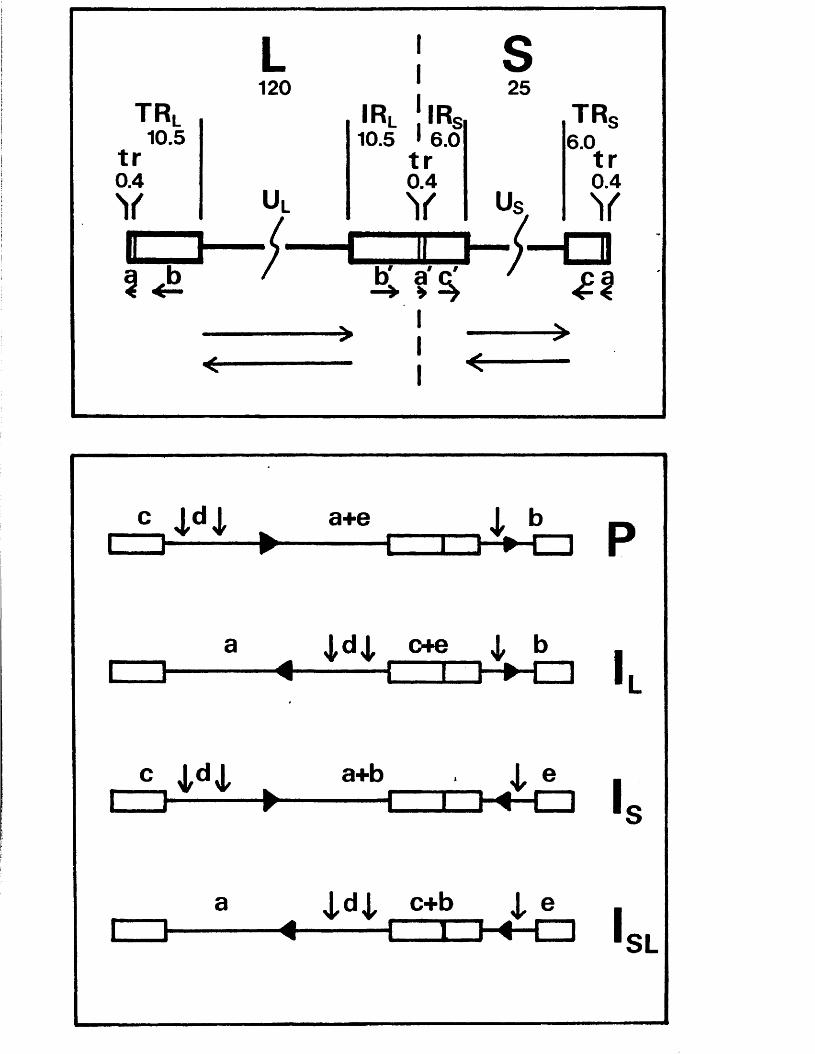
L II
IRl [iRs10.5 • 6.0
s120 25
TRl TRS10.5 6.0
t r t r t r0.4
Ul0.4 0.4
V Y Us# YI
a+e
a+b
SL

Figure A1.2 ( o v e r le a f )
S tr u c tu r e s o f h e r p e s v ir u s genom es and g e n e r a l
m odel f o r in te r c o n v e r s io n o f l i n e a r and c i r c u l a r fo r m s .
L in e a r genome s t r u c t u r e s o f th e fo u r main
groups ( I - I V ) a re shown to s c a le o n th e r ig h t hand p a g e .
Two sub grou p s have been t e n t a t i v e l y in c lu d e d in group I .
In each c a s e th e m .w t. (x 10~^) and o b serv ed o r ie n t a t io n s ( f i l l e d
arrow heads) o f and, where a p p r o p r ia te , Ug, a re in d ic a t e d .
R epeated se q u e n c e s a re shown a s r e c t a n g le s , and t h e ir
r e l a t i v e o r i e n t a t io n s by arrow s and by u se o f th e l e t t e r s
a , b, c and t h e i r com plem ent a 1, b 1, c 1 ( —A a n d a^ , bn
s i g n i f y m u lt ip le tandem r e p e a t s ) . The u se o f l e t t e r s d o es
n o t d en o te s i m i l a r i t y o f seq u en ce o r f u n c t io n in th e s e
r e g io n s betw een d i f f e r e n t genom es.
I t i s known th a t th e genom es i l l u s t r a t e d are
t e r m in a lly red u n d a n t, a lth o u g h th o s e o f PRV and EHV-1 have
n o t y e t been d i r e c t l y shown to be s o . N e v e r t h e le s s , t h i s
f e a tu r e makes i t p o s s i b l e , a t l e a s t in p r i n c i p l e , to
c i r c u l a r i s e h e r p e s v ir u s genom es by l i g a t i o n su b seq u en t to
th e a d t io n o f an e x o n u c le a s e . The fo u r g rou p s o f p u ta t iv e
c ir c u la r form s a re shown on th e l e f t hand p a g e , n o t drawn
to s c a l e . O r ie n ta t io n s o f r e p e a te d se q u e n c es ( r e c t a n g le s )
have been o m itte d f o r c l a r i t y . P o s i t i o n s o f l i g a t i o n on
o p p o s ite s tr a n d s o f th e d u p lex a r e in d ic a t e d ( ▼ ) .
C le a r ly , th e r e v e r s e p ro ce ss ., s in g l e s tra n d ed c le a v a g e on
each s id e o f th e te r m in a l r e p e a t w ith in a c i r c u l a r DNA
m o le c u le fo llo w e d by r e p a ir o f th e s in g l e s tra n d ed
te r m in i, would g e n e r a te a l i n e a r genom e. C lea v a g e may
o ccu r a t s e v e r a l p o s s ib l e l o c a t io n s w ith in th e tandem
r e i t e r a t i o n s o f grou p s I I and I I I , g e n e r a t in g th e
h e te r o g e n e o u s te r m in i o b serv ed in v ir io n DNA.
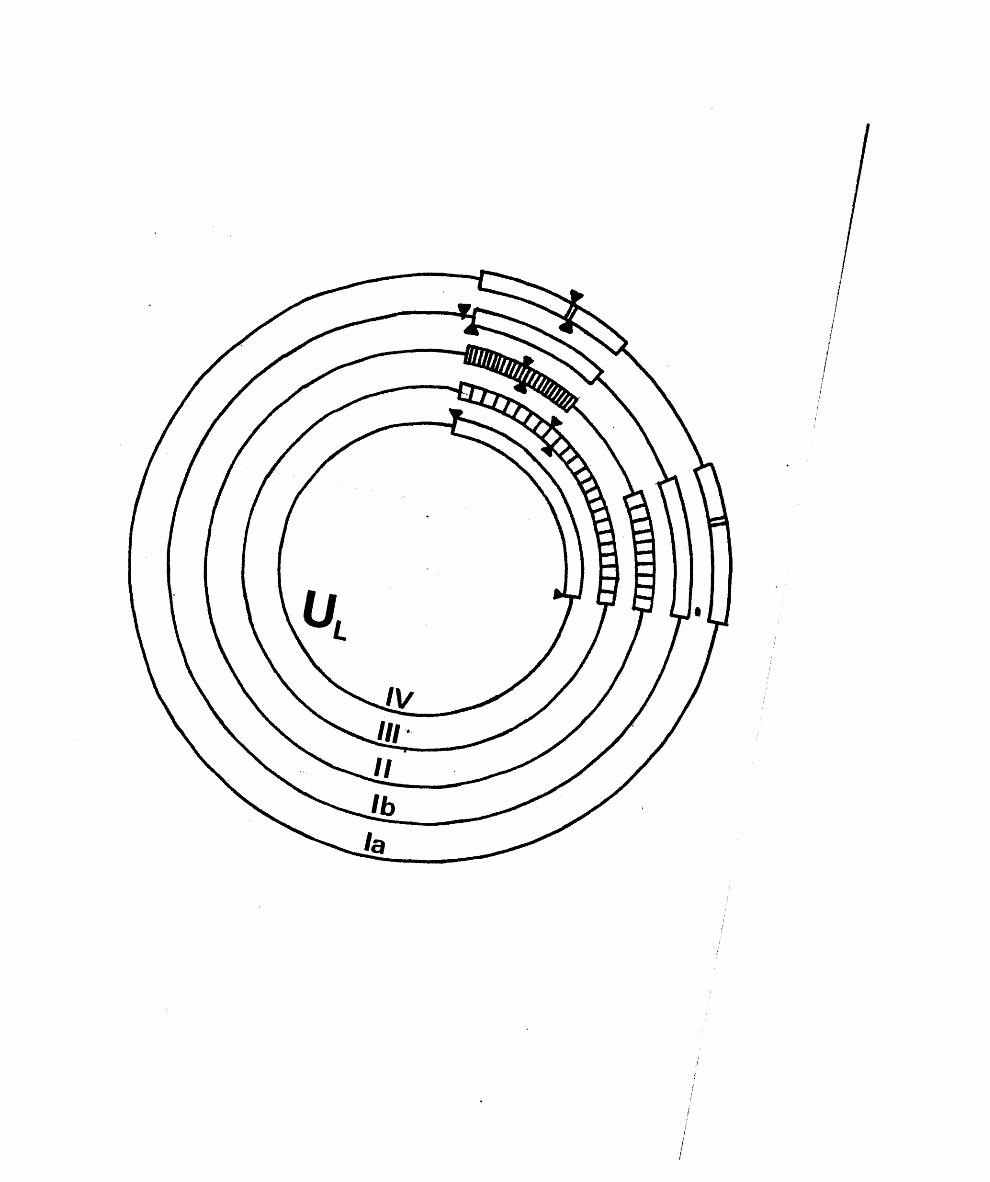
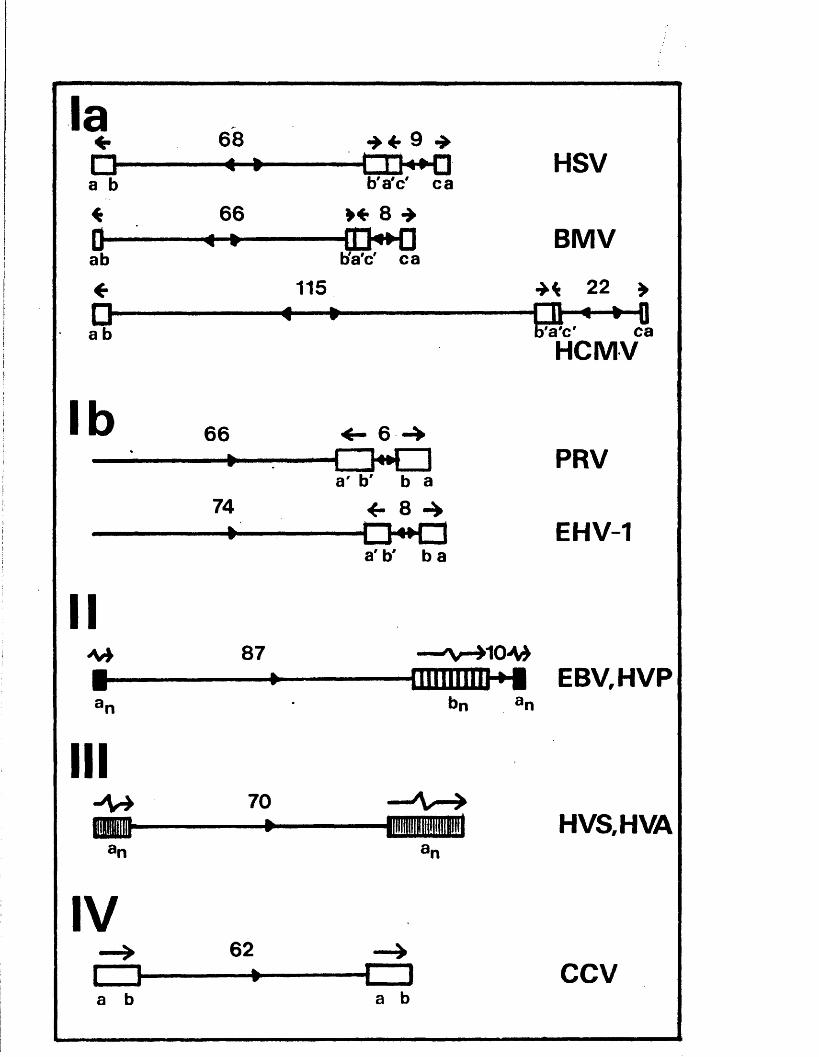
6 8 * * 9 *D - .<■ »■ - ■CEKK3 HSVa b b'a'c' ca
« 6 6 »«■ 8 *
D <->------------QU*HU BMVab ba'c' ca«■ 115 22 >□ ----------------------------------- «— »------------------------------------ c n « » - qa b B’a’c' ca
HCMV
lb
II
IIIan
I V
66 6— • — — i u * n p r v
a' b' b a 74 4" 8
— ■ 0 * 0 EHV-1a' b' b a
M 87 —i ------------------ » nnnnnw i ebv ,h v pa n b n a n
-V> 70»------------ j I HVS,HVA
62 ____I— I------------ *------------[ZD CCVa b a b

Figure A1.3
Mean GC c o n t e n t s o f h e r p e s v ir u s DNAs, tak en
from H oness and Watson (1 9 7 7 b ) . F i l l e d c i r c l e s in d ic a t e
human h e r p e s v ir u s e s .
A b b r e v ia t io n s o f v ir u s n om en cla tu re o th e r th an
th e o b v io u s a re a s f o l l o w s .
SpM - s p id e r monkey h e r p e s v ir u s(c e b id h e r p e s v ir u s 3)
IBR - i n f e c t i o u s b o v in e r h i n o t r a c h e i t i sv ir u s
SqM - s q u ir r e l monkey h e r p e s v ir u s(h e r p e s v ir u s ta m a rin u s)
GCMV - g u in e a p ig c y to m e g a lo v ir u s
MaCMV - marmoset c y to m e g a lo v ir u s
SA6 - s im ia n c y to m e g a lo v ir u s
HhCMV - r h e su s c y to m e g a lo v ir u s
VeCMY - v e r v e t c y to m e g a lo v ir u s
FelLT - f e l i n e i n f e c t i o u s la r y n g o t r a c h e i t i sv ir u s
AvILT - a v ia n i n f e c t i o u s la r y n g o t r a c h e i t i s
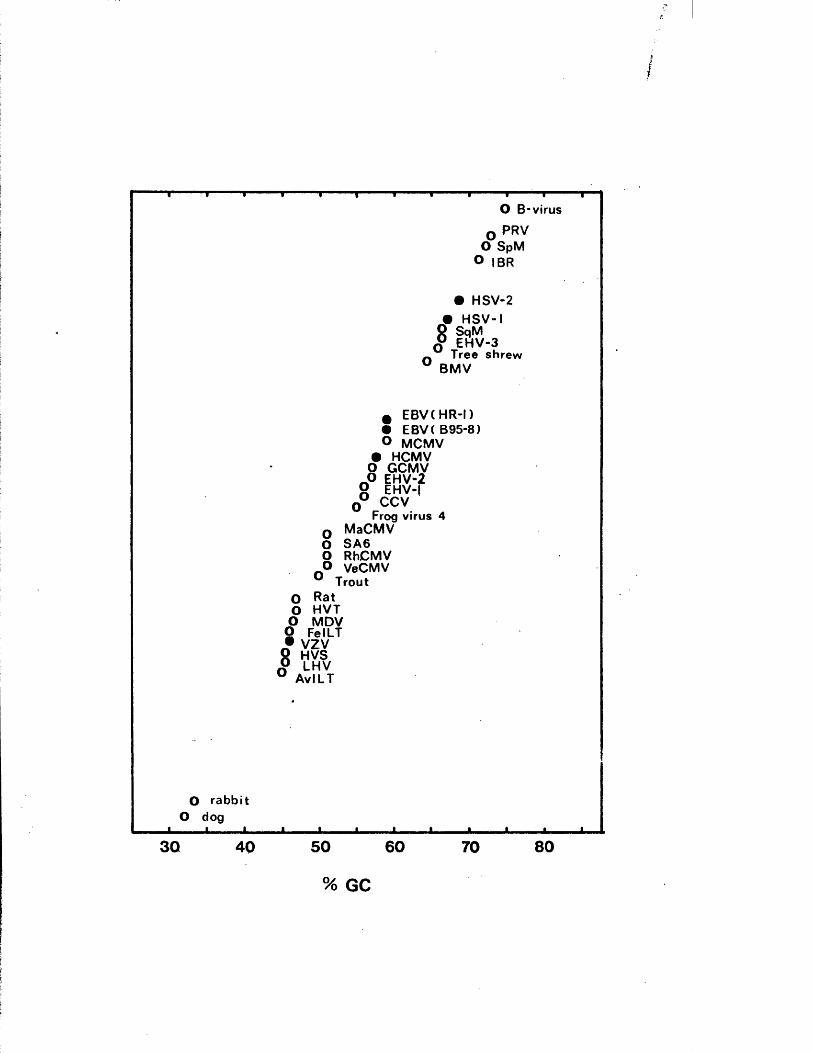
oO SpM
O IBR
O B-virus
PRV
8
• HSV-2HSV-I
SqM EHV-3
Tree shrewBMV
OOoo•
8
# EBV(HR-I)• EBV( B95-8) O MCMV
• HCMV8 GCMV EHV-2 « EHV-I
0° c c vFrog virus 4
MaCMV SA6 RhCMV VeCMV
u Trout Rat HVT MDV
FelLT VZV HVS LHV
AvILT
Oooo
O rabbi t O dog L 1, J.
3Q 40 50 60
% GC
70 80

Figure A1.4 (overleaf)
Immediate e a r ly (IE) t r a n s c r ip t io n a l program o f
HSV-1. The genome i s shown to s c a le in the c o n v e n tio n a l
manner, e x ce p t th a t two r e g io n s in have been om itted
f o r c l a r i t y ( A r ) , The s c a le a t the top o f th e F igure
shows c o o r d in a te s o f f r a c t io n a l genome le n g th .
The lo c a t io n s and d ir e c t io n s o f t r a n s c r ip t io n
o f IE mRNAs 1 -5 are shown (C lem ents e t a l . , 1977; Jones
e t a l . , 1977; C lem ents e t a l . , 1979; Anderson e t a l* , 1980a;
F .J . R ixon , p e r so n a l com m unication). A lso in c lu d e d are
s im ila r d a ta fo r th e 5 .0 kb mRNA cod ing fo r Vmw IE 136*(143)
(Anderson e t a l . , 1981; J . McLauchlan and J .B . C lem ents,
p e r so n a l com m unication). The p o s i t io n o f the in tr o n
common to the gen es fo r IE mRNAs 4 and 5 i s in d ic a te d
(Watson e t a l . , 1981; F .J . R ixon, p erso n a l com m unication).
I t i s su sp ec ted th a t th e gene fo r IE mRNA 1 a ls o c o n ta in s
an in tr o n , because th e mRNA i s c o n s id e r a b ly sm a ller than
th e r e g io n o f ENA encoding i t , whereas th e gen es fo r
IE mRNAs 2 and 3 c o n ta in no d e te c ta b le in tr o n s (F .J . R ixon,
p e r so n a l com m unication). The r e g io n s in the ENA encoding
th e 5 1 term in i o f IE mRNAs 1 , 3 , 4 and 5 have been lo c a te d
(Mackern and Roizman, 1980) and n u c le o t id e seq u en ces
determ ined fo r th e se r e g io n s o f IE mRNAs 3 , 4 and 5
(M-J. M urchie, p erso n a l com m unication). S im ila r ly th e
r e g io n s encod in g the 3' term in i o f IE mRNAs 1 , 2 and 5 are
known (S e c t io n 3 o f R e su lts ; M-J. Murchie and F .J . R ixon,
p e r so n a l com m u nication s). The sequence cod in g f o r th e
tr a n s la te d r eg io n o f IE mRNA 5, which l i e s co m p lete ly
w ith in Ug, has been determ ined (M-J. M urchie, p erso n a l
com m unication). The 5 .0 kb mRNA cod in g fo r Vmw IE 1 3 6 '(1 4 3 )

i s ex p ressed m in im ally under IE c o n d it io n s and th e r e fo r e
th ere i s some h e s i t a t io n in i t s in c lu s io n in the IE
c l a s s . A 1 .2 Jcb mRNA 3 1 -c o te r m in a l w ith th e 5*0 kb mRNA
encodes a d i f f e r e n t p o ly p e p tid e , th e co d in g r e g io n fo r
which l i e s o u ts id e th a t fo r Vmw IE 136* (143 )
(J . M clauchlan and J .B . C lem ents, p e r so n a l com m unication).
Below each mRNA i s in d ic a te d th e s i z e in kb o f the
p o ly a d en y la ted mRNA determ ined by m ethylm ercurie hydroxide
agarose g e l e le c tr o p h o r e s is (Watson e t a l . , 1979;
T alley-B row n and M i l l e t t e , 1979; Anderson e t a l . , 1980a and b ) .
In v i t r o t r a n s la t io n o f i s o la t e d mRNAs perform ed in
the s tu d ie s m entioned above, and th e mapping o f
p o ly p e p tid e s u t i l i s i n g HSV-l/HSV-2 in t e r t y p ic recom binants
(P resto n e t a l . , 1978; Marsden e t a l . , 1978; Morse e t a l . ,
1 9 7 8 ), have a llo w ed i d e n t i f i c a t io n o f the p o ly p e p tid e
encoded by each IE mRNA. At th e f o o t o f th e F igure th e
e q u iv a le n t p o ly p e p tid e s in the nom enclature o f H oness
and Roizman (1973 and 1974) are g iv e n .

CO CM(O CM
" ICO N-
cm (D CM
i

Figure A1.5
Map lo c a t io n s o f HSV-induced p o ly p e p tid e s
(Marsden e t a l , , 1978; P resto n e t a l . , 1978; H .S . Marsden,
p e r so n a l com m u nication ).
Each number (X) in d ic a t e s the apparent m.wt.
(x 10"^ ); f u l l nom enclature i s V X o r , in th e c a se o f
im m ediate e a r ly (IE ) p o ly p e p t id e s , Vmw IE X. P o ly p e p tid e s
w ith th e s u b s c r ip t "1" have no r e a d ily r e c o g n isa b le type 2
e q u iv a le n t , and th o se w ith th e su b s c r ip t 112 ” have no
r e a d ily r e c o g n is a b le type 1 e q u iv a le n t . Other p o ly p e p tid e s
are in d ic a te d by the m .wt. o f the HSV-1 form and have, no
s u b s c r ip t . A ccurate lo c a t io n s o f IE p o ly p e p tid e s from
t r a n s c r ip t mapping d a ta are shown in F igu re A 1 .4 .
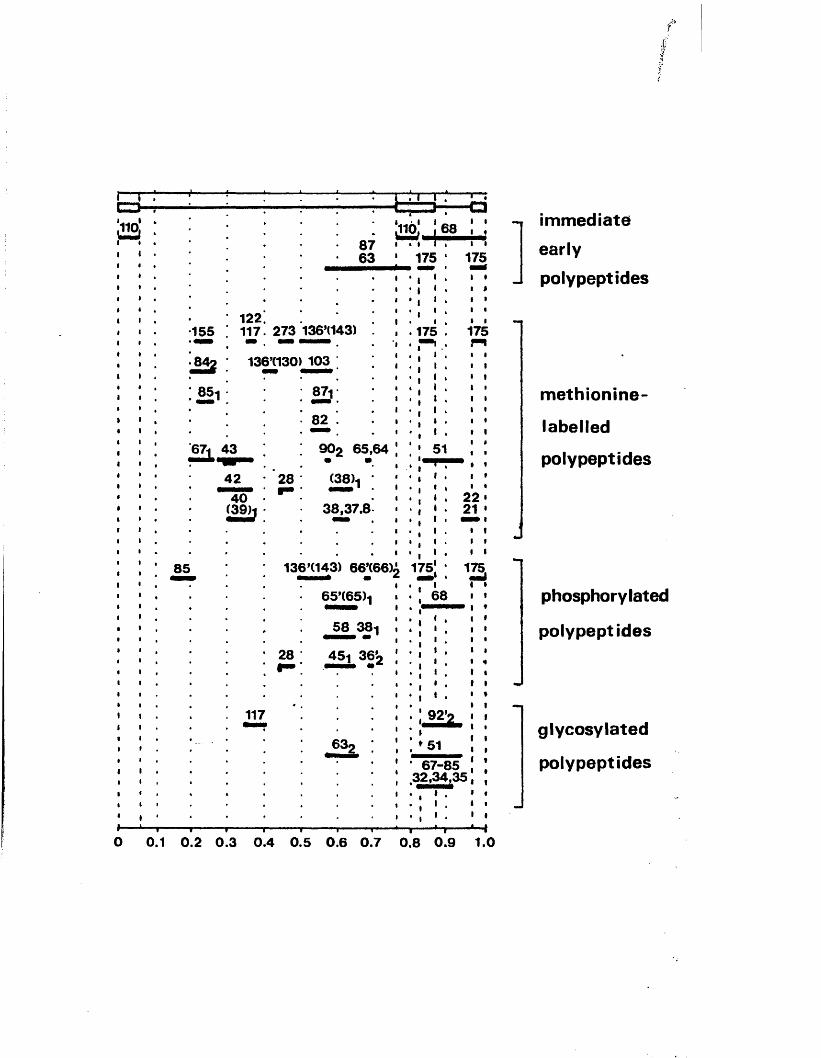
immediateearlypolypeptides
methionine-
labelled
polypeptides
phosphorylated
polypeptides
glycosylated
polypeptides
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.0
273 136’(143)
136T130) 103
90o 65,6467i 43
38,37.8
136’(143) 66’(66)
65’(65)<i
58 381
45-j 36o
67-8532,34,35

P A l l T B
M A T E R I A L S
A N D
M E T H O D S
i

- 38 -
MATERIALS
Vir u s e s
HSV-1 Glasgow s t r a in 17 syn* was i s o la t e d in Glasgow by' *4* ^])r C .A .P. R oss, and m utants syn tsB and syn tsD were in d ir e c t ly
•, - ^ - - - 4,
d er iv ed therefrom (Brown et; a l . , 1 9 7 3 ) .
HSV-1 s t r a in USA-8 syn* was i s o la t e d in P h ila d e lp h ia from
the l e f t tr ig e m in a l g a n g lio n o f a fem ale who d ied from
lym ph ocytic lymphoma (L onsdale e t a l . , 1 9 7 9 ) .
HSV-2 s t r a in HG52 was i s o la t e d from an a n a l l e s io n o f a
fem ale p a t ie n t in Edinburgh, and m utants t s l and t s 6 were
d er iv ed from t h i s s t r a in (Timbury, 1 9 7 1 ) .
liSV-l/HSV-2 in t e r t y p ic recom binant v ir u s e s are d e sc r ib ed
in F igure C 4 .3 .
The p seu d o ra b ies v ir u s s t r a in used was th a t o r ig in a l ly
i s o la t e d by Dr. A .S . Kaplan (Kaplan and V a tte r , 1 9 5 9 ) .
Ce l l s
The con tin u ou s c e l l l in e used was baby ham ster k idney
c lo n e 13 (BHK C13; MacPherson and S to k er , 1 9 6 2 ) .
B a c te r ia
E sc h e r ic h ia c o l i K12 s t r a in HB101 was used as h o s t
to p lasm id s (Boyer and R o iillan d -D u sso ix , 1 9 6 9 ) .
E sc h e r ic h ia c o l i met~"r~~m~" ( cI q^ S 7) used f o r the
growth o f b acter iop h age lambda was g iv en by Dr D. R i t c h ie .
B a c te r ia fo r the p rod u ction o f r e s t r i c t i o n en d o n u clea ses
are d e sc r ib e d in Table B l . l .
T issu e c u ltu r e media and s o lu t io n s
C e l ls were grown in th e Glasgow m o d if ic a t io n o f E a g le ’ s
medium (Busby e t a l . , 1964) supplem ented w ith 100 u n it s /m l

- 39 -p e n i c i l l i n , 1 0 0 /Mg/ml strep to m y cin , 0 . 2 yMg/ml a n tim y co tic
a g en t n -b u ty l-p -h y d ro x y b en zo a te , and 0 . 0 0 2 $ w t /v o l phenol r e d .
C a lf serum and p oo led human serum were prepared in the
I n s t i t u t e o f V ir o lo g y .
The fo l lo w in g com p osite s o lu t io n s were u sed .
E a g le ’ s medium c o n ta in in g x $ v o l /v o l c a l f serum;* _
ECx c o n ta in in g 10$ v o l /v o l D ifco tr y p to se phosphate
broth (E P );
E a g le 's medium c o n ta in in g s.% v o l /v o l human serum;
E a g le ’ s medium la c k in g orth oph osp hate c o n ta in in g
1 $ v o l /v o l c a l f serum.
P hosphate b u ffered s a l in e (PBS) co n ta in ed 0 .1 7 M NaCl, 0*0034
M KC1, 0 .0 0 1 M Na^HPO ., 0 .0 0 2 M KH2 PO4 in d i s t i l l e d w ater,
pH7• 2 .
PBSCa was PBS c o n ta in in g 5$ v o l /v o l c a l f serum.4 ^
V ersene c o n s is te d o f 0 .0 0 6M EDTA in PBS c o n ta in in g 0 .0 0 1 5 $m 4
w t/v o l phenol r e d .
T r is s a l in e was 0 .1 4 M NaCl, 0 .0 3 M KC1, 0 .0 0 0 2 8 M NaH2 ^ 4 >
1 mg/ml d e x tr o s e , 0 .0 0 1 5 $ w t /v o l p h en ol red , 0 .0 2 5 M T ris-H C l,r
pH 7 .4 supplem ented w ith 100 u n its /m l p e n i c i l l i n and 100 /<g/m l
strep to m y cin .
T ryp sin -versen e com prised one volume o f 0 .2 5 $ w t /v o l D ifco* ... *
tr y p s in in T r is s a l in e p lu s fou r volum es o f v e r se n e .
Giemsa s t a in c o n s is te d o f 1 .5 $ v o l /v o l su sp en sion o f Giemsa
in g ly c e r o l , heated to 5 6 ° fo r 90-120 min and d ilu te d w ith
an equal volume o f m ethanol.
R ad iochem icals
The fo l lo w in g r a d io is o to p e s were o b ta in ed from The
R a d io c h e m ic a l C e n t r e , Amersham, B u ck s .
ECx
ETCx
EHux
EC l-Pi

- 4 0 -
d eo x y n u c leo sid e (<*->2P) tr ip h o sp h a te s (dATP, PB164; dCTP, PB165
dGTP, PB166; dTTP, PB167; each a t 3 1 4 .8 TBq/mmol);
ad en osin e ( y - 52P) tr ip h o sp h a te (PB10218; > 185 TBq/mmol);
^2P orthophosphate (PBS11).
C hem icals
The f o l lo w in g firm s su p p lied th e l i s t e d c h e m ic a ls .
Agarose (ty p e 1 ) , Trizma b a se , eth id ium brom ide, 5$ v o l /v o l
s t e r i l e bovine serum album in, p o ly v in y lp y r o ll id o n e , bovine
serum albumin (RIA g r a d e ), r ib o n u c le a s e s A and Tl f y e a s t
RNA and lysozym e - Sigma (London) Chem ical Company, K in gston -
upon-Thames, London.
N ,N ,N ’ ,N 1-te tr a m e th y le th y le n e d ia m in e (TEMED), h y d r o x y la p a t ite ,
ammonium p ersu lp h a te and B iorex 70 (100-200 mesh) - B iorad
L a b o r a to r ie s , Richmond, C a l i fo r n ia .
Dextran su lp h a te , F i c o l l 400 and Sephadex G50 P ine -
Pharm acia F ine C hem icals, U ppsala, Sweden.
U ltr o g e l AcA 34 - LKB, Bromma, Sweden.
D iam inoethyl c e l lu l o s e (DE52) -Whatman L td ., M aidstone, Kent.
B r a in -h e a r t in fu s io n - Oxoid L td ., B a s in g sto k e , H ants.
2 , 5 -d ip h e n y lo x a zo le (PPO), l - 4 - d i - ( 2 - ( 5 - p h e n y lo x a z o ly l ) )
benzene (POPOP), t r ic h lo r o a c e t ic a c id and b o r ic a c id -
K och-Light L a b o ra to r ie s , Colnbrook, B erk s.
D im ethyl su lp h a te (DMS) - A ld r ich Chem ical Co. L td .,
G illin gh am , D o rse t.

- 41 -
H ydrazine - Eastman-Kodak, R och ester , New York.
P ip e r id in e - Pluorochem L td ., G lossop , D erb y sh ire .
M ithramycin (" M ith racin ") - P f iz e r L td ., Sandwich, Kent.
A m p ic illin - Beecham R esearch L a b o r a to r ie s , B ren tfo rd ,
M id d lesex .
O ther c h e m ic a ls , where n ot s t a t e d , were o b ta in ed where
p o s s ib le in Analar grade from BDH C hem icals L td ., P o o le , D o r se t .
jdna
C a lf thymus DNA and salmon sperm DNA were o b ta in ed from
Sigma L td ., and 0X i?4 RP I DNA from N ew England B io la b s ,
B e v er ly , M a ssa c h u sse tts .
P r o fe s so r D. S h e r r a tt k in d ly prov id ed pAT153 DNA.
Human cy to m eg a lo v iru s DNA ( s t r a in AD169) was g iv en by
Dr M. V/e s t s tr a t e .
DNA was prepared and g e n e ro u sly su p p lie d by Dr J . M.
W halley from a p la q u e -p u r if ie d A u str a lia n i s o l a t e (HVS 25)
o f equid h e r p e sv ir u s type I ob ta in ed from Dr II. Sabine o f
th e Department o f V eter in a ry M ed icin e, U n iv e r s ity o f Sydney.
Enzymes
R e s t r ic t io n en d o n u c lea ses n o t prepared from b a c te r ia
in t h i s s tu d y , and o th e r enzymes a c t in g upon DNA (T4 DNA
l i g a s e , b a c t e r ia l a lk a l in e p h osp h atase , E, c o l i DNA
polym erase I and th e Klenow fragm ent th e r e o f ) were purchased
from New England B io la b s , or from B ethesda R esearch
L a b o r a to r ie s , R o c k v il le , Maryland, or from th e B oehringer
C orporation (London) L td ., Lewes, E ast S u sse x .
D eoxyrib on u clease I was ob ta in ed from the W orthington
B ioch em ica l C orp oration , F reeh old , New J e r s e y .

- 42 -
T4 p o ly n u c le o t id e k in a se was su p p lie d by P-L /
B io ch em ica ls I n c . , M ilwaukee, Y /isconsin .
R e s t r ic t io n en d on u cleases Xhol and Smal were th e
generou s g i f t s o f Dr J . Arrand, and S s t I o f Dr P . R igby.
Other m a te r ia ls
P e t r i d is h e s - Plow la b o r a t o r ie s , I r v in e , S c o tla n d .
K odirex X -ray f i lm , X-Omat H X -ray f i lm and 1X80
d e v e lo p er - Kodak L td ., London.
Amfix - May and Baker L td ., Dagenham.
T eflo n tape - th e M innesota M ining and M anufacturing
Company, USA.
N it r o c e l lu lo s e membrane f i l t e r in s h e e t (BA85) and
grid d ed d is c form s - S c h le ic h e r and S c h i i l l , D a sse l, West Germany.
Commonly used s o lu t io n s and b u ffe r s
SSC 0 .1 5 M NaCl, 0 .0 1 5 M sodium o i t r a t e , pH 7 -5 ;
NTE 0 .0 1 M T ris-H C l, 0 .1 M NaCl, 0 .0 0 1 M EDTA,
pH 7 . 4 }
s c i n t i l l a t i o n 5 g PPO and 0 .3 g POPOP per l i t r e o f to lu e n e , f l u i d

METHODS
Growth o f c e l l s
BHK c e l l s were grown to co n flu e n c e in 80 oz r o l l e r
b o t t l e s a t 37° i a 200 ml ETC10 under 5$ 002 in a i r . C e l ls*
were h a rv ested by w ashing tw ic e w ith tr y p s in -v e r s e n e and
reseed ed a t 3 x 10^ c e l l s / r o l l e r b o t t le in ETC10. 50 mm- 6
d iam eter p l a s t i c P e t r i d is h e s were seeded a t 2 x 10
c e l l s / d i s h in 4 ml ETC10, th e m onolayers a t t a in in g c o n flu en ce
a f t e r 1 6 -2 4 t o a t 37°•
P rod u ction o f v ir u s s to c k s
80 oz r o l l e r b o t t le s c o n ta in in g a lm ost c o n f lu e n t BHK
c e l l s were in f e c t e d w ith v ir u s a t a m .o . i . o f 1 p . f . u . /
300 c e l l s . V irus was added in 40 ml EC5 per b o t t le and
in cu b a ted a t 3 1 ° . A fte r 2 -4 days in f e c t e d c e l l s were
3haken in to the medium, p e l le t e d a t 2000 r .p .ra . f o r 10 min
in an MSB C oo lsp in c e n tr i fu g e , and resuspended in 5 ml
PBSCa. The c e l l s were ruptured U3ing a C ole-P alm er
u lt r a s o n ic bath and c e n tr ifu g e d a t 2000 r .p .m . fo r 10 min
to remove c e l lu l a r d e b r is . The su p ern atan t was s to r e d in
s t e r i l e v i a l s a t - 7 0 ° . S t e r i l i t y ch eck s were c a r r ie d out
u s in g blood agar p la t e s and tr y p to se phosphate b ro th .
V irus t i t r a t i o n
V irus was d i lu t e d s e r i a l l y a t te n fo ld d i lu t io n s in
PBSCa. Drained BHK c e l l m onolayers i n 50 mm P e t r i d is h e s
were in o c u la te d w ith 0 .1 ml o f v ir u s d i lu t io n . V irus was
absorbed f o r 40 min a t th e tem perature o f subsequent
in c u b a tio n (3 1 ° or 3 7 °) and th en 4 ml EHu5 was added.
A fte r in c u b a tio n f o r 2 -3 days th e m onolayers were f ix e d and
s ta in e d w ith Giemsa s t a in f o r 15 min a t room tem p erature,
and p la q u es counted u s in g a d i s s e c t in g m icroscop e .

- 44 -
P laque p u r i f i c a t io n o f HSY
The medium (EHu5) was removed from P e tr i d is h e s
c o n ta in in g w e ll- s e p a r a te d in d iv id u a l p la q u es and th e
m onolayers were washed tw ic e w ith PBSCa. I n fe c te d c e l l s
form ing a s in g le p laque were taken up in to a f i n e l y drawn
P a steu r p ip e t t e and tr a n s fe r r e d to 0*5 ml PBSCa in a
s t e r i l e v i a l . Removal o f th e p laque from th e d is h was
c a r r ie d o u t under the dark f i e l d i l lu m in a t io n o f a Wild
M7A s te r e o s c o p ic d i s s e c t in g m icro sco p e . The v i a l was
so n ic a te d in an u lt r a s o n ic bath and r e le a s e d v ir u s
in o c u la te d in t e n fo ld d i lu t io n s onto BHK m onolayers in
P e t r i d is h e s , and p la q u es a llo w ed to d ev e lo p under 4 ml
EHu5. V irus was p laq ue p u r if ie d tw ic e more in th e same
way, and th e progeny from th e f i n a l p laque were used to
i n f e c t a s in g le 50 mm P e tr i d ish m onolayer. V irus from the
in f e c t e d m onolayer in cu b ated under 4 ml EC5 was t i t r a t e d
under a p p ro p r ia te c o n d it io n s and in some c a s e s used to
i n f e c t 80 oz r o l l e r b o t t l e s f o r p ro d u ctio n o f working
s to c k s o f v i r u s .
G en eration o f HSV-l/HSV-2 in t e r t y p ic recom binant v ir u s e s
The marker r e sc u e method o f Stow and W ilk ie (1976) was
u sed , in v o lv in g p r e c ip i t a t io n o f th e i n f e c t in g DNA a s a
ca lc iu m phosphate complex and osm otic sh ock in g o f th e c e l l s
by treatm en t w ith 25# d im eth y lsu lp h o x id e (DMSO) to s t im u la te
uptake o f th e DNA.1
I n ta c t DNA from a te m p e r a tu r e -s e n s it iv e m utant o f
HSV was m ixed ’a t 0 .4yu g /m l w ith 10 /Vg/ml c a l f thymus c a r r ie r
DNA and w ild type DNA r e s t r i c t i o n fragm ents i n H epes-
b u ffered s a l in e (HeBS: 8 .0 g /1 NaCl, 0 .3 7 g /1 KC1, 0 .1 g /1
Na2 HP04 , 1 .0 g /1 I - g lu c o s e , 5 .0 g /1 N -2 -h y d ro x y eth y lp ip era z in e

- 45 -N1-2 -e th a n e su lp h o n ic a c id , pH a d ju sted to 7 .0 5 w ith NaOH).
The amount o f fragm ent p resen t per ml was th a t gen era ted
from 1 Ag i n t a c t w ild type DNA. 2 M CaClg was added to a
f i n a l c o n c e n tr a t io n o f 0 .1 3 M, and 0 .5 ml sam ples added to
a lm ost c o n f lu e n t dra in ed BHK c e l l m on o layers. The d is h e s
were in cu b a ted a t 3 8 .5 ° from the tim e o f in f e c t i o n , 4 ml
EG5 b e in g added a f t e r 45 m in. At 4 hr p . i . th e c e l l s were
washed w ith E a g le 1s medium and exposed to 1 ml 25# v o l /v o l DMSO* ■■■» ^
i n HeBS fo r 5 min a t room tem p eratu re. The c e l l s were
c a r e f u l ly washed tw ic e w ith E a g le ’ s medium and in cu b a ted a t_ r
3 8 .5 in 4 ml EHu5 f o r 3 d a y s . Then th e m onolayers were
washed tw ic e w ith 4 ml PBSCa, 1 ml PBSCa added, and p la q u es
p ic k e d .
P rep a ra tio n o f HSV DNA
C on flu en t m onolayers o f BHK c e l l s i n 80 oz r o l l e r
b o t t l e s were in f e c t e d a s d e sc r ib ed and in cu b a ted a t 31°•
The in fe c te d c e l l s were detached by a g i t a t io n a f t e r 2 -3 days
and p e l le t e d by c e n tr ifu g a t io n a t 2000 r .p .m . fo r 10 m in.
The medium was r e ta in e d in th e c a se o f HSY-1. The c e l l s
were resuspended in RSB (0 .0 1 M T ris-H C l pH 7*4 , 0 .0 1 M
KC1, 0 .0 0 1 5 M MgCl2 ) c o n ta in in g 0 .5 # v o l /v o l N on id et P40• * _ •
(NP40) and k ep t on i c e fo r 10 m in. N u c le i were p e l l e t e d
by c e n tr i fu g a t io n a t 2000 r .p .m . fo r 5 min and th e
cy to p la sm ic su p ern atan t r e ta in e d . The n u c le i were r e
e x tr a c te d w ith 0 .5 # NP40 in RSB and the second c y to p la sm ic
su pern atant r e ta in e d . V irus was p e l le t e d by c e n tr ifu g a t io n
o f the combined su p ern a ta n ts a t 12000 r .p .m . fo r 3 hr i n an
MSE 6 x 250 ml r o to r .
The p e l l e t was resuspended in NTE, so n ic a te d in an
u ltr a s o n ic bath , 20# w t /v o l sodium d od ecy l su lp h a te (SDS)

- 46 -
added to a f i n a l c o n c e n tr a tio n o f 0 * 5 and e x tr a c te d $$ th
two volum es o f N T E -eq u ilib rated r e d i s t i l l e d phenol by g e n t le
a g i t a t io n on a r o p k -a n d -r o lle r . The upper aqueous phase
was removed and r e -e x tr a c te d w ith two fu r th e r volum es o f
N T E -eq u ilib rated phenol a f t e r c e n tr ifu g a t io n a t 2000 r .p .m .
fo r 10 m in. A fte r a second c e n tr ifu g a t io n s te p the upper
phase was e x tr a c te d f o r 10 min w ith isoam yl a lc o h o l in
chloroform (1 :2 4 v o l /v o l ) and a g a in c e n tr ifu g e d . The upper
aqueous phase c o n ta in in g HSV DNA was d ia ly s e d o v e rn ig h t a t
4° a g a in s t 0 .1 x SSC, and then sto r ed a t - 2 0 ° .
In some in s ta n c e s HSV DNA was p u r if ie d fu r th e r by3
iso p y k n ic banding on caesium c h lo r id e g r a d ie n ts . S o lid
CsCl was added to th e DNA s o lu t io n to g iv e a r e f r a c t iv e in d ex
o f 1 .4 0 1 -1 .4 0 2 . C e n tr ifu g a tio n was f o r 3 days a t 36000 r .p .m .
in a Beckman Ti50 r o to r a t 1 5 ° , or f o r 24 hr a t 40000 r .p .m .
in a S o r v a ll T7865B v e r t i c a l r o to r a t 15° • The g r a d ie n ts* •
were fr a c t io n a te d by p ie r c in g th e tube and c o l l e c t i n g d rop s.
5 / a sam ples o f each f r a c t io n were assayed by e le c tr o p h o r e s is
on a 1 $ ag a ro se g e l c o n ta in in g 0 . 5 yug/ml eth id ium bromi d e >
and n u c le ic a c id s v i s u a l i s e d under lo n g w avelen gth UV
r a d ia t io n (365 nm). F r a c tio n s c o n ta in in g HSV DNA, which
i s c le a r ly d is t in g u is h e d from degraded c e l lu l a r DNA and
RNA, were com bined, d ia ly s e d o v ern ig h t a g a in s t 0 .1 x SSC
a t 4 ° , and s to r e d a t - 2 0 ° .
HSV DNA s u i ta b le fo r n ic k t r a n s la t io n was prepared by
the method o f W ilk ie and C o r t in i (1976^. l in e a r CsCl
g r a d ie n ts o f r e f r a c t iv e in d ic e s 1 .3 8 (bottom ) to 1 .3 6 ( t o p ) ,
c o n ta in in g 20 yt*g/ml eth id ium bromide, were prepared .
0 .2 - 0 .3 ml o f p h e n o l-e x tr a c te d or iso p y k n ic banded HSV-1
DNA a t 100-500 ^Ag/ml was la y e r e d onto each g r a d ien t and th e

- 47 -
tu b es c e n tr ifu g e d a t 40000 r .p .m . fo r 110 min in a Beckman
SW50.1 r o to r . The band o f p u r if ie d v ir u s DNA was v i s u a l i s e d
u s in g a lo n g w avelen gth (365 nm) UV M in er a iig h t handlamp,
and removed v ia an 18-gauge sy r in g e n e e d le . The DNA
s o lu t io n was sa tu r a te d w ith CsCl and e x tr a c te d tw ic e w ith
aqueous p r o p a n -2 -o l sa tu ra te d w ith CsCl to remove eth id ium
brom ide. The low er aqueous phase was d ia ly s e d a g a in s t NTE
and p r e c ip i ta t e d w ith two volum es o f e th a n o l a t - 2 0 ° . The
DNA p e l l e t was r e d is s o lv e d in a sm all volume o f 0 .1 x SSC
and sto r e d a t -2 0 .
P rep a ra tio n o f PRV DNA
BHK c e l l m onolayers in 80 oz r o l l e r b o t t l e s were
in f e c t e d w ith PRV a t a m .o . i . o f 1 in 300 and in cu b a ted a t
37° u n t i l com p lete c . p . e . was ob served . V iru s was h a rv ested
by th e method o f B en-Porat e t a l . (1 9 7 4 ) . The in f e c t e d c e l l
medium was c e n tr ifu g e d a t 5000 r .p .m . fo r 10 min in a
S o r v a ll GSA r o to r , and v ir u s in th e su p ern atan t was
sedim ented onto a 30# w t /v o l su cro se ‘’cu sh io n 11 by c e n tr ifu g a t io n
a t 13500 r .p .m . fo r 1 hr in a S o r v a ll AH627 r o to r . The
v ir u s was la y e r e d onto a l in e a r su cro se g r a d ie n t
(1 5 -3 0 # w t /v o l su cro se in TBSA: 0 .0 1 M T ris-H C l pH 7 .5i - . . .
c o n ta in in g 8 g /1 NaCl, 0 .2 g /1 KC1, 0 .1 g /1 MgC^^HgO,
0 .1 g /1 C a C ^ .i^ O and 10 g /1 bovine serum album in) and
c e n tr ifu g e d in a S o r v a ll AH627 r o to r a t 10000 r .p .m . fo r
140 m in. The v i s i b l e v ir u s band was removed by p ie r c in g th e
tube w ith an 18-gauge sy r in g e n e e d le , d i lu te d s ix f o ld w ith
TBSA and c e n tr ifu g e d a t 13500 r .p .m . fo r 1 hr in a S o r v a ll
AII650 r o to r . The v ir u s p e l l e t was resuspended in 5-10 ml
NTE by s o n ic a t io n , and DNA prepared by phenol e x tr a c t io n
a3 d e sc r ib ed f o r HSV.

P rep a ra tio n o f b acter iop h age lambda DNA
E sc h e r ic h ia c o l i m e t 'r 'n f ( cI q^ S 7) c o n ta in s a lambda
prophage which i s h ea t in d u c ib le and l y s i s d e f i c i e n t . A
s t a r t e r c u ltu r e in TP broth was grown a t 31° and in o c u la te d
in to a u to c la v ed H -broth (5 g /1 D ifco b actop ep ton e, 1 g /1
g lu c o s e , 5 g /1 NaCI, 8 g ? l D ifco n u tr ie n t b ro th , pH 7«2)
a t a d i lu t io n o f 1 :5 0 . The b a c te r ia were grown a t 31° in
a Gallenkamp o r b i t a l in c u b a to r . At a c e l l d e n s ity o fQ
2 -4 x 10 per ml th e c u ltu r e tem perature was in c r e a se d to
42° by th e a d d it io n o f b o i l in g H -broth, and fu r th e r
in c u b a tio n was a t 42° fo r 10 m in. The c u ltu r e temperature*
was then reduced to 37° fo r 1 -2 h r . The b a c te r ia were
c e n tr ifu g e d a t 8000 r .p .m . fo r 2 min in an MSE 6 x 250 ml
r o to r and resuspended in 20 -50 ml T2 b u ffe r (0 .0 5 M Na2HP04 ,
0 .022 M KH2P04 , 0 .0 8 5 M NaCI, 0 .0 0 1 M MgS04 ,~ 0 .0 0 0 1 M CaCl2 ,
0 .0 0 1 # w t /v o l g e l a t i n ) . Chloroform was added to a f i n a l
c o n c e n tr a tio n o f 1 # v o l /v o l to ly s e th e c e l l s , then DNase
to 2 mg/ml. To each 10 ml o f su sp en sio n 0 .7 1 g p o ly e th y le n e
g ly c o l 6000, 0 .1 9 2 g NaCl and 0 .0 2 3 g d ex tran su lp h a te were
added w ith s t i r r i n g , and t h i s was l e f t o v e rn ig h t a t 4° to
a llo w the b acter iop h age to p r e c ip i t a t e . The m a jo r ity o f th e
upper phase was removed*and th e rem ainder c e n tr ifu g e d a t
2000 r .p .m . f o r 5 min. The m a ter ia l a t th e in t e r f a c e was
removed and resuspended in about 2 ml o f 1 # dextran su lp h a te ,
adding 0 .1 5 ml 3 M KC1 per ml o f su sp en s io n . A fte r 2 hr a t
4° th e su sp en sio n was c e n tr ifu g e d a t 2000 r .p .m . fo r 5 min,
and th e su p ern atan t c o n ta in in g th e v ir u s removed. The v ir u s
was p u r if ie d by c e n tr ifu g a t io n on a d is c o n tin u o u s caesium
c h lo r id e g r a d ie n t ( s p e c i f i c d e n s i t i e s o f 1 .7 , 1*5 and 1 .3
g/m l o f CsCl in 0 .0 0 5 M Tris-H Cl pH 7 .2 , 0 .0 0 5 M MgS04 ) a t
35000 r .p .m , fo r 1 hr in a Beckman SW50.1 r o to r . The band

- 49 -
o f v ir u s , in th e phase o f s p e c i f i c d e n s ity 1.5> was d ia ly s e d
a g a in s t 0 .5 x T2 b u ffe r . B acteriop hage DNA was prepared tjy
ph en ol e x tr a c t io n a s d escr ib ed fo r HSV.
E stim a tio n o f DMA c o n c e n tr a t io n s ^
Approximate e s t im a t io n s o f DNA c o n c e n tr a t io n s were
o b ta in ed by v i s u a l com parison o f sam ples w ith known amounts
o f DNA on e th id iu m -sta in e d agarose g e l s under UV i l lu m in a t io n
(365 nm). DNA c o n c e n tr a t io n s o f RNA-free sam ples were more- .rt
a c c u r a te ly e stim a te d by o p t ic a l d e n s ity measurement a t
260 nm, assum ing th a t an o p t ic a l d e n s ity o f 1 .0 corresp ond s
to a DNA c o n c e n tr a t io n o f 40 yug/m l. DNA c o n c e n tr a t io n s o f
sam ples a ls o c o n ta in in g RNA were e stim a te d by th e method
o f H i l l and Whatley (1975)* A s o lu t io n o f m ithram ycin a t
200 yvg/ml was made in 0 .3 M M gC^. 15 yul was mixed w ith
0 .3 ml DNA s o lu t io n and f lu o r e s c e n c e measured a t 550 nm upon
e x c i t a t io n a t 440 nm. DNA o f known c o n c e n tr a t io n , e i t h e r
lambda or c a l f thymus DNA, was used to c o n s tr u c t a standard
curve in th e range 0 -10 /4g/m l.
Pu r i f i c a t io n o f r e s t r i c t i o n en d o n u clea ses
S e v e r a l o f th e r e s t r i c t i o n en d o n u clea ses used in th e
i n i t i a l s ta g e s o f t h i s work were prepared from b a c te r ia l
c e l l p a s te s (T able 3 1 .1 ) . B r a in -h e a rt in f u s io n (BHI) a t
37 g /1 was used f o r bulk p r e p a r a tio n s , which were seeded a t
a d i lu t io n o f 1 :5 0 or 1:100 and grown in a Gallenkamp
o r b i t a l in cu b a to r a t 200-300 r .p .m . B a c te r ia were p e l le t e d
a t e a r ly s ta t io n a r y phase by c e n tr i fu g a t io n in a 6 x 250 ml
MSE r o to r a t 8000 r .p .m . fo r 2 min. They were resuspended
in a sm all volume o f BHI and fr o z e n a t - 2 0 ° .
10-20 g o f c e l l p a s te was hom ogenised in a S e r v a ll
Omnimixer blade hom ogeniser w ith an equal volume o f a c id -

- 50 -
washed g la s s beads (150-200 yum). H om ogenisation was perform ed
on i c e fo r 6 x 2 min p e r io d s a t 10000 r .p .m . w ith 2 min
in t e r v a l s , and th e e x te n t o f hom ogen isation was checked by
m icro sco p ic exam in ation . The g la s s beads were p e l le t e d by
c e n tr i fu g a t io n a t 1000 r .p .m . fo r 5 min, and the su p ern atant
c e n tr ifu g e d a t 40u0Q r .p .ra . fo r 1 hr in a Beckman SW40Ti r o to r
The su p ern atan t was made 1 M in NaCI, 10?° v o l /v o l in g ly c e r o l4 _
and im m ed iate ly su b jec ted to column chrom atography.
The fo l lo w in g column chrom atographic' p roced u res were
em ployed.
a) M olecu lar s i e v e . P r o te in s were sep a ra ted a cco rd in g to
m olecu lar w eight by p a ss in g th e c e l l e x tr a c t through a 2 .5 x
100 era column o f U ltr o g e l AcA34 a t 4 ° . The e lu t in g b u ffe r
(CB) was 0 .02 M T ris-H C l pH 7 .5 , 0 .0 0 1 M EDTA, 0 .0 1 M
2 -m erca p to eth a n o l, 10^ v o l /v o l g ly c e r o l , and an LKB f r a c t io n
c o l l e c t o r was used w ith a flo w r a te o f 2 0 -50 m l/h r . 5 yul
sam ples o f a l t e r n a te 6 ml f r a c t io n s were a ssa y ed fo r n u c le a se
a c t i v i t y as d e sc r ib ed below, and f r a c t io n s c o n ta in in g
r e s t r i c t i o n en d on uclease a c t i v i t y were p o o led and d ia ly s e d
a g a in s t CB.
b) Ion exchange. D iam inoethyl c e l l u l o s e (DE52) or B iorexA 1
70 colum ns o f 5 -20 ml bed volume were poured i n 20 ml sy r in g e
b a r r e ls and e q u il ib r a te d w ith CB. D ia ly se d U ltr o g e l f r a c t io n s
were a p p lie d , washed w ith CB, and e lu te d w ith KC1 g r a d ie n ts•»
in CB. G rad ients o f 0 - 0 .6 M KC1 were r o u t in e ly u sed .
A gain , 5 / * a l iq u o t s o f a lt e r n a te 3 ml f r a c t io n s were a ssayed
fo r n u c le a se a c t i v i t y , and f r a c t io n s c o n ta in in g r e s t r i c t i o n
en d on u clease a c t i v i t y were d ia ly se d a g a in s t CB. Both ty p e s
o f io n exchange column were used f o r m ost en d o n u clea ses
(T able H I.2 ) . The enzymes were co n cen tra ted by a p p lic a t io n .

- 51 -
to sm all B iorex columns (2 -5 m l) , e lu t io n w ith 10 ml o f
1 M KC1 in CB, and d i a l y s i s a g a in s t 50$ v o l /v o l g ly c e r o l in * *
CB. 1 $ w t /v o l s t e r i l e bovine serum album in was added to *
th e enzyme to a f i n a l c o n c e n tr a tio n o f 50 ywg/ml, and s to r a g e
was a t - 2 0 ° .
R e s t r i c t i o n e n d o n u c le a se d i g e s t i o n
A ll r e s t r i c t i o n d ig e s t s were c a r r ie d out in 0 .0 0 6 M
T ris-H C l pH '7.5, 0 .0 0 6 M MgC^, 0 .0 0 6 M 2 -m ercap toeth an o l,
0 .0 2 M KC1, and 0 .0 2 or 0 .1 $ w t /v o l s t e r i l e bovine serum , *• —
album in. R e s t r ic t io n s i t e s are g iv e n in T able EL.4 .
In the p rep a ra tio n o f r e s t r i c t i o n e n d o n u c lea ses , 5 jjOl
a l iq u o t s o f a lt e r n a te f r a c t io n s were in cu b ated a t 37° fo r
3 hr w ith 0 .5 yug lambda DNA in 50 yul r e s t r i c t i o n b u ffe r .
P u r if ie d en d on u cleases were t i t r a t e d a g a in s t 1 yug lambda
DNA in 5 0 1 fo r 3 hr a t 3 7 ° , in order to determ ine th e
amount o f enzyme req u ired fo r com plete d ig e s t io n o f th e DNA.
DNA, e it h e r u n la b e lle d or n ic k t r a n s la t e d , was in cu b ated
a t 10-100 yug/ml fo r 1 -3 hr a t 37° w ith th e req u ired amount
o f enzyme. TagI. BstNI and B e ll d ig e s t io n s were done a t
65°• 0 .1 5 yug lambda DNA was added to r e s t r i c t i o n d ig e s t s
c o n ta in in g sm all amounts o f in v iv o v P - la b e l le d HSV DNA
or r e s t r i c t i o n fragm ents i s o la t e d from n ic k tr a n s la te d HSV
DNA.
R e s tr ic t io n d ig e s t s were term in ated by th e a d d it io n
o f 10 yul o f d y e - P ic o l l , which co n ta in ed 10$ w t /v o l F ic o l l
400, 0 .1 M EDTA and 0 .1 $ w t /v o l bromophenol b lue in 5 x E
b u f f e r ( s e e b e lo w ) .
Agarose g e l e l e c t r o p h o r e s i s
Agarose g e l c o n c e n tr a t io n s between 0 .4 and 2 .0 $ w t /v o l
were em ployed, in E b u ffer (0 .0 3 6 M Trizma, 0 ,0 3 6 M Nai^PO^,

- 52 -
0 .001 M EDTA, pH 7 .8 ) c o n ta in in g 0 .5 yUg/ml eth id ium brom ide.
The agarose was d is s o lv e d by b o i l in g in a Tappan microwave
oven and c o o le d to about 50° b efore p o u rin g . H o rizo n ta l
g e l s were r o u t in e ly u sed , and DHA sam ples w ith added d y e -
P ic o l l were su b je c te d to e le c tr o p h o r e s is a t 2 V/cm fo r
1 6 -2 0 hr a t room tem perature in E b u ffe r c o n ta in in g 0 .5 1
eth id ium brom ide. G els were photographed under lo n g w avelen gth
(365 nm) UV ir r a d ia t io n u s in g P o la r o id ty p e 105 or ty p e 107C
f i lm . G els fo r autorad iograph y were d r ie d on p la in g la s s
p la t e s in an oven a t 6 0 ° .
P o lyacry lam id e g e l e le c tr o p h o r e s is
A s to c k s o lu t io n o f 29 > w t /v o l acry lam id e and 1 # w t /v o lm — <
N N '-m eth y len eb isacry lam id e in w ater was d i lu te d to th e
req u ired c o n c e n tr a tio n w ith th e a d d it io n o f 10 x THE to a
f i n a l c o n c e n tr a tio n o f 1 x TBE (0 .0 8 9 M Trizma, 0 .0 8 9 M b o r ic
a c id , 0 .0 0 2 5 M EDTA) and 10?* w t/v o l ammonium p ersu lp h a te toT ■ . *
0 . 0 6 7 6 0 ml o f t h i s s o lu t io n was d egassed u s in g a vacuum
pump and 30 yul N ,N ,N *,N f -te tr a m e th y le th y le n e d ia m in e (TEMED)
added, and th e g e l poured (265 mm x 165 mm x 1 .5 mm).
E le c tr o p h o r e s is was c a r r ie d ou t in 1 x TBE a t 3 V/cm fo r
10-20 hr a t room tem p erature, and th e g e l s were d r ied on
Whatman 3MM paper under vacuum in a B iorad h eated g e l d r ie r .
G eneral l a b e l l i n g o f DNA w ith ^2P
N ick t r a n s la t io n ’o f DNA i n , v i t r o was done e s s e n t i a l ly
a cco rd in g to th e method o f Rigby e t a l . (1 9 7 7 ) . 180 pm oles
o f each o f * - 52P dATP, dGTP, dCTP and dTTP (350 Ci/mmole), . . ^ —•
were added to a tube and d r ied in an Edwards l y o p h i l i s e r .
1 yug DNA in 100 yul n ic k t r a n s la t io n b u ffe r (0 .0 5 M Tris-H C l
pH 7 .8 , 0 .0 0 5 M MgC^, 58 yug/ml bovine serum album in, 0 .0 0 5 M

- 53 -
2-m ercap toeth an o l) was added to th e tube on i c e . 2*5 u n i t s
DNA polym erase I were added and tube in cu b ated a t 1 4 -1 5 °
f o r 90 m in. DNA p o s s e s s in g few er n ic k s than HSV DNA, suoh
a s th a t from b acter iop h age lambda and $X174, or piasm id DNA,— 3 m m /L '
was in cu b ated w ith th e a d d it io n o f DNase to 10 J or 10 m g/m l.
Iso to p e in c o r p o r a tio n was m onitored by s p o t t in g 2 yul sam ples
onto f i l t e r d i s c s and p r e c ip i t a t in g th e DNA w ith 5# w t /v o li *
t r i c h lo r a c e t i c a c id f o r 15 min on i c a . A fte r w ashing in
e th a n o l then a ce to n e th e d i s c s were d r ied and counted f o r
^2P in s c i n t i l l a n t . T h is method produced n ic k tr a n s la te d' 8DNA o f s p e c i f i c a c t i v i t y 1 -2 x 10 c .p .m ./iu g , maximal
in c o r p o r a tio n (38-70/*) b e in g reached a t 90 m in. The n ic k
t r a n s la t io n r e a c t io n was term in ated e i t h e r by adding EDTA
to a f i n a l c o n c e n tr a tio n o f 0 .0 1 M fo llo w e d by h e a t in g 1a t
65° fo r 10 min, or by phenol e x tr a c t io n . L a b e lled DNA was
sep arated from u n in corp orated d eo x y n u c leo sid e tr ip h o sp h a te s
by e lu t io n through a 15 cm column o f Sephadex G50 in NTE.32HSV DNA was P - la b e l le d in v iv o a s d e sc r ib ed by P resto n
n ' '■ ' ' - ~ 6e t a l . (1 9 7 8 ). 50 mm P e tr i d is h e s were seed ed w ith 2 x 10
c e l l s / p l a t e in E C l-P i and in cu b a ted a t 37* o v e r n ig h t . V irus
was added in 0 .2 ml E C l-P i a t a m .o . i . o f 5 p . f . u . / c e l l ,
in cu b ated a t 31° fo r 1 h r , th e m onolayers washed tw ic e w ith
E C l-P i, and 2 ml E C l-P i added. A fte r a fu r th e r 2 -3 hr a t
3 1 ° , 0 .2 5 - 0 .5 mCi o f c a r r ie r - f r e e ^2P -orth op h osp h ate was
added in 0 .5 ml E C l-P i, and the p la t e s were in cu b a ted a t 31°
f o r 2—3 d a y s . The in f e c t e d c e l l s were removed w ith a rubber
policem an or bent P a steu r p ip e t t e and c e n tr ifu g e d a t 2000
r .p .m . fo r 5 m in. Each c e l l p e l l e t was e x tr a c te d once w ith
5 ml 0*5/^ NP40 in RSB. The cy to p la sm ic su p ern atan t was4
combined w ith th e medium and c e n tr ifu g e d a t 3 5 0 0 0 r .p .m .
fo r 1 hr a t 4° in an Oakridge tube in a Beckman T i50 r o to r .

- 54 -The v ir u s p e l l e t was resuspended by s o n ic a t io n in 0 .5 ml
NTE, th en 50 yul 25?* w t /v o l sodium d od ecy l su lp h a te and
0 .5 ml N T E -saturated phenol were added to th e v ir u s su sp en sion
in a 1 .5 ml Eppendorf capped p l a s t i c tu b e . A fte r g e n t le
a g i t a t io n th e tube was c e n tr ifu g e d fo r 5 min a t 2000 r .p .m .,
and th e upper phase removed and e x tr a c te d w ith 0 .5 ml
isoam yl a lc o h o l in ch loroform ( 1 : 2 4 ) . B o iled RNase A and T1
were added to th e removed upper phase to f i n a l c o n c e n tr a t io n s
o f 25 yug/ml and 50 U/ml r e s p e c t iv e ly , and th e DNA d ia ly s e d
o v e r n ig h t a g a in s t 0 .1 x SSC a t room tem p eratu re. 10 yul
sam ples were s c i n t i l l a t i o n counted i n 2 ml w ater in order
to e q u a lis e co u n ts b efore r e s t r i c t i o n and g e l a n a ly s is .
I s o la t i o n o f r e s t r i c t i o n fragm ents
The method o f Wil'kie and C o r tin i (1976) was used to
i s o l a t e r e s t r i c t i o n fragm ents from agarose g e l s . U n la b e lled
fragm ents were lo c a te d by s ta in in g e i t h e r th e whole
h o r iz o n ta l g e l or a s t r ip from each edge w ith 0 . 5 yug/ml
eth id ium bromide in E b u ffe r , and n ic k tr a n s la te d fragm ents
were e x c is e d from agarose tube g e l s ( 1 .5 x 18 cm) which had
been e le c tr o p h o r e se d in th e p resen ce o f 0 .5 >ug/ml eth idium
brom ide. Agarose fragm ents were d is s o lv e d i n 2 -10 ml 5 M
sodium p e r c h lo r a te a t 60° and a p p lied to h y d ro x y la p a tite
columns o f bed volum es 0 .2 - 1 .0 ml in sy r in g e b a r r e ls .
A p p lic a tio n and e lu t io n o f th e DNA were done a t 6 0 ° . The
colum ns were washed w ith 5 x 1 ml 5 M sodium p e r c h lo r a te and
then w ith 2 x 1 ml 0 .1 4 M sodium phosphate pH 6 .8 . Double
strand ed DNA was e lu te d w ith 4 x 0 .5 ml 0 .4 M sodium
phosphate pH 6 .8 , and d ia ly s e d o v e rn ig h t a g a in s t 0 .1 x SSC.
R a d io a c tiv e sam ples were s c i n t i l l a t i o n counted f o r32Cerenkov r a d ia t io n from P.

- 55 -
T ran sfer o f DNA fragm ents to n i t r o c e l lu lo s e
The b lo t t in g tech n iq u e o f Southern (1975) in v o lv e s
th e im m o b ilisa t io n o f s in g le stranded DNA on a n i t r o c e l lu lo s e
f i l t e r . R e s tr ic te d HSV DNA was e le c t r o p h o r e t ic a l ly
sep ara ted on an agarose g e l (betw een 0 .4 and 1 .0 $ w t /v o l)
in a s in g le s l o t spanning th e g e l . The g e l was soaked f o r
1 hr in g e l soak I (0*2 M NaOH, 1 M NaCI) and th en f o r 1 hr
in g e l soak I I (1 M T ris-H C l pH 8 , 1 M N aC I). The g e l was
tr a n s fe r r e d to two s h e e ts o f Whatman 3MM paper on a g la s s
p la t e supported in a tr a y o f 2 1 o f 6 x SSC. A sh e e t o f
m oistened n i t r o c e l l u l o s e th en a sh e e t o f 3MM paper were
a p p lie d to th e g e l su r fa c e , fo llo w e d by a w eighted s ta c k o f
c u t paper to w e ls . T ran sfer o f th e s in g le strand ed DNA to
th e n i t r o c e l lu lo s e was a llow ed to occu r o v e rn ig h t a t room
tem p eratu re. The n i t r o c e l lu lo s e sh e e t was washed in 3 x SSC,
d r ied in a i r , then baked in a vacuum oven a t 70° fo r 2 h r .
I t was th en used e i t h e r fo r s t r ip o r c r o s s - b lo t h y b r id is a t io n .
B lo t h y b r id is a t io n
H y b r id isa tio n a t 75° in aqueous s o lu t io n (W ilk ie , 1976)
or a t 45° in 50>* v o l /v o l formamide (McConaughy e t a l . , 1969)
was u sed . N ick tr a n s la te d probe DNA was denatured by h e a tin g
a t 100° fo r 5 min in 0 .1 M NaOH, and th en n e u tr a l is e d on i c e
w ith 1 M HC1 a f t e r th e a d d it io n o f 1 M T ris-H C l pH 7*5 to
a c o n c e n tr a tio n o f 0 .0 5 M. 10^-10^ c .p .m . o f ^2P-DNA was
r o u t in e ly used per h y b r id is a t io n . 5 mm s t r i p s were cu t from
a n i t r o c e l lu lo s e b lo t and in cu b ated in a sh ak in g w ater bath
a t 75° fo r 1 hr in 15 ml p l a s t i c screw -top tu b es c o n ta in in g
5 ml o f 3 x SSC and D enhardt1s s o lu t io n (D enhardt, 1966;
0.02>* w t /v o l F ic o l l 400, 0 .0 2 $ w t/v o l p o ly v in y lp y r r o lid o n e ,* 4
0 .0 2 $ w t /v o l bovine serum alb u m in ). H y b r id isa t io n s c a r r ie d
out in 5 0 $ formamide in 3 x SSC and Denhardt*s s o lu t io n were

- 56 -
in cu b ated a t 45°• The tu b es were drained and th e denatured
probe added a f t e r b ein g made up to 5 ml i n 3 x SSC and
D enhardt1s s o lu t io n , or 50$ formamide in 3 x SSC and D enhardtf s
s o lu t io n . H y b r id isa tio n was a llow ed to occu r f o r 2 -4 days
a t 75° or 45° i n a shaking w ater bath , and th en th e s t r i p s
were washed w ith 2 x SSC a t 60° f o r a t l e a s t s i x hours w ith
se v e r a l changes o f s o lu t io n . The s t r i p s were d r ied in a i r ,
a p p lie d to a g la s s p la t e w ith d o u b le -s id e d S e l lo ta p e , baked
a t 70° fo r 30 min, and autorad iograp h ed .
The method o f c r o s s - b lo t h y b r id is a t io n , d evelop ed by
H utchison (S a to e t g A ., 1 9 7 7 ), in v o lv e s th e sim u ltan eou s
t r a n s fe r o f denatured n ic k tr a n s la te d DNA fragm ents from an
agarose g e l to a n i t r o c e l lu lo s e sh e e t c o n ta in in g im m ob ilised
u n la b e lle d DNA fra g m en ts . H y b r id isa tio n o ccu rs in s i t u ,i ■» ■ "
and homology between fragm ents i s d e te c te d a s s p o ts on an
a u torad iograp h . A square, n i t r o c e l lu lo s e b lo t was prepared
a s d e sc r ib ed and soaked fo r 1 hr a t 75° in 4 x SSC and
0 .1 $ w t /v o l sodium d od ecy l su lp h a te . 1 -2 /*g n ic k tr a n s la te d
HSV DNA was r e s t r i c t e d and e le c t r o p h o r e t ic a l ly sep a ra ted on
an agarose g e l o f e q u iv a le n t d im ensions to th e f i r s t g e l , and
then au torad iograp h ed . The g e l was tr e a te d w ith g e l soak I
then I I , and DNA fragm ents tr a n s fe r r e d to th e n i t r o c e l lu lo s e
sh e e t turned through 90° so th a t the DNA bands on th e sh e e t and
on th e g e l were p e r p e n d ic u la r . T ran sfer was c a r r ie d ou t
o v ern ig h t in 4 x SSC and 0 .1 $ sodium d od ecy l su lp h a te a t 75°
in an oven . A ll components o f th e system e x cep t th e g e l were
heated to 75° before commencing t r a n s f e r . The n i t r o c e l lu lo s e
sh e e t was th en washed e x te n s iv e ly w ith 3 x SSC a t 6 0 ° . In
order to o r ie n ta t e th e u n la b e lle d d im ension , a s t r ip was
removed from th e sh e e t b efore c r o s s - b lo t t in g and h y b r id ised

- 57 -
w ith 10^ c .p .m . o f u n r e s tr ic te d n ic k tr a n s la te d HSV DNA.
The s t r ip was washed w ith the sh e e t and reassem b led a lo n g s id e
i t . The assem bly was autoradiographed a f t e r d ry in g and
b a lin g . The la b e l le d dim ension was o r ie n ta te d lay comparing
th e autorad iograp h o f th e c r o s s - b lo t w ith th a t o f th e wet g e l
c o n ta in in g th e n ic k tr a n s la te d fragm en ts .
Autoradiography
G els were autoradiographed e i t h e r a t room tem perature
w ith Kodak K odirex f i lm or w ith Kodak X-Omat H f i lm , or a t
-7 0 ° w ith p r e fla sh e d Kodak X-Oraat f i lm and a Dupont Cronex
p h osp h otu n gsta te sc r e e n ,
R e s t r ic t io n fragm ent nom enclature
Gel e le c tr o p h o r e s is o f r e s t r i c t e d DNA g iv e s a*?
c h a r a c t e r i s t i c number o f bands which are sep ara ted a cco rd in g
to m o lecu lar w e ig h t. Bands are g iv e n l e t t e r s a lp h a b e t ic a l ly
or numbers s e q u e n t ia l ly from slow er to f a s t e r m ig ra tin g
bands ( la r g e r to sm a ller fr a g m e n ts ) . T h is a ls o a p p lie s to „
bands which are l e s s than m olar, such a s th o se gen erated
from th e ter m in a l and j o in t fragm ents o f HSV DNA. The o n ly
exceptions to t h i s are bands r e s u l t in g from sequence a d d it io n s
to o th e r fra g m en ts . For exam ple, th e jo in t fragm en ts HSV-1
BamHI k and IISV-2 BamHI g c o n s i s t o f HSV-1 BamHI k and HSV-2
BamHI g , r e s p e c t iv e ly , w ith an a d d it io n a l a sequence ( s e e ,
fo r exam ple, F igu re C 1 .3 ) . A band which i s more than m olar,
and y e t r e p r e s e n ts o n ly one sequence because th e c o n s t i tu e n t
fragm ents o r ig in a te from e n t ir e ly w ith in a r e p e a t seq u en ce,
i s g iv en o n ly one d e s ig n a tio n ( e . g . HSV-2 BamHI p , g ' , mf
in F igu re C l .2 0 ) . Fragments which com igrate in th e g e l
system used are d esig n a ted from l e f t to r ig h t on th e genome
map ( e .g . HSV-2 BamHI h ij in F igu re C 1 .2 o ).

- 58 -
C a lc u la t io n o f r e l a t iv e m o la r it ie s o f r e s t r i c t i o n fragm ents
A utoradiographs were scanned u s in g a J o y c e - lo e b l
a u to d e n s ito m e te r , and peaks were cu t o u t and w eighed .
M olar r a t i o s o f in d iv id u a l fragm ents were c a lc u la te d from
th e fo l lo w in g form u la .
M olar r a t io — wt o f peak__________ ^ m .w t. o f genome__t o t a l wt o f peak8 m .wt. -of^ fragm ent
R a tio n a le f o r mapping r e s t r i c t i o n s i t e s
The purpose o f r e s t r i c t i o n mapping i s to r e la t e the
r e s t r i c t i o n p r o f i l e to th e p h y s ic a l lo c a t io n o f r e s t r i c t i o n
fragm en ts w ith in th e genome. In t h i s stu d y th e fo l lo w in g
m ethods were u sed .
1 . R ecleavage exp erim en ts. In d iv id u a l ^ P - l a b e l l e d
r e s t r i c t i o n fragm en ts were i s o la t e d and r e c le a v e d w ith a
second enzyme, and th e p rod u cts su b jeo ted to e le c tr o p h o r e s is
a lo n g s id e th e d ig e s t o f th e whole genome w ith th e second
enzyme. T h is method shows which fragm ents produced by th e
second enzyme are w ith in th e r e c le a v e d fragm ent (p rod u cts
co rresp o n d in g to bands in th e r e s t r i c t i o n p r o f i l e o f th e
genome w ith th e second enzym e), and which o f th e produ ct
fragm en ts are a t th e ends o f the r e c le a v e d fragm ent (bands
n o t c o r r e sp o n d in g ) . Large p a r ts o f th e p h y s ic a l map fo r
th e second enzyme may then be d e r iv e d , e s p e c ia l ly whsJSfr th e
map f o r th e f i r s t enzyme i s a lrea d y known. T his method may
be r ev e rse d tjy r e c le a v in g fragm ents produced by th e second
en d on u clease w ith the f i r s t .

- 59 -
2 . Double d ig e s t exp erim en ts, P - la b e l le d DNA was c le a v e d
s im u lta n eo u sly w ith two e n d o n u c lea ses . I f a band i s p resen t
in th e s in g le d ig e s t p r o f i l e but n o t in th e double d ig e s t
p r o f i l e , t h i s in d ic a t e s th a t the fragm ent c o n ta in s a r e s t r i c t i o n
s i t e fo r th e second enzyme. The apparent p resen ce o f a band
in th e double d ig e s t does n o t n e c e s s a r i ly show th a t th e fragm ent
was u n c lea v ed , a s a fragm ent o f la r g e r m o lecu la r w eigh t may have
been c le a v ed to g iv e a product which f o r t u i t o u s ly com igrated
w ith th e band under c o n s id e r a t io n .
3 . H y b r id isa tio n exp er im en ts. ^ P - l a b e l l e d i s o la t e d fragm ents
were h y b r id ised to a n i t r o c e l lu lo s e b lo t o f sep a ra ted
r e s t r i c t i o n fragm ents produced by an o th er en d o n u c lea se .
A lte r n a t iv e ly th e c r o s s - b lo t method was u sed , but th e d a ta
o b ta in ed were e s s e n t i a l l y th e same. These m ethods, taken in
c o n ju n ctio n w ith r e c le a v a g e exp erim en ts, show which fragm ents
are w ith in th e complem entary fragm ent and which extend
p a s t i t s en d s.
C ontam ination o f r e s t r i c t i o n fragm ents i s o la t e d from g e ls
D ig e sted n ic k tr a n s la te d DNA produced a background o f
con tam in atin g seq u en ces in p r e p a r a tiv e g e l s , e s p e c ia l ly i n
the h igh m olecu lar w eigh t r e g io n . T h is i s a fe a tu r e o f
th e n ic k t r a n s la t io n r e a c t io n , but was som etim es due to
con tam in ation o f the r e s t r i c t i o n en d on u clease w ith non
s p e c i f i c n u c le a s e s . Bands in c lo s e p ro x im ity to o th e r s
tended to be contam inated w ith th o se s p e c i f i c seq u en ces .
T h erefo re , i s o la t e d fragm ents were e le c t r o p h o r e t ic a l ly
sep arated both u n c lea v ed , and c le a v ed w ith an endon u clease
o f known p h y s ic a l map, in order to ensure id e n t i t y and
e stim a te p u r ity .

- 60 -
P a in t bands on au torad iograp h s were presumed to r e s u l t
e ith e r from g e n e r a l sequence con tam in ation or from proxim al
band con tam in ation ( e . g . in F igu re 0 1 .8 , Kpnl k was n o t
c le a v ed by Hindl l l , but i t was contam inated both g e n e r a l ly ,
and s p e c i f i c a l l y by Kpnl 1 ) . In n e a r ly a l l r e c le a v a g e
experim en ts some o f th e parent fragm ent was r e fr a c to r y to
d ig e s t io n , d e s p ite com plete d ig e s t io n o f c a r r ie r lambda DNA,
and th e ab sen ce o f bands r e s u l t in g from p a r t ia l d ig e s t io n o f
th e fragm en t. The reason f o r t h i s o b se r v a tio n i s unknown.
Sequence con tam in ation was a ls o d e te c te d in h y b r id is a t io n
experim en ts in v o lv in g i s o la t e d n ic k tr a n s la te d fragm en ts.
G eneration o f b a c t e r ia l c lo n e s b earin g recom binant p lasm id s w ith HSV DNA i n s e r t s - - - -
HSV DNA fragm ents were l ig a t e d in to th e v e c to r pAT153
(Twigg and S h e r r a tt , 1 9 8 0 ), which i s a d e le t io n mutant o f
pBR322 unable to undergo co n ju g a l t r a n s fe r and which c a r r ie s
gen es c o n fe r r in g r e s i s ta n c e to t e t r a c y c l in e and a m p ic i l l in .
Recombinant p lasm id s were tr a n s fe c te d in to th e h o st
E sc h e r ic h ia c o l i K12 s t r a in HHL01 (Boyer and R ou llan d -D u sso ix ,
1 9 6 9 ) . The h o s t was grown in L -broth (0 ,1 7 M NaCI, 10 g /1
D ifco b a c to try p to n e , 5 g /1 y e a s t e x tr a c t ) a t 37° in a
Gallenkamp o r b i t a l in c u b a to r . Transformed b a c te r ia b earin g
pAT153 were grown in 1 -b r o th c o n ta in in g 100 ^ug/ml a m p ic i l l in .
S tock s were prepared by p e l l e t in g b a c te r ia from a 20 ml
o v ern ig h t c u ltu r e l?y c e n tr ifu g a t io n a t 8000 r .p .m . f o r 2 min,
and re su sp en d ing in 10 ml 40# v o l /v o l g ly c e r o l in 1 # w t /v o l
D ifco bactopeptone b e fo re s to r in g a t - 2 0 ° . in 1 ml a l iq u o t s .
HSV-2 HindI I I fragm ents were l ig a t e d in t o pAT153
e s s e n t i a l ly by the method o f Tanaka and Weisblum (1975)*
3 /'g HSV-2 s t r a in HG52 DNA and .l /< g pAT 153 DNA were

- 61 -
s e p a r a te ly d ig e s t e d w ith Hind I I I and th en h e a te d a t 7 0 ° f o r
10 min to i n a c t i v a t e th e en d o n u clea se# The two r e a c t io n s
were mixed and a p p r o p r ia te s o lu t io n s added to b r in g th e f i n a l
c o n d it io n s to 0 .0 2 M T ris-H C l pH 7 .5 , 0 .0 1 M MgCl2 , 0 .0 0 0 5 M
ATP, 0 .0 1 M d i t h i o t h r e i t o l , and 10 U/m l T4 DNA l i g a s e i n
a f i n a l volum e o f 50 z*1 , The m ix tu re was in c u b a te d a t 4°
f o r 16 h r .
The j o in t fra g m en ts HSV-1 BamHI k ( s t r a i n s 17 and USA-8)
and HSV-2 BamHI g ( s t r a i n HG52), and a l s o HSV-1 BamHI y
( s t r a i n 1 7 ) , were i s o l a t e d by h y d r o x y la p a t it e chrom atography
(HSV-1 BamHI y was i s o l a t e d by M -J. M urchie) a f t e r
e le c t r o p h o r e s i s o f 100 yug d ig e s t e d DNA on a g a r o se g e l s
i n th e a b sen ce o f e th id iu m brom ide. The fra g m en ts were
l i g a t e d in 20 yul r e a c t io n volum es a s d e s c r ib e d a b o v e , u s in g
betw een 0 .1 and 0 ,5 yug Bam H I-cleaved pATl53 DNA and th e
amount o f HSV fragm en t p o s s e s s in g th r e e to f i v e t im e s a s many
BamHI te r m in i a s th e pAT153 DNA.
L ig a te d m ix tu r e s w ere t r a n s f e c t e d i n t o th e h o s t
b acteriu m e s s e n t i a l l y a s d e s c r ib e d by Cohen e t a l . ( 1 9 7 2 ) ,
a f t e r c h e c k in g l i g a t i o n by g e l e l e c t r o p h o r e s i s a lo n g s id e
u n lig a te d and r e c le a v e d r e a c t io n m ix tu r e s . A l l p r o ce d u r es
in v o lv in g th e u se o f l i v i n g b a c t e r ia su b seq u en t to l i g a t i o n
o f DNA were perform ed under C ategory I I c o n d i t io n s , h a v in g
o b ta in ed th e a p p ro v a l o f th e G en etic M a n ip u la tio n A d v isory
Group. Jg. c o l i KL2 s t r a in HB101 c e l l s w ere grown to an
o p t i c a l d e n s i t y o f 0 .6 a t 630 nm, h a r v e s te d by c e n t r i f u g a t io n
a t 8000 r .p .m . f o r 2 m in, washed by r e su sp e n d in g in 400 ml
0 .0 1 M NaCI and a g a in c e n t r i f u g in g . The p e l l e t was
resu sp en d ed in 400 ml 0 .0 3 M CaCl2 , iiep t on i c e f o r 20 m in,
and h a r v e s te d a s b e fo r e . The b a c te r ia were re su sp en d ed in 9 .4 ml

- 62 -
o f 1 5 $ v o l / v o l g ly c e r o l in U .03 M CaCl2 , and 0 .2 ml a l iq u o t s4 ~ o
s to r e d a t -7 0 . Under C ategory I I co n ta in m en t c o n d it io n s
a l i q u o t s w ere m ixed in 15 ml capped p l a s t i c tu b e s w ith
l i g a t e d DNA sa m p les , k ep t on i c e f o r 30 m in , th en in c u b a te d
a t 37° f o r 5 m in. 3 ml L -b ro th was added and th e tu b e s
a e r a te d by b u b b lin g a t 37° f o r 90 m in . 0 .1 ml a l iq u o t s were
sp read on 90 mm L -b r o th a g a r p l a t e s c o n ta in in g 1 0 0 yug/m l
a m p ic i l l i n , and in c u b a t io n was a t 37° o v e r n ig h t . A p p ro p ria te
c o n t r o l s were in c lu d e d in th e t r a n s f e c t i o n s : u n d ig e s te d pAT153
-DNA, c le a v e d pATl53 DNA, and c le a v e d pATl53 DNA w hich had■* ■» _ • ibeen l i g a t e d .
The h y b r id is a t io n te c h n iq u e o f G ru n ste in and H ogness
(1 9 7 5 ) was u sed to i d e n t i f y c o lo n ie s b ea r in g recom b in an t
p la s m id s . S t e r i l e g r id d ed n i t r o c e l l u l o s e f i l t e r s were b o ile d
t h r ic e i n w a te r , a u to c la v e d and baked . B a c t e r ia l c o lo n ie s
were t r a n s fe r r e d u s in g s t e r i l e c o c k t a i l s t i c k s on to th e g r id s
on L -b ro th a g a r p l a t e s c o n ta in in g 100 yug/ml a m p ic i l l i n , and
a l s o to a m ic r o t i t r e p la t e c o n ta in in g 0 .2 m l/w e l l L -b ro th
p lu s a m p ic i l l i n . G rids were in c u b a te d a t 3 7 ° o v e r n ig h t and
th e m ic r o t i t r e p l a t e s were k ep t a t room te m p e r a tu r e . The
n i t r o c e l l u l o s e f i l t e r s were t r a n s f e r r e d w ith c o lo n ie s
upperm ost i n tu rn a t room tem p era tu re to f i l t e r p a p ers soaked
w ith 0 .1 U HC1 (1 m in ), 0 .5 M NaOH (15 m in ) , 1 M T ris-H C l
pH 7 .5 (5 m in ), and 1 .2 M NaCI (15 m in ), r e s p e c t i v e l y . The
f i l t e r s were d r ie d i n a i r and baked a t 7 0 ° i n vacuo f o r 2 h r .
P r e in c u b a t io n and h y b r id is a t io n o f th e f i l t e r s were c a r r ie d
ou t in 50/* form am ide a t 45° a s d e s c r ib e d f o r b lo t h y b r id is a t io n .
N ick t r a n s la t e d 1ISV-2 DNA was used a s probe f o r HSV-2 Hind i I I
c lo n e s , and th e i n d iv id u a l ly i s o l a t e d r e s t r i c t i o n fr a g m en ts
l o r HSV-1 HamHI k and y and HSV-2 BamHI g . C o lo n ie s b ea r in g
p la sm id s c o n ta in in g IISV DNA i n s e r t s were i d e n t i f i e d by

- 63 -
a u to r a d io g r a p h y . S to c k s were p rep ared from th e m ic r o t i t r e
p la t e c u l t u r e s a f t e r a f u r th e r s te p o f c o lo n y - p u r i f i c a t io n .
The i d e n t i t y o f p la sm id DNA was a s c e r ta in e d t»y h y b r id is a t io n
o f n ic k t r a n s la t e d p la sm id DNA to b lo t s t r i p s o f HSV
r e s t r i c t i o n d i g e s t s , and by r e s t r i c t i o n m apping o f th e p lasm id
in com p arison w ith HSV genome r e s t r i c t i o n m aps.
Between 1 and 1 0 # o f a m p i c i l l i n - r e s i s t a n t c o lo n ie s were
p o s i t i v e f o r HSV DNA i n s e r t s . 37 HSV-2 H in d l l l c lo n e s were
c h a r a c t e r is e d , 8 HSV-1 s t r a in 17 BamHI k c lo n e s , 3 HSV-1
s t r a in USA-8 BamHI k c lo n e s , 9 HSV-2 BamHI g c lo n e s , and 1
HSV-1 s t r a i n 17 BamHI y c lo n e . T able B I .3 shows the e q u iv a le n t
sta n d a rd n o m en cla tu re f o r t r i v i a l names o f th e c lo n e s u sed
i n t h i s work.
P lasm id DNA was i s o l a t e d e s s e n t i a l l y a s d e s c r ib e d by
C le w e ll and H e lin s k i (1 9 7 0 ) , a f t e r l y s i s o f b a c t e r ia w ith
a n e u tr a l d e te r g e n t (Komano and S in sh e im e r , 1 9 6 8 ) . 1 -2 1
o f l - b r o t h c o n t a in in g 100 yi^;/ml a m p ic i l l in was in o c u la te d w ith
an o v e r n ig h t c u ltu r e o f 1 0 -2 0 ml b a c t e r ia , and grown o v e r n ig h t
a t 37° i n a Gallenkamp o r b i t a l in c u b a to r . C hloroform was
added to 1^ v o l / v o l and th e b a c te r ia h a r v e s te d a f t e r 20 min4
by c e n t r i f u g a t io n a t 8000 r .p .m . f o r 2 m in . The p e l l e t was
resu sp en d ed i n 2 5> w t /v o l su c r o se in 0 .0 5 M T ris-H C l pH 8 ,
10 ml o f 5 mg/ml ly sozym e was added, and th e m ix tu re was k ep t
on i c e f o r 30 m in. In c u b a t io n on i c e was co n tin u e d f o r 30
min a f t e r th e a d d i t io n o f 10 ml 0 .2 h EDTA, and th en 133 /*1
NP4-0 was ad d eu . The tu b e was m ixed g e n t ly , k ep t on i c e f o r
10 m in , and c e n tr i fu g e d a t 20000 r .p .m . f o r 90 min in a
S o r v a l l SS34 r o t o r . The su p er n a ta n t was e x tr a c te d tw ic e by
v ig o r o u s sh a k in g w ith N T E -eq u ilib ra ted p h e n o l, e x tr a c te d
once w ith c h lo r o fo rm , and 3 M sodium a c e t a t e pH 7 .5 was added

- 64 -
to a f i n a l c o n c e n tr a t io n o f 0 .3 M. N u c le ic a c id s weref
p r e c ip i t a t e d by th e a d d it io n o f h a l f a volum e o f propan-2-r-ol,
and p e l l e t e d by c e n t r i f u g a t io n a t 2000 r .p .m . f o r 30 min
a f t e r 1 hr a t room te m p e ra tu r e . The p e l l e t was resu sp en d ed
i n 20 ml w a te r , and 2 5 .5 g s o l i d caesiu m c h lo r id e and nil
eth id iu m brom ide a t 10 mg/ml added, and w a ter added to a volum e
o f 34 m l. T h is was d iv id e d betw een two 17 ml tu b e s and
c e n t r i fu g e d f o r 20 hr a t 1 5 ° in a S o r v a l l TV865B v e r t i c a l
r o t o r . The r e s u l t i n g band o f p lasm id DNA i n ea ch tu b e was
v i s u a l i s e d by lo n g wave UV i r r a d i a t i o n (365 nm) and removed
by p ie r c in g th e tu b e w ith a s y r in g e n e e d le . The volume o f
th e DNA s o lu t io n was in c r e a s e d to 17 ml by th e a d d it io n o f
0 .7 5 g /m l caesiu m c h lo r id e c o n ta in in g 0 .5 mg/ml eth id iu m
brom ide. P lasm id DNA was banded a secon d tim e by c e n t r i f u g a t io n
a s d e s c r ib e d a b o v e , removed from th e tu b e v ia a s y r in g e
n e e d le , s a tu r a te d w ith C sC l, and e x tr a c te d tw ic e w ith aqueous
p r o p a n -2 -o l s a tu r a te d w ith CsCl to remove e th id iu m brom ide.
P u r i f ie d p la sm id DNA was d ia ly s e d o v e r n ig h t a g a in s t NTE,n i
e th a n o l p r e c ip i t a t e d , and r e d is s o lv e d in 1 -2 ml 0 .1 x SSC.
One l i t r e o f b a c t e r ia l c u l tu r e g e n e r a l ly y ie ld e d 0 .5 - 1 mg
o f p la sm id DNA.
La b e l l i n g o f DNA te r m in i w ith
R e s t r i c t io n fra g m en ts from c lo n e d DNA w ere la b e l l e d a t
t h e ir 5* te r m in i e s s e n t i a l l y a c c o r d in g to th e method o f
Maxam and G ilb e r t (1 9 8 0 ) . 1 -4 /*g o f a DNA fra g m en t i s o l a t e d
from an a g a r o se g e l was d ig e s t e d in 50 / a1 o f r e s t r i c t i o n
b u ffe r w ith th e r e s t r i c t i o n en d o n u c lea se c le a v in g a t th e
s i t e s to be l a b e l l e d ( s e e T ab le B 1 .4 ) . 50 jXL o f 0 .0 5 M
T ris-H C l pH 8 and 20 U b a c t e r ia l a l x a l i n e p h o sp h a ta seo
were added and in c u b a te d a t 65 f o r 1 h r . The DNA was

e x tr a c te d w ith 100 o f N T E -satu rated p h e n o l, e th a n o l
p r e c i p i t a t e d , washed w ith 9 5 $ v o l / v o l e t h a n o l, and l y o p h i l i s e d .
The DNA was d i s s o lv e d in 20 yul o f 0 .0 5 M T ris-H C l pH 7 .6 ,
0 .0 1 M MgCl2 , 0 .0 0 5 M d i t h i o t h r e i t o l , 0 .0 0 0 1 M sp e r m id in e ,
0 .0 0 0 1 M EDTA. In th e c a s e o f a r e s t r i c t i o n s i t e n o t g iv in g
a p r o tr u d in g 5' te r m in u s , th e r e a c t io n m ix was h e a te d a t 65°
f o r 2 min and quenched on i c e . 5 /*1 y’-^ P -A T P ( 5000 C i/m m ole)
and 20 U T4 p o ly n u c le o t id e k in a se were added and th e
r e a c t io n in c u b a te d a t 37° f o r 30 m in . The r e a c t io n m ix tu re
was in c r e a s e d i n volum e to 50 yul w ith NTE, p h en o l e x t r a c te d ,
ethanol p rec ip ita ted , washed with 955* ethanol, and ly o p h ilis e d .
U s u a lly th e DNA was th e n r e s t r i c t e d in 20 yul w ith a secon d 1
r e s t r i c t i o n e n d o n u c le a se to g iv e u n iq u e ly l a b e l l e d fr a g m e n ts ,
20 y 1 d y e s in g ly c e r o l ( 0 .0 2 5 $ w t /v o l brom ophenol b lu e ,
0 .0 2 5 $ w t /v o l x y le n e c y a n o l FP i n 1 0 $ v o l / v o l g l y c e r o l )4 , 4 4
added , and th e fra g m en ts se p a r a te d by e l e c t r o p h o r e s i s on a
4 $ p o ly a c r y la m id e g e l i n 0 .5 5 x TBE a t 20 V/cm.4
In one ex p erim en t ( s e e P ig u re 0 3 .1 6 ) , TagI s i t e s were
l a b e l l e d a t 3* t e r m in i . 2 j tg DNA c le a v e d w ith TagI was
in c u b a te d a t 1 5 ° f o r 1 hr i n 50 /* ! o f 0 .0 5 M T ris-H C l pH 7 .8 ,
0 .0 0 5 M MgCl2 » 0 .0 0 1 M d i t h i o t h r e i t o l c o n t a in in g 5 />*1
c*-^P-dC TP and 5 U o f th e Klenow fragm en t o f E . c o l i
DNA p o ly m era se I . The 3 1 l a b e l l e d TagI fra g m en ts w erei f r* rse p a r a te d by e le c t r o p h o r e s i s on an 8 $ p o ly a c r y la m id e 9*3 M
4u rea s e q u e n c in g g e l (d e sc r ib e d b e lo w ) . The r e q u ir e d fragm en t
f o r t u i t o u s l y m ig ra ted a s two s e p a r a te s tr a n d s w hich were
i s o l a t e d d i r e c t l y from th e g e l .
P o ly a c r y la m id e g e l s c o n ta in in g t e r m in a l ly - la b e l l e d
r e s t r i c t i o n fra g m en ts were a u to ra d io g ra p h ed w et w ith
r a d io a c t iv e in k m arkers, and bands e x c is e d and tr a n s fe r r e d
to 1 .5 ml Eppendorf tu b e s . The p o ly a c r y la m id e s l i c e s were

- 66 -
cru sh ed w ith s i l i c o n ! s e d g l a s s r o d s , 0 .6 ml o f 0 .1 $ w t /v o l
sodium d o d e c y l su lp h a te in NTE added to each tube and
in c u b a te d o v e r n ig h t a t 3 7 ° . The tu b e s were c e n tr ifu g e d in
an Eppendorf c e n t r i f u g e f o r 5 m in, th e su p e r n a ta n t removed
and th e p e l l e t r e - e x t r a c t e d w ith 0 .6 ml 0 .1 $ sodium d o d ec y l
s u lp h a te in NTE. The two su p e r n a ta n ts were com bined i n a
15 ml capped p l a s t i c tu b e , and e th a n o l p r e c ip i t a t e d in th e
p r e se n c e o f 30 Mg y e a s t RNA. A fte r c e n t r i f u g a t io n a t 2000
r .p .m . f o r 30 min th e r a d io a c t iv e p e l l e t s w ere r e d is s o lv e d
in 0 .2 ml 0 .1 M sodium a c e t a t e pH 7 .5 , and 0 .1 ml 1 $ w t /v o l
cetyltrim ethylam m onium bromide was added to ea ch . The
1 .5 ml Eppendorf tu b es were k ep t on i c e f o r 15 min and then
c e n t r i fu g e d a t 15000 r .p .m . f o r 15 min a t 4° in an MSE
c e n t r i f u g e . The p e l l e / t s were washed tw ic e w ith 0 .1 M
sodium a c e t a t e pH 7 .5 in 7 0 $ e th a n o l, once w ith 7 0 $ e th a n o l,
once w ith 9 5 $ e th a n o l, and th en l y o p h i l i s e d . The DNA was* T
d is s o lv e d in w ater p r io r to th e c h em ica l d e g r a d a tio n
r e a c t io n s f o r n u c le o t id e se q u e n c in g . In some ex p er im en ts
i s o l a t e d fr a g m en ts w ere la b e l l e d a t both t e r m in i and so th e y
were d ig e s t e d w ith a secon d e n d o n u c lea se a f t e r th e e th a n o l
p r e c i p i t a t i o n s t e p , and u n iq u e ly l a b e l l e d fra g m en ts were
i s o l a t e d a f t e r e le c t r o p h o r e s i s on a secon d 4 $ p o ly a c r y la m id e4
g e l . In a l l th e s t e p s d e s c r ib e d above th e w h ereab ou ts o f
th e l a b e l l e d DNA was fo llo w e d u s in g a r a d ia t io n m o n ito r .

- 67 -
D e te r m in a tio n o f th e n u c le o t id e seq u en ce o f DNA
The c h em ica l d e g r a d a tio n method was u sed (Maxam and
G ilb e r t , 1 9 8 0 ) . U n iq u ely 5 ’ - l a b e l l e d DNA was d is s o lv e d i n
25 /11 w a ter and a l iq u o t s put in to f i v e 1 .5 ml E ppendorf tu b e s
on i c e . R ea g en ts were th en added a s d e s c r ib e d below in o rd er
to m od ify gu an in e (G ), gu an in e and a d en in e (G+-A), a d en in e and
to a l e s s e r e x te n t c y t o s in e (A>C), c y t o s in e and thym ine (C+T),
o r c y t o s in e (C) r e s id u e s . A fte r n e u t r a l i s i n g th e r e a g e n t s ,
th e DNA was t r e a te d in each c a s e w ith p ip e r id in e i n o rd er to
c le a v e th e DNA stra n d a t th e p o s i t i o n s o f m o d if ie d r e s id u e s .
The t r e a te d DNA sam p les were then e l e c t r o p h o r e t i c a l l y
s e p a r a te d . For th e ch em ica l d e g r a d a tio n r e a c t io n s a l l r e a g e n ts
were i c e - c o l d , and m a n ip u la t io n s were c a r r ie d o u t s p e e d i ly
and , where a p p l ic a b le , on i c e . The w h ereab ou ts o f th e
r a d io a c t iv e DNA was fo llo w e d a t a l l s t a g e s w ith a hand
r a d ia t io n m o n ito r . R e a c tio n t im e s were e s t a b l i s h e d e m p ir ic a l ly .
1 . G r e s id u e s . 2 0 0 / i l DMS b u ffe r ( 0 .0 5 M sodium c a c o d y la te
pH 8 . 0 , 0 .0 1 M MgCl2 , 0 .0 0 1 M EDTA) and 1 /CL s o n ic a te d c a l f
thymus DNA a t 1 mg/ml were added to 5 /*1 e n d - la b e l le d DNA.
1 / i l d im e th y lsu lp h a te (DMS) was added in a fume hood to th e
tu b e , w h ich was mixed and th en in c u b a te d a t 25° f o r 2 -1 0 m in .
The tu b e was t r a n s fe r r e d to i c e and 50 /*1 DMS s to p added
( 1 .5 M sodium a c e t a t e pH 7 .0 , 1 M 2 -m e r c a p to e th a n o l, 100 /x g /m l
tRNA) and r a p id ly m ixed . 750 yul e th a n o l was added and m ixed ,
and th e tu b e p u t in an e t h a n o l- s o l id CO2 b a th f o r 5 min
( f i r s t e th a n o l p r e c i p i t a t i o n ) . The tu be was c e n t r i fu g e d f o r
5 min a t 4° in an Eppendorf 5412 c e n t r i f u g e , and th en
t r a n s f e r r e d to i c e . The su p er n a ta n t was removed w ith a
P a s te u r p i p e t t e anu th e p e l l e t d i s s o lv e d i n 250 /CL 0 .3 M
sodium a c e t a t e pH 6 .5 c o n ta in in g 40 / ig/m l y e a s t RNA.

- 68 -
750 yul e th a n o l was add ed , and th e tu b e m ixed and p u t f o r 5 min
i n an e t h a n o l - s o l id CO2 b ath (seco n d e th a n o l p r e c i p i t a t i o n ) .
The su p e r n a ta n t was removed a f t e r 5 min c e n t r i f u g a t io n , and
750 yul e th a n o l c a r e f u l l y added to th e p e l l e t . The tu b e was
c e n t r i fu g e d f o r 2 min a t 4° and th e su p e r n a ta n t d is c a r d e d .
The p e l l e t was d r ie d in a l y o p h i l i s e r . 100 /□ . 1 M p ip e r id in e
was added and th e tu b e h ea ted a t 9 0 ° f o r 30 m in . The s o lu t io n
was fr o z e n on s o l i d CO2 and l y o p h i l i s e d u n t i l no i c e
rem ain ed . 1 0 / i l w a ter was added to th e d r ie d p e l l e t , m ixed ,
f r o z e n and l y o p h i l i s e d , and th en t h i s s te p was r e p e a te d . The
f i n a l p e l l e t was d is s o lv e d in 5 -2 0 yul form am id e-d yes
(8 0 $ v o l / v o l form am ide, 0*55 x TBE, 0 .1 $ w t /v o l x y le n e c y a n o l4 ^ 4
FF, 0 .1 $ w t /v o l brom ophenol b lu e ) .4
2 . G+A r e s id u e s . 17 /»1 w a te r , 1 yul s o n ic a te d c a l f thymus
DNA a t 1 m g/m l, and 2 ^ 1 M p y r id in iu m fo rm a te pH 2 were
added to 3 / I e n d - la b e l le d DNA. The tu b e was in c u b a te d a t
2 5 ° f o r 3 0 -1 2 0 m in, and th en fr o z e n and l y o p h i l i s e d . The
p e l l e t was d is s o lv e d i n 20 yul w a ter , fr o z e n th en l y o p h i l i s e d .
Treatm ent w ith p ip e r id in e and su b seq u en t s t e p s were u n d ertak en
a s f o r th e G r e a c t io n .
3 . A>C r e s id u e s . 1 0 0 yd. 1 .2 M NaOH, 0 .0 0 1 M EDTA and 1 / I
s o n ic a te d c a l f thymus DNA a t 1 mg/ml were added to 5 / L en d -
l a b e l l e d DNA, and in c u b a te d a t 90° f o r 2 -1 0 m in. 150 yul
1 i.i a c e t i c a c id , 1 yul y e a s t KNA a t 10 m g/m l, and 750 ywl
e th a n o l were added . A fte r th e norm al e th a n o l p r e c i p i t a t i o n
p r o c ed u r e , tr ea tm en t w ith p ip e r id in e and su b seq u en t s t e p s were
c a r r ie d o u t a s f o r th e G r e a c t io n .
4 . C-KF r e s id u e s . 15 / - I w a ter , 1 yul s o n ic a te d c a l f thymus
DNA a t 1 m g/ml, and 30 / - l h y d ra z in e were added w ith m ix in g
to 5 /* ! e n d - la b e l le d DNA, and in cu b a ted a t 25° f o r 2 -1 0 m in .

200 jx l h y d r a z in e s to p ( 0 ,3 M sodium a c e t a t e pH 6 .5 , 0 .0 0 0 1 M
EDTA, 50 jug/m l tRNA) was th en added and th e tu b e m ixed.
750 jjOl e th a n o l was added and th e f i r s t e th a n o l p r e c i p i t a t i o n
c a r r ie d o u t . The secon d e th a n o l p r e c i p i t a t i o n and su b seq u en t
s t e p s were done a s f o r th e G r e a c t io n .
5 . C r e s id u e s . The s t e p s f o r th e C+T r e a c t io n were c a r r ie d
o u t e x c e p t t h a t th e 15 >ul w a ter in th e h y d ra z in e r e a c t io n
was r e p la c e d by 15 /d. 5 M NaCI.
A liq u o ts o f 3 -5 1 o f r e a c te d la b e l l e d DNA d is s o lv e d
in form am id e-d yes were h ea ted a t 1 0 0 ° f o r 2 min and quenched
on i c e , b e fo r e lo a d in g on to se q u e n c in g g e l s . For g e l
p r e p a r a t io n , s to c k s o lu t io n s o f d e io n is e d 2 0 # w t /v o l
a cry la ra id e (2 9 :1 w t:w t a c r y la m id e :N N '-m e th y le n e b isa c r y la m id e )
in 8 M u r e a , 10 x TBE, 10 M u r e a , and 1 0 # w t /v o l ammonium4 ^
p e r s u lp h a te were u s e d . S o lu t io n s o f th e a p p r o p r ia te
a c r y la m id e c o n c e n tr a t io n were p rep ared by d i l u t i n g th e 2 0 #
a c r y la m id e s o lu t io n w ith 10 M u r e a . 10 x TBE and 1 0 #4
ammonium p e r s u lp h a te were added to f i n a l c o n c e n tr a t io n s o f
0 .5 5 x TBE and 0 .0 6 7 # , r e s p e c t i v e l y , and th e p o ly m e r is in g4
c a ta ly s t TEMED added to 0 .0 0 0 5 # v o l /v o l . Sequencing g e ls
o f dimensions 450 x 230 x. 0 .5 mm were poured, using 60 ml
so lu tio n for each g e l . Three concentrations o f g e l were
used: 6# polyacrylam ide, 9*3 M urea; 8# p61yacrylamide,4 4
9 .3 M u r e a ; 1 9 # p o ly a c r y la m id e , 8 M u r e a . P r e - e le c t r o p h o r e s is4
o f g e l s was c a r r ie d o u t f o r 1 - 1 .5 h r a t 40 W ( 1 .5 - 2 kV)
b e fo r e lo a d in g th e sa m p les . G els were s u b je c te d to
e l e c t r o p h o r e s i s a t 40 W such th a t 1 9 # p o ly a c r y la m id e r e s o lv e d
1 -3 0 n (brom ophenol b lu e marker 15 cm down g e l ) , 0 #
p o ly a c r y la m id e r e s o lv e d 2 0 -8 0 n (x y le n e c y a n o l marker 15 cm
down g e l ) and 8 0 -1 2 0 n (x y le n e cy a n o l marker to bottom o f g e l ) ,

- 7 0 -
and 6?o p o ly a c r y la m id e r e s o lv e d 1 2 0 -1 5 0 n (x y le n e c y a n o l to
bottom o f g e l ) and 150 n and la r g e r (se co n d x y le n e c y a n o l
marker a p p lie d when th e f i r s t had rea ch ed th e bottom o f th e
g e l , th en a llo w e d to m ig r a te 15 cm down g e l ) .
A fte r e le c tr o p h o r e s is one g la s s p la t e was removed.
The g e l was wrapped in a p la in S t e r i l i n bag and a u to ra d io g ra p h ed
f o r 1 -3 0 d ays a t -7 0 ° u s in g p r e f la s h e d Kodak X-Omat H f i lm*
and a Dupont Cronex p h o sp h o tu n g sta te s c r e e n .

Table .Bl.l
Source b a c te r ia fo r r e s t r i c t i o n en d on u cleases
used in t h i s s tu d y , w ith growth tem peratures
fo r batch l iq u id c u l t u r e s .
Those b a o te r ia n o t a ss ig n e d a growth
tem perature were su p p lie d a s fr o z e n c e l l
p a s te s by th e M ic r o b io lo g ic a l R esearch
E sta b lish m e n t, P orton Down, W ilt s h ir e , and
by Dr J . Arrand.

-p
c~\
O I
0) Xt

Table B1.2
Elution of restrlotlon endonucleases from ion exchange columns*The molarity of TCC1 In the first ooluisn fraotlon containing restriction activity was calculated from the linear KC1 gradient used for elution*

Table B1.2
restrictionendonuclease
approx* eluting molarities (KCl)DE52 Biorex
Xba I 0.2 _ _
Hind III 0 0.3BcoR I 0.13 0.45Bgl II 0.18 0.3Hna I 0.15 0.2Kpn I Q.1 0.15BamH I 0.1 0.3Pvu II 0.15 0.4

Table B1.5
N om en clatu re o f recom b in an t p la sm id s b ea r in g HSV-1 (pGX)
o r HSV-2 (pGZ) DNA i n s e r t s . T r iv ia l names a re u sed in
th e t e x t .
The i n s e r t in pGZ12 c o n s i s t s o f HSV-2 H in d III £ and
a b o u t 1 x 10 d a l to n s o f a d ja c e n t seq u en ce from
HSV-2 H in d l l l 1 ( s e e P ig u re C 2 .4 ) . I t may have
o r ig in a t e d from a d e f e c t i v e DNA m o le c u le .
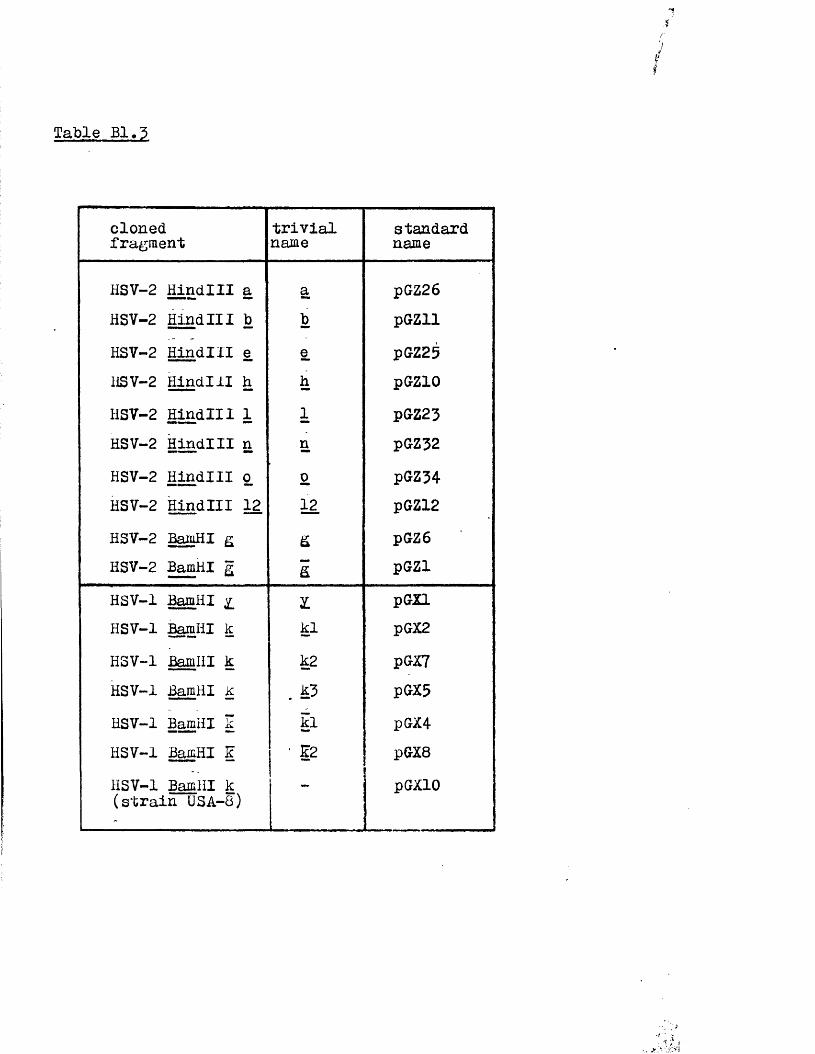
c lo n e dfragm en t
t r i v i a lname
sta n d a rdname
HSV-2 H in d III a a pGZ26
HSV-2 H in d III to to pG Zll
HSV-2 H in d iI I e e pGZ25
HSV-2 H in d iI I h h pGZIO
HSV-2 H in d III 1 1 pGZ23
HSV-2 H in d iI I n n pGZ32
HSV-2 H in d iI I o 0 pGZ34
HSV-2 H in d III 12 12 pGZ12
HSV-2 BarnHI g £ pGZ6
HSV-2 BamHI g 1 pGZl
HSV-1 BamHI £ I pGXl
HSV-1 BamHI k k l pGX2
HSV-1 BamHI k k2 pGX7
HSV-1 BamHI k . & PGX5
HSV-1 BamHI k k l PGX4
HSV-1 BamHI I ' g2 pGX8
HSV-1 BamHI k ( s t r a i n USA-8)
i pGXIO

Table 3 1 .4
S i t e s f o r r e s t r i c t i o n e n d o n u c le a se s u sed in t h i s
Work (from R o b e r ts , 1 9 8 1 ) . C leavage p o s i t i o n s are shown
by a p o s tr o p h e s .

T a b l e B l . 4
A lu l AGr* CT Kind I I I A1 AGCTT
A val C'ByCGPuG H ln f l G1 AN TC
A v a il G* GA/TCC Hpal GTT1 AAC
Bam lil G» GATCC Kpnl GGTAC’ C
B e l l T’ GATCA MboII G AA GAN u N NN N NN1
B ^ l l GCCKKNN'NGGC M nll CCTC (5 -•10 n . ~ 3 ' )
B ^ lI I A’ GATC'if P vu II CAG'CTG
BstN I CC1 A/TOC Sac I GAGO T1C
C la l AT'CGAT Sau3AI 1GATCf
D del C’ TNAG S s t I GAGCT'C (a s S a o l)
gcoKI G! AATTC S s t l l CCGC1 GG
l l a e l l PuCCGC'Py Sraal CCC1GGG
lia o I I I GG'CC Tag I T»CGA
n h a l GCG’C Xbal T’CTAGA
H ln cII GTPyPuAC Xhol C * TC GAG

P A & T C
it i!i S U L i1 S
A N D
D I S C U S S I O N

SECTION 1
liAPPIHG OP RESTRICTION ENDONUCLEASE SITES
ON HSV-1 AND HSV-2 DNA

- 71 -
INSULTS
I n tr o d u c t io n
R e s t r ic t io n maps f o r th e genom es o f HSV-1 s t r a in 17
and HSV-2 s t r a in HG52 which were a v a i la b le a t th e
commencement o f t h i s work were f o r X b al, H i n d l l l , BcoRI,
B g l l l and Hpal , and a l s o Kpnl f o r s t r a i n HG52 (W ilk ie , 1976;
C o r t in i and W ilk ie , 1 9 7 8 ) . These e n d o n u c le a se s produce
r e l a t i v e l y few r e s t r i c t i o n fra g m en ts from HSV DNA, and so
.the o b j e c t o f th e work d e s c r ib e d h e re was t o map HSV-1 and
HSV-2 DNA f o r e n d o n u c le a se s w hich produce many fra g m en ts , in
o rd er to be a b le to d e f in e more c l o s e l y th o se s t r u c t u r a l
and f u n c t io n a l a s p e c t s o f th e genome u nder i n v e s t i g a t i o n .
M o lecu la r w e ig h t c a l ib r a t io n
The s ta n d a r d s u sed to d eterm in e th e m o le c u la r w e ig h ts
o f r e s t r i c t i o n fra g m en ts were th e EcoRI d i g e s t o f lambda
DNA (Thomas and D a v is , 1 9 7 5 ) , th e H in d ll d i g e s t o f jrfx 174
nJMA (Mania i i s e t a l . , 1 9 7 5 ) , and th e Hind I I d i g e s t o f lambda
DNA (M a n ia tis e t a l . , 1 9 7 5 ) . These d i g e s t s were u sed to£
c a l ib r a t e m o le c u la r w e ig h ts i n th e r a n g e s o f 1 -1 0 x 106 6
0 .1 - 1 x 1 0 , and 0 .0 7 - 0 .1 x 10 , r e s p e c t i v e l y .
F ig u re C l . l shows th e g e l u sed to c a l ib r a t e
th e la r g e r fra g m en ts o f th e Kpnl , BamHI, X h o l, and Hpal
d i g e s t s o f HSV-1 DNA. The la r g e r fra g m en ts o f HSV-1 P vuII
were c a l ib r a t e d u s in g HSV-1 Hpal fragm en t m o le c u la r w e ig h ts
(W ilk ie , 1 9 7 6 ) . HSV-2 B g ll l fragm en t m o le c u la r w e ig h ts
(C o r t in i and W ilk ie , 1978) were u sed to c a l i b r a t e la r g e r
HSV-2 BamHI fr a g m e n ts .
F ig u re C l .2 shows th e a u to ra d io g ra p h u sed to c a l ib r a t e
th e sm a lle r fra g m en ts o f HSV-1 BamHI , HSV-1 X h o l, and HSV-2
BamHI. Com parison o f HSV-1 Xhol w ith HSV-1 P v u II a llo w ed

- 72 -
m o le c u la r w e ig h ts o f th e s m a lle r P vu II fra g m en ts to be
d e te rm in ed , HSV-1 Kpnl b 1 and c 1 were c a l ib r a t e d a g a in s t
th e HSV-1 BamHI d i g e s t , and HSV-I BamHI k '-n * a g a in s t lambda
H indl l fra g m en ts on a p o ly a c r y la m id e g e l .
The d is t a n c e m ig ra ted by s ta n d a rd bands was p lo t t e d
a g a in s t th e lo g a r ith m o f th e m o le c u la r w e ig h t , and t h i s cu rv e
was u sed to deduce m o le c u la r w e ig h ts o f r e s t r i c t i o n fra g m en ts
(Tables C 1.1-C 1.5). I t was assumed that the standard curve
was v a l i d f o r a l l r e s t r i c t i o n fr a g m e n ts; h ow ever, th e
n u c le o t id e c o m p o s it io n o f DNA m ight a f f e c t i t s e le c t r o p h o r e t ic*
m o b il i t y and h en ce th e a p p a ren t m o le c u la r w e ig h ts o f some
fr a g m e n ts .
R e la t iv e m o la r i t i e s o f r e s t r i c t i o n fra g m en ts
F ig u re C l . 3 show3 r e s t r i c t i o n p r o f i l e s o f
in v i v o ^ P - l a b e l l e d HSV-1 and HSV-2 DNA. A u to d en sito m etry
o f t h e s e , and o th e r , a u to r a d io g r a p h s was c a r r ie d o u t to
d e term in e th e r e l a t i v e m o la r i t i e s o f bands in th e p r o f i l e s
(T a b le s C 1 .1 -C 1 .5 ) . P o t e n t ia l s o u r c e s o f e r r o r in such
e s t im a t io n s in c lu d e in a c c u r a te m o le c u la r w e ig h ts , d e g r a d a tio n
o f DNA, a n o n - l in e a r r e sp o n se o f f i lm to r a d ia t io n ex p o su re ,
and fra g m en t h e t e r o g e n e i ty ( f o r in s t a n c e , a p r o p o r t io n o f
th e HSV-1 BamHI j o in t fragm en t k c o n ta in s two a seq u e n c es
in s t e a d o f o n e , and t h e r e fo r e co m ig ra ted w ith BamHI .
N e v e r t h e le s s , in g e n e r a l th e m easured r e l a t i v e m o la r i t i e s
a re in agreem en t w ith th o se p r e d ic te d from th e p h y s ic a l maps
( b e lo w ) . Bach o f th e r e s t r i c t i o n e n d o n u c le a se s f o r which
HSV-1 o r HSV-2 DNA was mapped in t h i s work h as a s i t e i n one
or b o th o f th e s e t s o f in v e r te d r e p e t i t i o n s and th e r e fo r e
p ro d u ces no q u a r ter -m o la r fr a g m e n ts , s in c e o n ly e n d o n u c le a se s
w ith no s i t e s in r e p e t i t i v e se q u e n c e s show t h i s c h a r a c t e r i s t i c .

- 73 -HSV-1 Kpnl p h y s ic a l map: r e c le a v a g e d a ta
F ig u r e s C 1 .4 -C 1 .5 show th e p r o d u c ts o f Kpnl c le a v a g e
o f i s o l a t e d HSV-1 r e s t r i c t i o n fra g m en ts o f known map
p o s i t i o n s . F ig u re C l . 6 shows a summary o f th e s e d a ta .
In th e f o l lo w in g argum ent m o le c u la r w e ig h ts a re g iv e n in m i l l i o n s .
1 . The S seg m en t.
.Both Hind i I I g and B g l l l 1 gave K£nl k a s an S te r m in a l
fra g m en t, and BcoRI h p r o d u c ts show Kpnl h to map in Us-
C lea v a g e o f KcoKI h and k in d ic a t e a th a t S c o n ta in s th r e e
fr a g m e n ts . The t h ir d Kpnl fragm en t i s la r g e r than
H indl l l m ( 4 . 6 ) , s in c e t h i s fragm en t was n o t c le a v e d oy
K pnl, and th e p r o d u c ts o f H indI I I n s u g g e s t th a t th e m o le cu la r
w e ig h t o f th e th ir d fragm en t i s 5*5 ( 4 .6 + 0 . 9 ) . S im i la r ly ,
c le a v a g e o f BcoRI h and k im p l ie s a s i z e o f 5 .4 ( 3 .5 + 1 .9 )
f o r th e t h ir d fra g m en t. T h is s i z e c o r r e sp o n d s to th a t o f
Kpnl j , w hich in d eed was a p ro d u ct o f th e c le a v a g e o f
Xbal a , X bal be, Hpal c d e , and JBglll g h . B oth Kpnl j. and k
w ere h a l f m olar in th e HSV-1 Kpnl d ig e s t (T a b le C l . l ) .
T h e re fo r e th e o rd er o f Kpnl fra g m en ts in S i s j. h k .
T h is c o n c lu s io n was u sed to deduce w h ich p r o d u c ts o f
Hpal gh were produced froip Hpal g and w hich from h , and th e
p r o d u c ts a re shown s e p a r a te ly in F ig u re C l. 6,. S im ila r
r e a s o n in g was a p p lie d w ith Hpal c d e , B g ll l g h , and X bal be
to d eterm in e th e c le a v a g e p r o d u c ts o f in d iv id u a l fr a g m e n ts .
2 . THL/IH L.
Fragm ents c o n ta in in g th e 1 term in u s gave Kpnl r a s a
common p r o d u c t . T h ere fo re Kpnl r maps a t th e 1 te r m in u s .
T n is a l lo w s th e p r o d u c ts o f H in d l l l h and i to be
deduced from H in d l l l h i .

- '74 -
3 . J o in t fr a g m e n ts .
The s i z e s o f L and S te r m in a l fr a g m e n ts in d ic a t e t h a t
th e two h a l f m olar j o i n t fra g m en ts have m o le c u la r w e ig h ts o f
'7.9 ( 5 .5 + 2 .4 ) and 7 .2 ( 4 .8 + 2 . 4 ) . Hpal a gave Kpnl a ( 8 . 0 ) ,
and Kpnl e (7 * 0 ) was a p rod u ct o f r e le v a n t c le a v a g e s ( Hpal
c d e . B g ll l e , Xbal q , Xbal b e ) • The j o in t fra g m en ts a r e
t h e r e fo r e a and e .
4 . J u n c t io n s o f TRj/ I R j, w ith Uj,.
There i s only"one" Kpnl s i t e i n TR j/IR ^, s in c e o n ly one
Kpnl fragm en t was common to b o th 1 te r m in i ( Kpnl r ) .
T h ere fo re fra g m en ts a d ja c e n t to t h i s s i t e m ust ex ten d in t o
UL, and h en ce must be g r e a te r in s i z e th an 3 .6 ( 6 . 0 - 2 . 4 ) .
The o n ly fragm en t from B g ll l f and Xbal d w h ich m eets t h i s
req u irem en t i s Kpnl g . S im i la r ly , th e r e s u l t s from B g l l l q
and k g iv e a s i z e o f 8 .1 ( 7 .0 + 1 .1 ) f o r th e o th e r ju n c t io n
fra g m en t, and t h i s c o rresp o n d s to Kpnl b . T h ere fo re th e
fra g m en ts so f a r deduced a r e , a s sp aced a lo n g th e genom e,
r b . . . . g r / j h k .
5 . Ujj from l e f t to r i g h t .
R e c lea v a g e o f H indl l l h and B g l l l k show s t h a t K gnl x
i s a d ja c e n t to Kpnl b . C leavage o f B g l l l o and k , b e a r in g
in mind t h a t Hpal k and u were n o t c le a v e d by K pnl, in d ic a t e s
th e s i z e o f th e n e x t fragm en t i s 6 .9 ( 4 . 6 + 2 . 3 ) - T h is
co rresp o n d s to Kpnl f , w hich was produced from Xba I c .
Comx)arison o f B g l l l p and Hpal j, c le a v a g e show s th a t th e n e x t
two fra g m en ts a re Kpnl q then w. In l i k e manner th e p r o d u c ts
o f Xbal c , B g l l l m and Hpal i d em on stra te t h a t th e n e x t th r e e
fra g m en ts a r e Kpnl i , b ’ th en m. T h ere fo re th e o rd er so f a r i s
r b y f q w i b ’ m . . . . g r / q h k .

- 75 -
The n e x t r e g io n c o n ta in s Kpnl n , p , t , v , x and a 1 ,
a c c o r d in g to c le a v a g e o f X bal f . The fragm en t a t th e l e f t
hand end o f t h i s group i s shown by c le a v a g e o f Hpal b to be
Kpnl t . P r o d u c ts o f B g ll l i in d ic a t e t h a t Kpnl n and p a re
th e n e x t two fr a g m e n ts , but t h e s e ca n n o t be ord ered w ith
r e s p e c t to ea ch o th e r by r e c le a v a g e m ethods b ecau se none o f
th e o th e r e n d o n u c le a se s c l e a v e s w ith in them . The n e x t fragm ent
i s l a r g e r in s i z e t h a t 1 * 0 , and c le a v a g e o f EcoRI m th e r e fo r e
show s th e o rd er o f Kpnl v , x , §t' i s y a* x . So th e o r d e r o f
fra g m en ts w ith in Xbal f i s Kpnl t (nq) v a* x .
The n e x t fragm en t i s la r g e r in m o le c u la r w e ig h t than •
6 .0 , a s shown by com p arison o f th e p r o d u c ts o f Hpal h and
EcoRI 1 . Kpnl c , a p ro d u ct o f B g l l l d , i s th e o n ly fragm ent
m eetin g t h i s r e q u ir e m e n t. C lea v a g e o f Xbal e , H in d l l l k ,
EcoRI a , B g l l l d and Hpal f show t h a t th e n e x t th r e e
fr a g m en ts a re Kpnl 1 c* s , and com parison o f th e B g l l l g ,, - . . * . . . . .
Hpal e and EcoRI i r e s u l t s d em o n stra te t h a t th e s e a re
fo l lo w e d by Kpnl d z u . The f i n a l fra g m e n t, from c le a v a g e
o f B g l l l f and Xbal d , i s Kpnl p .
In c o n c lu s io n , th e o rd er o f Kpnl fr a g m en ts w ith in
th e HSV-1 genome i s
r b y f q w i b ' m t (nq) v a 1 x c 1 c 1 s d z u o g r / p h k ,
w ith Kpnl a and e r e s u l t i n g from f u s io n a t th e j o i n t o f
Kpnl p and r , and k and r , r e s p e c t i v e l y . T h is c o n c lu s io n
a c c o u n ts f o r a l l th e bands in th e HSV-1 Kpnl r e s t r i c t i o n
p r o f i l e in t h e ir r e s p e c t iv e m o la r i t i e s . No fragm en t s m a lle r
than Kpnl c 1 h as been d e t e c t e d .
P ig u re C l . 6 , show ing a summary o f c le a v a g e d a ta ,
g iv e s th e p r o d u c ts o f in d iv id u a l fra g m en ts w ith in c o m ig r a tin g
bands a s a lr e a d y d e s c r ib e d , in c lu d in g Hpal o p q r .

- 76 -HSV-1 Kpnl p h y s ic a l map: d ou b le d i g e s t d a ta
P ig u re C l . 7 shows th e r e s u l t s o f s i n g l e and d o u b le
d ig e s t io n o f HSV-1 BNA. P ig u r e s C l . 8 - C l . 9 . show th e
r e s u l t s o f c le a v in g i s o l a t e d Kpnl fra g m en ts w ith o th e r
e n d o n u c le a s e s . These d a ta a re sum m arised in T ab le C l . 6 .
The r e s u l t s a re c o n s i s t e n t w ith th e p h y s ic a l map deduced
from r e c le a v a g e d a ta .
HSV-1 Kpnl p h y s ic a l map: h y b r id is a t io n d a ta
H y b r id is a t io n d a ta a r e p ro v id ed by th e c r o s s - b l o t
shown i n P ig u r e C l ,1 0 . Each s p o t on th e a u to r a d io g ra p h
i n d i c a t e s a r e g io n o f seq u en ce hom ology betw een th e two
fra g m en ts co n c er n e d . Por exam p le , Kpnl wx h y b r id is e d to
Hpal b and q , and r e c le a v a g e d a ta show t h a t Kpnl w maps
in Hpal q and Kpnl x i n Hpal b . The d a ta a re sum m arised in
P ig u re C l .1 1 , u s in g th e p h y s ic a l map a lr e a d y d ed u ced , w ith
w hich th e y a r e c o n s i s t e n t . Homology betw een th e 1 and S
te r m in i i s e x p la in e d by th e p r e se n c e a t both te r m in i o f
th e te r m in a l r e p e t i t i o n , or a seq u e n c e , w hich i s a l s o
p r e s e n t a t th e j o in t o f th e L and S se g m e n ts .
HSV-1 Kpnl p h y s ic a l mapt r e l a t i v e o rd er o f fra g m en ts n and p
None o f th e p r e v io u s m ethods a llo w e d th e o r d e r in g o f
Kpnl n and q , a s no r e le v a n t r e s t r i c t i o n s i t e i s p r e s e n t
w ith in th e r e g io n c o n ta in in g them . Work d e s c r ib e d i n S e c t io n
2 show s th a t th e genom es o f HSV-1 and HSV-2 a r e g e n e r a l ly
c o l in e a r , and s t u d ie s o f in t e r t y p ic reco m b in a n ts i s in
a cco rd w ith t h i s h y p o th e s is ( e . g . P r e s to n e t a l . , 1 9 7 8 ) .
T h erefo re a r e s t r i c t i o n s i t e w ith in HSV-2 seq u e n c es hom ologous
to HSV-1 Kpnl n and q co u ld be used to o r ie n t a t e th e s e two
fr a g m e n ts . The r e s t r i c t i o n s i t e u sed was th e B g ll l s i t e
betw een HSV-2 B g ll l q and o (C o r t in i and W ilk ie , 1 9 7 8 ) .

- 77 -
In P ig u re C l .12 th e r e s u l t i s shown o f h y b r id is in g
Kpnl no and q to n i t r o c e l l u l o s e b lo t s t r i p s c o n ta in in g
HSV-1 Hpal or HSV-2 B g ll l r e s t r i c t i o n fr a g m e n ts . Kpnl no
h y b r id is e d to HSV-1 Hpal b, e and q . P resum ably th e form er
was ow ing to h y b r id is a t io n o f Kpnl n and th e l a t t e r two to
Kpnl o . H y b r id is a t io n to Hpal i was p ro b a b ly due to
c o n ta m in a tio n o f Kpnl nq by Kpnl m. Kpnl q h y b r id is e d to
Hpal b o n ly , show ing th a t t h i s fra g m en t was r e l a t i v e l y p u re .
With HSV-2 B g ll l b l o t s t r i p s Kpnl no h y b r id is e d to B g ll l i
'and but n o t o . H y b r id is a t io n to B g ll l i was presum ably
due to Kpnl o and to B g ll l j. by Kpnl n , a s th e y are
d i f f e r e n t i a t e d by p h y s ic a l map p o s i t i o n . Kpnl q h y b r id is e d
to B g l l l q and o . These r e s u l t s s u g g e s t t h a t Kpnl n and q
a re in th e o rd er n q in th e HSV-1 genom e.
HSV-1 Kpnl p h y s ic a l map
The p h y s ic a l map c o n s i s t e n t w ith a l l th e d a ta i s
shown in P ig u re C l .1 8 .
P h y s ic a l maps f o r o th e r r e s t r i c t i o n e n d o n u c le a se s
The d e r iv a t io n o f th e HSV-1 Kpnl p h y s ic a l map h a s been
d e s c r ib e d , and maps o f HSV-1 DNA f o r BamHI . PvuI I and X h o l,
and o f HSV-2 DNA f o r BamHI were c o n s tr u c te d in s im i la r m anner.
A u torad iograp h s o f r e c le a v a g e , d o u b le d ig e s t io n and
h y b r id is a t io n ex p er im en ts have n o t been in c lu d e d , but in s t e a d
a re summarised in d iagram m atic (F ig u r e s C 1 .1 3 -C 1 .1 7 ) o r
ta b u la r form (T a b le s C 1 .7 -C 1 .1 2 ) . P h y s ic a l maps deduced
from r e c le a v a g e d a ta were con firm ed by d o u b le d i g e s t i o n and
h y b r id is a t io n e x p e r im e n ts , e x c e p t f o r HSV-1 P v u II , f o r w hich
h y b r id is a t io n ex p er im en ts were n o t c a r r ie d o u t . A ll
d e t e c ta b le HSV-1 Kpnl and BamHI fra g m en ts were mapped, but
in th e o th e r d i g e s t s an undeterm ined number o f v e ry sm a ll

- 78 -
fra g m en ts were n o t r e s o lv e d and t h e r e f o r e rem ained unmapped,
P in a l r e s t r i c t i o n maps a r e shown in P ig u r e s C 1.18 and C l .1 9 ,
and a l ig n e d HSV-1 and HSV-2 maps in P ig u r e C l .2 0 .
The m ethods d e s c r ib e d above f a i l e d to unam biguously
map a few fra g m en ts w hich were su b se q u e n tly a s s ig n e d a s f o l l o w s .
1 . K ec lea v a g e ex p e r im en ts showed th a t HSV-1 BamHI 1* and n f
a re lo c a t e d a d ja c e n t to BamHI z . The i n t e r t y p i c recom b in ant
HE4 (P ig u re 04*26) p o s s e s s e s a c r o s s o v e r in HSV-1 BamHI z ,
and produced BamHI n' but n o t l f , and t h e r e fo r e th e o rd er
HSV-1 BamHI n v z 1 ! i s th e m o s t ' ' l ik e ly .
2 . HSV-2 BamHI i and i 1 were ord ered by in f e r e n c e from d a ta
co n c er n in g th e i n t e r t y p i c recom b in ant RE4 (P ig u r e C 4 .2 6 ) .
T h is recom b in an t p o s s e s s e s an in s e r t i o n o f th e HSV-2 seq u en ce
sp an n in g th e HSV-2 BamHI i - r r e g io n in a n o th e r r e g io n o f th e
genome, a s d e s c r ib e d in S e c t io n 4 . The in s e r t e d p ie c e o f DNA
d o es n o t in c lu d e HSV-2 BamHI i 1, and th e r e fo r e i t i s l i k e l y
th a t i and r a re c o n t ig u o u s .
3 . I n t e r t y p ic reco m b in a n ts w ith a c r o s s o v e r in HSV-1 BamHI y
a s shown below la c x e d HSV-2 BamHI g ' , but produced a
c h a r a c t e r i s t i c a l l y s m a lle r fragm en t and HSV-2 BamHI m*
( e . g . HE4 s u b c lo n e s 1 , 5 , 7 , 1 5 , 1 6 , 17 in P ig u r e C 4 .2 2 ) .
These r e s u l t s s u g g e s t t h a t g ’ i s a d ja c e n t to u in th e HSV-2
BamHI map.HSV-1 DNA
x yL
— —T—— ——————1•m' q4
H i'Kg
T TTa 1 m1 g
•.
tu
HSV-1 BamHI s i t e s
HSV-2 BamHI s i t e s
HSV-2 DNA

- 79 -
Discussion
The g e n e r a l arrangem ent o f th e HSV genome h as been
a d e q u a te ly d em on stra ted and th e r e fo r e was assum ed in
d ed u c in g th e r e s t r i c t i o n maps p r e se n te d h e r e . The p r e se n c e
o f TRj/ I R jq and TRg/IRg was co n fir m ed . BamHI i s th e o n ly
r e s t r i c t i o n e n d o n u c le a se , f o r w hich th e e n t i r e HSV-2 genome
h a s been mapped, w hich c le a v e s w ith in b o th s e t s o f r e p e a t s ,
th u s d e m o n str a tin g t h e i r p r e s e n c e .
The s u b se q u e n tly p u b lish e d Kpnl and BamHI maps o f
HSV-1 s t r a i n s P and J u s t in (L ocker and P r e n k e l, 1 9 7 9 -b )d iffe r
from th o s e o f HSV-1 s t r a in 17 o n ly in th e p r e se n c e o r a b sen c e
o f a few r e s t r i c t i o n s i t e s , but in g e n e r a l a l l th r e e s t r a i n s
nave v e r y s im i la r r e s t r i c t i o n m aps. In c o n t r a s t , few i f
any r e s t r i c t i o n s i t e s a re i n e q u iv a le n t p o s i t i o n s in HSV-1
and HSV-2 DMA, even i n th e c a s e o f BamHI w h ich has many
more s i t e s in HSV DNA than p r e v io u s ly mapped e n d o n u c le a s e s .
T h is i s t r u e even o f th e more hom ologous r e g io n s betw een
th e two genom es ( s e e S e c t io n 2 o f R e s u l t s ) . Such a la r g e
d i f f e r e n c e i s o f ad v a n ta g e in d e te r m in in g th e genome
s t r u c t u r e s o f HSV-l/HSV-2 in t e r t y p ic reco m b in a n ts by
r e s t r i c t i o n en d o n u c le a se a n a l y s i s . I t ca n n o t be assum ed
th a t any r e s t r i c t i o n s i t e s in th e one genome a r e in
e q u iv a le n t p o s i t i o n s in th e o th e r , and t h e r e f o r e
r e s t r i c t i o n maps o f com parable r e g io n s o f th e two genom es
m ust be d eterm in ed s e p a r a te ly in ea ch c a s e .
S e v e r a l l i n e s o f e v id e n c e s u g g e s t t h a t th e KSV-1 and
HSV-2 genom es a re c o l in e a r , a s d is c u s s e d in S e c t io n 2 .
T h is v/as assum ed to a p p ly to th e r e g io n o f c o n ta in in g
HSV-1 Kpnl n and £ in d e te r m in in g t h e i r r e l a t i v e o r d e r
by h y b r id is a t io n to HSV-2 r e s t r i c t i o n fra g m en ts (P ig u r e
C l . 1 2 ) . I t i s p o s s ib le t h a t c o l i n e a r i t y d o es n o t a p p ly

- 80 -
in t h i s r e g io n , a lth o u g h th e s im p le s t in t e r p r e t a t io n o f
th e recom b in an t genome a n a ly s e s d e s c r ib e d by C hartrand e t a l .
(1979 and 1 980) i s t h a t c o l i n e a r i t y d o e s a p p ly .
R e s t r i c t io n fra g m en ts o r ig in a t in g from th e j o i n t
and L term in u s o f both HSV-1 and HSV-2 ENA d em o n stra ted an
a d d i t io n a l d i s c r e t e but f a i n t band m ig r a t in g a t a p o s i t i o n
3 0 0 -4 0 0 bp la r g e r th an th e m ajor s p e c ie s ( e . g . HSV-1 BamHI
k above BamHI k in P ig u re C1.3> and a f a i n t band above
HSV-1 Hpal m in P ig u re C l .7 tr a c k 1 1 ) . In some a u to r a d io g r a p h s
v e r y f a i n t bands r e p r e s e n t in g fu r th e r d i s c r e t e a d d i t io n s w ere
o b se r v e d . Pragm ents from th e S term in u s were p r e s e n t a t
t h i s r e s o lu t io n a s a s in g l e s i z e c l a s s . T hese o b s e r v a t io n s
co n firm th o s e o f Wagner and Summers (1 9 7 8 ) f o r HSV-1 and
W ilk ie e t a l . (1 9 7 7 a )fo r HSV-2. Wagner and Summers (1 9 7 8 )
co n c lu d ed t h a t th e a d d i t io n a l DNA c o m p r ise s th e a seq u en ce
and p erh ap s p a r t o f th e b s e q u e n c e . I t c o n t a in s , in f a c t ,
p r e c i s e l y th e a seq u en ce ( S e c t io n 3 o f R e s u l t s ) .
The m .w t. o f th e HSV genome can be c a lc u la t e d by
summation o f m .w t s .o f th e in d iv id u a l r e s t r i c t i o n fr a g m e n ts .
The mean m .w t. o f HSV-1 DNA, from th e fo u r d i g e s t s d e a l t w ith
h e r e , i s 9 6 . 4 ± 2 . 5 x 10 . B ea r in g in mind th e p r e se n c e o f
an unknown number o f sm a ll unmapped r e s t r i c t i o n fra g m en ts in
th e Xhol and P v u II d i g e s t s w hich have n o t been ta k en in to6 6
a c c o u n t , th e Kpnl (9 6 .7 x 10 ) and BamHI (9 7 * 4 x 10 ) m .w ts .
g iv e th e m ost r e l i a b l e e s t im a t e s . T hese s i z e s a r e i n a cco rd
w ith t h a t o f 98 x 10^ r e p o r te d by W ilk ie ( 1 9 7 6 ) . The m .w t.a ■
o f HSV-2 DNA, from th e BamHI d i g e s t , i s 97*6 x 10 > w hich- - c.
compares w ell with the s iz e o f 98.5 x 10 reported by
C o r t in i and W ilk ie ( 1 9 7 8 ) . T h is f ig u r e in c lu d e s th r e e sm a ll
fra g m en ts w hich were r e s o lv e d by p o ly a c r y la m id e g e l
e le c t r o p h o r e s i s but n o t mapped.

- t i l -
The mean m .wt. o f th e S segm ent o f th e HSV-1 genome
i s 1 6 .8 ± 0 . 5 x 1 0 ° , and tha/t f o r HSV-2 i s 1 8 .6 x 1 0 °
(compare C o r t in i and W ilk ie , 1978: 1 7 .9 x 10 b f o r HSV-2).
On th e b a s i s o i t n e s e r e s u l t s i t seems t h a t th e two s e r o t y p e s
d i f f e r i n th e m .wt. o f S by 1 -2 x 10^ (1 5 0 0 -3 0 0 0 b p ) . I t i s
p o s s i b l e t h a t th e HSV-1 and HSV-2 L seg m en ts are o f eq u a l
s i z e , and t h e r e f o r e t h a t th e HSV-2 genome i s l a r g e r tn an t h a t
o f HSV-1 by 1 -2 x 10 i n m.wt* Morse e t a l . (1977) l i k e w i s e
m entioned t h a t HSV-2 DNA i s 3 x 1 0 ° l a r g e r i n m .w t. than
HSV-1 DNA, and R eyes e t a l . (19 7 9 ) i n d i c a t e d t h a t th e e x t r a
DNA i s i n S . The p r e c i s e l o c a t i o n o f th e e x t r a DNA i n th e
HSV-2 S segm ent ca n n o t be d eterm in ed from th e r e s u l t s
p r e s e n te d h e r e , but th e a l ig n m e n t o f r e s t r i c t i o n s i t e s
con c lu d ed from th e recom b in ant a n a l y s i s summarised i n F ig u re
C 2.13 s u g g e s t s t h a t th e major p r o p o r t io n i s l o e a t e u i n
HSV-2 BamHI 1 i n th e m idd le o f Ug. The f u n c t i o n o i tn e
a d d i t i o n a l DNA i s unknown.
The p h y s i c a l maps a l s o p r o v id e in fo r m a t io n r e g a r d in g
th e s i z e s o f in v e r t e d r e p e a t s , which i s summarised i n Table
C l . 1 3 . These r e s u l t s are i n agreem ent w ith p r e v io u s ly
p u b l is h e d e s t i m a t e s f o r HSV-1 and HSV-2. The s i z e o f
HSV-1 TRg ( i . e . c a ) has s in c e been more a c c u r a t e l y e s t im a te d
a s 6720 bp (m .w t. 4 .3 7 x 1 0 ° ) , from d e t a i l e d r e s t r i c t i o n
mapping and n u c l e o t i d e se q u e n c in g e x p e r im e n ts ( S e c t i o n 3
o f R e s u l t s ; M-J. M urchie, p e r s o n a l com m u n ica tio n ) .

Tables Cl.1-01.5
M o lecu la r w e ig h t s and r e l a t i v e m o l a r i t i e s o f
r e s t r i c t i o n fr a g m e n ts o f HSV-1 DNA produced by
Kpnl, BamHI, Khol and P v u I I , and o f HSV-2 DNA
produced by Bamli l .
A l l r e s t r i c t i o n fra g m en ts o f a d i g e s t co u ld n o t
be r e s o lv e d on a g e l o f s i n g l e c o n c e n t r a t io n , so
r e l a t i v e m o l a r i t i e s o f l a r g e r fra g m en ts were
e s t im a te d from lo w er c o n c e n t r a t io n g e l s , and
th o s e o f s m a l le r fra g m en ts from h ig h e r c o n c e n t r a t io n
g e l s . The r e s u l t s were l in k e d u s in g fra g m en ts o f
in t e r m e d ia t e s i z e w hich were r e s o lv e d on both
g e l c o n c e n t r a t i o n s .
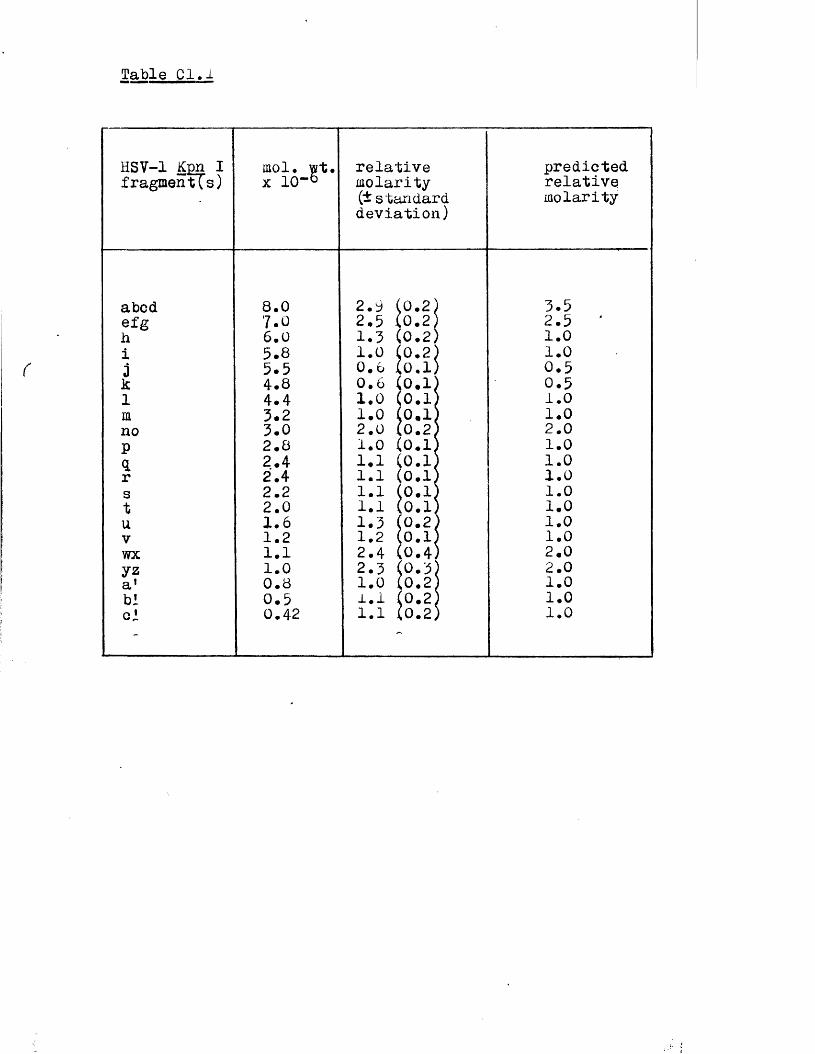
Table Cl.l
HSV-1 t o I fr a g m e n t(s )
mol* w t. x 10“ 6
r e l a t i v e m o la r ity (± stan d ard d e v i a t i o n )
p r e d ic t e dr e l a t i v em o la r ity
abed 8 .0 2 .9 ( 0 .2 ) 3 .5e f g '7.0 2 .5 ( 0 . 2 ) 2 .5 'h 6 .0 1 .3 ( 0 .2 ) 1 .0i 5 .8 1 .0 ( 0 . 2 ) 1 .0d 5 .5 0 . 6 ( O . l ) 0 . 5k 4 .8 0 . 6 ( 0 . 1 ) 0 .51 4 .4 1 . 0 ( 0 . 1 ) 1 .0m 5 .2 1 .0 ( O . l ) 1 .0no 3 .0 2 . 0 ( 0 . 2 ) 2 .0p 2 . 8 1 .0 ( O . l ) 1 .0q 2 .4 1 .1 ( O . l ) 1 .0r 2 .4 1 .1 ( O . l ) 1 .0s 2 .2 1 .1 ( O . l ) 1 .0t 2 .0 1 .1 ( O . l ) 1 .0u 1 . 6 1 .3 ( 0 . 2 ) 1 .0V 1 .2 1 .2 ( O . l ) 1 .0wx 1 .1 2 . 4 ( 0 .4 ) 2 .0yz 1 .0 2 .3 ( 0 .3 ) 2 .0a 1 0 . 8 1 .0 ( 0 .2 ) 1 .0bl 0 . 5 l . l ( 0 .2 ) 1 .0c l 0 .4 2 1 .1 ( 0 . 2 ) 1 .0

Ta b le C l . 2
HSV-1 BamH I f r a g m e n t ( s )
m ol. wt, x 10“ 6
r e l a t i v e m o la r ity (± stan d ard d e v ia t io n )
p r e d ic t e dr e l a t i v em o la r i t y
abcdefghiJk1mnopars tuvwxyza'b!c ld!e 1£ ’g 1h i .i l<j~
1*mln l
8.06.26 .05 .65 .45 .0 4 .94 .8 4 .34 .23 .93 .53 .33 .22 . 42 . 22 . 0 1.81 .71 . 5 1 .3 1.1 0 .9 0 0 .6 3 0 . 5 8 0 .5 5 0 .5 1 0 .4 8 0 .4 5 0 .3 5 0 .1 4 0 .1 3 0.10 0 .0 6
1.01.01.01.01 .01 . 01 .01 . 01 . 01 . 01 . 01.01 . 01.01.02.01.02.02 . 01 . 04 .01.0 1.0 1.0 1 . 0 1.0 2 . 0 1.0 1.0 1.0 1.0 1.0 2 . 0 1.0

Table Cl. 3
HSV-1 Xho I f r a g m e n t ( s )
m ol. w t. x l c T 8
r e l a t i v em o la r ity(-s ta n d a r dd e v i a t i o n )
p r e d ic t e dr e l a t i v em o la r ity
a 1 0 .0 0 .7 ( 0 .1 ) 1 . 0b y.o 0 .9 ( 0 .1 ) 1 . 0c 6 .6 0 .7 ( 0 .0 ) 0 .5CL 6 .2 0 . 5 ( 0 .0 ) 0 .5e 5 .7 1 .3 ( 0 .0 ) 1 . 0f 5 .2 1 . 1 ( 0 .1 ) 1 . 0
g h i 4 .9 2 . 5 ( 0 . 2 ) 2 .5i 4 .6 0 . 6 ( 0 .0 ) 0 .5k 4 .0 1 .2 ( 0 . 1 ) 1 . 0run 2 .5 1 .8 ( 0 .2 ) 2 .00 2 .3 1 .2 ( 0 .1 ) 1 . 0p q r s t 2 . 1 4 .5 ( 0 .7 ) 5 .0u 1 .8 1 .0 ( 0 .1 ) 1 . 0
V 1 .5 1 .0 ( 0 .1 ) 1 . 0w 1 . 4 1 .1 ( 0 .1 ) 1 . 0
xyz 1 . 4 3 .0 ( 0 .1 ) 3 .0a 1 1 .4 0 .9 ( 0 .2 ) 1 . 0b* 1 .3 0 .9 ( 0 .1 ) 1 . 0c l 1 .2 0 .9 (0 .2 ) . 1 . 0
d* 1 .0 1 .1 ( 0 .3 ) 1 . 0e ! f 1 0 .8 3 2 .2 ( 0 .1 ) 2 .0:r 1e> - 0 .7 3 1 .1 ( 0 .2 ) 1 . 0hi i ' 0 .6 2 1 .7 ( 0 .1 ) 2 .0j ' 0 .5 4 1 . 0 ( 0 .1 ) 1 . 0k I 0 .4 3 0 . 8 ( 0 .0 ) 1 . 0l l m ’n 1 0 .3 7 2 .8 ( l .O ) 3 .0o ' p i
*i.
0 .3 0 1 .4 ( 0 .2 ) 2 .0

Table 01*4
riSV-1 Pvu I I f r a g m e n t ( s )
abcdef
h1jklmnoPq r s tuvwxyza 1b i c 1die*f ' .glh*i 1 .
j ' k ' l *m’n i o lp ’q l -
r * s 1t !u*V ’
w’x 1
m ol, wt, x 10“ b
17141295 .34 .23 .83 .73 .53 .22 .72 .2 2 . 01 .91 .71 .51 . 51 .41 .4 1 .3 1 .2 5 1 .11 .0 5 0 .9 0 .8 3 0 .8 0 0 .7 0 0 .6 0 0 .4 0 0 .3 8 0 .3 5 0 .3 3 0 .3 0 0 .2 8
r e l a t i v e m o la r ity (±stan d ard d e v i a t i o n )
p r e d ic t e d r e l a t i v e m o la r ity
0 .3 ( 0 .1 )0 .5 ( 0 .1 )0 .4 ( 0 .1 )0 .5 ( 0 .1 )1 .2 ( 0 .2 )0 .9 ( 0 .1 )1 .0 ( 0 .1 )0 , 6 ( o . i )1 .1 ( 0 .1 )3 .2 ( 0 .5 )1 .1 ( 0 .1 )1 .0 ( 0 .1 )1 .0 ( 0 .1 )4 .0 ( 0 .6 )1 .3 ( 0 . 2 )1 .2 ( 0 . 2 )1 .5 ( 0 .2 )0 .9 ( 0 . 1 )0 .9 ( 0 . 2 )0 .9 ( 0 .1 )0 .9 ( 0 . 1 )2 .1 ( 0 .2 )1 .8 ( 0 .1 )1 .0 ( 0 .1 )2 .1 ( 0 .1 )1 .0 ( 0 .0 )2 .9 ( 0 .4 )3 .3 ( 0 .2 )
0 . 50 .50 .50 . 51 .01 .01 .01 .01.04 .01.0 1.0 1.04 .01.0 1.0 1.0 1.0 1.0 1.0 1.0 2 . 0 2 . 0 1.0 2 . 0 1.03 .03 .02 .0 2 . 0 1.0 1.0 1.0 2 . 0

Table 01.5
HSV-2 JBamll I fr a g m e n t (s )
m ol. w t. x 10~5
r e l a t i v e m o la r ity ( t s ta n d a r d d e v i a t i o n )
p r e d ic t e dr e l a t i v em o la r i t y
abcdef6h i j k 1 m n op q.rs tuvwxyza 1b 1c 1 a 1
h'
J'k! l 1m* .
8 . 05 .24 .94 .74 .33 .73 .33 .12 .8 2 . 62 . 42 .32 .2 2 . 01 .9 1 .81 .71 .7 1 .61 .51 .3 1 .2 1 .1 51 .0 5 0 .9 6 0 .8 0 0 .7 0 0 .5 8 0 .4 7 0 .3 9
2 . 01 .01 .01 .01 .01 .03 .01 .0 1.0 1 .0 1.0 1.03 .01 .0 2 . 0 1 .0 2 . 0 1 .0 2.0 1 .0 1 .0 2 . 0 1 .0 1.0 2.0 1.0 1.0 1.0 2 . 0 2 . 0

Table Cl.6
D i g e s t i o n o f HSV-1 Kpn I fra g m en ts by o th e r
r e s t r i c t i o n e n d o n u c le a s e s . The summarised d a ta
a r e ta k en from F ig u r e s C l . 7 - 9 .
M o le c u la r w e ig h ts r e f e r to th e p ro d u ct fra g m en ts
o f r e c l e a v a g e o f p u r i f i e d Kpn I fr a g m e n ts .
L e t t e r s a r e g iv e n when a p ro d u ct fragm ent
c o r r e s p o n d s to a fragm en t produced by th e
r e s t r i c t i o n o f HSV-1 DhA w it h th e r e c l e a v i n g
e n d o n u c le a se a l o n e .
x t h i s fragm en t i s n o t r e c le a v e d
* t h i s fragm en t i s r e c l e a v e d , but t h e p ro d u ct
s i z e s a r e n o t known

nSo
■pHCtiVatboa>s
»■pC©
H
cdP*
K
O cn • •-d- cm -d-
VO HH X VO CNi 0 |
VO
H > |
»l 3lPtfiA
X ^ | H
O
CM
o
•5f
orH
iHcl
O n
oin
X cn
O
X OJ K
at
Ch•pop*0o(4a
©ata)
Ho33o
•oa0*0
-H►at©HO©u
HH
tjDm
o
X X X
CM
CM
VO
CM
00X X - t f
ocm
iH
r>-
oi n
CM X X
H
«oa
W
o o • ♦-d- cn H
i—( |cm oo « «
l A O v o o • •
NO -d" CM 0 | X X VO X
CM
Ml M\
00oOn
«O
c |
O
X cm
HHH
TJC
•H«
j - o |«
movo • •* t> m o XXX
VO
CM
1T\cn
CM
cn
voo
S|
inoOn
cn x x x
at&X
vo no • • •_3- m rH
in cm -d-
X vo -d- c n X X X
CM
oc
x cn x x
•d da a ► Xas ' ©■H *0 O g © © fc JO as O o TJ c o
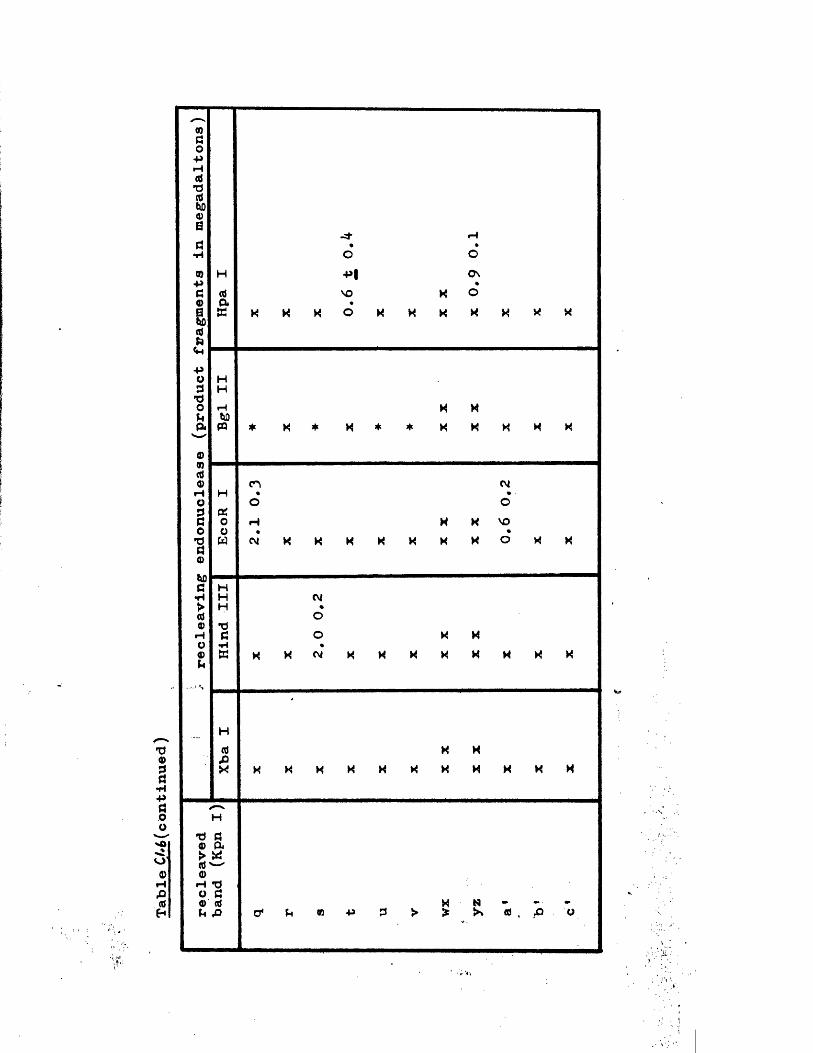
e CJ
+ 6 (c
ont
inue
d )
<oSo•pHflj•OctftD4)a
©U<H+>oTlouft
0)<n©©rlO33o*3a©toa•H>©©iHa©u
•it
o+*1VO©o
oON
K o
c i%orl<vl H
cv9 ■
O
X VO
K O M
WOo
K cvi K M
©
H
•d d © ft > US ds-' ©H *0a 3 © © U P
N>»

Table 01,7
D i g e s t i o n o f HSV-1 BamH I fra g m en ts T?y o t h e r
r e s t r i c t i o n e n d o n u c le a s e s . The summarised d a ta
a re from d o u b le d i g e s t i o n and r e c le a v a g e
e x p e r im e n ts . M o le c u la r w e ig h ts r e f e r to th e
p r o d u c t fr a g m e n ts o f r e c le a v a g e o f i s o l a t e d
BamH I f r a g m e n ts . L e t t e r s a r e g iv e n when a
p r o d u c t fra g m en t co rr e sp o n d s to a fragm en t
produced by th e r e s t r i c t i o n o f HSV-1 DNA w ith
the r e c l e a v i n g e n d o n u c le a se a lo n e .
x t h i s fragm en t was n ot c le a v e d
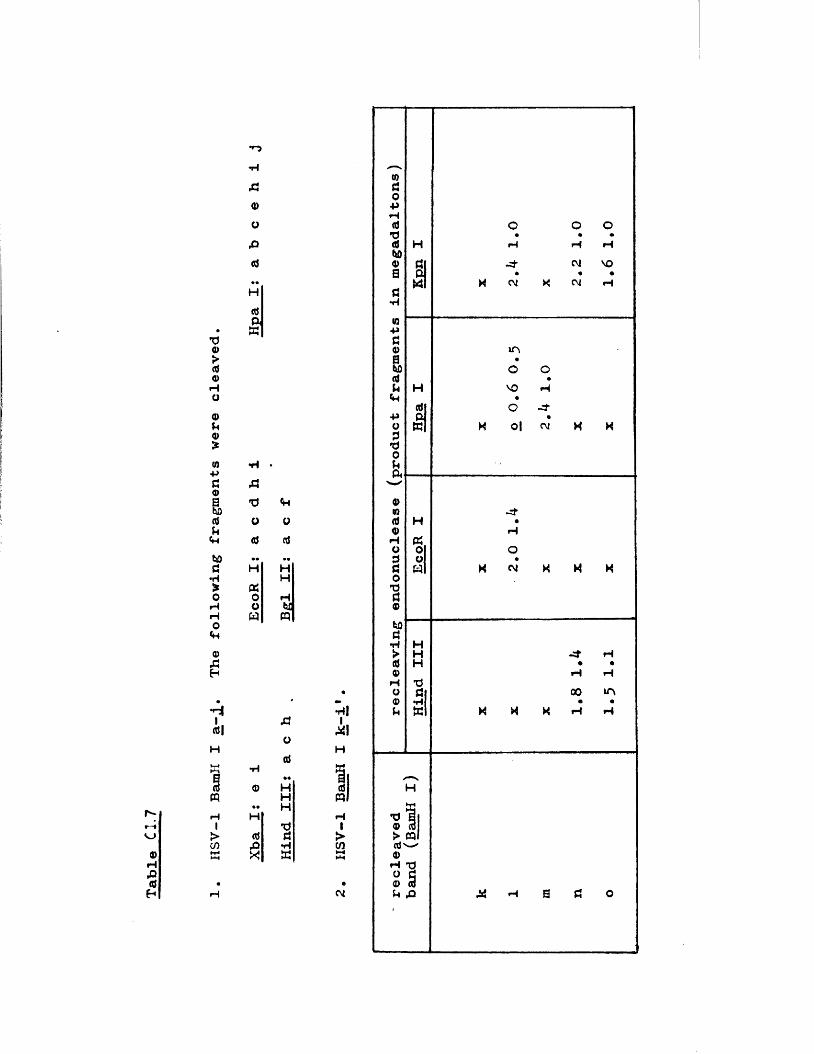
Table
CU
tf>a0■pHcd 000
•d • • •<d H l l ldo© 3( -3" CM SOS P| • • •
wl H CM X CM HG•H(0•Pa© »na •do o oa •u H VO H•
cdl o ^-p •o wl H Ol cm H X
•d0uQi
©«<d H •© HHO od •a Hi K <M K M H0■da«doa•H H> H Hflj H • •© H HH •dO at 00© •H • •a wl X X X 1 1
H
•d© Cut> cql<ti—©H T>O a© ©a & k 1 m n o
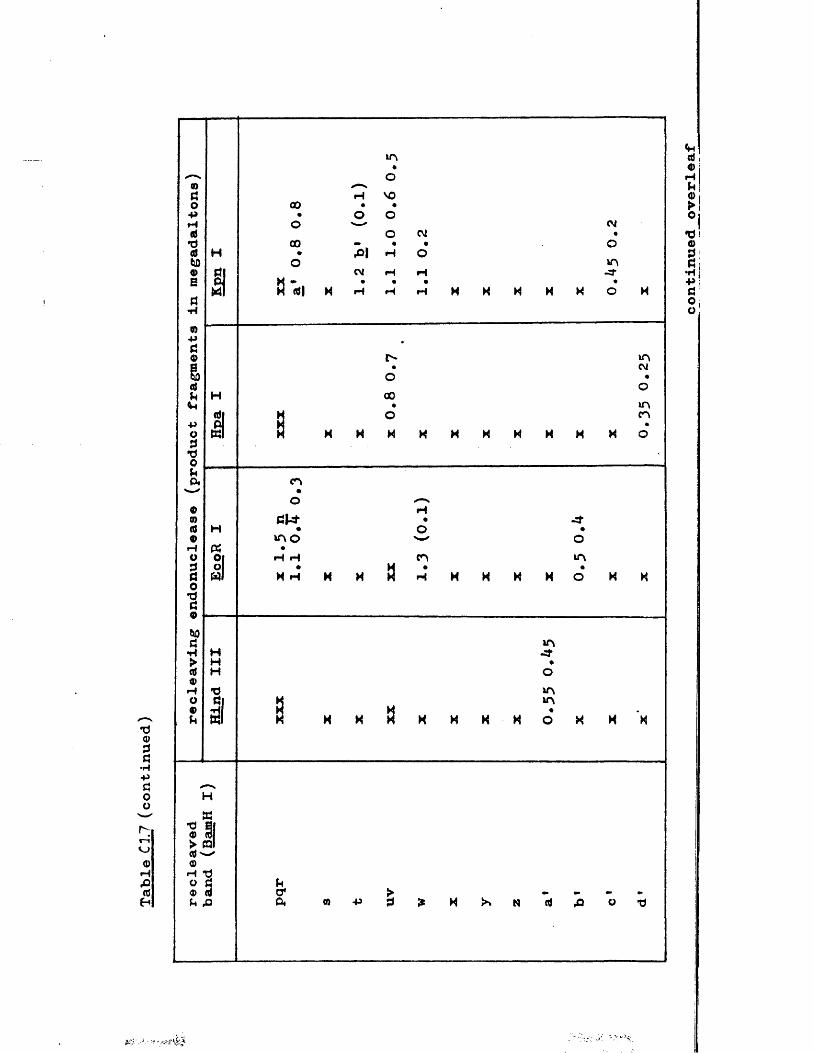
e C
l.7 (c
onti
nued
)
ndo4*rH(0•d<0toEd•Hm-pd®
<0U
+jodTJobp«
90}<09Oddo•dd®tod•H>(0<DHO•U
H
a
H
00o00o
a*.
HO
i lcm
1T\oVOooH
CM
OCM
Om-=r
(01 X X X
oooo
X X X
mCM
mcn
cn•o dPt m oH H
X H a
H
Ocn
x x
oin
X o x
a
m•if
inmX
&(0
H8
? 9> col <0"— ©H *2 dc*
Pi
■■i r'-ZS'iki
continued
over
leaf

Table
C1.7
(con
tinu
ed)
ndo■pH(4•da} H® G|B w H
wl X M X Xd•HQQ-Pa© in& c\t00 •d oH<H
<01 CM-P Pi •OH wl M H O H MPTJo(hn.
a)ma H0H «O °ld °\ Hd ml M M M Mo•dd<Ddod >-V•H H H► H «a) H o©H TfO -3“<D •HI M •U ml X X 0
j
X
-iK
•a 0|a) dl> mld^-'©rH d •*O d 110© du m e f h i
•HH
a
no>do•HV-POa<0•H
o|
I

Table Cl. 8
Summary o f HSV-1 Xho I d o u b le d i g e s t i o n d a t a .
Numbers i n p a r e n t h e s e s r e f e r t o th e number o f
m olar fr a g m e n ts w hich were c l e a v e d w i t h in a
m u lt i p l e band.

j/
Table Cl.8
secondendonuclease
Xho fragments which were cleaved in double digest
Xba I (a f h d')Hind III a e m ghi(0.5) pqrs(2) a* c*EcoR I a b c d f ghi(l.5) j mn(l)
pqrs(l) uBgl II a e f ghi(l) 1 mn(l) pqrs(3) uHpa I a b e f ghi(2) k 1 a» c* d*k e s I a b f k ran(l) o pqrs(3)
v w x y z e *BamH I a b o d e f g h i . j l m n
pqrs(3) v w xyz(2)

Table Cl.9
Summary o f HSV-1 Xho I fr a g m en ts to w hich i s o l a t e d
n i c k t r a n s l a t e d HSV-1 BamH I fra g m en ts h y b r id i s e d .
H y b r id i s a t io n o f BamH I k was n o t a s s p e c i f i c a s
f o r t h e o t h e r fr a g m e n ts , presum ably owing to
c o n ta m in a t io n by o t h e r fr a g m e n ts .

Table Cl,9
HSV-1 BamH I fragment
homologous fragments in HSV-1 Xho I digest
k (c d ghi j pqrs)1 bm 1n xyz c e io a xyzpq r c d f ghi j pqrs xyz e 1s o d pqrst f pqrsuv f pqrs j *w xyz ghi

Table Cl.10
Summary o f HSV-1 Pvu I I d ou b le d i g e s t i o n d a t a ,
lum bers i n p a r e n t h e s e s r e f e r to th e number o f
fr a g m e n ts w i t h in a m u l t ip l e band w hich were c l e a v e d .

Table Cl,10
secondendonuclease
Pvu XX bands which were cleaved in the double digest
Xba X c d f g j«k*l»(1)
Hind III c d g rst(l) x b*c’(l) d * e ' ( l )
EcoR X a b c d e f p q w y J ,k ,l ,(l)
Bgl II(bands larger than u)
a b e g h q rst(l)
Sea I a b c d e f g h l n b ,c ,(l)

Table Cl.11
D i g e s t i o n o f HSV—2 BamH I fra g m en ts by o t h e r
r e s t r i c t i o n e n d o n u c le a s e s . M o lecu la r w e ig h ts
r e f e r t o th e p r o d u c t fra g m en ts o f r e c le a v a g e
o f i s o l a t e d BamH I fr a g m e n ts , l e t t e r s a r e g iv e n
when a p r o d u c t fragm en t c o r r e sp o n d s to a fragm ent
produced by th e r e s t r i c t i o n o f HSV-2 DNA w ith
th e r e c l e a v i n g e n d o n u c le a se a l o n e .
x t h i s fragm en t was n o t r e c l e a v e d .
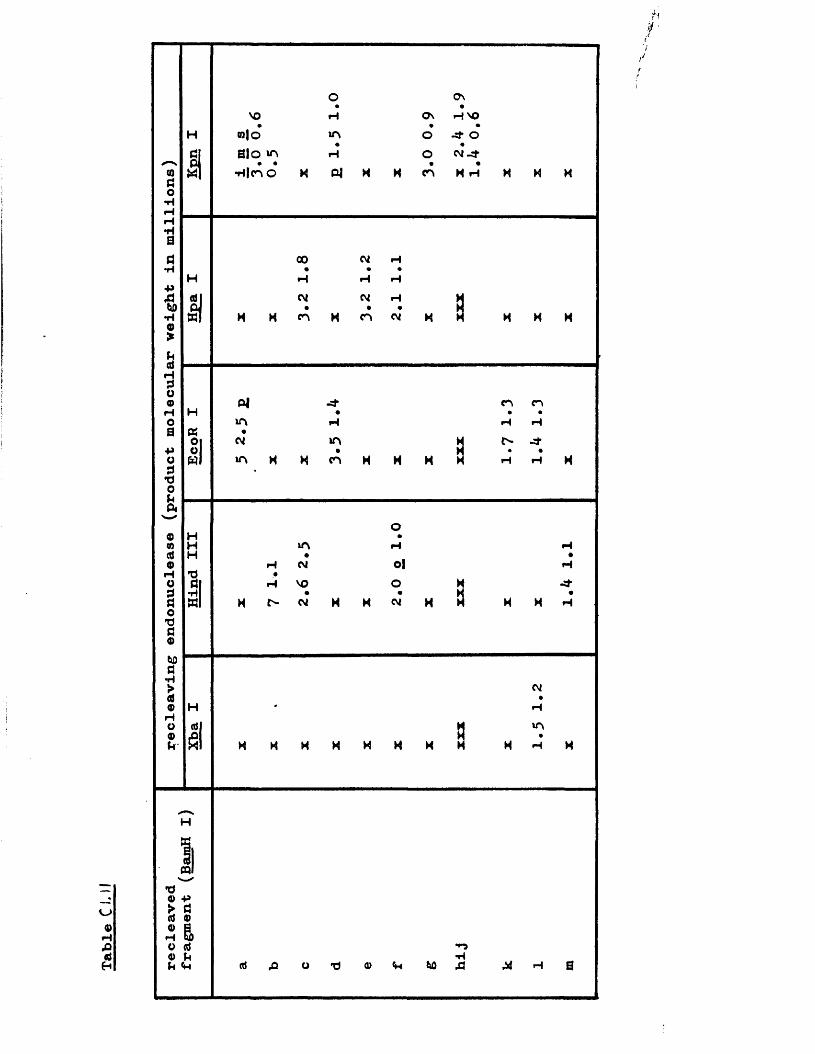
a)3o•H
H
3VO
n|o Bio »n •Hlcno
o Os• •H OS
9 H VO •ir\ O -=f O• «H O• CM-tf•P I H H cn H H H H
•H0a
•H
«P
•H9>Uat
o©
HO0■MO3*douPi
9<099
HO33o•d3©
fcjD3•rl>99Ho©U
300 c\* H
• • •H H H
CM CM H• • •
c n H c n CM
PIir\
<\*HlOcn
c n cn
-3-
HHH-d3•H«
Hr -
lT\CM
VO
CNJ
OloCM
HflS8 a
Ci
VTy
Cto •HA
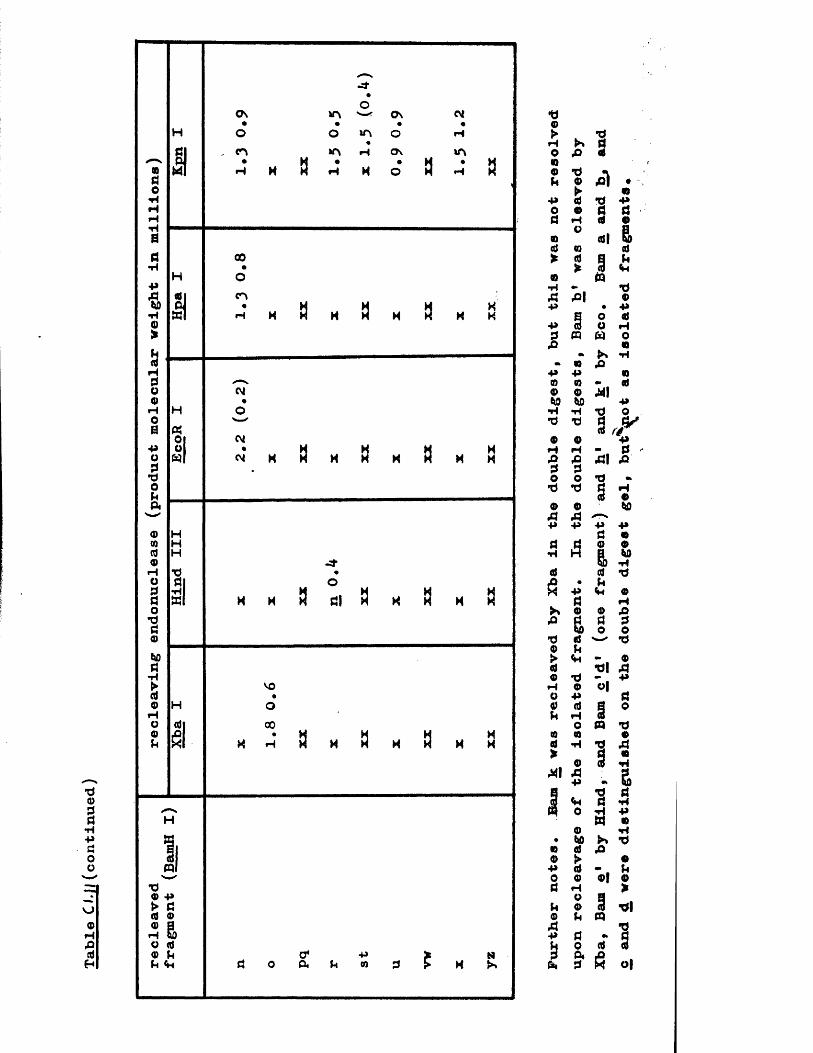
Tab
le
C/JI
(co
nti
nu
ed)
ado
• r t
HH•rtBd•rtPf t•rt©>(44Hdo©HOa+>od*dodft
(D(04©HO§o•dd©
ood•H►cd©HO©
H
H
II
H
Os
rs
V\
-=f•
OOs CM
• « •O ITv o iH
»fN•
H OS• • »4 •
H H O ►4 r t a
00
ors
a
cm
o(VI
cm a h a a
a a> a n a a
SO
00 a
a* ft
p0) N
•d®>HOo®U
POd
x>
•d®
®
00d a 4 *e•rt
A-P 43|
-p 4 d ®jo •k- e
p p to n ® ® to to ■rt *H •d Tl© ©
•dd4
A *e•d p d d 4 ®a) | 6>
48
•d ® p 4 H Oa
CO
oo«
4>
*1Po
JOdo•d®Apd■rt4
43•d4}>4®
'do•d©Ap
pd©
&4h<H•d rt ©
O P © 4h H
Oa a 4 *H
441 Ap
•d
- d -431 43
4 •to
p pd •© ©I toto *rt 4 *d h<H ©
H© 43d d o o— 73
- , ©•dl Ap
« « 9 °« *d •*d 44 a
» S)
1 Vi dO •rt P
• ©to
H►>
4•rt*da 4 43© >* •p 4 •* u
0 © ©1 ©d rt >d
O© 9 *1© U CP
jdp d 4» 19ft
0ftd
4 4Ol

Table Cl>12
Summary o f HSV-2 BamHI fr a g m en ts to w hich i s o l a t e d
n ic k t r a n s l a t e d HSV-2 Hind I I I / BeoRI fr a g m e n ts
h y b r i d i s e d . The HSV-2 Hindi I I / EcoRI map i s from
W ilk ie e t a l . (1 9 7 9 b ) .

Table 01.12
HSV-2Hind/Bcofragment
homologous fragments in HSV-2 BamH I digest
1-6>
ab o e f g hij m o pst u vw yz a* e* h'
7-8 ab d hij n q9 ab k v10-11 c f g p vw y z
12 d hij i»13 g p u vw y z a * e 114-15 c g p u vw x yz a*16 1» c *d*17 1 e * f«18 f k19 st20 r21 b*22 ab23 f
0 0.2 0.4 0.6 0.8 1.1 1 1 1 1, li
l l l II i II i it i it • i i i i10 15 8 12 2019 7 22 6 5 24 9 18 23 11 13 16 21 14

Table Cl.15
Estimates of upper and lower limits for molecular weights of the repeat sequences of HSV-1 and HSV-2.Molecular weights are given in millions and the relevant fragments are quoted.
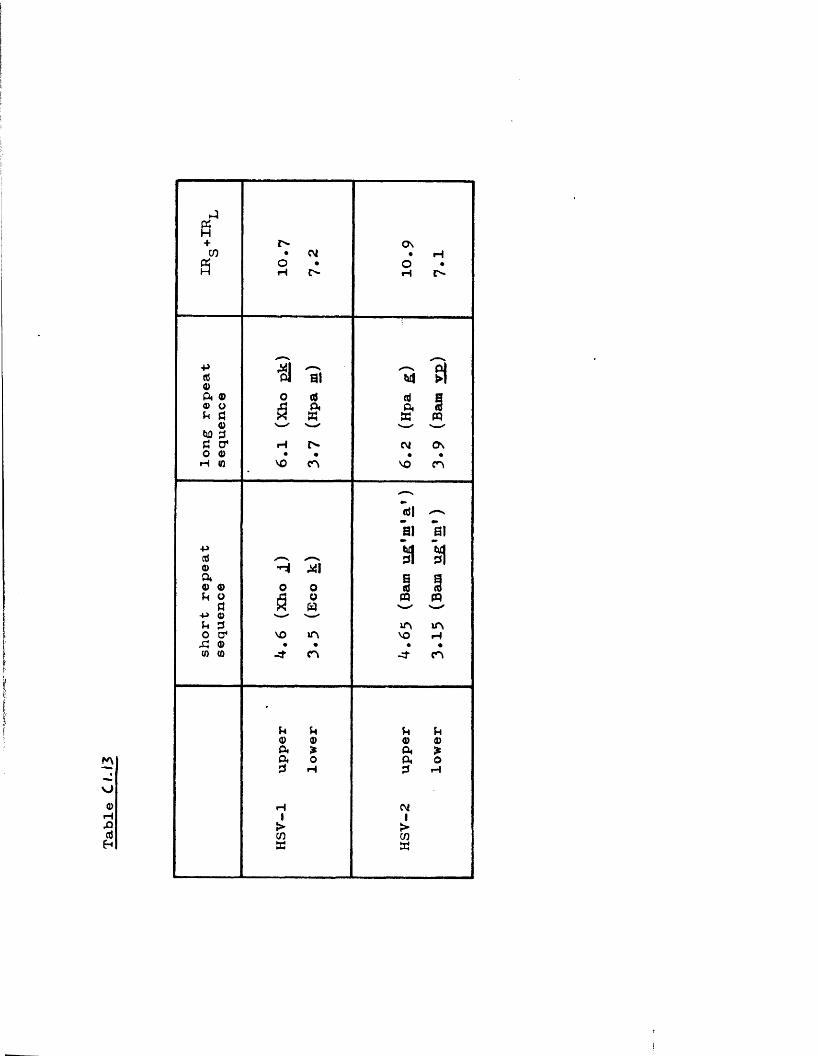
ftH+ r On
CO • CM • Hft o • o •H iH H C>*
•P S \ y,-^ Qfl5 0) ol 51 04 Mft © 0 (0 <d 5© o jq ft ft ©u c >< B B «© V Voo 3ti a* H c\i ONo © • • • •H (0 vo vo ca
— sr tl
91 S I+> *A fcdcd 51 51<1) - 4 * 1a 9 s© © o 0 4 (0fn O fl o a PQ
a W H w V-/4J © v_Ph 3 IT\ XAo a* VO IT\ vo rHB © • • • •(0 © ca CA
u u u© © © ©A > ft0< o ft O3 H 3 H
H e*1 1f>co COB B

llgure 01>1
M o le c u la r w e ig h t c a l i b r a t i o n o f HSV-1 DNA r e s t r i c t i o n
fr a g m e n ts (1 -1 0 x 10^ d a l t o n s ) .
1 . Lambda DNA c le a v e d w ith Hind I I I .
2 . Lambda DHA c le a v e d w ith KcoK I .
3 . HSV-1 DMA c le a v e d w ith Xho I .
4 . HSVr-1 DNA c l e a v e d w ith BamH I .
5 . HSV-1 DNA c l e a v e d w ith Kpn I .
6 . HSV-1 DNA c le a v e d w ith Hpa I .
Ihe 0.5>» a g a r o se g e l c o n t a in in g e th id iu m bromide
was photographed under UV i l l u m i n a t i o n . M o lecu la r ]
w e ig h t s (x 10"6 ) o f lambda DNA fr a g m e n ts a r e i n d i c a t e d j
(Thomas and D a v is , 1975; Murray and Murray, 1 9 7 5 ) .
1 2 3 4 5 6
13.7 -
6 .4 -
4 .7 -4 .3 -3.7 _ 3.5 “3 .0 -2.9 -
2.1 -
1.6 -1.4 -

Pigure Cl.2
M o le c u la r w e ig h t c a l i b r a t i o n o f HSV-1 and HSV-2 DNA
r e s t r i c t i o n f r a g m e n ts (Q .1 - 1 * 0 x 10 d a l t o n s ) .
M. J& 174 DNA c l e a v e d w i t h H i n d l l .
1 . HSV-tl DNA c l e a v e d w i t h BamHI .
2 . HSV-1 DNA c l e a v e d w i t h X h o l .
3 . HSV-2 DNA c l e a v e d w i t h BamHI.
)&C174 DNA was ^ P - l a b e l l e d by n i c k t r a n s l a t i o n and
HSV-1 and HSV-2 DMA w ere 32P - l a b e l l e d i n v i v o .
E l e c t r o p h o r e s i s was c a r r i e d o u t on a 1 . 7 # a g a r o s e
g e l w h ich was d r ie d t h e n a u t o r a d io g r a p h e d .
M o le c u la r w e i g h t s (x 10 o f 0X174- DNA fr a g m e n t s
a r e i n d i c a t e d .
0.681-
0.501-
0.396-0.322-0.255-0 . 220 -0.190-
0.137-i
0.681
0.501
0.3960.3220.255
- 0.220-0.190
0.137

Figure Cl. 5
K e s t r ic t io n p r o f i l e s o f HSV-1 (u p p er) and HSV-2
DNA (lo w er ) w hich were P - la b e l l e d i n v iv o .
E le c tr o p h o r e s is was c a r r ie d ou t on 0 .5 # (upper4 ^
p o r t io n s ) and 1 .2 # a g a r o se ( lo w er p o r t io n s ) .4 ^
Bands a re in d ic a te d o n ly f o r th o se d i g e s t s f o r w hich
r e s t r i c t i o n maps are a lr e a d y known, o r were
deduced i n t h i s work.

EcoR
1
Bgl
II
Hpa
1
Kpn
1 ZIm Xh
o 1
cf• I 9
II IB = ab
cde — a— b9hi ® J j — f a d — a
Jk1
— k1 ----i 'j = § S
■ h P . III!
m— i f— m «• jk1- k Jk — n — m Jk
1 o nm *mn P — no — n i
m P9r■ ■ or s[
s topq
_mno- P t s
o
— n
u
- - V— wx
st
— uv— w
u__ Vw • xyz a'
b— c
o U — VZ V a
ab d
— e' f
cdghi'
bhf9i
ic' k
j I'm'q
n'op
%cd
f9fh
oSsw*Vab'c'de
-&•Amno'pqr . s t
I I

Figure Cl«4
R e s u lt s o f r e c le a v a g e o f i s o l a t e d HSV-1 X b a l,
H in d III and EcoRI r e s t r i c t i o n fragm en ts by KpnI .
N ick t r a n s la t e d fragm en ts were c le a v e d and th e
p r o d u cts e l e c t r o p h o r e t ic a l ly se p a r a ted on 0 . 5 %
(upper p o r t io n s ) and 1 *5# a g a r o se g e l s ( lo w er
p o r t io n s ) , w hich were th en d r ied and a u to r a d io
graphed . The i d e n t i t y o f the p a ren t fragm ent
i s g iv e n a t th e to p o f each tr a c k , and p o s i t i o n s
o f HSV-1 KpnI fragm en ts are in d ic a te d in th e
d im en sion o f e le c t r o p h o r e s i s . C leavage p r o d u cts
are marked to th e r ig h t o f each t r a c k .

m
i i
£9
CCcuUJ
5 -x (0
i
f t
i9
I II 1 I I35—— Jt —I t
III II I 1£ 0 a sv*..
si i i M i i i l l i i i i£g a -M-
25 a?(5
— c ° P Nre a; x — Ji — E c a. crj *- 3 > > >> J. >
0)■3
<0X)X
' i i ' "i 0 i 6 ' 1A >0 0 0 0 00 0 0 0
0 0 0 o 0 0 0
ooo 0

Figure Cl,5
R e s u l t s o f r e c le a v a g e o f i s o l a t e d HSV-1 B g l l l
and Hpal r e s t r i c t i o n fr a g m e n ts by KpnI ,
D e t a i l s a r e g iv e n i n th e le g e n d t o F ig u r e C l . 4 .

Figure Cl.6
Combined r e c le a v a g e d a ta o f t h e HSV-1 genome f o r
Kpn I . The s c a l e i s o f m o le c u la r w e ig h t (x 1 0 “ ^ ) .
HSV-1 Kpn I fr a g m e n ts p rod u ced by r e c le a v a g e w ith
Kpn I a r e l i s t e d u n d er ea ch p a r e n t fr a g m e n t , a s
w e l l a s th e m o le c u la r w e ig h t s (x 10~^ ) o f r e c le a v a g e
fr a g m e n ts n o t c o r r e s p o n d in g to HSV-1 Kpn I fr a g m e n ts .
The sym b ol □ i n d i c a t e s fr a g m e n ts n o t c le a v e d by
Kpn I . R e c le a v a g e p r o d u c ts o f j o i n t fr a g m e n ts a re
shown on th e r i g h t . The Kpn I map d ed u ced from
t h e s e d a ta i s shown a t th e b o tto m .
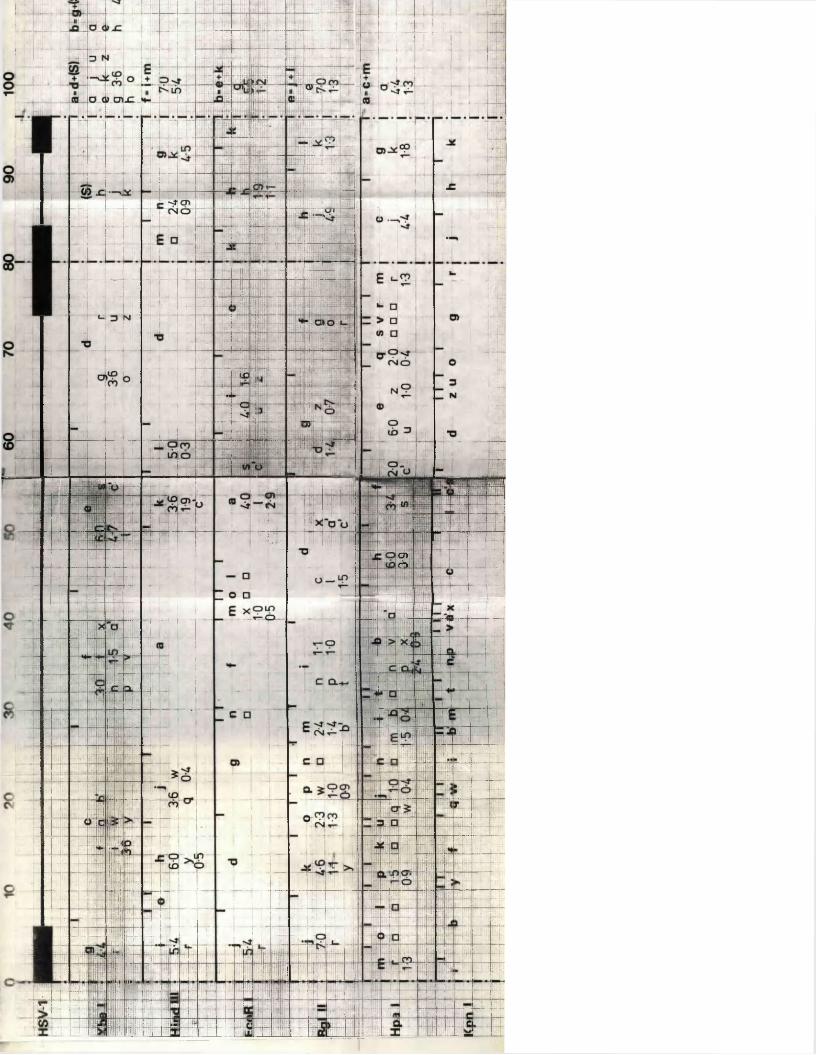
cn ' r~> ,
1—i-rrt f
^

Figure 01.7
D ou b le d i g e s t s o f i n v iv o -*2P - l a b e l l e d HSV-1 DNA.
1 . Xba I .
2 . Xba i/K p n I .
3 . Kpn I .
4 . Hind I I I /K p n I
5 . Hind I I I . '
6 , EcoR I .
'7. EcoR I/K pn I .
8 . Kpn I .
9 . i t e l II/K p n I .
1 0 . B g l I I .
1 1 . Hpa I .
1 2 . Hpa 1 / Kpn I .
1 3 . Kpn I .
E le c t r o p h o r e s i s was c a r r ie d o u t on a 0 . 6 # a g a r o s e4
g e l w h ich was th e n d r ie d and a u to r a d io g r a p h e d .
HSV-1 Kpn I r e s t r i c t i o n fr a g m e n ts a r e i n d i c a t e d .
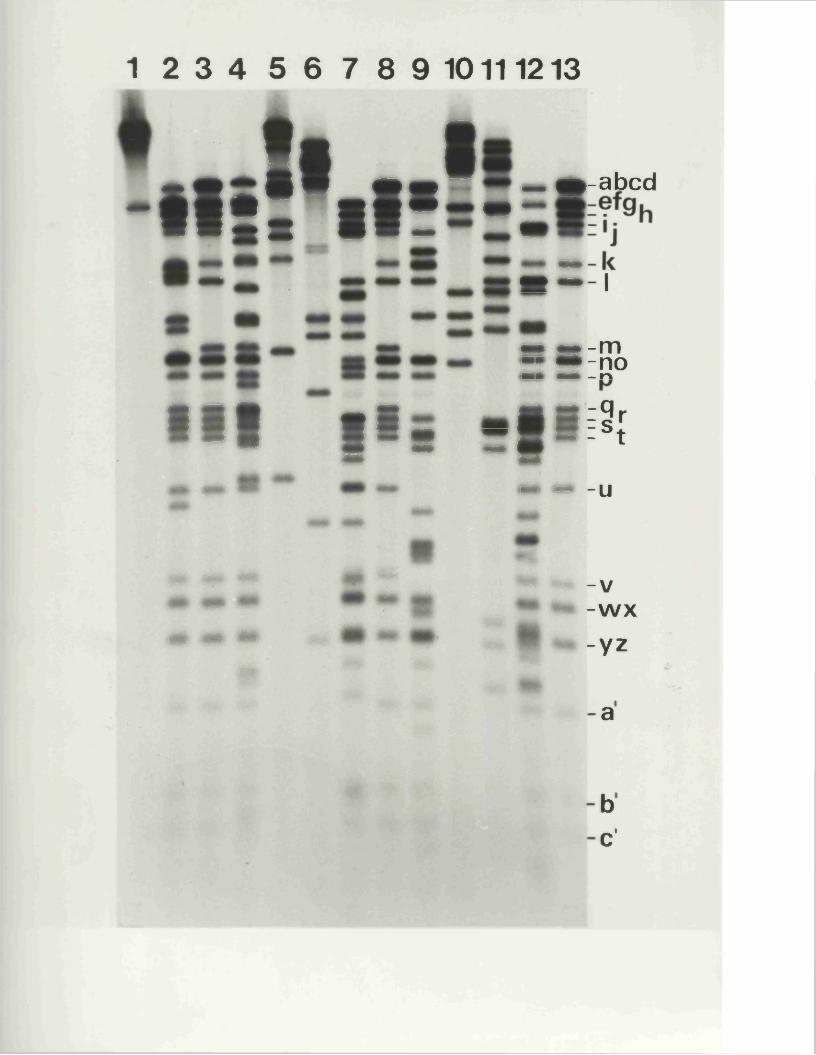
1 2 3 4 5 6 7 8 9 10111213
- • I s ! 1 ! ! ? * - ; f p - a b c d
m Z \
i n i i s r
mm K - m
r r S i r0
z ^ r = s t! i
•' — -U
m * »
-v-wx-yz
-a
b
c1

Figure Cl ♦ 8
C le a v a g e o f HSV-1 Kpn I r e s t r i c t i o n fr a g m e n ts
w ith v a r io u s r e s t r i c t i o n e n d o n u c le a s e s .
H ick t r a n s la t e d fr a g m e n ts w ere c le a v e d and th e
p r o d u c ts e l e c t r o p h o r e t i c a l l y s e p a r a te d on 1 #
a g a r o s e g e l s , w h ich w ere th e n d r ie d and a u to r a d io
g r a p h ed . The Kpn I fr a g m e n ts c le a v e d a r e
in d ic a t e d a t th e t o p * o f th e t r a c k s , w ith th e
r e c l e a v i n g e n d o n u c le a se a b o v e ea ch p a n e l . The
m arker d i g e s t o f th e r e c l e a v in g e n d o n u c le a se i s
in d ic a t e d i n th e d im e n s io n o f e l e c t r o p h o r e s i s .
H ot a l l r e c le a v a g e s a xe show n, b u t a l l th e d a ta
h a s b een in c lu d e d i h T ab le C l . 6 .
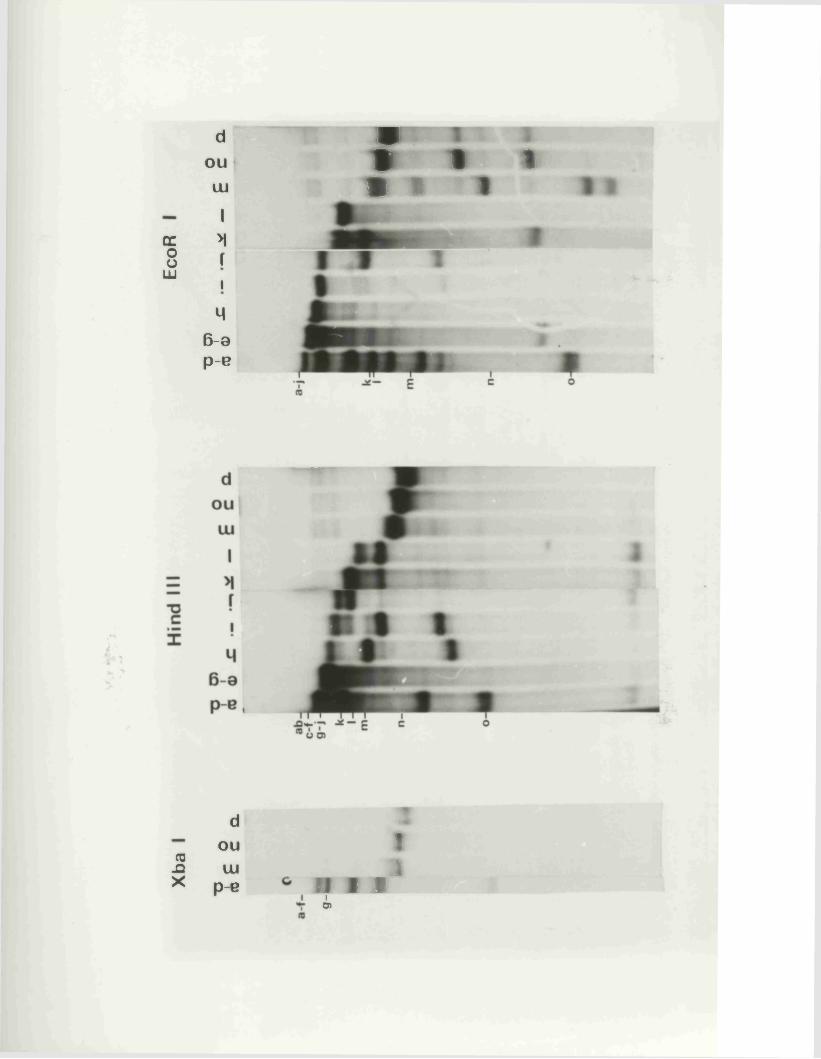
ccooLU
do uLU
I>1ri
M6 -8p-B
m
l i tB 1 1
■
CO-QX
do uLU
P-B ■ ■ aIi
To>

Figure Cl,.9
C le a v a g e o f HSV->1 Kpn I r e s t r i c t i o n fr a g m e n ts w ith
v a r io u s r e s t r i c t i o n e n d o n u c le a s e s . D e t a i l s a r e
g iv e n i n th e L egend to F ig u r e C l . 8 .
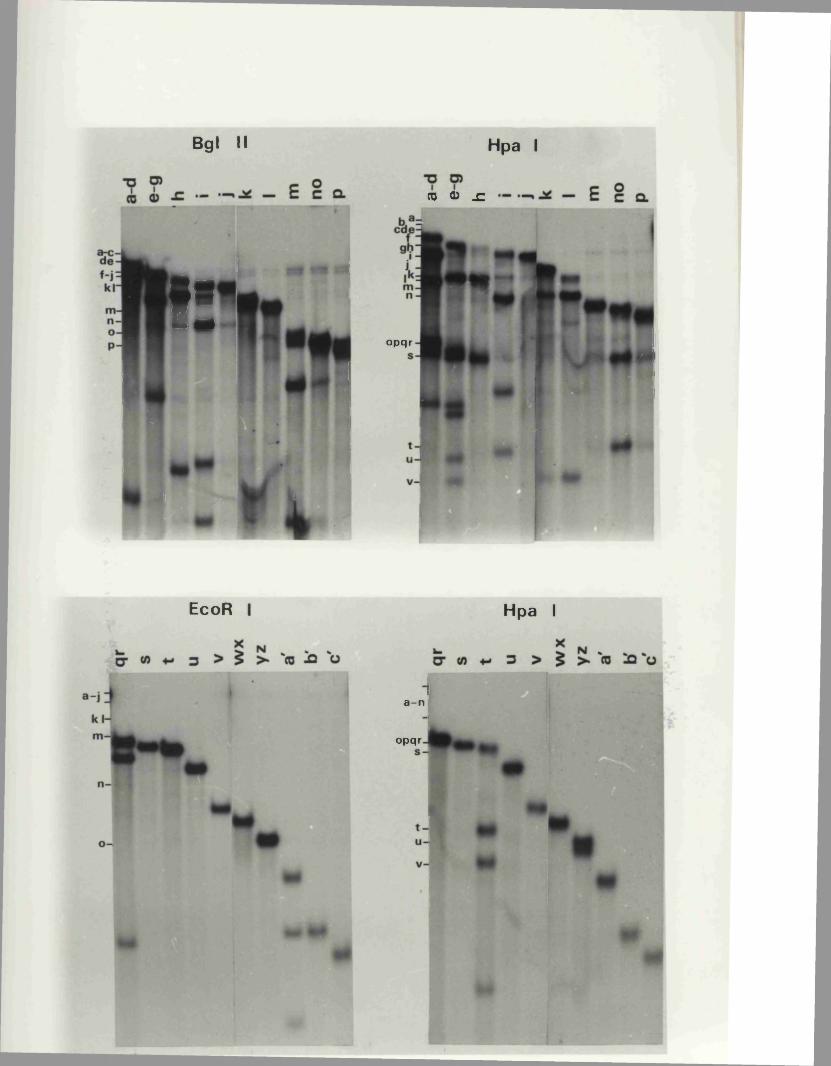
Bgl II Hpa I
CU <D _C
I !opqr
.
" •
EcoR I Hpa I
a-n
opqr

Figure 01,10
C ro ss-" b lo t o f HSV-1 Hpal a g a in s t n ic k t r a n s la t e d
HSV-1 KpnI . The u n la b e l l e d d im e n s io n i s
in d ic a t e d by th e m arker s t r i p t o th e r i g h t , and
th e l a b e l l e d d im e n s io n a c r o s s th e t o p . F a in t
s p o t s n o t c o r r e s p o n d in g t o KpnI bands a r e due
t o some fr a g m e n ts r e s u l t i n g from s l i g h t
u n d e r d ig e s t io n o f th e DHA.

— O f- u.I cc-Q O s - O) — — — fc C o
i w n 1 1 1 * i i •
> N -
5* -> -
Ca

Figure Cl.11
Summary o f HSV-1 KpnI h y b r i d i s a t i o n d a t a , u s in g* - -
th e p h y s i c a l map d ed u ced from r e c le a v a g e
e x p e r im e n ts . B e n e a th e a c h fra g m en t i s w r i t t e n
th e fr a g m e n t o f th e o t h e r d i g e s t w ith w h ich i t
h a s h om ology , a s d ed u ced from F ig u r e C l . 1 0 .—6The s c a l e i s m o le c u la r w e ig h t (x 10 ) .

iRZHfffrvuRgmnnn; innru-drawni t i i i i m f i n

Figure 01>12
D e te r m in a t io n o f th e r e l a t i v e o r d e r o f HSV-1
Kpn I n and p . The r e s t r i c t i o n d i g e s t p r e s e n t on
th e n i t r o c e l l u l o s e s t r i p s i s n o te d a t th e to p , and
th e fr a g m e n ts o f th e d i g e s t t o th e l e f t o f e a ch
p a n e l . Bands to w h ich h y b r id i s a t io n o f th e
n ic k t r a n s la t e d DHA p rob e o c c u r r e d a r e shown to
th e r i g h t o f ea ch p a n e l .
The p r o b e s w ere a s f o l l o w s . .
1 . HSV-1 ( l e f t p a n e l) o r HSV-2.DNA ( r i g h t p a n e l ) .
2 . HSV-1 Kpn I n o .
3 . HSV-1 Kpn I p .
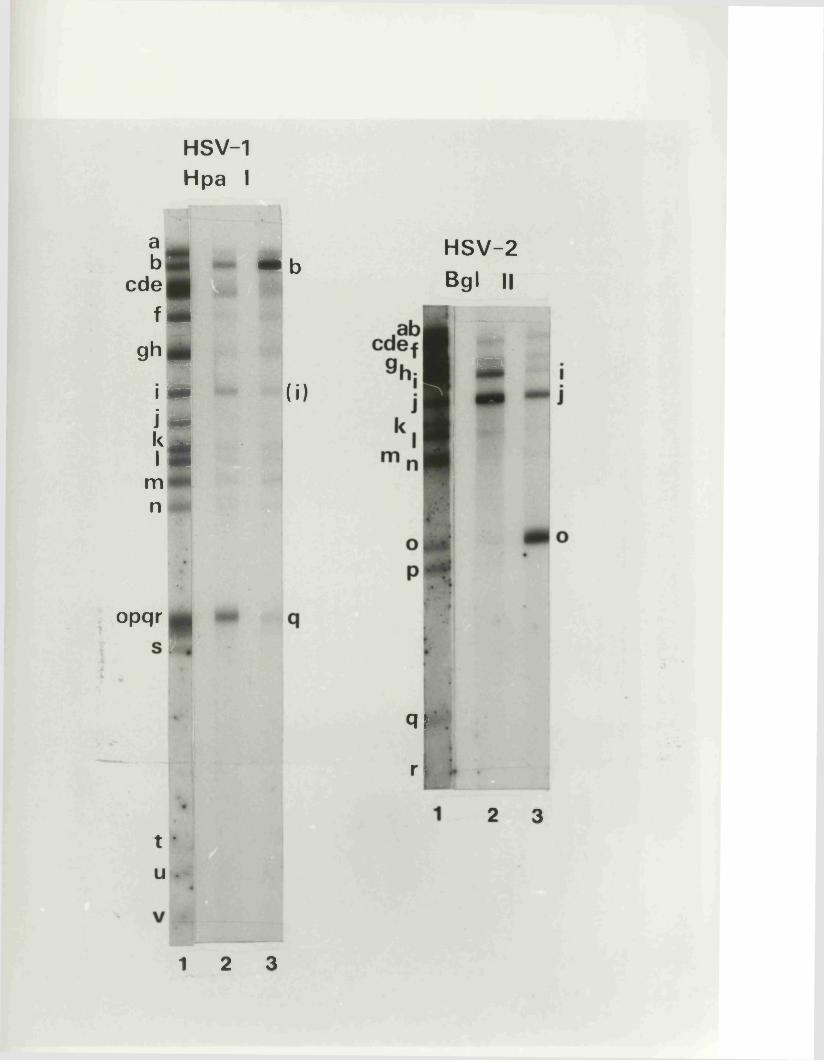
HSV-1 Hpa I
ab
cdef f c
9 h _> 3j m •<- ■I
m n
opqr
• b
(i)
t • u ■
HSV-2 Bgl II
q|
r i
L *.
2 3
1 2 3

Figure Ql.13
Combined r e c le a v a g e d a ta o f th e HSV-1 genome f o r
BamH I . The s c a l e i s o f m o le c u la r w e ig h t (x 1 0 " ^ ) .
HSV-1 BamH I fr a g m e n ts p rod u ced by r e c le a v a g e w ith
BamH I a r e l i s t e d u n d er ea ch p a r e n t fr a g m e n t , a s
w e l l a s th e m o le c u la r w e ig h ts (x 1 0 ““^) o f r e c le a v a g e
fr a g m e n ts n o t c o r r e s p o n d in g t o HSV-1 BamH I fr a g m e n ts .
The sym bol □ i n d i c a t e s fr a g m e n ts n o t c le a v e d by
BamH I . R e c le a v a g e p r o d u c ts o f j o i n t fr a g m e n ts
a r e shown on th e r i g h t . The BamH X map ded u ced
from t h e s e d a ta i s shown a t th e b o tto m .

LO LO *“iOmm r \io co <n *— o
04 on CO 04

Figure Cl.14
Summary o f HSV-1 BamH I h y b r id i s a t io n d a ta , u s in g
th e p h y s i c a l map d educed from r e c le a v a g e
e x p e r im e n ts . B en eath e a c h fra g m en t i s w r i t t e n th e
fra g m en t o f th e o th e r d i g e s t w ith w h ich i t h a s
h o m ology . . For th e BamH I map, th o s e fr a g m e n ts
u n d e r lin e d r e f e r to hom ology w ith Hpa X fr a g m e n ts ,
and th e o t h e r s r e f e r to th e Kpn I map.

.
—

Figure Cl.15
Combined r e c le a v a g e d a ta o f th e HSV-1 genome £ o r
Xho I . The s c a l e i s o f m o le c u la r w e ig h t (x 1 0 ““^*).
HSV-1 Xho I fr a g m e n ts p rod u ced by r e c le a v a g e w ith
Xho I a r e l i s t e d u n d er e a c h p a r e n t fr a g m e n t , a s
w e l l a s th e m o le c u la r w e ig h ts (x 10~^ ) o f r e c le a v a g e
fr a g m e n ts n o t c o r r e s p o n d in g to HSV-1 Xho I f r a g m e n ts .
The sym bol 0 i n d i c a t e s fr a g m e n ts n o t c le a v e d by
Xho I . H e c le a v a g e p r o d u c ts o f j o i n t fr a g m e n ts
a r e shown on th e r i g h t . The Xho I map d ed u ced
from t h e s e d a ta i s shown a t th e b o tto m .

ip- «> h
CMt O
-..CpCM^oo
cnc m *"—■
oo in> p o

Figure 01.16
Combined r e c le a v a g e d a ta o f th e HSV-1 genome f o r
Pvu I I . The s c a l e i s o f m o le c u la r w e ig h t (x 1 0 ~ b) •
HSV-1 Pvu I I fr a g m e n ts p rod u ced by r e c le a v a g e w ith
Pvu I I a r e l i s t e d u n d er ea c h p a r e n t fr a g m e n t , a s
w e l l a s th e m o le c u la r w e ig h t s (x 1 0 ““^) o f r e c le a v a g ea
fr a g m e n ts n o t c o r r e s p o n d in g to HSV-1 Pvu I I fr a g m e n ts .
The sym bol C3 i n d i c a t e s fr a g m e n ts n o t c le a v e d by
Pvu I I . K e c le a v a g e p r o d u c ts o f j o i n t fr a g m e n ts
a r e shown on th e r i g h t . The Pvu I I map d educed
from t h e s e d a ta i s sh o rn a t th e b o tto m .

>N N
to pp. CNI

Figure 01.17
Combined r e c le a v a g e d a ta o f th e HSV-2 genome f o r
BamH I . The s c a l e i s o f m o le c u la r w e ig h t (x 10” ^ )•
HSV-2 BamH I fr a g m e n ts p rod u ced by r e c le a v a g e w ith
HatnH I a r e l i s t e d u n d er e a c h p a r e n t fr a g m e n t , a s
w e l l a s th e m o le c u la r w e ig h t s (x 1 0 “ ^) o f r e c le a v a g e
fr a g m e n ts n o t c o r r e s p o n d in g to HSV-2 BamH I fr a g m e n ts .
The sym bol O i n d i c a t e s fr a g m e n ts n o t c le a v e d by
BamH I . K e c le a v a g e p r o d u c ts o f j o i n t fr a g m e n ts
a r e shown on th e r i g h t . The BamH I map d ed u ced
from t h e s e d a ta i s shown a t th e b o tto m .

C A J MVf'AN): ?:■] r-"T
HSV-2
a i L i £ |:- T-ri; -.: ~;— j b ______ 1. ; . . . . . J , , . b
! — =i-— ,L.....
I___ I__V.___ i---- A... ..i------l ...;—.—>r..w - !_*— --- X ___p 0*1 3 2 r i ’
..... .. V . I I . .2 2 r
tc?
jiIv ; :V. "q . s( :
m1 ■; -
J __
B a t II
: m i
d r p g
...... . , .„'... . □ i s
r ii o
4 - 3 1’ □
ic
■
Ii_ i 1 !
2 1 i ’I - I I . / 1 - 9 n ’I -! -i ■ •. ~- L r u- i f
i ^ i n12 2019 yHind 111/
L _ .^ L □ S1 -6 0 - 4 0 - 2
0-1
9Hind HI/ B g t l l o 0-62 - 2 0 4
7
m1 - 5 1>2
1- 1-6i 1 i i in
l i i ‘) r s l n n T b
tkuUUJ
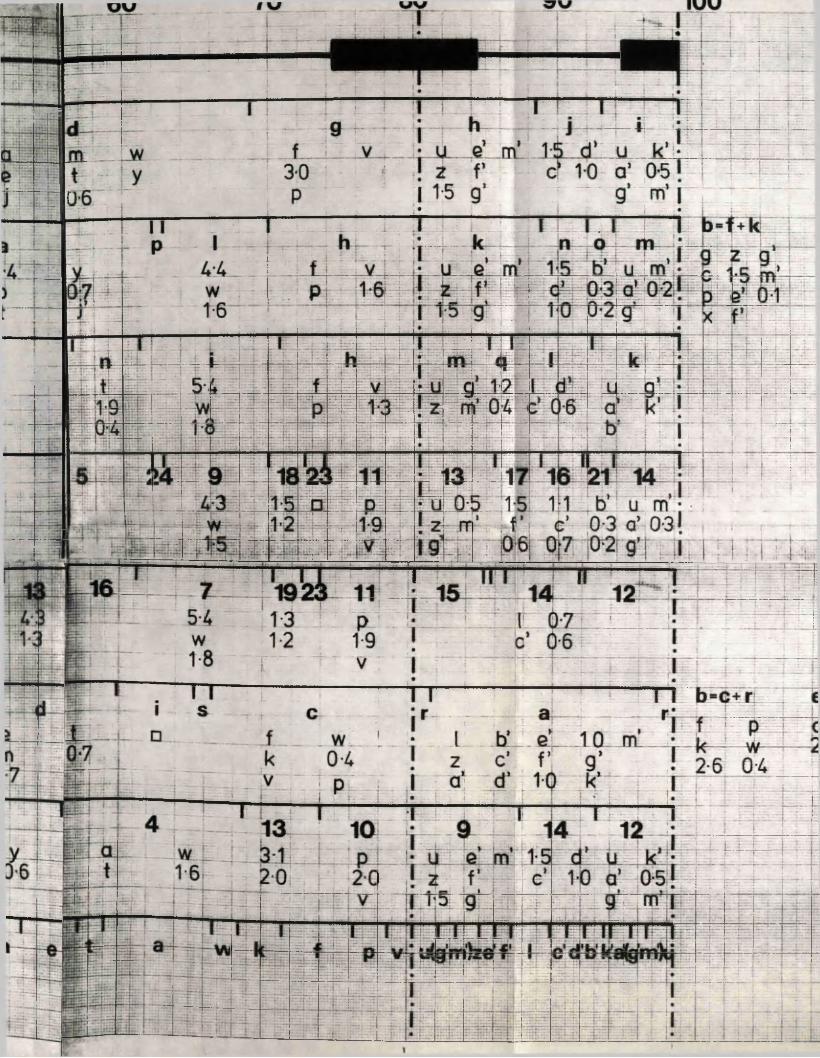

Figure Cl .18
A lig n e d r e s t r i c t i o n maps f o r HSV-1 BNA.
The s c a l e i s f r a c t i o n a l genom e l e n g t h .
The maps f o r Kpn I , BamH I , Xho I and Pvu I I
w ere d e te r m in e d a s a r e s u l t o f t h i s s tu d y ,
and t h e o t h e r s w ere th e work o f W ilk ie ( 1 9 7 6 ) .


Figure 01.19
A lig n e d r e s t r i c t i o n maps f o r HSV-2 DNA.
The s c a l e i s f r a c t i o n a l genome l e n g t h .
The map f o r BamH I was d e te r m in e d a s a r e s u l t
o f t h i s s t u d y , and th e o t h e r s w ere th e work
o f C o r t in i and W ilk ie ( 1 9 7 8 ) .

L ___
— ----------------
78
57
82
43

R e s t r i c t i o n maps f o r HSV-1 BNA a l ig n e d w it h
t h o s e f o r HSV-2 BNA. B a ta u se d t o d e te r m in e th e
b e s t a l ig n m e n t in c lu d e d r e s t r i c t i o n fr a g m e n t
s i z e s , i n t e r t y p i c h y b r i d i s a t i o n ( S e c t io n 2 o f
R e s u l t s ) , and a n a l y s i s o f i n t e r t y p i c r e c o m b in a n ts
( P r e s t o n e t a l . , 19785 Stow and W ilk ie , 1 9 7 8 ;
M arsden e t a l . , 1 9 78;. C h artran d e t a l . , 1979
and 1 9 8 0 ; S e c t io n 2 o f R e s u l t s ) .£
S in c e th e HSV-2 genpme h a3 an e x t r a 1 - 2 x 10
d a l t o n s o f BNA i n th e S seg m en t com pared w it h
t h a t o f HSV-1, th e f o l l o w i n g fr a g m e n ts h ave
b een s h o r te n e d i n th e HSV-2 m aps: Xba I J,,
Hind. I l l 1 , Ecoii I a , B g l I I 1 , Haa I ( g j ,
Kpn I &, and BamH I 1 . The s c a l e show s
f r a c t i o n a l genom e l e n g t h , and th e n a tu r e o f
j o i n t fr a g m e n ts i s shown on th e r i g h t .


S3CTI0H 2
HOBO 10 GY BiSWiSJSN HiSETBSVIHUS G3N0M3S

- 82 -
RESULTS
I n tr o d u c t io n
HSV-1 and HSV-2 sh are hom ology in ab ou t 5 0 £ o f t h e i r
DM se q u e n c e s ( K ie f f aJ a i « , 1 9 7 2 ) , w h ereas th e DM o f
p se u d o r a b ie s v ir u s (PKV) h as been r e p o r te d to be l e s s than
8 # hom ologous to HSV-1 and HSV-2 DM (B ronson e t a l . , 1 9 7 2 ) .
The p u rp o se o f th e work d e s c r ib e d h e r e was f i r s t l y to
d eterm in e th e genom ic d i s t r ib u t io n o f hom ologous seq u e n c e s
betw een HSV-1 and HSV-2, and to a s c e r t a in w h eth er th e genom es
a re c o l i n e a r . Second , e x p er im en ts were c a r r ie d ou t to
d eterm in e w h eth er r e g io n s o f g r e a te r hom ology betw een HSV-1
and HSV-2 DM a re a ls o hom ologous to PRV DNA and eq u in e h er p e s
v ir u s ty p e 1 (EHV-1) DNA to a g r e a te r e x te n t than o th e r r e g io n s .
Homology betw een th e HSV and human c y to m e g a lo v ir u s (CMV)
genom es was a ls o in v e s t i g a t e d . T h ird , c o l i n e a r i t y o f th e
genom es o f IISV and EHV-1 or PRV was c o n s id e r e d . L a s t , a
s tu d y was made o f a la r g e number o f HSV-l/HSV-2 in t e r t y p i c
reco m b in a n ts in o rd er to d eterm in e th e r e la t io n s h ip betw een
c r o s s o v e r lo c a t io n and r e g io n s o f h ig h e r hom ology .
D is t r ib u t io n o f hom ologous seq u en ces betw een HSV-1 and HSV-2 DNA
B lo t h y b r id is a t io n .p r o v id e s a good method f o r sem i-
q u a n t i t a t iv e e s t im a t io n o f th e p r o p o r t io n o f probe w hich
h y b r id is e s to each r e s t r i c t i o n fragm en t o f a genom e.
H y b r id is a t io n in s o lu t io n i s q u a n t i t a t iv e , but r e q u ir e s each
fragm en t to be i s o l a t e d and h y b r id is e d s e p a r a t e ly , and
th e r e fo r e i s much l e s s c o n v e n ie n t f o r i n i t i a l d e te r m in a t io n
o f th e d i s t r i b u t i o n o f hom ology betw een r e g io n s o f HSV-1 and
nSV-2 DNA.
F ig u r e s C 2 .1 -C 2 .2 show exam ples o f th e r e s u l t s o f
h y b r id is in g HSV-1 or iISV-2 p ro b es to n i t r o c e l l u l o s e f i l t e r s
c o n ta in in g fra g m en ts o f HSV-1 or HSV-2 DNA. The p rob es in

- 83 -
t h e s e , and su b se q u e n t, ex p er im en ts were p rep ared by n ic k Q
t r a n s la t io n i n th e p r e se n c e o f 10 g /m l DNase, and under
th e s e c o n d it io n s th e mean s i z e o f s in g le - s t r a n d e d DNA
fra g m en ts i s 400 n (R igby e t a l . , 1 9 7 7 ) . Each b lo t s t r i p
o f a p a ir c o n ta in e d an eq u a l amount o f DNA ( 0 .5 - 1 .0 yug), and
r e s u lt e d from t r a n s f e r o f DNA from th e same g e l . An eq u a l
amount o f probe DNA ( 0 .1 - 0 .2 yug) was added to each s t r i p -» ^
and a llo w e d to a n n e a l to s a tu r a t io n (5 -5 d a y s) under s t r in g e n t
h y b r id is a t io n c o n d it io n s (3 x SSC and D en h a rd t’ s s o lu t io n in
50# form am ide a t 45 ) • The e x p e r im e n ta l d e s ig n en sured th a t «
p o t e n t i a l c a u s e s o f a r t e f a c t s , su ch a s i n e f f i c i e n t t r a n s f e r
o f some r e s t r i c t i o n fra g m en ts in th e b l o t t i n g p r o ced u re ,
were c a n c e l le d o u t .
The e x t e n t o f in t e r t y p i c hom ology can in p r in c ip le be
d eterm in ed f o r each r e s t r i c t i o n fragm en t by com parison o f
th e band i n t e n s i t i e s produced by hom ologous and h e te r o lo g o u s
p r o b e s . In F ig u re C 2 .1 , f o r exam ple, HSV-1 BamHI o and r
are more in t e n s e than piq w ith HSV-2 p r o b e . A se m i-
q u a n t i t a t iv e e s t im a te o f h y b r id is a t io n to each fragm en t was
made by a u to d e n s ito m e tr ie sca n n in g o f s e v e r a l a u to r a d io g r a p h s .
Peaks were c u t o u t and w eigh ed , and th e f o l lo w in g r a t i o s
c a lc u la t e d .
R1 - wt o f peak w ith h e te r o lo g o u s probe wt o f peak w ith hom ologous probe
^2 — t o t a l wt o f p eak s w ith h e te r o lo g o u s probe t o t a l wt o f peaks w ith hom ologous probe
K3 = % /H g .
R'5 i s in d e p en d e n t o f s p e c i f i c a c t i v i t i e s o f th e p rob es and
d u r a tio n o f a u to r a d io g r a p h ic ex p o su re in th e l in e a r range o f
th e f i l m , and i s an a r b it r a r y v a lu e f o r r e l a t i v e hom ology.

- 84 -
A ll th e fra g m en ts o f r e s t r i c t i o n d i g e s t s c o u ld n o t be r e s o lv e d
on a s in g l e g e l , so i t was n e c e s s a r y to scan two a u to r a d io g r a p h s!
one from a g e l o f lo w er a g a ro se c o n c e n tr a t io n f o r bands o f
la r g e r m o le c u la r w e ig h ts , and one o f h ig h e r c o n c e n tr a t io n f o r
bands o f s m a lle r m o le c u la r w e ig h t . T h is approach le d to two
i n t e r n a l l y c o n s i s t e n t s e t s o f R^ v a lu e s i n each c a s e . A few
bands o f in te r m e d ia te m o le c u la r w e ig h t were r e s o lv e d on b oth
g e l s , so R v a lu e s f o r s m a lle r fr a g m en ts w ere m u lt ip l ie d by
a f a c t o r w hich e q u a lis e d th e two s e t s o f R^ v a lu e s f o r th e
in te r m e d ia te bands, th e re b y l in k in g th e two s e t s o f d a ta .
R- v a lu e s a r e , o f c o u r s e , m erely an a p p rox im ate i n d ic a t io n o f
r e l a t i v e hom ology, th e in t e r t y p ic genom e-to-genom e hom ology
b e in g u n i t y .
F ig u r e C 2 .3 shows h i s t o g r a p h ic a l ly th e r e s u l t s o f
s e v e r a l e x p e r im e n ts , and sum m arises th e r e g io n s o f g r e a t e r
hom ology betw een HSV-1 and HSV-2 DNA. There i s one r e g io n
o f g r e a t e r hom ology in TR g/lR g, and s i x , o r p erh ap s s e v e n ,
i n U^. R eg io n s o f l e a s t hom ology in c lu d e f e j / l R ^ , th e
ju n c t io n s betw een Ug and TRg/IRg, and th e c e h t r a l p o r t io n o f Ug.
F ig u re C2.2 shows th e e f f e c t o f tem p era tu re o f
h y b r id is a t io n upon h y b r id is a t io n o f HSV-2 probe to HSV-1
r e s t r i c t i o n fr a g m e n ts . The p a t t e r n s a t t h r e e d i f f e r e n t
tem p era tu res a re s im i la r , e x c e p t t h a t HSV-2 DNA h y b r id is e d
more e f f i c i e n t l y to th e j o in t and S te r m in a l fra g m en ts o f
HSV-1 DNA (BamHI k and g,) a t th e h ig h e r te m p e r a tu r e s . The
g e n e r a l s a tu r a t io n l e v e l o f HSV-2 probe to HSV-1 DNA
d e c r e a se d w ith in c r e a s in g te m p e r a tu r e .
C o l in e a r i t y o f th e genomes o f HSV-1 and HSV-2
I t h as been assumed in in t e r p r e t in g th e r e s u l t s o f
hom ology e x p er im en ts th a t th e genom es o f HSV-1 and HSV-2 a re
c o l in e a r . To t e s t t h i s assu m p tion p la sm id s c o n ta in in g HSV-2

- 85 -
H in d l l l fra g m en ts were n ic k t r a n s la t e d and h y b r id is e d to b lo t
s t r i p s c o n ta in in g BamHI fra g m en ts o f HSV-1 or HSV-2 DNA
(F ig u r e C 2 .4 ) . Fragm ents com plem entary to ea ch probe a re
l i s t e d in T ab le C 2 .1 . No h y b r id is a t io n o f p ro b es to r e g io n s
o f th e genome in w hich th e y do n o t map was d e t e c te d in t h i s
e x p e r im e n t. The few f a i n t bands o b serv ed when HSV-1 BamHI
b lo t s t r i p s were u sed (F ig u r e C 2 .4 ) were p ro b a b ly due to
s l i g h t u n d e r - d ig e s t io n o f th e HSV-1 DNA.
These r e s u l t s a re c o n s i s t e n t w ith c o l i n e a r i t y o f th e
HSV-1 and HSV-2 genom es. There i s no e v id e n c e from t h e s e
ex p e r im e n ts f o r hom ology betw een one r e g io n o f a genome and
a r e g io n o f th e same o r th e h e te r o lo g o u s genome m apping a t
a d i f f e r e n t p o s i t i o n .
N o n -c o lin e a r hom ology betw een th e genom es o f HSV-1 and HSV-2
The r e s u l t s o f c r o s s - b l o t e x p er im e n ts s u g g e s te d t h a t
c e r t a in fra g m en ts o f one genome a l s o h y b r id is e d to d i s t a l
fra g m en ts o f th e same, or th e h e t e r o lo g o u s , genom e. In
F ig u re C l .10 th e r e i s e v id e n c e o f hom ology betw een HSV-1
Kpnl i and k ( th e S te r m in i) and HSV-1 Huai m ( th e L term in u s)
and o p q r . In F ig u re C 2 .5 , HSV-1 Kpnl ^ and k h y b r id is e d to
HSV-1 BamHI d , j and s , and p erh ap s to b , c and e . Homology
betw een L and S te r m in i i s e x p la in e d by th e p r e se n c e to th e
a seq u en ce a t both te r m in i and a t th e L-S j o i n t . H y b r id is a t io n
to HSV-1 BamHI ^ was th e r e s u l t o f BamHI E (BamHI k w ith
an a d d i t io n a l a seq u en ce ) w hich co m ig ra ted w ith t h i s fra g m en t.
F ig u re C 2.5 shows a c r o s s - b l o t o f HSV-1 BamHI a g a in s t HSV-1
BamHI, and d em o n stra tes some h y b r id is a t io n o f HSV-1 BamHI
k and pq, to d . F ig u re C 2.6 shows some h y b r id is a t io n o f
KcoHI k to 1 . These r e s u l t s s u g g e s t a r e g io n in HSV-1 BamHI
d and BcoRI 1 to w hich th e S term in u s h as some seq u en ce
hom ology. H y b r id isa t io n betw een th e S term in u s and th e

- 8 6 -
ju n c t io n s betw een U- and TR^/IR^ was d e te c te d in some, but
n o t a l l , e x p e r im e n ts .
F ig u re C 2 .6 shows a c r o s s - b l o t betw een HSV-1 BamHI
and HSV-2 BamHI . The m ost o b v io u s a b e r r a n t h o m o lo g ie s a re
th o s e betw een HSV-1 BamHI k and pq and HSV-2 BamHI p and y z .
F a in te r s p o t s in c lu d e HSV-1 BamHI k and pq w ith HSV-2 BamHI
c , f , 1 and a 1, and HSV-2 BamHI a 1 w ith HSV-1 BamHI b, d and e .
Many, o f t h e s e a b er r a n t h o m o lo g ie s in v o lv e r e p e t i t i v e seq u e n c es
o r t h e i r ju n c t io n s w ith u n iq u e r e g io n s .
Homology betw een th e HSV genome and th e genom es o f EHV-1 and PRV
F ig u re C 2.7 shows th e r e s u l t s o f h y b r id is in g n ic k
t r a n s la t e d HSV-1, HSV-2, EHV-1 o r PRV DNA to b lo t s t r i p s
c o n t a in in g r e s t r i c t i o n fra g m en ts o f HSV-1 o r HSV-2 DNA.
T a b les C 2 .2 -C 2 .3 sum m arise th e HSV fra g m en ts to w hich EHV-1
and PRV probe h y b r id is e d . The r e g io n s o f hom ology o f e i t h e r
EHV-1 o r PRV DNA w ith th e genom es o f HSV-1 and HSV-2 are
shown in th e upper p a n e l o f F ig u r e C 2 .1 0 , and a r e l a r g e ly
s im i la r in p o s i t i o n to th e more hom ologous r e g io n s betw een
HSV-1 and HSV-2 DNA.
In o rd er to d eterm in e th e r e l a t i v e mapping p o s i t i o n s o f
HSV DNA se q u e n c e s hom ologous w ith EHV-1 and PRV DNA, n ic k. i
t r a n s la t e d p la sm id s c o n ta in in g HSV DNA fra g m en ts were
h y b r id is e d a t 40° to b lo t s t r i p s c o n t a in in g EHV-1 ..or PRV
r e s t r i c t i o n d i g e s t s . The r e s u l t s a r e shown in F ig u r e s C 2 .8 -
C 2.9 , and a r e summarised i n T a b le s C 2 .4 -C 2 .5 . The m id d le
and lo w e r p a n e ls o f F ig u re C 2.10 show th e p o s i t i o n s in th e
genom es o f EHV-1 and PRV to w hich ea ch c lo n e d fragm ent
h y b r id is e d . In in s t a n c e s where h y b r id is a t io n o ccu rred to a
band c o n ta in in g more than one r e s t r i c t i o n fra g m en t, r e s u l t s
o f th e o th e r d i g e s t s were u sed to d e term in e to w hich o f th e
fra g m en ts h y b r id is a t io n had o c c u r r e d . For exam ple, HSV-2

- 87 -HindI I I h h y b r id is e d to PRV Kpnl j k , but to BamHI k, in d ic a t in g
th a t h y b r id is a t io n was to Kpn l k r a th e r than to £ . The f a i n t
bands p r e s e n t on th e PRV DNA b lo t s t r i p s (P ig u r e C 2.9
such a s th o se j u s t below BamHI i . i and Kpnl 1 , w hich have n o t
been in c lu d e d in T ab le C 2 .5 , were due to th e p r e se n c e o f
m inute am ounts o f c o n ta m in a tin g pAT153 DNA in th e d y e - P ic o l l
w hich was added to th e d ig e s t e d PRV DNA p r io r to e le c t r o p h o r e s i s
th e p ro b es c o n ta in e d n ic k t r a n s la t e d pAT153 DNA a s w e l l a s
th e c lo n e d HSV DNA fr a g m e n ts , and th e r e fo r e r e v e a le d t h e s e
bands, w h ich were n o t d i s c lo s e d i n th e c o n t r o l s t r i p s .
Homology betw een th e HSV and CMV genomes
S im ila r ex p er im en ts to th o s e a lr e a d y d e s c r ib e d were
c a r r ie d o u t by p ro b in g b lo t s t r i p s c o n ta in in g r e s t r i c t i o n
d i g e s t s o f HSV-1 o r HSV*-2 DNA w ith human c y to m e g a lo v ir u s
(CMV) s t r a in AD169 DNA. The r e v e r s e ex p er im en t o f p ro b in g
b lo t s t r i p s c o n ta in in g CMV Hind I I I fra g m en ts w ith HSV-1 or
HSV-2 DNA was a l s o p erform ed . No h y b r id is a t io n was
d e te c te d under c o n d it io n s w hich a llo w e d EHV-1 and PRV hom ology
to HSV DNA to be d e t e c t e d . However, when e x c e s s probe8( 1 /ug a t a s p e c i f i c a c t i v i t y o f 2 x 10 c .p .m ./y tg ) was u se d ,
and lo n g ex p o su r e s made o f a u to r a d io g r a p h s , some h y b r id is a t io n
was d e te c te d (P ig u re C 2 . l l ) . CMV DNA h y b r id is e d to HSV-2
BamHI £ , and HSV-2 DNA to CMV H in d l l l no and h i ; th e s e r e s u l t s
were rep rod u ced in a seco n d e x p er im e n t. No hom ology betw een
HSV-1 and CMV DNA was d e t e c t e d .
S tu d ie s o f reco m b in a tio n betw een th e HSV-1 and HSV-2 genomes
The tech n iq u e o f marker r e s c u e o f HSV m utants (Stow
and W ilk ie , 1978) h as been v ery s u c c e s s f u l l y u sed to
g e n e r a te HSV-1/HSV-2 i n t e r t y p i c reco m b in a n ts (P r e s to n e t a l . ,
1 78; Stow and W ilk ie , 1978; Marsden e t a l . , 1978; Knipe

- 8 8 -
e t a l . , 1978; C hartrand e t a l . , 1979 and 1 9 8 0 ) . S tr u c tu r e s
o f th e recom b in ant genom es were r o u t in e ly deduced by
com paring t h e i r DNA r e s t r i c t i o n p r o f i l e s w ith th o se o f th e
p a r e n ta l v i r u s e s , and s c o r in g f o r th e p r e s e n c e or a b sen ce o f
r e s t r i c t i o n s i t e s . The ex p er im en ts d e s c r ib e d h ere were aimed
a t th r e e a s p e c t s o f r e c o m b in a tio n . The f i r s t aim was to
i n v e s t i g a t e th e p o s s i b i l i t y th a t o n ly one o f th e fo u r
iso m e r s o f th e HSV genome i s in v o lv e d in r e c o m b in a tio n . The
secon d aim was to d eterm in e w h eth er r e c o m b in a tio n te n d s to
o ccu r in r e g io n s o f h ig h e r in t e r t y p i c h om ology . The th ir d
aim was to a ttem p t t o c o n s tr u c t reco m b in a n ts w ith com p lete
t y p e - s p e c i f i c h e t e r o lo g y betw een r e p e t i t i v e r e g io n s ; i t
was p r e d ic te d th a t such reco m b in a n ts would n o t in v e r t
n o rm a lly in th e s e g m e n t(s ) bounded by h e t e r o t y p ic r e p e t i t i o n s .
T h is a s p e c t i s d e a l t w ith in S e c t io n 4 .
The m u ta tio n i n HSV-1 tsD maps i n TRg/IRg (BamHI q;
P r e s to n , in p r e s s ) , and may c o n v e n ie n t ly be u sed to produce
i n t e r t y p i c reco m b in a n ts p o s s e s s in g HSV-2 DNA in r e p e t i t i v e
r e g io n s . C e l l m on olayers were c o in f e c t e d w ith tsD DNA and
HSV-2 EcoRI bcde (fra g m en ts sp a n n in g th e j o i n t r e g io n )
i s o l a t e d from a g a r o se g e l s , by th e method o f Stow and
W ilk ie ( 1 9 7 6 ) . The m on olayers were in c u b a te d un d er EHu5
a t 3 8 .5 ° , and s i x p la q u e s p ic k ed from ea ch o f f i f t e e n P e t r i
d i s h e s . The n in e ty p la q u es were p u r i f i e d by two f u r th e r
rou n d s o f p la q u e -p ic k in g a t 3 8 .5 ° . V ir u s s to c k s were grown
a t 31° on P e t r i d i s h e s , t i t r a t e d , and in v iv o ^P-DNA was
p r ep a r ed . Genome s t r u c t u r e s were a n a ly se d by r e s t r i c t i o n
en d o n u c le a se d ig e s t io n w ith Xbal , H in d iI I , EcoRI, B g l l l ,
Hpal , Kpnl and BamHI. A l l c r o s s o v e r s mapped w ith in th e
r e s c u in g HSV-2 j o in t fra g m e n ts , and genome s t r u c t u r e s o f
th e s ix i s o l a t e s from any s in g le d is h s u g g e s te d t h a t in

- 89 -
g e n e r a l th e y were n o t r e la t e d .
The re co m b in a tio n e v e n ts w hich o ccu r in an ex p er im en t
o f t h i s ty p e are r a th e r com plex ow ing to th e f a c i l i t y w ith
w h ich seco n d a ry r e c o m b in a tio n e v e n ts occu r in r e p e t i t i v e
r e g io n s . T h is i s d is c u s s e d in d e t a i l in S e c t io n 4 . In
s h o r t , a prim ary c r o s s o v e r in one r e p e t i t i v e r e g io n i s a l s o
p r e s e n t i n th e o th e r r e p e t i t i o n a t th e s ta g e o f genome a n a l y s i s ,
due to seco n d a ry re co m b in a tio n e v e n t s . T h is means t h a t i f th e
p rim ary c r o s s o v e r o ccu rred i n a r e p e t i t i v e r e g io n o f a
segm en t, i t i s , in th e g r e a t m a jo r ity o f c a s e s , n o t p o s s ib le
t o s t a t e th e o r i e n t a t io n o f th a t segm ent d u r in g r e c o m b in a tio n .
N e v e r t h e le s s , r e co m b in a tio n e v e n ts in o r Ug a r e p r e ser v e d
u n am b igu ou sly r e g a r d le s s o f fu r th e r c r o s s o v e r s in r e p e t i t i v e
r e g io n s . T h e r e fo r e , c a r e f u l a n a ly s i s o f th e r e s u l t i n g
genom es from marker r e s c u e o f tsD a l lo w s e i t h e r th e*
o r ie n t a t io n o f b oth L and S a t prim ary rec o m b in a tio n to be
deduced ( c r o s s o v e r s in U- and U g ) , o r th e o r i e n t a t io n o f S
o n ly ( c r o s s o v e r s in TRj/UE^ and U g ) , o r th e o r ie n t a t io n o f
1 o n ly ( c r o s s o v e r in U£ and T R g/IR g), or n e i t h e r o r ie n t a t io n
( c r o s s o v e r s in TRj/lR-^" and TRg/IRg, o r TRg/IRg and Ug, o r
TRs /I K g o n ly ) .
T able C 2.6 shows th e numbers o f reco m b in a n ts
r e p r e s e n t in g each o r i e n t a t io n o f th e r e sc u e d genom e. I t i s
c l e a r from th e s e r e s u l t s t h a t tsD was r e sc u e d w ith th e
genome in b oth th e P and th e 1^ o r i e n t a t io n s , and a l l th e
o b se rv ed c r o s s o v e r s co u ld h ave" occu rred betw een tsD and
HSV-2 EcoRI b or c . I t can n ot be c o n c lu d e d , h ow ever, t h a t
r eco m b in a tio n in th e Ig or Ig^ o r ie n t a t io n s (w ith HSV-2
EcoRI d and e ) d id n o t o c c u r ,' b ecau se th e g r e a t m a jo r ity
o f reco m b in a n ts p o s s e s s e d sy m m etr ica l c r o s s o v e r s in TRg / I H S
and th e r e fo r e m ight e q u a lly w e l l have been th e r e s u l t o f
r e s c u e by HSV-2 EcoRI d or e . The p o s i t io n o f tsD in

- 90 -
TRg/IRg means th a t upon r e sc u e o f t h i s m u ta tio n a t l e a s t one
c r o s s o v e r ev en t in S i s o b l ig a t o r y , w hereas a co n cu rren t
c r o s s o v e r e v e n t in L i s o p t io n a l . I t may be co n clu d ed t h a t
a t l e a s t th e P and 1^ o r ie n t a t io n s o f th e genome p a r t ic ip a t e
in r e c o m b in a tio n .
A secon d f e a t u r e o f reco m b in a tio n i s r e v e a le d by
th e s e r e s u l t s . About te n t im e s a s many reco m b in a n ts
r e s u l t e d from marker r e s c u e o f th e 1^ o r ie n t a t io n compared
w ith th e P o r i e n t a t io n , a lth o u g h EcoRI b and c d i f f e r i n
s i z e by o n ly about 5^ . The d i s t r i b u t i o n o f prim arym
c r o s s o v e r p o s i t i o n s i n a re shown in P ig u re C 2 .1 2 . Two
reco m b in a n ts p o s s e s s e d sy m m etr ica l c r o s s o v e r s in TR / I rL’
so i t was assum ed f o r th e p u rp o ses o f th e diagram th a t th e
prim ary e v e n t co u ld have o ccu rred w ith eq u a l p r o b a b i l i t y
in e i t h e r TR or IH^. R ecom b in ation seem s to have o ccu rred
p r e f e r e n t i a l l y in th e more hom ologous r e g io n s between th e
HSV-1 and HSV-2 genom es.
In a secon d ex p er im e n t, tw e n ty - e ig h t reco m b in a n ts
were i s o l a t e d by marker r e s c u e o f tsD w ith HSV-2 Hindl l l
c d e fg ( c d fg a re th e j o in t fr a g m e n ts ) , o f w h ich i t was
p o s s ib l e to d eterm in e th e s t r u c t u r e s o f tw e n ty -s e v e n .
A l l c r o s s o v e r s in Ug ( 7 /2 7 ) o ccu rred i n r e g io n s o f HSV-1
HRA com plem entary to HSV-2 HindI I I k; o th e r c r o s s o v e r s
were w ith in T R g /I R g , and in a l l e x c e p t one c a s e were
sy m m e tr ic a lly p o s i t io n e d in both r e p e t i t i o n s . O ther
c r o s s o v e r p o s i t i o n s were e i t h e r in TR^/IR^ c l o s e to th e
j o i n t , or in TRg/IRg* th e s e two p o s s i b i l i t i e s co u ld n o t be
d is t in g u is h e d from ea ch o th e r . A lth ough th e r e s c u in g
fra g m en ts ex ten d ed i n t o U^, no c r o s s o v e r s i n were
d e t e c t e d . Prim ary c r o s s o v e r p o s i t i o n s in S from both

- 91 -
ex p er im en ts a re shown in P ig u re C 2 .1 3 . Only HSV-2 EcoRI
j o i n t fra g m en ts produced c r o s s o v e r s a t th e l e f t hand s id e
o f U , and o n ly HSV-2 H in d l l l j o i n t fra g m en ts recom bined a tiD
th e r ig h t hand s i d e . The com p arison o f reco m b in a tio n
f r e q u e n c ie s on th e two s id e s o f Ug was c o n s tr u c te d on th e
assu m p tio n t h a t prim ary r e c o m b in a tio n w ith in T R g /IR g
c o u ld o ccu r w ith e q u a l p r o b a b i l i t y i n e i t h e r ; th e r e fo r e
i n ea ch o f th e two c a s e s th e number o f prim ary re co m b in a tio n
e v e n ts o c c u r r in g in T R g /I R g was d iv id e d by tw o. The r e s u l t s
a re e x p r e s se d a s c r o s s o v e r f r e q u e n c ie s r e l a t i v e to th o se
w ith in HSV-1 BamHI A gain , reco m b in a tio n e v e n ts o ccu rred
p r e f e r e n t i a l l y w ith in th e m ost hom ologous r e g io n s .

- 92 -
JOISCUS5IOH
F e a tu r e s o f n u c le ic a c id h y b r id i s a t io n
H y b r id is a t io n i n s o lu t io n h a s been u sed s u c c e s s f u l l y f o r
many y e a r s to d eterm in e r e la t i o n s h i p s betw een n u c le ic a c id s ,
and th e f e a t u r e s o f t h i s a s s a y h ave been w e l l c h a r a c te r is e d
b oth t h e o r e t i c a l l y and e m p ir ic a l ly . H y b r id is a t io n r a t e i s
m axim al a t a p p r o x im a te ly 2 5 ° below th e h y b r id m e lt in g
tem p era tu re (Tm) , and v a r i e s m in im a lly o v e r a ran ge o f 1 0 -1 5 °
i n t h a t r e g io n . Tm i s r e la t e d to io n c o n c e n tr a t io n and OC
c o n te n t (S c h ild k r a u t and L if s o n , 1 9 6 5 ) , and i s d e c r e a se d by .
a p p r o x im a te ly 1 ° p e r 1 # b a se m ism atch in g (H u tton and Wetmur,
1 9 7 3 ) . The u se o f s in g le stra n d ed DNA bound to a n i t r o c e l l u l o s e
f i l t e r i n th e h y b r id is a t io n r e a c t io n i s a l e s s w e l l -
c h a r a c t e r is e d sy ste m , s in c e under c e r t a in c o n d it io n s th e r a t e
o f r e a c t io n i s l im i t e d by a d i f f u s i o n f a c t o r , and a l s o
c o n c a te n a t io n o f p a r t i a l l y r e a n n e a le d probe to th e f i l t e r - b o u n d
DNA may o ccu r ( F l a v e l l e t a l . , 1 974a and b ) . The u se o f
form am ide i n f i l t e r h y b r id is a t io n s t i l l f u r t h e r r e d u c es th e
p o s s i b i l i t y o f m ea n in g fu l q u a n t i f i c a t io n o f r e s u l t s . In
s o lu t io n h y b r id is a t io n , formam ide r e d u c e s Tm by 0*6° p er 1 $
form am ide, and -the h y b r id is a t io n r a t e by h a l f , a lth o u g h th e
r a t e r e t a i n s an ap p roxim ate in d ep en d en ce o f tem p eratu re in th e
r e g io n o f Tm-2 5 ° (H u tton , 1 9 7 7 ) . Formamide e f f e c t u a l l y
in c r e a s e s th e s t a b i l i t y o f m ism atched h y b r id s on f i l t e r s when
compared w ith what a re presum ably com parable c o n d it io n s i n th e
a b se n c e o f form am ide (Schm eckpeper and S m ith , 1 9 7 2 ) .
T h ere fo re f i l t e r h y b r id is a t io n i n 5 0 # form am ide, w hich0
was th e method u sed in t h i s s tu d y , can p r o v id e an i n d ic a t io n -
o f r e l a t i v e h y b r id is a t io n betw een r e s t r i c t i o n fra g m en ts o f
h e te r o lo g o u s DNAs, but n o t r e l i a b l e q u a n t i t a t iv e e s t im a t e s .
However, q u a n t i t a t iv e e s t im a t io n s o f th e d e g r ee o f hom ology

- 93 -
b etw een h e te r o lo g o u s DNAs d e r iv e d from h y b r id is a t io n
e x p e r im e n ts bear no d i r e c t r e la t io n s h ip to s i m i l a r i t y o f
n u c le o t id e se q u e n c e s , and t h i s i s o f c o u r se th e more
m ea n in g fu l d e s c r ip t io n o f hom ology. For exam p le , i t i s n o t
known w h eth er hom ologous se q u e n c es d e t e c te d i n t h i s s tu d y a re
w id e ly d i s t r ib u t e d w ith in th e hom ologous r e g io n s , or w hether
th e y c o n s i s t o f l o c a l r e g io n s o f v e r y h ig h h om ology . These
a l t e r n a t i v e s co u ld be d is t in g u is h e d by e s t im a t in g th e s t a b i l i t y
o f th e h y b r id s . A la r g e d i f f e r e n c e i n th e e x t e n t o f ap p aren t
hom ology b etw een two s t r a i n s o f v e s i c u l a r s t o m a t i t i s v ir u s
d eterm in ed by th e two m ethods o f h y b r id is a t io n and n u c le o t id e
se q u e n c in g was d is c u s s e d by McGeoch e t a l . ( 1 9 8 0 ) .
In h y b r id is in g h e te r o lo g o u s DNAs, th e Tm o f h y b r id s , and
h en ce t h e ir r a t e o f fo r m a tio n and r e s u l t i n g s t a b i l i t y , i s
d ep en d en t upon th e GC c o n te n t o f hom ologous r e g io n s and th e
e x t e n t o f b ase m ism a tch in g . U sin g th e e q u a t io n o f S c h ild k r a u t
and L if s o n (1 9 6 5 ) and assu m in g t h a t 5 0 # form am ide red u o es Tm
by 30° (H u tton , 1 9 7 7 ) , th e DNAs o f HSV-1, HSV-2, PRV and EHV-1
u nder th e c o n d it io n s u sed have Tm- 2 5 ° v a lu e s o f 4 8 .9 , 4 9 .7 ,
5 1 .4 and 4 4 .4 ° , r e s p e c t i v e l y . T hese a re mean v a lu e s , and Tm
w i l l d i f f e r f o r r e g io n s o f th e genome w ith GC c o n te n ts
d i f f e r e n t from th e mean. In g e n e r a l , h ow ever, h y b r id is a t io n o f
HSV-1 and HSV-2 DNA a t 50° a l lo w s up to 1 0 -1 5 # m ism atch in g ,
45 ° a l lo w s 1 5 -2 0 # , and 40 a l lo w s 2 0 -2 5 # , assu m in g t h a t th e4 4
e f f e c t o f m ism atch in g on Tm i s th e same i n s o lu t io n h y b r id is a t io n
and under th e c o n d it io n s u sed h e r e . H y b r id is a t io n o f HSV-1
and HSV-2 DNA a t t h e s e th r e e tem p era tu res r e s u l t e d i n s im i la r ,
b u t n o t i d e n t i c a l , p a t te r n s (P ig u r e 0 2 . 2 ) . T h is i s i n agreem ent
w ith K ie f f e t a l . (1 9 7 2 ) , who co n clu d ed t h a t th e hom ologous
se q u e n c es p o s s e s s a low d eg ree o f m ism a tch in g . H y b r id is a t io n
o f EHV-1 DNA to HSV-1 o r HSV-2 DNA was much more tem p era tu re -

- 94 -
d ep en d en t in th e 4 0 -5 0 ° range ( f i g u r e C 2 .7 ) , presum ably a s a
r e s u l t o f g r e a t e r m ism a tch in g .
The e q u a t io n o f S c h ild k r a u t and L if s o n (1 9 6 5 ) shows t h a t
hom ologous r e g io n s may v a r y in GC c o n te n t o v e r a 35?* ra n g e ,
o th e r f a c t o r s b e in g e q u iv a le n t , w ith o u t g r e a t ly a f f e c t i n g th e
r a t e o f h y b r id i s a t io n . The d i s t r i b u t i o n o f th e more hom ologous
r e g io n s b etw een HSV-1 DNA and th e o th e r h e r p e s v ir u s DNAs d o es
n o t o b v io u s ly r e f l e c t th e d i s t r i b u t i o n o f G C -rich r e g io n s in
th e HSV-1 genome lo c a t e d by th erm al d e n a tu r a t io n ex p er im en ts
( D e l iu s and C lem en ts , 1 9 7 6 )* I t i s t h e r e f o r e p ro b a b le t h a t th e
r e g io n s o f g r e a t e r hom ology d e t e c te d i n t h i s s tu d y a re l a r g e l y
th e r e s u l t o f g r e a t e r s i m i l a r i t y o f n u c le o t id e se q u e n c e s
r a th e r th a n m erely a r e f l e c t i o n o f l o c a l GC c o n t e n t .
C o l in e a r i t y o f th e genom es o f HSV-1 and HSV-2
H y b r id is a t io n o f c lo n e d DNA fra g m en ts to n i t r o c e l l u l o s e
s t r i p s c o n t a in in g HSY-1 o r HSV-2 r e s t r i c t i o n fra g m en ts showed
t h a t , w i t h in th e r e s o lu t io n a t t a in e d , th e two genom es a re
c o l in e a r ( f ig u r e C2.4» T ab le C 2 * l) . H y b r id is a t io n o f HSV-1
and HSV-2 BamHI fra g m en ts in th e c r o s s b lo t o f f ig u r e C2*6
a l s o d em o n stra ted c o l i n e a r i t y , w ith th e e x c e p t io n o f some
n o n -c o l in e a r hom ology a s d is c u s s e d p r e v io u s ly . E sp arza e t a l .
(1 9 7 6 ) co n c lu d ed t h a t th e r e i s a t l e a s t some d e g r e e o f
c o l i n e a r i t y betw een HSV-1 and HSV-2 DNA from th e o b s e r v a t io n
t h a t h e te r o lo g o u s m utant p a ir s w hich f a i l e d to com plem ent a l s o,V4V-
f a i l e d to reco m b in e . A n a ly se s o f i n t e r t y p io reco m b in a n ts
have shown t h a t c r o s s o v e r s o cc u r th ro u g h o u t th e genome,
r e f l e c t i n g f u n c t io n a l c o l i n e a r i t y . S p e c i f i c f u n c t io n s , su ch
a s th e dPyK gene (H a ll ib u r to n e t a l . , 1 9 8 0 ) , th e DNA p o lym erase
gene (C hartrand e t a l . , 1 9 7 9 ) , and th e IE g e n e s (C lem en ts e t a l . ,
1 9 7 7 ;~ E a sto n and C lem en ts , 1 9 80) a re in e q u iv a le n t l o c a t io n s
in th e two genom es. The m o r p h o lo g ic a l tr a n sfo r m in g r e g io n s o f

- 95 -HSV-1 and HSV-2 a re n o t e q u iv a le n t ly lo c a t e d , but th e DNA in
ea ch r e g io n i s c o l in e a r w ith th e a n a lo g o u s r e g io n i n th e o th e r
genome (R eyes e t a l . , 1 9 7 9 ) . Ruyechan e t a l . (1 9 7 9 ) co n clu d ed
t h a t th e g e n e s s p e c i f y in g gC a r e n o t c o l i n e a r , a lth o u g h t h i s
was n o t su p p o rted by H a ll ib u r to n (1 9 8 0 ) . The HSV-1 BamHI/
HSV-2 BamHI c r o s s b lo t o f F ig u r e C2*6 d id n o t s u g g e s t any
n o n - c o l in e a r i t y i n t h i s r e g io n (0 * 5 3 -0 .6 9 f r a c t i o n a l genome u n i t s ) .
C onserved r e g io n s i n th e genom es o f HSV-1. HSV-2, EHV-1 and FRY
Seven m ajor r e g io n s o f h ig h e r hom ology b etw een HSV-1 and
HSV-2 DNA w ere d em o n stra ted ( I - V I I i n l i g u r e C 2 .1 0 ) , and o f
t h e s e I , ' I I I , V, VI and V II were a l s o hom ologous to PRV and
EHV-1 DNA, a lth o u g h a t a much lo w er l e v e l . The e x t e n t o f
r e g io n ’ l l was more l im i t e d u s in g EHV-1 DNA a s p ro b e , and
r e g io n IV was d e t e c te d w ith PRV a s probe o n ly when HSV-1 DNA
f i l t e r s w ere u se d . A lth ou gh th e same r e g io n s a re con served "
b etw een th e fo u r genom es, th e r e s u l t s d id n o t show w h eth er
th e same DNA se q u e n c e s a r e co n se r v ed in ea ch c a s e . The
o b s e r v a t io n th a t n e a r ly a l l HSV-1 and HSV-2 r e s t r i c t i o n
fra g m en ts showed some l e v e l o f h y b r id is a t io n to th e h e te r o lo g o u s
probe (F ig u r e s C 2.1 and C 2 .2 ) , w h ereas o n ly c e r t a in fra g m en ts
h y b r id is e d to PRV o r EHV-1 p ro b es (F ig u r e C 2 * 7 ), may be
a t t r ib u t e d t o th e much lo w er l e v e l o f h y b r id is a t io n in g e n e r a l
in th e l a t t e r c a s e , and hence a lo w e r l e v e l o f s t r in g e n c y may
be r e q u ir e d to d e t e c t h y b r id is a t io n to th e o th e r fr a g m e n ts .
The sev en r e g io n s o f h ig h e r hom ology p ro b a b ly r e f l e c t c o n s t r a in t s
on DNA se q u e n c es im posed by c o n s e r v a t io n o f amino a c id
se q u e n c e s i n th e p r o t e in s encoded by th e DNA. The f o l lo w in g
p aragrap h s d is c u s s th e i d e n t i t i e s and f u n c t io n s o f p u t a t iv e
co n se r v ed p r o t e in s .
R eg ion I maps w ith in th e HSV-2 m o r p h o lo g ic a l tr a n sfo r m in g
r e g io n (R eyes e t a l . , 1 9 7 9 ) . One p o s s i b i l i t y f o r th e co n serv ed

- 96 -
p r o t e in i s gC, th e s t r u c t u r a l gen e f o r w hich i s lo c a t e d in
t h i s r e g io n but h a s n o t been p r e c i s e l y mapped (M arsden e t a l * ,
1 9 7 8 ; Ruyechan e t a l . , 1 979; H a ll ib u r to n , 1 9 8 0 ? . uyp e-
s p e c i f i c a n t ig e n ic s i t e s have been r e p o r te d f o r t h i s
g ly c o p r o t e in (N o r r ild and V e ste r g a a r d , 1 9 7 9 ) . P erh ap s a more
l i k e l y c a n d id a te i s Vmw IE 1 3 6 * (1 4 3 ) , th e gen e f o r w hich i s
t r a n s c r ib e d i n sm a ll q u a n t i t i e s i n th e p r e s e n c e o f
c y c lo h e x im id e , and i n g r e a t ly in c r e a s e d am ounts a f t e r rem oval
o f th e in h i b i t o r (C lem en ts e t a l * , 1977; J . M cLauchlan and
J .B . C lem en ts , p e r s o n a l co m m u n ica tio n ). The mRNA Jaas a s i z e
o f 5*0 kb and h as been a c c u r a t e ly lo c a t e d on th e genome
w ith in r e g io n I (A nderson e t a l l . 1 9 8 1 ; J . M cLauchlan and J .B .
C lem en ts , p e r s o n a l co m m u n ica tio n ). The f u n c t io n o f th e
p r o t e in i s unknown, a lth o u g h th e HSV-2 f u n c t io n c a u s in g
i n h i b i t i o n o f h o s t m acrom olecu lar s y n t h e s is maps i n th e same
l o c a t io n ( 0 .5 6 - 0 .5 7 ) •
R eg ion I I i s lo c a t e d i n th e p o s i t i o n o f th e s t r u c t u r a l
gen e c o d in g f o r th e HSV DNA p o ly m era se (C hartrand e t a l * , 1 9 7 9 ) .
H a llib u r to n and Andrew ( i9 ? 6 ) were a b le to d e t b c t a d eg ree
o f c r o s s - n e u t r a l i s a t i o n betw een th e DNA p o ly m er a se s o f HSV-1
and HSV-2, but n o t betw een th e s e two v i r u s e s and PRV.
The d a ta o f C on ley .e t a l . (1 9 8 1 ) s u g g e s t th a t r e g io n I I I
c o n t a in s th e gen e c o d in g f o r th e m ajor D N A -binding p r o t e in
(MDBP) o f HSV-1. Yeo e t a l . (1 9 8 1 ) showed t h a t a n t i s e r a to
c e l l s in f e c t e d w ith HSV-1, HSV-2, BMV, EHV-1 and PRV r e a c t
h e t e r o lo g o u s ly p red o m in a n tly w ith MDBP, and th e y p rop osed t h i s
p r o t e in a s a p u ta t iv e h e r p e s v ir u s group s p e c i f i c a n t ig e n .
A lth ou gh MDBP i s l i k e l y to have a c e n t r a l r o l e i n DNA
r e p l i c a t i o n , i t s p r e c i s e fu n c t io n i s unknown. The l o c i f o r
gA and gB a l s o map i n t h i s r e g io n , and t h e s e g ly c o p r o t e in s have
been r e p o r te d to be s e r o l o g i c a l l y co n serv ed i n HSV-1 and HSV-2
(N o r r ild e t a l . , 1978; E b er le and C ou rtn ey , 1 9 8 0 a ) . O th ers

- 97 -
h ave r e p o r te d t h a t gA and gB p o s s e s s t y p e - s p e c i f i c a n t ig e n ic
s i t e s (P o w e ll and W atson, 1975; C ourtney and P o w e ll, 1 9 7 5 ) .
Yeo e t a l . (1 9 8 1 ) a l s o r e p o r te d l im i t e d c r o s s - r e a c t i v i t y
b etw een th e m ajor c a p s id p r o t e in s (MCP) o f th e f i v e h e r p e s v ir u s e s
th e y c o n s id e r e d . C ourtney and P o w e ll (1 9 7 5 ) showed ty p e -
common a n t ig e n ic s i t e s on th e MCP o f HSV-1 and HSV-2 u s in g
s p e c i f i c a n t i s e r a , and Cohen e t a l , ( 1 9 8 0 b )d em o n stra ted
c o n s e r v a t io n o f MCP i n HSV-1 and HSV-2 by s e r o l o g ic a l and
p r o t e o l y t i c m eth od s. MCP has a m .w t. o f 1 4 -0 ,0 0 0 -1 6 0 ,0 0 0 i n a
w ide ra n g e o f h e r p e s v ir u s e s (S p ea r e t a l * , 1 9 7 8 ) and th e genea - - -
h a s been p r e c i s e l y lo c a t e d f o r HSV-1 (C o sta e t a l . , 1 9 8 1 ) ,^ - - -
c o r r e sp o n d in g w e l l w ith th e p o s i t i o n o f r e g io n V.
R eg ion V II i s c e r t a i n l y l o c a t e d , f o r HSV-1, w ith in th e
c o d in g r e g io n o f th e Vm IE 175 gen e (M -J. M urchie and P*J.
R ix o n , p e r s o n a l c o m m u n ica tio n s). D e t a i le d hom ology s t u d ie s o f
th e j o i n t r e g io n s o f HSV-1 and HSV-2 DNA, a s d is c u s s e d in
S e c t io n 3 , in d ic a t e t h a t hom ology e x te n d s th ro u g h o u t a t l e a s t
th e p o ly p e p t id e co d in g r e g io n o f th e g en e : th e arrangem ent o f
r e s t r i c t i o n s i t e s f a l s e l y g iv e s th e im p r e s s io n t h a t HSV-l/HSV-2
hom ology i n t h i s r e g io n i s c o n f in e d to HSV-1 BamHI y , w hereas
i t d o es ex te n d in t o BamHI k . An IE p o ly p e p t id e th o u g h t to
be a n a lo g o u s to HSV-1 Vmw IE 175 h as been mapped i n a s im i la r
p o s i t i o n i n TRg/IRg o f PRV t(]? o w ell, 1 9 7 9 ) . VBW IE 175
f u n c t io n s i n prom otin g t r a n s c r ip t io n o f e a r ly g e n e s . I t i s
n o te d t h a t , a lth o u g h hom ology betw een HSV-1 and HSV-2 DNA i n
t h i s r e g io n o f th e genome i s g r e a t e r th a n i n a d ja c e n t r e g io n s ,
i t seem s n o t to be a s g r e a t a s in r e g io n s I -V I (P ig u r e C 2 .3 )»
C ourtney and P o w e ll (1 9 7 5 ) r e p o r te d t y p e - s p e c i f i c s i t e s on
t h i s p o ly p e p t id e in HSV-1 and HSV-2, u s in g a s p e c i f i c a n tiseru m
to Vmw IE 1 7 5 .
Knowledge o f th e lo c a t io n s o f gen e f u n c t io n s on th e HSV

- 98 -
genome i s n o t y e t s u f f i c i e n t l y advanced to a l lo w th e
i d e n t i f i c a t i o n o f th e p r o t e in s w hich may be r e s p o n s ib le f o r
th e g r e a te r hom ology i n r e g io n s IV and V I. W ith th e e x c e p t io n
o f th e p u t a t iv e a n a lo g u e o f Vmw IE 175 i n PRV, n o th in g i s
d i r e c t l y known o f th e p r o t e in s coded by th e hom ologous
r e g io n s i n th e PRV and EHV-1 genom es.
The p r o t e in s d e s c r ib e d above com p rise e a r ly and l a t e ,
s t r u c t u r a l and n o n - s t r u c tu r a l members. I t i s n o ted ;th at th e
dPyK gen e d o e s n o t r e s id e w ith in a r e g io n o f g r e a t e r hom ology
in HSV-1 and liSV -2. The r e g io n s o f l e a s t hom ology betw een
HSV-1 and HSV-2 DNA encode HSV-1 Vmw IE 1-2 and 68 a t th e
TRg/us and IRg/Ug j u n c t io n s , and Vmw IE 110 i n T R i/lR ^.
A s t r i c t com p arison betw een s e r o l o g ic a l and h y b r id is a t io n
s t u d ie s i n th e se a r c h f o r co n serv ed p r o t e in s i s n o t
s tr a ig h t fo r w a r d , s in c e gD h as been r e p o r te d to be co n serv ed
in HSV-1 and HSV-2, b oth s e r o l o g i c a l l y (Sim and W atson, 1973;
H oness and W atson, 1974) and by t r y p t ic ~ p e p t id e mapping
(E ise n b e r g e t a l . , 1 9 8 0 ) . The gen e c o d in g f o r t h i s g ly c o p r o t e in
r e s id e s w ith in Ug (Ruyechan f i l . , 1 9 7 9 ) , none o f th e seq u e n c e s
o f w hich were d e te c te d a s b e in g p a r t ic u la r ly hom ologous in
HSV-1 and HSV-2.
Homology b etw een th e genomes o f HSV and HOMV
A v e r y sm a ll amount o f hom ology b etw een HSV-2 DNA and
a t l e a s t two l o c i in o f th e CMV genom e, and betw een CMV DNA
and TRj/IRjj o f th e HSV-2 genome was d e t e c te d a t th e
tem p era tu re o f h y b r id is a t io n u s e d . I t i s q u e s t io n a b le
w hether su ch a low l e v e l o f h y b r id is a t io n i s m ea n in g fu l a t a l l ,
s in c e i t may r e s u l t from n o n - s p e c i f i c h y b r id is a t io n o f DNA o f
u n u su a l c o m p o s it io n ( e . g . h ig h GC c o n t e n t ) . T hese r e s u l t s
a re i n agreem en t w ith th o se o f Huang and Pagano (1 9 7 4 ) , who
f a i l e d to d e t e c t s i g n i f i c a n t h y b r id is a t io n o f HSV-1 or HSV-2

- 99 -
DNA to CMV DNA. S e v e r a l o th e r l i n e s o f in v e s t i g a t i o n
h ave d em o n stra ted t h a t a p a r t from v ir io n m orphology, g r o s s
genome s t r u c t u r e and some a s p e c t s o f v ir u s grow th , HSV and
CMV show l i t t l e s i m i l a r i t y . For exam ple , CMV d o e s n o t p o s s e s s
a dPyK o r th y m id in e k in a se gen e ( M i l le r e t a l . , 1977;
J e r k o fsk y e t a l . , 1 9 8 0 ) , and e x h i b i t s an IE t r a n s c r ip t io n a l
p a t te r n m arkedly d i f f e r e n t from t h a t o f HSV (Wathen e t a l . ,
1 9 8 1 ) .
D is t r ib u t io n o f r e g io n s hom ologous to HSV DNA i n th e genom es o r " E H V ^ i '" a n a :“F R r “ ----------------------------------: —
The r e s u l t s sum m arised i n l i g u r e C 2.10 show th a t th e '
hom ologous r e g io n s in th e L segm ent o f EHV-1 DNA a re c o l in e a r
w ith th o s e in th e L segm ent o f HSV DNA i n t h e ’ l i , o r i e n t a t io n .
I t m ust be s t r e s s e d th a t th e e x p e r im en ts a llo w e d o n ly
hom ologous r e g io n s to be lo c a t e d . A lth ou gh i t i s r e a s o n a b le to
p ro p o se t h a t o th e r g e n e s a r e c o l in e a r i n th e two v ir u s genom es,
th e r e i s no e v id e n c e to su p p o rt t h i s h y p o t h e s is . Some hom ology
was d e t e c te d w hich d o e s n o t map c o l i n e a r l y . HSV-2 H in d l l l e
h y b r id is e d to some e x t e n t to th e same r e g io n &s HSV-2 Hindi H h .
T h is m igh t r e p r e s e n t a l im i t e d r e g io n o f g e n e t ic n o n - c o l in e a r i t y ,
o r m ight be an a b er ra n t r e s u l t i n t h a t i t d o e s n o t s i g n i f y
f u n c t io n a l c o l i n e a r i t y . A p p a ren tly a b e r r a n t h y b r id is a t io n o f
HSV ( e s p e c i a l l y HSV-2) fra g m en ts c o n ta in in g th e S term in u s to
th e m id d le o f L i n EHV-1 i s d is c u s s e d b e lo w .
The c o n c lu s io n s f o r PRV a r e b ro a d ly i n agreem ent w ith
th o s e o f Rand and B en -P o ra t (1 9 8 0 ) , who d e s c r ib e d th e p a t te r n
o f h y b r id is a t io n betw een th e HSV- and PRY genom es u s in g v i r io n
DNA, n o t in g t h a t m ost o f th e hom ologous se q u e n c e s a re s i t u a t e d
in th e L segm en t. The p a t te r n o f hom ologous r e g io n s in th e
L segm ent o f PRV DNA shown in F ig u re C2*10 d o e s n o t f i t a
c o l in e a r arrangem ent w ith r e s p e c t to HSV DNA. I n v e r s io n o f th e
r e g io n h y b r id is in g to HSV-2 H in d iII h - e - g would le a d to

- 100 -
ap p rox im ate c o l i n e a r i t y betw een th e hom ologous r e g io n s in th e
1 segm ent o f PRY DNA and th e I^ o r i e n t a t io n o f HSY DNA
( i . e . a n a lo g o u s to th e c o l i n e a r i t y o f EHV-1 and HSV DNA).
T h ere fo re th e p o s s i b i l i t y e x i s t s t h a t t h i s r e g io n o f th e PRV
genome i s in v e r t e d w ith r e s p e c t to HSV. The a c t u a l s i t u a t io n
may b e , o f c o u r s e , more com p lex . As w ith EHV-1 DNA, fra g m en ts
from HSV TRg/lR g h y b r id is e d to e q u iv a le n t p o s i t i o n s in PRV DNA,
and th e o r i e n t a t io n o f HSV-1 BamHI k and £ w ith r e s p e c t to ih©
PRV genome i s in agreement with the o r ien ta tio n o f the PRV gene
analogous to HSV-1 Vmw IE 175 (Pow ell, 1979; P.J. Rixon,
P e r so n a l co m m u n ica tio n ).
N o n -c o lin e a r hom ology
N o n -c o lin e a r hom ology was d e te c te d w it h in th e HSV-1
genome and betw een HSV-1 and HSV-2 DNA u s in g th e c r o s s b lo t
t e c h n iq u e . Such hom ology was n o t d e te c te d in ex p er im en ts
where c lo n e d DNA p ro b es and n i t r o c e l l u l o s e s t r i p s were u s e d .
The d i f f e r e n c e i s p ro b a b ly a r e s u l t o f th e d i f f e r e n t t e c h n iq u e s .
The c r o s s b lo t method a l lo w s th e d e t e c t io n o f r e g io n s o f poor
hom ology s in c e a t t h a t p o in t i n th e n i t r o c e l l u l o s e f i l t e r th e r e
i 3 no c o m p e t it io n f o r th e probe by f i l t e r - b o u n d seq u e n c es o f
g r e a t e r hom ology. The s ig n i f i c a n c e o f n o n -c o l in e a r hom ology
i s i n d o u b t. M ost in s t a n c e s in v o lv e th e in v e r t e d r e p e a t s o r
t h e i r ju n c t io n s w ith u n iq u e r e g io n s , s e q u e n c e s known to p o s s e s s
r e g io n s o f i n t r a - o r i n t e r - s t r a i n s i z e v a r i a b i l i t y and, a t
l e a s t i n some in s t a n c e s , to c o n ta in m u lt ip le r e i t e r a t i o n s o f
s h o r t G C -rich seq u e n c e s (L o n sd a le e t a l . , 1 9 8 0 ; S e c t io n p o f
R e s u lt s ; M -J. M urchie and P .J . R ixon , p e r s o n a l c o m m u n ica tio n s).
I t i s p o s s i b l e , t h e r e f o r e , t h a t n o n -c o lin e a r hom ology r e f l e c t s
th e l o c a t io n s o f su ch se q u e n c e s w ith in th e genom e. A s im i la r
c o n s id e r a t io n may be a p p lie d to th e o b serv ed h y b r id is a t io n o f
HSV fra g m e n ts c o n ta in in g th e S term in u s to r e g io n s in th e L

- 101 -
segm ent o f b oth PRY and EHV-1 DNA, and o f s e v e r a l HSV fra g m en ts
to r e p e t i t i v e r e g io n s o f th e PRV and EHV-1 genom es (T a b le s* ^ *
C 2 .4 -C 2 .5 ) . The n o n -c o lin e a r hom ology in t h e s e c a s e s was f a r
more p rom in en t th an t h a t d e t e c te d w ith in HSV-1 DNA or betw een*
th e HSV-1 and HSV-2 genom es, p ro b a b ly a s a r e s u l t o f th e
g e n e r a l ly low l e v e l o f h y b r id is a t io n o f PRV and EHV-1 DNA to
HSV DNA.
G e n e tic r e la t e d n e s s o f h e r p e s v ir u s e s
The main approach to s t u d ie s o f g e n e t ic r e la t e d n e s s among
h e r p e s v ir u s e s h as been an im m u n o lo g ica l o n e . A number o f
t e c h n iq u e s a re w id e ly u se d , in c lu d in g v ir u s n e u t r a l i s a t i o n ,
immune p r e c i p i t a t i o n and im m u n o flu o rescen ce . A n t is e r a may be
r a is e d a g a in s t v i r i o n s , n u c le o c a p s id s , v i r u s - i n f e c t e d c e l l s o r
p u r i f i e d p r o t e in s . The c r i t e r i o n o f r e la t e d n e s s i s w hether
an a n tiseru m produced a g a in s t one v ir u s c r o s s - r e a c t s w ith
a n o th e r v ir u s i n th e t e s t u s e d . The p e r t in e n t p o in t h as been
made, h ow ever, t h a t a n t ig e n s w ith t y p e - s p e c i f i c d e te r m in a n ts
a l s o p o s s e s s type-com mon s i t e s , th u s l i m i t i n g th e c o n c lu s io n s
w h ich may be drawn by t h i s ap proach to h e r p e s v ir u s
r e la t e d n e s s ( K i l l in g t o n e t a l . , 1 9 7 8 ; Yeo e t § 1 . , 1 9 8 1 ) .
The s e a r c h f o r a h e r p e s v ir u s group s p e c i f i c a n t ig e n has
n o n e t h e le s s p roceed ed from th e MCP (Kirkwood e t a l . , 1972;
H oness e t a l . , 1 9 7 4 ) to th e v i r i o n g ly c o p r o t e in s (N o r r ild e t a l . ,
1 9 7 8 ) and a t p r e s e n t c e n t r e s on th e MDHP (Yeo e t a l . , 1 9 8 1 ) .
K il l in g t o n e t a l . (1 9 7 8 ) have p rop osed an im m u n o lo g ica l
c l a s s i f i c a t i o n o f h e r p e s v ir u s e s w ith two m ain c l a s s e s :
a n e u tr o s e r o n , c o n ta in in g v ir u s e s w hich c r o s s - n e u t r a l i s e , and
a s e r o n , c o n ta in in g v ir u s e s w hich c r o s s - r e a c t a n t ig e n i c a l l y
but w hich do n o t n e c e s s a r i ly c r o s s - n e u t r a l i s e . T h is group h as
p r e se n te d e v id e n c e , from p erh ap s th e c l e a r e s t work on th e
s u b j e c t so f a r , th a t HSV-1, HSV-2 and EMV form a n e u tr o se r o n ,

- 102 -
and t h a t th e s e th r e e v i r u s e s p lu s PR? and EHV-1 form a se ro n
( K i l l in g t o n e t a l . , 1977 aad 1978; Yeo e t a l . , 1 9 8 1 ) . The
im p l ic a t io n th a t HSV-2 i s more c l o s e l y r e la t e d to HSV-1 th a n i s
EMV, and t h a t PRV and EHV-1 a re more d i s t a n t l y r e la t e d t o HSV-1,
has th e su p p o rt o f n u c le ic a c id h y b r id is a t io n s t u d ie s (R L eff
e t a l . , 1 9 72; Bronson e t a l . , 1 9 7 2 ; S te r z e t a l . , 1 9 7 4 ) . :
N u c le ic a c id h y b r id is a t io n le n d s i t s e l f a s an a l t e r n a t iv e
ap p roach to th e s tu d y o f g e n e t ic r e la t e d n e s s betw een h e r p e s v ir u s e s .
In th e p a s t , how ever, t h i s te c h n iq u e h as been u n s a t i s f a c t o r y
i n t h a t o n ly g r o s s genom e-to-genom e hom ology was m easured .
M oreover, m ost h e r p e s v ir u s DNAs ch are l e s s th an 5 -1 0 # hom ology,
a l e v e l w h ich by t h i s method i s d i f f i c u l t to d i s t i n g u i s h from
z e r o . The a v a i l a b i l i t y o f c lo n e d DNA fra g m en ts and th e u se o f. .
b lo t h y b r id is a t io n c l e a r l y en h an ces th e u s e f u ln e s s o f
h y b r id is a t io n , a s d em on strated i n t h i s s tu d y . L iq u id
h y b r id is a t io n e x p er im en ts would a l lo w b e t t e r q u a n t i f i c a t io n o f
r e s u l t s , a lth o u g h n u c le o s id e se q u e n c in g o f a n a lo g o u s g e n e s i n
d i f f e r e n t h e r p e s v ir u s genom es w i l l su p p ly th e u l t im a te
e s t im a t io n o f hom ology.
One a p p l i c a t io n o f th e s tu d y p r e s e n te d h er e i s t h a t th e
l o c a t io n s o f EHV-1 g e n e s can be p r e d ic t e d by v i r t u e o f th e
c o l i n e a r i t y betw een hom ologous r e g io n s i n EHY-1 and HSV DNA.
F u r th e r , an unmapped gen e i n e i t h e r EHV-1 o r PRV DNA c o u ld ‘ be
lo c a t e d by u s in g a s a probe th e HSV-1 g e n e , p r o v id in g th e r e i s
s u f f i c i e n t hom ology betw een th e r e s p e c t iv e DNA s e q u e n c e s . For
exam p le , th e MCP gene i n EHV-1 and PRV c o u ld be lo c a t e d u s in g
an HSV-1 MCP gene p ro b e . T h is would o b v ia te th e need to l o c a t e
t h i s f u n c t io n by marker r e s c u e o f a m u ta tio n i n th e g en e , w hich
r e q u ir e s a m utant, o r by a n a ly s i s o f i n t e r t y p i c reco m b in a n ts ,
w hich r e q u ir e s two s e r o ty p e s w ith d i s t i n c t i v e r e s t r i c t i o n and
p o ly p e p t id e p r o f i l e s w hich are a b le to reco m b in e . I t may
p rove p o s s ib l e to l o c a t e w ith HSV-1 p ro b es even a p p a r e n t ly

- 103 -
n on -h om ologou s g e n e s i n PRV o r EHV-1 DNA, su ch a s th e th ym id in e
k in a se g e n e , by d e c r e a s in g th e s tr in g e n c y o f h y b r id is a t io n .
Homolo/ry and re co m b in a tio n i n HSV
I t was co n c lu d ed from th e s t r u c t u r e s o f HSV-l/HSV-2
in t e r t y p i c reco m b in a n ts produced by m arker r e s c u e o f HSV-1 tsD
t h a t r e c o m b in a tio n te n d s to o ccu r in r e g io n s o f g r e a te r
h om ology. In a t te m p tin g to o b ta in th e d i s t r i b u t i o n o f c r o s s o v e r s
betw een th e two ends o f L and S i t was assum ed , and p rob ab ly
n o t u n r e a so n a b ly , t h a t re co m b in a tio n w ith in e i t h e r TRg or IRg
and e i t h e r TR or IR^ o c c u r s w ith eq u a l p r o b a b i l i t y w ith th e
a p p r o p r ia te s&gment i n e i t h e r o r i e n t a t i o n . Even i f t h i s were
n o t th e c a s e , th e c o n c lu s io n would be u n a f f e c t e d . Morse e t a l .
(1977 ) s u g g e s te d from t h e i r l im i t e d s tu d y t h a t c r o s s o v e r s
ten d ed to c l u s t e r i n th e r e g io n o f 0 .4 and 0 .6 2 - 0 .6 9 f r a c t i o n a l
genome u n i t s . An a n a ly s i s o f reco m b in a n ts from s e v e r a l
l a b o r a t o r ie s rev iew ed "by H a ll ib u r to n (1 9 8 0 ) show s a r a th e r good
f i t betw een th e d i s t r i b u t i o n o f c r o s s o v e r s th ro u g h o u t th e
genome and th e d i s t r i b u t i o n o f hom ology p r e se n te d h e r e , w ith th e
e x c e p t io n o f a h ig h fre q u en cy o f c r o s s o v e r s a t 0 .6 2 - 0 .6 9
o b se rv e d by Morse e t a l . (1 9 7 7 a ) but n o t by o th e r g ro u p s.- - - ^ ^
No reco m b in a n t h as y e t been d e s c r ib e d , in c lu d in g th o se
a n a ly se d h e r e , w hich h as a c r o s s o v e r in th e r e g io n o f l e a s t
hom ology betw een th e two genom es, t h a t i s , a t th e TRS/U g and
IRg/Us j u n c t io n s .
Do e s o n ly one HSV genome arrangem ent p a r t ic ip a t e i n reco m b in a tio n ?
The r e s u l t s o f th e work p r e se n te d h ere a re im p o rta n t
w ith r e s p e c t to th e h y p o th e s is t h a t n o t a l l f o u r genome
arran gem en ts may p a r t ic ip a t e i n th e g e n e r a t io n o f v ia b le
in t e r t y p i c rec o m b in a n ts . A n a ly se s o f th e numbers o f c r o s s o v e r s
in i n t e r t y p i c reco m b in a n ts have su g g e s te d t h a t o n ly one o r both
o f th e P and Ig arran gem en ts p a r t ic ip a t e s in reco m b in a tio n

- 104 -
(M orse e t a l . , 1977 ; P r e s to n e t a l . , 1 9 7 8 ; Roizman e t a l . ,
19 7 9 ; W ilk ie e t a l . , 1 9 7 9 b ) . The e v id e n c e i s t h a t genomes
p o s s e s s in g an odd number o f c r o s s o v e r s in UL d is p la y th e l e a s t
number o f c r o s s o v e r s i n th e P o r I g a rra n g em en ts . In d eed ,
t h i s h y p o th e s is f in d s su p p ort i n th e o b s e r v a t io n th a t th e
m a jo r ity o f v i r i o n DNA o f reco m b in a n ts B x l ( 2 8 - l ) and RE4
i s i n th e P o r I g , and I g , a rra n g em en ts , r e s p e c t i v e l y
(P r e s to n e t a l . , 1978; S e c t io n 4 o f R e s u l t s ) . The argum ent
h as th r e e w ea k n esses w hich were em p h asised by i t s p r o p o se r s .
F i r s t l y , i t i s based on a sm a ll number o f reco m b in a n ts w ith an
odd number o f c r o s s o v e r s i n U^: 9 /6 2 reco m b in a n ts produced from
g e n e t ic c r o s s e s in th e s tu d ie d o f Morse e t a l . (1977 .) ,
P r e s to n e t a l . (1 9 7 8 ) , Marsden e t aJL. (1 9 7 8 ) and H a llib u r to n
e t a l . ( 1 9 8 0 ) . The a ssu m p tio n t h a t an^odd number (n ) o f- - • ~ tK * rs
r e c o m b in a tio n e v e n ts i n th e genome i s more l i k e l y to o ccu r
than an even number ( n - f l ) m ust be v iew ed w ith c a u t io n s in c e
th e m a jo r ity o f reco m b in a n ts ( 44/ 6 2 ) p o s s e s s even numbers o f
c r o s s o v e r s i n th e genom e. S e c o n d ly , u n d e te c te d c r o s s o v e r s i n
th e reco m b in a n ts w ith odd numbers o f c r o s s o v e r s i n co u ld
red u ce th e number p u t forw ard a s e v id e n c e . At l e a s t two h^ve been
shown to have an a d d i t io n a l c r o s s o v e r n ea r th e 1 term in u s
( S e c t io n 4 o f R e s u l t s ) . 'T h ir d ly , th e way in w hich reco m b in a n ts
were made may have b ia se d t h e ir o r ie n t a t io n d u rin g
r e c o m b in a tio n . R ecom b in ation i s h o m o lo g y -d ep en d en t, and
th e r e fo r e recom b in ant genome s t r u c t u r e s a r e d ic t a t e d n o t o n ly
by th e p a r t ic u la r m utants u sed i n a g e n e t ic c r o s s and by th e
seq u e n c e s a v a i la b le f o r r e c o m b in a tio n i n th e c a s e o f
marker r e s c u e , but a l s o by th e e x t e n t and d i s t r i b u t i o n o f
hom ology w ith in th o s e se q u e n c e s .
The secon d argum ent in fa v o u r o f th e h y p o th e s is t h a t
no t a l l fo u r arran gem en ts ta k e p a r t in rec o m b in a tio n stem s

- 105 -
from g e n e t ic m apping o f m utant m ark ers. C lem en ts e t a l . (1 9 7 6 )
have made th e p o in t t h a t in v e r s io n o f 1 and S would be
e x p e c te d to le a d to eq u a l rec o m b in a tio n f r e q u e n c ie s betw een
any m arker in L and any marker i n S . That i s , i t would n o t be
p o s s ib l e to a l i g n m arkers i n L w ith r e s p e c t to th o s e i n S .
Brown a t a l . (1 9 7 3 ) o b ta in e d a l i n e a r g e n e t ic map betw een
s e v e r a l m arkers i n and one i n T R g/lR g. U n fo r tu n a te ly ,
Stow e t a l . (1 978) were u n a b le to c o r r e la t e t h i s map w ith th e
p h y s i c a l ly d eterm in ed l o c a t io n s o f th e m ark ers, and t h e r e fo r e i t
was n o t p o s s ib l e to s t a t e w hich o r ie n t a t io n o f th e g e n e t ic
map r e p r e s e n t s . P a r r is e t a l . (1 9 8 0 ) c la im e d t o ” be a b le to
c o r r e la t e th e g e n e t i c a l l y and p h y s i c a l ly d eterm in ed mapping
p o s i t i o n s o f s e v e r a l m u ta t io n s . T h e ir in t e r p r e t a t io n was th a t
m arkers i n L and S were, n o t e q u id is t a n t b eca u se o n ly one o r
b oth o f th e I L and Ig-^ arran gem en ts p a r t i c ip a t e s in g e n e t ic
re c o m b in a tio n ; C le a r ly t h i s i s th e o p p o s it e c o n c lu s io n from
th a t a t t a in e d by argum ent from recom b in an t genome s t r u c t u r e s .
The d a ta p r e se n te d h e r e show t h a t a t l e a s t th e P and 1^
arran gem en ts a re a b le to p a r t ic ip a t e i n t h e g e n e r a t io n o f
v i a b l e r e co m b in a n ts , but t h a t a v a i la b le hom ologous seq u e n c es
p r o fo u n d ly a f f e c t th e fr e q u e n c y o f re co m b in a tio n i n each
o r ie n t a t i o n . Taken t o g e th e r w ith th e above c o n tr a d ic to r y
c o n c lu s io n s , th e r e i s a t p r e s e n t no good r e a so n f o r su p p o sin g
t h a t n o t a l l fo u r arran gem en ts a re a b le to reco m b in e . A
r e c e n t c o n t r ib u t io n i s th e h y p o th e s is o f H oness e t a l . ( 1 9 8 0 ) ,
who p r e s e n te d a d e t a i l e d g e n e t ic a n a ly s i s o f s e v e r a l HSV-1
m ark ers, and co n clu d ed t h a t rec o m b in a tio n i n HSV o c c u r s by a
c i r c u la r o r co n ca te m e r ic m echanism . T h is c o n c lu s io n i s n o t
n eg a ted by in c lu s io n i n th e s tu d y o f HSV-1 t s c 7 5 , w hich had
p r e v io u s ly been e r r o n e o u s ly mapped in each a seq u en ce (K hipe
2-k 9 1 9 7 9 ) , but has s in c e been shown to map o n ly in th e

- 106 -
c se q u e n c e s (P r e s to n , i n p r e s s ) . Two f e a t u r e s o f t h i s m odel
a r e t h a t i t b e t t e r accom m odates i n c l u s i o n o f a l l fo u r
arra n g em en ts i n r e c o m b in a tio n , and t h a t a l l reco m b in a n ts
c o n c e p tu a lly have an even number o f c r o s s o v e r s .

Table 0 2 .1
H y b r id is a t io n o f n ic k t r a n s la t e d recom b in ant p lasm id
c o n t a in in g HSV i)HA fra g m en ts to n i t r o c e l l u l o s e b lo t
s t r i p s o f HSV-1 and HSV-2 BamH I d i g e s t s .
The d a ta a re ta k en from F ig u re C2 A .
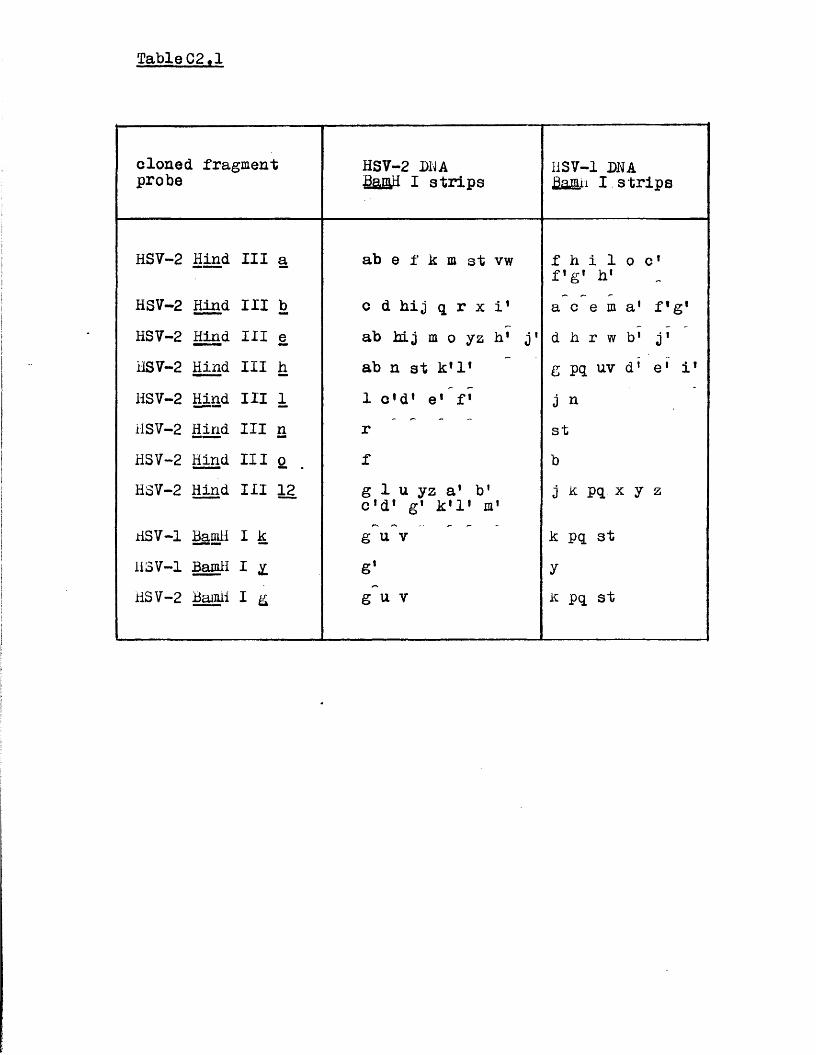
TableC2.1
c lo n e d fragm en t probe
HSV-2 PHA BamH I s t r i p s
HSV-1 DNA flamh I . s t r i p s
HSV-2 Hind I I I a ab e f k m s t vw f h i 1 o c !f ! g ' H1
HSV-2 Hind I I I b c d h i j q r x i* a c e m a' f 'g *
HSV-2 Hind I I I e ab M j m o yz h' j 1 d h r w b 1 j 1
HSV-2 Hind I I I h ab n s t k ! l* g pq uv d ’ e 1 i 1
HSV-2 Hind I I I 1 1 c 1d f e 1 f* j n
HSV-2 Hind I I I n r s t
HSV-2 Hind I I I o f b
HSV-2 Hind I I I 12 g 1 u yz a 1 b' j k pq x y zc ‘d* g 1 k ' l 1 m1
HSV-1 BamH I k g u v k pq s t
HSV-1 BamH I y g* y
HSV-2 BamH I g g u v k pq s t

Table C2.2
H y b r id is a t io n ol' n ic k t r a n s la t e d EHV-1 and PRV
HRA to n i t r o c e l l u l o s e b lo t s t r i p s o f th e HSV-1
BamH I d i g e s t . B ata are tak en from F ig u re C 2 .7 .
BamH I fra g m en ts to which the p rob es h y b r id is e d ar
in d ic a t e d , th e number o f c i r c l e s d i s p la y in g an
e s t im a t io n o f th e amount o f h y b r id is a t io n .
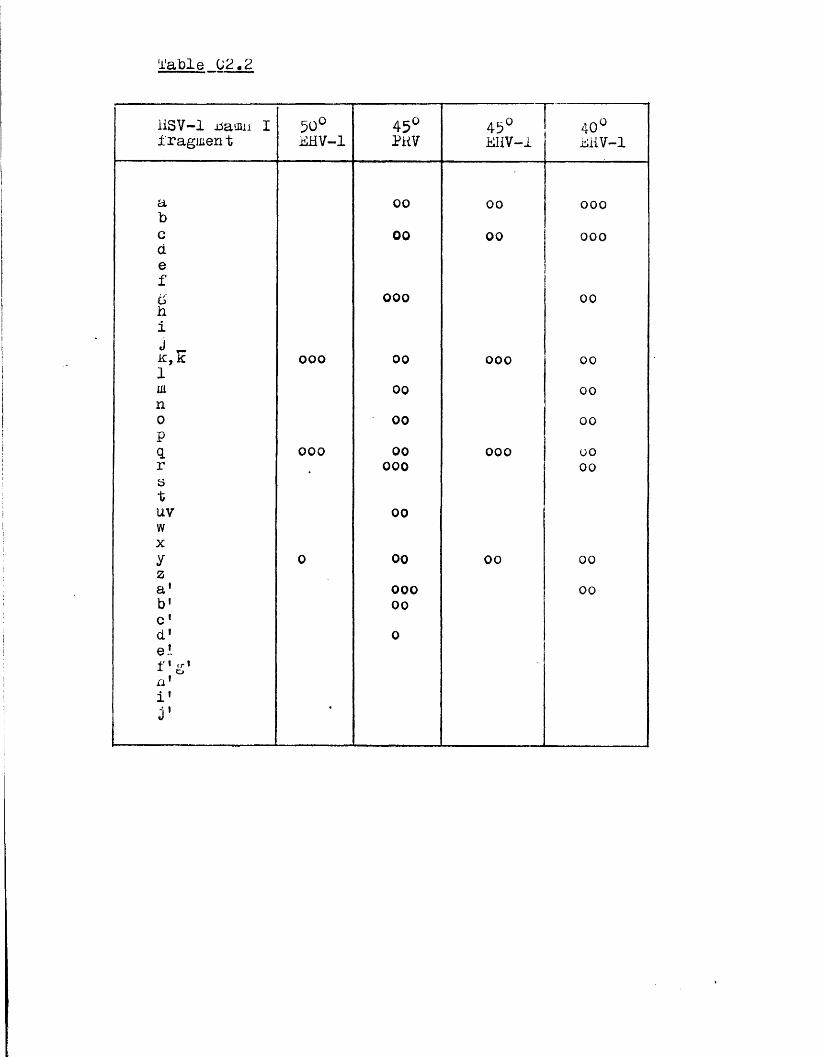
Table 02«2
llSV-1 Haaui I 50° 45° 45° -£* o c
i'r agmen t EHV-1 PRV EIIV-1 EHV-1
ci 00 00 000bc 00 00 000aefiS 000 00hij _ic,k ooo 00 000 001m 00 00n0 00 00Pa 000 00 000 00r 000 00stuv 00wX
y 0 00 00 00za' 000 00b’ 00c 1d.1 0e!T’g 1a 1i fj ’
•

Table C2.5
H y b r id is a t io n o f n ic k t r a n s la t e d EHV-1 and PRV
HislA to n itr o c e l- j -u lo s e b lo t s t r i p s o f th e HSV-2
BamH I d i g e s t . D ata a re tak en from F ig u re C 2 .7 .
BamH I fra g m en ts to w hich th e p ro b es h y b r id ise d
a re in d ic a t e d , th e number o f c i r c l e s d i s p la y in g
an e s t im a t io n o f th e amount o f h y b r id is a t io n .
The u se o f th e H i n d l l l / BcoHI d i g e s t o f HSV-2 DNA
in a n o th e r ex p er im en t con firm ed th a t h y b r id is a t io n
was to HSV-2 BamHI t and n o t a*

Table U2.3
HSV-2 Bamli I fragm en t
50°EHV-1
45°PRV
45°EHV-1
40°EilV-1
a
i1
b 000 0 0c 00 00 000a 000 0e 0f 0 0 0&‘>g 000 00 000 00ni;J 000 000k1m 0n0 00 0P 0 0a • 0r3 t 00 0U 000 000 000 00vwX oyza ’D*c ’ d fe*f '• i6 00 00
hi 000 00i !j* * 000k i i*m’ .
L_ . . . . . . .
00

Table C2.4
H y b r id is a t io n o f n ic k t r a n s la t e d recom binant
p la sm id 3 c o n ta in in g HSV DNA fra g m en ts to n i t r o c e l l u l o s e
b l o t ’s t i i p c o f EHV-1 DNA d i g e s t s .
D ata f o r B g l l l and B g l l l / EooRI d i g e s t s are
ta k en from F ig u r e C2*8. Fragm ents i n p a r e n th e se s
h y b r id is e d more w ea k ly . The tem p eratu re o f h y b r id is a t io n
was 40° e x c e p t f o r EHV-1 BamHI , when i t was 4 5 ° . In th e
l a t t e r ex p er im en t th e in c r e a s e d tem p era tu re s i g n i f i c a n t l y
red u ced h y b r id is a t io n when HSY-2 H ind l l l a , b and h were
u sed a s p r o b e s .
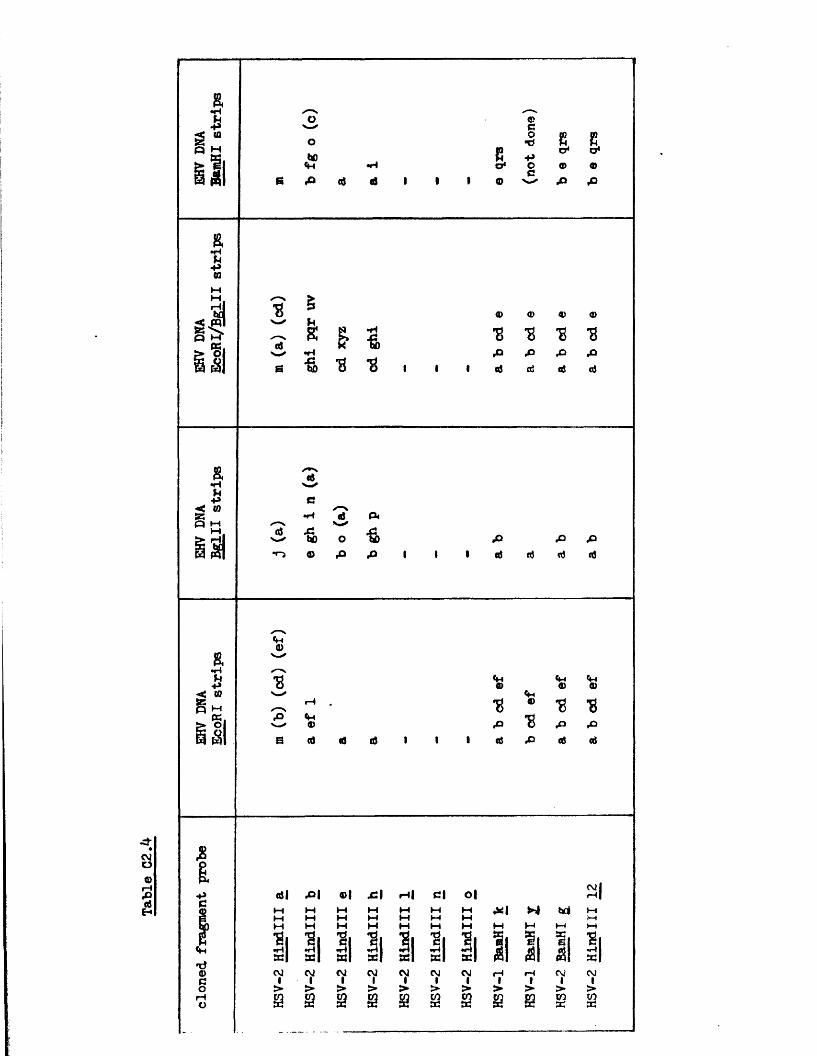
Tabl
e C
ZA
IIto«H
0
©co
•© 8 88 ■p
O1o* ©Cl 0 00 .o ,o
•p03
< p51IS> 3
8 %e. 0 0 0 0
Bs g,
cd ghi *8 *8 ■8 ■8PS•H
ft -8 i.o
1 0&0 0
.©0
&up
■< to
S SSi
/ » \0
' r f 'NwX
c•H
f t
y~\0
o
Pi
f t . o■ o 0 .o 1 1 1 0 0
.©0
.on5
&+» c (0
g £cc> o
Vt0
■8 n—«*
'iov—*B
Vi00 0
%V
Vi0
V0
■80
? *8.© •8 .©
0 ,© 0 0
M
X
r O | 0 1 j e l
M «—* ML J wt jMM
P iM
P IH
' S i d |• g .
OJ CJI I> >
XI(ViI>a a a a

Table C2.5
H y b r id is a t io n o l‘ n ic k t r a n s la t e d recom binant
p la sm id s c o n ta in in g HSV DNA fra g m en ts to
n i t r o c e l l u l o s e b lo t s t r i p s o f PHV d i g e s t s .
D ata are tanen from F ig u re C 2 .9 . Fragm ents in
p a r e n th e se s snowed l e s s h y b r id is a t io n .

Table C2.H
c lo n e d fragm en t p ro be
PRV DMA BamH I s t r i p s
PRV HR A Kpn I s t r i p s
HSV-2 Hind I I I a t> (1 1 ) (o ) a (d )
HSV-2 Hina I I I b c d g c i gh rn
HSV-2 Hind 111 e a (o ) d
HSV-2 Hind I I I h a k c jk
HSV-2 Hind I I I 1 - -
HSV-2 liinct I I I n - -
HSV-2 Hind I I I 0 - -
HSV-2 Hind I I I 2£ i j k o ( a ) (b )
g S i !ef)e HHJk (a )u ; (d) u ) ( i ) -
(b)
llS V -l BamH I k ±1 k 0 (a ) (b )
i l i S i ' 10"
e jk (a )ic 7 (d ) ( i )(1 )
(b )
HSV-1 Hamll i 1 i t e gh
HSV-2 Barnll I & i j k o (a ) (b )
S i S i ' l ° "
e » (a )u ; (a ) u )U ) ■
(b )

Table C2 .6
Genome arran gem en ts o f recom b in an ts produced by
m arker r e s c u e o f 1ISV-1 ts l) w ith HSV-2 EcoR I
j o i n t fr a g m e n ts . The o r ig in a l arran gem en ts o f the
t s l) genome in v o lv e d in reco m b in a tio n were deduced
from th e r e s u l t i n g recom binant s t r u c t u r e s , a c c o r d in
to argum ents d e s c r ib e d in th e t e x t , and a re l i s t e d
w ith th e numbers o f recom b in an ts r e f l e c t i n g
each p a r t ic u la r arrangm ent.
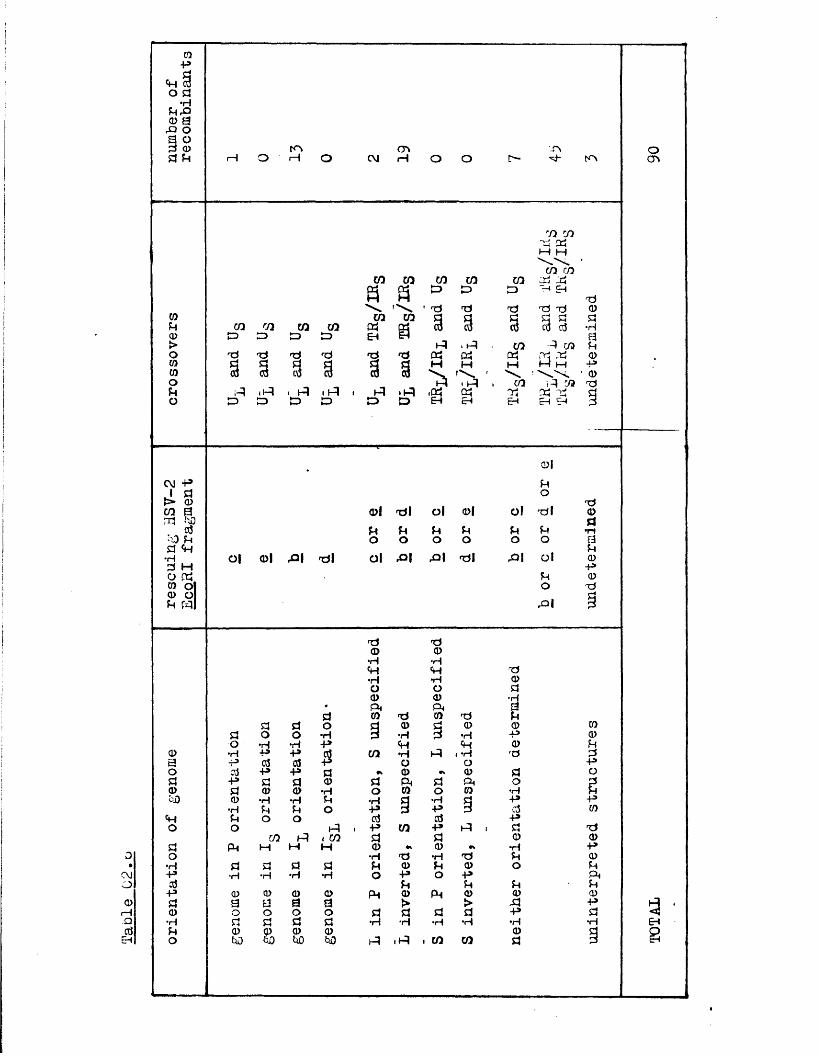
Tabl
e 02
o
COp
<H So a
•HUrOo) arQ Oa o3 O KN 03a u i—1 o i—1 o CM rH O O r - H-
73 03a aM Hi
03 0303 03 03 03 03 'V* wPJu _M
aCD CD CD 'H £h
a"v. \ 1 a a a 73 7j 0to 03 03 q q q rH rH aFi m CQ 03 03 ca Pi 3 3 3 3 a •H0 to CD CD CD EH EH a> h-H . hH 03 a 03 ao a a 1=2 a 'a a « ca cd vd a 0CO q S3 a a q a M M M M M pto 3 3 3 3 3 N ' \ : — • 0o H • iH 03 i a 73 aFi 1h-H . t-H i hI hH .ca cd a a ao CD CD CD CD C3 CD EH EH EH EH EH
---------
olCM P fHJ ^ Ot=> 0 acn a o l a l Ol 0l Ol a l 0rd is a
aj Fi Fi Fh u u a •ri50 Fhf*4 ft_| O O o o o o at .£J HW
•H Ol 0 l 431 a l Ol 43| 431 a l 431 o lH0
3 H po ca Fh 0to ol o a0 o piFh f^l a l
a ao 0•H •H
'H a•H •H 0O O a0 0 •H
• f t f t da to a 0 a Fh
a a o d 0 Si 0 0 0a o o •H 3 •H 3 •H P 0o •H •H P <h <H 0 Fh
0 •H p P a 03 •H i *ri a aa p cd C4 p O O po Cti p P a •t 0 •V 0 a oa P a a 0 a ft a f t o a0 a 0 0 *H o CO o 0 •H FhCjQ 0 •H •H Fi •rH rt •H Si P p
•H fh a O P 3 P 3 a 0«H u o o cd a pO o »-h P 03 p hH , a a
03 h=\ i 03 a a 0 0a f t M M M 0 •* 0 *v a Po •rH a a a Fh 0•H a a a a in 0 Fh 0 o FhP •H •H •H ♦H O p o p ftcd Fh Fh Fh Fhp 0 0 0 0 ft 0 ft 0 0 0d a a a a > > s i P0 o o o o a a a a p a■H a a a a *H •H •H •H •H •HFh 0 0 0 0 0O w 50 50 50 it-H . 03 03 a
oCP>
3EH
I

Figure C2.1
HSV-l/HSV-2 i n t e r t y p i c hom ology. R e s t r ic t io n d ig e s t
o f HSV-1 or HSV-2 D M , in d ic a te d a t th e top o f each
p a ir o f t r a c k s , were t r a n s fe r r e d from th e same g e l
t o n i t r o c e l l u l o s e . The b lo t s t r i p s were h y b r id ise d
w ith n ic k t r a n s la t e d HSV-1 (1 ) or HSV-2 (2 ) DNA, and
a u to r a d io g r a p h e d . The f i v e p a ir s o f tr a c k s on the
l e f t r e p r e s e n t 0 .5 ^ (upper p a n e ls ) and 1 f* a garose4 ^ 4
g e l s ( lo w e r p a n e l s ) . The f i v e p a ir s o f tr a c k s on
th e r ig h t r e p r e s e n t 1 # a g a ro se g e l s , th e low er4
p a n e ls show ing a lo n g e r exp osure o f th e bottom
p a r t o f th e g e l . Tracks o f a p a ir were n o t
a u to ra d io g ra p h ed f o r eq u a l d u r a t io n s . The
tem p era tu re o f h y b r id is a t io n was 4 5 ° .
C/5 9" CM X ^
CM “! > - C/5 CT _ , X CO ^
5 E ^s i -
*■ 0-J*£ o CT « ♦*a
d u ^ M l c i rM R A
T - c o a « t
a *-
« i i l l • I I I ' _ Jk w m » <
rco<u.c :=> —c og-w </i*-M i l 1
> J">N ■« x u i v o f : . -
5 c Nc/5 a X * i -
o "X T -
3 W>0- T-
_ II E CM
>CO m - I
c "a* i-
“ CM<0aX T-
■ M l 1 M l■ M l 1 M
19 uI I f il .
CO -u u u 1) H-C-—.05 ■* - £ 0 £ « - 3E S * 5 > « J o TJ *■ a , j c
X 0) -C
M i l M i l M 1 0 1H H M i l 11 H I M
i u u *o a> t ■fljk
lm
n o Pq
rst u X N -jg -y '0» i - X ' - — *°
* x - o D) c" E
M ■ 1 « « «M M l III
RJ X -Q >*-X • - o « Ol
_ j* —_ C 0 c > - tn«-E a § J X>^, <0 UTOOJ.O^C —
■ M■ M B I"O 0 5 * y —J?S-CD
£ o a crw^ 3 2 . eg -O «
■ •~ - _____ _ ____ " ii -■m m
• l II Idl 00
CD

F ig u r e C 2.2
LEFT HAND FIVE PAIRS OF TRACKS
1ISV-1/HSV-2 i n t e r t y p i c hom ology. R e s t r i c t io n d i g e s t s
o f HSV-2 DNA, in d ic a te d a t th e to p o f each p a ir o f
t r a c k s , were tr a n s fe r r e d from a 0 .5 /2 a g a r o se 0 e l so4
n i t r o c e l l u l o s e . The b lo t s t r i p s were h y b r id is e d
w ith n ic k t r a n s la t e d HSV-1 (1 ) o r HSV-2 (2 ) DKA
a s in d ic a t e u , and a u to ra d io g r a p h e d . Tracks o f
a p a ir were n o t a u to ra d io g ra p h ed f o r eq u a l
d u r a t io n s . The tem p eratu re o f h y b r id is a t io n
was 4 5 ° .
RIGHT HAND SIX PAIRS OF TRACKS
A f f e c t o f tem p eratu re on the h y b r id is a t io n o f
n ic k t r a n s la t e d HSV-2 DNA to b lo t s t r i p s
c o n ta in in g Kpn I or liamH I d i g e s t s o f HSV-1
DNA.
jejI o t s t r i p s were h y b r id is e d w ith HSV-1 (1 ) or
HSV-2 (2 ) proue a t th ree d i f f e r e n t te m p e ra tu r es ,
th en autox’a u io g r a p h e d . Tracks o f a p a ir were n o t
a u to ra d io g ra p n ed fo r eq u a l d u r a t io n s .
Fragm ents o f tnu Kpn I d ig e s t are in d ic a te d on
tn e l e f t , ana damn I on the r i g h t .
IE CM
o COO 00L0
c CMa
IE CMCD mmmO
LA CQ<fr _
Ca CM
IE CMCD
o 00—ca CM
— I
-□■a "av-, E eo o a — . x — c
« • •
■ M S i i a ift P i » 4 l ■ ■ t" •
I *"§ CMm x o o) r'jj: — c a >.*< >x N<o T3 x E o o’ « > >
M R « n 1SOTJJl*. O) .* £X -*-■ _
co co
N CO O ) O * - <N CON W S C O O -J J ' J IA D*-c4 <0 —. 'rn'~ —0) *-
S p i n 0 'T t ® CO o> 5 2 « “ ? x £ ® ®Wirt *"’ Tf

Figure C2.5
Summary o f HSV-l/HSV-2 in t e r t y p i c hom ology d a ta .
A u to ra d io g ra p h s o f n i t r o c e l l u l o s e b lo t s t r i p s o f
HSV-1 or HSV-2 d i g e s t s to w hich n ic k t r a n s la t e d
HSV-1 o r HSV-2 DMA had been h y b r id is e d a t 45° were
a u t o d e n s i t o m e t r ic a l ly scanned and R^ v a lu e s c a lc u la t e d
( s e e t e x t ) . In t h i s diagram mean v a lu e s a re
r e p r e s e n te d h i s t o g r a p h ic a l ly w ith r e s p e c t to
r e s t r i c t i o n maps o f the- HSV genome in th e c o n v e n t io n a l
o r ie n t a t io n ( P ) . Fragm ents w hich c o u ld n o t be
sc o r e d a r e n o t in c lu d e d i n th e h is to g r a m . The
number o f a u to r a d io g ra p h s scanned f o r ea ch d i g e s t i s
shown on th e r ig h t o f each map, and th e v e r t i c a l l i n e
on th e l e f t in d ic a t e s an R^ v a lu e o f 1 . 0 . An
appx*oximate d i s t r i b u t i o n o f i n t e r t y p i c hom ology i s
g iv e n above th e genome a t th e bottom , where la r g e R
v a lu e s in d ic a t e r e g io n s o f h ig h e r hom ology; th e y a r e
c l a s s i f i e d in t o th r e e l e v e l s o n ly .
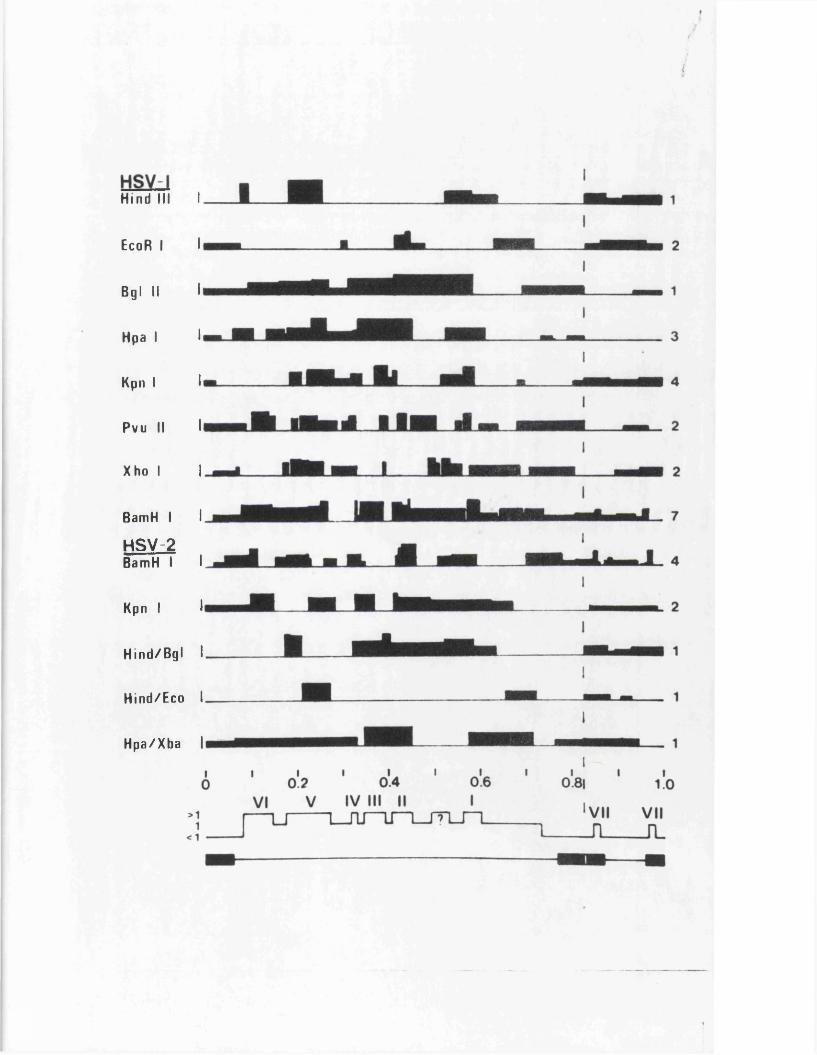
H i n d Hi
E c o R I
Bgl II
H p a I
Kp n I
P v u II
X h o I
B a m H I
HSV-2 B a m H I
Kp n I
H i n d / B g l
H i n d / E c o
H p a / X b a

Figure C2.4
A u to ra d io g ra p h s o f th e r e s u l t s o f h y b r id is a t io n
a t 4 0 ° o f n ic k t r a n s la t e d recom b in an t pAT153
p la sm id 3 c o n t a in in g i n s e r t s o f HSV-1 o r HSV-2 DNA
to b l o t s t r i p s o f HSV-2. BamH I ( l e f t ) o r HSV-1
BamH I ( r i g h t ) . The n a tu r e o f th e HSV DNA i n s e r t
i s in d ic a t e d a t th e top o f each, s t r i p .
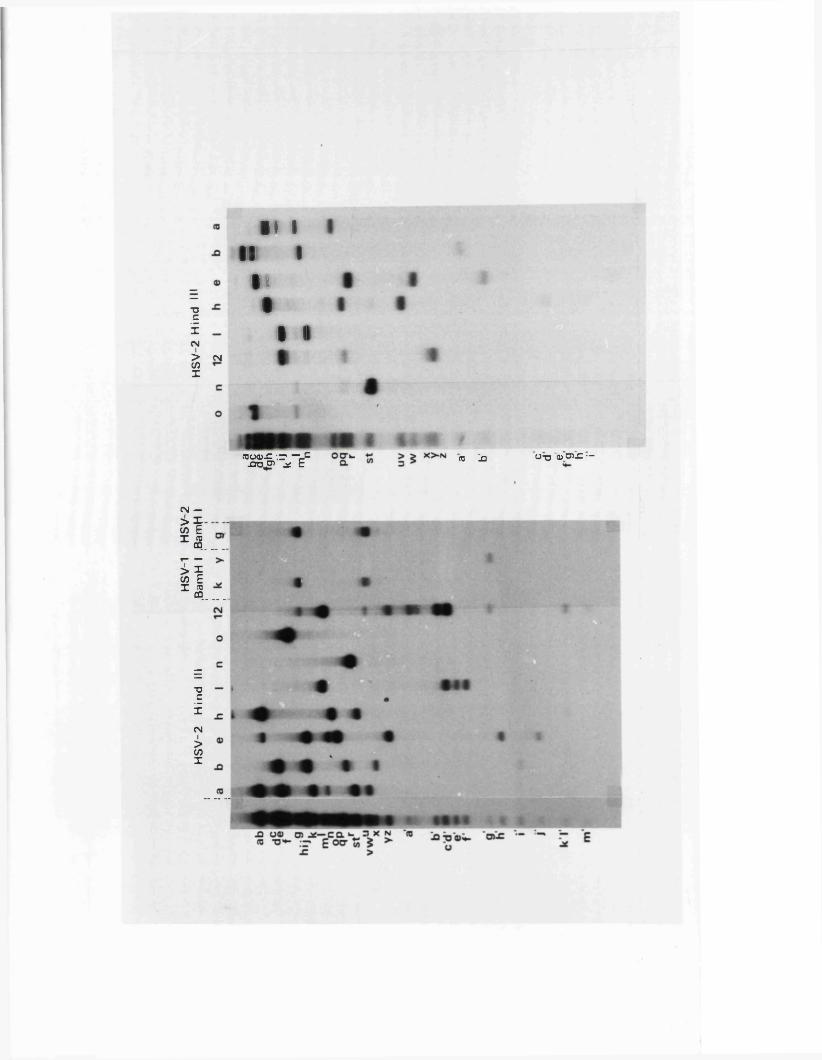
HSV-
1 H
SV-2
HSV-
2 Hi
nd
III Ba
mH
I Ba
mH
I HS
V-2
Hind
III
CM
I I I
I I I
I t •
I I
roO<D-C '.Z' ~ r-C- -CO,™ J* C ro lo ots ’a) oilc —

Figure C2#5
C ross b lo t s o f HSV-1 Kpn I/HSV-
and HSY-1 BamH 1 / HSV-1 BamH I .
d im en sio n i s v e r t i c a l , and th e
d im en sion i s h o r iz o n ta l*
■1 BamH I (u pp er)
The u n la b e l le d
32p_iabelled
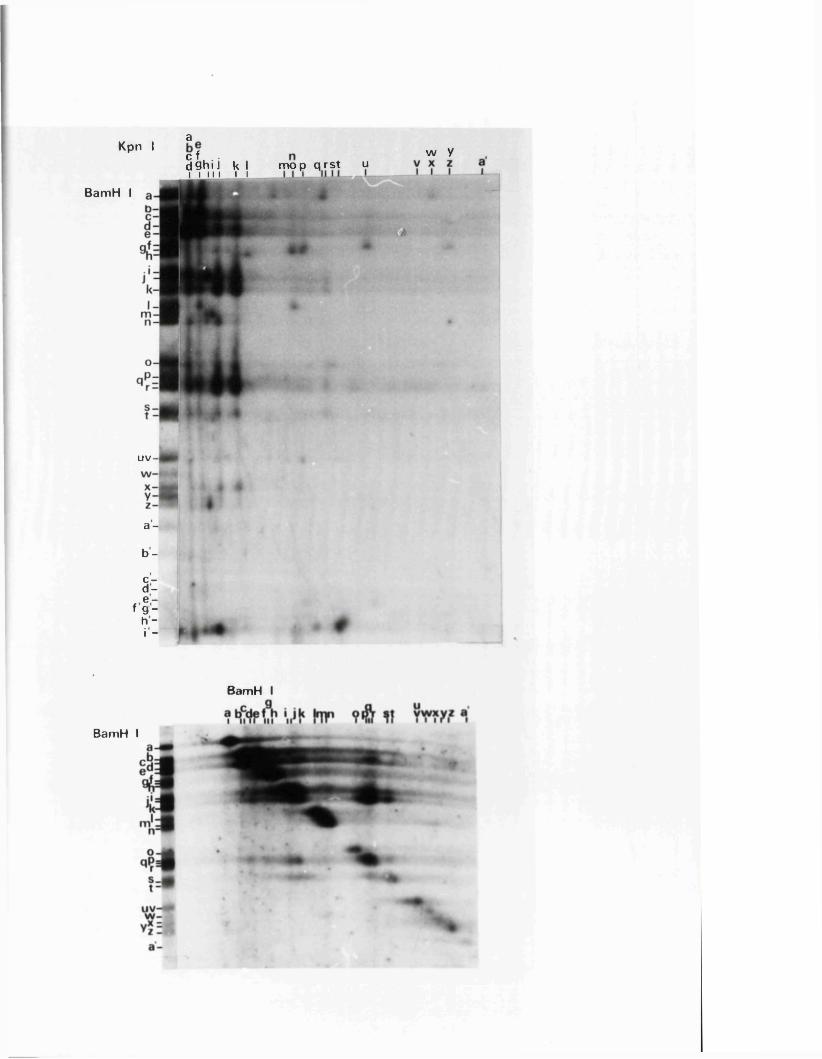
Kpn Ia
c fd 9hi j k Ii it 11 i i mop qrst u
BamH I
w y
. -
uv-
a -b -
c - d - e -
f 9 - h - i -
BamH I
BamH I

• Fig u r e C2»6
C ross b l o t s o f HSV-1 KcoK I/H SV -1 EooR I (u p p er)
and HSV-1 BamH l/H SV -2 BamH I ( lo w e r ) . The
u n la b e l le d d im en sio n i s v e r t i c a l , and th e ^ P - l a b e l l e d
d im en sio n i s h o r iz o n t a l .
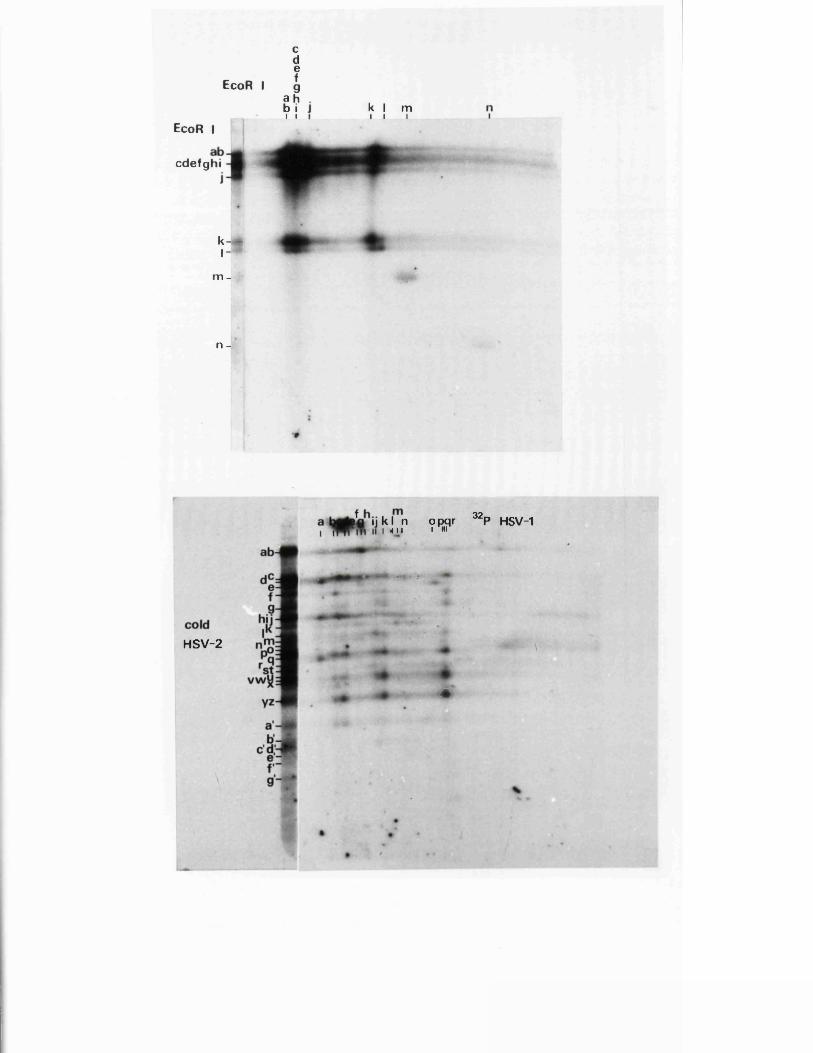
EcoR I
EcoR I
c d e f 9
a h b ji i k Ii i mi
cdefghi
k—#I-*
m -
n -
HSV-2
f i] k I nI 411 opqri Mir
32 P HSV-1
*

Figure C2.7
Homology b etw een HSV and PRV o r EHV-1 UNA.
N ick t r a n s la t e d HSV-1 ( l ) , HSV-2 ( 2 ) , PRV (P) or
EHV-1 (E) DNA v/as h y b r id is e d a t two tem p era tu res
to b lo t s t r i p s o f HSV-1 BamH I o r HSV-2 BamH I ,
a s in d ic a t e d below th e p a n e ls . Exposure tim e
d i f f e r s betw een t r a c k s .

c
LU 1 $ I I i j
▼-
CM 0 #
# # i #
i f i |
c jp
o
§
*£ c a - 3X n <o« ^ r - p o c S 5 >
c >-O "O 03 *»-
ob» A '■— — 'c
-*
LU M i * t ■
CM •• • L t 1
p m K E 5 m •M B 0a o o S ^ ^ c o O’ »- V)~ s r v o t _* £ a > > X>N 3 > OTD03.O)IC —H-
LU i •Q . • • • • I I I 1 1
i si it m * iCM 0 * * *
oin
-Q 0 03 0 3 C O . 3 xw "O'4- A ~ OO" V) $ C >
03 03'o M- Oi'A '■-
E
LU II 9 t I0 - M l M ' • l •CM M i t • • iT— • 1 — m • • » 1
as 00303 •—> ~ CA -a -c ^ E
> 4 - —
O 0"i_ V)*->a uv w X V z '03 ~_Q "O T3 03030
HSV-
1 HS
V-2
HSV-
1 HS
V-2

Fig u r e C 2,8
A u to ra d io g ra p h s o f th e r e s u l t s o f h y b r id is a t io n
a t 40° o f n ic k t r a n s la t e d recom b in an t pAT153
p la sm id s c o n t a in in g i n s e r t s o f HSV-1 o r HSV-2 DMA
to b lo t s t r i p s o f EHV-1 B g l >II/EcoH I ( l e f t ) or
B gl I I ( r ig h t ) d i g e s t s . The n a tu r e o f th e HSV
DiJA i n s e r t i s in d ic a t e d a t th e to p o f each s t r i p .
E xp erim en ts wei’e done t w ic e .

cc— + o o>T o CO LU

figure C2,9
A u to ra d io g ra p h s o f th e r e s u l t s o f h y b r id is a t io n
a t 40° o f n ic k t r a n s la t e d recom b in ant pAT153
p la sm id s c o n t a in in g i n s e r t s o f HSV-1 or HSV-2 DNA
to b l o t s t r i p s o f PRV BamH I . ( l e f t ) or Kpn I
( r ig h t ) d i g e s t s . The n a tu r e o f th e HSV DNA
i n s e r t i s in d ic a t e d a t th e top o f each s t r i p .
E x p erim en ts were done t w ic e .
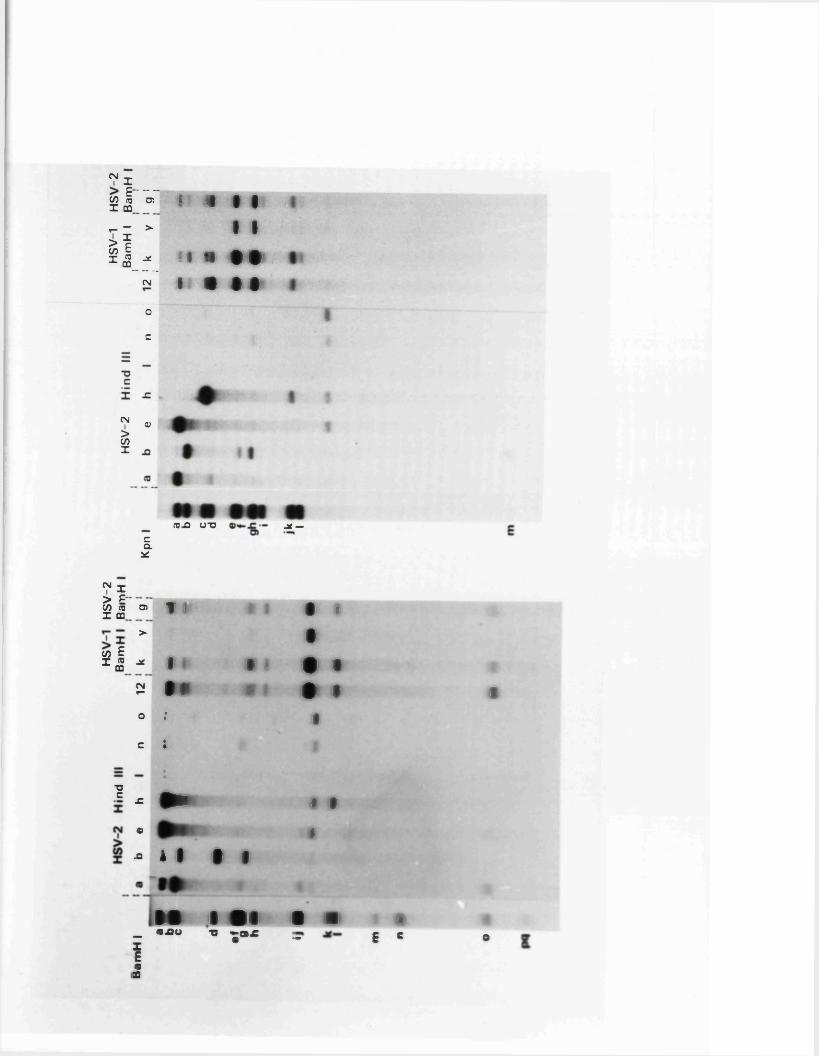
7 1 .......w i 05 rX CO
t- “ >-
MI ,2 -*GO<\l
OC
4 • • I I
l> • t tI • • •
0)CNI>C/)X
Ca*
<b.q o " o je —
1 1 in <o X co
SiCO
■oc
o> \ , • 1> •-* I • 1 • •5! t t • • •o ; •c ;
- m* • f• ••° A • • •
M 1 • • t * •__ l i ) UX
"O "JW-C - E e e
£•CO

F igure C2.10
Summary o f th e d i s t r i b u t i o n o f hom ologous r e g io n s
betw een th e genom es o f HSV-1, HSV-2, EHV-1 and PRV.
The upper p a n e l show s r e s t r i c t i o n maps o f th e HSV-1
and HSV-2 genom es i n th e 1^ a rran gem en t. B la ck r e c t a n g le s
in d ic a t e r e g io n s o f s i g n i f i c a n t l y g r e a t e r hom ology betw een
th e probe and th e b lo t s t r i p s to w h ich i t was h y b r id is e d
( p r o b e /b lo t ) , and h o r iz o n t a l l i n e s f o r EHV-1 and PRV p ro b es
show r e g io n s o f w eaker but d e t e c t a b le h y b r id i s a t io n .
R e s u lt s f o r HSV, EHV-1 and PRV p ro b es a re ta k en from
F ig u r e C 2 .3 , and T a b les C 2*2-C 2*3, u s in g h y b r id is a t io n
tem p er a tu re s o f 45° > 40° and 45° r e s p e c t i v e l y . The sev en
r e g io n s ( I - V I I ) o f g r e a te r hom ology betw een th e HSV-1 and
HSV-2 genom es a re in d ic a t e d , and th e c lo n e d HSV-2 H in d iI I
fra g m en ts w ith in w hich th e y a re lo c a t e d ( a , b , e , h , 1 2 ) .
The two lo w er pa n e l s show th e lo c a t io n s o f
r e s t r i c t i o n fra g m en ts w ith in th e PRV (P o w e ll , 1 9 7 9 ) and
EHV-1 genom es (J.M . W halley, p e r s o n a l com m unication) to
w hich c lo n e d HSV DNA fra g m en ts h y b r id is e d ( f o r n om en cla tu re
s e e T able B 1 . 3 ) . Data a re ta k en from T a b le s C 2 .4 -C 2 .5 ,
and fra g m en ts i n p a r e n th e se s d em o n stra ted w eaker h y b r id is a t io n .
Homologous r e g io n s i n th e EHV-1 genome a l ig n w ith
r e s p e c t to th o se in th e HSV genome i n th e 1^ (o r 13^)
arran gem en t, w h ereas th e PRV and IISV genom es a p p e a r 'n o t to
be c o l in e a r .
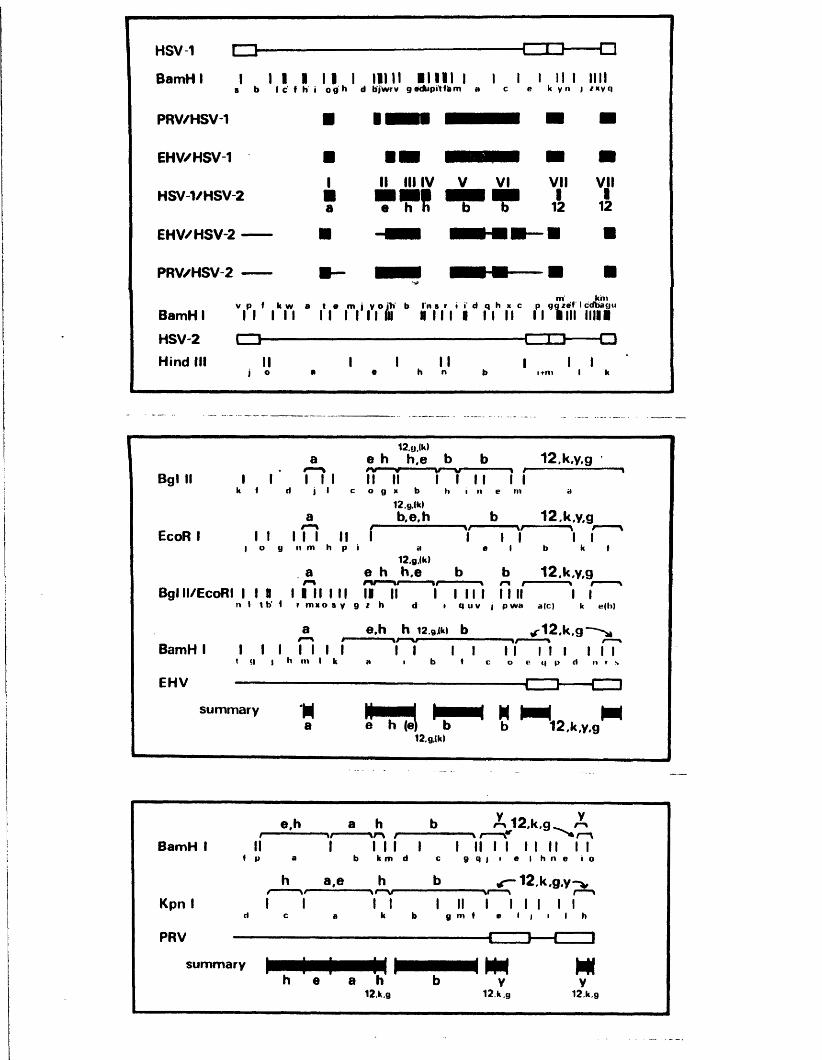
h s v -1 m i — — —------------------------------------------------ 1 n — c
BamH I I I I I I I I III 11 *1111 I I I I II I Mils b I c
PRV/HSV-1
EH V/HSV-1
HSV-VHSV-2
EHV/HSV-2-------
PRV/HSV-2 -------
I■a
■
II III IV V
m m f " be h
k y n i **vn
■ ■
■ ■
VII VII1 i
12 12
■
■ ■
m ^ k inv p t k w a t e m i y o m b I n s r i i d q h x c pgg i conaguBamH I I I I II I I I I 11 III II III I II II II l l l l Hill
HSV-2 C=J------------------------------------------ ----------------- [— 17)------- aHind III II I
i oII
h ni I I
i+nt I k
12,y.|k)a e h h,e b b 12,k,y,g
r - 1 »v 1 v v """i /' ■' "iBgl II I I I I I II II I I I I I I
k I d | I c o g x b h i n e m a12.g.(k)
a b.e.h b 12,k,y.gr-n /—---------------- —\/------------w a / >
EcoR I I I I I I II I I I I I Ij o g n m h p i a e l b k l
12.fl.lk). a e h h.e b b 1 2 (k,y,g
Bgl ll/EcoRI I I fi I Til I II !■* H V I I I I f i M ' I I ‘n I l b' I r m x o s y g / h d i q u v j p w a a(c) k e(b)
a e.h h 12.9jM b________ ^ 1 2 .k ,g—BamH I I I I M I I T l I I M I I I l l T
I <1 | h m I k a ■ b 1 c o e t | p d n r s
EHV
summary * M N p N M N * Ia e h (e) b b 12.k.y.g
12,g,(k)
BamH I
Kpn I
PRV
e.h
II* p
b ^ \ 1 2 . k . g ^ A
i i i 1 P ii i i 11 ii f ?b k m d c g q j i e i h n e i o
a h \n
h a.e/——~-\f — 1 ■I I
b ^ -^ .k .g .y -v11 ii i' 1i i i i r
k b g m f e I j i I h
---------------------------c
summary
d— m
MV
12.k ,g

Figure C 2 . l l
A u torad iograp h o f th e r e s u l t s o f h y b r id is a t io n
. a t 40° o f n ic k t r a n s la t e d HSV-2 ( 1 ,4 ) o r Cl.iV ( 2 ,3 )
BN A to n i t r o c e l l u l o s e b lo t s t r i p s c o n ta in in g th e
HSV-2 BamH I d i g e s t ( 1 ,2 ) or th e CMV Hind I I I
d i g e s t (3 > 4 ) . H e lev a n t r e s t r i c t i o n fra g m en ts
a re i n d ic a t e d , and r e s t r i c t i o n maps o f CMV
(AB169) BNA from W e s ts tr a te e t a l . (1 9 8 0 ) a re
shown, w ith a m o le c u la r w eig h t s c a le (x 1 0 ~^ d a l t o n s ) .
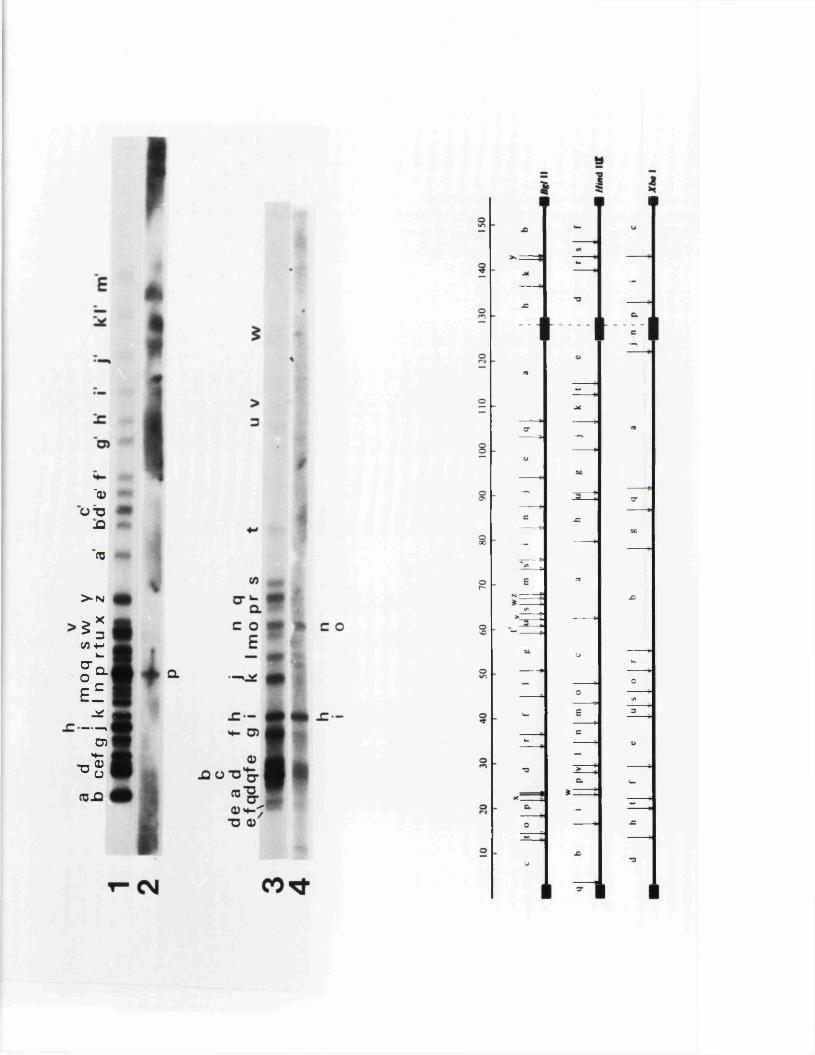
d imoqswy
c cef g
jkln
prtu
x z
a' bd*
e’

Figure C2«12
H istogram o f c r o s s o v e r p o s i t i o n s in th e L segm ent o f
reco m b in a n ts produced by marker r e s c u e o f HSV-1 tsD
w ith HSV-2 EcoRI j o in t fr a g m e n ts . 33 c r o s s o v e r s in
87 reco m b in a n ts o ccu rred a t th e l e f t hand s id e o f 1 ,
and 4 c r o s s o v e r s a t th e r ig h t hand s i d e .
HSV-2 r e s t r i c t i o n s i t e s a re shown a l ig n e d w ith r e s p e c t
to th o s e f o r HSV-1 below th e r e p r e s e n ta t io n o f th e
p o r t io n o f th e L segm ent co v ered by th e r e s c u in g
HSV-2 fr a g m e n ts . The 1 segm ent i s shown r e l a t i v e
t o th e P o r ie n t a t io n of- th e genom e. An e s t im a t io n
o f th e d e g r e e o f in t e r t y p i c hom ology i s shown by th e
t h ic k n e s s o f th e b la ck l i n e .
1 X bal
2 H indiX I
3 EcoR I
4 .B g l U
5 Hpal
6 Kpnl
7 BamHI
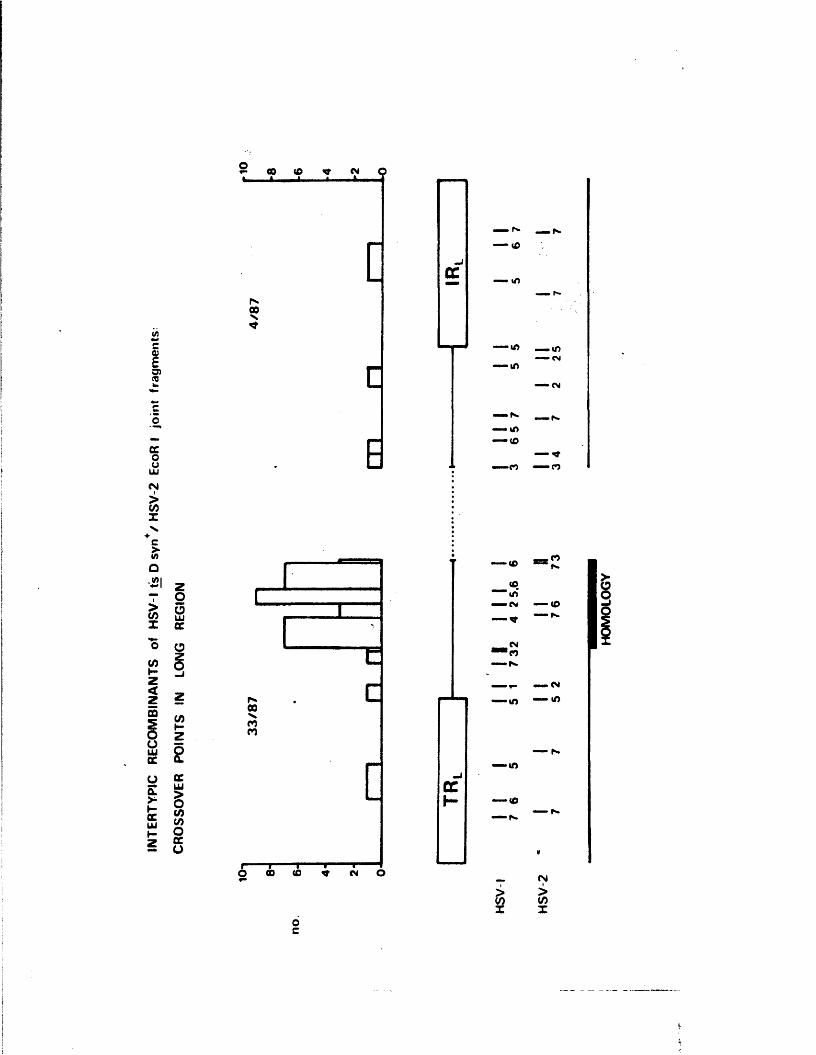
INTE
RTY
PIC
RECO
MBI
NA
NTS
of
HSV
-1 t
s D
syn*
/ HS
V-2
EcoR
I
join
t fr
agm
ents
;
CRO
SSO
VER
PO
INTS
IN
LONG
RE
GIO
N
oc

SECTION 3
NUCLEOTIDE SEQUENCES 01' THE JOINT
BETWEEN THE L AND S SEGMENTS
OP THE GENOMES OP HSV-1 AND HSV-2

- 107 -
RESULTS
I n tr o d u c t io n
The a seq u en ce i s p r e s e n t a s a d i r e c t r e p e t i t i o n a t
th e p h y s ic a l t e r m in i o f th e IISV genome, and a l s o in in v e r te d
o r ie n t a t io n a t th e j o in t between L and S . The o b j e c t iv e o f
t n i s work was to d eterm in e the DNA seq u en ce o f th e j o in t
r e g io n s o f th e genomes o f HSV-1 and HSV-2, in o rd er to
l o c a t e seq u en ce arran gem en ts w hich m igh t be in v o lv e d in th e
f u n c t i o n ( s ) o f th e a seq u e n c e , to e v a lu a te th e co d in g
p o t e n t i a l o f t h i s r e g io n , and to u n d ersta n d th e p r e v io u s ly
o b se rv ed s i z e h e t e r o g e n e i ty in th e HSV-1 j o i n t r e g io n
(Wagner and Summers, 1 9 7 8 ) .
S s t I r e s t r i c t i o n maps o f the HSV-1 and HSV-2 .jo in t r e g io n s
K eclea v a g e ex p er im en ts were perform ed to map th e
r e s t r i c t i o n s i t e s o f S s t I (or i t s is o s c h ia o m e r , S a cI )
w ith in th e llSV-1 and HSV-2 BamHI j o i n t and te r m in a l fr a g m e n ts .
I m p l i c i t in th e f o l lo w in g argum ents i s th e a ssu m p tio n th a t
r e s t r i c t i o n s i t e s in IR^ or IRg are p r e s e n t in i d e n t i c a l
l o c a t io n s in TR o r TRg', r e s p e c t iv e ly .
F ig u re C l ,1 8 shows th a t HSV-1 BamHI k c o n s i s t s o f a
f u s io n o f BamHI q and q , and th a t EcoRI k i s an S ter m in a l
fragm en t w hich e n t i r e l y c o n ta in s BamHI q . The s i z e s o f th e
fo u r S s t I fra g m en ts produced from HSV-1 BamHI k a re g iv e n in
T able C 3 * l, and no s m a lle r fragm en ts were d e te c te d on
p o ly a c r y la m id e g e l s . The r e s u l t s shown in F igu re C 3.1 p a n e l
A a llo w the p o s i t i o n s o f S s t I s i t e s w ith in EcoRI k and BamHI k
to be d e term in ed . Bamll l s gave two fr a g m e n ts , th e la r g e r
o f w hich co m ig ra ted w ith S s t I B ( tr a c k 2 ) . T h erefore th e
s in g l e S s t I s i t e w ith in BamHI q d e f in e s S s t I B a t th e l e f t
hand end o f BamHI k , w ith r e s p e c t to th e c o n v e n t io n a l
4

- 108 -
arrangem ent o f th e genom e, EcoRI k produced th r e e fra g m en ts ,
th e s m a l le s t o f w hich co m ig ra ted w ith S s t I C ( tr a c k 3 ) ,
Only th e la r g e r o f th e o th e r two fr a g m en ts co m ig ra ted w ith
a fra g m en t in th e HSV-1 S s t I d i g e s t (n o t show n), and so
t h i s i s th e S te r m in a l S s t I fra g m en t. T h ere fo re S s t I A
m ust be th e j o in t fra g m en t, c o m p r is in g a f u s io n o f th e
s m a lle r S s t I fragm en t o f BamHI s p lu s th e l a r g e s t S s t I
fragm en t o f EcoRI k . To th e r ig h t o f S s t I A i s C, w hich
wa3 common to th e BamHI k and EcoRI k d i g e s t s . To th e r ig h t
o f S s t I C i s B, th e r ig h t hand end fragm en t o f BamHI k .
T h ere fo re th e o rd er o f S s t I r e s t r i c t i o n fra g m en ts w ith in
HSV-1 BamHI k i s B A C B. F ig u re C 3.3 show s th e S s t I
p h y s ic a l map o f HSV-1 BamHI k .
HSV-2 BamHI g i s th e j o in t fragm en t c o n s i s t in g o f a
f u s io n o f BamHI u and v , EcoRI k and m a r e th e two S te r m in a l
fr a g m e n ts , and EcoRI f i s an 1 te r m in a l fr a g m e n t. T h erefore
BamHI u i s c o n ta in e d e n t i r e l y w ith in EcoRI k and m, and
BamHI v w ith in EcoRI f (F ig u r e C 1 .1 9 ) . BamHI g produced
e ig h t S s t I fr a g m e n ts , o f w h ich £ and 8 w ere more r e a d i ly
r e s o lv e d on p o ly a c r y la m id e g e l s ; fragm en t s i z e s a r e g iv e n in
T able C 3 .1 . F ig u re C 3.1 shows d a ta f o r th e d e d u c tio n o f
S s t I s i t e s w ith in BamHI g., BamHI u produced S s t I 1 , Jj, 6 ,
and, ta k in g in t o a cc o u n t band i n t e n s i t i e s , a fragm en t w hich
c o m ig r a te d w ith £ (F ig u r e C 3.1 p a n e l B tr a c k 2 ) . The
l a t t e r fragm en t m ust be th e S te r m in a l S s t I fra g m en t.
S s t I 6 was n o t produced by EcoRI k o r m and t h e r e fo r e m ust be
th e r ig h t hand fragm en t o f BamHI g , w ith r e s p e c t to th e
c o n v e n t io n a l r e p r e s e n ta t io n o f th e genom e. C lea v a g e o f
HSV-2 Kpnl a , w hich c o n ta in s th e e n t ir e S segm ent e x c e p t fo r
th e te r m in a l 1 x 1 0 6 d a lto n s ( i . e . Kpnl r , s e e F ig u re C l .1 9 ) ,
gave S s t I £ (F ig u re C 3.1 p a n e l D ), w hereas Kpnl r s g a v e ,

- 109 -
among o th e r p r o d u c ts , S s t I 8 (p a n e l E ) . T h ere fo re th e
o rd er o f S s t I fra g m en ts a t th e r ig h t hand s id e o f BamHI g
i s 8 1 £ 6 . S s t I £ a*1*1 1 o r ig in a t e , th e n , from th e l e f t
hand s id e o f BamHI g , and 2 was n o t produced by r e c le a v a g e
o f la r g e r te r m in a l fr a g m e n ts , s u g g e s t in g t h a t i t i s th e S s t I
j o in t fr a g m e n t. EcoRI f gave S s t I 2. an(1 Z n o t £> 311(1
t h e r e fo r e £ maps a t th e l e f t hand end o f BamHI g . In
c o n c lu s io n , th e o rd er o f S s t I fra g m en ts w ith in BamHI g i s
£ (2>Z) 2 8 1 ^ 6 , w ith 2 mapping a t th e j o i n t . The r e l a t i v e
o rd er o f 2 and Z was a s s ig n e d i n th e e lu c id a t io n o f th e
Smal map, and th e f u l l S s t I map i s shown in F ig u re C 3 .3 .
O ther r e s t r i c t i o n mans o f th e HSV-1 and HSV-2 j o in t r e g io n s
O ther maps w ere d eterm in ed l a r g e l y u s in g BamHI j o in t
fra g m en ts i s o l a t e d from recom b in ant p la s m id s . The major
form o f j o in t p r e s e n t in HSV-1 and HSV-2 DNA c o n ta in s a
s in g l e a se q u e n c e , but m inor form s c o n ta in two or more tandem
r e p e a te d a s e q u e n c e s . T h is d i s t r i b u t i o n was r e f l e c t e d in
c lo n e d fr a g m e n ts . The two p la sm id s u sed f o r d ed u c tio n o f
r e s t r i c t i o n maps were k l (HSV-1) and g l (HSV-2) (F ig u r e C 3 .6 ) .
The form er h a s a s in g l e a seq u en ce and th e l a t t e r has tw o .
F u rth er e x p e r im e n ts , u s in g a p la sm id w ith two a seq u en ces
i n th e HSV-1 DNA i n s e r t (1E2 in F ig u re C3#6) and a p lasm id
w ith a s in g l e a seq u en ce i n th e HSV-2 DNA i n s e r t , con firm ed
t h a t m u lt ip le a se q u e n c e s a re p r e s e n t i n each c a s e a s tandem
d i r e c t r e p e a t s . T h is i s d is c u s s e d e ls e w h e r e . R e s t r ic t io n
maps shown in F ig u r e C 3.3 show s in g le a se q u e n c e s a t th e j o i n t .
P la sm id DNA was n ic k t r a n s la t e d , c le a v e d w ith BamHI ,
and th e r e le v a n t j o i n t fragm en t i s o l a t e d from an a g a ro se g e l .
A number o f s in g le and d ou b le d ig e s t io n s were th en done to
map r e s t r i c t i o n s i t e s , a s w e l l a s some p a r t i a l d ig e s t io n s
and d ig e s t io n s o f 5 1- t e r m in a l ly l a b e l l e d DNA. P rod u ct s i z e s

- 11 u -
were c a l ib r a t e d u s in g th e H in fI and H a e l l l d i g e s t s o f
pBK322 DNA ( S u t c l i f f e , 1 9 7 8 ) . The d a ta i s shown in T a b les
C 3.2 and 0 3 -3 , from w hich r e s t r i c t i o n maps were deduced
(F ig u r e C 3 .3 ) . D e t a i l s o f th e argum ent f o r th e Smal map
o f HSV-1 BamHI k a re d e s c r ib e d b elow .
The H in c II and Sau3AI s i t e s in BamHI k were lo c a t e d
(T a b le C 3*2). P a n e ls A and B o f F ig u re 03*2 show d ou b le
d ig e s t io n s o f HSV-1 BamHI k in c lu d in g Sm al. H inc I I c le a v e d
Smal B, S s t I c le a v e d Smal B, E and N, and Sau3AI c le a v e d
Smal A and B. C lea v a g e o f i s o l a t e d S s t I and Sau3AI
fra g m en ts a llo w e d Smal fra g m en ts to be a s s ig n e d to r e g io n s
o f BamHI k a s f o l lo w s (T ab le C 3 ,2 ) .
i f W fi A (F ,R ) B (Mf fl) E 0
32The o rd er w ith in Sau3AI A was d eterm in ed by P - l a b e l l i n g
t h i s fragm en t a t th e 5 1 te r m in i w ith T4 p o ly n u c le o t id e k in a s e ,
d ig e s t in g w ith S s t I and s e p a r a t in g th e two u n iq u e ly la b e l l e d
fra g m en ts by a g a ro se g e l e l e c t r o p h o r e s i s . The two fra g m en ts
were th en p a r t i a l l y d ig e s t e d w ith Smal and s i z e s o f p ro d u c ts
d eterm in ed by com fjarison w ith m o le c u la r w e ig h t m arkers
(F ig u r e C 3.2 p a n e l F ) , The p ro d u ct s i z e s co rresp o n d to
s e q u e n t ia l a d d it io n s o f Sima I fra g m en ts to th e l a b e l l e d
te r m in a l Smal fra g m en t, so th a t th e s i z e in c r e m e n ts co rresp o n d
to Smal fragm en t s i z e s . The p a r t i a l d ig e s t io n p ro d u c ts a re
shown in T able C 3 .2 , and g iv e th e o rd er o f Smal fra g m en ts
w ith in Sau3AI A a s J C L D N H I K P G , w ith 430 bp o f
Smal A betw een Smal G and th e Sau3AI s i t e ( th e Sau3AI s i t e
c o in c id e s w ith th e BamHI s i t e a t th e l e f t s id e o f J ) .
The o n ly a m b ig u ity i s in th e p o s i t i o n s o f Smal G and H,
s in c e th ey a re s im i la r i n s i z e . They were unam biguously
a s s ig n e d by f i n e mapping o f th e H in f l fragm en t c o n ta in in g

- I l l -
Smal A, G, K, P and p a r t o f I , w hich i s d e s c r ib e d l a t e r .
The o n ly fragm en t common to th e j o i n t and b oth te r m in i was
Smal G (T ab le C 3 .2 ) , s u g g e s t in g th a t t h i s fragm en t i s
c o n ta in e d c o m p le te ly w ith in th e q se q u e n c e .
The o rd er o f Smal F and R was a s c e r ta in e d by p a r t i a l l y
d ig e s t in g Sau3AI B (F ig u re C 3.2 p a n e l C ), The o rd er o f
fra g m en ts w ith in Sau3AI B from d ou b le d ig e s t io n i s , in bp,
6 4 0 - (2 4 ,4 5 0 ) - 4 9 5 , where th e unordered fra g m en ts Smal F and R
a re 450 and 24 bp r e s p e c t iv e ly (a s c a l ib r a t e d on a g a r o se ;
F m ig ra ted a s 410 bp on p o ly a c r y la m id e ) . The
p r e d ic te d p a r t i a l d ig e s t io n p r o d u c ts from th e two p o s s ib l e
arran gem en ts a re shown in T ab le C3*2 p a r t I . The th r e e
la r g e r p a r t i a l d ig e s t io n p ro d u c ts were 1 1 1 5 , 970 and 945 bp
(F ig u r e C 3.2 p a n e l C ), .su p p o rtin g th e o r d e r o f th e two
fra g m en ts a s Smal R F. The o th e r two e x p e c te d p a r t i a l
d ig e s t io n p ro d u cts o f 665 and 475 bp were n o t d i s t in g u is h e d
p ro b a b ly b ecau se 665 bp d id n o t r e s o lv e from 640 bp, and
475 bp d id n o t r e s o lv e from 495 bp. The i n t e n s i t i e s o f th e
bands a t 640 and 495 bp were n o t d e c r e a se d to th e same
e x t e n t a s th e 450 bp (Sm al F) band by u s in g l e s s Smal in th e
d i g e s t i o n . The o rd er was con firm ed a s R F by p a r t ia l
d ig e s t io n o f S s t I A, when F-R was d e t e c te d b ut n o t 750-R ,
where 750 bp i s th e p a r t o f Smal B c o n ta in e d w ith in S s t I A
(F ig u r e 0 3 ,2 p a n e l E ) .
Smal M and £ were o rd ered by p a r t i a l d ig e s t io n o f S s t I C.
The o rd er o f fra g m en ts w ith in S s t I C from d o u b le d ig e s t io n
i s , in bp, 7 8 - ( 9 1 ,3 8 ) - 2 6 0 , where th e u n ord ered fra g m en ts
Smal M and a re 91 and 38 bp, r e s p e c t i v e l y . The p a r t i a l
d ig e s t io n p r o d u c ts p r e d ic te d from th e two p o s s ib l e arran gem en ts

- 112 -t
a r e shown in T ab le C 3.2 p a r t J . The a c t u a l p r o d u c ts , shown
in F ig u re C 3.2 p a n e l D, g iv e th e o rd er a s Smal M The f u l l
o rd er o f Smal fra g m en ts in HSV-1 BamHI k i s shown in F ig u re
C 3 .3 . The o th e r r e s t r i c t i o n maps shown in t h i s F ig u re were
d eterm in ed s im i l a r l y by c o n s id e r a t io n o f th e d a ta in T a b le s
C 3.2 and C 3 .3 .
Si z e v a r i a b i l i t y in th e .jo in t r e g io n
R e c e n t ly p la q u e -p u r if ie d s to c k s o f HSV-1 s t r a in 17 and
HSV-2 s t r a in HG52 were p la te d on BHK c e l l s , grown under
Biiu2, and a s in g le p laq u e o f each p ic k e d . Ten su b p la q u es
o f each were i s o l a t e d , p la q u e -p u r if ie d t w ic e m ore, and a
v ir u s s to c k p rep ared a t 37° £ o r each p a r e n t and progeny■50
p la q u e . V ir io n DMA was P - la b e l l e d in v i v o and BamHI
r e s t r i c t i o n p r o f i l e s compared (F ig u r e C 3 .4 ) . T erm inal
and j o i n t fra g m en ts o f HSV-1 DNA w ere th e m ost v a r ia b le in
s i z e (BamHI k , q , s ) , and v a r ia t io n s i n th e j o i n t and S
te r m in a l fra g m en ts (BamHI k and q , r e s p e c t i v e l y ) seemed
c o r r e la t e d . V a r ia t io n was shown to be in q and n o t p ,
w hich co m ig ra ted i n some c a s e s , by d ig e s t io n w ith o th e r
r e s t r i c t i o n e n d o n u c le a se s . L ess o b v io u s v a r ia t io n was
ob serv ed i n th e m o b i l i t i e s o f HSV-1 BamHI n and x , w hich
span th e ju n c t io n s betw een Ug and TRg/IRg ( e . g . compare
tr a c k s 2 and 5)., .and in BamHI z , w h ich maps i n Ug ( e . g .
compare tr a c k s 5 and 6 ) . In c o n t r a s t , HSV-2 i s o l a t e s
v a r ie d o n ly in BamHI z and a 1, w hich span th e ju n c t io n s
betw een U3 and TR^/IR^, and no v a r ia t io n i n HSV-2 j o in t
(BamHI g) o r te r m in a l fra g m en ts (BamHI u and v ) was
d e t e c t e d . V a r ia b i l i t y was shown to r e s id e in z and n o t
y by d ig e s t io n w ith o th e r r e s t r i c t i o n e n d o n u c le a se s .
The r e g io n o f v a r i a b i l i t y in th e HSV-1 j o in t r e g io n
was lo c a t e d more p r e c i s e l y by Smal d ig e s t io n o f BamHI

- 113 --• . 32j o i n t fra g m en ts i s o l a t e d from in v iv o P—l a b e l l e d v ir io n .
Dll A (P ig u re C 3 .5 )« The j o in t r e g io n o f c l o n a l l y r e la t e d
HSV-2 i s o l a t e s was a n a ly se d s im i la r ly (P ig u r e C 3 .6 ) .
I1SV—1 Smal G v a r ie d in s i z e betw een 275 and 400 bp see m in g ly
by th e in s e r t i o n or d e l e t io n o f m u lt ip le s o f a p p ro x im a te ly
1 1 -1 2 bp. HSV-1 Smal A e x h ib ite d s i z e v a r i a b i l i t y o f up to
400 bp w hich was in d ep en d en t o f th a t in Smal G. D ig e s t io n
o f BamHI k from f i v e o f th e HSV-1 i s o l a t e s w ith Sau3AI and
Tag I a llo w e d th e v a r ia b le r e g io n i n Smal A to be more
a c c u r a t e ly lo c a t e d betw een th e Sau3AI A-B and Tag I A-D s i t e s
(P ig u r e 0 3 * 5 , maps in P ig u re C3*3)* R e c lea v a g e o f BamHI
fra g m en ts c o n ta in in g th e genom ic t e r m in i show ed, f o r each
i s o l a t e , th e same s i z e d i s t r i b u t i o n o f v a r ia b le fra g m en ts
a s in th e j o in t r e g io n . No v a r ia t io n i n th e BamHI j o i n t
fra g m en ts o f th e HSV-2 i s o l a t e s was d e t e c te d by c le a v a g e
w ith Smal (P ig u re 0 3 .6 ) or S s t I .
HSV-1 BamHI j o i n t fra g m en ts i s o l a t e d from p la sm id s ,
l i k e th o se i s o l a t e d d i r e c t l y from v ir io n DNA, showed s i z e
v a r ia t io n in Smal A and G (P ig u r e C 3 .6 ) , w hereas no v a r ia t io n
was o b serv ed in c lo n e d HSV-2 j o i n t fr a g m e n ts , e x c e p t th a t
due to th e p r e se n c e o f an e x tr a a seq u en ce a s d e s c r ib e d
b e lo w . One o f th e HSV-1 c lo n e s , k 2 , had a s m a lle r Smal I
fra g m en t, a lth o u g h v a r i a b i l i t y in t h i s fragm en t was n o t
o b serv ed in v ir io n DNA.
A lthough HSV-1 and HSV-2 d i f f e r in th a t th e form er
showed s i z e v a r i a b i l i t y in th e j o i n t r e g io n and th e l a t t e r
d id n o t , th e y both p o s s e s s a secon d ty p e o f h e t e r o g e n e ity
in th e j o in t r e g io n and a t th e L term in u s w hich c o n s i s t s o f
i n s e r t i o n s o f a p p r o x im a te ly 300 bp th o u g h t to c o n ta in th e
a s e q u e n c e . The m a jo r ity o f HSV DNA m o le c u le s h as a s in g l e
a seq u en ce a t th e j o i n t , w h ile a s m a lle r p r o p o r t io n h a s two

- 114 -
or m ore. T h is i s r e f l e c t e d in c lo n ed HSV-1 j o in t fra g m en ts ,
m ost o f w hich had a s in g le a seq uence (kl, k 2 , k3 in P igu re
C 3 .6 ) , but some o f w hich had two (El, 3E2). P la sm id s El and
§2 each had two Smal G fra g m en ts , th e l a t t e r w ith two o f th e
same s i z e but th e form er w ith two o f d i f f e r e n t s i z e s . Smal
p r o f i l e s were c o n s is t e n t w ith the p r e se n c e o f th e two a
se q u e n c e s a s tandem d i r e c t r e p e a ts ( s e e P ig u re C 3 .1 3 ) .
S im ila r r e s u l t s were o b ta in ed w ith c lo n e d HSV-2 j o in t fr a g m e n ts ,
and an i s o l a t e w ith two a seq u en ces ( g l ) i s in c lu d e d in
P ig u re C3*6.
Homology betw een th e .jo in t r e g io n s o f HSV-1 and HSV-2 DNA
N it r o c e l lu lo s e b lo t s c o n ta in in g Smal fra g m en ts o f
HSV-1 and HSV-2 BamHI j o in t fragm en ts were p rep ared from
p la sm id DNA (k l and g l .in P igu re C 3 .6 ) by t r a n s f e r from 2 f*
a g a ro se or 5# p o ly a cry la m id e g e ls * S t r ip s were probed w ith
n ic k t r a n s la t e d p lasm id DNA c o n ta in in g th e j o i n t fragm en ts*
R e s u lt s from p o ly a cry la m id e g e l s a re shown in P ig u re C 3 .7 .
fra g m en ts la r g e r than 1000 bp and s m a lle r th an 150 bp d id n o t
t r a n s f e r e f f i c i e n t l y , and th e r e fo r e o n ly HSV-1 Smal B -J and
HSV-2 Smal 2 -6 were r e s o lv e d a f t e r h y b r id is a t io n . The
l a r g e r fra g m en ts t r a n s fe r r e d s a t i s f a c t o r i l y from a g a ro se
g e l s . Homologous p rob es f a i l e d to show e x p e c te d h y b r id is a t io n
to fra g m en ts mapping in th e a seq u e n ces (HSV-1 Smal G, HSV-2
Smal £ and J) and so i t was n o t p o s s ib le to compare hom ology
betw een th e iiSV-1 and HSV-2 a s e q u e n c e s . T h is e f f e c t co u ld
have been due e i t h e r to i n e f f i c i e n t t r a n s f e r o f th e s e
fra g m en ts to n i t r o c e l l u l o s e or to i n e f f e c i e n t h y b r id is a t io n .
HSV-1 p lasm id probe h y b r id ise d to HSV-2 Smal 1 (a g a r o se
g e l s ) and £ , and HSV-2 probe h y b r id is e d to HSV-1 Smal
B, S, P and H (th e S s t I/S m al double d i g e s t showed th a t
h y b r id is a t io n d id n o t occur to D ). These r e s u l t s are

- 115 -
in te r p r e te d , in term s o f a r e g io n o f i n t e r t y p i c hom ology
e x te n d in g th ro u g h HSV-1 Smal P, B and B (HSV-2 Smal 1 ) in
th e c s e q u e n c e s , and a r e g io n o f l e s s hom ology betw een HSV-1
Smal H and HSV-2 Smal ± in th e b s e q u e n c e s .
P in e r e s t r i c t i o n maps o f th e .jo in t r e g io n s o f HSV-1 and HSV-2
B e fo r e a t te m p tin g to d eterm in e th e n u c le o t id e seq u e n ces
o f th e j o in t r e g io n s o f HSV-1 and HSV-2 DNA i t was n e c e s sa r y
to map f u r t h e r r e s t r i c t i o n s i t e s i n t h e s e r e g io n s . N ick
t r a n s la t e d HSV-1 BamHI k Sau5AI A from p la sm id k l was
c le a v e d w ith a number o f e n d o n u c le a se s and a l s o d ou b le
d ig e s t e d w ith Smal . H in fl a p p a r e n t ly d id n o t c le a v e Smal
G, K, P and 430 bp (p a r t o f Smal A) b ut d id c le a v e Smal I
(P ig u r e C 3.8 p a n e l A ). I s o l a t io n o f th e l a r g e s t Sau3 A l/H in fI
fragm en t o f 1130 bp and* d ig e s t io n w ith Smal showed t h a t i t
c o n ta in e d Smal G, K, P and two fr a g m en ts o f 410 bp (430 bp
on a g a r o se ; p a r t o f Smal A) and 160 bp (p a r t o f Smal I ) .
The l a r g e s t Hinf l / BamHI fragm en t from p la sm id k l (a H in fl
fragm en t o f 1760 bp) c o n ta in e d Smal G, K, P and two fra g m en ts
o f 1040 bp (n e a r ly a l l o f Smal A) and 160 bp (p a r t o f Smal I ) .
A number o f s in g le and d o u b le d i g e s t s were done to map t h i s
fra g m en t, w hich c o n ta in s th e HSV-1 j o i n t a se q u e n c e . D ata a re
summarised in Table 0 3 .4 , and r e s t r i c t i o n maps a re
shown i n P ig u re C 3 .9 . HSV-2 BamHI g S s t I 2 and Smal 2 were
mapped s im i la r ly ( s e e P ig u r e 03*3 f o r th e l o c a t io n s o f t h e s e
fr a g m e n ts ) . D ata a re summarised i n T ab le 0 3 . and maps
a r e shown in P ig u re 0 3 .9 .
Lo c a t io n o f th e v a r ia b le r e g io n w ith in th e HSV-1 a seq u en ce
P ig u re 0 3 .1 0 shows a com parison o f d i g e s t s o f th e
Sau3 A l/Hinf l j o in t fragm en t from p la sm id s k l and k 2 , in th e

- 116 -
l a t t e r o f w hich Smal G i s la r g e r (P ig u r e C 3 .6 ) . The fra g m en ts
w hich a re la r g e r in k2 than in k l a r e , in hp, Smal - 340 ,
A v a il - 680 , B stN I - 1 0 0 0 , A val - 3 4 0 , Hael l - 9 8 0 , B g l l - 9 3 0 ,
S s t l l - 320 ( k l s i z e s g iv e n ) . The s i z e d i f f e r e n c e i s ab ou t
40 bp. The S s t l l r e s u l t p la c e s th e r e g io n o f v a r i a b i l i t y
w ith in th e a seq u en ce in th e 320 bp fr a g m e n t. Taken in
c o n ju n c t io n w ith th e Smal r e s u l t , th e v a r ia b le r e g io n r e s id e s
i n th e 270 bp betw een th e Smal s i t e a b o u t 40 bp from th e
l e f t hand end o f th e a seq u en ce and th e S s t l l s i t e ab ou t
100 bp from i t s r ig h t hand end , a c c o r d in g to th e maps shown
in P ig u re C 3 .9 .
De f i n i t i o n o f th e genom ic te r m in i and lo c a t io n o f th e a seq u en ce
P ig u re 0 3 .9 show s th e s t r a t e g i e s a d op ted to d eterm in e
th e n u c le o t id e se q u e n c e s shown i n P ig u re 0 3 * 1 1 . S eq u en ces
were d eterm in ed f o r th e j o i n t r e g io n s o f HSV-1 s t r a i n s 17
and USA-8 and HSV-2 s t r a in HG52, and a l s o f o r th e r e g io n
sp an n in g th e ju n c t io n betw een two d i r e c t l y r e p e a te d a se q u e n c es
in HSV-1 s t r a in 17 and HSV-2 s t r a in HG52.
The a seq u en ce i s d e f in e d a s th e seq u en ce p r e s e n t a s
d i r e c t r e p e a t s a t th e genom ic te r m in i and a s an in v e r te d
r e p e a t a t th e th e j o i n t betw een th e 1 and S seg m en ts . Hence
th e te r m in i may be a l ig n e d w ith th e j o in t r e g io n a s shown in
P ig u re 0 3 .1 3 . The p o s i t i o n o f th e a seq u en ce a t th e j o i n t
ca n n o t be d eterm in ed d i r e c t l y from th e j o i n t n u c le o t id e
se q u e n c e , but i t can be d eterm in ed by l o c a t in g th e t e r m in i
w ith r e s p e c t to th e j o in t n u c le o t id e se q u e n c e , u s in g t h e i r
common r e s t r i c t i o n s i t e s . I t i s assum ed, a lth o u g h i t h as
n o t been p ro v en , t h a t th e te r m in a l and j o i n t a se q u e n c e s are
i d e n t i c a l .
ilSV-1 DIM A was ^ P -^ L ab elled s p e c i f i c a l l y a t th e 5 1

- 117 -
te r m in i and s i z e s o f genom ic te r m in a l r e s t r i c t i o n fra g m en ts
d eterm in ed (P ig u r e C 3 .1 2 ) . T h is method l o c a t e s th e 5 *
te r m in i o f th e genome, and n o t th e 3 ' t e r m in i , w ith r e s p e c t
to th e j o i n t s e q u e n c e . The s i z e s in n u c le o t id e s f o r th e
S term in u s o f s t r a in 17 w ere: Smal 3 6 -3 7 , H a e l l l 2 3 -2 4 .
The d i f f e r e n c e in s i z e o f t h e s e two fra g m en ts i s two
n u c le o t id e s l e s s th an would be p r e d ic te d from th e j o in t
n u c le o t id e seq u en ce in P ig u re 0 3 * 1 1 , in w hich th e Smal and
H a e l l l s i t e s are lo c a t e d a f t e r n u c le o t id e s 367 and 352
r e s p e c t i v e l y . T h is d isc r e p a n c y i s p ro b a b ly a r e s u l t o f a
h ig h p r o p o r t io n o f C m o ie t ie s (1 0 /1 5 ) in th e l a b e l l e d s tra n d
betw een th e two s i t e s , w hich in c r e a s e s th e o v e r a l l ch arge to
mass r a t i o o f th e la r g e r fra g m en t, c a u s in g i t to m ig ra te
f a s t e r in th e g e l . I f t h i s i s s o , th e s m a lle r Hael l l
te r m in a l fragm en t p r o v id e s th e more a c c u r a te e s t im a t e ,
p la c in g th e S term in u s a t 3 2 9 -3 3 0 on th e j o in t se q u e n c e .
The s i z e s in n u c le o t id e s f o r th e L term in u s i n s t r a in 17
w ere: S s t l l 1 5 -1 6 , Hael l l 9 -1 0 , Smal 6 -7* T hese r e s t r i c t i o n
s i t e s a re lo c a t e d in th e j o i n t seq u en ce a f t e r n u c le o t id e s
7 1 1 , 719 and 722 r e s p e c t i v e l y , b e a r in g in mind t h a t in
t h i s c a s e th e l a b e l l e d s tr a n d i s com plem entary to t h a t
shown in P ig u re C 5 . l l . Por th e r e a so n d e s c r ib e d above th e
sm a lle r fra g m en ts m igh t p r o v id e a b e t t e r e s t im a te o f th e
n u c le o t id e s d e f in in g th e 1 term in u s (7 2 8 -7 2 9 ) . The HSV-1
a seq u en ce in p la sm id k l t h e r e fo r e has a s i z e o f 3 9 8 -4 0 0 bp
(betw een 3 2 9 -3 3 0 and 7 2 8 -7 2 9 ) .
The genom ic te r m in i o f HSV-1 s t r a i n USA- 8 were d e f in e d
by th e same c r i t e r i a (P ig u r e C 3 .1 2 ) and a r e shown in P ig u re
C3 . l l a t 3 3 0 -3 3 1 and 7 2 8 -7 2 9 w ith r e s p e c t to th e s t r a i n 17
se q u e n c e . Prom th e a u to ra d io g r a p h rep rod u ced in P ig u re

- 118 -03*12 i t co u ld o© se e n t h a t S te r m in a l fra g m en ts from s t r a in
USA—8 D1VJA were two n u c le o t id e s s m a lle r th an th o s e from s t r a in
17 DNA, w hereas B te r m in a l fra g m en ts were th e same s i z e i n
b o th s t r a i n s . The S and L te r m in i o f HSV-2 s t r a in HG52
were lo c a t e d a t 6 2 -6 4 and 309 -3 1 9 r e s p e c t i v e l y (P ig u r e C 3 .H )
by Smal c le a v a g e o f n ic k t r a n s la t e d HSV-2 BamHI u and v
i s o l a t e d from v ir io n DNA (T a b le C3*3 p a r t s B and C ). The
HSV-2 a seq u en ce th e r e fo r e has a s i z e o f 2 4 5 -2 5 7 bp.
In b oth HSV-1 s t r a in s and th e HSV-2 s t r a in th e genom ic
te r m in i a re lo c a te d c l o s e to th e r ig h t hand end o f d ir e c t
r e p e a t s in th e j o in t s e q u e n c e s . The d i r e c t r e p e a t in s tr a in -
USA- 8 c o m p r ise s th e f i r s t 17 n u c le o t id e s o f th e 21 n u c le o t id e
d i r e c t r e p e a t in s t r a i n 17 a t 3 1 1 -3 3 1 and 710- 7 3 0 .
However, th e prim ary seq u e n ces o f th e HSV-1 d i r e c t r e p e a t s
a re d i f f e r e n t from t h a t o f HSV-2.
The n u c le o t id e seq u en ce o f p la sm id s c o n ta in in g j o i n t
fra g m en ts o f HSV-1 3t r a in 17 o r HSV-2 s t r a i n HG52 w ith two
a se q u e n c es showed th a t th e a se q u e n c es a re tandem d i r e c t
r e p e a t s w ith a s in g l e copy o f th e a p p r o p r ia te 21 o r 17
n u c le o t id e d i r e c t r e p e a t a t th e a - a ju n c t io n (P ig u r e 03*15)*
Tandem r e i t e r a t i o n s in th e j o i n t r e g io n
The HSV-1 s t r a in 17* seq u en ce shown in P ig u re Q 3*H
was d eterm in ed u s in g p lasm id k l , but k2 was a l s o seq uenced
in th e r e g io n s where i t d i f f e r e d in s i z e from k l . Three
r e g io n s o f r e i t e r a t i o n were found in th e HSV-1 s t r a in 17
j o i n t r e g io n , one lo c a t e d in th e a seq u en ce and two in
a d ja c e n t c seq u e n c es ( r e i t e r a t i o n s I , I I and I I I i n P ig u r e s
C 3.9 and C 3 .H ) . The th r e e r e p e a te d u n i t s o f 1 2 , 16 and 17
n u c le o t id e s o f w hich th e r e i t e r a t i o n s c o n s i s t have d i f f e r e n t
prim ary s e q u e n c e s , a lth o u g h th e y a l l c o n ta in a predom inance

- 119 -
o f C r e s id u e s in th e same s tr a n d o f th e d u p le x . The seq u en ce
o f k2 in two o f th e r e i t e r a t i o n s ( I and I I I ) d i f f e r e d from
t h a t o f k l o n ly in th e number o f r e i t e r a t e d u n i t s .
R e i t e r a t io n I (4 0 8 -6 2 3 ) iu k l c o n ta in e d 18 c o p ie s o f
CCGCTCCTCCCC w hereas k2 c o n ta in e d 21 c o p ie s (P ig u r e C 3 .1 4 ) .
S im i la r ly , r e i t e r a t i o n I I I (1 1 6 7 -1 4 8 9 ) c o n s is t e d o f 19
c o p ie s o f CCGCCCCTCGCCCCCTC in k l and 11 c o p ie s in k2
(P ig u r e 0 3 * 1 5 ) . T hese two r e i t e r a t i o n s a re c o n ta in e d
w ith in Smal G and Smal A r e s p e c t i v e l y , and p r o v id e an
e x p la n a t io n fo r th e o b serv ed s i z e v a r i a b i l i t y in th e a and
c seq u e n c e s o f d i f f e r e n t p la q u e i s o l a t e s o f HSV-1 s t r a in 1 7 .
R e i t e r a t io n I I (9 2 8 -1 0 5 5 ) d id n o t c o n ta in a s im p le
r e p e a te d u n i t . F u rth erm ore, th e d i f f e r e n c e in seq u en ce
betw een k l and k2 was n o t r e f l e c t e d in d i f f e r e n t copy
numbers but in th e o r g a n is a t io n o f th e r e p e a te d u n i t s , th u s:
k l (CCCTCCCCAGCCCCAG)2 (CCCTCCCCGGCCCCAGJs
k2 (CCCTCCCCAGCOGCAG)! (CCCTCCCCGGCCCCAGjy.
The two 16 n u c le o t id e r e p e a te d u n i t s d i f f e r in a s in g l e
n u c le o t id e , and th e seq u e n c es o f r e i t e r a t i o n I I in k l and k2
d i f f e r by a s in g l e n u c le o t id e (P ig u r e C 3 .1 4 ) .
R e i t e r a t io n s I and *11 were a l s o i d e n t i f i e d in , th e
j o in t r e g io n o f liSV-1 s t r a in USA-8. R e i t e r a t io n I I
com p rised a t l e a s t 15 c o p ie s o f th e f i r s t 16 n u c le o t id e
v a r ia n t o f th e r e p e a te d u n i t o f s t r a in 17 (shown a b o v e ) .
R e it e r a t io n I i s r e la t e d to t h a t o f s t r a in 17 in a more
com plex way (P ig u r e 0 3 .1 4 ) . P ig u re C 3 . l l shows t h a t i t
c o n ta in s 8 c o p ie s o f a u n i t w hich c o n s i s t s o f th e f i r s t
11 n u c le o t id e s o f th e s t r a i n 17 12 n u c le o t id e r e p e a te d u n i t .
A d ja cen t to t h i s a re 6 c o p ie s o f a u n it w hich c o m p r ises th e

- 120 -
I I n u c le o t id e s w ith the seq u en ce TCTGTGGGTGGGG in s e r t e d
a f t e r th e th ir d n u c le o t id e . R e i t e r a t io n I I I was a l s o
i d e n t i f i e d in s t r a in USA-8, but th e seq u en ce was n o t
a n a ly se d in d e t a i l .
A p a r t i a l o v e r la p o f th e r e p e a te d u n i t o f r e i t e r a t i o n s
I , I I and I I I i s p r e s e n t im m ed ia te ly to th e r ig h t o f ea ch
r e i t e r a t i o n d e f in e d by th e b r a c k e ts in P ig u r e C 3 . l l . The
lo c a t io n o f ea ch p a r t i a l o v e r la p i s a r b it r a r y and depends
on th e d e f i n i t i o n o f ea ch r e p e a te d u n i t . The p a r t ia l
o v e r la p s c o n s i s t o f th e f i r s t 7 , H and 10 n u c le o t id e s o f
th e r e p e a te d u n i t s o f r e i t e r a t i o n s I , I I and I I I r e s p e c t i v e l y .
No tandem r e i t e r a t i o n s e q u iv a le n t to th o s e p r e s e n t in
th e HSV-1 a seq u en ce were found in th e th e HSV-2 seq u en ce
(P ig u r e C 3 . l l ) . I t i s n o t known w h eth er r e i t e r a t i o n s
e q u iv a le n t to I I and I I I e x i s t i n HSV-2 DNA.
Com parison o f n u c le o t id e seq u en ces
The r e g io n o f g r e a t e s t d i f f e r e n c e i n th e j o i n t s eq u e n c es
o f HSV-1 s t r a i n s 17 and USA-8 i s in th e c seq u en ce im m ed ia te ly
a d ja c e n t to th e a se q u e n c e . The d i f f e r e n c e s in t h i s r e g io n ,
w hich i s AC r ic h on th e s tra n d shown in P ig u re C 3 * l l ,
r e p r e s e n t both s u b s t i t u t io n o f n u c le o t id e s and d e l e t io n o r
in s e r t io n o f s h o r t s e q u e n c e s . Only one n u c le o t id e d i f f e r s
betw een th e 310 a n a ly se d n u c le o t id e s o f th e b se q u e n c e s to
th e l e f t o f th e f i r s t 17 n u c le o t id e r e p e a t a t 3 1 1 -3 2 7 , and
s t r a in s 17 and USA-8 a re i d e n t i c a l i n 166 com parable n u c le o t id e s
a t th e r ig h t hand end o f th e c seq u en ce shown (1 6 0 1 -1 7 6 6 ) .
R e it e r a t io n I i s th e r e g io n o f g r e a t e s t d i f f e r e n c e betw een
th e a se q u e n c e s o f th e two HSV-1 s t r a i n s , and th e r e a re
ten b ase s u b s t i t u t io n s and one d e le t e d n u c le o t id e in th e
rem ainder o f th e a se q u e n c e . S in c e r e i t e r a t i o n s I , I I and
I I I were i d e n t i f i e d in both s tr a in s * a lth o u g h th e s e d i f f e r

- 121 -
w id e ly in tim e and geography o f i s o l a t i o n , i t seem s l i k e l y
th a t th e y w i l l be a g e n e r a l f e a tu r e o f HSV-1 i s o l a t e s .
The a se q u e n c e s o f HSV-1 and HSV-2 b oth h ave a h ig h
GC c o n te n t (835* and 845* r e s p e c t i v e l y ) , but th e seq u e n c es f . A
show l i t t l e s i m i l a r i t y . Y /ith in th e a se q u e n c e s th e lo n g e s t
s im i la r s e q u e n c e , w hich i s a l s o h ig h ly asym m etric betw een
th e two s tr a n d s o f th e d u p lex , i s 3 6 2 -2 8 9 o f HSV-1 s t r a in
17 and 9 4 -1 2 3 o f HSV-2 s t r a in HG52. U n lik e HSV-1, HSV-2
d o es n o t p o s s e s s r e i t e r a t i o n I , but b oth s e r o ty p e s have a
d ir e c t r e p e a t a t th e b -a and a - c j u n c t io n s .
B x p r e ss io n o f th e HSV-1 j o i n t r e g io n
There i s a s y e t no e v id e n c e t h a t t r a n s c r ip t io n s t a r t s
in th e n u c le o t id e seq u en ce o f th e HSV-1 j o i n t r e g io n shown
in P ig u re 0 3 .1 1 , and no. seq u e n c es c o n s i s t e n t w ith th e
c o n sen su s "H ogness box" (Gannon e t a l • , 1 9 7 9 ) a re p r e s e n t .
The h e x a n u c le o t id e AAUAAA has been fou nd i n c l o s e p r o x im ity
to th e 3' te r m in i o f th e m a jo r ity o f p o ly a d e n y la te d
e u k a r y o t ic mRNAs y e t a n a ly se d (B e n o is t e t a l . » 1 9 8 0 ) . A
few e x c e p t io n s , w hich p o s s e s s a seq u en ce c l o s e l y r e la t e d
to AAUAAA, have been n o ted (MacDonald e t a l * , 1980;
Hobart e t a l . , 1980; G oeddel e t a l . , 1 9 8 1 ) . The DNA seq u en ce
AATAAA i s lo c a t e d a t two p o s i t i o n s in th e s t r a in 17r j o in t
r e g io n , a t 1 5 0 0 -1 5 0 5 and on th e o p p o s i t e s tr a n d a t 1 5 4 6 -1 5 4 1 .
B e n o is t e t a l . (1 9 8 0 ) have o b serv ed a c o n ser v e d -sequence/ T T ~ T C . ’ T C C
r e la t e d to TTTTCACTGC (more a c c u r a t e ly g$ t gaa *at$ )
lo c a t e d 3 1 to th e AATAAA c lo s e to th e p o ly a d e n y la t io n s i t e
o f s e v e r a l , but n o t a l l , mRNAs, and th e r e la t e d seq u en ce
TTTGCAGTAG i s p r e s e n t a t 1 5 2 0 -1 5 1 1 3 ’ to th e AATAAA a t
1 5 46-1541 ( c o n s id e r in g th e o th e r s tr a n d o f th e d u p lex from
th a t show n). No such seq u en ce i s p r e s e n t 3* to th e o th e r

- 122 -
AATAAA.
These r e s u l t s s u g g e s t th a t a mRNA i s t r a n s c r ib e d from
r ig h t t o l e f t and i s p o ly a d e n y la te d in th e r e g io n 1 5 3 0 -1 5 2 0 .
E xp erim en ts were c a r r ie d o u t in c o l la b o r a t io n w ith Dr F .J .
R ixon to a s c e r t a in w hether t h i s i s in d e e d th e c a s e . The
Tag I D fragm en t (F ig u r e s C 3.3 and C 3 .9 ) , c o n s i s t in g o f
n u c le o t id e s 1 4 9 8 -1 6 0 8 , was l a b e l l e d a t th e 3' t e r m in i by32in c o r p o r a t io n o f o<- P-dCTP u s in g th e Klenow fragm en t o f
E . c o l i DNA p o ly m e r a se . The two s tr a n d s were s e p a r a te d by
p o ly a c r y la m id e g e l e l e c t r o p h o r e s i s under d e n a tu r in g c o n d it io n s
(8 M u r e a ) . H S V - l- in fe c te d c e l l mRNA was produced under
im m ediate e a r ly c o n d it io n s (c y c lo h e x im id e b lo c k ; C lem ents
£ £ 1 9 7 7 ) , from w hich th e 4*7 kb mRNA was i s o l a t e d from
m eth y lm ercu rie h y d ro x id e a g a r o se g e l s (W atson e t a l . , 1 9 7 9 ) .
A liq u o ts o f e i t h e r t o t a l im m ediate e a r ly RNA or th e 4*7 kb
mRNA were h y b r id is e d s e p a r a te ly w ith th e two u n iq u e ly -
l a b e l l e d DNA s in g le s tr a n d s (5 7 ° in 3 x SSC and 9 0 #^ 4
form am ide f o r 5 h r) a c c o r d in g to th e method o f Berk and
Sharp (1 9 7 7 ) . The sam p les were t r e a te d w ith S I n u c le a s e
to d egrad e s in g le - s t r a n d e d n u c le ic a c id s , and th e n s u b je c te d
to p o ly a c r y la m id e g e l e l e c t r o p h o r e s i s under d e n a tu r in g
c o n d it io n s (8 M u r e a ) . Only one o f th e DNA s tr a n d s was
p r o te c te d by RIM A, so t h i s was compared w ith th e n u c le o t id e
seq u en ce o f th a t fragm en t on th e same g e l (P ig u r e C 3 # l6 ) .
T h is r e s u l t shows t h a t th e 3 1 te r m in a l coded n u c le o t id e o f
th e mRNA c o d in g f o r Vmw IE 175 i s lo c a t e d i n th e r e g io n
1 5 3 0 -1 5 2 5 . No t r a n s c r ip t s from th e o th e r s tr a n d te r m in a t in g
c l o s e to th e AATAAA a t 1 5 0 0 -1 5 0 5 were d e t e c t e d , e i t h e r i n
im m ediate e a r ly RNA o r in RNA i s o l a t e d from c e l l s h a r v e s te d
6 hr p o s t i n f e c t i o n a t 3 7 °•
I t i s n o t y e t known w hether j o in t se q u e n c e s o th e r

- 123 -
than th o s e c o d in g f o r Vmw IE 175 mRNA a re r e p r e s e n te d in
s t a b le t r a n s c r i p t s , o r where prim ary t r a n s c r ip t io n t e r m in a te s .
The r e g io n c o n t a in s s e v e r a l p o t e n t i a l s p l i c e donor (PuGTXXG)
and a c c e p to r (PyPyXPyAG) s ig n a l s ( S e i f e t a l . , 1 9 7 9 ) , but
th e s e ca n n o t be in t e r p r e t e d in th e a b se n c e o f t r a n s c r ip t
mapping d a ta . Three f e a t u r e s o f th e DNA seq u en ce make i t
l i k e l y t h a t a c o n s id e r a b le p a r t o f th e j o i n t r e g io n d o es n o t
code f o r p o ly p e p t id e , n o ta b ly th e g and a d ja o e n t c s e q u e n c e s .
F i r s t , t r a n s la t io n i s u n l ik e ly to o ccu r th rou gh r e i t e r a t i o n I
in th e a se q u e n c e . V a r ia t io n in th e copy number o f th e 11
n u c le o t id e r e p e a te d u n i t o f t h i s r e i t e r a t i o n in s t r a i n USA-8
co u ld r e s u l t in t r a n s l a t io n o f ea ch o f th e th r e e p o s s ib le
downstream r e a d in g fr a m e s . S im ila r argum ents a p p ly to th e
c seq u en ce a d ja c e n t to a , w hich c o n t a in s r e i t e r a t i o n s I I and
I I I . S econ d , i f t r a n s la t io n o f mRNA r e p r e s e n t in g r e i t e r a t i o n
I o f s t r a in 17 o c c u r r e d , th e p o ly p e p t id e would c o n ta in th e
u n u su a l s t r u c t u r e o f 18 tandem r e p e a t s o f 4 amino a c id s
(p la sm id k l ) . S im i la r ly , r e i t e r a t i o n I I I would code f o r a
p o ly p e p t id e c o n ta in in g 6 tandem r e p e a t s o f 17 amino a c i d s .
T hird , th e a b se n c e o f n u c le o t id e s from one s t r a in compared
w ith th e o th e r would g e n e r a te t r a n s l a t io n a l f r a m e s h i f t s .
S tr a in USA-8 h as one n u c le o t id e in s e r t e d betw een 328 and
329 o f s t r a in 1 7 , one n u c le o t id e d e le t e d a t 3 4 0 , and f i v e
n u c le o t id e s in s e r t e d betw een 740 and 741 w hich would produce
t h i s e f f e c t .

- 124 -
DISCUSSION
F e a tu r e s o f r e i t e r a t i o n s
D eterm in a tio n o f th e DuA seq u e n c es a t th e j o in t r e g io n
betw een th e 1 and 3 segm en ts o f th e genom es o f HSV-1 and HSV-2
h as r e v e a le d s e v e r a l f e a t u r e s . The p r e se n c e o f r e i t e r a t i o n s i n
HSV-1 p r o v id e s a read y e x p la n a t io n f o r th e w id e ly o b serv ed s i z e
v a r i a b i l i t y in th e a and c s e q u e n c e s . The HSV genome i s known
r e a d i ly to undergo reco m b in a tio n d u r in g r e p l i c a t i o n , and u n eq u a l
c r o s s o v e r e v e n ts oetw een tandem r e i t e r a t i o n s would g iv e r i s e to
s i z e v a r i a b i l i t y in th e s e r e g io n s . S im ila r tandem r e i t e r a t i o n s
m ight be r e s p o n s ib le f o r such v a r i a b i l i t y e lse w h e r e i n th e
genome. A f u n c t io n a l r o le f o r th e r e i t e r a t i o n s h a s n o t y e t
been i d e n t i f i e d , and i t i s p o s s ib le t h a t th e y encode no
e s s e n t i a l f u n c t io n s but r e p r e s e n t r e g io n s in w hich e r r o r s in
DNA r e p l i c a t i o n can a ccu m u la te . The a b se n c e o f r e i t e r a t i o n I
in th e a seq u en ce o f HSV-2 s t r a in HG52 a r g u e s a g a in s t an
e s s e n t i a l r o l e f o r th a t tandem r e i t e r a t i o n i n v ir u s
r e p l i c a t i o n . The 11 n u c le o t id e s CCCGCTCCTCC w hich form th e
f i r s t r e p e a te d u n i t o f r e i t e r a t i o n I in HSV-1 s t r a i n USA-8
( in t h i s c a s e r e d e f in in g th e u n it shown i n H g u r e C 3*H by
m oving th e b r a c k e ts one n u c le o t id e to th e l e f t ) a re a l s o
p r e s e n t a t 8 7 1 -8 8 1 (w ith r e s p e c t to th e s t r a i n 17 s e q u e n c e ) ,
show ing t h a t t h i s seq u en ce i s n o t a m p lif ie d o f n e c e s s i t y i n
th e HSV-1 genom e. The r e p e a te d u n i t s o f th e r e i t e r a t i o n s ,
a lth o u g h d i f f e r e n t , have s i m i l a r i t i e s such a s a p rep on d eran ce
o f C r e s id u e s on one s tra n d o f th e d u p lex and G r e s id u e s on
th e o th e r , and i t i s p o s s ib le t h a t such se q u e n c e s a r e more
r e a d i ly a m p lif ie d th an o t h e r s . T h is s im i l a r i t y ex te n d s a l s o
to r e i t e r a t i o n s IV and V w hich a re lo c a t e d e lse w h e r e i n th e
iiSV-1 genome, a s sum m arised in F ig u r e C 3 .1 8 . I t i s o f
in t e r e s t ' to n o t e t h a t th e seq u en ce CPyCCXCCC i s p r e s e n t in

- 125 -
r e i t e r a t i o n s I and I I I o f both HSV-1 s t r a i n s , and in
r e i t e r a t i o n IV; in d eed i t i s p r e s e n t on b oth s tr a n d s o f th e
24 n u c le o t id e r e p e a te d u n i t o f r e i t e r a t i o n I i n s t r a i n USA-8 .
GGCCTCCC i s p r e s e n t in r e i t e r a t i o n I I and CCCCACTC (CPyCCXCTC)
i n r e i t e r a t i o n V. The SV40 genome p o s s e s s e s s i x c o p ie s o f
PyPyCCGCCC c l o s e to th e o r ig in o f DNA r e p l i c a t i o n (Dhar e t
a l . , 1 9 7 7 ) , two o f w hich a re w ith in T -a n t ig e n b in d in g s i t e
I I I ( T j ia n , 1 9 7 8 ) . However, th e p r e se n c e o f t h i s b in d in g s i t e
i s n o t e s s e n t i a l f o r SV40 DNA r e p l i c a t io n (Subram anian and
Shenk, 1 9 7 8 ) . T -a n t ig e n b in d in g s i t e I I , . th e p r e se n c e o f w hich
i s r e q u ir e d f o r DNA r e p l i c a t io n (Shenk , 1 9 78; Gluzman e t a l . ,
1 9 8 0 ) , p o s s e s s e s two p a lin d ro m es o f GCCTC (T j ia n , 1 9 7 8 ) , and
r e i t e r a t i o n s I -I V c o n ta in PyCCTC and r e i t e r a t i o n V c o n ta in s
CACTC. Such se q u e n c e s , • how ever, a re s u f f i c i e n t l y s h o r t to be
p r e s e n t commonly i n HSV-1 DNA ( e . g . PyPyCCGCCC a t 1 6 8 8 -1 6 9 5
and GCCTC a t 1 6 8 6 -1 6 9 0 ) , and so i t rem ain s to be shown t h a t
th e se q u e n c e s o f th e r e p e a te d u n i t s a re s p e c i f i c f o r
p r e f e r e n t i a l a m p l i f i c a t io n .
C oding p o t e n t i a l a t th e 3* end o f Vmw IE 175 mRNA
The g en es co d in g f o r dPyK (M cK night, 1 9 8 0 ) , Vmw IE 175
( t h i s s tu d y ; M-J. M urchie, p e r s o n a l co m m u n ica tio n ), Vmw IE 12
(M -J. M urchie and F .J . R ixon , p e r s o n a l com m u n ication s) and
Vffiw 1 3 6 ’ (1 4 3 ) (J . M cLauchlan, p e r s o n a l com m unication) have
5 ! and 3 ’ seq u e n c es c o n s i s t e n t w ith co n serv ed f e a t u r e s o f
e q u iv a le n t r e g io n s o f o th e r e u k a r y o t ic g en es c o d in g f o r
p o ly a d e n y la te d mRNAs. The o n ly in t r o n s y e t d e s c r ib e d f o r HSV-1
r e s id e in TR5/I R 3 in th e g en es co d in g f o r Vmw IE 12 and 68
(Y/atson e t a l . , 1981; P .J . R ixon and M-J. M urchie, p e r s o n a l
0 0JU:i»Ui xca ijiuhs) .
The seq uence a t th e 3 1 term in u s o f th e Vmvv IE 175 mRNA
(P ig u re C 3 . l l ) c o n ta in s two p o s s ib le te r m in a t io n cod on s in

- 126 -
d i f f e r e n t fra m es, a t p o s i t i o n s c o r r e sp o n d in g to 156 2 -1 5 6 0
(UGA) and 1 5 9 1 -1 5 8 9 (UAA). The h y p o t h e t ic a l amino a c id
seq u en ce o f th e carb oxy term in u s o f Vmw IE 175 u t i l i s i n g each
o f t h e s e te r m in a t io n codons i s g iv e n i n T ab le C% 6. A lso
shown i s th e o c cu r r en ce in each c a s e o f th e fo u r amino a c id s
coded by t r i p l e t s c o n ta in in g c o m b in a tio n s o f G and C m o ie t ie s
o n ly , and th e d i s t r i b u t i o n o f th e fo u r n u c le o t id e s in th e
th ir d codon p o s i t i o n . Two o b s e r v a t io n s from t h e s e d a ta
s u g g e s t t h a t , i f e i t h e r te r m in a t io n codon i s u se d , th e UAA
fram e i s th e more l i k e l y . F i r s t , t h i s fram e c o n ta in s f a r
few er a r g in in e r e s id u e s in t h i s r e g io n and t h e r e fo r e m ight be
th ou gh t to code f o r a more r e a s o n a b le p r o t e in s t r u c t u r e .
S econd , th e g r e a t m a jo r ity o f cod on s i n t h i s fram e have G or
C r e s id u e s in th e th ir d p o s i t i o n . T h is i s c o n s i s t e n t w ith
th e e x p e c t a t io n th a t t h i s r e g io n o f th e DNA would p o s s e s s a
h ig h GC c o n te n t e s p e c i a l l y i n th e t h ir d codon p o s i t i o n , s in c e
i t i s under s e l e c t i v e p r e s s u r e s to m a in ta in b oth a h ig h GC
c o n te n t ( f o r unknown r e a s o n s ) and a f u n c t io n a l Vmw IE 175 g e n e .
F u l l e lu c id a t io n o f th e c o d in g r e g io n o f t h i s gene a w a it s
co m p le te seq u en ce d e te r m in a t io n .
R ole o f th e a seq u en ce in c o d in g f o r p o ly p e p t id e
I t seem s l i k e l y t h a t th e HSV-1 a seq u en ce d o es n o t code
f o r p o ly p e p t id e . P r e v io u s ly , th e l e s i o n in an HSV-1 t s
mutant ( t s c 7 5 ) had been i n c o r r e c t ly mapped i n th e a seq u e n c e ,
and ex a m in a tio n o f th e p o ly p e p t id e s in d u ced by t h i s m utant a t
th e n o n -p e r u .is s iv e tem p era tu re prompted th e c o n c lu s io n t h a t th e
a seq u en ce form s p a r t o f th e Vmw IE 175 gen e (K nipe e t a l . , 1 9 7 9 ) ,
T h is s tu d y shows t h a t th e a seq u en ce i s n o t in c lu d e d i n th e
g en e . P r e s to n ( in p r e s s ) h as shown t h a t th e m u ta tio n maps
in th e c seq u en ce some 2 kbp away from th e a se q u e n c e . T hese
f in d in g s have r e p e r c u s s io n s d e tr im e n ta l to th e h y p o th e s is t h a t

- 127 -
a l l a s e q u e n c e s w ith in a genome a r e o b l i g a t o r i l y i d e n t i c a l ,
a s d is c u s s e d i n S e c t io n 4 .
Homology betw een th e j o i n t r eg io n s o f th e HSV-1 and HSV-2 genom es
H y b r id is a t io n ex p er im en ts (F ig u r e C 3 .7 ) d em on stra ted
hom ology betw een HSV-1 BamHI k and th e c seq u en ce o f HSV-2
BamHI g , and betw een HSV-2 BamHI g and th e c seq u en ce o f HSV-1
BamHI k . These r e g io n s co rresp o n d i n l o c a t io n to th e 3 !
p o r t io n s o f th e g e n e s c o d in g f o r HSV-1 V IE 175 andmwHSV-2 Vffiw IE 1 8 2 . A more l o c a l i s e d r e g io n o f hom ology i n th e
b seq u e n c es (HSV-1 Smal H and HSV-2 Smal 4 ) was a ls o
d e t e c t e d . I t was n o t shown th a t hom ology e x i s t s betw een th e s e
two fr a g m e n ts , a lth o u g h t h i s i s th e s im p le s t e x p la n a t io n o f
th e r e s u l t s . HSV-1 Smal H i s a d ja c e n t to se q u e n c e s c o d in g f o r
th e 5 1 term in u s o f Vmw IE 110 mRNA (Mackem and Roizm an, 1 9 8 0 ) ,
and t h e r e f o r e i t i s p o s s ib l e t h a t HSV-1 and HSV-2 sh are
hom ology in t r a n s c r ip t io n a l c o n t r o l seq u e n c e s a t th e 5 1
te r m in i o f th e g en es co d in g f o r Vmw IE 110 (HSV-1) and
\ w IE 118 (H SV -2). Fragm ents c o n ta in in g th e a seq u en ce d id
n o t h y b r id is e w e l l even to hom ologous p r o b e , and i t was
th e r e fo r e n o t p o s s ib le to e s t im a te th e e x t e n t o f hom ology
betw een th e HSV-1 and IISY-2 a se q u e n c e s .
R ole o f th e a seq u en ce in m a tu ra tio n o f genome le n g t h DNA
In both iISV-1 and HSV-2 th e S term in u s i s e q u iv a le n t
to a p o s i t i o n c lo s e to th e 'r ig h t band end o f th e 17 and 21
n u c le o t id e d i r e c t r e p e a t a t th e b -a j u n c t io n , and th e L
term in u s i s e q u iv a le n t to th e r ig h t hand end o f th e o th e r
d i r e c t r e p e a t a t th e a -c ju n c t io n . A lth ough th e lo c a t io n s o f
th e te r m in i w ith r e s p e c t to th e j o in t seq u en ce have n o t been
d e f in e d to s in g le n u c le o t id e s , F ig u r e 03*11 shows t h a t th e
HSV-1 s t r a in USA-8 S term in u s i s one n u c le o t id e to th e r ig h t
o f t h a t i n ..SV -l s t r a in 1 7 , w hereas th e L te r m in i c o in c id e .

- 128 -
A m in o r ity o f v i r io n UNA m o le c u le s c o n ta in more than one copy o f
th e a seq u en ce a t th e 1 term in u s and a t th e j o i n t . D e te r m in a tio n
o f th e n u c le o t id e seq u e n c es betw een two tandem IISV-l o r HSV-2
a seq u e n c e s h as shown th a t th e a se q u e n c es a r e arran ged a s
h e a d - t o - t a i l d im ers w hich have one copy o f th e d i r e c t r e p e a t
a t t h e i r p o in t o f f u s io n (P ig u r e 0 3 .1 7 )* T h is arrangem ent
c o u ld a r i s e e i t h e r by m is a lig n e d g e n e t i c re co m b in a tio n a t th e
d i r e c t r e p e a t s , o r by d i r e c t l i g a t i o n o f th e L and S te r m in i*
The f a c t t h a t m u lt ip le a se q u e n c es have n o t been o b se rv ed a t
th e S term in u s o f HSV-1 s t r a i n 17 and HSV-2 s t r a in HG52
presum ably r e f l e c t s th e m echanism o f DNA r e p l i c a t i o n and
m a tu r a tio n . I t i s p o s s ib le t h a t th e d i r e c t r e p e a t s s e r v e a s
r e c o g n i t io n s i t e s f o r th e m a tu ra tio n o f u n i t l e n g th DNA
from co n ca tera ers . S in c e th e HSV genom ic t e r m in i a r e ’
s p e c i f i c a l l y lo c a t e d , i t i s l i k e l y t h a t c o n c a te m e r ic DNA i s
s p e c i f i c a l l y c le a v e d a t a l t e r n a t e j o i n t r e g io n s to produce
DNA o f genome l e n g t h , b u t th e r e i s a s y e t i n s u f f i c i e n t e v id e n c e
upon w h ich to b ase d e t a i l e d m o d e ls . N e v e r t h e le s s , th e
co n seq u en ce o f th e m a tu ra tio n m echanism i s t h a t th e L term in u s
e s s e n t i a l l y c o n ta in s b oth c o p ie s o f th e d i r e c t r e p e a t , w h ereas
th e S term in u s c o n t a in s o n ly th e a -c copy (P ig u r e C 3 .1 7 ) .
R ole o f th e a seq u en ce i n segm ent in v e r s io n
D ata w hich s u g g e s t t h a t th e a seq u en ce h a s a r o l e i n
segm ent in v e r s io n by a s i t e - s p e c i f i c m echanism a re d is c u s s e d i n
S e c t io n 4 . P r e c i s e ly w hich se q u e n c es a r e in v o lv e d in th e
p rop osed s i t e - s p e c i f i c reco m b in a tio n e v e n ts i s unknown. One
p o s s i b i l i t y i s t h a t th e r e g io n w hich i s co n serv ed betw een th e
HSV-1 and HSV-2 a seq u en ces (3 6 2 -3 8 9 in P ig u r e C3*HA and
9 4 -1 2 3 -in P ig u re C 3.11B) i s a reco m b in a to r seq u en ce a n a lo g o u s
to c h i in p r o k a r y o te s (Sm ith e t a l . , 1980) and co g in fu n g i
(C a tc h e s id e and C orcoran , 1 9 7 3 ) , p rom otin g h ig h l o c a l r a t e s o f9

- 129 -
recom bination, or that i t i s equivalent to a t t in lambda
(Dandy and Ross, 1977)> promoting recombination in a sp e c if ic sequence. These hypotheses could be te s te d by tran sferrin g
th is sequence to any other rea d ily s t u d i e d genome. Purther
aspeots o f the ro le o f the a sequence in DNA r e p lic a t io n are
discussed in Section 4-

Table 03*1
S iz e s (bp) o f r e s t r i c t i o n fra g m en ts o f HSV-1 BamH I k
and 11SV-2 BamH I g . These fra g m en ts c o n ta in one a
seq u e n ce and , s in c e th e r e a r e r e g io n s o f s i z e
v a r i a b i l i t y i n HSV-1 BamH I k , s i z e s r e f e r
s p e c i f i c a l l y to th e recom b in an t p la sm id k l 3hown
in P ig u re 0 3 * 6 , T o ta l s i z e s o f th e fra g m en ts
a re g iv e n , and r e s t r i c t i o n maps a r e shown in
P ig u re 0 3 .3

Table C 3.1
IiSV-1 BamH I k
S g t I Sma I
A 3500 A 1100
B 1600 B 840
C 510 C 700
G 330 D. 560
£ 560
F ' 410
G 340
H 350
I 275
i 240
K 150
L 143
Ivl 91
N 86
0 48
P 43
Q 38
R 24
5940 6015"
HSV-2 Bamii I £
s s i x Sma I
1 2020 1 2650
■2 1310 2 1050
3 960 3 540
4 740 4 510
5 390 5 440
6 290 6 300
7 160 7 112
8 130 8 87
9 76
10 67
11 63
12 14
6000 5909

Table 03.2
D ata f o r th e r e s t r i c t i o n mapping o f HSV-1 BamHI k .
M ost e x p e r im e n ts u sed t h i s fragm en t i s o l a t e d from
p la sm id k l (F ig u r e 0 3 . 6 ) , e x c e p t in p a r t s B, G, F
and G, when th e fra g m en ts were i s o l a t e d from v ir io n
DBA. N ick t r a n s la t e d DNA was u sed e x c e p t i n p a r t H,
when a fragm en t ^2P - la b e l l e d a t th e 5 f te r m in i was
u s e d . Fragm ents i n p a r e n th e s e s were n o t r e s o lv e d ,
and so in d ic a t e p r e d ic t e d r a th e r th an a c t u a l s i z e s .
E le c t r o p h o r e s is was c a r r ie d o u t on 2 # a g a r o s e ,
7 .5 ,‘* p o ly a c r y la m id e o r 15/^ p o ly a c r y la m id e g e ls *- *
A garose g e l s c o n s i s t e n t l y gave la r g e r v a lu e s f o r
s i z e s o f fr a g m en ts g r e a t e r th a n 500 bp than
p o ly a c r y la m id e g e l s . T h ere fo re s i z e s above 500 bp
a re q u oted from a g a r o se g e l s , and below 500 bp from
p o ly a c r y la m id e g e l s . T he.deduced r e s t r i c t i o n maps
a re shown i n F ig u re C3*3*

Table 05.2
A. Fragment s i z e s (bp) o f HSV-1 BamH I k .
S s t I 3500 1600 510 330
Sau3A I 3300 1600 900 170
Tag. I 3700 1270 900 110 26
H inc I I 4600 1400
Sma I 1100 840 700 560 560 410 340 350 275240 150 143 91 86 48 43 38 24
B. Fragment s i z e s (bp) o f HSV-1 BamH I q .
S s t I 2700 510 330
Sma I 1100 840 560 410 340 91 48 3b 37 24
C. Fragment s i z e s (bp) o f HSV-l BamH I s ,
S s t I 1600 1300
Sma I 700 560 350 340 275 240 150 143 86 43
D. Fragment s i z e s (bp) o f d ou b le d i g e s t s o f HSV-1 BamH I k .
Sma l/S s_ t 1 1100 760 700 560 410 35Q 340 275260 250 240 150 143 91 86 78 70(48 43 38 2 4 )
Sma 1 / Sau3A I 700 660 560 560 510 420 410 350 340275 240 170 150 150 143 91 86(48 43 38 2 4 )
Sma I/Tag. I 820 680 700 560 560 410 350 340 275240 150 150 143 140 110 91 86 4338 26 24 22
Sma i / l l i n c I I 1100 700 680 560 560 410 350 340275 240 160 150 143 91 86 48 4338 24 V
S s t l/S a u 3 A I 1700 1600 1600 510 330 170 70
S s t I/Tag. I ( 200u 1600 / 1270) 510 300 110 70 26
S s t I /H in c I I 2900 1600 6O0 510 330
Sau3A I /T a q 1 (3300 1100 900) 400 170 1J0 26

Table 03.2
£ . fra g m en t s i z e s (bp) o f d ig e s t s o f fra g m en ts o f HSV-1 BamH I k .
fragm en t d ig e s t e d w ith p ro d u cts
S s t I A Sma I 1100 720 410 350 150 80 43 24
S s t I B Sma I 700 560 240 143
S s t I C Sma I 260 91 78 38
S s t I £ * Sma I 250 48
Sau3A I A Sma I 700 560 430 350 240 150 86 43
Sau3A I B Sma I 640 495 410 24
Sau3A I C Sma I 560 150 91 48 38
S s t I A Sau3A I (1650 8 4 0 ) 560 75
Sst; I C Sau3A I 510
S s t I A Tag 1 (2000 8 4 0 ) HO 70
S s t I C Tag. I 510
Sau3A I A Tag I (3 3 0 0 )
Sau3A 1 B Taa I (1 1 0 0 ) 350 110
P. Pragiaent s i z e s (bp) o f d i g e s t s o f fra g m en ts o f 1I3V-1 BamLi I cl# -
f ragm ent
S s t I A
S s t I C
S s t I 1)
d ig e s t e d , w ith
Sma I
Sma I
Sma I
p r o d u c ts
1100 '720 410 340 40 24
260 91 78 38
250 48

Table 03.2
G. Ifragment s i z e s (bp) o f d i g e s t s o f fragments? o f HSV-1 Bamil I s . .
fragm en t dig e s t e d w ith produc t s
Sma I 700 560 240 143
Sma I 350 340 275 150 80 43
H. Sma I p a r t i a l d ig e s t io n p r o d u c ts (bp) from ea ch o f th e two h a lv e s o f HSv-1 BamH I k Sau3A I k , w hich had been 5 ' - l a b e l e d a t th e Sau3A I sltes*~and tEe two h a l v e s ’ s e p a r a te d by d ig e s t io n w ith S s t I , and t h e ir in t e r p r e t a t io n a s Sma I fr a g m e n ts .
l e f t hand fragm en t o f 5 a u 3 r r r ---------------
240 (J)
950 (J-C)
1100 (J-C -L )
1650 (J -C -L -h )
I . Sma I p a r t i a l d ig e s t io n p r o d u c ts (bp) o f HSV-1 .BamH I k Sau3A I B w hich would be p r e d ic te d from th e tw o ~ p o s s ib le a rra n g em en ts .
f ragm ent o rd er pr e d ic t e d p a r t i a l d ig e s t io n p ro d u cts
640-K -JM 95 1 H 5 970 945 665 475
640-F -R -495 1115 i09Q 970 520 475
r ig h t hand fragm en t o f'S e f i S r T T . -------------
430 (p a r t o f A)
770 (4 3 0 -£ )
820 (430-G -P )
1 0 0 0 * (430-G -P -K )
1290 ( 430-G -P-K -X )
1630 ( 430-G -P -iC -I-H )
1700 (430—G -P -K -I-H -p a rt o f N)
S s t I £
S s t X X

Table 03.2
J . Sma I p a r t i a l d i g e s t i o n p r o d u c ts (bp) o f HSV-1 BamH I k S s t I C w hich would be p r e d ic te d , from tEe two“ p o s s i b l e a rra n g em en ts .
fragm en t o r d e r p r e d ic t e d p a r t ia l d i g e s t i o n p ro d u cts
78—M—(J—260 389 298 207 169 129
7 8-£}-M-2 60 389 351 207 129 116

Table C3.3
B ata f o r th e r e s t r i c t i o n mapping o f fiSV-2 Bamiil g .
M ost e x p er im e n ts u sed t h i s fragm en t i s o l a t e d from
p la sm id g l (F ig u r e 0 3 . 6 ) , e x c e p t in p a r t s 3 , C, 3,
G and H, when th e fra g m en ts were i s o l a t e d from v ir io n
BNA. The p la sm id fragm en t c o n ta in s two tandem
r e p e a te d a seq u e n c e s in s t e a d o f th e s in g l e a seq u en ce
p r e s e n t in th e m ajor j o i n t fragm en t from v i r i o n BNA.
T h is e x t r a 250 bp r e s u l t s , f o r exam p le , in two
a d d i t io n a l fra g m en ts in . th e Smal d i g e s t w hich a re
n o t produced from th e norm al j o i n t (230 and 14 b p ).
T h is was ta k en i n t o a cco u n t in d ed u cin g th e
r e s t r i c t i o n maps shown in F ig u re C 3 .3 .
N ick t r a n s la t e d BNA was u se d , and fra g m en ts in
p a r e n th e se s in d ic a t e p r e d ic te d r a th e r than a c t u a l
s i z e s s in c e th e y were n o t r e s o lv e d on th e g e l u s e d .
E le c t r o p h o r e s is was c a r r ie d o u t on 25* a g a r o s e ,
7 .5 /* p o ly a c r y la m id e o r 155* p o ly a cr y la m id e g e l s .-
A garose g e l s c o n s i s t e n t l y gave la r g e r v a lu e s than
p o ly a c r y la m id e g e l s f o r s i z e s o f fra g m en ts g r e a te r
than 500 bp. T h ere fo re s i z e s above 500 bp are
q u oted from a g a r o se g e l s , and below 500 bp from
p o ly a c r y la m id e g e l s .

Table C3«3
A, Fragm ent s i z e s (bp,) o f HSV—2 BamH I w hich c o n ta in s two tandem ly r e p e a te d a s e q u e n c e s .
S s t I 2020 1550 960 740 390 290 160 130
Sau3A I 4100 1250 900
Tag I 1800 1600 1050 770 660 85 25
H inc I I 2600 1800 1300 250
Kffi I 4700 1500
Sma I 2650 1050 540 510 440 500 230 87 76 67 63 14 14
112
B, Fragment s i z e s (bp) o f HSV-2 BamH I u .
S s t I . 2020 390 390 290 130
Sma I 2650 440 87 22 14
0 . Fragment s i z e s (bp) o f HSV-2 BamH I v .
S s t I 1550 1200 960 160
Sma I 1050 540 510 300 215 112 76 67 6;
B. Fragment s i z e s (bp) o f d o u b le d i g e s t s o f HSV-2 BamH I g .
. •'* T'U■ WSma I / S s t I 2020 1000 540 460 390 350 . 290 230 ----- 210 112 87 87 76 67 63 62 40 23
Sma I/S au 3A I 1600 1050 900 540 460 440 300 230112 87 76 67 63 53
—Sma I/T ag. I 510 480 440 290 250 230 112 85 7667 63. 57 44 25
Sma I /H in c I I 1400 1100 1050 540 510 440 300 230112 87 76 67 63
Sma I/Krm I 1400 1100 1050 540 510 440 300 230112 87 76 67 63

Table C3.3
B. fragm en t s i z e s (bp) o f d o u b le d i g e s t s o f HSV-2 BamH I g
( c o n t in u e d ) .
S s t 1 / Sau3A 1 1600 1550 740 600 450 390 290 200160 130
— S s t i/T a a I 560 390 280 160 130 85 48 41 25
S s t I / H inc I I 1300 1000 960 740 460 390 360 290 250160 130
S s t I/K on I 1550 1050 960 750 740 390 290 160130— Sau3A I /H in c I I 290 250
— Sau3A I/K pn I 500
— Tac I /H in o I I 540 460 400 250 85 25
— Tag I/K pn I 85 25
- - H inc II/K pn I 2600 1800 1300 250 250
ita g m e n ts s m a lle r than 20 bp were n o t r e s o lv e d . For th o se d i g e s t s marked — fra g m en ts l a r g e r than 6Q0 bp were n o t r e s o lv e d .
IS. 'Fragment s i z e s (bp) o f d i g e s t s o f S s t I fra g m en ts o f HSV-2 BamH I g . .
fragm en t d ig e s t e d w ith p r o d u c ts
1 Sma I 2020
2 Sma I 1000 350 230
1 Sma I 460 290 112
1 Sma I 540 63
£ Sma I 390
6 Sma I 210 87
1 Sma I 76 62 23
8 Sma I 8 7 40

Table 05*3
E. fragm en t s i z e s (Jap) o f d i g e s t s o f S s t I fra g m en ts o f HSV-2 BamH I~ g ( c o n t in u e d ) . “
fragm en t d ig e s t e d w itb p r o d u c ts
1 Tag I (1050 1000)
2 Tag I (8 7 0 ) 530
1 Tdq I (9 2 0 ) 39
i Tag. I VJI o 00 VJI
Tag. I 390
6 Tag I 260 25
I Tag I 160
8 Tag I 130
Bo fragm en t s i z e s (bp) o f Sma I d i g e s t s o f S s t I fra g m en ts o f IISV-2 BamH - r B. —
S a t I 1 2020
S s t I 390 350 25 14
S s t I 6 210 8'7
S s t 1 8 87 40
0 . i'ragment s iz e s - (bp) o f Sma I d i g e s t s o f S s t I fra g m en ts o f HSV-2 BamH I v . ! -
S s t I X 1000 220 67 14
S s t I 2 460 290 112
S s t X 2 540 63
S s t I I 76 62 23

Table C?.3
H. fragm en t s i z e s (bp) o f Sma I p a r t i a l d i g e s t i o n o f S s t I fra g m e n ts , o f HSV-?"lkmH I g .
S s t I 1050 365 80
S s t I i 570 410
S s t I i 600 67

Table U3.4
i)a ta f o r th e r e s t r i c t i o n mapping o f th e 1760 bp
H in f I fragm en t w hich c o n ta in s th e j o i n t betw een HSV-1
1 and S . A lso u sed was th e Sau3A I /H in f I 1130 bp
fragm en t w hich sp a n s tn e j o i n t , and co m p r ise s th e
l e f t hand end o f th e 1760 bp fr a g m e n t•
K ick t r a n s la t e d DNA was p rep ared from recom b in an t
p lasm id k l (F ig u r e 0 3 . 6 ) , E le c t r o p h o r e s is was
c a r r ie d o u t on 2 y* a g a r o se o r 7*57* p o ly a c r y la m id e g e ls>« 4 s
and fragm en t s i z e s g r e a te r than 500 bp a re q uoted
from a g a r o se g e l s and l e s s than 500 bp from
p o ly a c r y la m id e g e l s . The deduced r e s t r i c t i o n maps
a re shown in F ig u re 0 3 .9 .

Table C?«4
A. fragm en t s i z e s (bp) o f th e HSV-1 JBamH I k Sau3A I /H in f I 1130 bp fra g m en t, w hich c o n t a in s tEe a se q u e n c e ,
Sma I 410 340 160 150 43
Ava I I 680 m
BstN I 1000 130
Ava I 360 340 160 150 50 43
Hae I I 980 150
B gl I 930 170 25
S s t I I 420 320 190 85 80 18
Tag. I 1130
U n d er lin ed fra g m en ts a r e th o se w hich were l a b e l l e d when th e 1130 bp fragm en t was p rep ared from 5* - l a b e l le d HSV-1 BamH I k Sau3A I 'A. ,
B. Pragment s i z e s (bp) o f th e HSV-1 BamH I k H in f I 1760 bp fra g m en t, w hich c o n t a in s th e a seq u e n c e .
Sma I 1040 340 160 150 43
Ava I I 1080 680
BstN I 1500 130 130
Ava I 680 350 340 160 150 43
Hae I I 1000 430 150 12 5 70
B £l I 1460 170 70 26 25
S s t I I 950 320 190 95 85 80 18
Tag I 1500 150 110
U n d erlin ed fra g m en ts were d e f i n i t e l y n o t p r e s e n t in th e H in f I d i g e s t o f n ic k t r a n s la t e d HSV-1 BamH I k .

C. Fragm ent s i z e s (bp) o f Bau4A I d o u b le d i g e s t s o f th e HSV-1 BamH I k H in f I 1760 bp fra g m en t.
Sau3A l / Sma I 630 410 340 160 150 43
Sau3A I / Ava I I 680 650 450
Sau3A I/BstIM I 1000 500 130 130
Sau3A I / Ava I 630 350 340 160 150 50 43
Sau3A I/H a e I I 980 430 150 125 70
Sau3A I / B gl I 930 530 170 70 26 25
Sau3A I / S s t I I 570 420 320 190 95 05 80 18
Sau3A I/T a g I 1130 370 150 110
D. Fragm ent s i z e s (b p) o f d o u b le d i g e s t s o f th e HSV-1BamH I k Sau3A I / H in f I 1130 bp fra g m en t.
Sma I / Ava I I
Sma I /B stN I
Sma I /H a e I I
Sma I /B g l I
Sma I / S s t I I
Ava I I /B s tN I
Ava I I /H a e I I
Ava I l / B g l I
Ava I I / S s t I I
BstN I /B g l I
BstN I / S s t I I
B g l I / Hae I I
B gl I / S s t I I
S s t I I / Ava I
S s t I I / f la e I I
B g l 1 / Ava I
410 310 160 150 45 43
410 340 150 130 43 32
410 340 150 150 43
410 340 160 125 43 27
260 130 65 80 58 - H CO
550 450 130
530 450 150
530 400 1'70 25
420 320 190 65 41 32 18
930 130 35 25
420 320 170 65 80 27 18
930 150 25 13
420 320 120 65 80 62 18
350 250 135 65 80 61 48
420 320 140 65 80 50 18
350 340 160 125 50 43 27

T a b l e 0 5 « 4
ii. ij'Tagment s i z e s (bp) o f d ou b le d i g e s t s o f HSV-1 BamH I k H in f I 1760 bp fra g m en t.
1 * 1 I /H ae I I 980 350 150 125 62 38 26
Tag. I /B g l I 1300 170 110 70 53 26 25
Tag I / S s t I I 800 320 190 110 95 85 60 50 18
m l I /H ae I I 930 430 150 70 70 29 26 25

Table 03.5
Bata f o r th e r e s t r i c t i o n mapping o f the 1310 bp HSV-2
S s t I fra g m en t w hich c o n ta in s bhe j o i n t betw een
L and S (HSV-2 BamH I & S s t I 2 in F ig u re 0 3 . 3 ) .
A lso u sed was th e Sma I 900 bp fragm en t w hich does'
n o t span th e j o i n t , b u t o v e r la p s a lm o st e n t i r e l y th e
1310 bp fragm en t a t i t s l e f t hand end (HSV-2 BamH I g
Sma I 2 in F ig u re C3o3J* N ick t r a n s la t e d BNA was
p rep ared from recom b in an t p la sm id g l (F ig u r e 0 3 ..6 ) '■
w hich c o n t a in s two tandem r e p e a te d a se q u e n c e s
in s t e a d o f th e one a seq u en ce p r e s e n t in th e m ajor
j o in t fra g m en t o f v i r i o n BNA, and so th e S s t I
fragm en t i s 1550 bp in t h i s c a s e ( s e e T ab le 0 3 . 3 ) .
T h is f a c t has been tak en in t o a cco u n t in d ed u c in g th e
r e s t r i c t i o n maps shown i n F ig u re 0 3 .9 .
E le c t r o p h o r e s is was c a r r ie d o u t on 2 °/o a g a r o se or 7 * 5 $
p o ly a c r y la m id e g e l s , and fragm en t s i z e s g r e a t e r4
than 500 bp a r e q u oted from a g a r o se g e l s and l e s s
than 500 bp from p o ly a c r y la m id e g e l s . The deduced
r e s t r i c t i o n mai)s a re shown in F ig u r e 03«9

Table 05.5
A. Fragment s i z e s (bp) o f HSV-2 BamH I g S s t I 2 , w hich c o n ta in s two tandem ly r e p e a te d a s e q u e n c e s .
Dde I 1200 360
A lu I 1440 125
H inc I I 960 550 250
Sma I 680 360 230 67 14 14
Ava I 500 360 280 230 130 67 14 14
Ava I I 830 550 100 70 27
BstN I 1000 250 170 70 29 15
S s t I I 480 540 310 210 200 28 28 14 14
H in f I 600 360 300 220 29 29 26
B. fragm en t s i z e s (bp) < HSV-2 BamH I g S s t I
o f Sma I 2 .
d ou b le d i g e s t s o f
Dde I 500 360 360 230 67
A lu I 880 240 230 125 67
Hinc II 880 360 230 67
BstN I 360 330 250 230 170 70 67 29
S s t I I 540 310 290 200 110 110 78 67 28
H in f I 600 360. 250 210 44 26 25 25
Fragm ents sm a lle r than 20 bp were n o t r e s o lv e d .

B ata c o n c e r n in g th e p o s s ib le amino a c id seq u e n c es
a t th e carb o x y term in u s o f Vmw IE 1 7 5 .

Table C5.6
A. COOING POTENTIAL OP V TEEMINUS OP Vmw IE 175 mKNA- - — r ^ — — ~ , - . . . - _ , ^
UGA s to p codon
v a l - s e r - l e u - g l y - l e u - a l a - h i s - g l y - p r o - g l n - a r g - h i s - a r g - a l a - g l y -
g l y - g l y - g l y - g l y - a r g - g l y - g l y - a l a - g l y - g l y - g l y - g l y - g l y - l e u - g l y -
h i s - a l a - p r o - a l a - a l a - g l y - s e r - c y s - g l y - l e u - g l y - a r g - a r g - l e u - g l y -
a r g - a r g - a r g - a r g - a r g - a r g - v a l - a r g - g ly - g ly - a r g - g ly - a la - v a l - t h r -
g ly -a r g -a s p -g ly -a la -g ly -a r g - le u -C O O H
UAA s to p codon
i l e - a r g - t r p - a l a - c e r - a l a - t h r - a l a - a r g - s e r - g l y - t h r - v a l - l e u - a l a -
a l a - a l a - g l y - a l a - v a l - g l u - v a l - l e u - g l y - a l a - g l u - a l a - g l y - l e u - a l a -
t h r - p r o - p r o - a r g - a r g - g lu - v a l - v a l~ a s p - t r p - g lu - g ly - a la - t r p - a s p -
g lu -a s p -a s p -g ly - g ly -a la - p h e -g lu - g ly -a s p -g ly - v a l - le u - C O O H
B. AMIBO ACIB USAGB
amino a c id UGA s to p codon UAA s to p codon
pro 2 2
a rg 15 4
a la 8 12
g !y 24 9
o th e r s 29 31
to bal 68 5B
( c o n t i n u e d o v e r l e a f )
T able C5»6 co n tin u ed
G. THItfD CODOLi POSITION USAGE
n u c le o t id e UGA sto p codon UAA sto p codon
G 2 4 1 3° 1' 44
1[52C
t .
oCM 22 J
A 4 ]' 25 ’ 8
T 12 j 4 j

Figure C3.1
D ata f o r S s t I r e s t r i c t i o n maps o f HSV-1 BamH I k and HSV-2 BamH I g .
A 1 HSV-1 ■BamH I k2 HSV-1 BamH I 33 HSV-1 BcoR I k
D ig e s ted[ w ith S s t I ; a g a ro se g e l .4
B 1 HSV-2 BamH I2 HSV-2 BaraH I u3 HSV-2 BcoR I k4 HSV-2 BcoR I ra!? HSV-2 Bamli I uvw6 HSV-2 BcoR I £-~
D ig e s te d . w ith S s t I ; 2 ‘» a g a r o se g e l .
C 1 HSV-2 BamH I £2 HSV-2 BamH I u3 HSV-2 BcoR I k4 HSV-2 BcoR I m
D ig e s ted w ith S s t I ; '7 • 5; * p o ly a c rylam:
D 1 i.iSV-2 Damn I u2 HSV-2 npn I a
D ig e s te u w ith S s t I ; 1 .3 > a g a r o se g e l
B 1 HSV-2 Damn I2 HSV-2 Kpn I r s
D ig e s te d WitxJ S s t I ; 2?* a g a r o se g e l .
Bach o f t lis aoove fr a g m en ts was prex>ared from n ic k t r a n s la t e d nSV-1 o r nSV—2 DBA.
S o t I fra g m en ts o f Damn I j o i n t or te r m in a l fra g m en ts a re i n d ic a t e d , A .d e n o te s a genome te r m in a l fra g m en t.

* I H H ! !CO
CN
Ol
m 1m cb
i ioo
t- CM CO ^ in co
Is-
CD
in
co
CN
^ _ i _____ i i 1 1^ It *•- » .'•-4
i x a t I9
I * 1
6-|

Fig u re 0 3 . 2
Some d a ta f o r Sma I r e s t r i c t i o n maps o f HSV-1 BamH I k.
1 HSV-1 BamH I k2 HSV-1 BamH I k3 IISV-1 BamH I g4 HSV-1 BamH I k5 HSV-1 BamH I k6 HSV-1 BamH I g Sau5A
I A d ig e s t e d w ith Sma I B d ig e s t e d w ith Sma I G d ig e s t e d w ith Sma
2fs> a g a ro se g e l .
B 1 pBR322 H in f :2 HSV-1 BamH I3 HSV-1 BamH I4 HSV-1 BamH I
■ 5 HSV-1 BamH I6 HSV-1 BamH I7 HSV-1 BamH I8 HSV-1 BamH I
S s tS s t
5/^ p o ly a c r y la m id e g e l .
C 1 pBB322 H inf I m arkers2 -6 IISV-1 BamH I k Sau3A I A d ig e s t e d w ith
in c r e a s in g am ounts o f Sma I
2 fa agaro s e g e1 .
[ m arkersk d ig e s t e d w ith Sma I 1 Sau3A I A d ig e s t e d w ith Smak Sau3A I B d ig e s t e d w ith Smak Sau3A I C d ig e s t e d w ith Sma
B,B d ig e s t e d w ith SmaA d ig e s t e d w ith Sma I C d ig e s t e d w ith Sma I
JJ 1 HSV-1 BamH I k S s t w ith Sma I~
I C p a r i i a l l y d ig e s t e d
5 ft p o ly a c r y la m id e g e l .
B 1 iiSV-1 BamH I m Sau3A I B p a r t i a l l y d ig e s t e d w ith Sma I
3; p o ly a c r y la m id e g e l .
Sma I r e s t r i c t i o n fra g m en ts o f HSV-1 BamH I k a re in d ic a te u in p a n e ls A and 3 . S i z e s o f p a r t i a l d i g e s t i o n p r o d u c ts in bp a re in d ic a te d in p a n e ls C-B w itii Sma I r e s t r i c t i o n fra g m en ts in l e t t e r s .H ick t r a n s la t e d p lasm id nil A was U3ea.Sma I fra g m en ts o f iiSV-1 BamHI x Sau3AI A (430 bp i sp a r t o f Smal A) and a 1631 bp maricer a re in d ic a t e d in p a n e l F,
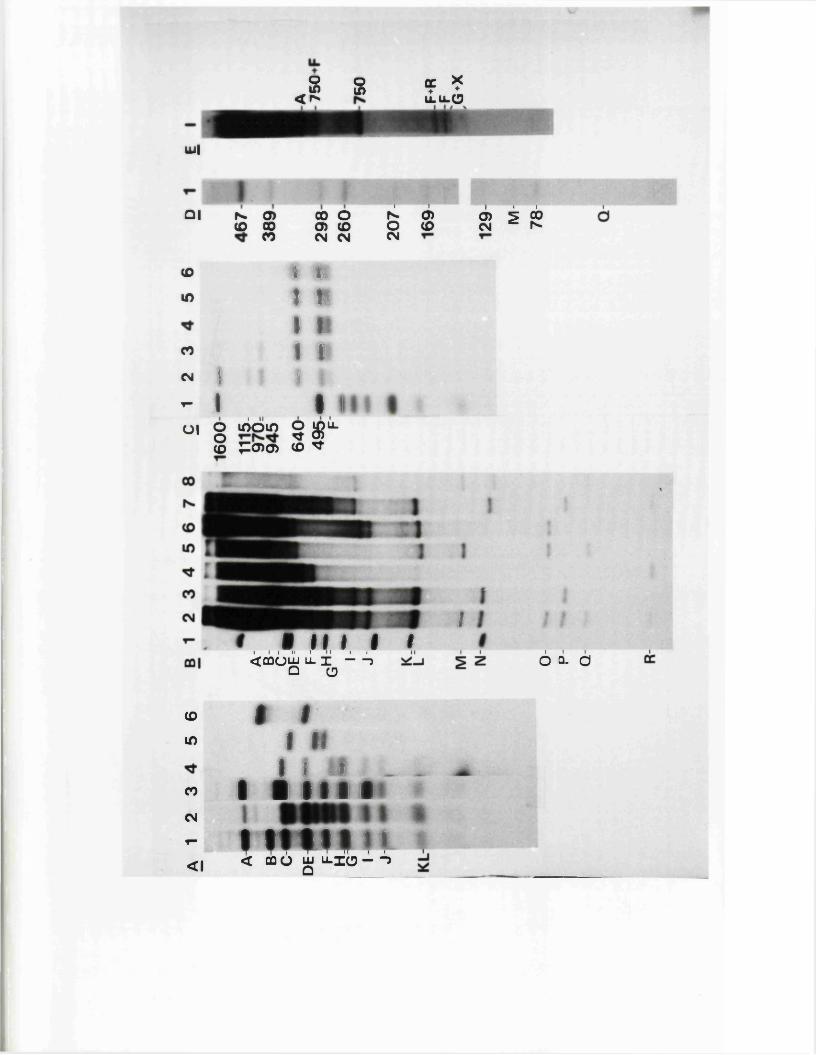
§ g< l" " f *L L U .O
Ui|
□ I
CO
If)
CO
CM
rs. O) CO CO
CO
CO O o> CO CM CM
I 1
I II
I II
I IJ I M
’■ I t n I Iui o m o m o mu-
1 o ^ 2(O ^0)0) co ^
I Ih- O)O CO CM
1 II I f Ii i i ii r ~ i i i<C O O L U LU I )O o
1 I "T ~05 g COCM Is*
/
I I Io q- a
CO
m
CO
CM
1—
<1
I II If
M i t I 1 I I I •
i t t f i i n< DQ O LU U - I 0 — “ )
Q

Figure C3.5
R e s t r i c t i o n maps o f HSV-1 BamH I R (u pp er p a n e l)
and. HSV-2 BamH I g ( lo w er p g n e l ) , w hich c o n ta in
th e j o i n t o f L and S . The r e c t a n g le in d ic a t e s
th e £_>osition o f th e a se q u e n c e , and b oth fra g m en ts
a r e ab ou t 6000 bp in s i z e . .
N om en cla tu re has been a s s ig n e d to th e fr a g m en ts o f
some d i g e s t s . Sac I and S s t I a re is o s c h iz o r a e r s .
R e s t r i c t io n maps f o r genome t e r m in i, ea ch o f
w hich h as a te r m in a l a seq u e n c e , may r e a d i ly be
deduced from th o se o f th e j o i n t r e g io n s ( s e e
i l g u r e 0 3 .1 3 ) .
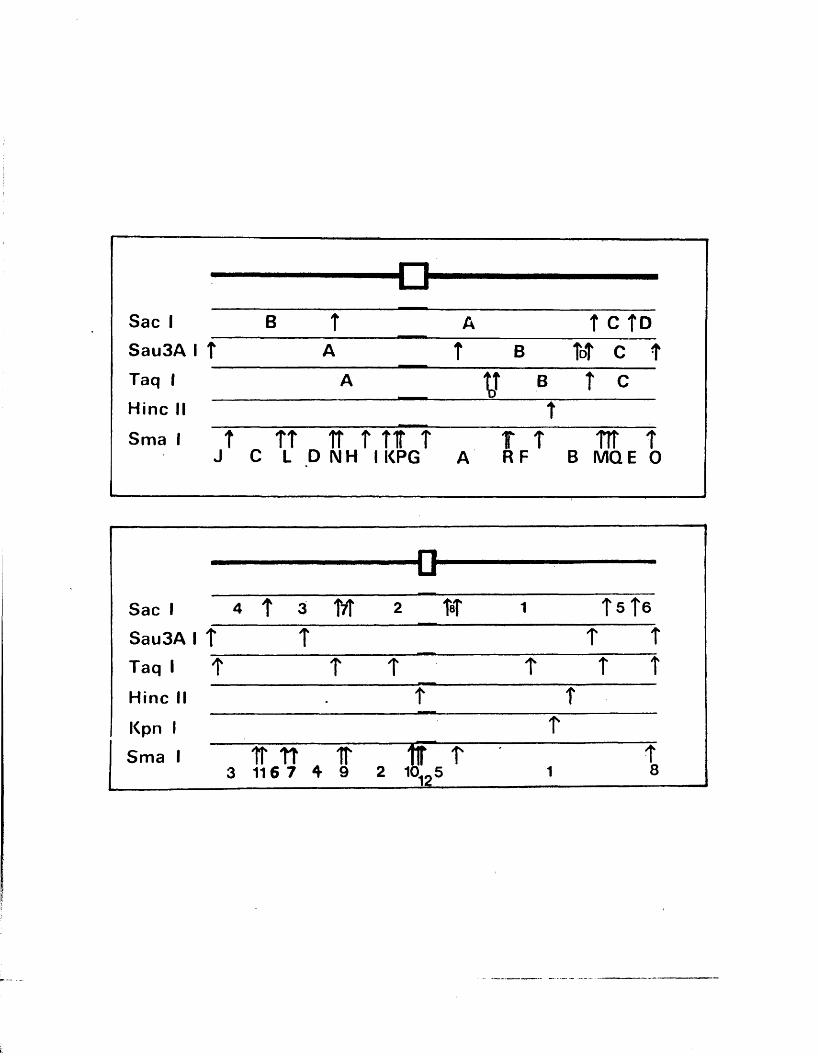
I- 1L_JSac 1 B T A t c ToSau3A 1 T A T B Tot c tTaq 1 A 1f f B T cHinc II tSma 1 t TT TT t TT T T T trr t
J C L D IMH 1 KPG A R F B MQE O
nUSac 1 4 T 3 2 Tat 1 T sSau3A 1 T T T TTaq 1 t T T T T THinc II t TKpn 1 rSma 1 TT TT3 116 7 4 ir
9i f r
2 1°125T1 8

figure C3.4
A u to ra d io g ra p h sh ow in g BamHI r e s t r i c t i o n p r o f i l e s o f
i n v iv o ^ P - l a b e l l e d v i r i o n DNA from te n s t o c k s o f
IiSV-1 s t r a i n 17 and t h e i r p a r e n t s to c k ( P ) , and from
te n s t o c k s o f HSV-2 s t r a in HG52 and t h e i r p a r e n t
s to c k ( P ) . Gel e l e c t r o p h o r e s i s was c a r r ie d o u t
on 0 .7 $ (HSV-1) and 1 $ a g a r o se (H SV -2). J o in t and
te r m in a l fr a g m e n ts , and fra g m en ts e x h ib i t in g s i z e
v a r i a b i l i t y , a r e i n d ic a t e d . R e s t r i c t i o n maps a re
shown in F ig u r e s C l018 and C l .1 9 .
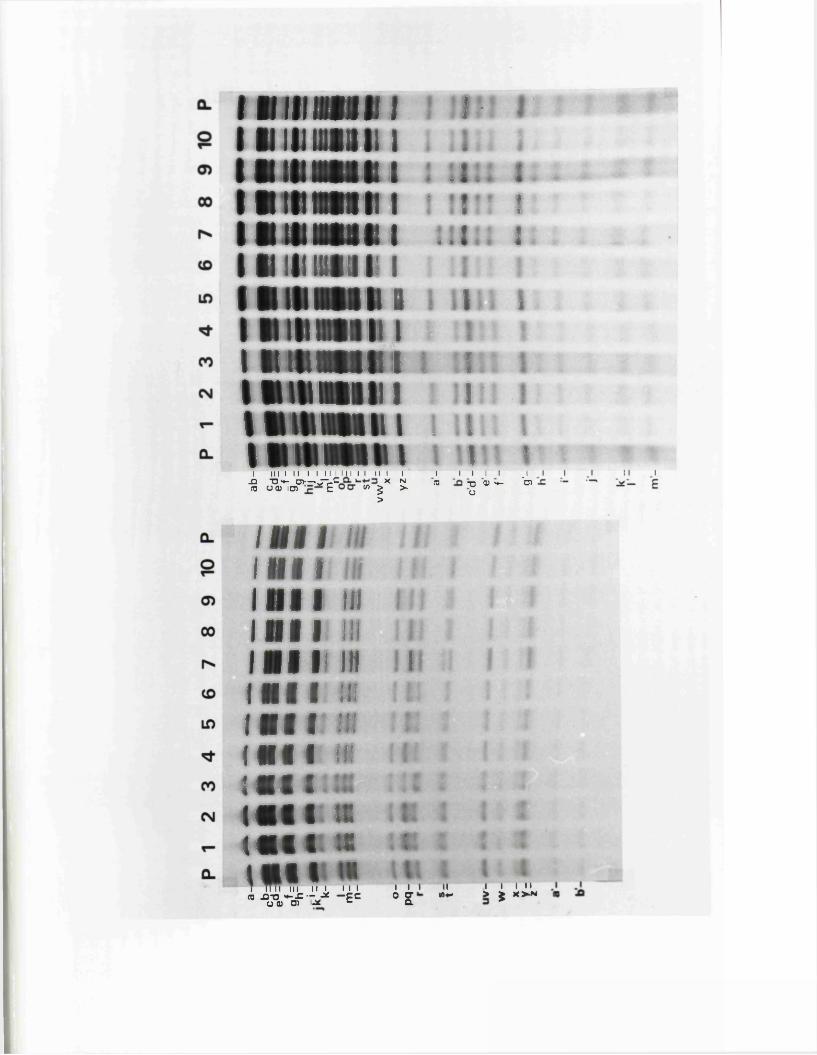
i n 111 u i iii I Ii f *
IUIUIMIIII1 1 1 I ! 1 1
m iu a u u i ' ! 1 * f f4 4 • 1 4 • *
mitt n u n ! Ill* Imui nuii i UUl 1m m uni r im i n i i 1 till 11 it in man i! itnitiuatii i f t K f 1I I I I I I
m m n u n 1 Ii nut man ii i 1 ? I 1 ’ 1 I 1 1 I 1m m ua in i r tilt i
I || I 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1a -a>*- o v - ' ^ -r c x n CO UD 105 E w £ >
>
i i i i i _ i ro "o m h- o) .
o
u / m i m
i i i i i i n
m u inm i l h i
mmiH i n
m u inm u inm u i n
m«i:&m e i m
m e t w t
m « i i «T TTTi in it i 11 i » i i m a -v *-a ■ - -* —g c 0 ?OO O ) W w Q .
II 1 1 1 II M «- > jg K > N

F igu re C 3.5
LEFT HAND PANEL
Autorad&graph show ing Smal r e s t r i c t i o n p r o f i l e s o f
Bam HI j o in t fra g m en ts i s o l a t e d from i n v iv o ^ P -
l a b e l l e d LNA from te n HSV-1 s t r a in 17 s t o c k s , t h e ir
p a r e n t ( P ) , and a m ix tu re o f th e te n fra g m en ts (M).
Gel e le c t r o p h o r e s i s was c a r r ie d o u t on a 7 * 5 $
p o ly a c r y la m id e g e l , and th e p o r t io n below J
r e p r e s e n ts a lo n g e r ex p o su re o f th e g e l . Fragment
s i z e s (bp) were c a l ib r a t e d w ith H a e l l l and H in fl
d i g e s t s o f pBR322 DNA ( S u t c l i f f e , 1 9 7 8 ) , and s i z e
r a n g es o f th e v a r ia b le fra g m en ts Smal A and G a re
in d ic a t e d . The Smal map i s shown in F ig u re C 3 .3 .
KIGIIT I1ANP PANEL
A u torad iograp h show ing TagI and Sau3AI p r o f i l e s o f' ' ' 32BamHI j o in t fra g m en ts i s o l a t e d from in v iv o P -
l a b e l l e d DNA from f i v e o f th e ten HSV-1 s t r a in 17
s t o c k s , w ith t h e i r p a r e n t ( P ) . E le c t r o p h o r e s is was
c a r r ie d o u t on a 1 .5 # a g a r o se g e l . TagI fra g m en ts4 _
a re in d ic a t e d on th e l e f t and Sau3AI fra g m en ts on
th e r i g h t . The p o s i t i o n s o f m o le c u la r w e ig h t
m arkers o f 1631 and 516 bp a r e shown. Maps a re
g iv e n in F ig u re C3.3*

< CQ O
*tO I
_ 00 I
S " I3 T-(0
CO 0 .
05 | |
to I ICO I
CM § |
« *“ ^I - 0 .
I I I In CD CJ £<o K
o
2 O O o O n O o o2 CO *fr£ ooIs- mm ^ to cm cmi i i ii i i i i -1
50”
143
t « I i t « %G ) 1 II 1 1 ! i0 0 • H 1 1 II I t •
1 II 1 111 I I iCO 1 II 1 1 1 ii o “ M l 1 1 1 1 l l
1 II 1CO u t i i i i t •CM i i i i i i i i ir - i i i i i i i
i i i i i i i i f0 - ' i i i i /
i ) i i ii i i i i VflOO IU U_ I -----5
Q g 111111111 jii_j
r - <£> 00 COO) CO Tt TtI I I I
I I
I I
I I I IO Q -O CC
<3
-38

F igu re C3»6
LEFT HAND PANEL
A u to ra d io g ra p h show in g Smal r e s t r i c t i o n p r o f i l e s o f32
BamHI j o i n t fr a g m en ts i s o l a t e d from i n v i v o P -
l a b e l l e d LNA from te n HSV-2 s t r a in HG52 s t o c k s , t h e ir
p a r e n t ( P ) , and a m ix tu r e o f th e te n fra g m en ts (M ).
E le c t r o p h o r e s is was c a r r ie d o u t on a 7.57*
p o ly a c r y la m id e g e l , and th e p o r t io n below 6
r e p r e s e n t s a lo n g e r ex p o su re o f th e g e l . Fragment
s i z e s a r e g iv e n in T ab le C 3 * l, and th e Smal map in
F ig u re C 3 .3 .
EIGHT HAND PANEL
Smal / -BamHI r e s t r i c t i o n p r o f i l e s o f recom b in an t
p la sm id s b ea r in g th e BamHI j o in t fra g m en t o f HSV-1
s t r a i n 17 or HSV-2 s t r a i n HG52 LNA* 2 j*g LNA was
a p p lie d to ea ch t r a c k o f a p o ly a c r y la m id e g e l ,4
w h ich a f t e r e l e c t r o p h o r e s i s was s ta in e d w ith e th id iu m
brom ide and p h otograp h ed under u l t r a v i o l e t i l lu m in a t io n .
The p la sm id v e c t o r pATl53 c o n ta in s no Smal s i t e s and
th e r e fo r e m ig r a te s a s a la r g e fragm en t in ea ch t r a c k .
C lo n es k l , k2 and k3 c o n ta in th e HSV-1 j o i n t fragm en t
w ith a s in g l e a se q u e n c e , x l and §2 c o n ta in th e same
fragm en t w ith two a se q u e n c e s , and g l c o n ta in s th e
nSV-2 j o i n t fragm en t w ith two a seq u en ces* HSV-1
Smal fra g m en ts a r e in d ic a t e d on th e l e f t , w ith s i z e
r a n g e s o f A and G, and s i z e s i n bp o f 0X174 Hael l l
m arxers on th e r i g h t . S tandard r e f e r e n c e num bers o f
th e c lo n e s a re g iv e n in Table B 1 .3 -
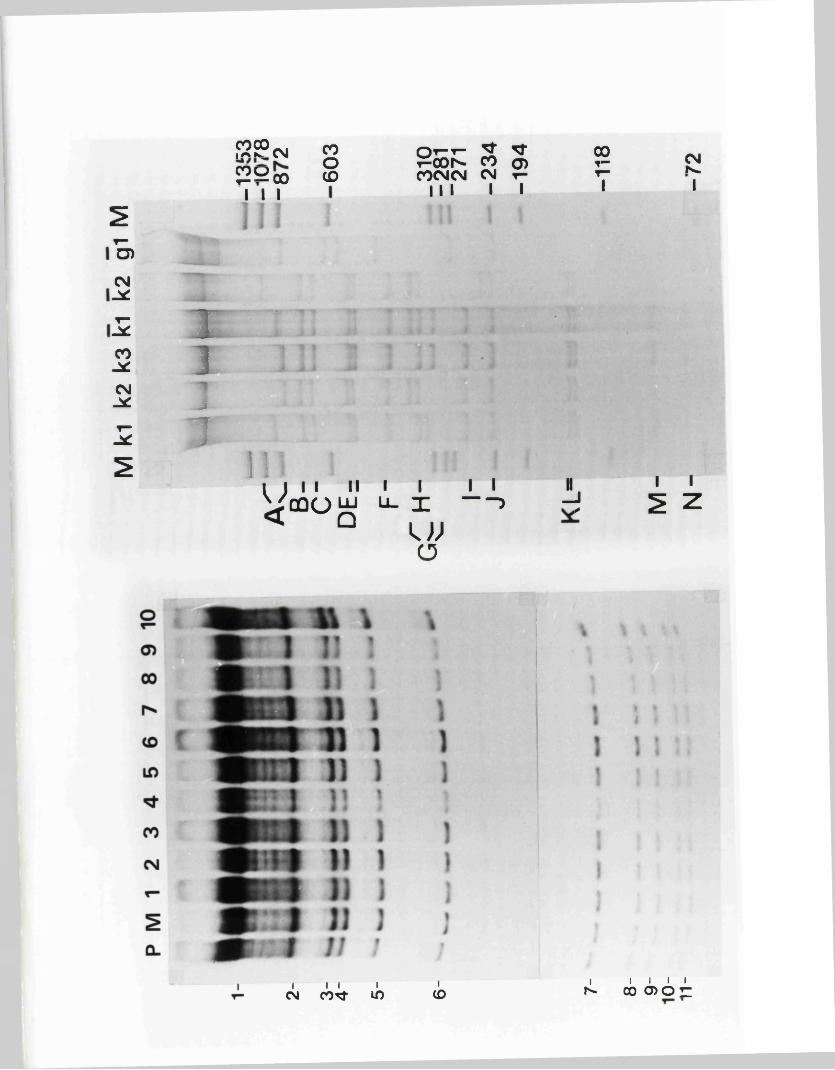
M k1
k2 k3
k1 k2
gl M
f2£?C M tO O ’- ’- St ^ 00 _ .o T-oor^ to 01 ’- £
S 2 2 o o tO COCMCM N r 7I I I I I I I I ' 1
I------- w
3
J
- -"T*
v _ »
7 1
i n
u<
I I II I I I I H, 1 1COO^ L*- X 3 ~} ^ Z
Q U Jo
\
i i i ieg coTt in
iCO
I I I I IIs- co o o n

Homology betw een th e j o in t r e g io n s o f th e
genomes o f HSV-1 and HSV—2 .
Recom binant p la sm id s c o n ta in in g th e HSV-1 or HSV-2
3amH I j o in t fra g m en ts were d ig e s t e d and tr a n s fe r r e d
to n i t r o c e l l u l o s e from a 4 $ p o ly a c r y la m id e g e l .
B lo t s t r i p s were th en h y b r id is e d w ith n ic k
t r a n s la t e d recom b in ant p la sm id DIJA.
1 k l Barnh I/Sm a I h y b r id is e d w ith k l
2 k l Barnh I/Sm a I n y o r id is e d w ith g l
3 . k l Bamil I / Sma I / S s t I h y b r id is e d w ith k l
4 k l Bamh I / Sma I / S s t I h y b r id is e d w ith g l
3 g l Bam a I / Sma I n y O rid ised w ith g l
6 g l Barnh i / Sma I n y O rid ised w ith k l
hSV-1 Barnh I k Sma I fra g m en ts are in d ic a t e d f o r
tr a c k s 1 - 4 , tn o se fra g m en ts in p a r e n th e s e s i n d ic a t in g
th e new p o s i t i o n s o f m ig r a tio n o f p o r t io n s o f Sma I
fra g m en ts a f t e r c le a v a g e w ith S s t I . HSV-2
Bamii I g Sma I fra g m en ts a re in d ic a te d f o r t r a c e s 3 - 6 .
Smal maps a r e snown in f ig u r e s C 3.3 and C 3 .1 3 .
fragm ent J o f g l , s in c e t h i s p lasm id h as two a
se q u e n c e s , maps in a l o c a t io n sp a n n in g th e a -a
ju n c t io n ( f ig u r e C 3 .1 3 ) .

3 4
G = *
(E)l J -

Some d a ta f o r the r e s t r ie t iu n m apping o f th e 1760 bp H in f I fragm en t w hich c o n t a in s th e j o i n t betw een HSV-1 L a n d S.-
A 1 Sau3A I A d ig e s t e d w ith H in f I2 Sau3A I A d ig e s t e d w ith HTnf I/Sm a I3 Sau3A I A d ig e s t e d w ith Sma I .
B *1 BamH I k d ig e s t e d w ith Sma I2 S /U d ig e s t e d w ith Sma I3 Sau3A I A d ig e s te d ~ w ith Sma I4 §7H d ig e s t e d w ith Ava I5 S/H d ig e s t e d w ith Ava I I6 S/H d ig e s t e d w itn Ava I l / Sma I7 S /n d ig e s t e d w ith BstN I8 S/II d ig e s t e d w ith BstN i/S m a I9 S/H d ig e s t e d w ith S ae I I
10 S /ii d ig e s t e d w ith Hae I I / Sma IM pBH322 Hae I I I m arkers ~*
C M pBR322 Hae I I I m arkers1 S/H d ig e s t e d w ith Sma I2 S /ii d ig e s t e d w ith S s t I I / Sma I3 S/H d ig e s t e d w ith S s t I I4 S/H d ig e s t e d w ith BgT I / Sma I5 s / i i d ig e s t e d w ith B gl I
A l l th e fra g m en ts were p rep ared from n ic k t r a n s la t e d p la sm id Li\lA c o n ta in in g HSV-1 BamH I k ( k l in P ig u re 0 3 . S ) . ,S/H r e f e r s to th e Sau3A I /H lh f I 1130 bp fra g m en t w hich c o n ta in s th e j o in t betw een L and S i n th e HSV-1 genom e. 7*b<<> p o ly a c r y la m id e g e l s were u se d .In p a n e l A the Sma I fra g m en ts o f Sau3A I A are in d ic a t e d 7 in c lu d in g th e 430 bp fra g m en t wKich i s partv o f Sma I A. The Sma I fra g m en ts o f BamH I k a r e in d ic a t e d in p a n e l and th e s i z e s o f pBB322r llae I I I m arkers in bp in p a n e ls B and C.
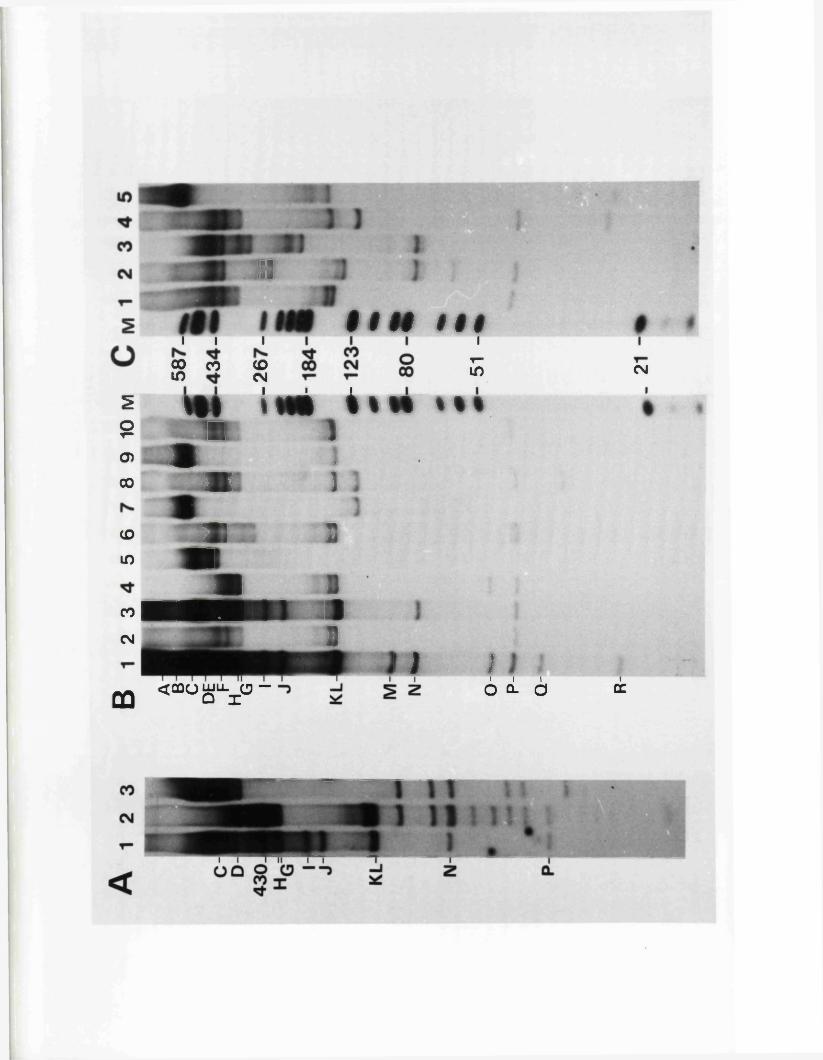
:::
i i i00 co coin CM
9 I H I I II
T—in
5 m i M i 1£ V . iO) 3 m
CO X )
CO X ; i iin S t
■ Bco H E f l H B ■ i i 3cm 2)
i i i i «%«
I I ;CQ < “ 0 ^ 0 -
i n io q- a
CM
Icc

Figure C5«9
R e s t r ic t io n e n d o n u c lea se maps o f th e j o i n t r e g io n s
o f HSV-1 s t r a in 17 and HSV-2 s t r a in HG52, and th e
n u c le o t id e seq u e n c in g s t r a t e g i e s em ployed . The
HSV-1 fragm en t (H in fI 1760 bp) c o n t a in s th r e e
r e i t e r a t i o n s shown in b la c k ( I , I I , I I I ) . R e s t r ic t io n
s i t e s in t h i s r e g io n o f th e DHA o f HSV-1 s t r a i n
USA-H were i d e n t i c a l to th o s e in s t r a in 1 7 ,
e x c e p t f o r an e x t r a Hinf I s i t e i n th e c seq u en ce
50 bp from th e a se q u e n c e . The HSV-2 fragm en t i s
S s t I 1310 bp ( BamHI g S s t I 2 ) .
L o c a t io n s o f th e d i r e c t r e p e a t s a t th e b -a and a - c
ju n c t io n s a re in d ic a t e d .
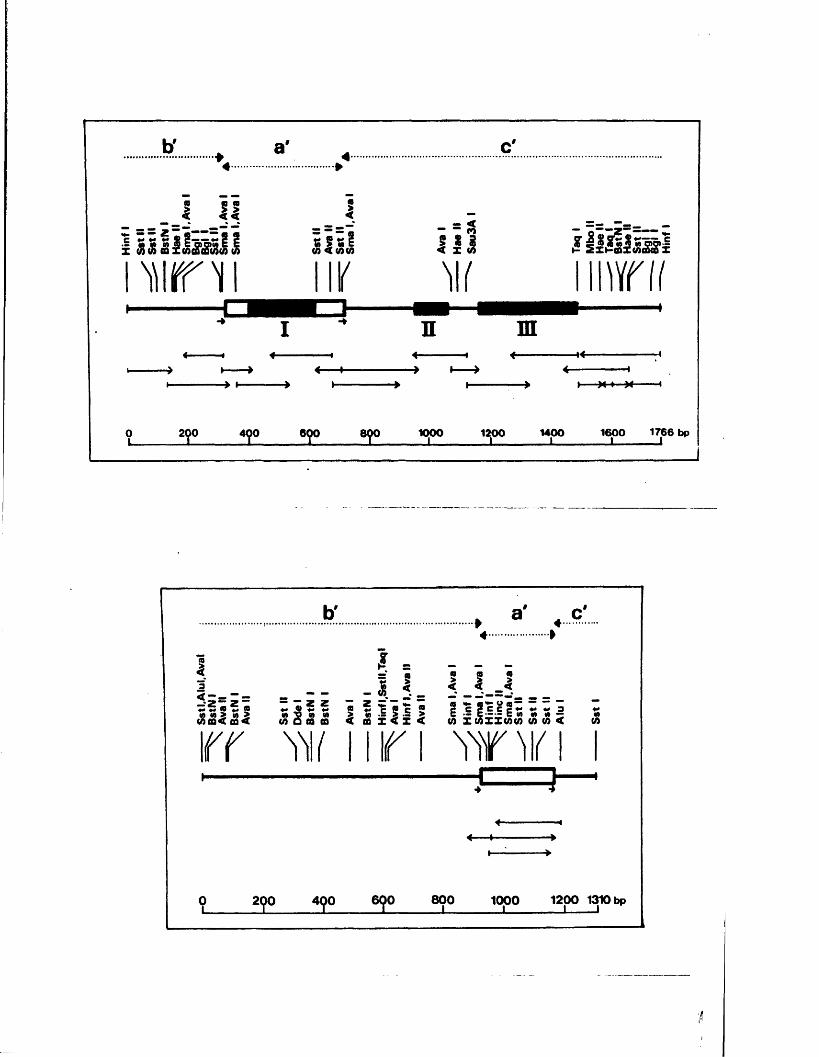
...........b ; .......... a ' 4 ............. C '* 4 .........
^ .............................. ►
i I I i
Hin
f I
Sst
II Ss
t II
BstN
1
Has
II Sm
a \J
Bg
l I
Bgl
1 Ss
t II
Sma
\J
Sma
1./
Sst
II Av
a II
Sst
IISm
a l.i
Ava
1 Ha
e II
Sau3
A
Taq
1M
bo
II Ha
e II
Taq
1 B
sfN
I Ha
e II
Sst
II
if!
iHi
nf
1
1 W l l n W ______________ I I I V O
->i n m
8-
*i
oo-
CMo- « f o a f o 1000 1200 1400 1 6 0 0 1766 bp
M 2 «C - Z ^** 3t +* +* 5 c fi cM j (0 (A > « ^ 3 2o o to m < to z < z
w i i n w w i
(0 X » I Z ( A ( 0 <0 <0 <
1200 1310 bp800 1000

F igure 0 3 . IQ
F in er d e f i n i t i o n o f the lo c a t io n o f HSV-1 a sequence
v a r i a o i l i t y .
Sau3A I A was i s o l a t e d from n ic k t r a n s la t e d p la sm id s
k l (1) and k 2 ( 2 ) , which c o n ta in i n s e r t s o f
hSV-1 BamH I k, and the l a t t e r o f which has a
l a r g e r Sma I (J fragment (F igure 03# 6 ) .
The two Sau3A I A fragm ents were c le a v e d as
i n d ic a t e d , and in eacn case the fragm ent which i s
la r g e r in k2 i s shown. The pBH322 Hae I I I marker
fragm ents (h) are in c lu d e d .
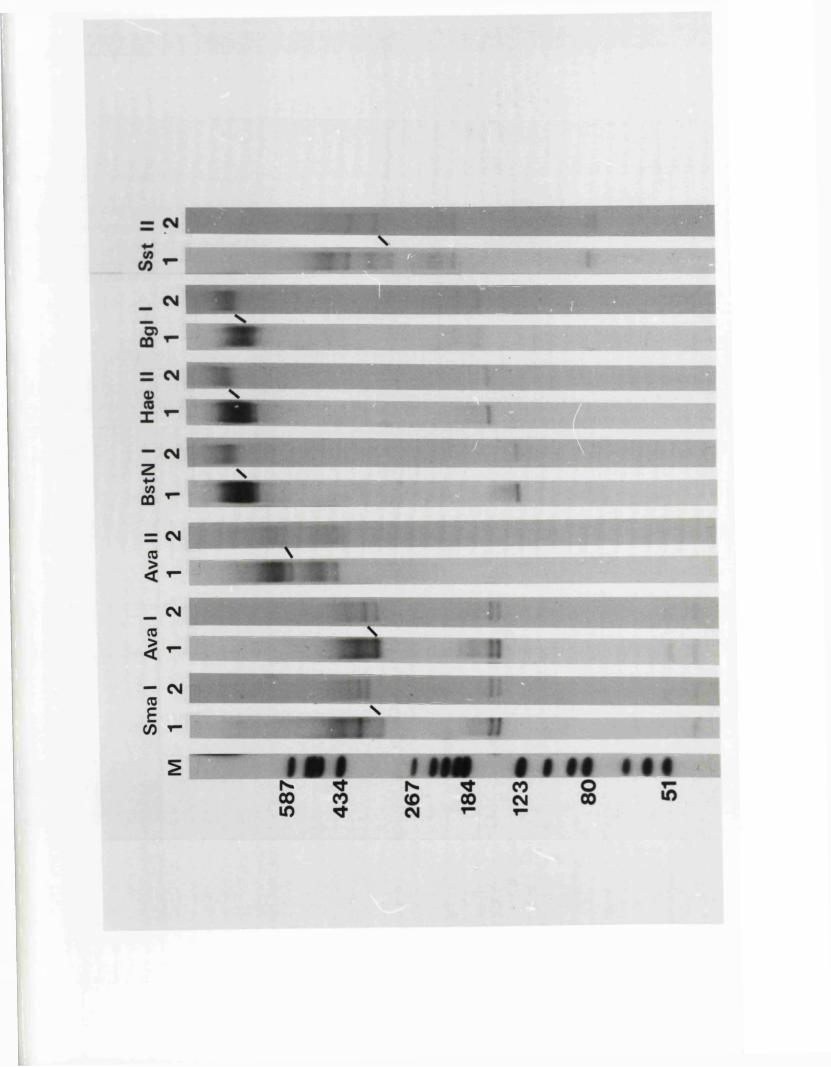

F igure C 5.l l
N u c le o t id e seq u en ces o f th e j o i n t r e g io n s o f HSV-1 and
HSV-2 Dl\iA, shown c o n v e n t io n a l ly 5 ' - 3 ' in the b ' - a ' - c 1
o r i e n t a t i o n .
A HSV-1 s t r a i n 17 c lo n e h i , w ith the p o s i t i o n s where
HSV-1 s t r a i n USA-8 d i f f e r s in d ic a t e d below the sequ en ce .
A dash r e p r e s e n t s the absen ce o f a n u c l e o t id e , and s t r a in
USA-8 was n o t sequenced between th e two s e t s o f th ree
d o t s . The l o c a t i o n s o f the genomic ter m in i w ith r e s p e c t
to th e j o i n t seq u en ces are shown ( s t r a i n 17 above and
s t r a i n USA-8 below the s e q u e n c e ) . These d e f in e th e
b -a and a -c ju n c t io n s and are c l o s e to the r i g h t hand
ends o f d i r e c t r e p e a t s (21 n u c l e o t id e s in s t r a i n 1 7 ,
17 in s t r a i n USA-8), the common 17 n u c l e o t id e s o f which
are boxed (311 -32 7 and 7 1 0 - 7 2 6 ) . R e i t e r a t io n s I , I I
and I I I are i n d ic a t e d , and the two AATAAA h e x a n u c le o t id e s
are boxed (1500 -1 505 and 13 4 6 -1 5 4 1 )• The r e g io n in
wnich the te r m in a l coded n u c le o t id e a t the 3' term inust ^
o f Vmw ih 175 mHHA was mapped i s shown (1530-1525)*
a number o f r ep ea ted s t r u c t u r e s o f unknown s i g n i f i c a n c eit
are p r e se n t but are n o t d i s c u s s e d in th e t e x t ( e . g . an* t
a lm o st e x a c t 18 n u c l e o t id e in v e r te d r e p e a t a t 243-260
and 2 8 1 -2 9 2 and a d i r e c t r e p e a t o f 20 n u c le o t id e s in
s t r a i n U3a- 8 te r m in a t in g a t 778 and 8 6 1 ) . The e n t i r e
sequence lias a GO c o n te n t o f 7 8 N.
H HSV-2 s t r a i n nGt>2. Tne l o c a t i o n s o f th e genomic
term in i w ith r e s p e c t to tn e j o i n t sequence are shown.
Tney are c l o s e to the r ig h t nand ends o f 17 n u c le o t id e
d i r e c t r e p e a t s ^ooxed) ana d e f in e tne p -a ana a -c
j u n c t io n s . The e n t i r e sequence has a GC c o n te n t o f 80?*.

GACTCGGGAA CGTGGAGCCA CTGGCGCAGC AGCAGCGAAC AAGAAGGCGG GGGCCCACCG GCGGGGGGCG 701 A
GCGGCGGGGC GGCCGCGGGC GCGCTCCTGA CCGCGGGTTC CGAGTTGGGC GTGGAGGTTA CCTGGGACTG 140
TGCGGTTGGG ACGGCGCCCG TGGGCCCGGG CGGCCGGGGG CGGCGGGGGC CGCGATGGCG GCGGCGGCGG 210
GCCATGGAGA GAGAGAGCGT GCCGGGGTGG TAGAGTTTGA CAGGCAAGCA TGTGCGTGCA GAGGCGAGTA
b'/la'GTGCTTGCCT GTCTAACTCG CTAGTCTCGG .CCGCGGGGGG CCCGGGqT-GC CCGCCGCCAC CGCTTTAAAG
G-
GGCCGCGCGC GACCCCCGGG GGGTGTGTTT TGGGGGGGGC CCGTTTTCGG GGTCTGG C C T
reiteration I( CCGCTCCTCCCC)18 CCGCTCC CGCGGCCCCG CCCCCCACGC(CCGCTCCTCCC—) g ( CCGTCTGTGGGTGGGGCTCCTCCC) g
280
350
407
650
CCGCCGCGCG CGCGCACGCC GCCCGGACCG CCGCCCGCCT TTTTTGCGCG CGCGCGCGqT'cGCGGGGGGC I G CGC C
S m al a f \ °GCGGGCJGCC ACAGGTGAAA----------- CCAACAG-----------------AGC ACGGCGCACT CCGCACGT-----------------------CA
C T CAACA AACACCAAC C GGCG TCCAGTTCCTCATCVCACGTCACGT CATCCACCAC ACCTGCCCAA CAACACAACT CACAGCGACA ACTCACCGCG CAACAACTCC
720
770
840
TGTTCCTCAT CCACACGTCA CCGCGCACCT CCCGCTCCTC CAGACGTACC CCGGCGCAAC ACACCGCTCCC A C G
reiteration IITGCTACACAC CACCGCC ( CCCTCCCCAGCCCCAG) 2 ( CCCTCCCCGGCCCCAG) g CCCTC
TC CCAG ( CCCTCCCCAGCCCCAG)15+ . . .
910
1060
CCCGGCGCGT CCCGCGCTCC CTCGGGGGGG TTCGGGCATC TCTACCTCAG TGCCGCCAAT CTCAGGTCAG 1130
reiteration IIIAGATCCAAAC CCTCCGGGGG CGCCCGCGCA CCACCA (CCGCCCCTCGCCCCCTC)19 C CGCCCCTCC^
V,. IE 175 mRNA 3 '-end_ _ _ _ 4 ...................................ATAA^CAACG CTACTGCAAA ACTTAATCAG* GTTGTTGCCG [TTTATTGCGT CTTCGGGTCT CACAAGCGCC
1500
1570
CCGCCCCGTC CCGGCCCGTT ACAGCACCCC GTCCCCCTCG AACGCGCCGC CGTCGTCTTC QTCCCAGGCG 1640
CCTTCCCAGT CCACAACTTC CCGCCGCGGG GGCGTGGCCA AGCCCGCCTC CGCCCCCAGC ACCTCCACGG 1710
CCCCCGCCGC CGCCAGCACG GTGCCGCTGC GGCCCGTGGC CGAGGCCCAG CGAATC 1766
Bb'A a'
TGGCGTGCGC AGCCCGGGCC GTGTTGCGGG CCCTCTTAAG GGGCGGCGGC AGGACGGGGA CTpCCGCCCC 70
GCCTCTTTTC CCCCGGGGAG TCAACCCCCG GGGGGGGTGT TTTTTGGGGG GGGGCGCGAA GGCGGGCGGC 140
GGCGGCGGGC GGGCGGCAGG GCAGCCCCGC GCGCCCCCTT CCCCGTCCCT CCCCCGGAGC CGGCCGCTCC 210
CCCGCGGGCG CCGCCCCTCC CCCCGCGCGC CGCGGGCTGC CTTCCGCGGC GCCCCCGCGC G.G
a > — ^ — iC'280
CCGCGCCCGC CCCCGCgCGC^jAGGACGGGG^CJAGCAGGC TGTGCCGCAG ACCACCACAC ACTCCCAA 348

Figure C3.12
A u to ra d io g ra p h show ing s i z e s o f genom ic te r m in a l
r e s t r i c t i o n fra g m en ts o f HSV-1 s t r a in s 17 and USA-8.3220 /*g HSV-1 UNA was P - l a b e l l e d a t 5 1 te r m in i i n a
volum e o f 2 0 y u l, and th e BamHI S and L te r m in a l
fra g m en ts i s o l a t e d . These w ere r e c le a v e d a s
in d ic a t e d and e l e c t r o p h o r e s i s c a r r ie d ou t on a
1 6 # p o ly a c r y la m id e (8 M u r e a ) se q u e n c in g g e l , u s in g
a s s i z e m arkers G-A and C-T c h e m ica l d e g r a d a tio n
n u c le o t id e se q u e n c in g r e a c t io n s (k in d ly p ro v id ed
by M -J. M u rch ie ), th e fra g m en t s i z e s o f w hich a r e
g iv e n in n u c l e o t i d e s . The S te r m in a l BamHI fragm en t
was co n ta m in a ted w ith th e L te r m in a l fragm en t
p o s s e s s in g an a d d i t io n a l a se q u e n c e .
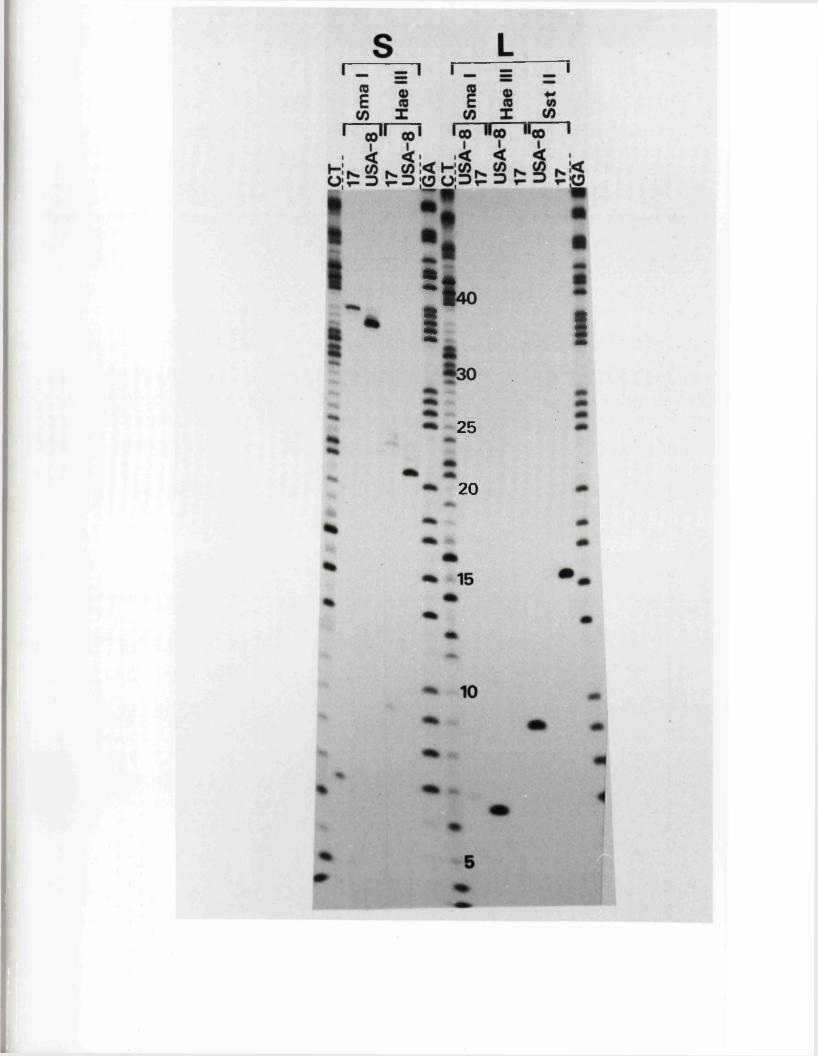
5 I
I I
25
20

Figure C3.13
H e s t r ic t io n maps o f th e BamHI fr a g m en ts o f HSV-1
UNA (u p p er p a n e l) and HSV-2 UNA ( lo w e r p a n e l)
w hich c o n ta in th e j o in t (6 kbp) and te r m in a l
r e g io n s . The two te r m in i a re a l ig n e d w ith th e
j o i n t so th a t th e p o l a r i t i e s o f th e th r e e r e g io n s ,
in c lu d in g th e a se q u e n c es ( r e c t a n g l e s ) , a re
e q u iv a le n t . The rem ainder o f th e genome i s
in d ic a t e d by a d o tte d l i n e . The S s t I map i s
shown above th e genom e, and th e Smal map b elow .
and kg d en o te S s t I and Smal te r m in a l fra g m en ts
o f th e genom e.
The p r e se n c e o f two tandem d i r e c t l y r e p e a te d a
se q u e n c es a t th e HSV-1 j o i n t , a s shown a t th e
bottom o f th e upper p a n e l , in tr o d u c e s a secon d
Smal G fra g m en t, and a fragm en t P 1 o f 43 bp w hich
i s th e same s i z e a t Smal P . The p r e se n c e o f two
tandem d i r e c t l y r e p e a te d a seq u e n c es a t th e HSV-2
j o i n t , a s snown a t th e bo i/fcom o f th e lo w er p a n e l ,
in tr o d u c e s a secon d Smal 12 fra g m en t, and a
fragm en t J o f 2^0 bp.
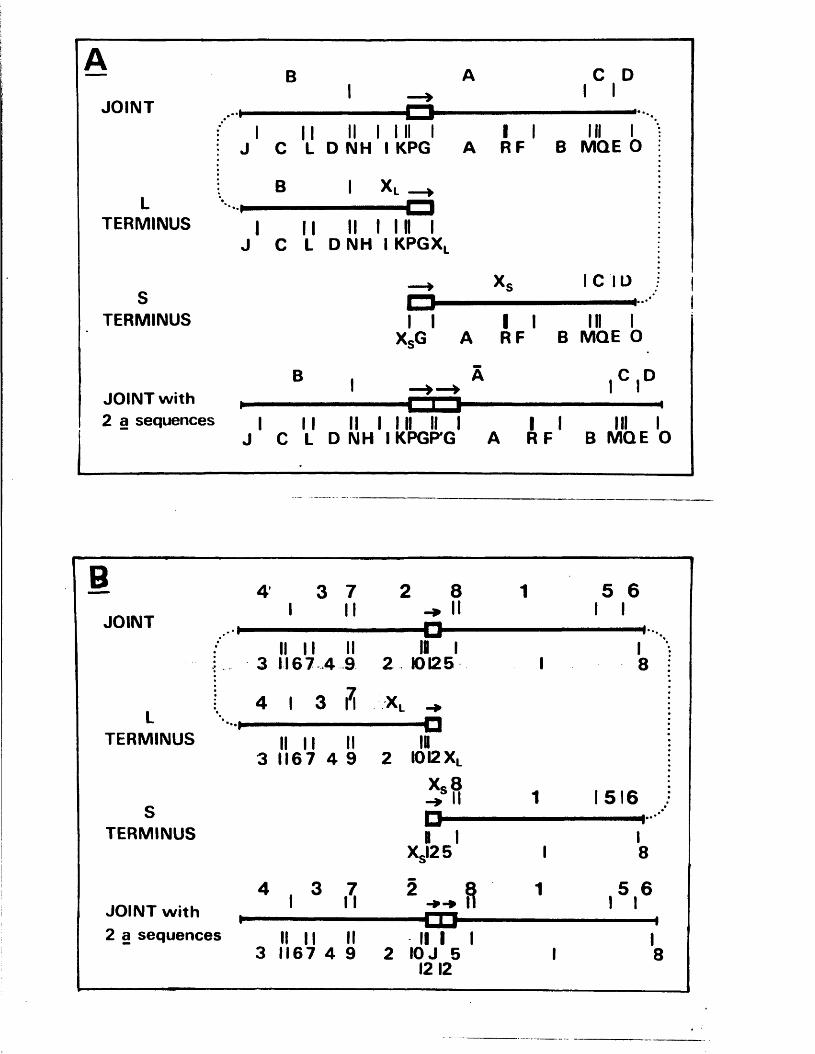
B C D
JOINTI | | II I III I
J C L D NH I KPGI I III I R F B M Q E O
TERMINUS
B X L
I II II I III I J C L D NH I KPGXl
TERMINUS
C I D
I I I IXSG A RF B MQE O
JOINT with 2 a sequences
B C D I I
I II II I III II I I I III IJ C L D NH I KPGP'G A R F B MQE O
BJOINT
4 3 7I II
II II II 3 1167 4 9
8
IB I 2 10125
5 6
84 I 3 n X L
TERMINUS
TERMINUS
JOINT with 2 a sequences
II II II 3 1167 4 9
i7;
n M M3 1167 4 9
III2 K)I2Xl
Xs 8-* II □ — I I
X sl25
c u ll I I
2 10 J 5 12 12
5 1 6
8
I 5 . 6
I8

X
gigure Q3.14
LEPT HAND TWO PANELS
A u to ra d io g ra p h o f 8 $ p o ly a c r y la m id e se q u e n c in g g e l s
show in g r e i t e r a t i o n I o f HSV-1 s t r a i n s 17 and USA-8.
Prom l e f t to r ig h t th e fragm en t seq u en ce shown i n ea ch group i s a s f o l l o w s .
1 . S t r a in 17 (p la sm id k l ) from Aval 370 to Avai l 67 0 .
2 . S t r a in 17 (p la sm id k l ) from Avai l 670 to B stN I 1 3 0 .
3 . S t r a in 17 (p la sm id k2) from Avai l 670 to BstN I 1 3 0 .
4 . S tr a in USA-8 from A v a il 670 to Aval 3 7 0 .
The r e i t e r a t e d seq u en ce i s shown i n ea ch c a s e , and t h e i r cop y number in p la sm id s k l and k 2 .
RIGHT HAND TWO PANELS
A u to ra d io g ra p h o f 6 ‘/Z p o ly a c r y la m id e se q u e n c in g g e l s4
show in g r e i t e r a t i o n I I o f HSV-1 s t r a in 17*
Prom l e f t to r ig h t th e fragm en t seq u en ce in ea ch group i s a s fo l lo w s *
1 . S t r a in 17 (p la sm id k l ) from S s t l l 710 to Sau3AI 1 1 3 0 .
2 . S t r a in 17 (p la sm id k2) from Avai l 670 to Sau3AI 1 1 3 0 .
Copy numbers o f th e two r e p e a te d u n i t s , w hich d i f f e r
by a s i n g l e n u c le o t id e , a re shown by l e t t e r s and num bers.
The main r e i t e r a t e d seq u en ce i s n o te d .
The tr a c k s a re in grou p s o f f i v e a s f o l l o w s .
G - c le a v e d a t G r e s id u e sA - c le a v e d a t G and A r e s id u e sAC - c le a v e d a t A and, to a l e s s e r e x t e n t , C r e s id u e s T - c le a v e d a t C and T r e s id u e sC - c le a v e d a t C r e s id u e s .
The r e s t r i c t i o n s i t e s and n u c le o t id e p o s i t i o n s ( to th e n e a r e s t 10 bp) r e fe x r e d to a r e shown in P ig u re C3*9.
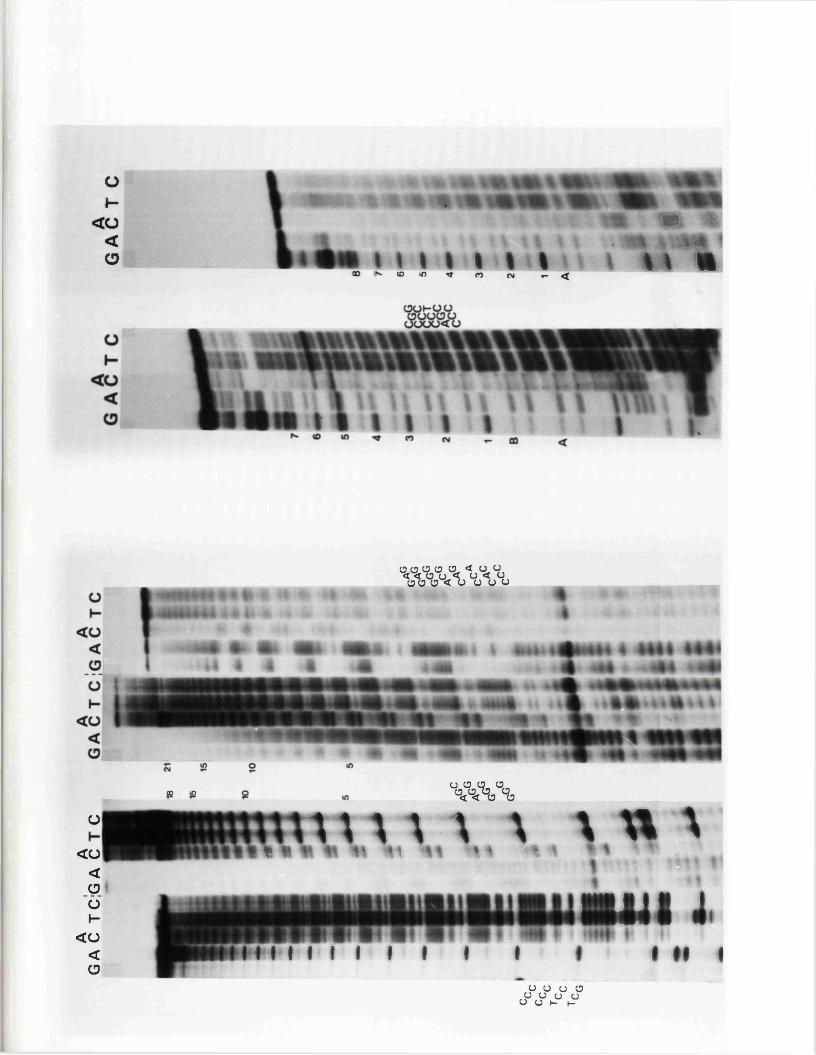
oI-
< o<o I I I I I I ICO K- (O in tT CO CM » - <
■ V I V\ v 1 4I I *
o o u o o < 0 0 < < o o < CJ < o (30 <J < OOOO
0
< 0<ooI-
< 0<e?
eg 10 0 0 0 O a o a o < < a a
1 1 a ; \ \ \ x i i1 1 * v " '
w u i 1
h 1 1 1 i 1 1 1 1 1 f 91 I
O O U C30 0 0 0 o o (- H

F igure 0 3 .1 5
LEFT HAND TWO PANELS
A u to ra d io g ra p h o f 8/» p o ly a c r y la m id e se q u e n c in g g e l s
sh ow in g r e i t e r a t i o n I I I o f HSV-1 s t r a in 1 7 .
From l e f t to r ig h t th e fra g m en t seq u en ce i n each group i s a s f o l l o w s .
1 . S t r a in 17 (p la sm id k l ) from TagI 1500 to Sau3AI 1 1 3 0 .
2 . S t r a in 17 (p la sm id k l ) from Sau3AI 1130 to TagI 1 5 0 0 .
3 . S tr a in 17 (p la sm id k2) from Sau3AI 1130 to TagI 1 5 0 0 .
The r e i t e r a t e d seq u en ce i s n o te d , and th e copy numbersin p la sm id s k l and k 2 .
RIGHT HAND THREE PANELS
A u to ra d io g ra p h o f 6 $ p o ly a c r y la m id e se q u e n c in g g e l s
show ing th e d i r e c t r e p e a t s a t th e a - c and a - a ju n c t io n s
o f HSV-1 s t r a i n 17 and a t th e c - a ju n c t io n o f HSV-1
s t r a i n USA-8.
From l e f t t o r ig h t th e fragm en t seq u en ce i n ea ch group i s a s f o l l o w s .
1 . S t r a in 17 (p la sm id k l ) from Avai l 670 to Sau3AI 1 1 3 0 .
2 . S t r a in 17 (p la sm id £2 ) from Avai l 670 to M nll 4 2 0 . T h is p la sm id h as two a s e q u e n c e s , so th e n u c le o t id e seq u en ce p ro ce ed s from th e Avai l s i t e i n one a se q u e n c e , a c r o s s th e ju n c t io n betw een th e tw o“ a s e q u e n c e s , to th e Mnll s i t e in th e n e x t .
3 . S t r a in USA-8 fro m -H in fI 770 to H a e ll 1 6 0 .
The r e p e a te d seq u en ce i s n o te d ; t h a t in (3 ) i s from th e com plem entary s tr a n d o f th e d u p lex to t h a t shown in F ig u r e 0 3 .1 1 * from r ig h t to l e f t . The lo c a t io n s o f M nll s i t e s were deduced from F ig u re 03*11• S t r a in USA-8“h a s an a d d i t io n a l H in fI s i t e a t 770 compared w ith s t r a i n 1 7 .
Track d e s ig n a t io n i s th e same a s f o r F ig u re 0 3 * 1 4 .The r e s t r i c t i o n s i t e s and n u c le o t id e p o s i t i o n s( to th e n e a r e s t 10 bp) r e f e r r e d to a re shown in F ig u re 0 3 .9 .

< o<o
(3 U U a O O (DO CD O OCT O CTO
O O O CD O CD
* * l |
\ M O A W W
O O 0,00000000 0 00O H o ,P o o p
O ' % ' % Xi - \ \% \ \ \ v N V
< o m \ \\\\ \ V \kO 0 0 J J O O O O O O O o o o
(Jh*
< o<o i i i i u m
.\V 9\V\ \ ‘ * I
oI-
< o<CDoI-
< o<C3
0> tf)
M M H I I I IM M M I I « I I I I I
' • 4 • ♦ ♦ » • #t ♦ ♦- i M i l t • I* # •M t l M I I I ♦ I I I I
o o o o o o o o t- o o
I - O (3 O U OM l ‘ II I
A ► Hi «•#* M< « 4 tit i » | |1 1 1 I I 1 •1 « « f « 1
4 4 « *# # - * ♦ > « M 9 * 4 < m• 0 • *• 0 N 9 I >♦ • * # * * 9# » 9 •
o o o o o0 0 , 0 0 0 rfO o o o o

Figure 03.16
L o c a t io n o f th e 3' term in u s o f HSV-1 Vmw IE 175
mRNA w ith r e s p e c t to th e DNA se q u e n c e .
The Taq.I D fragm en t c o n s i s t in g o f n u c le o t id e s
1 4 9 3 -1 6 0 8 was l a b e l l e d a t th e 3 ! te r m in i and s e p a r a te
s tr a n d s w ere i s o l a t e d . The s tr a n d com plem entary to
Vmw IE 175 mRNA ( t h a t shown i n F ig u r e 0 3 *11) was
i -y b f id is e d to RNA a s d e sc r ib e d i n th e t e x t , t r e a te d w ith
SI n u c le a s e and e le c tr o p h o r e s e d a lo n g s id e th e DNA seq u en ce
o f th e same fragm en t ( 3 f - 5 f r e a d in g from th e bottom ) on
an 8 fo p o ly a c r y la m id e (8 M u re a ) g e l . The tr a c k s a r e a s
f o l l o w s .
1 - u n tr e a te d s in g l e s tra n d ed DNA fragm en t
2 - a n n ea led w ith 10 fAg y e a s t RNA ( c o n t r o l )
3 - a n n ea led w ith 5 j jg II3V-1 p o ly a d e n y la te d IE mRNA
4 - a n n ea led w ith 10 iISV-1 p o ly a d e n y la te d IE mRNA
5 - a n n ea led w itn 15 HSV-1 p o ly a d e n y la te d IE mRNA
6 - a n n ea led w ith i s o l a t e d 4 .7 kb IE mRNA w hich c o d e s f o r
T racks G, A, T and C in d ic a t e th e DNA se q u e n c in g r e a c t io n s
a s d e s ig n a te d in F ig u re 0 3 .1 4 .
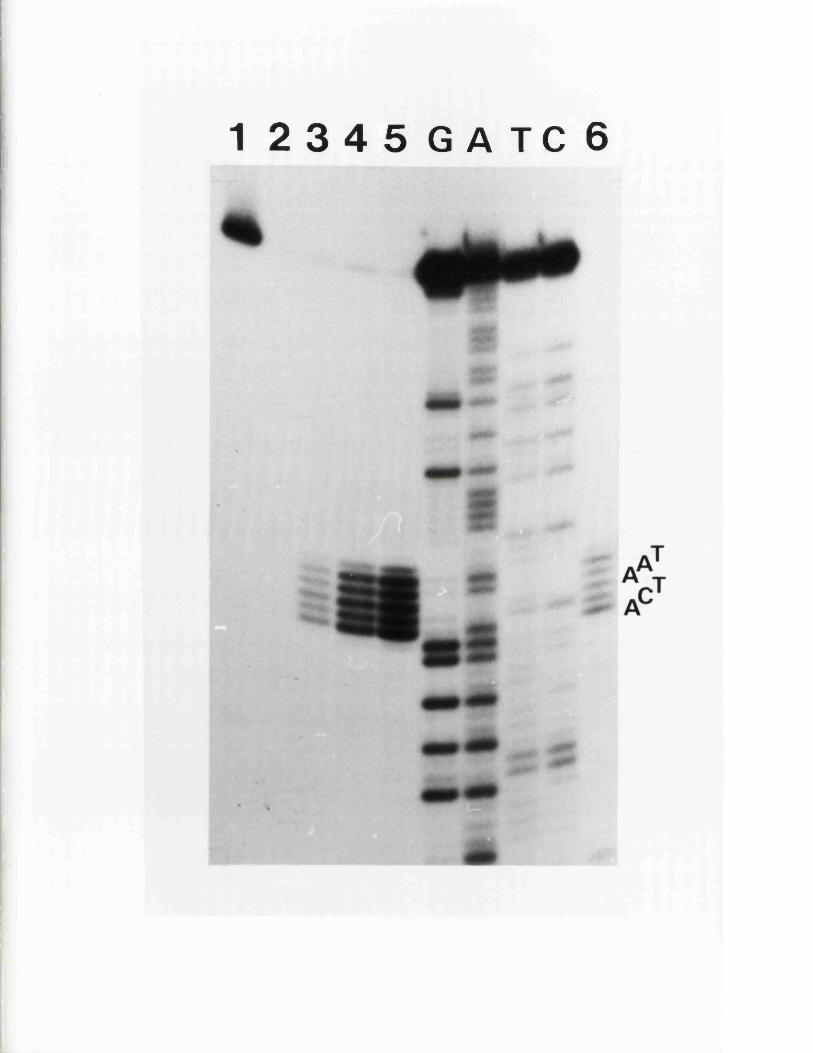
1 2 3 4 5 G A T C 6

S t r u c tu r a l arran gem en ts a t th e j o in t and te r m in i
o f th e HSV genome (a b o v e ) , and a t j o i n t s c o n ta in in g two
a seq u e n c e s ( b e lo w ) . The a se q u e n c e s a r e shown a3
r e c t a n g le s and t h e i r o r ie n t a t io n and th e l o c a t i o n s . o f
th e d i r e c t r e p e a t s a t th e b -a and a - c ju n c t io n s a re
i n d i c a t e d .
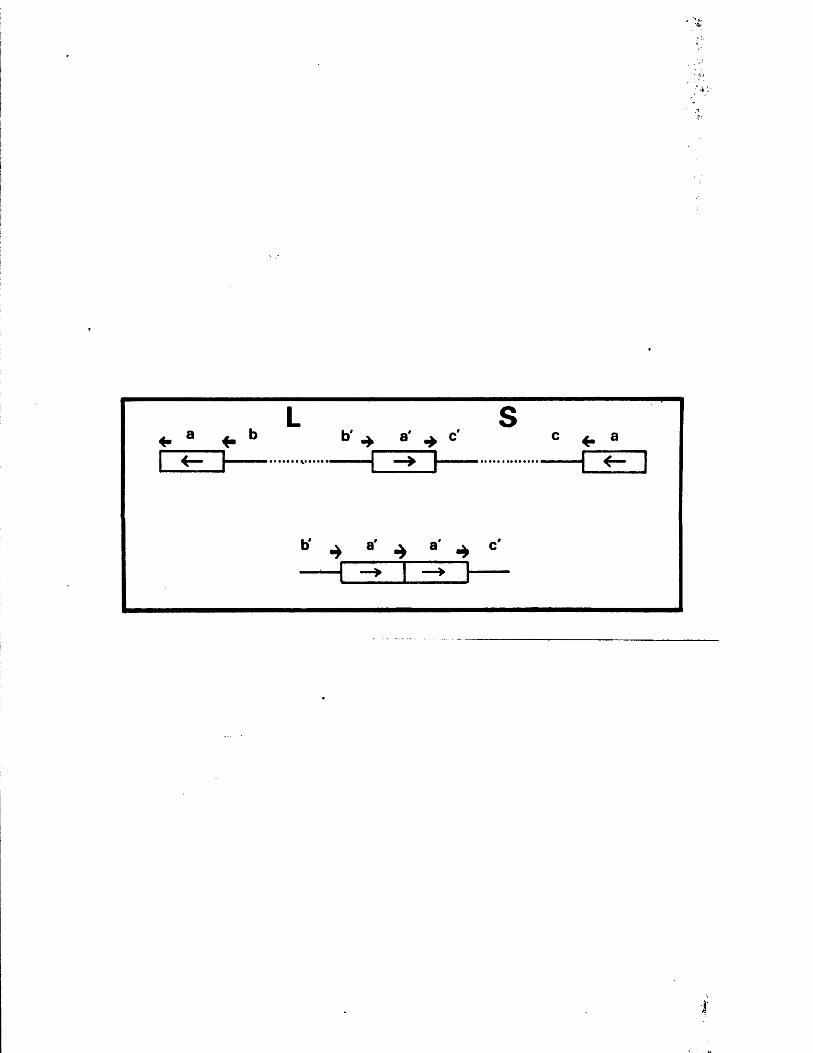

SECTION 4
INVERSION OP THE TWO SEGMENTS
OP THE HSV GENOME

- 130 -
RESULTS
In tr o d u c t io n
doth o r ie n t a t io n s o f th e L and S segm en ts o f HSV-1 and
113V-2 v ir io n DNA a re p r e s e n t i n eq u a l am ounts, g iv in g r i s e to
fo u r genome arra n g em en ts . T h is i s a l s o a f e a t u r e o f HSV-l/HSV-2
i n t e r t y p ic reco m b in a n ts . The mechanism by w hich segm ent
in v e r s io n o c cu r s i s unknown. V ir io n DNA o f th e recom b in ant
B x l^ 2 8 ), a r e s u l t o f g e n e t ic r e c o m b in a tio n betw een HSV-1 tsB
and IISV-2 t s l , was shown by P r e s to n e t a l . (1 9 7 8 ) to be a t y p ic a l
in p o s s e s s in g 1 p red o m in a n tly in one o r i e n t a t io n , w hereas
S was p r e s e n t e q u a lly in b oth o r i e n t a t io n s (P ig u r e C 4 .3 ) .
T h is p henotype was d e f in e d a s Mf ix e d " , norm al in v e r s io n b e in g
termed " fr e e " . I t m ust be s t r e s s e d t h a t i n f ix e d reco m b in a n ts
th e m a jo r ity , but n o t 'the e n t i r e t y , o f v i r i o n DNA m o le c u le s
p o s s e s s th e a p p r o p r ia te segm ent i n th e m ajor o r i e n t a t io n .
P r e s to n e t a l . (1978) co n clu d ed from g r o s s r e s t r i c t i o n
a n a ly s i s th a t B x l(2 8 ) p o s s e s s e s HSV-2 seq u en ce in TR and HSV-1
seq u en ce in IR^. One p o s s i b i l i t y , t h e r e f o r e , i s t h a t a r e g io n
o f t y p e - s p e c i f i c hom ology betw een TR and IRj, i s n e c e s s a r y f o r
normal in v e r s io n o f 1 . However, o th e r reco m b in a n ts a ls o
seemed h e t e r o ty p ic f o r TR j/lR^ and y e t in v e r t e d n o r m a lly .
I t i s n o t p o s s ib le by g r o s s r e s t r i c t i o n a n a l y s i s to d i s t in g u i s h
betw een HSV-1 and HSV-2 DNA seq u e n c es in r e g io n s c l o s e r to th e
j o in t or te rm in i th a n 4000 bp, so in o r d er to l o c a t e p r e v io u s ly
u n d e te c te d c r o s s o v e r s th e j o i n t and te r m in a l r e g io n s were
a n a ly sed in d e t a i l f o r B x l ( 2 8 ) , f o r B x l(2 8 ) s u b c lo n e s , and
fo r a number o f o th e r reco m b in a n ts .
The recom b in an t KH4 was g e n e r a te d by marker r e sc u e o f
HSV-1 t s B DNA by HSV-2 DNA fra g m en ts (W ilk ie e t a l . , 1 9 7 9 b ) .
KH/| was a n a ly se d in d e t a i l in th e work d e s c r ib e d h ere and
shown to oe f ix e u in L and S, a l lo w in g in v e r s io n o f both

- 131 -
segm en ts to be i n v e s t i g a t e d .
The g e n e s c o d in g f o r V ^ IE 110 (H SV -l) and i t s c o u n te r p a r t
in II3V-2, Vfi.w IE 1 1 8 , map e n t i r e l y w ith in TR and IHj,. P r e s to n
e t a l . (1 9 7 8 ) showed t h a t reco m b in a n ts w M ch 'a re h e t e r o t y p ic
f o r t h i s d ip lo id gene in d u ced b oth IE 110 and IE 1 1 8 . Two
s u b c lo n e s o f B x l(2 8 ) a re shown h ere to be d e le t e d in TRj, or IRj,,
and th e s e a llo w e d i n v e s t i g a t i o n o f th e e x p r e s s io n o f t h i s gene"
by genomes w ith o n ly one f u l l co p y .
In th e f o l lo w in g d e s c r ip t io n s i t i s ap p aren t t h a t th e
i n t e r p r e t a t io n o f d e t a i l e d r e s t r i c t i o n a n a ly s e s o f recom b in ant
genome s t r u c t u r e s i s v er y com p lex . T h e r e fo r e , a lth o u g h a
s i g n i f i c a n t p r o p o r t io n o f d a ta h a s been in c lu d e d i n th e
P ig u r e o , i t s in t e r j j r e t a t io n h as been l im i t e d to a few exam p les.
Emphases a r e p la c e d on s t r u c t u r e s c r i t i c a l t o th e c o n c lu s io n s
drawn, on d e m o n str a tin g th e l i m i t s o f th e a n a l y s i s , and on
show ing th e s t r u c t u r a l s u b t l e t i e s o f w h ich HSV recom b in an t
genom es a re c a p a b le .
S tr u c tu r e s o f th e .jo in ts and te r m in i o f B x l ( 2 8 ~ l )- • ^ • - -
The w ork ing s to c k u sed in t h e s e ex p er im e n ts was named
B x l ( 2 8 - 1 ) , th e term B x l(2 8 ) b e in g r e t a in e d f o r th e p u ta t iv e
o r i g i n a l recom b in an t a s d e s c r ib e d b e lo w . The DNAs o f t s B , t s l
and B x l (2 8 -1 ) were ^2P - la b e l l e d by n ic k t r a n s l a t io n and c le a v e d
w ith S s t I o r Smal and th e p r o d u c ts se p a r a te d by e le c t r o p h o r e s i s
on 2 $ a g a r o s e , 7 .5 $ p o ly a c r y la m id e o r 1 5 $ p o ly a c r y la m id e g e l s .
The r e s u l t i n g p r o f i l e s were a n a ly se d u s in g th e r e le v a n t
r e s t r i c t i o n maps o f th e s e r e g io n s o f th e genome (P ig u r e C3#3)*
C lea v a g e p r o d u c ts o f te r m in i w hich do n o t co rresp o n d to
fra g m en ts from th e j o in t r e g io n , and a re th e r e fo r e th e
te r m in a l Ss t I or Smal fra g m en ts o f th e genom e, a re r e f e r r e d to
a s X ( s e e P ig u re C 3 .1 3 ) . I t was som etim es n e c e s s a r y to c le a v e
c o n ig r a t in g fra g m en ts ( e . g . HSV-2 BamHI v w ), and in t h e s e

- 132 -J
in s t a n c e s a t t e n t io n i s drawn o n ly to c le a v a g e p r o d u c ts from /ir
j o in t or te r m in a l r e g io n s . P a in t bands a r i s i n g from
c o n ta m in a tin g se q u e n c e s , a fe a tu r e o f th e u se o f n ic k
t r a n s la t e d DUA, a re a l s o n o t d is c u s s e d .
F ig u re C 4.1 p a n e l A shows th e S s t I c le a v a g e p r o f i l e s o f
B x l(2 8 -1 ) j o i n t fr a g m e n ts . Track 2 shows th e m ajor j o in t
fra g m en t, w h ich i s ty p e 1 , and a ls o HSV-2 S s t I fra g m en ts from
th e b and c seq u e n c es i n l e s s e r abundance. The la r g e r j o in t
fragm en t ( tr a c k 3 ) h as bands c h a r a c t e r i s t i c o f both th e HSV-1
and IISV-2 j o in t r e g io n s , and p rob ab ly c o m p r ise s m a in ly th e ty p e
1 j o in t w ith an e x tr a a seq u en ce (HSV-1 S s t I A i s la r g e r ) and
ty p e 1 IHl jo in e d to ty p e 2 IRg ( th e n o v e l j o i n t fragm en t
p o s s ib ly m ig ra ted above IISV-2 S s t I 1 , p erh a p s w ith two a
s e q u e n c e s ) . Two r a r e r form s may a l s o be p r e s e n t : one w ith
ty p e 2 IR^ jo in e d to ty p e 1 IRg, and th e o th e r w ith HSV-2 DNA
'throughout th e j o i n t . These r e s u l t s show t h a t th e m ajor j o i n t
s p e c ie s i s ty p e 1 th ro u g h o u t, but t h a t o th e r s p e c ie s due to
in v e r s io n a r e a l s o p r e s e n t . P a n el B tr a c k 2 shows a Smal
d i g e s t o f th e m ajor j o i n t o f B x l ( 2 8 - 1 ) , in w hich a g a in th e
m ajor bands a re ty p e 1 . HSV-1 Smal G, b e in g a fragm en t v a r ia b le
in s i z e , i s s m a lle r in t h i s recom binant th a t in t s B . Track 3
o f p a n e l C and tr a c k 3 of* p a n e l D show t h a t th e m ajor L
term in u s i s ty p e 2 , in c lu d in g th e a se q u e n c e . The HSV-1 m inor
L term in u s i s b a r e ly d e t e c ta b le in p a n e l C tr a c k 4 , s in c e L
i s f i x e d .
P r e s to n e t a l . (1 9 7 8 ) d em on strated t h a t th e HSV-2 Kpnl
s i t e in TRg/IRg was to some e x te n t s e g r e g a te d betw een b oth
TRg and IRg in B x l ( 2 8 - 1 ) , and so th e two S te r m in i were
i s o l a t e d s e p a r a t e ly . The un ique HcoRI S term in u s o f B x l ( 2 8 - 1 ) ,
c o n v e n t io n a l ly th e r ig h t hand s id e o f S , c o n ta in s HSV-1 and

- 133 -
HSV-2 S s t I fragm en ts c o n s i s t e n t w ith i t b e in g a m ix tu re o f
th e norm al IISV-2 S te r m in a l BcoRI fra g m en t and a fragm en t o f
s im ila r s i z e w ith a c r o s s o v e r in TRg (F ig u r e C 4.2 p a n e l A;
summarised in F igu re C 4 .1 8 ) . S m all s i z e d i f f e r e n c e s betw een
some fra g m en ts from p a r e n ta l and recom b in an t genom es ( e . g .
HSV-2 S s t I 1) a re p ro b a b ly th e r e s u l t o f s i z e v a r i a b i l i t y in
th e s e r e g io n s . The S s t I p r o f i l e o f th e B g ll l S te r m in u s ,
c o n v e n t io n a l ly th e l e f t hand s id e o f S , i s c o n s i s t e n t w ith i t
b e in g a m ixtu re o f th e norm al HSV-2 S te r m in a l B g ll l fragm en t
and a fragm en t w ith a c r o s s o v e r i n TRg in an e q u iv a le n t
l o c a t io n to th a t d e s c r ib e d above (F ig u r e C 4.2 p a n e l C ).
P a n e ls B and 1) o f F ig u r e C4 .2 d em o n stra te th e p r e se n c e o f th e
Kpnl s i t e i n HSV-2 TR3/IK 2 in a t l e a s t a p r o p o r t io n o f both
S term in a l fra g m en ts of- B x l ( 2 8 - l ) . Smal d ig e s t io n o f th e
BamHI S te r m in a l fra g m en ts o f B x l ( 2 8 - l ) (F ig u r e C 4.2 p a n e l E)
shows th e p re sen c e o f an HSV-1 a seq u en ce a t th e ty p e 1
term in u s and an HSV-2 a seq u en ce a t th e ty p e 2 te rm in u s ,
deduced in t h i s c a s e from th e p r e se n c e o f HSV-2 Smal £ .
These r e s u l t s co n firm th a t B x l ( 2 8 - l ) i s f i x e d in L and
show t h a t , in th e m ajor o r ie n t a t io n o f 1 , TR and IR-^,
in c lu d in g txie a se q u e n c e s , a re h e t e r o t y p ic . "S c o n s i s t s o f a
m ixtu re o f a t l e a s t two s t r u c t u r e s , th e c r o s s o v e r i n one o f
w hich i s p rob ab ly m ost a c c u r a t e ly lo c a t e d w ith in HSV-1 BamHI m! ,
b ecau se su b c lo n e s o f B x l(2 8 -1 ) w hich p o s s e s s HSV-1 BamHI q
la c i: t h i s fragm en t (F ig u r e C 4 . l l p a n e l C ). A summary o f th e
s tr u c t u r e s p r e s e n t in B x l ( 2 8 - l ) , and t h a t o f th e p u t a t iv e
o r ig in a l recom b in ant B x l ( 2 8 ) , a re i l l u s t r a t e d in F ig u re 0 4 .1 8 .
G ross s t r u c t u r e s o f B x l ( 2 8 - l ) s u b c lo n e 3
TWenty su b c lo n e s o f 8 x 1 (2 8 -1 ) were i s o l a t e d by Mrs
m. Murphy by th re e c y c le s o f p laq u e p u r i f i c a t i o n a t 31°>

- 134 -
and in v iv o ^2P - la b e l l e d DNA was a n a ly se d u s in g X b a l, Hindl l l ,
BcoRI, B g l l l , Hpal , Kpnl and BamH I. R e s u lt s f o r s e v e r a l o f th e
s u b c lo n e s w ith fo u r e n d o n u c le a se s a re shown in F ig u r e s C4«4 and
0 4 .3 . Genome s t r u c t u r e s deduced from t h i s g r o s s a n a ly s i s a re
shown in F ig u re 0 4 .6 .
More th an h a l f o f th e s u b c lo n e s a re f r e e i n L, and a l l
a re f r e e i n S . Those su b c lo n e s f ix e d i n L a re r e a d i ly
i d e n t i f i e d , a s i s B x l ( 2 8 - l ) , by th e overabundance o f th e two
j o in t fr a g m en ts HSY-2 EcoRI c and e o v e r th e o th e r tw o, b and
d, and by th e e x c e s s o f th e L te r m in a l fra g m en t HSV-2 Hpal g
o v er HSV-1 Hpal m (F ig u r e C 4 .4 ) . The o r i g i n a l c r o s s o v e r in
i s c o n ser v e d in a l l s u b c lo n e s , and a l l e x c e p t perhaps
su b c lo n e 27 have sym m etric S seg m en ts: t h a t i s , e i t h e r S
c o n s i s t s e n t i r e l y o f HSV-2 DNA se q u e n c e , o r i t h a s HSV-1 DNA•»
a t both S te r m in i w ith c r o s s o v e r s w ith in th e two HSV-1 BamHI m1
fra g m en ts (F ig u r e 0 4 .1 1 p a n e l C ). The d a ta s u g g e s t t h a t two
su b c lo n e s a re d e le t e d i n T R j/lR ^; su b c lo n a 14 w ith 3 x 10^
d a l to n s d e le t e d in ty p e 1 su b c lo n e 22 w itn 5 x 10
d a lto n s d e le t e d i n ty p e 2 IR^. R e s t r i c t i o n p r o f i l e s o f
th e s e two s u b c lo n e s show n o v e l bands in t e r p r e t e d a s j o in t and
te r m in a l fra g m en ts r e s u l t i n g from th e d e l e t i o n s . Only su b c lo n e
27 has i d e n t i c a l r e s t r i c t i o n p a t t e r n s to B x l ( 2 8 - 1 ) .
The a n a ly s i s a l s o s u g g e s t s t h a t some f r e e su b c lo n e s such
a s 1 0 , 11 and p erh ap s 9 have a d d it io n a l c r o s s o v e r s in TR o r
IRl . T h ere fo re th e j o in t and te r m in a l fra g m en ts o f c e r t a in
s u b c lo n e s were a n a ly se d in d e t a i l by c le a v a g e w ith S s t I and Smal .
S tr u c tu re s o f th e .jo in ts and te r m in i o f B x l(2 8 -1 ) s u b c lo n e s
F ig u r e s 0 4 .7 and C 4.0 show d a ta f o r j o in t r e g io n s ,
F ig u r e s C 4.3 and C 4.10 f o r 1 t e r m in i , and F ig u r e s C 4 . l l and
C 4.12 f o r 3 t e r m in i. To a la r g e e x t e n t o n ly th e m ajor genome

- 135 -
form s w ere a n a ly sed in 'these e x p e r im e n ts .
The m ajor j o in t o f su b c lo h e 1 , w hich i s f ix e d in L, i s
ty p e 1 th rou ghou t (F ig u r e C 4.7 p a n e l A tr a c k 3 , p a n e l C tr a c k
3 , F ig u re C 4.8 p a n e l A tr a c k 2 ) , in c lu d in g th e a seq u en ce from
w hich HSV-1 Smal G com igrated w ith Smal I . The m ajor L
term in u s i s typ e 2 , in c lu d in g th e a seq u en ce (F ig u r e 04*9
p a n e l A tr a c k 2 , F ig u re 0 4 .1 0 p a n e l A tr a c k 6 j . The m inor
ty p e 1 L term in u s i s b a r e ly d e t e c t a b le i n th e S s t I p r o f i l e
(F ig u r e 0 4 .9 p a n e l A tr a c k 3 ) , a s e x p e c te d s in c e L i s f i x e d .
The S term in u s i s typ e 1 (F ig u r e C 4«H p a n e l A tr a c k 3 ,
F ig u re 0 4 .1 2 p a n e l A tr a c k 5 ) . T h ere fo re su b c lo n e i s
h e t e r o ty p ic f o r TR and IRj,, in c lu d in g th e a s e q u e n c e s , w hereas
TRg and IRg a re id e n t i c a l ."
S u b clon e 5 i s a ls o f ix e d i n L and s im i la r i n s tr u c tu r e to
su b c lo n e 1 e x c e p t th a t th e c r o s s o v e r s i n TRg/lRg a r e i n HSV-1
Bamli l q Smal F r a th e r th an in HSV-1 BamHI mf » ( e .g . F ig u re
C 4.7 p a n e l A tr a c k 4 , F ig u re 0 4 .1 2 p a n e l A tr a c k 6 ) .
S u b clon e 9 i 3 f r e e i n L and h a s a more com plex s t r u c t u r e .
HSV-2 BamHI v , th e norm al ty p e 2 L te rm in u s , was n o t d e te c te d
in th e g r o s s a n a ly s i s (F ig u r e 0 4 .5 ) , s u g g e s t in g th a t b oth L
te r m in i a re typ e 1 a s in d ic a te d i n F ig u re 0 4 * 6 . In d ee d , t h i s
wan su p p orted by th e f in d in g t h a t th e m ajor j o in t o f norm al
s i z e i s type 1 th ro u g h o u t (F ig u re 0 4 .7 p a n e l A tr a c k 5 , p a n e li—>
C tr a c k 4 , F ig u re 0 4 .8 i^anel A tr a c k 3)> and t h a t th e L
term in u s w hich com igrated w ith HSV-1 BamHI s i s ty p e 1
(F ig u r e C 4.9 i)an el A tr a c k 6 , F ig u re 0 4 .10 . p a n e l A tr a c k 2 ) .
I t i s n o ted th a t HSV-1 Smal P i s a few bp s m a lle r i n t h i s
su b c lo n e than in tsB . Bands r e p r e s e n t a t iv e o f th e ty p e 2
L term in u s v/ere d e te c te d i n th e S s t I d i g e s t o f th e S term in u s
(F ig u r e 0 4 .1 1 p a n e l A tr a c k 5 ) . T h e r e fo r e , a lth o u g h th e
norm al typ e 2 L term in u s i s n o t p r e s e n t , t h i s seq u en ce i s

- 136 -
r e p r e s e n te d in a fragm en t w hich m ig ra ted a t a p o s i t i o n
0 .3 x 10^ d a lto n s l a r g e r . In o rd er t o c l a r i f y t h e s e r e s u l t s ,
th e L te r m in i were i s o l a t e d s e p a r a t e ly v i a th e u n iq u e Hpal
fra g m en ts and c le a v e d w ith S s t I (F ig u r e C 4.9 p a n e l B t r a c k s
3 and 4)» The BamHI L term in u s from HSV-1 Hpal ift i s ty p e 1
a s e x p e c te d , but t h a t from HSV-2 Hpal g c o n s i s t s o f a m ix tu re
o f two s p e c i e s . The f i r s t i s a norm al ty p e 1 BamHI s fra g m en t,
and th e secon d i s a ty p e 2 term in u s v /ith an a d d i t io n a l 0 .3 x 10
d a lto n s o f DNA a t th e te r m in u s . In d e e d , HSV-2 Hpal g o f
su b c lo n e 9 c o n s i s t s o f a fragm en t o f norm al s i z e and one a
l i t t l e la r g e r (F ig u r e 0 4 * 5 ) . The p r e c i s e n a tu r e o f th e la r g e r
fragm en t was n o t d e term in ed , but i t seem s from Smal a n a ly s i s
n o t to te r m in a te i n a ty p e 2 a s e q u e n c e . The m ost l i k e l y
e x p la n a t io n o f th e s e r e s u l t s i s t h a t su b c lo n e 9 c o n s i s t s o f a
m ix tu re o f two s t r u c t u r e s d i f f e r i n g a t one L te r m in u s , a s
i n d i c a t e d in F ig u re C 4 .1 7 . The ty p e 1 a seq u en ce i s a l i k e l y
c a n d id a te f o r th e a d d i t io n a l DNA a t th e ty p e 2 term in u s o f
su b c lo n e 9b , b ut t h i s was n o t p ro v en . The a c t u a l e x p la n a t io n
may be .more com p lex , but i t i s c l e a r t h a t f u r t h e r re c o m b in a tio n
h as o ccu rred a t th e 1 te r m in i o f th e su b c lo n e 9 genom e,
g e n e r a t in g r e g io n s o f t y p e - s p e c i f i c hom ology betw een TR and IR^.
S u b clon e 29 h as 'two* ty p e s o f L t e r m in i . One i s th e
norm al ty p e 2 term in u s and th e o th e r c o n t a in s HSV-1 S s t I B
but a s m a lle r te r m in a l fragm en t S s t I X (F ig u r e C 4.9 p a n e l A
tr a c k s 7 - 9 , F ig u r e C 4.10 p a n e l A tr a c k 3 J . T h is i s c o n s i s t e n t
w ith th e o b s e r v a t io n o f HSV-2 BamHI y and a s l i g h t l y s m a lle r
fragm en t in th e g r o s s BamHI p r o f i l e (F ig u r e C 4 .5 ) . F ig u re
C 4.10 p a n e l A tr a c k 4 shows t h a t th e s m a lle r term in u s c o n s i s t s
o f HSV-1 DNA e x c e p t t h a t th e HSV-1 a seq u en ce h as been
r e p la c e d by an HSV-2 a seq u en ce ( s e e a l s o F ig u re C 4.9 p a n e l
13 tr a c k s 3 -6 and F ig u r e C 4.13 p a n e l D t r a c k s 3 - 4 ) . ^ne gmai i e r

- 137 -
s i z e i s e x p la in e d by th e f a c t th a t th e HSV-2 a seq u en ce i s
a p p r o x im a te ly lpO bp s m a lle r than th a t o f HSV***l ( s e e S e c t io n 3 ) .
The c r o s s o v e r i s lo c a t e d in HSV-1 Smal P , w hich sp an s th e
b -a j u n c t io n , g e n e r a t in g a new fragm en t o f 28±2 bp a s shown
i n F ig u re 0 4 .1 8 . Both th e norm al ty p e 2 term in u s and th e
n o v e l one a r e r e f l e c t e d i n p r o f i l e s o f j o in t fra g m en ts
(F ig u r e C4*7 p a n e l A tr a c k 6 , p a n e l C t r a c k 5 , F ig u r e C4*8
p a n e l B t r a c k s 2 - 3 ) . The S term in u s i s ty p e 2 (F ig u r e C 4 .H
p a n e l A tr a c k 6 , F ig u re 0 4 .1 2 p a n e l A tr a c k 8 ) . S u b clon e 2 9 ,
w hich i s f r e e i n 1 , has t y p e - s p e c i f i c hom ology betw een
TRl and IR^ w h ich i s co n fin e d e s s e n t i a l l y to th e ty p e g a
se q u e n c e s .
F ig u re C 4.8 p a n e l C shows t h a t th e j o in t r e g io n o f
su b c lo n e 14 h a s HSV-1 a and c se q u e n c e s , but fr a g m e n ts ,
t o t a l l i n g o n ly 600 bp o f HSV-1 Hpal m a d ja c e n t to th e Hpal
s i t e i n th e b seq u en c e . I n te r v e n in g b seq u en ce fr a g m en ts a r e
a b s e n t . These r e s u l t s con firm th a t 3 x 10 d a lto n s o f DNA
h ave been d e le t e d from IR- a d ja c e n t to th e a s e q u e n c e . The
predom inant L term in u s i s " t y p e 2 (F ig u r e 04*9 p a n e l C t r a c k 3>
F ig u re C 4.10 p a n e l B tr a c k 4 ) , and th e S ter m in u s i s ty p e 1
(F ig u r e C 4 . l l p a n e l B t r a c k 4 , F ig u re C 4.12 p a n e l B t r a c k 4 ) .
S u b clon e 22 has a Hpal L term in u s o f a p p r o x im a te ly
1 .4 x 10^ d a lto n s (2400 b p ), w hich i s ab ou t 5 x 10^ d a lto n s
s m a lle r than th e e q u iv a le n t fragm ent in t s l . C lea v a g e o f
t h i s fragm en t w ith Smal produced fo u r d e te c te d fra g m en ts o f
1 0 0 0 , 8 5 0 , 300 and 55 bp. The o rd er o f t h e s e fra g m e n ts from
th e L term in u s i s 3 0 0 -1 0 0 0 -5 5 -8 5 0 bp, and was deduced a s f o l l o w s .
The 300 bp fragm en t co rresp o n d s to HSV-1 Smal G from th e norm al
HSV-1 1 term in u s o f su b c lo n e 22 (F ig u r e 0 4 .1 0 p a n e l C );
th e r e fo r e th e n o v e l term in u s has a 'type 1 and n o t a ty p e 2 a
se q u e n c e . The 850 bp fragm ent i s s im i la r in , s i z e to th e o n ly

- 138 -
Smal fragm en t from IISV-2 Hpal g w hich was n o t produced from
HSV-2 Hpal f (F ig u r e 0 4 .1 0 p a n e l D t r a c k s 5 - 6 ) , and t h e r e fo r e
sp an s th e TR^-U^ J u n c t io n . I t i s n o t e x a c t ly th e same s i z e
b ecau se TR^-U^ ju n c t io n fra g m en ts a re somewhat v a r ia b le in s i z e
i n in t e r t y p ic 'r e c o m b in a n t s . The 55 bp fra g m en t i s common to
HSV-1 Hpal f and g and t h e r e f o r e i s c o n ta in e d w ith in TRj/ I R j .
The 1000 bp fragm en t m ust span th e d e le t e d r e g io n . I t i s ~
co n c lu d ed t h a t su b c lo n e 22 c o n ta in s no more than 1 .2 x 10
d a lto n s o f HSV-2 TR^, th e d e l e t io n e x te n d in g o v e r a p p ro x im a te ly
5 x 1 0 6 d a lto n s w ith in TR^. H y b r id is a t io n d a ta shown in
F ig u re 0 4 .1 2 co n firm ed t h e “ i d e n t i t y o f n o v e l J o in t and
te r m in a l fra g m en ts in r e s t r i c t i o n p r o f i l e s o f su b c lo n e s 14 and
2 2 , and d em on stra ted t h e ir freedom from c o n ta m in a tio n by
n o n -d e le te d genom es.
S tr u c tu r e s o f th e . jo in t s and te r m in i o f o th e r reco m b in a n ts
P r e v io u s ly d eterm in ed genome s t r u c t u r e s a r e shown in
F ig u re 0 4 .3 o f s e v e r a l reco m b in a n ts w h ich a p p a r e n t ly a re
h e t e r o ty p ic f o r a l l or p a r t o f TRL and IR- QTlTRg a n d ,IR g ,
and w hich y e t in v e r t n o r m a lly . T hese w ere a n a ly se d f o r
u n d e te c te d c r o s s o v e r s .
The BamHI p r o f i l e o f B x l ( 3 l - 2 ) i s shown in F ig u re 0 4 * 1 4 .
The J o in t fra g m en t i s la r g e r th a n t h o s e o f th e p a r e n ta l v i r u s e s ,
and when c le a v e d w ith S s t I produced HSV-2 IRj, and HSV-1 IRg
fra g m en ts (F ig u r e 04*13 p a n e l A t r a c k 2 ) . The ty p e 2 1— ~ 6 term in u s c o r r e sp o n d in g to t h i s J o in t fragm en t i s 0*3 x 10
d a lto n s la r g e r th an HSV-2 BamHI v , and d id n o t produce th e
u s u a l ty p e 2 te r m in a l Smal X fragm en t (F ig u r e 0 4 .1 3 p a n e l A
tr a c k 3 , p a n e l B tr a c k 3 ) . The o th e r L term in u s ( t r a c k 5)
i s ty p e 1 , w ith a la r g e r Smal G fragm en t th a n tsB w hich i s
a l s o p r e s e n t a t th e ty p e 2 1 term in u s and a t th e J o in t
(p a n e l n tra ck s 2 , ; j , 5 ) . Tuese r e s u l t s s u g g e s t th a t one

- 139 -
L term in u s o f B x l(3 1 -2 ) i s ty p e 1 , and th a t th e o th e r i s ty p e 2
w ith th e a d d it io n o f a ty p e 1 a seq u en ce a t th e term in u s
a d ja c e n t to a ty p e 2 a se q u e n c e . I s o l a t i o n o f th e two L
te r m in i s e p a r a te ly and d ig e s t io n w ith S s t I showed a norm al
IISV-1 1 term in u s (F ig u r e 0 4 .1 3 p a n e l C tr a c k 4 ) , and a fragm en t
c o n t a in in g , among c o n ta m in a tin g fr a g m e n ts , HSV-2 S s t I 2, &&& i
hut a la r g e r S s t I te r m in a l fragm en t ( t r a c k 3 ) . I s o l a t i o n o f
th e two S s t I L te r m in a l fra g m en ts and d i g e s t i o n w ith Smal
showed th e norm al HSV-1 term in u s and one c o n s i s t in g o f th e
IISV-2 term in u s w ith th e a d d it io n o f an HSV-1 a seq u en ce
a d ja c e n t to th e HSV-2 a seq u en ce a t th e term in u s (F ig u r e 04*13
p a n e l D t r a c k s 3 - 4 ) . The J o in t fragm en t c o r r e sp o n d in g to th e
norm al ty p e 1 L term in u s was n o t a n a ly s e d , so i t i s shown w ith
a s in g l e a seq u en ce in F ig u r e C 4.17 a lth o u g h i t p ro b a b ly
c o n ta in s more than o n e , in v iew o f th e a b se n c e o f HSV-1 BamHI k
o f norm al s i z e i n F ig u re 04*14 p a n e l A# I t i s co n c lu d ed from
th e s e r e s u l t s th a t B x l ( 3 1 - 2 ) , w hich in v e r t s n o r m a lly ,, h a s
t y p o - s p e c i f i c hom ology betw een HSV-1 a se q u e n c es a t th e J o in t
and 1 te r m in u s .
3 x 6 ( 1 7 - 6 ) , RS6 and £ 1 3 -2 were a n a ly se d s im i l a r l y , and
d a ta a rc shown in F ig u r e s 04*14 and 04*15* The r e s t r i c t i o n s i t e
betw een HSV-2 S s t I 2 ajad i s a b se n t i n B x 6 (1 7 -6 ) , b u t S s t I 2
i s p r e s e n t . The c r o s s o v e r in IRj, o f R 13-2 i s lo c a t e d w ith in
HSV-1 Smal N.
D x l(2 ) was i n i t i a l l y th o u g h t to be h e t e r o t y p ic f o r
TKj/IR^ and T R g/lR g. The BamHI J o in t fragm en t i s 0 .3 x 10^
d a lto n s la r g e r than t h a t o f HSV-1 o r HSV-2 DNA, and Smal
d i g e s t i o n snows i t to c o n s i s t o f HSV-1 IR^ seq u e n c es a d ja c e n t
to HSV-2 IRg se q u e n c es w ith one in te r v e n in g a seq u en ce o f each
ty p e (F ig u r e 0 4 .1 6 p a n e l A tr a c k 3 , F ig u r e s 0 4 .1 7 and C 4 I1 8 ).
in o x e io r e a sequcx.ce hom ology betw een J o in t and te r m in i i s

- 140 -
m a in ta in ed f o r both L and S . A s im ila r s t r u c t u r e w ith th e a
se q u e n c e s r e v e r se d in o rd er was i d e n t i f i e d i n a su b c lo n e o f
su b c lo n e 2 o f B x l ( 2 8 - l ) (d e sc r ib e d below ; F ig u re C 4.16 p a n e l
A tr a c k 4 ) , and i s a ls o i l l u s t r a t e d in F ig u r e C 4*18. The b a s ic
s tr u c t u r e o f D x l(2 ) shown in F ig u re C4*17 i s f u r th e r c o m p lica te d
by th e p r e se n c e o f a p r o p o r tio n o f L te r m in i w ith an a d d i t io n a l
ty p e 2 a seq u en ce a t th e term in u s (F ig u r e 0 4 * 1 6 p a n e l C tr a c k
3 , p a n e l I) t r a c k 3 ) . T h is i s even fu r th e r c o m p lic a te d by
s e g r e g a t io n o f th e se q u e n c e s in TR g/lR g. For in s t a n c e i n F ig u r e
04*16 p a n e l B HSV-1 S s t I C and £ and HSV-2 S s t I £ and 6 a re
p r e s e n t in TRg and IRg, w hich were i s o l a t e d s e p a r a t e ly .
These f e a t u r e s o f th e L and S te r m in i a r e a l s o ap p a ren t in th e
d a ta o f F ig u re 04*13 p a n e l H tr a c k s 7 -1 0 , b u t have n o t been
in c lu d e d in F ig u re 04*17.
E x p r e ss io n o f IE 1 1 0 /1 1 8 by B x l(2 8 -1 ) s u b c lo n e s
To i n v e s t i g a t e th e e x p r e s s io n o f IB 1 1 0 /1 1 8 by su b c lo n e s
14 and 2 2 , th e genomes of. w hich a re d e le t e d in TR^ o r Ifi^ ,
in f e c t e d c e l l x^ olypeptid es were prepared from t h e s e , and”
o th e r reco m b in a n ts , and su b je c te d to p o ly a c ry la m id e g e l
e le c t r o p h o r e s i s by Hr II .S . Marsden and Mr G-. Hope.
Im m ediate e a r ly p o ly p e p t id e s were ind u ced i n HFL c e l l s
(a l i n e o f human f o e t a l lu n g c e l l s e s t a b l i s h e d by Hr B. C a r r i t t
in th e I n s t i t u t e o f G e n e t ic s , G lasgow) a s d e s c r ib e d by
P r e s to n e t a l , (1 9 7 8 ) . The recom b in an ts u se d produced
e q u iv a le n t p laq u e numbers on b oth BHK 013 and HFI c e l l s .
The a e in o d in v o lv e d i n f e c t i o n o f c e l l s a t h ig h m u l t i p l i c i t y
and in c u o a t io n fo r 3 hr a t 37° in th e p r e s e n c e o f
c y c lo h e x im id e . The drug was removed and p o ly p e p t id e s l a b e l l e dd 9w ith ^ b -m etn ion in e in th e p resen ce o f a c t in o m y c in H. Whole

- 141 -in f e c t e d c e l l p o ly p e p t id e s were p rep ared by l y s i s i n s i t u
o f th e c e l l s (M arsden e t a l . , 1 9 7 8 ) . S in c e IE 110 h as been
shown to p a r t i t i o n w ith th e n u c le u s ( P e r e ir a e t a l . , 1 9 7 7 ) , in
some e x p er im e n ts n u c le i were p rep ared by tre a tm e n t o f c e l l s
w ith 0 .6 5 $ NP40 in i c e - c o l d i s o t o n i c b u f f e r , b e fo r e b o i l in g4
i n d e n a tu r in g b u f fe r .
F ig u r e C 4 .2 0 , w hich i s a co m p o site o f S H S -p o lyacry lam id e
g e l s from s e v e r a l e x p e r im e n ts , show s im m ed iate e a r ly
p o ly p e p t id e s in d u ced by HSV-1 s t r a in 1 7 , HSV-2 s t r a i n HG52
$nd s e v e r a l reco m b in a n ts , th e genome s t r u c t u r e s o f w hich are
shown in F ig u re 0 4 .1 9 . L anes 1 -1 0 show w hole in f e c t e d c e l l '
p o ly p e p t id e s , and la n e s 1 1 -2 1 show n u c le a r p o ly p e p t id e s .
S ix HSV-1 coded (1 7 5 , 1 3 6 ' ( 1 4 3 ) , H O , 8 7 , 6 8 , 63) and s ix
HSV-2 coded (182~ 1 3 8 , 1 3 4 -1 3 2 , 1 1 8 , 6 7 , 6 4 ) IE p o ly p e p t id e s
can be i d e n t i f i e d i n t h e s e e x p e r im e n ts .
As o b serv ed p r e v io u s ly (P r e s to n e t a l . , 1 9 7 8 ) ,
reco m b in a n ts B x l(3 1 -2 ) and B x l ( 2 8 - 1 ) , w h ich a r e h e t e r o ty p ic f o r
th e r e p e a t s bounding UL, in d u ced b oth IE 110 (HSV-1) and th e
e q u iv a le n t HSV-2 p o ly p e p t id e IE 1 1 8 . S im i la r ly , su b c lo n e s 1
and 2 9 , w h ich a re h e t e r o ty p ic f o r TR and IR^, in d u ced both
IE 110 and IE 1 1 8 . The l a t t e r i n v e r t s n o rm a lly in L and S
b u t su b c lo n e 1 i s f ix e d , in L, and t h e r e fo r e w hether o r n o t th e
L segm ent in v e r t s n o rm a lly seem s n o t to a f f e c t e x p r e s s io n o f
th e IE 1 1 0 /1 1 8 g e n e s . S u b clon e 1 4 , w h ich la c k s HSV-1 seq u en ce
from IRT ( in c lu d in g a r e g io n from w hich th e 5 1 end o f th e mRNA
f o r IE 110 i s tr a n s c r ib e d ; F .J . R ixon , p e r s o n a l com m u n ica tion ),
in d u ced o n ly th e HSV-2 IE 1 1 8 . C o n v e r se ly , su b c lo n e 2 2 , w hich
la c k s HSV-2 seq u en ce from TR ( in c lu d in g th e w hole o f th e gene
f o r IE 118; A .J . E a sto n , p e r s o n a l co m m u n ica tio n ), in d u ced o n ly
th e HSV-1 IE 1 1 0 . S u b clon e 11 in d u ced IE 118 and a p o ly p e p t id e
w hich m igrate^ f a s t e r than IE 1 1 0 . F ig u r e 04*19 shows th a t a

c r o s s o v e r i s lo c a te d , w ith in th e DNA se q u e n c es c o d in g f o r th e
HSV-1 IE 110 icRNA. The a l t e r e d p o ly p e p t id e p ro b a b ly a r o se a s a
co n seq u en ce o f th e c r o s s o v e r p o s i t i o n i n IRj,. The a p p a r e n tly
s m a lle r s i z e may r e s u l t from n o n - c o l in e a r i t y o f i n i t i a t i o n or
te r m in a t io n cod on s f o r th e two polypeptides, a d e l e t io n o f some
c o d in g seq u en ce a s a r e s u l t o f th e c r o s s o v e r , o r from a l t e r e d
p r o c e s s in g o f th e h y b r id p o ly p e p t id e .
S t r u c tu r e s o f s u b c lo n e s o f B x l(2 8 -1 ) s u b c lo n e s
S ev en tee n su b c lo n e s each were i s o l a t e d by th r e e c y c l e s o f
p la q u e p u r i f i c a t i o n a t 31° from B x l(2 8 -1 ) s u b c lo n e s 2 and 29-,
in o rd er to d e term in e w h eth er a f ix e d su b c lo n e would g iv e r i s e
to f r e e p ro g en y , and w h eth er a f r e e su b c lo n e would g iv e r i s e
to f ix e d p ro g en y . The g r o s s s t r u c t u r e s o f th e progen y su b c lo n e s
were a n a ly se d u s in g Xbal , Hind l l l , EcoRI , B g ll l , Hpal , Kpnl and
BamH I, and a r e shown in F ig u re 0 4*21 .
S tr u c tu r e o f th e genome o f RE4
The r e s t r i c t i o n s i t e s in RE4 DNA were a n a ly se d i n th e
u s u a l way. H ind l l l and BamHI d i g e s t s a re shown i n F ig u r e
C 4.22 in t r a c k s l a b e l l e d R, The r e s u l t s o f t h e s e d i g e s t s and
o th e r s su g g e s te d t h a t th e KE4 genome i s d e le t e d in IR^ and th a t
a seq u en ce from e lse w h e r e h a s been in s e r t e d in p la c e Of IR^*
A l i k e l y e x p la n a t io n was th a t th e in s e r t e d DNA c o n t a in s
r e s t r i c t i o n s i t e s f o r H in d III and BamHI . th u s g e n e r a t in g
n o v e l fra g m en ts from one L term in u s ( i . e . th e f a i n t band
s l i g h t l y la r g e r th an HSV-2 H in d iII o , and th e band in te r m e d ia te
in s i z e betw een HSV-1 BamHI t and u v ) . The fragm en t s l i g h t l y
s m a lle r than HSV-1 H in d l l l o and th e fra g m en t in te r m e d ia te in
s i z e oetw een HSV-1 BamHI p and r may c o n ta in th e J u n ctio n
betw een norm al seq u en ce and the in s e r t e d seq u en ce
(fra g m en t T J ) . T hese fra g m en ts co u ld n o t be e x p la in e d a s

- 143 -r e s u l t i n g from norm al c r o s s o v e r s i n th e RE4 genome, and
n e i t h e r co u ld o th e r n o v e l fragm en ts th o u g h t to be J o in t
fra g m en ts r e s u l t i n g from th e n o v e l 1 te r m in u s , such a s th e
fragm en t w hich i s s l i g h t l y sm a lle r th an HSV-1 -BamHI k .
The p rop osed genome s tr u c tu r e o f RE4 i s i l l u s t r a t e d in
F ig u r e 0 4 .2 6 , and ex p erim en ts are d e sc r ib e d below i n su p p ort
o f t h i s u n u su a l arrangem ent.
The r e s u l t s o f ex p er im en ts in v o lv in g h y b r id is a t io n o f
HSV-2 H in d l l l n and o to n i t r o c e l l u l o s e s h e e t s c o n ta in in g
HSV-1, HSV-2 and RE4 Hindl l l o r BamHI d i g e s t s a r e shown in
F ig u re 0 4 .2 3 . HSV-2 Hind l l l n h y b r id is e d n o t o n ly a s
e x p e c te d to HSV-1 H in d l l l a p r e s e n t i n RE4 DNA, but a l s o to
th e n o v e l J o in t and te r m in a l fr a g m e n ts . HSV-2 H in d in o
h y b r id is e d i n each c a se to fragm en t TJ, th o u g h t to r e p r e s e n t
th e J u n ctio n betw een norm al and in s e r t e d s e q u e n c e s . The
i s o l a t e d RE4 H ind l l l TJ fragm en t h y b r id is e d to HSV-1
Hind l l l ab ed and BamHI b and f 1, and to HSV-2 H ind l l l b and o
and BamHI f , i and r . The h y b r id is a t io n r e s u l t s c e r t a i n l y
ca n n o t be e x p la in e d in term s o f l e g i t im a t e c r o s s o v e r s , b ecau se
RE4 H ind l l l TJ c o n ta in s seq u en ces from HSV-2 H in d l l l o
(HSV-2 BamHI f ) and IISV-2 H in d iII b (HSV-2 BamHI i r ) , w hich a re
n o t c o n t ig u o u s in th e HSV-2 genome. T hese e x p e r im e n ts
su p p ort th e h y p o th e s is th a t RE4 DNA i s d e le t e d in a t l e a s t
th e m a jo r ity o f IR^, w ith th e in s e r t io n o f an HSV-2 seq u en ce
from a s shown i n F ig u re 0 4 .2 6 .
To l o c a t e more p r e c i s e ly th e J u n ctio n betw een norm al and
in s e r t e d DNA th e BamHI and BamH I/H in d III TJ fra g m en ts were
i s o l a t e d and c le a v e d w ith a number o f r e s t r i c t i o n e n d o n u c le a se s
(F ig u r e 0 4 .2 4 ) . The r e s u l t s show t h a t Hind l l l TJ c o n ta in s
a lm o s t a l l o f HSV-2 Hind l l l o , w ith th e e x c e p t io n o f n o t more
than 25 bp a t the r ig h t hand end ( s e e Hhal d i g e s t s ) . R e s u lt s
w ith Hhal and A lu l show th a t n o t more than 260 bp o f HSV-2

- 144 -
BamHI i a re a d ja c e n t to HSV-2 H in d l l l o s e q u e n c e s . The r e s u l t s
o f t h i s exp erim en t have been in c lu d e d in F ig u r e C 4 .2 6 .
C lea v a g e o f th e RE4 BamHI n o v e l j o in t fragm en t
d em o n stra tes th e p r e se n c e o f HSV-2 IRg fra g m en ts b u t th e
a b sen c e o f IR- fra g m en ts o f e i t h e r HSV-1 o r HSV-2 (F ig u r e C 4.25
p a n e l A t r a c k '2 , p a n e l C t r a c k 2 , p a n e l D tr a c k 2 ) . I n s t e a d ,
Smal fra g m en ts c h a r a c t e r i s t i c o f HSV-2 BamHI r w ere produced
(p a n e l C t r a c k 2 ) . HSV-2 Smal £ aM 12 p r e s e n t , s u g g e s t in g
a ty p e 2 a seq u en ce a t t h e " j o in t ; th e a b sen ce o f HSV-2 Smal 10
shows t h a t n o t more th an a p p ro x im a te ly 45 bp o f th e HSV-2
b seq u en ce a r e a d ja c e n t to th e a se q u e n c e . These r e s u l t s and
th o s e from th e n o v e l L term in u s (p a n e l C t r a c k s 5 -6 ) show
th a t th e n o v e l L term in u s co m p rises in s e r t e d HSV-2 seq u en ce
w ith a te r m in a l ty p e 2 a seq u e n ce , and t h a t th e n o v e l j o i n t
c o m p r ises th e in s e r t e d seq u en ce jo in e d to th e HSV-2 c seq u en ce
v i a a ty p e 2 a se q u e n c e . The two L te r m in i o f RE4 a re
h e t e r o t y p ic , in c lu d in g th e a seq uence (com pare n o v e l term in u s
w ith p a n e l E t r a c k l ) . T h is i s tr u e a l s o o f th e S t e r m in i ,
th e ty p e 1 term in u s b e in g shown i n p a n e l F t r a c k 2 . A
fu r th e r c o m p le x ity in th e genome s tr u c tu r e o f RE4 i s t h a t th e
BamHI ty p e 1 S term in u s , i s o l a t e d from u n iq u e fr a g m e n ts , a l s o
c o n ta in s ty p e 2 S te r m in a l fra g m en ts (p a n e l B t r a c k s 2 - 3 ) .
T h is s u g g e s t s t h a t , l i k e B x l ( 2 8 - 1 ) , a t l e a s t th e ty p e 1 S
term in u s o f RB4 DBA c o n s i s t s o f a m ix tu re o f s t r u c t u r e s
presum ably g en e r a te d by secon d ary r ec o m b in a tio n in v o lv in g th e
r e p e t i t i v e r e g io n s .
R e s t r ic t io n p r o f i l e s o f RE4 and th e r e c le a v a g e ex p er im en ts .
u e s c r ib e d above s u g g e s t th a t th e RE4 genome i s f ix e d i n I and S
in th e Ig arrangem ent (F ig u re C 4 .2 6 ) . For exam ple ,
a u to d e n s ito m e tr ic sca n n in g o f F ig u re C 4.22 t r a c k s R r e v e a le d
th a t HSV-1 H in d l l l m, c o n ta in in g th e ty p e 1 S te r m in u s , i s in

- 145 -
t h r e e - to f o u r - f o ld e x c e s s o v er HSV-1 H in d l l l g , w hich c o n ta in s
th e ty p e 2 S te r m in u s . Two d i s c e r n ib le j o in t fr a g m e n ts , th e
secon d and th ir d bands i n th e Hind l l l p r o f i l e , were un d er
r e p r e s e n te d . The ty p e 1 L term in u s HSV-1 BamHI s was
predom inant o v er th e f a i n t n o v e l term in u s w hich m ig ra ted below
HSV-1 BamHI t , and th e ty p e 1 S term in u s HSV-1 BamHI q was
predom inant o v er th e f a i n t ty p e 2 S term in u s w hich m ig ra ted below
i t . The norm al ty p e 1 j o i n t HSV-1 BamHI k was f a i n t compared
w ith th e n o v e l j o in t o f f a s t e r m o b i l i t y .
The RE4 genome la c k s 10*5 kbp o f DNA in c lu d in g IR^ andJ "I
a t l e a s t 300 bp o f a d ja c e n t seq u e n ce , and h a s an i n s e r t i o n
o f 2 .8 kbp o f HSV-2 DNA a t t h i s l o c a t io n from th e BamHI i - r
( H in d l l l b -n ) r e g io n . M oreover th e r e i s co m p le te t y p e - s p e c i f i c
h e t e r o lo g y betw een IRg and TRg and betw een TR^ and th e j o in t
r e g io n , and th e genome i s f ix e d i n L and S in " th e Ig arran gem en t.
S ev e n te en su b c lo n e s o f RE4 were i s o l a t e d b y 'th r e e c y c l e s o f
p la q u e - p u r i f ic a t io n a t 3 1 ° , and t h e ir genome s t r u c t u r e s a n a ly s e d .
H in d l l l and BamHI p r o f i l e s o f s e v e r a l s u b c lo n e s a re shown in
F ig u re C 4 .2 2 . A l l su b c lo n e s r e t a in th e d e l e t io n and i n s e r t i o n ,
and th e c r o s s o v e r s in and Ug p r e se n t i n RE4. E ig h t o f th e
s u b c lo n e s had r e s t r i c t i o n p r o f i l e s i d e n t i c a l t o th a t o f RE4,
but some more c l e a r l y showed th e f ix e d n a tu re o f L and S
( e . g . su b c lo n c 6 ) , im p ly in g t h a t RE4 i s n o t a pure s t r u c t u r e .
The o th e r n in e su b c lo n e s were p laq u e p u r i f ie d tw id e more
b eca u se s e v e r a l seemed s t i l l to com p rise m ix tu r e s o f genome
s t r u c t u r e s a f t e r th e f i r s t th r e e p laq u e p u r i f i c a t i o n s .
The p r o f i l e s o f m ost o f t h e s e su b c lo n e s are in c lu d e d i n F ig u re
C 4 .2 2 . They a re f r e e in S , w ith th e p o s s ib le e x c e p t io n o f
su b c lo n e 3 , and m ost a re f r e e a ls o in 1 w ith th e e x c e p t io n s o f
17 and p erh ap s 3 , 4 and 5* The d i s t i n c t i o n betw een f ix e d and
f r e e , how ever, i s n o t c l e a r in every c a s e and i t i s l i k e l y t h a t

- 146 -
some s u b c lo n e s may n o t be pure even a f t e r f i v e rou n d s o f p laq u e
p u r i f i c a t i o n . The n in e su b c lo n e s have seco n d a ry c r o s s o v e r s in
r e p e t i t i v e r e g io n s w hich were n o t a n a ly se d f u r t h e r . For exam ple,
s u b c lo n e s 1 , 5 , 7 , 1 5 , 16 and 17 each have a c r o s s o v e r i n HSV-2
BamHI g ’ , r e s u l t i n g in a s m a lle r fra g m en t, w h ich s u g g e s t s th a t
b oth S te r m in i i n t h e s e su b c lo n e s a re ty p e 1 . S u b clon e 1
p o s s ib ly h a s a c r o s s o v e r i n ty p e 1 TR^, s in c e i t . la c k s HSV-1
BamHI s and in s t e a d h a s a te r m in a l fragm en t la r g e r th an HSV-1
BamHI o , a s w e l l a s two o th e r fra g m en ts r e s u l t i n g from th e
la r g e r L te r m in u s . S u b c lo n es 7 , 1 2 , 15 and 16 have a BamHI .
j o in t fragm en t 0 .3 x 10^ la r g e r in m .w t. th an t h a t o f RE4,
p ro b a b ly r e s u l t i n g from a n .e x tr a a seq u en ce in th e j o in t
fragm en t c o n ta in in g in s e r t e d DNA. I t i s co n c lu d ed from t h e s e
i n i t i a l s t u d ie s t h a t in 'R E 4 , a s in B x l ( 2 8 - l ) , th e c o n v e r s io n
o f th e f ix e d p h en otyp e to th e f r e e o c c u r s by a d d i t io n a l
re co m b in a tio n e v e n ts i n r e p e t i t i v e r e g io n s w hich g e n e r a te
t y p e - s p e c i f i c hom ology betw een th e s e r e g ib n s * ., D e t a i le d
r e s t r i c t i o n a n a ly s i s o f th e genomes o f RE4 s u b c lo n e s i s
n e c e s s a r y to more c l e a r l y d e f in e th e secon d ary rec o m b in a tio n
e v e n t s .

- 147 -
DISCUSSION
F ix ed and f r e e reco m b in a n ts
B x l (2 8 -1 ) c o n ta in s a m ixtu re o f a t l e a s t two genome
s t r u c t u r e s d i f f e r i n g i n TR g/lR g. T hese s t r u c t u r e s , and th e
f a c t th a t th e recom b in ant was i n i t i a l l y i s o l a t e d by s i x c y c l e s
o f p la q u e p u r i f i c a t i o n , s u g g e s t th a t th e o r i g i n a l r e c o m b in a tio n
e v e n t produced th e recom b in ant genome s tr u c tu r e shown i n
F ig u re C4. 18. F u rth er in te r m o le c u la r r e c o m b in a tio n p ro b a b ly
th en produced th e s t r u c t u r e s p r e s e n t i n B x l ( 2 8 - 1 ) . I n t r a - or
in te r -m o le c u la r r ec o m b in a tio n e v e n ts c o u ld th en have g iv e n r i s e
to th e s u b c lo n e s . The s t r u c t u r e s p r e s e n t in su b c lo n e 27
presum ably a r o se by s u r v iv a l o f th e i n i t i a l recom b in an t B x l(2 8 )
d u r in g p la q u e p u r i f i c a t i o n , in s im ila r f a s h io n t o th e i s o l a t i o n
o f B x l( 2 8 - 1 ) •
The s u b c lo n e s o f B x l(2 8 -1 ) f a l l in t o two c l a s s e s , th o s e
w hich a re f i x e d in L and th o se in w hich L in v e r t s n o r m a lly .
Each f ix e d su b c lo n e had a fo u r fo ld e x c e s s o f one o r i e n t a t io n o f
L o v er th e o t h e r , and a s im i la r r a t io was o b serv ed f o r L and S
i n RE4. I t i s im p o rta n t to n o te t h a t i n no c a s e was in v e r s io n
o f a segm ent e n t i r e l y a b s e n t , and i t seem s from th e a n a ly s i s
t h a t r e s id u a l in v e r s io n i s a p ro p er ty o f th e f ix e d genome
r a th e r than a r e s u l t o f th e p r e sen ce o f co n ta m in a n ts w ith
seco n d a ry c r o s s o v e r s . The f ix e d p h en otyp e may r e p r e s e n t e i t h e r
p r e f e r e n t i a l r e p l i c a t io n or p r e f e r e n t ia l p a ck a g in g in t o v i r i o n s
o f DNA w ith one o r ie n t a t io n o f L. A n a ly s is o f n u c le a r DNA d id
n o t su p p ort th e l a t t e r (d a ta n o t show n). A l t e r n a t iv e ly , f i x e d
reco m b in a n ts co u ld be th e r e s u l t o f a b ia se d mechanism f o r
m a tu ra tio n o f u n i t le n g t h DNA from c o n c a te m e r s .
The s t r u c t u r e s o f B x l(2 8 -1 ) s u b c lo n e s and o th e r
reco m b in a n ts show t h a t norm al in v e r s io n o f L depends on a
r e g io n o f t y p e - s p e c i f i c hom ology betw een TR^ and IR^. A l l

- 148 -
n o rm a lly in v e r t in g su b c lo n e s p o s s e s s seco n d a ry c r o s s o v e r s i n
TR^ or IR- w hich g iv e r i s e to such hom ology. S u b clo n e 2 9 ,
B x l ( 3 1 - 2 ) 'an d perhaps su b c lo n e 9b show t h a t hom ology betw een th e
a seq u e n c es i s s u f f i c i e n t to a llo w norm al in v e r s io n . I t h as n o t
been d em o n stra ted , how ever, th a t a seq u en ce hom ology i s
n e c e s s a r y f o r norm al in v e r s io n , a s a recom b in an t w ith
hom ologous r e g io n s o f TR^ and IR^ but h e te r o lo g o u s a seq u e n c es
was n o t o b se r v e d . A n a ly s is o f su b c lo n e s o f th e s u b c lo n e s
su p p o r ts th e h y p o th e s is o f hom ology req u irem en ts a f ix e d form
(h e te r o lo g o u s ) gave r i s e to some n o rm a lly in v e r t in g p rogen y
(h o m o lo g o u s), but th e r e v e r s e c o n v e r s io n vras n o t o b se r v e d .
T h is i s e x p l ic a b le in th a t c r o s s o v e r s i n th e f ix e d form can
g e n e r a te hom ology betw een TR and IR^, and t h e s e w ere in d e ed
d e t e c t e d , w hereas a n o r m a lly " in v e r t ih g form ca n n o t g e n e r a te
h e t e r o lo g y s in c e i t d o e s n o t c o n ta in th e n e c e s s a r y s e q u e n c e s .
RE4, i t s u n u su al genome s tr u c tu r e a s id e , i s a n a lo g o u s to
B x l(2 8 -1 ) b ecau se i t i s f ix e d i n L and S i n th e I g o r i e n t a t io n .
I n i t i a l s t u d ie s o f th e s tr u c t u r e s o f RE4 and i t s su b c lo n e s
s u g g e s t t h a t th e hom ology req u irem en t a p p l ie s to b o th L and S ,
s in c e n o rm a lly in v e r t in g progeny p o s s e s s seco n d a ry c r o s s o v e r s
i n T R g/lR g. The s t r u c t u r e s o f reco m b in a n ts w ith two a d ja c e n t
h e t e r o t y p ic a seq u en ces a^ th e j o in t in d ic a t e e i t h e r t h a t
th e q seq u en ce i s c r u c ia l i n segm ent in v e r s io n o r t h a t such an
arrangem ent i s f o r t u i t o u s l y m a in ta in ed . I t i s i n t e r e s t i n g
to n o te th a t th e L term in u s o f D x l(2 ) had a s in g l e ty p e 1
a seq u en ce in about h a l f th e m o le c u le s and an a d d i t io n a l ty p e 2
a seq u en ce in th e o th e r h a l f (o n ly th e form er i s shown in
i i g u r e 0 4 * 1 7 ) .
I t i s proposed from th e se r e s u l t s th a t in v e r s io n o f th e
1 and S segm en ts o f th e HSV genome i s ca u sed by a s i t e - s p e c i f i c
r ec o m b in a tio n ev en t i n , or n e a r , th e a s e q u e n c e . T h is

- 149 -
h y p o th e s is h a s been s tr e n g th e n e d by th e o b s e r v a t io n t h a t
i n s e r t i o n o f a 4 .2 kbp fragm en t c o n ta in in g th e HSV-1 a seq u en ce
in t o th e HSV-1 dPyK gen e ca u sed a d d i t io n a l in v e r s io n s ab ou t
th e n o v e l j o i n t (M ocarsk i e t a l . , 1 980; J . S m ile y , p e r s o n a l
co m m u n ica tio n ). I n s e r t io n o f a fragm en t o f s im i la r s i z e from
e lse w h e r e i n th e genome ca u sed no a d d i t io n a l in v e r s io n s
(M ocarsk i e t a l . , 1 9 8 0 ) .
M odels f o r segm ent in v e r s io n
M odels p r e v io u s ly p rop osed f o r th e m echanism o f in v e r s io n
in v o lv e i n t e r - o r i n t r a - t y p i c r e c o m b in a tio n , and a r e i l l u s t r a t e d
i n F ig u re C 4*27. M odel 1 , w h ich p r o c e e d s v ia p h y s ic a l
s e p a r a t io n o f L and S and t h e i r su b seq u en t r e j o in in g t o g iv e
th e fo u r a rra n g em en ts , h a s n o t been su p p o rted e le c t r o n
m ic r o sc o p ic o r g e n e t ic s t u d ie s (Jacob e t a l . , 1 9 7 9 ) , a lth o u g h
th e r e j o in in g would be s p e c i f i c f o r th e a seq u e n o e . S im i la r ly
m odel 2 , w h ich p r o p o se s t h a t in v e r s io n o c c u r s by in te r m o le c u la r
r e c o m b in a tio n , co u ld in v o lv e a s i t e - s p e c i f i c e v e n t , b u t h a s n o t
th e su p p o rt o f g e n e t ic s t u d ie s (H oness e t a l . , 1 9 8 0 ) . In
m odel in v e r s io n o c c u r s by DNA s y n t h e s i s w ith c o n c u r r e n t
in tr a m o le c u la r r e c o m b in a tio n , but d o e s n o t f i t in w ith th e
c u r r e n t ly a c c e p te d m odel f o r DNA r e p l i c a t i o n . I n tr a m o le c u la ra *
r e co m b in a tio n betw een in v e r t e d r e p e a t s i n l i n e a r o r c i r c u l a r
form (m odel 3B) was o r i g i n a l l y p rop osed a s a n o n - s p e c i f i c
e v e n t , but i n th e l i g h t o f th e above r e s u l t s t h i s i s u n l ik e ly
to be th e c a s e . However, th e m odel can e q u a l ly w e l l
accommodate s i t e - s p e c i f i c re co m b in a tio n a t th e a s e q u e n c e s .
M odel 30 , i s e x p la in e d i n th e le g e n d to F ig u re C 4 .2 7 .
I t h as th e s t r e n g t h s th a t i t accom m odates th e c u r r e n t schem e
f o r DNA r e p l i c a t i o n , and th a t in v e r s io n can be in t e r p r e t e d i n
term s o f a s i t e - s p e c i f i c e v e n t a t th e a s e q u e n c e . One

- 150 -
p r e d ic t io n o f th e m odel i s th a t a l l th e a s e q u e n c e s , and p erh ap s
a d ja c e n t r e g io n s o f th e b and c se q u e n c e s , a re o b l i g a t o r i l y
i d e n t i c a l w ith in a r e p l ic a t e d DNA m o le c u le . T h is f e a tu r e
a r o se b eca u se o f th e r a th e r o v e rr a ted h y p o th e s is t h a t n o t a l l
fo u r genome arrangem ents p a r t ic ip a t e in r e c o m b in a tio n o r
r e p l i c a t i o n , and t h e r e fo r e t h a t g e n e r a t io n o f th e fo u r
arran gem en ts must in v o lv e an o b l ig a to r y e v e n t . T h is h y p o th e s is
i s d is c u s s e d in S e c t io n 2 . B en -P orat e t a l . (1 9 8 0 ) have
r e c e n t ly shown t h a t both arran gem en ts o f th e PRV genome e n te r
t h e n u c le u s , c i r c u l a r i s e , r e p l i c a t e , in v e r t and a re m atured .
Knipe e t a l . (1978) a ttem p ted to make a c a s e f o r o b l ig a t o r y
i d e n t i t y from th e o b s e r v a t io n t h a t i n t e r t y p i c reco m b in a n ts
p o s s e s s hom otypic seq u e n ces in th e a , and p erh a p s a d ja c e n t
r e g io n s o f th e c , seq u en ces bounding Ug . T h is f e a t u r e was
common i n th e reco m b in a n ts d e sc r ib e d i n S e c t io n 2 and i n t h i s
S e c t io n . A l e s s a ssu m p tiv e c o n c lu s io n i s t h a t th e ten d en cy f o r
i d e n t i t y o f t h e s e r e g io n s i s th e r e s u l t o f f a c i l e r e c o m b in a tio n
w ith in r e p e a te d r e g io n s , r a th e r th an th e r e s u l t o f an o b l ig a t o r y
e v e n t i n DNA s y n t h e s i s . A second p ie c e o f e v id e n c e stem s from
th e c la im e d mapping o f an HSV-1 t s n u ta t io n ( t s c 7 5 ) in ea ch a
se q u e n c e . P a r e n t h e t ic a l ly , t h i s d o es n o t d i c t a t e t h a t a
se q u e n c es m ust be o b l i g a t o r i l y i d e n t i c a l , s in c e th e i n i t i a l
m u ta tio n in one a seq u en ce co u ld have been t r a n s fe r r e d to th e
o th e r a seq u e n ces by f a c i l e reco m b in a tio n a t an e a r ly s t a g e .
R e g a r d le s s o f t h i s , P r e s to n ( in p r e s s ) has shown t h a t t h i s
l e s i o n maps w ith in th e Vmw IE 175 gene some 2 kbp away from th e
a seq u en ce i n th e c se q u e n c e . There i s t h e r e fo r e no c o n c lu s iv e
e v id e n c e f o r o b l ig a to r y i d e n t i t y o f th e a s e q u e n c e s . The
o b s e r v a t io n t h a t th e s i z e d i s t r ib u t io n p a t te r n o f th e HSV-1 a
se q u e n c es i s th e same a t th e j o in t and both te r m in i ( S e c t io n 3
o f R e s u lt s ) cou ld a g a in be e x p la in e d by f a c u l t a t i v e i d e n t i t y o f

- 151 -
'these r e g io n s . I*t i s c l e a r from th e work p r e se n te d h ere t h a t
some reco m b in a n ts a re a b le to r e p l i c a t e w ith o u t a req u irem en t
f o r o b l ig a t o r y i d e n t i t y o f a seq u e n c e s ( e . g . su b c lo n e 1 o f
B x l(2 8 -1 ) and RE4). T h ere fo re e i t h e r t h i s i 3 n o t th e
m echanism o f in v e r s io n , or th e s e reco m b in a n ts r e p l i c a t e by an
a l t e r n a t i v e m echanism . The number o f modes o f HSV DNA
s y n t h e s is i s unknown.
The s ta g e a t w hich in v e r s io n o c c u r s d u r in g v ir u s grow th
h as n o t been i d e n t i f i e d , a lth o u g h i s o m e r is a t io n p r o c e e d s to
e q u ilib r iu m a t an e a r ly s ta g e i n p la q u e fo r m a tio n (B en -P o ra t
e t aJL., 1 9 8 0 ) . The e v e n t c o u ld ta k e p la c a e s s e n t i a l l y b e fo r e
th e p r o d u c t io n o f co n c a tem e r s , o r co u ld be p o s t - r e p l i c a t i v e a s
in m odel 3C. I t i s p o s s ib l e t h a t in v e r s io n i s n o t o n ly a
r e s u l t o f r e c o m b in a tio n , but a l s o o f th e mode o f c l e a v in g
u n i t - l e n g t h m o le c u le s from c o n c a te m e r s . I f HSV co n ca tem er s
are produced from a c i r c u l a r in te r m e d ia t e , t h e i r s t r u c t u r e
in th e j o i n t r e g io n s would be d eterm in ed by th e s t r u c t u r e o f th e
j o in t r e g io n s o f th e c i r c u l a r m o le c u le . T h ere fo re th e
mechanism by w hich l i n e a r in p u t DNA c i r c u l a r i s e s would have
a d i r e c t e f f e c t upon con catem er j o i n t s t r u c t u r e .
Mech an ism s f o r genome c i r c u l a r i s a t i o n and m a tu ra tio n
There i s s tr o n g e v id e n c e from e le c t r o n m icro sco p y t h a t
HSV and PRV DNA r e p l i c a t i o n in v o lv e s c o n c a te m e r ic , and p erh a p s
c i r c u l a r , in t e r m e d ia t e s . Jean and B en -P orat (1 9 7 6 ) o b serv ed
l i n e a r PRV DNA m o le c u le s w ith s in g l e s tra n d ed te r m in i and u n i t -
le n g th c i r c l e s in DNA from in f e c t e d c e l l n u c le i i n th e a b se n c e
o f de novo p r o t e in s y n t h e s i s , and p rop osed th a t ex p o su re o f
com plem entary s in g l e stra n d ed te r m in i by an e x o n u c le a se p r e c e d e s
c i r c u l a r i s a t i o n . D e s p ite a s tr o n g b e l i e f in th e te r m in a l
redundancy o f PRV DNA, B en -P orat e t a l . (1 9 7 9 ) w ere u n a b le to
d em o n stra te a te r m in a l r e p e t i t i o n by e x o n u c le a se tr e a tm e n t and

- 152 -
a n n e a lin g i n v i t r o . N e v e r t h e le s s , i t i s p ro b a b le t h a t PRV DNA
i s c i r c u la r a t l e a s t d u r in g th e i n i t i a l s t a g e s o f DNA
r e p l i c a t io n (B en -P o ra t and Veach, 1 9 8 0 ) . On th e o th e r hand,
a lth o u g h HSV-1 DNA i s w ith o u t doubt t e r m in a l ly red u n d a n t, i t
h as n o t been shown th a t c i r c u l a r i s a t i o n i n v iv o o c c u r s by
a n n e a lin g o f exp osed s in g l e stra n d ed t e r m in i . The m a jo r ity o f
B x l (2 8 -1 - 1 ) DNA h as a ty p e 1 a seq u en ce a t th e S term in u s and a
’type 2 a seq u en ce a t th e 1 term in u s, and w ould be u n l ik e ly to
c i r c u l a r i s e by t h i s m echanism s in c e th e HSV-1 and HSV-2 a
seq u e n c e s a r e n o t com plem entary ( S e c t io n .3 o f R e s u l t s ) .
T h e r e fo r e , i f r e p l i c a t i o n p ro ceed s v i a a c i r c u l a r in te r m e d ia t e ,
e i t h e r c i r c u l a r i s a t i o n o c c u r s by an a l t e r n a t i v e m echanism , such
a s b lu n t-e n d l i g a t i o n , o r B x l(2 8 -1 -1 ) DNA c o n t a in s a m inor
p o p u la t io n o f m o le cu le s- w ith hom otypic t e r m in i ( e . g . an e x tr a
ty p e 1 a seq u en ce a t th e ty p e 2 1 term in u s) w hich a lo n e
r e p l i c a t e t o produce f o r unknown r e a s o n s a la r g e p r o p o r t io n o f
n o n - r e p l i c a t in g p rogen y w ith h e te r o ty p ic te r m in i ( i . e . th e
m ajor form o f B x l ( 2 8 - l - l ) genom e). The b i o l o g i c a l p r o p e r t ie s
o f t h i s recom b in an t have n o t been e lu c id a t e d , and o n ly th e
m ajor genome s tr u c t u r e was a n a ly se d , so i t i s n o t a t p r e s e n t
p o s s ib l e to d i s t i n g u i s h betw een th e s e a l t e r n a t i v e s .
The two form s o f c i r c u la r HSV-1 DNA w h ich would be
produced by com plem entary-end and b lu n t-e n d l i g a t i o n , and th e
co n ca tem er s g e n e r a te d th e re fro m , a r e shown i n F ig u r e C 4*28.
I t i s n o te d t h a t b lu n t-e n d l i g a t i o n p ro d u ces co n ca tem er s i n
w h ich a l t e r n a t e j o i n t s may be d i s t in g u is h e d by a c le a v a g e p r o c e s s
to produce u n i t - l e n g t h l i n e a r m o le c u le s . I t i s n e c e s s a r y f o r
b o th segm en ts to in v e r t in d e p e n d e n tly to p rod u ce th e fo u r
a rra n g em en ts . T h is f e a tu r e i s in c lu d e d in m odel 3C i n F ig u r e
C 4 .2 7 , o n th e in c o r r e c t assu m ption th a t th e HSV-1 genome has
two a se q u e n c e s and would th e r e fo r e d i s p la y two d i s s i m i l a r

- 153 -
j o in t s t r u c t u r e s a f t e r corapXementary-end. l i g a t i o n . As shown i n" # T
S e c t io n 3 o f R e s u l t s , th e m a jo r ity o f HSV-1 DNA m o le c u le s
c o n ta in a s i n g l e a seq u en ce a t th e j o in t and t e r m in i , and by
com plem entary-end l i g a t i o n would produce co n ca tem ers w ith
i d e n t i c a l j o i n t s a t a l l l o c a t io n s (F ig u r e C 4 .2 8 ) . C lea v a g e o fr s
u n i t - l e n g t h m o le c u le s from su ch co n ca tem ers would th en
n e c e s s i t a t e a secon d c r i t e r i o n , such a s an ap p rox im ate le n g t h
m easurem ent. The i n i t i a l c h o ic e o f c le a v a g e s i t e i n t h i s c a s e
may be a f a c t o r in in v e r s io n , a s e x p la in e d i n F ig u r e C 4*28.
Jacob e t a l . (1 9 7 9 ) commented t h a t c le a v a g e must be s e p a r a te
from e n c a p s id a t io n i n o rd er to a l lo w p rogen y DNA to e n te r th e
r e p l i c a t i v e p o o l* t h i s would n o t be th e c a s e i f c i r c u la r
monomers a r e r e a d i ly produced from co n ca tem ers by r e c o m b in a tio n .
L adin e t a l . (1 9 8 0 ) have shown t h a t m a tu ra tio n o f PRV DNA i s
d ep en d en t upon o a p s id a sse m b ly . The n a tu r e o f th e c le a v a g e
e v e n t , w h eth er i t i s a d o u b le s tra n d e d c u t o r a s in g l e s tr a n d ed
n ic k a t ea ch end o f th e a seq u en ce fo llo w e d by r e p a ir o f s in g l e
stra n d ed DNA, i s unknown.
F ix ed reco m b in a n ts a re r a r e . In d eed , th e marker r e s c u e
ex p e r im en ts d e s o r ib e d i n S e c t io n 2 w ere aim ed a t g e n e r a t in g
su ch reco m b in a n ts and y e t f a i l e d to do s o . The f ix e d
reco m b in a n ts s tu d ie d r a i s e a number o f q u e s t io n s r e g a r d in g
IISV DNA r e p l i c a t i o n , such a s th e mechanism by w hich su b c lo n e
1 o f B x l ( 2 8 - l ) , t h a t i s B x l( 2 8 - 1 - 1 ) , c i r c u l a r i s e s . More
p r o b le m a t ic a l i s th e way i n w hich th e DNA o f t h i s reco m b in a n t
i s m atu red . W ith r e s p e c t to a se q u e n c e s , c i r c u l a r o r
c o n c a te m e r ic B x l( 2 8 -1 - 1 ) DNA i s s t r u c t u r a l l y i d e n t i c a l to
B x l(3 1 -2 ) DNA, and y e t th e fo rm er h as a f ix e d p h en otyp e and
th e l a t t e r i n v e r t s n o r m a lly . F u rth er a n a ly s e s o f m inor DNA
p o p u la t io n s and o f th e c i r c u l a r s t r u c t u r e s o f th e s e
reco m b in a n ts i s n e c e s s a r y to an sw er su ch q u e s t io n s .

- 154 -
E x p r e s s io n o f B x l(2 8 -1 ) s u b c lo n e s d e le t e d i n TR^/IR^
The r e s u l t s o f ex p er im en ts co n d u cted w ith su b c lo n e s o f
B x l(2 8 -1 ) w hich a r e d e le t e d in e i t h e r TR^ o r IR^ showed th a t
e x p r e s s io n o f o n ly one o f th e two c o p ie s" 'o f Vmw"IE 1 1 0 /1 1 8 i s
s u f f i c i e n t f o r a p r o d u c t iv e i n f e c t i o n in v i t r o . Even"
e x t e n s iv e d e l e t io n o f TR^ r e s u l t e d i n v ia b le v i r u s , a s e v id en ced
by su b c lo n e 22 w hich la c k s a c o n s id e r a b le seq u en ce beyond th e
Vmw IE 118 g e n e . C le a r ly , any o th e r g e n e t ic in fo r m a t io n in
TR j/IR ^ i s n o t r e q u ir e d i n d ip lo id am ounts f o r c o n t in u e d v ir u s
r e p l i c a t io n in t i s s u e c u l t u r e . RE4 h a s a d e l e t io n e x te n d in g
from th e j o i n t , th ro u g h IR-^, and a t l e a s t 300 bp in t o U^,
but th e p o ly p e p t id e s in d u ced by t h i s recom b in an t have n b t y e t
been exam ined . The o b s e r v a t io n t h a t a l l r eco m b in a n ts exam ined,
in c lu d in g th o s e w ith d e le t e d s e q u e n c e s , have r e t a in e d a se q u e n c e s
a t th e j o i n t and te r m in i may be a r e s u l t o f a req u irem en t f o r ,
or r e g e n e r a t io n o f , th e a se q u e n c es i n DNA r e p l i c a t i o n .
D e le t io n s i n TRg/lRg a n a lo g o u s to th o s e i n TR j/lR ^ have n o t been
o b se r v e d , p erh ap s b ecau se o f a req u irem en t foi* or" g r e a t e r
s e l e c t i v e ad v a n ta g e o f f u n c t io n a l d ip lo id y o f TRg/lRg i n
p r o d u c t iv e i n f e c t i o n , or b eca u se o f th e m echanism o f d e l e t io n
w hich i s a t p r e s e n t unknown.
There a re s e v e r a l p o s s ib l e e x p la n a t io n s o f th e d ip lo id y
f o r T R j/IR ^ and TRg/IRg d is p la y e d by a l l f i e l d i s o l a t e s
exam ined to d a t e . Two o u t o f tw e n ty B x l( 2 8 -1 ) s u b c lo n e s were
d e le t e d i n TR or IR^, and y e t no e q u iv a le n t d e l e t io n s have been
d e te c te d in o v er a hundred i n t e r t y p i c reco m b in a n ts i s o l a t e d so
f a r i n G lasgow . As w ith n a tu r a l i s o l a t e s , th e fre q u e n cy w ith
w hich d e l e t io n s o ccu r or a re i s o l a t e d in th e l a t t e r c a s e m ust
be lo w e r . The d e le t e d genom es r e s u l t e d from reco m b in a n ts w hich
a re c o m p le te ly h e te r o lo g o u s f o r one o r b oth s e t s o f r e p e a t s .
No o th e r recom b in an t or f i e l d i s o l a t e has been shown to p o s s e s s

- 155 -c o m p le te ly h e t e r o ty p ic d ip lo id r e g io n s ; th u s , i f d e l e t io n s a re
n a t u r a l ly b e in g g e n e r a te d , t h e i r fr e q u e n c y m ust be so low a s to
have escap ed i s o l a t i o n in th e r e l a t i v e l y sm a ll number o f
f i e l d i s o l a t e s exam ined to d a t e . I t i s n o t known why
h e te r o ty p ic reco m b in a n ts w ith d e l e t io n s in TR^/IR-^ were so
r e a d i ly i s o l a t e d , and n e i t h e r i s i t known w h e th e r " th is i s
p r im a r ily a r e s u l t o f th e fr e q u e n t g e n e r a t io n o f d e l e t io n s o r
o f t h e i r su b seq u en t s e l e c t i v e a d v a n ta g e . I t sh o u ld be n o ted
th a t d e le t e d TR or IR^ seq u e n c e s can be r e g e n e r a te d in
hom otypic genombs by r e c o m b in a tio n , w h ereas t h i s i s n o t
p o s s ib le in h e t e r o ty p ic r ec o m b in a n ts .
I f v ir u s e s w ith d e l e t io n s i n th e genome were a t a
s e l e c t i v e d isa d v a n ta g e f o r grow th , n a t u r a l ly o c c u r r in g d e l e t io n
m utants would n o t s u r v iv e . The d e t a i l e d grow th c h a r a c t e r i s t i c s
o f d e le t io n m u tan ts have y e t to be i n v e s t ig a t e d i n v i t r o and
in v i v o . I t i s p o s s ib l e t h a t in c r e a s e d g en e d o sa g e r e s u l t i n g
from d ip lo id seq u e n c es c o n f e r s an a d v a n ta g e i n e s t a b l i s h i n g an
i n f e c t i o n a t low m u l t i p l i c i t i e s , o r i n c e l l t y p e s n a t u r a l ly
r e s i s t a n t to v ir u s i n f e c t i o n o r r e p l i c a t i o n , and t h i s m ight
be p a r t ic u la r ly im p o rta n t i n th e n a tu r a l h o s t .
I t i s a l s o p o s s ib l e t h a t th e p r e s e n c e o f in v e r t e d d ip lo id
r e g io n s , o r th e e x p r e s s io n o f g e n e s c o n ta in e d in them , i s
e s s e n t i a l f o r an u n id e n t i f i e d f u n c t io n o f th e Y ir u s , f o r
exam ple i n th e p r o c e s s e s o f la t e n c y o r c e l l u l a r tr a n s fo r m a t io n .
An u n l ik e ly a l t e r n a t i v e i s t h a t th e p r e s e n c e o f d ip lo id r e g io n s
i s o f no c r u c ia l con seq u en ce to HSV, but m erely r e f l e c t s a
p r o p e n s ity o f DNA to r e p l i c a t e w h erever p o s s i b l e .i
Genome s tr u c tu r e o f RE4
F in a l ly , i t i s w o rth w h ile com m enting on th e e x tr a o r d in a r y
genome s tr u c tu r e o f RB4. T h is v ia b le recom b in an t la c k s 10*5 kbp
o f DNA, in c lu d in g IKL and a s h o r t a d ja c e n t r e g io n o f UL.

- 156 -
2 .8 kbp o f HSV-2 DNA have been in s e r t e d i n t o th e p o s i t i o n o f
th e d e l e t i o n , an e v e n t w h ich presum ably o ccu rred i n th e same
c e l l a s th e i n i t i a l re co m b in a tio n e v e n t , r e n d e r in g th e genome
h e t e r o t y p i c a l l y " d ip lo id 1* f o r t h i s r e g io n o f U^. I t i s n o t
y e t known w h eth er th e in s e r t e d seq u e n c e s a r e f u n c t io n a l i n RE4,
even to th e p o in t o f f u l f i l l i n g a f u n c t io n o f d e le t e d
se q u e n c e s . I t i s o f i n t e r e s t to n o te t h a t th e in s e r t e d -
seq u en ce a lm o s t c e r t a i n l y c o n t a in s u pstream seq u e n c e s and
p erh a p s a p a r t o f th e t r a n s c r ib e d seq u en ce f o r th e gen e c o d in g
' fo r HSV—2 MOP, a m ajor l a t e p o ly p e p t id e . T h is i n t e r e s t i n g
d e le t e d reco m b in a n t w hich i s a l s o f ix e d i n both segm en ts
f u r n is h e s o p p o r tu n ity f o r f u r t h e r s tu d y .

Figure C4.1
Structures of the joints and long termini of Bxl(28-1).
A 1 tsB BamH I k“ 2 B x l(2 E P l) BamH I m ajor j o in t
3 B x l (2 8 -1 ) BamH I m inor j o in t4 t s l BamH I g
D ig e s te d w ith S s t I ; 2 $ a g a r o se g e l .
B 1 tsB BamH I k2 B x l (2 8 -1 ) BamH I ma^or j o in t3 t s l - BamH I g and g
D ig e s te d w ith Sma I ; 7 . 5 % p o ly a cr y la m id e g e l«
D. 1 tsB BamH I ^ t2 t s l BamH I vw3 B x l(2 8 -1 ) BamH I major typ e 2 lo n g term in u s4 B x l(2 8 -1 ) BamH I m inor typ e 1 lo n g te r m in u s ,
w ith HSV-2 Bamll I &
D ig e s te d w ith S s t I ; 2^ a g a r o se g e l .
D 1 tsB BamH I s t2 t s l BamH I vw3 B x l(2 8 -1 ) BamH I m ajor ty p e 2 lo n g term in u s
D ig e s te d w ith Sma I ; 7 .5 # p o ly a c r y la m id e g e l .
S s t I or Sma I r e s t r i c t i o n fra g m en ts o f j o in t o r lo n g te r m in a l fra g m en ts a re in d ic a t e d to th e l e f t (HSV-1) and r ig h t (HSV-2) o f each p a n e l .X d e n o te s a genome te r m in a l fra g m en t, and J th e Sma I fra g m en t sp an n in g th e ju n c t io n betw een th e two tandem a se q u e n c e s in HSV-2 BamH I g .
With r e s p e c t to th e t e x t , th e term s lo n g ter m in u s and 1 te r m in u s , and s h o r t term in u s and S te r m in u s , a re e q u iv a le n t .
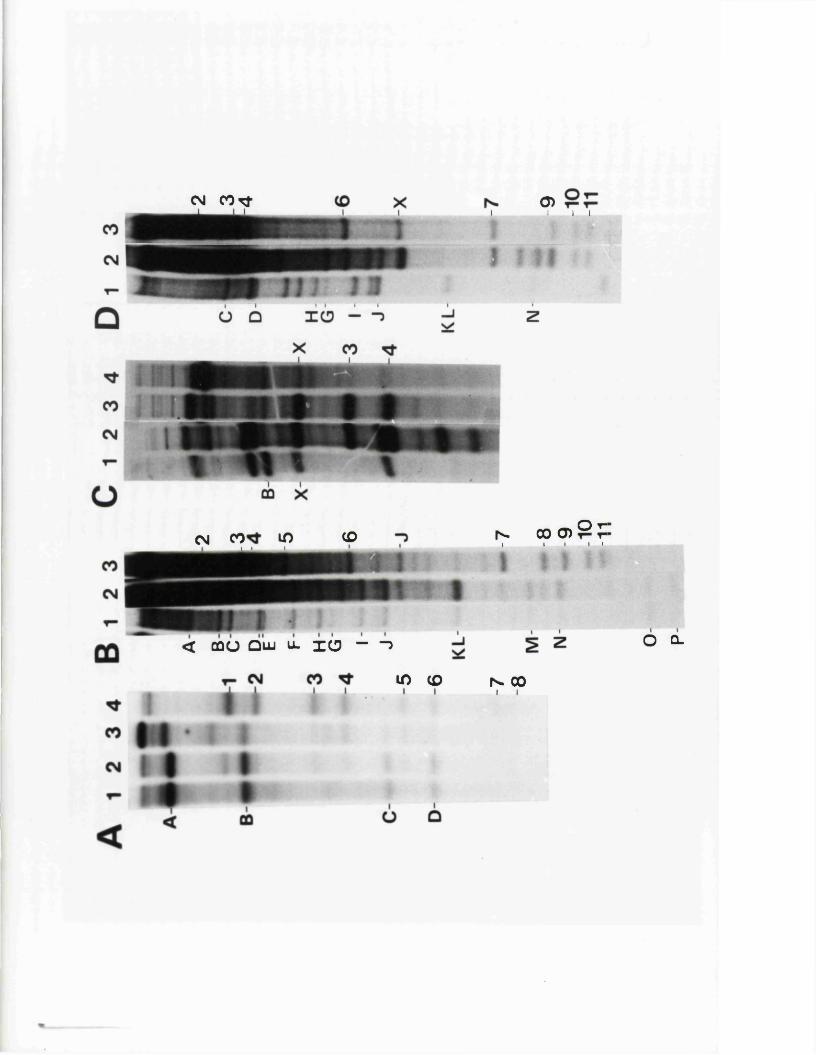
CM OO^f
CM 00 lOi i i
co
O)I I 1
< COO Qu-I ll XCO ” “ 5
lO CO
h- oo a> ° £■ i i i i
O Q-
h - CDi i

F igure C4.2
Structures of the short termini of Bxl(28-l).
A 1 t s B he pit I h2 J3xl(28-rl) hcoK I r ig h t hand S term in u s3 t s l . hooK I m
D ig e s te d w ith S s t I ; 1 .5 •'* a g a r o se g e l .
B As panel A but digested with Kpn I .
C 1 tsB Hind I I I m" 2 Bxl ( 2 8 -1 ) Bgl~~II l e f t hand S term in u s
3 t s l Bffl II~ra
D ig e s te d w ith S s t I ; 1 .5 /* a g a r o se g e l .
D As p a n e l C b u t d ig e s t e d w ith Kpn I .
h 1 tsB BamH I £gi~ 2 t s l BamH I r .
3 TqI BamH I s t4 B x l (2 8 -1 ) BamH I type 1 short terminus,
w ith HSV-2 BamH I r^5 B x l (2 8 -1 ) BamH I type 2 sEort terminus6 tsl-Bam H I u
D ig e s te d with Sma I ; 7 .5 /* polyacrylamide gel.
S s t I , Knn I o r Sma I r e s t r i c t i o n fra g m en ts o f s h o r t te r m in a l fra g m en ts a re in d ic a t e d to th e l e f t (HSV 1 ' ana r ig h t(H S V -2 ) o f each p a n e l . In p a n e ls A and o n ly th o se S3 t I fra g m en ts mapping i n th e BamH I te r m in a l fra g m en t a re in d ic a t e d .A u e n o te s a genome te r m in a l fra g m en t.
I o
l
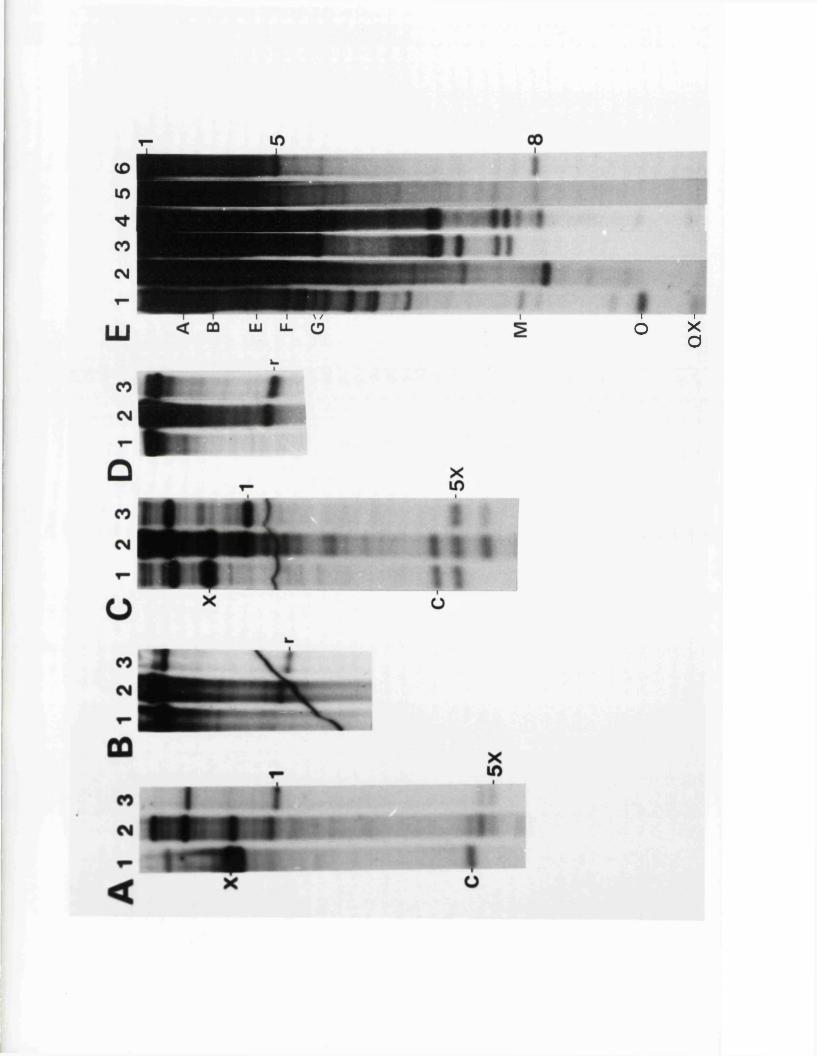
To
QX
^

Figure C4.2
Structures of the short termini of Bxl(28-1).
A 1 t s B Bcoit I k2 B x l (28^*1) BcoK I r i g h t hand S term in u s3 t s l . EcoH I m
D ig e s te d w ith S s t I ; 1 .5 '* a g a r o se g e l .
B As p a n e l A h ut d ig e s t e d w ith Kpn I .
C 1 tsB Hind I I I m~ 2 B x l (2 8 -1 ) B g l~ II l e f t hand S term in u s
3 t s l B g l I l ” m
D ig e s te d w ith S s t I ; lo5 /* a g a r o se g e l .
JJ As p a n e l C but d ig e s t e d w ith Kpn I .
K 1 tsB BamH I pp.~ 2 t s l BamH I r .
3 t s l BamH I s t4 B x l(2 8 -* l) BamH I ty p e 1 s h o r t term in u s ,
w ith HSV-2 BamH I r s5 B x l(2 8 -1 ) BamH I ty p e 2 sE o r t term in u s6 t s l - BamH I u
D ig e s te d w ith Sma I ; 7*5/* p o ly a c r y la m id e g e l .
S s t I , Kpn I or Sma I r e s t r i c t i o n fra g m en ts o f s h o r t te r m in a l fra g m en ts a re in d ic a t e d to th e l e f t (HSV-1) and r ig h t (liSV -2) o f each p a n e l . In p a n e ls A and 0 o n ly th o se S s t I fra g m en ts m apping i n th e BamH I te r m in a l fragm en t a re i n d ic a t e d . j£ u e n o te s a genome te r m in a l fr a g m en t.

CO
CM
I
XLO
■
QX
h

P igure C4» 3
P r e v io u s ly p u b lish e d recom b in ant genome s t r u c t u r e s .
HSV-1 seq u e n c es a re in d ic a t e d above and HSV-2 seq u e n ces below
th e r e p r e s e n t a t iv e genome i n each c a s e . C r o ss -h a tc h in g
d e n o te s th e r e g io n o f u n c e r ta in ty i n c r o s s o v e r p o s i t i o n .
R e fe r e n c e s i
B x l ( 2 o - l ) , 8 x 1 (3 1 -2 ) and 8 x 1 (2 ) - P r e s to n e t a l . (1 9 7 8 ) ,
RS6 - Stow and W ilk ie (1 9 7 8 ) ,
8 x 6 (1 7 -6 ) - iviarsden e t a l . (1 9 7 8 ) ,
10.35-2 - C hartrand e t a l . (1 9 8 1 ) ,
Sach recom b in an t in v e r t s n o rm a lly in L and S e x c e p t B x l ( 2 8 - l ) ,
i n w hich L i s f ix e d in th e o r ie n t a t io n p r e s e n t in th e P and
Ig genome a rra n g em en ts .
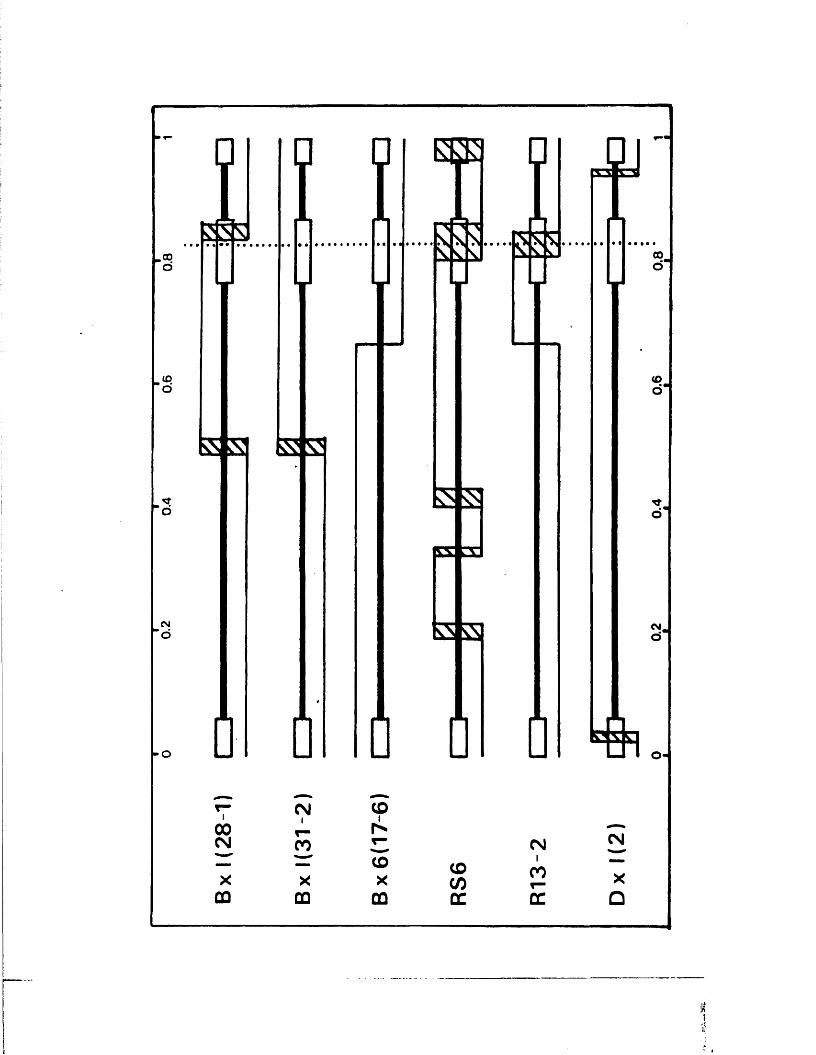
.COd
lOo
r fO
CJ'o
-o
j
o
(Do'
• « o
o
00CM
X00
CMir -00
X00
coi
cox
00
COCOoc
CMi
00
CM
X
Q

11,-Ture 04>4
R e s t r i c t io n p r o f i l e s o f in v iv o ^2P - la b e le d DNA
from s u b c lo n e s o f B x l (2 8 -1 ) f o r BcoR I (A ,B ) and
Hpa I (C ,D ). The su b c lo n e numbers a r e g iv e n a t
th e top o f th e t r a c k s , and a l s o in c lu d e d a re
HSV-1 tsB ( T l ) , HSV-2 t s l (T2) and B x l (2 8 -1 ) (R ) .
R e s t r i c t io n fra g m en ts are in d ic a t e d to th e l e f t
(iiS V -l) and r ig h t (R3V-2) o f each p a n e l .
0 .4 # a g a r o se g e l s were u se d .

A n R 1 2 102729 T2 B T1 R 5 9 11 14 22 T2
* ®d _ — . . _ _ r
de f g B » S - - ? 5 * g . h i :.
• * mm
I *m
m *•
l
C T1 R 1 2 10 2729 T2 D T1 R 5 9 11 14 22 T2abed

Pigure C4.5
R e s t r ic t io n p r o f i l e s o f in v iv o ^2P - la b e le d DNA
from s u b c lo n e s o f B x l (2 8 -1 ) f o r Kpn I (A ,B) and
Bamll I (Cf D ). The su b c lo n e numbers a re g iv e n a t
th e to p o f th e t r a c k s , and a l s o in c lu d e d a re
liSV-1 tsB ( 1 1 ) , HSV-2 t s l (1 2 ) and B x l ( 2 8 - l ) (R ).
R e s t r i c t io n fra g m en ts a r e in d ic a te d to th e l e f t
(HSV-1) and r ig h t (HSV-2) o f each p a n e l .
0#7r* and 1 .0 ^ a g a r o se g e l s were u se d .
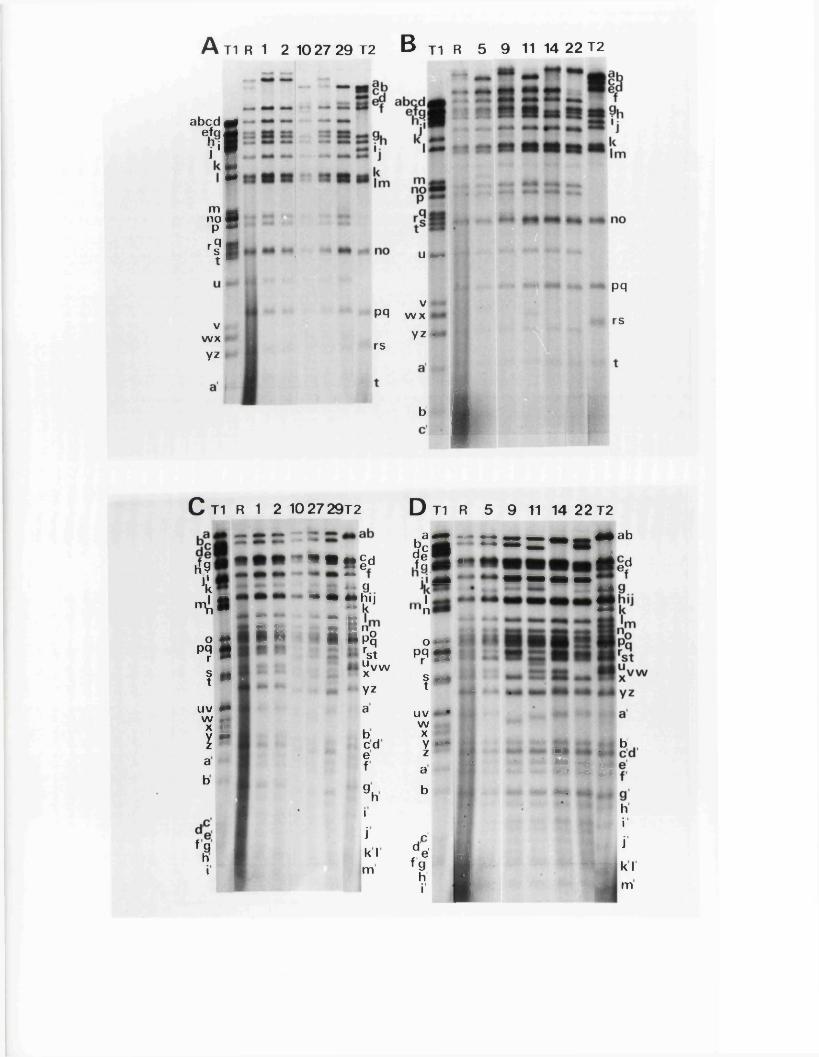
A
abedeJgj j
T1R 1 2 10 27 29 T2 B T1 R 5 9 11 14 22 T2
!
m — n o P P *■
rci ms+l u —
vwxyz
Vpq w x
rsyz «a
b I
pq
rs
C n R 1 2 102729T 2 D
hfg4
"V>*
8f •
opqr
PIPi» i i
uv •»w m x Il mab
' h9i'
• • | i d
; gWi ^ ft* hijfc. k
"o Pq• rstu
X
yzvw
*_ I
n
T1 R 5 9 11 14 22 T2
r - = = - - :^ W « i « M
I
ab
edef
o —• Ip ? f*
?• Ia uv • < ^ PM| 1 I
bc de
wXy —z . •
f a■ M w I
f * i ^ M
g hi
i
b
p '
• * ■-
k lm
i 1
b
hi'j’k rm

Fig u r e C4.6
G ross s t r u c t u r e s o f B x l (2 6 -1 ) s u b c lo n e s . B lack
r e p r e s e n t s HSV-2 DNA, and w n ite HSV-1 DMA, u s in g
th e c o n v e n t io n a l r e p r e s e n ta t io n o f th e 1ISV genom e.
H egion s o f u n c e r ta in ty o f c r o s s o v e r have been
o m itte d s in c e p r e c i s e d e t a i l s a r e g iv e n in
F ig u re C 4 .1 7 . A c r o s s i n d i c a t e s d e le t e d s e q u e n c e s .
The in v e r s io n p ro p e r ty o f L i s n o te d , and the
su b c lo n e number i s g iv e n on th e r i g h t .
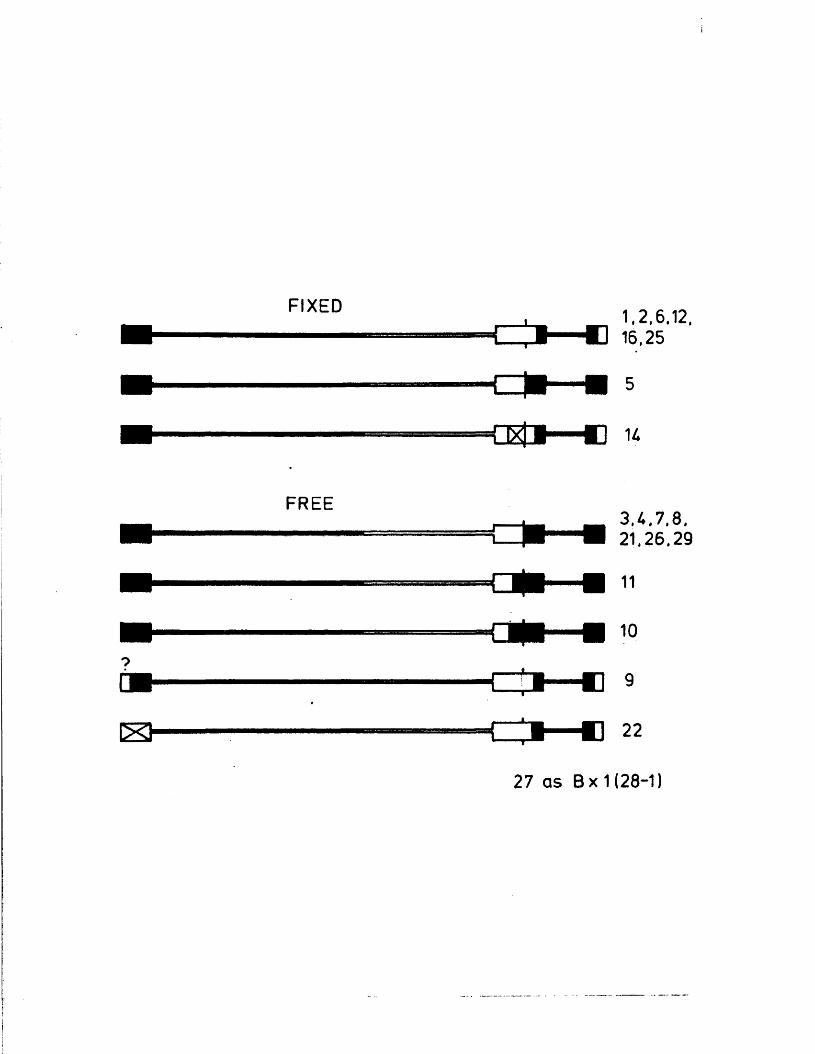
FIXED
FREE
9
m
1. 2 . 6 . 12.16.25
lO — E U
3.4.7.8. 21.26.29
11
10
1— E 9
a E 2 2
27 as Bx 1(28-1)

P ig u re jC iil
Structures of the joints of Bxl(28-1) suhclones
A 1 tsB BamH I k2 J s l BaraH I g3 su b c lo n e 1 BamH I j o in t4 su b c lo n e 5 BamH I j o in t5 su b c lo n e 9 BamH I j o in t6 su b c lo n e 29 BamH I j o in t
D ig e s te d w ith S s t I ; 2 $ a g a r o se g e l .4
B 1 tsB BamH I k"" 2 su b c lo n e 10 Bamli I j o in t
3 t s l BamH I4 t s l BamH I g
D ig e s te d w ith S s t I ; 2 $ a g a r o se g e l .
G 1 tsB BamH I k~ 2 t s l BaraH I g
3 su b c lo n e 1 BaraH I j o in t4 su b c lo n e 9 BamH I j o in t3 su b c lo n e 29~BamiI I j o in t
In v i v o - la b e le d DMA c le a v e d w ith Sraa I ; 7 ,3 /* p o ly a c r y la m id e g e l .
S s t I or Sma I r e s t r i c t i o n fra g m en ts o f j o i n t fra g m en ts a re in d ic a te d to th e l e f t (HSV-1) and r i g h t (lISV-2) o f each p a n e l .
i - cvjco^f in co h* ooi i i i i
I I l ii i i i i i i i i
O l < 0Q(J LU LL X U —
h- CO
S I S
<o
m
Tf
COCM
<1

figure C4»8
Structures of the joints of Bxl(28-1) subclones.
A 1 t sB BamH I k~ 2 su b c lo n e 1 BaraH I j o in t
3 su b c lo n e 9 BamH I j o in t4 t s l BamH I g
D ig e s te d w ith Sma I ; 7 . 5 ‘* (u p p er) and 1 5 & ( lo w e r ) p o ly a c r y la m id e g e l s . * -
B 1 tsB BamH I k2 su b c lo n e 29~BaraH I la r g e r j o in t3 su b c lo n e 29 SaraH I s m a lle r j o in t4 t s l BamH I g
D ig e s te d w ith Sma I ; (u p p er) and 1 5 # (lo w e r )p o ly a c r y la m id e ~ g e ls .
0 1 tsB BamH I k2 su b c lo n e 14 BamH I / Hpa I j o in t3 t s B Hna I m
D ig e s te d w ith Sma I ; 7 . 5 ‘* p o ly a c r y la m id e g e l .
D 1 tsB BamH I k2 su b c lo n e 10 BaraH I j o in t3 t s l Damn I g
D ig e s te d w ith Sma I ; 7 .5 # p o ly a c r y la m id e g e l .
Sma I r e s t r i c t i o n fra g m en ts o f j o in t fra g m en ts a re in d ic a t e d to th e l e f t ( HSV-1) and r ig h t (I1SV-2) o f eacn p a n e l .
A 1 2 3 4 B 1 2 3 4 C 1 2 3 D l 2 3
M «

figure 04.9
Structures of the long termini of Bxl(28-1) subclon
A 1 tsB Bamh I s2 su b c lo n e 1 Bamh I3 su b c lo n e 1 Bamh I
wi th iiSV-24 su b c lo n e 5 Bamli I5 su b c lo n e 5 Bamh I
w ith “ iiSV-26 su b c lo n e 9 Bamli I
w ith “ h3V-27 su b c lo n e 29 Bamh
w ith HSV-28 su b c lo n e 29 ilSV-29 su b c lo n e 29 Bamil
w ith lioV-210 B xl (3 1 -2 ) Bamil I
w ith 113 V-211 t s l Bamli I uvw12 T s l Ba;uh I wx
typ e 2 lo n g term in u s ty p e 1 'lo n g term in u s , Bamil I xtyp e 2 lo n g term in u s typ e 1 lo n g term in u s, Bamh I xtyp e 1 lo n g term in u s , BamH I x
I typ e 2 ~ lo n g term in u s , BaraH I u Bamil I x
I typ e l~ lo n g term in u s , Bamil I x
typ e 1 lo n g term in u s,Bamh I x
D ig e s te d w ith S s t I ; a g a ro se g e l .
tsB BaraH t s l Bamil
s ivw
su b clon e 9 5amn I type 2 lo n g term inus su bclone 9 Bamh I type 1 lo n g term inus su b clon e 29 Bamh I type 2 lo n g term inus 3Ubclone 29 Bamh I type 1 lon g term inus
D ig e s te d w ith S s t I ; 2 ‘« a g a ro se g e l . Term ini in t r a c x s 3 -6 were i s o l a t e d v ia th e u n iq u e lipa I lo n g te r m in a l fr a g m e n ts .
C 1 t s l Bamli I uvw2 su b c lo n e 10 Bamh I typ e 2 lo n g term in u s3 su b e lo n e 14 Bamh I ty p e 2 lo n g term in u s
D ig e s te d w ith S s t I ; a g a r o se g e l .
3 j t I r e s t r i c t i o n fra g m en ts o f lo n g te r m in a l fragm en ts a r e in d ic a t e d to the l e f t (h o V -l) and r ig h t (HSV-2) o f each p a n e l .A u e n o te s a genome term in a l fra g m en t.
to 1m 1 1
1
CO 1 1(N 9 1
CQ|
I
I
XI COI I
CO x
i
i
CD X<1

Pigure C4.1Q
Structures of the long termini of Bxl(28-1) subclones,
A 1 tsB BamH I s2 subcTone 9 BamH I ty p e 1 lo n g term in u s ,
w ith IiSV-2 BamH I x3 su b c lo n e 29 BamH I ty p e 2~"long term in u s4 su b c lo n e 29 Bamll I ty p e 1 lo n g term in u s5 t s l BamH I vw
'6 su b c lo n e 1 BamH I ty p e 2 lo n g term in u s
n ig e s t e a w ith Sma I ; 7•5!* (u p p er) and 15> (lo w e r )p o ly a c r y la m id e g e l s . * -
B 1 tsB Bamll I k.2 t s l BamH I vw3 su b c lo n e 10 BamH I typ e 2 lo n g term in u s4 su b c lo n e 14 Bamll I ty p e 2 lo n g term in u s
D ig e s te d w ith Sma I ; 7*5# p o ly a c r y la m id e g e l .
0 1 tsB Hoa I m2 su b e lo n e 22 typ e 2 Hpa I d e le t e d lo n g term in u s
D ig e s te d w ith Bamll I / Sma I ; 7.51* p o ly a c r y la m id e g e l .
D 1 tsB BamH I s~ 2 TBB Hpa I nf~
3 su b c lo n e 22 typ e 1 Hpa I lo n g term in u s4 su b c lo n e 22 ty p e 2 Hpa I d e le t e d lo n g term in u s3 t s l Hpa I g6 Tsl Hpa I x7 t s l BamH I vyy
D ig e s te d w ith Sma I ; 2 ‘t a g a r o se (u p p er) and 7 .5 ^ p o ly a c r y la m id e ( l o w e r ) - g e l s .
Sma I r e s t r i c t i o n fr a g m en ts o f lo n g te r m in a l fra g m en ts a re in d ic a t e d to th e l e f t (hSV -1) and r ig h t (HSV-2) o f each pari e l .A d e n o te s a genome te r m in a l fra g m en t.
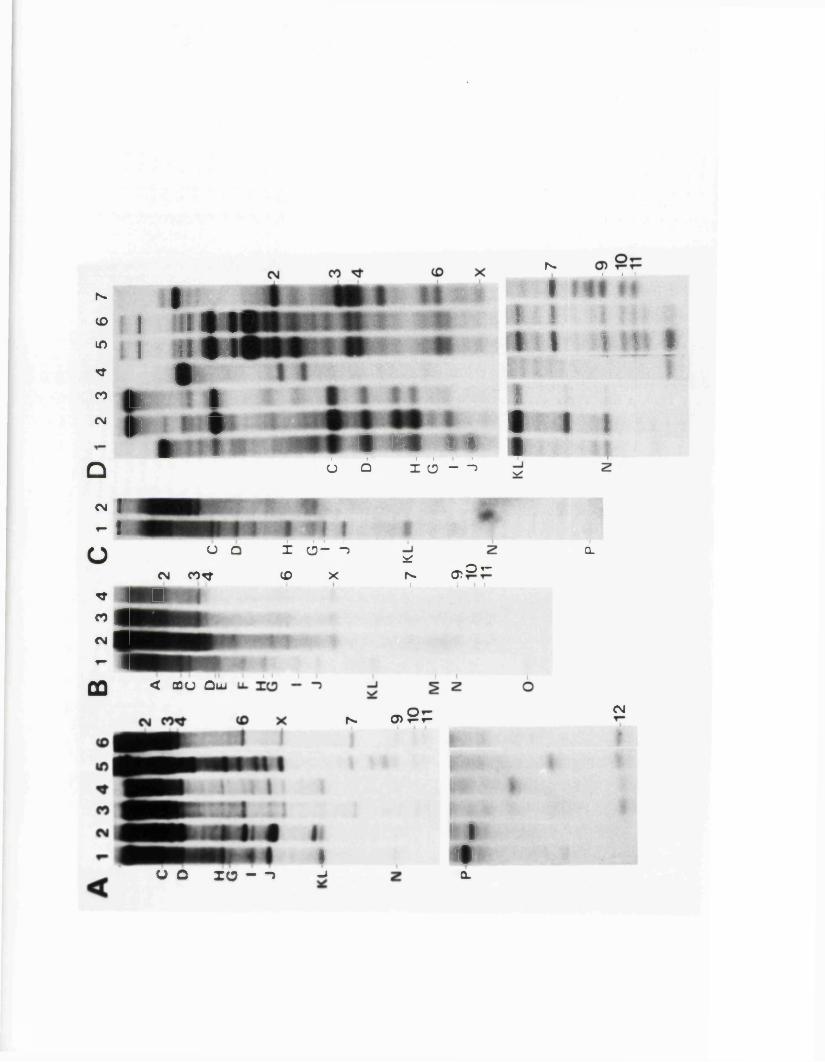
2 3
4 5
6 7 1
1 1 * 11 II $ 1
•m ! i
> • ? !m iQ
CM f- 1
o o c
cm corf^ I VCO ^CM f l
«- |00 < COO Oku 1
f N i » •I 1 I I I !
CJ Q I O - “3
1-
•1 1w1 II
1C•V
i < i >X u ~ - )
tO XXIs-I 0) o * -
Ti
I I I
I I I I I
rs. 0)2^
U O I O — >I-J
I • fQ.
I I
r0-
CM

figure C4.ll
Structures of the short termini of Bxl(28-1) subclones.
1 tsB BamH I2 t s l BamH I u3 su b c lo n e 1 Hamil I s h o r t te rm in u s ,
w ith HSV-2 BamH I r4 su b c lo n e 5 BamH I s h o r t te r m in u s ,
with""il3V-2 BamH I s5 su b c lo n e 9 BamH I s h o r t te r m in u s ,
w itlT iISV -2 BaraH I r6 su b c lo n e 29 BamH iI s n o r t term in u s7 B x l (3 1 -2 ) BamFTl :sh o r t term in u s ,
w ith HSV-2 Bamll I rco t s l BamH I pq t s l BamH 1 r
D ig e s te d w ith S s t I ; 2 # a g a r o se g e l*
B 1 t s l Bamll I r2 t s B BamH I3 su b c lo n e 10 Bamll I s n o r t term in u s4 su b c lo n e 14 BamH I s h o r t te r m in u s ,
w ith HSV-2 BamH I r5 tQ l BamH I u ~ ~
D ig e s te d w ith S s t I ; 2 fa a g a r o se g e l .
C R e s t r i c t io n p r o f i l e s o f in v iv o ^2P - la b e le d~ DNA from s u b c lo n e s o f BXIT28-T7 fo r BamH I ,
in th e r e g io n o f th e v e r y sm a ll fr a g m e n ts .The su b c lo n e numbers are g iv e n a t th e top o fth e t r a c e s , and a l s o in c lu d e d a re HSV-1 tsB (11 )and H3V-2 t s l ( 1 2 ) . R e s t r ic t io n fra g m en ts are in d ic a te d to th e l e f t (HSV-1) and r ig h t (H SV -2). A p o ly a c r y la m id e g e l was used •
S s t I r e s t r i c t i o n fra g m en ts o f s h o r t te r m in a l fragm en ts a re in d ic a t e d to th e l e f t (HSV-1) and r i g h t (HSV-2) o f p a n e ls A and B.X d e n o te s a genome te r m in a l fra g m en t. A lso marked in p a r e n tn e se s in p a n e l A a re S s t I 2 an(i 4. °?HSV-2 BamH I v , tn e lo n g ter m in u s .
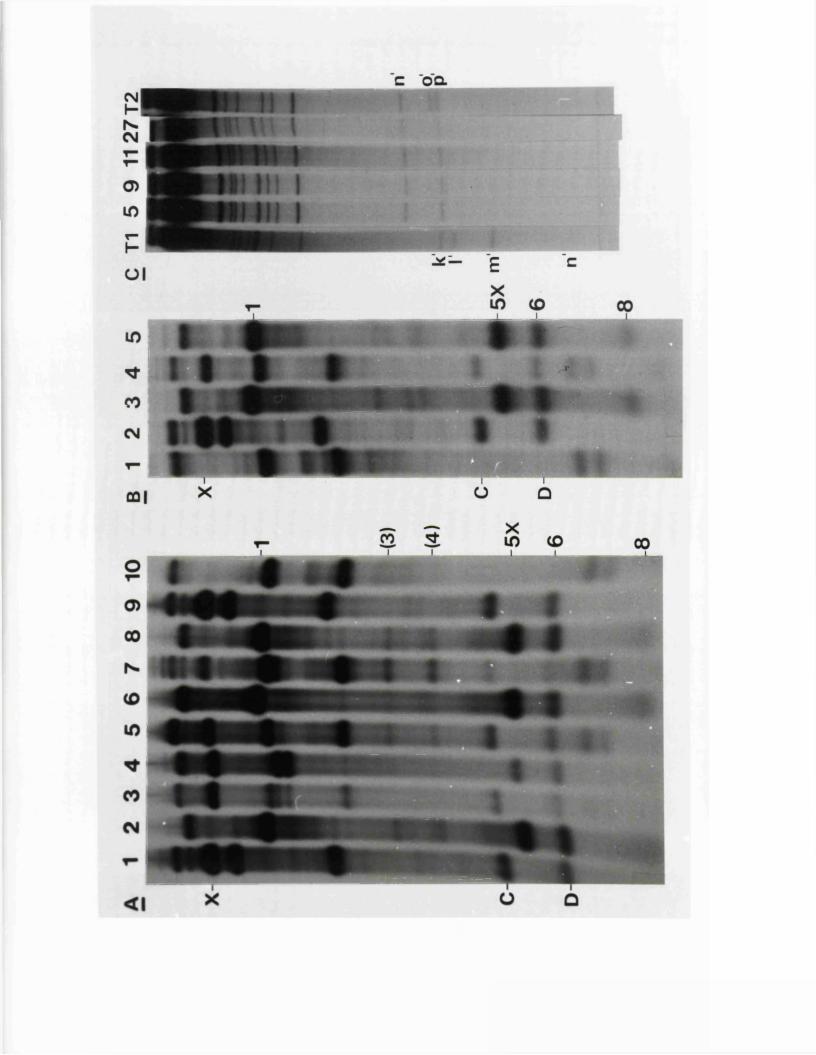

figure C4.12
LEPT HAND TWO PANELS
S tr u c tu r e s o f th e s h o r t te r m in i o f B x l(2 8 -1 ) s u b c lo n e s .
A 1 tsB BamH I pg_2 t s l BamH I r3 t s l BamH I s t4 t s l BamH I u3 su b e lo n e 1 BamH I sn ^ rt te rm in u s ,
w ith HSV-2 BamH I r s6 su b c lo n e 5 BamH I s h o r t te r m in u s ,
with~’HsV-2 BamH I s7 su b c lo n e 9 BamH I s h o r t te r m in u s ,
w ith HSV-2 Bamu I r8 su b c lo u e 29 Bamii I~sITort term in u s
D ig e s te d w ith Sma I ; 7 .5 /* p o ly a c r y la m ia e g e l .
B 1 t s l BamH I r2 tsB Bamli I £9.3 su b c lo n e 10 Bamli I s a o r t term in u s4 su b c lo n e 14 Bamli I s n o r t term in u s 3 t s l Bamll I u
D ig e s te d w ith Sma I ; 7 . 9 p o ly a c r y la m id e g e l .
Sma I r e s t r i c t i o n fra g m en ts o f s h o r t te r m in a l fragm en ts a r e in d ic a t e d to th e l e f t ( ilS V -l) ana r i g h t (HSV-2) o f th e p a n e l3 .X d e n o te s a genome te r m in a l fra g m en t.
RIGHT HAND TWO PANELS
A u torad iograp h o f n i t r o c e l l u l o s e b l o t s t r i p s c o n ta in in g Kpnl or BamHI r e s t r i c t i o n fra g m en ts o f su b c lo n e 14 (S lJT or 2<TX322) LNA., to w h ich n ic k t r a n s la t e d BNA p ro b es had been h y b r id is e d .1 1The p ro b es w ere: HSV-1 DNA (H ), and a m ixtu re o f p la sm id s c o n ta in in g liS V -l BamHI k and. HSV-2 BamHI g ( J ) . HSV-1 DNA h y b r id is e d s t r o n g ly ..to HSV-1 fra g m en ts, in th e recom b in an t genom es, and to a l e s s e r e x t e n t to HSV-2 fr a g m e n ts . An e x p la n a t io n o f th e bands to w hich th e J probe h y b r id is e d i s g iv e n (NLT - n o v e l L term in u s; HI -.H SV -1 fragm en t;H2 - HSV-2 fra g m en t; S . - S segm en t, s in c e th e su b c lo n e genom es have no Kpnl s i t e s in S ) .
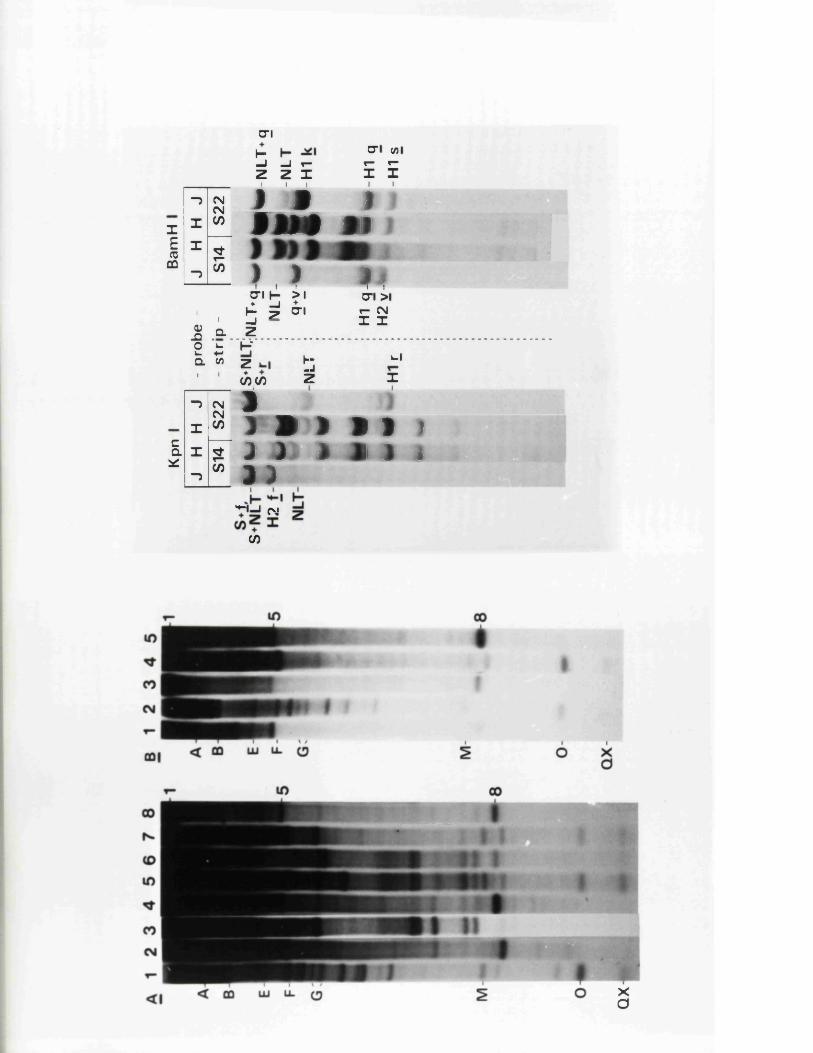
IECOco
ca*
□■ih*-J _l r-Z Z XI I 1
H1
q -H
1 s
“3I S
22 I 1 VIM
1
• 1 II
“) S14
» ? l i
iCD-Qo
iCJI f- >1♦ _j +h- 7 0-1 -J H1
q-
H2
v
aL_
ZKk.
a-*->(/) z t]
i.ii 1 *,co z Xi
“)
S22 >
X I I I 1 1 )X-) S1
4 J ) ] )
) )M1 1 i
*-|_l fVl
(/)
QXH

in j u r e C4.13
Structures of the joints and termini of Bxl(31-2).
A i tsB BamH I s t7 2 B x l (3X72) BamH I j o i n t ,
w ith o th e r fragm ents..3 B x l (3 1 -2 ) BamH I s h o r t term in u s and ty p e 2
la r g e r lo n g te rm in u s , w ith HSV-2 BamH I4 B x l (3 1 -2 ) HSV-2 BamH I s -----5 B x l£ 3 1 -2 ) BamH I ty p e l~ lo n g ter m in u s ,
w ith HSV-2 BamH I x6 t s l BamH I vw ~ ~
D ig e s te d w ith S s t I ; 2>« a g a ro se g e l .
B As f o r psu*el A but d ig e s t e d w ith Sma I ; 7 .5 # p o ly a c r y la m id e g e l .
0 1 t s B Bamii I s t2 t s l BamH I vw3 B x l (3 1 -2 ) BamH I type 2 lo n g term in u s4 B x l (3 1 -2 ) Bamli I typ e 1 lo n g term in u s5 D x l(2 ) BamH I lo n g term in u s from th e r ig h t
Haiid end o f L6 D x l(2 ) BamH I lo n g term in u s from th e l e f t
hand end o f L7 D xl (2 ) Bamii I s h o r t- te r m in u s from th e
r ig h t hand end o f S8 D x l(2 ) BamH I s h o r t term in u s from th e
,X eft hand end o f S9 tsB BamH I pq
10 t s l BamH I u
Fragm ents fo r tracK s 3 -3 i s o l a t e n from la r g e r u n iq u e fr a g m e n ts . D ig e s te d w ith S s t I ; 255 a g a r o se g e l .
D 1 KSV-1 S s t I lo n g term in u s~ 2 t^ 6 S s t 1 lo n g term in u s
3 B x l (3 1 -2 ) S s t I ty p e 2 la r g e r lo n g term in u s4 B x l i3 1 - 2 ) S s t I ty p e 1 lo n g term in u s5 D x l(2 ) S st~ T ~ Ion g term in u s from th e r ig h t
hand end o f L6 D x l(2 ) S s t I lo n g term in u s from th e l e f t
Hand end o f L7 t s 6 S s t I lo n g term in u s
Ss t I fra g m en ts i s o l a t e d from la r g e r fr a g m e n ts , w hich were u n iq u e f o r t r a c e s 3 - 6 . D ig e s te d w ith Sma I ;7.3,-® p o ly a c r y la m id e 0 e l
ites t r i e t io n fra g m en ts o f Bamii I te r m in a l fra g m en ts a re in d ic a t e d to th e l e f t (HSV-1) and r ig h t (HSV-2) o f th e p a n e ls e x c e p t in p a n e l 0 where lo n g te r m in a l fra g m en ts a re in d ic a t e d on th e l e f t and s n o r t te r m in a l fra g m en ts
w ith nSVr2 fra g m en ts in p a r e n th e s e s , x d e n o te s a genome UH’M inal fT agm cnt.
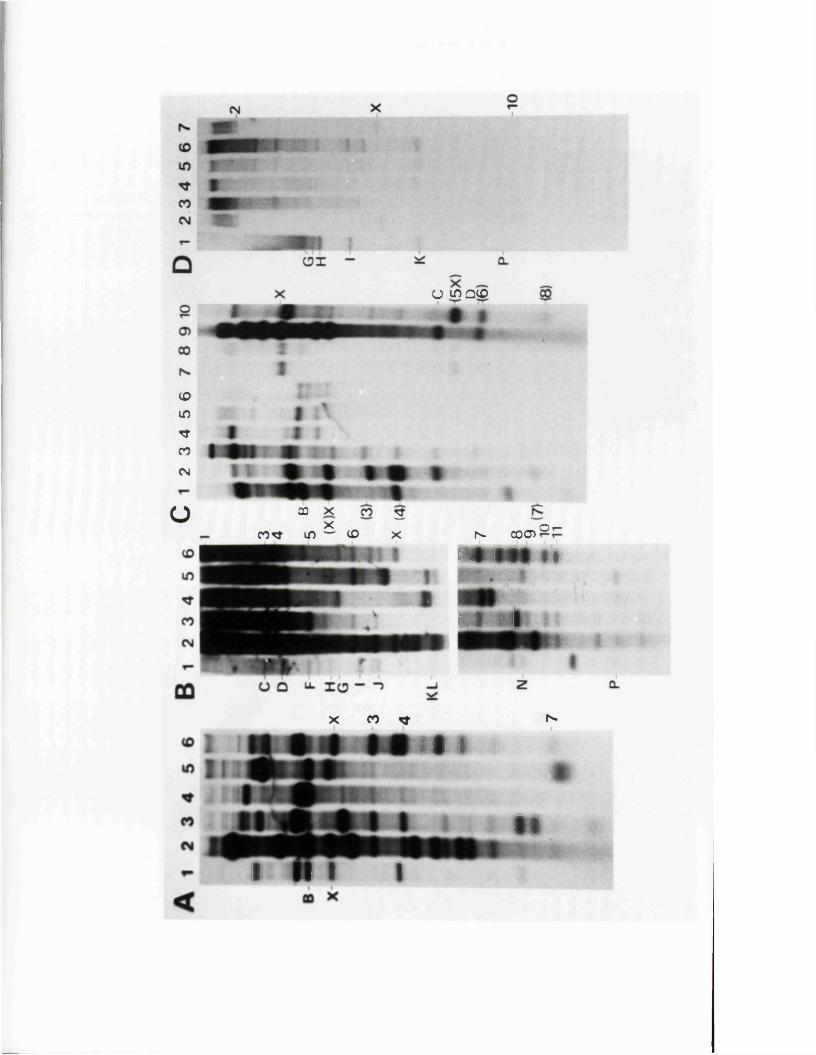
u mQlO
<Dintj-coCNJ
o 00 X co 5 Xco f m m x fs. CO G)
O t-
141 M t f t
H i I I
X CO Tf ^
if k i
i n i iit

Figure C4.14
Structures of the joints and termini of Bx6(17-6).
1 tsB2 tsB3 tsB4 5 x 65 Bx66 Bx6
7 t s l8 T s l9 t s l
10 t s l
A 1 ta l i2 B x l( 3 1 -2 )3 Bx 6 (1 7 -6 )4 t s l -
In v iv o ^ P ~ la b e le d j>i>[a d ig e s t e d w ith BamH I ; If* a g a r o se g e l .
B 1 tsB BamH I s t |sB BamH I s tsB BamH I k
BamH I j o i n t B x 6 ( l7 -6 ) Bamii I s h o r t term in u s B x 6 ( l7 -6 ) Bamii I lo n g te rm in u s ,
w ith H3V-2 BamH I w
t s l Bamli I vw t s l Bamii I x
D ig e s te d w ith S s t I ; 2f» a g a r o se g e l
G 1 tsB Bamii I k2 5 x 6 ( 1 7 -6 ) Bamii I jo in t3 t 3 l - BamH I~ g
D ig ested w ith Sma I ; 7 * 5 # p olyacry lam id e g e l .
D 1 tsJi Bamii I s2 J sB Bamii I ^3 Bx6 ( 1 7 -6 ) BamH I s h o r t term in u s4 B x 6 (1 7 -6 ) BamH I lo n g te rm in u s ,
w itE H3V-2 BamH I w5 t s l BamH I u ~6 t s l Bamii i vw7 t s T BamH I x
D ig e s te d w ith Sma I ; 7 « 5 ‘* p o ly a c r y la m id e g e l .
HSV-1 and HSV-2 r e s t r i c t i o n fragm en ts a re in d ic a t e d to th e l e f t and r i g h t , r e s p e c t i v e l y , o f each p a n e l .In p a n e l A th e y r e f e r to th e Bamll I d i g e s t o f HSV-1 and HSV—2~DJMA, and in p a n e ls B-D to th e S 3 t I o r Sma I d i g e s t o f Bauni I j o i n t fr a g m e n ts .

r- CM OOTfr 10 CD h* COO^r-I I I I !
J
1
i - ? oQ | < CQO Q lu Li. X C 3 — ")
O l < 00 (J Q lu ll i ( D — - )
_ D X >- CD !fi U Q)H- CTLC
• - * — C > £ X > ^ CD iQ - O t j c 1-

Figure C4.15
Structures of the joints and termini of HS6 and HI3-2.
A 1 tsB BamH I k2 TsB BamH I a3 HS6 BamH I j o in t4 KS6 BamH I lo n g term in u s5 t s l BamH I g6 T s l BamH I u7 t s l BamH I vw
D ig e s te u w ith S s t I ; 2>'« a g a ro se g e l .
B As f o r p a n e l A but d ig e s t e d w ith Sma I ; 7 .5 # p o ly a c r y la m id e g e l .
C 1 tsB BamH I k~ 2 t s l BamH I g
3 RT3-5"T?amIl I la r g e r j o in t4 H13-2 BamH I s m a lle r j o in t5 HI3—2 BamH I typ e 2 lo n g term in u s6 H 13-2 Bamli I ty p e 1 lo n g term in u s ,
w ith HSV-2 BamH I x7 t s B BamH I s ”* *"8 t s l BamH I vw
D ig e s te d w ith Sma I ; 7 .5 # (u p p er) and 1 5 # ( lo w e r )p o ly a c r y la m id e g e l s . - -
S s t I or Sma- I r e s t r i c t i o n fra g m en ts o f BamH I " jo in t fra g m en ts a r e in d ic a t e d to th e le ft^ T E S V -l) and r ig h t (liSV -2) o f each p an el.
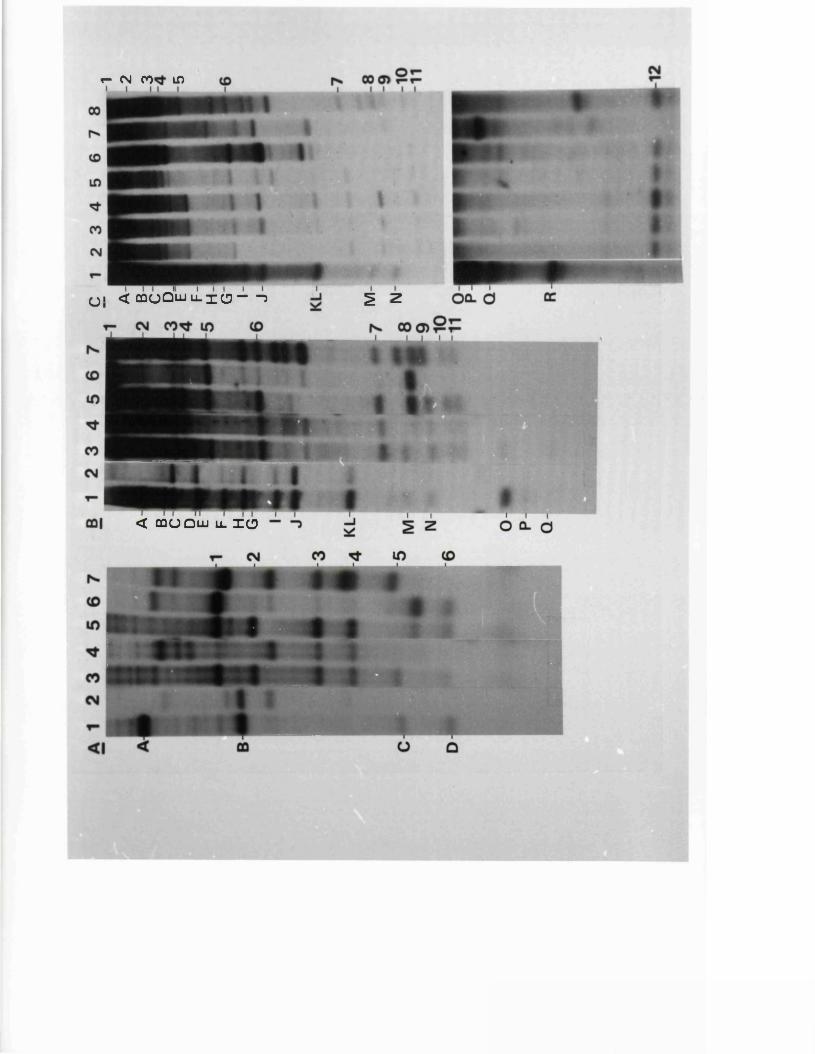
t - CM CO If)
QI < OQOQuj U-XCJ ) ^ 2
< COO O ld ll I O

Figure C4.16
Structures of the joints and termini of Dx 1(2)-.
A 1 tsB BamH I k2 t s l) Bamk I k3 B xl (2T""BamH~I j o in t4 s u b c lo n e o f su b c lo n e 2 o f B x l( 2 8 -1 ) BamH I j o in t5 t s l Bamll I g
D ig e s te d w ith Sma I ; 7 .5 # (u pp er) and 1 5 # ( lo w e r ) p o ly a c r y la m id e g e l s . - _ « „
B 1 t s B S s t I C and D ( i s o l a t e d )2 Bxl(2j~BamH I s h o r t term in u s from th e
r ig h t hand end o f S3 B x l (2 ) BamH I s h o r t term in u s from th e
l e f t hand end o f S4 t s l BamH I u
B ig e s te d w ith S s t I , e x c e p t f o r t r a c k 1$7 .5 # p o ly a c r y la m id e g e l .
C 1 . tsB BamH I pg.2 tsB BamH I s3 B xl (2 ) Bamli I s h o r t term in u s and la r g e r
lo n g term in u s4 B xl (2 ) Bamll I lo n g term in u s5 t s 6 BamH~~T~u6 t s 6 BamH I vw
B ig e s te d w ith S s t I ; 2 # a g a r o se g e l .
B 1 tsB BamH I pc2 TsB BamH I s3 jJ x l(2 j” BamH I s h o r t term in u s and la r g e r
lo n g term in u s '4 B x l (2 ) BamH I lo n g term in u s5 t s 6 . Bamii I. u6 t s 6 ’ Bamii I vw
B ig e s te d w itn Sma I ; 7 .5 # (u pp er) and 1 5 # ( lo w e r ) p o ly a c r y la m id e g e l s . * * -
H e s t r ic t io n fra g m en ts o f BamH I j o i n t (p a n e l A) and lo n g and s h o r t te r m in a l fra g m en ts (p a n e ls B-D) a re in d ic a t e d to th e l e f t (H SV -l) and r i g h t (HSV-2) o f each p a n e l . Xp d e n o te s th e te r m in a l S s t I or Sma I iragm en t o f L, and th e te r m in a l fragm en t o f S .

< CDCJ Q lu ll — -5
^ cm in to
<C 0U O u iU -X O ” ->
r» C0O>9t1 ‘ 1
_j- X
m ID
9 §9 f
i 1
t fO Q
00 o 9
11i
_jx 5 z
CM
1 co0 1 x
O
CO
CM
0 0 -0

Figure C 4 .17
D e ta ile d genome s tr u c tu r e s o f B x l(2 8 -1 ) su b c lon es
and o th e r recom binants: (a ) f ix e d in L, (b) f r e e in L.
and Ug have been sh o rten ed , and the c r o sso v e r in
fo r B x l(2 8 -1 ) su b c lo n es i s in the reg io n 0 .4 9 0 -
0^515. R ecta n g le s r e p r e se n t r e p e t i t iv e r e g io n s in the
c o n v e n tio n a l manner, b lack in d ic a t in g type 2 and
w h ite type 1 DNA. The a sequ en ces are shown on
a la r g e r s c a le fo r c l a r i t y . R e s t r ic t io n s i t e s
d e f in in g c r o ss o v e r s are shown a t the to p and bottom
o f th e diagram , and a ls o above (HSV-1) and below
(HSV-2) each r e p r e s e n ta t iv e genome (B BamH I ,
K Kpn I , R BcoR I , H Hpa I ) . C rosses in d ic a te
d e le te d seq u en ces and c r o s s -h a tc h in g shows r e g io n s
o f u n c e r ta in ty fo r c r o sso v e r s where g r e a te r than
300 bp. In some c a s e s c r o sso v e r s were determ ined
by f in e r e s t r i c t i o n mapping in the j o in t and
term in a l r e g io n s . Recombinant D x l(2 ) has two
h e te r o ty p ic a seq u en ces a t th e j o in t and so IRg
has been s h i f t e d r ig h tw a rd s.
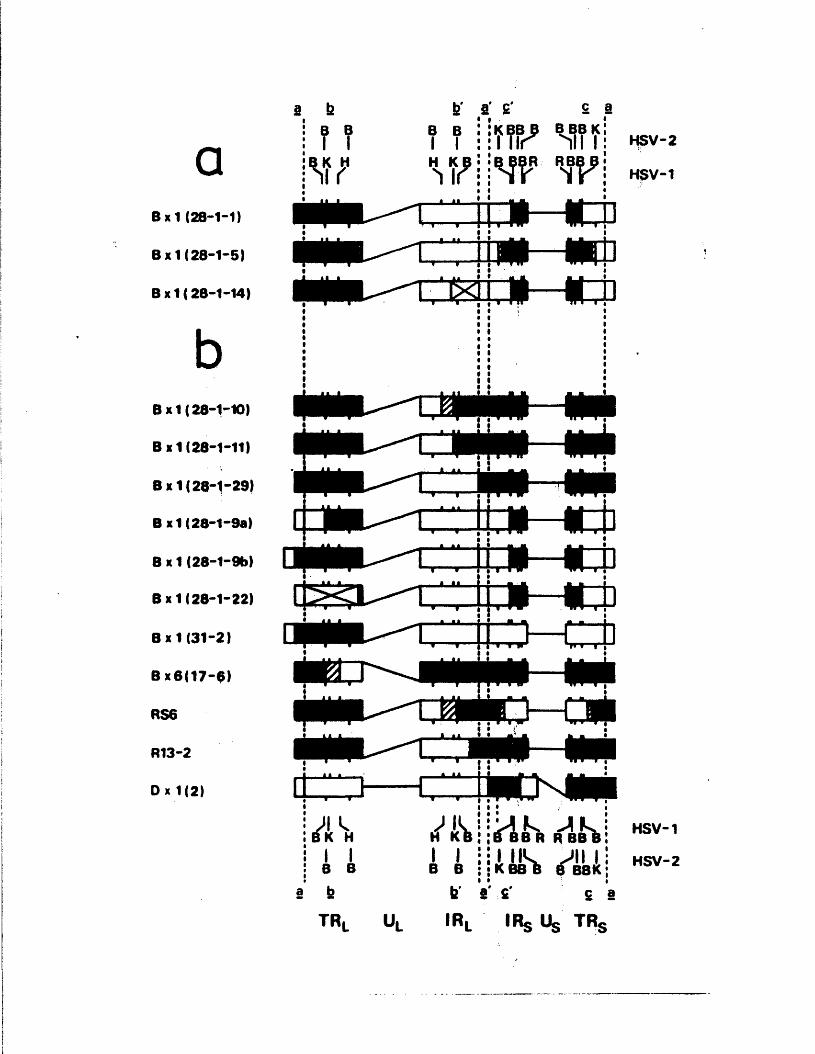
aB x 1 (28-1-1)
8 x 1 ( 2 8 - 1 - 5 )
B x l (2 8 -1 - 1 4 )
bB x 1 (28-1-10)
B x 1 (28-1-11)
B x 1 ( 2 8 - 1 - 2 9 )
B * 1 ( 2 8 - 1 -9a)
B x 1 ( 2 8 -1 -95 )
B x l ( 2 8 - 1 - 2 2 )
B x 1 ( 3 1 - 2 )
B * 6 ( 1 7 - 6 )
RS6
R13-2
Dx 1(2)
fe' 2' 2'B B
Vt? W \
■ m — i
• — I C P
t w — I W
a —* 3
Uil
LOio — i d ]
H a n d ]a —id )
i " : l j . : . v - c 3
TR, U,
m
• t f ^ B R R B B b !
1 y s r f u iI
£ 2
, r l IRs Us TRs
HSV-2HSV-1
HSV-1
H SV-2

Figure 04.18
Upper p an el
Genome s tr u c tu r e s p o s s ib ly p r e se n t in B x l ( 2 8 - l ) ,
and th a t o f th e p u ta t iv e o r ig in a l recom binant B x l(28 ) from
which th ey a r o s e , B lack in d ic a t e s HSV-2 and w h ite HSV-1
seq u en ces.
Lower p an el
S tr u c tu r e s o f th e 4 o in t r e g io n s o f th r e e
recom binant genom es. B lack in d ic a t e s HSV-2 and w h ite HSV-1
seq u en ces, and c r o s s -h a tc h in g d e n o te s th e r e g io n o f
u n c e r ta in ty o f c r o sso v e r p o s i t io n . The a sequ en ces are
shown a s broader r e c ta n g le s and th e ju n c tio n between two
a seq u en ces by v e r t i c a l l i n e s ex ten d in g above and below them.
S iz e s o f n o v e l Smal fragm en ts c o n ta in in g th e c r o sso v e r s are
givien.
a Subclone 29 o f B x l(2 8 -1 ) (F igu re 04*10 p an el A
tr a c k 4; e n t ir e genome s tr u c tu r e in F igu re 0 4 * 1 7 ),
b B x l(2 ) (F igu re 04*16 p an el A tr a c k 3 ; e n t ir e
genome s tr u c tu r e in F igure 0 4 * 1 7 ),
c Subclone o f su b clon e 2 o f B x l(2 8 -1 ) (F igu re 04*16
p an el A tr a c k 4; e n t ir e genome s tr u c tu r e in
F igu re C 4 .21 , second typ e o f p ro g en y ).
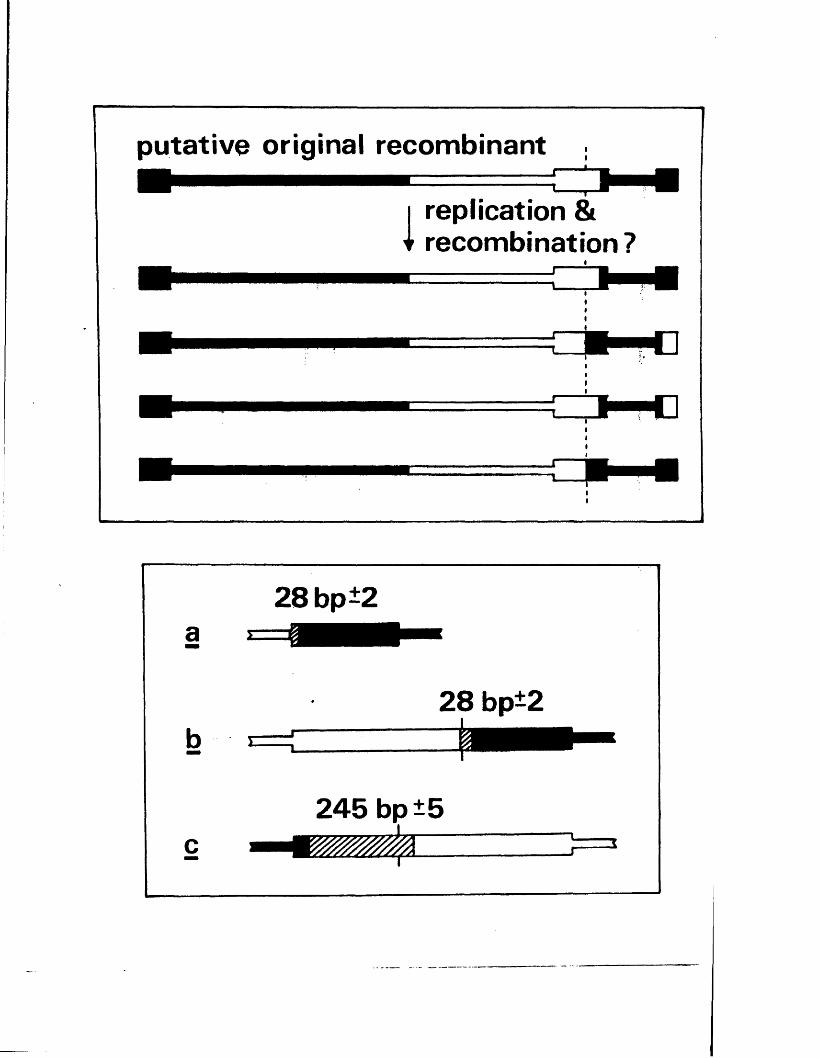
putative original recombinant
I replication & t recombination?
28 bp-2a £
28 bp-2b = [
245 bp ±5c —m m

Figure 04*19
Genome s tr u c tu r e s o f su b c lo n es o f B x l(2 8 -1 ) used to
in v e s t ig a t e e x p r e ss io n o f IB 1 1 0 /1 1 8 .
Ujj and Ucj have been sh orten ed to em phasise d e t a i l s
o f the r e p e t i t iv e r e g io n s . R e s t r ic t io n s i t e s
w ith in r e p e t i t i v e r e g io n s are shown above the
genome fo r iiSV-1 and below fo r HSV-2 (B « BamH I ,
K « Kpn I , II *= Hpa I , H - KooB I ) . B lack d en otes
HSV-2 DNA and w h ite HSV-1 DBA, c r o s s -h a tc h in g
in d ic a t e s u n c e r ta in ty o f c r o sso v e r p o s i t io n where
g r e a te r than 3 0 0 bp, ana a c r o s s s i g n i f i e s
a e le te d se q u en ce s . The c r o sso v e r p o s i t io n in
i s between 0 .4 9 0 and 0 .515 f r a c t io n a l genome u n it s
fo r each su b c lo n e . The p o s i t io n s and d ir e c t io n o f
t r a n s c r ip t io n o f the genes cod in g fo r IB 110 and
IB l i b are shown (A. B aston , F. Rixon and J .B .
C lem ents, p e r so n a l com m unication).

I.E. 118 I.E. 110
8x1(28-1)
S1
S11
S29
S14
S 2 2
H
MJ ___ IKl
H
H K BJ L i -
t H TK B BB B B B K •
I)
1 1 B BB R R BB B j
BB B K
BB B KK B BB

F igure 0 4 .2 0
A utoradiograph o f IE p o ly p e p tid e s induced in HFL
c e l l s by HSV-1 tsB ( 1 1 ) , HSV-2 t s l (T 2),
recom binants B x l(3 1 -2 ) and B x l(2 8 -1 ) and su b c lo n es
(S) d er iv ed from B x l ( 2 8 - l ) . A lso in c lu d ed are
m o ck -in fec ted (MI) sam p les. IE p o ly p e p tid e s from
whole in f e c t e d c e l l s were sep ara ted on 5 - 1 2 ^
p olyacry lam id e g r a d ie n t g e l s ( la n e s 1 - 1 0 ) or
n u c lea r e x t i 'a c ts were made and sep arated on
7.5;^ p o lyacry lam id e g e l s ( la n e s 1 1 -2 1 ) . Numbers to
the l e f t o f la n e 1 and to th e r ig h t o f la n e 2 1 show
the apparent m olecu lar w e ig h ts (x 1CT3 ) o f HSV-1
IE p o ly p e p t id e s . Numbers between la n e s 10 and 11
show th e apparent m olecu lar w e ig h ts (x 1 0 *“^) o f
HSV-2 IE p o ly p e p t id e s .
Symbols are p la ced to th e l e f t o f the la n e to which
th ey r e f e r .
• HSV-1 polypex>tide
o HSV-2 p o ly p ep tid e
4 - a l t e r e d p o ly p ep tid e
I n is experim ent was perform ed by Dr H .S. Marsden.
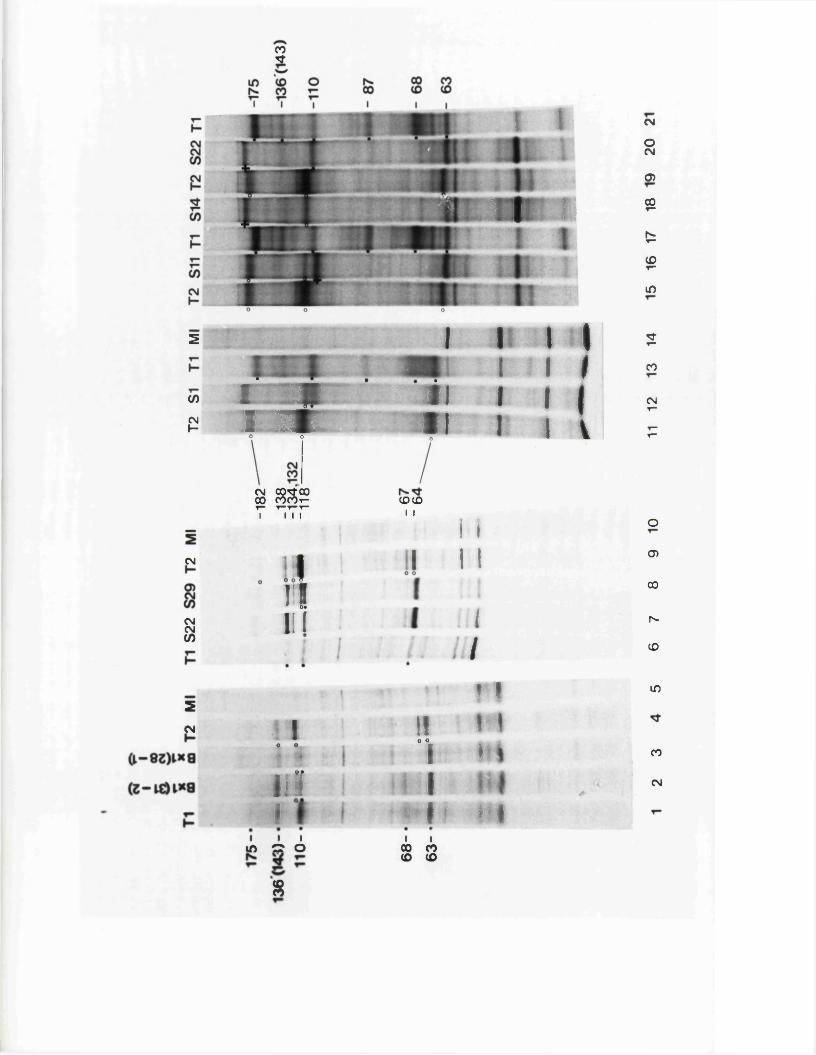
CO*a-
eg
eg cn cn
2
eg
i i i
a i !i
s SRS00co cocoi-7 777
I!'!1!!
! Io o My m.
1>?*
I I Iin 2 o
CD CD I I
I
I iI I
AI
1!o o
iI
i §ifi i
CO COco co
t I
f Il
h i
egOego>
coT-
£
COy—
in
2 1 i 1 1 ^X1 :i 1 1 «■T" " 1wt cn 1 .JP ii 1 1 | ?
p I 71"o o i in 1 1 V ?
oo>
co
r ^
CD
in
co
eg
8

Figure C4.21
Genome s tr u c tu r e s o f th e progeny o f two su b c lo n es
o f B x l(2 8 -1 ) : (a ) su b clon e 2 9 , (b) su b clon e 2 .
Schem atic d e t a i l s are th e same a s in F igure
C4»17, e x ce p t th a t a seq u en ces are n o t em phasised .
rih e p a r e n ta l s tr u c tu r e s are shown, and the number
o f progeny w ith each r e s u l t in g genome s tr u c tu r e .
F in d ic a t e s f ix e d in L and N in d ic a t e s norm ally
in v e r t in g ( f r e e ) in L.
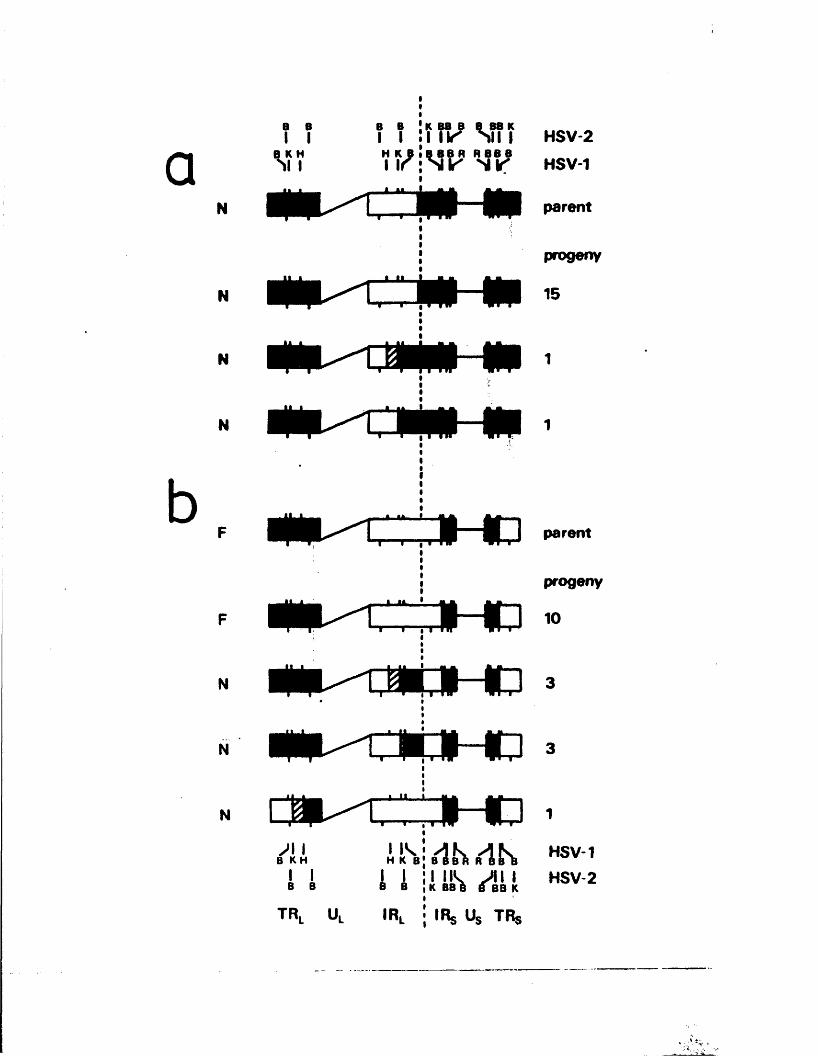
Cv\
H DX J U H D
a t a » 1 H 0
parent
progeny
15
1
1
parent
progeny
10
3
3
1

ffi&ure C4.22
R e s t r ic t io n i^ r o f ile s o i in v iv o ^ P - la b e le d DNA from
RE4 (R) and some o f i t s su b c lo n es (numbered)
compared w ith HSV-1 (T l) and IISV-2 ( i ‘2) fo r Hind I I I
( l e f t ; 0.4/S agarose g e l) and HainH I ( r ig h t ; 1>
agarose g e l ) . HSV-1 r e s t r i c t i o n fragm ents are
in d ic a te d to the l e f t o f each p a n e l, and IISV-2
r e s t r i c t i o n fragm ents to the l e f t o f th e v e r t i c a l
l i n e .
COLO
St
C\J
coLO
St00
CSIH
< • « « < «I M K I IBM 4i i i i i iifliiii
( A f t «( « < < i ( I l f ( Im i «
I «l l t l <1 I B I « BUtlHMBMIiiiura in i
14— *—
IBI1 i
• m i
i m i l ■ v i aI * B 1 f "
II l l t l
OaC/3-M
I I I I I
I I I I «
2 5 x>n 0)0)--
.Q O 0 a ) - 3 'v ' _ £ Q .H .D X N 03-d * - _ c — E o ct£ £ >
>
^2-D 0' a
4- c n -c ■-
r» 1 III I I1 I 1 •
CO 1 1 1 111 II 1 ! 1 1 flfliLO 1 II III I I1 1 1 1 I
1 1 III 111 1 1 1 IBBCM 111 III I I 1 1 1 I I B
11 I I S II 1 1 1 warnCO IB I I1 1 I IBBCO 1 1 1 IB ■ 1 1 1 IBM
1 I 1 III B 1 1 1 1 IMCO *J 1 IB B 1 1 1 I B M
B O B B i 1 81- mcc 1 E 1 III I I1 1 1 i nCMH R B B 1 1 11K I I 1 IB III 1 1 m«oo 0 *•- o > - c c <0*0 -E c o
▼ T " I T T T^■Sfx i r Eic io

Figure C4.23
H y b r id isa tio n o f s p e c i f i c probes to Southern b lo t s
o f HJS4 r e s t r i c t i o n d ig e s t s .
'i'he tra c k s are Hind I I I (upper h a lf o f F igu re) or
Bamli I (low er h a l f ) d ig e s t s o f HSV-1 ( T l) , HE4 (K)
or HSV-2 (T2) HNA tr a n s fe r r e d to n i t r o c e l lu lo s e from
agarose g e l s . A-C were tr a n sfe r r e d s im u lta n eo u sly
from the same g e l , and l ik e w is e H-E. The n i t r o c e l lu lo s e
f i l t e r s were c u t in to p o r t io n s c o n ta in in g th ree
tra ck s and h y b r id ised w ith n ic k tr a n s la te d BNA
a s f o l lo w s .
A HE4 HNA . . . -H recom binant p lasm id c o n ta in in g HSV-2 Hind I I I nC recom binant p lasm id c o n ta in in g HSV—2 HXnd I I I oH HE4 HNAS’ Ilind I I I TJ fragm ent i s o la t e d from HE4 HNA
( s l i g h t l y contam inated w ith RE4 Hind I I I x)
Those fragm ents which h y b r id ised to probes H, C and E
are in d ic a te d : HSV-1 fragm ents on the l e f t o f th e
tracic, HSV-2 on th e r ig h t . Fragments n o v e l to HE4
are in d ic a te d below th e band*. The in te r p r e ta t io n o f
the n o v e l fragm ents i s a s fo l lo w s ( s e e F igu re C 4 .2 6 ).
TJ Hind I I I or Hamll I fragm ent c o n ta in in g th e ju n c tio nbetween H3V-2 Hind I I I o (HSV-2 BarnH I f ) and HSV-2 Hind III~ b ( HSV-2~BamH I iT "
x Iiind I I I or Bamii I term in a l fragm ent o f Lgen era ted by tEe tr a n s p o s it io n
x x w ith an a d d it io n a l a sequence
j Hind I I I or Bamh I j o in t fragm ents c o n ta in in g x ,gen erated by the tr a n s p o s it io n
TI R T2
S I
§ c D ETl R T2 TI R T2 Tl R T2 TI R T2
a bed H t -
>n
ATl R T2
SB
X
Xn
TJ
o
Tl R T2 Tl R T2 Tl R T2 Tl
I 5 i
ER T2
TJ
cr
f f

Figure C4.24
Location of the inserted DNA in RE4.
A M pBR322 H a e l l l markers1 HSV-2 BamHI i d ig e s te d w ith Hhal2 RE4 BamHT U n d ig e s te d w ith HEaT3 RE4 BlndllTZBamHI TJ d ig e s te T ”w ith Hhal
. 4 HSV-2 H in d III-o - d ig e s te d w ith Hhal5 HSV-2 BamHI i H ig ested w ith AIuTT6 RE4 BamHT. U n d ig e s te d w ith Alul7 RE4 Hind I IlZBamHI TJ d ig e s te d -w ith Alul8 HSV-2-Hindl l l o- d ig e s te d w ith A lul - - -
7 . 57* (upper) and 1 5 7 * (lo w er) po lyacry lam id e g e l s .
B 1 HSV-2 BamHI 1 d ig e s te d w ith Smal2 HE4 BamHI U n d ig e s te d w ith SmaT3 HE4 HindIIl7BamHI TJ d ig e s te d w ith Smal4 HSV-2 .BindI I I o, d ig e s te d w ith Smal .7 M pBR322’~nTnfI markers
2 $ a g arose g e l .
Marker s i z e s are in d ic a te d in bp.
TJ d e n o te s a r e s t r i c t i o n fragm ent c o n ta in in g th e ju n c tio n Eetween norm al HSV-2 and in s e r te d HSV-2 seq u en ces.The lo c a t io n s o f BamHI TJ and th e double d ig e s t fragm ent HindI I l 7 BamHI TJ are. shown in F igu re C4#26.
A M 1 2 3 4 5 6 7 8 M B 1 2 3 4 M
•—« H —
■ - #
- 4
-1631
• - 5 0 6
- 3 9 6- 3 4 4
• - 2 9 8
m -220

Figure 0 4 .2 5
Structures of the joints and termini o f RE4.
A 1 tsB BamH I k2 RE4 BamH I major j o in t3 t g l BamH I g
D ig e sted w ith S s t I ; 2/« agarose g e l .
B 1 tsB BamH I c[* 2 RE4 BamH I major sh o r t term inu s, i s o la t e d
from type 1 EcoR I &3 RE4 BamH I major sh o r t term in u s, i s o la t e d
from type 1 Hind I I I m
D ig e sted w ith S s t I ; 2)* agarose g e l .
C 1 tsB BamH I k“ 2 1124 BamH I major j o in t
3 t s l BamH I g4 t s l BamH I r3 RE4 BamH I n o v e l lo n g term inus6 RE4 Hind I I I n o v e l lo n g term inus
D ig e sted w ith Sma I ; 7#5‘* p o lyacry lam id e g e l .
D 1 t s B Bamii I k2 RE4 BamH I major j o in t3 t s l Bamii I g
D ig e sted w ith Sma I ; 15/* p o lyacry lam id e g e l .
E 1 RE4 BamH I type 1 lo n g term inus 2 tsB Damn I . s
D ig ested w ith Sma I ; 7 #5/* p o lyacry lam id e g e l .
B 1 tsB BamH I c[2 RE4 BamH I major sn o r t term inus
D ig e sted w ith Sma I ; 7 .5 ^ p o lyacry lam id e g e l .
S s t I or Sma I frag iaen ts o f the j o in t or term in a l fragm ents o f IiSV-1 ( l e f t ) and HSV-2 ( r ig h t ) are in d ic a te d . X d en o tes a genome term in a l fragm ent.
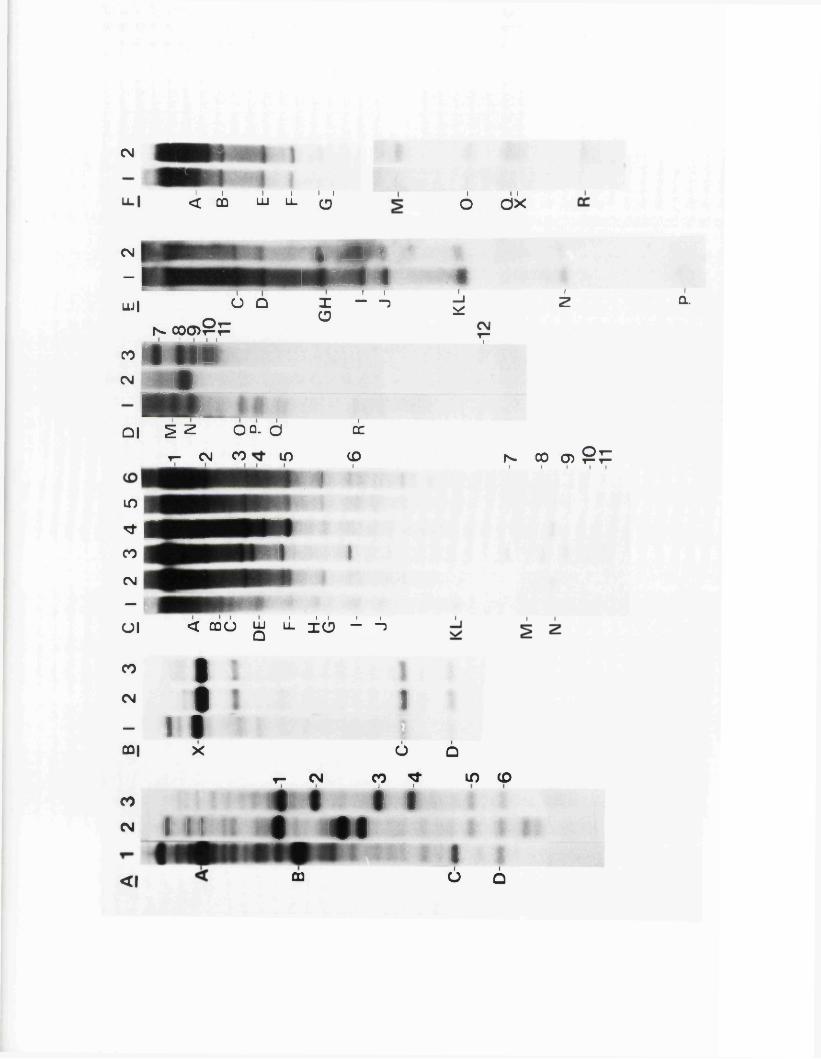
CM
i i i i i i< 0 0 LU U_ (J I i i
o a x
cm I 4* 1— | M 1 •UJ| 6 Q
S.COO)9^'_I 1 1« » * •« «
i i iI ---- 5CD KL
--1
2
N-
w mqi ^ z oq. a Icc
r - cm co in CDI -7 8 9 10
11
t f im^ I f l H H i B Mco ' 1 1cm t-
oi < coo g ii II 1 1I CD “ “5
« 1 I#- I
■ ■1a
- 1 iC 0 | X 6 io
« S™ I I ! |
CM 0 0 ^A A 1
• • l # «
5 6
i
iO.
< 1 cni i
O Q

figure C4.26
Genome structure of RE4.
■BamHI fragm ents p r e se n t in RE4 DHA are shown a t
th e top o f th e F ig u r e . B lach in d ic a t e s HSV-2 and w h ite
USV-1 seq u en ce . R egions o f u n c e r ta in ty o f c r o sso v e r
lo c a t io n are shown by h o r iz o n ta l h a tch in g and th e in s e r te d
HSV-2 sequence from i s denoted by c r o s s -h a tc h in g .
The n atu re o f th e a seq u en ces (HSV-1 or HSV-2: 1 or 2)
i s shown in s id e th e s t a r s . RE4 i s f ix e d in L and S in the
1,3 arrangem ent a s i l l u s t r a t e d .
The low er p a r t o f th e F igu re shows HSV-2 Hind iI I
and Hamill fragm en ts which are p r e se n t in th e r e g io n o f th e
in s e r t io n , v e r t i c a l arrows in d ic a t in g r e s t r i c t i o n s i t e s .
S iz e s in bp o f Sm al. Alul and Hhal fragm ents c o n ta in in g
the ju n c tio n between normal and in s e r te d ( 11 tra n sp o sed ” )
HSV-2 sequ en ces arc g iv e n . To th e r ig h t in p a re n th e se s
are g iv e n s i z e s in bp o f fragm ents produced by d ig e s t io n
o f th e H in d i! I / Bamiil fragm ent c o n ta in in g th e ju n c tio n
which were a ls o produced from HSV-2 BamHI i .
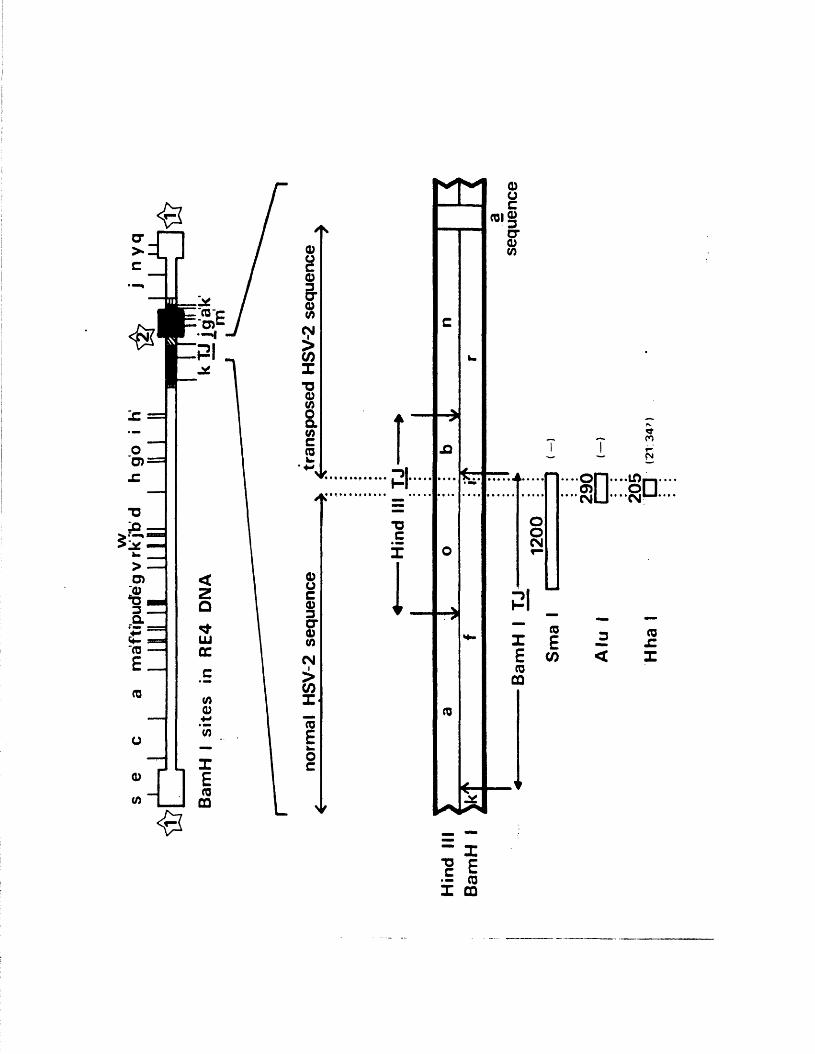
O)
'CO
Qg0&0CO
sC/>x“O0coa</>cCO
0)0 c 0) 3 O ’ 0) CO
CM
1x"coEoc
I3
“DC
JL
Ico
0 o c (01 §
O’0CO
ooCM
XECO
GO
coE
(/)3<
"" X? i•— co X CD
UfrC
-lZl
□ I
bm
h

Figure 04.27 (4 pages)
Models for the inversion of L and S in HSV PNA.
Model 1 shows s e p a r a tio n o f 1 and S, in l in e a r
or c ir c u la r form , and t h e ir su bsequent recom b in ation in the
a seq u en ces (Hayward e t a l . , 1975b; Share and Summers, 1 9 7 7 ) .
Model 2 shows in te r m o le c u la r r ecom bination o f
l in e a r or c ir c u la r m o lec u le s in a r e p e a t sequence to
p ro d u ce 'th e o th e r arrangem ents a f t e r a p p ro p ria te c le a v a g e
e v e n ts (Share and Summers, 1 9 7 7 ) .
Model 3A shows in tr a m o le c u la r recom b in ation
a f t e r MA s y n th e s is u s in g a s in g le stran d o f th e genome
a s tem p la te (Share and Summers, 1 9 7 7 ) .
Model 3B shows in tr a m o le c u la r recom b in ation
in l in e a r or c ir c u la r form in a r e p e a t sequence
(S h e ld r ic h and B e r th e lo t , 19745 Share and Summers, 1977;
Sm iley e t a Z ,, 1 9 8 0 ) .
Model 30 shows in tr a m o le c u la r recom b in ation
in v o lv in g r e p a ir o f a seq u en ces (Jacob and Roizman, 1979;
Roizman, 1 9 7 9 ) . B r ie f ly , the model in v o lv e s c ir c u la r i s a t io n
o f th e l in e a r DNA m olecu le by th e a c t io n o f an ex o n u c lea se
fo llo w e d by l i g a t i o n o f com plem entary s in g le stranded
te r m in i, DNA i s r e p l ic a t e d by a r o l l i n g c i r c l e mechanism
to g en era te con catem ers, which are c le a v ed in a lt e r n a te
j o in t r e g io n s to produce u n i t - 1 ength m o lecu les la c h in g

an a sequence a t one term in u s. The m iss in g a sequence
i s reg en era ted by "cop y in g 11 from the j o in t r e g io n .
At t h i s p o in t , n ic k in g a t th e j o in t a sequence and
branch m ig ra tio n c a u se s in v e r s io n o f th e segm ent to
produce th e P, Ig and 1 ^ arrangem ents. The Igj,
arrangem ent i s produced"by a second round o f ”
r e p l i c a t io n o f Ig or 1^, w ith con cu rren t in v e r s io n .
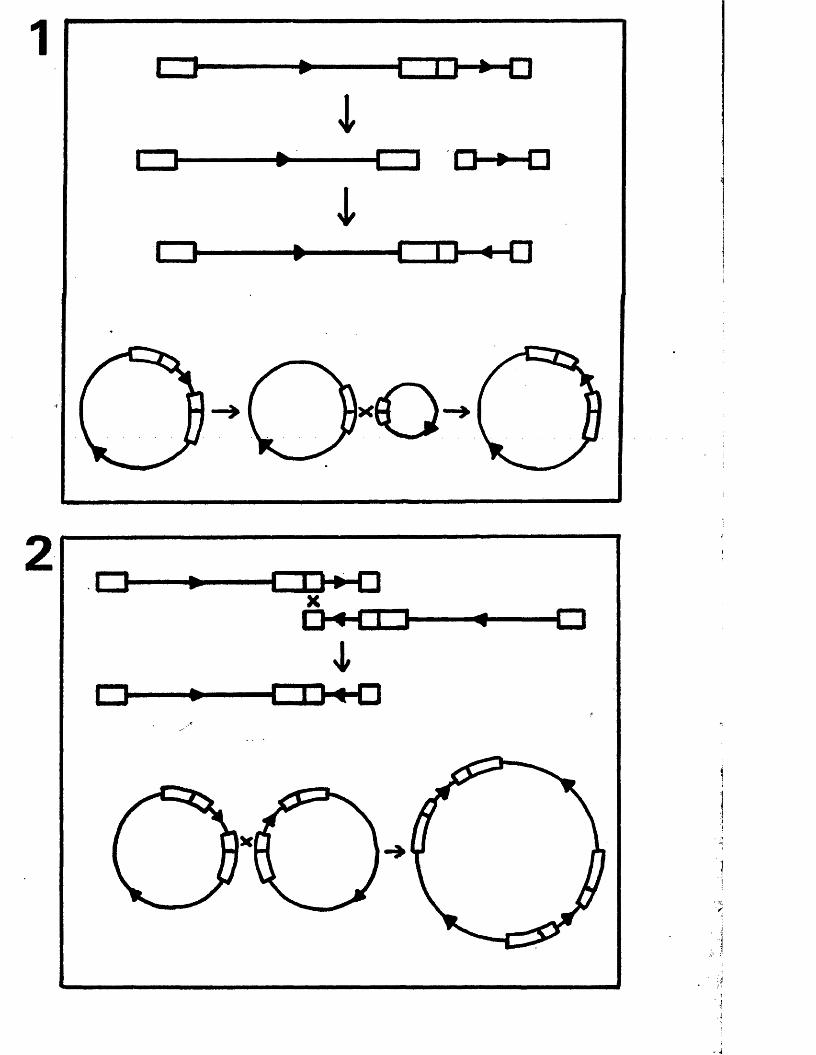
1
I— I-
I
I
(=3
C Z T J-M 3
d l O - ^ - D
o
2o cd-hd
CM-CO a
o
4m - « - n
o o
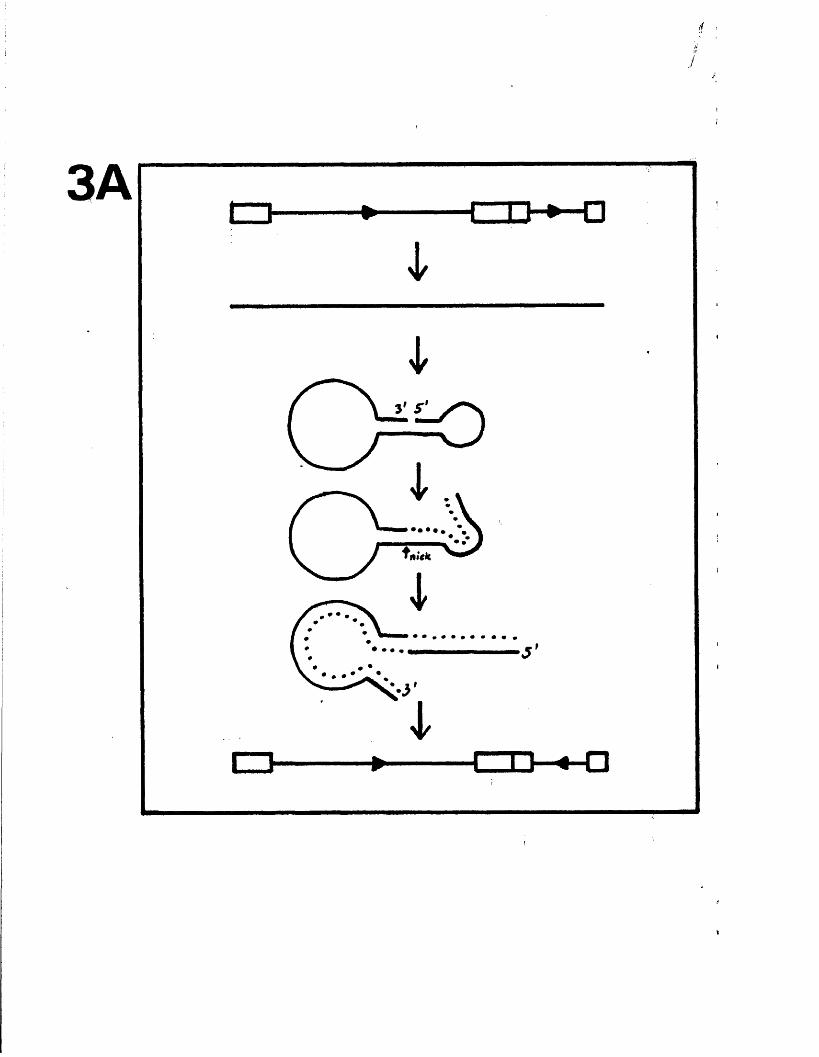
3Aa * cd-*-o
I
I
• • • •
I— V
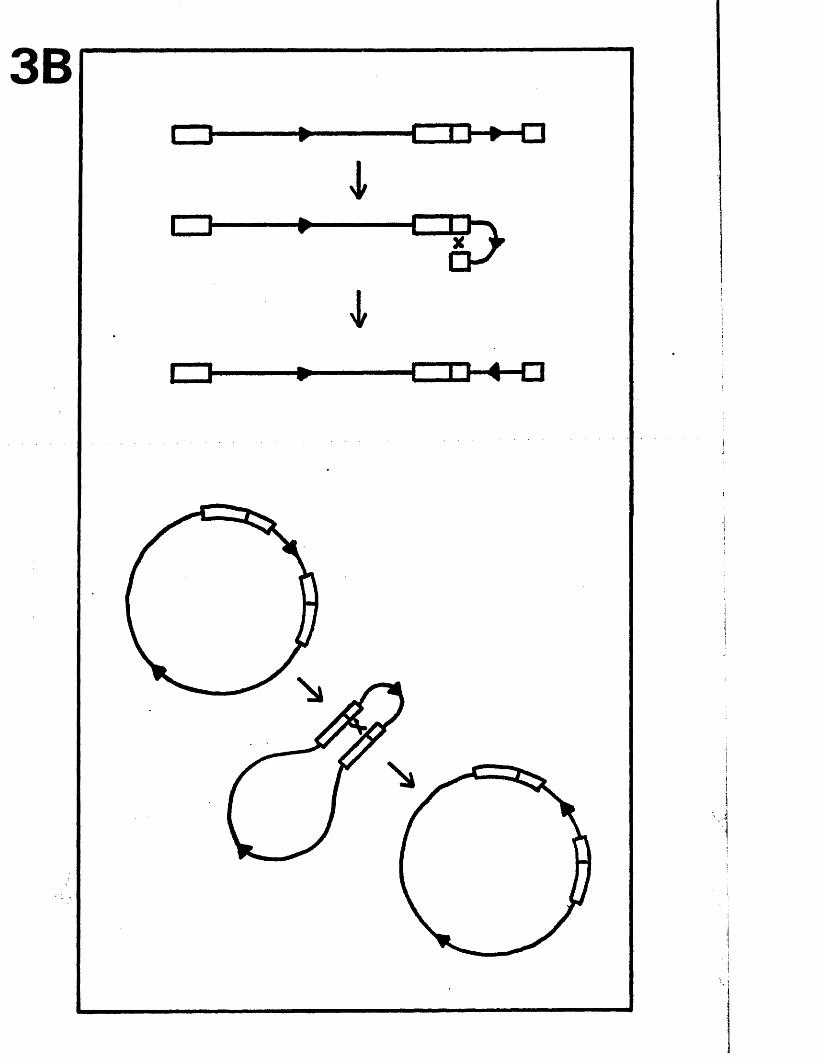
i
4
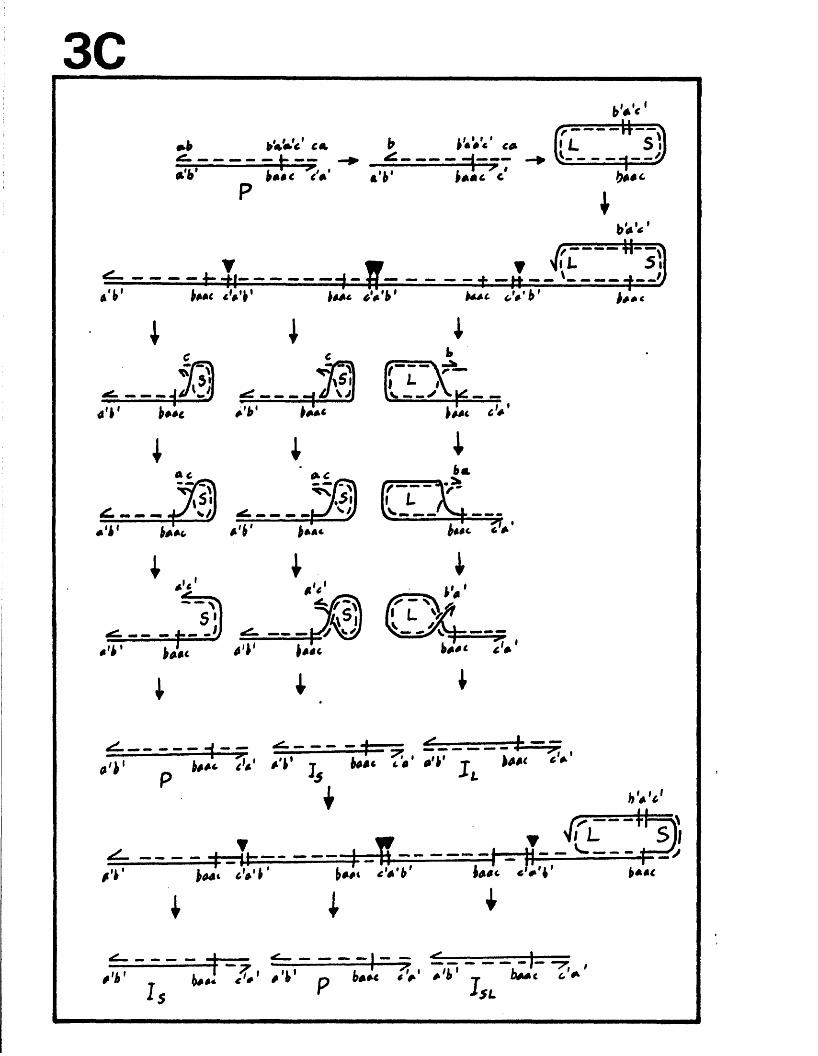
3C> — w—
It • » •
c
6*A t
5L

Comparison o f the e f f e c t s upon m aturation o f
u n it - le n g th genomes o f two mechanisms fo r g e n e ra tio n
o f c ir c u la r HSV DNA.
B lun t-en d l ig a t io n produces a c i r c l e w ith d is s im ila r
j o in t s , whereas complem entqry-end l i g a t i o n produces a
c i r c l e w ith id e n t i c a l j o i n t s . I n i t i a l progeny m o lecu les
may in v e r t n e i th e r , e i t h e r or both segm ents to produce
the fo u r c ir c u la r arrangem ents. In t h i s diagram model 3B
fo r s i t e - s p e c i f i c in tr a m o le c u la r recom b in ation h as been
invoked (F igu re C 4 .2 7 ). Four arrangem ents o f concatem ers
are then gen erated by r o l l i n g c i r c l e DHA s y n t h e s is .
C leavage may occu r a t any j o in t o f concatem ers
r e s u l t in g from com plem entary-end l i g a t i o n , so th a t each
concatem er arrangem ent produces two l in e a r genome
arrangem ents. C leavage cou ld r e s u l t frpm a s in g le
strand n ic k a t the ends o f the a seq u en ce , fo llo w e d by
r e p a ir o f th e s in g le stranded a sequence a t the term in i
o f u n it - le n g th m o le c u le s . I t i s n e c e ssa r y f o r o n ly one
segment to in v e r t to produce the fo u r genome arrangem ents,
and iso m e r isa t io n i s a fu n c t io n o f both in v e r s io n and
c le a v a g e .
C on verse ly , c le a v a g e may occur o n ly a t a l t e r n a te
j o in t s o f concatem ers r e s u l t in g from b lu n t-en d l i g a t i o n ,
and each concatem er produces o n ly one l in e a r genome
arrangem ent. C leavage in t h i s ca se cou ld r e s u l t frpm a
double stranded cu t between two a seq u en ces ( i . e . a t th e
j o in t which i s n o t c le a v ed in model 3C o f F igure C 4*27).
I t i s n e c essa ry fo r both segm ents to in v e r t to g en era te
the fou r arrangem ents. I so m e r isa t io n i s a fu n c t io n
only o f in v e r s io n . A lt e r n a t iv e ly , th e c lea v a g e
mechanism may r e c o g n ise both ty p e s p f j o i n t , and in t h i s
ca se the two m odels fo r c ir c u la r i s a t io n have id e n t i c a l
e f f e c t s upon m aturation ( i . e . th a t o f com plem entary-end l ig a t io n
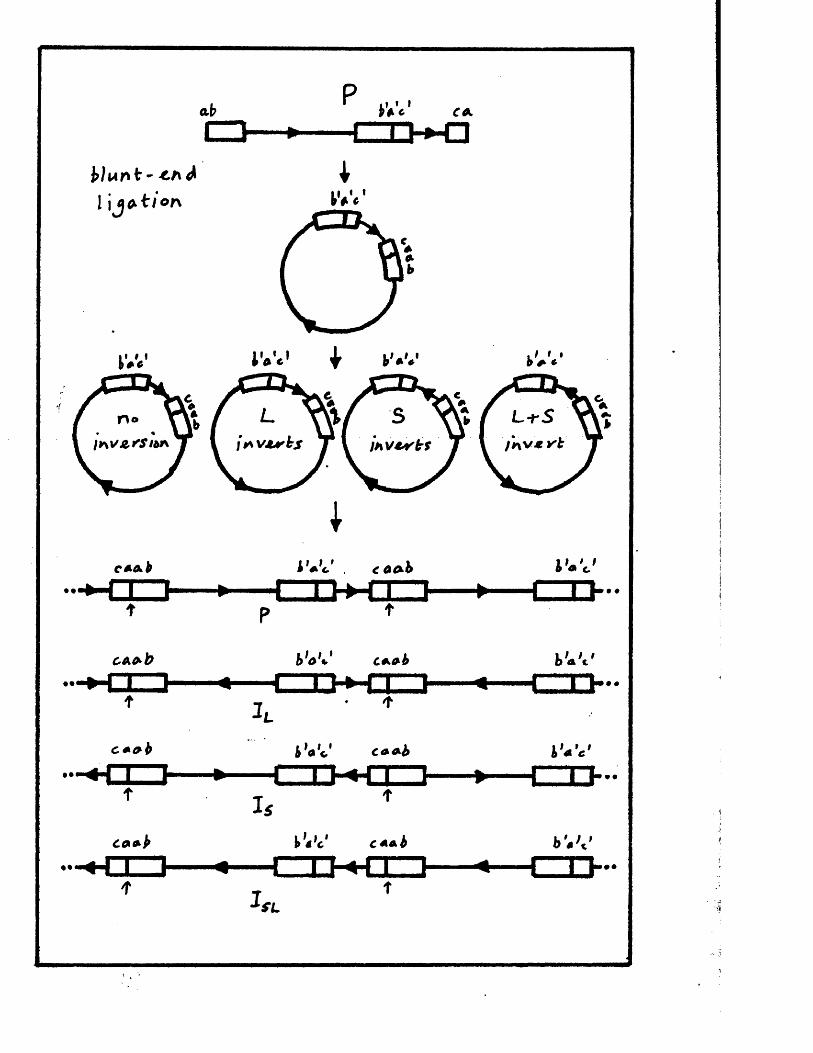
ab
( = }
• i » iP f i C C A
C Z O - H D
ILI 1 1 »# t
it # I ,/ # » b +■ *
, r
n o \ IM&rSti*
ie * * b
-* -C E 3t
c+ob~ T —
iV c ' <<IAi
- Q M Dp ♦
t V * 1 ca*£
Z D -H X I3 +
v * ' j
d D -
V c*&b i V e '
C Z Q -K IZ 3 — » 1— n -t
bVc* caa£ b # V
Z T H C E Z I -------- *— C H >
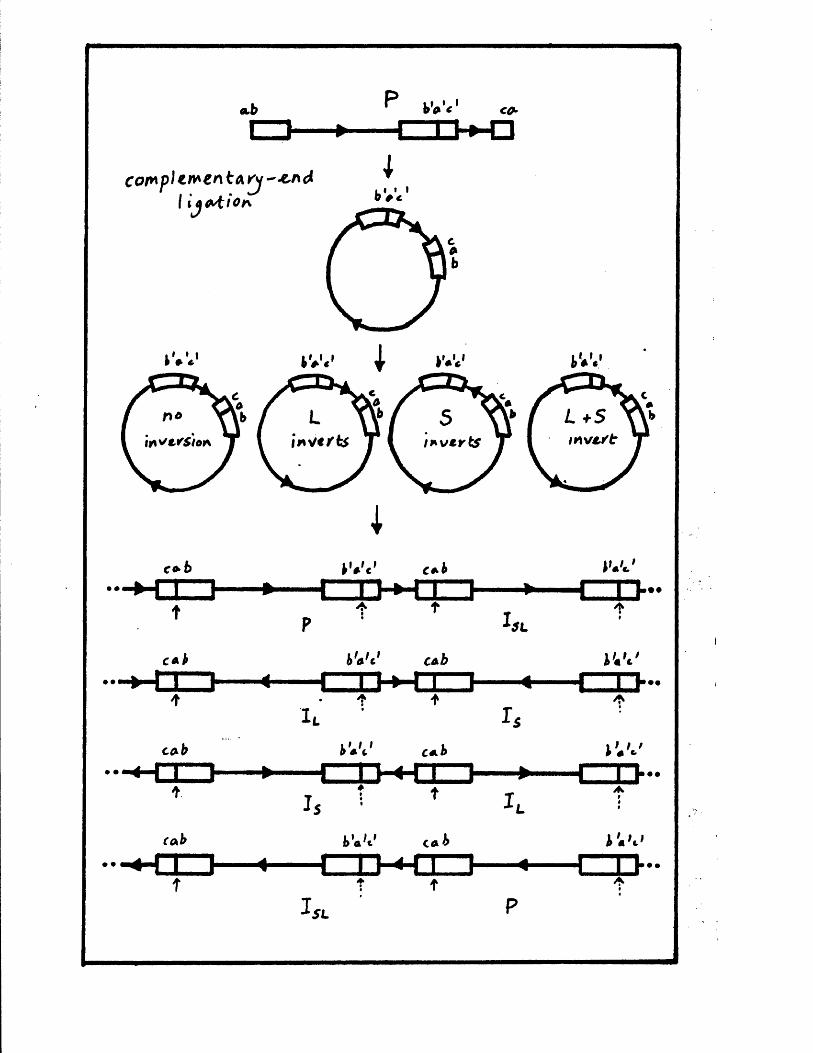
a.b
C D d W D
> V 1
4c a b
-H Z Z Dt
c a b
3 D
c a b
r v » - n I— *t - t
c a b
□ZD
bVe1 cab
4 C Z D -H mI t
b V c 1 c a b
Z T K C E
fab
□ZD
t I -r rJ 5 J-t
b W c a b
—CZE3-K3ZD---- *T t
l i t ' P
L*-U
mVC/fe m v t f bl a v e r t smy/efStoa,
J V c '
D D • •
i ; v
D D
I 1 I.' h a «-
C D
c d d

l l E J E E E N C i S

ALWINE, J .C . , STEINHART, W.L. and HILL, C.W. (1 9 7 4 ).
T r a n s c r ip t io n o f h e rp e s s im p le x v i r u s ty p e 1 ENA i n
n u c le i i s o l a t e d from i n f e c te d HEp-2 and KB c e l l s .
V iro lo g y 60, 302-307 .
ANDERSON, k .P . , STRINGER, J .R . , HOLLAND, L .E . and WAGNER, E .K .
(1 9 7 9 ). I s o l a t i o n and l o c a l i z a t i o n o f h e rp e s s im p lex
v i r u s ty p e 1 mRNA. J . V i r o l . £0 , 8 0 5 -8 2 0 .
ANDERSON, K .P ., COSTA, R .H ., HOLLAND, I .E . and WAGNER, E.K .
(1 9 8 0 a ) , C h a r a c te r iz a t io n o f h e rp e s s im p lex v i r u s type
1 RNA p r e s e n t i n th e a b sen c e o f de novo p r o t e in s y n th e s i s .
J . V i r o l . 5 4 , 9 -2 7 .
ANDERSON, K .P ., HOLLAND, L .E ., GAYLORD, B.H. and WAGNER, E .K .
(1 9 8 0 b ). I s o l a t i o n and t r a n s l a t i o n o f mRNA encoded by
a s p e c i f i c r e g io n o f th e h e rp e s s im p le x ty p e 1 genome.
J . V i r o l . 5 5 , 749.-759.
ANDERSON, K .P ., PRINK, R . J . , DEVI, G .B ., GAYLORD, B .H ., COSTA,
R.H, and WAGNER, E .K . (1 9 8 1 ) . D e ta i le d c h a r a c t e r i z a t i o n
o f th e mRNA m apping i n th e Hind fra g m e n t K r e g io n o f th e
h e rp e s s im p lex v i r u s ty p e 1 genome. J . V i r o l . 2 1 0 1 1 - 1 0 2 7 #
ARON, G.M., SCHAPPEN, P .A ., COURTNEY, R . J . , BENYESH-MEINICK, M.
and KIT, S. (1 9 7 3 ) . Thym idine k in a s e a c t i v i t y o f
h e rp e s s im p lex v i r u s t e m p e r a tu r e - s e n s i t iv e m u ta n ts .
I n t e r v i r o lo g y 1 , 9 6 -1 0 9 .
ARON, G . E . , PURIFOY, D .J . M . and SCHAPFER, P . A. (1 9 7 5 ) . DNA
syn th esis and DNA polymerase a c t iv i ty o f herpes simplex
v i r u s type 1 tem perature-3ensitive mutants.
J . V i r o l . 16 , 4 98 -507 .
ASHER, Y ., HELLER, M. and BECKER, Y. (1 9 6 9 ) . I n c o r p o r a t io n
o f l i p i d s i n to h e rp e s s im p lex v i r u s p a r t i c l e s .
J . g en . V i r o l . ± , 65 -76 .
ATKINSON, L .A ., BARR, S . and TB1BURY, W.C. (1 9 7 8 ) . The f i n e
s t r u c tu r e o f c e l l s i n f e c te d w ith t e m p e r a tu r e - s e n s i t iv e
m u tan ts o f h e rp e s sim p lex ty p e 2 . J . g e n . V i r o l . 40, 103-119

BACHENHEIMER, S .L . and ROIZMAN, B. (1 9 7 2 ) . R ib o n u c le ic a c id
s y n th e s i s i n c e l l s in f e c te d w ith h e rp e s s im p lex v i r u s .
V I. P o ly a d e n y lic a c id se q u e n c e s i n v i r a l m essenger
r ib o n u c le i c a c i d . J . V i r o l . 10 , 8 7 5 -8 7 9 .
BACHENHEIMER, S .L . , KIEFF, E .D ., LEE, L .F . and ROIZMAN, B. (1 9 7 2 ) .
C om para tive s t u d i e s o f DNAs o f M a re k 's d i s e a s e and
h e rp e s s im p le x v i r u s . In "O n co g en es is and H e rp e s v iru s e s " ,
PP. 7 4 -8 1 . E d ite d by P.M . B ig g s , G. de The and L .N .
P ay n e . IARC S c i e n t i f i c P u b l i c a t io n s , n o . 2 , Lyon.
BARAHONA, H ., MELENDEZ, L .V . and MELNICK, J .L . (1 9 7 4 ) . A
compendium o f h e r p e s v i r u s e s i s o l a t e d from non-hum an
p r im a te s . I n t e r v i r o lo g y 2 , 1 7 5 -1 9 2 .
BARINGER, J .R . and SWOVELAND, P . (1 9 7 3 ) . R ecovery o f h e rp e s
s im p le x v i r u s from human t r ig e m in a l g a n g l io n s . New
E ng land J o u rn a l o f M ed ic ine 288, 64 8 -6 5 0 .
BARTKOSKI, M. and ROIZMAN, B. (1 9 7 6 ) . RNA s y n th e s i s i n c e l l s
i n f e c t e d w ith h e rp e s s im p le x v i r u s . X I I I . D i f f e r e n c e s
i n th e m e th y la t io n p a t t e r n s o f v i r a l RNA d u r in g th e
r e p l i c a t i v e c y c le . J . V i r o l . 20 , 583- 5 8 8 .
BARTKOSKI, M .J. J r . and ROIZMAN, B. (1 9 7 8 ) . R e g u la tio n o f
h e r p e s v i r u s m ac ro m o le cu la r s y n t h e s i s . V II . I n h i b i t i o n
o f i n t e r n a l m e th y la t io n o f mRNA l a t e i n i n f e c t i o n .
V iro lo g y 8 2 , 1 4 6 -1 5 6 .
BAUCKE, R .B . and SPEAK, P .G . (1 9 7 9 ) . Membrane p r o t e in s
s p e c i f i e d by h e rp e s s im p le x v i r u s . V. I d e n t i f i c a t i o n
o f an P c -b in d in g g ly c o p r o te in . J . V i r o l . 2 7 7 9 - 7 8 9 .
BAYLISS, G .J . , UARSDEN, i i .S . and HAY, J . (1 9 7 5 ) . H erpes
s im p le x v i r u s p r o t e i n s : D N A -binding p r o t e i n s in
i n f e c t e d c e l l s and i n th e v i r u s s t r u c t u r e .
V iro lo g y 6 8 , 1 2 4 -1 3 4 .

BECKER, Y ., DYM, H. and SAROV, I . (1 9 6 8 ) . H erpes s im p lex
v i r u s DNA. V iro lo g y 2 6 , 1 8 4 -1 9 2 .
BECKER, Y. and ASHER, Y. (1 9 7 5 ) . In v i t r o s y n th e s i s o f DNA
i n n u c le i i s o l a t e d from h e rp e s s im p le x v i r u s - i n f e c t e d
c e l l s , u n t r e a te d and t r e a t e d w ith m e ta b o lic i n h i b i t o r s .
V iro lo g y 62> 2 0 9 -2 2 0 .
BECKER, Y ., ASHER, Y ., WEINBERG-ZAHLERING, E . , RABKIN, S . ,
FRIEDMANN, A. and KESSLER, E .~ (1 9 7 8 ) . D e fe c tiv e h e rp e s
s im p lex v i r u s DNA: c i r c u l a r and c i r c u l a r - l i n e a r m o le c u le s
re s e m b lin g r o l l i n g c i r c l e s . J . g e n . V i r o l , 42. > 319-335*
BECKER, Y ., CHEJANOVSKY, N. and FRIDLENDER, B. (1 9 8 0 ).
DNA-binding p r o t e i n s i n th e n u c le i o f h e rp e s s im p lex
v i r u s - i n f e c t e d , a r g in in e - d e p r iv e d , BSC-1 c e l l s .
J . g en . V i r o l . 2 0 , 2 5 9 -2 7 8 .
BELL, D. (1 9 7 4 ). DNA s y n th e s i s i n ’n u c le a r m o n o la y e rs ’ from
BSC-1 c e l l s i n f e c t e d w ith h e rp e s v i r u s .
N a tu re (London) 2 4 8 . 505-508 .
BEN01ST, C ., O’HARE, K ., BREATHNACH, R. and-CHAMBON, P . (1 9 8 0 ) .
The ovalbum in gene - seq u en ce o f p u t a t i v e c o n t r o l
r e g io n s . N u c le ic A cids R e se a rc h 8 , 1 2 7 -1 4 2 .
BEN-PORAT, T. and KAPLAN, A .S . (1 9 6 2 ) . The c h e m ic a l c o m p o s itio n
o f h e rp e s s im p lex and p s e u d o ra b ie s v i r u s e s .
V iro lo g y 16 , 26 1 -2 6 6 .
BEN-PORAT, T. and KAPLAN, A .S . (1 9 6 3 ) . The s y n th e s i s and f a t e
o f p s e u d o ra b ie s v i r u s DNA i n i n f e c t e d mammalian c e l l s
i n th e s t a t i o n a r y p h ase o f g ro w th . V iro lo g y 20 , 31 0 -3 1 7 .
BEN-PORAT, T ., DEMARCHI, J.M . and KAPLAN, A .S . (1 9 7 4 ) .
C h a r a c te r iz a t io n o f d e f e c t iv e i n t e r f e r i n g v i r a l p a r t i c l e s
p r e s e n t i n a p o p u la t io n o f p s e u d o ra b ie s v i r i o n s .
V iro lo g y 61, 2 9 -3 7 .

BEN-BOBAT, T ., S'i'EHN, B. and KAPLAN, A .S . (1 9 7 6 a ) . F a te o f
p a r e n t a l h e r p e s v i r u s DNA. V iro lo g y 71 , 41 2 -4 2 2 .
BEN-POHAi1, T . , KAPLAN, A .S .,' STEM, B. and HUBENSl’EIN, A .S . (1976b)
C o n ca tem eric fo rm s o f i n t r a c e l l u l a r h e r p e s v i r u s DNA.
V iro lo g y 69 , 547 - 5 6 0 .
BEN-PORAT, T. and TOKAZEWSKI, S .A . (1 9 7 7 ) . R e p l ic a t io n o f
h e r p e s v i r u s DNA. I I . S e d im e n ta tio n c h a r a c t e r i s t i c s o f
new ly s y n th e s iz e d DNA. V iro lo g y 1 9 , 2 9 2 -3 0 1 .
BEN-PORAT, T ., BLANKENSHIP, M .L ., DEMARCHI, J.M . and KAPLAN,
A .S . (1 9 7 7 ) . R e p l ic a t io n o f h e r p e s v i r u s DNA. I I I .
R a te o f DNA e lo n g a t io n . J . V i r o l . 2 2 , 7 3 4 -7 4 1 .
BEN-PORAT, T . and RIXON, P . J . (1 9 7 9 ) . R e p l i c a t io n o f h e rp e s v i r u s
DNA. IV . A n a ly s is o f c o n c a te m e rs . V iro lo g y 6 1 -7 0 .
BEN-PORAT, T ., RIXON, P . J . and BLANKENSHIP, M.L. (1 9 7 9 ) .
A n a ly s is o f th e s t r u c t u r e o f th e genome o f p s e u d o ra b ie s
v i r u s . V iro lo g y 92 , 285 -294 .
BEN-PORAT, T. and VEACH, R.A. (1 9 8 0 ) . O r ig in o f r e p l i c a t i o n
o f th e DNA o f a h e r p e s v i r u s ( p s e u d o r a b ie s ) .
P ro c . N a t l . A cad. S c i . USA H , 1 7 2 -1 7 5 .
BEN-PORAT, T . , VEACH, R .A . and LADIN, B .P . (1 9 8 0 ) . R e p l i c a t io n
o f h e r p e s v i r u s DNA. VI. V ir io n s c o n ta in in g e i t h e r
iso m e r o f p s e u d o ra b ie s v i r q s DNA a r e i n f e c t i o u s .
V iro lo g y lu 2 , 370- 3 8 0 .
BeN YBSH-i.iELN IC K, ivi. , SCHAFFER, P . A ., COURTNEY, R . J . , ESPARZA, J .
and KIMURA, S. (1 9 7 4 ) . V ir a l gene f u n c t io n s e x p re s s e d
and d e te c te d by t e m p e r a tu r e - s e n s i t iv e m u ta n ts o f h e rp e s
s im p le x v i r u s . C old S p rin g H arb o r Symp. Q uan t. B io l .
29 , 7 3 1 -7 4 6 .
BEN-ZEEV, A. and DECKER, Y. (1 9 7 5 ). S ym m etrica l t r a n s c r i p t i o n
o f h e rp e s s im p le x v i r u s DNA i n i n f e c t e d BSC-1 c e l l s .
N a tu re (London) 2 5 4 , 719-722 .

BEN-ZEEV, A ., ASHER, I . and BECKER, Y. (1 9 7 6 ) . S y n th e s is o f
h e rp e s s im p le x v i r u s - s p e c i f i e d RNA by an RNA p o ly m erase I I
i n i s o l a t e d n u c le i i n v i t r o . V iro lo g y 2 1 , 3 0 2 -311 .
BEN-ZEEV, A. and BECKER, Y. (1 9 7 7 ). R equ irem en t o f h o s t c e l l
RNA p o ly m e rase I I i n th e r e p l i c a t i o n o f h e rp e s s im p lex
v i r u s i n o ( -a m a n it in ^ s e n s it iv e and - r e s i s t a n t c e l l l i n e s .
V iro lo g y £ 6 , 2 4 6 -2 5 3 .
BERK, A .J . and SHARP, P .A . (1 9 7 7 ). S iz in g and m apping o f
e a r l y a d e n o v iru s mRNAs by g e l e l e c t r o p h o r e s i s o f S I
e n d o n u c le a s e -d ig e s te d h y b r id s . C e l l 1 2 , 72 1 -7 3 2 .
BIEGELEISEN, K. and RUSH, M.G. (1 9 7 6 ) . A s s o c ia t io n o f h e rp e s
s im p le x v i r u s ty p e 1 DNA w ith h o s t chrom osom al DNA d u r in g
p r o d u c t iv e i n f e c t i o n . V iro lo g y 69» 2 4 6 -2 5 7 .
BIEGELEISEN, K ., YANAGI, K. and RUSH, M.G. (1 9 7 7 ) . F u r th e r
s t u d i e s on th e a s s o c i a t io n o f h e rp e s s im p lex v i r u s ty p e 1
DNA w ith h o s t DNA d u r in g p ro d u c t iv e i n f e c t i o n .
V iro lo g y 8 2 , 22 1 -2 2 5 .
BISY/AL, N. and MURRAY, B.K. (1 9 7 4 ). S y n th e s is o f h e rp e s
s im p le x v i r u s ty p e i (HSV-1) DNA i n i s o l a t e d n u c l e i .
I . C o n d it io n s f o r i s o l a t i o n o f n u c le i c a p a b le o f
v i r a l DNA s y n t h e s i s . In t e r v i r o lo g y 4 , 1 -1 3 .
BISWAL, N ., MURRAY, B.K. and BENYESH-MELNICK, M. (1 9 7 4 ) .
R ib o n u c le o t id e s i n new ly s y n th e s iz e d DNA o f h e rp e s
s im p le x v i r u s . V iro lo g y 61, 8 7 -9 9 .
B10MBERG, J . (1 9 7 9 ) . S tu d ie s on th e a t ta c h m e n t o f h e rp e s
s im p le x v i r u s . E f f e c t o f t r y p s in t r e a tm e n t .
A rch . V i r o l . 61, 2 01 -206 .
BLUE, W.T. and STOBBS, D.G. (1 9 8 1 ). I s o l a t i o n o f a p r o t e in
k in a s e in d u c e d by h e rp e s s im p lex v i r u s ty p e 1 .
J. Virol. 28, 383-388.

BOLDEN, A ., AUCKEN, J . and iVEISSBACH, A. (1 9 7 5 ) . S y n th e s is o f
h e rp e s s im p le x v i r u s , v a c c in ia v i r u s and a d e n o v iru s DMA
i n i s o l a t e d HeLa c e l l n u c l e i . I . E f f e c t o f v i r a l -
s p e c i f i c a n t i s e r a and p h o sp h o n o a c e tic a c id .
J . V i r o l . 16 , 1 5 8 4 -1592 .
BONE, D .R ., BROWN, M ., CROMBIE, I . and FRANCKE, B. (1 9 7 8 ) .
V i r a l DNA s y n th e s i s i n c e l l s i n f e c t e d w ith te m p e ra tu re -
s e n s i t i v e m u ta n ts o f h e rp e s s im p le x ty p e 1 .
J . V i r o l . 2 8 , 1 4 -1 9 .
BOOKQUT, J . , 11IRSCH, I , , PURIFOY, D .J.M . and BISWAL, N. (1 9 7 9 ) .
H erpes s im p le x v i r u s ty p e s 1 and 2 : com parison o f th e
d e f e c t iv e genomes and v i r u s s p e c i f i c p o ly p e p t id e s .
V iro lo g y 23., 5 9 8 -6 0 4 .
BOOKOUT, J .B . and LEVY, C .C . (1 9 8 0 ) . C om para tive e x a m in a tio nr
o f th e p o ly p e p t id e s o f h e rp e s s im p le x v i r u s : ty p e s 1
and 2 . V iro lo g y 1 0 1 , 1 9 8 -2 1 6 .
BORNKAMM, G.W., DELIUS, H ., FLECK12NSTEIN, B . , WERNER, F . J . and
MULDER, C. (1 9 7 6 ) . S t r u c tu r e o f h e r p e s v i r u s s a i m i r i
genom es: a rra n g e m e n t o f heavy and l i g h t se q u en c es i n th e
M genome. J . V i r o l . 1 9 , 1 54 -161 .
BORNKAMM, G.W., DELIUS, H ., Z ImBER, U ., HUDEWENTZ, J . and
EPSTEIN, M.A. (1 9 8 0 ) . Com parison o f E p s te in - B a r r
v i r u s s t r a i n s o f d i f f e r e n t o r i g in by a n a l y s i s o f th e
v i r a l DNAs. J . V i r o l . 605 -618 . »
BOYER, H.W. and ROULLAND-DUSSOIX, D. (1 9 6 9 ) . A c o m p lem en ta tio n
a n a l y s i s o f th e r e s t r i c t i o n and m o d if ic a t io n o f DNA in
E s c h e r ic h ia c o l i . J . m ol. B io l . ^ 1 , 459-472 .
BRONSON, D .L ., GRAHAM, B . J . , LUDWIG, H ., BENYESH-MELNICK, M.
and BISWAL, N. (1 9 7 2 ) . S tu d ie s on th e r e l a t e d n e s s o f
h e rp e s v i r u s e s th ro u g h DNA-RNA h y b r i d i s a t i o n .
B ioch im . laiophys. A c ta 259 . 2 4 -3 4 .

BRONSON, D .L ., DREESMAN, O .R ., BISWAL, N. and BENYESH-MELNICK, M.
(1973)* D e fe c tiv e v i r i o n s o f h e rp e s s im p lex v i r u s ,
i n t e r v i r o l o g y 1 , 1 4 1 -1 5 3 .
BROWN, S .M ., RITCHIE, D.A. and SUBAK-SHARPE, J .H . (1 9 7 3 ).
G e n e tic s t u d i e s w ith h e rp e s s im p le x ty p e 1 . The
i s o l a t i o n o f te m p e ra tu re s e n s i t i v e m u ta n ts , t h e i r
o r d e r in g in to co m p lem en ta tio n g ro u p s and recoin b in a t io n a l
a n a l y s i s l e a d in g to a l in k a g e map. J . g e n . V i r o l . 1 8 ,
3 29 -346 .
BROWN, S.M. and RITCHIE, D.A. (1 9 7 5 ) . G e n e tic s t u d i e s w ith
h e rp e s s im p le x v i r u s ty p e 1 . A n a ly s is o f m ixed p la q u e -
fo rm in g v i r u s and i t s b e a r in g on g e n e t ic r e c o m b in a t io n .
V iro lo g y 64 , 3 2 -4 2 .
BROWN, S.M. and JAMIESON, A .T. (1 9 7 7 ) . L o c a tio n o f non
te m p e r a tu r e - s e n s i t iv e g e n e 3 on th e g e n e t ic map o f
h e rp e s s im p lex v i r u s ty p e 1 . In 11 Oncogene s i s and
H e rp e s v iru s e s 111" , p a r t I , p p . 3 5 -3 9 . E d ite d by
G. de The, W. H enle and F. R app. IARC S c i e n t i f i c
P u b l i c a t io n s , n o . 24» Lyon.
BUCHAN, A. and WATSON, D.H. (1 9 6 9 ) . The im m u n o lo g ica l
s p e c i f i c i t y o f th y m id in e k in a s e s i n c e l l s in f e c te d
by v i r u s e s o f th e h e rp e s g ro u p . J . g e n . V i r o l . £ , 46 1 -4 6 3 .
BUCHAN, A ., WATSON, D .H ., DUBBS, D.R. and KIT, S . (1 9 7 0 ) .
S e r o lo g ic a l s tu d y o f a m utan t h e r p e s v i r u s u n a b le to
s t im u la te th y m id in e k in a s e . J . V i r o l . £ , 8 1 7 -8 1 8 .
BUCHMAN, T.G. and ROIZMAN, B. (1 9 7 8 a ). Anatomy o f bovine
m a m ra ilitis v i r u s DNA. I . R e s t r i c t i o n e n d o n u c le a se
maps o f f o u r p o p u la t io n s o f m o le c u le s t h a t d i f f e r i n
th e r e l a t i v e o r i e n t a t i o n o f t h e i r lo n g and s h o r t
components. J. Virol. 2j>, 395-407.

BUCHMAN, T.G . and RQIZMAN, B. (1 9 7 8 b ). Anatomy o f bov ine
m a m m ilitis v i r u s DNA. I I . S iz e and a rra n g e m e n ts o f
th e d e o x y n u c le o tid e s e q u e n c e s . J . V ir o l . 22 , 2 3 9 -2 5 4 .
BUCHMAN, T . G . , ROIZMAN, B ., ADAIRS, G. and STOVER, H. (1 9 7 8 ) .
R e s t r i c t i o n e n d o n u c le a se f i n g e r p r i n t i n g o f h e rp e s s im p le x
DNA: a n o v e l e p id e m io lo g ic a l t o o l a p p l ie d to a n o so c o m ia l
o u tb r e a k . J . i n f e c t . D is . 1 3 8 », 488-498 .
BUCHMAN, T .G ., SIMPSON, T . , NOSAL, C ., ROIZMAN, B. and
NAHMIAS, A. (1 9 8 0 ) . The s t r u c t u r e o f h e rp e 3 s im p lex
v i r u s DNA and i t s a p p l i c a t i o n to m o le c u la r e p id e m io lo g y .
A nnals o f New York A cad. S c i . 354 , 27 9 -2 9 0 .
BUSBY, D.W .G., HOUSE, W. and MacDONALD," J .R . (1 9 6 4 ) . In
" V i r o lo g ic a l T e c h n iq u e s" , C h u r c h i l l , London.
BZIK, D .J . and PERSON, S . (1 9 8 1 ) . D ependence o f h e rp e s s im p le x
v i r u s ty p e 1 - in d u c e d c e l l f u s io n on c e l l ty p e .
V iro lo g y 110 , 3 5 -4 2 .

CABRAL, G.A. and. SCHAFFER, P .A . (1 9 7 6 ) . E le c t r o n m icro sco p e
s t u d i e s o f t e m p e r a tu r e - s e n s i t iv e m u ta n ts o f h e rp e s
s im p le x v i r u s ty p e 2 . J . V i r o l . 18 , 7 2 7 -7 3 7 .
CABRAL, G .A ., COURTNEY, R . J . , SCHAFFER, P .A . and MARCIANO-
CABRA1, F. (1 9 8 0 ) . U l t r a s t r u c t u r a l c h a r a c t e r i z a t i o n o f
an e a r l y , n o n - s t r u c t u r a l p o ly p e p t id e o f h e rp e s s im p lex
v i r u s ty p e 1 . J . V i r o l . 2 1 , 1 1 9 2 -1 1 9 8 .
CAMACHO, A. and SPEAR, P .O . (1 9 7 8 ) . T ra n s fo rm a tio n o f h a m ste r
embryo f i b r o b l a s t s by a s p e c i f i c f ra g m e n t o f th e h e rp e s
s im p le x v i r u s genome. C e ll 1£ , 9 9 3 -1 0 0 2 .
CATCHESIDE, D.G. and CORCORAN, D. (1 9 7 3 ) . C o n tro l o f n o n - •
a l l e l i c re c o m b in a tio n i n N e u ro sn o ra c r a s s a .
A u s t r a l ia n J . b i o l . S c i . 26 , 1 3 3 7 -1353 .
CHAI, L . (1 9 7 1 ) . A r c h i te c tu r e o f th e DNA m o le c u le w ith in
E p s te in -B a r r v i r u s (EBV). P ro c . Amer. A ssoc , f o r
C ancer R es. 12 , a b s t r a c t n o . 7 1 .
CHAN, T -S . (1 9 7 7 ). In d u c t io n o f d e o x y c y tid in e deam inase
a c t i v i t y i n mammalian c e l l l i n e s by i n f e c t i o n w ith
h e rp e s s im p lex v i r u s ty p e 1 . P ro c . N a t l . Acad. S c i .
USA 1734-1738 .
CHARTRAND, P . , STOW, N .D ., TIMBURY, M.C. and WILKIE, N.M. (1979)
P h y s ic a l m apping o f p aa r m u ta t io n s o f h e rp e s s im p lex
v i r u s ty p e 1 and 2 by i n t e r t y p i c m ark e r r e s c u e .
J . V i r o l . 2 i , 26 5 -2 7 6 .
CHARTRAND, P . , CRUMPACKER, C .S . , SCHAFFER, P .A . and WILKIE, N.M.
(1 9 8 0 ) . P h y s ic a l and g e n e t ic a n a l y s i s o f th e h e rp e s
s im p le x v i r u s DNA p o ly m erase lo c u s . V iro lo g y 1 0 3 , 311-326
CHARTRAND, P . , WILKIE, N.M. and TIMBURY, M.C. (1 9 8 1 ) .
P h y s ic a l m apping o f t e m p e r a tu r e - s e n s i t iv e m u ta t io n s o f
h e rp e s s im p lex v i r u s ty p e 2 by m arker r e s c u e .
J . g e n . V i r o l . £2 , 121-133 .

CHOUSTERMAN, S . , LACASA, M. and SHELDRICK, P . (1 9 7 9 ) .
P h y s ic a l map o f th e c h a n n e l c a t f i s h v i r u s genome:
l o c a t i o n o f s i t e s f o r r e s t r i c t i o n e n d o n u c le a se s EcoRI ,
H in d I I I . Hpal and X b a l. J . V i r o l . 21 , 7 3 -8 5 .
CHU, C -T ., PARRIS, D .S ., DIXON, R .A .P . , PARSER, P .E . and
SCHAFFER, P .A . (1 9 7 9 ) . H ydroxylam ine m u ta g e n e s is o f
HSV DNA and DNA f ra g m e n ts : i n t r o d u c t i o n o f m u ta t io n s
i n t o s e l e c te d r e g io n s o f th e v i r a l genome.
V iro lo g y 28 , 1 6 8 -1 8 1 .
CHURCHILL, A .E. and BIGGS, P.M. (1 9 6 7 ) . A gent o f M a re k 's
d i s e a s e i n t i s s u e c u l t u r e . N a tu re (London) 2 1 5 , 52 8 -5 3 0 .
CLEMENTS, J . B . , CORTINI, R. and WILKIE, n Tm. (1 9 7 6 ) . A n a ly s is
o f h e r p e s v i r u s DNA s u b s t r u c tu r e by means o f r e s t r i c t i o n
e n d o n u c le a s e s , J . g e n . V ir o l . 2 4 3 -2 5 6 .
CLEMENTS, J . B . , WATSON, R .J . and WILKIE, N.M. (1 9 7 7 ) .
Tem poral r e g u l a t i o n o f h e rp e s s im p lex v i r u s ty p e 1
t r a n s c r i p t i o n : l o c a t i o n o f t r a n s c r i p t s on th e v i r a l
genome. C e l l 1 2 , 2 7 5 -2 8 5 .
CLEMENTS, J . B . , McLAUCHLAN, J . and McGEOCH, D .J . (1 9 7 9 ) .
O r i e n t a t i o n o f h e rp e s s im p lex v i r u s ty p e 1 im m ed ia te
e a r ly rnRNAs. N u c le ic A cids R e se a rc h 2» 7 7 -9 1 .
CLEWELL, D.B. and HELINSICI, D.R. (1 9 7 0 ) . P r o p e r t i e s o f a
s u p e r c o i le d d e o x y r ib o n u c le ic a c i d - p r o t e i n r e l a x a t i o n
com plex and s t r a n d s p e c i f i c i t y o f th e r e l a x a t i o n e v e n t .
B io c h e m is try 2.> 4428-4440 .
cohen , g . h . , vaughan', r . k . and L aw rence, w .c . (1971 ) .
D e o x y r ib o n u c le ic a c id s y n th e s i s i n sy n c h ro n iz e d
mammalian KB c e l l s i n f e c te d w ith h e rp e s s im p le x v i r u s .
. J. Virol. 2, 783-791.

COHEN, G .H ., KATZE, M., HYDREAN-STERN, C. and EISENBERG, R .J .
(1 9 7 8 ) . Tyije-common CP-1 a n t ig e n o f h e rp e s s im p lex
v i r u s i s a s s o c ia t e d w ith a 5 9 ,0 0 0 -m o le c u la r w e ig h t
e n v e lo p e g ly c o p r o te in . J . V i r o l . 22, 1 7 2 -1 8 1 .
COHEN, G .H ., LONG, D. and EISENBERG, R .J . (1 9 8 0 a ) . S y n th e s is
and p r o c e s s in g o f g ly c o p r o te in s gD and gC o f h e rp e s
s im p le x v i r u s ty p e 1 . J . V i r o l . ^ 6 , 42 9 -4 3 9 .
COHEM, G .H ., POMCE HE LEON, M., BIGGELM ANN, H ., LAWRENCE, W.C.,
VERHON, 3 .K . and EISENHERG, R .J . (1 9 8 0 6 ). S t r u c t u r a l
a n a l y s i s o f th e c a p s id p o ly p e p t id e s o f h e rp e s s im p lex
v i r u s ty p e s 1 and 2 . J . V i r o l . 24* 521-531
COHEN, S .N ., CHANG, A.C.Y. and HSU, L. (1 9 7 2 ) . Nonchromosomal
a n t i b i o t i c r e s i s t a n c e i n b a c t e r i a : g e n e t ic t r a n s f o r m a t io n
o f E s c h e r i c h ia c o l i by R - f a c to r DNA.- ^
P ro c . N a t l . A cad.' S c i . USA 6g, 2110-2114 .
COLBERE-GARAPIN, F . , C IiOUS TERM AN, S . , HQRODNICEANU, F . ,
KOURILSKY, P . and GARAPIN, A-C. (1 9 7 9 ) . C lo n in g o f th e
a c t i v e th y m id in e k in a s e gene o f h e rp e s s im p le x v i r u s
ty p e 1 i n E s c h e r ic h ia c o l i K -12.
P ro c . N a t l . Acad. S c i . USA 2 § , 3755-3759*
CONLEY, A .J . , KNIPE, D.M ., JONES, P . and ROIZMAN, B. (1 9 8 1 ) .
M o le c u la r g e n e t ic s o f h e rp e s s im p lex v i r u s . V II .
C h a r a c te r iz a t i o n o f a t e m p e r a tu r e - s e n s i t iv e m u tan t
p ro d u ced by in v i t r o m u ta g e n e s is and d e f e c t iv e i n DNA
s y n th e s i s and a c c u m u la tio n o f y p o ly p e p t id e s .
J . V i r o l . 32* 19 1 -2 0 6 .
COOPER, G .E. (1 9 7 3 ) . P h o s p h o ry la t io n o f 5 -b ro m o d eo x y c y tid in e
i n c e l l s i n f e c te d w ith h e rp e s s im p lex v i r u s .
P ro c . N a t l . Acad. S c i . USA 20 , 3788-3792 .
CORTINI, R. and WILKIE, N.M. (1 9 7 8 ). P h y s ic a l maps f o r HSV
ty p e 2 DNA w ith f i v e r e s t r i c t i o n e n d o n u c le a se s .J. g e n . ’V ir o l . 32* 259-280 .

COSTA, R .I I . , DEVI, E .G ., ANDERSON, K .P ., GAYLORD, B.H. and
WAGNER, E .K . (1 9 8 1 ) . C h a r a c te r iz a t i o n o f a m a^or l a t e
h e rp e s s im p lex v i r u s ty p e 1 mRNA. J . V i r o l . 2 8 , 483-496
COSTANZO, P . , CALPADELLI-PIUME, G ., POA-TOMASI, L . and
CASSAI, E. (1 9 7 7 ) . E v id en ce t h a t h e rp e s s im p le x v i r u s
DNA i s t r a n s c r i b e d by c e l l u l a r RNA p o ly m erase B.
J . V i r o l . 21 , 9 9 6 -1 0 0 1 .
COURTNEX, R .J . and POWELL, K .L . (1 9 7 5 ) . Im m uno log ica l and
b io c h e m ic a l c h a r a c t e r i z a t i o n o f p o ly p e p t id e s in d u ce d
by h e rp e s s im p le x v i r u s ty p e 1 and 2 . In "O n co g en esis * *■>
and H e rp e s v iru s e s I I " , p a r t I , p p . 6 3 -7 3 . E d ite d by
G. de The, M.A. E p s te in and H. z u r H ausen . IARC
■ S c i e n t i f i c P u b l i c a t i o n s , n o . 11 , Lyon.
COURTNEY, R . J . , SCHAPPER, P .A . and POWELL, K .L . (1 9 7 6 ) .
S y n th e s is o f v i r u s s p e c i f i c p o ly p e p t id e s by te m p e ra tu re -
s e n s i t i v e m u ta n ts o f h e rp e s s im p lex v i r u s ty p e 1 .
V iro lo g y 3 0 6 -3 1 8 .
CREMEH, N . J . , SUMMERS, W.C. and GESTELAND, R .P . (1 9 7 7 ) .
C e l l - f r e e s y n th e s i s o f h e rp e s s im p le x v i r u s p r o t e i n s .
J . V i r o l . 22 , 7 5 0 -7 5 7 .
C REM EH, K ., BODElViER, M. and SUMMERS, W.C. (1 9 7 8 ) .
C h a r a c te r i z a t i o n o f th e mRNA f o r h e rp e s s im p le x v i r u s
thym ine k in a s e by c e l l - f r e e s y n th e s i s o f a c t i v e enzyme.
N u c le ic A cid s R e se a rc h £ , 2333 -2344 .
CHCMBIE, I .E . (1 9 7 5 ) . G e n e tic and b io c h e m ic a l s t u d i e s w ith
h e rp e s s im p le x v i r u s ty p e 1 . PhD t h e s i s , U n iv e r s i ty
o f G lasgow .
CRUMPACKER, C .S . , CJAHTRAND, P . , SUBAK-SHARPE, J .H . and WILKIE.
N.M. (1 9 3 0 ) . R e s is ta n c e o f h e rp e s s im p le x v i r u s to
a c y c lo g u a n o s in e - g e n e t ic and p & y sica l a n a l y s i s .
V iro lo g y 1 0 5 . 1 7 1 -1 8 4 .

DALES, S. and SILVERBERG, H. (1 9 6 9 )• V iro p e x is o f h e rp e s
s im p le x v i r u s by HeLa c e l l s . V iro lo g y 2 2 , 475-480*
DARAI, G. and MUNK, K. (1 9 7 3 ) . Human em bryonic lu n g c e l l s
a b o r t i v e l y i n f e c t e d w ith h e r p e s v i r u s h o m in is
ty p e 2 show some p r o p e r t i e s o f c e l l t r a n s f o r m a t io n .
N a tu re New B io l . 2 4 1 * 268-269*
DAKBY, G ., LARDER, B .A ., BASTOW, K .P . and PIELD, H .J . (1 9 8 0 ) .
S e n s i t i v i t y o f v i r u s e s to p h o s p h o ry la te d 9 - (2 -h y d ro x y -
e th o x y m e th y l) g u a n in e r e v e a le d i n T K -tran sfo rm ed c e l l s .
J . g e n . V i r o l , £8 , 451-454*
DARBY, G ., FIELD, H .J . and SALISBURY, S .A . (1 9 8 1 ) . A l te re d
s u b s t r a t e s p e c i f i c i t y o f h e rp e s s im p le x v i r u s th y m id in e
k in a s e c o n fe r s a c y c l o v i r - r e s i s t a n c e . N a tu re (London)
289 , 81-83*
DARLINGTON, R.W. and MOSS, L.H* I I I (1 9 6 8 ) . H e rp e s v iru s
en v e lo p m e n t. J . V i r o l . 2 , 48-55*
DELIUS, H. and CLEMENTS, J .B . (1 9 7 6 ) . A p a r t i a l d e n a tu r a t io n
map o f h e rp e s s im p le x v i r u s ty p e 1 DNA: e v id e n c e f o r
i n v e r s io n s o f th e u n iq u e DNA r e g io n s . J . g e n . V iro l*
21 , 1 2 5 -1 3 3 .
DEMARCHI, J . , BLANKENSHIP, M., BROWN, G. and KAPLAN, A .S . (1 9 7 8 ) .
S iz e and c o m p le x ity o f human c y to m e g a lo v iru s DNA.
V iro lo g y 82., 643 -646 .
DKNHARDT, D .T. (1 9 6 6 ) , A membrane f i l t e r te c h n iq u e f o r th e
d e te c t io n o f com plem entary DNA. B iochem . b io p h y s .
R es. Cominun. 22 , 641-646 .
DHAR, R ., SUBRAmANIAN, K .N ., PAN, J . and WEISSMAN, S.M. (1 9 7 7 ) .
S t r u c tu r e o f a l a r g e segm ent o f th e genome o f s im ia n
v i r u s 40 t h a t does n o t encode known p r o t e i n s .
P ro c . N a t l . Acad. S c i . USA 8 2 7 -8 3 1 .

DIXON, R .A .F . and FARBErt, F .E . (1 9 8 0 ) . C hannel c a t f i s h
v i r u s : p h y s ic o c h e m ic a l p r o p e r t i e s o f th e v i r a l genome
and i d e n t i f i c a t i o n o f v i r a l p o ly p e p t id e s .
V iro lo g y 102 , 2 6 7 -2 7 8 .
DIXON, R .A .F . and SCHAFFERr P .A . (1 9 8 0 ) . F i n e - s t r u c t u r e m apping
and f u n c t i o n a l a n a l y s i s o f t e m p e r a tu r e - s e n s i t iv e
m u ta n ts i n th e gene e n co d in g th e h e rp e s s im p le x v i r u s
ty p e 1 im m e d ia te - e a r ly p r o t e i n VP175*
J . V i r o l . 2 i» 1 8 9 -2 0 3 .
D01YNIUK, M ., PRITCHETT, R. and KIEFF, E. (1 9 7 6 ) . P r o t e in s o f
E p s te in -B a r r v i r u s . I . A n a ly s is o f th e p o ly p e p t id e s
o f p u r i f i e d e n v e lo p ed E p s te in - B a r r v i r u s .
J . V i r o l . 1 2 , 9 3 5 -9 4 9 .
DUFF, R. and RAPP, F. (1 9 7 1 ) .. P r o p e r t i e s o f h a m s te r embryo
f i b r o b l a s t s tra n s fo rm e d in v i t r o a f t e r e x p o su re to
u l t r a v i o l e t - i r r a d i a t e d h e rp e s s im p le x v i r u s ty p e 2 .
J . V i r o l . 8 , 46 9 -4 7 7 .
DUMAS, A.M ., GEELEN, J .L .M .O ., MARIS, W. and VAN DER NOORDAA, J .
(1 9 8 0 ) . I n f e c t i v i t y and m o le c u la r w e ig h t o f v a r i c e l l a -
z o s t e r v i r u s DNA. j . gen . V i r o l . ££ , 2 3 3 -2 3 5 .

EASTON, A .J . and CLEMENTS, J .B . (1 9 8 0 ) . Tem poral r e g u la t i o n
o f h e rp e s s im p le x v i r u s ty p e 2 t r a n s c r i p t i o n and
c h a r a c t e r i z a t i o n o f v i r u s im m e d ia te -e a r ly mRNAs.
N u c le ic A cids R e se a rc h 8 , 2627-2645*
EBERLE, R. and COURTNEY, R .J . (1980a) • P r e p a r a t io n and
c h a r a c t e r i z a t i o n o f s p e c i f i o a n t i s e r a to i n d iv i d u a l
g ly c o p r o te in a n t ig e n s c o m p ris in g th e m ajo r g ly c o p r o te in
r e g io n o f h e rp e s s im p lex v i r u s ty p e 1 . J . V i r o l . 25> 902-917
EBERLE, R. and COURTNEY, R .J . (1 9 8 0 b ) . gA and gB g ly c o p r o te in s
o f h e rp e s s im p lex v i r u s ty p e 1 : two fo rm s o f a s in g le
p o ly p e p t id e . J . V i r o l . 6 6 5 -6 7 5 .
ECKER, J .R . and HYMAN, R.W. (1 9 8 1 ) . A n a ly s is o f i n t e r r u p t i o n s
i n th e p h o s p h o d ie s te r backbone o f h e rp e s s im p lex
v i r u s DNA. V iro lo g y 1 1 0 , 2 1 3 -2 1 6 .
EGLIN, R . P . , ’SHARP, F . , MacLEAN, A .B ., MacNAB, ^ .C .M ., CLEMENTS,
J .B . and WILKIE, N.M. (1 9 8 1 ) . D e te c t io n o f RNA
com plem entary to h e rp e s s im p le x v i r u s DNA i n human
c e r v i c a l squam ous c e l l n e o p la sm s . C an cer R e se a rc h ,
in p r e s s (S ep tem ber 1 9 8 1 ) .
EISENBERG, R . J . , iiYDREAN-STERN, C. and COHEN, G.H. (1 9 7 9 ).
S t r u c t u r a l a n a l y s i s o f p r e c u r s o r and p ro d u c t fo rm s o f
type-common en v e lo p e g ly c o p r o te in D (CP-1 a n t ig e n ) o f
h e rp e s s im p lex v i r u s ty p e 1 . J . V i r o l . 21 , 608-620 .
EISENBERG, R . J . , PONCE DE LEON, M. and COHEN, G.H. (1 9 8 0 ) .
C om parative s t r u c t u r a l a n a l y s i s o f g ly c o p r o te in gD o f
h e rp e s s im p le x v i r u s ty p e s 1 and 2 . J . V i r o l . 2 £ , 42 8 -4 3 5 .
ELION, G .B ., FURMAN, P .A ., FYFE, J . A . , DE MIRANDA, P . ,
BEAUCHAMP, L . and SCHAEFFER, H .J . (1 9 7 7 ) . S e l e c t i v i t y
o f a c t io n o f an a n t i h e r p e t i c a g e n t , 9 - (2 -h y d ro x y -
e th o x y m eth y l) g u a n in e . P ro c . N a t l . Acad. S c i . USA
24, 5716-5720.

ENQUIST, L ,W ., VANLE WOUDE, G .F ., WAGNER, M ., SMILEY, J .R .
and SUMMERS, W.C. (1 9 7 9 ) . C o n s tru c t io n and
c h a r a c t e r i z a t i o n o f a rec o m b in a n t p la sm id en co d in g th e
gene f o r th e th y m id in e k in a s e o f h e rp e s s im p le x ty p e
1 v i r u s . Gene 2 , 3 35 -342 .
EPSTEIN, M.A. (1 9 6 2 a ) . O b s e rv a tio n s on th e f i n e s t r u c t u r e o f
m atu re h e rp e s s im p le x v i r u s and on th e c o m p o s itio n o f
i t s n u c le o id . J . e x p . Med. 1 1 5 , 1 -9 .
EPSTEIN, M.A. (1 9 6 2 b ). O b s e rv a tio n s on th e mode o f r e l e a s e o f
h e rp e s v i r u s from i n f e c te d HeLa c e l l s .
J . c e l l . B io l . 1 2 , 589-597 .
EPSTEIN, M.A. and HOLT, S . J . (1 9 6 3 ). E le c t r o n m ic to s c o p ic
o b s e r v a t io n s on th e s u r f a c e a d e n o s in e t r ip h o s p h a t a s e -
l i k e enzym es o f HeLa c e l l s i n f e c t e d w ith h e rp e s v i r u s .
J . c e l l . B io l . 1 9 , 337-347 .
ERICKSON, J .S . and KAPLAN, A .S . (1 9 7 3 ) . S y n th e s is o f p r o t e i n s
i n c e l l s i n f e c t e d w ith h e r p e s v i r u s . IX . S u l f a te d
p r o t e i n s . V iro lo g y ££ , 9 4 -1 0 2 .
ESPARZA, J . , PURIFOY, D .J .M ., SCHAFFER, P .A . and BENYESH-
MELNICK, M. (1 9 7 4 ) . I s o l a t i o n , co m p lem en ta tio n and
p r e l im in a r y p h e n o ty p ic c h a r a c t e r i z a t i o n o f te m p e ra tu re -
s e n s i t i v e m u ta n ts o f h e rp e s s im p lex v i r u s ty p e 2 .
V iro lo g y £2> 55 4 -5 6 5 .
ESPARZA, J . , BENYESH-MELNICK, M. and SCHAFFER, P .A . (1 9 7 6 ) .
I n t e r t y p i c c o m p lem en ta tio n and re c o m b in a tio n betw een
t e m p e r a tu r e - s e n s i t iv e m u ta n ts o f h e rp e s s im p le x v i r u s
ty p e s 1 and 2 . V iro lo g y 22* 3 72 -384 .

FALK, ! . , DEINHARDT, F . , NONOYAMA, M ., WOLFE, L .G ., BERGHOLTZ, C
LAPIN, B ., YAKOVLEVA, L . , AGRBA, V ., HENLE, G. and
HENLE, W. (1 9 7 6 ) . P r o p e r t i e s o f a baboon ly m p h o tro p ic
h e rp e s v i r u s r e l a t e d to E p s te in -B a r r v i r u s .
I n t . J . C ancer I £ , 798 -807 .
FALKE, D*, SIEGER T, R. and VOGEL, W. (1 9 5 9 ) . E le k tro n e n m ik ro -
s k o p isc h e Befunde z u r F rag e d e r D oppelm em branbildung
d e s H e rp e s - s im p le x -v i ru s . A rch . g e s . V i r u s f o r s c h .
% 48 4 -4 9 6 .
FALKE, D ., NEHRBASS, W., BRAUER, L . and MULLER, W.E.G. (1 9 8 1 ) .
A d en y lic a c id :d e o x y th y m id in e 5 ' - p h o s p h o t r a n s f e r a s e :
e v id e n c e f o r th e e x is te n c e Qf a n o v e l h e ry e s s im p lex
v i r u s - in d u c e d enzyme. J . g en . V i r o l . 22* 2 4 7 -2 5 5 .
FAR HER, F .E . (1 9 7 6 ) . Com parison o f DNA f a c i l i t a t o r s i n th e
u p ta k e and i n t r a c e l l u l a r f a t e o f i n f e c t i o n s h e rp e s
s im p le x v i r u s ty p e 2 DNA. B ioch im . b io p h y s . A cta 454 . 410
FARNHAM, A .E. and NEWTON, A.^A. (1 9 5 9 ) . The e f f e c t o f some
e n v iro n m e n ta l f a c t o r s on h e rp e s v i r u s grown i n HeLa
c e l l s . V iro lo g y 2 , 449-461 .
FA7/CETT, D.W. (1 9 5 6 ) . E le c tro n m ic ro sco p e o b s e r v a t io n s o f
i n t r a c e l l u l a r v i r u s - l i k e p a r t i c l e s a s s o c i a t e d w ith th e
c e l l s o f th e Lucke r e n a l ad en o carc in o m a .
J . b io p h y s . b iochem . C y to l . 2 , 72 5 -7 4 2 .
FENWICK, M.L. and WALKER, M .J. (1 9 7 8 ) . S u p p re ss io n o f th e
s y n th e s i s o f c e l l u l a r m acrom olecu les by h e rp e s s im p lex
v i r u s . J . g en . V i r o l . £1 , 3 7 -5 1 .
FENWICK, M .L ., WALKER, M .J. and PETKEVICH, J.M . (1 9 7 8 ).
On th e a s s o c i a t i o n o f v i r u s p r o t e i n s w ith th e n u c l e i o f
c e l l s in f e c te d w ith h e rp e s s im p lex v i r u s .
J. gen. Virol. 29, 519-529.

FENWICK, M.L. and WALKER, M .J. (1 9 7 9 ) . P h o s p h o ry la t io n o f a
r ib o s o m a l p r o t e i n and o f v i r u s - s p e c i f i c p r o t e i n s i n c e l l s
i n f e c t e d w ith h e rp e s s im p le x v i r u s . J . g e n . V i r o l .
45. 3 9 7 -4 0 5 .
FENWICK, M., MORSE, L .S . and ROIZMAN, B. (1 9 7 9 ) . Anatomy o f
h e rp e s s im p le x v i r u s DNA. X I. A pparen t c l u s t e r i n g o f
f u n c t i o n s e f f e c t i n g r a p id i n h i b i t i o n o f h o s t DNA and
p r o t e i n s y n t h e s i s . J . V i r o l . 29 , 8 2 5 -8 2 7 .
FENWICK, M ., WALKER, M. and MARSHALL, 1 . (1 9 8 0 ) . Some
c h a r a c t e r i s t i c s o f an e a r l y p r o t e i n (IC P22) s y n th e s iz e d
i n c e l l s i n f e c t e d w ith h e rp e s s im p le x v i r u s .
J . g e n . V i r o l . £ £ , 3 3 3 -3 4 2 .
FIELD, H .J . and DARBY, G. (1 9 7 8 ) . The p a th o g e n ic i ty o f
th y m id in e k i n a s e - d e f i c i e n t m u tan t o f h e rp e s s im p le x v i r u s
i n m ic e . J . H ygiene (C am bridge) 8 1 , 2 6 7 -2 7 7 .
FIELD, H .J . and DARBY, G. (1 9 8 0 ) . The p a th o g e n ic i ty f o r m ice
o f s t r a i n s o f h e rp e s s im p le x v i r u s w hich a r e r e s i s t a n t
to a c y c lo v i r i n v i t r o and in v iv o . A n t im ic ro b ia l A gents
and C hem otherapy 1 2 , 2 0 9 -2 1 6 .
FINNIE, B .P . , LITTLEJOHNS, I .R . and ACLAND, H.M. (1 9 7 6 ) .
M o r t a l i t i e s i n Parm a w a l la b ie s (M acropus parm a) a s s o c ia te d
w i th p ro b a b le h e r p e s v i r u s . A u s t r a l ia n V e te r in a r y J . . ......
52 , 2 9 4 .
FLANAGAN, J .F . (1 9 6 7 ) . V i r u s - s p e c i f i c r i b o n u c le i c a c id
s y n th e s i s i n KB c e l l s i n f e c t e d w ith h e rp e s s im p le s v i r u s .
J . V i r o l . 1 , 5 8 3 -5 9 0 .
, FLAVELL, R .A ., BIRFELDER, E . J . , SANDERS, J .P .M . and BORST, P .
(1 9 7 4 a ) . DNA-DNA h y b r i d iz a t i o n on n i t r o c e l l u l o s e f i l t e r s .
I . G e n e ra l c o n s id e r a t i o n s and n o n - id e a l k i n e t i c s .
B ur. J . B iochem . ££ , 535-543-

PLAVELL, R .A ., BORST, P . and BIRFELDER, E .J . (1 9 7 4 b ). DNA-DNA
h y b r i d i s a t i o n on n i t r o c e l l u l o s e f i l t e r s . 2 . C o n c a te n a tio n
e f f e c t s . B ur. J . Biochem . ££ , 545-548 .
FLEC KEN STEIN, B. and WOLF, H. (1 9 7 4 ) . P u r i f i c a t i o n and p r o p e r t i e s
o f h e r p e s v i r u s s a im i r i LNA. V iro lo g y £8 , 5 5 -6 4 .
FLECKENSTEIN, B ., BORNKAMM, G. W. and LUDWIG, H. (1 9 7 5 ) .
R e p e t i t i v e se q u en c es i n co m p le te and d e f e c t iv e genomes
o f h e r p e s v i r u s s a i m i r i . J . V i r o l . 1£ , 39 8 -4 0 6 .
FLECKENSTEIN, B . , BORNKAMM, G.W., MULDER, C ., WERNER, F - J . ,
DANIEL, M .D ., FALK, L .A. and DELIUS, H. (1 9 7 8 ) .
H e rp e s v iru s a t e l e s DNA and i t s hom ology w ith h e rp e s v i r u s
s a i m i r i n u c le ic a c i d . J . V i r o l . 2 ^ , 3 6 1 -3 7 3 .
FRANCKE, B ., MOSS, H ., TIMBURY, M.C. and HAY, J . (1 9 7 8 ).
A lk a lin e DNase a c t i v i t y i n c e l l s i n f e c t e d w ith a
t e m p e r a tu r e - s e n s i t iv e m u tan t o f h e rp e s s im p lex v i r u s
ty p e 2 , J . V i r o l . 26 , 209-213*
FRANCKE, B. and MARGOLIN, J . (1 9 8 1 ) . E f f e c t o f n o v o b io c in and
o th e r DNA g y ra s e i n h i b i t o r s on v i r u s r e p l i c a t i o n and DNA
s y n th e s i s i n h e rp e s s im p lex ty p e 1 - i n f e c t e d BHK c e l l s .
J . g e n , V i r o l . £2 , 401-404*
FRENKEL, N. and ROIZMAN, B. (1 9 7 2 a ) . S e p a ra t io n o f th e
h e r p e s v i r u s d e o x y r ib o n u c le ic a c id d u p le x in to u n iq u e
f ra g m e n ts and i n t a c t s t r a n d s on s e d im e n ta t io n i n
a l k a l i n e g r a d i e n t s . J . V i r o l . 10 , 565 -572 .
FRENKEL, N. and ROIZMAN, B. (1 9 7 2 b ). R ib o n u c le ic a c id
s y n th e s i s i n c e l l s i n f e c t e d w ith h e rp e s s im p le x v iru s *
c o n t r o l s o f t r a n s c r i p t i o n and RNA ab u n d an ce .
P ro c . N a t l . Acad. S c i . USA 62., 2654-2658 .
FRENKEL, N ., ROIZMAN, 3 . , CASSAI, E. and NAHMIAS, A . (1 9 7 2 ) .
A h e rp e s s im p lex 2 DNA fra g m e n t and i t s t r a n s c r i p t i o n
in human c e r v i c a l c a n c e r t i s s u e . P ro c . N a t l . Acad.
S c i . USA 62, 3784-3789 .

FRENKEL, N . , SILVERSTEIN, S . , GASSAI, E. and ROIZMAN, B. ( 1 9 7 3 ) .
RNA s y n t h e s i s i n c e l l s i n f e c t e d w ith h e rp e s s im p lex v i r u s .
V II . C o n tro l o f t r a n s c r i p t i o n and o f t r a n s c r i p t a b u n d a n c ie s
o f u n iq u e and common se q u e n c e s o f h e rp e s s im p lex v i r u s
1 and 2 . J . V i r o l . 11 , 8 8 6 -8 9 2 .
FRENKEL, N ., JACOB, R . J . , HONESS, R .W ., HAYWARD, G .S ., LOCKER,
H. and ROIZMAN, B. (1 9 7 5 ) . Anatomy o f h e rp e s s im p lex
v i r u s DNA. I I I . C h a r a c te r iz a t i o n o f d e f e c t iv e DNA
m o le c u le s and b i o lo g i c a l p r o p e r t i e s o f v i r u s p o p u la t io n s
c o n ta in in g them . J . V i r o l . 1 6 , 1 5 3 -1 6 7 .
FRENKEL, N ., LOCKER, H ., BATTERSON, W., HAYWARD, GwS. and
ROIZMAN, B. (1 9 7 6 ) . Anatomy o f h e rp e s sim p lex v i r u s DNA.
V I. D e f e c t iv e DNA o r i g i n a t e s from th e S com ponent.
J . V i r o l . 2 0 , 52 7 -5 3 1 .
i'KEHKEL, H ., LOCKEH, H. and V1AZNY, D.A. (1 9 8 0 ) . S tu d ie s o f
d e f e c t i v e h e rp e s s im p lex v i r u s e s . A nnals o f NOW York
Acad. S c i . 254* 3 47 -370 .
FRIEDMANN, A ., COV/ARD, J . E . , ROSENKRANZ, H .S . and MORGAN, C. (1 9 7 5 ) .
E le c t r o n m ic ro sc o p ic s t u d i e s on assem b ly o f h e rp e s s im p lex
v i r u s upon rem oval o f h y d ro x y u rea b lo c k .
J . g e n . V i r o l . 2 6 , 1 7 1 -1 8 1 .
FRIEDMANN, A ., SHLOMAI, J . and BECKER, Y. (1 9 7 7 ) . E le c t r o n
m ic ro sco p y o f h e rp e s s im p le x v i r u s DNA m o le cu le s i s o l a t e d
from i n f e c t e d c e l l s by c e n t r i f u g a t i o n i n CsCl d e n s i ty
g r a d i e n t s . J . g e n . V i r o l . 24.* 5 0 7 -5 2 2 .
FURLONG, D ., SWIFT, H. and ROIZMAN, B. (1 9 7 2 ) . A rrangem ent o f - — —
h e r p e s v i r u s d e o x y r ib o n u c le ic a c id i n th e c o r e .
J . V i r o l . 10 , 107 1 -1 0 7 4 .
FURMAN, P .A ., ST. CLAIR, M .H., FYFE, J .A . , RIDEOUT, J . L . , KELLER,
P.M. and ELION, G.B. (1 9 7 9 ) . I n h i b i t i o n o f h e rp e s s im p lex
v i r u s - in d u c e d DNA p o ly m erase a c t i v i t y and v i r a l DNA
r e p l i c a t i o n by 9 - (2 -h y d ro x y e th o x y m e th y l) g u an in e and i t a
t r ip h o s p h a te . J . V iro l . ^ 2 , 7 2 -7 7 .

GALLAHER, W.R., LEVITAN, D.B. and BLOUGH, H.A. (1 9 7 3 ) . E f f e c t o f
2-d e o x y -D -g lu c o se on c e l l f u s io n in d u c e d by N ew castle
d i s e a s e and h e rp e s s im p le x v i r u s e s . V iro lo g y 1 9 3 -2 0 1 .
GALLOWAY, D.A. and SWAIN, M. (1 9 8 0 ) . C lo n in g o f h e rp e s s im p lex
v i r u s ty p e 2 DNA f ra g m e n ts i n a p la sm id v e c to r .
Gene 11 , 2 5 3 -2 5 7 .
GALLOWAY, D.A. and McDOUGALL, J .K . (1 9 8 1 ) . T ra n s fo rm a tio n o f
r o d e n t c e l l s by a c lo n e d DNA fra g m e n t p f h e rp e s s im p lex
v i r u s ty p e 2 . J . V i r o l . 7 4 9 -7 6 0 .
GANNON, F . , O'HARE, K ., PERRIN, P . , LEPENNEC, H .P . , BENOIST, C .,
COCHET, M ., BREATHNAQH, R . , ROYAL, A ., GARAPIN.* A ., CAMI,
B. and CHAM BON, P . (1 9 7 9 ) . O r g a n is a t io n and seq u en ces
a t th e 5 ' end o f a c lo n e d co m p le te ovalbum in g e n e .
N a tu re (London) 2 7 8 . 4 2 8 -4 3 3 .
GARFINKLE, B. and McAUSLAN, B .R. (1 9 7 4 ) . R e g u la tio n o f h e rp e s
s im p lex v i r u s - in d u c e d th y m id in e k in a s e .
B iochem . b io p h y s . R es. Commun. £8 , 8 2 2 -8 2 9 .
GEELEN, J .L .M .C ., WALIG, C ., WERTHEIM, P . and VAN DER NOORDAA,
J . (1 9 7 8 ) . Human c y to m e g a lo v iru s DNA. I . M o le cu la r
w e igh t and i n f e c t i v i t y . J . V i r o l . 2j6, 8 i3 -8 1 6 .
GELB, L .D ., HUANG, J . J . and WELLINGHOFF, W .J. (1 9 8 0 ) .
V a r i c e l l a - z o s t e r v i r u s t r a n s f o r m a t io n o f h a m s te r embryo
c e l l s . J . g e n . V i r o l . gl» 1 7 1 -1 7 7 .
GERBER, P . , PRITCHETT, R .F . and KIEFF, E. (1 9 7 6 ) . A n tig e n s
and DNA o f a ch im panzee a g e n t r e l a t e d to E p s te in -B a r r
v i r u s . J . V i r o l . 19 , 1090-1099 .
GIBSON, W. and ROIZMAN, B. (1 9 7 1 ) . C o m p a r tm e n ta liz a tio n o f
sperm ine and sp e rm id in e i n th e h e rp e s s im p le x v i r i o n .
P ro c . N a t l . Acad. S c i . USA 68, 2 818 -2821 .

GIBSON, W, and ROIZMAN, B. (1 9 7 2 ) . P r o t e in s s p e c i f i e d by
h e rp e s s im p le x v iru s * V I I I . C h a r a c te r iz a t io n and
c o m p o s itio n o f m u l t i p le c a p s id fo rm s o f su b ty p e s 1
and 2 . J . V i r o l . 1 0 , 104 4 -1 0 5 2 .
GIBSON, W. and ROIZMAN, B. (1 9 7 4 ) . P r o t e in s s p e c i f i e d by
h e rp e s s im p le x v i r u s . X. S ta in in g and r a d i o l a b e l l i n g
p r o p e r t i e s o f B c a p s id and v i r i o n p r o t e i n s in
p o ly a c ry la m id e g e l s . J . V i r o l . 1 J , 1 55 -165 .
GIRA1D0, G ., BETH, E . , KOURILSKY, G ., HENLE, W., HENLE, G .,
MIKE, V ., HURAUX, J .M ., ANDERSEN, H .K ., GHARBI, M*R.,
KYALWAZI, S .K . and PUISSANT, A. (1 9 7 5 ) . A ntibody
p a t t e r n s to h e r p e s v i r u s e s i n K a p o s i1 s sarcom a:•t
s e r o l o g i c a l a s s o c i a t i o n o f E uropean K aposi*s sarcom a• * r*
w ith c y to m e g a lo v iru s . I n t . J . C ancer 1£ , 83 9 -8 4 8 .
GliUN, D. and KEjSPP, S . (1 9 7 8 ) . DNA o f E p s te in -B a r r v i r u s .^ *1
IV . L in k ag e map o f r e s t r i c t i o n en d o n u c lea se f ra g m e n ts
o f th e B95-8 and W91 s t r a i n s o f E p s te in - B a r r v i r u s .
J . V i r o l . 2 8 , 524 -542 .
GIVEN, D. and KIEFF, E. (1 9 7 9 ) . DNA o f E p s te in -B a r r v i r u s .
V I. M apping o f th e i n t e r n a l tandem r e i t e r a t i o n .
J . V i r o l . 21, 3 15 -324 .
GIVEN, D ., YEE, D ., GRIMM,' K. and KIEFF, E. (1 9 7 9 ). DNA o f
E p s te in - B a r r v i r u s . V. D i r e c t r e p e a t s a t th e ends o f
E p s te in - B a r r v i r u s DNA. J . V i r o l . 20 , 8 52 -862 .
GLUZMAN, Y ., SAMBROOK, J .F . and FRISQUE, R .J . (1 9 8 0 ). E x p re s s io n
o f e a r ly g e n e s o f o r i g i n - d e f e c t i v e m u ta n ts o f s im ia n
v i r u s 4 0 . P ro c . N a t l . Acad. S c i . USA XL* 3898-3902 .
GOEDDEL, D .V ., LEVING, D.W ., DULL, T . J . , GROSS, M., LAWN, R .M .,
McCANDLISS, R ., SEEBURG, P .H ., ULLRICH, A ., YELVERTON, E.
and GRAY, P . J . (1 9 8 1 ) . The s t r u c t u r e o f e ig h t d i s t i n c t
c lo n e d human le u k o c y te i n t e r f e r o n cDNAs.
N a tu re (London) 2 9 0 . 2 0 -2 6 ,

GOLDIN, A .L ., SANDRI-GOLDIN, f i . M . , LEVINE, M. and GLORIOSO,
J .C . (1 9 8 1 ). C lo n in g o f h e rp e s s im p le x v i r u s ty p e 1
se q u en c es r e p r e s e n t in g th e w hole genome. J . V i r o l . 22* 50-58
GOQDIIEART, C .R ., PLUMMER, G. and WANER, J .L . (1 9 6 8 ) . D e n s ity
d i f f e r e n c e s o f DNA b f human h e rp e s s im p le x v i r u s e s ,
ty p e s 1 and 2 . V iro lo g y 22* 4 7 3 -4 7 5 .
GORDIN, H ., OLSHEVSKY, U ., ROSENKRANTZ, H .S . and BECKER, Y. (1 9 7 3 ) .
S tu d ie s on h e rp e s s im p le x v i r u s DNA: d e n a tu r a t io n
p r o p e r t i e s . V iro lo g y 22* 2 8 0 -2 8 4 .
GRAFSTROM, R .H ., ALWINE, J .C . , STEINHART, W.L. and HILL, C.W.
(1 9 7 4 ) . T erm inal r e p e t i t i o n s i n h e rp e s s im p le x v i r u s *
ty p e 1 DNA. Cold S p r in g H arbo r Symp. Q u an t. B io l . 22.* 879-68:
GRAFSTROM, R .H ., ALWINE, J .C . , STEINHART, W .L., HILL, C.W. and
HYMAN, fi.W. (1 9 7 5 ) . The te r m in a l r e p e t i t i o n q f h e rp e s
s im p lex v i r u s DNA. V iro lo g y 67. 1 4 4 -1 5 7 .
GRAHAM, B . J . , BENGALI, Z. and VANDE WOUDE, G .F. (1 9 7 8 ) .
P h y s ic a l map o f th e o r i g i n o f d e f e c t iv e DNA i n h e rp e s
s im p lex v i r u s ty p e 1 DNA. J . V i r o l . 22 , 8 7 8 -8 8 7 ,
GRAHAM, F .L . , VELDHUISEN, G and WILKIE, N.M. (1 9 7 3 ) .
I n f e c t io u s h e rp e s v i r u s DNA. N a tu re New B io l . 2 4 5 , 2 6 5 -2 6 6 .
GRAY, A ., TOKUMARU, T. and SCOTT, T .F . (1 9 5 8 ) . D i f f e r e n t
c y to p a th o g e n ic e f f e p t s o b se rv e d i n HeLa c e l l s i n f e c t e d
w ith h e rp e s s im p lex v i r u s . A rch . g e s . V i r u s f o r s c h . 8 , 6 0 -7 6 .
GRUNSTEIN, M. and HOGNESS, D .S . (1 9 7 5 ) . Colony h y b r i d i z a t i o n :
a m ethod f o r th e i s o l a t i o n o f c lo n e d DNAs t h a t c o n ta in
a s p e c i f i c g en e . P ro c . N a t l . Acad. S c i . USA 72 , 3961-3965 .

HAARR, L. and IvlAKSDEN, U .S . (1 9 8 1 ) . T w o-d im ensional g e l
a n a l y s i s o f HSV ty p e 1 - in d u c e d p o ly p e p t id e s and
g ly c o p r o te in p r o c e s s in g . J . g e n . V i r o l . £2 , '77-92.
HALLIBURTON^ I.W . (1 9 7 2 ) . B io lo g ic a l c o m p a riso n s o f ty p e 1
and ty p e 2 h e rp e s s im p le x v i r u s e s . In "O n co g en es is and
H e r p e s v iru s e s 1*, p p . 4 3 2 -4 3 8 . E d ite d by P.M . B ig g s,
G. de The and L .N . P ay n e . IARC S c i e n t i f i c P u b l i c a t i o n s ,
n o . 2 , Lyon.
HALLIBURTON, I.W . and TIMBURY, M.C. (1 9 7 3 ) . C h a r a c te r iz a t i o n
p f t e m p e r a tu r e - s e n s i t iv e m u ta n ts o f h e rp e s s im p le x v i r u s
ty p e 2 . Growth and DNA s y n t h e s i s . V iro lo g y 6 0 -6 8 .
HALLIBURTON, I.W . and ANDREW, J .C . (1 9 7 6 ) . DNA p o ly m erase i n
p s e u d o ra b ie s v i r u s i n f e c t e d c e l l s . J . g e n . V i r o l .
20 , 1 4 5 -1 4 3 .
HALLIBURTON, I.W . and TIMBURY, M.C. (1 9 7 6 ) . T em p era tu re -
s e n s i t i v e m u ta n ts o f h e rp e s s im p le x v i r u s ty p e 2 ;
d e s c r i p t i o n o f t h r e e new co m p lem en ta tio n g ro u p s and
s t u d i e s on th e i n h i b i t i o n o f h o s t c e l l DNA s y n t h e s i s .
J . g e n . V i r o l . 22* 2 0 7 -2 2 1 .
HALLIBURTON, I.W . (1 9 8 0 ) . I n t e r t y p i c re c o m b in a n ts o f h e rp e s
s im p le x v i r u s e s . J . g e n . V i r o l . 4§.» 1 -2 3 .
HALLIBURTON, I .W ., MORSE, L . S . ,. ROIZMAN, B. and QUINN, K .E.
(1 9 8 0 ) . M apping o f th e th y m id in e k in a s e g en es o f ty p e
1 and ty p e 2 h e rp e s s im p le x v i r u s e s u s in g i n t e r t y p i c
re c o m b in a n ts . J . g e n . V i r o l . 49 , 235-254*
HAY, J . , KOTELES, G . J . , KEIR, H.M. and SUBAK-SHARPE, H. (1 9 6 6 ) .
H erpes v i r u s s p e c i f i c r i b o n u c le i c a c i d s .
N a tu re (London) 2 1 0 , 3 8 7 -3 9 0 .
IiAY, J . and SUBAK-SHARPE, J .I i . (1 9 7 6 ) . M u tan ts o f h e rp e s s im p lex
v i r u s ty p e s 1 and 2 t h a t a r e r e s i s t a n t to p h o sp h o a o a c e tic
a c id in d u c e a l t e r e d DNA p o ly m erase a c t i v i t i e s i n i n f e c te d
c e l l s . J . g e n . V i r o l . 2i» 1 4 5 -1 4 8 .

HAY, R .T . and HAY, J . (1 9 8 0 ). P r o p e r t i e s o f h e r p e s v i r u s -
in d u c e d " im m ed ia te e a r ly " p o ly p e p t id e s .
V iro lo g y 1 0 4 , 2 3 0 -2 3 4 .
HAYWARD, G .S ., FRENKEL, N. and ROIZMAN, B. (1975a)* The
anatom y o f h e rp e s s im p lex v i r u s DNA: s t r a i n d i f f e r e n c e
and h e te r o g e n e i ty i n th e l o c a t i o n o f r e s t r i c t i o n
e n d o n u c le a se s i t e s . P ro c . N a t l . A cad. S c i . USA J 2 , 1768-1772
HAYWARD, G .S ., JACOB, R . J . , WADSWORTH, S .C . and ROIZMAN, B. *
(1 9 7 5 b ) . Anatomy o f h e rp e s s im p le x v i r u s DNA: e v id e n c e
f o r f o u r p o p u la t io n s t h a t d i f f e r i n th e r e l a t i v e
o r i e n t a t i o n s o f t h e i r lo n g and s h o r t com ponen ts .
P ro c . N a t l . Acad. S c i . USA £2 , 4243-4247*
HAYWARD, S .D ., NOGEE, L. and HAYWARD, G .S . (1 9 8 0 ) .
O rg a n iz a t io n o f r e p e a te d r e g io n s w i th in th e E p s te in -
B a rr v i r u s DNA m o le c u le . J . V i r o l . 22* 5 07 -521 .
H E IN E ,'J .W ., HONESS, R .W ., CASSAI, E . and ROIZMAN, B. (1 9 7 4 ) .
P r o t e in s s p e c i f i e d by h e rp e s s im p le x v i r u s . X I I .
The virion polypeptides of type 1 strains.J . V i r o l . 1 4 , 640 -651 .
HEINE, V. and COTTLER-FOX, M. (1 9 7 5 ) . E le c t r o n m ic ro sc o p ic
o b s e r v a t io n s on th e c o m p o s itio n o f h e rp e s - ty p e v i r u s e s .
In "O n co g en esis and#H e rp e s v iru s e s I I " , p a r t I , p p . 1 03 -
110 . E d ite d by G. de The, M.A. E p s te in and H. z u r
H ausen . IARC S c i e n t i f i c P u b l i c a t io n s , n o . 1 1 , Lyon.
HELLER, M. and KIEFF, E. (1 9 8 1 ). C o l in e a r i t y betw een th e DNAs - - ~ **
o f E p s te in - B a r r v i r u s and h e r p e s v i r u s p a p io .
J . V i r o l . 21* 8 2 1 -8 2 6 .
HELLER, M., DAMBAUGH, T. and KIEFF, E. (1 9 8 1 a ) . E p s te in -B a r r
v i r u s DNA. IX. V a r ia t io n among v i r a l DNAs from** 1
p ro d u c e r and n o n p ro d u ce r i n f e c t e d c e l l s .
J . V i r o l . 28, 632-648 .

HELLER, Ivl., GERBER, P . and KIEFF, E. (1 9 8 1 b ) . H e rp e s v iru s
p a p io DNA i s s i m i l a r i n o r g a n iz a t io n to E p s te in -B a r r
v i r u s DNA. J . V i r o l . 21* 698-709 .
HENLE, G ., HENLE, W. and DIEHL, V. (1 9 6 8 ) . R e la t io n o f
B u r k L t f s tu m o r - a s s o c ia te d h e rp e s - ty p e v i r u s to
i n f e c t i o u s m o n o n u c le o s is . P ro c . N a tl* Acad. S c i . USA
2 2 , 9 4 -1 0 1 .
HENLE, G ., HENLE, W., CLIFFORD, P . , DIEHL, V ., KAFUKO, G.W.,
KIRYA, B .G ., KLEIN, G ., . MORROW, R .H ., MUNUBE, G .M .R .,
PIKE, M .C ., TUKEI, P.M . and ZIEGLER, J .L . (1 9 6 9 ) .
A n tib o d ie s to EB v i r u s i n B u r k i t t f s lymphoma and c o n t r o l
■ g ro u p s . J . N a t. C an cer I n s t . 4 3 ." H 4 7 - H 5 7 .
HILL, B .T . and WHATLEY, S. (1 9 7 5 ) . A s im p le , r a p id m ic ro a s s a y
f o r DNA. FEBS L e t t e r s £6 , 2 0 -2 3 .
HINZE, H.C. (1 9 7 1 ) . In d u c t io n o f a lym phoid h y p e r p la s i a and
ly m phom a-like d i s e a s e i n r a b b i t s by h e rp e s v i r u s
s y l v i l a g u s . I n t . J . C ancer 8 , 5 1 4 -5 2 2 .
IilRAI, K ., IKUTA, K. and KATO, S . (1 9 7 9 ) . C om para tive s tu d i e s
on M a re k 's d i s e a s e v i r u s and h e r p e s v i r u s o f tu r k e y DNAs.
J . g e n . V i r o l . 4 5 , 1 1 9 -1 3 1 .
HIRSCH, I . and VODKA, V. (1 9 7 4 ) . R ib o n u c le o t id e s l in k e d to
DNA o f h e rp e s s im p le x v i r u s ty p e 1 . J . V i r o l . 1 2 , 1162-1168
HIRSCH," I . , RE ISC HI G, - J . , ROUBAL, J . and VONKA, V. (1 9 7 5 ) .
S t r u c tu r e o f h e rp e s s im p le x v i r u s DNA: to p o g rap h y o f
th e m o le c u le . I . A bsence o f c i r c u l a r l y perm uted sequences*
V iro lo g y 62.* 4 9 6 -5 0 5 .
HIRSCH, I . , ROUBAL, J . and VONKA, V. (1 9 7 6 ) . R e p l i c a t in g DNA
o f h e rp e s s im p le x v i r u s ty p e 1 . I n t e r v i r o lo g y £ , 1 5 5 -1 7 5 .
HIRSCH, I . , CABRAL, G ., PATTERSON, M. and BISWAL, N. (1 9 7 7 ) .
S tu d ie s on th e i n t r a c e l l u l a r r e p l i c a t i n g DNA o f" h e rp e s
s im p le x v i r u s ty p e 1 . V iro lo g y 8 1 , 4 8 -6 1 .

HO BART, P . , CRAWFORD, R ., SHED, L . , PICTET, R. and RUTTER,
W .J. (1 9 8 0 ). C lo n in g and seq u en ce a n a l y s i s o f cDNAs
en co d in g two d i s t i n c t s o m a to s ta t in p r e c u r s o r s found in
th e e n d o c rin e p a n c re a s o f a n g l e r f i s h .
N a tu re (London) 2 8 8 . 1 3 7 -1 4 0 .
HOCHBERG, E. and BECKER, Y. (1 9 6 8 ) . A d s o rp tio n , p e n e t r a t i o n
and u n c o a tin g o f h e rp e s s im p le x v i r u s .
J . g en . V i r o l . 2 , 231-241*
HOGGAN, M.D. and ROIZMAN, B. (1 9 5 9 ) . The i s o l a t i o n and
p r o p e r t i e s o f a v a r i a n t o f h e rp e s s im p le x p ro d u c in g
m u lt in u c le a te d g i a n t c e l l s i n m ono layer c u l t u r e s i n
th e p re se n c e o f a n t ib o d y . A m erican J . H ygiene JO, 2 0 8 -2 1 9 .
HOLLAND, L .E . , ANDERSON, K .P ., STRINGER, J .R . 'a n d WAGNER, E .K .
(1 9 7 9 ) . I s o l a t i o n and l o c a l i z a t i o n o f h e rp e s s im p lex
v i r u s ty p e 1 mRNA a b u n d an t b e fo re v i r a l DNA s y n th e s i s .
J . V i r o l . £ 1 , 447 -462 .
HOLLAND, L .E . , ANDERSON, K .P ., SHIPMAN, C. J r . and WAGNER, E .K .
(1 9 8 0 ) . V i r a l DNA s y n th e s i s i s r e q u i r e d f o r th e
e f f i c i e n t e x p re s s io n o f s p e c i f i c h e rp e s s im p le x v i r u s
ty p e 1 mRNA s p e c i e s . V iro lo g y 1 0 1 , 10-24*
HOLMES, I .H . and WATSON, D.H. (1 9 6 3 ) . An e le c t r o n m ic ro sco p e
s tu d y o f th e a tta c h m e n t and p e n e t r a t i o n o f h e rp e s
v i r u s in BHK 21 c e l l s . V iro lo g y 2 1 , 1 1 2 -1 2 3 .
HONESS, R.W. and ROIZMAN, B. (1 9 7 3 ) . P r o t e in s s p e c i f i e d by
h e rp e s s im p lex v i r u s . X I. I d e n t i f i c a t i o n and r e l a t i v e
m olar r a t e s o f s y n th e s i s o f s t r u c t u r a l and n o n - s t r u c t u r a l
h e rp e s vir^us p o ly p e p t id e s in th e i n f e c t e d c e l l .
J . V i r o l . 12 , 1347-1365 .
HONESS, R.W. and ROIZMAN, B. (1 9 7 4 ) . R e g u la tio n o f h e rp e s v i r u s
m acro m o lecu la r s y n th e s i s . I . C ascade r e g u l a t i o n o f th e
s y n th e s is o f t h r e e g ro u p s o f v i r a l p r o t e i n s . *
J . V i r o l . 14 , 8 -1 9 .

HONESS, R.W. and WATSON, D.H, (1 9 7 4 ). H erpes s im p lex v i r u s -
s p e c i f i c p o ly p e p t id e s s tu d ie d by p o ly a c ry la m id e g e l
e l e c t r o p h o r e s i s o f immune p r e c i p i t a t e s .
J . g e n . V i r o l . 22 , 1 7 1 -1 8 5 .
HONESS, R .W ., POWELL, K .L ., ROBINSON, D .J . , SIM, C. and
WATSON, D.H. (1 9 7 4 ) . Type s p e c i f i c and ty p e common
a n t ig e n s i n c e l l s i n f e c t e d w ith h e rp e s s im p lex v i r u s
ty p e 1 and on th e s u r f a c e s o f naked and en v e lo p ed p a r t i c l e s
o f th e v i r u s . J . g en . V i r o l . 22 , 15 9 -1 6 9 .
HONESS, R.W. and ROIZMAN, B, (1 9 7 5 a ) . R e g u la tio n o f h e r p e s v i r u s
m ac ro m o le cu la r s y n t h e s i s : s e q u e n t i a l t r a n s i t i o n o f
p o ly p e p t id e s y n th e s i s r e q u i r e s f u n c t i o n a l p o ly p e p t id e s .
P ro c . N a t l . A cad. S c i . USA J 2 , 1276-1280 .
HONESS, R.W. and ROIZMAN, B. (1 9 7 5 b ). P r o t e in s s p e c i f i e d by
h e rp e s s im p le x v i r u s . X I I I . The g ly c o s y la t io n o f
v i r a l p o ly p e p t id e s . J . V i r o l . 1 6 , 1308-1326 .
HONESS, R.W. and WATSON, D.H. (1 9 7 7 a ) . H erpes s im p lex v i r u s
r e s i s t a n c e and s e n s i t i v i t y to p h o sp h o n o a c e tic a c id .
J . V i r o l . 2 1 , 5 8 4 -600 .
HONESS, R.W. and WATSON, D.H. (1 9 7 7 b ). U n ity and d i v e r s i t y i n
th e h e r p e s v i r u s e s . J . g en . V i r o l . XL* 1 5 -3 7 .
HONESS, R .W ., BUCHAN, A ., HALLIBURTON, I.W . and WATSON, D.H. ( 1 9 8 0 i .
R eco m b in a tio n and l in k a g e betw een s t r u c t u r a l and
r e g u l a to r y g e n es o f h e rp e s s im p lex v i r u s type 1 .
J . V i r o l . 21* 7 1 6 -7 4 2 .
HUANG, A .S . and WAGNER, R .R . (1 9 6 4 ). P e n e t r a t i o n o f h e rp e s
s im p lex v i r u s i n to human ep id e rm o id c e l l s .
P ro c . Soc. exp . B i o l . Med. 1 1 6 , 8 63 -869 .
HUANG, E -S ., CHEN, S .T . and PAGANO, J . (1 9 7 3 ) . Human
c y to m e g a lo v iru s . I . P u r i f i c a t i o n and c h a r a c t e r i z a t i o n
o f v i r a l DNA. J . V i r o l , 12 , 1473-1481 .

HUANG, E -S . and PAGANO, J . (1 9 7 4 ). Human c y to m e g a lo v iru s .
I I . Lack o f r e l a to d n e s s to DNA o f h e rp e s s im p le x I and
I I , E p s te in -B a r r v i r u s and non-hum an s t r a i n s o f
c y to m e g a lo v iru s . J . V ir o l . 1 ^ , 642-645 .
HUANG, E -S ., KILPATRICK, B ., LAKEMAN, P . and ALFORD, C.A. (1 9 7 8 ) .
G e n e tic a n a l y s i s o f a c y to m e g a lo v ir u s - l ik e a g e n t
i s o l a t e d from human b r a in . J . V iro l# 26 , 71 8 -7 2 3 .
HUMMELER, K ., TOMASSINI, N, and ZAJAC, B. (1 9 6 9 ) . E a r ly e v e n ts
i n h e rp e s s im p lex v i r u s i n f e c t i o n s a r a d io a u to g ra p h ic
s tu d y . J . V i r o l . ± , 67 -7 4 .
HUSZAR, D. a n d ’BACCHETTI, S . (1 9 8 1 ) . P a r t i a l p u r i f i c a t i o n .
and c h a r a c t e r i z a t i o n o f th e r i b o n u c le o t id e r e d u c ta s e
in d u ce d 6y h e rp e s s im p le x v i r u s i n f e c t i o n o f mammalian
c e l l s . J . V i r o l . 21* 580 -588 .
liUTTON, J . i t . and YYETMUR, J .G . (1 9 7 3 ) . E f f e c t o f b h em ica l
m o d if ic a t io n on th e r a t e o f r e n a t u r a t i o n o f d e o x y r ib o n u c le ic
a c id . D eam inated and g ly o x a la te d d e o x y r ib o n u c le ic a c i d .
B io c h e m is try 12, 558 -563 .
HUTTON, J .R . (1 9 7 7 ) . R e n a tu ra t io n k i n e t i c s and th e rm a l
s t a b i l i t y o f DNA i n aqueous s o l u t io n s o f form am idet
and u r e a . N u c le ic A cids R e se a rc h 4 , 3537-3555 .
HYMAN, d. ' i l . , jiUriiui, S . and KUDLER, 1 . (1 9 7 6 ) . A n e a rb y
in v e r t e d r e p e a t o f th e t e r m in a l seq u en ce o f h e rp e s
s im p lex v i r u s DNA. Biochem. b io p h y s . R es. Commun. 68 , 609-61
iiYmAN, R . w. , OAKES, J . E . and KUDLER, L . (1 9 7 7 ) . In v i t r o
r e p a i r o f th e p r e - e x i s t i n g n ic k s and g ap s i n h e rp e s
s im p lex v i r u s DNA. V iro lo g y J2* 2 8 6 -2 9 4 .
HYMAN, R.W. (1 9 8 0 ). I d e n t i f i c a t i o n o f p r o t e i n s t i g h t l y bound
to h e rp e s s im p lex v i r u s DNA. V iro lo g y 1 0 5 , 2 54 -255 .

IM I S , J . P . , OAKES, J . E . , HYMAN, R.W. and RAPP, P . (1 9 7 7 ) .
C om parison o f th e DNAs o f v a r i c e l l a z o s t e r v i r u s e s
i s o l a t e d from c l i n i c a l c a s e s o f v a r i c e l l a and h e rp e s
z o s t e r . V iro lo g y 8 2 , 3 4 5 -352 .
INGLIS, M.M. and NEWTON, A.A. (1 9 8 1 ) . C om parison o f th e
a c t i v i t i e s o f HSV-1 and c e l l u l a r mRNAs a s te m p la te s
f o r i n v i t r o t r a n s l a t i o n . V iro lo g y 1 1 0 , 1 -1 5 .
IWASAKA, T . , ODA, H ., MORI, R . , MIYAZOZO, J . and NAGAFUCHI, S .^ • i , • -
(1 9 7 9 ) . P ro d u c t io n o f t u b u la r s t r u c t u r e s i n Vero c e l l s
i n f e c t e d w ith h e rp e s s im p lex v i r u s ty p e 2 t e f f e c t s o f
u l t r a v i o l e t l i g h t i r r a d i a t i o n and a n t i m e t a b o l i t e s .
' J . g e n . V i r o l . 44 , 5 7 -6 7 .

JACOB, R .J . and ROIZMAN, B. (1 9 7 7 ). Anatomy o f herpes sim plex
v ir u s DNA. V III , P r o p e r t ie s o f the r e p l ic a t in g DNA.
J . V ir o l . 22, 3 94 -411 .
JACOB, R .J . , MORSE, L .S . and ROIZMAN, B. (1 9 7 9 ) . Anatomy o f
h erpes sim plex v ir u s DNA. X II. Accum ulation o f head-
t o - t a i l concatem ers in n u c le i o f in f e c t e d c e l l s and t h e ir
r o le in th e g e n e r a tio n o f the fo u r iso m e r ic arrangem ents
o f v i r a l DNA. J . V ir o l . 2%, 44 8 -4 5 7 .
JACQUEMONT, B. and ROIZMAN, B. (1 9 7 5 a ). RNA s y n th e s is in c e l l s
in f e c t e d w ith h erpes sim plex v ir u s . X. P r o p e r t ie s o f
v i r a l symmetric t r a n s c r ip ts and o f d o u b le -stra n d ed RNA
prepared from them. J . V ir o l. 1 £ , 707-713-
JACQUEMONT, B. and ROIZMAN, B. (1 9 7 5 b ). R ib o n u c le ic a c id
s y n th e s is in c e l l s in fe c te d w ith h erp es pim plex v ir u s i
c h a r a c te r is a t io n o f v i r a l h igh m o lecu lar w eight
n u c lea r RNA. J . gen . V ir o l. 2£ , 1 5 5 -1 6 5 .
JACQUEMONT, B. and HUPPERT, J . (1 9 7 7 ). I n h ib it io n o f v i r a l
RNA m eth y la tio n in herpes sim plex v ir u s type 1 - in f e c t e d
c e l l s by 5 ’ -S - is o b u ty l-a d e n o s in e . J . V ir o l . 22 , 160-167•
JACQUEMONT, B ., GARCIA, A. and HUPPERT, J . (1 9 8 0 ) . N uclear
p r o c e ss in g o f v i r a l h ig h -m o lecu ia r -w e ig h t RNA in c e l l s
in fe c te d w ith herp es sim plex v ir u s typ e 1 .
J . V iro l.- 3 8 2 -3 8 9 .
JAMIESON, A.T. and SUBAK-SHARPE, J.H . (1 9 7 4 ) . B ioch em ical
s tu d ie s on h erpes sim plex v ir u s - s p e c i f i e d deoxypyrim id ine
k in a se a c t i v i t y . J . gen . V ir o l . 2 4 , 4 8 1 - 4 9 2 .
JAMIESON, A .T ., GENTRY, G.A. and SUBAK-SHARPE, J .H . (1 9 7 4 ) .
In d u ction o f both thym idine and d e o x y c y tid in e k in a se
a c t i v i t y by herpes v ir u s e s . J . gen . V ir o l . 24 , 4 65 -480 .

JAMIESON, A .T ., HAY, J . and SUBAK-SHARPE, J .H . (197o a j.
H erp esv iru s p r o te in s : in d u c tio n o f n u c le o s id e
p h o sp h o tra n sfera se a c t i v i t y a f t e r h erp es s im p le x '
v ir u s i n f e c t i o n . J . V ir o l . 1£ , 10 5 6 -1 0 5 9 .
JAMIESON, A .T ., MacNAB, J .C .M ., PERBAl, B. and CLEMENTS, J .B .
(1 9 7 6 b ). V iru s s p e c i f ie d enzyme a c t i v i t y and RNA
s p e c ie s in h erp es sim p lex v ir u s typ e 1 transform ed
mouse c e l l s . J . gen . V ir o l . 22 ., 4 9 3 -5 0 8 .
JAMIESON, A. T. and SUBAK-SHARPE, J.H . (1 9 7 8 ) . I n t e r a l l e l i c
com plem entation o f m utants o f h erp es s im p lex v ir u s
d e f i c i e n t in d eoxypyrim id in e k in a se a c t i v i t y .
V iro lo g y 8 £ , 1 0 9 -1 1 7 .
JARIWALLA, R .J . , AURELIAN, I . and TS'O, P .O . (1 9 8 0 ).I
Tum origenic tra n sfo rm a tio n induced by a s p e c i f i c
fragm ent o f DNA from h erp es sim p lex type 2 .
P ro c . N a t l . Acad. S c i . USA J J f 2279-2283 .
JEAN, J-H . and BEN-PORAT, T. (1 9 7 6 ) . Appearance In v iv o o f
s in g le - s tr a n d e d complem entary ends on h erp esv iru s DNA.
P ro c . N a t l . Acad. S c i . USA 22» 2674 -2678 .
JEAN, J -II ., BLANKENSHIP, M.L. and BEN-PORAT, T. (1 9 7 7 ).
R e p lic a t io n o f h e r p e sv ir u s DNA. I . E lec tro n
m icro sco p ic a n a ly s is o f r e p l i c a t iv e s tr u c tu r e s .
V ir o lo g y 22> 2 8 1 -2 9 1 .
JEAN, J -H ., VOSUIMUKA, N ., E ARUM A, T. and POLTKIN, S .A . (1 9 7 8 ).. «■* “I ' 1
I n t r a c e l lu la r form s o f p a r e n ta l HCMV genome a t e a r ly
s ta g e s o f th e in f e c t i o u s p r o c e s s . V iro lo g y 8 6 , 2 8 1 -2 8 6 .
JERKQPSKY, in. A ., DOEERSEN, M.J. and GREER, S . (1 9 8 0 ).
I n h ib it io n o f th e r e p l i c a t io n o f human cy to m eg a lo v iru s
by brom odeoxycytid ine in th e absen ce o f d e te c ta b le
in c r e a s e s in brom odeoxycytid ine k in a se a c t i v i t y .
I n te r v ir o lo g y 1±, 2 3 3 -2 3 8 .
JOERE, J .T . , SCRAPPER, P i . and PARRIS, D .S . (1 9 7 7 ) . G en etics
o f r e s is ta n c e to p h osp h on oacetic a c id in s t r a in KOS o f
h erpes sim p lex v ir u s typ e 1 . J . V ir o l . 22 , 8 3 3 -8 3 6 .

JOPRE, J .T . , COURTNEY, R .J . and SCRAPPER, P .A . (1 9 8 1 ).
A dominant l e t h a l te m p e r a tu r e -s e n s it iv e mutant o f
herpes sim p lex v ir u s type 1 . V iro logy 111 , 1 7 3 -1 9 0 .
JONES, P .C ., HAYWARD, G .S. and ROIZMAN, B. (1 9 7 7 ) . Anatomy
o f herpes sim p lex v ir u s DNA. V II. <x RNA i s homologous
to n on con tiguou s s i t e s in both th e L and S components
o f v i r a l DNA. J . V ir o l . 2 1 , 2 6 8 -2 7 6 .
JONES, P .C . and ROIZMAN, B. (1 9 7 9 ) . R eg u la tio n o f h e r p e sv iru s
m acrom olecular s y n t h e s is . V III . The tr a n s c r ip t io n
program c o n s i s t s o f th r e e p h ases du rin g which both
e x te n t o f t r a n s c r ip t io n and accu m u lation o f RNA in th e
cytop lasm are r e g u la te d . J . V ir o l . 2i> 299-314-
JONGENEEL, C.V. and BACHENHEIMER, S .L . (198Q ). R e p lic a t io n
o f herpes sim p lex v ir u s ty p e 1 DNA in p erm ea b ilized
in fe c te d c e l l 3 . N u c le ic A oids R esearch 8 , 1661-1673 .

KAERNER, H .C ., MAICHLE, I . B . , OTT, A. and SCHRODER, C.H/' (1 9 7 9 ) .
O rig in o f two d i f f e r e n t c la s s e s o f d e f e c t iv e HSV-1
A n g e lo t t i DNA. N u c le ic A cids R esearch 6 , 1467-1478 .
KAPLAN, A .S . and VATTER, A.E. (1 9 5 9 ) . A com parison o f herpes
sim p lex and p seu d o ra b ies v ir u s e s . V ir o lo g y 2> 3 9 4 -4 0 7 .
KAPLAN, A .S . and BEN-PORAT, T. (1 9 6 4 ) . Mode o f r e p l ic a t io n o f
p seu d o ra b ies v ir u s DNA. V iro lo g y 2 £ , 9 0 -9 5 .
KAPLAN, A .S . and BEN-PORAT, T. (1 9 7 6 ) . S y n th e s is o f p r o te in s
in c e l l s in f e c t e d w ith h e r p e sv ir u s . X I. S u lfa te d ,
s tr u c tu r a l g ly c o p r o te in s . V iro lo g y 22* 5 61 -563 .
KAZAMA, F.Y. and SCHDRNSTEIN, K.L. (1 9 7 2 ) . H erp es-typ e v ir u s
p a r t i c l e s a s s o c ia te d w ith a fu n g u s . S c ien ce 1 7 7 . 6 9 6 -6 9 7 .
KEIR, H.Ll. and GOLD, E. (1 9 6 3 ) . DNA n u c le o t id y l tr a n s fe r a s e
and d eo x y r ib o n u c lea se from c u ltu r e d c e l l s in fe c te d w ith
h erp es s im p lex v ir u s . B iochim . b iop h ys. A cta 22> 2 6 3 -2 7 6 .
KEIR, H.M., SUBAK-SHARPE, H ., SHEDDEN, W .I.H ., WATSON, D.H.
and WILDY, P . (1 9 6 6 ) . Im m unological ev id en oe f o r a
s p e c i f i c DNA polym erase produced a f t e r in f e c t io n w ith
herp es sim p lex v ir u s . V iro lo g y JO, 1 5 4 -1 5 7 .
KELLER, J.ivi., SPEAR, P.G . and ROIZMAN, B. (1 9 7 0 ) . P r o te in s
s p e c i f ie d by h erpes sim p lex v ir u s . I I I . V iru ses
d i f f e r in g in t h e i r e f fe c t* o n the s o c ia l behaviour o f
in f e c t e d c e l l s s p e c i f y d i f f e r e n t membrane g ly c o p r o te in s .
P ro c . N a t l . Acad. S c i . USA 6 J , 8 6 5 -8 7 1 .
KELLER, J.M. (1 9 7 6 ) . The e x p r e ss io n o f th e syn gene o f herpes
sim p lex v ir u s typ e 1 . I I . R equirem ents o f m acrom olechlar
s y n t h e s i s . V iro lo g y 2 2 , 40 2 -4 0 9 .
KHAN, N .C ., MELNICK, J .L . and BISWAL, N. (1 9 7 8 ) . The f a t e o f
p a r e n ta l h e r p e sv ir u s DNA a f t e r in f e c t io n w ith normal and
p h o to in a c tiv a te d v ir io n s . J . gen . V ir o l . J2., 1 9 5 -2 0 0 .

lOEPP, E .D ., BACHENHEIMER, S .L . and ROIZMAN, B. (1 9 7 1 ) .
S iz e , com p osition and s tr u c tu r e o f th e d e o x y r ib o n u c le ic
a c id o f Herpes sim plex v ir u s su b ty p es 1 and 2 .
J . V ir o l . 8 , 1 2 5 -1 3 2 .
KIEPF, E .D ., IIOYER, B ., BAC HEN HEIM ER, S .L , and ROIZMAN, B. (1 9 7 2 ) .
G en etic r e la te d n e s s o f ty p e 1 and ty p e 2 h erpes sim p lex
v ir u s e s . J . V ir o l . 2., 7 3 6 -7 4 5 .
KILLINGTON, R .A ., YEO, J . , HONESS, R.W ., WATSON, D .H ., DUNCAN,
B .E ., HALLIBURTON, I.\Y. and MUMPORD, J . (1 9 7 7 ) .
Com parative a n a ly s is o f th e p r o te in s and a n t ig e n s o f
f i v e h e r p e s v ir u s e s . J . gen . V iro l* J2 , 2 9 7 -3 1 0 ,
KILLINGTON, R .A ., RANDALL, R .E ., YEO, J . , HONESS, R.W .,tHALLIBURTON, I.W .'and WATSON, D.H. (1 9 7 8 ) . O b serva tion s
on a n t ig e n ic r e la te d n e s s between v ir u s e s o f th e herp es
sim p lex " n eu troseron " . In "O ncogenesis and H erp esv iru ses H I
p a r t I , pp. 1 8 5 -1 9 4 . E d ited by G. de (fh6 , W. H enle and
P. Rapp. IARC S c i e n t i f i c P u b lic a t io n s , n o . 24 , Lyon.
KILPATRICK, 3 . A ., HUANG, E -S. and PAGANO, J .S . (1 9 7 6 ) . A n a ly s is
o f cy to m eg a lo v iru s genomes w ith r e s t r i c t i o n en d o n u clea ses
H in d ll l and EcoRI. J . V ir o l . 18 , 1 095 -1105 .
KILPATRICK, 3 . A. and HUANG, E -S . (1 9 7 7 ) . Human cy to m eg a lo v iru s
genome: p a r t ia l d en a tu ra tio n map and o r g a n iz a tio n o f
genome se q u en ce s . J . V ir o l . 2£ , 2 6 1 -2 7 6 .
KIM, K .S ., SAPIENZA, V .J . , CARP, R .I . and MOON, H.M. (1 9 7 6 a ).
A n a ly s is o f s tr u c tu r a l p r o te in s o f p u r if ie d murine
cy to m e g a lo v ir u s . J . V ir o l . 1 2 , 906-915*
KIM, K .S ., SAPIENZA, V .J .’, CARP, R .I , and MOON, H.M. (1 9 7 6 b ).
A n a ly s is o f s tr u c tu r a l p r o te in s o f p u r if ie d human
c y to m e g a lo v ir u s . J . V ir o l. 2 0 , 6 0 4 -6 1 1 . .

KINTNER, C.R. and SUGDEN, B. (1 9 7 9 ) . The s tr u c tu r e o f th e
t e n c in i o f th e DNA o f E p ste in -B a rr v ir u s . C e ll 17., 661-671
KIRKWOOD, J . , GEARING, G. and ODD, L .J . (1 9 7 2 ) . D em onstration
o f group and t y p e - s p e c if ic " a n t ig e n s o f h erp es v ir u s e s .
In "O ncogenesis and H erp esv iru ses" , pp . 479.
E d ited by P.M. B ig g s , G. de The and L.N. Payne. IARC
S c i e n t i f i c P u b lic a t io n s , n o . 2 , Lyon.
KIT, S . and DUBBS, D.H. (1 9 6 3 a ) . A c q u is it io n o f thym idine
k in a se a c t i v i t y by h erp es sim p lex in f e c t e d mouse
f ib r o b la s t c e l l s . Biochem. b iop h ys. R es. Commun. 1 1 , 55-59
KIT, S . and DUBBS, D.R. (1 9 6 3 b ). N o n -fu n c tio n a l thym idine
k i n a s e i s t r o n in brom odeoxyuridine r e s i s t a n t s t r a in s
o f herp es s im p lex v ir u s . Biochem. b iop h ys. R es. Commun.
1 J , 5 0 0 -5 0 4 .
KLEMPERER, H .G ., HAYNES, G .R ., SHEDDEN, W1 I .H . and WATSON, D.H.
(1 9 6 7 ) . A v ir u s s p e c i f i c thym idine k in a se in BHK 21
c e l l s in f e c t e d w ith h erp es sim p lex v ir u s .
V iro lo g y J l , 1 2 0 -1 2 8 .
KNIPE, D.M ., RUYECIIAN, W.T., ROIZMAN, B. and HALLIBURTON, I.W .
(1 9 7 8 ) . M olecu lar g e n e t ic s o f h erp es sim p iex v ir u s :
dem on stration o f r e g io n s o f o b lig a to r y and n o n o b lig a to r y
id e n t i t y w ith in d ip lo id r e g io n s o f th e genome by sequence
rep lacem ent and in s e r t io n . P roc . N a t l . Acad. S c i . USA
U f 3896-3900 .
KNIPE, D.M ., RUYECHAN, W.T., HONESS, R.W. and ROIZMAN, B. (1 9 7 9 ) .
M olecu lar g e n e t ic s o f herp es sim p lex v ir u s : th e term in a l
3 seq u en ces o f th e L and S components are o b l ig a t o r i ly
id e n t i c a l and c o n s t i t u t e a p a r t o f a s t r u c tu r a l gene
mapping predom inantly in th e S com ponent. P roc . N a t l .
Acad. S c i . USA £ 6 , 4534 -4538 .

KNIPE, D.i»i., BATTEHSON, W., NOSAL, C ., ROIZMAN, B. and BUCHAN, A.
(1 9 8 1 ). M olecu lar g e n e t ic s o f h erp es sim p lex v ir u s . VI.
C h a r a c te r iz a tio n o f a te m p e r a tu r e -s e n s it iv e mutant
d e fe c t iv e in th e e x p r e ss io n o f a l l e a r ly v i r a l gene
p ro d u cts . J . V ir o l . J8 , 5 3 9 -5 4 7 .
KNOPP, K-W, (1 9 7 9 ) . S y n th e s is o f h erp es sim p lex v ir u s DNA in «* * ^ ^
i s o la t e d chrom atin . B ioch em istry 18 , 1776 -1780 .
KNOWLES, fi.W. and PEESOH, S . (1 9 7 6 ) . E f f e c t o f 2 -d e o x y g lu c o se ,
glucosam ine and mannose on c e l l f u s io n and th e g ly c o p r o te in s
o f h erp es sim p lex v ir u s . J . V ir o l . 1 8 , 644 -651 .
KOLBER, A.R. (1 9 7 5 ) . In v i t r o s y n th e s is o f DNA In n u c le i
i s o la t e d from human lu n g c e l l s in fe o te d w ith h erp es
sim plex typ e I I v ir u s . J . V ir o l . JJ5, 3 2 2 -3 3 1 .
KOMANO, T. and SINSHEIMER, R.L. (1 9 6 9 ) . P rep a ra tio n and
p u r i f ic a t io n o f 0X-RP component I .
Bfochim. b iop h ys. Acta 1 5 5 # 2 9 5 -2 9 8 .
KOUSOUIAS, K .G ., PERSON, S . and HOLLAND, T.C. (1 9 7 8 ) . Timing
o f some o f th e m olecu lar e v e n ts req u ired f o r c e l l fu s io n
induced by h erp es sim p lex v ir u s typ e 1 . J . V ir o l. 2 J , 505-51^
KOZAK, M. and ROIZMAN, B. (1 9 7 4 ) . R e g u la tio n o f h erp q sv iru s
m acrom olecular s y n th e s is : n u c le a r r e te n t io n o f non
tr a n s la te d v i r a l RNA seq u en ces . P ro c . N a t l . Acad. S c i .
USA 21 , 4322-4326 .
KOZAK, M. and ROIZMAN, B. (1 9 7 5 ) . RNA s y n th e s is in c e l l s
in fe c te d w ith h erpes sim p lex v ir u s . IX. E vidence fo r
accum ulation o f abundant symmetric t r a n s c r ip t s in
n u c le i . J . V ir o l . 1 J , 3 6 -4 0 .
KUDLER, L. and HYmAN, R.W. (1 9 7 9 ) . E xonuclease I I I d ig e s t io n
o f herpes sim p lex v ir u s DNA. V iro lo g y £2!, 6 8 -8 1 .

LADIN, B .E ., BLANKENSHIP, w .L . and BEN-PORAT, T. ( I 9 6 0 ) .
R e p lic a t io n o f h e r p e sv ir u s DNA. V. m aturation o f
con ca tem eric DNA o f p seu d o ra b ies v ir u s to genome le n g th
i s r e la te d to c a p s id fo rm a tio n . J . V ir o l . JJ , I I 5 I - H 6 4 .
LAKEMAN, A.B. and OSBORN, J .E . (1 9 7 9 ) . S iz e o f in f e c t io u s DNA
from human and murine c y to m e g a lo v ir u s . J . V ir o l . JO,
4 1 4 -4 1 6 . :
LANDO, D.L. and RYHINER, M-L. (1 9 6 9 ) . P ou rvo ir in f e c t ie u x
du DNA d * h erp esv iru s hom in is en c u ltu r e c e l l u l a i r e .
Compte Kendu de 1 'Academic de S c ie n c e s de P a r is
' 26^, 5 2 7 -5 3 0 .
DANDY, A. and ROSS, V/. (1 9 7 7 ) . V ir a l in te g r a t io n and e x c is io n ;
s tr u c tu r e o f th e lambda a t t s i t e s . S c ie n c e 197 , 1 1 4 7 -1160 .
LANGELIER, Y ., DECHAMPS, M. andBtJTTIN, G. (1 9 7 8 ) . A n a ly s is o f- — ^
dCMP deam inase and CDP r ed u c ta se l e v e l s in ham ster c e l l s
in f e c t e d by h erp es sim p lex v ir u s . J . V ir o l . 26 , 5 4 7 -555 .
LEE, G.T-Y. and SPEAR, P .G . (1 9 8 0 ) . V ir a l and c e l lu l a r f a c t o r s
th a t in f lu e n c e c e l l fu s io n induced by herpes sim p lex
v ir u s . V ir o lo g y 1 0 7 , 40 2 -4 1 4 .
LEE, L .P . , KIEPP, E .D ., BAC HEN HEIMER, S .L . , ROIZMAN, B ., SPEAR,
P .G ., BURMESTER, B.R. and NAZERIAN, K. (1 9 7 1 ) . S iz e and
co m p o sitio n o f Marek’ s d is e a s e v ir u s d e o x y r ib o n u c le ic
a c id . J . V ir o l . J , 2 8 9 -2 9 4 .
LEE, Y .S . , TANAKA, A ., LAU, R .Y ., NONOYAIIA, M.. and RABIN, H. (1 9 8 0 ) .
Com parative s tu d ie s o f h e r p e sv ir u s papio (baboon
h e r p e sv ir u s ) DNA and E p ste in -B arr v ir u s DNA.
J . gen . V ir o l . J l , 2 4 5 -2 5 3 .
■LEE, Y .S ., NONOYAMA, M. and RABIN, H. (1 9 8 1 ) . C o lin ear
r e la t io n s h ip s o f h e r p e sv ir u s papio DNA to E p ste in -B arr
v ir u s DNA. V ir o lo g y 1 1 0 , 24 8 -2 5 2 .

LEINBACH, S .S and SUMMERS, W.C. (1 9 8 0 ) . The s tr u c tu r e o f
h erp es sim p lex v ir u s type 1 DNA as probed by m icro co cca l
n u c le a se d ig e s t io n . J . gen . V ir o l . J l , 4 5 -6 0 .
LEMASTER, S . and ROIZMAN, B. (1 9 8 0 ) . herpes sim plex v ir u s
p h o sp h o p ro te in s . I I . C h a r a c te r iz a t io n o f th e v ir io n
p r o te in k in a se and o f th e p o ly p e p tid e s p h osp horylated
in th e v ir io n . J . V ir o l . JJ , '798-811,
LEUNG, W-C., DB10CK, K ., SMILEY, J . and BACCHETTI, S . (1 9 8 0 ) .
Herpes sim p lex v ir u s thym idine k in a se t r a n s c r ip t s are
ab sen t from both n u c le u s and cytop lasm during in f e c t io n
in th e p resen ce o f c y c lo h e x im id e . J . V ir o l . 26, 361 -365 .
LINDAHL, T ., ADAMS, A ., BJURSELL, G ., BORNKAMM, G.W.,
KASCHAA-DIERICH, C. and JEHN, V. (1 9 7 6 ) . C o v a len tly
c lo s e d c ir c u la r duplex DNA o f E p stein rB arr v ir u s in a
human lymphoid c e l l l i n e . J . m ol. B io l . 1 0 2 , 5 1 1 -530 .
LINNEMANN, C .C .J r ., BUCHMAN, T .G ., LIGI-IT, I , J . , BALLARD, J .L .
and ROIZMAN, b . (1 9 7 8 ) . T ransm ission o f h erp es sim p lex
v ir u s type 1 in a n u rsery fo r the newborn: i d e n t i f i c a t io n
o f v i r a l i s o l a t e s by DNA f in g e r p r in t in g . L ancet 1 , 96 4 -9 6 6 .
LOCKER, H. and PRENKEL, N. (1 9 7 8 ) . The DNA o f s e r i a l l y
p assaged h erpes sim plex v ir u s : o r g a n iz a t io n , o r ig in ,
and homology to v i r a l RNA. In "O ncogenesis and
H erp esv iru ses I I I " , p a r t I , pp . 7 5 -8 5 . E d ited by
G. de The, Y/. Henle and P. Rapp. I ARC S c i e n t i f i c
P u b lic a t io n s , n o . 24, Lyon.
LOCKER, H. and PEENKEL, N. (1 9 7 9 a ). S tru c tu re and o r ig in o f
d e f e c t iv e genomes con ta in ed in s e r i a l l y passaged herpes
sim p lex v ir u s type 1 ( J u s t in ) , J . V ir o l . 29, 1065-1077 .

MACKEM, S . and ROIZMAN, B. ( I 9 6 0 ) . R eg u la tio n o f h e r p e sv iru s
m acrom olecular s y n th e s is : t r a n s c r ip t io n - in i t i a t io n s i t e s
and domains o f <* g e n e s . P roc . N a t l . Acad. S c i . USA
TLf 7122-7126 .
MAITLAND, N .J . and McDOUGALL, J.K . (1 9 7 7 ) . B ioch em ica l
tra n sfo rm a tio n o f mouse c e l l s by fragm ents o f h erp es
sim p lex v ir u s DNA. C e l l 11, 2 3 3 -2 4 1 .
MANIATIS, T ., JEPEREY, A. and VAN DE SANDE, H. (1 9 7 5 ) . Chain
le n g th d e term in a tio n o f sm all d o u b le - and s in g le -s tr a n d e d
DNA m o lecu les by p o lyacry lam id e g e l e le c t r o p h o r e s is .
B io ch em istry 1£ , 3787-3794 .
MANSBRVIGI, R ., SPEAR, P.G . and BUCHAN, A. (1 9 7 7 ) . C e l l fu s io n
induced by herp es sim plex v ir u s i s promoted and su pp ressed
by d i f f e r e n t g ly c o p r o te in s . P ro c . N a t l . Acad, S c i . USA
'1±, 3913-3917 .
MANTEI, N ., BOLL, W. and WEISSMAN, C. (1 9 7 9 ) . R abbit /3 -g lo b in
mRNA p rod u ction in mouse L c e l l s transform ed w ith c lon ed
r a b b it /J -g lo b in chromosomal DNA. Nature (London) 281, 4 0 -4 6 .
MAO, J.C .H . and ROEISHAW, E .E . (1 9 7 5 ) . Mode o f in h ib i t io n o f
h erp es sim plex v ir u s DNA polym erase by p h osp h on oaceta te .
B ioch em istry 1 £ , 5475-5479 .
MARC IANO -C A6R AL, R .,D I X , K .D .,‘ CABRAL, G.A. and COURTNEY, R .J .
(1 9 7 7 ) . E f f e c t s o f c y to c h a la s in B on the m aturation o f
h erp es sim p lex v ir u s typ e 1: an u l t r a s t r u e t u r a l
in v e s t ig a t io n . V iro lo g y 76 , 8 6 0 -8 6 5 .
mAREK, J . (1 9 0 7 ) . M u ltip le nervenentzundung ( p o ly n e u r it is )
b e i hu hn em . D eutsche T ie r a r z t l ic h e VYochenschrift
12, 41 7 -4 2 3 .

MARSDEN, U .S ., CROMBIE, I .E . and SUBAK-SHARPE, J .N . (1 9 7 6 ).
C ontrol o f p r o te in s y r t h e s i s in h e r p e s v ir u s - in fe c te d
c e l l s : a n a ly s is o f the p o ly p e p tid e s induced by w ild -
type and s ix te e n tem pera1t u r e - s e n s i t iv e m utants o f
HSV s tr a in 1 7 . J . gen . V ir o l . £ 1 , 3 4 7 -3 7 2 .
MARSDEN, H .S ., STOW, N .D ., PRESTON, V .G ., TIMBURY, M.C. and
WILKIE, N.M. (1 9 7 8 ) . P h y s ic a l mapping o f herp es sim plex
v iru s-in d u c ed p o ly p e p t id e s . J . V ir o l . 28 , 624-642 .
MATHEWS, R .E .P . (1 9 7 9 ) . Third r ep o r t o f th e in te r n a t io n a l
com m ittee on taxonomy o f v ir u s e s . C la s s i f i c a t io n and
nom enclature o f v ir u s e s . I n te r v ir o lo g y 1 2 , 1 2 9 -2 9 6 .
MAXAM, A.M. and GILBERT, W. (1 9 8 0 ). Sequencing e n a - la b e lle d
DNa w ith b a s e - s p e c i f i c chem ical c le a v a g e s .
Methods in Enzymology 6J>, 4 9 9 -5 6 0 .
MELENDEZ, L .V ., HUNT, R .D ., DANIEL, M .D., GARCIA, E.G. and
ERASER, c .E .O . (1 9 6 9 ) . H erp esv iru s s a im ir i . I I .
E xp erim en ta lly induced m alignant lymphoma in p r im a tes.
Laboratory Animal Care 12., 3 7 8 -3 8 6 .
MELENDEZ, L .V ., RUNT, R .D ., KING, N.W., BARAHONA, H .H ., DANIEL
M J)., ERASER, C.E.O. and GARCIA, F.G. (1 9 7 2 ).
H erpesv iru s a t e l e s , a new lymphoma v ir u s o f monkeys.
Nature New B io l . 2 5 5 * 1 8 2 -1 8 4 .
MELNICK, J .L . , RABIN, E.R. and JENSON, A .B. (1 9 6 8 ) .
I n tr a c e l lu la r h e r p e sv iru s a g g reg a te in th e form o f a
pentagon al d ipyram idal c r y s t a l - l i k e s tr u c tu r e .
J . V ir o l . 2 , 7 8 -8 0 .
MILLER, R .H . , KUELER, L ., PRYSTOWSKY, J . and HYMAN, R .J . (1980)
Comparison o f fo ld b a ck seq u en ces o f herpes sim plex ty p e s
1 and 2 DNA. J . gen . V ir o l. 46, 5 1 -6 2 .

MILLER, R .L ., IL fIS , J . r . and RAPP, P. (1 9 7 7 ) . D i f f e r e n t ia l
e f f e c t o f a r a b in o sy lfu r a n o sy l-th y m in e on th e
r e p l ic a t io n o f human n e r p e s -v ir u s e s . J . V ir o l . 2J2, o79-684 .
MILLETTE, R .L . and KLAIBER, R. (1 9 8 0 ) . Gene e x p r ess io n o f
h erp es sim p lex v ir u s . I I . UV r a d io lo g ic a l a n a ly s is o f
v ir a l t r a n s c r ip t io n u n i t s . J . V ir o l . 2 ! , 604 -614 .
MIOVIC, Ivi.L. and PIZER, L .I . (1 9 8 0 ) . C h a r a c te r iz a tio n o f RNA
sy n th e s iz e d in i s o la t e d n u c le i o f h erp es sim plex v ir u s
type 1 - in f e c t e d KB c e l l s . J . V ir o l . 22* 567 -571 .
MIYAMOTO, K. and MORGAN, 0 . (1 9 7 1 ) . S tru c tu re and developm ent
o f v ir u s e s a s ob served in the e le c tr o n m icroscop e . IX.
Entry and u n co a tin g o f h erp es sim p lex v ir u s .
J . V ir o l . 8 , 9 1 0 -9 1 8 .
MOCARSKI, E .S ., POST, L .E . and ROIZMAN, B. (1 9 8 0 ) . M olecular
e n g in e e r in g o f the h erp es sim plex v ir u s genome:
in s e r t io n o f a second L-S ju n c tio n in to th e genome
ca u ses a d d it io n a l in v e r s io n s . C e l l 22, 243-255*
MORGAN, 0 . , JONES, E .P ., HOLDEN, M. and ROSE, H.M. (1 9 5 8 ) .
In tra n u c le a r c r y s t a l s o f h erp es sim p lex v ir u s observed
w ith the e le c tr o n m icroscope* V iro lo g y 568-571 .
MORGAN, C ., ROSE, H.M., HOLDEN, M. and JONES, E .P . (1 9 5 9 ) .
E le c tr o n m icro sco p ic o b se r v a t io n s on the developm ent
o f h erp es sim p lex v ir u s . J . exp . Med. 1 1 0 , 643-656 .
MORGAN, C ., ROSE, H.M. and MEDNIS, B. (1 9 6 8 ) . E lec tro n
m icroscopy o f h erp es sim p lex v ir u s . I . E n try .
J . V ir o l . 2 , 5 0 7 -5 1 6 .
MOHRlSOn, J.*.i. and 1CEIR, H.M. (1 9 6 8 ) . A new DNA-endonuclease
in c e l l s in f e c t e d w ith h erp es v ir u s : p a r t ia l
p u r i f ic a t io n and p r o p e r t ie s o f the enzyme.
J . gen . V ir o l . £ , 3 3 7 -3 4 7 .

MORSE, L .S . , BUGhi»iAN, T .G ., ROIZMAN, B. and SCHAEFER, P .A . (1 9 7 7 ) .
Anatomy o f h erp es sim plex v ir u s DNA. IX. Apparent
e x c lu s io n o f some p a r e n ta l DNA arrangem ents in th e
g en era tio n o f in t e r t y p ic (HSV-1 x HSV-2) recom b in an ts.
J . V ir o l. 2R, 2 3 1 -2 4 8 .
MORSE, L ., PEREIRA, L ., ROIZMAN, B. and SCHAFFER, P .A . (1 9 7 8 ) .
Anatomy o f h erp es sim p lex v ir u s (HSV) DNA. X. Mapping
o f v ir a l genes by a n a ly s is o f p o ly p e p tid e s and fu n c t io n s
s p e c i f ie d by HSV-1 x HSV-2 recom b in an ts.
J . V ir o l. 26 , 3 89 -410 .
MOSMANN, f .K . and HUDSON, J .3 . (1 9 7 3 ) . Some p r o p e r t ie s o f
th e genome o f murine cy to m eg a lo v iru s (MCV).
V iro lo g y ££ , 13 5 -1 4 9 .
MOSS, B ., GERSCHOV/ITZ, A ., STRINGER, J .R ., HOLLAND, L .E. and
WAGNER, E.A. (1 9 7 7 ) . 5 f -te r m in a l and in t e r n a l
m ethylated n u c le o s id e s in h erp es sim p lex v ir u s typ e 1
mRNA. J . V ir o l. §2 , 2 3 4 -2 3 9 .
MOSS, H., CII ARTE AND, P . , TIMBURY, M.C. and HAY, J . (1 9 7 9 ) .
Mutant o f h erp es sim p lex typ e 2 w ith tem p eratu re-
s e n s i t iv e l e s io n s a f f e c t in g v ir io n th e r m o s ta b i l ity and
DNase a c t i v i t y : i d e n t i f i c a t io n o f th e l e t h a l m utation
and p h y s ic a l mapping o f the nuc~~ l e s i o n .
J . V irol.- j32, 140—146 .
IviOUTTET, M-E., GUETARD, D. and BECHET, J.M. (1 9 7 9 ) . Random
c lea v a g e o f in tr a n u c le a r h erp es sim plex v ir u s DNA by
m icrococca l n u c le a s e . FEBS L e t te r s 1 0 0 , 10 7 -1 0 9 .
MULLER, W.E.G., ZAHN, R .R ., ARENDES, J. and FALKE, D. (1 9 7 9 a ).
O lig o r ib o n u c le o t id e i n i t i a t o r s fo r h erp es sim p lex v ir u s
DNA s y n th e s is i n v iv o and in v i t r o . V iro lo g y 98 , 2 0 0 -2 1 0 .

MULLER, W.K.G., ZAHN, K .A ., ARENDES, J . and PALKE, D. (1979b ).
P h o sp h o ry la tio n o f arab in o fu ran osy lth ym in e in
n o n - in fe c te d and h e r p e sv ir u s (TK and TK ) - in f e c t e d
c e l l s . J . gen . V ir o l . 42 , 2 6 1 -2 7 1 .
M UN YON, W., KRAISEDBURD, E . , DAVIS, D. and MANN, J . (1 9 7 1 ) .
T ran sfer o f thym idine ldlnase to thym idine k in a s e le s s
L c e l l s by in f e c t io n w ith u l t r a v io l e t ir r a d ia te d
h erp es sim p lex v ir u s . J . V ir o l. 2 , 8 1 3 -8 2 0 .
MURPHY, F .A ., HARRISON, A.K. and WHITFIELD, S.G . (1 9 6 7 ) .
In tr a n u c le a r form ation o f f i la m e n ts in h erpes v ir u s .
hom in is in f e c t io n o f m ice . Arch. g e s . V ir u sfo r sc h .
21 , 46 3 -4 6 8 .
MUitRAY, H.n. and BISV/AL, N. (1 9 7 4 ) . S y n th e s is o f h erp es
sim p lex v ir u s typ e 1 (HSV-1) DNA in i s o la t e d n u c le i .
I I . C ovalen t l in k a g e o f RNA to n a sce n t v i r a l DNA.
I n te r v ir o lo g y £ , 1 4 -2 2 .
MURRAY, B. K ., BENYESli-MELNICK, M. and BISWAL, N. (1 9 7 4 ) ._ *» - '-V
E arly and l a t e v i r a l - s p e c i f i c p o ly r ib o so m a l RNA in
h e r p e sv ir u s -1 and -2 in f e c t e d r a b b it k id ney c e l l s .
B iochim . b io p h y s . Acta 361 , 2 0 9 -2 2 0 .
MURRAY, B .k . , BISWAL, N ., BOOKQUT, J .B . , LANFORD, R .J . ,
COURTNEY, R .J . and MELNICk, J .L . (1 9 7 5 ) . C y c lic
appearance o f d e f e c t iv e in t e r f e r in g p a r t i c l e s o f
herpes sim p lex v ir u s and th e con com itan t accum ulation
o f e a r ly p o ly p e p tid e VP175. I n te r v ir o lo g y 1 7 3 -1 8 4 .
MURRAY, K. and MURRAY, N .E. (1 9 7 5 ) . Phage lambda r e c e p to r
chromosomes f o r DNA fragm ents made w ith r e s t r i c t i o n
endon u clease I I I o f Haemophilus in f lu e n z a e and
r e s t r i c t i o n en d on u clease I o f E sc h e r ic h ia c o l i .
J , m ol. B io l . 98 , 531 -564 .

NAIB, Z.M., NAHMIAS, A .J . and JOSEY, W.E. (1 9 6 6 ) . C ytology
and h is to p a th o lo g y o f c e r v ic a l h erp es sim p lex in f e c t io n .
Cancer 19 , 1026-1031*
NAZERIAN, K. (1 9 7 4 ) . DNA c o n f ig u r a t io n in th e core o f Marek*s
d is e a se v ir u s . J . V ir o l . 12 , 1 1 4 8 -1150 .
NEUBAUER, R .II., RABIN, H ., STRAND, B .C ., NONOYAMA, M. and
NBLSON-REES, W.A. (1 9 7 9 ) . E sta b lish m en t o f a
lym ph oblasto id c e l l l in e and i s o l a t i o n o f an E p ste in -B arr
(EBV) r e la te d v ir u s o f g o r i l l a o r ig in . J . V ir o l . 31, 8 4 5 -8 4 8 .
N i l , S . , MORGAN, 0 . and ROSE, H.M. (1 9 6 8 a ) . E lec tro n m icrosoopy
o f herpes sim p lex v ir u s . I I . Sequence o f developm ent.
J . V ir o l. 2 , 517-536 .
N i l , S . , ROSENKRANZ, H .S ., MORGAN, C. and ROSE, H.M. (1 9 6 8 b ).
E lec tro n m icroscopy o f herp es sim p lex v ir u s . I I I .
E f fe c t o f hydroxyurea. J . V ir o l . 2 , 1163 -1171 .
N i l , S . (1 9 7 1 ). E lec tro n m icro sco p ic o b s e r v a t io n s on PL c e l l s
in fe c te d w ith herpes sim plex v ir u s . I I . Envelopm ent.
Biken J . 1 4 , 3 2 5 -348 .
N i l , S. and YASUDA, I . (1 9 7 5 ) . D e te c t io n o f v i r a l c o r e s having
to r o id s tr u c tu r e s in e ig h t h e r p e s v ir u s e s .
Biken J . 18 , 4 1 -4 6 .
NISHIOKA, Y. and SILVERSTEIN, S . (1 9 7 7 ) . D egradation o f
c e l lu la r mRNA during in f e c t io n lay h erpes sim plex v ir u s .
P roc. N a t l . Acad. S c i . USA ££» 2370-2374*
NISHIOKA, Y. and SILVERSTEIN, S . (1 9 7 8 ) . Requirement o f
p r o te in s y n th e s is fo r th e d egrad ation o f h o s t mRNA in
Friend erythroleu kem ia c e l l s in f e c t e d w ith h erpes
sim plex v ir u s typ e 1 . J . V ir o l . 2 7 , 619-627 .

N0RHI1D, B . , LUDWIG, H. and ROTT, R. (1 9 7 8 ). I d e n t i f i c a t io n
o f a common a n tig e n o f herpes sim plex v ir u s , bovine
h erp es m am m ilitis v ir u s and B v ir u s . J . V ir o l . 26 , 7 1 2 -717 .
NORRILD, B. and VESTRRGAARD, B .F . (1 9 7 9 ) . Im m unoelectrophoretic
i d e n t i f i c a t io n and p u r i f ic a t io n o f h erp es sim p lex v ir u s
a n t ig e n s r e le a s e d from in f e c t e d c e l l s in t i s s u e c u ltu r e .
I n te r v ir o lo g y 1 1 , 1 0 4 -1 1 0 .

OAiCiiS, J.±S., ILTIS, J .P . , dIMAN, R.tf. and RAPP, P. (1 9 7 7 ) .
A n a ly s is by r e s t r i c t i o n enzyme c le a v a g e o f human
v a r ic e l la - z o s t e r v ir u s DNAs. V iro lo g y 82 , 3 53 -361 .
ODA, H ., MORI, R ., MIIAZOiiO, j . and IWASAKA, T. (1 9 7 9 ) .
E f f e c t s o f a n t im e ta b o lit e s on th e p rod u ction o f
tu b u la r s tr u c tu r e s in Vero c e l l s in f e c t e d w ith herpes
sim plex v ir u s typ e 2 . Arch. V ir o l . 62, 17 5 -1 8 7 .
0*DONOVAN, C. and ROIZMAN, B. (1 9 6 1 ). d i f f e r e n t ia t io n o f th e
p ro cess o f c e l l i n f e c t io n w ith f r e e h erp es sim plex
v ir u s from th e recru itm en t o f c e l l s in to v ir u s - in d u c e d
s y n c y t ia in X -rayed c e l l c u l t u r e s . V iro logy 15., 3 7 4 -3 7 6 .
OKADA, K., FUJBlO'iO, Y ., NAKANISHI, Y .H ., ONUlviA, M. and
I.IIKAMI, T. (1 9 7 9 ) . F urther study on th e th r e e -
d im en sion a l s tr u c tu r e o f the core o f M arek's d is e a s e
and h e r p e sv ir u s o f tu rk ey . Arch. V ir o l . £9, 137-144 .
OLD, L .J . , BOYSE, E .A ., OETTGEN, H .F ., BE HARVEN, E . ,
GEER IN G, G., WILLIAMSON, B. and CLIFFORD, P . (1 9 6 6 ).
P r e c ip i t a t in g an tib od y in human serum to an a n tig e n
p r e sen t in c u ltu r e d B u r k it t ’ s lymphoma c e l l s .
P roc . N a t l . Acad. S c i . USA £6 , 1699 -1704 .
OLSHEVSKY, U. and BECKER, Y. (1 9 7 2 ). S u rface g ly c o p e p tid e s
in the en velop e o f herpes sim p lex v ir io n s .
V iro lo g y 2 0 , 27 7 -2 7 9 .

PARA, M.S'., BAUCKE, R .n . arid SPEAR, P.G. (1 9 8 0 ).
Immunoglobulin G (F c)-b in d in g r e c e p to r s on v ir io n s o f
h erp es sim plex type 1 and tr a n s fe r o f th e se r e c e p to r s
to th e c e l l su r fa c e by i n f e c t i o n . J . V ir o l 24, 512-520 .
PARRIS, D .S . , DIXON, R .A .F . and SCHAFFER, P .A . ( I 9 6 0 ) .
P h y s ic a l mapping o f h erp es sim plex v ir u s type 1 t s
m utants by marker r e sc u e : c o r r e la t io n o f th e p h y s ic a l
and g e n e t ic maps. V iro lo g y 1 0 0 , 2 7 5 -2 8 7 .
PAULS, F .P . and DOWDLE, W.R. (1 9 6 7 ) . A s e r o lo g ic a l stu dy o f
h e r p e sv ir u s hom in is s t r a in s by m ic r o n e u tr a lisa t io n
t e s t s . J . Immun. 9 8 , 9 4 1 -9 4 7 .
PEDERSEN, M ., TALLEY-BROWN, S . and MILLETTE, R.L. (1 9 8 1 ) .
Gene e x p r e ss io n o f h erp es s im p lex v ir u s . I I I . E f fe c t
o f a r a b in o sy l-a d e n in e on v i r a l p o ly p e p t id e .s y n th e s is .
J . V ir o l . 7 1 2 -7 1 9 .
PB1LICKH, A ., V/IGLKH, U ., AXEL, R. and SILVERSTEIN, S . (1 9 7 8 ) .
The t r a n s f e r and s ta b le in te g r a t io n o f th e HSV
thym id ine k in a se gene in to mouse c e l l s . C e l l 1 4 , 1 3 3 -141 .
PENNINGTON, T.H. and McCREA, M.A. (1 9 7 7 ) . ' P r o c e s s in g o f a
p seu d o ra b ie s v ir u s - in d u c e d p r o te in which i s g ly c o s y la te d ,
su lp h a ted and e x c r e te d . J . gen . V ir o l. 24 , 1 5 5 -1 6 5 .
PERDUE, M .L., KEMP, M .C., RANDALL, C.C. and O’CALLAGHAN, D .J .
(1 9 7 4 ) . S tu d ie s on th e m olecu lar anatomy o f the L-M
s t r a in o f equ ine h erp es v ir u s typ e 1 . P r o te in s o f the
n u c le o c a p s id and in t a c t v ir io n . V iro lo g y g ) , 2 0 1 -2 1 6 .
PEREIRA, 1 . , WOLFF, K .ii. , FENWICK, M. and ROIZMAN, B. (1 9 7 7 ) .
R eg u la tio n o f hei’p e s v ir u s m acrom olecular s y n th e s is . V.
P r o p e r t ie s o f p o l y p e p t i d e s made in HSV-1 and HSV-2
in f e c t e d c e l l s . v ir o lo g y 72, 73 3 -7 4 9 .

PIGNATTI, P .P ., CASSAI, E. and BER'TAZZONI, U. (1 9 7 9 ) . Herpes
sim plex v ir u s DNA s y n th e s is in a p a r t i a l l y p u r if ie d
so lu b le e x ti'a c t from in f e c t e d c e l l s . J . V ir o l.
22 , 1033-1036 .
PIGNATTI, P .P . and CASSAI, E. (1 9 8 0 ) . A n a ly s is o f h erpes
sim plex v ir u s n u c le o p r o te in com plexes e x tr a c te d from
in fe c te d c e l l s . J . V ir o l. 2S> 8 1 6 -8 2 8 .
PIZER, L . I . , COHEN, G.H. and EISENBERG, R .J . (1 9 8 0 ) . E f fe c t
o f tunicam ycin on h erp es sim p lex v ir u s g ly c o p r o te in s
and in f e c t io u s v ir u s p ro d u ctio n . J . V ir o l . 24 > 1 4 2 -1 5 3 .
PLUMMER, G. (1 9 6 4 ) . S e r o lo g ic a l com parison o f the h erpes
v ir u s e s . B r it i s h J . exp . P a th o l. 42> 1 3 5 -1 4 1 .
PONCE DE LEON, M., EISENBERG, R .J . and COHEN, G.H. (1 9 7 7 ) .
R ib o n u c leo tid e r ed u c ta se from h erp es s im p les v ir u s
(ty p e s 1 and 2) in fe c te d and u n in fe c te d KB c e l l s :
p r o p e r t ie s o f the p a r t ia l l y p u r if ie d enzym es.
J . gen . V ir o l . 26 , 1 6 3 -1 7 3 .
POST, I . E . , COLLEY, A .J . , MOCARSKI, E .S . and ROIZMAN, B. (1 9 8 0 ) .
C loning o f r e i t e r a te d and n o n r e ite r a te d h erp es sim p lex
v ir u s 1 sequ en ces a s BamHI fragm en ts. P roc . N a t l . Acad.
S c i . USA XL* 4201-4205.'
POST, I . E . , luACKEM, S. and ROIZMAN, B. (1 9 8 1 ) . R eg u la tio n o f
qc genes o f h erpes sim plex v ir u s : e x p r e ss io n o f ch im eric
genes produced by fu s io n o f thym idine k in a se w ith o<
gene prom oters. C e ll 2£, 555 -566 .
POWELL, D .J . (1 9 7 9 ). S tru c tu re and e x p r ess io n o f th e
p seu d orab ies v ir u s genome. PhD t h e s i s , U n iv e r s ity o f
Glasgow.

POWELL, K .L ., BUCHAN, A ., SIM, C. and WATSON, D.H. (1 9 7 4 ) .
T y p e -s p e c if ic p r o te in in herpes sim p lex v ir u s en velop e
r e a c t s w ith n e u t r a l i s in g a n tib o d y . Nature (London)
2 4 9 . 3 6 0 -3 6 1 .
POWELL, K.L. and WATSON, D.H. (1 9 7 5 ) . Some s tr u c tu r a l
a n t ig e n s o f h erp es s im p lex v ir u s typ e 1 .
J . g en . V ir o l . 2 £ , 1 6 7 -1 7 8 .
POWELL, K.L. and PURIFOY, D.J.M . (1 9 7 6 ) . DNA-binding p r o te in s
o f c e l l s in f e c t e d by h erp es sim p lex v ir u s type 1 and
type 2 . I n te r v ir o lo g y 2 2 5 -2 3 9 .
POWELL, K.L. and PURIFOY, D.J.M . (1 9 7 7 ) . N o n stru c tu ra l p r o te in s
o f h erp es s im p lex v ir u s . I . P u r if ic a t io n o f thei
induced DNA p o lym erase . J , V ir o l. 2^, 618-626 .
PRESTON, C.M. and NEWTON, A.A, (1 9 7 6 ) . The e f f e c t s o f herp es
sim p lex v ir u s typ e 1 on c e l lu l a r DNA dependent RNA.i
polym erase a c t i v i t i e s . J . gen . V ir o l . 22* 47 1 -4 8 2 .
PRESTON, C.M. (1 9 7 7 ) . C e l l - f r e e s y n th e s is o f h erp es sim p lex
v ir u s-c o d e d p yrim id in e d e o x y r ib o n u c leo s id e k in a se
enzyme. J . V ir o l . §2 , 4 5 5 -4 6 0 .
PRESTON, C.M. (1 9 7 9 ) . C ontrol o f h erpes sim plex v ir u s type 1
mRNA s y n th e s is in c e l l s in f e c t e d w ith w ild -ty p e v ir u s
or tn c te m p e r a tu r e -s e n s it iv e mutant tsK .
J . V ir o l . 22., 2 7 5 -2 8 4 .
PRESTON, C.M. and McGEOCIi, D .J . (1 9 8 1 ) . I d e n t i f i c a t io n and
mapping o f th e two p o ly p e p tid e s encoded w ith in th e
h erp es s im p lex v ir u s typ e 1 thym idine k in a se gene
se q u en ces . J . V ir o l . 593 -605 .
PRESTON, V .G ., DAVISON, A .J . , MARSDEN, H .S ., TIMBURY, M .C.,
SUBAK-SHARPE, J .H . and WILKIE, N.M. (1 9 7 8 ). Recom binants
between h erp es sim p lex v ir u s ty p e s 1 and 2: a n a ly s e s o f
genome s tr u c tu r e s and e x p r ess io n o f im m ed ia te -ear ly
p o ly p e p t id e s . J . V ir o l . 28 , 4 9 9 -517 .

PRINGLE, C .R ., HOWARD, D.K. and HAY, J* (1 9 7 3 ) . Tem perature-
s e n s i t iv e m utants o f p seu d o ra b ies v ir u s w ith
d i f f e r e n t i a l e f f e c t s on v i r a l and h o s t DNA s y n t h e s is .
V iro lo g y ££, 495 -505 .
PRITCHETT, R .F ., HAYWARD, S .D . and KIEFF, E. (1 9 7 5 ) . DNA o f
E p stein -B arr v ir u s . I . Comparative s tu d ie s o f th e
DNA o f E p ste in -B arr v ir u s from HR-1 and B95-8 c e l l s :
s i z e , s tr u c tu r e and r e la t e d n e s s . J . V ir o l . 12 , 556r569.
PRITCHETT, R .F . (1 9 8 0 ) . DNA n u c le o t id e sequence h e te r o g e n e ity. * - ^
between th e Towne and AD169 s t r a in s o f cy to m e g a lo v ir u s .
J . V ir o l . £6 , 1 5 2 -1 6 1 .
PURCHASE, H .G ., OKAZAKI, W. and BURMESTER, B.R. (1 9 7 1 ) . F ie ld
t r i a l s w ith th e h erp es v ir u s o f tu rk ey s (HVT), s t r a in
FC 126, a s a v a c c in e a g a in s t Marek*s d i s e a s e .
P o u ltr y S c ien ce 20.* 7 7 5 -7 8 3 .
PURIFOY, D.J.M . and BENYESH-MELNICK, M. (1 9 7 5 ) . DNA-* ■* /-s
polym erase in d u c tio n by DNA-negative tem p eratu re-
s e n s i t iv e m utants o f herp es sim p lex v ir u s type. 2 .
V iro logy 68, 3 7 4 -386 .
PURIFOY, D.J.M . and POWELL, K.L. (1 9 7 7 ) . H erpes sim p lex v ir u sip. -■* ■» ” " ^ , T
! DNA polym erase a s the s i t e o f ph osp hon oacetate
s e n s i t i v i t y : te m p e r a tu r e -s e n s it iv e m utan ts.
J . V i r o l . - 2 4 , 4 70 -477 .
PURIFOY, D.J.M . and POWELL, K.L. (1 9 8 1 ) . T e m p e ra tu r e -sen sitiv e
mutants in two d i s t i n c t com plem entation groups o f
herpes sim plex v ir u s typ e 1 s p e c ify th e r m o la b ile DNA
polym erase. J . gen . V ir o l . 2 1 7 -2 2 2 .

RAAB-TRAUB, N ., FRITCHETT, it. and KIEFF, E. (1 9 7 8 ) . D> A o f
E p ste in -B a rr v ir u s . I I I . I d e n t i f i c a t io n o f r e s t r i c t i o n
enzyme fragm en ts which c o n ta in DNA seq u en ces which d i f f e r
among s t r a in s o f EBV. J . V ir o l . 2J , 38 8 -3 9 8 .
RAAB-TRAUB, N . , DAMRAUGH, T. and KIEFF, E. (1 9 8 0 ) . DNA o f■« * - —
E p ste in -B a rr v ir u s . V III . A n a ly s is and m olecu lar
ep id em io logy o f th e " a d d itio n a l" DNA o f two B u rk itt tumor
i s o l a t e s o f EBV. C e l l 22 , 2 5 7 -2 6 7 .
RABIN, II ., NEUBAUER, R .H ., HOPKINS, R .F . I l l and NONOYAMA, M.
(1 9 7 8 ) . F urther c h a r a c te r iz a t io n o f a h e r p e sv ir u s -
p o s i t iv e orangutan c e l l l i n e and com parative a s p e c ts o f
v it r o tra n sfo rm a tio n w ith lym photrop ic o ld world
prim ate h e r p e s v ir u s e s . I n t l . J . C ancer. 21 , 7 6 2 -7 6 7 .
RAND, T.H. and BEN-PORaT, T. (1 9 8 0 ) . D is t r ib u t io n o f sequ en ces
homologous to th e DNA o f herp es sim plex v ir u s , ty p e s 1
and 2 , in th e genome o f p seu d o ra b ies v ir u s .
I n te r v ir o lo g y 1 2 , 4 8 -5 3 .
RAPP, F . , I1T IS, J .P . , OAKES, J .E . and HYMAN, R.V/. (1 9 7 7 ) .
A n o v e l approach to stu d y the DNA o f h erp es z o s te r
v ir u s . I n te r v ir o lo g y 8 , 2 7 2 -2 8 0 .
RAWLS, W.E., LAUREL, D ., MELNICK, J .L . , GLICKSMAN, J.M. and
KAUFMAN, R.H. (1 9 6 8 ) . A sea rch f o r v ir u s e s in smegma,
prem alignan t and early m alignant c e r v ic a l t i s s u e s . The
i s o l a t i o n o f h e r p e sv ir u se s w ith d is ,in c t a n t i0
p r o p e r t ie s . American J . E p id em io l. 8 £ , 647-655*
REYES, G .E ., LA DEMINA, R ., HAYWARD, b .D . and HAYWARD, G.S. (1 9 7 9 ).
M orp h olog ica l tra n sfo rm a tio n by DNA fragm ents o f human
h erpes v ir u s e s : ev id en ce fo r two d i s t i n c t tran sform in g
r e g io n s in HSV-1 and HSV-2 and la c k o f c o r r e la t io n w ith
b ioch em ica l tr a n s fe r o f the thym idine niriase g en e .
Cold Spring Harbor Symp. Quant. B io l . £4, 629-641 .

RICE, M., STRINGER, J . , SWANSTROM, R. and WAGNER, E.K. (1 9 7 6 ) .
The a s s o c ia t io n o f th e i n f e c t i n g DNA o f herpes sim plex
v ir u s w ith c e l lu l a r com ponents. V iro lo g y 20 , 1 8 5 -1 8 9 .
RICHARDS, J .C . , HYMAN, R.W. and RAPP, F. (1 9 7 9 ) . A n a ly s is o f
th e DNAs from seven v a r io e l la - z o s t e r v ir u s i s o l a t e s .
J . V ir o l . 22 , 8 1 2 -8 2 1 .
RIGBY, P .W .J ., DIECKMANN, M., RHODES, C. and BERG, P. (1 9 7 7 ) .
L a b e llin g d e o x y r ib o n u c le ic a c id to h igh s p e c i f i c a c t i v i t y
in v i t r o by n ic k - t r a n s la t io n w ith DNA polym erase I .
J . m ol. B io l . 112, 2 3 7 -2 5 1 .
RITCHIE, D .A ., BROWN, S .M ., SUBAK-SHARPE, J .H . and JAMIESON,'
A.T. (1 9 7 7 ) . H ete ro z y g o sis and g e n e t ic recom b ination
in h erp es sim plex type 1 v ir u s . V iro lo g y 8 2 , 3 23 -333 .
RIXON, F .J . (1 9 7 7 ) . S tu d ie s on h erpes sim p lex v ir u s DNA
s y n t h e s is . PhD t h e s i s , U n iv e r s ity o f Glasgow.
ROBERTS, R .J . (1 9 8 1 ) . R e s t r ic t io n and m o d if ic a t io n enzymes and
t h e ir r e c o g n it io n seq u en ces . N u c le ic A cids R esearch 9 ,
r 7 5 -r 9 6 .
ROIZMAN, B. and ROANE, P .R .J r . (1 9 6 4 ) . The m u lt ip l ic a t io n o f
herpes sim p lex v ir u s . I I . The r e la t io n between p r o te in
s y n th e s is and th e d u p lic a t io n o f v i r a l DNA in in fe c te d
IIEp-2 c e l l s . V iro logy 22 , 2 6 2 -2 6 9 .
ROIZMAN, B ., BORMAN, G.S. and KAMALI-ROUSTA, M. (1 9 6 5 ) .
M acrom olecular s y n th e s is in c e l l s in f e c t e d w ith h erpes
sim p lex v ir u s . Nature 2 0 6 , 1374-1375 .
ROIZMAN, B. and FUrLONG, D. (1 9 7 4 ) . The r e p l ic a t io n o f
h e r p e s v ir u s e s . In "Comprehensive V iro logy" , v o l . 3 ,
pp. 2 2 9 -4 0 3 . E dited by H. F raenkel-C onrat and R.R.
Wagner. Plenum P r e s s , New York.

ROIZMAN, B ., KOZAK, M., HONESS, R.W. and IIAYWARD, G. (1 9 7 4 ) .
R eg u la tio n o f h e r p e sv ir u s m acrom olecular s y n th e s is :
ev id en ce fo r m u lt i le v e l r e g u la t io n o f h erp es sim plex
1 RNA and p r o te in s y n t h e s is . Cold Spring Harbor
Symp. Quant. BLol. 2 2 ., 687-701 .
ROIZMAN, B ., CARMICHAEL, S . , DE THE, G .B ., MASIC, M.,
NAHMIAS, A ., PLOY/RIGHT,'w., RAPP, P . , SHELDRICK, P . ,
TAKASHI, M., TERNI, M. and WOLFE, K. (1 9 7 8 ) .
P r o v is io n a l c l a s s i f i c a t i o n o f h e r p e s v ir u s e s . In
"O ncogenesis and H erp esv iru ses I I I " , v o l . I I , pp.
£ 0 7 9 -1 0 8 2 . E d ited by G. de The, F, Rapp and W. H en le .jf _
i IARC S c i e n t i f i c P u b lic a t io n s , n o . 24 , Lyon.
ROIZMAN, B. (1 9 7 9 ) . The s tr u c tu r e and iso m e r iz a t io n o f
h erp es s im p lex v ir u s genom es. C e l l 1 6 , 4 8 1 -4 9 4 .
ROIZMAN, B.* JACOB, R .J . , KNIPE, D.M ., MORSE, L .S . and
RUYECHAN, L .S . (1 9 7 9 ) . On th e s tr u c tu r e , fu n c t io n a l
e q u iv a le n c e , and r e p l i c a t io n o f th e fo u r arrangem ents
■ o f herp es sim p lex v ir u s DNA. Cold Spring Harbor
Symp. Quant. B io l . 42 , 8 0 9 -8 2 6 .
ROLTON, H.A. and KEIR, H.M. (1 9 7 4 ) . D e o x y c y tid y la te deam inase.
E vidence fo r a new enzyme in c e l l s in f e c t e d by the
v ir u s o f h erp es s im p le x . Biochem. J . 1 4 3 , 403 -409 .
ROYSTON, I . and AURElIAN, L. (1 9 7 0 ). Im m unofluorescent
d e te c t io n o f h e r p e sv ir u s a n t ig e n s in e x fo l ia t e d c e l l s
from human c e r v ic a l carcinom a. P roc . N a t l . Acad. S c i .
USA 62, 2 0 4 -2 1 2 .
RUBENSTEIN, A .S ., GRAVELh, M. and DARLINGTON, R. (1 9 7 2 ) .
P r o te in k in a se in enveloped h erpes sim p lex v ir io n s .
V iro lo g y 22., 2 8 7 -2 9 0 .

RUBENSTEIN, A .S. and KAPLAN, A .S . (1 9 7 5 ) . E lec tro n
m icroscop ic s tu d ie s o f th e DNA o f d e f e c t iv e and
standard pseuc’o r a b ie s v ir io n s . V iro logy 66, 385-392 .
RUSSELL, YMC., GOLD, E ., KEIR, H.M., OMURA, H ., WATSON, D.H.
and WILDY, P . (1 9 6 4 ) . The growth o f h erp es sim p lex
v ir u s and i t s n u c le ic a c id . V iro logy 2 2 , 1 0 3 -1 1 0 .
RUYECHAN, W.T., MORSE, L .S . , KNIPE, D.M. and ROIZMAN, B. (1 9 7 9 ) .
M olecular g e n e t ic s o f herpes sim plex v ir u s . I I .
Mapping o f th e major v i r a l g ly c o p r o te in s and o f th e
g e n e tio l o c i s p e c ify in g th e s o c ia l behaviour o f
in fe c te d c e l l s . J . V ir o l . 67 7 -6 9 7 .
RYMO, L. and FORSHLUM, S. (1 9 7 8 ) . C leavage o f E p ste in -B a rr
v ir u s DNA by r e s t r i c t i o n en d o n u clea ses EcoRI , HindI I I
and BamHI. N u c le ic A cids R esearch 2 , 1387-1402 .

SANDRI-GOId)IN, R.M. , LEVINE, M. and GLORIOSO, J . C. (1 9 8 1 ) .
Method f o r in d u c t io n o f m u tation s in p h y s ic a l ly d e fin e d
r e g io n s o f th e h erp es sim p lex v ir u s genome.
J . V ir o l . 2 8 , 4 1 -4 9 .
SAROV, I . and FRIEDMANN, A. (1 9 7 6 ) . E lec tro n m icroscopy o f
human cy to m eg a lo v iru s DNA. Arch. V ir o l . 22* 3 4 3 -3 4 7 .
SARMIENTO, M., HAFFEY, M. and SPEAR, P .A . (1 9 7 9 ) . Membrane
p r o te in s s p e c i f ie d by h erp es sim p lex v ir u s e s . I I I .
R ole o f g ly c o p r o te in VP7 (B2) in v ir io n i n f e c t i v i t y .
J; V ir o l . 29 , 1 1 4 9 -1 1 5 8 .
SASAKI, Y ., SASAKI, R ., COHEN, G.H. and PIZER, i . I . (1 9 7 4 ) .
Rl\iA polym erase a c t i v i t y and in h ib i t io n in h e r p e sv ir u s -
in f e c t e d c e l l s . I n te r v ir o lo g y 2? 1 4 7 -1 6 1 .
SATO, S . , HUTCHISON, C.A. I l l .and HARRIS, J . I . (1 9 7 7 ) .
A th erm ostab le s e q u e n c e -s p e c if ic endon u clease from
Thermus a q u a t ic u s . P roc . N a t l . Acad. S c i . USA 74 , 542-546 .
SCHAFFER, P .A ., ARON, G.m., EISWA1, N. and BENYESH-MELNICK, M.
. (1 9 7 3 ) . T e m p e ra tu r e -sen sit iv e m utants o f herpes sim plex
v ir u s type I s i s o l a t i o n , com plem entation and p a r t ia l
c h a r a c te r iz a t io n . V iro logy £2 , 5 7 -7 1 .
SCHAFFER, P .A ., TEVETHIA, ivi.J. and BENYESH-MELNICK, M. (1 9 7 4 ) .
R ecom bination betv.een tem pera‘t u r e - s e n s i t iv e m utants o f
herp es sim p lex v ir u s type 1 . V iro lo g y £8 , 21 9 -2 2 8 .
SCHAFFER, P .A ., CARTER. V.C. and TIMBURY, to.C. (1 9 7 8 ) .
C o lla b o r a t iv e com plem entation study o f tem p eratu re-
s e n s i t i v e m utants o f h erp es sim p lex v ir u s ty p e s 1
and 2 . J . V ir o l . 22 , 4 9 0 -504 .
SCHILDKRAUT, C. and LIFSON, S . (1 9 6 5 ) . Dependence o f th e
m e ltin g tem perature o f DNA on s a l t c o n c e n tr a t io n .
B iopolym ers 2» 19 5 -2 0 8 .

SCHILDKRAUf, I . , COOPER, G.M. and GREER, S . (1 9 7 5 ) . S e le c t iv e
in h ib i t io n o f the r e p l i c a t io n o f h erpes sim p lex v ir u s
by 5 -h a logen ated an a logu es o f d e o x y c y tid in e .
M olecu lar Pharm acology 1 1 , 1 5 3 -1 5 8 .
SCHLEHOPER, J .R ., IIAMPl, H. and HABERMEH1, K-0. (1 9 7 9 ) .
D if fe r e n c e s in the morphology o f h erp es sim p lex v ir u s
in f e c t e d c e l l s : I . Comparative scan n in g and tr a n sm iss io n
e le c tr o n m icro sco p ic s tu d ie s on HSV-1 in f e c t e d HEp-2
and c h ick embryo f ib r o b la s t c e l l s . J . gen . V ir o l ,
i i , 433 -442 .
SCHMECKPEPER, B .J . and SMITH, K.D. (1 9 7 2 ) . Use o f formamide in
n u c le ic a c id r e a s s o c ia t io n . B ioch em istry 11 , 1319 -1326 .
SCHWARTZ, J . and ROIZMAN, B. (1 9 6 9 ) . C oncerning th e e g r e s s o f
herpes sim p lex v ir u s from in f e c t e d c e l l s : e le c tr o n and
l i g h t m icro sco p ic o b s e r v a t io n s . V ir o lo g y 28 , 4 2 -4 9 .
SEIF, I . , KHOURY, G. and DHAR, R. (1 9 7 9 ) . BKV s p l ic e sequ en ces
based on a n a ly s is o f p r e fe rr e d donor and a c ce p to r s i t e s .
N u c le ic A cids R esearch 6, 3387-3398 .
SHELDRICK, P . , LAITHIER, M., LANDO, D. and RYHINER, M.L. (1 9 7 3 ) .
I n fe c t io u s DNA from h erpes sim plex v ir u s : i n f e c t i v i t y o f
double and s in g le -s tr a n d e d m o le c u le s . P roc . N a t l .
Acad. S o l . USA 'JO, 362 1 -3 6 2 5 .
SHElDRICK, P . and BERTIIE10T, N. (1 9 7 4 ) . In ven ted r e p e t i t io n s
in the chromosome o f h erp es sim p lex v ir u s . Cold Spring
Symp. Quant. B io l . £9 , 667 -678 .
SHENK, T. (1 9 7 3 ) . C o n stru ctio n o f a v ia b le SV40 v a r ia n t
c o n ta in in g two fu n c t io n a l o r ig in s o f DNA r e p l i c a t io n .
C e ll 12, 791 -798 .
SH10MAI, J . and BECKER, Y. (1 9 7 5 ) . A n a ly s is o f h erp es sim plex
v ir u s DNA sy n th e s iz e d in n u c le i i s o la t e d from 3SC-1 c e l l s .
V iro logy 68, 2 75 -279 .

SHIOMAI, J . , ERIEBMAMH, A. and BECK3E, X. (1 9 7 6 ) . R ep /.ica tiv e
in te r m e d ia te s o f h erp es sim p lex v ir u s DNA.
V iro lo g y 6g , 647-659 .
SHLOMAI, J . , ASHER, Y. and BECKER, Y. (1 9 7 7 ) . In v i t r o
s y n t h e s is o f h erp es sim p lex v ir u s DNA in n u c le i i s o la t e d
from in f e c t e d BSC-1 c e l l s . J . g en • V ir o l . 24 , 2 2 5 -2 3 4 .
SILVERSTEIN, S . , BACHENHEIMER, S .L ., PRENKEL, N. and ROIZMAN, B
(1 9 7 3 ) . R e la t io n sh ip between p o s t - t r a n s c r ip t io n a l
a d e n y la tio n o f herp es sim p lex RNA and m essenger RNA
abundance. P ro c . N a t l . Acad. S c i . USA JO, 2101-2104 .
SILVERSTEIN, S . , MILLETTE, R ., JONES, P . and ROIZMAN, B. (1976)«. tj , .. ^
i RNA s y n th e s is in c e l l s in f e c t e d w ith h erp es sim plex
v ir u s . X II . Sequence co m p lex ity and p r o p e r t ie s o f
. RNA d i f f e r in g in ex ten t, o f a d e n y la t io n .
J . V ir o l . 1 8 , 9 7 7 -9 9 1 .
SILVERSTEIN, S . and ENGE11IARDT, D .L. (1 9 7 9 ) . A lte r a t io n s in
th e p r o te in s y n th e t ic apparatu s o f o e l l s in f e c t e d w ith
. h erp es s im p lex v ir u s . V iro lo g y 334-342 .
SIM, C. and WATSON, D.H. (1 9 7 3 ) . The r o le o f typ e s p e c i f i c
and e r o s s -r e a o t in g s t r u c tu r a l a n tig e n s in th e
n e u t r a l iz a t io n o f h erp es sim p lex v ir u s ty p e s 1 and 2 .
J . g en . V ir o l . 1 £ , 21 7 -2 3 3 .
SKARE, J . , SUMMERS, J .P . and SUMMERS, W.C. (1 9 7 5 ) . S tru ctu re and funcrbion or h erp esv iru ^ geuv I . C. o£
f i v e HSV-1 and two HSV-2 s t r a in s by c le a v a g e s a t e s on
herp es sim p lex v ir u s typ e 1 DNA. J . V ir o l . 2 £ , 7 2 6 -7 3 2 .
SKARE, J . and SUMMERS, W.C. (1 9 7 7 ) . S tru c tu re and fu n c t io n o f
h e r p e sv ir u s ge:vu:es. I I . EcoRI, Xbal . and Hind I I I
end on u clease c le a v a g e s i t e s on herp es sim p lex v ir u s
typ e 1 DNA. V iro lo g y £ 6 5 581 -595 .

SMILEY, J .R . ( I9 6 0 ) , C o n stru etio n in v i t r o and rescu e o f
thym idine k in a s e -d e f ic ie n t d e le t io n m u tation o f herp es
sim p lex v i i u s . Nature (London) 2 8 5 . 5 3 3 -3 3 5 .
SMILEY, J .R . , WAGNER, M .J ., SUMMERS, W.P. and SUMMERS, W.C. (1 9 8 0 ) .
G en etic and p h y s ic a l ev id en ce f o r th e p o la r i t y o f
t r a n s c r ip t io n o f th e thym idine k in ase gene o f herpes
s im p lex v ir u s . V iro lo g y 1 0 2 , 8 3 -9 3 .
SMITH, G .R ., SCHULTZ, D.W. and CRASEMAN, J.M. (1 9 8 0 ).
G en era lized recom b in ation : n u c le o t id e sequence homology
betw een Chi recom b in a tion a l h o ts p o ts . C e ll 1 9 , 7 8 5 -7 9 3 .
SMITH, J .D . and HE HAKVEN, E. (1 9 7 3 ) . Herpes sim p lex v ir u s and
cy tom ega lov iru s r e p l ic a t io n in WI-38 c e l l s . I .
Sequence o f v ir u s r e p l i c a t io n . J . V ir o l . 12 , 9 1 9 -9 3 0 .
SMITH, K.O. (1 9 6 4 ). R e la tio n sh ip between the en velop e and
th e i n f e c t i v i t y o f herp es sim p lex v ir u s . P ro c . Soc.
exp . B io l . Med. 115» 8 1 4 -8 1 6 .
SOEHNER, R .L ., GENTRY, G.A. and RANDALL, C.C. (1 9 6 5 ) . Some
p h y sico ch em ica l c h a r a c t e r i s t i c s o f equine a b o r tio n v ir u s
n u c le ic a c id . ^V irology 2 6 , 3 9 4 -405 .
SOUTHERN, E.M. (1 9 7 5 ). D e te c t io n o f s p e c i f i c seq u en ces among
D N ^fragm ents sep arated by g e l e le c t r o p h o r e s is , '
J . m ol. B io l . 98 , 503 -517 .
SPEAR, P.G . and-ROIZMAN, B. (1 9 6 8 ) . The p r o te in s s p e c i f ie d by
h erpes sim plex v ir u s . I . Time o f s y n t h e s is , t r a n s fe r
in to n u c le i , and p r o p e r t ie s o f p r o te in s made in
p r o d u c t iv e ly in fe c te d c e l l s . V iro logy £6 , 5 45 -555 .
SPEAR, P.G . and ROIZMAN, B. (1 9 7 0 ) . The p r o te in s s p e c i f ie d
by h erp es sim plex v ir u s . IV. The s i t e o f g ly c o s y la t io n
and accum ulation o f v i r a l membrane p r o te in s . P roc.
N a t l . Acad. S c i . USA 66, 7 3 0 -7 3 7 .

SPEAR, P .G ., KELLER, J.Jki. and ROIZMAN, B. (1 9 7 0 ) . P r o te in s
s p e c i f ie d by h erp es s im p lex v ir u s . I I . V ir a l
g ly c o p r o te in s a s s o c ia te d w ith c e l lu l a r membranes.
J . V ir o l . 2 , 1 2 3 -1 3 1 .
SPEAR, P .G . and ROIZMAN, B, (1 9 7 2 ) . P r o te in s sp e c if ie d * b y
h erp es sim p lex v ir u s . V. P u r if ic a t io n and s tr u c tu r a l
p r o te in s o f th e h e r p e s v ir io n . J . V ir o l . % 14-3-159.
SPEAR, P.G . (1 9 7 6 ) . Membrane p r o te in s s p e c i f ie d by h erp es
sim p lex v ir u s e s . I . I d e n t i f i c a t io n o f fo u r g ly c o p r o te in
p r e cu rso r s and t h e ir p rod u cts in ty p e 1 in f e c t e d c e l l s .
J . V ir o l . 12 , 9 9 1 -1 0 0 8 .
SPEAR, P .G ., SARMIENTO, M. and MANSERVIGI, R. (1 9 7 8 ) . The• ^
s tr u c tu r a l p r o te in s and g ly c o p r o te in s o f h e r p e sv ir u se s :
a r e v ie w . In “O ncogen esis and H erp esv iru ses I I I ” , v o l l I ,
PP. 1 5 7 -1 6 7 . E d ited by G. de The, W. H enle and P . Rapp#
IARC S c i e n t i f i c P u b lic a t io n s , no . 2 4 , Lyon.
SPRING, S .B . and ROIZMAN, B. (1 9 6 8 ) . H erpes sim p lex v ir u s
p rod u cts in p ro d u ctiv e and a b o r t iv e i n f e c t io n . I I I .
D if f e r e n t ia t io n o f in f e c t io u s v ir u s d er iv ed from n u c leu s
and cytop lasm w ith r e s p e c t to s t a b i l i t y and s i z e .
J . V ir o l . 2 , 9 7 9 -9 8 5 .
STACKPQLE, C.W. (1 9 6 9 ) . H erp es-typ e v ir u s o f th e fr o g r e n a l
adenocarcinom a. I . V irus developm ent in tumo-
t r a n s p la n ts m ain ta ined a t low tem p eratu res.
J . V ir o l . £ , 7 5 -9 3 .
STEGMAN, B ., ZENTGRAP, H ., OTT, A. and SCHRODER*. C.H. (1 9 7 8 ) .
S y n th e s is and packagin g o f h erp es sim p lex v ir u s DNA in
th e cou rse o f v ir u s p a ssa g e s a t h igh m u l t ip l i c i t y .
I n te r v ir o lo g y 10 , 2 2 8 -2 4 0 .

STEIN, S . , TODD, P . and MAHONEY, J . (1 9 7 0 ) . I n f e c t i v i t y o f
h erp es sim p lex v ir u s w ith and w ith o u t e n v e lo p e s .
Canadian J . M ic r o b io l. 1 6 , 953-957*
STERZ, H ., LUDWIG, H. and ROTT, R. (1 9 7 4 ) . Immunologic and
g e n e t ic r e la t io n s h ip between h erp es sim p lex v ir u s and
bovine m am m ilitis v ir u s . I n te r v ir o lo g y 2 , 1 -1 3 .
STEVELY, W.S. (1 9 7 5 ) . V iru s-in d u ced p r o te in s in p seu d o r a b ie s-
in fe c te d c e l l s . I I . P r o te in s o f th e v i s io n and
n u c le o c a p s id . J . V ir o l . 1 6 , 9 4 4 -9 5 0 .
STEVELY, W.S. (1 9 7 7 ) . In v er ted r e p e t i t io n in th e chromosome
o f p seu d orab ies v ir u s . J . V ir o l . 22 , 2 3 2 -2 3 4 .
STINSKI, M .P., MOCARSKI, E .S . and THOMSEN, D.R. (1 9 7 9 ) . DNA o f** ' n 'I
human cy tom ega lov iru s: s i z e , h e te r o g e n e ity and
d e fe c t iv e n e s s r e s u l t in g from s e r i a l u n d ilu te d p a ssa g e ,
J . V ir o l . 2 3 1 -239 .
STOKER, M.G.P, (1 9 5 8 ). Mode o f i n t e r c e l l u l a r tr a n s fe r o f
herpes v ir u s . Nature (London) 1 8 2 . 1525 -1526 .
STOKER, M.G.P. and NEWTON, A.A. (1 9 5 9 ) . The e f f e c t o f h erp es
v ir u s on HeLa c e l l s d iv id in g p arasyn ch ron ou sly .
V iro logy 2> 4 38 -448 .
STOW, N.D. and WILKIE, N.M. (1 9 7 6 ) . An improved tech n iq u e fo r
o b ta in in g enhanced i n f e c t i v i t y w ith herpes- sim plex v ir u s
type 1 DNA* J . gen . V ir o l . 22* 4 47 -458 .
STOW, N.D. and WILKIE, N.M. (1 9 7 8 ) . P h y s ic a l mapping o f
te m p e r a tu r e -s e n s it iv e m u tation s o f h erp es sim plex v ir u s
type 1 by in t e r t y p ic marker r e s c u e . V iro lo g y 22> 1 -1 1 .
STOW, N .D ., SUBAK-SHARPE, J.H . and WILKIE, N.M. (1 9 7 8 ) .
P h y s ic a l mapping o f herpes sim p lex v ir u s typ e 1 m u tation s
by marker r e sc u e . J . V ir o l . 23 , 1 8 2 -1 9 2 .

STRINGER, J .R . , HOLLAND, L .E ., SWANSTROM, R .I . , PIVO, K. and
WAGNER, E.K. (1 9 7 7 ) . Q u a n tita tio n o f h erp es sim plex
v ir u s ty p e 1 RNA in in f e c t e d c e l l s . J , V ir o l . 21, 8 8 9 -9 0 1 .
STRINGER, J .R . , HOLLAND, L .E . and WAGNER, E.K. (1 9 7 8 ) . Mapping
e a r ly t r a n s c r ip t s o f h erp es sim p lex v ir u s typ e 1 by
e le c tr o n m icroscop y . J . V ir o l . 2J , 5 6 -7 3 .
STRNAD, B.C. and AURRLIAN, L. (1 9 7 6 ) . P r o te in s o f h e r p e sv ir u s
type 2 . I . V ir io n , n o n v ir io n and a n t ig e n ic p o ly p e p tid e s
in in f e c t e d c e l l s . V iro lo g y 6^, 4 3 8 -4 5 2 .
STROBEL-PIDLER, M. and PRANCKE, B. (1 9 8 0 ) . A lk a lin e
d eo x y r ib o n u c lea se induced by h erp es sim p lex v ir u s type 1:
co m p o sitio n and p r o p e r t ie s o f th e p u r if ie d enzyme.
V ir o lo g y 1 03 , 4 9 3 -5 0 1 .
SUBAK-SHARPE, H ., BURK, R.R, CRAWPORD, L .V ., MORRISON, J .M .,
HAY, J . and KEIR, II.M. (1 9 6 6 ) . An approach to
e v o lu t io n a r y r e la t io n s h ip s o f mammalian DNA v ir u s e s
through a n a ly s is o f the p a tte r n o f n e a r e s t neighbour
- base se q u en ce s . Cold Spring Harbor Symp. Quant. B io l .
7 3 7 -7 4 8 .
SUBAK-SHARPE, J.H . (1 9 6 9 ) . In “P ro ceed in g s o f th e P ir s t
I n te r n a t io n a l C ongress f o r V ir o lo g y , H e ls in k i , 1968” ,
p . 252 , E d ited by J .L . M eln ick . K arger, B a se l.
SUBAK-SHARPE,. J .H . , BROWN, S .M ., RITCHIE, D .A ., TIMBURY, M.C.,
MacNAB, J .C .M ., MARSDEN, U .S . and HAY, J . (1 9 7 4 ) .
G en etic and b ioch em ica l s tu d ie s w ith h e r p e s v ir u s e s .
Cold S p rin g Harbor Symp. Quant. B io l . 7 1 7 -7 3 0 .
SUBRAMANIAN, K.N. and SHEIMK, T. (1 9 7 8 ). D e f in i t io n o f the
b ou n d aries o f th e o r ig in o f DNA r e p l ic a t io n in sim ian '
v ir u s 40 . N u c le ic A cids Research £ , 3635-3642 .

SUGDEN, B . , SUITERS, W.C. and KLEIN, G. (1 9 7 6 ) . N u c le ic a c id
r e n a tu r a t io n and r e s t r i c t io n endon u clease c lea v a g e
a n a ly s e s show th a t the DNAs o f a tran sform in g and a
n on tran sform in g s t r a in o f E p stein -B arr v ir u s share
ap p rox im ately 90# o f t h e ir n u c le o t id e seq u en ces .
J . V ir o l . 18 , 765 -775 .
SUGDEN, B. (1 9 7 7 ) . Comparison o f E p ste in -B arr v i r a l DNAs in
B u r k itt lymphoma b iop sy c e l l s and in c e l l s c lo n a l ly
transform ed in v i t r o . P roc. N a t l . Acad. S c i . USA
2 £ , 4651 -4655 .
SUGINO, W.M. and KINGSBURY, D.T. (1 9 7 6 ) . DNA h om olog ies between
s t r a in s o f h erp es sim plex v ir u s . V iro lo g y 2i> 605-608 .
SUMMERS, W .P., WAGNER, M. and SUMMERS, W.C. (1 9 7 5 ) . P o s s ib le
p e p tid e ch a in term in ation m utants in thym idine k in a se
gene o f a mammalian v ir u s , herpes sim plex v ir u s .
P ro c . N a t l . Acad. S c i . USA 22, 4081-4084 .
SUTCLIFFE, J .G . (1 9 7 8 ) . pBR322 r e s t r i c t i o n maps d er iv ed from
th e DNA sequence: accu rate DNA s i z e markers up to 4361
n u c le o t id e p a ir s lo n g . N u cle ic A cids R esearch £ , 2721 -2728 .
SWANSTROM, R .I . and WAGNER, E.K. (1 9 7 4 ) . R eg u la tio n o f
s y n t h e s is o f h erp es sim plex type 1 v ir u s mRNA during
p ro d u ctiv e in f e c t io n . V irology, 6 0 , 52 2 -5 3 3 .
SWANSTROM, R .I .- , PIVO, K. and WAGNER, E.K. (1 9 7 5 ) . R e s tr ic te d
t r a n s c r ip t io n o f herpes sim plex v ir u s genome o ccu rr in g
e a r ly a f t e r in f e c t io n and in the p resen ce o f m etab o lic
in h ib i t o r s . V iro logy 66, 1 40 -150 .
SYDISKIS, R .J . and ROIZMAN, B. (1 9 6 6 ). Polysom es and p r o te in
s y n th e s is in c e l l s in fe c te d w ith a DNA v ir u s .
S c ie n c e 153 . 7 6 -7 8 .
SYDISKIS, R .J . and ROIZMAN, B. (1968) . The sed im en ta tio n
p r o f i l e s o f cy top lasm ic polyrib osom es in mammalian c e l l s
p r o d u c t iv e ly and a b o r t iv e ly in fe c te d w ith h erp es sim plexv i r u . c j . V i r n l n rrir ^ A g £ o r r / - t r _ ___________________________________________________________________

TALLEY-BROWN, S . and MILLETTE, R.L. (1 9 7 9 ) . Gene e x p r e ss io n o f
h erp es sim p lex v ir u s . I . A n a ly s is o f cy to p la sm ic RNAs in
in f e c t e d Xeroderma Pigmentosum c e l l s . J . V ir o l . 2i> 7 3 3 -7 4 0 .
TANAKA, T. and WEISBLUM, B. (1 9 7 5 ) . C o n stru ctio n o f a c o l i c i n
El-R f a c to r com p osite p lasm id in v i t r o : means f o r
a m p lif ic a t io n o f d e o x y r ib o n u c le ic a c id . J . B a ct. 1 2 1 , 35 4 -3 6 2 .
THOMAS, M. and DAVIS, H.'.V. (1 9 7 5 ) . S tu d ie s on th e c le a v a g e o f
b acter iop h age lambda DNA w ith EcoRI r e s t r i c t i o n
en d o n u c lea se . J . m ol. B io l . g l , 3 1 5 -3 2 8 .
THOULESS, M.E. (1 9 7 2 ) . S e r o lo g ic a l p r o p e r t ie s o f thym idine
k in a se produced in c e l l s in f e c t e d w ith typ e 1 or type 2
h erp es v ir u s . J . g en . V ir o l . 12 , 3 0 7 -3 1 5 .
THOULESS, i,i.E. and WILDY, P . (1 9 7 5 ) . D eoxypyrim idine k in a se s o f
h erp es sim p lex v ir u s e s ty p e s 1 .and 2: com parison o f
s e r o lo g ic a l and s t r u c tu r a l p r o p e r t ie s . J . gen . V ir o l.
26 , 1 5 9 -1 7 0 .
TIMBURY, i.l.C. (1 9 7 1 ) . Temperature s e n s i t i v e m utants o f h erp es
sim p lex v ir u s type 2 . J . gen . V ir o l . 12 , 37 3 -3 7 6 .
TIMBURY, i.j.C. and SUBAK-SHARPE, J.H . (1 9 7 3 ) . G en etic in t e r a c t io n s
betw een te m p e r a tu r e -s e n s it iv e m utants o f ty p e s 1 and 2
h erpes sim plex v ir u s . J . gen . V ir o l . 1 8 , 3 4 7 -357 .
TIMBURY, J/J.C. and CALDER, L. (1 9 7 6 ) . T e m p e ra tu r e -sen sit iv e
m utants o f h erp es s im p lex v ir u s type 2: a p r o v is io n a l
l in k a g e map based on recom b in ation a n a ly s is .
J . gen . V ir o l . 2Q.» 1 7 9 -1 8 6 .
TJIAN, R. (1 9 7 3 ) . Protein-DNA in t e r a c t io n s a t the o r ig in o f
sim ian v ir u s 40 DNA r e p l i c a t io n . Cold Spring Harbor
Symp. Quant. B io l . £2* 655-662 .
TWIGG, A .J . and SiiERHATT, u .J . ( I 9 6 0 ) . Trans-com plem entable
copy-number m utants o f p lasm id Col E l. Nature (London)
262 , 2 1 6 -2 1 6 .

VAHLNE, A ., SVENNEMIOLM,. B. and 1YCKE, E. (1 9 7 9 ) . Evidence
f o r h erp es sim p lex t y p e - s e le c t iv e r e c e p to r s on c e l lu l a r
plasm a membranes. J . gen . V ir o l . 4 4 , 21 7 -2 2 5 .
VERNON, S .K ., LAWRENCE, W.C. and COHEN, G.H. (1 9 7 4 ) .
M orp hologica l components o f h e r p e sv ir u s . I .
In tercap som eric f i b r i l s and the geom etry o f th e c a p s id .
I n te r v ir o lo g y £ , 2 3 7 -2 4 8 .
VLAZNY, Df A. and FRENKEL, N. (1 9 8 1 ) . R e p lic a t io n o f herpes
sim p lex v ir u s DNA: lo c a l i z a t i o n o f r e p l i c a t iv e
r e c o g n it io n s ig n a ls w ith in d e f e c t iv e v ir u s genom es.
P ro c . N a t l . Acad. S c i . USA 28 , 7 4 2 -7 4 6 .

WADSWOR331, S . , JACOB, R .J . and ROIZMAN, B. (1 9 7 5 ) . Anatomy o f
h erp es sim p lex v ir u s SNA. I I . S iz e , com p osition and
arrangem ent o f in v e r te d term in a l r e p e t i t io n s .
J . V ir o l . 12 , 1487 -1497 .
WADSWORTH, S . , HAYWARD, G .S. and ROIZMAN, B. (1 9 7 6 ) . Anatomy o f
h erp es sim p lex v ir u s DNA. V. T erm inally r e p e t i t iv e
seq u en ces . J . V ir o l . 12 , 5 0 3 -512 .
WAGNER, E.K. and ROIZMAN, B. (1 9 6 9 a ). RNA s y n th e s is in c e l l s
in f e c t e d w ith h erp es sim p lex v ir u s . I I . E vidence th a t
a c la s s o f v i r a l mRNA i s d er iv ed from a h ig h m olecu lar
w eigh t p recu rsor sy n th e s iz e d in th e n u c le u s .
P ro c . N a t l . Acad. S c i . USA 64, 626-633 .
WAGNER, E.K. and ROIZMAN, B. (1 9 6 9 b ). R ib o n u c le ic a c id
s y n th e s is in c e l l s in f e c t e d w ith h erp es sim p lex v ir u s .
I . P a tte r n s o f r ib o n u c le id a c id s y n th e s is in
p r o d u c t iv e ly in fe c te d c e l l s . J . V ir o l. 4 , 3 6 -4 6 .
WAGNER, E.K. (1 9 7 2 ) . E vidence f o r t r a n s c r ip t io n a l c o n tr o l o f
, th e h erp es sim p lex v ir u s genome in in f e c t e d human c e l l s *u
V iro lo g y £ £ , 5 0 2 -5 0 6 .
WAGNER, E .K ., SWANSTROM, R .I . and STAFFORD, M.G. (1 9 7 2 ) .
T r a n sc r ip tio n o f h erpes sim p lex v ir u s genome in human
c e l l s . J . V ir o l . 1 0 , 675 -682 .
WAGNER, M .,. SKARE, J . and SUMMERS, W.C. (1 9 7 4 ) . A n a ly s is o f
DNA o f d e f e c t iv e h erp es sim plex v ir u s type 1 by
r e s t r i c t i o n end on u clease c lea v a g e and n u c le ic a c id
h y b r id iz a t io n . Cold Spring Harbor Symp. Quant. B io l .
39 , 683-686 .
WAGNER, M .J. and SUMMERS, W.C. (1 9 7 8 ). S tru c tu re o f the j o in t
r e g io n and th e term in i o f th e DNA o f herpes sim plex
v ir u s typ e 1 . J . V ir o l. 22, 37 4 -3 8 7 .

WAGNER, M .J ., SHARP, J .A . and SUMMERS, W.C. (1 9 8 1 ).
N u c le o tid e sequence o f th e thym idine k in ase gene o f
h erp es sim plex v ir u s 1 . P roc . N atl* Acad. S c i . USA
28 , 1441 -1445 .
WARD, R .L. and STEVENS, J .G . (1 9 7 5 ) . L ife t im e s o f mRNA
m o lecu les d ir e c t in g th e s y n th e s is o f v i r a l p r o te in s in
h erp es sim p lex v ir u s - in f e c t e d c e l l* J . V ir o l . 1£ , 81-89*
WATHEN, M.W., THOMSEN, D.R. and STINSKI, M. (1 9 8 1 ) .
Temporal r e g u la t io n o f human cy to m eg a lo v iru s tr a n s c r ip t io n
a t im m ed ia te -ea r ly and e a r ly tim es a f to r in f e c t io n .
J . V ir o l . 28, 446 -459 .
WATSON, R .J . and CLEMENTS, J .B . (1 9 7 7 ) . V irus t r a n s c r ip t
mapping s tu d ie s in c e l l s in f e c t e d w ith tem p eratu re-
s e n s i t i v e mutants- o f h erp es sim p lex v ir u s type 1 .
In "O ncogenesis and H erp esv iru ses I I I " , v o l . I , pp. 313 -326 .
E dited by G. de The, W. Henle and F . Rapp. IARC
S c i e n t i f i c P u b lic a t io n s , no* 24 , Lyon.
WATSON, R .J . and CLEMENTS, J .B . ( 1 9 7 8 ) / C h a r a c te r iz a tio n o f
t r a n s c r ip t io n - d e f ic ie n t te m p e r a tu r e -s e n s it iv e m utants o f
h erpes sim plex v ir u s typ e 1 . V iro lo g y 91 , 3 6 4 -3 7 9 .
WATSON, R .J . , PRESTON, C.M. and CLEMENTS, J .B . (1 9 7 9 ) .
S ep ara tion and c h a r a c te r iz a t io n o f h erp es sim p lex v ir u s
type 1 im m ed ia te -ea r ly mRNAs. J . V ir o l, 4 2 -5 2 .
WATSON, R .J . and CLEMENTS, J .B . (1 9 8 0 ) . A h erp es sim plex v ir u s
type 1 fu n c t io n c o n tin u o u sly req u ired fo r e a r ly and l a t e
v ir u s RNA s y n th e s is . N ature (London) 2 8 5 . 329 -331 .
WATSOK, R .J . , SULLIVAN, M. and VANDE WOUDE, G.F. (1 9 8 1 ) .
S tr u c tu r e s o f two s p lic e d h erp es sim p lex v ir u s type 1
im m ed ia te -ea r ly mRNAs which map a t the ju n c tio n s o f the
unique and r e i t e r a te d r e g io n s o f th e v ir u s DNA 3
component. J . V ir o l . 431 -444 .

WAUBKfi, R ., ZUR IIAUSEN, H. and HENLE, W. (1 9 6 8 ) . Chromosomal
and a u to ra d io g ra p h ic s tu d ie s o f c e l l s in fe c te d w ith
h erp es s im p lex v ir u s . J . V ir o l . 2 , 1047-1054 .
WERNER, F - J . , BORNKAUIvi, G.W. and FLECKENSTEIN, B. (1 9 7 7 ) .
Episom al v i r a l DNA in a h e r p e sv iru s sa im ir i-tra n sfo r m e d
lym phoid c e l l l i n e . J . V ir o l. 22., 7 9 4 -8 0 3 .
WESTSTRATE, M.W., GEE1EN, J .l .M .C . and VAN DER NOORDAA, J . (1980)
Human cy to m eg a lo v iru s DNA: p h y s ic a l maps f o r th e*»
r e s t r i c t i o n en d o n u c lea ses B g ll l . Hindi I I and X bal.
J . g en . V ir o l . £9 , 1 -2 2 .
WHARTON, J .H . , HENRY, B .E. and O’CALLAGHAN, B .J . (1 9 8 1 ) .
Equine c y to m e g a lo v ir u s i c u l tu r a l c h a r a c t e r i s t i c s and
p r o p e r t ie s o f v i r a l DNA. V iro lo g y 109 , 1 0 6 -1 1 9 .
WIGLER, M ., SILVERSTEIN, S . , LEE, L .S ., PELLICER, A ., CHENG,
Y.C. and AXEL, R. (1 9 7 7 ) . T ran sfer o f p u r if ie d herpes
v ir u s thym id ine k in a se gene to c u ltu r e d mouse c e l l s .
C e ll 1 1 , 2 2 3 -2 3 2 .
WIGLER, M., SWEET, R ., SIM, G .K ., WOLD, B . , PELLICER A ., LACY,
E ., MANIATIS, T ., SILVERSTEIN,' S . and AXEL, R. (1 9 7 9 ) .
T ransform ation o f mammalian c e l l s w ith gen es from
p r o c a r y o te s and e u c a r y o te s . C e ll 1 6 , 7 7 7 -7 8 5 .
WI1C0X, K.W., KOHK, A ., SKLYANSKAYA, E. and EOIZMAN, B. (1 9 8 0 ) .
H erpes sim p lex v ir u s p h o sp h o p ro te in s . I . Phosphate
c y c le s on and o f f some v i r a l p o ly p e p tid e s and can a l t e r .
t h e ir a f f i n i t y f o r DNA. J . V ir o l . 1 6 7 -1 8 2 .
W1LDY, P . (1 9 5 5 ) . R ecom bination w ith h erpes sim p lex v ir u s .
J . gen . M ic r o b io l. 12 , 3 4 6 -3 6 0 .
WILDY, P . , RUSSELL, W.C. and HORNE, R.W. ( I 9 6 0 ) . The morphology
o f h erp es v ir u s . V iro lo g y 12 , 2 0 4 -2 2 2 .

WILDY, P . , SMITH, C ., NEWTON, A.A. and DENDY, P . (1 9 6 1 ).
Q u a n tita tiv e c y t o lo g ic a l s tu d ie s on HeLa c e l l s in f e c t e d
w ith h erp es v ir u s . V iro lo g y 1 £ , 486 -500 .
WILKIE, N.M. (1 9 7 3 ) . The s y n th e s is and su b stru ctu re o f
h e r p e sv ir u s DNA: th e d i s t r ib u t io n o f a l k a l i - l a b i l e s in g le
strand in te r r u p t io n s in HSV-1 DNA. J . g en . V ir o l.
21 , 4 5 3 -4 6 7 .
WILKIE, N.M ., CLEMENTS, J .B . , MacNAB, J.C.M . and SUBAK-SHARPE,
J.H . (1 9 7 4 ) . The s tr u c tu r e and b io lo g ic a l p r o p e r t ie s o f
h erp es sim p lex v ir u s DNA. Cold Sp rin g Hafrbor Symp.
Quant. B io l . 22> 657-^66.
WILKIE, N.M. (1 9 7 6 ) . P h y s ic a l maps f o r h erp es sim p lex v ir u s
type 1 DNA fo r r e s t r i c t i o n en d o n u clea ses Hind I I I ,
Hpa-1 and X .bad. - J* V ir o l. 2 0 , 2 2 2 -2 3 3 .
WILKIE, N.M. and CORTINI, R. (1 9 7 6 ) . Sequence arrangem ent in - ■» ^
h erp es simxYLex v ir u s type 1 DNA; i d e n t i f i c a t i o n o f
term in a l fragm ents in r e s t r i c t i o n en d on u clease d ig e s t s
and ev id en ce fo r in v e r s io n in redundant and unique
seq u en ces . J* V ir o l . 20 , 2 1 1 -2 2 1 .
WILKIE, N.M ., CORTINI, R. and CLEMENTS, J .B . (1 9 7 7 a ) .
S tr u c tu r a l s tu d ie s and p h ysica l, maps fo r th e herpes
sim p lex v ir u s genome. J . a n t im ic r o b ia l Chemotherapy
2 (Suppl.- A ), 4 7 -6 2 .
WILKIE, N .M ., STOW, N .D ., MARSDEN, H .S ., PRESTON, V ., CORTINI;
R ., TIMBURY, M.C.' and SUBAK-SHARPE, J .H . (1 9 7 7 b ).
P h y s ic a l mapping o f h erp es sim p lex v ir u s coded fu n c t io n s
and p o ly p e p tid e s by marker r escu e and a n a ly s is o f
HSV-l/HSV-2 in t e r t y p ic recom b inan ts. In "O ncogenesis
and Herx^esviruses I I I " , v o l . I , pp. 1 1 -3 1 . E d ited by
G. de The, W. Henle and P. Rapp. IARC S c i e n t i f i c
P u b lic a t io n s , no . 24, Lyon.

WILKIE, N .M ., CLEMENTS, J .B . , BOLL, W., MANTEI, N . , LONSDALE,
D. and WEISSMANN, C. (1 9 7 9 a ) . Hybrid p lasm id s
c o n ta in in g an a c t iv e thym idine k in a se gene o f herpes
sim p lex v ir u s 1 . N u c le ic A cids R esearch 2> 85 9 -8 7 7 .
WILKIE, N.M ., DAVISON, A ., CHARTRAND, P . , STOW, N .D .,
PRESTON, ’v .G . and TIMBURY* M.C. (1 9 7 9 b ). Recom bination
in h erp es s im p lex v ir u s : mapping o f m u tation s and a n a ly s is
o f in t e r t y p ic recom b inan ts; Cold Spring Harbor Symp.
Quant. B io l . £ 2 , 8 2 7 -8 4 0 .
WILKIE,’ N.M ., EGLIN, R .P . , SANDERS, P.G . and CLEMENTS, J .B .• — H — -» -v t
(1 9 8 0 ) . The a s s o c ia t io n o f h erpes sim p lex v ir u s w ith
w squamous carcinom a o f the c e r v ix , and s tu d ie s o f the
v ir u s thym idine k in a se g en e . P ro c . Royal Soc. 2 1 0 , 411-421 .
WOHLRAHB, P. and PRANCKE, B. (1 9 8 0 ) . D eoxyribopyrim idine
tr ip h o sp h a ta se a c t i v i t y s p e c i f i c f o r c e l l s in fe c te d w ith
h erp es sim p lex v ir u s typ e 1 . P ro c . N a t l . Acad. S c i . USA
2 2 , 1872 -1876 .
W0LPr H. and ROIZMAN, B. (1 9 7 8 ) . The r e g u la t io n o f ( s t r u c tu r a l)
p o ly p e p tid e s y n th e s is in h erp es sim p lex v ir u s ty p e s 1
and 2 in f e c t e d c e l l s . In '’O n cogen esis and H erp esv iru ses
I I I ” , v o l . I , pp. 5 2 7 -3 3 6 . E d ited by G. de The, W. Henle
and P. Rajjp. IARC S c i e n t i f i c P u b lic a t io n s , n o . 24, Lyon.
W(iLP, K. and DARLINGTON, R.W. (1 9 7 1 ) . Channel c a t f i s h v ir u s :
a new h e r p e sv iru s o f i c t a l u r id f i s h . J . V ir o l . 8 , 525-533 .
WU, M ., HYMAN, R.W. and DAVIDSON, N. (1 9 7 9 ) . E lectron "
m icro sco p ic mapping o f p r o te in s bound to h erp es sim plex
v ir u s DNA. N u c le ic A cids R esearch 6 , 3427-3441 .

YAMAMOTO, S . , KABUTA, J . , IMAMOTO, M. and MATSUMOTO, H. (1 9 7 5 ) .
G en etic a n a ly s is o f p o ly k a r y o c y to s is by herpes sim plex
v ir u s . I . Experim ents u s in g recom binants from
in t e r t y p ic c r o s s . Kurume Med. J . 22 , 7 1 -7 8 .
YAMAMOTO, S . and KABUTA, H. (1 9 7 7 ) . G enetic a n a ly s is o f
p o ly k a r y c y to s is by h erp es sim p lex v ir u s . I I I .
Com plem entation and recom b in ation between n o n -fu s in g
m utants and c o n s tr u c t io n o f a l in k a g e map w ith regard
to th e fu s io n fu n c t io n . Kurume Med. J . 2±, 16 3 -1 7 2 .
YAMANISHI, K ., OGINO, T. and TAKAHASHI, M. (1 9 7 5 ) . In d u ctio n
o f c e l l u l a r DNA s y n th e s is by a te m p e r a tu r e -s e n s it iv e
mutant o f h erp es sim p lex v ir u s typ e 2 . V iro lo g y
62, 4 5 0 -4 6 2 .
YANAGI, K ., RUSH, M.G. and BIEGELEISEN, K. (1 9 7 9 ) . In te g r a t io n
o f h erp es sim p lex v ir u s type 1 DNA in to th e DNA o f■*
g ro w th -a rrested BHK-21 c e l l s . J . gen . V ir o l . fH, 6 5 7 -6 6 7 .
YEO, J . , KILLINGTON, R .A ., WATSON, D.H. and POWELL, K.L. (1 9 8 1 ) .
S tu d ie s on c r o s s - r e a c t iv e a n t ig e n s in th e h e r p e sv ir u se s .
V ir o lo g y 108, 25 6 -2 6 6 .
ZlfVEIG, M• , HEILMAN, C .J .J r . and HAMPAR, B. (1 9 7 9 ) .
I d e n t i f i c a t io n o f d is u lp h id e - l in k e d p r o te in com plexes
in the n u c le o c a p s id s o f herpes sim plex v ir u s type 2 .
V iro logy 91, 442-450 .
| G L A S G O W I U N I V B R S m i




















