
Mechanisms of adrenomedullin antimicrobial action
6
Mechanisms of adrenomedullin antimicrobial action Robert P. Allaker a, * , Paul W. Grosvenor b , David C. McAnerney a , Barry E. Sheehan a , Bakula H. Srikanta a , Keith Pell c , Supriya Kapas a a Oral Microbiology, Institute of Cell and Molecular Science, Queen Mary, University of London, Newark Street, London E1 2AT, UK b Bioinformatics Group, Centre for Infectious Disease, Institute of Cell and Molecular Science, Queen Mary, University of London, Newark Street, London E1 2AT, UK c Imaging Unit, Institute of Cell and Molecular Science, Queen Mary, University of London, Newark Street, London E1 2AT, UK 1. Introduction The epithelium represents the first barrier against infection with pathogenic bacteria. In health, surfaces of higher eukaryotes, such as plants, invertebrates and vertebrates, including humans, possess a normal microflora which causes no damage to its host. The reason, apart from physical barriers, includes the production of gene-encoded antimicro- bial peptides by epithelial cells [6]. Adrenomedullin (AM) is a 52 amino acid multifunctional peptide [8] produced by a wide variety of tissues and cells. This peptide has many properties in common with other cationic antimicrobial peptides including human b-defensin-2. Such molecules are defined as peptides of 12–50 amino acids with a net positive charge of +2 to +7 [6]. Apart from human AM being 52 amino acid residues in length, AM also has a net positive charge owing to an excess of basic amino acids (arginine and lysine). The molecule also has a single intramolecular disulphide bond between residues 16 and 21, along with an amidated tyrosine at the carboxyl terminus. In common with other antimicrobial peptides including human b-defensin-2, the amphipathic structure characterized by spatially sepa- rated hydrophobic and charged regions, permits bacterial membrane intercalation. Chemically, therefore AM does resemble an antimicrobial peptide, however, its mode of interaction with bacteria is unknown. A variety of modes of action with respect to antimicrobial activity have been proposed for cationic antimicrobial pep- tides, including critical membrane depolarization, creation of physical holes in membranes, induction of hydrolytic enzymes that degrade the cell wall, disturbance of membrane function and disruption of critical intracellular processes [16]. The aim of this study was to gain insights into the mechanism of antimicrobial action of adrenomedullin against the Gram-negative bacterium Escherichia coli and the Gram- positive bacterium Staphylococcus aureus. peptides 27 (2006) 661–666 article info Article history: Received 14 July 2005 Accepted 2 September 2005 Published on line 13 October 2005 Keywords: Adrenomedullin Antimicrobial peptide Structure–function abstract The mechanism of antimicrobial action of the multifunctional peptide adrenomedullin (AM) against Escherichia coli and Staphylococcus aureus was investigated. AM (52 residues) and AM fragments (1–12, 1–21, 13–52, 16–21, 16–52, 22–52, 26–52 and 34–52 residues) were tested for activity. Carboxy-terminal fragments were shown to be up to 250-fold more active than the parent molecule. Minimum inhibitory concentration values of the most active fragments (13–52 and 16–52) and the parent molecule were 4.9 10 2 and 12.5 mg/ml, respectively, with E. coli. Ultrastructural analyses of AM treated cells demonstrated marked cell wall disruption with E. coli within 0.5 h. Abnormal septum formation with no apparent peripheral cell wall disruption was observed with S. aureus after 2 h. Outer membrane permeabilisation assays with E. coli confirmed that the C-terminal fragments were significantly (P < 0.05) more active. It is suggested that postsecretory processing may generate multiple AM congeners that have enhanced antimicrobial activities against a range of potential targets. # 2005 Elsevier Inc. All rights reserved. * Corresponding author. Tel.: +44 20 7882 2388; fax: +44 20 7882 2191. E-mail address: [email protected] (R.P. Allaker). available at www.sciencedirect.com journal homepage: www.elsevier.com/locate/peptides 0196-9781/$ – see front matter # 2005 Elsevier Inc. All rights reserved. doi:10.1016/j.peptides.2005.09.003
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Mechanisms of adrenomedullin antimicrobial action

Mechanisms of adrenomedullin antimicrobial action
Robert P. Allaker a,*, Paul W. Grosvenor b, David C. McAnerney a, Barry E. Sheehan a,Bakula H. Srikanta a, Keith Pell c, Supriya Kapas a
aOral Microbiology, Institute of Cell and Molecular Science, Queen Mary, University of London, Newark Street, London E1 2AT, UKbBioinformatics Group, Centre for Infectious Disease, Institute of Cell and Molecular Science, Queen Mary, University of London,
Newark Street, London E1 2AT, UKc Imaging Unit, Institute of Cell and Molecular Science, Queen Mary, University of London, Newark Street, London E1 2AT, UK
p e p t i d e s 2 7 ( 2 0 0 6 ) 6 6 1 – 6 6 6
a r t i c l e i n f o
Article history:
Received 14 July 2005
Accepted 2 September 2005
Published on line 13 October 2005
Keywords:
Adrenomedullin
Antimicrobial peptide
Structure–function
a b s t r a c t
The mechanism of antimicrobial action of the multifunctional peptide adrenomedullin (AM)
against Escherichia coli and Staphylococcus aureus was investigated. AM (52 residues) and AM
fragments (1–12, 1–21, 13–52, 16–21, 16–52, 22–52, 26–52 and 34–52 residues) were tested for
activity. Carboxy-terminal fragments were shown to be up to 250-fold more active than the
parent molecule. Minimum inhibitory concentration values of the most active fragments
(13–52 and 16–52) and the parent molecule were 4.9 � 10�2 and 12.5 mg/ml, respectively, with
E. coli. Ultrastructural analyses of AM treated cells demonstrated marked cell wall disruption
with E. coli within 0.5 h. Abnormal septum formation with no apparent peripheral cell wall
disruption was observed with S. aureus after 2 h. Outer membrane permeabilisation assays
with E. coli confirmed that the C-terminal fragments were significantly (P < 0.05) more active.
It is suggested that postsecretory processing may generate multiple AM congeners that have
enhanced antimicrobial activities against a range of potential targets.
# 2005 Elsevier Inc. All rights reserved.
avai lab le at www.sc iencedi rect .com
journal homepage: www.elsev ier .com/ locate /pept ides
1. Introduction
The epithelium represents the first barrier against infection
with pathogenic bacteria. In health, surfaces of higher
eukaryotes, such as plants, invertebrates and vertebrates,
including humans, possess a normal microflora which causes
no damage to its host. The reason, apart from physical
barriers, includes the production of gene-encoded antimicro-
bial peptides by epithelial cells [6].
Adrenomedullin (AM) is a 52 amino acid multifunctional
peptide [8] produced by a wide variety of tissues and cells. This
peptide has many properties in common with other cationic
antimicrobial peptides including human b-defensin-2. Such
molecules are defined as peptides of 12–50 amino acids with a
net positive charge of +2 to +7 [6]. Apart from human AM being
52 amino acid residues in length, AM also has a net positive
charge owing to an excess of basic amino acids (arginine and
lysine). The molecule also has a single intramolecular
* Corresponding author. Tel.: +44 20 7882 2388; fax: +44 20 7882 2191.E-mail address: [email protected] (R.P. Allaker).
0196-9781/$ – see front matter # 2005 Elsevier Inc. All rights reserveddoi:10.1016/j.peptides.2005.09.003
disulphide bond between residues 16 and 21, along with an
amidated tyrosine at the carboxyl terminus. In common with
other antimicrobial peptides including human b-defensin-2,
the amphipathic structure characterized by spatially sepa-
rated hydrophobic and charged regions, permits bacterial
membrane intercalation. Chemically, therefore AM does
resemble an antimicrobial peptide, however, its mode of
interaction with bacteria is unknown.
A variety of modes of action with respect to antimicrobial
activity have been proposed for cationic antimicrobial pep-
tides, including critical membrane depolarization, creation of
physical holes in membranes, induction of hydrolytic
enzymes that degrade the cell wall, disturbance of membrane
function and disruption of critical intracellular processes [16].
The aim of this study was to gain insights into the
mechanism of antimicrobial action of adrenomedullin against
the Gram-negative bacterium Escherichia coli and the Gram-
positive bacterium Staphylococcus aureus.
.

p e p t i d e s 2 7 ( 2 0 0 6 ) 6 6 1 – 6 6 6662
2. Materials and methods
2.1. Organisms and culture conditions
E. coli BUE55 was used as an indicator organism to assess the
structure–function relationship of AM and AM fragments. This
strain was originally isolated because of its increased
sensitivity to polymyxin B and has been used in other studies
of antimicrobial peptides [13]. E. coli (NCTC 9001), S. aureus
(Oxford antibiotic sensitive strain; NCTC 6571) and C. albicans
(ATCC 24433) were also used. Microorganisms were grown in
Tryptone Soy Broth (TSB; Lab M) at 37 8C in air with 5% CO2.
2.2. Peptides
Synthetic AM and AM fragments (residues 1–12, 1–21, 13–52,
16–21, 16–52, 22–52, 26–52 and 34–52) were obtained from
Phoenix Pharmaceuticals (Karlsruhe, Germany). Fragmenta-
tion positions in the AM molecule are shown as follows:
YRQSMNNFQGLR12S13FGC16RFGTC21T22VQKL26AHQIYQFT34
DKDKDNVAPRSKISPQGY52.
2.3. Minimum inhibitory concentration (MIC) andminimum bactericidal concentration (MBC) determinations
MIC determinations of peptides and fragments were deter-
mined in a broth micro-dilution assay against E. coli BUE55 [3].
Serial 1 in 2 dilutions of peptides were made in TSB medium.
Wells were inoculated with a 1 in 10 dilution of an overnight
bacterial culture (final concentration approximately 5 � 105
colony forming units/ml). After incubation at 37 8C for 24 h,
MICs were recorded as the lowest concentration of peptide
inhibiting growth as measured by optical density (540 nm).
MBCs of peptides were determined by inoculating samples
from MIC determination wells onto blood agar base (Oxoid
CM271) with defibrinated horse blood (5%, v/v). Plates were
then incubated at 37 8C in air with 5% CO2 for 24 h.
2.4. Transmission electron microscopy of bacteria
S. aureus and E. coli cultures were centrifuged (10,000 � g for
5 min) to pellet cells and phosphate buffered saline added to
provide a final concentration of approximately 108 colony
forming units/ml. These suspensions were then treated with
AM (final concentration of 500 mg/ml) for various lengths of
time (30–120 min) at 37 8C. Peptide concentration and expo-
sure time were based upon the characterisation of human b-
defensin-3 [7]. The bacteria were then centrifuged (10,000 � g
for 5 min), cold glutaraldehyde (0.5%, v/v) in 0.1 M sodium
cacodylate buffer (pH 7.4) was added, and samples were held
for 2 h at 4 8C. After removal of fixative, 4% (w/v) molten
agarose was added to provide a solid pellet. Cacodylate buffer
was then added and samples kept on ice. Each pellet was cut
into 1 mm cubes and post-fixed for a further 2 h in 1% osmium
tetroxide. Dehydration was carried out in ascending concen-
trations of ethanol. Samples were immersed in a mixture of
propylene oxide and TAAB resin. Samples were then finally
embedded in TAAB resin and polymerized at 60 8C for 48 h.
Sections were stained for 20 min with 2% (w/v) aqueous uranyl
acetate followed by 6 min with Reynolds lead citrate. Sections
were examined in a Jeol JEM 1200 transmission electron
microscope operating at 80 kV.
2.5. Cell-bound 1-N-phenylnaphthylamine measurements
The mode of interaction of full length AM and AM fragments
with the surface of E. coli was studied with the hydrophobic
fluorescent probe 1-N-phenylnaphthylamine (NPN) [10]. Mid-
log-phase cells were centrifuged (10,000 � g for 5 min) and
suspended in 5 mM HEPES buffer (pH 7.2) at an optical density
of 0.5 at 540 nm. Cells were then exposed to peptide/peptide
fragments (100 mg/ml) for 1 h at 37 8C. NPN (dissolved in
acetone at a concentration of 500 mM) was added at a final
concentration of 10 mM. Samples were centrifuged (10,000 � g
for 1 min) and the NPN concentration of the supernatant, i.e.
dye not bound to permeabilized cells was determined by
measuring the fluorescence in a Fluostar Optima plate reader
(BMG Lab Tech). Excitation and emission wavelengths for NPN
were set at 350 and 420 nm, respectively. Controls without
added cells or AM were included.
3. Results
The pattern of E. coli growth inhibition by the complete AM
peptide is illustrated in Fig. 1. The lowest concentration of AM
to prevent bacterial growth was found to be 12.5 mg/ml
(2.07 mmol l�1). Carboxy-terminal AM fragments demon-
strated increased activity (up to �250) when compared to
whole AM peptide against E. coli BUE55 (Fig. 2). Fragments
demonstrated varying degrees of inhibition. MIC values of
0.049, 0.049, 0.098 and 0.39 mg/ml were demonstrated with
fragments 13–52, 16–52, 22–52 and 34–52, respectively (molar
values of 0.011, 0.012, 0.029 and 0.184 mmol l�1, respectively).
No inhibition was observed with AM fragments 1–12, 1–21, 16–
21 or 26–52. Corresponding MBC values of 0.049, 0.049, 0.19 and
1.56 mg/ml were demonstrated with the active fragments.
With the Oxford strain of S. aureus MIC values of 12.5, 0.098,
0.098, 0.19 and 0.19 mg/ml were demonstrated with whole AM
and AM fragments 13–52, 16–52, 22–52 and 34–52, respectively
(molar values of 2.07, 0.022, 0.023, 0.057 and 0.09 mmol l�1,
respectively). No inhibition was observed with the other four
fragments tested. Both the whole peptide and eight fragments
were shown to be inactive against the yeast C. albicans.
Morphological changes occurring upon exposure of E. coli
and S. aureus to 500 mg/ml AM were examined by transmission
electron microscopy. Marked disruption to the integrity of the
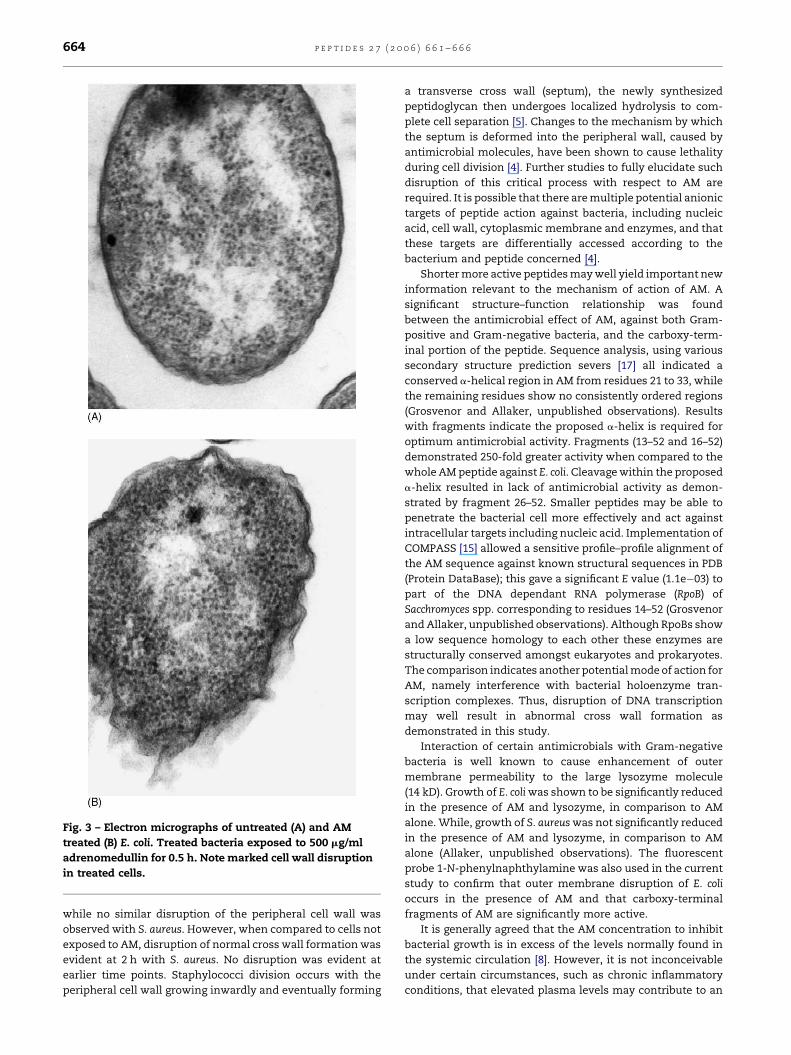
outer and cytoplasmic membranes with E. coli was observed in
50/100 of exposed bacteria examined within 0.5 h (Fig. 3). In the
corresponding control (0.5 h incubation with no peptide) only
10/100 demonstrated any evidence of membrane disruption.
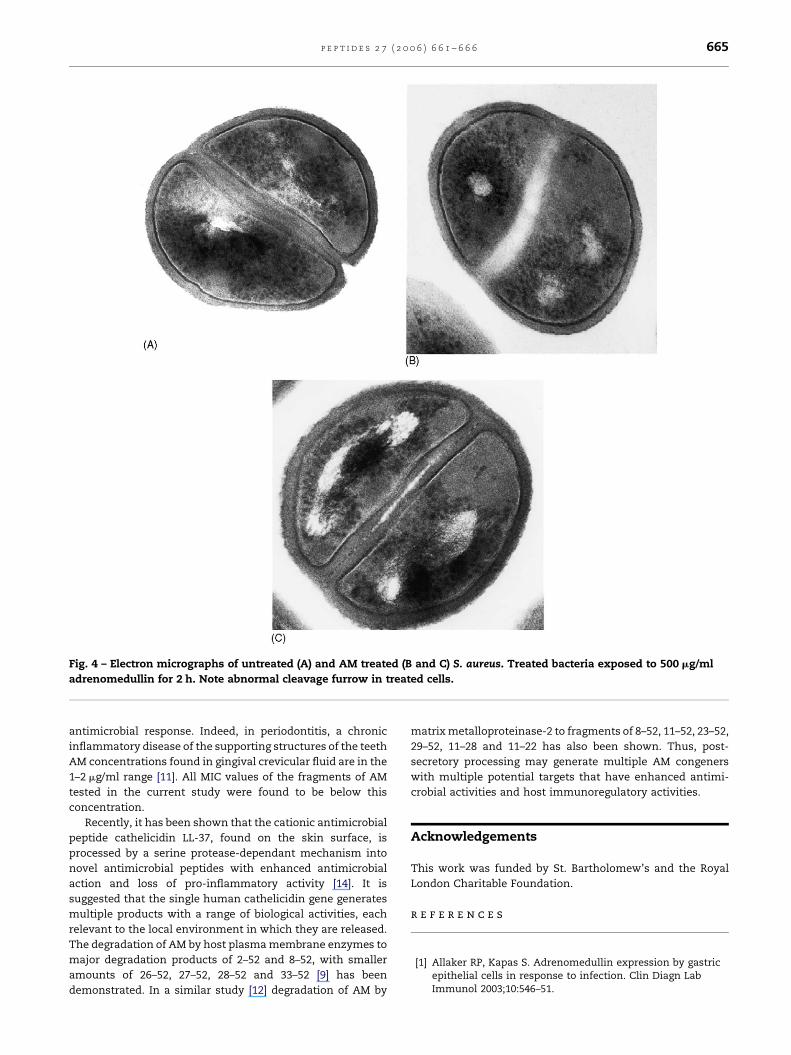
With S. aureus no apparent lysis or gross leakage of cellular
cytoplasmic contents was observed within 2 h. However,
evidence of abnormal septum (cross wall) formation was
observed in 20/100 of exposed bacteria examined (Fig. 4). In
the corresponding control (2 h incubation with no peptide)
only 2/100 demonstrated any evidence of abnormal septum
formation.
With the fluorescent probe 1-N-phenylnaphthylamine, the
mean amount (% �S.E.M.) of dye remaining in the super-

p e p t i d e s 2 7 ( 2 0 0 6 ) 6 6 1 – 6 6 6 663
Fig. 1 – Growth inhibition of E. coli BUE55, as measured by optical density in a broth micro-dilution assay, with decreasing
concentrations (mg/ml) of adrenomedullin (mean W S.E.; n = 9).
natant, as an indication of outer membrane permeabilisation,
for the parent molecule, inactive fragments (1–12, 1–21, 16–21
and 26–52) and active C-terminal fragments (13–52, 16–52, 22–
52 and 34–52) was 89.89 � 4.80, 99.65 � 6.92 and 77.71 � 4.02,
respectively. A significant difference was found between the
active C-terminal and other fragments (P < 0.05; Student’s
independent t-tests).
4. Discussion
AM has 30% homology at the nucleotide level to the cecropin
group of antimicrobial peptides, and therefore, might have a
similar mechanism of action. The mechanism of action of
cecropins is thought to involve channel formation in mem-
Fig. 2 – Growth inhibition of E. coli BUE55, as measured by optic
concentrations (mg/ml) of adrenomedullin fragments (mean W S
branes and subsequent lysis [2]. It has been shown that AM has
high affinity with E. coli lipopolysaccharide (LPS) [1], and
therefore, it might be expected that Gram-negative bacteria
would be more susceptible than Gram-positive organisms.
However, as clearly shown by the minimum inhibitory
concentrations, this was not the case. While, C. albicans was
shown to be resistant to both the whole AM molecule and
fragments. It is suggested that the binding capacity of AM may
be absent or reduced in this yeast thus making it resistant to
the antimicrobial action of AM.
To help elucidate how E. coli and S. aureus are possibly
killed, morphological changes were examined by transmis-
sion electron microscopy following exposure to AM at a
concentration above the MIC for a short period of time. Marked
changes were observed in the cell wall of E. coli within 0.5 h,
al density in a broth micro-dilution assay, with decreasing
.E.; n = 6).
p e p t i d e s 2 7 ( 2 0 0 6 ) 6 6 1 – 6 6 6664
Fig. 3 – Electron micrographs of untreated (A) and AM
treated (B) E. coli. Treated bacteria exposed to 500 mg/ml
adrenomedullin for 0.5 h. Note marked cell wall disruption
in treated cells.
while no similar disruption of the peripheral cell wall was
observed with S. aureus. However, when compared to cells not
exposed to AM, disruption of normal cross wall formation was
evident at 2 h with S. aureus. No disruption was evident at
earlier time points. Staphylococci division occurs with the
peripheral cell wall growing inwardly and eventually forming
a transverse cross wall (septum), the newly synthesized
peptidoglycan then undergoes localized hydrolysis to com-
plete cell separation [5]. Changes to the mechanism by which
the septum is deformed into the peripheral wall, caused by
antimicrobial molecules, have been shown to cause lethality
during cell division [4]. Further studies to fully elucidate such
disruption of this critical process with respect to AM are
required. It is possible that there are multiple potential anionic
targets of peptide action against bacteria, including nucleic
acid, cell wall, cytoplasmic membrane and enzymes, and that
these targets are differentially accessed according to the
bacterium and peptide concerned [4].
Shorter more active peptides may well yield important new
information relevant to the mechanism of action of AM. A
significant structure–function relationship was found
between the antimicrobial effect of AM, against both Gram-
positive and Gram-negative bacteria, and the carboxy-term-
inal portion of the peptide. Sequence analysis, using various
secondary structure prediction severs [17] all indicated a
conserved a-helical region in AM from residues 21 to 33, while
the remaining residues show no consistently ordered regions
(Grosvenor and Allaker, unpublished observations). Results
with fragments indicate the proposed a-helix is required for
optimum antimicrobial activity. Fragments (13–52 and 16–52)
demonstrated 250-fold greater activity when compared to the
whole AM peptide against E. coli. Cleavage within the proposed
a-helix resulted in lack of antimicrobial activity as demon-
strated by fragment 26–52. Smaller peptides may be able to
penetrate the bacterial cell more effectively and act against
intracellular targets including nucleic acid. Implementation of
COMPASS [15] allowed a sensitive profile–profile alignment of
the AM sequence against known structural sequences in PDB
(Protein DataBase); this gave a significant E value (1.1e�03) to
part of the DNA dependant RNA polymerase (RpoB) of
Sacchromyces spp. corresponding to residues 14–52 (Grosvenor
and Allaker, unpublished observations). Although RpoBs show
a low sequence homology to each other these enzymes are
structurally conserved amongst eukaryotes and prokaryotes.
The comparison indicates another potential mode of action for
AM, namely interference with bacterial holoenzyme tran-
scription complexes. Thus, disruption of DNA transcription
may well result in abnormal cross wall formation as
demonstrated in this study.
Interaction of certain antimicrobials with Gram-negative
bacteria is well known to cause enhancement of outer
membrane permeability to the large lysozyme molecule
(14 kD). Growth of E. coli was shown to be significantly reduced
in the presence of AM and lysozyme, in comparison to AM
alone. While, growth of S. aureus was not significantly reduced
in the presence of AM and lysozyme, in comparison to AM
alone (Allaker, unpublished observations). The fluorescent
probe 1-N-phenylnaphthylamine was also used in the current
study to confirm that outer membrane disruption of E. coli
occurs in the presence of AM and that carboxy-terminal
fragments of AM are significantly more active.
It is generally agreed that the AM concentration to inhibit
bacterial growth is in excess of the levels normally found in
the systemic circulation [8]. However, it is not inconceivable
under certain circumstances, such as chronic inflammatory
conditions, that elevated plasma levels may contribute to an
p e p t i d e s 2 7 ( 2 0 0 6 ) 6 6 1 – 6 6 6 665
Fig. 4 – Electron micrographs of untreated (A) and AM treated (B and C) S. aureus. Treated bacteria exposed to 500 mg/ml
adrenomedullin for 2 h. Note abnormal cleavage furrow in treated cells.
antimicrobial response. Indeed, in periodontitis, a chronic
inflammatory disease of the supporting structures of the teeth
AM concentrations found in gingival crevicular fluid are in the
1–2 mg/ml range [11]. All MIC values of the fragments of AM
tested in the current study were found to be below this
concentration.
Recently, it has been shown that the cationic antimicrobial
peptide cathelicidin LL-37, found on the skin surface, is
processed by a serine protease-dependant mechanism into
novel antimicrobial peptides with enhanced antimicrobial
action and loss of pro-inflammatory activity [14]. It is
suggested that the single human cathelicidin gene generates
multiple products with a range of biological activities, each
relevant to the local environment in which they are released.
The degradation of AM by host plasma membrane enzymes to
major degradation products of 2–52 and 8–52, with smaller
amounts of 26–52, 27–52, 28–52 and 33–52 [9] has been
demonstrated. In a similar study [12] degradation of AM by
matrix metalloproteinase-2 to fragments of 8–52, 11–52, 23–52,
29–52, 11–28 and 11–22 has also been shown. Thus, post-
secretory processing may generate multiple AM congeners
with multiple potential targets that have enhanced antimi-
crobial activities and host immunoregulatory activities.
Acknowledgements
This work was funded by St. Bartholomew’s and the Royal
London Charitable Foundation.
r e f e r e n c e s
[1] Allaker RP, Kapas S. Adrenomedullin expression by gastricepithelial cells in response to infection. Clin Diagn LabImmunol 2003;10:546–51.

p e p t i d e s 2 7 ( 2 0 0 6 ) 6 6 1 – 6 6 6666
[2] Christensen B, Fink J, Merrifield RB, Mauzerall D. Channel-forming properties of cecropins and related modelcompounds incorporated into planar lipid membranes.Proc Natl Acad Sci USA 1988;85:5072–6.
[3] Dykhuizen RS, Frazer R, Duncan C, Smith CC, Golden M,Benjamin N, et al. Antimicrobial effect of acidified nitrite ongut pathogens: importance of dietary nitrate inhost defence. Antimicrob Agents Chemother1996;40:1422–5.
[4] Friedrich CL, Moyles D, Beveridge TJ, Hancock REW.Antibacterial action of structurally diverse cationicpeptides on Gram-positive bacteria. Antimicrob AgentsChemother 2000;44:2086–92.
[5] Giesbrecht P, Kersten T, Maidhof H, Wecke J.Staphylococcal cell wall: morphogenesis and fatalvariations in the presence of penicillin. Microbiol Mol BiolRev 1998;62:1371–414.
[6] Hancock REW, Diamond G. The role of cationic peptides ininnate host defences. Trends Microbiol 2000;8:402–10.
[7] Harder J, Bartels J, Christophers E, Schroder JM. Isolationand characterisation of human b-defensin-3, a novelhuman inducible peptide antibiotic. J Biol Chem2001;276:5707–13.
[8] Hinson JP, Kapas S, Smith DM, Adrenomedullin, amultifactorial paracrine regulator. Endocrinol Rev2000;21:138–67.
[9] Lewis LK, Smith MW, Brennan SO, Yandle TG, Richards AM,Nicholls MG. Degradation of human adrenomedullin (1–52)by plasma membrane enzymes and identification ofmetabolites. Peptides 1997;18:733–9.
[10] Loh B, Grant C, Hancock RE. Use of the fluorescent probe 1-N-phenylnaphthylamine to study the interactions ofaminoglycoside antibiotics with the outer membrane ofPseudomonas aeruginosa. Antimicrob Agents Chemother1984;26:546–51.
[11] Lundy FT, O’Hare MMT, Briggs JE, McKibben BM, Linden GJ.Adrenomedullin in gingival crevicular fluid in periodontalhealth and disease. J Dent Res 2001;80:1176.
[12] Martinez A, Oh H-R, Unsworth EJ, Bregonzio C, Saaverda JM,Stetler-Stevenson WG, et al. Matrix metalloproteinase-2cleavage of adrenomedullin produces a vasoconstrictor outof a vasodilator. Biochem J 2004;383:413–8.
[13] Moore AJ, Beazley WD, Bibby MC, Devine DA. Antimicrobialactivity of cecropins. J Antimicrob Chemother1996;37:1077–89.
[14] Murakami M, Lopez-Garcia B, Braff M, Dorschner RA, GalloRL. Postsecretory processing generates multiplecathelicidins for enhanced topical antimicrobial defense. JImmunol 2004;172:3070–7.
[15] Sadreyev RI, Baker D, Grishin NV. Profile–profilecomparisons by COMPASS predict intricate homologiesbetween protein families. Protein Sci 2003;12:2262–72.
[16] Yeaman MR, Yount NY. Mechanisms of antimicrobialpeptide action and resistance. Pharmacol Rev2003;55:27–55.
[17] Secondary structure analysis web references:;PSIPRED, http://bioinf.cs.ucl.ac.uk/psipredJNET,http://www.compbio.dundee.ac.uk/�www-jpred/jnetSAM,http://www.cse.ucsc.edu/research/compbio/HMM-apps/T02-query.htmlJUFO, http://www.jens-meiler.de/jufo.html.




















