
Vinculin Promotes Cell Spreading by Mechanically Coupling Integrins to the Cytoskeleton
Upload
independentCategory
view
1download
0
A
cstmbucm©
K
1
hpadaa[slc
vT
0d
Available online at www.sciencedirect.com
Journal of Chromatography A, 1175 (2007) 187–196
Mechanically stable porous cellulose media for affinity purificationof family 9 cellulose-binding module-tagged fusion proteins
Mojgan Kavoosi a,b, Dexter Lam a,b, Jenny Bryan a,c,Douglas G. Kilburn a, Charles A. Haynes a,b,∗
a The Michael Smith Laboratories, The University of British Columbia, Vancouver, BC V6T 1Z4, Canadab Department of Chemical and Biological Engineering, The University of British Columbia, Vancouver, BC V6T 1Z4, Canada
c Department of Statistics, The University of British Columbia, Vancouver, BC V6T 1Z4, Canada
Received 4 May 2007; received in revised form 17 July 2007; accepted 23 July 2007Available online 7 August 2007
bstract
A mechanically stable cellulose-based chromatography media was synthesized to permit inexpensive affinity purification of recombinant proteinsontaining the family 9 carbohydrate-binding module (CBM9) fused to either the N- or C-terminus of the target protein. A second-order responseurface model was used to identify optimal concentrations of the primary reactants, epichlorohydrin and dimethyl sulfoxide (DMSO), requiredo cross-link the starting material, Perloza MT100, a compressible but inexpensive cellulose-based chromatography resin. This resulted in a
echanically stable cross-linked affinity chromatography media capable of operating at an order-of-magnitude higher linear velocity than permittedy unmodified MT100. Moments and Van Deemter analyses were used to show that rates of solute mass transfer within the column are largely
naffected by the cross-linking reaction, while the binding capacity decreased by 20% to 7.1 �mol of protein/g resin, a value superior to mostommercial affinity chromatography media. In sharp contrast to MT100, the mechanical stability and purification performance of the cross-linkededia are not diminished by scale-up or repeated column use. 2007 Published by Elsevier B.V.ked c
ebbmtAstol
eywords: Protein purification; Affinity chromatography; Fusion tag; Cross-lin
. Introduction
Recombinant DNA technology and affinity chromatographyave advanced the field of protein purification such that manyroteins and peptides can now be purified without extensivepriori knowledge of the physicochemical properties of the
esired product. By designing a chimeric protein incorporatingknown affinity tag, the protein of interest can be purified fromculture supernatant in a single, highly selective capture step
1,2]. Affinity tags have been used extensively at the laboratory
cale [3,4], yet their application at the preparative scale has beenimited, due in large part to the cost and stability (both chemi-al and mechanical) of the associated affinity matrices [5]. For∗ Corresponding author at: Michael Smith Laboratories, 2185 East Mall, Uni-ersity of British Columbia, Vancouver, BC V6T 1Z4, Canada.el.: +1 604 822 5136; fax: +1 604 822 2114.
E-mail address: [email protected] (C.A. Haynes).
sFeTwcm
h
021-9673/$ – see front matter © 2007 Published by Elsevier B.V.oi:10.1016/j.chroma.2007.07.082
ellulose; Epichlorohydrin; Preparative-scale purification
xample, immobilized metal affinity chromatography (IMAC),ased on the (His)6 affinity tag, relies on a stationary phaseearing iminodiacetate groups to which a transition metal ionay complex. The complexed metal ions act as the coordina-
ion sites for selective binding of (His)6-tagged proteins [6,7].lthough a popular and effective technology for laboratory-
cale applications, the cost associated with the synthesis ofhe stationary phase along with concerns related to leachingf the bound metal ion into the eluent make this technologyess attractive at preparative scales. Other popular affinity-tagystems such as the glutathione S transferase tag [8,9] and theLAG (N-DYKDDDDK-C) tag [10,11] likewise exhibit prop-rties that limit their scalability to preparative applications [12].he successful scale-up of tag-based affinity chromatographyould therefore benefit from an affinity tag that provides effi-
ient capture and elution on an inexpensive, yet chemically andechanically robust stationary phase.Kavoosi et al. [13] recently introduced novel vectors for
igh-level production in E. coli of soluble chimeric proteins

1 atog
cf1lccucpci[tatmrhs
[snpthscpcbdMtpcmnisSs
gisccaacupdabp
lreeosp[
oscnucrmpfitmiaeMid
2
2
lCg5cU
2
b9ltaMaees
88 M. Kavoosi et al. / J. Chrom
omprised of a target protein or peptide fused to CBM9, theamily 9 carbohydrate-binding module (CBM9) of xylanase0A of T. maritima. CBM9 strongly binds both insoluble cellu-ose and soluble polysaccharides [14], allowing for the specificapture of CBM9-tagged fusion proteins on a cellulose-basedapture column and quantitative elution of the purified proteinsing an inexpensive soluble sugar such as glucose. The CBM9-ellulose affinity cassette, and CBM tags in general [15], areotentially attractive for preparative-scale applications becauseellulose is inexpensive, chemically inert, and can be machinednto well-defined geometries offering high specific surface area16]. Cellulose has also been approved for many pharmaceu-ical and human uses and is unlikely to introduce any harmfulgents into the purified product [17,18]. Cellulose beads haveherefore been widely utilized as packing materials in chro-atography, including serving as the base chemistry for a wide
ange of size-exclusion and ion-exchange media [19]. Kuga [20]as reviewed preparation methods of cellulose beads and theireparation properties.
CBM9 binds directly to the reducing ends of cellulose chains14], permitting most commercially available cellulose-basedtationary phases to serve as the affinity media without theeed for additional chemical processing. As an example, theorous cellulosic media Perloza MT100 (henceforth referredo as MT100) binds CBM9 strongly (Ka ∼ 106 M−1), with veryigh capacity (qmax
i ∼ 500 mg protein/g dry media), and withoutignificant fouling, so that repeated purification cycles from alarified bacterial cell extract consistently result in high producturities and yields [21]. However, a limitation of highly porousellulose-based matrixes such as MT100 is the tendency fored compression at elevated linear velocities or at larger bedimensions [22,23]. Conventional cellulose beads, includingT100, are characterized by relatively large particle-size dis-
ributions, resulting in an additional flow resistance in columnsacked with these media that is associated with smaller parti-les occupying void spaces among larger ones. Pure celluloseedia are also susceptible to compressive deformation, which
ot only increases flow instabilities within the packed bed, reduc-ng dynamic capacity and column performance, but also canignificantly increase band broadening in extreme cases [24,25].cale-up of columns utilizing these media therefore requires atrategy to mechanically stabilize the packed bed.
Sephadex and other popular hydrophilic gel-like chromato-raphic media typically achieve mechanical strength throughnterchain cross-linking [26–28]. In contrast, the mechanicaltability of Perloza MT100, a highly porous, hydrophilic mediaomposed of regenerated cellulose, is derived only from inter-hain hydrogen bonds and dispersion forces between disorderednd partially crystalline regions of its regenerated celluloserchitecture (Iontosorb, CZ). This suggests that the mechani-al stability of MT100 may be improved through cross-linkingsing an appropriate epoxide or halohydrin [29]. A particularlyromising cross-linking agent for this purpose is epichlorohy-
rin, which yields hydrophilic linkages, is industrially proven,nd is readily available and inexpensive. The resulting epoxideond is stable at high pH, permitting stringent clean-in-placerotocols using caustic. Epichlorohydrin is therefore the cross-fpmw
r. A 1175 (2007) 187–196
inking agent used in the preparation of Sephadex [30] and itseaction chemistry is compatible with cellulose. Guo and Ruck-nstein [31], for example, have shown that cross-linking withpichlorohydrin can be used to increase the mechanical stabilityf cellulose-based filter paper, while Wang et al. have used aimilar strategy to improve the mechanical strength of a super-orous cellulose bead for high-speed protein chromatography32].
This paper focuses on the use of response surface method-logy (RSM) to identify optimal reaction conditions forynthesis of a mechanically stable, porous cellulose affinityhromatography media for use with CBM9 fusion-tag tech-ology. Intra-particle cross-links are introduced into MT100sing epichlorohydrin-based epoxide chemistry with the con-entrations of epichlorohydrin and dimethyl sulfoxide (DMSO)epresenting the two input variables. Treated and untreatededia are compared by monitoring both bed height and
ressure drop across the column to identify the critical super-cial velocity ucrit for column compression, which is used as
he response variable. A mechanically enhanced, cross-linkededium is identified and moments analysis, equilibrium binding
sotherms and height-equivalent-to-a-theoretical-plate (HETP)nalysis are used to compare the binding and transport prop-rties of this mechanically stabilized media against untreatedT100. The application of this cross-linked media to the affin-
ty purification of a recombinant protein from E. coli is alsoemonstrated.
. Materials and methods
.1. Reagents
Epichlorohydrin, glucose and all other reagents were ana-ytical grade and purchased from Sigma–Aldrich (Mississauga,anada) unless otherwise stated. Perloza MT100 chromato-raphic media with a nominal particle diameter distribution of0–80 �m was purchased from Iontosorb (Czech Republic). E.oli BL21 (DE3) was obtained from Novagen (Madison, WI,SA).
.2. Cross-linking of MT100
Cross-linking reactions were carried out in a 200 mL round-ottom flask immersed in a water-filled jacket. A PolyScience10 constant-temperature water pump (Preston, IN, USA) circu-ated heated water through the jacket, maintaining the reactionemperature at 50 ◦C. The reaction flask was charged with
magnetic stir bar and appropriate volumes (mL per gramT100 cellulose) of 5.0 M sodium hydroxide, epichlorohydrin
nd DMSO. The reagents were allowed to mix and thermallyquilibrate before the addition of an aliquot of MT100 slurryquivalent to 2 g of dry media. The resulting mixture was welltirred throughout the reaction, which was allowed to proceed
or 2 h before being quenched with acid addition to neutralizeH and immediate immersion in an ice bath. The cross-linkededia was then filtered and repeatedly washed with 200 mL ofater before being degassed as a dilute slurry.
omat
vaii
2
Pcioth
btsstra
2
gcsPmvwct(AoftbtG
2
hiawD3bwp
abfiHr2B1t
um5fldaTSt
3
3p
uaAbcmt
The epichlorohydrin-based cross-linking chemistry proposedto stabilize the mechanical properties of MT100 against pneu-matic compression is described in Fig. 2. Cross-linking reactionsinvolving epoxides such as epichlorohydrin are typically carried
M. Kavoosi et al. / J. Chr
The cross-linking reactions were continuously mixed to pre-ent formation of inter-particle linkages and to ensure thatll cross-linking occurred inside the resin particle. Lack ofnter-particle cross-linking was verified by optical microscopemaging of the reaction products.
.3. Hydrodynamic characterization
MT100 or cross-linked MT100 media were packed into aharmacia (now GE Healthcare; Baie d’Urfe, Canada) HR10olumn (I.D. = 1.0 cm) at an elevated flow rate using standardnclined pouring techniques. A silica filter pad was positionedn top of the bed and the flow distributor lowered and securedo achieve finger-pressure compression of the bed. The final bedeights ranged between 8.5 and 10.6 cm.
The flow curve for each column was measured by pumpinguffer (50 mM potassium phosphate, 100 mM NaCl, pH 7.0)hrough the column at specified superficial velocities and mea-uring the pressure drop at steady state. Compression of thetationary phase, measured as the separation of the bed fromhe flow distributor, was also monitored at each flow rate byecording time-lapse images of the top surface of the bed usingLogitech Quickcam Pro 4000 web camera.
.4. Measurement of binding isotherms
Equilibrium adsorption isotherms for binding of CBM9-reen fluorescent protein (CBM9-GFP) to MT100 and eachross-linked MT100 preparation were performed at 22 ◦C in lowalt buffer (50 mM potassium phosphate, 100 mM NaCl, pH 7.0).urified protein at concentrations ranging from 1 to 30 �M wasixed with media (1 mg dry weight) in low salt buffer to a final
olume of 1 mL. Samples were then incubated overnight at 22 ◦Chile mixing end-over-end. The supernatant was collected after
entrifugation at 27,000 × g for 20 min at 22 ◦C and the concen-ration of unbound protein was determined by UV absorbance280 nm) using a Cary 100 spectrophotometer (Varian, Palolto, CA, USA) and a calculated molar extinction coefficientf 62,870 M−1 cm−1 [33]. The above protocol was repeatedor binding of ovalbumin to identify any non-specific bindinghat may arise due to the cross-linking reaction. Langmuir-typeinding parameters were determined by non-linear regression ofhe Langmuir isotherm equation to the experimental data usingraphPad Prism 3.0 software.
.5. Affinity purification of CBM9-GFP
An overnight 5 mL culture of E. coli BL21 (DE3) cellsarbouring the pET28-CBM9-GFP vector [21] was used tonoculate 500 mL of lauryl broth (LB). Cells were then grownt 37 ◦C to an A600 nm of about 0.8 and protein productionas induced by the addition of 0.1 mM isopropyl-1-thio-�--galactoside (IPTG). The culture was further incubated at
0 ◦C for another 10–12 h after which the cells were harvestedy centrifugation for 20 min at 8500 × g and 4 ◦C. The cellsere resuspended in high salt buffer (50 mM potassium phos-hate, 1 M NaCl, pH 7.0) and ruptured by two passes throughFb
ogr. A 1175 (2007) 187–196 189
French pressure cell (1448 bar). The cell debris was removedy centrifugation at 27,000 × g for 30 min and 4 ◦C. The clari-ed cell extract was frontal loaded onto an XK26 column (GEealthcare) packed with MT100 or a cross-linked MT100 prepa-
ation. Unbound proteins were washed through the column using10 mL of high salt buffer followed by 150 mL of low salt buffer.ound CBM9-GFP was eluted by a step gradient of 100 mL ofM glucose in low salt buffer; 1 mL fractions were collected
hroughout the process.The chromatogram for the purification process was detected
sing in-line UV absorbance at 280 nm and off-line measure-ent of the fluorescence intensity (395 nm excitation of GFP,
09 nm emission) of each collected fraction using a Cary Eclipseuorescence spectrophotometer (Varian). Process yields wereetermined by comparing the fluorescence intensity of appropri-tely diluted elution fractions to that of the original cell lysate.he purity of the combined product peak was analyzed by 12%DS-PAGE (sodium dodecyl sulfate-polyacrylamide gel elec-
rophoresis).
. Results and discussion
.1. Response surface methodology to improve mechanicalroperties
Both flow curve and bed-compression visualization data weresed to evaluate the mechanical stability of unmodified MT100nd of each chemically modified cellulose stationary phase.s illustrated in Fig. 1, significant compression of a packeded of unmodified Perloza MT100 is observed at all superfi-ial velocities above 0.64 × 10−4 m s−1, indicating the need forechanical stabilization of this media prior to column scale-up
o permit acceptable throughput without column compression.
ig. 1. Picture of an MT100 column pre- and post-compression. Picture showsed compression at a superficial velocity of 1.06 × 10−4 m s−1.

190 M. Kavoosi et al. / J. Chromatogr. A 1175 (2007) 187–196
based
oestbssittet
iceDfcccvfbc
u
wbwvsaT0eGfi
fvrsfpmertdccooo
mtrm
u
wfiraastrt
Fig. 2. Schematic representation of the proposed epoxide-
ut under strong caustic conditions to deprotonate all solvent-xposed hydroxyl groups and increase their reactivity. Previoustudies have shown that the reaction product is relatively insensi-ive to NaOH provided the concentration of NaOH is sufficient toring the reaction pH above 12 [31]. A polar organic co-solventuch as methanol, or in this case DMSO, is generally used towell the matrix and improve accessibility of reactive hydrox-des to the cross-linking reagent, and the mechanical stability ofhe reaction product is therefore likely sensitive to the concen-ration of the co-solvent ([DMSO]) used. The concentration ofpichlorohydrin ([Epi]) is also expected to be a determinant ofhe characteristics of the reaction product.
We investigated the dependence of the mechanical stabil-ty (response variable ucrit, detailed below) of epichlorohydrinross-linked media on the MT100-normalized concentration ofpichlorohydrin (input variable ξ1 in mL Epi/g MT100) andMSO (input variable ξ2 in mL DMSO/g MT100) used in the
ormulation reaction. For each column prepared at a given ξ1, ξ2ondition, pressure drop was measured as a function of superfi-ial velocity u to determine ucrit, the value of u at which the flowurve deviates from linearity, which was used as our responseariable quantifying mechanical stability. In all cases, ucrit dif-ered by less than 15% from the linear velocity at which incipiented collapse was observed, suggesting that the onset of flowurve non-ideality is directly related to bed compression.
A general response model for this system can be written as:
crit ≡ y = f (ξ1, ξ2) + ε (1)
here ε represents random variation (e.g., measurement error orackground noise) and is assumed to have a normal distributionith mean zero and variance σ2. To identify a range of ξ1, ξ2alues for which model (1) is applicable, we first determined aet of ξ1, ξ2 pairs that yielded a well-defined reaction productnd for which a value of ucrit could be measured reproducibly.he resulting values suggested an input variable space of 0,
≤ ξ1, ξ2 ≤ 12, 12 mL/g, which is consistent with the range ofpichlorohydrin and DMSO concentrations previously used byuo and Ruckenstein [31] to optimize cross-linking of celluloselters. As the true response function f was unknown, we first per-
enmo
reaction for cross-linking cellulose with epichlorohydrin.
ormed a nonparametric exploration of the observed responsealues. To obtain a flexible and smooth representation of theesponse surface, we used locally weighted polynomial regres-ion, in which the fitted response surface for a target point resultsrom a weighted, least squares fit of a polynomial to the dataoints in a neighborhood of the target [34]. The resulting esti-ate of the response surface f, depicted in Fig. 3, showed strong
vidence for substantial curvature; specifically, a mound-shapedesponse surface was suggested by the data. This fit implies thathe mechanical stability ucrit is maximized when [epichlorohy-rin] and [DMSO] are 4.08 and 8.2 mL/g, respectively, which isonsistent with maximum observed in our experiment at inputoncentrations of ξ1 = 2.78 mL/g and ξ2 = 8.33 mL/g. The plotsf the response surface also suggest that mechanical stability atr near this maximum is only achieved in a relatively small setf DMSO and epichlorohydrin concentration pairings (Table 1).
For the purposes of conducting inference and interpretingodel parameters, it was also desirable to pose a more struc-
ured, global model. Interestingly, the standard second-orderesponse surface model can be seen as simply a global imple-entation of the polynomial fits employed above [35]:
crit = f (ξ1, ξ2)
= β0 + β1ξ1 + β2ξ2 + β11ξ21 + β22ξ
22 + β12ξ1ξ2 + ε
(2)
here β1 and β11 are the linear and quadratic regression coef-cients for the main effect of [Epi] on ucrit, and β12 is theegression coefficient for linear interaction effect between ξ1nd ξ2. The results of fitting model (2) are presented in Table 2nd Fig. 4, and the interaction effect β12 is seen to be highly non-ignificant (p = 0.8664 � 0.05), whereas all other non-intercepterms are highly significant (maximum p = 0.0134 < 0.05). Theesults of fitting a version of model (2) without the interactionerm are also presented in Table 2 and we find that both the
stimates and the strong statistical significance of the remainingon-intercept terms are essentially unchanged. The simplifiedodel therefore permits a direct and unambiguous analysisf the dependence of ucrit on the input variables ξ1 and ξ2,

M. Kavoosi et al. / J. Chromat
Table 1Critical linear velocity ucrit data for cross-linked media prepared under variousreaction conditions
Resin Epichlorohydrin(ml/g cellulose)
DMSO (ml/gcellulose)
ucrita × 104
(m s−1)
MT100 0 0 0.60/0.68CRL100-1 0.28 0 0.98/0.87CRL100-2 0.96 0 1.71/1.45CRL100-3 2.78 0 1.43/1.68CRL100-4 5.56 0 1.24/1.20CRL100-5 11.11 0 0.96/0.91CRL100-6 0 5.56 0.87/1.06CRL100-7 2.78 5.56 2.94/2.56CRL100-8 5.56 5.56 4.88/4.60CRL100-9 8.33 5.56 1.49/1.72CRL100-10 11.11 5.56 1.35/1.29CRL100-11 0 8.33 1.10/1.26CRL100-12 0.28 8.33 1.48/1.34CRL100-13b 0.96 8.33 2.85/2.71CRL100-14 2.78 8.33 6.74/6.49CRL100-15 5.56 8.33 4.66/4.38CRL100-16 8.33 8.33 3.42/3.30CRL100-17 11.11 8.33 1.39/1.51CRL100-18 0 11.11 1.67/1.53CRL100-19 0.96 11.11 1.13/1.28CRL100-20 5.56 11.11 2.24/1.86CRL100-21b 11.11 11.11 1.15/1.07
a Values for duplicate runs shown as run 1 value/run 2 value.b The first and second reactions for CRL100-13 and CRL100-21 were con-
dr
eiiii8icM
lhstcmcchb
adξ
[ttdrvRna
Cm(pspmdsdthroughput and mass-transfer related reductions in column effi-
TR
C
Rβ
β
β
β
β
β
Rβ
β
β
β
β
ucted at 1.4 and 2.8 ml/g of cellulose of 5 M NaOH, respectively. All othereactions had 1.4 ml/g of cellulose of 5 M NaOH.
ach of which was found to influence the mechanical stabil-ty. At low DMSO concentrations, the observed improvementn ucrit with increasing ξ2 is consistent with the known solvat-ng effects of DMSO. The quadratic dependence of ucrit on ξ2s then observed because DMSO concentrations greater than.33 mL/g result in significant swelling of the particles, weaken-
ng interchain hydrogen bonds between disordered and partiallyrystalline regions of the regenerated cellulose architecture ofT100 even in the presence of a large concentration of cross-cfc
able 2esults of fitting quadratic response surface models to the data set reported in Table 1
oefficient Estimate Standard error
egression equation: ucrit = β0 + β1ξ1 + β2ξ2 + β11ξ21 + β22ξ
22 + β12ξ1ξ2 + εa
0 0.548 0.425
1 0.710 0.163
2 0.432 0.140
11 −0.068 0.013
22 −0.033 0.013
12 0.002 0.010
egression equation: ucrit = β0 + β1ξ1 + β2ξ2 + β11ξ21 + β22ξ
22 + β12ξ1ξ2 + εb
0 0.512 0.363
1 0.720 0.148
2 0.436 0.135
11 −0.068 0.013
22 −0.033 0.013
a Residual standard error = 1.079 on 38 d.f.; multiple R-squared = 0.5281; adjustedb Residual standard error = 1.066 on 39 d.f.; multiple R-squared = 0.5278; adjusted
ogr. A 1175 (2007) 187–196 191
inking agent. Similarly, excessive cross-linking of the media atigh epichlorohydrin loads (i.e., >3 mL/g) results in significantwelling and mechanical breakdown of the media, presumablyhrough the high osmotic pressures and new interchain forcesreated through the cross-linking reaction. Thus, the experi-ental and RSM results suggest that chemical cross-linking
an improve the mechanical stability of MT100 only underonditions where the reaction does not disrupt the interchainydrogen-bonding forces that naturally provide mechanical sta-ility to the unmodified MT100 media.
Finally, a two-fold change in the volume of 5.0 M NaOHdded was also applied at two randomly chosen reaction con-itions (ξ1 = 0.96 mL/g, ξ2 = 8.33 mL/g; and ξ1 = 11.11 mL/g,2 = 11.11 mL/g) to verify, as reported by Ruckenstein and Guo31], that the NaOH concentration does not influence the reac-ion product provided the concentration of NaOH is sufficiento bring the reaction pH above 12. We further note that reactionuration and temperature can also affect the properties of theeaction product. The dependence of product quality on theseariables was explored in a less systematic manner by Guo anduckenstein [31] and, to maintain an experimentally reasonableumber of input variables, we chose to fix the reaction durationnd temperature at the values recommended in [31].
As quantified by ucrit, the mechanical stability ofRL100-14 (ucrit = 6.62 × 10−4 m s−1) is ca. an order ofagnitude greater than that of untreated Perloza MT100
ucrit = 0.64 × 10−4 m s−1). This is verified in Fig. 5, which com-ares the measured flow curve for a packed bed of each media tohow the significant extension of the desired linear response inressure drop to increases in flow. As a result, mechanically per-issible linear velocities within the CRL100-14 column extend
eep onto the linear region of the Van Deemter plot (as will behown later in this manuscript), allowing column operating con-itions to be selected based on the traditional trade-off between
iency. Thus, cross-linked media CRL100-14 was selected andurther characterized as a potentially scalable matrix for affinityapture and purification of CBM9-tagged fusion proteins.
t-Statistic p-Value
1.289 0.2054.353 9.79 × 10−5
3.094 0.0037−5.085 1.02 × 10−5
−2.595 0.01340.169 0.8664
1.410 0.1674.851 2.00 × 10−5
3.223 0.0026−5.152 7.75 × 10−6
−2.622 0.0124
R-squared = 0.466; F-statistic = 8.506 on 5 and 38 d.f.; p-value = 1.784 × 10−5.R-squared = 0.4793; F-statistic = 10.9 on 4 and 39 d.f.; p-value = 4.997 × 10−6.

192 M. Kavoosi et al. / J. Chromatogr. A 1175 (2007) 187–196
Fig. 3. Imageplot (A) and wireframe surface (B) for a locally weighted poly-nomial regression fit to the measured ucrit (in m s−1 × 104) data set reported inTable 1. Created with the loess, levelplot, and wireframe functions in R [48] andtat
3
motiericit
Frd
semccwpb
3
Using methods described previously [16], moments analy-sis [37,38] was combined with Van Deemter theory [39] todetermine parameters characterizing the geometric properties of
he add-on package Lattice (Lattice Graphics R package, version 0.14–16), withspan of 0.65 and a degree of 2 (i.e., local fits are quadratic). The location of
he maximum is indicated by (⊗).
.2. Comparison of sorption properties
Equilibrium adsorption studies were performed to deter-ine the influence of cross-linking on the binding properties
f the media. Adsorption isotherms for CBM9-GFP bindingo untreated MT100 and to the cross-linked media are shownn Fig. 6. Both data sets are well described by the Langmuirquation, which shows that the epichlorohydrin cross-linkingeaction does not alter binding affinity, but reduces static capac-
ty by ca. 25% (Table 3). The epichlorohydrin cross-linkinghemistry is expected to target free hydroxyls (Fig. 2), includ-ng the reducing ends of solvent-exposed cellulose chains withinhe media, leading to the observed decrease in the ligand den-Fov
ig. 4. Wireframe surface of the results obtained from the standard quadraticesponse surface model (2) when applied to the measured ucrit (in m s−1 × 104)ata set reported in Table 1.
ity for CBM9 binding. Nevertheless, the static capacity, whichquates to 370 mg CBM9-GFP bound per gram cross-linkededia, remains significantly higher than that reported for most
ommercially available media used for fusion-tag based affinityhromatography [36]. Finally, negligible binding of ovalbuminas observed (Fig. 6), indicating that cross-linking of the mediaroduced no chemical alterations that enhance non-specificinding of proteins.
.3. Comparison of column parameters and properties
ig. 5. Flow curve for solvent passage through an MT100 (filled triangles)r CRL100-14 (filled circles) column monitored over a range of superficialelocities.
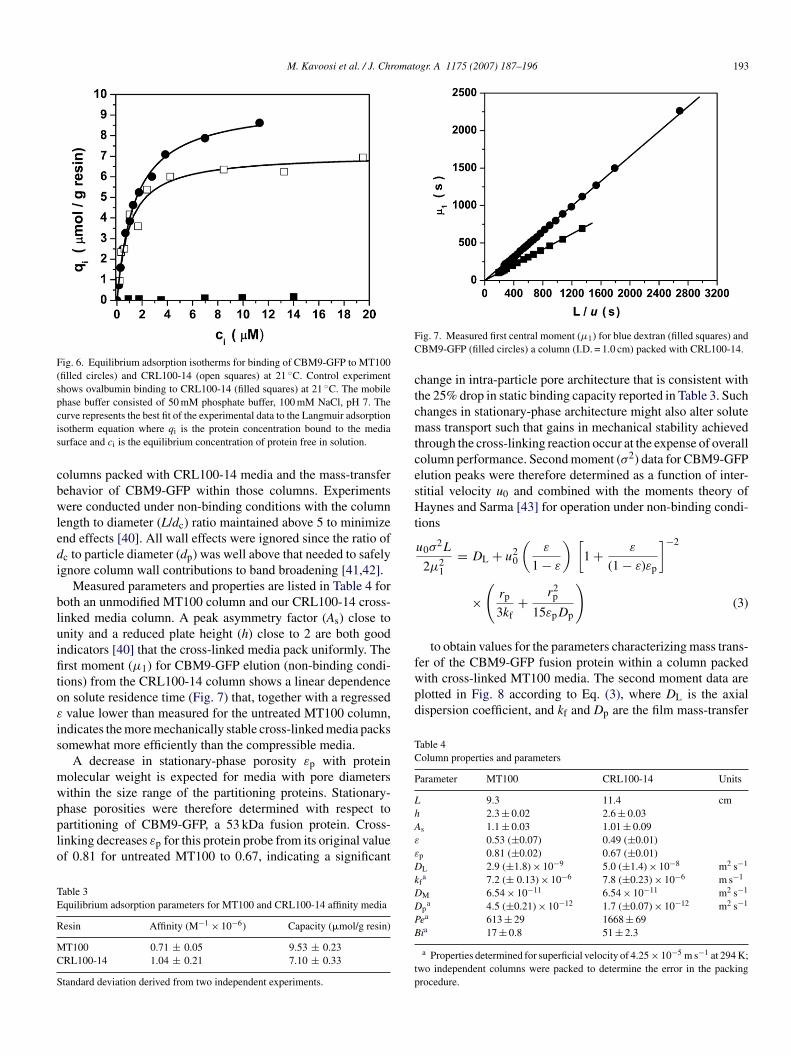
M. Kavoosi et al. / J. Chromatogr. A 1175 (2007) 187–196 193
Fig. 6. Equilibrium adsorption isotherms for binding of CBM9-GFP to MT100(filled circles) and CRL100-14 (open squares) at 21 ◦C. Control experimentshows ovalbumin binding to CRL100-14 (filled squares) at 21 ◦C. The mobilephase buffer consisted of 50 mM phosphate buffer, 100 mM NaCl, pH 7. Thecis
cbwledi
bluifitoε
is
mwpplo
TE
R
MC
S
FC
ctcmtcesHt
fer of the CBM9-GFP fusion protein within a column packedwith cross-linked MT100 media. The second moment data areplotted in Fig. 8 according to Eq. (3), where DL is the axialdispersion coefficient, and kf and Dp are the film mass-transfer
Table 4Column properties and parameters
Parameter MT100 CRL100-14 Units
L 9.3 11.4 cm
urve represents the best fit of the experimental data to the Langmuir adsorptionsotherm equation where qi is the protein concentration bound to the mediaurface and ci is the equilibrium concentration of protein free in solution.
olumns packed with CRL100-14 media and the mass-transferehavior of CBM9-GFP within those columns. Experimentsere conducted under non-binding conditions with the column
ength to diameter (L/dc) ratio maintained above 5 to minimizend effects [40]. All wall effects were ignored since the ratio ofc to particle diameter (dp) was well above that needed to safelygnore column wall contributions to band broadening [41,42].
Measured parameters and properties are listed in Table 4 foroth an unmodified MT100 column and our CRL100-14 cross-inked media column. A peak asymmetry factor (As) close tonity and a reduced plate height (h) close to 2 are both goodndicators [40] that the cross-linked media pack uniformly. Therst moment (μ1) for CBM9-GFP elution (non-binding condi-
ions) from the CRL100-14 column shows a linear dependencen solute residence time (Fig. 7) that, together with a regressedvalue lower than measured for the untreated MT100 column,
ndicates the more mechanically stable cross-linked media packsomewhat more efficiently than the compressible media.
A decrease in stationary-phase porosity εp with proteinolecular weight is expected for media with pore diametersithin the size range of the partitioning proteins. Stationary-
hase porosities were therefore determined with respect toartitioning of CBM9-GFP, a 53 kDa fusion protein. Cross-inking decreases εp for this protein probe from its original valuef 0.81 for untreated MT100 to 0.67, indicating a significantable 3quilibrium adsorption parameters for MT100 and CRL100-14 affinity media
esin Affinity (M−1 × 10−6) Capacity (�mol/g resin)
T100 0.71 ± 0.05 9.53 ± 0.23RL100-14 1.04 ± 0.21 7.10 ± 0.33
tandard deviation derived from two independent experiments.
hAε
ε
DkDDPB
tp
ig. 7. Measured first central moment (μ1) for blue dextran (filled squares) andBM9-GFP (filled circles) a column (I.D. = 1.0 cm) packed with CRL100-14.
hange in intra-particle pore architecture that is consistent withhe 25% drop in static binding capacity reported in Table 3. Suchhanges in stationary-phase architecture might also alter soluteass transport such that gains in mechanical stability achieved
hrough the cross-linking reaction occur at the expense of overallolumn performance. Second moment (σ2) data for CBM9-GFPlution peaks were therefore determined as a function of inter-titial velocity u0 and combined with the moments theory ofaynes and Sarma [43] for operation under non-binding condi-
ions
u0σ2L
2μ21
= DL + u20
(ε
1 − ε
) [1 + ε
(1 − ε)εp
]−2
×(
rp
3kf+ r2
p
15εpDp
)(3)
to obtain values for the parameters characterizing mass trans-
2.3 ± 0.02 2.6 ± 0.03
s 1.1 ± 0.03 1.01 ± 0.090.53 (±0.07) 0.49 (±0.01)
p 0.81 (±0.02) 0.67 (±0.01)
L 2.9 (±1.8) × 10−9 5.0 (±1.4) × 10−8 m2 s−1
fa 7.2 (± 0.13) × 10−6 7.8 (±0.23) × 10−6 m s−1
M 6.54 × 10−11 6.54 × 10−11 m2 s−1
pa 4.5 (±0.21) × 10−12 1.7 (±0.07) × 10−12 m2 s−1
ea 613 ± 29 1668 ± 69ia 17 ± 0.8 51 ± 2.3
a Properties determined for superficial velocity of 4.25 × 10−5 m s−1 at 294 K;wo independent columns were packed to determine the error in the packingrocedure.

194 M. Kavoosi et al. / J. Chromatogr. A 1175 (2007) 187–196
Fig. 8. Second moment analysis under non-binding conditions for a column(I.D. = 1.0 cm) packed with CRL100-14 media. Pulse injections of CBM9-G4s
cttEBttmconTrticcbm
ccocp
3o
CCsii
F1p
tttttaovbprconnecting CBM9 to GFP [47]. Two wash steps were used toeffectively remove most of the contaminating proteins presentin the column. As evident by the overlapping A280 and fluores-
Fig. 10. Chromatogram for purification of CBM9-GFP on a 60-mL CRL100-14 column at 21 ◦C. Clarified cell extract from E. coli BL21 were loadedonto a CRL100-14 column (2.6 cm I.D. × 11.8 cm) at a superficial velocity of
−4 −1
FP were used over the superficial velocity range 4.25 × 10−5 m s−1 to.25 × 10−4 m s−1. The mobile phase consisted of 2 M glucose in 50 mM potas-ium phosphate, 100 mM NaCl (pH 7, 21 ◦C).
oefficient and pore diffusivity, respectively. Standard correla-ions for the bulk molecular diffusivity DM [44] and kf [45]herefore permit estimation of DL and Dp through regression ofq. (3). The subsequent calculation of the column Peclet (Pe) andiot (Bi) numbers then provides a sound basis for determining if
he cross-linking reaction has negatively impacted solute massransport within the column. In a previous study [46], we deter-ined that uptake of CBM9-tagged proteins within a MT100
olumn is limited by the rate of solute diffusion within the poresf the stationary phase, with axial dispersion providing no sig-ificant contribution to elution band broadening. As shown inable 4, cross-linking of the MT100 media changes this resultelatively little. Axial dispersion remains insignificant, and massransfer across the fluid film is enhanced by the small reductionn column voidage. Dp decreases by ca. 60% compared to theolumn utilizing untreated MT100, so that solute mass transferontinues to be limited by the rate of pore diffusion as indicatedy the small increase in both Pe and Bi for the cross-linkededia.Finally, as noted previously, the Van Deemter plot (Fig. 9)
haracterizing CBM9-GFP transport within the CRL100-14olumn confirms that this cross-linked media can be reliablyperated far into the flow regime where reduced plate height h isontrolled by solute mass transfer kinetics within the stationaryhase.
.4. Preparative-scale affinity purification of CBM9-GFPn CR1
Fig. 10 shows the chromatogram for the purification ofBM9-GFP from clarified E. coli BL21 cell lysate on a 60 mL
RL100-14 column, which represents an eightfold columncale-up by volume. Pressure drops for a given linear veloc-ty in the scaled-up column were statistically identical to thosen the original 8 mL column. No protease inhibitors were added
1ccfa
ig. 9. Van Deemter plot describing CBM9-GFP transport within the CRL100-4 column (I.D. = 1.0 cm × 11.4 cm); the reduced plate height h(= σ2/(Ldp)) islotted as a function of the reduced linear velocity, given by u/(dpDL).
o the cell extract and the purification was performed at roomemperature (21 ◦C). Although u in this separation is ca. threeimes larger than ucrit for untreated Perloza MT100, the purifica-ion on the CRL100-14 column proceeded in a similar mannero that on the MT100 column [21] when both chromatogramsre reported in terms of elution volume. Initial breakthroughf contaminating proteins was observed after ca. one columnoid volume. The small fluorescence peak observed within thereakthrough profile for contaminating proteins indicates theresence in the clarified cell lysate of free GFP, released as aesult of a small amount of degradation within the linker region
.5 × 10 m s , washed with ca. 3 column volumes (CVs) of high salt buffer,a. 2.5 CVs of low salt buffer and bound CBM9-GFP was eluted with 1 M glu-ose in low salt buffer, all at a superficial velocity of 1.5 × 10−4 m s−1. 10 mLractions were collected and analyzed by both absorbance at 280 nm (solid line)nd fluorescence emission at 509 nm (filled circles).

M. Kavoosi et al. / J. Chromat
Table 5Yield and purity for consecutive purification cycles of CBM9-GFP on a CRL100-14 column
Column cycle Yield (%) Puritya (%)
1 74.8 >952 88.7 >953
cC
8Cyyaoc
(niiebrtrus
3
aupoft
Ftdmfli
ocflitoocCp
4
mrmfptf16mrsbosilsd
A
83.5 >95
a Determined by SDS-PAGE analysis.
ent signals, addition of 1 M glucose to the mobile phase elutesBM9-GFP as a single sharp peak.
The product yield obtained in the pooled elution peak was2 ± 7% (Table 5). Most of the lost product can be attributed toBM9-GFP degradation products in the feed. Nevertheless, theield from the cross-linked column is slightly lower than the 86%ield previously recorded for the same separation performed onn untreated MT100 column at a 3.6-fold lower linear velocityf 0.41 × 10−4 m s−1, indicating a small reduction in dynamicapacity with increased u.
SDS-PAGE gel documentation of the scaled-up purificationFig. 11) shows quantitative flow-through (Lane 2) of contami-ating proteins with minimal presence of CBM9-GFP, resultingn elution of the 53 kDa CBM9-GFP product (Lane 5) character-zed by a purification factor greater than 95%. The weak 26 kDalution band in Lane 5 corresponds to CBM9 affinity tag releasedy proteolytic degradation of the fusion protein within the linkeregion. As the purification was performed at 21 ◦C with no pro-ease inhibitors, activity from endogeneous proteases was likelyesponsible for the observed product degradation. Higher prod-ct yields may therefore be realized through application of auitable protease inhibitor.
.5. Column reusability
The ability of a CRL100-14 column to provide acceptablend predictable purification performance with repeated columnse was also investigated. Three consecutive purifications were
erformed on the scaled-up column at a superficial velocityf 1.5 × 10−4 m s−1 to identify any changes in column per-ormance with increasing number of purification cycles. In allhree purification cycles, very high product purity (>95%) wasig. 11. 12% SDS-PAGE documentation for preparative-scale affinity purifica-ion of CBM9-GFP on a CRL100-14 column (I.D. = 2.6 cm). All samples wereissolved in sample buffer containing 10% SDS. Lanes: M = molecular weightarkers in kg/mole; 1 = clarified cell extract prior to column loading; 2 = columnow through; 3 = high salt wash; 4 = low salt wash; 5 = pure CBM9-GFP eluted
n low salt buffer containing 1 M glucose.
feENBS
R
[
ogr. A 1175 (2007) 187–196 195
bserved. Interestingly, product yield increased after the firstolumn use (Table 5) to a value comparable to that observedor the untreated MT100 column. This suggests that the cross-inking reaction introduces a small number of sites that exhibitrreversible binding to CBM9-GFP. These sites are blocked inhe first cycle, leading to the higher observed yields in the sec-nd and subsequent cycles. While the large quantity (ca. 15 g)f fusion protein required to load the scaled-up column pre-luded additional cycle studies, multiple runs on the small-scaleRL100-14 column showed no significant change in columnerformance.
. Conclusions
The mechanical stability of porous Perloza MT100 cellulosicatrix was enhanced through an RSM optimized cross-linking
eaction using epichlorohydrin to create a chromatographicedia for affinity purification of recombinant CBM9-tagged
usion proteins. The chemistry used to cross-link MT100 is sim-le to conduct and inexpensive, yielding a stable affinity mediahat is ca. 1/65 the cost of popular affinity media currently usedor industrial processing. The mechanically enhanced CRL100-4 media can withstand mobile-phase superficial velocities up to.6 × 10−4 m s−1, representing an order of magnitude improve-ent in throughput over unmodified MT100. The cross-linking
eaction alters pore architecture, reducing the porosity of thetationary phase by 20% and thereby lowering binding capacityy ca. 25%. Despite this loss in capacity, CRL100-14 is capablef binding 7.1 �mol CBM9-GFP/g media (376 mg/g), a valueuperior to most commercial affinity-tag systems. The mechan-cal stability of the cross-linked media is maintained during (ateast) modest column scale-up. Consecutive purifications on acaled-up column showed yields equal to or greater than 82%,emonstrating reliable column performance from cycle to cycle.
cknowledgements
The authors would like to thank Professor Tony Warrenor helpful discussions and use of his laboratory space andquipment. This work was supported by grants from Proteinngineering Network of Centres of Excellence (PENCE) andSERC. C.A.H. is the Canada Research Chair in Interfacialiotechnology and J.B. is a principal scholar of the Michaelmith Foundation for Health Research.
eferences
[1] C.F. Ford, I. Suominen, C.E. Glatz, Protein Expr. Purif. 2 (1991) 95.[2] C.R. Lowe, A.R. Lowe, G. Gupta, J. Biochem. Biophys. Methods 49 (2001)
561.[3] J. Nilsson, S. Stahl, J. Lundeberg, M. Uhlen, P.-A. Nygren, Protein Expr.
Purif. 11 (1997) 1.[4] M.T. Hearn, D. Acosta, J. Mol. Recognit. 14 (2001) 323.[5] K. Terpe, Appl. Microbiol. Biotechnol. 60 (2003) 523.
[6] L. Rulisek, Z. Havlas, J. Am. Chem. Soc. 122 (2000) 10428.[7] J. Porath, J. Carlsson, I. Olsson, G. Belfrage, Nature 258 (1975) 598.[8] K.L. Guan, J.E. Dixon, Anal. Biochem. 192 (1991) 262.[9] D.B. Smith, K.S. Johnson, Gene 67 (1988) 31.10] A. Einhauer, A. Jungbauer, J. Biochem. Biophys. Methods 49 (2001) 455.
1 atog
[
[[
[
[
[
[
[
[[[
[
[
[
[
[[
[[[
[[[[[
[
[[[
[[[[[[[
96 M. Kavoosi et al. / J. Chrom
11] T. Hopp, K.S. Pricket, V.L. Price, R.T. Libby, C.J. March, D.P. Ceretti, D.L.Urdal, P.J. Conlon, Bio/Technology 6 (1988) 1204.
12] M. Linhult, S. Guelich, S. Hober, Protein Peptide Lett. 12 (2005) 305.13] M. Kavoosi, J. Meijer, E. Kwan, A.L. Creagh, D.G. Kilburn, C.A. Haynes,
J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 807 (2004) 87.14] A.B. Boraston, A.L. Creagh, M.M. Alam, J.M. Kormos, P. Tomme, C.A.
Haynes, R.A. Warren, D.G. Kilburn, Biochemistry 40 (2001) 6240.15] Y. Cao, Q. Zhang, C. Wang, Y. Zhu, G. Bai, J. Chromatogr. A 1149 (2007)
228.16] B. Rodriguez, M. Kavoosi, J. Koska, A.L. Creagh, D.G. Kilburn, C.A.
Haynes, Biotechnol. Prog. 20 (2004) 1479.17] N.A. Hoenich, C. Woffindin, P.J. Cox, M. Goldfinch, S.J. Roberts, Clin.
Nephrol. 48 (1997) 253.18] M.P. Varela, P.L. Kimmel, T.M. Phillips, G.J. Mishkin, S.Q. Lew, J.P.
Bosch, Blood Purif. 19 (2001) 370.19] Y. Motozato, C. Hirayama, J. Chromatogr. 298 (1984) 499.20] S. Kuga, Kami Pa Gikyoshi 38 (1984) 166.21] M. Kavoosi, J. Meijer, E. Kwan, A.L. Creagh, D.G. Kilburn, C.A. Haynes,
J. Chromatogr. B 807 (2004) 87.22] C.B. Colby, B.K. ONeill, F. Vaughan, A.P.J. Middelberg, Biotechnol. Prog.
12 (1996) 662.23] K.C.E. Ostergren, A.C. Tragardh, G.G. Enstad, J. Mosby, AIChE J. 44
(1998) 2.24] C.B. Colby, B.K. ONeill, A.P.J. Middelberg, Biotechnol. Prog. 12 (1996)
728.25] A.W. Mohammad, D.G. Stevenson, P.C. Wankat, Ind. Eng. Chem. Res. 31
(1992) 549.26] M. Leonard, J. Chromatogr. B 699 (1997) 3.27] S. Hjerten, B. Wu, J. Liao, J. Chromatogr. 396 (1987) 101.
[
[
r. A 1175 (2007) 187–196
28] T. Laas, Protides Biol. Fluids 23 (1976) 495.29] S.R. Narayanan, L.J. Crane, Trends Biotechnol. 8 (1990) 12.30] L. Holmberg, B. Lindberg, B. Lindqvist, Carbohydr. Res. 272 (1995)
203.31] W. Guo, E. Ruckenstein, J. Membr. Sci. 182 (2001) 227.32] D.-M. Wang, G. Hao, Q.-H. Shi, Y. Sun, J. Chromatogr. A 1146 (2007) 32.33] H. Mach, C.R. Middaugh, R.V. Lewis, Anal. Biochem. 200 (1992) 74.34] W.S. Cleveland, S.J. Devlin, J. Am. Stat. Assoc. 83 (1988) 596.35] G.E.P. Box, J.S. Hunter, W.G. Hunter, Statistics for Experimenters: Design,
Innovation and Discovery, 2nd ed., Wiley, New Jersey, 2005.36] J.J. Lichty, J.L. Malecki, H.D. Agnew, D.J. Michelson-Horowitz, S. Tan,
Protein Expr. Purif. 41 (2005) 98.37] M. Kubin, Collect. Czech. Chem. Commun. 30 (1965) 2900.38] E. Kucera, J. Chromatogr. 19 (1965) 237.39] J.J. Van Deemter, F.J. Zuiderweg, A. Klinkenberg, Chem. Eng. Sci. 5 (1956)
271.40] A. Jungbauer, J. Chromatogr. 639 (1993) 3.41] J.H. Knox, G.R. Laird, P.A. Raven, J. Chromatogr. 122 (1976) 129.42] P.A. Bristow, J.H. Knox, Chromatographia 10 (1977) 279.43] H.W. Haynes, P.N. Sarma, AIChE J. 19 (1973) 1043.44] M.E. Young, P.A. Carroad, R.L. Bell, Biotechnol. Bioeng. 22 (1980) 947.45] E.J. Wilson, C.J. Geankoplis, Ind. Eng. Chem. Fundam. 5 (1966) 9.46] M. Kavoosi, N. Sanaie, F. Dismer, J. Hubbuch, D.G. Kilburn, C.A. Haynes,
J. Chromatogr. A 1160 (2007) 137.
47] M. Kavoosi, A.L. Creagh, D.G. Kilburn, C.A. Haynes, Biotechnol. Bioeng.98 (2007) 599.48] R Development Core Team, R: A language and environment for statistical
computing. R Foundation for Statistical Computing, Vienna, 2006 (URL:http://www.R-project.org).
Copyright © 2022 FDOKUMEN




















