
Active Kallikrein Response to Changes in Sodium-Chloride Intake in Essential Hypertensive Patients1
Upload
independentCategory
view
1download
0
356
Journal of Andrology. Vol. 20. No. 3. May/June 1999
Copyright © American Society of Andrology
The Effects of Aging on the Seminiferous Epithelium and theBlood-Testis Barrier of the Brown Norway Rat
SHIRLEY LEVY,* VALERIE SERRE,* LOUIS HERMO,j- AND BERNARD ROBAIRE*
From the *Department of Pharmacology and Therapeutics and the tDeparrmenr of Anatomy and Cell Biology,
McGill University.
ABSTRACT: Steroidogenesis and spermatogenesis decrease in ag-ing Brown Norway rats. We therefore hypothesized that there must
be accompanying morphological changes taking place in the seminif-erous tubules of the aging testis. The testes of Brown Norway ratsranging in age from 3 to 24 months were prepared for lightand elec-
tron microscopy. To assess the integrity of the blood-testis barrier withage, a lanthanum nitrate study was done. The normal seminiferous
tubules present in rats at 3 and 12 months of age were largely re-placed at 24 months by fully regressed tubules that were virtually de-void of germ cells and contained large intercellular spaces. An elec-
tron-microscopic study of these regressed tubules showed a completeloss of cyclical variations of the organelles of the Sertolicells.The
nucleus was more irregularly shaped and was present at various lev-els in the epithelium. The endoplasmic reticulum was a loose, vesic-ulated network that was unlike the elaborate, tubular, anastomotic net-work noted in young animals. The lysosomes were large, oddly-shaped, and contained lipidic inclusions, in contrast to the distinct
membrane-bound lysosomes and dense core bodies found in the
young animals. Adjacent Sertoli cell processes encompassed large,empty intercellular spaces, possibly occupied previously by germ
cells. The typical Sertolr-Sertoli junctions of the blood-testis barrier inthe young animal were rarely seen at 24 months and were replaced
by focal contact points, usually between three Sertolicell processes.In the aged animals, lanthanum nitrate permeated the basal and ad-luminal compartments, extending between Sertoli cellprocesses and
entering the intercellularspaces and lumen. Insummary, during aging,
there isa breakdown of the blood-testis barrier, and there are striking
changes in the appearance of Sertolicells.These results suggest a
possible intrinsiclimitationthat prevents stem cells from renewing
themselves, whether because of a degeneration of immunological or-
igin or because of a lack of Sertoli cell support.Key words: Sertoli cell, germ cells, spermatogenesis, tight junc-
tions, lanthanum nitrate, testicular morphology.J Androl 1999;20:356-365
T he functions and histology of the testis during devel-
opment and in the adult are well described (Leblond
and Clermont, 1952; Br#{246}kelmann, 1963; Elftman, 1963;
Burgos et al, 1970; Wartenberg, 1989), but the effects of
aging on the testis have not been as intensively studied.
Light-microscopic analyses of the aging human and rat
testis have revealed partial and/or total regression of the
seminiferous tubules (Paniagua et al, 1991; Wright et al,
1993; Gruenewald et al, 1994). In the few studies on mor-
phological changes of the aging human testis, partially
regressed tubules, with spermatogenesis arrested at the
level of spermatocytes or spermatogonia, contained Ser-
toli cells with a variable appearance. These Sertoli cells
could be multinucleated, with normal or degenerating or-
ganelles; dedifferentiated, with the appearance of prepu-
bertal Sertoli cells; or filled with numerous polymorphic
Supported by a program grant from NIH AG0832 1, by the Fonds pour
Ia Formation de Chercheurs et l’Aide a Ia Recherche (FCAR), Quebec,
and the Medical Research Council of Canada (to L.H.).
Correspondence to: Dr. Bernard Robaire, Department of Pharmacology
and Therapeutics, McGill University,3655 Drummond Street,Montreal.
Quebec, H3G lY6, Canada. E-mail: [email protected]
Received for publication September 18, 1998; accepted for publication
October 19, 1998.
mitochondria (Schulze and Schulze, 1981; Paniagua et al,
1985).
The abnormalities and loss of Sertoli cells seen in the
aging human testis may be due to a disruption of the
blood-testis barrier (Johnson, 1986; Paniagua et al, 1987).
The incidence of antibodies against spermatozoa in serum
has been reported to increase with advancing age in men
(Fjalbrant, 1975), and antigens may arise from degener-
ating tubules because of an altered blood-testis barrier
(Johnson et al, 1984). Tracer studies in normal adult rats
(Dym and Fawcett, 1970; Neaves, 1973) have shown that
lanthanum nitrate can penetrate the myoid cell layer and
surround germ cells in the basal compartment. However,
lanthanum is stopped at the Sertoli-Sertoli tight junctions
and cannot enter the adluminal compartment. To date, no
tracer studies have been conducted to determine whether
there were changes in the blood-testis barrier with age.
The Brown Norway rat is a remarkable aging model
because it does not exhibit many of the age-related pa-
thologies seen in other rat strains, nor does it become
obese during its long life span (Zirkin et al, 1993). There
are striking changes intrinsic to the aging Brown Norway
rat testis, such as a decrease in the ability of Leydig cells
to produce testosterone (Zirkin et al, 1993; Chen et al,
1994) and a reduction in total sperm count (Wang et al,

Levy et al . Seminiferous Epithelium of the Aging Rat 357
1993). The large variability observed in human studies,
attributable to individual stress, diet, or underlying dis-
ease (Vermeulen, 1988) is eliminated by using this animal
model. Significantly, aging of the reproductive system in
man is also associated with a reduction in steroidogenesis
and spermatogenesis (Neaves et al, 1984; Vermeulen,
1991), making the Brown Norway rat ideal for this study.
Steroid and sperm production are two of the most cru-
cial roles of the testis. Therefore, we undertook an in-
depth light- and electron-microscopic analysis of the mor-
phological changes in the seminiferous epithelium that
occurred as these functions decreased in aging Brown
Norway rats. In addition, to assess the age-related chang-
es in the blood-testis barrier, a lanthanum nitrate tracer
study was done. We report striking ultrastructural changes
with age in Sertoli cells that correlate with a disappear-
ance of germ cells and with a disruption of the functional
integrity of the blood-testis barrier.
Materials and Methods
Animals
Brown Norway rats aged 3, 12. 18. and 24 months were pur-chased from the National Institute on Aging (Bethesda, Mary-land) and were supplied by Charles River Breeding Laboratories(Wilmington, Massachusetts). Animals used in the morphologi-
cal study were housed at the Johns Hopkins School of Hygiene
and Public Health in a temperature- (22#{176}C)and light- (14L: 1OD)controlled room where food and water was available ad libitum.
Animals used in the lanthanum tracer study were housed at the
McGill University McIntyre Animal Centre under the same con-ditions.
Tissue Preparation for Light and Electron Microscopy
Brown Norway rats aged 3 (n = 4), 12 (n = 4), 18 (n = 3), and
24 (n = 5) months were anesthetized with an IP injection of
sodium pentobarbital (Somnotol; Steris Laboratories Inc., Phoe-
nix, Arizona). The testes were fixed by a retrograde perfusion
through the abdominal aorta with 5% glutaraldehyde buffered in
sodium cacodylate (0.1 M) containing 0.05% CaC12, pH 7.4. The
testes were removed, cut into 1-mm3 pieces, and placed in the
same fixative for 2 hours at 4#{176}C.The tissues were washed three
times in sodium cacodylate buffer (0.1 M) containing 4% su-
crose, pH 7.4. They were then postfixed in 1% osmium tetroxide
and 1.5% potassium ferrocyanide and were embedded in epoxyresin. Semi-thin (l-m) and thin (75-nm) sections were cut with
an ultramicrotome. The semi-thin sections were stained with to-luidine blue and were examined with the light microscope (LeitzWetzlar, Laborlux D, Montreal, Quebec, Canada). The thin sec-
tions were counterstained with uranyl acetate and lead citrateand examined with a Philips 400 electron microscope (Philips,Eindhoven, The Netherlands).
Lanthanum Tracer Study
Brown Norway rats aged 3 (n = 6) and 24 months (n = 6) wereanesthetized with an IM injection of a cocktail of ketamine hy-
drochloride (Ketalean; MTC Pharmaceuticals, Cambridge, On-
tario, Canada), xylazine (Rompun; Bayer Inc., Etobicoke, On-tario, Canada), acepromazine maleate (Atravet; Ayerst Labora-tories, Montreal, Quebec, Canada), and 0.9% sodium chloride
(Baxter Corporation, Toronto, Ontario, Canada). This cocktail isused because it decreases the risk of respiratory depression that
can occur with sodium pentobarbital, especially in aged animals.A retrograde perfusion through the abdominal aorta was used to
fix the testes. The fixative consisted of 5% glutaraldehyde buff-
ered with 0.16 M collidine buffer (pH 7.4) containing a final
concentration of 2% lanthanum nitrate (Marivac, Nova Scotia,Canada) and 2.5% polyvinylpyrrolidone and a final pH of 7.3.
The solution was filtered through a Millipore filter using a 5-micron membrane (MicronSep Membrane Filters, Westborough,Massachusetts) prior to use. After perfusion, the testes were re-moved, cut into 1-mm3 pieces. and left for 2 hours at 4#{176}C,eitherin the same fixative or in the same fixative without lanthanum.The tissues were washed quickly three times in a 0.16 M colli-
dine buffer with or without 2% lanthanum nitrate and 2% su-crose, pH 7.4, and were subsequently washed three times in 0.1
M sodium cacodylate buffer with or without 2% lanthanum ni-trate and 2% sucrose, pH 7.4 (this last wash was done simplyto remove any toxic collidine which remained). Lanthanum was
omitted from the immersion solutions in some cases to confirm
that the observed path of the tracer was due exclusively to ex-travasated lanthanum. Tissues were postfixed in 1% osmium te-troxide containing the same cacodylate buffer mixture used forwashing, stained en bloc with uranyl acetate, and embedded in
epoxy resin. Thin sections were cut with an ultramicrotome;
some sections were left unstained, others were stained with ura-
nyl acetate and lead citrate, and all were examined with a Philips410 electron microscope.
Results
Light Microscopy
The seminiferous epithelium of the Brown Norway rats
of 3 and 12 months of age exhibited active spermatogen-
esis; all steps of germ cell development were present (Fig.
lA). Sertoli cell nuclei were located at the base of the
epithelium, and the germ cell arrangement was morpho-
logically normal. In contrast, the testes of 18-month-old
rats contained a mixture of normal, partially regressed,
and fully regressed seminiferous tubules (Fig. lB). In the
partially regressed tubules, some germ cells sloughed off
into the lumen, and many of the spermatids that remained
were detached from the epithelium. The fully regressed
tubules were smaller in size, with a thickened basement
membrane and a disrupted epithelium. There was a de-
crease in germ cells present in the regressed tubules, with
a concurrent appearance of spaces within the epithelium
that were identified with the electron microscope as in-
tercellular spaces.
By 24 months, the majority of tubules were fully re-
gressed (Fig. lC). At times, normal tubules undergoing
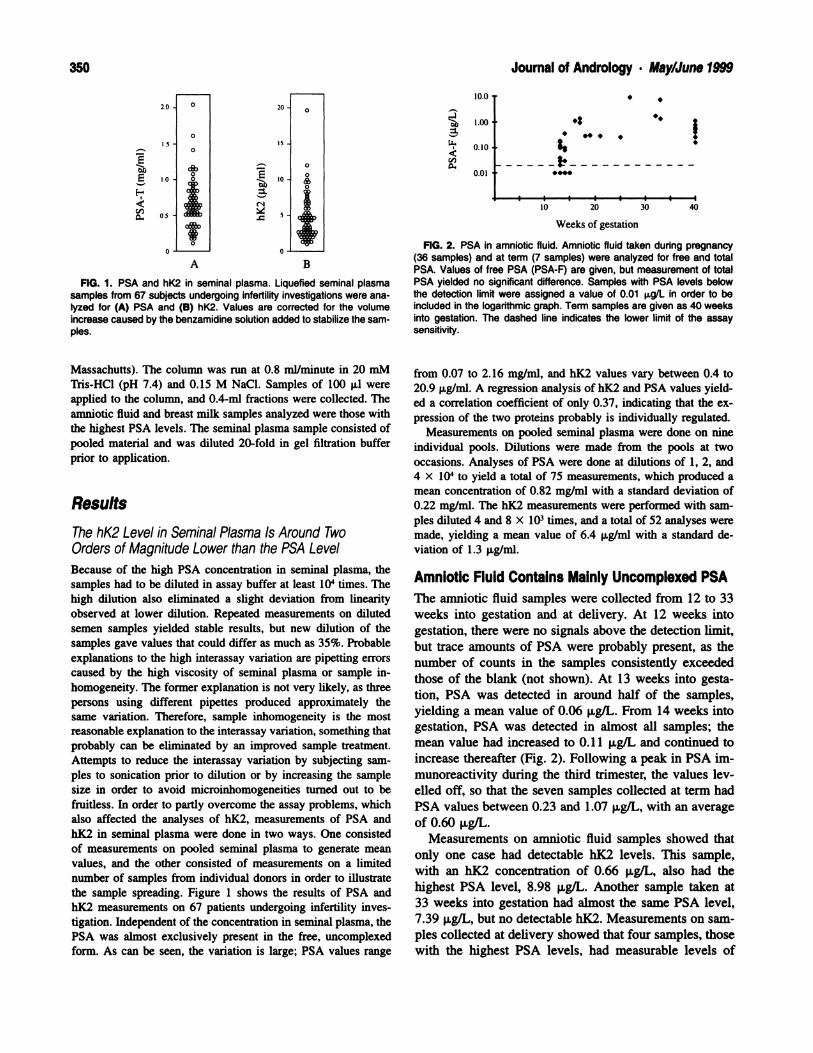
358 Journal of Andrology . May/June 1999
FIG. 1. Light micrographs showing the effects of age on the mor-phology of the seminiferous epithelium of the Brown Norway rat. (A)Normal seminiferous tubules in a 3-month-old rat. (B) Regressed tubulesare seen adjacent to normal tubules in a testis of an 18-month-old rat,whereas in (C), a majority of regressed tubules are present in a 24-month-old rat. Note Sertoli cells and large, empty intercellular spaceswithin the epithelium of the regressed tubules in (B) and (C). Lu, lumen;S, Sertoli cell; IV, VII, VIII, IX, XI, stages; asterisks, intercellular spaces.Scale bar: A = 35 p.m; B = 44 m; C = 31 pm.
complete spermatogenesis appeared directly adjacent to
regressed tubules. The regressed tubules contained few
germ cells, and there were many more intercellular spaces
than at 18 months (Fig. 1 C). These intercellular spaces
were seen in all regressed tubules, and their location and
size varied within the epithelium; larger spaces were pre-
sent closer to the lumen. As the majority of tubules were
small and were devoid of germ cells, it was impossible
to assign a stage to them. Sertoli cells made up the ma-
jority of cells in the fully regressed seminiferous tubules.
The Sertoli cell nuclei were present at all levels of the
epithelium, including near the lumen (Fig. 1C).
Electron Microscopy
Because of the insufficient amount of regressed tubules
in our sample of 18-month-old animals, the electron-mi-
croscopic study consisted of a comparison of seminifer-
ous tubules of 3-month-old and 12-month-old rats, which
were identical, and of regressed tubules of 24-month-old
rats. In the seminiferous epithelium of Brown Norway
rats of 3 months of age, at stage VII, Sertoli cells and
preleptotene spermatocytes resided on the basement
membrane (Fig. 2A). The endoplasmic reticulum was a
highly anastomotic, tubular network, whereas the Golgi
apparatus showed several large stacks of saccules. Clus-
ters of small, membrane-bound lysosomes were found at
the base of the Sertoli cell. The elaborate blood-testis
barrier, composed of adjacent plasma membranes of Ser-
toli cells with tight and gap junctions, subsurface endo-
plasmic reticulum, and filaments, separated the basal from
the adluminal compartment (Fig. 2A).
At 24 months, most of the tubules were regressed;
some were virtually devoid of germ cells, while in others,
several could be found. These germ cells consisted of a
darker, more granular cytoplasm than was seen in the 3-
month-old animals, and they were separated from the
thickened basement membrane (Fig. 2B). In marked con-
trast to what was seen in the 3-month-old animals, Sertoli
cells predominated in the epithelium and showed more
lobulated, irregularly shaped nuclei, which were now
closer to the lumen (Fig. 2B). In all regressed tubules,
Sertoli cells did not show any cyclical variation with re-
gard to their organelles. The endoplasmic reticulum was
represented as an irregular, loose, vesiculated network
(Fig. 2B), unlike the tubular network seen at certain stag-
es of spermatogenesis in the young animals. The Golgi
apparatus was formed of several small stacks of saccules,
unlike the large, developed stacks present in the young
animals (Fig. 2A). The lysosomes in the tubules of 24-
month-old rats were large, irregular in shape, and few in
number (Fig. 2B). These lysosomes showed a heteroge-
neous composition, representing lipofuscin-like inclusions
(Fig. 2B), in contrast to the numerous, small lysosomes
observed at specific stages at 3 months (Fig. 2A).
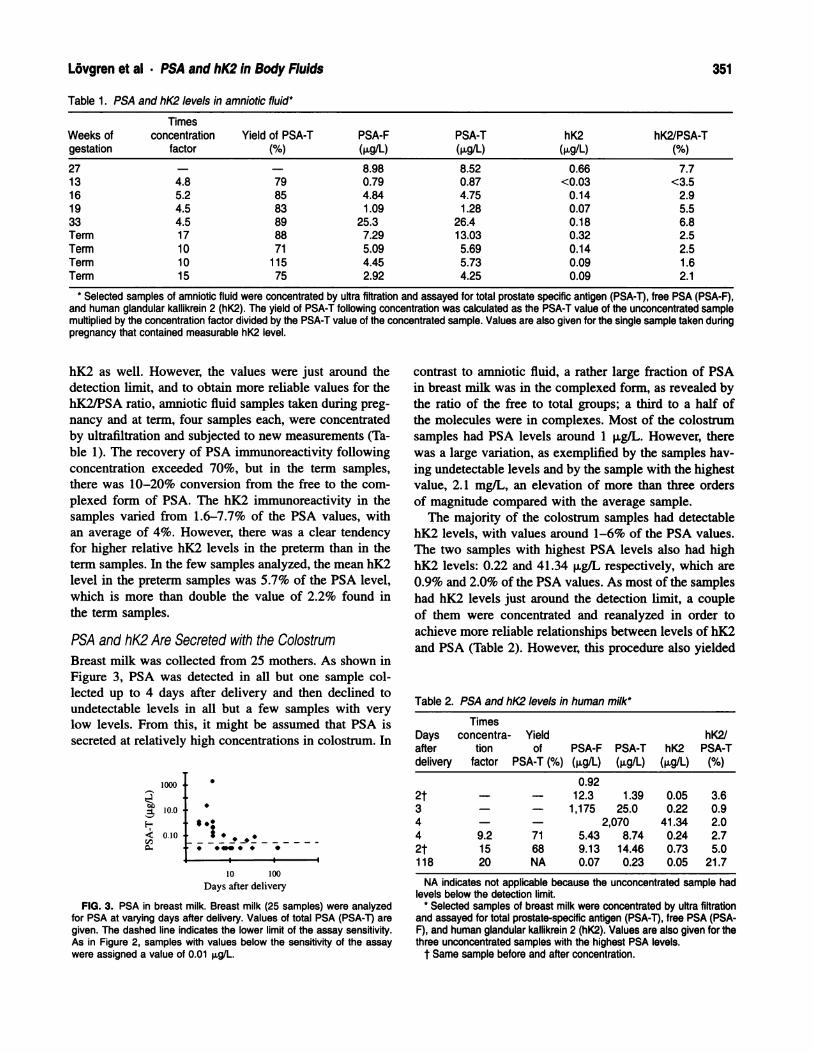
Levy et at . Seminiferous Epithelium of the Aging Rat 359
FIG. 2. Electron micrographs showing the effects of age on the morphology of the seminiferous epithelium of the Brown Norway rat. (A) Seminiferoustubules at stage VII of a 3-month-old rat. G, Golgi apparatus; arrowheads, ER cisternae; arrows, lysosomes; open arrow, junctional complex; PL,preleptotene spermatocyte; N, Sertoli cell nucleus; MY, myoid cell; asterisk, basement membrane; E, endothelial cell; Ley, Leydig cell. Scale bar =
1 .3 p.m. (B) Seminiferous tubule of a 24-month-old rat. Few germ cells can be found; Sertoli cell processes enclose large, empty intercellular spacesin the epithelium where germ cells may have resided. In all tubules devoid of germ cells, Sertoli cells do not show any cyclical variation with regardto their organelles. N, Sertoli cell nucleus; n, nucleolus; arrowheads, ER cistemae; G, Golgi apparatus; L, irregular lysosomes; curved arrows, focalcontact points; asterisks, intercellular spaces; BM, basement membrane; Gc, germ cell. Scale bar = 1 .8 p.m.
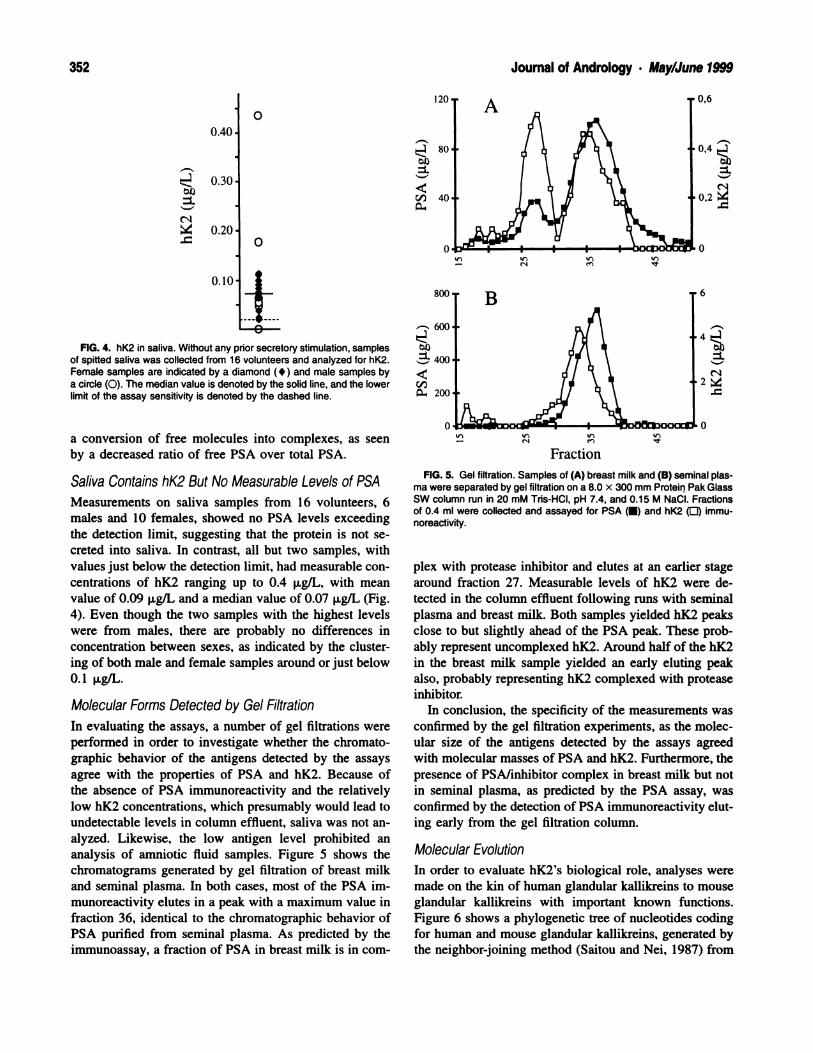
360 Journal of Andrology . May/June 1999
The typical morphological configuration of the blood-
testis barrier seen in young animals was not conspicuous
in aged animals (Figs. 2B, 3A). The vast majority of ap-
posing Sertoli cell plasma membranes were devoid of the
apposing layers of endoplasmic reticulum and bundles of
filaments characteristic of this barrier, although an occa-
sional unapposed endoplasmic reticulum cisterna could be
seen (Fig. 2B, 3A). However, numerous focal contact
points were observed, especially in areas where the pro-
cesses of three Sertoli cells joined one another (Fig. 3A).
In the epithelium, large intercellular spaces were encom-
passed by Sertoli cell processes connected by these focal
points (Fig. 3A and inset). The intercellular spaces took
on various shapes and sizes and appeared to be main-
tained by the Sertoli cell processes (Fig. 3A). They did
not have a content and may represent sites formerly oc-
cupied by germ cells.
In 3-month-old animals, ectoplasmic specializations
consisted of a layer of endoplasmic reticulum and fila-
ments that was oriented parallel to the Sertoli cell plasma
membrane and apposed to the spermatid’s plasma mem-
brane facing the acrosome (not shown). In 24-month-old
animals, ectoplasmic specializations showed a variable
orientation and appearance (Fig. 3B). In some cases, ec-
toplasmic specializations faced the lumen and were ori-
ented parallel to the Sertoli plasma membrane, whereas
in other cases, they were perpendicular to it and in mul-
tiple rows (Fig. 3B). Some ectoplasmic specializations
closely apposed the plasma membrane of adjacent Sertoli
cells but with varied orientations. Occasionally, multiple
rows of ectoplasmic specializations were stacked parallel
to each other and to the Sertoli cell plasma membrane
(Fig. 3B, inset). It should be noted that microtubules con-
tinued to be abundant in the apical processes of Sertoli
cells of the 24-month-old Brown Norway rats (Fig. 3B).
Lanthanum Tracer
In 3-month-old Brown Norway rats, lanthanum nitrate
lined the capillaries and penetrated the tunica propria and
the basement membrane to enter the seminiferous tubules
(Fig. 4A). There were regions, however, where lanthanum
nitrate was prevented from entering the epithelium by the
junctions in the myoid cell layer (not shown). When lan-
thanum entered the seminiferous epithelium, the tracer
could be seen surrounding germ cells in the basal com-
partment (Fig. 4A). In most cases, lanthanum nitrate was
seen encompassing spermatogonia and entering the inter-
cellular space up to the tight junctions between adjacent
Sertoli cells (Fig. 4A). At 24 months, lanthanum nitrate
was seen in the epithelium of the basal (Fig. 4B and inset)
and adluminal regions of the regressed seminiferous tu-
bules (Fig. 5). The lanthanum nitrate was not seen as a
continuous line but rather was intermittently noted be-
tween adjacent Sertoli cell plasma membranes at various
levels of the epithelium (Figs. 4B, 5). The tracer also
appeared in the intercellular spaces (Fig. 5, inset). Lan-
thanum nitrate was seen between adjacent plasma mem-
branes of Sertoli cells near the lumen, and the tracer was
observed in the lumen as small, dark speckles (Fig. 5 and
inset).
Discussion
The observations made with the light microscope were in
agreement with previous studies on the aging Brown Nor-
way rat testis (Wright et al, 1993; Richardson et al, 1995).
There were progressive alterations in the seminiferous ep-
ithelium with increasing age. Normal seminiferous tu-
bules seen in 3-month-old and 12-month-old Brown Nor-
way rats were intermixed with partially regressed tubules
at 18 months. By 24 months, while some normal tubules
were still present, they were adjacent to numerous fully
regressed tubules that were virtually devoid of germ cells
and contained large intercellular spaces. Thus, since the
vast majority of seminiferous tubules in our study were
fully regressed by 24 months, they were the focus of the
electron-microscopic analysis.
In the present study, we did not observe multinucleated
(Schulze and Schulze, 1981) or dedifferentiated (Paniagua
et al, 1985) Sertoli cells that fit the description of those
seen in the aged human testis in tubules with arrested
maturation of spermatogenic cells at spermatogonium or
spermatocyte stages (Paniagua et al, 1991). The dediffer-
entiated Sertoli cells reported in the aged human testis
lacked the abundant smooth endoplasmic reticulum and
irregularly outlined nucleus with the large, centrally lo-
cated tripartite nucleolus characteristic of adult Sertoli
cells (Paniagua et al, 1991). In addition, extensive junc-
tional specializations and abundant lipid droplets were ob-
served (Paniagua et al, 1991). From our analysis, we re-
port a loss in the cyclic modifications of the organelles
of Sertoli cells at 24 months. It has been proposed that
structural changes in Sertoli cells during the cycle of the
seminiferous epithelium (Fawcett, 1975; Kerr et al, 1979;
Russell, 1993; Hermo et a!, 1994) were due to the capac-
ity of the germ cells to influence the cytoplasmic orga-
nization of the Sertoli cells (Parvinen, 1982). In our ex-
periment, it is possible that the loss of cyclical changes
in the organelles of the Sertoli cell was a result of the
depletion of germ cells.
The complete loss with age of cyclical variations of the
organelles of Sertoli cells may be the cause of major al-
terations in the functions of these Sertoli cells. The en-
doplasmic reticulum and Golgi apparatus were generally
abundant in stages VII and VIII of the young Brown Nor-
way rat and were necessary for the synthesis and secre-
tion of many of the proteins required by germ cells (Skin-

Levy et al . Seminiferous Epithelium of the Aging Rat 361
FIG. 3. (A) Several large, empty intercellular spaces enveloped by Sertoli cell processes in the seminiferous tubules of a 24-month-old BrownNorway rat. Typical Sertoli-Sertoli junctional complexes are not seen; one unapposed ER cistema can be observed at times. However, numeroussmall focal contact points join the Sertoli cell processes together. Note ectoplasmic specializations in Sertoli cell process (open arrow). (A) Inset:
High-power electron micrograph of the focal contact points joining the Sertoli cell processes. Asterisks, intercellular spaces; 5, Sertoli cell processes;curved arrows, focal contact points; small arrows, unapposed ER cistema. Scale bar: A = 2.0 pm; inset, A = 0.7 p.m. (B) Apical region of Sertolicells of a 24-month-old Brown Norway rat. Ectoplasmic specializations persist in Sertoli cell processes even in the absence of germ cells. They varyin their distribution and orientation within these processes. (B) Inset: High-power electron micrograph of ectoplasmic specializations oriented perpen-dicular to the plasma membrane. S. Sertoli cells; arrowheads, ectoplasmic specializations between Sertoli cells; open arrows, ectoplasmic speciali-zations free of contact with any cells; large arrowheads, ER cistemae; box, microtubule; Lu, lumen. Scale bar: B = 0.4 p.m; inset, B = 0.5 p.m.

362 Journal of Andrology . May/June 1999
FIG. 4. Stained and unstained electron micrographs showing the ef-fects of age on the permeation of lanthanum into the seminiferous epi-thelium of the Brown Norway rat. (A) Seminiferous tubule of a 3-month-old rat. Lanthanum nitrate can be seen crossing the basement mem-brane, the tunica propria, and the junctions of the myoid cell layer toenter the seminiferous tubules. Lanthanum surrounds the spermatogoniabut is stopped by the Sertoli-Sertoli cell tight junction. Stained sections.BM, basement membrane; SG, spermatogonia; arrow, tight junction.Scale bar = 1.4 p.m. (B) Lanthanum nitrate can be observed in the basal
ner and Griswold, 1982; Kerr 1988; Griswold, 1988; Her-
mo et a!, 1994). However, in aged animals, certain pro-
teins such as CP-2/cathepsin L made by Sertoli cells are
reduced (Wright et al, 1993); this is consistent with the
reduction noted in endoplasmic reticulum and Golgi ap-
paratus.
In addition, the numerous, small lysosomes present in
specific stages in the young animal were replaced by
large, heterogeneous lysosomes seen scattered in the Ser-
toli cell cytoplasm of aged animals. The reduction in the
number of lysosomes in the aging Brown Norway rats
may be due to the fact that the Sertoli cells have less of
a need for these organelles since there are no residual
bodies to eliminate in the aged testes. The large size of
the lysosomes could result from their inability to break
down waste products such as lipofuscin. An accumulation
of lipofuscin has also been seen in many aging tissues
(Sohal and Brunk, 1990; Von Zgliicki et al, 1995; Weis-
se, 1995), including the epithelium of the aging Brown
Norway epididymis (Sene and Robaire, 1998), for which
it was proposed that this accumulation may be caused by
a decline in the degradation pathways or by a buildup of
oxidative stress (Brunk et a!, 1992; Von Zglinicki et al,
1995).
An accumulation of lipids with age has been seen in
previous studies and was suggested to be due to degen-
erating germ cells (Paniagua et a!, 1987). Studies on var-
ious seasonal breeders have also shown a buildup of lipid
in the Sertoli cells during testicular regression (Sinha Hik-
im et al, 1989; Hodgson et al, 1979). Vitamin E deficien-
cy, involving incomplete spermatogenesis, also results in
germ cell degeneration and in lipid buildup in Sertoli cells
(Bensoussan et al, 1998). However, in the present study,
we did not note lipid accumulation in Sertoli cells in fully
regressed tubules.
Interestingly, the morphology of the Sertoli cells in the
aged testis resembled, in part, that reported for the tran-
sitional cells (modified Sertoli cells) of the intermediate
region of seminiferous tubules of the adult rat (Hermo
and Dworkiri, 1988). The transitional cells showed an
abundance of microtubules and showed single or stacked
ectoplasmic specializations oriented parallel and perpen-
dicular to the apical Sertoli plasma membrane in the ab-
sence of spermatids. In this respect, they have the same
characteristics that were seen in the aged testis of the
Brown Norway rat. It has been suggested that the ecto-
plasmic specializations serve as adhesion sites for sper-
matids (Vogi, 1988); our results suggest that in the ab-
compartment of the seminiferxus tubule of the 24-month-old rat betweenadjacent Sertoli cells. Inset High-power electron mkrograph of lanthanumnitrate in the basal region. N, Sertoli cell nucleus; arrows, lanthanum nitrate;
BM, basement membrane. Scale bar: B = 0.9 p.m; inset, B = 0.3 p.m.

Levy et al Seminiferous Epithelium of the Aging Rat363
Lu4,
, .r_
1’p
pp,.
- .* 1y
-
Lu
FIG. 5. Seminiferous tubule of a 24-month-old Brown Norway rat. Lanthanum nitrate can be seen intermittently between Sertot cell plasma mem-branes in the adluminal compartment of the seminiferous epithelium. The tracer surrounds the intercellular spaces and enters the lumen. Unstainedsection. Inset: Lanthanum nitrate can also be seen within the intercellular spaces. Large, irregularly shaped lysosomes are observed in close proximityto these areas. Unstained section. Asterisks, intercellular spaces; S, Sertoli cells; arrows, lanthanum nitrate; L, lysosomes; Lu, lumen. Scale bar: 0.8p.m; inset: 2.1 p.m.
sence of spermatids they lose their orientation. The tran-
sitional cells were described as undergoing fluid-phase
and adsorptive endocytosis (Hermo and Dworkin, 1988)
and have been implicated in the phagocytosis of sper-
matozoa (Dym, 1974; Osman 1978; Nykanen, 1979).
These functions could also be postulated to be the ones
of aged Sertoli cells, as we suspect that the luminal milieu
may be modified because of the alterations in the blood-
testis barner and because of the depletion of germ cells
in these aged animals.
In the present study, the intercellular spaces are located
in regions where spermatocytes and spermatids appeared
to have resided. These spaces are surrounded by Sertoli
cell processes that are usually connected by focal contact
points and consist of three Sertoli cells. Similar intercel-
lular spaces were observed in the rat testes after surgical
induction of cryptorchidism (Kerr et al, 1979) and in Va-
sectomized hamsters (Flickinger, 1981), where it was as-
sumed that these spaces developed from premature ex-
foliation of germ cells. In our study, such spaces suggest
the location of germ cells that have undergone degener-
ation and are lost from the epithelium. They also suggest
that the integrity of the Sertoli cell processes persisted
around these spaces because of an intact cytoskeletal sys-
tem. The abundance of microtubules supports the hypoth-
esis that the cytoskeleton of the Sertoli cells maintained
Copyright © 2022 FDOKUMEN




















