
Mapping Putative Contact Sites Between Subunits in a Bacterial ATP-binding Cassette (ABC)...
14
Mapping Putative Contact Sites Between Subunits in a Bacterial ATP-binding Cassette (ABC) Transporter by Synthetic Peptide Libraries Bettina Blüschke 1 , Viola Eckey 1 , Britta Kunert 1 , Susanne Berendt 1 Heidi Landmesser 1 , Michael Portwich 2 , Rudolf Volkmer 2 and Erwin Schneider 1 ⁎ 1 Humboldt Universität zu Berlin, Institut für Biologie/Bakterienphysiologie, Chausseestr. 117, D-10115 Berlin, Germany 2 Institut für Medizinische Immunologie, Charité- Universitätsmedizin Berlin, Schumannstr. 20-21, D-10098 Berlin, Germany The maltose ATP-binding cassette transporter of Salmonella typhimurium is composed of the soluble periplasmic receptor, MalE, and a membrane- associated complex comprising one copy each of the pore-forming hydrophobic subunits, MalF and MalG, and of a homodimer of the ATP- hydrolyzing subunit, MalK. During the transport process the subunits are thought to undergo conformational changes that might transiently alter molecular contacts between MalFG and MalK 2 . In order to map sites of subunit–subunit interactions we have used a comprehensive peptide map- ping approach comprising large-scale microsynthesis of labelled probes and array techniques. In particular, we screened the binding of (i) MalFG- derived soluble biotinylated peptides to immobilized MalK, and (ii) radiolabelled MalK to MalFG-derived cellulose membrane-bound peptides. The first approach identified seven peptides (10mers) each of MalF and MalG that specifically bound to MalK. The peptides were localized to TMDs 3 and 6, periplasmic loop P4 and cytoplasmic loops C2 and C3 of MalF, while MalG-derived peptides localized to the N terminus, TMDs 4–6, periplasmic loop P1 and cytoplasmic loop C2. Peptides from C3 and C2, respectively, of MalF and MalG partially encompass the conserved EAA- motif, known to be crucial for interaction with MalK. These results were basically confirmed by screening MalFG-derived peptide arrays consisting of 16mers or 31mers with radiolabelled MalK. This approach also allowed us to perform complete substitutional analyses of peptides in question. The results led to the construction of MalFG variants that were subsequently analyzed for functional consequences in vivo. Growth experiments revealed that most of the mutations had no phenotype, suggesting that the mutated residues themselves are not critical but part of a discontinuous binding site. However, two novel mutations affecting residues from the EAA motifs of MalF (Ile417Glu) and MalG (Phe203Gln/Asn), respectively, displayed severe growth defects, indicating their functional importance. Together, these experimental outcomes identify specific molecular contacts made between MalK and MalFG that extend beyond the well-characterized EAA motif. © 2007 Elsevier Ltd. All rights reserved. *Corresponding author Keywords: ATP-binding cassette; maltose transport; Salmonella typhimurium; protein–protein interactions; synthetic peptide arrays Abbreviations used: ABC, ATP-binding cassette; CAPE, cellulose-amino-hydroxypropyl ether; DIPEA, diisopropylethylamine; Fmoc-, 9-fluorenyl methoxy carbonyl-; RP-HPLC, reverse phase high pressure liquid chromatography; MALDI-TOF, matrix-assisted laser desorption ionisation-time of flight; TBTU, O-(benzotriazol-1-yl)-N, N,N′,N′-tetramethyluronium-tetrafluoroborate. E-mail address of the corresponding author: [email protected] ARTICLE IN PRESS YJMBI-59255; No. of pages: 14; 4C: 4, 5 doi:10.1016/j.jmb.2007.03.043 J. Mol. Biol. (2007) xx, xxx–xxx 0022-2836/$ - see front matter © 2007 Elsevier Ltd. All rights reserved. Please cite this article as: Blüschke, B. et al., Mapping Putative Contact Sites Between Subunits in a Bacterial ATP-binding Cassette (ABC) Transporter by Synthetic Peptide Libraries, J. Mol. Biol. (2007), doi:10.1016/j.jmb.2007.03.043
-
Upload
charite-de -
Category
Documents
-
view
3 -
download
0
Transcript of Mapping Putative Contact Sites Between Subunits in a Bacterial ATP-binding Cassette (ABC)...

ARTICLE IN PRESSYJMBI-59255; No. of pages: 14; 4C: 4, 5
doi:10.1016/j.jmb.2007.03.043 J. Mol. Biol. (2007) xx, xxx–xxx
Mapping Putative Contact Sites Between Subunits in aBacterial ATP-binding Cassette (ABC) Transporter bySynthetic Peptide Libraries
Bettina Blüschke1, Viola Eckey1, Britta Kunert1, Susanne Berendt1
Heidi Landmesser1, Michael Portwich2, Rudolf Volkmer2
and Erwin Schneider1⁎
1Humboldt Universitätzu Berlin, Institut fürBiologie/Bakterienphysiologie,Chausseestr. 117,D-10115 Berlin, Germany2Institut für MedizinischeImmunologie, Charité-Universitätsmedizin Berlin,Schumannstr. 20-21,D-10098 Berlin, GermanyAbbreviations used: ABC, ATP-bidiisopropylethylamine; Fmoc-, 9-fluchromatography; MALDI-TOF, matrN,N′,N′-tetramethyluronium-tetrafluE-mail address of the correspondi
0022-2836/$ - see front matter © 2007 E
Please cite this article as: Blüschke, B. e(ABC) Transporter by Synthetic Peptid
The maltose ATP-binding cassette transporter of Salmonella typhimurium iscomposed of the soluble periplasmic receptor, MalE, and a membrane-associated complex comprising one copy each of the pore-forminghydrophobic subunits, MalF and MalG, and of a homodimer of the ATP-hydrolyzing subunit, MalK. During the transport process the subunits arethought to undergo conformational changes that might transiently altermolecular contacts between MalFG and MalK2. In order to map sites ofsubunit–subunit interactions we have used a comprehensive peptide map-ping approach comprising large-scale microsynthesis of labelled probes andarray techniques. In particular, we screened the binding of (i) MalFG-derived soluble biotinylated peptides to immobilized MalK, and (ii)radiolabelled MalK to MalFG-derived cellulose membrane-bound peptides.The first approach identified seven peptides (10mers) each of MalF andMalG that specifically bound toMalK. The peptides were localized to TMDs3 and 6, periplasmic loop P4 and cytoplasmic loops C2 and C3 of MalF,while MalG-derived peptides localized to the N terminus, TMDs 4–6,periplasmic loop P1 and cytoplasmic loop C2. Peptides from C3 and C2,respectively, of MalF and MalG partially encompass the conserved EAA-motif, known to be crucial for interaction with MalK. These results werebasically confirmed by screening MalFG-derived peptide arrays consistingof 16mers or 31mers with radiolabelled MalK. This approach also allowedus to perform complete substitutional analyses of peptides in question. Theresults led to the construction of MalFG variants that were subsequentlyanalyzed for functional consequences in vivo. Growth experiments revealedthat most of the mutations had no phenotype, suggesting that the mutatedresidues themselves are not critical but part of a discontinuous binding site.However, two novel mutations affecting residues from the EAA motifs ofMalF (Ile417Glu) and MalG (Phe203Gln/Asn), respectively, displayedsevere growth defects, indicating their functional importance. Together,these experimental outcomes identify specific molecular contacts madebetween MalK and MalFG that extend beyond the well-characterized EAAmotif.
© 2007 Elsevier Ltd. All rights reserved.
Keywords: ATP-binding cassette; maltose transport; Salmonella typhimurium;protein–protein interactions; synthetic peptide arrays
*Corresponding authornding cassette; CAPE, cellulose-amino-hydroxypropyl ether; DIPEA,orenyl methoxy carbonyl-; RP-HPLC, reverse phase high pressure liquidix-assisted laser desorption ionisation-time of flight; TBTU, O-(benzotriazol-1-yl)-N,oroborate.ng author: [email protected]
lsevier Ltd. All rights reserved.
t al., Mapping Putative Contact Sites Between Subunits in a Bacterial ATP-binding Cassettee Libraries, J. Mol. Biol. (2007), doi:10.1016/j.jmb.2007.03.043

2 Contact Sites in a Bacterial ABC Transporter
ARTICLE IN PRESS
Introduction
ATP-binding cassette (ABC) transporters exist inall living organisms and form one of the largestsuperfamilies. They are integral to almost everybiological process and physiological system. ABCtransport systemsmediate the uptake or export of anenormous variety of substances across cell mem-branes, from small ions to large polypeptides, atthe expense of ATP.1 They share a common archi-tectural organization comprising two hydrophobictransmembrane domains (TMDs) that form thetranslocation pathway and two hydrophilic nucleo-tide binding (ABC) domains (NBDs) that hydrolyzeATP. In prokaryotes, these domains are mostlyexpressed as separate protein subunits, whereas ineukaryotes, especially in mammalian cells, they areusually fused into a single polypeptide chain.The maltose ABC transporter of Escherichia coli/
Salmonella typhimurium is one of the best character-ized systems that can serve as a model for ABCimporters.2,3 The transporter is composed of theextracellular (periplasmic) receptor, the maltosebinding protein (MBP or MalE), and the mem-brane-bound complex comprising the hydrophobicsubunits, MalF and MalG, and two copies of theATPase (ABC) subunit, MalK. MalF has a molecularmass of 55 kDa and is predicted to span themembrane eight times. It is further characterizedby a large periplasmic loop (20 kDa) of, thus far,unknown function, connecting putative TMDs 3 and4. MalG is predicted to consist of six TMDs and has amolecular mass of 33 kDa (see also Figure 3). MalKand closely related ABC subunits contain a uniqueC-terminal extension which, in the crystal structureof the MalK dimer, contributes substantially tomonomer–monomer contacts.4
Transport of maltose is assumed to be initiated byinteraction of substrate-loaded MalE with periplas-mic loops of MalFG, thereby triggering conforma-tional changes that result in ATP hydrolysis at theMalK subunits and eventually in substrate trans-location.5 Binding of liganded MalE and ATP occursrather simultaneously and a transient stable complexof MalFGK2 with MalE is formed.6 ATP-dependentclosure of the MalK dimer interface coincides withopening of MalE and maltose release to thetransporter.7 ATP hydrolysis is required to reset thetransporter to the resting state by opening thecontacts between the MalK subunits.8
ATP-induced conformational changes of the iso-lated MalK dimer4,9 as well as of the subunits in theassembled complex10–12 have been well documen-ted. However, little is known on the binding sites bywhich the subunits contact each other. It is wellestablished that association of the MalK subunitswith MalF and MalG requires at least the so-calledEAA sequence motifs (consensus EAAx3Gx9Ix1LP;x, any amino acid) which are conserved in the lastcytoplasmic loop regions of TMDs of ABC importsystems10,13 (see also Figure 3). The role of theEAA motif in subunit–subunit interactions was con-firmed by the crystal structure of the E. coli vitamin
Please cite this article as: Blüschke, B. et al., Mapping Putative Cont(ABC) Transporter by Synthetic Peptide Libraries, J. Mol. Biol. (2007
B12 transporter (BtuCD), thus far the only ABCimport system for which structural information isavailable.14 Nonetheless, it is unclear whether theseinteractions are sufficient to allow formation of astable MalFGK2 complex that requires treatmentwith 6.6 M urea to release MalK.15 Moreover,contact sites might be altered transiently as a resultof conformational changes during the transportcycle. Thus, here, we have employed biotinylatedsoluble and cellulose membrane-bound syntheticpeptides derived from MalFG to map their interac-tions with the ATP-hydrolyzing MalK subunits. Thisapproach has been widely used to map antibodyepitopes and to identify protein interaction domains,to name just a few applications.16 Our study,complemented by in vivo data of selected mutantsreveals contact sites that extend beyond the EAAmotif, including peptide regions localized to TMDsand periplasmic loops of MalFG.
Results
Enzyme-linked absorbance studies: binding ofMalFG-derived biotinylated soluble peptides toimmobilized MalK
As an initial approach to map the putative contactsites between MalK and MalFG we analyzedMalFG-derived soluble peptides for their capabilityto bind to purified MalK. To this end, 15mersrepresenting the complete primary structures ofMalF and MalG (total of 168 and 96 peptides,respectively) that overlapped by nine amino acidresidues were synthesized on a cellulose membrane,biotinylated and subsequently cleaved off the solidphase as described.17,18 Mixtures of two consecutivepeptides were then incubated with purified MalKpreviously immobilized to the wells of a microtiterplate. After washing off unbound peptides, theamounts of retained peptides were quantifiedenzymatically after incubation with peroxidase-conjugated avidin. In order to account for unspecificbinding the obtained read-outs were corrected bysubtracting absorbance values of the respectivepeptide mixes with immobilized bovine serumalbumin (BSA). This unspecific reaction was in therange from zero to 40% of the absorbance measuredwith MalK. A total of 16 and 12 putative bindingregions were identified in MalF and MalG, respec-tively (not shown).Subsequently, in order to narrow the number of
possibly interacting peptides, a new set of peptidesrepresenting the above identified binding regionswas synthesized. This time the peptides were10mers overlapping by nine amino acid residuesand each was incubated separately with immobi-lized MalK. Again, immobilized BSAwas used as acontrol. As a representative example of the dataobtained, Figure 1 shows the absorbance values ofpeptides representing the region Cys304 to Val327 ofMalF. Peptide 9 (K312-GKAIYRVL-L321) displayed
act Sites Between Subunits in a Bacterial ATP-binding Cassette), doi:10.1016/j.jmb.2007.03.043

Figure 1. Binding of soluble biotinylated peptide 9 ofMalF to immobilized MalK. Peptide binding assays wereperformed in microtiter plates in the presence (blackbars) or absence (grey bars) of ATP/Mg2+ as describedin Materials and Methods. Absorbance readings werecorrected for unspecific binding by substracting absor-bance values obtained with BSA in parallel assays. Arepresentative data set out of three independent assaysis shown. While absolute absorbance values varied bet-ween experiments the ratio of read-outs of each peptideto the strongest binder (K312-L321) did not changesignificantly (<30%).
Figure 2. Comparative binding of soluble biotinylatedMalFG-derived peptides to MalK that displayed strongestbinding intensities in the individual assays. Assay condi-tions were as described in Materials and Methods and inthe legend to Figure 1. (a) and (b) MalF-derived peptides.(c) MalG-derived peptides. Peptide MalF-6 was analyzedin the absence of ATP/Mg2+ only. The absorbance valuesof each peptide represent the average of three ((a) and (b))and two (c) experiments, respectively. Deviations from themean were <39% ((a) and (b)) and <30% (c).
3Contact Sites in a Bacterial ABC Transporter
ARTICLE IN PRESS
the strongest signal while the readings for bothneighboring peptides were substantially lower.These data indicate that in particular residuesK312 and L321 are crucial for binding to MalK inthe context of peptide 9 (see also below). Thepresence of ATP/Mg2+ in the incubation bufferseemed to slightly weaken the binding.In Figure 2 a comparative binding assay is shown
with those peptides from each of the analyzedregions of MalF and MalG that exhibited strongestbinding to MalK. Each column represents theaverage of three experiments that were correctedfor unspecific binding to BSA as described above. Byusing a cut-off of 0.2, ten out of 17 MalF-derivedpeptides were considered to exhibit significantbinding activities, with peptides 4, 5, 8, 9 and 11 ofMalF giving the most intense signals. Among those,peptides 3 and 5 also displayed substantial bindingto BSA and thus were not studied further. Onlybinding of peptides 12 and 14 was strongly affectedby ATP/Mg2+. Interestingly, peptide 13 encompass-ing the N-terminal part of the conserved EAA motifexhibited almost no binding activity for MalK.In case of MalG, eight out of 14 peptides gave
absorbance values of 0.2 or higher. The most intensesignals were obtained with peptides 1 and 12–14.With the exception of peptide 12, all displayedsomewhat weaker binding in the presence of ATP/Mg2+. Peptide 11 that comprises the N-terminal partof the EAA motif was only weakly reacting, whilepeptide 12 representing the C-terminal region of themotif displayed the strongest signal of all peptidesanalyzed. Peptide 5 exhibited some binding to BSAand was thus not further considered. Table 1compiles those MalFG-derived peptides that exhib-ited specific binding to MalK. The positions of each
Please cite this article as: Blüschke, B. et al., Mapping Putative Cont(ABC) Transporter by Synthetic Peptide Libraries, J. Mol. Biol. (2007
of the peptides within the topological models ofMalF and MalG are shown in Figure 3(a) and (b)(residues in blue). As expected from the localizationof the MalK dimer, peptides within cytoplasmicallyexposed loop regions of both proteins (MalF: C2 andC3; MalG: N terminus and C2) are involved inbinding. However and most surprisingly, peptidesfrom transmembrane and periplasmic domains
act Sites Between Subunits in a Bacterial ATP-binding Cassette), doi:10.1016/j.jmb.2007.03.043

Table 1.MalFG-derived peptides with strong and specificbinding affinity for immobilized MalK
PeptideaEffect ofMgATP
Topologicallocalizationb Sequence
MalF4 − TMD 3 V79-LFPLVCTI-A888 − (TMD 4), C2 M300-VLACLVQW-E3099 − C2, (TMD 5) K312-GKAIYRVL-L32111 − TMD 6 L379-GYPYMMIL-C38812 + TMD 6, C3 C388-MGLLKAIP-D39714 + C3 Q412-NFFKITLP-L42117 − P4 L467-VSYTYRIA-F476
MalG1 (+) N terminus K7-SQKLRLLI-T166 − P1 F79-PVLLWLWN-S889 (+) TMD 4 Y166-LGGIALHV-W17510 (+) TMD 4, C2 H173-VWTIKGYF-E18212 − C2, (TMD 5) W200-QAFRLVLL-P20913 (+) (TMD 5), P3 I220-LSFIAAIT-E22914 (+) TMD 6 D262-FAAAAVLS-A271
a Numbering is according to Figure 2.b The topological localization is according to Figure 3.
4 Contact Sites in a Bacterial ABC Transporter
ARTICLE IN PRESS
were also identified. These peptides localize toTMDs 3–5 and periplasmic loop P4 of MalF, and toTMDs 4–6 and loop P1 of MalG.
Please cite this article as: Blüschke, B. et al., Mapping Putative Cont(ABC) Transporter by Synthetic Peptide Libraries, J. Mol. Biol. (2007
Array studies: probing MalF and MalG-derivedpeptide arrays for MalK binding
In order to obtain further support for thesefindings, we wished to perform detailed substitu-tional analyses of the identified peptides. However,using again soluble peptides would have been toolaborious and time-consuming even with the nano-synthesis approach. Thus, we took advantage of theSPOT synthesis technology that permits parallelsynthesis and screening of a large number ofcellulose membrane-bound peptides. First, cellu-lose-bound peptide arrays representing the com-plete MalF and MalG sequences were screened forbinding of MalK. The peptide arrays consisted of16mers or 31mers, that overlapped with adjacentpeptides by 15 and 30 amino acid residues, res-pectively. The cellulose membranes were incubatedwith 35S-labelled MalK, washed to remove excesslabel, and retained radioactivity was visualized byphosphoimaging. The results obtained with peptidearrays containing 16mers are shown in Figure 4(a)(MalF) and (b) (MalG). For both proteins identicalsignal patterns were obtained with 31mers (notshown). Thirteen and ten signal rows, respectively,were found for MalF and MalG, albeit with different
Figure 3. Topological models ofMalF (a) and MalG (b) according toFroshauer et al.37 and Boyd et al.,38
respectively. Residues from peptideregions identified by enzyme-lin-ked binding assays and arrays areshown in blue. Residues suggestedfrom substitutional analyses to becrucial for interactionwithMalK areshown in red. Residues discussed inthe text are indicated by their codonnumber. Residues that are part ofthe EAA consensus sequence areunderlined. P, Periplasmic loops; C,cytoplasmic loops.
act Sites Between Subunits in a Bacterial ATP-binding Cassette), doi:10.1016/j.jmb.2007.03.043
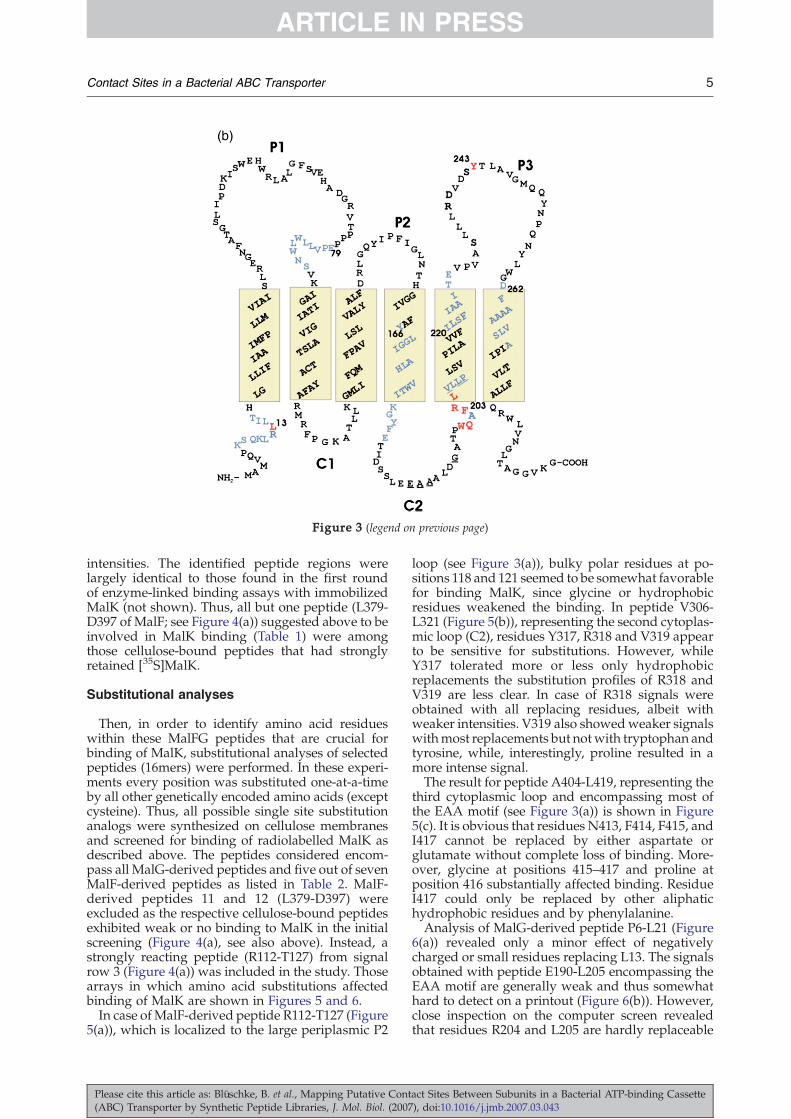
Figure 3 (legend on previous page)
5Contact Sites in a Bacterial ABC Transporter
ARTICLE IN PRESS
intensities. The identified peptide regions werelargely identical to those found in the first roundof enzyme-linked binding assays with immobilizedMalK (not shown). Thus, all but one peptide (L379-D397 of MalF; see Figure 4(a)) suggested above to beinvolved in MalK binding (Table 1) were amongthose cellulose-bound peptides that had stronglyretained [35S]MalK.
Substitutional analyses
Then, in order to identify amino acid residueswithin these MalFG peptides that are crucial forbinding of MalK, substitutional analyses of selectedpeptides (16mers) were performed. In these experi-ments every position was substituted one-at-a-timeby all other genetically encoded amino acids (exceptcysteine). Thus, all possible single site substitutionanalogs were synthesized on cellulose membranesand screened for binding of radiolabelled MalK asdescribed above. The peptides considered encom-pass all MalG-derived peptides and five out of sevenMalF-derived peptides as listed in Table 2. MalF-derived peptides 11 and 12 (L379-D397) wereexcluded as the respective cellulose-bound peptidesexhibited weak or no binding to MalK in the initialscreening (Figure 4(a), see also above). Instead, astrongly reacting peptide (R112-T127) from signalrow 3 (Figure 4(a)) was included in the study. Thosearrays in which amino acid substitutions affectedbinding of MalK are shown in Figures 5 and 6.In case ofMalF-derived peptide R112-T127 (Figure
5(a)), which is localized to the large periplasmic P2
Please cite this article as: Blüschke, B. et al., Mapping Putative Cont(ABC) Transporter by Synthetic Peptide Libraries, J. Mol. Biol. (2007
loop (see Figure 3(a)), bulky polar residues at po-sitions 118 and 121 seemed to be somewhat favorablefor binding MalK, since glycine or hydrophobicresidues weakened the binding. In peptide V306-L321 (Figure 5(b)), representing the second cytoplas-mic loop (C2), residues Y317, R318 and V319 appearto be sensitive for substitutions. However, whileY317 tolerated more or less only hydrophobicreplacements the substitution profiles of R318 andV319 are less clear. In case of R318 signals wereobtained with all replacing residues, albeit withweaker intensities. V319 also showedweaker signalswithmost replacements but notwith tryptophan andtyrosine, while, interestingly, proline resulted in amore intense signal.The result for peptide A404-L419, representing the
third cytoplasmic loop and encompassing most ofthe EAA motif (see Figure 3(a)) is shown in Figure5(c). It is obvious that residues N413, F414, F415, andI417 cannot be replaced by either aspartate orglutamate without complete loss of binding. More-over, glycine at positions 415–417 and proline atposition 416 substantially affected binding. ResidueI417 could only be replaced by other aliphatichydrophobic residues and by phenylalanine.Analysis of MalG-derived peptide P6-L21 (Figure
6(a)) revealed only a minor effect of negativelycharged or small residues replacing L13. The signalsobtained with peptide E190-L205 encompassing theEAA motif are generally weak and thus somewhathard to detect on a printout (Figure 6(b)). However,close inspection on the computer screen revealedthat residues R204 and L205 are hardly replaceable
act Sites Between Subunits in a Bacterial ATP-binding Cassette), doi:10.1016/j.jmb.2007.03.043

Figure 4. MalK binding to cellulose-bound peptide arrays. Peptide arrays (16mers) derived from the sequences ofMalF (a) andMalG (b) were probed for MalK binding. Peptide-bound cellulose membranes were prepared and incubatedwith 35S-labelled MalK as described in Materials and Methods. Last spots of rows (right) and N-terminal residues ofpeptides of the first spots of rows (left) are indicated.
6 Contact Sites in a Bacterial ABC Transporter
ARTICLE IN PRESS
without loss of binding activity. R204 tolerated onlysubstitution by lysine and arginine as does L205.The same holds true for F203. Interestingly enough,lysine or arginine at position 201 substantiallyincreased binding of MalK. Together, these findingsmight indicate that a positively charged residue iscrucial in this region of the peptide chain.Peptide V223-V247 (Figure 6(c)), representing the
third periplasmic loop of MalG, showed clearsubstitutional effects on binding of MalK at position243. Residue Y243 was found to be irreplaceable bynegatively charged residues and asparagine but alsoby small hydrophobic residues.
Effect of selected point mutations in malFmalGon maltose transport in vivo
Do these results reflect contact sites of MalK andMalFG in the context of the assembled proteincomplex in vivo? To address this question, we
Please cite this article as: Blüschke, B. et al., Mapping Putative Cont(ABC) Transporter by Synthetic Peptide Libraries, J. Mol. Biol. (2007
introduced selected single, and double mutationsin malF and malG and analyzed their consequenceson transporter activity. To this end, E. coli strain ES66(pFS1)was transformedwith plasmids harboring therespective malF and malG alleles and the resultingtransformants were assayed for growth on maltoseas sole source of carbon and energy. Strain ES66carries a deletion of the entire malB region encodingthe maltose transport proteins while plasmid pFS1provides the malK and malE genes in trans. Residuesthat were mutated according to the results of thesubstitutional analyses included K118(→G), Y317(→Q), K416(→E), and I417(→E) of MalF, and L13(→A), F203(→N,Q) and Y234(→A,N) of MalG (seeFigure 3(a) and (b)). Residues from TMDs were notconsidered in order to exclude possible effects oncorrect membrane insertion and folding. The resultsexpressed in doubling times of cells grown in liquidmedium are summarized in Table 3. Most of themutations allowed growth of the transformant very
act Sites Between Subunits in a Bacterial ATP-binding Cassette), doi:10.1016/j.jmb.2007.03.043

Table 2. Peptides subjected to complete substitutionalanalysis
Peptide
Number ofcorresponding
peptidesfrom Table 1
Topologicallocalization
MalFF78-VLFPLVCTIAIAFT-N93 4 TMD 3R112-SYQAGKTYNFGLYP-T127 − P2V306-QWEALKGKAIYRVL-L321 8+9 C2Q412-NFFKITLPLLIKPL-T427 14 C3G462-YTDLLVSYTYRIAF-E477 17 P4
MalGP6-KSQKLRLLITHLGL-L21 1 N terminus,
TMD 1T75-PPPFPVLLWLWNSV-K90 6 P1G168-GIALHVWTIKGYFE-T183 9+10 TMD 4, C2E190-AAALDGATPWQAFR-L205 12 C2I220-LSFIAAITEVPVAS-L235 13 TMD 5, P3D262-FAAAAVLSAIPITL-V277 14 TMD 6
The peptides identified by binding to immobilized MalK (seeTable 1) are in bold.
7Contact Sites in a Bacterial ABC Transporter
ARTICLE IN PRESS
similar to control cells carrying the malFG wild-typealleles. A slightly slower growth was consistentlyobserved with MalG-Y243A or MalG-L13A, how-ever combination of both mutations did not result inan additive effect. Interestingly, while MalF-K416Gdid not cause a growth defect, although thesubstitution was shown to reduce binding of MalK(Figure 5(c)), the I417E mutant exhibited muchslower growth than wild-type. Likewise, replacingMalG-F203 by asparagine or glutamine substantiallyaffected growth of the transformants. In both cases,this was not caused by dislocalization of MalK asassayed by cell fractionation and subsequent immu-noblotting. About the same amounts of MalK werefound associatedwith the cytoplasmicmembrane forwild-type and both mutants. Furthermore, localiza-tion ofMalF to themembrane indicated that complexassembly hadmost likely occurred (Figure 7; data forI417E not shown). This conclusion is based on theearlier observation that assembly defects causeproteolytic cleavage of MalF.19 Possible effects on
Figure 5. Substitutional analyses of MalF-derived peptid(16mers) corresponding to spots 112 (a), 306 (b) and 404 (c) in Fall other 20 L-amino acids (except cysteine) in alphabetical ordeof 35S-labelled MalK as described in Materials and Methods. A(wt) of the peptides. Amino acids important for binding are u
Please cite this article as: Blüschke, B. et al., Mapping Putative Cont(ABC) Transporter by Synthetic Peptide Libraries, J. Mol. Biol. (2007
the interaction with MalE can of course not be ruledout.Two mutants were subsequently selected for
further characterization. MalF(I417E)GK2 andMalFG(Y243A)K2 were purified from overprodu-cing strains and incorporated into liposomes accord-ing to standard protocols.15 The resulting proteolipo-somes were assayed for MalE-maltose-stimulatedand vanadate-sensitive ATPase activity.15 Both cri-teria are widely used as a measure for transportactivity in vitro.5,6,11,12,15 In accordance with thegrowth experiments, MalF(I417E)GK2 displayed nomeasurable ATPase activity, while MalFG(Y243A)K2exhibited 86% of wild-type activity (100%=2.1 μmolPi/min per mg). Moreover, the latter activity wascompletely inhibited by vanadate (0.1 mM) (data notshown).Together, the observation that most of the mutants
exhibited no phenotype is consistent with the notionthat discontinuous binding sites exist betweenMalFG and MalK. If only of structural importancereplacing one or two residues from these sites byunfavorable residues as predicted from the peptidearrays should not cause a functional defect. In con-trast, the severe growth defects of mutants produ-cing the variant MalG-F203N/Q and MalF-I417E,respectively, are likely suggesting that these residuesare functionally critical for the transport process. Theobservation that complex assembly is unchangedcompared towild-type provides further evidence forthese residues being also part of a discontinuouscontact site.
Discussion
Evaluation of the methods applied to studymembrane protein complexes
We have studied the contact sites of the mem-brane-integral subunits MalFG with the MalKATPase homodimer of the maltose ABC transporterby using MalFG-derived synthetic peptides: firstlyas soluble peptides in the nanomolar range and also
es recognized by MalK. Each amino acid of the peptidesigure 4(a) (right-hand side of membrane) is substituted byr (shown on top of each membrane) and tested for bindingll spots in the left column comprise the wild-type sequencenderlined.
act Sites Between Subunits in a Bacterial ATP-binding Cassette), doi:10.1016/j.jmb.2007.03.043

Figure 6. Substitutional analyses of MalG-derived peptides recognized by MalK. The peptides analyzed correspondto spots 6 (a), 190 (b) and 232 (c) in Figure 4(b). See the legend to Figure 5 for details.
8 Contact Sites in a Bacterial ABC Transporter
ARTICLE IN PRESS
as cellulose membrane-bound peptide arrays bothprepared by the SPOT synthesis approach. Prepara-tion of soluble peptides by the SPOT techniqueresulted approximately in 50–100 nmol peptide perspot (spot area 0.25 cm2).17,18,20–23 Over the pastyears it was found that SPOT-synthesized solublepeptides (up to 18mers) can in most cases be ob-tained with a purity above 60%.20,23 More recently,it was demonstrated that even the 34mericFBP28D15N WW domain could be SPOT synthe-sized with an overall purity of 65%.24 The highlyparallel character of the SPOT synthesis approach,the good synthesis quality of so prepared solublepeptides and the preparation of huge numbers ofpeptides at low costs make them ideal candidates forbiological or biochemical solution-based screeningstudies. SPOT-synthesized soluble peptides havebeen successfully applied to several biological assaysystems like T-cell epitope mapping,22 or mem-brane-integral receptor studies like the PACAP-VIPinteraction.18 However, it should also be mentionedthat the method, like all other solid phase peptidesynthesis approaches, has its limitations when
Table 3. Effects of mutations on cell growth on maltose
MalFG variantssynthesized
Topologicallocalization
Doublingtime (h)
Wild type 5.9
MalFK118G P2 5.9Y317Q C2 6.0K416E C3 5.3I417E C3 9.2
MalGL13A N terminus 6.3F203N C2 9.0F203Q C2 >>10Y243A P3 6.3Y243N P3 5.5L13A, Y243A N terminus, P3 6.3L13A, Y243N N terminus, P3 5.5
Cells of strain ES66(pFS1) harboring the respective pES62-97derivatives were grown in minimal salt medium supplementedwith 2% maltose and appropriate antibiotics at 37 °C. For detailssee Materials and Methods. Data represent average of threeindependent experiments.
Please cite this article as: Blüschke, B. et al., Mapping Putative Cont(ABC) Transporter by Synthetic Peptide Libraries, J. Mol. Biol. (2007
sequences that cannot be synthesized by a standardsynthesis protocol are concerned. Nevertheless, ourcontrol analysis by HPLC and MALDI-TOF massspectrometry of some selected MalFG-derivedsequences revealed peptide identity and purities inthe range of 60–90%.However, analytical control of SPOT synthesized
TMD-deduced peptides failed due to the fact thatthese peptides show extreme poor solubility inaqueous buffer systems. Therefore, standard HPLCand MALDI-TOF mass spectrometry analyses areunsuitable control instruments in this case. None-theless, it was demonstrated that the couplingefficiency of aliphatic and hydrophobic aminoacids is satisfactory.18,25 Furthermore and as demon-strated recently by the successful SPOT synthesis ofseveral 32meric leucine zipper analogs, amphipathichelical peptides can be synthesized with good yields
Figure 7. Subcellular localization of MalK (a) andMalF (b) in mutant strains. Cells of host strain ES66(pFS1)(see Materials and Methods) harboring derivatives ofplasmid pES67-92 that encode wild-type MalFMalG,MalFMalG(F403N), and MalFMalG(F403Q), respectively,were grown in LB medium (10 ml) supplemented withampicillin (0.1 mg/ml) and chloramphenicol (0.02 mg/ml)at 37 °C to late exponential phase. Subsequently, cells weredesintegrated by passage through a French press andfractions corresponding to low speed pellet (LSP), cytosol(Cy) and cytoplasmic membrane vesicles (CM) wereobtained as described.15 Proteins from each fraction(20 μg) were separated by SDS-PAGE, electro-transferredto nitrocellulose membranes and probed with specificantisera raised against MalK and MalF, respectively. St,molecular weight markers.
act Sites Between Subunits in a Bacterial ATP-binding Cassette), doi:10.1016/j.jmb.2007.03.043

9Contact Sites in a Bacterial ABC Transporter
ARTICLE IN PRESS
and quality.26 Taking into account that TMDs areamphipathic helical structures composed of alipha-tic amino acids it is likely that the SPOT synthesis ofthe TMD-deduced peptides of this study wassuccessful on the cellulose membrane.Cellulose membrane-bound peptide arrays pre-
pared by the SPOT synthesis technology haveemerged over the past decade as a powerful proteo-mics tool to study molecular recognition events.16
Well-known applications include the identificationof linear peptide epitopes that bind to antibodies27
or to protein interaction domains.28,29 Recently, acomprehensive and statistically significant studydemonstrated the reliability of array-based mea-surement of peptide binding affinity.30 The intra-membrane comparability was found to be reason-ably good and peptide affinities correlated well withthe measured spot signal intensities. In contrast, theinter-membrane comparability is quite low and adrawback of the method.In a previous study, we have successfully used
peptide arrays to map the binding surfaces of thecytosolic regulatory protein enzyme IIAGlc thatinteract with MalK.31 In contrast, peptide arrayshave not yet been employed to identify contact sitesbetween subunits in a membrane protein complex.Such interactions are likely to include linear as wellas conformational (discontinuous) binding sites,which make their identification more difficult.By pepscan experiments sequentially continuous
protein binding sites can be readily mapped, whichis one the strengths of SPOT technology.16 Discon-tinuous protein–protein contact sites are formed bysequentially distant fragments of the molecule thatare brought into spatial proximity by protein fol-ding. As a result, affinities of these separatedfragments are generally low. Therefore, it is a greatchallenge to map discontinuous contact sites bypepscan experiments. Nevertheless, the first map-ping of discontinuous protein–protein contact sitesapplying the SPOT technology was reported byReineke et al.,32,33 mapping the discontinuousbinding site of interleukin-10. However, because ofthe low affinity the peptides must be adjusted foroptimal length and their specificity has to be testedby additional substitutional analyses and by quan-titative affinity measurement by ELISA and/orsurface plasmon resonance experiments.Nonetheless, by combining data obtained with
soluble and cellulose-bound peptide arrays with theresults frommutational analyses, we have identifiedputative contact sites between MalFG and MalK.While binding assays employing biotinylated solu-ble peptides and immobilized MalK enabled us toaccount for unspecific binding events, cellulosebound peptide arrays allowed complete substitu-tional analyses of peptides in question. Togetherwith the in vivo analysis of mutants, both ap-proaches identified possible interaction sites loca-lized to periplasmic and cytoplasmic loop regions aswell as to TMDs of MalF and MalG.So far, only residues from the conserved EAA
motif have been demonstrated to be crucial for the
Please cite this article as: Blüschke, B. et al., Mapping Putative Cont(ABC) Transporter by Synthetic Peptide Libraries, J. Mol. Biol. (2007
interaction between MalFG and MalK.10,13 Peptidesderived from this motif, although representing theless well-studied carboxy-terminal end only, wereamong those that displayed strongest binding toMalK. Thus, this finding is indicative for thesuitability of the approach used here to map contactsites between subunits of a protein complex.Below we discuss in detail the identified putative
contact sites with respect to other reports.
Putative MalK binding sites in TMDs
No significant binding ofMalK to peptides derivedfrom the N-terminal TMDs 1and 2 of MalF weredetected, which is consistent with findings suggest-ing that both helices are dispensable for function.34,35Specific binding was however found for peptidesV79-A88 (TMD 3), M300-E309 (TMD 4/C2), andL379-D397 (TMD 6/C3). Mutational analysis bySteinke et al.36 identified TMD 3 as crucial for func-tion. Substitutions mainly affected MalF assembly,which does not exclude that contacts to othersubunits are disturbed. Furthermore, mutations ofT295, C304 andQ307, respectively, in TMD 4 ofMalFwere found to result in reduced transport activity.36
Also, a deletion mutant affecting TMD 6 causedcomplete dislocation of MalK.35
In case of MalG our results suggest that TMDs 4–6might participate in contacting MalK. Linker inser-tions at positions A171 (TMD 4) and I220 (TMD 5)(see Figure 3(b)), respectively, caused severe defectsin protein stability.19 Although speculative, thephenotype could be due to insufficient complexassembly and subsequent degradation of MalG. Noother evidence that TMD 6 is involved in MalKbinding has thus far been reported.A note of caution however seems appropriate
concerning the assignments of TMDs and connect-ing loop regions. Although the topological modelsof MalF and MalG (Figure 3(a) and (b)) are based onexperimental data using the phoA fusion techni-que37–39 the transition of the polypeptide chain fromTMD to loop and vice versa cannot be preciselydetermined. Thus, these models do not exclude thatpeptides derived from one end of a transmembranehelix according to Figure 3(a) and (b) are actuallypart of the connecting loop. In fact, alternativetopological models based on several criteria, includ-ing prediction algorithms and mutational data,40
predict TMDs 4 and 6 of MalF to have their carboxy-terminal ends at C300 and L387, respectively.Furthermore, according to this model TMD 4 ofMalG would encompass residues L167 to I184,thereby placing peptides 9 and 10 (see Table 1)entirely into the membrane. It should also be notedthat in two reports using alkaline phosphatasefusions TMD 4 of MalG was differently defined.38,39
Furthermore, our study did not include lengthanalyses of the identified peptides. Thus, it is notunlikely that the minimum length of a peptidedisplaying MalK binding affinity is smaller thanten. Consequently, peptides extending from loopregions into TMDs (see Figure 3(a) and (b)) might
act Sites Between Subunits in a Bacterial ATP-binding Cassette), doi:10.1016/j.jmb.2007.03.043

10 Contact Sites in a Bacterial ABC Transporter
ARTICLE IN PRESS
actually require only loop-derived sequences forbinding. Moreover, only those residues mightactually be involved in binding that lie on the sameface of the helix.Clearly, we will have to await the crystal structure
of the complete MalFGK2 complex to clarify thismatter. In case of the BtuCD complex for whichstructural data are available, predicted lengths andsequences of helical and loop regions in fact differsomewhat from those found in the crystal, therebysupporting the above notion.14
Taking these considerations into account, thecontact sites identified by our study that localize toTMDs might thus be restricted to regions in TMDs 3and 6 of MalF and TMDs 4 and 6 of MalG. In anycase, it is intriguing that transmembrane regions atall display binding activity for MalK. This mightindicate that MalK embeds itself into the membraneduring its interaction with MalFG. Protease studieswith spheroplasts containing the MalFGK2 complexsuggested such a scenario.41 Furthermore, an earlierreport on the protease susceptibility of HisP, theABC protein of the histidine transporter of S.typhimurium, arrived at the same conclusion,42
although it should be noted that the physiologicalsignificance of these results was questioned.43 Inaddition, transmembrane helices that make contactsto MalK might be tilted rather than being orientedperpendicularly to the membrane. The way bywhich the ten transmembrane helices of BtuC arepacked together is at least not in contradiction to thisnotion,14 also variations in helix packing are likely toexist in TMDs from different ABC transporters.In this respect it is also worth mentioning that
heterobifunctional crosslinking of the ABC proteinDrrA to its cognate membrane protein DrrB alsoidentified residues localized to TMDs of DrrB asbeing in close contact to DrrA.44
Putative MalK binding sites in periplasmic loopregions
Our study suggested two regions within peri-plasmic loops P2 (R112-T127) and P4 (L467-F476) ofMalF to interact with MalK. Since the P2-peptidealso displayed significant binding to BSA, a ratherunspecific interaction cannot be ruled out. How-ever, mutants carrying deletions of residues 112 to121 and 114 to 121 in P2, respectively, were found topartially accumulate MalK in the cytoplasm.35
Based on these and other mutations, these authorsspeculated that the large P2 loop might be incontact to MalK. Our data are consistent with thisnotion.A deletion mutant comprising residues 453 to 470
in P4 was defective in maltose transport and alsoaffected MalK localization, leading the authors tosuggest that P4 might be critical for both interactionwith the substrate binding protein MalE andMalK.35
Peptides identified within periplasmic loops ofMalG are part of P1 and P3, respectively. Again,support for the relevance of the region in P1 was
Please cite this article as: Blüschke, B. et al., Mapping Putative Cont(ABC) Transporter by Synthetic Peptide Libraries, J. Mol. Biol. (2007
obtained by linker insertion mutagenesis. In theirstudy, Nelson and Traxler19 found insertions atpositions W84, K90 and A92 (Figure 3(b)) to causeprotein instability. The identified peptide regionwithin the third periplasmic loop P3 was alsocharacterized by insertion mutagenesis (at T244and A246, see Figure 3(b)) and found to be partiallyassembly-deficient.19
Together, these results suggest that the MalKdimer might also contact extracytoplasmic domainsof MalFG. This could be explained by the assump-tion that the loops are not fully solvent-exposed butpartially embedded in the MalFG core. Interestingly,and at least not in contradiction to this notion, MalF-P2 and MalG-P1 are relatively resistant to trypsin inthe resting state but expose a cleavage site in thepresence of ATP. Thus, both loops are likelychanging their conformation during the transportcycle.12
Putative MalK binding sites in cytoplasmic loops
Our study identified peptides from the cytoplas-mically oriented loops C2 and C3 of MalF as well asfrom the N terminus and from loop C2 of MalG,respectively, as being crucial for MalK binding.MalF-C2 was suggested by Tapia et al.35 to partici-pate in the attachment of MalK to the membranebased on the phenotype of a mutant carrying adeletion of residues 312–318. Our data are perfectlyconsistent with this notion although a single pointmutation (Y317Q) exhibited no effect on cell growth(Figure 5(b)).The conserved EAA motif, which is localized to
the C3 and C2 loops, respectively, of MalF andMalGwas demonstrated by crosslinking experiments toreside in close proximity to the helical subdomain ofMalK.10 These data provide independent evidencefor our finding that peptides from both loops areinvolved in MalK binding. Moreover, the datapresented here suggest that residues preceedingthe core motif or at its C-terminal end are critical(Figure 3(a) and (b)). In MalF, especially residues inregion N413–L419 turned out to be sensitive toreplacements, as was the case for the correspondingpeptide W200-P209 in MalG. Significant sequencesimilarity of both peptides likely accounts for thisobservation. Interestingly, substitution of glutamatefor MalF-K416 or MalF-I417 caused different phe-notypes. While the strain carrying K416E grew likewild-type, the I417E mutant displayed a severegrowth defect (Table 3) and the purified variantlacked ATPase activity. Isoleucine is predominantlypresent at this position of the EAA motif in ABCmembrane proteins closely related to the maltosetransporter.45 Only conservative replacements byvaline and leucine are found, which might explainthe observed phenotype. To our knowledge, theresidue has not yet been included in a mutationalanalysis.With MalG-F203 we have identified another
residue from the EAA motif that is crucial forfunction as mutations to either Gln or Asn resulted
act Sites Between Subunits in a Bacterial ATP-binding Cassette), doi:10.1016/j.jmb.2007.03.043

11Contact Sites in a Bacterial ABC Transporter
ARTICLE IN PRESS
in severe growth defects. In related proteins theresidue aligns only with other aromatic residues,although it is not part of the consensus EAA se-quence.45 Since MalK was properly associatedwith the membrane (Figure 7) the residue mightalso be part of a discontinuous binding epitope forMalK. This is in contrast to mutations affectingthe highly conserved A192(→D) and the invariantG196(→P) that caused mislocalization of MalK.13
Here, our data suggest a role in complex assemblyrather than in direct MalK binding, since bindingaffinity of the respective biotinylated peptide toMalK was weak (Figure 2(c), compare peptides 11and 12).Interestingly, peptide C388-D397 derived from
the MalF-C3 loop displayed binding to immobilizedMalK only in the absence of ATP/Mg2+, suggestingthat the site of interaction is recognized when theMalK dimer resides in the open conformation.4 Thisis in perfect agreement with results from a pre-vious crosslinking study demonstrating that homo-bifunctional thiol-specific linkers paired MalF-S403C with A85C of MalK in the absence of ATPonly.10
Effect of ATP
A few biotinylated peptides displayed signifi-cantly weaker binding to MalK in the presence ofATP/Mg2+ (Figure 2). Crystal structures of thesoluble MalK dimer suggested that ATP but notADP binding triggers a transition from an open to aclosed state.4,8 Thus, our findings might imply thatthe closed MalK dimer has less contact sites on theTMDs. Furthermore, all but one of these peptideswere derived from MalG. Although the effect on asingle peptide is difficult to discuss in the absence ofa crystal structure of the complete transporter, thisobservation indicates that the closed MalK dimermight interact differently with MalF and MalG. Forthe EAA motifs of both subunits this has alreadybeen proposed on the basis of mutant analyses13 andcrosslinking experiments.10 Moreover, the structureof the multidrug ABC exporter Sav1866 revealeddifferent contacts between the NBDs and two helicesthat connect the NBDs to the TMDs.46
In conclusion, our data provide detailed insight onsubunit–subunit interactions that might occur dur-ing the transport cycle of an ABC import system andwill set the stage for experiments to further definethe contact sites, e.g. by means of crosslinking andsurface plasmon resonance.
Materials and Methods
Bacterial strains and plasmids
E. coli strain JM109 (Stratagene, La Jolla, USA) wasroutinely used for cloning purposes and as host forplasmid pGS91-1 (malKht).47 E. coli strain ES66 is aderivative of strain ED16913 and carries a malTc mutationthat causes constitutive synthesis of the positive regulator
Please cite this article as: Blüschke, B. et al., Mapping Putative Cont(ABC) Transporter by Synthetic Peptide Libraries, J. Mol. Biol. (2007
of the maltose regulon, MalT (laboratory collection).Plasmid pFS1 carries malE under the control of itschromosomal promoter and malK under control of theptrc promoter on vector pSE380 (laboratory collection).Derivatives of pES67-92 (malFG under the control of the trcpromotor on plasmid pSU1848 carrying single pointmutations) were obtained by site-directed mutagenesisusing Stratagene's Quikchange kit.
Growth experiments
Cells of E. coli strain ES66 carrying the indicatedplasmids were grown in 5 ml of M9 minimal medium49
supplemented with maltose (2%, w/v), ampicillin(0.1 mg/ml), and chloramphenicol (0.02 mg/ml) at 37 °Cunder vigorous agitation.
Protein purification
MalFGK2 (wild-type and variants) and MalE wereprepared as described.15 MalK was either purified as anN-terminal His6 fusion or tag-free as described.15,50
[35S]methionine-labelling of MalK
E. coli strain JM109(pGS91-1)15 (20 ml) was grown inminimal medium E51 supplemented with 0.2% Casa-mino acids (omitting methionine), 2 μg/ml thiamine,0.5% (w/v) glucose, 0.1 mg/ml ampicillin at 37 °C toA650nm=0.5. Cells were harvested by centrifugation for5 min at 5000 g, resuspended in pre-heated freshmedium and incubated with 0.5 mM IPTG at 37 °C for12 min under vigorous shaking. Subsequently, [35S]methionine (250 μCi) was added and incubationcontinued for 5 min. Cells were then cooled on ice for20 min, harvested by centrifugation as above, washedonce with cell lysis buffer (50 mM Tris-HCl (pH 8),150 mM NaCl, 20% (v/v) glycerol, 0.1 mM phenyl-methylsulfonylfluoride) and subsequently disrupted byultrasonication (3× for 1 min with 1 min intervals;output 60%; Branson sonifier). The cytosolic fraction wasthen recovered by centrifugation for 30 min at 12,000gand MalK was purified by Ni-NTA chromatography asdescribed elsewhere.15 The average specific radioactivitywas 5×105 cpm/μg MalK.
Peptide synthesis on cellulose membranes(SPOT synthesis)
Cellulose-bound peptide libraries were prepared bysemi-automatic SPOT synthesis using a SPOT robot(INTAVIS AG, Köln, Germany). SPOT synthesis wascarried out as described in the SPOT synthesis protocol52
and arrays were synthesized on modified Whatman 50cellulose membranes (Whatman, Maidstone, UK). Se-quence files and design of the arrays were generatedwith the in house software LISA. Peptides derived from S.typhimurium MalF (GenBank accession no. CAA38190)and MalG (GenBank accession no. CAA38191) were usedfor pepscan analyses. To this end, two peptide arraysconsisting of 16meric or 31meric peptides, overlapping by15, and 30 amino acid residues, respectively, weresynthesized. Complete substitutional and length analysesof the interacting 16mer-peptides were generated usingthe software LISA and subsequently synthesized asdescribed.20,53
act Sites Between Subunits in a Bacterial ATP-binding Cassette), doi:10.1016/j.jmb.2007.03.043

12 Contact Sites in a Bacterial ABC Transporter
ARTICLE IN PRESS
Screening of cellulose membrane-bound peptidearrays
Before screening, the dried membranes were washed for10 min in ethanol, 3×10 min in TBS (50 mM Tris-HCl (pH8), 137 mM NaCl, 27 mM KCl) and subsequentlyincubated in TBS, supplemented with 5% blocking buffer(Sigma, München, Germany) and 5% (w/v) sucrose, for3 h at room temperature. After washing with TBS, peptidearrays were incubated with 35S-labelled MalK (105 cpm) inblocking buffer, supplemented with 20% glycerol for 6 h atroom temperature with gentle shaking. Unbound MalKwas removed with TBS and peptide-bound MalK wasvisualized and quantified using a phosphoimager andassociated software (Fuji, Japan).
Nano-synthesis of biotinylated peptides
The synthesis of cleavable biotinylated peptides on aCAPE (cellulose-amino-hydroxyl propylether) mem-brane was carried out as described.17 After completionof the synthesis, a spacer consisting of three β-alanineresidues was attached to the N terminus of eachpeptide. Subsequent biotinylation was achieved byusing 0.6 M Fmoc-lysine(biotinyl-ε-aminocaproyl)-OH,previously activated with TBTU and DIPEA. Briefly,Fmoc-lysine-Biotin, TBTU, and DIPEA were mixed inN-methyl-pyrrolidon, incubated for 5 min, and centri-fuged to remove undissolved Fmoc-lysine-Biotin. Thisstep was repeated three times for each peptide. Removalof Fmoc and side-chain deprotection was performed asdescribed.20 Biotinylated peptides were finally releasedby punching out the spots followed by treatment ofeach spot with 0.125 M NaOH in 50% acetonitrile for30 min. Subsequently, the pH was adjusted to 2–4 byadding hydrochloric acid, the peptides were dried ina vacuum centrifuge and then dissolved in 50 mMTris-HCl (pH 7.5), containing 50% (v/v) DMSO to afinal concentration of 1.2 mM. Some peptides werequality-controlled by RP-HPLC and MALDI-TOF massspectrometry.
Peptide binding assay
Binding of soluble biotinylated peptides derived fromMalF and MalG was assayed by incubation with purifiedMalK immobilized in the wells of a microtiter plate(Polysorb™, Nunc, Wiesbaden, Germany). For eachpeptide to be analyzed two wells were incubated overnight with purified MalK (1 μg) in 50 mM Tris-HCl (pH 8),0.1 mM phenylmethylsulfonylfluoride (PMSF), 20% gly-cerol (buffer 1) or with 0.5 μg MalK in buffer 1 containing5 mM MgCl2, 10 mM ATP at 4 °C (final volume: 100 μl).Under these conditions, MalK displayed a specific ATPaseactivity of 5 nmol Pi/min per mg (control value at 37 °C:377 nmol Pi/min per mg; see also Schmees et al.47). Thus,hydrolysis of ATP during the incubation time wasnegligible. Subsequently, the wells were washed threetimes with 50 mM Tris-HCl (pH 7.5), 200 mM NaCl, 10%glycerol (buffer 2). To block unspecific binding to thesurface of the Polysorb surface the wells were thenincubated with 0.5% (w/v) BSA in buffers 1 and 2,respectively, for 30 min at room temperature. A third wellwas also incubated with buffer 1 containing BSA and usedas reference. After three washing steps with buffer 2containing 0.05% (v/v) Tween 20, peptides (12 μM each)were added to wells 1 and 3 in 50 mM Tris-HCl (pH 7.5),
Please cite this article as: Blüschke, B. et al., Mapping Putative Cont(ABC) Transporter by Synthetic Peptide Libraries, J. Mol. Biol. (2007
200 mM NaCl, 1 mM dithiothreitol (DTT), 0.1 mM PMSF,20% glycerol, 0.05% Tween 20 (buffer 3) and to well 2 inbuffer 3 containing 5 mM MgCl2, 10 mM ATP (buffer 4).Incubation took place overnight at 4 °C under gentleshaking. Subsequently, the wells were washed withbuffer 3 and 4, respectively, and bound peptides weredetected enzymatically after reaction with peroxidase-conjugated avidin (1 μg/ml) for 20 min at roomtemperature. After five washing steps per well withbuffers 3 and 4, respectively, substrate solution (100 mMacetate/citrate buffer (pH 6), 8.5 mM H2O2, 5.5 mMtetramethylbenzidine) was added. The reaction wasterminated after 100 s by adding 0.05 M Na2SO4 in 2 MH2SO4 and absorbance readings for each well wererecorded with a microplate reader (SCT Labinstruments,Crailsheim, Germany).
Analytical procedures
Preparation of proteoliposomes, ATPase assay, proteindetermination, SDS-PAGE and immunoblotting wereperformed as described elsewhere.15,21
Acknowledgements
We thank H. Landmesser for excellent technicalassistance. This research was supported by theDeutsche Forschungsgemeinschaft (SFB 449, B14and Z1).
References
1. Holland, E. B., Cole, S., Kuchler, K. & Higgins, C. F.(eds) (2003). ABC Proteins: From Bacteria to Man,Academic Press, New York.
2. Schneider, E. (2003). Import of solutes by ABCtransporters-the maltose and other systems. In ABCProteins: From Bacteria to Man (Holland, E. B., Cole, S.,Kuchler, K. &Higgins, C., eds), pp. 157–185, AcademicPress, New York.
3. Davidson, A. L. & Chen, J. (2004). ATP-bindingcassette transporters in bacteria. Annu. Rev. Biochem.73, 241–268.
4. Chen, J., Lu, G., Lin, J., Davidson, A. L. & Quiocho,F. A. (2003). A tweezers-like motion of the ATP-binding cassette dimer in an ABC transport cycle.Mol.Cell, 12, 651–661.
5. Davidson, A. L., Shuman, H. A. & Nikaido, H. (1992).Mechanism of maltose transport in Escherichia coli:transmembrane signaling by periplasmic bindingproteins. Proc. Natl Acad. Sci. USA, 89, 2360–2364.
6. Chen, J., Sharma, S., Quiocho, F. A. & Davidson, A. L.(2001). Trapping the transition state of an ATP-bindingcassette transporter: evidence for a concerted mechan-ism of maltose transport. Proc. Natl Acad. Sci. USA, 98,1525–1530.
7. Austermühle, M. I., Hall, J. A., Klug, C. S. &Davidson, A. L. (2004). Maltose-binding protein isopen in the catalytic transition sate for ATP hydro-lysis during maltose transport. J. Biol. Chem. 279,28243–28250.
8. Lu, G., Westbrooks, J. M., Davidson, A. L. & Chen, J.(2005). ATP hydrolysis is required to reset the ATP-
act Sites Between Subunits in a Bacterial ATP-binding Cassette), doi:10.1016/j.jmb.2007.03.043

13Contact Sites in a Bacterial ABC Transporter
ARTICLE IN PRESS
binding cassette dimer into the resting-state confor-mation. Proc. Natl Acad. Sci. USA, 102, 17969–17974.
9. Schneider, E., Wilken, S. & Schmid, R. (1994). Nucleo-tide-induced conformational changes of MalK, abacterial ATP binding cassette transporter protein.J. Biol. Chem. 269, 20456–20461.
10. Hunke, S., Mourez, M., Jèhanno, M., Dassa, E. &Schneider, E. (2000). ATP modulates subunit-subunitinteractions in an ATP-binding cassette transporter(MalFGK2) determined by site-directed chemicalcross-linking. J. Biol. Chem. 275, 15526–15534.
11. Mannering, D. E., Sharma, S. & Davidson, A. L. (2001).Demonstration of conformational changes associatedwith activation of the maltose transport complex.J. Biol. Chem. 276, 12362–12368.
12. Daus, M. L., Landmesser, H., Schlosser, A., Müller, P.,Herrmann, A. & Schneider, E. (2006). ATP inducesconformational changes of periplasmic loop regions ofthe maltose ATP-binding cassette transporter. J. Biol.Chem. 281, 3856–3865.
13. Mourez, M., Hofnung, N. & Dassa, E. (1997). Subunitinteractions in ABC transporters: a conservedsequence in hydrophobic membrane proteins ofperiplasmic permease defines an important site ofinteractions with the ATPase subunit. EMBO J. 16,3066–3077.
14. Locher, K. P., Lee, A. T. & Rees, D. C. (2002). The E. coliBtuCD structure: a framework for ABC transportersand mechanism. Science, 296, 1091–1098.
15. Landmesser, H., Stein, A., Blüschke, B., Brinkmann,M., Hunke, S. & Schneider, E. (2002). Large-scalepurification, dissociation and functional reassemblyof the maltose ATP-binding cassette transporter(MalFGK2) of Salmonella typhimurium. Biochim. Bio-phys. Acta, 1565, 64–72.
16. Reineke, U., Volkmer-Engert, R. & Schneider-Mergener, J. (2001). Applications of peptide arraysprepared by the SPOT-technology. Curr. Opin. Bio-technol. 12, 59–64.
17. Licha, K., Bhargava, S., Rheinländer, C., Becker, A.,Schneider-Mergener, J. & Volkmer-Engert, R. (2000).Highly parallel nano-synthesis of cleavable peptide-dye conjugates on cellulose membranes. TetrahedronLetters, 41, 1711–1715.
18. Bhargava, S., Licha, K., Knaute, T., Ebert, B., Becker,A., Grotzinger, C. et al. (2002). A complete substitu-tional analysis of VIP for better tumor imagingproperties. J. Mol. Recogn. 15, 145–153.
19. Nelson, B. D. & Traxler, B. (1998). Exploring the role ofintegral membrane proteins in ATP-binding cassettetransporters: analysis of a collection of MalG insertionmutants. J. Bacteriol. 180, 2507–2514.
20. Wenschuh, H., Volkmer-Engert, R., Schmidt, M.,Schulz, M., Schneider-Mergener, J. & Reineke, U.(2000). Coherent membrane supports for parallelmicrosynthesis and screening of bioactive peptides.Biopolymers (Peptide Sci.), 55, 188–206.
21. Stein, A., Seifert, M., Volkmer-Engert, R., Siepelmeyer,J., Jahreis, K. & Schneider, E. (2002). Functionalcharacterization of the maltose ATP-binding-cassettetransporter of Salmonella typhimurium by means ofmonoclonal antibodies directed against the MalKsubunit. Eur. J. Biochem. 269, 4074–4085.
22. Kamradt, T. & Volkmer-Engert, R. (2004). Cross-reactivity of T lymphocytes in infection and auto-immunity. Mol. Diversity, 8, 271–280.
23. Ay, B., Streitz, M., Boisguerin, P., Schlosser, A., Kern, F.& Volkmer, R. (2007). Sorting and pooling strategy: A
Please cite this article as: Blüschke, B. et al., Mapping Putative Cont(ABC) Transporter by Synthetic Peptide Libraries, J. Mol. Biol. (2007
novel tool to map a virus proteome for CD8 T-cellepitopes. Biopolymers, 88, 64–75.
24. Przezdziak,J.,Tremmel,S.,Kretzschmar,J.,Beyermann,M., Bienert, M. & Volkmer-Engert, R. (2006). Probingthe ligand binding specificity and analyzing thefolding state of SPOT synthesized FBP28 WW domainvariants. ChemBioChem. 7, 780–788.
25. Ay, B., Volkmer, R. & Boisguerin, P. (2007). Synthesisof cleavable peptides with authentic C-termini: anapplication for fully automatic SPOT synthesis. Tetra-hedron Letters, 48, 361–364.
26. Portwich, M., Keller, S., Strauss, H. M., Mahrenholz,C. C., Kretzschmar, I., Kramer, A. & Volkmer, R.(2007). A network of coiled-coil associations derivedfrom synthetic GCN4 leucine-zipper arrays. Angew.Chem. Int. Ed. Engl. 46, 1654–1657.
27. Reineke, U., Kramer, A. & Schneider-Mergener, J.(1999). Antigen sequence- and library-based mappingof linear and discontinous protein-protein-interactionsites by spot synthesis. Curr. Top. Microbiol. Immunol.243, 23–36.
28. Boisguerin, P., Leben, R., Ay, B., Radziwill, G.,Moelling, K., Dong, L. & Volkmer-Engert, R. (2004).An improved method for the synthesis of cellulosemembrane-bound peptides with free C termini isuseful for PDZ domain binding studies. Chem. Biol. 11,449–459.
29. Landgraf, C., Panni, S., Montecchi-Palazzi, L., Castag-noli, L., Schneider-Mergener, J., Volkmer-Engert, R.& Cesareni, G. (2004). Protein interaction networksby proteome peptide scanning. PLoS Biol. 2, E14.
30. Weiser, A. A., Or-Guil, M., Tapia, V., Leichsenring, A.,Schuchhardt, J., Frommel, C. & Volkmer-Engert, R.(2005). SPOT synthesis: reliability of array-basedmeasurement of peptide binding affinity. Anal. Bio-chem. 342, 300–311.
31. Blüschke, B., Volkmer-Engert, R. & Schneider, E.(2006). Topography of the surface of the signaltransducing protein EIIAGlc that interacts with theMalK subunits of the maltose ATP-binding cassettetransporter (MalFGK2) of Salmonella typhimurium.J. Biol. Chem. 281, 12833–12840.
32. Reineke, U., Sabat, R., Volk, H. D. & Schneider-Mergener, J. (1998). Mapping of the interleukin-10/interleukin-10 receptor combining site. Protein Sci. 7,951–960.
33. Reineke, U., Sabat, R., Misselwitz, R., Welfle, H., Volk,H. D. & Schneider-Mergener, J. (1999). A syntheticmimic of a discontinuous binding site on interleukin-10. Nature Biotechnol. 17, 271–275.
34. Ehrmann, M. & Beckwith, J. (1991). Proper inser-tion of a complex membrane protein in the absenceof its amino-terminal export signal. J. Biol. Chem. 266,16530–16533.
35. Tapia, M. I., Mourez, M., Hofnung, M. & Dassa, E.(1999). Structure-function study of MalF protein byrandom mutagenesis. J. Bacteriol. 181, 2267–2272.
36. Steinke, A., Grau, S., Davidson, A., Hofmann, E. &Ehrmann, M. (2001). Characterization of transmem-brane segments 3, 4, and 5 of MalF by mutanionalanalysis. J. Bacteriol. 183, 375–381.
37. Froshauer, S., Green, G. N., Boyd, D., McGovern, K. &Beckwith, J. (1988). Genetic analysis of the membraneinsertion and topology of MalF, a cytoplasmicmembrane protein of Escherichia coli. J. Mol. Biol. 200,501–511.
38. Boyd, A., Traxler, B. & Beckwith, J. (1993). Analysisof the topology of a membrane protein by using a
act Sites Between Subunits in a Bacterial ATP-binding Cassette), doi:10.1016/j.jmb.2007.03.043

14 Contact Sites in a Bacterial ABC Transporter
ARTICLE IN PRESS
minimum number of alkaline phosphatase fusions.J. Bacteriol. 175, 553–556.
39. Dassa, E. & Muir, S. (1993). Membrane topology ofMalG, an inner membrane protein from the maltosetransport system of E. coli. Mol. Microbiol. 7, 29–38.
40. Ehrmann, M., Ehrle, R., Hofmann, E., Boos, W. &Schlösser, A. (1998). The ABC maltose transporter.Mol. Microbiol. 29, 685–694.
41. Schneider, E., Hunke, S. & Tebbe, S. (1995). The MalKprotein of the ATP-binding cassette transporter formaltose of Escherichia coli is accessible to proteasedigestion from the periplasmic side of the membrane.J. Bacteriol. 177, 5364–5367.
42. Baichwal, P. A., Liu, D. X. & Ames, G. F.-L. (1993). TheATP-binding component of a prokaryotic trafficATPase is exposed to the periplasmic (external)surface. Proc. Natl Acad. Sci. USA, 90, 620–624.
43. Blott, E. J., Higgins, C. F. & Linton, K. J. (1999).Cysteine-scanning mutagenesis provides no evidencefor the extracellular accessibility of the nucleotide-binding domains of the multidrug resistance trans-porter P-glycoprotein. EMBO-J. 23, 6800–6808.
44. Kaur, P., Rao, D. K. & Gandlur, S. M. (2005).Biochemical characterization of domains in the mem-brane subunit DrrB that interact with the ABC subunitDrrA: identification of a conservedmotif. Biochemistry,44, 2661–2670.
45. Saurin, W., Köster, W. & Dassa, E. (1994). Bacterialbinding protein-dependent permeases: charateriza-tion of distinctive signatures for functionally relatedintegral cytoplasmic membrane proteins. Mol. Micro-biol. 12, 993–1004.
46. Dawson, R. J. P. & Locher, K. P. (2006). Structure of a
Please cite this article as: Blüschke, B. et al., Mapping Putative Cont(ABC) Transporter by Synthetic Peptide Libraries, J. Mol. Biol. (2007
bacterial multidrug ABC transporter. Nature, 443,180–185.
47. Schmees, G., Stein, A., Hunke, S., Landmesser, H. &Schneider, E. (1999). Functional consequences ofmutations in the conserved ‘signature sequence’ of theATP-binding-cassette protein MalK. Eur. J. Biochem.266, 420–430.
48. Martinez, E., Bartolome, B. & de la Cruz, F. (1988).pACYC 184-derived cloning vectors containing themultiple cloning site and lacZα reporter gene ofpUC8/9 and pUC18/19. Gene, 68, 159–162.
49. Miller, J. H. (1972). Experiments in Molecular Genetics,Cold Spring Harbor Laboratory Press, Cold SpringHarbor, NY.
50. Schneider, E., Linde, M. & Tebbe, S. (1995). Func-tional purification of a bacterial ATP-binding cassettetransporter protein (MalK) from cytoplasmic fractionof an overproducing strain. Protein Expres. Purif. 6,10–14.
51. Roth, J. (1970). Genetic techniques in studies ofbacterial metabolism. Methods Enzymol. 17, 3–35.
52. Wenschuh,H., Gausepohl, H., Germeroth, L., Ulbricht,M., Matuschewski, H., Kramer, A. et al. (2000).Positionally addressable parallel synthesis on contin-uousmembranes. InCombinatorialChemistry:APracticalApproach (Fenniri, H., ed), pp. 95–116, Oxford Uni-versity Press, Oxford.
53. Volkmer-Engert, R., Ehrhard, B., Höhne, W., Kramer,A., Hellwig, J. & Schneider-Mergener, J. (1994).Preparation, analysis and antibody binding studiesof simultaneously synthesized soluble and cellulose-bound HIV-1 p24 peptide epitope libraries. Lett.Peptide Sci. 1, 243–253.
Edited by I. B. Holland
(Received 15 December 2006; received in revised form 12 March 2007; accepted 13 March 2007)
act Sites Between Subunits in a Bacterial ATP-binding Cassette), doi:10.1016/j.jmb.2007.03.043




















