
Manipulation of the Conformation and Enzymatic Properties of T1 Lipase by Site-Directed Mutagenesis...
11
1 23 Applied Biochemistry and Biotechnology Part A: Enzyme Engineering and Biotechnology ISSN 0273-2289 Volume 167 Number 3 Appl Biochem Biotechnol (2012) 167:612-620 DOI 10.1007/s12010-012-9728-2 Manipulation of the Conformation and Enzymatic Properties of T1 Lipase by Site- Directed Mutagenesis of the Protein Core Roswanira Abdul Wahab, Mahiran Basri, Raja Noor Zaliha Raja Abdul Rahman, Abu Bakar Salleh, Mohd Basyaruddin Abdul Rahman, et al.
-
Upload
teknologimalaysia -
Category
Documents
-
view
7 -
download
0
Transcript of Manipulation of the Conformation and Enzymatic Properties of T1 Lipase by Site-Directed Mutagenesis...

1 23
Applied Biochemistry andBiotechnologyPart A: Enzyme Engineering andBiotechnology ISSN 0273-2289Volume 167Number 3 Appl Biochem Biotechnol (2012)167:612-620DOI 10.1007/s12010-012-9728-2
Manipulation of the Conformation andEnzymatic Properties of T1 Lipase by Site-Directed Mutagenesis of the Protein Core
Roswanira Abdul Wahab, MahiranBasri, Raja Noor Zaliha Raja AbdulRahman, Abu Bakar Salleh, MohdBasyaruddin Abdul Rahman, et al.

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer
Science+Business Media, LLC. This e-offprint
is for personal use only and shall not be self-
archived in electronic repositories. If you
wish to self-archive your work, please use the
accepted author’s version for posting to your
own website or your institution’s repository.
You may further deposit the accepted author’s
version on a funder’s repository at a funder’s
request, provided it is not made publicly
available until 12 months after publication.

Manipulation of the Conformation and EnzymaticProperties of T1 Lipase by Site-Directed Mutagenesisof the Protein Core
Roswanira Abdul Wahab & Mahiran Basri &Raja Noor Zaliha Raja Abdul Rahman &
Abu Bakar Salleh &
Mohd Basyaruddin Abdul Rahman & Leow Thean Chor
Received: 12 July 2011 /Accepted: 1 May 2012 /Published online: 12 May 2012# Springer Science+Business Media, LLC 2012
Abstract In silico and experimental investigations were conducted to explore the effects ofsubstituting hydrophobic residues, Val, Met, Leu, Ile, Trp, and Phe into Gln 114 ofT1 lipase. The in silico investigations accurately predicted the enzymatic character-istics of the mutants in the experimental studies and provided rationalization for someof the experimental observations. Substitution with Leu successfully improved theconformational stability and enzymatic characteristics of T1 lipase. However, replacementof Gln114with Trp negatively affected T1 lipase and resulted in the largest disruption of proteinstability, diminished lipase activity and inferior enzymatic characteristics. These resultssuggested that the substitution of a larger residue in a densely packed area of the proteincore can have considerable effects on the structure and function of an enzyme. This is especiallytrue when the residue is next to the catalytic serine as demonstrated with the Phe and Trpmutation.
Appl Biochem Biotechnol (2012) 167:612–620DOI 10.1007/s12010-012-9728-2
R. A. Wahab :M. Basri (*) :M. B. A. RahmanFaculty of Science, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysiae-mail: [email protected]
R. N. Z. R. A. Rahman :A. B. Salleh : L. T. ChorFaculty of Biotechnology and Biomolecular Sciences, Universiti Putra Malaysia, 43400 Serdang,Selangor, Malaysia
M. Basri : R. N. Z. R. A. Rahman :A. B. Salleh : L. T. ChorInstitute of Bioscience, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
R. A. WahabFaculty of Science, Universiti Teknologi Malaysia, 81310 Skudai, Johor, Malaysia
Author's personal copy

Keywords Hydrophobic residue . Hydrophobic core . T1 lipase . Protein design .
Conformational stability
Introduction
Successful protein design should retain both the folding efficiency and functionality of anenzyme [28]. It is clear that protein’s sequence and the interactions between residues in theprotein core are not fully optimized in nature. Instead, organisms achieve the minimumrequirements for efficiency and stability that permit proper functioning in the cell. Thissituation leaves plenty of room for improvement by protein engineering [5, 14]. The majorstabilizing factor in protein folding is generally accepted to be the hydrophobic effect andefficient packing of residues in the protein’s core [21, 24]. The placement of hydrophobicamino acids within the protein is critical for both maintaining a highly ordered structure andas an important driving force for protein folding [1, 9, 11, 27]. Protein stabilities havebeen reported to increase linearly with increasing hydrophobicity of the substitutedresidues [29]. Given that there are many inherent uncertainties associated with proteinfolding, protein prediction software is used to obtain simplified folding pathways thatcan be investigated experimentally to confirm the predictions. Theoretical predictionsof the protein structure and stability invariably help focus the experimental work towardtheoretical protein sequences that may exhibit good functionality and eliminate inefficientmutations.
T1 lipase from Geobacillus zalihae is a good candidate for protein engineering because ithas been well studied and the structure is available. There are 388 residues in T1 lipase, andthe substitution site Gln114 was chosen because it is buried deep within the hydrophobiccore and is in the catalytic pocket near the substrate. In addition, Gln114 and Phe16 werethought to form the oxyanion hole of T1 lipase based on a previous report of BTL2 lipasethat was crystallized in an open conformation. BTL2 lipase shares 98 % similaritywith T1 lipase, and the oxyanion hole is formed by residues Phe17 and Gln115 [3].We hypothesized that the replacement of the hydrophilic Gln with a hydrophobicamino acid may improve both the conformational stability and the enzymatic charac-teristics of T1 lipase. Using this approach, improvement can be achieved in threeways. First, by improving the global protein structure through increasing hydrophobicinteractions, the enzyme can become more compact and stable. Second, increasing thehydrophobic interactions may cause a change in the shape of the catalytic pocket thatoptimized contact with the substrate. Finally, substitution of a residue with a smaller side-chainmay allow a closer interaction between the substrate and the mutated residue, which couldincrease the rate of catalysis.
As yet, there have been no reports on how the mutations of the hydrophobic residuesadjacent to the catalytic serine affect the conformational stability and enzymatic character-istics of T1 lipase or lipases from the closely related group 1.5. The rationale of this study isto demonstrate that the hydrophobicity of the residue at position 114 may serve threepurposes: protein stabilization, optimization of enzymatic activity, and improved hydrolysisof the substrate. In silico mutagenesis was performed to estimate the possible changes withreference to protein densities and the total energy of each mutation. Six variants of T1 lipasewere obtained using site-directed mutagenesis and were evaluated under variablehydrolysis conditions. The conformational stabilities of the proteins were also examined,and each amino acid uniquely affected the stability and enzymatic characteristics of T1lipase.
Appl Biochem Biotechnol (2012) 167:612–620 613
Author's personal copy

Methods
Materials
The growth components were procured from Difco Laboratories (Detroit, MI). The Quick-Change™ site-directed mutagenesis kit was purchased from Stratagene (La Jolla, CA). Agarplates were prepared by addition of tributyrin into Luria-Bertani agar (Oxoid, Basingstoke,UK) and antibiotics, chloramphenicol (35 mg/ml), and ampicillin (50 mg/ml) were suppliedby Amresco (Solon, OH). Corn oil, soy oil, and sunflower oil were acquired from Mazola,Malaysia. Coconut oil was obtained from Parachuta, Malaysia. Rice bran oil was purchasedfrom Pura Harvest, Malaysia. Olive oil was obtained from Bertolli, Italy. The solvents wereof HPLC grade and purchased from Sigma-Aldrich Inc. (St. Louis, MO). The surfactantswere procured from Merck (Darmstadt, Germany). The various metal ions were acquiredfrom Fluka, Buchs, Switzerland. Glycerol (99 %) was obtained from BDH, UK. Deionizedwater was produced in our laboratory.
In Silico Mutation and Estimation of Protein Structural Changes
For the in silico mutation and estimation of the protein stability of the crystal structure ofT1 lipase (PDB code, 2DSN), FoldX software version Beta 3.0 was used. Residueswith bad torsion angles were identified and repaired. Six mutants were prepared bysubstituting Gln114 with Val, Leu, Met, Ile, Trp, and Phe. Possible conformations of thesubstituted amino acids were analyzed and subjected to simple energy minimization toremove higher-energy local minima, which was followed by the calculation of proteinstability.
The protein packing of each lipase variant was further evaluated by estimating the proteindensity and number of cavities with Voronoia 1.0 software [22]. The three-dimensionalstructures of the lipase variants built in FoldX were submitted to the standalone Voronoia 1.0program.
Substitution of Residues by Site-Directed Mutagenesis
The reaction mixture for the PCR site-directed mutation was prepared according to protocolprovided by the manufacturer. The primers were designed so that the mutation pointwas positioned in the center of the primer with approximately 10 to 15 bases that actas a template with the complementary sequence on both sides. The segment DNA wasincubated with the restriction enzyme Dpn 1 at 37 °C for 1 h to digest the parentalmethylated DNA. The Dpn 1-digested DNA was transformed into competent Escher-ichia coli BL21 DE3 pLysS cells, and the tranformants were grown on LB-tributyrinagar media supplemented with antibiotics, ampicillin and chloramphenicol. Each muta-tion was confirmed by sequencing. Glycerol culture stocks were then prepared and keptat −80 °C.
Preparation of Crude Enzymes, Standard Lipase Assay, and Protein Concentration
The crude solutions of the lipase variants and wild-type enzyme were prepared according toprevious work. The standard assay of lipase activity was performed according to the methodof Kwon and Rhee [15]. The protein content was determined according to the method ofBradford [2].
614 Appl Biochem Biotechnol (2012) 167:612–620
Author's personal copy

Lipase Characterization
The experiments for the characterization of the lipase variants were performed in triplicatesunder two separate determinants. The standard deviation was calculated based upon specificactivity. Specific activity is the amount of substrate converted by the lipase per unit of timeper milligram of protein in the sample.
Optimum Temperature The enzymes were evaluated at temperatures ranging from 40 to 80 °Cusing the standard lipase assay.
Effect of Various Oils Lipase assays were performed with different oils to determine ifmutations affected the preference for hydrolysis. Eight oils were tested, corn, sunflower,coconut, soy, rice bran, and olive oil. The standard lipase assay was performed using 1 ml ofculture (30 μl enzyme in 970 μl of 50 mM glycine–NaOH at pH 9.0) combined by shakingwith 2.5 ml of various different natural oils and glycine–NaOH (1:1, v/v) to form anemulsion.
Effect of Organic Solvents The lipase activity of the enzymes in organic solvents withdifferent values of log P was also evaluated. The solvents were added to the enzymes at aratio of 1:3 and incubated for 30 min at 30 °C with agitation at 200 rpm. The standard lipaseassay was then used to measure activity of the enzyme.
Effect of Surfactants The lipase activity of the enzymes was investigated in the presence ofsurfactants (0.1 %, v/v) at 65 °C for 30 min. The surfactants were added directly to thestandard lipase assay.
Effect of Metal Ions The lipase activity of the enzymes was evaluated in the presence ofmetal ions, Li(I), Na(I), K(I), Ca(II), Mg(II), Sr(II), Fe(III), and Zn(II) at 65 °C for 30 min.The metal ions were added directly to the standard lipase at final concentration of 1 mM.
Results and Discussion
In Silico Mutation and Estimation of Protein Structural Changes
FoldX predicted, in descending order, that Met, Leu, Ile, and Val mutants will be more stablethan the wild-type enzyme (Table 1). The lower values of the total energies could becorrelated to the more hydrophobic nature of the substituting residues according to theEngelman-Steitz hydrophobicity scale [8]. However, prediction by Voronoia 1.0 showed thatthe Phe variant will be more stable with a marginally higher packing density than the wild-type lipase, even though the number of cavities in the Phe variant increased by one. Thelower total-energy values predicted by FoldX for some of the variants suggest that amutation at Gln114 could produce variants with improved characteristics.
Lipase Characterization
Optimal Temperature The Val, Met, and Trp variants showed a reduction in the optimumtemperature, while the Ile and Phe variants were similar to the wild-type at 70 °C (Fig. 1).The Leu variant had significantly higher optimum temperature of 80 °C, which could be
Appl Biochem Biotechnol (2012) 167:612–620 615
Author's personal copy
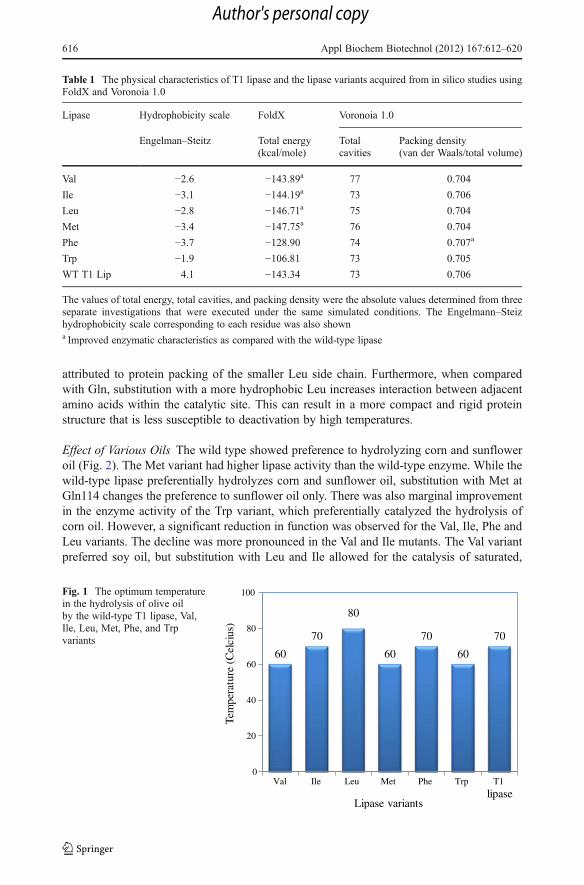
attributed to protein packing of the smaller Leu side chain. Furthermore, when comparedwith Gln, substitution with a more hydrophobic Leu increases interaction between adjacentamino acids within the catalytic site. This can result in a more compact and rigid proteinstructure that is less susceptible to deactivation by high temperatures.
Effect of Various Oils The wild type showed preference to hydrolyzing corn and sunfloweroil (Fig. 2). The Met variant had higher lipase activity than the wild-type enzyme. While thewild-type lipase preferentially hydrolyzes corn and sunflower oil, substitution with Met atGln114 changes the preference to sunflower oil only. There was also marginal improvementin the enzyme activity of the Trp variant, which preferentially catalyzed the hydrolysis ofcorn oil. However, a significant reduction in function was observed for the Val, Ile, Phe andLeu variants. The decline was more pronounced in the Val and Ile mutants. The Val variantpreferred soy oil, but substitution with Leu and Ile allowed for the catalysis of saturated,
Table 1 The physical characteristics of T1 lipase and the lipase variants acquired from in silico studies usingFoldX and Voronoia 1.0
Lipase Hydrophobicity scale FoldX Voronoia 1.0
Engelman–Steitz Total energy(kcal/mole)
Totalcavities
Packing density(van der Waals/total volume)
Val −2.6 −143.89a 77 0.704
Ile −3.1 −144.19a 73 0.706
Leu −2.8 −146.71a 75 0.704
Met −3.4 −147.75a 76 0.704
Phe −3.7 −128.90 74 0.707a
Trp −1.9 −106.81 73 0.705
WT T1 Lip 4.1 −143.34 73 0.706
The values of total energy, total cavities, and packing density were the absolute values determined from threeseparate investigations that were executed under the same simulated conditions. The Engelmann–Steizhydrophobicity scale corresponding to each residue was also showna Improved enzymatic characteristics as compared with the wild-type lipase
60
70
80
60
70
60
70
0
20
40
60
80
100
Val Ile Leu Met Phe Trp T1 lipase
Tem
pera
ture
(C
elci
us)
Lipase variants
Fig. 1 The optimum temperaturein the hydrolysis of olive oilby the wild-type T1 lipase, Val,Ile, Leu, Met, Phe, and Trpvariants
616 Appl Biochem Biotechnol (2012) 167:612–620
Author's personal copy

short-chain coconut oil with high enzyme activity. Conversely, a higher hydrolysis rate wasobserved for sunflower and rice bran oil when Gln114 was mutated to Phe.
Effect of Organic Solvents The log P is a term that is used to demonstrate the solvent-effecton the activity or stability of enzymes. It is defined as the logarithm of the partitioncoefficient in a standard n-octanol/water two phase system [16]. Figure 3 depicts the effectof the solvent (log P) on the activities of the lipase variants and the wild type. Each lipasevariant was found to show a distinctive tolerance profile towards solvents with variable log Pvalues. All variants afforded their highest lipase activity in solvents with log P valuesbetween 2.50 and 8.80 for the exception of Trp. Interestingly, the Leu variant had a profile
0
100
200
300
400
500
Val Ile Met Leu Phe Trp T1
Spec
ific
act
ivity
(U
/mg)
Lipase variants
Corn oil
Sunflower oil
Coconut oil
Soy oil
Rice bran oil
Olive oil
Fig. 2 The various natural oils catalyzed by the wild-type T1 lipase, Val, Ile, Met, Leu, Phe, and the Trp enzymevariants. Each lipase variant was found to show a varied preference in catalyzing the various natural oils
0
50
100
150
200
250
300
Val Ile Leu Met Phe Trp T1 lipase
Spec
ific
act
ivity
(U
/mg)
Lipase variants
DMSO (-1.38)
Acetonitrile (-0.40)
Ethyl acetate (1.8)
Toluene (2.50)
Hexane (3.50)
Isooctane (4.40)
N-tetradecane (7.60)
N-hexadecane (8.8)
Fig. 3 The effect of the various log P of organic solvents on the activity of wild-type T1 lipase, Val, Ile, Met,Leu, Phe, and Trp variants in the hydrolysis of olive oil. Each lipase variant was found to exhibit a distinctivetolerance profile towards the different solvents of variable log P values
Appl Biochem Biotechnol (2012) 167:612–620 617
Author's personal copy

that is opposite from the wild-type lipase with a significant improvement of hydrolysis inethyl acetate and N-hexadecane. The increase stability of the Leu variant toward the water-miscible solvents can be correlated to an increased hydrophobic interaction at the catalyticsite [13]. Similarly, the Met and Phe variants exhibited enhanced activity in the samesolvents as seen in Leu. Substitution with Ile, Phe, or Leu led to an improved activity inN-hexadecane only. However, enzymes with Val and Trp at position 114 showed signifi-cantly reduced activity. Generally, solvents with log P values in the range of 1.5 to 4.0 areseverely toxic because they are preferentially partitioned in the cytoplasmic membrane [4].These solvents also interfere with the intricate balance between the hydrophobic interactions,hydrogen bonds, and van der Waals interactions that contribute to protein folding andnative protein structure [25]. Additionally, enzyme molecules are less susceptible to strip-ping of essential water molecules when exposed to hydrophobic solvents versus hydrophilicones [12].
Effect of Various Surfactants Surfactants are amphipathic molecules with hydrophilic head-group and a hydrophobic tail [26]. The Leu variant had improved protein stability in fourdifferent surfactants (Table 2). The highest activity of Leu variant was recorded in Span 20,and the mutant was significantly more active than the wild-type enzyme. Conversely, theactivity of the Leu mutant in Tween 20 and the Val mutant in SDS was almost completelydiminished. A similar trend of surfactant-induced deactivation was observed for the Met andPhe mutants, but the latter was more susceptible to Tween 20–80 that almost diminishedlipase activity. All variants showed decreased activity in Tween 20–80 except for the enzymewith Leu at position 114. The subtle structural changes in the Leu variant induced by thesurfactants may have increased the adsorption of lipase molecules to the interface andenhanced catalysis [7]. Improved stability in SDS was observed for the Ile, Met and Leuvariants. SDS is more disruptive than Tween 20–80 or Span 20 because it can causeunfolding of the protein’s tertiary structure by binding to both the hydrophobic and hydro-philic residues of the enzyme and interfering with the interactions [18]. This phenomenon isnot seen in Tween 20–80 and Span 20 because their rigid and bulky non-polar heads cannotenter into the water-soluble protein [10, 23].
Table 2 The effect of various metal ions (1 mM) and surfactants (0.1 %, v/v) on characteristics of the wild-type T1 lipase and the lipase variants, Val, Ile, Met, Leu, and Phe in catalyzing the hydrolysis of olive oil
Enzymevariants
Preferredmetal ions
Improved catalysisover wild type
Activity in surfactant Improved catalysisover wild type
Val Sr(II) and Li(I) Nil Tween 80 and Span 20 Tween 80
Ile Sr(II) and Mn(II) Nil SDS SDS
Leu Sr(II) and Mn(II) Sr(II) and Mn(II) Span 20, Tween 40,and Tween 60
Span 20, Tween 40,Tween 60, and SDS
Met Sr(II) and Ca(II) Zn(II) (marginal) Span 20 and Tween 60 SDS
Phe Ca(II) and Sr(II) Nil Span 20 Nil
Trp Mg(II) and Ca(II) Nil Span 20 Nil
T1 lipase Sr(II) and Li(I) – Tween 20, Tween 60,and Span 20
–
The lipase variants showing improved catalysis upon supplementation with certain metal ions and improvedtolerance towards some surfactants were also shown
618 Appl Biochem Biotechnol (2012) 167:612–620
Author's personal copy

Effect of Metal Ions Investigation on the activity of the mutant enzymes revealed that thelipase activity of each variant is activated by a different metal ion as shown in Table 2. Wild-type lipase catalyzed the hydrolysis of olive with the highest activity when supplementedwith Sr(II) ions instead of Ca(II) as previously reported [17]. In general, the mutants of T1lipase were activated by divalent metals, namely, Sr(II), Ca(II), and Mn(II), which agreedwith our earlier results [19]. Interestingly, we found that Sr(II) enhanced hydrolysis in thewild-type enzyme and in the Val, Ile, Met, and Phe mutants. Meanwhile, a general loss ofactivity was observed in reactions supplemented with ions Li(I) and K(I), which could beattributed to the interaction between the side chains of the surface amino acids and thecharged salt ions that could influence the conformation and stability of the variants [4, 20].Additionally, interaction between salt ions and the enzyme surface may have affectedionization of certain amino acids and completely altered enzyme activity [6].
Conclusions
The substitution of Gln with hydrophobic residues improved the protein stability of somemutants and introduced unique enzymatic characteristics in each variant. Substitution of Leuresulted in improvements in three distinct properties of T1 lipase, conformational stability,enzymatic characteristics and enzyme activity. This study demonstrates that the replacementof a Leu residue at Gln114 can improve the protein folding of T1 lipase. However, replacinga residue with a small side chain with a larger amino acid is not recommended in denselypacked areas of the protein core. This is especially true when the residue is near the catalyticserine. This situation was observed in the Phe and Trp mutants, which both resulted inreduced protein stability and inferior enzymatic characteristics. The effect was more pro-nounced in the Trp variant due to a large side chain with restricted rotameric forms thatinterfere with the favorable packing of the protein core of T1 lipase. We believe our findingsmay also be applicable to the protein engineering of other hydrolases with similar folds toachieve improvements in both protein stability and enzymatic properties.
Acknowledgments The authors are indebted to the Ministry of Science, Technology and Innovation forfinancial support (grant no. 5487729). We would like to thank Prof. Dr. Romas Kazlauskas for helpfuldiscussions.
References
1. Anil, B., Sato, S., Cho, J. H., & Raleigh, D. P. (2005). Fine structure analysis of a protein foldingtransition state: distinguishing between hydrophobic stabilization and specific packing. Journal ofMolecular Biology, 354, 693–705.
2. Bradford, M. M. (1976). A rapid and sensitive method for the quantification of microgram quantities ofprotein using the principle of protein-dye binding. Analytical Biochemistry, 72, 248–254.
3. Carrasco-Lopez, C., Godoy, C., de las Rivas, B., Fernandez-Lafuente, G., Palomo, J. M., Guisan, J. M., etal. (2009). Activation of bacterial thermoalkalophilic lipases is spurred by dramatic structural rearrange-ments. The Journal of Biological Chemistry, 284, 4365–4372.
4. Dandavate, V., Jayesh, J., Keharia, H., & Madamwar, D. (2009). Production, partial purification oforganic solvent-tolerant lipase from Burkholderia multivorans V2 and its application for ester synthesis.Bioresource Technology, 100, 3374–3381.
5. Demetrius, L. (2002). Thermodynamics and kinetics of protein folding an evolutionary perspective.Journal of Theoretical Biology, 217, 397–411.
Appl Biochem Biotechnol (2012) 167:612–620 619
Author's personal copy

6. Don, H., Ga, S., Han, S. P., & Cao, S. G. (1999). Purification and characterization of a Pseudomonas sp.Lipase and its properties in non-aqueous media. Biotechnology and Applied Biochemistry, 30, 251–156.
7. Eltaweel, M. A., Rahman, R. N. Z., Salleh, A. B., & Basri, M. (2005). An organic-solvent stable lipasefrom Bacillus sp. strain 42. Analytical Biochemistry, 53, 187–192.
8. Engelman, D. M., Steitz, T. A., & Goldman, A. (1986). Identifying non-polar transbilayer helices inamino acid sequences of membrane proteins. Annual Review of Biophysics and Biophysical Chemistry,15, 321–353.
9. Farber, P. J., & Mittermaier, A. (2008). Side chain burial and hydrophobic core packing in protein foldingtransition states. Protein Science, 4, 644–651.
10. Hermoso, J., Pignolo, D., Kerfelec, B., Crenon, I., Chapus, C., & Fontecilla-Camps, J. (1996). Lipaseactivation by non-ionic detergents. The crystal structure of porcine lipase-colipase-tetraethylene glycolmonooctyl ether complex. Journal of Biological Chemistry, 271, 18007–18016.
11. Hurley, J. H., Baase, W. A., & Matthews, B. W. (1992). Design and structural analysis of alternativehydrophobic core packing rearrangements in bacteriophage T4 lysozyme. Journal of Molecular Biology,244, 1143–1159.
12. Klibanov, A. M. (1997). Why are enzymes less active in organic solvents than in water? Trends inBiotechnology, 15, 77.
13. Klibanov, A. M. (2001). Improving enzymes by using them in organic solvent. Nature, 409, 241–246.14. Kim, D. E., Gu, H., & Baker, D. (1998). The sequences of small proteins are not extensively optimized for
rapid folding by natural selection. Proceedings of the National Academy of Science USA, 95, 4981–4986.15. Kwon, D. K., & Rhee, J. S. (1986). A simple and rapid colorimetric method for determination of free fatty
acids for lipase assay. Journal of American Oil Chemistry Society, 63, 89–92.16. Laane, C., Boeren, S., Hilhorst, R., & Veeger, C. (1987). In C. Laane, J. Tramper, & M. D. Lilly (Eds.),
Optimization of biocatalysis in organic media. Biocatalysis in organic media (pp. 65–84). Amsterdam:Elsevier.
17. Leow, T. C., Rahman, R. N. Z., Basri, M., & Salleh, A. B. (2004). High level expression of thermostablefrom Geobacillus sp. Strain T1. Bioscience. Biotechnology and Biochemistry, 68, 96–103.
18. Leow, T. C. (2005). Ph.D. thesis, Universiti Putra Malaysia, Malaysia.19. Leow, T. C., Rahman, R. N. Z., Basri, M., & Salleh, A. B. (2007). A thermoalkaliphilic lipase of
Geobacillus sp. T1. Extremophiles, 11, 527–535.20. Rahman, R. N. Z., Baharum, S. N., Basri, M., & Salleh, A. B. (2005). High yield purification of an
organic solvent-tolerant lipase from Pseudomonas sp. strain S5. Analytical Biochemistry, 341, 267–274.21. Rose, G. D., Fleming, P. J., Banawar, J. R., & Maritan, A. (2006). A backbone-based theory of protein
folding. Proceedings of the National Academy of Science USA, 45, 16623–16633.22. Rother, K., Hildebrand, P. W., Goede, A., Gruening, B., & Preissner, R. (2009). Voronoia: analyzing
packing in protein structures. Nucleic Acids Reviews, 37, D393–D395.23. Ruiz-Pena, M., Oropesa-Nuriez, R., Pons, T., Louro, S. R., & Perez-Gamatges, A. (2010). Physico-
chemical studies of molecular interactions between non-ionic surfactants and bovine serum albumin.Colloids and Surfaces. B, Biointerfaces, 75, 282–289.
24. Saraboji, K., Michael Gromiha, M., & Ponnuswamy, M. N. (2005). Relative importance of secondarystructure and solvent accessibility to the stability of protein mutants. A case study with amino acidproperties and energetic on T4 and human lysozymes. Computational Biology and Chemistry, 29, 25–35.
25. Tanford, C. (1961). Physical chemistry of macromolecules. New York: Wiley.26. Valstar, A. (2000). Ph.D. thesis, Uppsala University, Sweden.27. Ventura, S., & Serrano, L. (2004). Designing protein from the inside out. Proteins: Structure, Function,
and Bioinformatics, 56, 1–10.28. Ventura, S., Vega, M. C., Lacroix, E., Angrand, I., Spagnolo, L., & Serrano, L. (2002). Conformational
strain in the hydrophobic core and its implications for protein folding and design. Nature StructuralBiology, 9, 485–493.
29. Yutani, K., Ogasahara, K., Tsujita, T., & Yoshinobu, S. (1987). Dependence of conformational stability onhydrophobicity of the amino acid residue in a series of variant proteins substituted at a unique position oftryptophan synthase α subunit. Proceedings of the National Academy of Science USA, 84, 4441–4444.
620 Appl Biochem Biotechnol (2012) 167:612–620
Author's personal copy




















