
Major and minor groups in evolution
51
Major and Minor Groups in Evolution Abstract Kerr and Godfrey-Smith argue that two mathematically equivalent, alternative formal representations drawn from population genetics, the contextualist and collectivist formalisms, may be equally good for quantifying the dynamics of some natural systems, despite important differences between the formalisms. I draw on constraints on causal representation from Woodward (2003) and Scheines and Eberhardt (2006) to argue that one or the other formalism will be superior for arbitrary natural systems in which individuals form different types of groups. Table of contents 0. Introduction 1. Causal and dynamical adequacy 2. Dynamical adequacy 3. Causal interpretability 3.1 Interventions 3.2 Parametric interventions 4. Mechanical adequacy 5. Intervention test vs. near-variant test 6. Dicrocoelium dendriticum forms minor groups 7. Organisms form major groups of sickle-cell gametes/alleles 8. Major and minor groups 9. The contextualist and collectivist formalisms 10. Applying the intervention test 10.1 Case 1: Minor groups modeled with contextualist formalism 10.2 Case 3: Major groups modeled with collectivist formalism 10.3 Case 3: Minor groups modeled with collectivist formalism 10.4 Case 4: Major groups modeled with contextualist formalism
Transcript of Major and minor groups in evolution

Major and Minor Groups in Evolution
Abstract
Kerr and Godfrey-Smith argue that two mathematically equivalent, alternative formal representations drawn from population genetics, the contextualist and collectivist formalisms, may be equally good for quantifying the dynamicsof some natural systems, despite important differences between the formalisms. I draw on constraints on causal representation from Woodward (2003) and Scheines and Eberhardt (2006) to argue that one or the other formalism will be superior for arbitrary natural systems in which individuals form different types of groups.
Table of contents
0. Introduction1. Causal and dynamical adequacy2. Dynamical adequacy3. Causal interpretability3.1 Interventions3.2 Parametric interventions4. Mechanical adequacy5. Intervention test vs. near-variant test6. Dicrocoelium dendriticum forms minor groups7. Organisms form major groups of sickle-cell gametes/alleles8. Major and minor groups9. The contextualist and collectivist formalisms10. Applying the intervention test10.1 Case 1: Minor groups modeled with contextualist formalism10.2 Case 3: Major groups modeled with collectivist formalism10.3 Case 3: Minor groups modeled with collectivist formalism10.4 Case 4: Major groups modeled with contextualist formalism

11. Monism and pluralism with respect to formal representations 12. Conclusion
0. Introduction
In a 2002 work, Kerr and Godfrey-Smith (KGS) argue that two mathematically equivalent, alternative formal representations drawn from population genetics, the contextualist and collectivist formalisms, may be equally good for quantifying the dynamics of natural systems, despite important differences between them (Kerr and Godfrey-Smith 2002). In a recent reconsideration of their views, the same authors consider a handful of different natural systems and argue that one or the other of the formalisms may better represent the causal structure of somenatural systems, though others remain equally well represented by each formalism (Godfrey-Smith and Kerr 2012).Their judgments of the adequacy with which the formalisms represent the causal structure of these systems turn on a test they call the ‘near variant’ test. Below I argue that for systems featuring individuals that form groups, only oneof the two formalisms is causally adequate in the sense of meeting several constraints for causal representation found in Woodward (2003) and Eberhardt and Scheines (Eberhardt and Scheines 2006; see also Glymour, Scheines, and Spirtes 1993; Pearl 2000).
I offer a general criterion for choosing between the contextualist and collectivist formalisms for arbitrary systems in which groups are formed. Groups whose contributions to future generations are caused by their genetic variations via the phenotypes of group members I call minor groups; groups whose contributions are caused by genes, but not via group member phenotypes, I call major groups. (The contrast is specified in a technically careful manner in section 8.) The contextualist formalism is appropriate for natural systems featuring minor groups whilethe collectivist formalism is appropriate for ones featuringmajor groups. I use a test similar to, but importantly
2

different from, KGS’s near variant test to show how the contextualist formalism fails to model major groups in a causally adequate fashion while the collectivist one fails to do so for minor groups.
I begin by discussing key notions of dynamical and causal adequacy, and in particular distinguishing types of interventions about which only causally adequate equations yield reliable information. I then set forth a test, the intervention test, that I later use to exhibit the causal inadequacy of the contextualist formalism for major groups and that of the collectivist one for minor groups. I give examples of each type of group, offer a rigorous specification of what makes them different, distinguish the contextualist and collectivist formalisms, and then deploy the intervention test. In a final section, I consider the ramifications of the results for the philosophical debate between monists and pluralists concerning formal representations.
1. Causal and dynamical adequacy
KGS contend, correctly, that the contextualist and collectivist formalisms can each be used to make inferences about how the same natural systems will behave. Both formalisms are dynamically adequate with respect to the same natural systems. Dynamically adequate equations can be used to make reliable predictive and retrodictive inferences about system dynamics. Two systems of equations that differ may nonetheless both by dynamically adequate with respect tothe same natural system or systems.1
The notion of causal adequacy is stronger than the notion of dynamical adequacy. Causally adequate systems of equations have two important features that systems of equations that are merely dynamically adequate lack. Causally adequate equations are both causally interpretable and mechanically adequate. Note especially that causal
1 Waters characterizes dynamically adequate equations as empirically sufficient for bookkeeping (Waters 2005, 315). Keeping good books is different, writes Waters, from correctly representing causal structures.
3

interpretability is only one component of causal adequacy. Next, I set out two simple causal systems, pictured in Figure 1, and then use them to illustrate the notions of dynamical adequacy, causal interpretability, and mechanical adequacy.
Consider the graphs below of two different systems (discussion here follows Woodward (2003, §2.3):
Figure 1: Two different natural systems featuring X as a cause of Y
In each graph, directed edges represent causal dependencies among causal variable nodes. For the system graphed on the left, the following equations are causally adequate (and hence dynamically adequate, too):
(1)
For the system graphed on the right, the following equationsare causally (and hence dynamically) adequate:
(2)
2. Dynamical adequacy
If we let a + bc = f · g , then both equations (1) and (2) are dynamically adequate with respect to both the natural systems pictured in Figure 1; each system of equations
4

yields the same value for Y given the same values for X. Equations are dynamically adequate for some system if, givenvalues for the right-hand side variables, the equations giveyou the correct values for the left-hand side ones. Note hownatural systems may vary in terms of causal structure, as the two natural systems pictured in Figure 1 do, but may nevertheless be modeled in a dynamically adequate fashion using the same systems of equations. Thus, a single system of equations may be dynamically adequate with respect to multiple natural systems, and multiple systems of equations may be dynamically adequate with respect to a single naturalsystem.
3. Causal interpretability
Causal interpretability is one component of causal adequacy.Causally interpretable systems of equations are presented with causes on the right-hand side and effects on the left. They may be distinguished from their non-causally interpretable counterparts by their additional implications.Only causally interpretable equations yield reliable information about the consequences of interventions.
For a single natural system, not all dynamically adequate systems of equations will also be causally interpretable. For the systems pictured in Figure 1, equations (1) are causally interpretable when matched with the left-hand side system while equations (2) are causally interpretable when matched with the right-hand side system, but not vice-versa.
3.1 Interventions
A system of equations is causally interpretable with respectto a given natural system provided it yields reliable information about the consequences of interventions. Interventions, thus, deserve some special discussion here. Here is Woodward’s definition of intervention, where I is anintervention variable:
5

I1. I causes X.I2. I acts as a switch for all the other variablesthat cause X. That is, certain values of I are such that when I attains those values, X ceases todepend on the values of other variables that causeX and instead depends only on the value taken by I.I3. Any directed path from I to Y goes through X. That is, I does not directly cause Y and is not a cause of any causes of Y that are distinct from X except, of course, for those causes, of Y, if any,that are built into the I-X-Y connection itself; that is, except for (a) any causes of Y that are effects of X (i.e. variables that are causally between X and Y) and (b) any causes of Y that are between I and X and have no effect on Y independently of X. I4. I is (statistically) independent of any variable Z that causes Y and that is on a directedpath that does not go through X. (Woodward 2003, 98)
It is perhaps worth emphasizing that intervention variables are such only with respect to a causal relationship: one uses intervention variable I to intervene upon causal variable X with respect to effect variable Y.
Woodward’s definition of intervention picks out structuralor hard interventions by virtue of including condition I2. Causes that meet all of Woodward’s conditions but I2 are softor parametric interventions. We consider specific instances ofboth sorts of interventions as part of the intervention test.
When active, or “on,” a structural intervention, say an intervention on Z, can be represented by eliminating any equation from a causally interpretable set of equations in which the target variable, Z, appears on the left, e.g., Z =fX, while inserting into the set of equations Z = I, where I is the (structural) intervention variable. In cases where there is no equation for Z (Z is exogenous), no equation is
6

eliminated; a new one is merely added to the system of equations. This eliminate-and-replace procedure is how structural interventions are modeled for a given system of equations that is causally interpretable. When the procedureis performed on causally interpretable systems of equations,the result is a new system of equations that is dynamically adequate with respect to the natural system in its post-intervention state.
Consider again equations (2) above, the ones that are causally interpretable with respect to the system on the right-hand side of Figure 1. Set values for variables as follows: f = 1, g = 2. With these values set, we can infer how the system will behave for various values of X: e.g., when X = 1, Y = 2. Because the equations are causally interpretable, we can also infer how the system will respondto an intervention on Z, such as the one contemplated in theprevious paragraph. The new system of equations (2*) can be used to infer how the system behaves post-intervention:
(2*)
Notice how, post-intervention, the system behaves differently: the value of X no longer matters to system dynamics, and the value of Y is determined by the value of anew variable, I, a variable that did not previously influence system dynamics. Given a value for the intervention variable I, e.g., I = 3, we can infer how the system will behave post-intervention using equations (2*): we know that Y will take value g • I = 6. Equations (6*) are thus dynamically adequate for the natural system post-intervention: they tell us the truth about Y given values for the right-hand side variables in (2*).
Consider now equations (1). If we carefully set the values of the variables in equations (5), we can generate a dynamically adequate system of equations for the natural system pictured on the right-hand side of Figure 1. Note that here we are mismatching the equations and the natural system: equations (1) are causally inadequate for the system
7

on the right-hand side of Figure 1. But, if we set values inequations (5) as follows: a = 1, b = 1, and c = 1, we infer the same value of Y from the same values of X as we do usingequations (2), at least in the absence of interventions. This illustrates how different systems of equations can be dynamically adequate for the same systems.
Notice, however, that equations (1) are causally inadequate for the natural system on the right-hand side of Figure 1 because they do not yield reliable information about the consequences of interventions. Suppose we intervene on the natural system as we did earlier, by changing the value of Z to 3. As we saw from the manipulating the causally adequate equations (2) by means ofthe eliminate-and-replace procedure, the contemplated intervention makes Y = 6. If we model an intervention on Z in equations (1) by means of the eliminate-and-replace procedure, the result is equations (1*)
(1*)
Notice that equations (1*) are dynamically inadequate for the system post-intervention. We know that Y takes value 6 when Z is set to 3 by means of an intervention, but if we infer Y from by means of equations (1*) we get Y = 4.This shows that, while equations (1) are dynamically adequate for the system on the right-hand side of Figure 1, they are causally inadequate for the same system. They do not yield reliable information about the consequences of interventions, where the measure for yielding such reliable information is this: the equations cannot be altered by means of the eliminate and replace procedure used for structural interventions such that the resulting system of equations is dynamically adequate for the intervened-upon system post-intervention.
To restore dynamical adequacy to equations (1) post-intervention, we could perform some operation other than theeliminate-and-replace procedure we use for structural interventions. We could modify the values of multiple right-
8

hand side variables in (1). Given that the intervention fixes Z at value 3 we would have to set X = 3, too, in orderto infer the right value for Y. This is important. Equations(1) can be modified such that they are dynamically adequate for the system on the right-hand side of Figure 1 after a structural intervention on Z. But the equations have to be modified in the wrong sort of way. The procedure by which we have to modify equations (1) such that they are dynamically adequate for the right-hand side system from Figure 1 post-intervention is not the procedure used to model interventions for causally interpretable equations. In particular, the necessary alterations to equations (1) fail to meet Woodward’s condition I3. The intervention is represented as changing the value to multiple right-hand side variables, while interventions are defined as changing only a single right-hand side quantity. That causally inadequate systems of equations cannot capture structural interventions as changes to the value of single causal variables is a feature of such equations that is used below in the intervention test to exhibit their causal inadequacy.
3.2 Parametric interventions
To run the intervention test below, we also need a definition of parametric intervention. Woodward’s definitionof intervention above picks out only a subset of interventions, hard or structural interventions. Causes that meet all of Woodward’s conditions but I2 are soft or parametric interventions. While hard interventions fix the value of a causal variable and eliminate all other influences upon it, soft interventions are added causal influences that leave other causal influences on the target variable intact. Critically, just as only causally interpretable equations yield reliable information about the consequences of hard interventions, only causally interpretable equations yield reliable information about the consequences of soft interventions.
We will be especially concerned below with parametric interventions on suppressed intermediaries. In directed
9

causal graphs, each edge in a causal graph represents the operation of a distinct causal mechanism. Such mechanisms may be, and indeed nearly always are, themselves complex, featuring causal relationships among variables not represented in the graph. The operation of such mechanisms is represented by coefficients that serve as weights on edges linking causes with their effects. For instance, friction coefficients quantify complicated interactions between two surfaces (see Woodward 2002). Interventions on the surfaces made, for instance, by sanding them, can be effectively quantified by altering the value of the frictioncoefficient in a physical model of the system.
To illustrate parametric interventions at greater depth,consider again the systems pictured in Figure 1. Suppose that W is an intermediary between X and Z for the right-hand side natural system in Figure 1. We can make this intermediary explicit by adding a node in the graph and a corresponding equation to the causally adequate set (2) to yield equations (3) below:
(3)
Suppose now we intervene upon the system by means of a parametric intervention P on W, producing the system illustrated in Figure 2.
Figure 2. A system equivalent to the one pictured on the right of Figure 1 but with a parametric intervention P on previously suppressed intermediary W
10

Associated with parametric interventions is a procedure for modifying causally interpretable systems of equations. As with structural interventions, we eliminate any equation in which W appears on the left-hand side and we replace it with another equation, where W is a function of the intervention variable (P in this case). The inserted equation is different, however, in the case of parametric equations. Instead of being a simple equation where the target variable is fixed by an intervention variable and nothing more, the inserted equation makes the target of the intervention a function of the intervention variable along with its other causes. Different sorts of parametric interventions can be modeled by different functions; we willbe concerned with cases in which the target variable is a multiplicative function of its previous cause and the intervention variable. Thus, our eliminate and replace procedure generates equations (3#) in the case at hand:
Y=gZZ=WW=fX·P (3#)
Equations (3#) are dynamically adequate for the natural system in Figure 2 after the parametric intervention P on W. Suppose, for instance, that X = 1 and the parametric intervention is set at P = 2 while variables are otherwise unchanged from the earlier discussion. We infer that Y = 4 for the intervened-upon system.
While equations (3#) correctly capture the impact of a parametric intervention upon W, an alternative way to capture such an intervention is simply to modify the value of the coefficient f in equations (2). Starting with equations (2), rather than first making the intermediary W explicit in the equations and then altering the new system of equations by means of the eliminate and replace procedurejust considered, we could start with equations (2) and quantify the impact of a parametric intervention on W by simply changing the value of the coefficient on the X Z
11

path. In particular, equations (3⌘) below quantify the impact of a parametric intervention on W provided that we set f* = f • P:
(3⌘)
Equations (3⌘) will be dynamically adequate for the naturalsystem pictured on the right-hand side of Figure 1 in the wake of a parametric intervention P on W. Thus, capturing the parametric intervention P on suppressed intermediary W can be most straightforwardly accomplished simply by changing the value of f to Pf. Generally, parametric interventions like P on suppressed intermediaries can be captured as changes to the values of coefficients that weight the causal mechanism that includes the suppressed causal variable that is the target of the intervention.
Consider now equations (1), which are causally inadequate for the system on the right-hand side of Figure 1. If we make the parametric intervention to the natural system just considered, one whereby we alter W without severing its connection to its previous causes, equations (1) will misrepresent the consequences of doing so when we use the procedure just discussed for altering equations in response to parametric interventions on suppressed intermediaries. Consider what happens if we make explicit the intermediary W on the XZ path by adding an equation for W to equations (1), and then let W be set by a new function that makes W a product of its pre-intervention cause and the parametric intervention variable P. Our new system of equations, then, will be these:
Y=aX+cZZ=WW=bX·P (1#)
Notice how, given P = 2, we do not infer the same value for Y that we inferred using the causally adequate equations (3#) and (3⌘). In particular, given P = 2 and X = 1, and
12

leaving the variables otherwise unchanged (i.e., a = b = c =1), we get Y = 3. Note, further, that we get this same result if we use the simpler procedure for quantifying parametric interventions on suppressed intermediaries, the procedure whereby we alter the coefficient along the manipulated causal path. Equations (1) misrepresent system dynamics if we replace b with b*, where b* = Pb.
As before, equations (1) can be modified such that dynamical adequacy is restored post-intervention, but the character of the necessary modifications exhibits the causalinadequacy of (1). In particular, if change both b to Pb anda to Pa, we thereby generate a causally adequate set of equations for the post-intervention system. However, this sort of procedure to restore dynamical adequacy is not the sort procedure used to quantify parametric interventions on suppressed intermediaries. That equations (1) can be alteredsuch that they are dynamical adequate post-intervention onlyby means of changing the values of multiple coefficients, or, equivalently, by eliminating and replacing multiple equations, is indicative of the causal inadequacy of the equations under consideration. This fact about causally inadequate equations is used in the intervention test below to detect equations causally inadequacy.
Mechanical adequacy
As noted earlier, the notion of causal adequacy has two components: causally adequate systems of equations must be both causally interpretable and mechanically adequate. To illustrate how systems of equations may fail to be mechanically adequate, consider how equation (4) might be used to model each of the natural systems from Figure 1:
Y = hX. (4)
(4) will be dynamically adequate provided we set values for variables in this manner: h = a + bc = f · g.
(4) fails to be mechanically adequate by dint of failingto represent the causal mechanisms that connect X to Y for
13

each natural system pictured in Figure 1. It would be a mistake to use the equation (4) to model either of the systems graphed in Figure 1 in the place of equations (1) or(2), which feature equations for Z. By featuring equations for Z, (1) and (2) make clear how many mechanisms connect X to Y. Making mechanisms explicit is valuable since the resulting distinct representations for the two systems issuein different sets of commitments about the consequences of combinations of interventions, and, accordingly, issue in different commitments about what explains the dynamics of the natural systems.
When an effect is produced by means of multiple mechanisms, that is, by means of multiple causal paths, a formal representation is a mechanically adequate representation of the causes that produce the effect only ifit includes equations that separately quantify the operationof each mechanism, at least provided that variables along the distinct causal paths are among the measured or known causes in the system.
By virtue of yielding information about the consequencesof interventions, causally adequate systems of equations canbe used to answer what Woodward dubs w questions, what-if-things-had-been-different questions. The potential for using causally adequate equations about the consequences of one or more interventions is, Woodward argues, what makes the equations explanatory (Woodward 2003, §5.1). Explanation, finally, is a basic goal of science, making causally adequate equations superior to their alternatives.
5. Intervention test vs. near-variant test
I seek to adjudicate whether formal representations that arecandidates for serving as causally adequate representations of natural systems really are such. I determine this by considering how the representations must be adjusted to remain dynamically adequate when hypothetical interventions are made upon natural systems for which they are dynamicallyadequate. If, to restore dynamical adequacy in the wake of an intervention on the natural system, a set of equations
14

must be manipulated in a way different from those formal manipulations that are appropriate for interventions, then the formalism is not causally adequate. As we just saw, a system of equations is causally inadequate if, to restore dynamical adequacy after a structural intervention, multiple values for right-hand side causal variables must be altered.Equally, a system of equations is causally inadequate if, torestore dynamical adequacy after a parametric intervention ona suppressed intermediary, multiple values for right-hand side coefficients must be altered. Thus, consideration how dynamical adequacy must be restored to a system of equationspost-intervention provides a means to detect causal inadequacy.
The test procedure just set forth, what I will henceforth call the intervention test, can be usefully contrastedwith the near variant test used by KGS. KGS are equally concerned with the restoration of dynamical adequacy in the face of changes to population genetics equations. But several important differences between KGS’s near variant test and my own intervention test account for differences inresults when the tests are run for the same systems.
Chief among the differences between the tests is the strength of the test procedures. KGS write that their near variant test will not always adjudicate between competing formalisms for given systems [2012, 10]. This is because they measure how many variables must be altered in a formal representation such that it governs a near variant of a natural system, and different formalisms may require the same number of changes from an original. In contrast, failing the intervention test is decisive: as we will see below, the intervention test rules that the contextualist formalism is causally inadequate for major groups while the collectivist formalism is causally inadequate for minor groups.
The tests vary in other ways. First, only some transformations that KGS consider in order to generate near variants are also the product of interventions. Second, KGS consider formal representations of natural systems that failto meet criteria for causal representation other than
15

criteria having to do with interventions. In particular, they consider systems featuring fitness coefficients that must replaced by fitness functions for the equations even tobe candidates for causal adequacy. The intervention test I deploy can only be used to distinguish between causally adequate and causally inadequate formal representations whenthe tested representations have met all the criteria for causal adequacy besides, possibly, the one used to run the test, that of capturing interventions as such. Accordingly, below, I sometimes specify functions to replace the coefficients used by KGS when we consider the same natural systems in order to generate a mechanically adequate set of equations.
As the discussion in the previous sections made clear, the intervention test is justified by contentions about causality that are characteristic of a specific approach to understanding that notion, one defended by a host of writersworking within the same general approach to causal modeling(Glymour, Scheines, and Spirtes 1993; Pearl 2000; Woodward 2003; Eberhardt and Scheines 2006). At the very least, the intervention test can be used to assess whether some formalism is causally adequate given the approach. In contrast, KGS’ near-variant test lacks a similar basis.
KGS write that their near-variant analysis is “related to a family of recent approaches to causal questions that emphasize manipulation and intervention, and also to some non-interventionist counterfactual approaches”; they cite works on causation by Pearl (2000), Woodward (2003), and Lewis (2000). Beyond this, KGS make no effort to ground their test in any particular approach to causation. They exhibit their test, illustrate its application, and make explicit a number of its assumptions, but they do not provide evidence that the results of the test yield facts about causal structures on any specific interpretation of what “cause” means. Moreover, Pearl and Woodward, on the onehand, and Lewis, on the other, take substantially different stances on just this issue. Woodward is clear that Lewis’ rules for evaluating causation generate mistaken evaluationsof causal claims in cases that Woodward’s rules handle
16

correctly (2003, 137). No single test could be justified by its relationship to competing theories of causation, since these will issue in different commitments concerning what causal claims are true of the systems over which the test isapplied.
In sum, the near-variant test and the intervention test are not on equal footing. The intervention test is based upon Woodward’s definition of structural intervention and Scheines and Eberhardt’s definition of parametric intervention. A formalism that fails the intervention test is not causally adequate when “cause” is understood in the sense of “cause” used by these authors. The near-variant test has no similar justification.
I turn now to consider the contrast between major and minor groups. I set forth an example of each first, and thenset forth the contrast in formal terms. Having done so, I distinguish between the contextualist and collectivist interpretations, set up some expectations for the results ofthe intervention test, and then run it. I then address the implications of the results for the debate over whether a plurality of formal representations may be each maximally adequate for a single system.
6. Dicrocoelium dendriticum forms minor groups
The behavior of the infamous D. dendriticum parasite is a compelling example of altruism in nature. At one stage of their lifecycle, the fluke parasites infect an ant host. Of the fifty odd parasites within the host, a single individualmigrates to the brain of the ant and causes it to spend timeon the tips of grass blades, where it is more likely to be eaten by livestock, the parasite’s next host. The parasite that infects the ant brain dies, while the rest survive to colonize the ungulate.
The case of the D. dendriticum parasite is often raised in discussions of multi-level selection and group selection(e.g., Sober and Wilson 1998; Sterelny 2001; Kerr and Godfrey-Smith 2002). Take the parasites within a single ant to form a group. Imagine a genetic variation among such
17

parasites that determines whether or not their bearer forms a brainworm provided no other group member does. Call such individuals altruists. The genetic variation that determinesaltruism has a clear influence on the performance of all themembers of a group, that is, all the parasites within a single ant: if no parasites within a group are altruists, then group members are less likely to make it to the next lifecycle stage. Accordingly, genes determine how groups of different types, ones featuring different numbers of altruists, perform better of worse.
Note how, in the D. dendriticum case, the influence of altruistic alleles within groups upon group contributions tofuture generations is mediated by the phenotypes of the parasites within the groups, specifically the behavior of the parasites: the genetic variations determine whether or not a fluke navigates to the brain of the ant to manipulate the ant’s behavior, which in turn causally influences next-generation fluke frequencies. According to the distinction made above, then, the groups formed by collections of parasites within distinct ant hosts are minor groups.
7. Organisms form major groups of sickle-cell gametes/alleles
Consider now an allele that controls some phenotypic featureof an organism. Let’s consider another classic case, that ofthe sickle-cell allele. Among humans who live in malarial environments, heterozygotes for the sickle-cell allele have higher fitness than does either type of homozygote. Homozygotes for the sickle-cell allele suffer from sickle-cell disease, while homozygotes for the non-sickle-cell allele are more vulnerable to malaria than heterozygotes are.2
2 The reality of sickle-cell disease and the causal influence of sickle-cell alleles is much more complex than the sketch here would suggest, but consideration of a simplified version of the sickle-cell case has become standard in arguments over formal representations (see Sober and Lewontin 1982; Sober 1987; Maynard Smith 1987a, 1987b; Kitcher and Sterelny 1988; Kitcher, Sterelny, and Waters 1990; Lloyd 2005; Lloyd et al. 2005; Waters 2005)).
18

Conceive of the human organism as a group made up of two gametes, a fused sperm and egg. (One can also think of it as a group of two alleles at the sickle-cell locus.) Onceagain, the genetic variations at the sickle-cell locus have a clear causal influence on the performance of members of the group: the alleles carried by the gametes will contribute more or fewer copies to subsequent generations depending upon their influence on organism viability. But, in this case, the genetic variations do not make the organisms more or less viable by controlling some or anotherfeature of the behavior (or more generally the phenotype) ofthe gametes. Equally, insofar as alleles may impact their future frequencies by means other than through their effectson gamete or organism phenotypes, as occurs in cases of meiotic drive, the same independence holds. The developmental pathway by which sickle-cell genes affect the viability of humans features neither gamete phenotypes nor (manipulation of) meiosis as an intermediary. Sickle-cell alleles affect the development of red blood cells, and theseare not formed from gamete phenotypes but are produced by anentirely separate developmental process, making organisms major groups of sickle-cell gametes and alleles.3
It is a little strange, maybe, to think of organisms asgroups of gametes or alleles, perhaps because they the groups’ constituents cease to exist upon group formation. The idea that diploids can be treated as groups of alleles is not a novel proposal; it goes back to Wilson (1975; see also Kerr and Godfrey-Smith 2002; Okasha 2006; Godfrey-Smithand Kerr 2012). What really matters is that organisms are clearly formed from gametes and the alleles they bear; persistence of those group members post-formation is irrelevant to the major/minor group distinction proposed here. That distinction is causal in character and depends onwhether genetic variations carried by group members affect group phenotypes by means of member phenotypes. Even if we decide that some or all of what are here called major and
3 Other genes may affect organism phenotypes by means of gamete phenotypes; in these cases, the organisms are not major groups of gametes differentiated by such genetic variations.
19

minor groups are not really groups after all but something else, the major/minor contrast will still apply. Moreover, the distinction will have the implications it is said to have: the dynamics of major groups-or-whatever will quantified in a causally adequate fashion by means of the collectivist formalism while minor groups-or-whatever will be quantified in a causally adequate fashion by the contextualist formalism.
8. Major and minor groups
The major/minor distinction is between two types of groups or collectives, which are formed from individuals or particles.4 Wecan define variables over collectives and distinguish between major and minor groups in terms of how these variables are causally related. Collectives contribute particles of different types to future generations. Let the number of (arbitrarily designated) A type particles contributed to future generations be represented by C, a property of groups that ranges over the natural numbers (we will consider only cases in which there are two types of particles, A and B types). C is caused by G, some natural number of alleles of some arbitrary type (e.g., altruistic alleles) contained within the collective. Collectives vary in phenotype as well, where Pc is a dummy coded variable that ranges over the natural numbers and picks out discrete phenotypic properties of collectives. The phenotypic properties of collectives may be emergent insofar as they may not be specifiable as a statistical function of group composition. Lastly, collectives may vary in the number of particles they contain featuring a given particle phenotype,Pp; for instance, particles may vary in their expression of,or disposition toward, altruistic behavior, and collectives may vary with respect to the count or frequency of particleswith a given degree of altruism.
Among minor groups, G causes C by means of Pc, where theGPc path features at least one particle phenotype intermediary, e.g., GPpPcC. (Henceforth I will 4 The collective/particle contrast is due to Okasha (2006).
20

concentrate on cases in which there is a single GPc path with a single intermediary particle phenotype Pp.) By contrast, among major groups, G causes C by means of at least one path, GPcC, where Pp does not lie on the path for any particle phenotype. Among major groups, either G does not cause Pp no matter what phenotypic characteristic of the particles is examined, or, if G does cause Pp, then Pp does not lie on the GPcC path. Major groups may feature multiple paths from G to C, and in particular may feature the path GPpC. But among major groups, there is no path connecting G and C featuring both Pp and Pc. (See Figures 3-6 below.)
Because genes affect major group phenotypes without doing so by means of particle phenotypes, one can intervene upon the mechanism connecting genes to collective phenotypeswithout intervening upon particles. Equally, among major groups, interventions on particle phenotypes will not alter collective phenotypes. In contrast, among minor groups, one can manipulate collective phenotypes by means of interveningupon particle phenotypes.
In the above definitions, I mean to use the term phenotype in a precise fashion. Particle phenotypes must be manipulable by means of interventions on particles while collective phenotypes must be manipulable by means of interventions on collectives. Moreover, being part of a collective with one or another phenotype is ruled out as a particle phenotype and features of genetic variations, including their number, frequency, and patterns of methylation or activation, are also ruled out as phenotypes of either particles or collectives (this runs contrary to standard usage). I rule these things out because the major/minor distinction has to do with how genes affect particles and collectives, rather than their locations or other features.
Figure 3: Causal relationships within a minor group
21
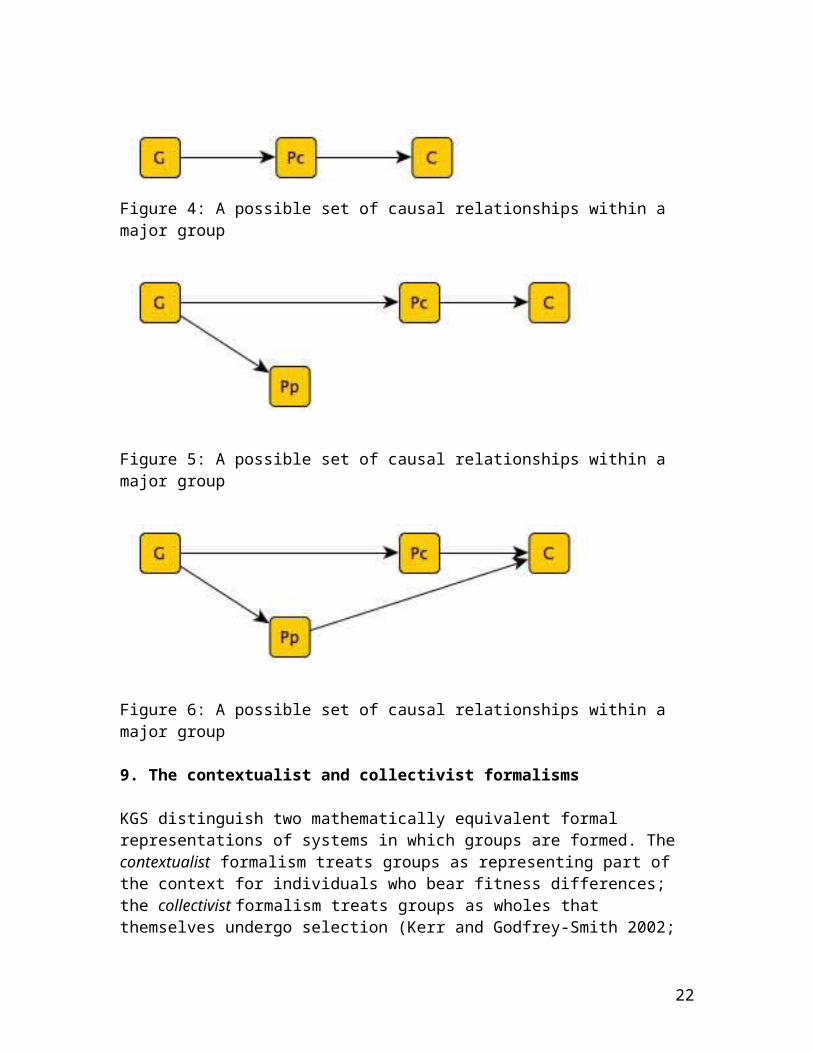
Figure 4: A possible set of causal relationships within a major group
Figure 5: A possible set of causal relationships within a major group
Figure 6: A possible set of causal relationships within a major group
9. The contextualist and collectivist formalisms
KGS distinguish two mathematically equivalent formal representations of systems in which groups are formed. The contextualist formalism treats groups as representing part of the context for individuals who bear fitness differences; the collectivist formalism treats groups as wholes that themselves undergo selection (Kerr and Godfrey-Smith 2002;
22

2012). Both the formalisms are Wright-Fisher discrete generation classical population genetics models in which fitness differences quantify the effects of genetic variations.
Here is the collectivist formalism, in which the next-generation frequencies p̄(t+1) and q̄(t+1) , of two types of individual, A and B, are calculated this way (Kerr and Godfrey-Smith 2002, 483-484; I have pulled the average fitness parameter to the right):
(5)
where fi(t) represents the frequency of a group with i A types, the πi are group specific fitness variables representing group productivity, and the Φi represent how much group productivity is claimed by A types (for the definition of the Φ parameter, see Kerr and Godfrey-Smith(2012, 3)).
Here is the contextualist formalism representation (Kerr and Godfrey-Smith 2002, 483-484; I have pulled the average fitness parameter to the right):
(6)
23

where n represents the total number of A and B type individuals in a group, while the α and β parameters represent the fitnesses of A and B types.
KGS set out a translation scheme for moving between the collectivist and contextualist representations (2002, 482):
(7)
The possibility of making translations across the formalismsby means of the above equations undergirds KGS’s assertion of mathematical equivalency between the formalisms.
Though the collectivist and contextualist formalisms aredynamically adequate with respect to the same systems, when causally interpreted, the two formalisms entail different commitments concerning how genes affect next-generation particle frequencies. In each formalism, fitnesses quantify how genetically variant individuals contribute to next-generation particle production. In the contextualist formalisms, fitness parameters, αi and βi, weight particle frequencies. A causal interpretation of the formalism implies that those fitnesses quantify the operation of causal mechanisms by which genetically variant particles affect next-generation particle frequencies. Genes make a difference to particle features and these then matter to next-generation particle frequencies in a manner quantified by fitnesses. By contrast, in the collectivist formalism, fitness parameters quantifying differential genetic effects,πi, weight collective frequencies. A causal interpretation of the formalism implies that those fitnesses quantify the operation of causal mechanisms by which genetically variant collectives affect next-generation particle frequencies. Genes make a difference to collective features and these then matter to next-generation particle frequencies in a
24

manner quantified by fitnesses.5
10. Applying the intervention test
In what follows, we consider natural systems featuring either major or minor groups, subject to either interventions on collectives or particles, and modeled by either collectivist or contextualist formalisms. The idea isto use the intervention test to show that collectivist systems of equations are appropriate for systems with major groups while contextualist ones are appropriate for systems featuring minor groups because the formalisms are causally adequate when paired with natural systems in this last fashion, but not vice-versa. I use the D. dendriticum and sickle-cell cases as definite scenarios for consideration. In the course of discussing several of the cases, I compare the results of the intervention test with the results of KGS’s near-variant test and explain whatever incongruities arise.
Prior to running these test cases, it is possible to setup some expectations on the basis of the commitments undertaken above, ones against which the outcomes of the tests can be measured. Recall from section 3 above that one sign that some system of equations is not causally adequate is this: the eliminate and replace procedure used to quantify structural interventions cannot be performed upon the system of equations such that the result of the procedure is a system of equations that is dynamically adequate for the intervened-upon natural system post-intervention. Briefly, if you have to change the value of more than one right-hand side causal variable to quantify
5 It worthwhile to note, too, that in the collectivist formalism, the f(i)t parameters take particle frequencies as inputs. They must do so forthe equations to have a recursive structure. Thus, not only do fitness parameters quantifying differential genetic effects appear weighting collective frequencies, they do so in equations featuring particle frequencies that are not weighted by fitness parameters. Even though particle frequencies could be weighted by fitness parameters, they are not, clearly implying that the differential genetic effects are effects on collectives rather than effects on particles.
25

the impact of an intervention, then the equations are not causally adequate for the intervened-upon natural system. Similarly, if, in the face of a parametric intervention on asuppressed intermediary like P above, you must change the value of more than a single right-hand side coefficient to quantify the intervention’s impact, then the equations are not causally adequate.
I claim that the contextualist formalism is causally adequate for natural systems in which minor groups are formed while the collectivist one is causally adequate for natural systems in which major groups are formed, but not vice-versa. So, we expect to be able to expose the causal inadequacy of each formalism for its mismatched natural system by considering interventions upon the natural system and then showing that these interventions cannot be quantified by means of changes to single right-hand side variables. In the four sub-section that follow, I show how two sorts of interventions, a structural intervention on brainworms’ destruction and a parametric intervention on human health deficits caused by malaria, are quantifiable aschanges to the values of single variables in causally adequate formalisms but not in causally inadequate ones.
10.1 Case 1: Minor groups modeled with contextualist formalism
I claim that the contextualist formalism is causally adequate for minor groups. Accordingly, restoring dynamical adequacy after an intervention upon particle phenotypes thatare caused by genes at the brainworm locus among D. dendriticum collectives should require changing the value of asingle quantity in the contextualist formalism. To run our test, we need a definite system of equations that meets test-irrelevant conditions for causal adequacy for a system.To generate such a system of equations, we begin with KGS’s discussion of a D. dendriticum type system.
KGS’ system features only six individuals per group (per ant), in which an altruist dies in each group in which there is one, and any group with an altruist contributes to
26

the next generation while groups without altruists contribute at a rate that is only a tenth that of groups with altruists (Kerr and Godfrey-Smith 2002, 503). They propose these fitness functions to set the values of the α and β parameters in the contextualist formalism (recall i isthe number of A types, altruists, in the group):
(8)
Because the above functions do not separate out the several mechanisms by which altruistic genes affect their future frequencies—the equations are mechanically inadequate—we must expand upon them to generate a causally adequate set. The fitnesses take the values they do because altruists benefit all group members, harm themselves, and can count ona base rate of contribution that all groups enjoy. We can state functions for the α and β parameters using the familiar breakdown that separates out the base rate (0.1), the benefit conferred by altruists (b), and the cost of altruism (c). b quantifies the influence that altruists haveover the brains of their hosts, and c quantifies how flukes invading the ant brain are destroyed by the mastication of ungulates:
(9)
We can use equations (6) and (9) together to evaluate the causal adequacy of the contextualist formalism for the D dendriticum system using the intervention test.
27

The intervention we consider is a structural intervention whereby we alter the system so that brainworms survive ungulate ingestion. Clearly, the destruction of the brainworms is a known and measured cause of system dynamics:altruistic alleles cause the death of those of their bearersthat form brainworms; they earn the moniker “altruistic” as a result. Causally adequate equations should have variable that represents this cause. That, or the variable should be suppressed intermediary along some causal mechanism whose operation is quantified by a coefficient in the equations.
The contextualist representation of the system, equations (6) together with equations (9), features a variable, c, that quantifies brainworm destruction. The proposed intervention can be quantified by means of the eliminate-and-replace procedure used for structural interventions by adding an equation to equations (9) that sets c = I (c was exogenous before, so we need not eliminateany equation in (9) setting its value). Post-intervention, the value of c is wholly determined by intervention variableI. By setting I = 0, and consequently c, too, we effectively destroy the cost associated with altruism and thereby allow altruists to survive to contribute descendants to the next generation, even if they form brainworms. Equations (9*) aredynamically adequate for the fluke case when the altruists from groups with an even number of altruists and nonaltruists (i =3), have been intervened upon such c is setto 0 by means of a structural intervention:
αi=(0.1+bi)−1i ci
βi=0.1+bibi¹0=0.9b0=0ci¹3=1ci=3=II=0 (9*)
The contextualist formalism, when rendered into a
28

mechanically adequate and hence causally adequate form for the D. dendriticum system, passes the intervention test when weconsider intervening on brainworm destruction.
Consider a specific system in which i = 32 and n = 64. In its pre-intervention state, the system will evolve such that i = 26 after a single generation, as can be inferred from (6) and (9). If we intervene upon the system in the manner just contemplated by setting c = 0, we change its dynamics such that i = 28 after one generation, as can be inferred from (6) and (9*).
10.2 Case 3: Major groups modeled with collectivist formalism
This is our second well-matched case. I allege that the collectivist formalism is causally adequate for systems featuring major groups, so restoring dynamical adequacy in response to interventions on group phenotypes should requirechanging the values of single fitness values in the collectivist formalism when that formalism is used for majorgroups. This time, we consider a parametric intervention on a suppressed intermediary.
Modeling simple cases of viability selection on diploid organisms is done using the genotypic selection model, whichis an instance of the collectivist formalism. Set the f(i)t parameters by the Hardy-Weinberg rule using p and q for normal and sickle-cell gametes respectively; set the πi parameters by fitnesses for each type of zygote, wD, wH, wR;and set the ϕ parameter quantify fair meiosis, such that ittakes value 1 in each homozygote and ½ for heterozygotes. The result is the familiar genotypic selection model:
(10)
29

In the systems modeled by the genotypic selection model, fitness coefficients sit as weights on edges connecting pre-selection frequencies for each of normal homozygotes, heterozygotes, and sickle-cell homozygotes (d, h, r, respectively) to post-selection frequencies for these (D, H,R). A full graph of a diploid system can be found in Appendix 1; here we concentrate on a portion of that graph featuring fitness variables serving as coefficients:
Figure 7 Causal mechanisms connecting pre-selection to post-selection frequencies of different zygotes in the sickle-cell case
For the intervention test, we consider parametric interventions upon health deficits suffered due to malaria infection (H). Because genotype affects the extent to whichindividuals suffer such health deficits, and the health deficits are in turn is relevant to survival to maturity, His a suppressde intermediary on the paths from initial frequency to mature frequencies for genetically variant diploid humans.
We might consider two sorts of interventions on H. A structural intervention on H would entirely determine the
30

value of that variable. For instance, a technique that ensured that individuals homozygote for the normal allele never suffered from any health deficits from malaria would count as a structural intervention on Hd. Distribution of a full-proof anti-malaria drug (G) to every homozygous individual with a 100% compliance rate would constitute a structural intervention on Hd. Such an intervention would destroy any causal relationship between genotype at the sickle-cell locus on health deficits suffered as a result ofmalaria infection (see Figure 8).
Figure 8: a structural intervention G on Hd
By contrast, an intervention that affected whether individuals suffered health deficits as a result of malaria infection, but did not entirely determine the value of H, would serve as a parametric intervention on that variable. As an example of a parametric intervention of this last sort, consider bednet distribution, B.
Distributing bednets to individuals in malaria-stricken regions affects health deficits suffered as a result of malaria. However, bednet distribution is not a structural intervention; it does not wholly determine whether individuals suffer health deficits from malaria infection. This is at least because bednet usage rates stand at roughly60% (Cohen and Dupas 2008, 16). Thus, even though bednet distribution affects the rates at which individuals suffer health deficits from malaria, it remains that genotype continues to play a role in determining the value of Hd, too. There are some individuals who do not properly hang
31

bednets, despite receiving them, and are infected with malaria as a result, and, for at least those individuals, their genotype will affect the health deficits they suffer owing to their infection.
We can graph the influence of the parametric bednet intervention on the fitness of dominant homozygotes by replacing the dD path in Figure 7 above, where the health deficient intermediary (Hd) is suppressed, with the new graph below:
Figure 9: A parametric intervention, B, on an intermediary, Hd, between d and D
Notice how, in the case of a parametric intervention, unlikethat of a structural intervention, the intervened-upon variable continues to be affected by the causes that affected it prior to the intervention.
Suppose that, in the collectivist formalism, we set values for zygote fitness in the pre-intervention state thisway: wD = 0.6, wH = 0.9, wR = 0.5. The system thus exhibits the predictable stable polymorphism typical for cases of heterozygote superiority, as is detectable by setting
in (10) and solving for p = 0.57.Recall how restoring dynamical adequacy in the face of a
parametric intervention may be accomplished most straightforwardly by altering the coefficient weighing the edge containing the suppressed variable that is intervened-upon, in this case w. To quantify the bednet intervention, we let wD = wD*, where wD* = wD B. A realistic pair of values for B is 1, in the case bednets are not distributed,
32

and 1.26 in the case that they are. This way, the bednets increase to 1 the fitness of 60% of those who receive them, thereby eliminating the detrimental effects of malaria for those humans entirely. We can use equations (10*) below to infer a new equilibrium for the system post-intervention:
(10*)
As a result of the intervention that enhances dominant homozygote fitness, the stable frequency of the normal allele shifts from p = 0.57 to p = 0.74.
Restoring dynamical adequacy in response to an intervention on the sickle-cell system requires a change in the value of a single quantity, and this is characteristic of causally adequate formalisms. The collectivist formalism passes the intervention test for the sickle cell case when we consider manipulating human health by means of bednet distribution.
10.3 Case 3: Minor groups modeled with collectivist formalism
The D. dendriticum system can be captured in a dynamically adequate fashion using the collectivist formalism. This
33

follows straightaway from the fact that it can be modeled inthe dynamically adequate fashion by the contextualist formalism, together with the equations (7) above, which set equivalencies between the variables from the different formalisms. Provided we set values for variables as mandatedby (7), the result is a dynamically adequate collectivist version of the fluke case considered earlier, in the absenceof interventions:
(11)
It remains to consider whether these equations are causally adequate; we expect that they are not.
Let us consider again the intervention whereby the system is altered such that the altruists who invade the antbrain are no longer destroyed as a result. The causal influence of altruist destruction is not quantified
34

explicitly in the collectivist formalism as it is, by c, in the contextualist formalism. This absence from the collectivist formalism of a parameter quantifying a known cause might be thought to indicate immediately that the collectivist formalism is causally inadequate. However, brainworm destruction might be quantified as a suppressed intermediary in the collectivist formalism, as health consequences of malaria are in the sickle-cell case. Thus, we must check whether, following an intervention on the brainworms’ destruction within a given group, dynamical adequacy can be restored to the collectivist formalism through the alteration of a single fitness variable quantifying genetic effects at the altruism locus in the collectivist formalism. In the collectivist formalism, the intervention of brainworm destruction might be quantified asa parametric intervention on a suppressed intermediary.
Considering a particular group type, one featuring an even split between altruists and non-altruists, we interveneupon brainworm destruction as we did earlier, and alter the system such that altruistic flukes in that group survive even if they form brainworms. To restore dynamical adequacy to the collectivist formalism post-intervention, we would have to alter both the π3 and ϕ3 variables, indicating causal inadequacy. To see this result, note that the intervention has two consequences: it affects the overall number of particles produced by the group, quantified by π3,and it affects the proportion of altruists produced, quantified by ϕ3.
Consider the π3 and ϕ3 variables in turn. The π3 variablequantifies the number of next-generation particles produced by the group with three altruists. The π3 variable must be altered post-intervention because the intervention causes anextra individual to survive and produce descendants. Restoring dynamical adequacy post-intervention would requireincreasing the value of π3 from 5 to 6 to adjust for this extra survivor. Consider now the ϕ3 variable. Recall that this variable quantifies the proportion of altruists among the particles contributed to the next-generation. To restoredynamical adequacy post-intervention, this quantity, too,
35

must be increased, from 0.4 to ½, because the extra particles contributed by the new survivor always bear altruistic genes.
We saw earlier that the result of the intervention on brainworm destruction using the contextualist formalism. In a single generation in the intervened-upon population, a population with 32 altruists evolved into one featuring 28 altruists. We can get the collectivist equations to reproduce this result for the post-intervention case, but only if we alter π3 by increasing it from 5 to 6 and altering ϕ3 by increasing it to 0.4 to ½. There is no way toreproduce the result by altering only one of these variables: if we try to quantify the intervention by manipulating only the collectivist fitness variable π3, by increasing it from 5 to 7.5 (the most plausible option), we get the right value in the numerator for , but the wrong value in the denominator.
Intervening on brainworm destruction alters both the number of flukes and the proportion of flukes who survive ungulate ingestion, and hence requires altering two quantities in the collectivist formalism to restore dynamical adequacy post-intervention. That two quantities must be changed to restore dynamical adequacy in response toan intervention indicates the causal inadequacy of the formalism. Brainworm destruction is not a suppressed intermediary between initial group frequency and next-generation particle production quantified by a fitness variable, π3, in the collectivist formalism. Instead, the collectivist formalism is simply causally inadequate for theD. dendriticum system.
The conclusions reached in my consideration of the flukecase run counter to what KGS conclude from their (2012) consideration of the trait-group model, of which the D. dendriticum model considered above is an instance. KGS claim that the collectivist and the contextualist formalisms are on roughly equal footing for a trait-group model [2012, 10].The reasons they reach a different conclusion from the one reached here are discussed in Appendix 2.
36

10.4 Case 4: Major groups modeled with contextualist formalism
Officially, we expect the contextualist formalism to be causally inadequate for the sickle-cell case, since the contextualist formalism is appropriate for minor groups, andhuman organisms form major groups of gametes/alleles. Consider the contextualist formalism in dynamically adequateform for the sickle-cell case. To generate it, we use the values used earlier for the collectivist formalism and inputthem into equations (7), yielding these values for contextualist variables:
Letting the f(i)t parameters be set, once again, by the Hardy-Weinberg rule, and setting values for frequencies of particles in group types at either 1 or ½, we get these dynamically adequate contextualist equations for the sickle-cell case in the absence of interventions:
(12)
Note especially how different fitness variables, α1 and β1, quantify genetic effects on particles that occur together inheterozygotes.
Since major groups are such that genes do not affect collectives by means of genetic effects on particles, it
37

should be possible to expose the inadequacy of the contextualist formalism for major groups by considering interventions that alter how genes affect next-generation particle frequencies. If the contextualist formalism is correct for major groups, then interventions on what genes do should be quantifiable as changes to the values of singlefitness variables, since these quantify differential geneticeffects on next-generation particle frequencies. But if we do alter change how genes exert their differential effects, for instance by means of the parametric bednet intervention considered earlier, we find that we cannot quantify the intervention by means of changes to the values of single fitness variables in equations (12).
In particular, restoring dynamical adequacy after bednetdistribution to heterozygotes would require changing two quantities, both and α1 and β1. Since even heterozygotes forthe sickle-cell allele suffer somewhat from malaria infection, they would profit from one or another sort of anti-malarial intervention, including bednet distribution (we might also think up more direct and brutal ways of altering heterozygote fitness). Restoring dynamical adequacyto equations (12) in response to an intervention altering how genes affect heterozygotes’ next-generation gamete production, as the bednet intervention does, would require changing both the α1 and β1 coefficients when the contextualist formalism is used, exhibiting the formalism’s causally inadequacy.
To see this point, note that heterozygotes produce two sorts of gametes, ones bearing sickle-cell alleles and ones bearing normal alleles. Thus, an increase in the productive capacity of heterozygotes, one that results from increased survival due to bednet distribution, will result in an increase the number of gametes/alleles of both sorts. In thecontextualist formalism, only the α1 and β1 coefficients, quantifying the relative viability of the two types of particles, could capture the influence of bednet provision on system dynamics.6 Restoring dynamical adequacy after the 6 The f(i)t parameters quantify group formation rates: they are set by the Hardy-Weinberg equation for the gamete-to-zygote lifecycle
38

bednet intervention requires increasing the value of both the α1 and β1 coefficients, because extra particles of both sorts are produced.
To examine the case in greater detail, recall that priorto the bednet intervention, heterozygote fitness is 0.9, reflecting relatively minor health deficits due to malaria, while normal homozygote fitness is 0.6 and sickle-cell homozygotes have fitness 0.5. Assuming heterozygotes are no more or less disposed to use bednets than homozygotes are, bednet distribution should raise to 1 the fitness of 60% of the heretozygotes. Thus, we model the parametric intervention by replacing heterozygote fitness wH with wH*, where wH* = wH • B, and B = . If calculate the stable frequency in the manipulated system using the collectivist equations, we get p = 0.56. Reproducing this result by modifying the contextualist equations is impossible unless we alter both the α1 and β1 coefficients. In particular, we must boost each from 0.45 to 0.48. Altering only one of these quantities fails to reproduce the correct stable frequency, since altering just one of α1 and β1 means takingan increase in heterozygote viability to be favorable to only some of the gametes/alleles that heterozygotes produce.We might try increasing, say, α1 to 0.51, since doing so would ensure that average fitness came out right, but it would leave the numerator of the equation for too high,while leaving the numerator for too low.
Only bumping up both α1 and β1 will restore dynamical adequacy to the collectivist equations. Restoring dynamical adequacy post-intervention cannot be achieved by means of formal techniques for quantifying either structural interventions or parametric interventions on suppressed intermediaries if we model the sickle-cell case using the contextualist formalism, exhibiting the causal inadequacy ofthe formalism.
A final way to expose the inadequacy of the contextualist formalism for major groups is to causally interpret the formalism and then use the formalism as a
transition. The i and n parameters are just counts of population members.
39

guide to what sorts of interventions it ought to be possibleto make. In the test just considered, we showed how an intervention that altered what genes do in the sickle-cell case cannot quantified as an intervention in the contextualist formalism. In this next test, we begin by causally interpreting the contextualist formalism. Specifically, we interpret the α1 and β1 coefficients as quantifying genetic effects on particles, thereby licensing the expectation that it should be possible to intervene on particles to alter next-generation particle frequencies. In particular, it should be possible to intervene upon how alleles manipulate meiosis or how they affect gamete phenotypes, given a causal interpretation of the contextualist formalism.
There is, however, no intervention that fits the bill for the case of diploid selection. There is no way we can intervene upon the particles such that the intervention is quantifiable by a change to just one of the α1 or β1 parameters. No intervention on gametes and no intervention on how alleles manipulate meiosis may be quantified as a change to either of the α1 and β1 parameters alone. We consider these in turn.
Interventions on the process of meiosis cannot be quantified by either the α1 or β1 parameter alone. If we introduce meiotic drive into a system in which selection occurs among organisms, we must alter both α1 and β1 (see appendix 3). Interventions that alter how particles affect meiosis are not quantifiable as such using the contextualistformalism.
Consider now the case of gametic selection occurring prior to zygote formation, such as occurs within systems with free-swimming gametes. An intervention that altered theprocess of gametic selection cannot be quantified in the contextualist formalism by means of the α and β parameters. The result here is even more dramatic.
Gametic selection prior to zygote formation requires the introduction of fitness weights on p and q in the haplo-diplont model used for such cases (Scudo 1967; see appendix 1). This is because genetic effects on gametes prior to
40

zygote formation make a difference to zygote frequencies. Since zygote frequencies are not a function of the α and β parameters, but are instead fixed by the f(i)t parameters in the contextualist formation, the α and β parameters cannot be used to quantify interventions on gametes that would affect their viability prior to zygote formation. KGS seem to overlook the impact of gametic selection on zygote frequencies in their discussion of this sort of case. They write:
Perturbations to the viability of one gamete type (e.g., α1) do not logically imply changes to the viability of the other (β1). Consequently a contextual parameterization will be superior” ((Godfrey-Smith and Kerr 2012, 212).
While it is true that perturbations to the viability of one gamete won’t affect the fitness of the other, such perturbations will matter to zygote frequencies, that is, what is quantified by the f(i)t parameters in the contextualist model. As a result, no change in the value of either the α1 and β1 parameter alone (or even both) in the contextualist model can quantify gametic selection. The contextualist model is causally inadequate for the gametic selection case not because gametic selection requires changing β1 parameter along with α1, but because it requires changes to the f)i)t parameters along with changes to fitnesses (see Appendix 1 for further discussion of gametic selection prior to zygote formation). 7
If we causally interpret the contextualist model, we license the expectation that particles may be intervened-upon such that the intervention is quantifiable by means of some change to one of the α and β parameters. However, no such interventions are possible. The contextualist formalismcannot be used to causally model populations of diploids
7 See Lloyd (2005, 306) for the dynamical insufficiency of contextualistmodels for cases where particles are sorted into genotypes non-randomly,as occurs as a result of gametic selection; she credits the point to Lewontin.
41

whenever genes affect particle phenotypes as well as zygote phenotypes.
11. Monism and pluralism with respect to formal representations
The contentions of this work are relevant to debate over howmany formal representations in population genetics form maximally adequate formal representations of systems in which groups are formed. In that debate, monists contend that, for at least some systems, a single formal representation is best because it represents the causal facts correctly. Lisa Lloyd and her co-authors, for instance, claim that the collectivist formalism is best for modeling diploid dynamics, as in the sickle-cell case (Lloyd2005; Lloyd et al. 2005; Lloyd, Lewontin, and Feldman 2008).By contrast to monists, pluralists contend that several formal representations may be equally good for a single system; genic pluralists contend that among these there willalways be a maximally adequate formal representation that attributes causal efficacy to genic properties (Kitcher and Sterelny 1988; Kitcher, Sterelny, and Waters 1990).8 The position taken here is closest to the view that Waters calls“multilevel monism,” the position he attributes, guardedly, to Lloyd (Waters 2005, 313). I am best construed as a multi-level monist because I contend that there is a single maximally adequate formal representation that may either be contextualist or collectivist, depending on whether major and minor groups are formed in the modeled population.
It is important to note, however, that the reasons I give for preferring the contextualist formalism for minor groups and the collectivist one for major groups have nothing to do with whether either sort of system exhibits selection at the group level. Suppose we assess whether or
8 Waters (2005, §2) does an excellent job distinguishing the various positions in the monism vs. pluralism debate; he disavows genic pluralism as an appropriate label for his (2005) view, though he acknowledges that “all selective episodes (or, perhaps, almost all) can be interpreted in terms of genic selection” (2005, 314).
42

not there is selection at level x by judging whether or not entities at the level x have features that are causally relevant to system dynamics. To follows Waters’ (2005) use of Sober’s (1984) language, there is selection at level x provided there is selection for, and not just selection of, characteristics of x-level entities. If we assess whether a system exhibits selection at the group level in these terms,then both major and minor groups undergo group selection. This is true by definition: both major and minor groups are defined as groups in which group-level properties cause next-generation particle frequencies. The contrast between major and minor groups, and the attendant implications aboutwhat formalism is best, have to do with how group-level causally efficacious properties are caused by genes, in particular, whether the connection occurs by means of particle phenotypes or not.
12. Conclusion
By distinguishing between major and minor groups, I seek chiefly to establish a connection between natural systems and formal population genetics models: minor groups may be modeled in a causally adequate fashion by the contextualist formalism and major groups by the collectivist one. But I hope to put the major/minor group distinction to work elsewhere. For instance, Dictostylium discoideum, the infamous slime mold, forms aggregates at a critical point in its lifecycle, and it remains unclear whether these should be deemed distinct individuals for the purposes of evolutionarymodeling. From the point of view of this work, we know that if we find out that Dictostylium discoideum aggregates form major groups, then these must be formally represented, with their own frequency terms and associated fitness variables, in population genetics models that have explanatory purport.Moreover, at least some hymenopteran colonies may be treated as major groups of female/male organism pairings, with sterile workers forming part of the collective phenotype of such groups, much like somatic cells in organisms form part of the phenotype of gamete pairs when these are treated as
43

groups. Thus, the major/minor group distinction has the potential to help distinguish explanatory models of non-standard systems in which groups are formed.
Lastly, the evolution of major groups may prove important to the study of evolutionary history because it represents the shedding of an important engineering limitation upon group-level adaptations. Minor groups will only feature adaptations that can be constructed from the raw materials of group member phenotypes, but in systems in which major groups are formed, genes may impact group development, and group adaptedness, directly. That means greater flexibility in terms of what causal effects genetic variations can have, because genetic variations need not have positive (or at least non-disastrous) effects on the development of particle phenotypes to positively affect collective performance among major groups. Indeed, many organisms carry many genes that have no effect on gametes ormeiotic division. At least one of what Maynard Smith and Szathmáry (1995) highlight as the major transitions in evolution, the transition to multicellularity, involves the creation of major groups.
Appendix 1
Below are equations appropriate for a haplo-diplont model(Scudo 1967), suitable for a system featuring genetic effects on viability at both the gamete and the organism lifecycle stages, along with coefficient k quantifying meiosis. The haplo-diplont model contains three separate sets parameters, v’s, w’s, and k, which quantify differential genetic effects on gametes, zygotes, and meiosis, respectively. The genotypic selection model is a determination of the haplo-diplont model in which v1 and v2 are set to the same value and k = ½.
Pre-selection A1 gametes = pPre-selection A2 gametes = q = (1 – p)Post-selection A1 gametes = P = Post-selection A2 gametes = Q = Average gamete fitness = V =
44

Mature A1 gametes = P’ =
Mature A2 gametes = Q’ =
Pre-selection A1A1 organisms = d = (P’)2
Pre-selection A1A2 organisms = h = (Q’)2
Pre-selection A2A2 organisms = r = 2P’Q‘Post-selection A1A2 organisms = D = Post-selection A1A2 organisms = H = Post-selection A2A2 organisms = R =
Average organism fitness = W =
Mature A1A2 organisms = D’ =
Mature A1A2 organisms = H’ =
Mature A2A2 organisms = R’ =
Next-generation A1 gametes = p’ = D’ + k’Next-generation A2 gametes = q’ = R’ + kH’
Reduced to a pair of equations, the haplo-diplont model is this:
p̄(t+1)=wD (v1p )2+wH2v1pv2qkwD (v1p )2+wH2v1pv2qk+wR (v2q)2
q̄(t+1)=wH2v1pv2q(1−k )+wR (v2q)2
wD (v1p )2+wH2v1pv2q+wR (v2q)2 (13)
Here is a causal graph for this sort of system:
45
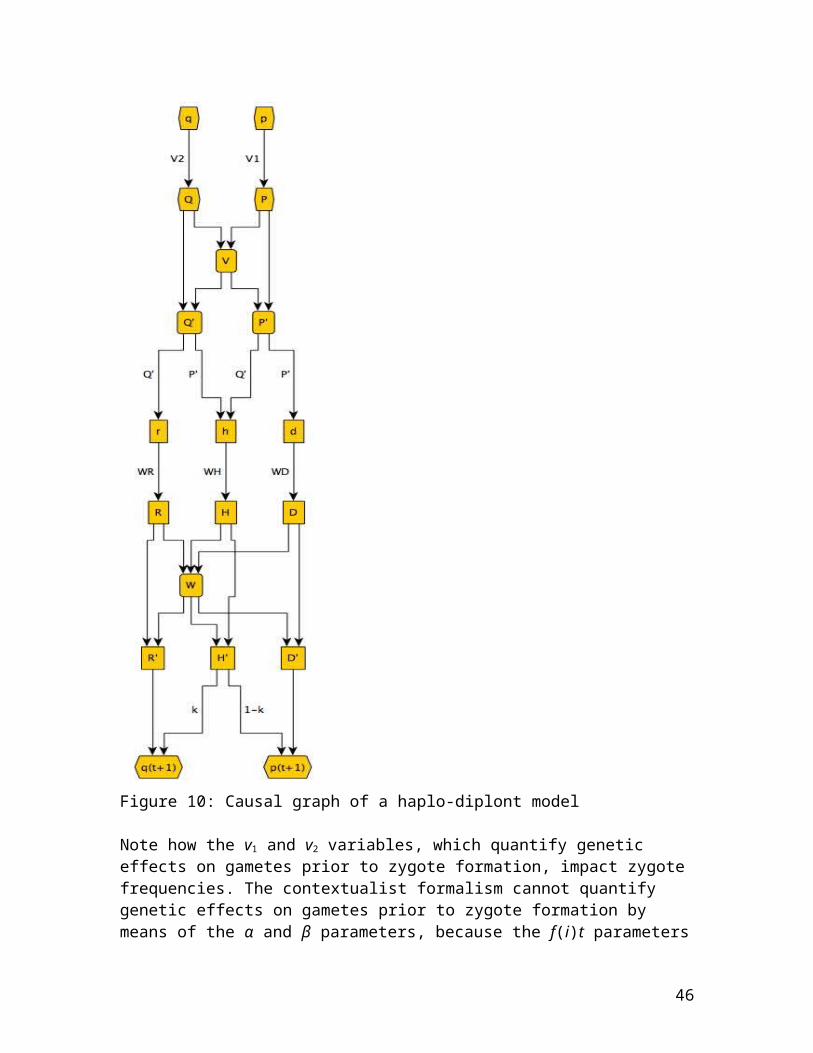
Figure 10: Causal graph of a haplo-diplont model
Note how the v1 and v2 variables, which quantify genetic effects on gametes prior to zygote formation, impact zygote frequencies. The contextualist formalism cannot quantify genetic effects on gametes prior to zygote formation by means of the α and β parameters, because the f(i)t parameters
46

are not functions of these.Even if we alter the f)i)t functions such that they are
a function of the α and β parameters in order to accommodate gametic selection prior to zygote formation, theresult is that the α and β parameters can no longer quantifydifferential zygote viability: poor gametic performance may demand a low value for, say, α2 while strong zygote performance among homozygotes for the dominant allele may demand a high value for the same quantity. Cases of antagonistic pleiotropy, in which gametic selection and zygotic selection each to favor rival alleles cannot be modeled using a single set of fitness parameters: instead two sets of fitness parameters quantify selection at each lifecycle phase, as are featured, in the form of both v’s and w’s, in the haplo-diplont model above.
Appendix 2
In their (2012) consideration of a trait-group model of altruism, an instance of a contextualist model, KGS considera system in which groups of three particles are formed, individuals have a baseline fitness of 2, altruists contribute 2 extra offspring each to fellow group members, and incur a cost of 1 offspring for their altruism. KGS set out fixed fitness parameters for both the contextualist and the collectivist representations:
KGS next run their near-variant test on the trait-group model, changing things so that altruists provide a benefit of 4 to fellow group members and incur a cost of 2. They conclude from their test that both the contextualist and thecollectivist parameterizations are on roughly equal footing since one must change the same number of parameters in each formalism to restore dynamical adequacy for the near-variant(Godfrey-Smith and Kerr 2012, 9).
47

KGS’s formal representation of the trait-group model uses fitness coefficients rather than fitness functions, such that their equations are not causally adequate. When their fitness coefficients are replaced by fitness functionsthat separate out the causal mechanisms into a baseline component, a benefit component, and a cost component, it becomes clear that KGS’s near variant test effectively involves changing multiple causal mechanisms at once, since they imagine altering both the benefit conferred by altruists (b) and the cost of altruism (c). Accordingly, thetransformation they use to generate a near variant is not produced by an intervention. This is why KGS draw a different conclusion than the one drawn here with respect tocontextualist representation of minor groups.
To see this in detail, replace KGS’s fixed fitness coefficients for the α and β parameters in the contextualistmodel with the following fitness functions (with k as baseline fitness):
αi=k+bi−ciβi=k+bik=2b=2ic=1
KGS generate a near variant by means of two simultaneous interventions on the system, one setting b = 4, and another setting c = 2. If we contemplate running these interventionsseparately, then we get results that parallel the ones we got above for the D. dendriticum case: the contextualist formalism handles informally described interventions by means of changes to the values of single quantities, while the collectivist one does not do so. Indeed, KGS themselves note that “if costs alone, or benefits alone, are altered, then the contextual parametrization fares better” (2012, 10).
Appendix 3
48

Consider a contextualist representation of viability selection among diploids, where, if modeled using the collectivist formalism, π0 = wD = 4, π1 = wH = 5, π2 = wR = 6.A dynamically adequate version of the contextualist formalism for the same case, where f(i)t parameters are once again set by the Hardy-Weinberg rule, looks like this (from equations (7)):
p̄(t+1)=α2p
2+α12pqα2p
2+α12pq+β12pq+β0q2
q̄(t+1)=α12pq+β0q
2
α2p2+α12pq+β12pq+β0q
2
α2=3 α1=2.5 β1=2.5 β0=2 (14)
If we use the haplo-diplont model (13) from Appendix 1 aboveto infer the dynamics of our diploid system with zygote fitness parameters set as above, no selection prior to zygote formation (v1 = v2), some drive k = 3/5, and p = 0.5,we infer that, after a single generation, the frequency of the A allele grows to 0.6. Reproducing this result using thecontextualist model requires bumping α1 to 3 and reducing β1
to 2 (Godfrey-Smith and Kerr 2012, 212). That both parameters must be altered shows the causal inadequacy of the contextualist formalism for the diploid selection case.
Works Cited
Cohen, Jessica, and Pascaline Dupas. 2008. "Free Distribution or Cost-Sharing? Evidence from a Randomized Malaria Prevention Experiment." On-Line Working Paper Series.
Eberhardt, Frederick, and Richard Scheines. 2006. "Interventions and Causal Inference." Philosophy of Science 74 (5):981-95.
Glymour, Clark, Richard Scheines, and Peter Spirtes 1993. Causation, Prediction, and Search. Cambridge, MA: MIT Press.
49

Godfrey-Smith, Peter, and Benjamin Kerr. 2012. "Gestalt-Switching and the Evolutionary Transitions." British Journal for the Philosophy of Science 0:1-18.
Kerr, Benjamin, and Peter Godfrey-Smith. 2002. "Individualist and Multi-level Perspectives on Selection in Structured Populations." Biology and Philosophy 17 (4):477-517.
Kitcher, Philip, and K. I. M. Sterelny. 1988. "The Return ofThe Gene." Journal of Philosophy 85:339-61.
Kitcher, Philip, K. I. M. Sterelny, and C. Kenneth Waters. 1990. "The Illusory Riches of Sober's Monism." Journal of Philosophy.
Lewis, David. 2000. "Causation as Influence." Journal of Philosophy 97:182-98.
Lloyd, Elisabeth. 2005. "Why the Gene Will not Return." Philosophy of Science 72 (2):287-310.
Lloyd, Elisabeth, Matthew Dunn, Jennifer Cianciollo, and Costas Mannouris. 2005. "Pluralism without Genic Causes." Philosophy of Science 72 (2):334-41.
Lloyd, Elisabeth, Richard C. Lewontin, and Marcus W. Feldman. 2008. "The Generational Cycle of State Spaces and Adequate Genetical Representation." Philosophy of Science 75:140-56.
Maynard Smith, John 1987a. "How to Model Evolution." In The Latest on the Best, ed. John Dupre, Cambridge, Mass,: MIT Press.
——— 1987b. "Reply to Sober." In The Latest on the Best, ed. John Dupre, Cambridge, MA: MIT Press.
Maynard Smith, John, and Szathmáry 1995. The Major Transitions in Evolution. Oxford: Oxford University Press.
Okasha, Samir 2006. Evolution and the Levels of Selection. New York: Oxford University Press.
Pearl, Judea 2000. Causality: Models, Reasoning, and Inference. Cambridge: Cambridge University Press.
Scudo, Francesco M. 1967. "Selection on Both Haplo and Diplophase." Genetics 56:693-704.
Sober, Elliott 1984. The Nature Of Selection: Evolutionary Theory In Philosophical Focus. Cambridge, Mass.: MIT Press.
50

——— 1987. "Comments on Maynard Smith." In The Latest on the best : essays on evolution and optimality ed. John Dupre, 359 Cambridge, Mass.: MIT Press.
Sober, Elliott, and Richard C. Lewontin. 1982. "Artifact, Cause And Genic Selection." Philosophy of Science.
Sober, Elliott, and David Sloan Wilson 1998. Unto Others: The Evolution and Psychology of Unselfish Behavior. Cambridge: HarvardUniversity Press.
Sterelny, Kim 2001. The Evolution of Agency and other essays. Cambridge: Cambridge University Press.
Waters, Kenneth C. 2005. "Why Genic and Multilevel SelectionTheories are Here to Stay." Philosophy of Science 72 (2):311-33.
Wilson, David Sloan. 1975. "A Theory of Group Selection." Proceedings of the National Academy of Sciences of the United States of America 72 (1):143-46.
Woodward, James. 2002. "What is a mechanism? A counterfactual Account." Philosophy of Science 69 (S3):S366-S77.
——— 2003. Making Things Happen. New York: Oxford University Press.
51




















