
Low Densities of Epiphytic Bacteria from the Marine Alga Ulva australis Inhibit Settlement of...
10
Published Ahead of Print 26 October 2007. 10.1128/AEM.01543-07. 2007, 73(24):7844. DOI: Appl. Environ. Microbiol. Case, Adrian Low, Peter Steinberg and Staffan Kjelleberg Dhana Rao, Jeremy S. Webb, Carola Holmström, Rebecca Settlement of Fouling Organisms Inhibit Ulva australis the Marine Alga Low Densities of Epiphytic Bacteria from http://aem.asm.org/content/73/24/7844 Updated information and services can be found at: These include: REFERENCES http://aem.asm.org/content/73/24/7844#ref-list-1 at: This article cites 70 articles, 20 of which can be accessed free CONTENT ALERTS more» articles cite this article), Receive: RSS Feeds, eTOCs, free email alerts (when new http://journals.asm.org/site/misc/reprints.xhtml Information about commercial reprint orders: http://journals.asm.org/site/subscriptions/ To subscribe to to another ASM Journal go to: on August 15, 2013 by UNIV OF ALBERTA http://aem.asm.org/ Downloaded from
Transcript of Low Densities of Epiphytic Bacteria from the Marine Alga Ulva australis Inhibit Settlement of...

Published Ahead of Print 26 October 2007. 10.1128/AEM.01543-07.
2007, 73(24):7844. DOI:Appl. Environ. Microbiol. Case, Adrian Low, Peter Steinberg and Staffan KjellebergDhana Rao, Jeremy S. Webb, Carola Holmström, Rebecca Settlement of Fouling Organisms
InhibitUlva australisthe Marine Alga Low Densities of Epiphytic Bacteria from
http://aem.asm.org/content/73/24/7844Updated information and services can be found at:
These include:
REFERENCEShttp://aem.asm.org/content/73/24/7844#ref-list-1at:
This article cites 70 articles, 20 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on August 15, 2013 by U
NIV
OF
ALB
ER
TA
http://aem.asm
.org/D
ownloaded from

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Dec. 2007, p. 7844–7852 Vol. 73, No. 240099-2240/07/$08.00�0 doi:10.1128/AEM.01543-07Copyright © 2007, American Society for Microbiology. All Rights Reserved.
Low Densities of Epiphytic Bacteria from the Marine AlgaUlva australis Inhibit Settlement of
Fouling Organisms�
Dhana Rao,1,2 Jeremy S. Webb,1,2† Carola Holmstrom,1,2 Rebecca Case,1,2 Adrian Low,1,2
Peter Steinberg,2,3 and Staffan Kjelleberg1,2*School of Biotechnology and Biomolecular Sciences,1 Centre for Marine Bio-innovation,2 and School of Biological, Earth and
Environmental Sciences,3 University of New South Wales, Sydney, New South Wales 2052, Australia
Received 9 July 2007/Accepted 15 October 2007
Bacteria that produce inhibitory compounds on the surface of marine algae are thought to contribute to thedefense of the host plant against colonization of fouling organisms. However, the number of bacterial cellsnecessary to defend against fouling on the plant surface is not known. Pseudoalteromonas tunicata and Phaeo-bacter sp. strain 2.10 (formerly Roseobacter gallaeciensis) are marine bacteria often found in association with thealga Ulva australis and produce a range of extracellular inhibitory compounds against common foulingorganisms. P. tunicata and Phaeobacter sp. strain 2.10 biofilms with cell densities ranging from 102 to 108 cellscm�2 were established on polystyrene petri dishes. Attachment and settlement assays were performed withmarine fungi (uncharacterized isolates from U. australis), marine bacteria (Pseudoalteromonas gracilis, Altero-monas sp., and Cellulophaga fucicola), invertebrate larvae (Bugula neritina), and algal spores (Polysiphonia sp.)and gametes (U. australis). Remarkably low cell densities (102 to 103 cells cm�2) of P. tunicata were effective inpreventing settlement of algal spores and marine fungi in petri dishes. P. tunicata also prevented settlement ofinvertebrate larvae at densities of 104 to 105 cells cm�2. Similarly, low cell densities (103 to 104cells cm�2) ofPhaeobacter sp. strain 2.10 had antilarval and antibacterial activity. Previously, it has been shown thatabundance of P. tunicata on marine eukaryotic hosts is low (<1 � 103 cells cm�2) (T. L. Skovhus et al., Appl.Environ. Microbiol. 70:2373–2382, 2004). Despite such low numbers of P. tunicata on U. australis in situ, ourdata suggest that P. tunicata and Phaeobacter sp. strain 2.10 are present in sufficient quantities on the plant toinhibit fouling organisms. This strongly supports the hypothesis that P. tunicata and Phaeobacter sp. strain 2.10can play a role in defense against fouling on U. australis at cell densities that commonly occur in situ.
Surfaces never remain pristine in the marine environmentbut become quickly colonized by a film of marine bacteria.Such biofilms serve as cues that modify the behavior of settlinginvertebrate larvae and algal cells (12, 73) and significantlyinfluence the final composition of the biofouling community.These cues can be either positive (11, 42, 43, 49, 57, 69) ornegative (19, 33, 50), depending on the species of foulingorganism and bacteria concerned. Thus, biofilms play an im-portant role in the development of fouling communities onmarine surfaces.
As well as being a common fouling organism, the intertidalgreen alga Ulva australis is itself susceptible to fouling becauseit is sessile and restricted to the photic zone, where conditionsfor fouling organisms are optimal (13). Many seaweeds haveevolved efficient strategies to combat epibiosis (14, 17, 41), butthese antifouling defenses can be energetically costly (72), andit has been suggested that epibiotic bacteria living on the sur-face of the alga may provide microbial defense (3, 18, 19, 33).Such interactions are not uncommon in the marine environ-
ment (16, 24, 25, 28). A number of inhibitory bacteria havebeen isolated from U. australis, and one of the best character-ized of these is Pseudoalteromonas tunicata (18, 20, 32). Thisbacterium produces a diverse range of biologically active com-pounds that specifically target marine fouling organisms (19,23, 30, 32, 33, 38) and gram-negative and gram-positive bacte-ria from a range of environments (48). Phaeobacter sp. strain2.10 (formerly Roseobacter gallaeciensis) is also frequently iso-lated from the surface of U. australis and has known antibac-terial activity (4, 58, 61). Members of the Roseobacter clade arefrequently associated with algae (8) and can comprise up to25% of microbial communities in coastal environments (71).
Molecular investigations based on real-time quantitativePCR have shown that while the genus Pseudoalteromonas iscommon throughout the marine environment, P. tunicatamostly inhabits living surfaces that are relatively free fromfouling such as green algae (Ulva lactuca and Ulvaria fusca)and tunicates (Ciona intestinalis) (65) and has low in situ den-sity (�1 � 103 cells cm�2) (66). Studies based on a methodcombining catalyzed reporter deposition with fluorescence insitu hybridization suggested that Phaeobacter sp. strain 2.10may be present in higher numbers, as the genus Roseobactercomprised 12% of the epiphytic bacterial community on U.australis (unpublished data).
Although the inhibitory effects of P. tunicata against a rangeof fouling organisms in the laboratory are well established (18,31), the observation that P. tunicata is present at such low
* Corresponding author. Mailing address: School of Biotechnologyand Biomolecular Sciences and Centre for Marine Bio-innovation,Biological Sciences Building, University of New South Wales, Sydney,NSW 2052, Australia. Phone: 61 (2) 9385 2102/2276. Fax: 61 (2) 93851779. E-mail: [email protected].
† Present address: School of Biological Sciences, University ofSouthampton, Southampton SO16 7PX, United Kingdom.
� Published ahead of print on 26 October 2007.
7844
on August 15, 2013 by U
NIV
OF
ALB
ER
TA
http://aem.asm
.org/D
ownloaded from

densities raises the question as to whether it could in fact haveantifouling activity at these densities in the marine environ-ment. In this study, we tested the density dependence of anti-fouling activity of P. tunicata and Phaeobacter sp. strain 2.10biofilms and show that they have inhibitory effects at ecologi-cally relevant densities. Furthermore, the inhibition of algalspores and larval settlement by P. tunicata biofilms is shown tobe due to the production of antifouling compounds, as mutantsdefective in the production of the extracellular inhibitors donot display inhibitory activity. Inhibition of settlement andattachment by Phaeobacter sp. strain 2.10 biofilms is through anunknown mechanism. However, Phaeobacter sp. strain 2.10produces the density-dependent signal molecules acyl homo-serine lactones (AHLs), which are also settlement cues forUlva sp. spores (40). This suggests that AHLs may play a rolein mediating the relationship between U. australis and quorum-sensing bacterial epiphytes capable of defending the alga. Ourdata support the hypothesis that P. tunicata and Phaeobactersp. strain 2.10 play a role in defending U. australis againstfouling in situ.
MATERIALS AND METHODS
Preparation of marine strains. Bacterial strains were isolated from the surfaceof U. australis as described previously by Rao et al. (58). The organisms selectedfor this study were P. tunicata, Phaeobacter sp. strain 2.10, P. gracilis, Alteromonassp., Cellulophaga fucicola; as controls, P. tunicata mutants defective in the pro-duction of the antibacterial protein (AlpP) (47), all inhibitory compounds(WmpR) (17), and the antifungal compound (FM3) (20) were used. Cultureswere stored at �80°C in 50% (vol/vol) glycerol in VNSS medium (52) andmaintained on VNSS agar plates. For visualization of bacterial cells using epi-fluorescence microscopy, P. tunicata, P. gracilis, Alteromonas sp., C. fucicola, andPhaeobacter sp. strain 2.10 were labeled with either DsRed or green fluorescentprotein (GFP) color tag as described previously by Rao et al. (58).
Establishment of biofilms. Bacteria were cultured for 24 h at 25°C in VNSSbroth for preparation of inocula. Cells were harvested by spinning down theculture and resuspending the pellet in seawater. The cell concentration wasestimated by counting using epifluorescence microscopy and a hemocytometerand adjusted by dilution with seawater to the desired end concentration. Biofilmswere established by inoculation from overnight precultures into 3.6-cm petridishes containing 3 ml of 10% VNSS medium (diluted in seawater) and incu-bated at 23°C for 24 h. Different densities of cells were inoculated so that thefinal density of attached cells on the surface of the petri dishes ranged from 102
to 108cells cm�2. After 24 h of incubation, growth medium was discarded andestablished biofilms were rinsed three times with sterile filtered seawater andincubated in fresh sterile seawater before the bioassays were conducted. Num-bers of attached cells were determined by counting the number of DsRed- orGFP-labeled cells with epifluorescence microscopy and an eyepiece grid. Twenty-four-hour-old biofilms were used for antifouling assays as older biofilmsundergo a certain amount of detachment and sloughing, which affected celldensities. The biofilm densities were measured after the assays to establishwhether the density had changed. Preliminary experiments indicated that celldensities in biofilms did not increase significantly over the course of theexperiment, particularly for the lower densities. For all assays, three experi-ments were carried out with at least four replicates in each treatment.
Antialgal bioassays. The effects of different cell densities of bacteria on at-tachment and survival of algal propagules were assessed by exposing gametes orspores directly to monoculture biofilms of P. tunicata and Phaeobacter sp. strain2.10 at densities ranging from 102 to 108 cells cm�2. Sterile seawater and P.tunicata WmpR mutant biofilms established at 106cells cm�2 served as controlsfor P. tunicata biofilms. Sterile seawater was the control for Phaeobacter sp. strain2.10.
U. australis gamete and Polysiphonia sp. spore bioassays were set up as de-scribed by Egan et al. (19). U. australis gamete settlement was assessed after 5days using an inverted light microscope (Zeiss). Counts were conducted in 10fields of view using a �40 magnification, and settlement was compared to that incontrols. Polysiphonia sp. spore settlement and subsequent development wereassessed after 24 h, the numbers of settled (i.e., attached) and unsettled spores
were counted using a dissecting microscope, and the percentage of settlementwas determined.
(i) Detection of AHLs. P. tunicata and Phaeobacter sp. strain 2.10 werescreened for AHL production using a streak assay and the AHL reporter strain,Agrobacterium tumefaciens A136. Briefly, plates were poured with one mediumand set, and half of the medium was aseptically removed and replaced with asecond medium. This was done because P. tunicata, Phaeobacter sp. strain 2.10,and A. tumefaciens A136 require different growth media. A. tumefaciens A136was grown on LB5 plates (2) supplemented with tetracycline (4.5 �g/ml), spec-tinomycin (50 �g/ml), and X-Gal (5-bromo-4-chloro-3-indolyl-�-D-galacto-pyranoside [50 �g/ml]). Marine strains grew on marine broth 2216 supplementedwith 1.5% agar.
Following a positive result in the AHL streak assay, AHLs were furthercharacterized with thin-layer chromatography (TLC) plates overlaid with A.tumefaciens A136. To extract AHLs from P. tunicata and Phaeobacter sp. 2.10,the strains were grown to stationary phase in 10 ml of marine broth 2216. These10-ml cultures were sterile filtrated (0.22 �m), and the supernatants were re-tained. AHLs were extracted from the supernatants as described by Ravn et al.(60), the AHL extracts were resuspended in 100 �l of acidified ethyl acetate, and40 to 80 �l of each sample was applied to C18 TLC plates (TLC aluminum sheets,10 by 10 cm2, Rp-18 F254 s,1.05559 [Merck 64271]) and developed in 60:40methanol–Milli-Q water as described by Shaw et al. (63). A lawn of A. tumefa-ciens A136 in AB medium was overlaid on the TLC plate as described by Ravnet al. (60). AHLs were compared against commercial standards 3-oxo-hexanoylhomoserine lactone (OHHL), N-oxo-octanoyl-L-homoserine lactone (OOHL),and N-octanoyl-homoserine lactone (OHL), and Rf values were calculated (56).AHLs were identified based on Rf values and the shape of spots.
(ii) AHLs as settlement cues. To determine if AHLs were involved in enhanc-ing settlement of U. australis gametes, an assay was conducted with differentconcentrations of the C8-AHL, OOHL. OOHL was used because unsubstitutedC7-AHL is not commercially available and AHLs with a carbon chain length of�6 are freely diffusible in water and those with a length of �10 have markedlylower solubility (55). OOHL was suspended in a 1% agarose–distilled watersupport matrix as described in reference 70 at concentrations of 5, 10, 20, and 50�M, respectively. A control of 50 �M methanol in 1% agarose was used. Gameteattachment was assessed after 1 h, using an inverted light microscope (Zeiss).Gametes were stained with crystal violet, counts were conducted in 10 fields ofview using a �40 magnification, and settlement was compared to that in controls.
Antilarval assays. P. tunicata and Phaeobacter sp. strain 2.10 at a range ofdensities were tested using standard settlement assays of larvae of the bryozoanBugula neritina (7). Adult broodstocks of B. neretina were collected from pilingsat Rose Bay, Sydney, Australia (33°87�52”S, 151°25�56”E) and larvae were ob-tained as described by de Nys et al. (13) and Bryan et al. (7). Only newly releasedlarvae were included in the bioassay (i.e., within 15 min of release). Biofilms wereestablished as described above, petri dishes were rinsed three times with 2 ml ofsterile seawater, and then 3 ml of filtered seawater containing approximately 20larvae was added to each petri dish and incubated at 25°C for 2 days. Larvae werecounted under a dissecting microscope, and the percentage of settlement wasdetermined. Control petri dishes contained either sterile seawater alone (forboth P. tunicata and Phaeobacter sp. strain 2.10) or the P. tunicata WmpRmutant’s biofilm with cell densities of 106 cells ml�1 (control for P. tunicata).
Antifungal assays. Both yeast and filamentous fungi were used to assess theantifungal activity of P. tunicata and Phaeobacter sp. strain 2.10. Unidentifiedmarine yeast (Y1, Y2, and Y3) and a filamentous fungal (Y4) strain previouslyisolated from U. australis (21) were used as target strains. Fungal strains weremaintained on VNSS plates and inoculated into VNSS broth. Bacterial biofilmswere established as described earlier, petri dishes were rinsed three times with 2ml of sterile seawater, and 2 ml of medium (10% VNSS–90% seawater) con-taining 105cells ml�1 of fungi was added and incubated for 48 h. The fungi werestained with Syto 59, and the percentage of fungal surface cover was comparedto that of the original inoculum by epifluorescence microscopy. Biofilms of P.tunicata FM3 and WmpR mutant cells established at 106 cells cm�2 served ascontrols. Counts were done on 10 fields of view using a �40 magnification.
Antibacterial assays. Marine strains isolated from U. australis were used astarget strains to test for antibacterial activity of P. tunicata and Phaeobacter sp.strain 2.10 biofilms established on plastic surfaces. The bacterial challenge strainsconsisted of C. fucicola, Alteromonas sp., Phaeobacter sp. strain 2.10, P. tunicata,and P. gracilis. Biofilms of P. tunicata, and Phaeobacter sp. strain 2.10 wereestablished and challenged with test bacteria added to the biofilms at 106 cellsml�1 and incubated for 48 h. P. tunicata AlpP and WmpR mutant biofilms andculture media were used as controls for P. tunicata, while culture medium was thecontrol for Phaeobacter sp. strain 2.10. DsRed-labeled challenge strains were
VOL. 73, 2007 BACTERIAL INHIBITION OF FOULING ORGANISM SETTLEMENT 7845
on August 15, 2013 by U
NIV
OF
ALB
ER
TA
http://aem.asm
.org/D
ownloaded from
readily distinguished from GFP-labeled biofilm bacteria by epifluorescence mi-croscopy. Counts were done on 10 fields of view using a �10 magnification.
Statistical analysis. One-way analysis of variance (ANOVA) followed byTukey’s pairwise comparisons were used to compare the settlement of spores orlarvae and attachment of bacteria or fungi in response to various densities of P.tunicata and R. gallaeciensis. ANOVA assumptions of normality and heteroge-neity of variance of the data were checked. Tests were conducted with Systat 10(SPSS).
RESULTS AND DISCUSSION
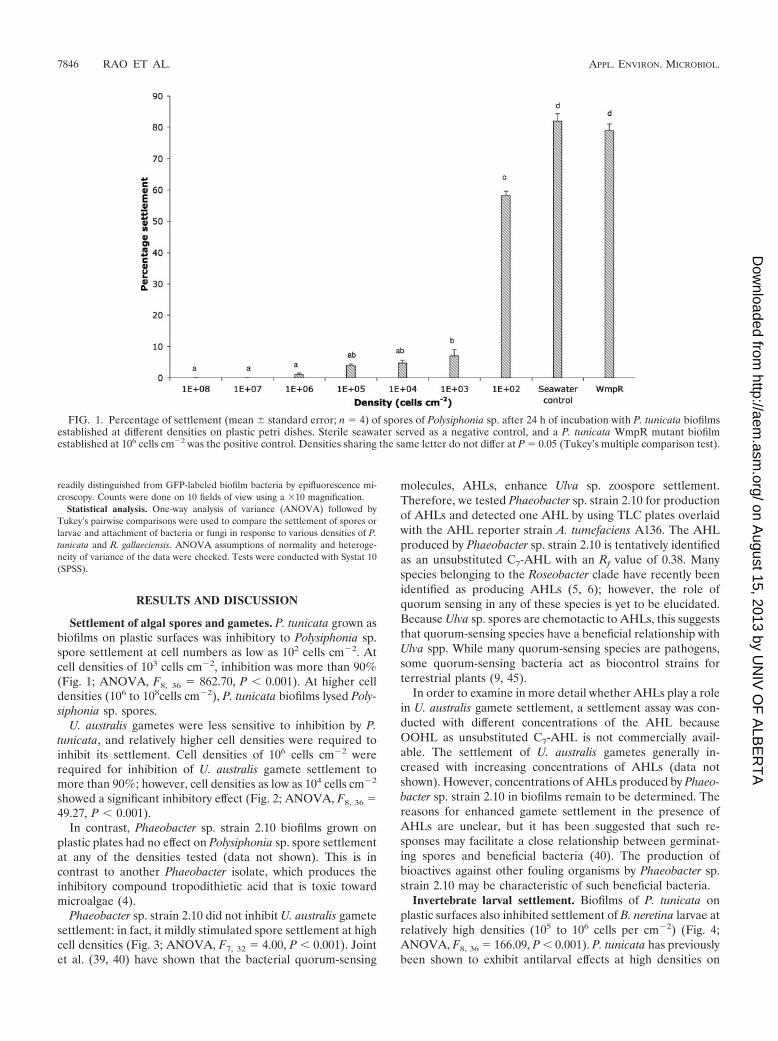
Settlement of algal spores and gametes. P. tunicata grown asbiofilms on plastic surfaces was inhibitory to Polysiphonia sp.spore settlement at cell numbers as low as 102 cells cm�2. Atcell densities of 103 cells cm�2, inhibition was more than 90%(Fig. 1; ANOVA, F8, 36 862.70, P � 0.001). At higher celldensities (106 to 108cells cm�2), P. tunicata biofilms lysed Poly-siphonia sp. spores.
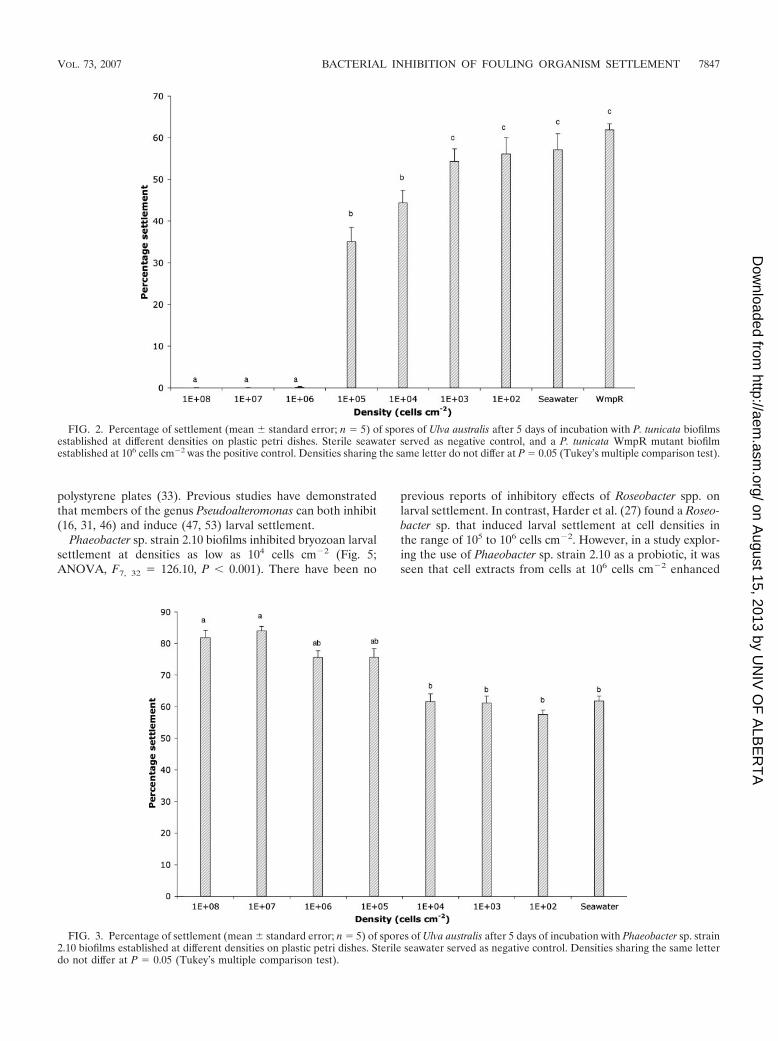
U. australis gametes were less sensitive to inhibition by P.tunicata, and relatively higher cell densities were required toinhibit its settlement. Cell densities of 106 cells cm�2 wererequired for inhibition of U. australis gamete settlement tomore than 90%; however, cell densities as low as 104 cells cm�2
showed a significant inhibitory effect (Fig. 2; ANOVA, F8, 36 49.27, P � 0.001).
In contrast, Phaeobacter sp. strain 2.10 biofilms grown onplastic plates had no effect on Polysiphonia sp. spore settlementat any of the densities tested (data not shown). This is incontrast to another Phaeobacter isolate, which produces theinhibitory compound tropodithietic acid that is toxic towardmicroalgae (4).
Phaeobacter sp. strain 2.10 did not inhibit U. australis gametesettlement: in fact, it mildly stimulated spore settlement at highcell densities (Fig. 3; ANOVA, F7, 32 4.00, P � 0.001). Jointet al. (39, 40) have shown that the bacterial quorum-sensing
molecules, AHLs, enhance Ulva sp. zoospore settlement.Therefore, we tested Phaeobacter sp. strain 2.10 for productionof AHLs and detected one AHL by using TLC plates overlaidwith the AHL reporter strain A. tumefaciens A136. The AHLproduced by Phaeobacter sp. strain 2.10 is tentatively identifiedas an unsubstituted C7-AHL with an Rf value of 0.38. Manyspecies belonging to the Roseobacter clade have recently beenidentified as producing AHLs (5, 6); however, the role ofquorum sensing in any of these species is yet to be elucidated.Because Ulva sp. spores are chemotactic to AHLs, this suggeststhat quorum-sensing species have a beneficial relationship withUlva spp. While many quorum-sensing species are pathogens,some quorum-sensing bacteria act as biocontrol strains forterrestrial plants (9, 45).
In order to examine in more detail whether AHLs play a rolein U. australis gamete settlement, a settlement assay was con-ducted with different concentrations of the AHL becauseOOHL as unsubstituted C7-AHL is not commercially avail-able. The settlement of U. australis gametes generally in-creased with increasing concentrations of AHLs (data notshown). However, concentrations of AHLs produced by Phaeo-bacter sp. strain 2.10 in biofilms remain to be determined. Thereasons for enhanced gamete settlement in the presence ofAHLs are unclear, but it has been suggested that such re-sponses may facilitate a close relationship between germinat-ing spores and beneficial bacteria (40). The production ofbioactives against other fouling organisms by Phaeobacter sp.strain 2.10 may be characteristic of such beneficial bacteria.
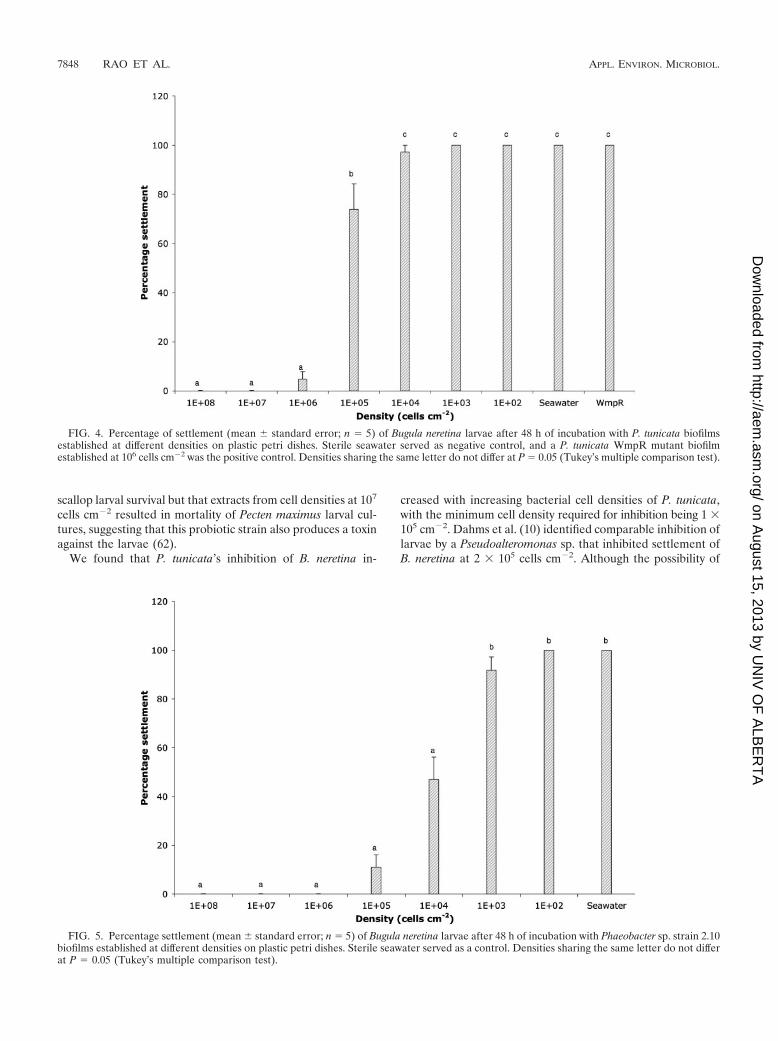
Invertebrate larval settlement. Biofilms of P. tunicata onplastic surfaces also inhibited settlement of B. neretina larvae atrelatively high densities (105 to 106 cells per cm�2) (Fig. 4;ANOVA, F8, 36 166.09, P � 0.001). P. tunicata has previouslybeen shown to exhibit antilarval effects at high densities on
FIG. 1. Percentage of settlement (mean standard error; n 4) of spores of Polysiphonia sp. after 24 h of incubation with P. tunicata biofilmsestablished at different densities on plastic petri dishes. Sterile seawater served as a negative control, and a P. tunicata WmpR mutant biofilmestablished at 106 cells cm�2 was the positive control. Densities sharing the same letter do not differ at P 0.05 (Tukey’s multiple comparison test).
7846 RAO ET AL. APPL. ENVIRON. MICROBIOL.
on August 15, 2013 by U
NIV
OF
ALB
ER
TA
http://aem.asm
.org/D
ownloaded from
polystyrene plates (33). Previous studies have demonstratedthat members of the genus Pseudoalteromonas can both inhibit(16, 31, 46) and induce (47, 53) larval settlement.
Phaeobacter sp. strain 2.10 biofilms inhibited bryozoan larvalsettlement at densities as low as 104 cells cm�2 (Fig. 5;ANOVA, F7, 32 126.10, P � 0.001). There have been no
previous reports of inhibitory effects of Roseobacter spp. onlarval settlement. In contrast, Harder et al. (27) found a Roseo-bacter sp. that induced larval settlement at cell densities inthe range of 105 to 106 cells cm�2. However, in a study explor-ing the use of Phaeobacter sp. strain 2.10 as a probiotic, it wasseen that cell extracts from cells at 106 cells cm�2 enhanced
FIG. 2. Percentage of settlement (mean standard error; n 5) of spores of Ulva australis after 5 days of incubation with P. tunicata biofilmsestablished at different densities on plastic petri dishes. Sterile seawater served as negative control, and a P. tunicata WmpR mutant biofilmestablished at 106 cells cm�2 was the positive control. Densities sharing the same letter do not differ at P 0.05 (Tukey’s multiple comparison test).
FIG. 3. Percentage of settlement (mean standard error; n 5) of spores of Ulva australis after 5 days of incubation with Phaeobacter sp. strain2.10 biofilms established at different densities on plastic petri dishes. Sterile seawater served as negative control. Densities sharing the same letterdo not differ at P 0.05 (Tukey’s multiple comparison test).
VOL. 73, 2007 BACTERIAL INHIBITION OF FOULING ORGANISM SETTLEMENT 7847
on August 15, 2013 by U
NIV
OF
ALB
ER
TA
http://aem.asm
.org/D
ownloaded from
scallop larval survival but that extracts from cell densities at 107
cells cm�2 resulted in mortality of Pecten maximus larval cul-tures, suggesting that this probiotic strain also produces a toxinagainst the larvae (62).
We found that P. tunicata’s inhibition of B. neretina in-
creased with increasing bacterial cell densities of P. tunicata,with the minimum cell density required for inhibition being 1 �105 cm�2. Dahms et al. (10) identified comparable inhibition oflarvae by a Pseudoalteromonas sp. that inhibited settlement ofB. neretina at 2 � 105 cells cm�2. Although the possibility of
FIG. 4. Percentage of settlement (mean standard error; n 5) of Bugula neretina larvae after 48 h of incubation with P. tunicata biofilmsestablished at different densities on plastic petri dishes. Sterile seawater served as negative control, and a P. tunicata WmpR mutant biofilmestablished at 106 cells cm�2 was the positive control. Densities sharing the same letter do not differ at P 0.05 (Tukey’s multiple comparison test).
FIG. 5. Percentage settlement (mean standard error; n 5) of Bugula neretina larvae after 48 h of incubation with Phaeobacter sp. strain 2.10biofilms established at different densities on plastic petri dishes. Sterile seawater served as a control. Densities sharing the same letter do not differat P 0.05 (Tukey’s multiple comparison test).
7848 RAO ET AL. APPL. ENVIRON. MICROBIOL.
on August 15, 2013 by U
NIV
OF
ALB
ER
TA
http://aem.asm
.org/D
ownloaded from
repellents was not discounted by Dahms and coworkers (10),they ascribed the higher settlement of larvae at lower densitiesof bacterial cells to bacterial free space and lower wettabilityon biofilm-coated dishes. Similarly, Olivier et al. (54) showedthat cyprid settlement in Balanus amphitrite was negativelycorrelated with total density in biofilms and attributed this tofree-space availability. The results described here suggest,however, that antifouling compounds produced by P. tunicataresult in inhibition of settlement, as the biofilm established bythe WmpR mutant was not inhibitory to larval settlement.
Many studies have stressed the importance of biofilm age ininfluencing the settlement of larvae or algal propagules (1, 26,34, 44, 64). However, the rate of settlement reported in thesestudies seems to correlate with the density of cells, whichgenerally increases with the age of the biofilm. Althoughchanges in cell densities were generally not monitored in thesestudies, as biofilms grew older, the older the biofilms, thehigher the level of induction (1, 26, 34, 44, 64) or inhibition (10,51). Thus, some invertebrate larvae are able to discriminatebetween the ages of biofilms as induction or inhibition levelscorrelate with bacterial cell densities.
A recent study demonstrated that bacterial strains that in-duce a high level of settlement in the sea urchin Heliocidariserythrogramma were dominated by the genera Pseudoalteromo-nas, Shewanella, and Vibrio. These genera were effective atinducing settlement, despite being present at low densities onseaweed surfaces (�1 � 105 cells cm�2) and representing onlya small percentage of the total bacterial community (36). Fieldrecruitment of H. erythrogramma correlated with laboratorysettlement assays, suggesting that cues were present at a con-centration that larvae were able to detect and respond to.
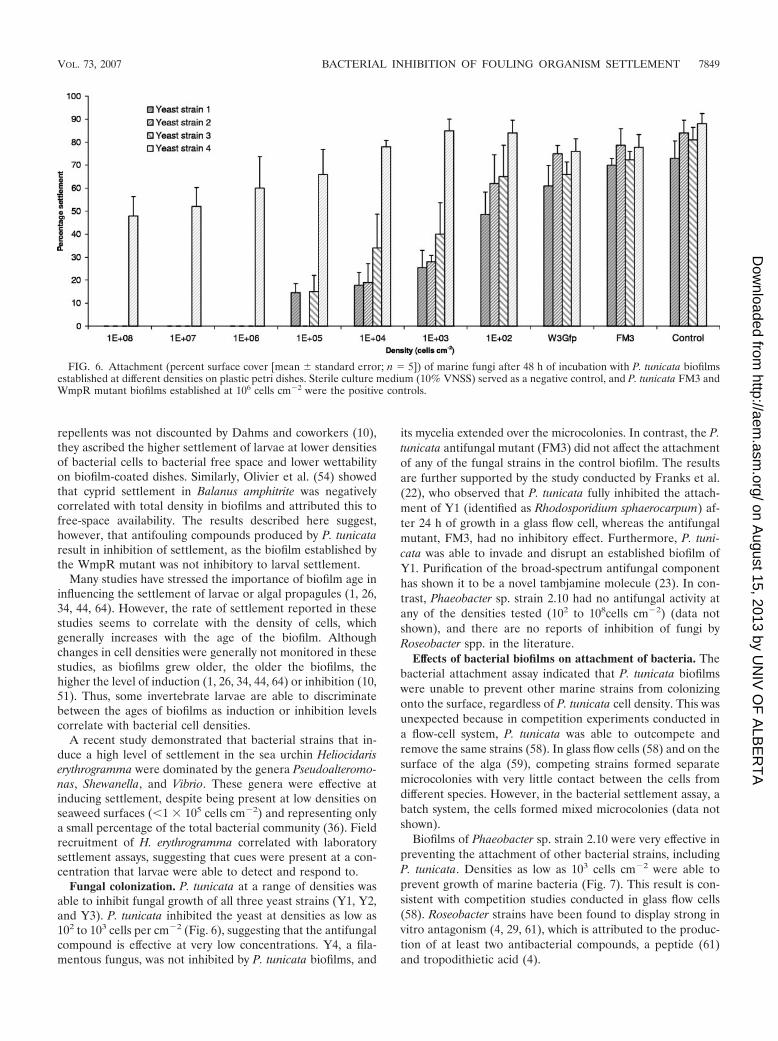
Fungal colonization. P. tunicata at a range of densities wasable to inhibit fungal growth of all three yeast strains (Y1, Y2,and Y3). P. tunicata inhibited the yeast at densities as low as102 to 103 cells per cm�2 (Fig. 6), suggesting that the antifungalcompound is effective at very low concentrations. Y4, a fila-mentous fungus, was not inhibited by P. tunicata biofilms, and
its mycelia extended over the microcolonies. In contrast, the P.tunicata antifungal mutant (FM3) did not affect the attachmentof any of the fungal strains in the control biofilm. The resultsare further supported by the study conducted by Franks et al.(22), who observed that P. tunicata fully inhibited the attach-ment of Y1 (identified as Rhodosporidium sphaerocarpum) af-ter 24 h of growth in a glass flow cell, whereas the antifungalmutant, FM3, had no inhibitory effect. Furthermore, P. tuni-cata was able to invade and disrupt an established biofilm ofY1. Purification of the broad-spectrum antifungal componenthas shown it to be a novel tambjamine molecule (23). In con-trast, Phaeobacter sp. strain 2.10 had no antifungal activity atany of the densities tested (102 to 108cells cm�2) (data notshown), and there are no reports of inhibition of fungi byRoseobacter spp. in the literature.
Effects of bacterial biofilms on attachment of bacteria. Thebacterial attachment assay indicated that P. tunicata biofilmswere unable to prevent other marine strains from colonizingonto the surface, regardless of P. tunicata cell density. This wasunexpected because in competition experiments conducted ina flow-cell system, P. tunicata was able to outcompete andremove the same strains (58). In glass flow cells (58) and on thesurface of the alga (59), competing strains formed separatemicrocolonies with very little contact between the cells fromdifferent species. However, in the bacterial settlement assay, abatch system, the cells formed mixed microcolonies (data notshown).
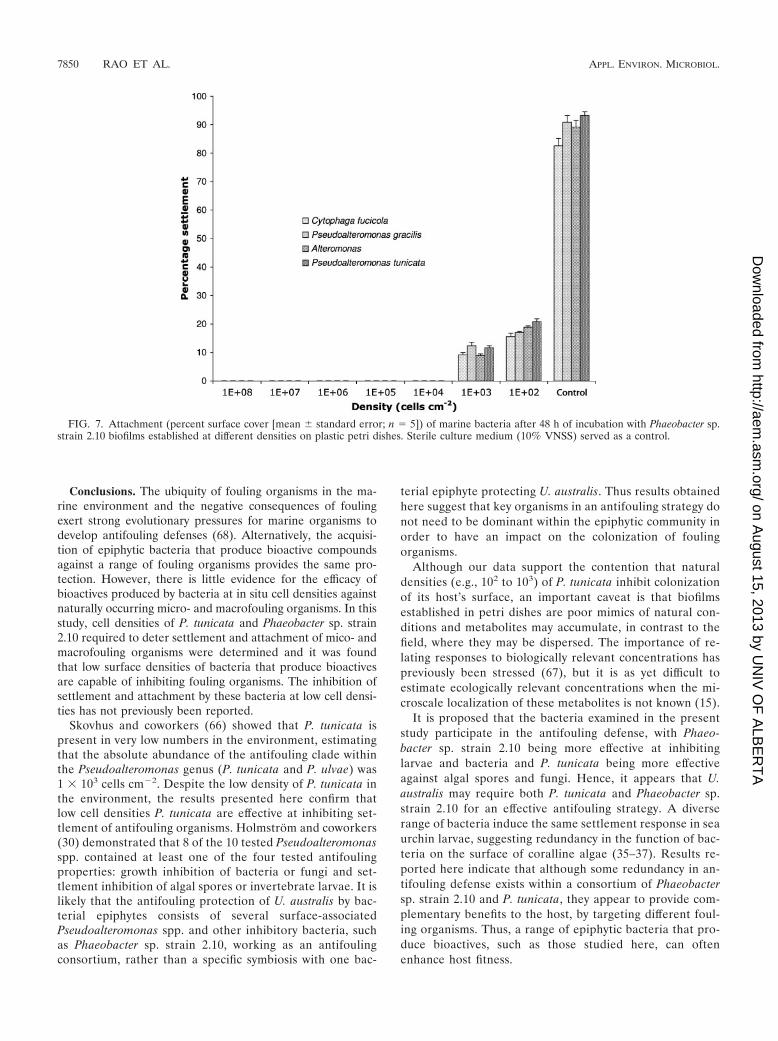
Biofilms of Phaeobacter sp. strain 2.10 were very effective inpreventing the attachment of other bacterial strains, includingP. tunicata. Densities as low as 103 cells cm�2 were able toprevent growth of marine bacteria (Fig. 7). This result is con-sistent with competition studies conducted in glass flow cells(58). Roseobacter strains have been found to display strong invitro antagonism (4, 29, 61), which is attributed to the produc-tion of at least two antibacterial compounds, a peptide (61)and tropodithietic acid (4).
FIG. 6. Attachment (percent surface cover [mean standard error; n 5]) of marine fungi after 48 h of incubation with P. tunicata biofilmsestablished at different densities on plastic petri dishes. Sterile culture medium (10% VNSS) served as a negative control, and P. tunicata FM3 andWmpR mutant biofilms established at 106 cells cm�2 were the positive controls.
VOL. 73, 2007 BACTERIAL INHIBITION OF FOULING ORGANISM SETTLEMENT 7849
on August 15, 2013 by U
NIV
OF
ALB
ER
TA
http://aem.asm
.org/D
ownloaded from
Conclusions. The ubiquity of fouling organisms in the ma-rine environment and the negative consequences of foulingexert strong evolutionary pressures for marine organisms todevelop antifouling defenses (68). Alternatively, the acquisi-tion of epiphytic bacteria that produce bioactive compoundsagainst a range of fouling organisms provides the same pro-tection. However, there is little evidence for the efficacy ofbioactives produced by bacteria at in situ cell densities againstnaturally occurring micro- and macrofouling organisms. In thisstudy, cell densities of P. tunicata and Phaeobacter sp. strain2.10 required to deter settlement and attachment of mico- andmacrofouling organisms were determined and it was foundthat low surface densities of bacteria that produce bioactivesare capable of inhibiting fouling organisms. The inhibition ofsettlement and attachment by these bacteria at low cell densi-ties has not previously been reported.
Skovhus and coworkers (66) showed that P. tunicata ispresent in very low numbers in the environment, estimatingthat the absolute abundance of the antifouling clade withinthe Pseudoalteromonas genus (P. tunicata and P. ulvae) was1 � 103 cells cm�2. Despite the low density of P. tunicata inthe environment, the results presented here confirm thatlow cell densities P. tunicata are effective at inhibiting set-tlement of antifouling organisms. Holmstrom and coworkers(30) demonstrated that 8 of the 10 tested Pseudoalteromonasspp. contained at least one of the four tested antifoulingproperties: growth inhibition of bacteria or fungi and set-tlement inhibition of algal spores or invertebrate larvae. It islikely that the antifouling protection of U. australis by bac-terial epiphytes consists of several surface-associatedPseudoalteromonas spp. and other inhibitory bacteria, suchas Phaeobacter sp. strain 2.10, working as an antifoulingconsortium, rather than a specific symbiosis with one bac-
terial epiphyte protecting U. australis. Thus results obtainedhere suggest that key organisms in an antifouling strategy donot need to be dominant within the epiphytic community inorder to have an impact on the colonization of foulingorganisms.
Although our data support the contention that naturaldensities (e.g., 102 to 103) of P. tunicata inhibit colonizationof its host’s surface, an important caveat is that biofilmsestablished in petri dishes are poor mimics of natural con-ditions and metabolites may accumulate, in contrast to thefield, where they may be dispersed. The importance of re-lating responses to biologically relevant concentrations haspreviously been stressed (67), but it is as yet difficult toestimate ecologically relevant concentrations when the mi-croscale localization of these metabolites is not known (15).
It is proposed that the bacteria examined in the presentstudy participate in the antifouling defense, with Phaeo-bacter sp. strain 2.10 being more effective at inhibitinglarvae and bacteria and P. tunicata being more effectiveagainst algal spores and fungi. Hence, it appears that U.australis may require both P. tunicata and Phaeobacter sp.strain 2.10 for an effective antifouling strategy. A diverserange of bacteria induce the same settlement response in seaurchin larvae, suggesting redundancy in the function of bac-teria on the surface of coralline algae (35–37). Results re-ported here indicate that although some redundancy in an-tifouling defense exists within a consortium of Phaeobactersp. strain 2.10 and P. tunicata, they appear to provide com-plementary benefits to the host, by targeting different foul-ing organisms. Thus, a range of epiphytic bacteria that pro-duce bioactives, such as those studied here, can oftenenhance host fitness.
FIG. 7. Attachment (percent surface cover [mean standard error; n 5]) of marine bacteria after 48 h of incubation with Phaeobacter sp.strain 2.10 biofilms established at different densities on plastic petri dishes. Sterile culture medium (10% VNSS) served as a control.
7850 RAO ET AL. APPL. ENVIRON. MICROBIOL.
on August 15, 2013 by U
NIV
OF
ALB
ER
TA
http://aem.asm
.org/D
ownloaded from

ACKNOWLEDGMENTS
This research was supported by a USP-AusAID scholarship, theAustralian Research Council, The Leverhulme Trust, United King-dom, and the Centre for Marine Bio-innovation.
We thank A. Franks for fungal strains, S. Egan for antifoulingmutant strains, N. Tujula for assistance in the field, and N. Paul forhelp with statistics.
REFERENCES
1. Bao, W. Y., C. G. Satuito, J. L. Yang, and H. Kitamura. 2007. Larvalsettlement and metamorphosis of the mussel Mytilus galloprovincialis in re-sponse to biofilms. Mar. Biol. 150:565–574.
2. Bertani, G. 1951. Studies on lysogenesis. I. The mode of phage liberation bylysogenic Escherichia coli. J. Bacteriol. 62:293–300.
3. Boyd, K. G., D. R. Adams, and J. G. Burgess. 1999. Antibacterial andrepellent activities of marine bacteria associated with algal surfaces. Biofoul-ing 14:227–236.
4. Brinkhoff, T., G. Bach, T. Heidorn, L. Liang, A. Schlingloff, and M. Simon.2004. Antibiotic production by a Roseobacter clade-affiliated species from theGerman Wadden Sea and its antagonistic effects on indigenous isolates.Appl. Environ. Microbiol. 70:2560–2565.
5. Bruhn, J. B., L. Gram, and R. Belas. 2007. Production of antibacterialcompounds and biofilm formation by Roseobacter species are influenced byculture conditions. Appl. Environ. Microbiol. 73:442–450.
6. Bruhn, J. B., K. F. Nielsen, M. Hjelm, M. Hansen, J. Bresciani, S. Schulz,and L. Gram. 2005. Ecology, inhibitory activity, and morphogenesis of amarine antagonistic bacterium belonging to the Roseobacter clade. Appl.Environ. Microbiol. 71:7263–7270.
7. Bryan, P. J., J. L. Kreider, and P. Y. Qian. 1998. Settlement of the serpulidpolychaete Hydroides elegans (Haswell) on the arborescent bryozoan Bugulaneritina (L.): evidence of a chemically mediated relationship. J. Exp. Mar.Biol. Ecol. 220:171–190.
8. Buchan, A., J. M. Gonzalez, and M. A. Moran. 2005. Overview of the marineRoseobacter lineage. Appl. Environ. Microbiol. 71:5665–5677.
9. Chin-A-Woeng, T. F., C. D. van den Broek, G. de Voer, K. M. van der Drift,S. Tuinman, J. E. Thomas-Oates, B. J. J. Lugtenberg, and G. V. Bloemberg.2001. Phenazine-1-carboxamide production in the biocontrol strain Pseudo-monas chlororaphis PCL1391 is regulated by multiple factors secreted intothe growth medium. Mol. Plant-Microbe Interact. 14:969–979.
10. Dahms, H. U., S. Dobretsov, and P. Y. Qian. 2004. The effect of bacterial anddiatom biofilms on the settlement of the bryozoan Bugula neritina. J. Exp.Mar. Biol. Ecol. 313:191–209.
11. Dahms, H. U., T. Harder, and P. Y. Qian. 2007. Selective attraction andreproductive performance of a harpacticoid copepod in a response to bio-films. J. Exp. Mar. Biol. 341:228–238.
12. Davis, A. R., N. M. Targett, O. J. McConnel, and C. M. Young. 1989.Epibiosis of marine algae and benthic invertebrates: natural products chem-istry and other mechanisms inhibiting settlement and overgrowth. Bioorg.Mar. Chem. 3:85–114.
13. de Nys, R., P. D. Steinberg, P. Willemsen, S. A. Dworjanyn, C. L. Gabelish,and R. J. King. 1995. Broad spectrum effects of secondary metabolites fromthe red alga Delisea pulchra in antifouling assays. Biofouling 8:259–271.
14. Dobretsov, S., H. U. Dahms, T. Harder, and P. Y. Qian. 2006. Allelochemicaldefense against epibiosis in the macroalga Caulerpa racemosa var. turbinata.Mar. Ecol. Prog. Ser. 318:165–175.
15. Dobretsov, S., H. U. Dahms, and P. Y. Qian. 2006. Inhibition of biofoulingby marine microorganisms and their metabolites. Biofouling 22:43–54.
16. Dobretsov, S. V., and P. Y. Qian. 2002. Effect of bacteria associated with thegreen alga Ulva reticulata on marine micro- and macrofouling. Biofouling18:217–228.
17. Dworjanyn, S. A., R. de Ny, and P. D. Steinberg. 1999. Localisation andsurface quantification of secondary metabolites in the red algae Deliseapulchra. Mar. Biol. 133:727–736.
18. Egan, S., S. James, C. Holmstrom, and S. Kjelleberg. 2002. Correlationbetween pigmentation and antifouling compounds produced byPseudoalteromonas tunicata. Environ. Microbiol. 4:433–442.
19. Egan, S., S. James, C. Holmstrom, and S. Kjelleberg. 2001. Inhibition ofalgal spore germination by the marine bacterium Pseudoalteromonas tu-nicata. FEMS Microbiol. Ecol. 35:67–73.
20. Egan, S., T. Thomas, C. Holmstrom, and S. Kjelleberg. 2000. Phylogeneticrelationship and antifouling activity of bacterial epiphytes from the marinealga Ulva lactuca. Environ. Microbiol. 2:343–347.
21. Franks, A. 2005. Identification and characterisation of an antifungal com-pound produced by the marine bacterium Pseudoalteromonas tunicata. Ph.D.thesis. University of New South Wales, Sydney, Australia.
22. Franks, A., S. Egan, C. Holmstrom, S. James, H. Lappin-Scott, and S.Kjelleberg. 2006. Inhibition of fungal colonization by Pseudoalteromonastunicata provides a competitive advantage during surface colonization. Appl.Environ. Microbiol. 72:6079–6087.
23. Franks, A., P. Haywood, C. Holmstrom, S. Egan, S. Kjelleberg, and N.
Kumar. 2005. Isolation and structure elucidation of a novel yellow pigmentfrom the marine bacterium Pseudoalteromonas tunicata. Molecules 10:1286–1291.
24. Gil-Turness, M. S., and W. Fenical. 1992. Embryos of Homarus americanusare protected by epibiotic bacteria. Biol. Bull. 182:105–108.
25. Gil-Turness, M. S., M. E. Hay, and W. Fenical. 1989. Symbiotic marinebacteria chemically defend crustacean embryos from a pathogenic fungus.Science 246:116–118.
26. Hamer, J. P., G. Walker, and J. W. Latchford. 2001. Settlement of Pomato-ceros lamarkii (Serpulidae) larvae on biofilmed surfaces and the effect ofaerial drying. J. Exp. Mar. Biol. Ecol. 260:113–132.
27. Harder, T., S. C. K. Lau, H. Dahms, and P. Qian. 2002. Isolation of bacterialmetabolites as natural inducers for larval settlement in the marinepolychaete Hydroides elegans (Haswell). J. Chem. Ecol. 28:2029–2043.
28. Harder, T., S. Dobretsov, and P. Qian. 2004. Waterborne polar macromol-ecules act as algal antifoulants in the seaweed Ulva reticulata. Mar. Ecol.Prog. Ser. 274:133–141.
29. Hjelm, M., A. Riaza, F. Formoso, J. Melchiorsen, and L. Gram. 2004.Seasonal incidence of autochthonous antagonistic Roseobacter spp. andVibrionaceae strains in a turbot larva (Scophthalmus maximus) rearing sys-tem. Appl. Environ. Microbiol. 70:7288–7294.
30. Holmstrom, C., S. Egan, A. Franks, S. McCloy, and S. Kjelleberg. 2002.Antifouling activities expressed by marine surface associated Pseudoaltero-monas species. FEMS Microbiol. Ecol. 41:47–58.
31. Holmstrom, C., S. James, S. Egan, and S. Kjelleberg. 1996. Inhibition ofcommon fouling organisms by marine bacterial isolates with special refer-ence to the role of pigmented bacteria. Biofouling 10:251–259.
32. Holmstrom, C., S. James, B. A. Neilan, D. C. White, and S. Kjelleberg. 1998.Pseudoalteromonas tunicata sp. nov., a bacterium that produces antifoulingagents. Int. J. Syst. Bacteriol. 48:1205–1212.
33. Holmstrom, C., D. Rittschof, and S. Kjelleberg. 1992. Inhibition of settle-ment by larvae of Balanus amphitrite and Ciona intestinalis by a surface-colonizing marine bacterium. Appl. Environ. Microbiol. 58:2111–2115.
34. Huang, S. Y., and M. G. Hadfield. 2003. Composition and density of bacterialbiofilms determine larval settlement of the polychaete Hydroides elegans.Mar. Ecol. Prog. Ser. 260:161–172.
35. Hugget, M. 2005. Settlement of generalist marine invertebrate herbivores inresponse to bacterial biofilms. Ph.D. thesis. University of New South Wales,Sydney, Australia.
36. Huggett, M. J., R. de Nys, J. E. Williamson, M. Heasman, and P. D. Stein-berg. 2005. Settlement of larval blacklip abalone, Haliotis rubra, in responseto green and red macroalgae. Mar. Biol. 147:1155–1163.
37. Huggett, M. J., J. E. Williamson, R. de Nys, S. Kjelleberg, and P. D. Stein-berg. 2006. Larval settlement of the common Australian sea urchin Helio-cidaris erythrogramma in response to bacteria from the surface of corallinealgae. Oecologia 149:604–619.
38. James, S. G., C. Holmstrom, and S. Kjelleberg. 1996. Purification and char-acterization of a novel antibacterial protein from the marine bacterium D2.Appl. Environ. Microbiol. 62:2783–2788.
39. Joint, I., M. E. Callow, J. A. Callow, and K. R. Clarke. 2000. The attachmentof Enteromorpha zoospores to a bacterial biofilm assemblage. Biofouling16:151–158.
40. Joint, I., K. Tait, M. E. Callow, J. A. Callow, D. Milton, P. Williams, and M.Camara. 2002. Cell-to-cell communication across the prokaryote-eukaryoteboundary. Science 298:1207.
41. Keats, D. W., M. A. Knight, and C. M. Pueschel. 1997. Antifouling effects ofepithelial shedding in three crustose coralline algae (Rhodophyta, Coralines)on a coral reef. J. Exp. Mar. Biol. Ecol. 213:281–293.
42. Keough, M. J., and P. T. Raimondi. 1996. Responses of settling invertebratelarvae to bioorganic films: effects of large-scale variation in films. J. Exp.Mar. Biol. Ecol. 207:59–78.
43. Lau, S. C. K., and P. Y. Qian. 1997. Phlorotannins and related compoundsas larval settlement inhibitors of the tube-building polychaete Hydroideselegans. Mar. Ecol. Prog. Ser. 159:219–227.
44. Lau, S. C. K., V. Thiyagarajan, S. C. K. Cheung, and P. Y. Qian. 2005. Rolesof bacterial community composition in biofilms as a mediator for larvalsettlement of three marine invertebrates. Aquat. Microb. Ecol. 38:41–51.
45. Laue, R. E., Y. Jiang, S. R. Chhabra, S. Jacob, G. S. A. B. Stewart, A.Hardman, J. A. Downie, F. O’Gara, and P. Williams. 2000. The biocontrolstrain Pseudomonas fluorescens F113 produces the Rhizobium small bacterio-cin, N-(3-hydroxy-7-cis-tetradecenoyl) homoserine lactone, via HdtS, a pu-tative novel N-acyl homoserine lactone synthase. Microbiology 146:2469–2480.
46. Lee, O. O., and P. Y. Qian. 2003. Chemical control of bacterial epibiosis andlarval settlement of Hydroides elegans in the red sponge Mycale adherens.Biofouling 19(Suppl.):171–180.
47. Leitz, T. 1997. Induction and metamorphosis of cnidarian larvae: signals andsignal transduction. Invertebr. Reprod. Dev. 31:109–122.
48. Mai-Prochnow, A., F. Evans, D. Dalisay-Saludes, S. Stelzer, S. Egan, S.James, J. S. Webb, and S. Kjelleberg. 2004. Biofilm development and celldeath in the marine bacterium Pseudoalteromonas tunicata. Appl. Environ.Microbiol. 70:3232–3238.
VOL. 73, 2007 BACTERIAL INHIBITION OF FOULING ORGANISM SETTLEMENT 7851
on August 15, 2013 by U
NIV
OF
ALB
ER
TA
http://aem.asm
.org/D
ownloaded from

49. Maki, J., and R. Mitchell. 1985. Involvement of lectins in the settlement andmetamorphosis of marine invertebrate larvae. Bull. Mar. Sci. 37:675–683.
50. Maki, J. S., D. Rittschof, J. D. Costlow, and R. Mitchell. 1988. Inhibition ofattachment of larval barnacles, Balanus amphitrite, by bacterial surface films.Mar. Biol. 97:199–206.
51. Maki, J. S., D. Rittschof, M.-O. Samuelsson, U. Szewzyk, A. B. Yule, S.Kjelleberg, J. D. Costlow, and R. Mitchell. 1990. Effect of marine bacteriaand their exopolymers on the attachment of barnacle cypris larvae. Bull.Mar. Sci. 46:499–511.
52. Marden, P., A. Tunlid, K. Malmcrona-Friberg, G. Odham, and S. Kjelle-berg. 1985. Physiological and morphological changes during short term star-vation of marine bacteria isolates. Arch. Microbiol. 142:326–332.
53. Negri, A. P., N. S. Webster, R. T. Hill, and A. J. Heyward. 2001. Metamor-phosis of broadcast spawning corals in response to bacteria isolated fromcrustose algae. Mar. Ecol. Prog. Ser. 223:121–131.
54. Olivier, F., R. Tremblay, E. Bourget, and D. Rittschof. 2000. Barnacle set-tlement: field experiments on the influence of larval supply, tidal level,biofilm quality and age on Balanus amphitrite cyprids. Mar. Ecol. Prog. Ser.199:185–204.
55. Patel, P., M. E. Callow, I. Joint, and J. A. Callow. 2003. Specificity in thesettlement-modifying response of bacterial biofilms towards zoospores of themarine alga Enteromorpha. Environ. Microbiol. 5:338–349.
56. Piper, K. R., S. B. Vonbodman, and S. K. Farrand. 1993. Conjugation factorof Agrobacterium tumefaciens regulates Ti plasmid transfer by autoinduction.Nature 362:448–450.
57. Rahim, S., J. Y. Li, and H. Kitamura. 2004. Larval metamorphosis of the seaurchins Pseudocentrotus depressus and Anthocidaris crassispina in response tomicrobial films. Mar. Biol. 144:71–78.
58. Rao, D., J. S. Webb, and S. Kjelleberg. 2005. Competitive interactions inmixed-species biofilms containing the marine bacterium Pseudoalteromonastunicata. Appl. Environ. Microbiol. 71:1729–1736.
59. Rao, D., J. S. Webb, and S. Kjelleberg. 2006. Microbial colonization andcompetition on the marine alga Ulva australis. Appl. Environ. Microbiol.72:5547–5555.
60. Ravn, L., A. B. Christensen, S. Molin, M. Givskov, and L. Gram. 2001.Methods for detecting acylated homoserine lactones produced by Gram-negative bacteria and their application in studies of AHL-production kinet-ics. J. Microbiol. Methods 44:239–251.
61. Ruiz-Ponte, C., V. Cilia, C. Lambert, and J. L. Nicolas. 1998. Roseobacter
gallaeciensis sp. nov., a new marine bacterium isolated from rearings andcollectors of the scallop Pecten maximus. Int. J. Syst. Bacteriol. 48:537–542.
62. Ruiz-Ponte, C., J. F. Samain, J. L. Sanchez, and J. L. Nicolas. 1999. Thebenefit of a Roseobacter species on the survival of scallop larvae. Mar.Biotechnol. 1:52–59.
63. Shaw, P. D., G. Ping, S. L. Daly, C. Cha, J. E. Cronan, K. L. Rinehart, andS. K. Farrand. 1997. Detecting and characterizing N-acyl-homoserine lac-tone signal molecules by thin-layer chromatography. Proc. Natl. Acad. Sci.USA 94:6036–6041.
64. Shikuma, N. J., and M. G. Hadfield. 2005. Temporal variation of an initialmarine biofilm community and its effect on larval settlement and metamor-phosis of the tubeworm Hydroides elegans. Biofilms 2:231–238.
65. Skovhus, T. L. 2004. Occurrence and function of marine antifoulingPseudoalteromonas species. Ph.D. thesis. University of Aarhus, Aarhus, Den-mark.
66. Skovhus, T. L., N. B. Ramsing, C. Holmstrom, S. Kjelleberg, and I. Dahllof.2004. Real-time quantitative PCR for assessment of abundance ofPseudoalteromonas species in marine samples. Appl. Environ. Microbiol.70:2373–2382.
67. Steinberg, P. D., R. de Nys, and S. Kjelleberg. 2002. Chemical cues forsurface colonisation. J. Chem. Ecol. 28:1935–1951.
68. Steinberg, P. D., R. Schneider, and S. Kjelleberg. 1997. Chemical defences ofseaweeds against microbial colonisation. Biodegradation 8:211–220.
69. Szewzyk, U., C. Holmstrom, M. Wrangstadh, M.-O. Samuelsson, J. S. Maki,and S. Kjelleberg. 1991. Relevance of the exopolysaccharide of marinePseudomonas sp. strain S9 for the attachment of Ciona intestinalis larvae.Mar. Ecol. Prog. Ser. 75:259–265.
70. Tait, K., I. Joint, M. Daykin, D. L. Milton, P. Williams, and M. Camara.2005. Disruption of quorum sensing in seawater abolishes attraction ofzoospores of the green alga Ulva to bacterial biofilms. Environ. Microbiol.7:229–240.
71. Wagner-Dobler, I., V. Thiel, L. Eberl, M. Allgaier, A. Bodor, S. Meyer, S.Ebner, A. Hennig, R. Pukall, and S. Schulz. 2005. Discovery of complexmixtures of novel long-chain quorum sensing signals in free-living and host-associated marine alphaproteobacteria. ChemBioChem 6:2195–2206.
72. Wahl, M. 1989. Marine epibiosis. 1. Fouling and antifouling: some basicaspects. Mar. Ecol. Prog. Ser. 58:175–189.
73. Wieczorek, S. K., and C. D. Todd. 1998. Inhibition and facilitation of settle-ment of epifaunal marine invertebrate larvae by microbial biofilm cues.Biofouling 12:81–118.
7852 RAO ET AL. APPL. ENVIRON. MICROBIOL.
on August 15, 2013 by U
NIV
OF
ALB
ER
TA
http://aem.asm
.org/D
ownloaded from




















