
Travels and adventures of an orchid hunter. An account of ...
Upload
independentCategory
view
5download
0
Environmental and Experimental Botany 41 (1999) 219–230
Changes in photosynthetic capability and carbohydrateproduction in an epiphytic CAM orchid plantlet exposed to
super-elevated CO2
Sok Siam Gouk a, Jie He b, Choy Sin Hew a,*a Department of Biological Sciences, National Uni6ersity of Singapore, Lower Kent Ridge Road, Singapore 119260, Singapore
b School of Science, National Institute of Education, Nanyang Technological Uni6ersity, 469 Bukit Timah Road,Singapore 259756, Singapore
Received 24 September 1998; received in revised form 20 January 1999; accepted 22 January 1999
Abstract
The effects on growth in super-elevated (1%) CO2 in terms of photosynthetic capability and carbohydrateproduction were studied in an epiphytic CAM (Crassulacean acid metabolism) orchid plantlet, Mokara Yellow(Arachnis hookeriana×Ascocenda Madame Kenny). The growth of the plantlets was greatly enhanced after growingfor 3 months at 1% CO2 compared with the control plantlets (0.035% CO2). CO2 enrichment produced more than a2-fold increase in dry matter production. The enhanced root growth at 1% CO2 led to a higher root:shoot ratio.Plantlets grown at super-elevated CO2 had higher Fv/Fm values, a higher photochemical quenching (qP) and arelatively lower non-photochemical quenching (qN). CO2 at 1% appeared to enhance the utilization of captured lightenergy in the orchid plantlets. CO2 enrichment also increased contents of soluble sugars (glucose and sucrose) andstarch in the orchid plantlets. The extra starch formed under 1% CO2 did not cause a disruption of the chloroplasts.Chlorophyll content was higher and a clear granal stacking was evident in young leaves and roots of plantlets grownat 1% CO2. An extensive thylakoid system was observed in the young leaf chloroplasts of the CO2-enriched plantletsindicating an improved development of the photosynthetic apparatus when compared to that of the control plantlets.The increased photosynthetic capacity and enhanced growth of the epiphytic roots under CO2 enrichment wouldfacilitate the generation of more photoassimilates and acquisition of essential resources, thereby increasing thesurvival rate of orchid plantlets under stressful field conditions. © 1999 Elsevier Science B.V. All rights reserved.
Keywords: Arachnis ; Carbohydrate production; Chlorophyll fluorescence; Chloroplast ultrastructure; CO2 enrichment; Epiphyte;Mokara Yellow
1. Introduction
Plantlets growing in vitro are considered tohave low photosynthetic ability which fails toprovide a positive carbon balance. Sugar is there-
* Corresponding author. Tel.: +65-8742713; fax: +65-7792486.
E-mail address: [email protected] (C.S. Hew)
S0098-8472/99/$ - see front matter © 1999 Elsevier Science B.V. All rights reserved.
PII: S0098 -8472 (99 )00006 -4

S.S. Gouk et al. / En6ironmental and Experimental Botany 41 (1999) 219–230220
fore required as a carbon and energy source fortheir mixotrophic growth (Grout and Ashton,1978). Enhancing photosynthetic ability of invitro plantlets would favor the success of theiracclimatization to ex vitro conditions (Kozai etal., 1990; Kozai, 1991). Recent research has re-vealed that the development of photosyntheticcharacteristics of in vitro plantlets could be im-proved by altering their environmental growthconditions, such as reducing the sucrose level inthe medium or increasing the light intensity orCO2 concentration (Kozai, 1991; Hdider and Des-jardins, 1994).
The absorption of excessive light energy byleaves has been recognized as a physiologicalstress not only in the field but also in vitro(Aitken-Christie et al., 1992). During conven-tional closed system of micropropagation, lowCO2 concentration occurs just a few hours afterthe onset of photoperiod (Fujiwara et al., 1987).This low CO2 availability in culture vessels maystimulate photoinhibition in the in vitro plantlets(Serret et al., 1996). It has been demonstrated thatan increase in the atmospheric CO2 concentrationduring growth would enhance the utilization ofcaptured light energy (Habash et al., 1995),thereby reducing the susceptibility to photoinhibi-tion in plants at elevated CO2. However, there islimited knowledge on the effect of CO2 enrich-ment on the susceptibility to light in in vitroplantlets, particularly with orchid plantlets grow-ing under limiting CO2 concentration.
Long-term exposure to elevated CO2 concentra-tion has been demonstrated to cause down-regula-tion of net photosynthesis in many plant species(Gunderson and Wullschleger, 1994). It has beensuggested that the photosynthetic down-regula-tion in plants growing at increased CO2 concen-trations was due to end-product inhibition bycarbohydrate accumulation as a result of changesin the carbohydrate source: sink inter-relation-ships (Stitt, 1991; Woodrow, 1994), and/or a rede-ployment of nitrogen (N) away from the CO2
fixation machinery (especially Rubisco) into sys-tems to acquire other resources (Besford et al.,1990; Bowes, 1991; Drake et al., 1997). The abilityof the plants to sustain an adequate sink activityhas been invoked as the main process that can
regulate the acclimation responses of plants toelevated CO2 (Stitt, 1991). The feedback inhibi-tion by carbohydrate production on photosynthe-sis is often related to chloroplast disruption. Caveet al. (1981) had reported a disruption of chloro-plast function by large starch granules.
In previous reports, we had demonstrated thatthe growth of epiphytic CAM orchid plantlets,Mokara White (Hew et al., 1995) and MokaraYellow (Gouk et al., 1997), was greatly enhancedunder super-elevated (1%) CO2. We hypothesizethat super-elevated CO2 would enhance the uti-lization of captured light energy, thereby enhancegrowth of orchid plantlets, particularly that of theroots. Enhanced photosynthetic capability inroots would lead to the generation of more pho-toassimilates. This study is of considerable valueto the orchid industry because enhanced rootgrowth would not only promote resource acquisi-tion but also reduce mortality rate of orchidplantlets when they are transferred from the cul-ture flasks to the field. In this study we examinethe effects of CO2 enrichment on the efficiency ofphotosystem II (PSII) and the chloroplast ultra-structure by the accumulation of nonstructuralcarbohydrates in Mokara Yellow plantlets.
2. Materials and methods
2.1. Plant material and treatments
The experiments were conducted on MokaraYellow (Arachnis hookeriana (Rchb. f.)×Asco-cenda Madame Kenny) plantlets, an epiphyticCAM orchid. The details of the experiment wasreported earlier (Hew et al., 1995; Gouk et al.,1997), and the system set-up was described brieflyas follows. Healthy plantlets (0.8–1.0 g) wereselected and placed in GA-7 culture vessels (Ma-genta™, 3×3×4¦, Sigma, St. Louis, MO) whichwere then arranged in transparent zipped plasticbags (540×380 mm, KH Plastics, Singapore).The GA-7 culture vessel was modified by drillinga hole (2 mm in diameter) at the center of the lidto allow gaseous diffusion. Each GA-7 culturevessel contained 5 ml of modified Vacin and Wentculture solution (Hew et al., 1995) where tryptone,

S.S. Gouk et al. / En6ironmental and Experimental Botany 41 (1999) 219–230 221
NAA, kinetin, sucrose and agar were excluded.Different bags were flushed with 0.035% (control)or 1% CO2 until fully inflated. CO2 was obtainedfrom gas tanks prepared commercially by Na-tional Oxygen (Singapore). Natural diffusion wasallowed to take place for 2 days before the nextflushing was carried out. Preliminary studies haveshown that similar levels of CO2 could be main-tained within the culture vessels by flushing every2 days (Hew et al., 1995). The plantlets weregrown for 3 months under the following condi-tions: 12 h photoperiod daily; temperature of2694°C; and photosynthetic photon flux densityof 200 mmol m−2 s−1 (Osram 400 W lamp).
2.2. Growth and carbohydrate measurements
After a 3-month culture period, 12 plantletswere harvested. Old leaves (OL), young leaves(YL) and roots (R) were separated, the freshweights (FW) of the tissues were measured. Leafarea was measured using a leaf area analyzer(Skye Instrument, UK). The dry weights (DW) ofplant tissues were determined after drying for 1week in an oven set at 70°C. Relative growth rate(RGR) was estimated using the formula,(ln W2− ln W1)/(t2− t1), where W1 and W2 areDW at time t1 and t2, respectively (Beadle, 1993).Specific leaf weight (SLW) was calculated as theratio between the DW and the area of the leaves(Wong, 1990). Leaf area ratio (LAR) was theratio of the leaf area per unit total DW (Beadle,1993). An estimate of leaf density was determinedby the ratio of leaf DW to leaf FW (Dijkstra,1989). Chlorophyll (Chl) content was determinedaccording to Moran (1982).
Nonstructural carbohydrates in plant tissueswere determined on samples harvested at 16:00 hlocal time. Soluble sugars (glucose and sucrose) inleaves and roots were extracted three times (2, 2and 18 h), each in 3 ml hot 80% ethanol (55°C).The extracts were pooled and evaporated to dry-ness in a vacuum evaporator (Vacucenter, Salvis,Switzerland). The residues were redissolved in 0.2ml distilled water and spun at 13 000 rpm at 4°Cfor 15 min. The clear supernatant was then usedfor nonstructural carbohydrate determination.The glucose content in 10 ml supernatant was
determined using glucose kit 115-A (Sigma). Su-crose content was determined as described byFarrar (1993) with slight modification. Sucrose in0.1 ml supernatant was hydrolyzed by adding 1ml 0.1 M citrate buffer (pH 4.6), followed by 24U invertase (EC 3.2.1.26, Sigma). The reactionmixture was incubated at 55°C for 18 h. Theglucose content was then measured using the glu-cose kit and the initial glucose content was sub-tracted. The sucrose content in the respectivesupernatant was then calculated. Starch in theremaining pellet was hydrolyzed for 3 h with 1 ml3% HCl in a boiling bath. The glucose content ofthe digest was assayed using the glucose kit.
2.3. In 6i6o measurements of chlorophyll afluorescence
Chl a fluorescence of attached leaves and rootswas measured at 23–25°C with a pulse amplitudemodulation fluorometer (PAM 2000, Walz, Effel-trich, Germany). The plantlets were kept in dark-ness for 15 min prior to the measurements. Theleaf or root was initially exposed to the weakmodulated measuring beam (0.1 mmol quantam−2 s−1) to estimate the minimal yield of Chlfluorescence of the dark-adapted plant tissue (Fo),when all the PSII reaction centers were open.Then, the respective plant part was exposed to a0.8 s saturation pulse of white light (3000 mmolm−2 s−1) to close the PSII reaction centers, atwhich point the maximal yield of Chl fluorescenceof the dark-adapted plant tissue (Fm) was as-sessed. The variable fluorescence yield, Fv, wasdetermined by Fm−Fo. The efficiency of excita-tion energy captured by open PSII reaction cen-ters in dark-adapted plant part was estimated bythe fluorescence ratio (Fv/Fm).
After the saturation pulse, continuous actiniclight (about 170 mmol m−2 s−1) was applied todetermine the steady-state fluorescence yield, Ft.Once Ft was obtained, a 0.8-s saturation pulse wasgiven to obtain estimates of Fm% . For Fo% -determi-nation, far-red illumination was activated aftertransient darkening (i.e. turning off 170 mmolm−2 s−1). The photochemical quenching (qP) andnon-photochemical quenching (qN) were calcu-lated on the basis of Fo% by using the data acquisi-
S.S. Gouk et al. / En6ironmental and Experimental Botany 41 (1999) 219–230222
tion software DA-2000 (Walz). qP denoted thefraction of open PSII reaction centers, whereas qN
represented a measure of non-radiative dissipationof the light energy.
2.4. Transmission electron microscopy (TEM)
Plant tissues were excised from the middle por-tion of leaf blades (avoiding the midrib) or rootsharvested between 09:30 and 11:00 h. Small seg-ments were excised under 2.5% glutaraldehyde in0.2 M phosphate buffer (pH 7.2). The segmentswere fixed in the above fixative buffer for 4 h atroom temperature (25°C). The tissue segmentswere then post-fixed in 1% osmium tetroxide in0.1 M phosphate buffer for 1.5 h. Post-fixed tissuesegments were rinsed in phosphate buffer anddehydrated in a graded ethanol series followed bytwo changes of absolute acetone. The tissue seg-ments were then embedded in Spurr resin, SPR-LV (Sigma). Ultrathin sections were cut using anultramicrotome (Reicherd, Austria) with a glassknife. Sections were collected on 200-mesh coppergrids and stained in uranyl acetate (55 min), fol-lowed by lead citrate (8–12 min) and subse-
quently viewed in a transmission electronmicroscope (Philips CM10) at 60 kV. Three repli-cates were used for each treatment and three tofive grids for each plant part were viewed.
2.5. Data analysis
The statistical differences between the CO2
treatments were studied using Minitab StatisticalSoftware Release 8.1 (Minitab, State College, PA,USA). Data were subjected to one-way analysis ofvariance (ANOVA) and presented as means9standard errors (SE).
3. Results
Growth of Mokara Yellow plantlets wasgreatly enhanced under super-elevated CO2 (Table1). Total DW was more than 2-fold that of thecontrol plantlets, and this was attributed to asignificant increase in the DW of young leaves(YL) and roots (R) (PB0.001). The enhancedroot growth accounted for the 70% higherroot:shoot ratio at 1% CO2 compared with the
Table 1Biomass, dry matter partitioning, and growth characteristics of Mokara Yellow plantlets grown for 3 months at ambient (0.035%)or super-elevated (1%) CO2
a
Growth characteristics 1% CO20.035% CO2
Dry wt (mg)48.5 (1.9)Old leaf (OL) 48.2 (3.5) ns
**20.9 (1.3)Young leaf (YL) 80.9 (5.4)22.8 (2.0)Root (R) 71.6 (3.8) **
201 (8)Total **92 (3)0.56 (0.03)Root: shoot ratio **0.33 (0.03)
**0.59 (0.01)Relati6e growth rate (mg g−1 week−1) 0.56 (0.01)Leaf area (mm2)
1193 (37)OL 1140 (63) ns**1171 (60)YL 595 (39)
19.5 (0.5)Leaf area ratio, LAR (mm2 mg−1) 11.6 (0.3) **Specific leaf weight, SLW (mg cm−2)
4.06 (0.07)OL 4.18 (0.13) nsYL 3.54 (0.09) 6.86 (0.18) **Leaf density, DW/FWOL 0.074 (0.001) 0.081 (0.001) *
0.067 (0.002) 0.093 (0.001)YL **
a Results are means with SE in parentheses (n=12). ns, * and ** mean, respectively, non-significant, PB0.005 and PB0.001 fordifference between plantlets grown in ambient and super-elevated CO2.

S.S. Gouk et al. / En6ironmental and Experimental Botany 41 (1999) 219–230 223
Table 2Chlorophyll (Chl) content in leaves and roots of Mokara Yellow plantlets grown for 3 months at ambient (0.035%) or super-elevated(1%) CO2
a
Old leaf Young leaf Root0.035% CO2 1% CO21% CO2 0.035% CO20.035% CO2 1% CO2
* 86 (8) 157 (14)Chl a 259 (10) 311 (23) ns 258 (26) **416 (43)**94 (11)39 (4)Chl b 110 (3) *129 (12) ns 105 (12) 181 (18)
** 2.17 (0.03) 1.92 (0.04)Chl a/b **2.36 (0.02) 2.44 (0.05) ns 2.45 (0.01) 2.30 (0.02)**273 (26)125 (12)Total Chl 369 (14) *440 (36) ns 363 (42) 597 (61)
a Chl a (mg g FW−1), Chl b (mg g FW−1), Chl a/b ratio, and total Chl (mg g FW−1) are means of three to four replicates withSE in parentheses. ns, * and ** mean, respectively, non-significant, PB0.05 and PB0.005 for difference between plantlets grown inambient and super-elevated CO2.
control. CO2 enrichment not only enhanced thegrowth of existing roots, it also induced rootinitiation from the internodes of Mokara Yellowplantlets (Figure not shown). The RGR for theCO2-enriched plantlets was 5% higher (PB0.001).Under CO2 enrichment, SLW for YL was 94%greater than that of ambient CO2 (Table 1, PB0.001). The density (DW/FW ratio) of the CO2-enriched old leaves (OL) and YL increased by 9and 39%, respectively (PB0.005). On the otherhand, LAR in Mokara Yellow plantlets grown at1% CO2 decreased compared to that at the ambi-ent CO2 (Table 1, PB0.001).
An increase of the CO2 concentration resultedin an increase in Chl contents in YL and Rcompared with the control (Table 2). The increaseranged from 61 to 141% (PB0.05 for YL andPB0.005 for R). Despite the increase in Chlcontent, there was a slight decline in the Chl a/bratio for both the YL (6%) and R (12%).
The Fv/Fm values of the leaves were consistentlyhigher throughout the day for plantlets grown atsuper-elevated CO2 compared to those grown atambient CO2. In OL, Fv/Fm was 0.76890.004 at1% CO2 and 0.71590.004 at 0.035% CO2, whilein YL, the values were 0.78890.008 and 0.73590.003, respectively. In general, YL had higherFv/Fm values than OL. An increase in CO2 con-centration also increased Fv/Fm values for R, i.e.0.69 at 1% CO2 compared to 0.60 at 0.035% CO2
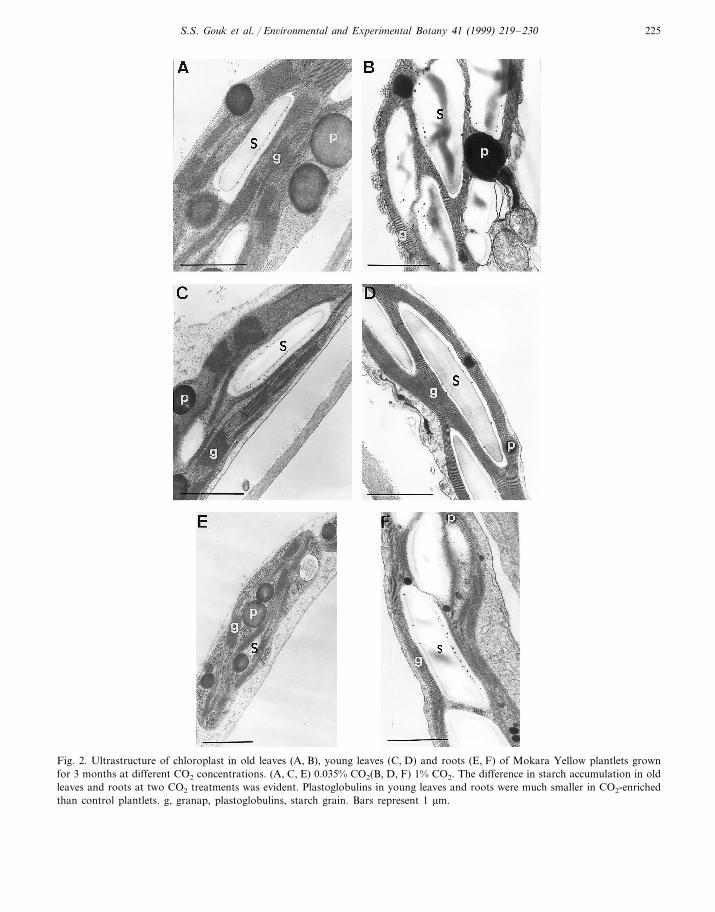
(PB0.001).The diurnal time course studies of qP and qN in
leaves showed a similar trend in both OL and YL(Fig. 1). Plantlets grown for 3 months at super-el-evated CO2 sustained a higher qP (about 0.74).
For the control plantlets, qP started to decreaseafter 13:00 h (average of 0.66) with a concomitantincrease in qN (around 0.81 at 19:00 h). qN wasmaintained at lower values (average of 0.63) at1% CO2 throughout the day, compared to thecontrol plantlets (about 0.75).
In general, more carbohydrate accumulated inplant tissues as growth CO2 increased from 0.035to 1% (Table 3). Super-elevated CO2 caused adrastic increase in starch content in R, about20-fold higher than in the control (PB0.01). Thisstarch level in the roots (8.0 mg glucose equivalentg FW−1) was comparable to that found in OL ofCO2-enriched plantlets. For plantlets grown atambient CO2, the starch level in R was onlyone-tenth that of OL. The starch content in YLwas not affected by elevated growth CO2. Thesucrose level was 6–13-fold higher (PB0.01) inthe CO2-enriched plantlets, with the highest in-crease found in R (4.5 mg g FW−1). Under 1%CO2, the YL had accumulated the highest amountof glucose (2.07 mg g FW−1), nearly 40-foldhigher compared with the respective counterpartat ambient CO2. This value was two to five timeshigher than OL and R of CO2-enriched plantlets.
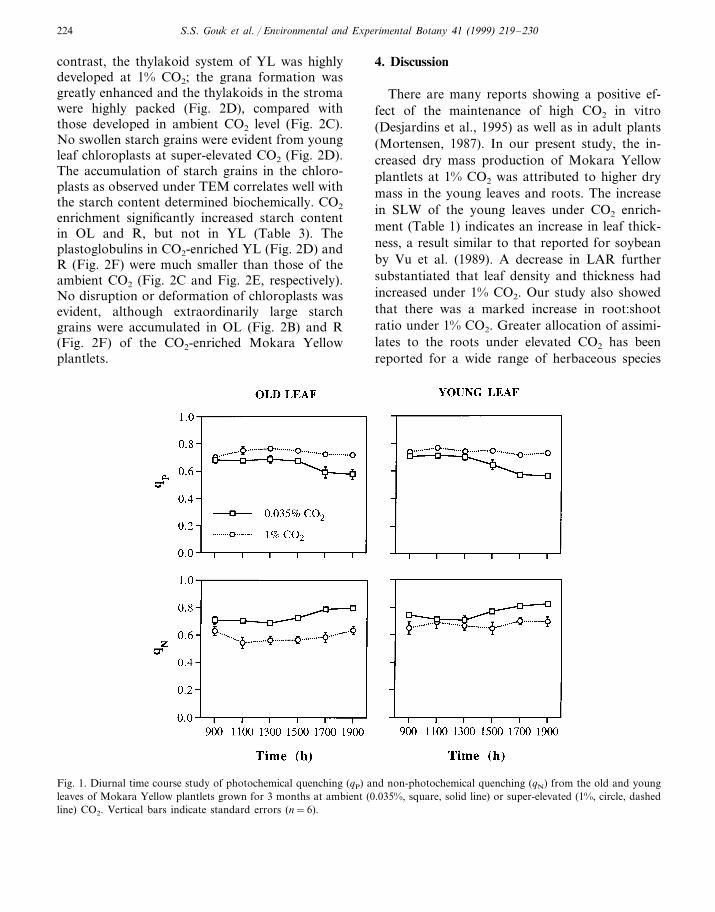
For TEM study of orchid plantlets, only somerepresentative data are shown (Fig. 2) as theelectronmicrographs (EMs) of the replicates weresimilar. The mesophyll cells from different plantparts showed normal chloroplasts from both CO2
treatments, although the relative development ofgrana as well as the accumulation of starchvaried. There was a reduction in the grana forma-tion in OL and R at high CO2 level (Fig. 2B, F) ascompared to that of ambient CO2 (Fig. 2A, E). In
S.S. Gouk et al. / En6ironmental and Experimental Botany 41 (1999) 219–230224
contrast, the thylakoid system of YL was highlydeveloped at 1% CO2; the grana formation wasgreatly enhanced and the thylakoids in the stromawere highly packed (Fig. 2D), compared withthose developed in ambient CO2 level (Fig. 2C).No swollen starch grains were evident from youngleaf chloroplasts at super-elevated CO2 (Fig. 2D).The accumulation of starch grains in the chloro-plasts as observed under TEM correlates well withthe starch content determined biochemically. CO2
enrichment significantly increased starch contentin OL and R, but not in YL (Table 3). Theplastoglobulins in CO2-enriched YL (Fig. 2D) andR (Fig. 2F) were much smaller than those of theambient CO2 (Fig. 2C and Fig. 2E, respectively).No disruption or deformation of chloroplasts wasevident, although extraordinarily large starchgrains were accumulated in OL (Fig. 2B) and R(Fig. 2F) of the CO2-enriched Mokara Yellowplantlets.
4. Discussion
There are many reports showing a positive ef-fect of the maintenance of high CO2 in vitro(Desjardins et al., 1995) as well as in adult plants(Mortensen, 1987). In our present study, the in-creased dry mass production of Mokara Yellowplantlets at 1% CO2 was attributed to higher drymass in the young leaves and roots. The increasein SLW of the young leaves under CO2 enrich-ment (Table 1) indicates an increase in leaf thick-ness, a result similar to that reported for soybeanby Vu et al. (1989). A decrease in LAR furthersubstantiated that leaf density and thickness hadincreased under 1% CO2. Our study also showedthat there was a marked increase in root:shootratio under 1% CO2. Greater allocation of assimi-lates to the roots under elevated CO2 has beenreported for a wide range of herbaceous species
Fig. 1. Diurnal time course study of photochemical quenching (qP) and non-photochemical quenching (qN) from the old and youngleaves of Mokara Yellow plantlets grown for 3 months at ambient (0.035%, square, solid line) or super-elevated (1%, circle, dashedline) CO2. Vertical bars indicate standard errors (n=6).
S.S. Gouk et al. / En6ironmental and Experimental Botany 41 (1999) 219–230 225
Fig. 2. Ultrastructure of chloroplast in old leaves (A, B), young leaves (C, D) and roots (E, F) of Mokara Yellow plantlets grownfor 3 months at different CO2 concentrations. (A, C, E) 0.035% CO2(B, D, F) 1% CO2. The difference in starch accumulation in oldleaves and roots at two CO2 treatments was evident. Plastoglobulins in young leaves and roots were much smaller in CO2-enrichedthan control plantlets. g, granap, plastoglobulins, starch grain. Bars represent 1 mm.

S.S. Gouk et al. / En6ironmental and Experimental Botany 41 (1999) 219–230226
Table 3Carbohydrate content of Mokara Yellow plantlets grown for 3 months at ambient (0.035%) or super-elevated (1%) CO2
a
Young leaf RootOld leaf0.035%CO20.035%CO2 1% CO2 1% CO20.035%CO2 1% CO2
0.06 (0.001) 0.36 (0.06) **Glucose 0.05 (0.003) 0.62 (0.21) * 0.05 (0.005) 2.07 (0.52) ****Sucrose 0.71 (0.04) 5.73 (0.74) ** 0.75 (0.07) 5.27 (0.81) ** 0.32 (0.08) 4.50 (0.44)
0.40 (0.07) 8.04 (2.97) *Starch 3.18 (0.51) 7.04 (0.51) ** 2.39 (0.54) 2.14 (0.34) ns
a Glucose (mg g FW−1), sucrose (mg g FW−1) and starch (mg Glu equ g FW−1) are means with SE in parentheses (n=4). ns,* and ** mean, respectively, non-significant, PB0.05 and PB0.01 for difference between plantlets grown in ambient andsuper-elevated CO2.
(Farrar and Williams, 1991; Prior et al., 1994;Rogers et al., 1994). Many aspects of root re-sponse to elevated CO2 have been discussed(Norby, 1994; Rogers et al., 1994). Very littleattention, however, was focused on the responseof epiphytic roots to elevated CO2 (Hew et al.,1995; Gouk et al., 1997). Epiphytic roots nor-mally have poor access to nutrients (Benzing etal., 1983; Pridgeon, 1987) and slow rate of min-eral uptake (Hew and Yong, 1997). Nevertheless,photosynthesis by the aerial roots of epiphyticorchids in contributing to the carbon economy ofthe whole plant is well documented (Benzing etal., 1983; Hew and Yong, 1997). The increase inphotosynthetic capability of the aerial roots ofepiphytic Mokara Yellow plantlets under super-elevated CO2 was indicated by an increase in Chlcontent and higher Fv/Fm values. Enhancedgrowth of the in vitro plantlets of Mokara Yellowas a result of CO2 enrichment has also proven tobe particularly beneficial during acclimatization toex vitro conditions (Lakso et al., 1986; Desjardinset al., 1987). An increase in root: shoot ratio ofEucalyptus plantlets correlated well with an in-crease in net photosynthetic rate suggesting thatthe development of root systems could be pro-moted by an increase in net leaf photosyntheticrate and translocation of photoassimilates toroots (Kirdmanee et al., 1995). It is likely thatwhen the CO2-enriched in vitro plantlets aretransferred to the field or greenhouse, the epi-phytic roots, with enhanced photosynthetic capac-ity, could generate more photoassimilates.Furthermore, an increased proportion of biomassdistribution to the root system may result in
increased acquisition of essential resources (Prioret al., 1994; Rogers et al., 1994). As a whole, thiswould enable the plantlets to adapt to a morestressful environment with reduced mortality rateand eventually leading to enhanced growth andyield. Further investigation into this area wouldbe of great significance to the orchid industry interms of yield production.
The ratio of variable fluorescence to maximalchlorophyll fluorescence (Fv/Fm) is an excellentindicator for the PSII quantum yield; reduction inFv/Fm might be associated with thermal damageof PSII reaction centers (Demmig and Bjorkman,1987; Weis and Berry, 1987). In our study, higherFv/Fm values were evident in CO2-enrichedMokara Yellow plantlets relative to that of thecontrol plantlets. Two possibilities could accountfor this; high CO2 enhanced the high absorptionrate of the leaves, affecting Fo, and/or increasedthe energy cycling between the reaction centersand the chlorophyll pool, affecting Fm (Havaux etal., 1991). An increase in quantum yield of PSIIhas been reported for Triticum aesti6um (Habashet al., 1995) and Scirpus olneyi (Long and Drake,1991) grown at elevated CO2. Higher Fv/Fm valuesin the Mokara Yellow plantlets grown at 1% CO2
are associated with an increase in Chl a and b(especially the young leaves and roots). Since bothChl a and b are associated with the light-harvest-ing complexes of PSII, an increase in these pig-ments would strongly suggest that 1% CO2 didnot impair the core complexes of PSII (vonWillert et al., 1995). Our findings do not corre-spond with that reported by Keutgen et al. (1997).They reported a reduction in Fv/Fm values on

S.S. Gouk et al. / En6ironmental and Experimental Botany 41 (1999) 219–230 227
strawberry grown at elevated CO2, with a con-comitant decline in Chl a, b and a/b ratio.
Excess light energy beyond that usable for pho-tosynthesis can lead to photoinhibition (Powles,1984). The orchid plantlets grown at ambient CO2
showed susceptibility to photoinhibition asreflected in a decline in Fv/Fm values and a con-comitant lower qP with consistently higher qN
(Fig. 1). During micropropagation, low CO2 con-centration inside the culture vessels may be at-tained a few hours after the onset of photoperiod.This may lead to photoinhibition (Solarova, 1989;Kozai, 1991). Reduced CO2 availability could af-fect electron transport by limiting consumption ofassimilatory power (Peterson, 1990). In the con-trol plantlets, a sharp decrease in qP with a con-comitant increase in qN were observed in theyoung leaves. On the other hand, because of highCO2 availability, plantlets grown at 1% CO2 couldutilize a greater proportion of the radiant energyand dissipated only a small proportion, mani-fested by a higher qP and a concomitant loweringof qN. From our study, it seems probable that thepossible photoinhibitory effect on in vitroplantlets could be alleviated by increasing theatmospheric CO2 concentration. Studies onpineapple also indicated a 45% lower qN forplants grown at elevated CO2 than those at ambi-ent CO2 (Zhu et al., 1997).
Various anatomical and morphological re-sponses to CO2 enrichment have been docu-mented (Allen, 1990). Cave et al. (1981) reportedan abnormal chloroplast structure and a disturbedconfiguration of granal stacking as a result ofexcessive accumulation of starch grains in Tri-folium subterraneum grown in elevated CO2. Wulffand Strain (1982) observed a reduction in thephotosynthetic rates of Desmodium paniculatumexposed to elevated CO2 level. They correlatedthis to increased starch accumulation and reducedgrana formation. The orchid plantlets grown un-der 1% CO2 did not display any morphologicalabnormality or chlorotic development of leaf tis-sues as reported by others (Madsen, 1973; Cave etal., 1981). From our TEM study, it appears thatthere was no disruption of chloroplasts from dif-ferent plant parts of the orchid plantlets whengrown for 3 months under super-elevated CO2.
No dilation or disorganization of thylakoid wasevident either. Although slightly swollen chloro-plasts were observed in CO2-enriched old leavesand roots as a result of starch accumulation, theirphotosynthetic apparatus did not seem to be af-fected. This is supported by the higher Fv/Fm
values and Chl contents in these tissues. Compari-son of grana to assess membrane stacking charac-teristics has been reported (Schiffgens-Gruber andLutz, 1992; Pritchard et al., 1997); however, wewere unable to quantify this because of excessiveaccumulation of starch grains in the orchidplantlets grown at 1% CO2. Bockers et al. (1997)have observed changes in the shape of chloro-plasts in photoautotrophic Marchantia polymor-pha culture cells maintained at 2% CO2. However,this did not affect its photosynthetic performance.In the chloroplasts of young leaves of CO2-en-riched Mokara Yellow plantlets, we observed anextensive granal stacking and more thylakoids inthe stroma. This could partly account for thehigher Chl content and Fv/Fm values in youngleaves of CO2-enriched orchid plantlets. This mayalso explain why the young leaves developedfaster and better when the plantlets were grown at1% CO2.
Orchid plantlets grown at 1% CO2 exhibitedmuch smaller plastoglobulins in young leaves androots than those at ambient CO2. Our results donot correspond with that reported by Pritchard etal. (1997) for the coniferous tree Pinus palustris.They reported a disruption of chloroplast in-tegrity accompanied by increased number of plas-toglobulins and shorter grana in pine seedlingsgrown at elevated CO2. The presence of largeplastoglobulins in the chloroplasts of MokaraYellow plantlets grown at ambient CO2 could beindicative of photoinhibitory stress as shown forGardenia plantlets (Serret et al., 1996).
Increased levels of leaf starch for plants grownin elevated CO2 are often associated with insuffi-cient sink demand (Stitt, 1991) and down-regula-tion of photosynthesis (Tissue et al., 1993; Rodenand Ball, 1996). Under CO2 enrichment, the su-crose and starch contents in the leaves and rootsof Mokara Yellow plantlets increased in tandem.The starch content in the young leaves of CO2-en-riched Mokara Yellow plantlets was much lower

S.S. Gouk et al. / En6ironmental and Experimental Botany 41 (1999) 219–230228
than that in the old leaves and the roots. Since theyoung leaves are still expanding, carbohydratesare being utilized for growth as well as mainte-nance. The amount of carbohydrates accumulatedin the roots was comparable to that in the oldleaves. This might indicate that there is sizabletranslocation of photosynthates from the sourceleaves to the roots as observed in adult orchidplants (Yong and Hew, 1995). It appears that theorchid plantlets developed large sink capacity as aresult of the formation of larger roots and in-creased root initiation when grown at super-ele-vated CO2, which allowed the utilization of extracarbon fixed. Our earlier work showed that orchidroots did fix carbon but did not show net photo-synthesis (Hew et al., 1984). Also, carbon fixed byroots was not exported (Yong and Hew, 1995).On the other hand, a considerable amount (10–20%) of carbon fixed in leaves was translocated tothe roots. The roots therefore behaved predomi-nantly as a sink.
In conclusion, an increase in CO2 concentrationmay not result in feedback inhibition of photosyn-thesis in Mokara Yellow plantlets, although thesugar and starch content were higher. Instead,super-elevated CO2 could enhance the utilizationof captured light energy which alleviates photoin-hibition and enhances the growth of Mokara Yel-low plantlets.
Acknowledgements
We thank Ong Tang Kwee and Chong Ping Leefor excellent technical assistance. Discussion withDr Wendell Sun and Jean Yong was much appre-ciated. This study was supported by the ResearchProject RP960305, National University of Singa-pore. SSG is grateful to the University for provid-ing scholarship during the project.
References
Aitken-Christie, J., Davies, H.E., Holland, L., Kubota, C.,Fujiwara, K., 1992. Effect of nutrient media compositionsugar-free growth and chlorophyll fluorescence of Pinusradiata shoots in vitro. Acta Hortic. 319, 125–130.
Allen, L.H. Jr., 1990. Plant responses to rising carbon dioxideand potential interactions with air pollutants. J. Environ.Qual. 19, 15–34.
Beadle, C.L., 1993. Growth analysis. In: Hall, D.O., Scurlock,J.M.O., Bolhar-Nordenkampf, H.R., Leegood, R.C.,Long, S.P. (Eds.), Photosynthesis and Production in AChanging Environment. A Field and Laboratory Manual.Chapman and Hall, London, pp. 36–46.
Benzing, D.H., Friedman, W.E., Peterson, G., Renfrow, A.,1983. Shootlessness, velamentous roots and the pre-emi-nence of Orchidaceae in the epiphytic biotope. Am. J. Bot.70, 121–133.
Besford, R.T., Ludwig, L.J., Withers, A.C., 1990. The green-house effect: acclimation of tomato plants growing in highCO2, photosynthesis and ribulose-1,5-bisphosphate car-boxylase protein. J. Exp. Bot. 41, 925–931.
Bockers, M., Capkova, V., Ticha, I., Schafer, C., 1997.Growth at high CO2 affects the chloroplast number butnot the photosynthetic efficiency of photoautotrophicMarchantia polymorpha culture cells. Plant Cell TissueOrgan Cult. 48, 103–110.
Bowes, G., 1991. Growth at elevated CO2: photosyntheticresponses mediated through Rubisco. Plant Cell Environ.14, 795–806.
Cave, G., Tolley, L.C., Strain, B.R., 1981. Effect of carbondioxide enrichment on chlorophyll content, starch contentand starch grain structure in Trifolium subterraneum leaves.Physiol. Plant. 51, 171–174.
Demmig, B., Bjorkman, O., 1987. Comparison of the effect ofexcessive light on chlorophyll fluorescence (77 K) andphoton yield of O2 evolution in leaves of higher plants.Planta 171, 171–184.
Desjardins, Y., Gosselin, A., Yelle, S., 1987. Acclimatizationof ex 6itro strawberry plantlets in CO2-enriched environ-ments and supplementary lighting. J. Am. Soc. Hortic. Sci.112, 846–851.
Desjardins, Y., Hdider, C., De Riek, J., 1995. Carbon nutri-tion in vitro-regulation and manipulation of carbon assim-ilation in micropropagated systems. In: Aitken-Christie, J.,Kozai, T., Smith, M.A.L. (Eds.), Automation and Envi-ronmental Control in Plant Tissue Culture. Kluwer, Dor-drecht, pp. 441–472.
Dijkstra, P., 1989. Cause and effect of differences in specificleaf area. In: Lambers, H., Cambridge, M.L., Konings, H.,Pons, T.L. (Eds.), Causes and Consequences of Variationin Growth Rate and Productivity of Higher Plants. SPBAcademic Publishing, The Hague, pp. 125–140.
Drake, B.G., Gonzaler-Meler, M.A., Long, S.P., 1997. Moreefficient plants: a consequence of rising atmospheric CO2?Annu. Rev. Plant Physiol. Plant Mol. Biol. 48, 609–639.
Farrar, J.F., 1993. Carbon partitioning. In: Hall, D.O., Scur-lock, J.M.O., Bolhar-Nordenkampf, H.R., Leegood, R.C.,Long, S.P. (Eds.), Photosynthesis and Production in AChanging Environment. A Field and Laboratory Manual.Chapman and Hall, London, pp. 232–246.
Farrar, J.F., Williams, M.L., 1991. The effects of increasedatmospheric carbon dioxide and temperature on carbon

S.S. Gouk et al. / En6ironmental and Experimental Botany 41 (1999) 219–230 229
partitioning, source-sink relations and respiration. PlantCell Environ. 14, 819–830.
Fujiwara, K., Kozai, T., Watanabe, I., 1987. Measurement ofcarbon dioxide gas concentration in closed vessel contain-ing tissue cultured plantlets and estimates of net photosyn-thetic rates of the plantlets. J. Agric. Meteorol. 43, 21–30.
Gouk, S.S., Yong, J.W.H., Hew, C.S., 1997. Effects of super-elevated CO2 on the growth and carboxylating enzymes inan epiphytic CAM orchid plantlet. J. Plant Physiol. 151,129–136.
Grout, B.W.W., Ashton, M.J., 1978. Transplanting ofcauliflower plants regenerated from meristem culture. II.Carbon dioxide fixation and the development of photosyn-thetic ability. Hortic. Res. 17, 65–71.
Gunderson, C.A., Wullschleger, S.D., 1994. Photosyntheticacclimation in trees to rising atmospheric CO2: a broaderperspective. Photosynth. Res. 39, 369–388.
Habash, D.Z., Paul, M.J., Parry, M.A.J., Keys, A.J., Lawlor,D.W., 1995. Increased capacity for photosynthesis in wheatgrown at elevated CO2: the relationship between electrontransport and carbon metabolism. Planta 197, 482–489.
Havaux, M., Strasser, R.J., Greppin, H., 1991. A theoreticaland experimental analysis of the qP and coefficients ofchlorophyll fluorescence quenching and their relation tophotochemical and nonphotochemical events. Photosynth.Res. 27, 41–55.
Hdider, C., Desjardins, Y., 1994. Effects of sucrose on photo-synthesis and phosphoenolpyruvate carboxylase activity ofin vitro culture strawberry plantlets. Plant Cell TissueOrgan Cult. 36, 27–33.
Hew, C.S., Yong, J.W.H., 1997. The Physiology of TropicalOrchids in Relation to the Industry. World Scientific,Singapore 331 pp.
Hew, C.S., Ng, Y.W., Wong, S.C., Yeoh, H.H., Ho, K.K.,1984. Carbon fixation in orchid aerial roots. Physiol. Plant.60, 154–158.
Hew, C.S., Hin, S.E., Yong, J.W.H., Gouk, S.S., Tanaka, M.,1995. In vitro CO2 enrichment of CAM orchid plantlets. J.Hortic. Sci. 70, 721–736.
Keutgen, N., Chen, K., Lenz, F., 1997. Responses of straw-berry leaf photosynthesis, chlorophyll fluorescence andmacronutrient contents to elevated CO2. J. Plant Physiol.150, 395–440.
Kirdmanee, C., Kitaya, Y., Kozai, T., 1995. Effects of CO2
enrichment and supporting material in vitro on photoau-totrophic growth of Eucalyptus plantlets in vitro and exvitro. In Vitro Cell Dev. Biol. 31, 144–149.
Kozai, T., 1991. Micropropagation under photoautotrophicconditions. In: Debergh, P.C., Zimmerman, R.H. (Eds.),Micropropagation: Technology and Application. Kluwer,Dordrecht, pp. 447–469.
Kozai, T., Oki, H., Fujiwara, K., 1990. Photosynthetic charac-teristics of Cymbidium plantlet in vitro. Plant Cell TissueOrgan Cult. 22, 205–211.
Lakso, A., Reisch, B.I., Mortensen, J., Roberts, M.H., 1986.Carbon dioxide enrichment for stimulation of growth of invitro-propagated grapevines after transfer from culture. J.Am. Soc. Hortic. Sci. 111, 634–638.
Long, S.P., Drake, B.G., 1991. Effect of the long-term eleva-tion of CO2 concentration in the field on the quantum yieldof photosynthesis of the C3 sedge, Scirpus olneyi. PlantPhysiol. 96, 221–226.
Madsen, E., 1973. Effect of CO2 enrichment on the morpho-logical, histological and cytological changes in tomatoplants. Acta Agric. Scand. 23, 241–246.
Moran, R., 1982. Formulae for determination of chlorophyl-lous pigments extracted with N,N-dimethylformamide.Plant Physiol. 69, 1376–1381.
Mortensen, L.M., 1987. Review: CO2 enrichment in green-house. Crop responses. Sci. Hortic. 33, 1–25.
Norby, R.J., 1994. Issues and perspectives for investigatingroot responses to elevated atmospheric carbon dioxide.Plant Soil 165, 9–20.
Peterson, R.B., 1990. Effects of water vapor deficit on photo-chemical and fluorescence yields in tobacco leaf tissue.Plant Physiol. 92, 608–614.
Powles, S.B., 1984. Photoinhibition of photosynthesis inducedby visible light. Annu. Rev. Plant Physiol. 35, 15–44.
Pridgeon, A.M., 1987. The velamen and exordemis of orchidroots. In: Arditti, J. (Ed.), Orchid Biology—Reviews andPerspectives, vol. IV. Cornell University Press, Ithaca, pp.139–192.
Prior, S.A., Rogers, H.H., Runion, G.B., Hendry, G.R., 1994.Free-air CO2 enrichment of cotton: Vertical and lateralroot distribution patterns. Plant Soil 165, 33–44.
Pritchard, S.G., Peterson, C.M., Prior, S.A., Rogers, H.H.,1997. Elevated atmospheric CO2 differentially affectsneedle chloroplast ultrastructure and phloem anatomy inPinus palustris : interactions with soil resource availability.Plant Cell Environ. 20, 461–471.
Roden, J.S., Ball, M.C., 1996. The effect of elevated [CO2] ongrowth and photosynthesis of two eucalyptus species ex-posed to high temperatures and water deficits. Plant Phys-iol. 111, 909–919.
Rogers, H.H., Runion, G.B., Krupa, S.V., 1994. Plant re-sponses to atmospheric CO2 enrichment with emphasis onroots and the rhizosphere. Environ. Pollut. 83, 155–189.
Schiffgens-Gruber, A., Lutz, C., 1992. Ultrastructure of meso-phyll cell chloroplasts of spruce needles exposed to O3, SO2
and NO2 alone and in combination. Environ. Exp. Bot. 32,243–254.
Serret, M.D., Trillas, M.I., Matas, J., Araus, J.L., 1996.Development of photoautotrophy and photoinhibition ofGardenia jasminoides plantlets during micropropagation.Plant Cell Tissue Organ Cult. 45, 1–16.
Solarova, J., 1989. Diurnal variation in CO2 concentration incultivation vessels resulting from photosynthetic activity.Photosynthetica 23, 100–107.
Stitt, M., 1991. Rising CO2 levels and their potential signifi-cance for carbon flow in photosynthetic cells. Plant CellEnviron. 14, 741–762.
Tissue, D.T., Thomas, R.B., Strain, B.R., 1993. Long-termeffects of elevated CO2 and nutrients on photosynthesisand Rubisco in loblolly pine seedlings. Plant Cell Environ.16, 859–865.

S.S. Gouk et al. / En6ironmental and Experimental Botany 41 (1999) 219–230230
von Willert, D.J., Matyssek, R., Herppich, W., 1995. Experi-mentelle Pflanzenokologie. Grundlagen und Anwendun-gen. Thieme, Stuttgart, New York.
Vu, J.C.V., Allen, L.H. Jr., Bowes, G., 1989. Leaf ultrastruc-ture, carbohydrates and protein of soybeans grown underCO2 enrichment. Environ. Exp. Bot. 29, 141–147.
Weis, E., Berry, J.A., 1987. Quantum efficiency of photosys-tem II in relation to ‘energy’-dependent quenching ofchlorophyll fluorescence. Biochim. Biophys. Acta 894,198–208.
Wong, S.C., 1990. Elevated atmospheric partial pressure ofCO2 and plant growth. II. Non-structural carbohydratecontent in cotton plants and its effect on growth. Photo-synth. Res. 23, 171–180.
Woodrow, I.E., 1994. Control of steady-state photosynthesisin sunflowers growing in enhanced CO2. Plant Cell Envi-ron. 17, 277–286.
Wulff, R.O., Strain, B.R., 1982. Effects of CO2 enrichment ongrowth and photosynthesis in Desmodium paniculatum.Can. J. Bot. 60, 1084–1091.
Yong, J.W.H., Hew, C.S., 1995. Partitioning of 14C assimilatesbetween sources and sinks during different growth stages inthe sympodial thin-leaved orchid Oncidium Goldiana. Int.J. Plant Sci. 156, 188–196.
Zhu, J., Bartholomew, D.P., Golstein, G., 1997. Effect ofelevated carbon dioxide on the growth and physiologicalresponses of pineapple, a species with Crassulacean acidmetabolism. J. Am. Soc. Hortic. Sci. 122, 233–237.
.
Copyright © 2022 FDOKUMEN




















