
Distribution of life cycle stages in a lithophytic and epiphytic orchid
14
Folia Geobotanica 41: 107–120, 2006 DISTRIBUTION OF LIFE CYCLE STAGES IN A LITHOPHYTIC AND EPIPHYTIC ORCHID Noel Rivera Gómez 1) , Raymond L. Tremblay *,1,2,4) & Elvia Meléndez-Ackerman 3,4) 1) Department of Biology, 100 Rd. 908, University of Puerto Rico, Humacao, Puerto Rico, 00791, *) Corresponding author: [email protected] 2) Department of Biology, PO Box 23360, University of Puerto Rico, Río Piedras, Puerto Rico, 00931-3360 3) Institute of Tropical Ecosystem Studies, University of Puerto Rico, Box 23341, Río Piedras, Puerto Rico, 00931-3341 4) Crest, Center for Applied Tropical Ecology and Conservation, PO Box 23341,University of Puerto Rico, Río Piedras, Puerto Rico, 00931-3341 Abstract: Density-dependent processes may have multiple effects on populations, which among other things include the regulation of population abundance and of the relative distribution of life-cycle stages within populations. The epiphytic habitat is often characterized as highly ephemeral and therefore epiphytic orchid populations may never achieve density-dependent regulation. In this study, we investigated the potential for density-dependent regulation in epiphytic and lithophytic orchids by examining the association between seedlings, juvenile and adult life-history stages in the Caribbean endemic orchid, Lepanthes rupestris in a cross-sectional study of 179 populations surveyed in the Luquillo National Forest along a riparian area where it is locally abundant. Under density-dependent regulation we expected a negative association between the ratio of seedling/adults and juveniles/adults and total population density. Population density was in the range of 140 individuals per m 2 , however patch sizes were small and mostly limited to less than 0.5 m 2 with a maximum of 3m 2 . We found no evidence of reduction of the ratio of seedlings or juveniles to adults as population size increased in either tree or boulder populations suggesting negative density dependence for population regulation in L. rupestris is either rare or occurs at even higher densities than those measured here. Moreover, we found positive (although weak) relationship between the ratio of seedlings and juveniles to adults and population size, suggesting that facilitation may be occurring. Keywords: Caribbean National Forest, Density-dependent competition, Lepanthes, Orchidaceae, Population dynamics, Pleurothallidinae, Puerto Rico INTRODUCTION Population density is one of many factors that can influence biotic interactions in natural plant populations in many ways. For example, an increase in the number of individuals could magnify negative interactions such as intraspecific competition (KERY et al. 2000), predation (e.g. JANZEN 1970, CONNELL 1971), disease incidence (e.g. AUSPURGER 1983). All these phenomena may in turn affect plant reproduction and survival and thus act as stabilizing mechanisms of population growth in plants (HARPER 1977). Increases in plant densities, however, need not necessarily lead to reductions in population growth rate. For example, increases in plant densities could augment pollinator activity (KRANNITZ &ANWAR 1992, CAMPBELL &WASER 1989), which in turn may increase plant reproduction (e.g. ROLL et al. 1997). In addition, increases in population density can also affect the mating structure of plant
Transcript of Distribution of life cycle stages in a lithophytic and epiphytic orchid

Folia Geobotanica 41: 107–120, 2006
DISTRIBUTION OF LIFE CYCLE STAGES IN A LITHOPHYTICAND EPIPHYTIC ORCHID
Noel Rivera Gómez1)
, Raymond L. Tremblay*,1,2,4)
& Elvia Meléndez-Ackerman3,4)
1) Department of Biology, 100 Rd. 908, University of Puerto Rico, Humacao, Puerto Rico, 00791,
*) Corresponding author: [email protected]
2) Department of Biology, PO Box 23360, University of Puerto Rico, Río Piedras, Puerto Rico, 00931-3360
3) Institute of Tropical Ecosystem Studies, University of Puerto Rico, Box 23341, Río Piedras, Puerto Rico,
00931-3341
4) Crest, Center for Applied Tropical Ecology and Conservation, PO Box 23341,University of Puerto Rico, Río
Piedras, Puerto Rico, 00931-3341
Abstract: Density-dependent processes may have multiple effects on populations, which among other thingsinclude the regulation of population abundance and of the relative distribution of life-cycle stages withinpopulations. The epiphytic habitat is often characterized as highly ephemeral and therefore epiphytic orchidpopulations may never achieve density-dependent regulation. In this study, we investigated the potential fordensity-dependent regulation in epiphytic and lithophytic orchids by examining the association betweenseedlings, juvenile and adult life-history stages in the Caribbean endemic orchid, Lepanthes rupestris ina cross-sectional study of 179 populations surveyed in the Luquillo National Forest along a riparian area where itis locally abundant. Under density-dependent regulation we expected a negative association between the ratio ofseedling/adults and juveniles/adults and total population density. Population density was in the range of 140individuals per m2, however patch sizes were small and mostly limited to less than 0.5 m2 with a maximum of3 m2. We found no evidence of reduction of the ratio of seedlings or juveniles to adults as population sizeincreased in either tree or boulder populations suggesting negative density dependence for population regulationin L. rupestris is either rare or occurs at even higher densities than those measured here. Moreover, we foundpositive (although weak) relationship between the ratio of seedlings and juveniles to adults and population size,suggesting that facilitation may be occurring.
Keywords: Caribbean National Forest, Density-dependent competition, Lepanthes, Orchidaceae, Populationdynamics, Pleurothallidinae, Puerto Rico
INTRODUCTION
Population density is one of many factors that can influence biotic interactions in naturalplant populations in many ways. For example, an increase in the number of individuals couldmagnify negative interactions such as intraspecific competition (KERY et al. 2000), predation(e.g. JANZEN 1970, CONNELL 1971), disease incidence (e.g. AUSPURGER 1983). All thesephenomena may in turn affect plant reproduction and survival and thus act as stabilizingmechanisms of population growth in plants (HARPER 1977). Increases in plant densities,however, need not necessarily lead to reductions in population growth rate. For example,increases in plant densities could augment pollinator activity (KRANNITZ & ANWAR 1992,CAMPBELL & WASER 1989), which in turn may increase plant reproduction (e.g. ROLL et al.1997). In addition, increases in population density can also affect the mating structure of plant

populations (e.g. ELLSTRAND et al. 1978, BRABANT et al. 1980) by increasing interplantpollinator movement (PYKE 1978, HEINRICH 1979) and outcrossing (MURAWSKI &HAMRICK 1991, VAN TREUREN et al. 1993, 1994). Thus, detection of patterns suggestive ofdensity-dependence is one step towards understanding how and at which scale bioticinteractions may regulate populations.
Stabilization of population sizes and population structure may be more the exception thanthe rule in rapidly fluctuating environments where drastic changes through abiotic factors ornatural disturbances may constantly shift population sizes from their equilibrium point(LUNDBERG et al. 2000). This phenomenon could make it difficult to detectdensity-dependent regulation. In orchids, the epiphytic environment is regarded as ephemeral(BENZING 1979, ACKERMAN 1983, CATLING et al. 1986) suggesting that environmentalstochasticity may play a larger role in abundance patterns of epiphytic plants thandensity-dependent processes. While long-term frequent monitoring of population abundancesis most likely the best option to detect density-dependent processes in changing environments(GILLMAN & DODD 2000), an alternative to long-term monitoring in unstable environmentsmay be available to plant species of small size that exhibit populations with patchydistribution and variable size. Species with such population characteristics may at least allowfor cross-sectional studies in which one could evaluate the relationship between demographicstructure and population density using multiple populations at any given time.
A large number of orchid species are indeed epiphytic (73%, ATWOOD 1986). However,studies that have focused on density-dependent processes in orchids within the epiphytichabitat are scarce and results are not consistent. The combined results of two studies with thetropical epiphytic orchid Tolumnia variegata suggest a positive association between increasesin flower density and fruit set (SABAT & ACKERMAN 1996) and in turn a positive associationbetween fruit set and seedling recruitment (ACKERMAN et al. 1996). In contrast, for theepiphytic orchid Ryncholaelia glauca in Mexico, FLORES-PALACIOS & GARCÍA-FRANCO
(2003) found at best a weak positive relationship between flower density on host trees andfruit set, and only in one of their two census years and conclude that there is no evidence ofpositive density dependence on reproductive success. As yet we have no evidence of whatlimits population density in epiphytic orchids.
This study focused on testing for the possibility of density-dependent processes inLepanthes rupestris STIMSON, an endemic miniature orchid of Puerto Rico that is a commonlithophyte and epiphyte along rivers of the northwestern slopes of the Luquillo Mountains ofPuerto Rico (ACKERMAN 1995, TREMBLAY 1997a). Populations of Lepanthes rupestris (aswell as those of most Lepanthes) are discrete and patchily distributed (TREMBLAY 1997a)with size distributions that are often leptokurtic with many populations of small size and fewlarge populations (TREMBLAY 1997a). Such population structure may allow for detection ofdensity-dependence with cross-sectional studies.
The different habitats (epiphytic and lithophytic) in which L. rupestris populations arefound may lead to differences in the opportunity for density dependence. Streams inhabitedby L. rupestris are subjected to frequent flash floods (JOHNSON et al. 1998) resulting inmortality of many if not most individuals (TREMBLAY et al., submitted). Epiphytic plants ofL. rupestris occur on the boles of trees, and thus may be a more stable environment as
108 N. Rivera Gómez et al.

compared to other epiphytic orchids that commonly inhabit branches or twigs and may showhigh mortality rates as a result of the frequent breakage of their substrate. Thus, wehypothesized that population sizes of L. rupestris would be more variable for lithophyticpopulations growing on boulders along the stream than for epiphytic orchids growing on theboles of trees along the riparian zone. In other words we expect to find substrate-related (treesvs. boulders) differences in population regulation by density dependent-processes if any aredetected.
The purpose of this study was to evaluate the potential for density-dependent populationregulation in L. rupestris by comparing the relative distribution of non-reproductive andreproductive individuals among many natural patches of varying size. We further evaluatedwhether or not the relative distribution of non-reproductive and reproductive individualsdiffered between two substrates (trees vs. boulders). We expected a negative relationshipbetween the ratio of seedlings and juveniles to adults and population density undera hypothesis of negative density-dependence. In contrast, a positive relation between theseparameters would suggest positive density dependence. Under the assumption that orchidsgrowing on tree trunks are more stable than those growing on boulders we expected densitydependent effects (if any) to be stronger on tree substrates.
METHODS
Study system
Lepanthes rupestris plants are caespitose with slender stems, erect with a solitary leaf(1.3–4.3 cm long) subtending one to a few inflorescences capable of producing many flowers.Flowers are minute (< 6 mm) and variable in coloration and morphology (Fig. 1; TREMBLAY
1997b). Plants are obligately cross-pollinated and partially protandrous (TREMBLAY et al.,unpubl.). Reproductive success in this orchid as in most orchids is pollinator-limited(TREMBLAY & ACKERMAN 2001, TREMBLAY et al. 2005). Previous studies have shown thatlimited gene flow is present among groups of individuals on different boulders and trees evenwhen physically close (TREMBLAY & ACKERMAN 2001), which is typical of many orchids(see TREMBLAY & ACKERMAN 2003, TREMBLAY et al. 2005). Fruits produce many seeds(approx. 3000, TREMBLAY, unpubl.) and require a mycorrhizal association for germinationand survival until plants start photosynthesis (BAYMAN et al. 2002). The pollinator ofL. rupestris is unknown but black-winged fungus gnats (Diptera: Sciaridae) are knownpollinators of Lepanthes elsewhere (BLANCO & BARBOZA 2005). Average and medianpopulation sizes are 50 and 26 individuals respectively with populations separated by 5 m onaverage (TREMBLAY 1997a). Estimated average life span is approximately 3.4 yrs(TREMBLAY 2000).
Study area
The study was carried out along a 1000 m section of Quebrada Sonadora in the LuquilloExperimental Forest (USDA Forest Service). Quebrada Sonadora is a first order tributary ofthe Espíritu Santo River, northeastern Puerto Rico, 18°18� N, 65°47� W. The section studiedfluctuated between 400 and 500 m in elevation and ran through an area of mature secondary“Tabonuco Forest”, dominated by the tree Dacryodes excelsa (WAIDE & REAGAN 1996).
Life cycle stages in a lithophytic orchid 109

110 N. Rivera Gómez et al.
Fig. 1. Lepanthes rupestris growing on a boulder among moss (scale 25 cent USA currency, photo NRG).

Life cycle stages in a lithophytic orchid 111
Fig 2. Lepanthes rupestris on the bole of the tree along Quebrada Sonadora in the Caribbean National Forest.S – seedling, J – juvenile and A – adult plants, photo by NRG.

112 N. Rivera Gómez et al.
Fig 3. Lepanthes rupestris on a boulder along Quebrada Sonadora in the Caribbean National Forest.S – seedling, J – juvenile and A – adult plants. S – seedling, J – juvenile and A – adult plants, photo by NRG.

Average annual precipitation of 3.6 m but rainfall is seasonal, with higher values from Maythrough December relative to the January to April period (BROWN et al. 1983, MCDOWELL &ESTRADA-PINTO 1988). Water discharge in this stream, however, is highly variable andclosely follows precipitation events (JOHNSON et al. 1998). Large storms have been shown toincrease water discharge to levels that are much higher than normal and capable of movingboulders and carrying large pieces of debris downstream (JOHNSON et al. 1998).
Data collection
We censused 250 patches of L. rupestris during the summer of 2003 and for each patch wecounted the number of individuals for all life cycle stages (seedlings, juveniles, and adults) inall populations. We eliminated all patches with less then 10 individuals to avoid the effect ofstochastic variation due to small sample size on the relative distribution of life cycle stagesand included 179 patches in this survey. Epiphytic plants were present on the bole and theexposed roots of trees and rarely on branches (Fig. 2), while lithophytic plants were located onboulders (Fig. 3) at different distances to the stream edge. Within each patch plants wereclassified into one of three life-cycle stages according to the following rules: (i) seedlingswere small plants without lepanthiform sheets on the petioles of any of the leaves,(ii) juveniles were individuals with at least one lepanthiform sheath on the petiole withoutevidence of past or current reproduction (iii) adults encompassed individuals that arecurrently reproductive (active inflorescences) or have been in the past (inflorescences maypersist for years after they have been active).
For each patch we calculated the ratio of seedling and juveniles to adults. We quantified thepatch occupancy area based on the perimeter formed by the outermost individuals in a patchwith the aid of a 150 cm2 mesh. We then used this area to calculate patch density as thenumber of individuals divided by patch occupancy area.
Statistical analysis
We evaluated the ratios of seedlings to adults and juveniles to adults as function of patchdensity with linear regression. For these analyses we transformed the data according to thefollowing equations: ln( )seedlings
adults+1 and ln( )juveniles
adults+1 .
We evaluated these functions for each substrate type separately as well as the pooled data.Third, we tested for differences in plant density between substrate types (i.e., tree vs. boulder)with a t-test. All tests and graphical outputs were performed in JMP (version 5.0, SAS Inc.).
RESULTS
Distribution of population size, patch size and population density
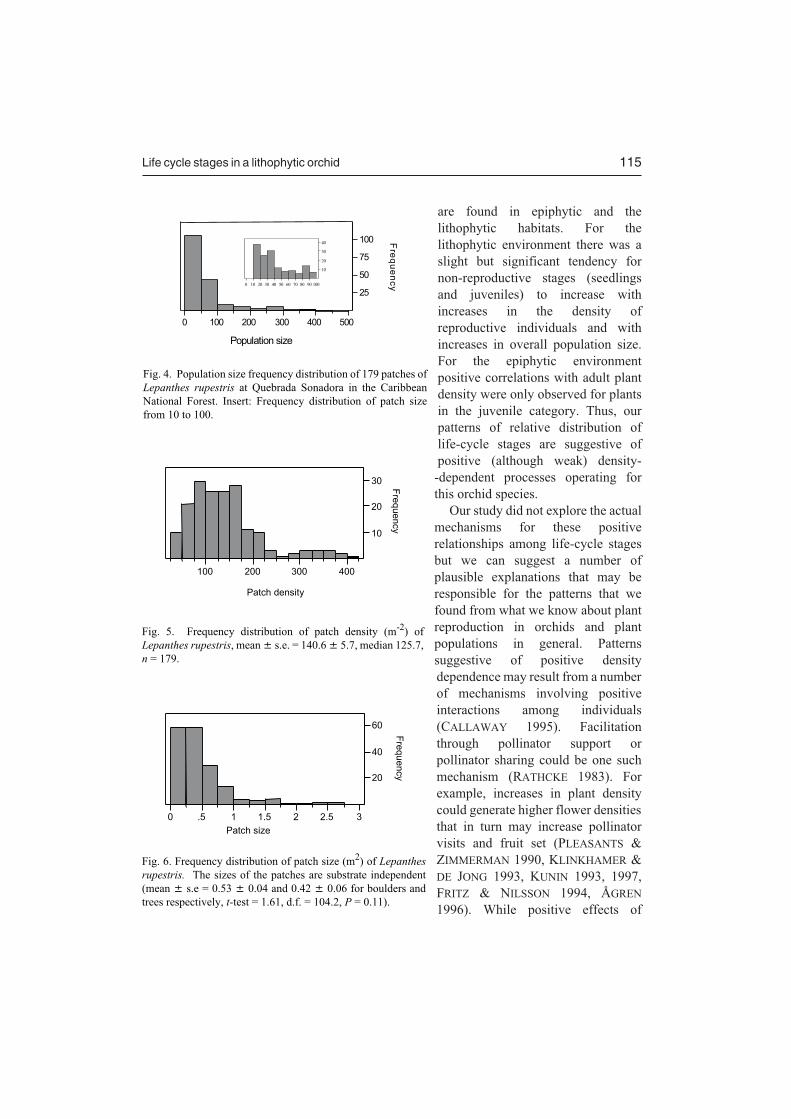
Distribution of population size is skewed towards many small patches and few largepatches, with a maximum of 496 individuals (Fig. 4). The median patches size was 39individuals with a mean (s.e.) of 69.7 (6.1). The population density of Lepanthes rupestris isapproximately 140 individuals/m2 (Fig. 5) with few populations having higher density then200 individuals/m2. Patch size varied from very small (0.1 m2) to a maximum of close to 3 m2
Life cycle stages in a lithophytic orchid 113

(Fig. 6), however patch distribution was skewed to the smaller size classes with most patchessized below 0.5/m2.
Relationships among life-cycle stages
We found a positive linear relationships in the ratios of juveniles to adults as function ofdensity for both the lithophytic and the epiphytic environments, but the ratios of seedlings toadults was positively significant only on trees. The amount of variation in the ratio ofseedlings to adults that was explained by variation in patch density fluctuated between 9.4 and3.2% (radj
2 ) for the lithophytic and epiphytic environment
ln( )seedlings
adults+1 = -0.09 + 0.04 density / m 2 , F1, 126 = 14.2, P = 0.0003,
ln( )seedlings
adults+1 = -0.02 + 0.06 density / m 2 , F1, 50 = 2.70, P = 0.11, respectively.
The amount of variation in the ratios of juveniles to adults showed positive increase withpopulation density for both the lithophytic and epiphytic environment
ln( )juveniles
adults+1 = 0.42 + 0.05 density / m 2 , radj
2 = 0.12, F1, 126 = 18.7, P < 0.0001,
ln( )juveniles
adults+ 1 = 0.42 + 0.06 density / m 2 , radj
2 = 0.12, F1, 50 = 18.7, P = 0.0001,respectively).
When the data for the different substrates were pooled the plant density explained a largerproportion of the variation in the ratios of seedlings and juveniles to adults
ln( )seedlings
adults+ 1 = -1.12 + 0.28 density / m 2 , radj
2 = 0.17, F1, 178 = 38.94, P < 0.0001,
ln( )juveniles
adults+1 = 0.10 + 0.47 density / m 2 , radj
2 = 0.19, F 1, 178 = 42.6, P < 0.0001).
Density of life-cycles stages between substrates
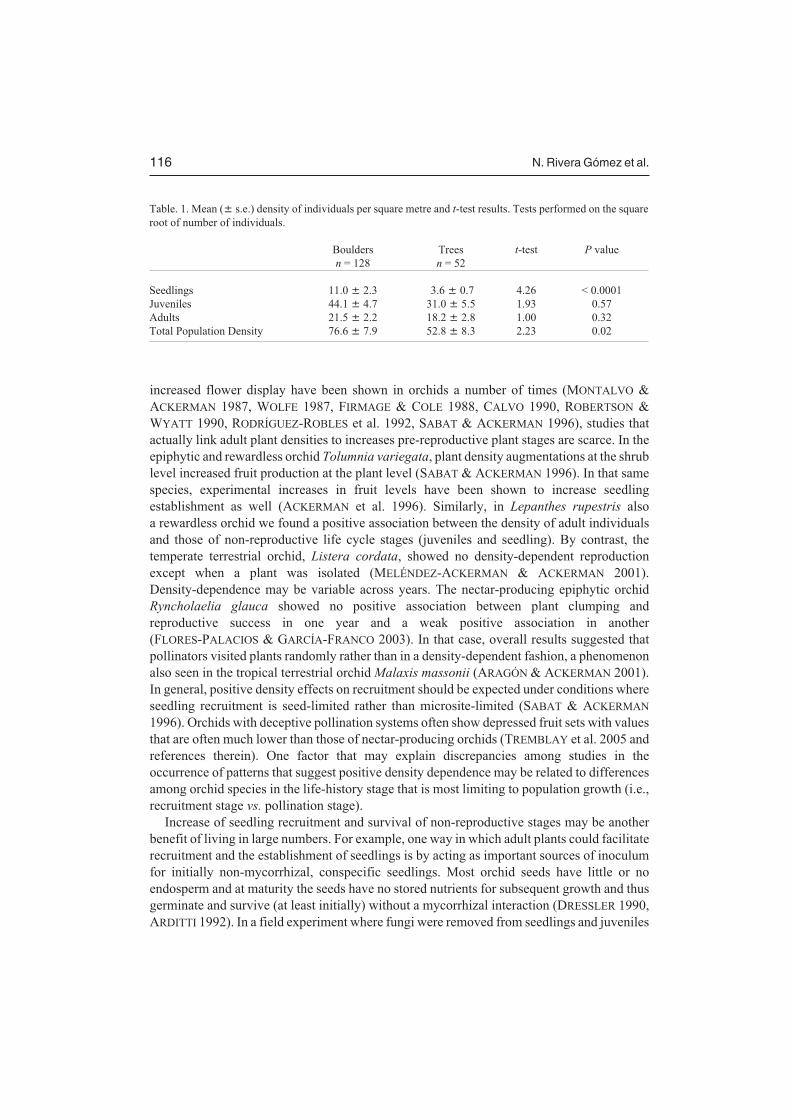
There were no differences in the mean density of juveniles or adults between patchesgrowing on boulders and trees (Table 1). However the mean density of seedlings wassignificantly lower on trees than on boulders. Consequently when all stages were pooledlarger densities were present on boulders (Table 1).
DISCUSSION
In general, studies on orchid demography often assume density-independent growth intheir predictive models (WILLEMS & BIK 1991, TREMBLAY 1997c, HUTCHINGS et al. 1998,WAITE & FARRELL 1998, WILLEMS & MELSER 1998, KULL 2002, TREMBLAY &HUTCHINGS 2004). This cross-sectional study does not support a hypothesis of negativedensity-dependent regulation of population density in the orchid Lepanthes rupestris, which
114 N. Rivera Gómez et al.
are found in epiphytic and thelithophytic habitats. For thelithophytic environment there was aslight but significant tendency fornon-reproductive stages (seedlingsand juveniles) to increase withincreases in the density ofreproductive individuals and withincreases in overall population size.For the epiphytic environmentpositive correlations with adult plantdensity were only observed for plantsin the juvenile category. Thus, ourpatterns of relative distribution oflife-cycle stages are suggestive ofpositive (although weak) density--dependent processes operating forthis orchid species.
Our study did not explore the actualmechanisms for these positiverelationships among life-cycle stagesbut we can suggest a number ofplausible explanations that may beresponsible for the patterns that wefound from what we know about plantreproduction in orchids and plantpopulations in general. Patternssuggestive of positive densitydependence may result from a numberof mechanisms involving positiveinteractions among individuals(CALLAWAY 1995). Facilitationthrough pollinator support orpollinator sharing could be one suchmechanism (RATHCKE 1983). Forexample, increases in plant densitycould generate higher flower densitiesthat in turn may increase pollinatorvisits and fruit set (PLEASANTS &ZIMMERMAN 1990, KLINKHAMER &DE JONG 1993, KUNIN 1993, 1997,FRITZ & NILSSON 1994, ÅGREN
1996). While positive effects of
Life cycle stages in a lithophytic orchid 115
25
50
75
100 Fre
qu
en
cy
0 100 200 300 400 500
Population size
10
20
30
40
0 10 20 30 40 50 60 70 80 90 100
Fig. 4. Population size frequency distribution of 179 patches ofLepanthes rupestris at Quebrada Sonadora in the CaribbeanNational Forest. Insert: Frequency distribution of patch sizefrom 10 to 100.
10
20
30
Fre
quency
100 200 300 400
Patch density
Fig. 5. Frequency distribution of patch density (m-2) ofLepanthes rupestris, mean � s.e. = 140.6 � 5.7, median 125.7,n = 179.
20
40
60
Fre
quency
0 .5 1 1.5 2 2.5 3
Patch size
Fig. 6. Frequency distribution of patch size (m2) of Lepanthes
rupestris. The sizes of the patches are substrate independent(mean � s.e = 0.53 � 0.04 and 0.42 � 0.06 for boulders andtrees respectively, t-test = 1.61, d.f. = 104.2, P = 0.11).
increased flower display have been shown in orchids a number of times (MONTALVO &ACKERMAN 1987, WOLFE 1987, FIRMAGE & COLE 1988, CALVO 1990, ROBERTSON &WYATT 1990, RODRÍGUEZ-ROBLES et al. 1992, SABAT & ACKERMAN 1996), studies thatactually link adult plant densities to increases pre-reproductive plant stages are scarce. In theepiphytic and rewardless orchid Tolumnia variegata, plant density augmentations at the shrublevel increased fruit production at the plant level (SABAT & ACKERMAN 1996). In that samespecies, experimental increases in fruit levels have been shown to increase seedlingestablishment as well (ACKERMAN et al. 1996). Similarly, in Lepanthes rupestris alsoa rewardless orchid we found a positive association between the density of adult individualsand those of non-reproductive life cycle stages (juveniles and seedling). By contrast, thetemperate terrestrial orchid, Listera cordata, showed no density-dependent reproductionexcept when a plant was isolated (MELÉNDEZ-ACKERMAN & ACKERMAN 2001).Density-dependence may be variable across years. The nectar-producing epiphytic orchidRyncholaelia glauca showed no positive association between plant clumping andreproductive success in one year and a weak positive association in another(FLORES-PALACIOS & GARCÍA-FRANCO 2003). In that case, overall results suggested thatpollinators visited plants randomly rather than in a density-dependent fashion, a phenomenonalso seen in the tropical terrestrial orchid Malaxis massonii (ARAGÓN & ACKERMAN 2001).In general, positive density effects on recruitment should be expected under conditions whereseedling recruitment is seed-limited rather than microsite-limited (SABAT & ACKERMAN
1996). Orchids with deceptive pollination systems often show depressed fruit sets with valuesthat are often much lower than those of nectar-producing orchids (TREMBLAY et al. 2005 andreferences therein). One factor that may explain discrepancies among studies in theoccurrence of patterns that suggest positive density dependence may be related to differencesamong orchid species in the life-history stage that is most limiting to population growth (i.e.,recruitment stage vs. pollination stage).
Increase of seedling recruitment and survival of non-reproductive stages may be anotherbenefit of living in large numbers. For example, one way in which adult plants could facilitaterecruitment and the establishment of seedlings is by acting as important sources of inoculumfor initially non-mycorrhizal, conspecific seedlings. Most orchid seeds have little or noendosperm and at maturity the seeds have no stored nutrients for subsequent growth and thusgerminate and survive (at least initially) without a mycorrhizal interaction (DRESSLER 1990,ARDITTI 1992). In a field experiment where fungi were removed from seedlings and juveniles
116 N. Rivera Gómez et al.
Table. 1. Mean (� s.e.) density of individuals per square metre and t-test results. Tests performed on the squareroot of number of individuals.
Boulders Trees t-test P valuen = 128 n = 52
Seedlings 11.0 � 2.3 3.6 � 0.7 4.26 < 0.0001Juveniles 44.1 � 4.7 31.0 � 5.5 1.93 0.57Adults 21.5 � 2.2 18.2 � 2.8 1.00 0.32Total Population Density 76.6 � 7.9 52.8 � 8.3 2.23 0.02

of Lepanthes rupestris, BAYMAN and collaborators (2002) showed that in one fungicidetreatment survivorship of both stages was reduced in the fungal-free treatments but thatseedling recruitment was independent of fungal treatment. Thus, it is possible that a scenariowhere adults of L. rupestris are acting as “nurse plants” for non-reproductive stages may alsobe responsible for the positive relationships between non-reproductive individuals and adultplants.
Regardless of fact that we found evidence that suggests positive interactions betweenlife-cycle stages in L. rupestris, these were weak and only explained a small percentage of thevariation in density. The effects of environmental stochasticity can also shift the sign of bioticinteractions among individuals within a plant community (CALLAWAY 1997). Indeed,vulnerability of the lithophytic habitat to flash floods of L. rupestris has been proposed toreduce density effects among adults and juveniles (TREMBLAY 2000). Some preliminaryevidence for this phenomenon comes from the related species, L. rubripetala. Specifically,a population of this species experienced a flash flood where only one of 35 adults survived yetalmost all juveniles and seedlings lived (TREMBLAY, unpubl.). Regardless, this would notexplain the similar relationship between adult plants and non-reproductive stages in theepiphytic environment. Thus, other factors are likely to have a larger influence on thedemography of this species. For example, intraspecific density-dependent interactions arewidespread within natural communities and can strongly influence the abundance anddistribution of plant species (CALLAWAY & WALKER 1997, BRUNO et al. 2003).
The lithophytic environment supported a higher density of seedlings than the epiphyticenvironment. All other stages presented similar densities across substrates. This fails tosupport the theory of higher vulnerability of the lithophytic habitat due to flash floods(TREMBLAY 2000). Our data actually suggest that plant recruitment may be more limited inthe epiphytic environment. Yet if this is the case, then how can we explain the lack ofdifferences between substrates in the density of juvenile and adult classes? The most likelyexplanation is that survivorship of seedlings is much higher in the epiphytic environment thanin the lithophytic environment, which would then compensate for the smaller seedling densityon trees relative to boulders.
We hypothesized that there should be no negative intraspecific interactions in theepiphytic-lithophytic environment under the generalized assumption that such environmentsare highly variable. Indeed, we found no evidence supporting density-dependent regulation ofpopulation size and life-cycle stages in L. rupestris in agreement with our initial hypothesisfor the lack of negative intraspecific interactions in the epiphytic-lithophytic environmentunder the generalized assumption that such environments are highly variable. Instead, weshowed positive relationships between life-cycle stages. While results would suggest theoccurrence of facilitative interaction among life-history stages, they only explained a minorcomponent of the variation of densities of life-cycle stages. Thus additional factors (such asthe occurrence of interspecific interactions or natural disturbances) may need to be exploredin order to understand the factors that regulated the patterns of abundance and distribution intropical orchid.
Acknowledgment: We would like thank J.D. Ackerman for discussion and comments on the most recentversion of the manuscript and greatly appreciate the anonymous reviewers’ comments.
Life cycle stages in a lithophytic orchid 117

REFERENCES
ACKERMAN J.D. (1983): On the evidence for a primitively epiphytic habit in orchids. Syst. Bot. 8: 474–477.ACKERMAN J.D. (1995): An Orchid Flora of Puerto Rico and the Virgin Islands. Mem. New York Bot. Gard. 73:
1–208.ACKERMAN J.D., SABAT A. & ZIMMERMAN J.K. (1996): Seedling establishment in an epiphytic orchid: an
experimental study of seed limitation. Oecologia 106: 44–49.ÅGREN J. (1996): Population size, pollinator limitation, and seed set in the self-incompatible herb Lythrum
salicaria. Ecology 77: 1779–1790.ARAGÓN S. & ACKERMAN J.D. (2001): Density effects on the reproductive success and herbivory of Malaxis
massonii. Lindleyana 16: 3–12.ARDITTI J. (1992): Fundamentals of orchid biology. John Wiley & Sons, New York.ATWOOD J.T. (1986): The size of the Orchidaceae and the systematic distribution of epiphytic orchids. Selbyana
9: 171–186.AUSPURGER C.K. (1983): Seed dispersal of the tropical tree Platypodium elegans, and the escape of its seedlings
from fungal pathogens. J. Ecol. 71: 759–771.BAYMAN P., GONZÁLEZ E., FUMERO J.J. & TREMBLAY R.L. (2002): Are fungi necessary? How fungicides
affect growth and survival of the orchid Lepanthes rupestris in the field. J. Ecol. 90: 1002–1007.BENZING D.H. (1979): Alternative interpretations for the evidence that certain orchids and bromeliads act as
shoot parasites. Selbyana 5: 135–144.BLANCO M. & BARBOZA G. (2005): Pseudocopulatory pollination in Lepanthes (Orchidaceae:
Pleurothallidinae) by fungus gnats. Ann. Bot. (Oxford) 95: 763–772.BRABANT PH., GOUYON P.H., LEFORT G., VALDEYRON G. & VERNET PH. (1980): Pollination studies in
Thymus vulgaris L. (Labiatae). Oecol. Pl. 1: 37–45.BROWN S., LUGO A.E., SILANDER S. & LIEGEL L.H. (1983): Research opportunities at the Luquillo
Experimental Forest. In: USDA Forest Service Southern Experiment Station General Technical Report
SO-44, New Orleans.BRUNO J.F., STACHOWICZ J.J. & BERTNESS M.D. (2003): Inclusion of facilitation into ecological theory.
Trends Ecol. Evol. 18: 119–125.CALLAWAY R.M. (1995): Positive interactions among plants. Bot. Rev. 61: 306–349.CALLAWAY R.M. (1997): Positive interactions in plant communities and the individualistic-continuum concept.
Oecologia 112: 143–149.CALLAWAY R.M. & WALKER L.R. (1997): Competition and facilitation: a synthetic approach to interactions in
plant communities. Ecology 78: 1958–1965.CALVO R.N. (1990): Inflorescence size and fruit distribution among individuals in three orchid species. Amer. J.
Bot. 77: 1378–1381.CAMPBELL D.R. & WASER N.M. (1989): Variation in pollen flow within and among populations of Ipomopsis
aggregata. Evolution 43: 1444–1455.CATLING P.M., BROWNELL V.R. & LEFKOVITCH L.P. (1986): Epiphytic orchids in a Belizean Grapefruit
orchard: Distribution, colonization, and association. Lindleyana 1: 194–202.CONNELL J.H. (1971): On the role of the natural enemies in preventing competitive exclusion in some marine
animals and in rain forest trees. In: DEN BOER P.J. & GRADWELL G. (eds.), Dynamics of populations,PUDOC, Wageningen, pp. 298–312.
DRESSLER R.L. (1990): The Orchids. Natural history and classification. Harvard University Press, Cambridge.ELLSTRAND N.C., TORRES A.M. & LEVIN D.A. (1978): Density and rate of apparent outcrossing in Helianthus
annus (Asteraceae). Syst. Bot. 3: 403–407.FIRMAGE D.H. & COLE F.R. (1988): Reproductive success and inflorescence size of Calopogon tuberosus
(Orchidaceae). Amer. J. Bot. 75: 1371–1377.FLORES-PALACIOS A. & GARCÍA-FRANCO J. G. (2003): Effects of floral display and plant abundance on fruit
production of Ryncholaelia glauca (Orchidaceae). Rev. Biol. Trop. 51: 71–78.FRITZ A.-L. & NILSSON L.A. (1994): How pollinator-mediated mating varies with population size in plants.
Oecologia 100: 451–462.
118 N. Rivera Gómez et al.

GILLMAN M.P. & DODD M. (2000): Detection of delayed density dependence in an orchid population. J. Ecol.
88: 204–212.HARPER J.L. (1977): Population biology of plants. Academic Press, New York.HEINRICH B. (1979): Resource heterogeneity and pattern of movement in foraging bumblebees. Oecologia 40:
235–245.HUTCHINGS M.J., MENDOZA A. & HAVERS W. (1998): Demographics properties of an outlier population of
Orchis militaris L. (Orchidaceae) in England. Bot. J. Linn. Soc. 126: 95–107.JANZEN D.H. (1970): Herbivores and the number of tree species in tropical forests. Amer. Naturalist 104:
501–529.JOHNSON S.L., COVICH A.P., CROWL T., ESTRADA-PINTO A., BITHORN J. & WURSBAUGH W.A. (1998): Do
seasonality and disturbance influence reproduction in freshwater atyind shrimp in headwater streams, PuertoRico? Int. Vereinigung Theor. Limnol. Verh. 26: 2076–2081.
KERY M., MATTHIES D. & LINKHAMER H.H. (2000): Reduced fecundities and offspring performance in smallpopulations of the declining grassland plants Primula veris and Gentiana lutea. J. Ecol. 88: 17–30.
KLINKHAMER P.G.L. & DE JONG T. (1993): Attractiveness to pollinators: a plant dilemma. Oikos 66: 108–114.KRANNITZ P.G. & MAUN M.A. (1992): An experimental study of floral display amd reproductive success in
Viburnum opulus: importance of grouping. Canad. J. Bot. 69: 394–399.KULL T. (2002): Ecology: Population dynamics of north temperate orchids. In: KULL T. & ARDITTI J. (eds.),
Orchid biology: reviews and perspectives VIII, Kluwer Academic Publishers, Dordrecht, pp. 139–165.KUNIN W.E. (1993): Sex and the single mustard: population density and pollinator behavior effects on seed-set.
Ecology 74: 2145–2160.KUNIN W.E. (1997): Population size and density effects in pollination: pollinator foraging and plant
reproductive success in experimental arrays of Brassica kaber. J. Ecol. 85: 225–234.LUNDBERG P., RANTA E., JÖRGEN R. & KAITALA V. (2000): Population variability in time and space. Trends
Ecol. Evol. 15: 460–463.MCDOWELL W.H. & ESTRADA PINTO A. (1988): Rainfall at the El Verde Field Station, 1964–1986. University
of Puerto Rico CEER Technical Report T-228.MELÉNDEZ-ACKERMAN E.J. & ACKERMAN J.D. (2001): Density-dependent variation in reproductive success
in a terrestrial orchid. Pl. Syst. Evol. 227: 27–36.MONTALVO A.M. & ACKERMAN J.D. (1987): Limitations to fruit production in Ionopsis utricularioides
(Orchidaceae). Biotropica 19: 24–31.MURAWSKI D.A. & HAMRICK J.L. (1991): The effect of the density of flowering individuals on the mating
system of nine tropical tree species. Heredity 67: 167–174.PLEASANTS J.M. & ZIMMERMAN M. (1990): The effect of inflorescence size and pollinator visitation of
Delphinium nelsonii and Aconitum columnianum. Collect. Bot. (Barcelona) 19: 21–39.PYKE G.H. (1978): Optimal foraging: movement patterns of bumble between inflorescences. Theor. Populat.
Biol. 13: 72–97.RATHCKE B. (1983): Competition and facilitation among plants for pollination. In: REAL L. (ed.), Pollination
biology, Academic Press, Inc., Orlando, pp. 305–327.ROBERTSON J.L. & WYATT R. (1990): Reproductive biology of the yellow-fringed orchid Platanthera ciliaris.
Amer. J. Bot. 77: 388–398.RODRÍGUEZ-ROBLES J.A., MELÉNDEZ E.J. & ACKERMAN J.D. (1992): Effects of display size, flowering
phenology, and nectar availability on effective visitation frequency in Comparettia falcata (Orchidaceae).Amer. J. Bot. 79: 1009–1017.
ROLL J., MITCHELL R.J., CABIN R.J. & MARSHALL D.L. (1997): Reproductive success increases with localdensity of conspecifics in a desert mustard (Lesquerella fendleri). Conservation Biol. 11: 738–746.
SABAT A.M. & ACKERMAN J.D. (1996): Fruit set in a deceptive orchid: the effect of flowering phenology,display size, and local floral abundance. Amer. J. Bot. 83: 1181–1186.
SOKAL R.R. & ROHLF F.J. (1997): Biometry: The principles and practice of statistics in biological research. Ed.3. W.H. Freeman and Company, New York.
TREMBLAY R.L. (1997a): Distribution and Dispersion patterns of individuals in nine species of Lepanthes
(Orchidaceae). Biotropica 29: 38–45.
Life cycle stages in a lithophytic orchid 119

TREMBLAY R.L. (1997b): Morphological variance among populations of three tropical orchids with restrictedgene flow. Pl. Spec. Biol. 12: 85–96.
TREMBLAY R.L. (1997c): Lepanthes caritensis, an endangered orchid: no sex, no future? Selbyana 18: 160–166.TREMBLAY R.L. (2000): Plant longevity in four species of Lepanthes (Pleurothallidinae, Orchidaceae).
Lindleyana 15: 257–266.TREMBLAY R.L. & ACKERMAN J.D. (2001): Gene flow and effective population size in Lepanthes
(Orchidaceae): A case for genetic drift. Biol. J. Linn. Soc. 72: 47–62.TREMBLAY R.L. & ACKERMAN J.D. (2003): The genetic structure or orchid populations and its evolutionary
importance. Lankersteriana 7: 87–92.TREMBLAY R.L. & HUTCHINGS M.J. (2004): Population dynamics in orchid conservation: a review of analytical
methods based on the rare species Lepanthes eltoroensis. In: DIXON K., CRIBB P., KELL S. & BARRETT R.(eds.), Orchid conservation, Kluwer Academic Press, Perth, pp. 183–204.
TREMBLAY R.L., ACKERMAN J.D., ZIMMERMAN J.K. & CALVO R. (2005): Variation in sexual reproduction inOrchids and its evolutionary consequences: A spasmodic journey to diversification. Biol. J. Linn. Soc. 84:1–54.
VAN TREUREN R., BIJLSMA R., OUBORG N. J. & VAN DELDEN W. (1993): The effects of population size andplant density on outcrossing rates in locally endangered Salvia pratensis. Evolution 47: 1094–1104.
VAN TREUREN R., BIJLSMA R., OUBORG N. J. & KWAK M.M. (1994): Relationships between plant density,outcrossing rates and seed set in natural and experimental populations of Scabiosa columaria. J. Evol. Biol. 7:287–302.
WAIDE R.B. & REAGAN D.P. (1996): The rainforest setting. In: REAGAN D.P. & WAIDE R.B. (eds.), The food
web of a tropical rainforest, The University of Chicago Press, Chicago, pp. 1–16.WAITE S. & FARRELL L. (1998): Population biology of the rare military orchid (Orchis military L.) at en
established site in Suffolk, England. Bot. J. Linn. Soc. 126: 109–121.WILLEMS J.H. & BIK L. (1991): Long-term dynamics in a population of Orchis simia in the Netherlands. In:
WELLS T.C.E. & WILLEMS J.H. (eds.), Population ecology of terrestrial orchids, SPB Academic Publishers.The Hague, pp. 33–45.
WILLEMS J.H. & MELSER C. (1998): Population dynamics and life history of Coeloglossum viride (L.) HATM.An endangered orchid species in the Netherlands. Bot. J. Linn. Soc. 126: 83–93.
WOLFE L.M. (1987): Inflorescence sizes and pollinaria removal in Asclepias curassavica and Epidendrum
radicans. Biotropica 19: 86–89.
Received 15 November 2004, revision received and accepted 29 August 2005
120 N. Rivera Gómez et al.




















