
Inorganic and organic nitrogen uptake by the toxigenic diatom Pseudo-nitzschia australis...
38
Accepted Manuscript Title: Inorganic and Organic Nitrogen Uptake by the Toxigenic Diatom Pseudo-nitzschia australis (Bacillariophyceae) Authors: William P. Cochlan, Julian Herndon, Raphael M. Kudela PII: S1568-9883(08)00100-5 DOI: doi:10.1016/j.hal.2008.08.008 Reference: HARALG 445 To appear in: Harmful Algae Received date: 8-5-2007 Revised date: 2-1-2008 Accepted date: 1-8-2008 Please cite this article as: Cochlan, W.P., Herndon, J., Kudela, R.M., Inorganic and Organic Nitrogen Uptake by the Toxigenic Diatom Pseudo-nitzschia australis (Bacillariophyceae), Harmful Algae (2007), doi:10.1016/j.hal.2008.08.008 This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Transcript of Inorganic and organic nitrogen uptake by the toxigenic diatom Pseudo-nitzschia australis...

Accepted Manuscript
Title: Inorganic and Organic Nitrogen Uptake by the ToxigenicDiatom Pseudo-nitzschia australis (Bacillariophyceae)
Authors: William P. Cochlan, Julian Herndon, Raphael M.Kudela
PII: S1568-9883(08)00100-5DOI: doi:10.1016/j.hal.2008.08.008Reference: HARALG 445
To appear in: Harmful Algae
Received date: 8-5-2007Revised date: 2-1-2008Accepted date: 1-8-2008
Please cite this article as: Cochlan, W.P., Herndon, J., Kudela, R.M., Inorganicand Organic Nitrogen Uptake by the Toxigenic Diatom Pseudo-nitzschia australis(Bacillariophyceae), Harmful Algae (2007), doi:10.1016/j.hal.2008.08.008
This is a PDF file of an unedited manuscript that has been accepted for publication.As a service to our customers we are providing this early version of the manuscript.The manuscript will undergo copyediting, typesetting, and review of the resulting proofbefore it is published in its final form. Please note that during the production processerrors may be discovered which could affect the content, and all legal disclaimers thatapply to the journal pertain.

Page 1 of 37
Accep
ted
Man
uscr
ipt
Inorganic and Organic Nitrogen Uptake by the Toxigenic Diatom 1
Pseudo-nitzschia australis (Bacillariophyceae) 2
3
4
William P. Cochlan1, Julian Herndon1, and Raphael M. Kudela2 5
6
1Romberg Tiburon Center for Environmental Studies, San Francisco State University, 3152 7
Paradise Drive, Tiburon, California 94920-1250, USA 8
9
2Ocean Science Department, University of California, Santa Cruz, 1156 High Street, Santa Cruz, 10
California 95064, USA 11
12
Corresponding Author: W. Cochlan 13
Phone: 415-338-3541 14
FAX: 415-435-7120 15
E-mail: [email protected] 16
Correspondence Address: as above 17
Manuscript submitted to Harmful Algae Special Issue on: May 8, 2007 18
Manuscript Revision on: December 21, 2007 19
20

Page 2 of 37
Accep
ted
Man
uscr
ipt
2
Abstract 1
2
The nitrogen uptake capabilities of the toxigenic diatom Pseudo-nitzschia australis (Frenguelli), 3
freshly isolated from Monterey Bay California, were examined in uni-algal laboratory cultures at 4
saturating photosynthetic photon flux densities (100 µmol photons m-2 s-1) and 15° C. The 5
kinetics of nitrogen (nitrate, ammonium, urea and glutamine) uptake as a function of substrate 6
concentration were estimated from short (20.5-min) incubations using the 15N-tracer technique. 7
Based on the estimated maximum specific uptake rates and measures of N affinity (the initial 8
slope of the uptake versus nutrient concentration curve), nitrate is the preferred nitrogen 9
substrate, followed by glutamine and ammonium, which are equivalent. Rates of urea uptake by 10
P. australis did not saturate at concentrations as high as 36 µg-at N L-1, and urea uptake as a 11
function of concentration could not be described by Michaelis-Menten kinetics over the 12
concentration gradient tested. Although there is a clear preference for nitrate at equivalent 13
concentrations (compared to ammonium, urea, and glutamine), these laboratory results 14
demonstrate the capability of this pennate diatom to utilize both inorganic and organic forms of 15
nitrogen, supporting field observations that P. australis blooms during both upwelling and non-16
upwelling conditions off the west coast of North America. Substantial differences in the 17
nitrogenous nutrition of P. australis can be expected in these environments, and anthropogenic 18
inputs of N substrates such as ammonium and urea can support growth and may contribute 19
significantly to both harmful diatom blooms and the maintenance of seed populations at non-20
bloom abundances, particularly during periods of reduced or absent upwelling. 21
22
Key Words: ammonium, Bacillariophyceae, glutamine, diatom, nitrate, nitrogen uptake kinetics, 23
Pseudo-nitzschia australis, urea 24

Page 3 of 37
Accep
ted
Man
uscr
ipt
3
1
1. Introduction 2
3
Coastal diatoms from the genus Pseudo-nitzschia Pergallo (Bacillariophyceae) have 4
received considerable attention since the discovery of domoic acid (DA) production by the 5
pennate diatom Pseudo-nitzschia multiseries (then termed Nitzschia pungens f. multiseries Hasle 6
1995) (Subba Rao et al., 1988; Bates et al., 1989). When accumulated through trophic transfer, 7
this neurotoxic amino acid is responsible for illness and mortality of marine mammals, seabirds 8
and humans, and has caused significant economic losses to shellfish and planktivorous fisheries 9
since its discovery in Atlantic Canada in 1987 (cf., review by Bates et al., 1998). Currently 10
eleven or twelve species of Pseudo-nitzschia are thought to synthesize DA (Bates and Trainer, 11
2006), and a number of factors have been identified that affect growth and toxin production in 12
Pseudo-nitzschia. Numerous laboratory experiments using isolates of Pseudo-nitzschia have 13
demonstrated that macronutrient (Si and P) limitation and micronutrient (Fe, Cu) limitation, as 14
well as elevated Cu conditions, can enhance the production of DA. In addition, elevated pH 15
(Lundholm et al., 2004), the phase of cell growth (stationary versus exponential), availability of 16
light, and appropriate temperature have all been shown to influence DA production in culture 17
(see review by Bates and Trainer, 2006). However, little is known about the nitrogenous 18
nutrition of Pseudo-nitzschia species, from either cultured populations or field assemblages. 19
Nitrogen is required for the growth of all phytoplankton, and is the macronutrient thought 20
to limit the growth of natural phytoplankton assemblages in most coastal regions (e.g., Ryther 21
and Dunstan, 1971). Nitrogen is also a necessary component for synthesis of domoic acid and all 22
other amino acids. Based on both the inherent requirement for nitrogen to synthesize domoic 23

Page 4 of 37
Accep
ted
Man
uscr
ipt
4
acid, and extensive laboratory experiments using predominantly P. multiseries, it is generally 1
assumed that plentiful N concentrations in the medium (or seawater) are a prerequisite for toxin 2
production (cf. review by Bates, 1998). Unlike P- or Si-limitation, growth limitation by N is 3
directly unfavorable for DA production in cultures (Bates et al., 1991; but see Kudela et al., 4
2004a,b for evidence of reduced but measurable DA production under N-limiting conditions in 5
the laboratory and field), and the form of nitrogen also appears to influence cell growth and DA 6
production. An early study showed that at concentrations above 200 µM, ammonium promotes 7
greater DA production than nitrate during stationary phase for P. multiseries, but at lower 8
concentrations (110 and 55 µM), cell yield and DA production were equivalent for both N 9
substrates (Bates et al., 1993). In a subsequent study, Hillebrand and Somer (1996) found that 10
inorganic (nitrate, nitrite) and organic (urea, glutamine) N, when supplied at 200 µM, supported 11
similar growth rates of P. multiseries at low light levels (25 µmol photons m–2 s–1); rates that 12
were consistently greater than those obtained using ammonium as the sole N source. Whereas at 13
high light (230 µmol photons m–2 s–1), both the carrying capacity and the specific growth rate 14
were severely depressed when grown on ammonium versus nitrate. At more ecologically 15
relevant N concentrations (50 µM), others have found that nitrate- and ammonium-grown cells 16
produce equivalent amounts of DA for P. australis in exponential phase, but that DA production 17
is enhanced when urea is the sole N source; the growth rate is significantly reduced when 18
provided with only urea-N, however (Cochlan et al., 2006a; Howard et al., 2007). 19
In coastal regions where freshwater inputs are substantial, it is not unusual to expect 20
elevated concentrations of ammonium, as well as dissolved organic N forms such as urea and 21
other less characterized DON (e.g., Berman and Bronk, 2003) from urban and agricultural 22
sources. The apparent increase in the magnitude and frequency of HABs around the world are 23

Page 5 of 37
Accep
ted
Man
uscr
ipt
5
frequently attributed to cultural eutrophication, specifically to the effects of anthropogenic 1
nutrient sources on the growth of harmful algal bloom species (cf. reviews by Hallegraeff, 1993; 2
Anderson et al., 2002; Glibert et al., 2005). However in the case of toxigenic diatom blooms, the 3
link between eutrophication and Pseudo-nitzschia is more difficult to discern. For example in 4
the Gulf of Mexico, although the sedimentary record reveals a correlation between nitrate fluxes 5
and Pseudo-nitzschia abundance (Parsons et al., 2002), observational associations of non-toxic 6
blooms of Pseudo-nitzschia in the early 1990s were negatively related to ambient concentrations 7
of all macronutrient concentrations, including nitrate and ammonium (Dortch et al., 1997). 8
Similarly the abundances of Pseudo-nitzschia spp. and the concentrations of DA in the Southern 9
California Bight were generally found to be inversely related to ambient concentrations of 10
phosphate, nitrate and silicic acid, despite sampling after major rainfall events which contributed 11
to substantial river discharge from this heavily urbanized region (Schnetzer et al., 2007). 12
On the west coast of the USA, the association of anthropogenic nutrients and toxigenic 13
diatom blooms are very rare, although there have been relatively few studies examining these 14
linkages (see review by Anderson et al., this volume). Elevated ammonium levels, ostensibly 15
from sewage inputs, have been suggested as the triggering factor for dense (up to 23 x 106 16
cells L-1) blooms of P. pseudodelicatissima in Sequim Bay, WA (Trainer et al., 2007), and 17
Ladizinsky (2003) suggested that enhanced domoic acid production in Monterey Bay, CA was 18
associated with excess Cu from anthropogenic sources. However coastal upwelling appears to 19
be the dominant forcing factor for toxigenic diatom bloom development along the USA west 20
coast (e.g., Horner et al., 2000; Trainer et al., 2000; Kudela et al., 2004a, 2005; Anderson et al., 21
2006) where frequent blooms of Pseudo-nitzschia spp., most commonly P. australis and P. 22
multiseries are found in nitrate-replete waters. Despite the importance of nitrogen dynamics in 23

Page 6 of 37
Accep
ted
Man
uscr
ipt
6
coastal phytoplankton ecology, the critical processes of nutrient uptake and growth are rarely 1
measured in the field even during such uni-algal blooms. Given the episodic nature of coastal 2
upwelling relative to the longevity of these blooms, it may prove difficult to establish a direct 3
relationship between Pseudo-nitzschia abundance and nutrients, based only on ambient nutrient 4
conditions. 5
To date our understanding of the nitrogenous nutrition of Pseudo-nitzschia spp. is 6
severely limited. Quantification of the physiological capacity for N-uptake by Pseudo-nitzschia 7
spp. under both N-replete and -depleted conditions is a necessary prerequisite for an 8
understanding of the success of Pseudo-nitzschia spp. relative to other phytoplankton in these 9
coastal systems, and will help to identify the proximate causes of toxin production by this 10
organism. Here we describe the inorganic and organic N uptake capabilities of P. australis 11
isolated from a well-known toxic ‘hot spot’ on the USA west coast – Monterey Bay, California. 12
This is the first study to provide a full suite of N uptake kinetic parameters for this toxigenic 13
diatom, which is often the dominant toxin-producing organism from the Pseudo-nitzschia genus 14
in California. 15
16
2. Materials and Methods 17
2.1 Cell culturing 18
Unialgal batch cultures of P. australis (culture AU-03184-5D, isolated from Monterey 19
Bay, CA, USA by G.J. Smith) were grown in filter-sterilized (0.2-µm, Polycap™, 150 TC, 20
Whatman®) Monterey Bay enriched seawater following Berges et al. (2001 and subsequent 21
Corrigendum 2004), with the following modifications. Metals Stock I: FeCl3•6H2O, 1.77 g L-1, 22
Na2-EDTA, 2.44 g L-1; Metals Stock II: Na2-EDTA, 3.09 g L-1, MnSO4•H2O, 0.409 g L-1, 23

Page 7 of 37
Accep
ted
Man
uscr
ipt
7
Na2MoO4•2H2O, 0.0148 g L-1, NiCl2•6H2O, 0.0149 g L-1. Copper, as CuSO4•5H2O, was added to 1
the medium to achieve a final concentration of 8 x 10-8 M. Selenium was prepared as a separate 2
stock (Na2SeO3, 0.0022 g L-1), and added to the medium to achieve a concentration of 6.36 x 10-9 3
M. Both metal stocks were added at double the concentrations of Berges et al. (2001); all other 4
enrichments were unchanged except silicic acid which was doubled to ensure a minimum Si:N 5
ratio of 2:1. Nitrate, the sole nitrogen source was reduced from 550 to 70 µg-at N L-1. 6
Cultures were maintained at 15° C (± 0.3° C) within an environmental chamber, and 7
continuously illuminated from one side by four Vita-Lite® Plus Power Twist fluorescent tubes 8
(Duro-Test® Corporation; Color Rendering Index-91). Average photosynthetic photon flux 9
density (PPFD), measured with a scalar collector (Biospherical Instruments QSL-100 quantum 10
scalar irradiance meter) immersed within medium-filled culture vessels, was 100 µmol photons 11
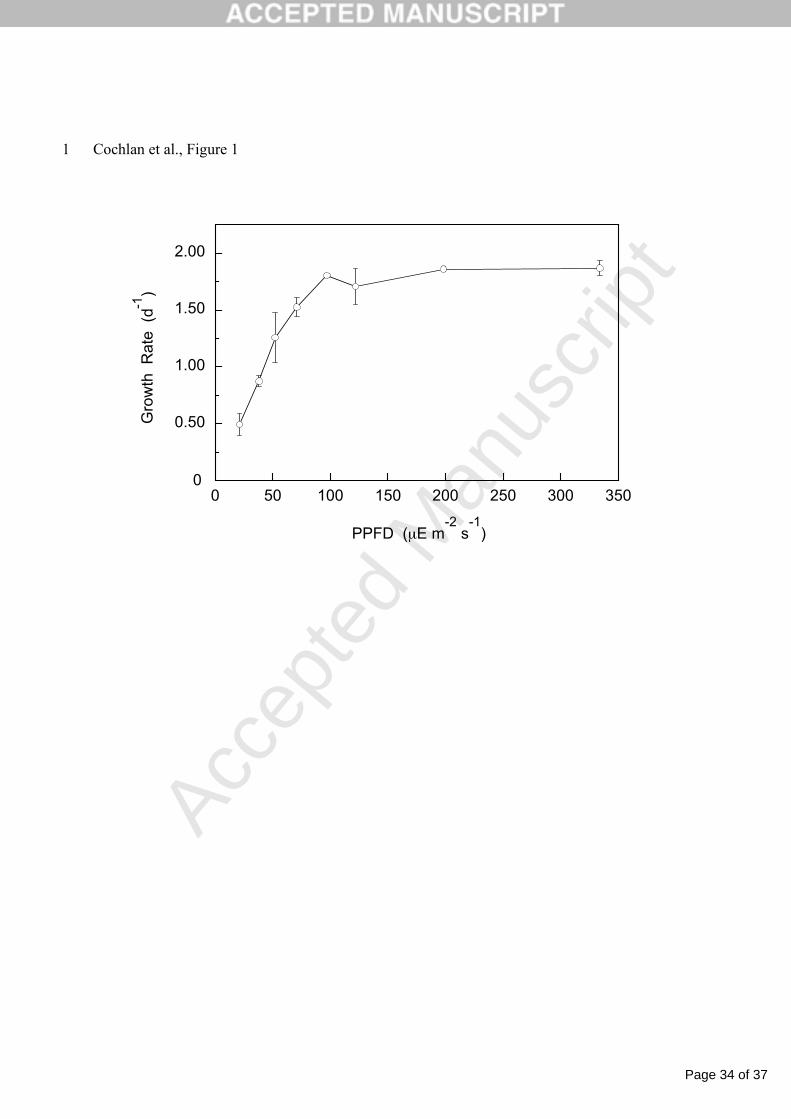
m–2 s–1. This irradiance level was experimentally determined to be saturating for the growth of P. 12
australis (UCSC culture 0771B) at 13-14° C (B.D. Bill, unpublished data; Figure 1), and is the 13
recommended minimum irradiance for optimizing growth and toxin production in P. multiseries 14
(Bates, 1998). Cell growth was monitored by abundance; daily samples were fixed in 1% acidic 15
Lugol’s solution and counted using a Palmer-Maloney nanoplankton chamber at 100X using a 16
phase contrast microscope (Eclipse E4000, Nikon®). Sterile technique was employed to prevent 17
fungal growth and to minimize bacterial contamination. Transfers and pre-experimental culture 18
manipulation were conducted in a positive pressure, laminar flow hood (Envirco Corporation 19
MAC 10® HEPA filtration unit). The glass flasks used for culturing the diatoms, and vessels 20
used for storing enriched seawater, were first washed in dilute detergent (Contrad 70®; Decon 21
Laboratories, Inc.), soaked in weak hydrochloric acid (5% v/v) for at least 24 hours, rinsed 22
thoroughly with ultra-pure water (18.2 MΩ·cm; Milli-Q®), and autoclaved prior to use. 23

Page 8 of 37
Accep
ted
Man
uscr
ipt
8
1
2.2 Analytical methods 2
Samples for macronutrients (nitrate plus nitrite [hereafter referred to as nitrate], silicate 3
and ortho-phosphate) were stored frozen, and analyzed with a Lachat Quick Chem 8000 Flow 4
Injection Analysis system using standard colorimetric techniques (Knepel and Bogren, 2001; 5
Smith and Bogren, 2001a, b). Ammonium and urea samples were collected using 50-mL, low-6
density polyethylene (LDPE) centrifuge tubes (Corning®). Ammonium samples were stored 7
refrigerated after the addition of the phenolic reagent; the addition of the phenolic reagent binds 8
NH4+ and eliminates the need to freeze samples (Degobbis, 1973). The remaining reagents were 9
added within 24 hours, and the samples were manually analyzed using a spectrophotometer 10
equipped with a 10-cm cell (Solórzano, 1969). Urea samples were frozen at -20° C, and 11
subsequently thawed at room temperature before manual analysis using the diacetyl monoxime 12
thiosemicarbizide technique (Price and Harrison, 1987), modified to account for a longer color 13
development period (72 h) and lower digestion temperature (22˚C). 14
15
2.3 Experimental procedures 16
N uptake kinetics were determined for four nitrogen substrates (NO3-, NH4
+, glutamine, 17
and urea), but all cells were grown on NO3- in duplicate, 3-L Erlenmeyer glass flasks (Pyrex®) 18
prior to the kinetic experiments. Cultures were sampled daily, using sterile, disposable 19
serological pipettes to monitor cell abundance and ambient nitrogen concentrations. Kinetic 20
experiments were initiated immediately after the ambient NO3- + NO2
- concentration in the 21
cultures was below detection limits (ca. 0.05 µg-at N L-1). Growth rates for cultures averaged 22
1.0-1.2 d-1 during the 4-5 days prior to N depletion and the initiation of kinetic experiments. 23

Page 9 of 37
Accep
ted
Man
uscr
ipt
9
Timing was important, because NO3- in the medium must be depleted to ascertain the uptake 1
response to low concentrations of N, and to eliminate any effects of simultaneous uptake of NO3- 2
on the uptake response of the N substrate being tested, but the cells must not be N starved to 3
avoid non-linearity in uptake over time observed previously for other diatoms (e.g., Conway et 4
al., 1976), including species of Pseudo-nitzschia (Auro, 2007). 5
Sub-samples (25 mL) were transferred from the 3-L growth flasks to a series of 50-mL 6
borosilicate (Pyrex®) culture tubes equipped with polypropylene screw caps and inoculated with 7
15N-ammonium chloride, (98.85 atom %; Cambridge Isotopes), 15N-sodium nitrate (98.25 atom 8
%), 15N-urea (98.2 atom %), or 15N-glutamine (98.4% atom %) at a range of initial substrate 9
concentrations: 0.1, 0.2, 0.4, 0.8, 1.6, 2.4, 4.2, 5.9, 12, 18, and 36 µg-at N·L-1 (plus 70 µg-at N L-10
1 for glutamine) with two replicates at each concentration. All N concentrations are reported in 11
N equivalent units (µg-at N L-1) to avoid confusion associated with molar units (i.e., 1 µM of 12
urea or glutamine = 2 µg-at N L-1). Incubations were terminated after 20.5 min by filtration (50-13
70 mm Hg) onto combusted Whatman GF/F filters (0.7 µm nominal pore size) and frozen in 14
polypropylene cryovials; the filtration period was consistently 30s or less. After thawing/drying 15
(<50° C for minimum of 24 h) the filters were analyzed for particulate nitrogen (PN) 16
concentration and isotopic enrichment with a Europa Scientific PDZ 20/20 mass spectrometer. 17
Due to mass spectrometry difficulties, many samples were lost and replicate kinetic experimental 18
data are only available for urea. 19
Based on the isotopic enrichment and concentration of the 15N-substrate added, and the 20
atom % excess and PN concentration collected at the end of the short incubations, it was 21
calculated that on average (±1 SD) 10.9 ± 7.7% (n = 18), 10.6 ± 6.1% (n = 19), 3.6 ± 4.8% (n = 22

Page 10 of 37
Accep
ted
Man
uscr
ipt
10
43), and 9.7 ± 13.9% (n = 22) of the 15N-substrate was incorporated into PN during the 15NH4+, 1
15NO3-, 15N-urea, and 15N-glutamine kinetic experiments, respectively. Even at the lowest N 2
substrate enrichment (0.10-0.20 µg-at N L-1), only an average of 19.3 ± 12.6% (n = 15) was used. 3
Substrate depletion was therefore not considered a problem in these experiments, even at the 4
lowest N enrichments. At the start of each kinetic experiment, heterotrophic bacteria in the GF/F 5
filtrate and unfiltered cultures were enumerated by epifluorescence microscopy using acridine 6
orange, and bacterial productivity was estimated from the incorporation rate of 3H-leucine (20 7
nM final concentration, 180 Ci mmol-1
, Amersham) following the procedures outlined in 8
Cochlan (2001). 9
10
2.4 Calculations 11
Nitrogen specific uptake rates (V [h-1], N taken up per unit PN) were estimated from the 12
accumulation of 15N in the particulate material, and calculated according to a constant specific 13
uptake model (equation #6 of Dugdale and Wilkerson, 1986). Rates were not corrected for the 14
effects of isotopic dilution (Glibert et al., 1982) as these are expected to be minimal during the 15
extremely short incubation times and in the absence of micro-grazers. Reported glutamine 16
uptake rates are conservative in that the ambient concentration of glutamine is assumed to be 17
zero (Eppley et al., 1977), and it is also assumed that there is no discrimination in the uptake of 18
the 15N-labeled amide N versus the non-labeled, amino-N of the glutamine molecule. 19
Curve fitting was completed using an iterative, non-linear least-squares technique 20
(KaleidaGraph®; Abelbeck Software), which utilizes the Levenberg-Marquardt algorithm (Press 21
et al., 1992). The N kinetics data were fitted directly to the Michaelis-Menten formulation: 22
23

Page 11 of 37
Accep
ted
Man
uscr
ipt
11
V =Vmax • SKs + S
(1) 1
2
Where: V is the PN specific uptake rate (h-1), Vmax is the maximal specific uptake rate (h-1), S is 3
the substrate concentration (µg-at N L-1), and Ks is the half-saturation constant for the N 4
substrate (µg-at N L-1). The substrate affinity at low concentrations (i.e., S < Ks) was determined 5
from the initial slope (α) of the Michaelis-Menten plot, and calculated as α = Vmax/Ks; the 6
derivative of Equation 1, with respect to S, as S approaches zero (Button, 1978; Healey, 1980). 7
8
3. Results 9
10
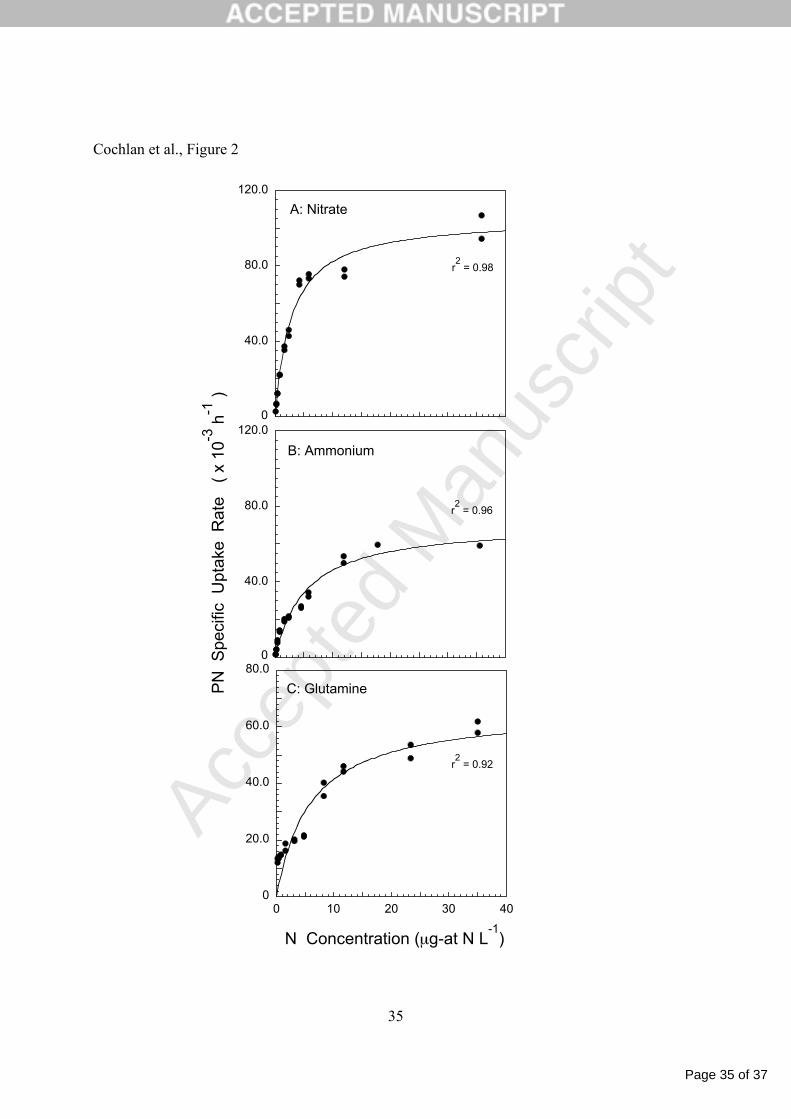
The uptake rates of NO3-, NH4
+ and glutamine by P. australis can be related to their 11
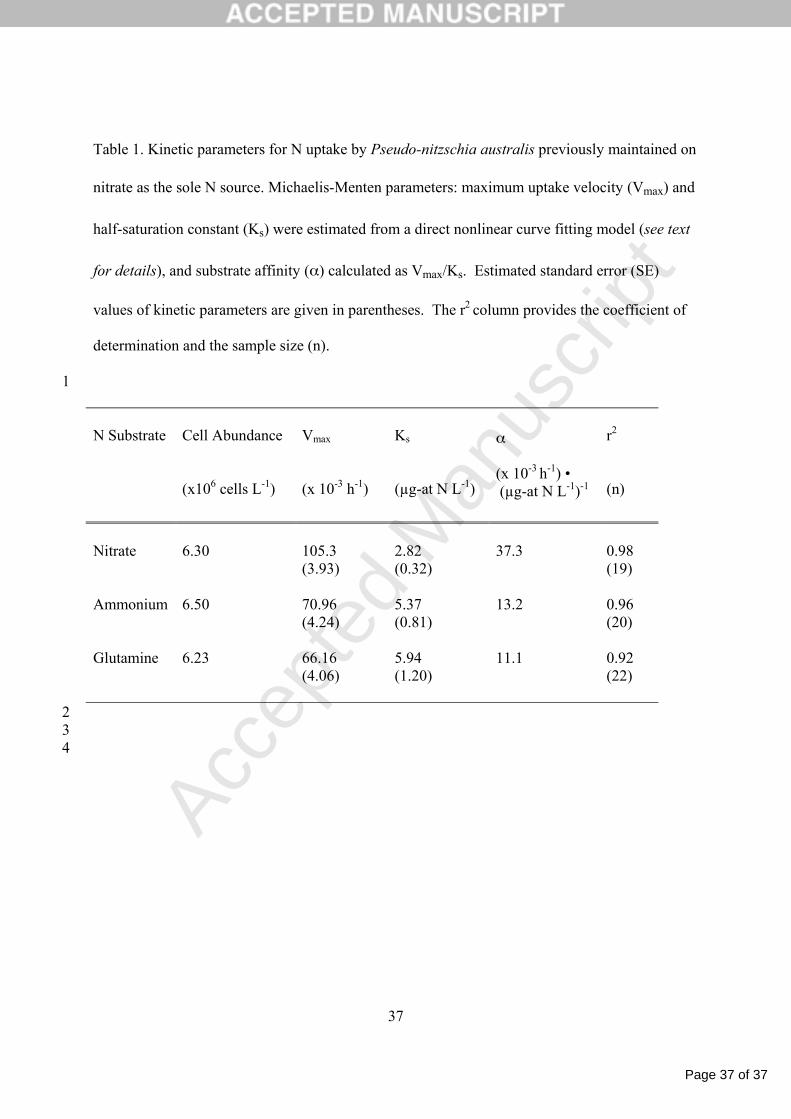
external substrate concentration following Michaelis-Menten type kinetics (Table 1, Figure 2). 12
In this study, during 20.5-min incubations, the specific NO3-, NH4
+, and glutamine uptake rates 13
can be effectively described as a hyperbolic function of their concentration, and saturate at 14
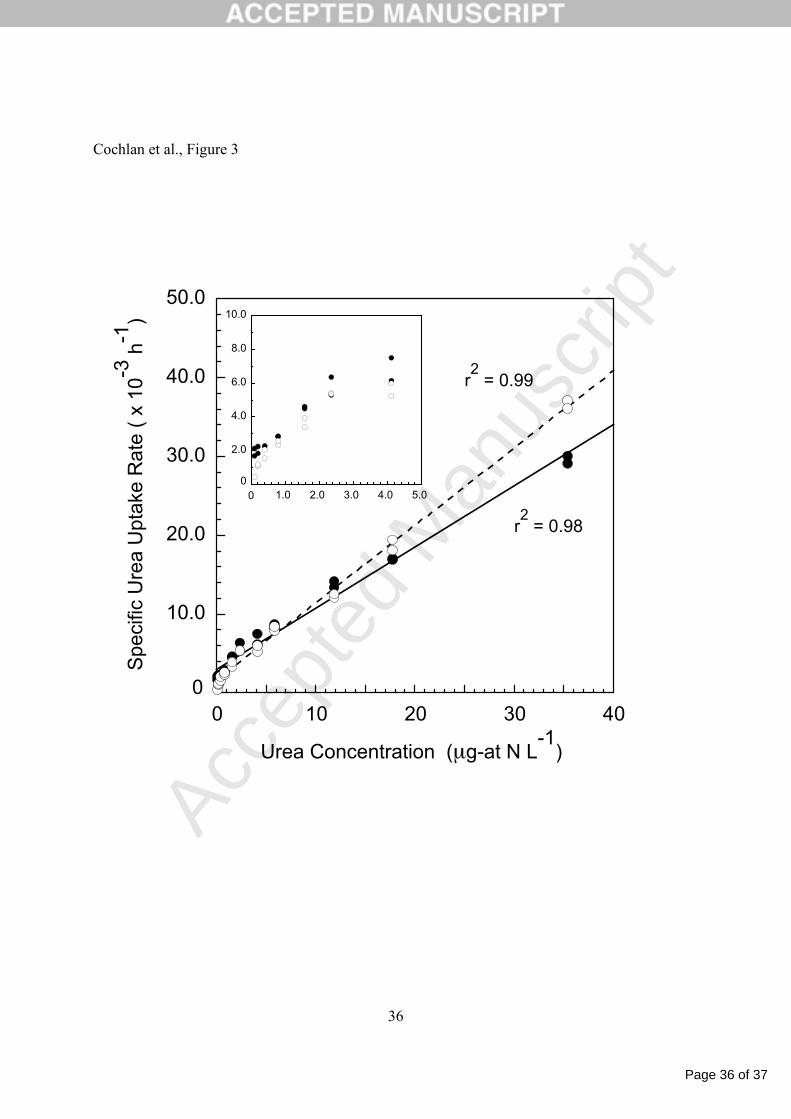
concentrations at or below the highest N concentration employed (35-36 µg-at N L-1). However, 15
uptake rates of urea could not be described similarly, and specific urea uptake rates are a linear 16
function of the ambient urea concentration over the range of substrate employed here; specific 17
rates do not demonstrate any trend toward saturation at the greatest urea concentrations (Figure 18
3). 19
A comparison of the maximum uptake rates (Vmax) indicate that the substrate-saturated 20
rate of NO3- uptake is greatest, followed by uptake rates of NH4
+ and glutamine, with PN-21
specific rates of 105.3, 71.0 and 66.2 x 10-3 h-1 for NO3-, NH4
+, and glutamine, respectively. For 22

Page 12 of 37
Accep
ted
Man
uscr
ipt
12
comparison, the uptake rate of urea at 40 µg-at N L-1, calculated from the least-squares linear 1
regression of uptake versus concentration data from replicate cultures, is 30.4 x 10-3 h-1. The 2
half-saturation constants (Ks) increase in order: NO3-, NH4
+, glutamine, and are 2.82, 5.37 and 3
5.94 µg-at N L-1, respectively, whereas a Ks value for urea could not be ascertained. The α 4
values for NO3-, NH4
+ and glutamine follow the same order as the Ks values, the traditional 5
measure of affinity, but decrease in value as 37.3, 13.2, 11.1 x 10-3 h-1/µg-at N L-1, respectively. 6
For comparison, the initial slope (urea < 2.4 µg-at N L-1) of the urea uptake vs. concentration 7
plot is only 2.8 x 10-3 h-1/µg-at N L-1, suggesting that the affinity of P. australis for urea is less 8
than for the other N substrates tested. 9
Although not quantified, it is possible that the overall lower uptake rates of urea observed 10
for P. australis were due to loss of labeled urea as NH3/NH4+ from the cells to the surrounding 11
medium. NH3/NH4+ release following urea uptake and assimilation has been observed for other 12
phytoplankton, including the dinoflagellate Prorocentrum micans (Uchida, 1976), and the 13
diatoms Phaeodactylum tricornutum (Rees and Bekheet, 1982) and Thalassiosira pseudonana 14
(Price and Harrison, 1988). In the present study, however, samples for NH4+ analyses were not 15
taken from the culture medium following the 15N-urea enrichments, so it is not known whether 16
loss of labeled N contributed to the lower uptake rates measured. 17
The combusted GF/F filters employed in this study appear very effective at retaining 18
bacteria enumerated using epifluorescence microscopy. At the initiation of each kinetic 19
experiment, the heterotrophic bacterial abundance in GF/F filtrates averaged only 3% of the raw 20
(unfiltered) abundance, indicating that 97% of the bacteria were captured during filtration of the 21
replicate 25-mL subsamples of P. australis cultures for 15N analyses. P. australis cell abundance 22
in these replicate cultures ranged from 5.59 to 6.50 x 106 cells L-1, and the rates of bacterial 23

Page 13 of 37
Accep
ted
Man
uscr
ipt
13
carbon production averaged 0.83 to 1.49 µg C h-1. The contribution of heterotrophic bacteria to 1
the specific uptake rates reported in this study can be estimated from the bacterial carbon 2
production rates, assuming a C:N ratio of 3.7 (5.6 fg N cell-1; Lee and Fuhrman 1987) and 3
correcting for the 97% retention rate of the combusted GF/F filters. With these assumptions, the 4
potential bacterial contribution to the reported uptake rates was minimal, and averaged only 4-5
6% of the measured NO3-, glutamine and NH4
+ uptake rates, but was a complicating factor in the 6
urea uptake rates, if one assumes that all the bacterial N demands are met by urea alone. 7
Bacterial N demand averaged 22% for the pooled urea-N experiments, but exhibited a 8
concentration-dependent effect; at higher urea concentrations (> 0.8 µg-at N L-1) only 7-14% of 9
the total urea-N uptake was potentially attributed to the heterotrophic bacterial fraction due to the 10
greater urea uptake rates measured at these concentrations. 11
12
4. Discussion 13
14
Diatoms of the genus Pseudo-nitzschia have received considerable attention in the past 15
two decades because of their production of the neurotoxin domoic acid, its potentially harmful 16
effects to marine mammals, birds and humans, and the resulting negative impacts on commercial 17
and recreational shellfish and crab fisheries. The west coast of the USA has been at the center of 18
this interest due to the development of large toxic blooms in central (e.g., Scholin et al., 2000; 19
Trainer et al., 2000), and southern California (e.g., Anderson et al., 2006; Schnetzer et al., 2007) 20
as well as off the coast of Washington (e.g., Trainer et al., 2002) many of which consist almost 21
entirely of P. australis. The role of macronutrients in supporting these blooms and triggering 22
toxin production has, surprisingly, received little attention besides the measurement of ambient 23

Page 14 of 37
Accep
ted
Man
uscr
ipt
14
nutrients and attempts to relate these to the magnitude of toxin production and/or Pseudo-1
nitzschia biomass. To date, the only strong correlate appears to be the association of elevated 2
biomass (and sometimes high DA) and silicic acid depletion (e.g., Anderson et al., 2006), 3
although this is not a uniform trend for all west coast Pseudo-nitzschia assemblages (e.g., 4
Cochlan et al., 2006b; Marchetti et al., 2004; Trainer et al., 2007). 5
The measurement of nutrient uptake rates by natural blooms of Pseudo-nitzschia are still 6
lacking, and besides the recent results of N-amended grow-out experiments (Howard et al., 7
2007), there are no published reports of differential growth and toxin response to inorganic and 8
organic N substrates in natural assemblages of Pseudo-nitzschia spp. Despite the importance of 9
quantifying N nutrition as a means to help explain HAB dynamics, there have also been no 10
previous studies of the kinetics of N utilization for any species of Pseudo-nitzschia. Our study 11
thus provides the first N kinetic parameters of inorganic and organic nitrogen use by P. australis, 12
and demonstrates the potential importance of organic N substrates in fueling toxigenic diatom 13
growth under eutrophied conditions. 14
The kinetic parameters of N uptake can be used to assess the relative preference (and 15
affinity) of one N substrate over another in low and high N environments, and species-specific N 16
uptake kinetic parameters may then be used to explain species competition when N is the 17
limiting nutrient. The reader is cautioned not to confuse these measures of N preference with the 18
relative preference index (RPI, McCarthy et al., 1977), which compares the relative utilization of 19
a particular N substrate to the relative availability of that N substrate in the water. In the present 20
study, the Vmax-NO3- for exponentially growing P. australis cultures were ~1.5 fold greater than 21
the Vmax of NH4+ and glutamine, which were equivalent, and triple the maximum uptake of urea. 22
The latter assumes that specific urea uptake rate at 40 µg-at N·L-1 reflects a maximal rate (but see 23

Page 15 of 37
Accep
ted
Man
uscr
ipt
15
below). Based on these individual N substrate uptake versus concentration experiments, the N 1
preference of P. australis follows the order: NO3- > (NH4
+, glutamine) > urea when exposed to 2
high N conditions; this may explain the repeated occurrence of Pseudo-nitzschia blooms in the 3
high NO3- waters associated with upwelling on the USA west coast. Concentrations of the 4
organic N substrates are generally low in such upwelling zones, but during periods of heavy 5
runoff, both agricultural and urban sources may result in elevated concentrations that could 6
support the maximal uptake rates reported here (see Kudela et al., 2007a this volume). 7
Glutamine, although not routinely measured in the field and expected to be present in very low 8
concentrations, was included in this study as a representative organic N substrate of 9
anthropogenic origin known to support Pseudo-nitzschia growth (Hillebrand and Sommer, 1996; 10
Mengelt, 2005) and when available, glutamine is utilized at equivalent rates (based on the Vmax 11
values) as NH4+ by this diatom. 12
The half-saturation constants, which are traditionally used to indicate nutrient affinity 13
under nutrient-limited conditions, are once again equivalent for NH4+ and glutamine, and double 14
the lower Ks values reported for NO3-. These Ks values could be interpreted as the cells’ more 15
effective capability to utilize low concentrations of NO3- than either NH4
+ or glutamine. 16
However, the interpretation of Ks as measure of affinity is complicated by the functional 17
relationship between Vmax and Ks. Combining them into a ratio (α = Vmax/Ks; the initial slope of 18
the uptake versus substrate concentration curve) emphasizes both factors, and provides a more 19
descriptive metric of N affinity at sub-saturating concentrations (< Ks) and when inter-species 20
competition is likely to occur (e.g., Healey, 1980; Harrison et al., 1989). 21

Page 16 of 37
Accep
ted
Man
uscr
ipt
16
In the present study, the affinity coefficient (α) for NO3- is three-fold higher than the α 1
values for NH4+ or glutamine, and 10-fold greater than the initial slope of the urea uptake curve. 2
These results demonstrate that P. australis can utilize low (sub-saturating) concentrations of 3
NO3- more effectively than equivalent concentrations of the other N substrates, and that N 4
preference under sub-saturating N concentrations also follows the order NO3- > (NH4
+ and 5
glutamine) > urea. Although it is difficult to directly ascribe results determined under optimal 6
laboratory conditions to those occurring during natural blooms of P. australis, these results 7
suggest that under both conditions of low and high ambient N representative of oligotrophic and 8
eutrophic areas, respectively, NO3- is acquired more readily than NH4
+ and glutamine, and urea 9
is the least preferred of the N substrates tested. However, all of the N substrates tested were 10
taken up, supporting the conclusion that Pseudo-nitzschia will potentially utilize multiple N 11
substrates, if present. 12
Other factors, however, should be considered when attempting to use such lab-derived 13
parameters in the field and include: the difference in uptake incubation periods (minutes vs. 14
hours; review by Harrison et al., 1989), N substrate interactions, particularly the inhibitory 15
effects of NH4+ on NO3
- uptake (e.g., Cochlan and Bronk, 2003), the N substrate used to 16
precondition cultures (Fan et al., 2003b), the physiological status of the community (i.e., N –17
replete vs. N- depleted cells), and its influence on transitory ‘surge’ uptake (e.g., Cochlan and 18
Harrison, 1991), and the differential effect of light on the uptake rates of the various N substrates 19
(e.g., Kudela and Cochlan, 2000). However, despite such factors that potentially limit the utility 20
of kinetics for direct (or universal) application to elucidate the nitrogenous nutrition of harmful 21
or benign phytoplankton assemblages, we do not feel that such kinetic relationships should be 22
abandoned. It has been suggested that Michaelis-Menten kinetics are not appropriate in coastal 23

Page 17 of 37
Accep
ted
Man
uscr
ipt
17
upwelling or other high NO3- waters based on linear relationships between specific NO3
- uptake 1
rates and NO3- concentrations observed from a compilation of spatially and temporally distinct 2
stations sampled off northern California and elsewhere (Dugdale et al., 2006). These authors 3
suggest that the lack of such a Michaelis-Menten type kinetic response, even at low 4
concentrations (near the expected Ks values), are due to the short time period (a few hours) in 5
which coastal phytoplankton (typically diatoms) experience low NO3- concentrations in 6
upwelling zones. However, during our experiments, P. australis NO3- kinetics were conducted 7
within minutes, not hours of NO3- depletion in the culture medium, and a classic rectangular 8
hyperbolic relationship was observed, with saturation clearly evident at the highest NO3- 9
concentration employed (35 µg-at N·L-1) - a value representative of maximal surface 10
concentrations reported for upwelling zones (e.g. review by Wilkerson and Dugdale, in press). 11
As mentioned above, kinetic parameters may vary as a function of the pre-conditioned 12
substrate used for growth, and the reader should be cautioned that all P. australis cells used in 13
the present study were pre-conditioned on NO3-. This could potentially affect the kinetic 14
parameters reported here. It may be particularly important for urea uptake, given that urease 15
activity is neither constitutively expressed in all phytoplankton, nor consistently expressed as a 16
function of N source, even within the same species. For example, Peers et al. (2000) found that 17
urease activity was present at high (and similar) constitutive levels regardless of the N sources 18
used for growth in N-replete cultures of the diatom Thalassiosira weissflogii, whereas Fan et al. 19
(2003a) found urease activities were depressed in NO3- -grown cultures compared to NH4
+- and 20
urea-grown cultures of T. weissflogii. Similar variability in urease regulation according to N 21
supply has been observed by others within a single species (e.g., the diatom Phaeodactylum 22
tricornutum: Rees and Bekheet, 1982; Lefty and Syrett, 1973), or among other marine 23

Page 18 of 37
Accep
ted
Man
uscr
ipt
18
phytoplankton species, including the cyanobacterium Synechococcus (Collier et al., 1999) and 1
some dinoflagellates (Alexandrium fundyense: Dyhrman and Anderson, 2003), but not others 2
(Prorocentrum minimum: Fan et al., 2003a). To date it is not known whether N growth source 3
has any affect on N uptake and assimilation in any of the species of Pseudo-nitzschia, including 4
P. australis. 5
The relationship between urea uptake by P. australis and the external urea concentration 6
cannot be described by Michaelis-Menten kinetics, in that our data show no evidence of 7
saturation at elevated concentrations (Figure 3). Although curvature is evident at the lower 8
concentrations (< 5 µg-at N L-1), a linear fit can be used to describe this relationship equally 9
well. Such biphasic or nonsaturating (linear) kinetics are not unusual for phytoplankton N 10
uptake (cf., review by Collos et al., 2005), and have been described for NO3- uptake by cultures 11
of centric diatoms (T. weissflogii: Collos et al., 1992; Skeletonema costatum: Serra et al., 1978), 12
natural assemblages in coastal lagoons in the Mediterranean Sea (Collos et al. 1997), and spring 13
diatom-dominated assemblages in temperate estuaries (Chesapeake Bay; Lomas and Glibert, 14
1999). Additionally, they have been mentioned (but data not presented) for NH4+ uptake within 15
CEPEX enclosures in Saanich Inlet, BC, Canada (Harrison et al., 1977), and for NH4+, amino 16
acid mixtures, and urea uptake by Choptank River assemblages (Fan et al., 2003). Recently, 17
linear uptake of urea also was demonstrated for two strains (isolated from Spain and Sardinia) of 18
the dinoflagellate A. catenella (Jauzein et al., 2006), and for a natural assemblage dominated by 19
the dinoflagellate Cochlodinium fulvescens from Monterey Bay, California (Kudela et al., 20
2007b). 21
Such linear relationships between uptake and concentration have been explained by 22
diffusion (Fan et al., 2003b) or a simultaneous combination of mediated and diffusive transfer 23

Page 19 of 37
Accep
ted
Man
uscr
ipt
19
(Serra et al., 1978), but it is not clear whether urea uptake by P. australis is controlled by urease 1
activity or urea carboxylase/allophanate hydrolase activities [adenosine triphosphate (ATP) urea 2
amidolyase (UAL-ase) pathway], or both – which is unlikely since an alga is thought to possess 3
either one enzyme system or the other, but not both (see review by Antia et al., 1991). However 4
urease activity has been confirmed in other members of the Bacillariophyceae, and all reported 5
kinetics of urease have been described by Michalis-Menten kinetics (Mobley et al., 1995). 6
In any case, despite our limited understanding of the physiological mechanism of urea 7
uptake by this diatom, and its apparent lack of preference for urea, it is clear that both organic 8
and inorganic N sources can be utilized by P. australis, and that N sources of anthropogenic 9
origin may be of particular importance during periods of relaxed upwelling and reduced NO3- 10
concentrations, possibly by sustaining non-bloom concentrations of P. australis until upwelling 11
resumes; these conditions have previously been associated with maximal DA production and 12
HAB formation in California (Kudela et al., 2004a). Although ambient concentrations of NH4+ 13
and urea are generally low in upwelling regions (but see Dugdale et al., 2006), periods of storm 14
runoff from urban and agricultural areas may result in elevated concentrations of both inorganic 15
and organic N substrates independent of upwelling (cf., Kudela et al, 2007 this volume), and 16
need to be considered when evaluating the potential effects of cultural eutrophication on the 17
growth of harmful diatoms. 18
19
5. Acknowledgements 20
21
This manuscript developed from a presentation given as part of the GEOHAB HABs and 22
Eutrophication workshop in 2005. We thank the organizers, and in particular Dr. Patricia 23

Page 20 of 37
Accep
ted
Man
uscr
ipt
20
Glibert, for the opportunity to participate. We acknowledge the expert assistance of N.C. 1
Ladizinsky (RTC) in conducting these experiments. The US ECOHAB Program grant 2
NA96OP0475 awarded to WPC and RMK supported this study. This is publication No. XX of 3
ECOHAB. 4
5
6. References 6
7
Anderson, C.R., Brzezinski, M.A., Washburn, L, Kudela, R., 2006. Circulation and 8
environmental conditions during a toxigenic Pseudo-nitzschia australis bloom in the Santa 9
Barbara Channel, California. Mar. Ecol. Prog. Ser. 327, 119-133. 10
11
Anderson, D.M., Glibert, P.M., Burkholder, J.M., 2002. Harmful algal blooms and 12
eutrophication: Nutrient sources, composition and consequences. Estuaries 25, 562-584. 13
14
15
Antia, N.J., Harrison, P.J., Oliveira, L., 1991. Phycological reviews 11. The role of dissolved 16
organic nitrogen in phytoplankton nutrition, cell biology and ecology. Phycologia, 30, 1-89. 17
18
Auro, M.E., 2007. Nitrogen dynamics and toxicity of the pennate diatom Pseudo-nitzschia 19
cuspidata: A field and laboratory study. M.S. Thesis, San Francisco State University, San 20
Francisco, California, USA, 91 p. 21
22
Bates, S.S., Bird, C.J., de Freitas, A.S.W., Foxall, R., Gilgan, M., Hanic, L.A., Johnson, G.R., 23
McCulloch, A.W., Odense, P., Pocklington, R., Quilliam, M.A., Sim, P.G., Smith, J.C., Subba 24

Page 21 of 37
Accep
ted
Man
uscr
ipt
21
Rao, D.V., Todd, E.C.D., Walter, J.A., Wright, J.L.C., 1989. Pennate diatom Nitzschia pungens 1
as the primary source of domoic acid, a toxin in shellfish from eastern Prince Edward Island, 2
Canada. Can. J. Fish. Aquat. Sci. 46, 1203-1215. 3
4
Bates, S.S., de Freitas, A.S.W., Milley, J.E., Pockington, R., Quilliam, M.A., Smith, J.C., 5
Worms, J., 1991. Controls on domoic acid production by the diatom Nitzschia pungens f. 6
multiseries in culture: nutrients and irradiance. Can. J. Fish. Aquat. Sci. 48, 1136-1144. 7
8
Bates, S.S., Worms, J., Smith, J.C., 1993. Effects of ammonium and nitrate on growth and 9
domoic acid production by Nitzschia pungens in batch culture. Can. J. Fish. Aquat. Sci. 50, 10
1248-1254. 11
12
Bates, S.S., 1998. Ecophysiology and metabolism of ASP toxin production. In: Anderson, D.M., 13
Cembella, A.D., Hallegraeff, G.M. (Eds.), The Physiological Ecology of Harmful Algal Blooms. 14
Springer-Verlag, Heidelberg, pp. 405-426. 15
16
Bates, S.S., Garrison, D.L., Horner, R.A., 1998. Bloom dynamics and physiology of domoic-17
acid-producing Pseudo-nitzschia species. In: Anderson, D.M., Cembella, A.D., Hallegraeff, 18
G.M. (Eds.), The Physiological Ecology of Harmful Algal Blooms. Springer-Verlag, Heidelberg, 19
pp. 267-292. 20
21
Bates, S.S., Trainer, V.L., 2006. The ecology of harmful diatoms. In: Granéli, E., Turner, J.T. 22
(Eds.), Ecology of Harmful Algae, Springer-Verlag, Heidelberg, pp. 81-93. 23

Page 22 of 37
Accep
ted
Man
uscr
ipt
22
1
Berges, J.A., Franklin, D.J., Harrison, P.J., 2001. Evolution of an artificial seawater medium: 2
Improvements in enriched seawater, artificial water over the last two decades. J. Phycol. 37, 3
1138-1145 [Correction addition, 40, 619, 2004]. 4
5
Berman, T., Bronk, D.A., 2003. Review. Dissolved organic nitrogen: a dynamic participant in 6
aquatic ecosystems. Aquatic Microbiol. Ecol. 31, 279-305. 7
8
Button, D.K., 1978. On the theory of control of microbial growth kinetics by limiting nutrient 9
concentrations. Deep Sea Res. 25, 1163-1177. 10
11
Cochlan, W.P., 2001. The heterotrophic bacterial response during a mesoscale iron enrichment 12
experiment (Iron Ex II) in the eastern equatorial Pacific Ocean. Limnol. Oceanogr. 46, 428-435. 13
14
Cochlan, W.P., Harrison, P.J., 1991. Uptake of nitrate ammonium and urea by nitrogen-starved 15
cultures of Micomonas-pusilla (Prasinophyeae): Transient responses. J. Phycol. 27, 673-679. 16
17
Cochlan, W.P., Bronk, D.A., 2003. Effects of ammonium on nitrate utilization in the Ross Sea: 18
Implications for ƒ-ratio estimates. In: G. DiTullio, G.R, Dunbar, R.B. [Eds.], Biogeochemistry of 19
the Ross Sea. AGU Antarctic Research Series 78, 159-178. 20
21

Page 23 of 37
Accep
ted
Man
uscr
ipt
23
Cochlan, W.P., J. Herndon, J., Ladizinsky, N.C., Kudela, R.M., 2006a. Nitrogen uptake by the 1
toxigenic diatom Pseudo-nitzschia australi. GEOHAB Open Science Meeting on HABs and 2
Eutrophication, Baltimore, MD, March 2005. EOS Trans. Am. Geophys. Union, 87, (36) suppl. 3
4
Cochlan, W.P., Wells, M.L., Trainer, V.L., Trick, C.G., Lessard, E.J., Hickey, B., 2006b. 5
Domoic acid production is not linked to silicate limitation in natural populations of Pseudo-6
nitzschia. Twelfth International Conference on Harmful Algae, Copenhagen, DK, September 7
2006 (Abstract). 8
9
Collier, J.L., Brahamsha, Palenik, B., 1999. The marine cyanobacterium Synechococcus sp. 10
WH7805 requires urease (urea amidohydrolase, EC 3.51.5) to utilize urea as a nitrogen source: 11
molecular-genetic and biochemical analysis of the enzyme. Microbiol. 145, 447-459. 12
13
Collos,Y. Siddiqi, M.Y., Wang, M.Y., Glass, A.D.M., Harrison, P.J., 1992. Nitrate uptake 14
kinetics by two marine diatoms using the radioactive tracer 13N. J. Exp. Mar. Biol. Ecol. 163, 15
251-260. 16
17
Collos, Y., Vaquer, A, Bibent, B., Slawyk, G., Garcia, N., Souchu, P., 1997. Variability in nitrate 18
uptake kinetics of phytoplankton communities in a Mediterranean coastal lagoon. Est. Coastal Shelf 19
Sci. 44, 369-375. 20
21
Collos, Y., Vaquer, A., Souchu, P., 2005. Review: Acclimation of nitrate uptake by 22
phytoplankton to high substrate levels. J. Phycol. 41, 466-478. 23

Page 24 of 37
Accep
ted
Man
uscr
ipt
24
1
Conway, H.L., Harrison, P.J., Davis, C.O., 1976. Marine diatoms grown in chemostats under 2
silicate or ammonium limitation. II. Transient response of Skeletonema costatum to a single 3
addition of a limiting nutrient. Mar. Biol. 35, 187-199. 4
5
Degobbis, D., 1973. On the storage of sea water samples for ammonia determination. Limnol. 6
Oceanogr. 18, 146-150. 7
8
Dortch, Q., Robichaux, R., Pool, S., Milsted, D., Mire, G., Rabalais, N.N., Soniat, T.M., Fryxell, 9
G.A., Turner, R.E., Parsons, M.L., 1997. Abundance and vertical flux of Pseudo-nitzschia in the 10
northern Gulf of Mexico. Mar. Ecol. Prog. Ser., 146, 249-264. 11
12
Dugdale, R.C., Wilkerson, F.P., 1986. The use of 15N to measure nitrogen uptake in the eutrophic 13
oceans; experimental considerations. Limnol. Oceanogr. 31, 673-689. 14
15
Dugdale, R.C., Wilkerson, F.P., Hogue, V.E., Marchi, A., 2006. Nutrient controls on new 16
production in the Bodega Bay, California, coastal upwelling plume. Deep-Sea Res II, 53, 3049-17
3062. 18
19
Dyhrman, S.T., Anderson, D.M., 2003. Urease activity in cultures and filed populations of the 20
toxic dinoflagellate Alexandrium. Limnol. Oceanogr., 48, 647-655. 21
22

Page 25 of 37
Accep
ted
Man
uscr
ipt
25
Eppley. R.W., Sharp, J.H., Renger, E.H., Perry, M.J., Harrison, W.G., 1977. Nitrogen 1
assimilation by phytoplankton and other organisms in the surface waters of the central North 2
Pacific Ocean. Mar. Biol. 39, 111-120. 3
4
Fan, C., Glibert, P.M.,Alexander, J., Lomas, M.W., 2003a. Characterization of urease activity in 5
three marine phytoplankton species, Aureococcus anophagefferens, Prorocentrum minimum, and 6
Thalassiosira weissflogii. Mar. Biol., 142, 949-958. 7
8
Fan, C., Glibert, P.M., Burkholder, J.M., 2003b. Characterization of the affinity for nitrogen, 9
uptake kinetics, and environmental relationships for Prorocentrum minimum in natural blooms 10
and laboaratory cultures. Harmful Algae, 2, 283-299. 11
12
Fryxell, G.A., Villac, M.C., Shapiro, L.P., 1997. The occurrence of the toxic diatom genus 13
Pseudonitzschia (Bacillariophyceae) on the West Coast of the USA, 1920-1996: A review. 14
Phycologia 36, 419-437. 15
16
Glibert, P.M., Lipschultz, F., McCarthy, J.J., Altabet, M.A., 1982. Isotope dilution models of 17
uptake and remineralization of ammonium by marine plankton. Limnol. Oceanogr. 27, 639-650. 18
19
Glibert, P.M., Anderson, D.M., Gentien, P., Granéli, E., Sellner, K.G., 2005. The global, 20
complex phenomena of harmful algal blooms. Oceanography 18, 136-147. 21
22

Page 26 of 37
Accep
ted
Man
uscr
ipt
26
Hallegraeff, G.M., 1993. A review of harmful algal blooms and their apparent global increase. 1
Phycologia 32, 79-99. 2
3
Harrison, P.J., Parslow, J.S., Conway, H.L., 1989. Determination of nutrient uptake kinetic 4
parameters: a comparison of methods. Mar. Ecol. Progr. Ser. 52, 301-312. 5
6
Harrison, W.G., Eppley, W.G., Renger, E.H., 1977. Phytoplankton nitrogen metabolism, 7
nitrogen budgets, and observations on copper toxicity: controlled ecosystem Pollution 8
experiment. Bull. Mar. Sci. 27, 44-57. 9
10
Hasle, G.R., 1995. Pseudo-nitzschia pungens and P. multiseries (Bacillariophyceae): 11
nomenclatural history, morphology and distribution. J. Phycol. 31, 428-435. 12
13
Hillebrand, H., Sommer, U., 1996. Nitrogenous nutrition of the potentially toxic diatom 14
Pseudonitzschia pungens f. multiseries Hasle. J. Plank. Res. 18, 295-301. 15
16
Healey, F.P., 1980. Slope of the Monod equation as an indicator of advantage in nutrient 17
competition. Microbial Ecol. 5 281-286. 18
19
Horner, R.A., Hickey, B.M., Postel, J.R., 2000. Pseudo-nitzschia blooms and physical 20
oceanography off Washington state. S. Afr. J. Mar. Sci. 22, 299-308. 21
22
Howard, M.D.A., Cochlan, W.P., Ladizinsky, N. and Kudela, R.M., 2007. Nitrogenous 23

Page 27 of 37
Accep
ted
Man
uscr
ipt
27
preference of toxigenic Pseudo-nitzschia australis (Bacillariophyceae) from field and laboratory 1
experiments. Harmful Algae 6 206-217. 2
3
Jauzein, C., Collos, Y., Garcés, E., Vila, M, Maso, M., 2006. Short-term temporal variability of 4
ammonium and urea uptake by Alexandrium catenella in cultures. Twelfth International 5
Conference on Harmful Algae, Copenhagen, DK, September 2006 (Abstract). 6
7
Knepel, K., Bogren, K., 2001. Determination of orthophosphorous by flow injection analysis in 8
seawaters: QuickChem Method 31-115-01-1-H. In: Saline Methods of Analysis. Lachat 9
Instruments, Milwaukee, WI, 14 pp. 10
11
Kudela, R.M., Cochlan, W.P., 2000. Nitrogen and carbon uptake kinetics and the influence of 12
irradiance for a red tide bloom off southern California. Aquatic Microbial Ecol. 21, 31-47. 13
14
Kudela, R., Cochlan, W., Roberts, A., 2004a. Spatial and temporal patterns of Pseudo-nitzschia 15
spp. in central California related to regional oceanography. In: Steidinger, K.A., Landsberg, J.H., 16
Tomas, C.R., Vargo, G.A. (Eds.) Harmful Algae 2002. Florida and Wildlife Conservation 17
Commission, Florida Institute of Oceanography, and Intergovernmental Oceanographic 18
Commission of UNESCO, pp. 347-349. 19
20
Kudela, R., Roberts, A., and Armstrong, M., 2004b. Laboratory analyses of nutrient stress and 21
toxin production in Pseudo-nitzschia spp. from Monterey Bay, California. In: Steidinger, K.A., 22
Landsberg, J.H., Tomas, C.R., Vargo, G.A. (Eds.) Harmful Algae 2002. Florida and Wildlife 23

Page 28 of 37
Accep
ted
Man
uscr
ipt
28
Conservation Commission, Florida Institute of Oceanography, and Intergovernmental 1
Oceanographic Commission of UNESCO, pp. 136-138. 2
3 Kudela, R., Pitcher, G., Probyn, T., Figueiras, F., Moita, T. and Trainer, V., 2005. Harmful Algal 4
Blooms in coastal upwelling systems. Oceanography 18 184-197. 5
6
Kudela, R.M., Lane, J.Q., Cochlan, W.P. 2007a. The potential role of anthropogenically derived 7
nitrogen in the growth of harmful algae in California, USA. Harmful Algae (In press). 8
9
Kudela, R.M., Ryan, J., Blakely, M., and Lane, J. 2007b. Linking the physiology and ecology of 10
Cochlodinium to better understand harmful algal bloom events: a comparative approach. 11
Harmful Algae (In press). 12
13
Ladizinsky, N.C., 2003. The influence of dissolved copper on the production of domoic acid by 14
Pseudo-nitzschia species in Monterey Bay, California: laboratory experiments and field 15
observations. MSc Thesis, California State University, Monterey Bay, CA, USA, 68 p. 16
17
Lee, S., Fuhrman, J.A., 1987. Relationships between biovolume and biomass of naturally derived 18
marine bacterioplankton. Appl. Environ. Microbiol. 53, 1298-1303. 19
20
Lomas, M.W., Glibert, P.A., 1999. Temperature regulation of nitrate uptake: A novel hypothesis 21
about nitrate uptake and reduction in cool-water diatoms. Limnol. Oceanogr. 44, 556-572. 22
23

Page 29 of 37
Accep
ted
Man
uscr
ipt
29
Lundholm, N., Hansen, P.J., Kotaki, Y., 2004. Effect of pH on growth and domoic acid 1
production by potentially toxic diatoms of the genera Pseudo-nitzschia and Nitzschia. Mar. Ecol. 2
Prog. Ser. 273, 1-15. 3
4
Marchetti, A., Trainer, V.L., Harrison, P.J., 2004. Environmental conditions and phytoplankton 5
dynamics associated with Pseudo-nitzschia abundance and domoic acid in the Juan de Fuca 6
eddy. Mar. Ecol. Prog. Ser. 281, 1-12. 7
8
McCarthy, J.J., Taylor, W.R., Taft, J.L., 1977. Nitrogenous nutrition of the plankton in the 9
Chesapeake Bay. I. Nutrient availability and phytoplankton preferences. Limnol. Oceanogr. 22, 10
996-1011. 11
12
Mengelt, C., 2005. A mechanism coupling sewage effluent to the dark survival of the toxic 13
diatom Pseudo-nitzschia sp. and subsequent initiation of toxic blooms by coastal upwelling. UC 14
Marine Council Coastal Water Quality Initiative, eScholarship repository, Univ. California, No. 15
012. 16
17
Mobley, H.L.T., Island, M.D., Hausinger, R.P., 1995. Molecular biology of microbial ureases. 18
Microbiol. Reviews 59, 451-480. 19
20
Parsons, M.L., Dortch, Q., Turner, R.E., 2002. Sedimentological evidence of an increase in 21
Pseudo-nitzschia (Bacillariophyceae) abundance in response to coastal eutrophication. Limnol. 22
Oceanogr. 47, 551-558. 23
24

Page 30 of 37
Accep
ted
Man
uscr
ipt
30
Peers, G.S., Milligan, A.J., Harrison, P.J., 2000. Assay optimization and regulation of urease 1
activity in two marine diatoms. J. Phycol. 36, 523-528. 2
3
Press, W.H., Teukolsky, S.A., Vetterling, W.T., Flannery, B.P., 1992. Numerical Recipes in C. 4
Cambridge University Press, New York, pp. 994. 5
6
Price, N.M., Harrison, P.J., 1987. A comparison of methods for the measurement of dissolved 7
urea concentration in seawater. Mar. Biol., 92, 307-319. 8
9
Price, N.M., Harrison, P.J. 1988. Uptake of urea C and urea N by the coastal marine diatom 10
Thalassiosira pseudonana. Limnol. Oceanogr. 33, 528-237. 11
12
Rees, T.A.V., Bekheet, I.A., 1982. The role of nickel in urea assimilation by algae. Planta 156, 13
385-387. 14
15
Ryther, J.H., Dunstan, W.M., 1971. Nitrogen, phosphorus and eutrophication in the coastal 16
marine environment. Science 171, 1008-1013. 17
18
Schnetzer, A., Miller, P.E., Schaffner, R.A., Stauffer, B.A., Jones, B.H., Weisberg, S.B., 19
DiGiacomo, P.M., Berelson, W.M., Caron, D.A., 2007. Blooms of Pseudo-nitzschia and domoic 20
acid in the San Pedro Channel and Los Angeles harbor areas of the Southern California Bight. 21
Harmful Algae 6, 372-387. 22
23

Page 31 of 37
Accep
ted
Man
uscr
ipt
31
Scholin, C.A., Gulland, F., Doucette, G.J., Benson, S., Busman, M., Chavez, F.P., Cordaro, J., 1
DeLong, R., DeVogelaere, A., Harvey, J., Haulena, M., Lefebvre, K., Lipsomb, T., Loscutoff, S., 2
Lowenstine, L.J., Marin III, R., Miller, P.E., McLellan, W.A., Moeller, P.D.R., Powell, C.L., 3
Rowles, T., Silvagni, P., Silver, M., Spraker, T., Trainer, V., Van Dolah, F.M., 2000. Mortality 4
of sea lions along the central California coast linked to a toxic diatom bloom. Nature 403, 80-84. 5
6
Serra, J.L., Llama, M., Cadenas, E., 1978. Nitrate utilization by the diatom Skeletonema 7
costatum. I. Kinetics of nitrate uptake. Plant Physiol. 62, 987-990. 8
9
Smith, P., Bogren, K., 2001a. Determination of nitrate and/or nitrite in brackish or seawater by 10
flow injection analysis colorimeter: QuickChem Method 31-107-04-1-E. In: Saline Methods of 11
Analysis. Lachat Instruments, Milwaukee, WI, 12 pp. 12
13
Smith, P., Bogren, K., 2001b. Determination of silicate in brackish or seawater by flow injection 14
analysis colorimeter: QuickChem Method 31-114-27-1-C. In: Saline Methods of Analysis. 15
Lachat Instruments, Milwaukee, WI, 12 pp. 16
17
Subba Rao, D.V., Quilliam, M.A.Pockington, R., 1988. Domoic acid – a neurotoxic amino aid 18
produced by the marine diatom Nitzschia pungens in culture. Can J. Fish. Aquatic Sci. 45, 2076-19
2079. 20
21
Solorzano, L., 1969. Determination of ammonium in natural waters by the phenolhypochlorite 22
method. Limnol. Oceanogr. 14,799-801. 23

Page 32 of 37
Accep
ted
Man
uscr
ipt
32
1
Trainer, V.L., Adams, N.G., Bill, B.D., Stehr, C.M., Wekell, J.C., Moeller, P., Busman, M., 2
Woodruff, D., 2000. Domoic acid production near California coastal upwelling zones, June 3
1998. Limnol. Oceanogr. 45, 1818-1833. 4
5
Trainer, V.L., Hickey, B.M., Horner, R.A., 2002. Biological and physical dynamics of domoic 6
acid production off the Washington coast. Limnol. Oceanogr. 47, 1438-1446. 7
8
Trainer, V.L., Cochlan, W.P., Erickson, A. Bill, B.D., Cox, F.H., Borchert J.A., Lefebvre, K.A., 9
2007. Recent domoic acid closures of shellfish harvest areas in Washington State inland 10
waterways. Harmful Algae 6, 449-459. 11
12
Uchida, T., 1976. Excretion of ammonium by Prorocentrum micans Ehrenburg in urea-grown 13
culture. Japan J. Ecol. 26, 43-44. 14
15
Wilkerson, F., Dugdale, R.C. Coastal Upwelling. In: Nitrogen in the Marine Environment, 16
Capone, D., Carpenter, E., Mulholland, M. Bronk, D. [Eds.], (In press). 17
18

Page 33 of 37
Accep
ted
Man
uscr
ipt
33
7. Figure captions 1
2
Figure 1. The specific growth rate (µ; d-1) as a function of photosynthetic photon flux density 3
(PPFD) for Pseudo-nitzschia australis (UCSC strain 0771B isolated by P. Hughes from Point 4
Reyes, CA) grown in ESAW-enriched, sterile-filtered seawater. Mean of duplicate (n =2) 5
cultures are plotted; error bars indicate range of duplicates. (courtesy of B.D. Bill, unpublished 6
data). 7
8
Figure 2. Particulate nitrogen (PN) specific uptake rates (h-1) of NO3- (A), NH4
+ (B), and 9
glutamine (C) for recently nitrate-replete cultures of Pseudo-nitzschia australis plotted versus 10
substrate concentration calculated at the beginning of each 20.5-min incubation period. Rate 11
estimates are fitted to the Michaelis-Menten equation (see text for details). For the glutamine 12
experiment, uptake was also determined at 70 µg-at N L-1, and is included in the curve fit shown. 13
Note the change in scale of ordinal axes. 14
15
Figure 3. Specific uptake rates (h-1) of urea for duplicate cultures of Pseudo-nitzschia australis 16
plotted versus urea concentration collected at the beginning of the incubation period. The lines 17
denote least-squares, linear regressions for culture one ( ) and culture two ( ) and their 18
corresponding coefficients of determination (r2). Cell abundance was 5.59 and 6.50 x 106 cells 19
L-1 for culture 1 and 2, respectively. Insert plot is specific urea uptake versus concentration at 20
low (< 5 µg-at N L-1) concentrations. 21
Page 34 of 37
Accep
ted
Man
uscr
ipt
Cochlan et al., Figure 1 1
0.50
1.00
1.50
2.00
0 50 100 150 200 250 300 350
Gro
wth
Rat
e (d
-1)
PPFD (µE m-2
s-1
)
0
Page 35 of 37
Accep
ted
Man
uscr
ipt
35
Cochlan et al., Figure 2
40.0
80.0
120.0
r2 = 0.98
0
A: Nitrate
40.0
80.0
120.0
r2 = 0.96
B: Ammonium
0
20.0
40.0
60.0
80.0
0 10 20 30 40
r2 = 0.92
0
C: Glutamine
N Concentration (µg-at N L-1
)
PN
Spe
cific
Upt
ake
Rat
e (
x 1
0-3 h
-1 )
Page 36 of 37
Accep
ted
Man
uscr
ipt
36
Cochlan et al., Figure 3
2.0
4.0
6.0
8.0
10.0
1.0 2.0 3.0 4.0 5.00
0
10.0
20.0
30.0
40.0
50.0
0 10 20 30 40
r2 = 0.99
r2 = 0.98
0
Urea Concentration (µg-at N L-1
)
Spe
cific
Ure
a U
ptak
e R
ate
( x 1
0-3 h
-1)
Page 37 of 37
Accep
ted
Man
uscr
ipt
37
Table 1. Kinetic parameters for N uptake by Pseudo-nitzschia australis previously maintained on
nitrate as the sole N source. Michaelis-Menten parameters: maximum uptake velocity (Vmax) and
half-saturation constant (Ks) were estimated from a direct nonlinear curve fitting model (see text
for details), and substrate affinity (α) calculated as Vmax/Ks. Estimated standard error (SE)
values of kinetic parameters are given in parentheses. The r2 column provides the coefficient of
determination and the sample size (n).
1
N Substrate
Cell Abundance (x106 cells L-1)
Vmax (x 10-3 h-1)
Ks
(µg-at N L-1)
α (x 10-3 h-1) • (µg-at N L-1)-1
r2
(n)
Nitrate
6.30
105.3 (3.93)
2.82 (0.32)
37.3
0.98 (19)
Ammonium 6.50 70.96 (4.24)
5.37 (0.81)
13.2 0.96 (20)
Glutamine
6.23
66.16 (4.06)
5.94 (1.20)
11.1 0.92 (22)
2 3
4




















