
Elucidating coral reef predator trophodynamics across an ...
Upload
independentCategory
view
4download
0
Estuarine, Coastal and Shelf Science 66 (2006) 634e642www.elsevier.com/locate/ecss
Living macromolluscs from a paleo-reef region on thenortheastern Venezuelan continental shelf
Joaquın Buitrago a,*, Juan Capelo a, Javier Gutierrez a, Martın Rada a,Ricardo Hernandez a, Sylvia Grune b
a Estacion de Investigaciones Marinas de Margarita, Fundacion La Salle de Ciencias Naturales, Punta de Piedras, Isla de Margarita,
Apartado 144, Porlamar 6301-A, Venezuelab Escuela de Ciencias Aplicadas al Mar, Universidad de Oriente, Nucleo Nueva Esparta, Venezuela
Received 9 February 2005; accepted 10 November 2005
Available online 27 December 2005
Abstract
Drowned reefs, fossil reefs or paleo-reefs, are important ecologically as areas of high biodiversity, foraging, shelter environment, and as areassupporting the spawning aggregations of economically important reef fish species. This is particularly significant when the structures are situatedin a wide soft-bottom continental shelf. The presence of limestone structures, fossil reefs and pinnacles dating from circa 8 to 9 ka, to the northof the Paria Peninsula in north-eastern Venezuela, has been known to local fishermen for decades. Using echograms obtained during acousticfisheries evaluations and the scarce previously available information, an improved location map of hard-bottom structures was made. Benthicsamples to study macromolluscs were taken at depths between 54 and 93 m using an unmodified 2-m beam trawl. Four trawl samples werelocated over fossil reef areas while another four were situated in soft-bottom valleys between limestone structures. Fossil reefs in the areashowed a highly patchy distribution. A total of 91 species from 43 Bivalvia, Gastropoda and Scaphopoda families were found, Gastropoda beingthe dominant class with 49 species. Paleo-reef-covered areas showed higher species richness and only 21% of the species found were common toboth substrates. Gastropods Tonna maculosa and Polystira albida were the most abundant species and occurred in both substrate types. Bivalvelife habits, a mixture of organismesubstrate relationships, shell fixation, mobility and feeding type, differed significantly according to bottomtype. Six species are recorded for the first time for eastern Venezuelan waters. Bottom heterogeneity plays an important role in marine ecosys-tems, providing shelter to fish populations and may be significant as breeding and nursery areas. Its presence in a region with biogeographicalinterest, situated in the confluence of three major provinces and with oceanographic conditions varying seasonally from upwelling dominated toOrinoco River discharges, makes this the area of interest and it should be evaluated as a possible Marine Protected Area.� 2005 Elsevier Ltd. All rights reserved.
Keywords: molluscs; fossil paleo-reefs; Venezuelan continental shelf; Caribbean
1. Introduction
Records of global sea level change provide important infor-mation on the dynamics of glaciers and ice sheets (Clark et al.,2002). Studies in different parts of the world, from coastal de-positional systems (Blum et al., 2002) to the base of sub-merged sea-cliffs (Blanchon et al., 2002), have provided
* Corresponding author.
E-mail address: [email protected] (J. Buitrago).
0272-7714/$ - see front matter � 2005 Elsevier Ltd. All rights reserved.
doi:10.1016/j.ecss.2005.11.006
crucial understanding of paleoclimatic events. Reef corals,by virtue of their close correlation to sea level, have been fav-oured study subjects in this aspect (Bloom, 1994). Sea levelindicators at locations around the world and glacio-hydro-isostatic models show that the pattern of relative sea levelrise during the Middle and Late Holocene was not uniformin time and space, but rather depended on the proximity tothe melting ice sheets and local tectonics (Shaked et al., 2002).
The northeastern Venezuelan shelf extends north of PariaPeninsula, and eastward from Margarita Island and Los Testi-gos archipelago to Trinidad and Tobago. The presence of

635J. Buitrago et al. / Estuarine, Coastal and Shelf Science 66 (2006) 634e642
limestone structures like fossil reefs and pinnacles, was firstdescribed during oil exploration and from recovered samples,C14 dated 8.1 to 9.9 ka (Milliman, 1980; Milliman et al., 1980,1982; Butenko et al., 1981), dates consistent with similar for-mations around the Caribbean (Bloom, 1994; Toscano andLundberg, 1998). Fossil coral species composition of Pariareefs has not been studied to date. Demise of these reefswas probably caused by some combination of drowning and/or smothering by mud as rising sea level opened connectionsnear Trinidad that permitted Holocene sediment transportfrom the Orinoco/Amazon systems to begin (Milliman et al.,1982). This view is supported by sedimentation studies inthe Cariaco Basin to the west (Peterson et al., 2000).
Drowned reefs, fossil reefs or paleo-reefs are importantecologically as areas of high biodiversity, foraging and shelterenvironment, and as source areas supporting the spawning ag-gregations of economically important fish species (Turgeonet al., 2002). This is particularly significant when the struc-tures are situated in a wide soft-bottom continental shelf likethe northern Paria area or the pinnacles region in northernGulf of Mexico (Ludwig and Walton, 1957). In these cases,not only the hard-bottom areas, but also nearby muddy orsandy areas, show increased benthic faunal richness and diver-sity, becoming important fish-feeding areas, like the ‘‘Bancode Las Animas’’ in the Colombian Gulf of Salamanca(Guzman-Alvis and Dıaz, 1993). Some of these regions, likethe Florida Middle Grounds, include areas that have been de-clared as Marine Protected Areas (Turgeon et al., 2002).
Fossil reef boulders, notches in fossil reef rock (Nagel-kerken et al., 2000) and other structures in substrate, as wellas the interactions between organisms and minerals (Baves-trello et al., 2000), are important determinants of the biotapresent in an area. Suitable hard substrate available for repro-duction (Swanson, 2004) and larval fixation affects not onlyinitial colonization but also later assemblages. Structurallycomplex habitats exhibit higher diversity, abundance and,a more complex trophic organization, in terms of more func-tional groups and guilds of fish assemblages, than less struc-tured environments (Angel and Ojeda, 2001). As in similarregions of continental margins, extensive systems of scarpedhard-bottom representing areas of natural relief, provide im-portant marine habitat (Obrochta et al., 2003).
The presence of limestone structures, fossil reefs and pin-nacles north of the Paria Peninsula in north-eastern Venezuela,has been known to local fishermen for decades. As a result ofacoustic fisheries studies, maps of bottom conditions reportedthem as ‘‘rough bottom’’ (Institute of Marine Research Bergen,1989). Grouper and snapper fishermen based in nearbyMargaritaIsland (Gonzalez, 1991) have traditionally used the region asfishing grounds. On the other hand, the presence of these struc-tures limits trawl fisheries distribution (Mendoza et al., 1994).However, their location, and the ecosystems they conform havenever been studied, and in Caribbean oceanographic literature,their existence is limited to a dashed area in a wide continentalshelf map (Butenko et al., 1981). Available navigation chartsfor the region are based on half a century old soundings and donot provide any reference to the existence of these structures.
Although the relationship between fish assemblages andtheir environment has been relatively well described over thelast two decades (Letourneur et al., 2003) and high regionalproductivity in north-eastern Venezuela is reflected in a largefishing industry (Muller-Karger et al., 1989), very little isknown about the north of Paria fossil reefs biota. In the past,the area was surveyed occasionally by research vessels cruises(Bayer, 1971), and some taxa, like molluscs, have been studiedbased on littoral and shallow water samples (Tello, 1975;Princz, 1982; Alvarez et al., 1992; Capelo and Buitrago,1998). Meanwhile, gas exploration in the region continuesand drilling, production platforms and pipeline settings are ex-pected to increase in intensity, representing a potential threatto an almost completely unknown, but probably regionally im-portant environment. This work presents the results of explor-atory mollusc dredging in the area and its relation to fossil reefdistribution inferred from echo-sounder records.
2. Methods
Location of limestone structures, fossil reefs and pinnacles,on the northern Paria continental shelf (Fig. 1) was knownonly from anecdotal references, their description as ‘‘roughbottom’’ (Institute of Marine Research Bergen, 1989) andfrom the Butenko et al. (1981) paper. During the last 25 years,Margarita’s Marine Research Station (EDIMAR) has carriedout frequent acoustic cruises for fish stock evaluation. Before1995, EDIMAR’s fisheries acoustic cruises were onboard R/V‘‘La Salle’’ using an SIMRAD� EKS, 120 kHz scientificecho-sounder (Gerlotto and Gines, 1988). Later the R/V‘‘Hermano Gines’’ used an SIMRAD� EK 500, 38 and120 kHz scientific echo-sounder (Cardenas and Achury,2000). Echograms obtained during these cruises were usedto locate hard-bottom structures. Most cruises were limitedto the Margarita Island region, but some of them extendedeast to the northern Paria shelf. Cruises ‘‘ECOVEN 3’’ (Sep-tember 1985), ‘‘ECOVEN 4’’ (April 1986), ‘‘FEPELOR’’(September 1988) and ‘‘EISAR’’ (March 1999) crossed, tosome extent, the Paria study area. Raw data from these cruisesin the form of echograms with graphic scale, route and dis-tance, were examined for bottom structures indicating fossilreefs, pinnacles and other hard-bottom structures present(Fig. 2) (Butenko et al., 1981). Echograms representing1536 km of survey along four basic routes were evaluated. Re-cords showing a structured hard-bottom were geo-referencedfrom the cruise echograms and logbook data, showing initialand final survey transect position, route, bearing and distance.Identified structured hard-bottom areas were mapped usingMAPINFO 6.0� software. Line maps of both structuredhard-bottom and soft-bottom were made for each cruise, giv-ing value one to sectors where structures were detected andvalue zero to soft smooth bottom areas. Route line mapsfrom cruises were converted to regions generating 1-km-widebuffer areas each side route and discretely joining regionswith same value to produce a general bottom type distributionmap for surveyed areas. Zones where (Institute of MarineResearch Bergen, 1989) reported ‘‘rough bottom’’ and
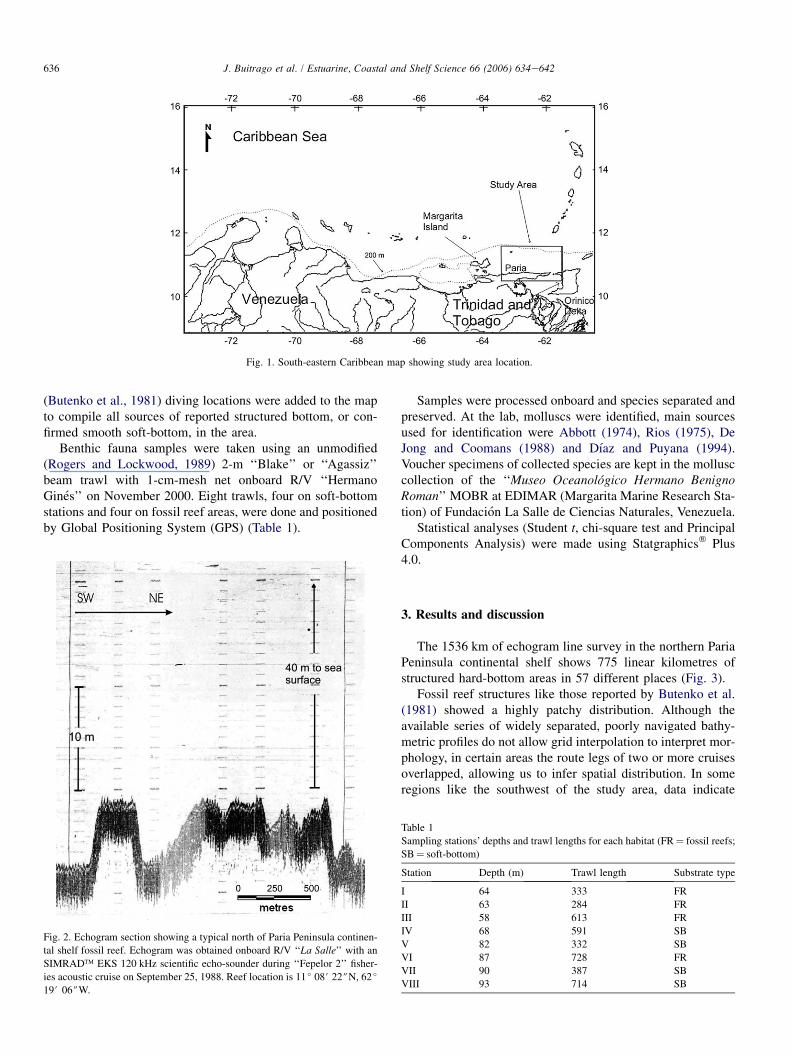
636 J. Buitrago et al. / Estuarine, Coastal and Shelf Science 66 (2006) 634e642
Fig. 1. South-eastern Caribbean map showing study area location.
(Butenko et al., 1981) diving locations were added to the mapto compile all sources of reported structured bottom, or con-firmed smooth soft-bottom, in the area.
Benthic fauna samples were taken using an unmodified(Rogers and Lockwood, 1989) 2-m ‘‘Blake’’ or ‘‘Agassiz’’beam trawl with 1-cm-mesh net onboard R/V ‘‘HermanoGines’’ on November 2000. Eight trawls, four on soft-bottomstations and four on fossil reef areas, were done and positionedby Global Positioning System (GPS) (Table 1).
Fig. 2. Echogram section showing a typical north of Paria Peninsula continen-
tal shelf fossil reef. Echogram was obtained onboard R/V ‘‘La Salle’’ with an
SIMRAD� EKS 120 kHz scientific echo-sounder during ‘‘Fepelor 2’’ fisher-
ies acoustic cruise on September 25, 1988. Reef location is 11 � 08# 22$N, 62 �
19# 06$W.
Samples were processed onboard and species separated andpreserved. At the lab, molluscs were identified, main sourcesused for identification were Abbott (1974), Rios (1975), DeJong and Coomans (1988) and Dıaz and Puyana (1994).Voucher specimens of collected species are kept in the mollusccollection of the ‘‘Museo Oceanologico Hermano BenignoRoman’’ MOBR at EDIMAR (Margarita Marine Research Sta-tion) of Fundacion La Salle de Ciencias Naturales, Venezuela.
Statistical analyses (Student t, chi-square test and PrincipalComponents Analysis) were made using Statgraphics� Plus4.0.
3. Results and discussion
The 1536 km of echogram line survey in the northern PariaPeninsula continental shelf shows 775 linear kilometres ofstructured hard-bottom areas in 57 different places (Fig. 3).
Fossil reef structures like those reported by Butenko et al.(1981) showed a highly patchy distribution. Although theavailable series of widely separated, poorly navigated bathy-metric profiles do not allow grid interpolation to interpret mor-phology, in certain areas the route legs of two or more cruisesoverlapped, allowing us to infer spatial distribution. In someregions like the southwest of the study area, data indicate
Table 1
Sampling stations’ depths and trawl lengths for each habitat (FR¼ fossil reefs;
SB¼ soft-bottom)
Station Depth (m) Trawl length Substrate type
I 64 333 FR
II 63 284 FR
III 58 613 FR
IV 68 591 SB
V 82 332 SB
VI 87 728 FR
VII 90 387 SB
VIII 93 714 SB

637J. Buitrago et al. / Estuarine, Coastal and Shelf Science 66 (2006) 634e642
Fig. 3. Limestone structures, fossil reefs and pinnacles distribution in northern Paria continental shelf. Structures registered in echograms from EDIMAR’s fish
stock assessment acoustic cruises are shown in dark grey and areas where soft-bottom was encountered are in light grey. Vertical dashed areas show Butenko
et al. (1981) fossil reefs diving locations and horizontal lines indicate rough bottom areas as reported by Institute of Marine Research Bergen (1989).
extreme habitat patchiness. Cruises routes crossing each othershowed alternatively rough and soft smooth bottom profiles.
The area of reef cover inferred from echogram studiesamounts to 2260 km2. Previously reported rough bottom(Institute of Marine Research Bergen, 1989) and fossil reefs(Butenko et al., 1981) cover an estimated 2700 km2, whilesoft-bottom areas account for 1780 km2 (Fig. 3).
In the northern legs of cruise routes, pinnacle-like bottomstructures were observed on the continental shelf border dur-ing ECOVEN 3 cruise (Fig. 4). These features are similar inmorphology to those reported by Butenko et al. (1981) andto those found in other regions like the northern Gulf of
Mexico (USGS, 2002), but their dimensions seem to be larger,reaching over 100 m in height and in 200 m deep water.
During the November 2000 cruise eight beam trawls sam-ples were taken. Four samples were located over fossil reefareas (mean length 490 m; mean depth 68 m), while otherfour were situated in the soft-bottom valleys between lime-stone structures (mean length 506 m; mean depth 83 m). AStudent’s t test showed no significant difference between fossilreef and soft-bottom trawls depths (t ¼�1.78, p ¼ 0.12) andlengths (t ¼�0.12, p ¼ 0.91).
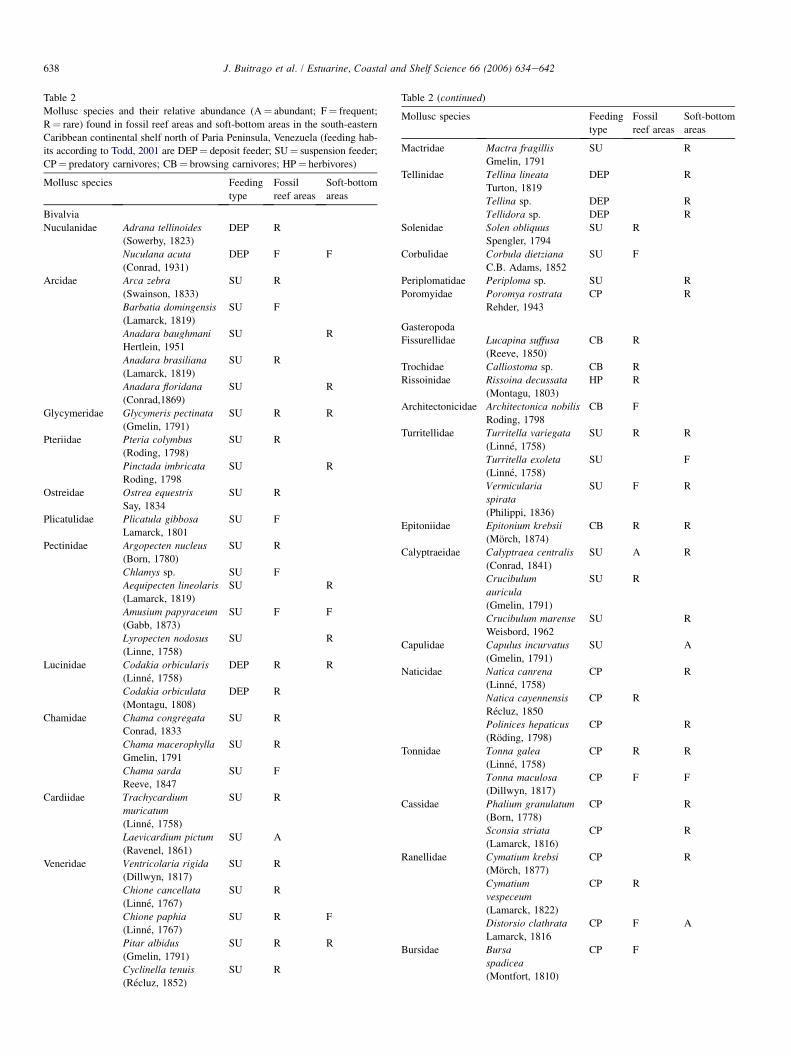
A total of 91 species from 43 families of Bivalvia, Gastro-poda and Scaphopoda were found (Table 2). Gastropoda was
Fig. 4. Echogram section showing pinnacles found in northern Paria continental shelf borders. Echogram was obtained onboard R/V ‘‘La Salle’’ with an SIM-
RAD� EKS 120 kHz scientific echo-sounder during ‘‘ECOVEN 3’’ fisheries acoustic cruise on September 25, 1985. Pinnacle location is 11 � 29# 15$N, 62 �
49# 10$W.
638 J. Buitrago et al. / Estuarine, Coastal and Shelf Science 66 (2006) 634e642
Table 2
Mollusc species and their relative abundance (A¼ abundant; F¼ frequent;
R¼ rare) found in fossil reef areas and soft-bottom areas in the south-eastern
Caribbean continental shelf north of Paria Peninsula, Venezuela (feeding hab-
its according to Todd, 2001 are DEP¼ deposit feeder; SU¼ suspension feeder;
CP¼ predatory carnivores; CB¼ browsing carnivores; HP¼ herbivores)
Mollusc species Feeding
type
Fossil
reef areas
Soft-bottom
areas
Bivalvia
Nuculanidae Adrana tellinoides(Sowerby, 1823)
DEP R
Nuculana acuta
(Conrad, 1931)
DEP F F
Arcidae Arca zebra
(Swainson, 1833)
SU R
Barbatia domingensis
(Lamarck, 1819)
SU F
Anadara baughmani
Hertlein, 1951
SU R
Anadara brasiliana
(Lamarck, 1819)
SU R
Anadara floridana
(Conrad,1869)
SU R
Glycymeridae Glycymeris pectinata
(Gmelin, 1791)
SU R R
Pteriidae Pteria colymbus
(Roding, 1798)
SU R
Pinctada imbricataRoding, 1798
SU R
Ostreidae Ostrea equestris
Say, 1834
SU R
Plicatulidae Plicatula gibbosaLamarck, 1801
SU F
Pectinidae Argopecten nucleus
(Born, 1780)
SU R
Chlamys sp. SU F
Aequipecten lineolaris
(Lamarck, 1819)
SU R
Amusium papyraceum(Gabb, 1873)
SU F F
Lyropecten nodosus
(Linne, 1758)
SU R
Lucinidae Codakia orbicularis(Linne, 1758)
DEP R R
Codakia orbiculata
(Montagu, 1808)
DEP R
Chamidae Chama congregataConrad, 1833
SU R
Chama macerophylla
Gmelin, 1791
SU R
Chama sardaReeve, 1847
SU F
Cardiidae Trachycardium
muricatum(Linne, 1758)
SU R
Laevicardium pictum
(Ravenel, 1861)
SU A
Veneridae Ventricolaria rigida(Dillwyn, 1817)
SU R
Chione cancellata
(Linne, 1767)
SU R
Chione paphia(Linne, 1767)
SU R F
Pitar albidus
(Gmelin, 1791)
SU R R
Cyclinella tenuis
(Recluz, 1852)
SU R
Table 2 (continued)
Mollusc species Feeding
type
Fossil
reef areas
Soft-bottom
areas
Mactridae Mactra fragillis
Gmelin, 1791
SU R
Tellinidae Tellina lineataTurton, 1819
DEP R
Tellina sp. DEP R
Tellidora sp. DEP R
Solenidae Solen obliquusSpengler, 1794
SU R
Corbulidae Corbula dietziana
C.B. Adams, 1852
SU F
Periplomatidae Periploma sp. SU R
Poromyidae Poromya rostrata
Rehder, 1943
CP R
Gasteropoda
Fissurellidae Lucapina suffusa
(Reeve, 1850)
CB R
Trochidae Calliostoma sp. CB R
Rissoinidae Rissoina decussata
(Montagu, 1803)
HP R
Architectonicidae Architectonica nobilis
Roding, 1798
CB F
Turritellidae Turritella variegata
(Linne, 1758)
SU R R
Turritella exoleta
(Linne, 1758)
SU F
Vermicularia
spirata
(Philippi, 1836)
SU F R
Epitoniidae Epitonium krebsii
(Morch, 1874)
CB R R
Calyptraeidae Calyptraea centralis
(Conrad, 1841)
SU A R
Crucibulum
auricula
(Gmelin, 1791)
SU R
Crucibulum marenseWeisbord, 1962
SU R
Capulidae Capulus incurvatus
(Gmelin, 1791)
SU A
Naticidae Natica canrena(Linne, 1758)
CP R
Natica cayennensis
Recluz, 1850
CP R
Polinices hepaticus
(Roding, 1798)
CP R
Tonnidae Tonna galea
(Linne, 1758)
CP R R
Tonna maculosa
(Dillwyn, 1817)
CP F F
Cassidae Phalium granulatum
(Born, 1778)
CP R
Sconsia striata
(Lamarck, 1816)
CP R
Ranellidae Cymatium krebsi(Morch, 1877)
CP R
Cymatium
vespeceum
(Lamarck, 1822)
CP R
Distorsio clathrata
Lamarck, 1816
CP F A
Bursidae Bursa
spadicea(Montfort, 1810)
CP F
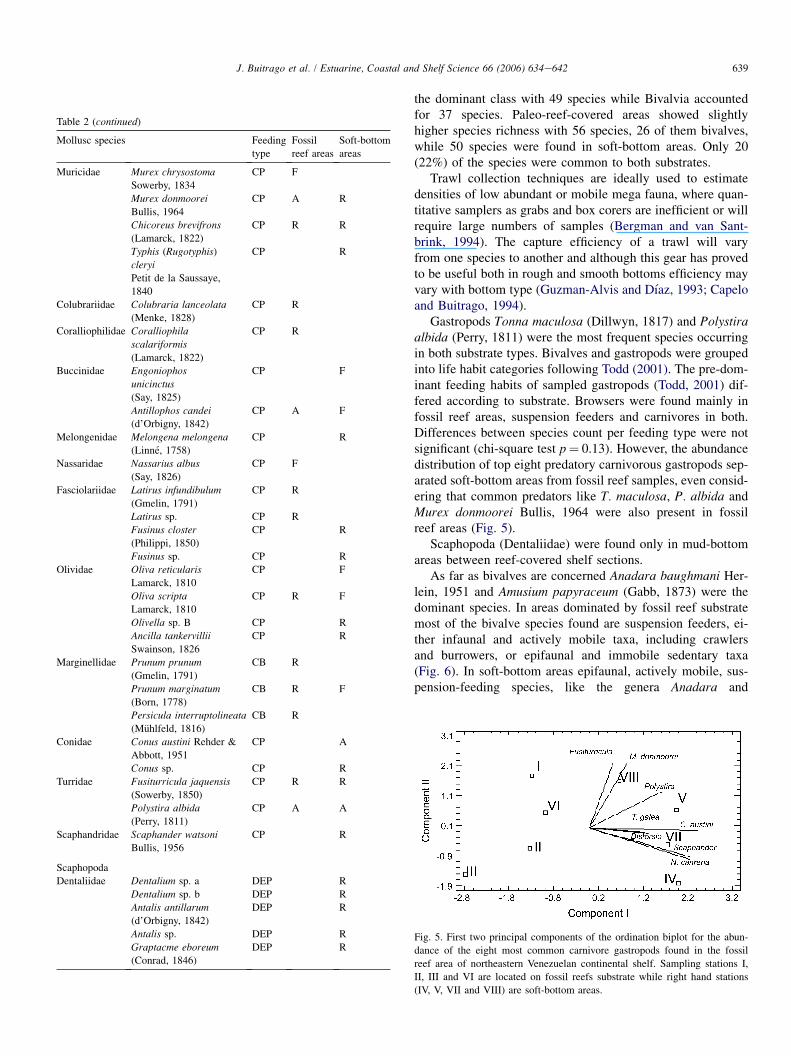
639J. Buitrago et al. / Estuarine, Coastal and Shelf Science 66 (2006) 634e642
Table 2 (continued)
Mollusc species Feeding
type
Fossil
reef areas
Soft-bottom
areas
Muricidae Murex chrysostoma
Sowerby, 1834
CP F
Murex donmoorei
Bullis, 1964
CP A R
Chicoreus brevifrons
(Lamarck, 1822)
CP R R
Typhis (Rugotyphis)
cleryiPetit de la Saussaye,
1840
CP R
Colubrariidae Colubraria lanceolata(Menke, 1828)
CP R
Coralliophilidae Coralliophila
scalariformis
(Lamarck, 1822)
CP R
Buccinidae Engoniophos
unicinctus
(Say, 1825)
CP F
Antillophos candei(d’Orbigny, 1842)
CP A F
Melongenidae Melongena melongena
(Linne, 1758)
CP R
Nassaridae Nassarius albus
(Say, 1826)
CP F
Fasciolariidae Latirus infundibulum
(Gmelin, 1791)
CP R
Latirus sp. CP R
Fusinus closter
(Philippi, 1850)
CP R
Fusinus sp. CP R
Olividae Oliva reticularis
Lamarck, 1810
CP F
Oliva scriptaLamarck, 1810
CP R F
Olivella sp. B CP R
Ancilla tankervillii
Swainson, 1826
CP R
Marginellidae Prunum prunum
(Gmelin, 1791)
CB R
Prunum marginatum
(Born, 1778)
CB R F
Persicula interruptolineata
(Muhlfeld, 1816)
CB R
Conidae Conus austini Rehder &
Abbott, 1951
CP A
Conus sp. CP R
Turridae Fusiturricula jaquensis
(Sowerby, 1850)
CP R R
Polystira albida
(Perry, 1811)
CP A A
Scaphandridae Scaphander watsoni
Bullis, 1956
CP R
Scaphopoda
Dentaliidae Dentalium sp. a DEP R
Dentalium sp. b DEP R
Antalis antillarum
(d’Orbigny, 1842)
DEP R
Antalis sp. DEP R
Graptacme eboreum(Conrad, 1846)
DEP R
the dominant class with 49 species while Bivalvia accountedfor 37 species. Paleo-reef-covered areas showed slightlyhigher species richness with 56 species, 26 of them bivalves,while 50 species were found in soft-bottom areas. Only 20(22%) of the species were common to both substrates.
Trawl collection techniques are ideally used to estimatedensities of low abundant or mobile mega fauna, where quan-titative samplers as grabs and box corers are inefficient or willrequire large numbers of samples (Bergman and van Sant-brink, 1994). The capture efficiency of a trawl will varyfrom one species to another and although this gear has provedto be useful both in rough and smooth bottoms efficiency mayvary with bottom type (Guzman-Alvis and Dıaz, 1993; Capeloand Buitrago, 1994).
Gastropods Tonna maculosa (Dillwyn, 1817) and Polystiraalbida (Perry, 1811) were the most frequent species occurringin both substrate types. Bivalves and gastropods were groupedinto life habit categories following Todd (2001). The pre-dom-inant feeding habits of sampled gastropods (Todd, 2001) dif-fered according to substrate. Browsers were found mainly infossil reef areas, suspension feeders and carnivores in both.Differences between species count per feeding type were notsignificant (chi-square test p ¼ 0.13). However, the abundancedistribution of top eight predatory carnivorous gastropods sep-arated soft-bottom areas from fossil reef samples, even consid-ering that common predators like T. maculosa, P. albida andMurex donmoorei Bullis, 1964 were also present in fossilreef areas (Fig. 5).
Scaphopoda (Dentaliidae) were found only in mud-bottomareas between reef-covered shelf sections.
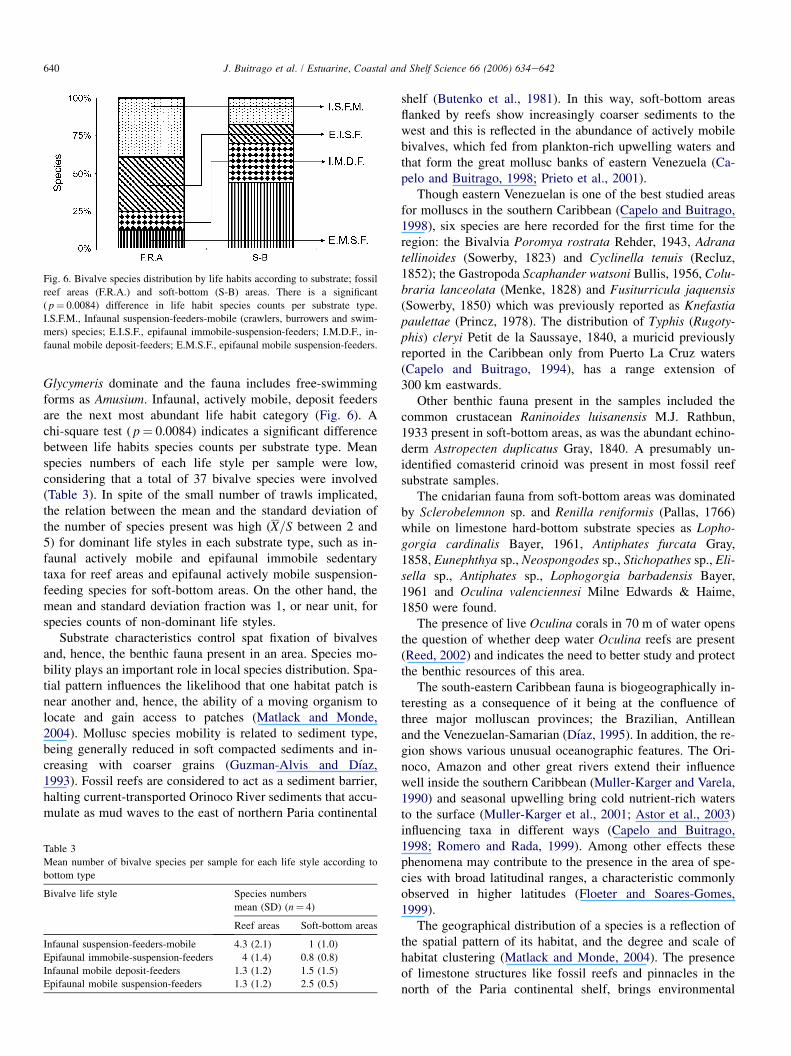
As far as bivalves are concerned Anadara baughmani Her-lein, 1951 and Amusium papyraceum (Gabb, 1873) were thedominant species. In areas dominated by fossil reef substratemost of the bivalve species found are suspension feeders, ei-ther infaunal and actively mobile taxa, including crawlersand burrowers, or epifaunal and immobile sedentary taxa(Fig. 6). In soft-bottom areas epifaunal, actively mobile, sus-pension-feeding species, like the genera Anadara and
Fig. 5. First two principal components of the ordination biplot for the abun-
dance of the eight most common carnivore gastropods found in the fossil
reef area of northeastern Venezuelan continental shelf. Sampling stations I,
II, III and VI are located on fossil reefs substrate while right hand stations
(IV, V, VII and VIII) are soft-bottom areas.
640 J. Buitrago et al. / Estuarine, Coastal and Shelf Science 66 (2006) 634e642
Glycymeris dominate and the fauna includes free-swimmingforms as Amusium. Infaunal, actively mobile, deposit feedersare the next most abundant life habit category (Fig. 6). Achi-square test ( p ¼ 0.0084) indicates a significant differencebetween life habits species counts per substrate type. Meanspecies numbers of each life style per sample were low,considering that a total of 37 bivalve species were involved(Table 3). In spite of the small number of trawls implicated,the relation between the mean and the standard deviation ofthe number of species present was high (X=S between 2 and5) for dominant life styles in each substrate type, such as in-faunal actively mobile and epifaunal immobile sedentarytaxa for reef areas and epifaunal actively mobile suspension-feeding species for soft-bottom areas. On the other hand, themean and standard deviation fraction was 1, or near unit, forspecies counts of non-dominant life styles.
Substrate characteristics control spat fixation of bivalvesand, hence, the benthic fauna present in an area. Species mo-bility plays an important role in local species distribution. Spa-tial pattern influences the likelihood that one habitat patch isnear another and, hence, the ability of a moving organism tolocate and gain access to patches (Matlack and Monde,2004). Mollusc species mobility is related to sediment type,being generally reduced in soft compacted sediments and in-creasing with coarser grains (Guzman-Alvis and Dıaz,1993). Fossil reefs are considered to act as a sediment barrier,halting current-transported Orinoco River sediments that accu-mulate as mud waves to the east of northern Paria continental
Fig. 6. Bivalve species distribution by life habits according to substrate; fossil
reef areas (F.R.A.) and soft-bottom (S-B) areas. There is a significant
( p ¼ 0.0084) difference in life habit species counts per substrate type.
I.S.F.M., Infaunal suspension-feeders-mobile (crawlers, burrowers and swim-
mers) species; E.I.S.F., epifaunal immobile-suspension-feeders; I.M.D.F., in-
faunal mobile deposit-feeders; E.M.S.F., epifaunal mobile suspension-feeders.
Table 3
Mean number of bivalve species per sample for each life style according to
bottom type
Bivalve life style Species numbers
mean (SD) (n ¼ 4)
Reef areas Soft-bottom areas
Infaunal suspension-feeders-mobile 4.3 (2.1) 1 (1.0)
Epifaunal immobile-suspension-feeders 4 (1.4) 0.8 (0.8)
Infaunal mobile deposit-feeders 1.3 (1.2) 1.5 (1.5)
Epifaunal mobile suspension-feeders 1.3 (1.2) 2.5 (0.5)
shelf (Butenko et al., 1981). In this way, soft-bottom areasflanked by reefs show increasingly coarser sediments to thewest and this is reflected in the abundance of actively mobilebivalves, which fed from plankton-rich upwelling waters andthat form the great mollusc banks of eastern Venezuela (Ca-pelo and Buitrago, 1998; Prieto et al., 2001).
Though eastern Venezuelan is one of the best studied areasfor molluscs in the southern Caribbean (Capelo and Buitrago,1998), six species are here recorded for the first time for theregion: the Bivalvia Poromya rostrata Rehder, 1943, Adranatellinoides (Sowerby, 1823) and Cyclinella tenuis (Recluz,1852); the Gastropoda Scaphander watsoni Bullis, 1956, Colu-braria lanceolata (Menke, 1828) and Fusiturricula jaquensis(Sowerby, 1850) which was previously reported as Knefastiapaulettae (Princz, 1978). The distribution of Typhis (Rugoty-phis) cleryi Petit de la Saussaye, 1840, a muricid previouslyreported in the Caribbean only from Puerto La Cruz waters(Capelo and Buitrago, 1994), has a range extension of300 km eastwards.
Other benthic fauna present in the samples included thecommon crustacean Raninoides luisanensis M.J. Rathbun,1933 present in soft-bottom areas, as was the abundant echino-derm Astropecten duplicatus Gray, 1840. A presumably un-identified comasterid crinoid was present in most fossil reefsubstrate samples.
The cnidarian fauna from soft-bottom areas was dominatedby Sclerobelemnon sp. and Renilla reniformis (Pallas, 1766)while on limestone hard-bottom substrate species as Lopho-gorgia cardinalis Bayer, 1961, Antiphates furcata Gray,1858, Eunephthya sp., Neospongodes sp., Stichopathes sp., Eli-sella sp., Antiphates sp., Lophogorgia barbadensis Bayer,1961 and Oculina valenciennesi Milne Edwards & Haime,1850 were found.
The presence of live Oculina corals in 70 m of water opensthe question of whether deep water Oculina reefs are present(Reed, 2002) and indicates the need to better study and protectthe benthic resources of this area.
The south-eastern Caribbean fauna is biogeographically in-teresting as a consequence of it being at the confluence ofthree major molluscan provinces; the Brazilian, Antilleanand the Venezuelan-Samarian (Dıaz, 1995). In addition, the re-gion shows various unusual oceanographic features. The Ori-noco, Amazon and other great rivers extend their influencewell inside the southern Caribbean (Muller-Karger and Varela,1990) and seasonal upwelling bring cold nutrient-rich watersto the surface (Muller-Karger et al., 2001; Astor et al., 2003)influencing taxa in different ways (Capelo and Buitrago,1998; Romero and Rada, 1999). Among other effects thesephenomena may contribute to the presence in the area of spe-cies with broad latitudinal ranges, a characteristic commonlyobserved in higher latitudes (Floeter and Soares-Gomes,1999).
The geographical distribution of a species is a reflection ofthe spatial pattern of its habitat, and the degree and scale ofhabitat clustering (Matlack and Monde, 2004). The presenceof limestone structures like fossil reefs and pinnacles in thenorth of the Paria continental shelf, brings environmental

641J. Buitrago et al. / Estuarine, Coastal and Shelf Science 66 (2006) 634e642
heterogeneity to the substrate of a region otherwise dominatedby soft-bottom, river-transported sediments. These remarkabletopographic features, in a region that supports intense fisheriesand is targeted for important oil industry activities, suggestthat it may be evaluated as a possible Marine ProtectedArea. Topographic heterogeneity plays an important role inmarine ecosystems (Turner et al., 1999); provides shelter tofish populations of regional importance and may be significantas breeding and nursery areas. Limestone substrate providesa habitat to benthic species that plays an important role in re-gional food webs. Conservation planning has long embracedspatial connectivity of habitat as a goal, hence the geographicconnections of this hard-bottom area with the nearby concen-trations of benthic molluscs like Arca zebra (Swainson, 1833)and Pinctada imbricata Roding 1798 that forms large oysterreefs and represent a regionally important fisheries resource,suggest that it should be considered as a potential bioticcorridor.
The effects of this ecologically significant zone on sur-rounding areas in terms of nursery habitat availability, poten-tial for spill-over of adult fish to other areas, importance andcapacity as a reproductive environment or feeding area, hencereducing the risk of population collapse in commercial fisher-ies species, should be evaluated.
Acknowledgments
This work was partially carried on during a service contractwith PDVSA (Petroleos de Venezuela, S.A.), to characterizethe VLNG (Venezuelan Liquefied Natural Gas) project area.The authors thank J. Guaiquirian, J.J. Cardenas, A. Achuryand all the personnel from acoustic surveys for use of rawechograms. The authors thank Y. Astor for improving the En-glish. An anonymous referee contributed greatly to manuscriptimprovement. Contribution N � 333 from Estacion de Investi-gaciones Marinas de Margarita de Fundacion La Salle deCiencias Naturales.
References
Abbott, R.T., 1974. American Seashells, second ed. Van Nostrand Reinhold
Co., New York, 663 pp., 24 pls.
Alvarez, M., Garcia, N., Prieto, A.S., 1992. Ecologıa de tres comunidades de
moluscos bentonicos en la costa noreste del estado Sucre. Acta Cientıfica
Venezolana 43 (Suppl. 1), 60.
Angel, A.F., Ojeda, P., 2001. Structure and trophic organization of subtidal fish
assemblages on the northern Chilean coast: the effect of habitat complex-
ity. Marine Ecology Progress Series 217, 81e91.
Astor, Y., Muller-Karger, F., Scranton, M., 2003. Seasonal and interannual var-
iation in the hydrography of the Cariaco basin: implications for basin ven-
tilation. Continental Shelf Research 23, 125e144.
Bavestrello, G., Bianchi, C.N., Calcinai, B., Cattaneo-Vietti, R., Cerrano, C.,
Morri, C., Puce, S., Sara, M., 2000. Bio-mineralogy as a structuring factor
for marine epibenthic communities. Marine Ecology Progress Series 193,
241e249.
Bayer, F.M., 1971. New and unusual mollusks collected by R/V Jhon Elliot
and R/V Gerda, in Tropical Western Atlantic. Bulletin of Marine Science
21, 111e236.
Bergman, M.J.N., van Santbrink, J.W., 1994. A new benthos dredge (‘triple-
D’) for quantitative sampling of infauna species of low abundance. Nether-
lands Journal of Sea Research 33, 129e133.
Blanchon, P., Jones, B., Ford, D.C., 2002. Discovery of a submerged relic reef
and shoreline off Grand Cayman: further support for an early Holocene
jump in sea level. Sedimentary Geology 147, 253e270.
Bloom, A., 1994. The coral record of late glacial sea level rise. In:
Ginsburg, R. (Ed.), Proceedings of the Colloquium on Global Aspects of
Coral Reefs: Health, Hazards and History, 1993. Rosentiel School of Ma-
rine and Atmospheric Science, University of Miami, pp. 1e6.
Blum, M.D., Carter, A.E., Zayac, T., Goble, R., 2002. Middle Holocene sea-
level and evolution of the Gulf of Mexico coast (USA). Journal of Coastal
Research, Special Issue 36, 65e80.
Butenko, J., Barbat, J.P., Daza, J., 1981. Singularidades geologicas exploradas
con el mini submarino del Calypso en Venezuela. Revista Tecnica de IN-
TEVEP 1, 27e35.Capelo, J., Buitrago, J., 1994. Presencia de Typhis (Rugotyphis) cleryi. Petit de
la Saussage, 1840. en las aguas costeras de Venezuela. Memoria de la So-
ciedad de Ciencias Naturales La Salle 141, 27e39.Capelo, J., Buitrago, J., 1998. Distribucion Geografica de los Moluscos Mar-
inos en el Oriente de Venezuela. Memoria de la Sociedad de Ciencias Nat-
urales La Salle 150, 109e160.
Cardenas, J.J., Achury, A., 2000. Acustica pesquera de los recursos marinos
del nororiente de Venezuela: evaluacion y regimen espacio-temporal del
stock de sardina (Sardinella aurita Valenciennes, 1847). Memoria de la So-
ciedad de Ciencias Naturales La Salle 154, 39e54.
Clark, P.U., Mitrovica, J.X., Milne, G.A., Tamisiea, M.E., 2002. Sea-level fin-
gerprinting as a direct test for the source of global meltwater pulse. Sci-
ence 295, 2438e2441.
De Jong, K.M., Coomans, H.E., 1988. Marine Gastropods from Curazao, Ar-
uba and Bonaire. J. Brill, Leiden, 261 pp.
Dıaz, J.M., 1995. Zoogeography of marine gastropods in the southern Carib-
bean. A new look at provinciality. Caribbean Journal of Science 31,
104e121.Dıaz, J.M., Puyana, M., 1994. Moluscos del Caribe Colombiano Colciencias.
Fundacion Natura. Invemar, Santa Marta, 277 pp.
Floeter, S.R., Soares-Gomes, A., 1999. Biogeographic and species richness
patterns of Gastropoda on the Southwestern Atlantic. Revista Brasileira
de Biologia 59, 567e575.
Gerlotto, F., Gines, H., 1988. Diez anos de ecointegracion en EDIMAR, re-
ferida a la sardina del oriente venezolano (Sardinella aurita). Memoria
de la Sociedad de Ciencias Naturales La Salle XLVIII (Suppl. 3),
311e324.
Gonzalez, L.W., 1991. Indice de abundancia de la pesquerıa de media altura
pargo-mero en la region oriental de Venezuela. Memoria de la Sociedad
de Ciencias Naturales La Salle 135&136, 159e168.
Guzman-Alvis, I., Dıaz, J.M., 1993. Distribucion especial de la Taxocenosis
Annelida-Mollusca en la Plataforma Continental del Golfo de Salamanca,
Caribe Colombiano. Anales del Instituto de Investigaciones Marinas y
Costeras de Punta Betin 22, 45e59.
Institute of Marine Research, 1989. Surveys of the Fish Resources in the Shelf
Areas Between Suriname and Colombia 1988. Institute of Marine Re-
search, Bergen, 141 pp.
Letourneur, Y., Ruitton, S., Sartoretto, S., 2003. Environmental and benthic
habitat factors structuring the spatial distribution of a summer infralit-
toral fish assemblage in the north-western Mediterranean Sea. Journal
of the Marine Biological Association of the United Kingdom 83,
193e204.
Ludwig, J.C., Walton, W.R., 1957. Shelf-edge calcareous brominences in
northeastern Gulf of Mexico. Bulletin of the American Association of Pe-
troleum Geologists 41, 2054e2101.
Matlack, G.R., Monde, J., 2004. Consequences of low mobility in spatially
and temporally heterogeneous ecosystems. Journal of Ecology 92,
1025e1035.Mendoza, J., Sanchez, L., Marcano, L., 1994. Variaciones en la distribucion y
abundancia de los principales recursos demersales del nororiente de Ven-
ezuela. I. Peces. Memoria de la Sociedad de Ciencias Naturales La Salle
142, 47e64.

642 J. Buitrago et al. / Estuarine, Coastal and Shelf Science 66 (2006) 634e642
Milliman, J.D., 1980. Nature and origin of limestones from the north of Paria
Continental Shelf. Informe a INTEVEP S.A.
Milliman, J.D., Butenko, J., Barbot, J.P., 1980. Holocene sediments from the
Orinoco River on the Paria Shelf, Northern Venezuela. 26th International
Geological Congress, Paris, July 1980.
Milliman, J.D., Butenko, J., Barbot, J.P., Hedberg, J., 1982. Depositional pat-
terns of modern Orinoco/Amazon muds on the northern Venezuelan shelf.
Journal of Marine Research 7, 643e657.Muller-Karger, F., McClain, C.R., Fisher, T.R., Esaias, W.E., Varela, R., 1989.
Pigment distribution in the Caribbean sea: observations from space. Prog-
ress in Oceanography 23, 23e64.
Muller-Karger, F., Varela, R., 1990. Influjo del rıo Orinoco en el mar Caribe:
observaciones con el CZCS desde el espacio. Memoria de la Sociedad de
Ciencias Naturales La Salle 131&134, 361e390.
Muller-Karger, F., Varela, R., Thunell, R., Scranton, M., Boher, R., Taylor, G.,
Capelo, J., Astor, Y., Tappa, E., Ho, T.Y., Walsh, J.J., 2001. Annual cycle of
primary production in the Cariaco basin: response to upwelling and implica-
tions for vertical export. Journal of Geophysical Research 106, 4527e4542.
Nagelkerken, I., Dorenbosch, M., Verberk, W.C.E.P., Cocheret, de la
Moriniere, E., Van der Velde, G., 2000. Importance of shallow-water bio-
topes of a Caribbean bay for juvenile coral reef fishes: patterns in biotope
association, community structure and spatial distribution. Marine Ecology
Progress Series 202, 175e192.Obrochta, S.P., Duncan, D.S., Brooks, R.G., 2003. Hardbottom development
and significance to the sediment-starved west-central Florida inner conti-
nental shelf. Marine Geology 200, 91e306.
Peterson, L.C., Haug, G.H., Murray, R.W., Yarincik, K.M., King, J.W.,
Bralower, T.J., Kameo, K., Rutherford, S.D., Pearce, R.B., 2000. 4. Late
Quaternary Stratigraphy and Sedimentation at Site 1002, Cariaco Basin
(Venezuela). In: Leckie, R.M., Sigurdsson, H., Acton, G.D., Draper, G.
(Eds.), Proceedings of the Ocean Drilling Program, Scientific Results,
vol. 165, pp. 85e99.
Prieto, A., Ruiz, L.J., Garcıa, N., Alvarez, M., 2001. Diversidad malacologica
en una comunidad de Arca zebra (Mollusca: Bivalvia) en Chacopata, Es-
tado Sucre, Venezuela. Revista de Biologıa Tropical 49, 591e598.
Princz, D., 1982. Lista y Bibliografıa de los Gastropodos marinos vivos de los
mares de Venezuela. Trinidad e islas vecinas de sotavento Boletın de la So-
ciedad de Ciencias Naturales La Salle 37, 103e107.
Reed, J.K., 2002. Deep-water Oculina coral reefs of Florida: biology, impacts,
and management. Hidrobiologıa 471, 43e55.
Rios, E.C., 1975. Brazilian Marine Mollusks Iconography. Fundacao Univer-
sidade do Rio Grande, Centro de Ciencias do Mar, Rio Grande, 331 pp.
Rogers, S.I., Lockwood, S.J., 1989. Observations on the capture efficiency of
a two-metre beam trawl for juvenile flatfish. Netherlands Journal of Sea
Research 23, 347e352.
Romero, N., Rada, M., 1999. Octocorales de las aguas costeras de la Isla de
Margarita, Estado Nueva Esparta, Venezuela. Memoria de la Sociedad
de Ciencias Naturales La Salle 59, 95e115.Shaked, Y., Marco, S., Lazar, B., Stein, M., Cohen, C., Sass, E., Agnon, A.,
2002. Late Holocene shorelines at the Gulf of Aqaba: migrating shorelines
under conditions of tectonic and sea level stability. EGU Stephan Mueller
Special Publication Series 2, 105e111.Swanson, C.A., 2004. Effect of substrate availability and conspecific cues on
communal oviposition in the apple murex snail Phyllonotus pomum.
Marine Ecology Progress Series 275, 175e184.Tello, J., 1975. Catalogo de la fauna venezolana. VII. Mollusca. C.O. III.
Conferencia de las Naciones Unidas sobre el Derecho del Mar, 599 pp.
Todd, J.A., 2001. Introduction to molluscan life habits databases. http://porites.
geology.uiowa.edu/database/mollusc/mollusclifestyles.htm. In: Budd, A.F.
(Ed.), NeogeneMarine Biota of Tropical America (NMITA) web systematic
database. http://nmita.geology.uiowa.edu/index.htm.
Toscano, M.A., Lundberg, J., 1998. Early Holocene sea-level record from
submerged fossil reefs on the southeast Florida margin. Geology 26,
255e258.
Turgeon, D.D., Asch, R.G., Causey, B.D., Dodge, R.E., Jaap, W., Banks, K.,
Delaney, J., Keller, B.D., Speiler, R., Matos, C.A., Garcia, J.R., Diaz, E.,
Catanzaro, D., Rogers, C.S., Hillis-Starr, Z., Nemeth, R., Taylor, M.,
Schmahl, G.P., Miller, M.W., Gulko, D.A., Maragos, J.E.,
Friedlander, A.M., Hunter, C.L., Brainard, R.S., Craig, P., Richond, R.H.,
Davis, G., Starmer, J., Trianni, M., Houk, P., Birkeland, C.E., Edward, A.,
Golbuu, Y., Gutierrez, J., Idechong, N., Paulay, G., Tafileichig, A., Vander
Velde, N., 2002. The State of Coral Reef Ecosystems of the United
States and Pacific Freely Associated States: 2002. National Oceanic and
Atmospheric Administration/National Ocean Service/National Centers for
Coastal Ocean Science, Silver Spring, MD, 265 pp.
Turner, S.J., Thrush, S.F., Hewitt, J.E., Cummings, V.J., Funnell, G., 1999.
Fishing impacts and the degradation or loss of habitat structure. Fisheries
Management and Ecology 6, 401e420.
United States Geological Services (USGS), 2002. Pinnacles Region, Northern
Gulf of Mexico. http://walrus.wr.usgs.gov/pacmaps/pn-index.html.
Copyright © 2022 FDOKUMEN




















