
Succession patterns in algal turf vegetation on a Caribbean coral reef
Upload
khangminh22Category
view
0download
0
www.elsevier.com/locate/jembe
Journal of Experimental Marine Biology and Ecology
300 (2004) 217–252
Experimental biology of coral reef ecosystems
Michael P. Lesser*
Department of Zoology and Center for Marine Biology, University of New Hampshire, Durham, NH 03824, USA
Received 23 November 2003; received in revised form 18 December 2003; accepted 28 December 2003
Abstract
Coral reef ecosystems are at the crossroads. While significant gaps still exist in our understanding
of how ‘‘normal’’ reefs work, unprecedented changes in coral reef systems have forced the research
community to change its focus from basic research to understand how one of the most diverse
ecosystems in the world works to basic research with strong applied implications to alleviate
damage, save, or restore coral reef ecosystems. A wide range of stressors on local, regional, and
global spatial scales including over fishing, diseases, large-scale disturbance events, global climate
change (e.g., ozone depletion, global warming), and over population have all contributed to declines
in coral cover or phase shifts in community structure on time scales never observed before. Many of
these changes are directly or indirectly related to anthropogenically induced changes in the global
support network that affects all ecosystems. This review focuses on some recent advances in the
experimental biology of coral reef ecosystems, and in particular scleractinian corals, at all levels of
biological organization. Many of the areas of interest and techniques discussed reflect a progression
of technological advances in biology and ecology but have found unique and timely application in
the field of experimental coral reef biology. The review, by nature, will not be exhaustive and reflects
the author’s interests to a large degree. Because of the voluminous literature available, an attempt has
been made to capture the essential elements and references for each topic discussed.
D 2004 Elsevier B.V. All rights reserved.
Keywords: Coral reef ecosystems; Experimental biology; Global climate change
Scleractinian, or reef-building corals, are a central component to coral reef ecosystems
worldwide between 30jN and 30jS latitude and contribute to thousands of square
kilometers of critical marine habitat. The prolific growth rates (3–15 cm year�1) of
reef-building corals in optically clear, oligotrophic tropical seas are responsible for the
three-dimensional framework of coral reef systems (Fig. 1). While other organisms serve
0022-0981/$ - see front matter D 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.jembe.2003.12.027
* Tel.: +1-603-862-3442; fax: +1-603-862-3784.
E-mail address: [email protected] (M.P. Lesser).

Fig. 1. Underwater photograph of coral reef in Indonesia with almost 100% cover of Acropora sp. (Photograph by
M. Lesser).
M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252218
to consolidate the framework of the reef structure together (e.g. calcareous algae) and use
it as essential habitat (e.g. fish, algae, invertebrates and bacteria), corals are the functional
group that has contributed significantly to coral reef ecosystems for at least 200 million
years (Veron, 1995) and have built the primary structure of entire reefs, islands and such
massive oceanic barriers as the barrier reefs of Mesoamerica and Australia. Coral reefs are
a source of food and livelihood for at least 100 million people worldwide, support major
industries (fishing and tourism), play a key role in stabilizing coastlines, and their high
species and genetic diversity rivals that of tropical rainforests (Connell, 1978; Hoegh-
Guldberg, 1999). This biodiversity is now just beginning to be exploited in the search for
bioactive compounds that could benefit humankind (Quinn et al., 2002).
Unfortunately, coral reefs are also experiencing unparalleled levels of anthropogenically
induced stress. Current estimates on the rate of decline in the health of coral reefs and the
loss or change in community structure of reefs are of worldwide concern (Wilkinson, 2000).
It is estimated that a combination of physical, chemical and biological stresses will cause the
decline of between 40% to 60% of the world’s coral reefs over the next 50 years unless
appropriate steps are taken (Wilkinson, 2000). Until recently, global climate change was
seen as just one of many factors (e.g., eutrophication, coastal development, sedimentation,
over-fishing) responsible for the decline in the health of coral reefs (Wilkinson, 1999) while
the time scales of change due to global climate effects were believed to be slow and other
anthropogenic causes a higher priority for study. In 1998, however, an estimated 16% of the
world’s living corals were eliminated in a single warming event related to El Nino
(Wilkinson, 2000). During this event, sea temperatures warmed to 2–3 jC above long-

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252 219
term average summer temperatures and resulted in a catastrophic ‘‘bleaching’’ event that
caused significant mortality of several species of coral (e.g., both the expulsion of
zooxanthallae and host tissue death occurred). The impact of this thermal event on the
percent cover of shallow coral reefs worldwide and the projection of continued rising sea
temperatures under greenhouse warming (Hoegh-Guldberg, 1999) has radically changed the
focus of a large proportion of the research community towards understanding the potential
impact of greenhouse-driven climate change on the world’s coral reefs. Bleaching as a result
of thermal stress is not the only threat from global climate change and coral reef biologists
from around the world have had to use new experimental tools at all levels of biological
organization in their efforts to understand how reefs work, determine which corals will
survive anthropogenically driven change, and predict what reefs will look like at the end of
the next century. In essence, who will be the winners and the losers (Loya et al., 2001)?
1. The coral–algal symbiosis
Coral reef communities contain a wide variety of mutualistic associations none more
important than the relationship between corals and their symbiotic dinoflagellates of the
genus Symbiodinium sp., commonly referred to as zooxanthellae. Scleractinian corals first
appeared in the Triassic (Veron, 1995), and it is widely accepted that their rapid ecological
success was directly related to the acquisition of dinoflagellate endosymbionts that enabled
the symbiosis to survive in oligotrophic and high solar irradiance habitats. Corals acquire
the majority of their energetic and nutrient requirements by two mechanisms: photosyn-
thesis by their zooxanthellae and heterotrophy, or the direct ingestion of zooplankton and
other organic particles in the water column by the cnidarian host. The zooxanthellae reside
within vacuoles in the cells of the host gastrodermis (Fig. 2a and b; Trench, 1979, 1987)
where they serve as primary producers and supply their coral host with up to 95% of their
photosynthetic products, such as sugars, amino acids, carbohydrates and small peptides
(Trench, 1979; Muscatine, 1990) making corals autotrophic with respect to carbon in most
habitats. These compounds provide the coral with energy for respiration, growth, and the
deposition of its CaCO3 skeleton (Muscatine, 1990).
Supplying translocated photosynthate to the host contributes significantly to the fitness
of the symbiosis (Muscatine, 1990; Mueller-Parker and D’Elia, 1997) while in return the
zooxanthellae receive essential nutrients such as ammonia, phosphate, and carbon dioxide
from the metabolic wastes of the coral (Trench, 1979; Mueller-Parker and D’Elia, 1997).
Additionally, photoautotrophy is not the only source of nutrition for corals. An increasing
amount of experimental evidence continues to document that heterotrophy in corals is
essential for providing nitrogen, phosphorus, and other nutrients which make it possible
for the coral host to use the available carbon skeletons for protein synthesis and other
essential metabolic requirements. Initially, the degree of heterotrophy appeared to be
positively correlated with coral polyp size (Porter, 1976). Porter (1976) described a
bathymetric gradient from autotrophy in shallow waters to heterotrophy in deeper waters
that was correlated with polyp size in the Caribbean. Species with small polyps that were
more dependent on autotrophy were found in shallow waters while more heterotrophic
large polyp species of coral were found in deep waters (Porter, 1976). Clearly, heterotro-

Fig. 2. (a) Electron micrograph of zooxanthellae in hospite. (b) Phase-contrast micrograph of zooxanthellae in
tentacle squash preparation (Photographs by M. Lesser and T. LaJeuness).
M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252220
phy in corals is important. Glynn (1973) described plankton depletion on a coral reef as
water flowed past and Wellington (1982) provided experimental, multifactorial, evidence
that supported Porter’s autotrophy to heterotrophy gradient, but also showed that
heterotrophy did not compensate for the decrease in solar irradiance with depth when
growth rates were measured. Recently, Sebens and colleagues (Sebens and Johnson, 1991;
Johnson and Sebens, 1993; Sebens et al., 1996, 1998) have shown quite convincingly that
both small and large polyped corals are successful at capturing certain size classes of
zooplankton and that any differences in the efficiency of capture were due largely to the
escape ability of the zooplankton.
Whether from autotrophy or heterotrophy, the tight recycling of nutrients within the
coral symbiosis and the close coupling between trophic levels at reasonably high

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252 221
efficiencies contribute to the very high productivity of corals (Muscatine and Porter, 1977;
Falkowski et al., 1984; Muscatine, 1990; Mueller-Parker and D’Elia, 1997). Nutrient
limitation imposed by the host on the algal symbionts is also believed to be part of a highly
regulated control mechanism on the growth of zooxanthellae that would otherwise out-
divide their host cells at rates approaching those of free-living phytoplankton (Muscatine
and Porter, 1977). From an organismal and experimental perspective, it would appear that
the role of autotrophy and heterotrophy in the energetics and nutrient metabolism of corals
should be vigorously revisited. This will require simultaneous and interdisciplinary studies
by groups of collaborators on a range of coral species in different habitats using a range of
tools (e.g., fluorescence measurements, feeding studies, stable isotopes) to fill in what
appear to be large gaps in our understanding. By definition mutualistic associations incur
both benefits and costs for the partnered species. For any mutualistic symbiosis to develop
and persist, a constant evaluation of the costs and benefits must be occurring such that the
selective pressure favors those associations where the benefit to both partners outweighs
the costs (Cushman and Beattie, 1991). Under the continuing scenario of rapid change on
coral reefs, it is important to understand, at an organismal level, which species will survive
in the broad range of trophic strategies that span the dependence on autotrophy versus
heterotrophy.
2. Hurricanes, overfishing, eutrophication, bleaching, and community phase shifts
Both the growth forms and species of corals show typical and well-described zonational
patterns on reefs worldwide (Loya, 1972; Huston, 1985; Done, 1995). While heterogeneity
exists, species diversity along a bathymetric gradient is predictable to a certain degree and
reflects both biotic and abiotic processes. Much of the recent ecological work on coral reefs
has been framed around the concept that reefs are non-equilibrium systems whose
community structure and diversity are largely determined by the intensity and rate of
disturbance as described in the intermediate disturbance hypothesis (Connell, 1978, 1997;
Connell et al., 1997). Additionally, strong latitudinal and bathymetric gradients in abiotic
factors such as solar irradiance, water flow, and calcium carbonate saturation state
significantly influence the community structure, growth forms, and state of photoacclima-
tization over both small and large spatial scales (Falkowski et al., 1990; Done, 1995;
Wilkinson, 1999; Lesser et al., 2000). The scale-dependent variability in coral reef
community structure continues to be an important area of study for understanding not
only the range of scales at which different patterns occur but also what processes at different
scales may be driving that variability (Murdoch and Aronson, 1999).
The current concern by coral reef biologists is that the periodicity and intensity of
disturbance events, which now include a suite of anthropogenic factors over large (e.g.,
kilometer) spatial scales, is rapidly changing coral reefs and threatening their existence
which is in juxtaposition to the long-term persistence of coral reefs over geological time
scales (Pandolphi, 1999). Most coral reef biologists do agree that coral reefs are changing
and will exist in the near future but they will not be the ‘‘coral reefs’’ we have come to know
inmany parts of the world (Knowlton, 2001). The outcome on each reef systemwill likely be
determined by a combination of the number and severity of insults, but also which set of the

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252222
unique and varied life-history traits will be able to cope with these stressors on ecological
and evolutionary time scales (Hughes et al., 1992; Done et al., 1996).
Jackson et al. (2001) demonstrate from several sources of historical data that a range of
disturbances including overfishing and coastal development have consistently led to major
changes in coral reefs ecosystem structure and health. The most poignant example of the
effects of anthropogenic influences is the state of reefs in the Caribbean. A recent meta-
analysis of coral cover throughout the Caribbean has shown an 80% decline in percent
coral cover that has been both long-term (e.g., decadal) and region-wide (Gardner et al.,
2003). Though many reefs worldwide have suffered similar reductions in coral cover
(McClanahan, 2002), most Caribbean reefs have undergone a shift from being coral-
dominated to algal-dominated in this time period (Hughes, 1994). The causes of this shift
vary from reef to reef but are the result of several types of disturbance that include
hurricane damage (Hughes, 1994; Hughes and Connell, 1999), eutrophication (Lapointe,
1997), thermal stress resulting in coral bleaching (Hoegh-Guldberg, 1999; Aronson et al.,
2000, 2002; Ostrander et al., 2000), coral diseases (Harvell et al., 2002; Richardson, 1998;
Rosenberg and Ben-Haim, 2002), the transport and deposition of sand and dust from the
Sahara in the Caribbean, which may be a factor that partially explains the increase in coral
diseases (Shinn et al., 2000), and reduced herbivory from over-fishing compounded by an
epizootic of unknown etiology that decimated Diadema populations in the 1980s
(Carpenter, 1988; Hughes, 1994).
Hughes (1994) described the rapid and significant ecological changes that occurred on
coral reefs in Jamaica when herbivores were removed by fishing, to the point where reef
resilience (i.e. ability to recover from a disturbance) was lost and a permanent phase shift to
algal-dominated communities began. Additionally, natural factors conspired with anthro-
pogenic stresses to produce this outcome. First, Hurricane Allen, a category five hurricane
struck Jamaica after almost 40 years without any significant storm damage to coral reefs.
While most of the damage occurred in shallow waters (<10 m) a period of recovery began
but was short-lived due to the loss of the sea urchin, Diadema antillarum, between 1982
and 1984 from an epizootic disease of unknown etiology. This Caribbean-wide loss of a
critical herbivore that controlled algal growth led to significant changes on these coral reefs.
Without urchin and fish herbivores on coral reefs around the Caribbean, large populations
of foliose macrophytes formed and prevented coral settlement and growth with the net
result that coral cover dramatically declined. Significant bleaching events beginning in the
late 1980s also caused coral mortality and further shifted the change to algal-dominated
reefs. This scenario has been repeated in many parts of the world as key elements of coral
reef communities, such as grazing fishes and invertebrates, have been removed resulting in
dramatic changes in community structure (Lewis, 1986; Jackson et al., 2001). Recently,
however, there is evidence that Diadema populations may be recovering. Edmunds and
Carpenter (2001) reported that at several sites along the North coast of Jamaica, urchin
populations have increased significantly by 10-fold and is correlated with an 11-fold
increase in the density of juvenile corals. Recovery of urchins may initiate beginning of a
shift from one alternative stable state to another throughout Jamaica and the rest of the
Caribbean.
While the work of Edmunds and Carpenter (2001) appears to clearly indicate that top-
down control of macroalgal growth is essential to ‘‘reef health’’ or maintenance of coral reef

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252 223
community structure as we know it, many coral reef biologists worldwide are still
discussing what controls macroalgal abundance on coral reefs. The two sides of this debate
have fallen along the ‘‘bottom-up’’ versus ‘‘top-down’’ dichotomy. The controversy is
centered around whether algal blooms are kept in check by herbivory as discussed in a
critique of Lapointe (1997) by Hughes et al. (1999), or responding to nutrient availability at
critical ‘‘threshold concentrations’’ that determine the balance between algal bloom and
non-bloom conditions Lapointe (1997,1999). In reading these papers, one can appreciate
the fact that each group recognizes the potential role of the ecological process discussed by
the other, but neither group has incorporated the opposing ecological process in their
experimental evaluation of this issue. Complicating the debate had been the lack of rigorous
multifactorial field experiments examining both herbivory and nutrient concentrations in
the field. Additionally, many of the sites examined (e.g., Jamaica and the Florida Keys)
have also experienced repeated disturbances, both natural and anthropogenic (see above),
that complicate the assessment of whether ‘‘bottom-up’’ or ‘‘top-down’’ processes from
either of these sites can be universally applied to all reefs.
Miller et al. (1999), however, provide us with experimental insight into what may be
occurring along the Florida reef tract. First, their design was multifactorial (herbivory and
nutrients), well replicated (i.e., without pseudoreplication), and appropriately analyzed
despite the complications associated with the field work that occurred during their study. An
aspect of their paper that will be of interest to experimental ecologists is the unique method
of nutrient addition amongst treatments. Fertilizer spikes commonly used in gardens were
partially covered in paraffin, and entombed in cinder blocks with holes to allow for the
diffusion of nutrients into the environment. The results of the field experiments by Miller et
al. were interpreted in the context of the Littler and Littler (1991) ‘‘relative dominance
model’’ for corals, turf algae, crustose coralline algae, and foliose macrophytes and the
‘‘threshold nutrient’’ model of Lapointe (1997). The Miller et al. paper provides a test of the
relative dominance model, that supports a strong role for herbivore control of algae on coral
reefs while not supporting the ‘‘threshold nutrient’’ hypothesis of Lapointe (1997). Another
paper by Jompa and McCook (2002) experimentally demonstrated the subtle affects of
herbivory when nutrients were also in abundance. Their experimental design clearly
revealed that herbivores are the single most important factor affecting coral growth and
mortality but that nutrients can also have an effect, but only if herbivory is weak and allows
nutrients to control algal growth and subsequently competitive interactions with corals.
Finally, a recent paper by Aronson and Precht (2000) examined the effects of herbivory on
Jamaican reefs over time in a nonmanipulative manner, essentially a ‘‘natural experiment’’.
Despite the shortfalls of a natural experiment, the Aronson and Precht (2000)results support
the experimental work of Miller et al. (1999) and Jompa and McCook (2002) which both
show that herbivory, or top-down control of macroalgae on coral reefs, has a greater impact
than nutrients on the community structure of coral reefs.
3. Global climate change
There can be little doubt at this point in time that global climate change, principally the
emission of greenhouse gases (e.g., CO2, CH4), and its subsequent effects on seawater

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252224
temperature, calcium carbonate saturation point, large-scale changes in atmospheric/
oceanic coupling [e.g., El Nino-Southern Oscillation (ENSO)], and changes in sea level,
is occurring (Smith and Buddemeier, 1992; Huppert and Stone, 1998; Hoegh-Guldberg,
1999; Kleypas et al., 1999; Wilkinson, 1999; Crowley, 2000; Stott et al., 2000; Urban et
al., 2000; Wellington et al., 2001). There is concern that within the framework of
evolutionary adaptation, scleractinian corals will not be able to physiologically adapt at
the current rates of environmental changes (Gates and Edmunds, 1999). In particular,
increases in seawater temperatures are thought to be the primary cause of the unprece-
dented number of ‘‘coral bleaching’’ events since the early 1980s (Brown, 1997; Glynn,
1991, 1993), with predictions for continued increases of seawater temperature in the future
(Hoegh-Guldberg, 1999). In 1998, coral reefs experienced the largest and most widespread
thermally induced mass bleaching of corals ever recorded with an estimated 16% mortality
of the world’s living corals as a result of an ENSO event (Wilkinson, 2000).
Even within tropical and sub-tropical environments, temperature is a pervasive abiotic
factor controlling the distribution and abundance of corals. Many species exhibit
population specific ranges of temperature where growth, reproduction, and survival will
occur, and is also related to the mean temperature of the warmest month in that
geographical location. It is generally believed that corals are living very close to their
upper thermal limits (Jokiel and Coles, 1990) where temperature effects on respiration are
much greater than on photosynthesis, resulting in a decrease in the P/R ratio and reduction
in net photosynthesis during exposure to elevated temperatures. When seawater temper-
atures warm anywhere from 2 to 3 jC above long-term average summer temperatures,
corals exhibit the stress response known as bleaching. Several field and laboratory studies
on bleaching in corals and other symbiotic cnidarians have established a causal link
between temperature stress and bleaching (Hoegh-Guldberg and Smith, 1989; Jokiel and
Coles, 1990; Lesser et al., 1990; Glynn, 1991; Fitt et al., 1993; Lesser, 1997; Hoegh-
Guldberg, 1999; Coles and Brown, 2003), and the extent of bleaching, subsequent
mortality, and the underlying mechanism (s) are related to the magnitude of temperature
elevation and the duration of exposure for any individual event. As is typical, and
consistent with experimental results, the severity for coral bleaching events varies in space
and time. For example, during the 1998 ENSO event 48% of corals in the Indian Ocean
died while only 3–5% of corals died on the Great Barrier Reef. The 1998 bleaching event,
unlike any previously observed, began a new appreciation for the projection of rapidly
rising sea temperatures under greenhouse warming (Fig. 3) and has increased the concern
of scientists, governments, and the general population about the potential impact of
greenhouse-driven climate change on the world’s coral reefs (Hoegh-Guldberg, 1999;
Wilkinson et al., 1999; McClanahan et al., 2002).
The number and severity of coral bleaching events are believed to be a ‘‘biological
signal’’ (sensu Hughes, 2000) for the consequences of global climate change on coral reefs
that is occurring worldwide, and is predicted to continue if current trends persist (Hoegh-
Guldberg, 1999; Sheppard, 2003). Coral bleaching is defined here as a response to
environmental stress that leads to a series of cellular responses that culminates in the
expulsion of the symbiotic zooxanthellae from the coral host tissues causing a paling or
whitening of the affected coral (Fig. 4a and b). Defining bleaching is not a trivial issue. It
is important to put bleaching in the context of the seasonal cycling of zooxanthellae

Fig. 3. Sea surface temperature data generated by the global-ocean-ice-model forced by greenhouse emissions that
conform to the IPCC scenario IS92a (Tahiti, 17.5jS, 149.5jW; Phuket, 7.5jN, 98.5jE; Jamaica, 17.5jN, 76.5jW).
With permission from Ove Hoegh-Guldberg and Marine and Freshwater Research (SO: 839–866, 1999).
M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252 225
densities in reef corals, which has recently been documented (Fagoonee et al., 1999; Fitt et
al., 2000).The annual high in sea surface temperatures coincides with yearly lows in
zooxanthellae densities in all studies made to date (Stimson, 1997; Fagoonee et al., 1999;
Fitt et al., 2000) suggesting the phenomenon is universal amongst symboitic corals.
Seasonal cycles in the quantum yields of chlorophyll fluorescence of corals have also been
observed (Warner et al., 2002), revealing seasonal acclimatization in solar irradiance and
seawater temperature.
While thermal stress is seen as the principal cause of coral bleaching, other environ-
mental factors, including those that are affected by anthropogenic influences, can act
synergistically by effectively lowering the threshold temperature at which coral bleaching
occurs. The principal abiotic factor that has significant influence on the severity of
thermally induced coral bleaching is solar radiation, both its visible (photosynthetically
active radiation, PAR: 400–700 nm; Hoegh-Guldberg and Smith, 1989; Dunne and
Brown, 2001) and ultraviolet (UVR: 290–400 nm, UVB: 290–320 nm, UVA: 320–400
nm; Shick et al., 1996) components.
The global decrease of stratospheric ozone from anthropogenic inputs of chlorinated
fluorocarbons has resulted in an increase in the amount of harmful UVB radiation reaching

Fig. 4. (a) Underwater photo of bleached M. faveolata in the vicinity of Lee Stocking Island (Rainbow Gardens),
Bahamas (Photograph by M. Lesser). (b) Underwater photograph of bleached Acropora sp. on the Great Barrier
Reef (Photograph by R. Berkelmans).
M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252226
the sea surface (Madronich et al., 1998). Although earlier concerns were centered on the
Antarctic, tropical ecosystems, with their smaller solar zenith angle and thinner layer of
ozone (Cutchis, 1982) have exposed tropical ecosystems over evolutionary time to higher
irradiances of UVR, and UVB in particular (Green et al., 1974; Frederick et al., 1989). In
absolute terms, even a small percentage decrease in stratospheric ozone over the tropics
would be important because the UVB irradiance there is already high. The optical
properties of tropical waters also result in low attenuation coefficients and allow UVR
to penetrate to depths of 15 m or more (Fig. 5, Smith and Baker, 1979; Gleason and
Wellington, 1993; Shick et al., 1996; Lesser, 2000; Lesser and Gorbunov, 2001). Although
tropical waters are generally more transparent to UVR than temperate waters, the water

Fig. 5. Depth profile of spectral irradiance data (300–700 nm) collected from the outer fore reef at Carrie Bow
Cay, Belize (17jN) in Spring 1996 using a LiCor 1800 UW scanning spectroradiometer. For details of instrument,
see Lesser (2000). Lesser, unpublished data.
M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252 227
column overlying coral reefs in coastal areas is susceptible to terrigenous inputs,
upwelling, and variations in dissolved organic matter that can affect its optical properties
(absorption and scattering) and increase the attenuation of UVR (Kirk, 1994).
Ultraviolet radiation is known to have a detrimental effect on photosynthesis and
growth in zooxanthellae (Shick et al., 1996) with the harmful effects of UVR involving
damage to DNA, proteins, and lipids. This damage may be the result of both the direct and
indirect effects of UVR on many cellular targets. For sessile corals, exposure to solar UVR
in shallow tropical waters is unavoidable and exposure to UVR is particularly important
during hyperoxic conditions (Dykens and Shick, 1982; Kuhl et al., 1995) that occur
intracellularly in corals during photosynthesis and leads to the photodynamic production
of reactive oxygen species (ROS) (Valenzeno and Pooler, 1987).
An important response of corals during exposure to UVR includes the synthesis of UVR-
absorbing compounds and enzymes involved in the protection of both the host and symbiont
from oxidative stress (Dykens, 1984; Dykens and Shick, 1984; Lesser and Shick, 1989;
Dykens et al., 1992; Shick et al., 1996; Lesser, 1996; Shick and Dunlap, 2000; Brown et al.,
2002; Lesser and Farrell, in press). UVR-absorbing compounds are believed to provide
protection from the high-energy wavelengths within the UVR part of the spectrum by
providing a broad-band filter as these compounds have absorption maxima in the UVR

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252228
portion of the spectrum from f310 to 360 nm. The concentration of UVR-absorbing
compounds [mycosporine-like amino acids (MAAs)] in corals shows an exponential
decrease with depth (Dunlap et al., 1986; Banaszak et al., 1998; Lesser, 2000) and
experimental evidence has shown that MAAs are produced by the symbiotic zooxanthellae
(Shick et al., 1999). MAA concentrations in corals, zoanthids and cultured zooxanthellae
have been shown to decrease upon exposure to elevated seawater temperatures (Lesser et al.,
1990; Glynn et al., 1992; Lesser, 1996; Shick and Dunlap, 2000), potentially leaving both
host tissues and symbiotic zooxanthellae more susceptible to biological damage caused by
exposure to UVR. Lastly, the MAA mycosporine-glycine is now know to also have
antioxidant activity (Dunlap and Yamamoto, 1995; Kim et al., 2001; Suh et al., 2003).
Other than Shick et al. (1999), there is no published work on the shikimic acid pathway in
zooxanthellae which synthesizes MAAs, and other environmental factors that could
influence carbon flux through this pathway. Jokiel et al. (1997) showed that differences in
the rates of water flow could influence MAA concentration under identical solar irradiances,
suggesting that photosynthetic rates andMAA synthesis are also under the control of carbon
delivery due to differences in the thickness of the diffusional boundary layer (Lesser et al.,
1994; Jokiel et al., 1997). In addtion to studies that show an increase in MAA concentration
upon exposure to UVR, Jokiel et al. (1997) suggested that the syntheses of MAAs may also
be controlled by the flux of carbon through the shikimic acid pathway which is in turn
affected by rates of photosynthesis. This MAAs follows carbon flux scenario has not
received general acceptance because Jokiel et al. (1997) did not describe a specific
stoichiometry between carbon flux and MAA concentration (Shick et al., 2000). Shick et
al. (2000), however, argues against MAAs synthesis being affected by carbon flux using the
calculations of photosynthesis and MAA concentration from laboratory experiments on a
free-living dinoflagellate (Neale et al., 1998). In the Neale et al. (1998) study the
concentration of MAAs was normalized to chlorophyll content, a co-varying factor that
overstates any differences inMAA concentration when cultures photoacclimated to different
irradiance regimes are compared. In any case the effects of changes in carbon flux on MAA
synthesis through the shikimic acid pathway has not received the appropriate experimental
attention to clearly show whether there is any relationship between carbon flux and MAA
concentrations and what other environmental factors affect the shikimic acid pathway and
subsequent biosynthesis of MAAs.
Exposure to elevated temperatures alone (Iglesias-Prieto et al., 1992), UVR alone
(Lesser and Shick, 1989), or in combination (Lesser, 1996, 1997) can result in photo-
inhibition of photosynthesis in zooxanthellae. Photoinhibition occurs as a result of the
reduction in photosynthetic electron transport, combined with the continued high absorp-
tion of excitation energy (Osmond, 1981). One consequence of reducing electron transport
is the production of ROS such as singlet oxygen [1O2] superoxide radicals [O2�], hydrogen
peroxide [H2O2], and hydroxyl radicals [OH], for which there are many cellular targets
including photosystem II and the primary carboxylating enzyme, Rubisco in zooxanthellae
(Lesser, 1996). The enzymes superoxide dismutase, catalase, and ascorbate peroxidase act
in concert to inactivate superoxide radicals and hydrogen peroxide, thereby preventing the
formation of the most reactive form of ROS, the hydroxyl radical, and subsequent cellular
damage (Fridovich, 1986). Enzymic defenses in the animal host occur in proportion to the
potential for photooxidative damage in symbiotic cnidarians (Dykens and Shick, 1982;

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252 229
Dykens et al., 1992). However, high fluxes of ROS in the host (Dykens et al., 1992; Nii
and Muscatine, 1997) or zooxanthellae (Lesser, 1996) can overwhelm the protective
enzymatic response and result in hydroxyl radical production via the Fenton reaction
(Asada and Takahashi, 1987). Oxidative stress has been proposed as a unifying mecha-
nism for several environmental insults that cause bleaching (Lesser, 1996). Oxidative
stress can lead to bleaching of zooxanthellae via exocytosis from coral host cells (Lesser,
1996, 1997) or apoptosis (Gates et al., 1992; Dunn et al., 2002; Lesser and Farrell, in
press). A cellular model of bleaching in symbiotic cnidarians has been developed and
includes oxidative stress, PSII damage, DNA damage, and apoptosis as underlying
processes (Lesser et al., 1990; Gates et al., 1992; Lesser, 1996, 1997; Warner et al.,
1999; Lesser and Farrell, in press). This model is consistent with a variety of biomarker
proteins expressed in corals during thermal stress (Brown et al., 2002; Downs et al., 2000,
2002).
Damage to photosystem II (PSII) reaction centers in the zooxanthellae, specifically at
the D1 protein of PSII, following exposure to elevated temperatures and solar radiation, is
believed to be an important factor leading to the bleaching of corals (Iglesias-Prieto et al.,
1992; Lesser, 1996; Warner et al., 1996, 1999), and caused by ROS (Lesser, 1997; Richter
et al., 1990). Damage or impairment of PSII function is easily detected using nondestruc-
tive active chlorophyll fluorescence techniques (Brown et al., 1999; Jones et al., 1998;
Gorbunov et al., 2001; Warner et al., 1999; Winters et al., 2003). Instruments have been
developed that incorporate protocols to measure the multiple photochemical turnover
[pulse amplitude-modulated (PAM)], and single photochemical turnover of PSII [fast
repetition rate (FRR)] in the laboratory and in the field (Schreiber et al., 1986; Gorbunov et
al., 2000). These instruments measure, nondestructively, fluorescent transients that provide
information on the efficiency of PSII and can discern chronic photoinhibition from
dynamic photoinhibition, the former representing damage to PSII and the latter a
protective regulatory response of the photosynthetic apparatus. The underwater FRR has
been used to examine diel cycling and dynamic versus chronic photoinhibition of corals in
shallow and deep waters (Gorbunov et al., 2001; Lesser and Gorbunov, 2001). One
advantage of the FRR versus the PAM instrument is that because of the protocol used to
measure fluorescent transients, a series of flashlets that saturate PSII in microseconds (Fig.
6a), this instrument can also measure the optical cross section of PSII which is a valuable
parameter for discerning dynamic versus chronic photoinhibition (Gorbunov et al., 2000).
An underwater version of the PAM instrument is commercially available and is capable of
examining the photoacclimatization state of corals by measuring the relationship between
electron transport rates and irradiance (Fig. 6b) which can be interpreted, with caution, to
traditional photosynthesis versus irradiance (P– I) curves. Using these data and non-linear
fitting techniques the user can then can fit curves to the data and calculate photosynthetic
parameters. The instrument has also been widely used to study diel changes in the
quantum yield of PSII fluorescence and its relationship to differences between photo-
chemical and non-photochemical quenching, or dynamic photoinhibition (Brown et al.,
1999; Hoegh-Guldberg and Jones, 1999). Brown et al. (1999) suggested that the diurnal
patterns in quantum yields of PSII and xanthophyll cycling they observed were suggestive
of photoinhibition followed by photoprotection. Hoegh-Guldberg and Jones (1999) and
Jones et al. (1998) observed similar patterns and suggested that sink limitations were also

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252230

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252 231
important in regulating the quantum yields of PSII. Warner et al. (1999) and Lesser and
Farrell (in press) have correlated changes in PSII fluorescence with changes in the
concentration of D1 protein during exposure to thermal stress and/or solar radiation.
Other models of thermally induced bleaching have suggested that the dark reactions of
photosynthesis are affected initially, leading to sink limitation, overreduction of photo-
synthetic electron transport, oxidative stress, and damage to PSII (Jones et al., 1998).
From the available data, Lesser and Farrell (in press) have proposed a model of
bleaching induced by damage to PSII that incorporates simultaneous damage to both
photochemistry and carbon fixation in a feedback loop that greatly enhances the damage to
PSII. There is evidence that PSII is already affected directly by high solar radiation without
any thermal stress (Gorbunov et al., 2001) and that thermal stress without high solar
irradiances affects PSII (Warner et al., 1999). Additionally, high levels of ROS are a
consistent feature of coral physiology, especially in the presence of thermal stress and UVR
in both the symbiont and host (Lesser, 1996, 1997; Lesser and Farrell, in press). Reactive
oxygen species, especially hydrogen peroxide, are well-known inhibitors of Rubisco
(Asada and Takahashi, 1987) and cause damage to PSII (Asada and Takahashi, 1987;
Richter et al., 1990). The work by Jones et al. (1998) illustrated the importance of carbon
sink limitation in exacerbating this damage. This observation is significant because carbon
limitation has been observed in shallow water corals (Muscatine et al., 1989), and can be
significantly effected by water flow (Lesser et al., 1994). Damage to PSII and sink
limitation under these conditions are likely to be occurring simultaneously with thermal
stress with the resulting formation of ROS which overwhelms all of the host and algal
antioxidant defense systems. The cascade of events that ultimately induces the expulsion of
zooxanthellae from their host could include the accumulation of ROS and damage to PSII
or a decrease in the amount of translocated photosynthate, or both (Lesser et al., 1990).
The host also responds to thermal stress. In particular, heat shock proteins (HSPs) are
up-regulated in response to thermal stress (Black et al., 1995; Fang et al., 1997; Sharp et
al., 1997). Heat shock proteins are inducible by a number of environmental factors and
appear to be a generalized stress response that is evolutionarily conserved. Under stressful
conditions, HSPs interact with proteins to maintain their conformation and function or in
targeting damaged proteins for degradation. This function is also consistent with patterns
of expression in markers of protein degradation observed in corals (Downs et al., 2000,
2002). Studies on the effect of UVR and thermal stress on corals have also shown
significant DNA damage in host tissues upon exposure to UVR (Anderson et al., 2001)
and thermal stress combined with exposure to solar radiation (Lesser and Farrell, in press).
DNA damage can lead to apoptosis or programmed cell death if not repaired. One of the
key cell cycle genes activated after DNA damage is p53. If DNA repair is not possible,
then p53-mediated apoptosis may be initiated. The expression pattern of a putative p53
protein in Montastraea faveolata after exposure to thermal stress and high irradiances of
Fig. 6. (a) Fast repetition rate fluorometer (FRRF) measurements on the coral M. faveolata from 1–2 m at Lee
Stocking Island, Bahamas. Note difference between dark adapted (Night) and steady state (Day) quantum yield
measurements of PSII fluorescence. Lesser, unpublished data. (b) Pulse amplitude-modulated (PAM) rapid light
curves on M. faveolata from different depths at Carrie Bow, Cay, Belize. Note the sensitivity of electron transport
rates (ETR) and depression of maximum rates at higher irradiances with depth as observed with photosynthesis–
irradiance ( P versus I ) curves. Lesser, unpublished data.
M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252232
solar radiation was consistent with the observed pattern of DNA damage (Lesser and
Farrell, in press).
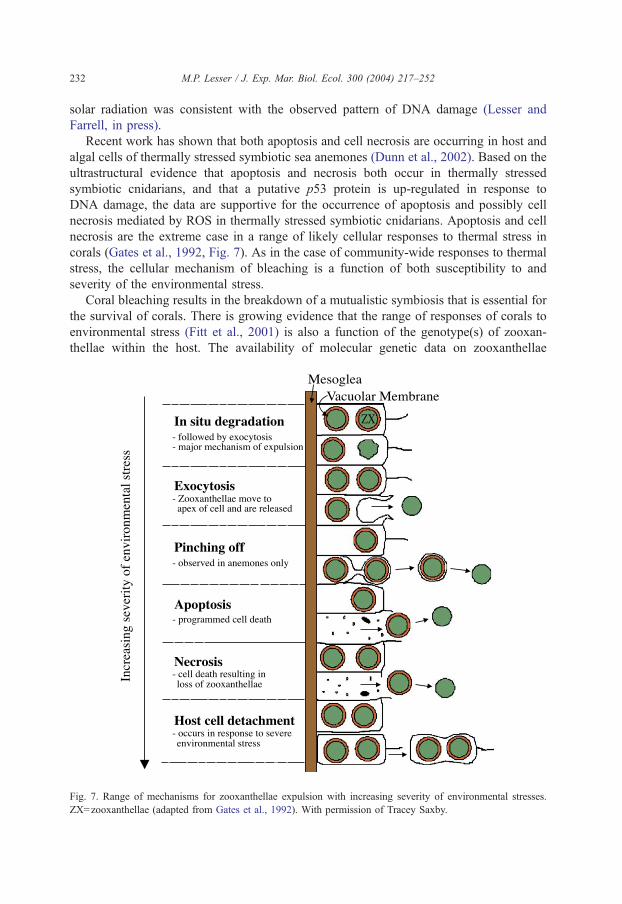
Recent work has shown that both apoptosis and cell necrosis are occurring in host and
algal cells of thermally stressed symbiotic sea anemones (Dunn et al., 2002). Based on the
ultrastructural evidence that apoptosis and necrosis both occur in thermally stressed
symbiotic cnidarians, and that a putative p53 protein is up-regulated in response to
DNA damage, the data are supportive for the occurrence of apoptosis and possibly cell
necrosis mediated by ROS in thermally stressed symbiotic cnidarians. Apoptosis and cell
necrosis are the extreme case in a range of likely cellular responses to thermal stress in
corals (Gates et al., 1992, Fig. 7). As in the case of community-wide responses to thermal
stress, the cellular mechanism of bleaching is a function of both susceptibility to and
severity of the environmental stress.
Coral bleaching results in the breakdown of a mutualistic symbiosis that is essential for
the survival of corals. There is growing evidence that the range of responses of corals to
environmental stress (Fitt et al., 2001) is also a function of the genotype(s) of zooxan-
thellae within the host. The availability of molecular genetic data on zooxanthellae
Fig. 7. Range of mechanisms for zooxanthellae expulsion with increasing severity of environmental stresses.
ZX=zooxanthellae (adapted from Gates et al., 1992). With permission of Tracey Saxby.

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252 233
genotypes (see below) and their micro- and macroscale distributions will very likely play a
significant role in who are the winners and losers under any continuing scenario of global
climate change.
4. Coral symbiont systematics
Until recently most symbiotic dinoflagellates were considered to be members of a
single pandemic species, Symbiodinium microadriaticum. Early work by Schoenberg and
Trench (1980a,b) showed clear physiological and biochemical differences between many
cultured zooxanthellae from different hosts which were characteristics of different species,
and culminating in work by Blank and Trench (1985) and Trench and Blank (1987) which
used many independent lines of evidence to show that several different species of
zooxanthellae do exist and that there were probably more undescribed species in this
genus. Rowan and Powers (1991a,b) then used molecular genetic tools, restriction
fragment length polymorphisms (RFLPs) of the small ribosomal subunit (ssRNA) and
sequencing of ssRNA, to show that the zooxanthellae of reef-building corals and other
symbiotic invertebrates are a highly diverse group of organisms organized at that time into
three major ‘‘clades’’; A, B, and C. Additionally, many corals contain as many as two or
three species per host that appear to be ecologically segregated based on small-scale
gradients in their physical environment (e.g., light) (Rowan and Knowlton, 1995; Rowan
et al., 1997). Additional work using RFLP genotypes using ssRNA and large subunit
ribosomal RNA (lsRNA) (Baker and Rowan, 1997), chloroplast 23S-rDNA sequencing
(Santos et al., 2002), and sequencing of the internal transcribed spacer regions (ITS)
(LaJeunesse, 2001) have all provided a wealth of information on the diversity of this genus
(Baker, 2003) and its seven clades (A–G) along with host diversity (Fig. 8a) to the point
where we can now begin to compare within and between regional diversity over large
spatial scales (LaJeuness, 2002; LaJeuness et al., 2003).
More interesting experimental work remains to be done on whether all of this genetic
diversity translates into physiological diversity and the ability to tolerate various types of
environmental stress. Different clades have already been assigned different functional
groupings (e.g., stress-tolerant generalists and narrowly adapted specialists) based on
latitudinal or bathymetric gradients (Fig. 8b) in clade type within a single host species
(Rodriguez-Lanetty et al., 2001; LaJeuness, 2002), on small-scale spatial patterns of
multiple genotypes within a single coral (Rowan and Knowlton, 1995; Toller et al.,
2001a), or on repopulation studies of bleached corals (Rowan et al., 1997; Toller et al.,
2001b). Much of the significance of these studies has been attributed to putative
physiological differences between clades. While simultaneous works on genetics and
temperature tolerances have revealed some differences (Kinzie et al., 2001; LaJeuness et
al., 2003), other works on general photosynthetic capabilities have shown less difference
between clades (Savage et al., 2002).
This brings us to the vigorously debated issue of the Adaptive Bleaching Hypothesis
(ABH) originally formulated by Buddemeier and Fautin (1993). While environmental
stress can reduce fitness, it is believed that under certain conditions the same stress can
result in positive, directional, selection for phenotypes that arose from the generation and

Fig. 8. (a) Cladogram of currently known groups of zooxanthellae (Symbiodinium sp.) based on ITS 2 sequence
data and associated hosts. Numbers at branch nodes are boot strap values. (b) Community structure of holobionts
and distribution of Symbiodinium sp. Types as determined by ITS 2 sequencing at Puerto Moreles, Mexico
(20j50VN, 86j52VW). Symbiont types are compared to the number of host species associated with that type at
each depth. With permission from T. LaJeuness and Marine Biology (141: 387–400, 2002).
M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252234

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252 235
maintenance of variability stimulated by that stress (Hoffmann and Hercus, 2000). Other
studies have shown generally that significant constraints to adaptive evolution in response
to global warming exist (Etterson and Shaw, 2001). The ABH contends that after bleaching
occurs, the ‘‘shuffling’’ of zooxanthellae genotypes, from the large number currently
available and presumably representing a range of thermal tolerances, is possible and
adaptive in an ecological and evolutionary sense for the holobiont (Buddemeier and Fautin,
1993; Ware et al., 1996). It is believed that corals bleach and can survive future episodes of
thermal stress by allowing zooxanthellae genotypes with greater thermal tolerances that
were not initially expelled to become the new dominant genotype residing in the host
tissues (Baker, 2001, 2003). Several of the assumptions of the ABH have been experi-
mentally tested on a limited number of zooxanthellae cultures in terms of thermal tolerance
and shown to be related to their placement in specific clades as discussed above and would
therefore appear to be consistent with the ABH (Kinzie et al., 2001). Recently, Toller et al.
(2001b) showed that Montastraea sp. corals experimentally bleached and placed back in
the field were repopulated with zooxanthellae from different clades. However, the same
occurred for field populations that had not bleached, suggesting that thermal stress, and
subsequent bleaching, is not the only mechanism for symbiont ‘‘shuffling’’.
As the number and severity of bleaching events increases, the percent mortality of
corals has increased significantly (Hoegh-Guldberg, 1999), and the change in temperature
required for bleaching in many species of coral, which is already small (Jokiel and Coles,
1990), will be exceeded for most species of corals. Under these conditions, any
catastrophic mortality that would occur might be perceived as a limitation for the
opportunity of zooxanthellae ‘‘shuffling’’ consistent with the ABH. While most critiques
of the ABH have suggested that there is little evidence in the last 20 years that bleached
corals (i.e., the holobiont) have adapted to thermal stress, there is some evidence that
suggests otherwise. A prediction of the ABH is that corals that have recovered from prior
bleaching should be more resistant to subsequent thermal stress because of a change in
their symbiont composition compared to corals that have not experienced bleaching. In the
tropical far eastern Pacific, severe coral bleaching during the 1982–1983 El Nino resulted
in coral mortality of 50–90% (Glynn, 1988). The survivors of this event avoided
significant bleaching or mortality during the 1997–1998 El Nino even though the
temperature anomalies were as high in 1997–1998 as they were in 1982–1983,
presumably as a result of changes in zooxanthellae genotype (Glynn et al., 2001). The
corals in the eastern Pacific are now reverting back to their original, temperature-sensitive,
genotype (Baker, personal communication). This potentially adds another complication
(i.e., time dependence of genotype stability) to the ABH. It may be that only corals
experiencing chronically high temperatures (e.g., Arabian Gulf) are able to maintain
zooxanthellae genotypes that exhibit greater tolerances to thermal stress.
The ABH provides an experimental framework to examine interesting questions about
the stress response of corals and their symbionts. A question consistently raised is whether
we are now dealing with time scales and rates of environmental change where the results
of testing the ABH would be useful in the context of large-scale changes on coral reefs.
There is no agreement on this currently; however, producing new experimental work on
the thermal tolerances, genetic diversity, and infection capabilities of zooxanthellae is
essential at this time. We now have the molecular genetic tools to identify and follow

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252236
zooxanthellae populations over space and time, but little knowledge about the range of
environmental tolerances of those symbionts.
5. Coral host systematics
Presently there are approximately 1300 species of scleractinian corals (Veron, 1995,
2000) that have been identified primarily by morphological characters (e.g., corallite
structure). Molecular data are now being applied routinely to answer questions regarding
taxonomic and evolutionary questions on corals. Molecular markers such as ribosomal
RNA, the internal transcribed region (ITS), or amplified fragment-length polymorphisms
(AFLP) have been used to answer interesting questions on systematics, presence of
sibling species, and hybridization. Recent molecular and morphological studies have
clearly placed the Anthozoa as the basal class within the phylum Cnidaria (Bridge et al.,
1995) with the Scleractinia embedded within the Class Anthozoa and probably evolved
from the Corallimorpharia (Fautin and Lowenstein, 1992; Stanley and Fautin, 2001).
Zooxanthellate scleractinian corals have been present since the mid-Triassic (Veron,
1995) and based on mitochondrial 16S rDNA, two distinct clades of scleractinian
families appear to have diverged before the appearance of modern scleractinian taxa in
the fossil record (Romano and Palumbi, 1996; Romano and Cairns, 2000). These results
do not support traditional morphological systematics, but instead a polyphyletic origin of
the scleractinian skeleton. While understanding the evolution of these higher taxonomic
levels is problematic, the identification of coral ‘‘species’’ is no less troublesome. For
several prominent and ecologically important species there are now several different lines
of evidence that support the presence of cryptic or sibling species. The most prominent
example is from the coral Montastrea annularis, which exhibits a high degree of
morphological variation that was ascribed to phenotypic plasticity in response to solar
irradiance or other abiotic factors. These sympatric morphotypes are now recognized as
at least three closely related species (Knowlton et al., 1992; Weil and Knowlton, 1994)
known as Montastraea annularis, M. faveolata, and M. franksi. While character analysis
and assessments of reproductive isolation have generally supported the existence of these
sibling species (Knowlton et al., 1997), it is also known that hybrid larvae can be
produced and that intermediate coral morphologies exist in the field (Szmant et al.,
1997). Molecular analyses have been equivocal with an analysis using AFLPs and
microsattelites supporting reproductive isolation and therefore some degree of divergence
(Lopez et al., 1999), and sequencing of the ITS region and mitochondrial cytochrome
oxidase (COI) showing no support for distinct species (Medina et al., 1999). Other
groups of closely related morphotypes present similar questions regarding plasticity
versus sibling species. One explanation for the speciose nature and morphological
variability of certain groups is ‘‘reticulate evolution,’’ where repeated episodes of
hybridization and fusion of lineages occur over evolutionary time leading to variants
whose species boundaries become ‘‘fuzzy’’ (Veron, 1995). Recent molecular analyses
(i.e., ITS sequences) on several species in the genus Madracis from the Caribbean have
shown high levels of intraspecific and intra-individual variability (Diekmann et al.,
2001). Based on these data, and paleontological data, the most parsimonious interpre-

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252 237
tation for these results was that reticulate evolution was the dominant mechanism
responsible for the observed variability. Probably the best example of reticulate evolution
in corals comes from the genus Acropora. In the Caribbean, three species of Acropora
exist; Acropora cervicornis, Acropora palmata, and Acropora prolifera. A. prolifera has
been described as an intermediate morphology and a hybrid between A. cervicornis and
A. palmata. Recent nuclear, mitochondrial, and ribosomal sequence data have supported
the hypothesis that A. prolifera is a hybrid and that the hybrids are long-lived and
propagate principally by asexual reproduction (Van Oppen et al., 2000; Vollmer and
Palumbi, 2002).
In the coming years we will see the next wave of molecular influence on coral biology
with the initiation of several coral genome projects. The case for a cnidarian genomics
database has been made (Ryan and Finnerty, 2003), and progress has been made on an
EST library for Acropora millepora (Kortschak, 2003) which has already revealed a
surprising number of conserved genes previously believed to have arisen during the
evolution of vertebrates. Which coral(s) to use as subjects for genomic sequencing, and
proteonomics as well, is under debate but the benefits for evolutionary, physiological, and
ecological studies on corals will be significant. The development of coral-specific micro-
and macroarrays for stress related markers, the continued identification of developmental
and cell cycle genes homologous with higher taxa in these tissue-grade diploblastic
animals, and DNA profiling to characterize genetic differences between individuals and
populations will benefit tremendously from the completion of one or more coral genome
projects (Ball et al., 2002; Gibson, 2002).
6. Fluorescent proteins
Recently, there has been a flurry of activity surrounding the identification of host
fluorescence (Kawaguti, 1969) in corals and other cnidarians as a group of homologous
fluorescent proteins related to green fluorescent protein (GFP; Fig. 9a and b; Matz et al.,
1999; Salih et al., 2000; Dove et al., 2001; Mazel et al., 2003) originally isolated and
described from the hydromedusae, Aequorea victoria (Tsein, 1998). The 238-amino-acid
protein, within which three residues at positions 65–67 form the active chromophore, is
extremely resistant to extremes in pH and temperature and requires the presence of oxygen
as it is translated and folds into its native configuration (Tsein, 1998). It requires no
cofactors for fluorescence, is not coupled to a bioluminescent (e.g., luciferin-luciferase)
system in corals, and in corals these fluorescent proteins are located principally in the
epithelial cells of the cnidarian host (Salih et al., 2000; Mazel et al., 2003), although they
can be observed in gastrodermal tissue as well (Salih et al., 2000). While considerable
interest in the utility of coral fluorescent proteins as transcription reporter genes has
dominated the literature, little is known about the function of this protein in corals. It has
been proposed that these fluorescent proteins provide photoprotection under high-light
conditions (Kawaguti, 1969; Salih et al., 2000), enhance photosynthesis under low-light
conditions (Salih et al., 2000), or both depending on the position of the fluorescent
pigment relative to the zooxanthellae (Salih et al., 2000; Dove et al., 2001). The
fluorescence can also contribute significantly to the spectral signature (e.g., reflectance)

Fig. 9. (a) Underwater photograph of Montastraea cavernosa taken under white light at Lee Stocking Island,
Bahamas. (b) Same coral taken under blue light excitation showing the dominant green light emission of a green
fluorescent protein in the epithelial cells of this coral (Mazel et al., 2003). Photographs by C. Mazel.
M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252238
of corals under daylight illumination (Dove et al., 2001; Mazel and Fuchs, 2003) and
therefore the perceived color of corals by the human eye.
Many fluorescent proteins, based on fluorescence emission spectra and molecular data,
have been identified in corals (Mazel, 1995; Matz et al., 1999; Dove et al., 2001; Labas et
al., 2002; Kelmanson and Matz, 2003; Mazel et al., 2003), and recent data suggests they
may have arisen through gene duplications at several loci (Kelmanson and Matz, 2003).
Of the proposed functions described above, recent data have clearly shown that there is no
role for fluorescent proteins in enhancing photosynthesis under low-light conditions
(Gilmore et al., 2003; Mazel et al., 2003). Fluorescence resonance energy transfer (FRET)
between fluorescent proteins occurs but transfer from fluorescent proteins to chlorophyll

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252 239
does not occur in corals. Additionally, for the Caribbean coral M. faveolata, there is no
evidence that fluorescent proteins protect the holobiont from the deleterious effects of
UVR (Mazel et al., 2003). While new fluorescent proteins continue to be discovered,
understanding their role in coral physiology and ecology remains elusive.
7. Coral diseases
One of the most significant changes on coral reefs along the Florida Keys Reef tract and
in the Caribbean generally has been the emergence of diseases and the potential
relationship to global climate change (Richardson, 1998; Rosenberg and Ben-Haim,
2002; Harvell et al., 2002). While coral bleaching is most commonly associated with
thermal stress and its physiological consequences, bleaching in at least one species of
coral, Oculina patagonica, is caused by the bacterium, Vibrio shiloi, subsequent to thermal
stress (Kushmaro et al., 1996). Additionally, the mortality of gorgonian corals, caused by a
fungal pathogen, Aspergillus sydowii, has been linked to transatlantic dust transport from
Africa that has increased in the last 25 years due to desertification (Shinn et al., 2000).
For many years the dominant, and ecologically significant, disease of corals was black
band disease (Rutzler et al., 1983; Kuta and Richardson, 1996), which is now known to be
a microbial consortium of cyanobacteria and sulfide-oxidizing bacteria that exploit
gradients of oxygen and sulfide while simultaneously producing anoxic zones that kill
the coral tissue (Richardson, 1998). Acropora cervicornis and A. palmata were once the
dominant species across the Caribbean including the Florida reef tract. In the 1980s,
however, disease (white band disease) resulted in almost the complete mortality of these
species changing the community structure of these reefs for the foreseeable future. White
band disease is now recognized as two variants of the same disease, type I and type II, with
the etiological agent believed to be a gram-negative bacterium (Richardson, 1998). Plague
or ‘‘white plague’’ is another disease of corals that also has two variants with plague type
II emerging in the mid-1990s and affecting mostly a single species of Caribbean coral,
Dichocoenia stokesi. The etiological agent for this disease has been identified as a new
species of Sphingomonas, a gram-negative bacterium (Richardson, 1998; Richardson et
al., 1998).
In the 1990s it appears that a suite of new coral diseases has emerged. Some of these
diseases are associated with elevated nutrients, either from agricultural runoff or from
human sewage. Recent evidence has shown that an enteric bacterium, Serratia marces-
cens, is responsible for the ‘‘white pox’’ disease of A. palmata (Patterson et al., 2002).
Continued development along coastal waterways paired with preexisting dwellings has
resulted in an enormous discharge of human sewage. While in many instances sewage is
first treated prior to discharge, evidence has demonstrated that not all of the infectious
organisms present in sewage are inactivated prior to their discharge. Several investigators
have noted alarming amounts of pathogenic organisms in and around the Florida canals
and around the Florida Keys (Lipp et al., 2002). Two issues have frustrated managers and
scientists interested in the study of coral diseases; nomenclature and a common set of
symptoms associated with a particular disease name, and isolation, identification, and
reinfection studies on the putative causative agents of these diseases. Fulfilling Koch’s

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252240
postulates has been a central tenant of studies with pathogenic microorganisms for over a
century and only the bacteria associated with black-band, white plague II, and white pox
coral diseases have fulfilled the requirements to be identified as the causative agent with a
high degree of confidence. An additional problem for these studies is culturability. Many
marine prokaryotes remain non-culturable unlike the majority of pathogens in clinical,
wildlife, or veterinarian settings. Molecular techniques such as quantitative PCR and
fluorescent in situ hybridization (FISH; Bythell et al., 2002) and others should provide
excellent proxies for the culturing usually required for fulfilling Koch’s postulates.
8. Remote sensing
Our understanding of coral reef ecology is still hampered by the inability to map and
monitor large expanses of reef area over any reasonable temporal scale. One way to assess
changes in the aerial coverage of coral reefs on large spatial and temporal scales is using
remote sensing imagery taken from airplanes or satellite (Fig. 10) platforms (Green et al.,
1996; Mumby et al., 1997). Several practical and analytical hurdles remain to be solved
before this approach can be widely used. There is a practical reason to examine reefs as
they relate to shallow water habitats and remote sensing. Coral reefs are generally found in
optically clear, Case I waters. Although chlorophyll concentrations of the waters over reefs
are typically 0.1 to 0.2 mg Chl a m�3, typical of open ocean waters, there is significant
absorption in the blue wavelengths because of benthic-derived CDOM (from corals and
Fig. 10. 1998 SeaWifs image of Florida, Florida Straights, and the Bahamas including the Bahama Banks and the
Tongue of the Ocean. (NASA archives).

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252 241
sea grasses), and there is often extra scattering due to mineral particles derived from water
column precipitation of aragonite or resuspension of sediments. An optical approach to
monitoring coral reefs should include an understanding of the underlying reasons for
changes in the optical signal(s) of choice and whether one can generalize these
mechanisms from reef to reef. One goal of this approach is to obtain optical closure,
assessing which photons are absorbed, reflected, or re-emitted as fluorescence, essentially
a photon budget. Areas of high absorbance reflect the presence of primary producers
containing photosynthetic and accessory pigments whereas areas of low absorbance or
high reflectance indicate areas of low coral or algal cover. Another product of an optical
approach is to utilize reflectance or fluorescence signatures of the benthic community to
establish a reef classification scheme that can then be used with remote sensing imagery
(Mazel, 1995; Hochberg and Atkinson, 2000). Coral reefs therefore provide an excellent
test of our abilities to extract both water column optical and benthic properties of reefs
from remote sensing reflectance data.
Up to this point, when we compare traditional classification schemes (e.g., SCUBA
transects) to remote sensing classifications, the range of agreement between the two
methods is anywhere from 30% to 85%. This is not very satisfying at the moment but new
sensors and analytical approaches continue to be developed, as this is a high priority area
for managers and ecologists. Most remote sensing approaches still require a significant
amount of sea truthing over large spatial and temporal scales to validate the use of the
imagery and analytical routines. Several investigators have constructed irradiance reflec-
tance libraries of various functional groups (e.g., corals, macrophytes, seagrasses,
microbial mats) that span the possible pigment compositions observed on a reef and
can be used to develop algorithms (Hochberg and Atkinson, 2000; Hedley and Mumby,
2003b). Additionally, not all images are optimal for use in classification schemes. One
consistent problem is sea surface glint. Because of consistent wind patterns, sea surface
glint from wavelets often introduces errors in the water leaving radiances or remote
sensing reflectances. Routines to remove sea surface glint from remote sensing imagery
have recently been developed (Hochberg et al., 2003) and should contribute to more
accurate benthic classifications.
There has been significant interest in coral reef mapping using multispectral platforms
such as Landsat 7 and Ikonos imagery. The advantage of these satellite platforms is that
they provide global synoptic coverage of coral reefs anywhere from once to several times
annually and there is already a large database that has recently, and continues to be,
collected as part of the Long Term Acquisition Program (LTAP). There are presently
significant limitations in using these data sets for detailed bottom classifications but
progress has been made using multispectral data from coral reef environments. A
comparative study using the compact airborne spectrographic imager (CASI) showed that
CASI consistently outperformed satellite sensors (e.g. Landsat, SPOT) and aerial photog-
raphy in classifying bottom features (e.g., live corals, sand, seagrass) (Mumby et al., 1997,
2000). Recently, a radiative transfer approach and multispectral imagery was used on reefs
in Hawaii down to a depth of 25 m with accuracy exceeding 85% (Isoun et al., 2003).
Hyperspectral remote sensing imagery from airborne (e.g., CASI) and satellite (e.g.,
Hyperion) platforms holds the promise of providing the detailed information required for
remotely mapping coral reefs at 1-m2 resolution and consistent capabilities to discriminate

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252242
between functional end-members such as macroalgae and corals. Additionally, hyper-
spectral imaging spectrometers are viewed as a potentially important tool for the
assessment and management of tropical coastal resources (Green et al., 1996). New and
novel analytical approaches will be needed to take advantage of the additional information
contained within hyperspectral imagery. One of the disadvantages of current algorithms is
that because of the spectral mixing that occurs on certain types of reefs (e.g., Caribbean),
spectral, and therefore classification, information is lost during conventional supervised
routines. This translates into pixel to pixel errors because current algorithms are con-
strained to one pixel-one end-member classifications. Recognizing this problem has
resulted in new approaches of analysis and classification. Hochberg and Atkinson
(2000) and Andrefouet et al. (2003) have used derivative analysis coupled with linear
discriminant function analysis to analyze hyperspectral data for coral reefs in Hawaii and
French Polynesia. Hedley and Mumby (2003a,b) have developed a mathematical approach
that resolves both depth and subpixel spectral composition which is suitable for hyper-
spectral imagery. Another method of spectral classification uses a ‘‘lookup table’’ (LUT)
approach where a library of end-member remote sensing reflectances was generated from
radiative transfer computations (Louchard et al., 2003). An initial use of this approach was
applied to shallow waters (<5 m) in the Bahamas with 60–80% accuracy in classification
and 83% accuracy in bathymetry (Lochard et al., 2003). Lastly, multiple spectral
signatures in a single pixel can be resolved using a blend of different algorithms in a
‘‘Fuzzy’’ logic classification scheme. Fuzzy logic classifications allow for multiple end-
members to be recognized in an individual pixel. This type of classification scheme should
reduce errors associated with one pixel-one end-member algorithms and better reflect the
heterogeneous nature of coral reef habitats on small scales. Fuzzy classification schemes
have been used for ocean color data (Moore et al., 2001) and on coral reefs (Andrefouet et
al., 2000; Andrefouet et al., 2003). In the future, both multispectral and hyperspectral
imagery will require the development of algorithms capable of consistently extracting
accurate bottom classifications from remote sensing reflectance (Rrs) or extracting enough
optical information from the original image to effectively ‘‘remove the water’’ to be able to
use reflectance libraries of different end-members.
9. Future exploration and experimentation
Despite the voluminous amount of data on the biology and ecology of shallow water
reef systems, there are significant gaps in our knowledge about coral reef communities
beyond the 30-m depth of most studies. Deep fore reef communities have largely escaped
the effects of global climate change as it relates to ultraviolet radiation and global
warming. Because of the close proximity of near-shore fringing reefs to human popula-
tions and their relatively shallow depths (5 to 30 m), these reefs are most susceptible to
harmful human activities (sedimentation, nutrient enrichment, physical damage, over-
fishing) but also to the effects of natural disturbances (storm wave damage, high sea
surface temperatures, high irradiance). The mid-shelf coral reefs, either fringing reefs
associated with the offshore cays or nonemergent linear reefs, are less susceptible to
human induced stresses as described above, but are exposed to similar natural impacts due

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252 243
to their comparable depths (5 to 30 m). The deep reefs (>30 m) are largely free from
human-induced stresses (excluding fishing and anchoring) and natural impacts due to their
greater distance from human populations and their greater depth, respectively. There is a
major gap in our overall understanding of coral reef communities and our lack of
understanding about deep reef environments, and in particular sponge and coral popula-
tions, has been keeping us from realizing an untapped resource of bioactive compounds,
understanding the ecology and biodiversity of deep reef communities, and assessing
whether deep reef communities are a potential source of larvae for shallow reef
communities. Most studies on coral reefs have focused on shallow reef (<30 m) systems
because of the technical limitations of conducting studies deeper than 30 m and the
expense of using submersible technology in relatively shallow depths (<150 m). The
technical limitations are slowly being overcome by the introduction of mixed gas technical
diving, both open-circuit SCUBA and closed-circuit rebreathers, to the scientific diving
community. A better understanding of deep reef ecology and biodiversity is critical if we
are to establish criteria that characterize the ‘‘health’’ of coral reefs and formulate
management plans in response to anthropogenic and natural disturbances.
Additionally, many cold, deep coral reefs composed of azooxanthellate taxa have
recently been discovered in deep continental shelf habitats. These communities appear to
be extremely fragile and may provide important three-dimensional habitat for juvenile fish
of commercial importance. In several locations these reefs have been severely damaged by
fish trawling activities, prompting fishing closures and proposals for marine protected
areas which include these reefs. These two unique communities, deep fore reef and deep
and cold reefs, will require experimental studies to understand their trophic and
reproductive biology, two critical life history traits which will help guide managers as
they attempt to formulate strategies for protecting these unique environments.
10. Conclusions
In closing, I want to restate that this review is not exhaustive and reflects the authors
bias of what represents the latest developments in the area of experimental biology of coral
reef ecosystems. It is essential for the coral reef community to work together for the
common good of the ecosystem. Time is short, as are finances, to conduct the integrative
studies required to understand the range of acclimative capabilities that the holobiont has
in the face of continued environmental change, and to potentially predict what reefs will
remain, and what will they look like in the future.
Acknowledgements
The author wishes to thank the editors of JEMBE for the invitation to write this review.
Support for the author’s work has been provided by NSF (Biological Oceanography),
ONR (Environmental Optics), NOAA (National Undersea Research Program and Ocean
Exploration), UNESCO, and the Smithsonian Institution (Caribbean Coral Reef
Ecosystems). Photographs or figures were generously provided by Charles Mazel, Todd

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252244
LaJeuness, NASA, Tracey Saxby, Ray Berkelmans, and Ove Hoegh-Huldberg. A special
thank you to Bill Fitt whose comments greatly improved this manuscript. [SS]
References
Anderson, S., Zepp, R., Machula, J., Santavy, D., Hansen, L., Mueller, D., 2001. Indicators of UV exposure
in corals and their relevance to global climate change and coral bleaching. Hum. Ecol. Risk Asses. 7,
1271–1282.
Andrefouet, S., Roux, L., Chancerelle, Y., Bonneville, A., 2000. A fuzzypossibilistic scheme of study for objects
with indeterminate boundaries: applications to French Polynesian reefscapes. IEEE Trans. Geosci. Remote
Sens. 38, 257–270.
Andrefouet, S., Payri, C., Hochberg, E.J., Che, L.M., Atkinson, M.J., 2003. Airborne hyperspectral detection of
microbial mat pigmetation in Rangiroa atoll (French Polynesia). Limnol. Oceanogr. 48, 426–430.
Aronson, R.B., Precht, W.F., 2000. Herbivory and dynamics on the coral reef at Discovery Bay, Jamaica. Limnol.
Oceanogr. 45, 251–255.
Aronson, R.B., Precht, W.F., Macintyre, I.G., Murdoch, T.J.T., 2000. Coral bleach-out in Belize. Nature 405,
36.
Aronson, R.B., Precht, W.F., Toscano, M.A., Koltes, K.H., 2002. The 998 bleaching event and its aftermath on a
coral reef in Belize. Mar. Biol. 141, 435–447.
Asada, K., Takahashi, M., 1987. Production and scavenging of active oxygen in photosynthesis. In: Kyle, D.J.,
Osmond, C.B., Arntzen, C.J. (Eds.), Photoinhibition. Elsevier, Amsterdam, pp. 228–287.
Baker, A.C., 2001. Reef corals bleach to survive. Nature 411, 765–766.
Baker, A.C., 2003. Flexibility and specificity in coral-algal symbiosis: diversity, ecology, and biogeography of
Symbiodinium. Ann. Rev. Ecolog. Syst. 34, 661–689.
Baker, A.C., Rowan, R., 1997. Diversity of symbiotic dinoflagellates (zooxanthellae) in scleractinian corals on
the Caribbean and eastern Pacific. Proc. 8th Int. Coral Reefs Sym. 2, 1301–1306.
Ball, E.E., Hayward, D.C., Reece-Hoyes, J.S., Hislop, N.R., Samuel, G., Saint, R., Harrison, P.L., Miller,
D.J., 2002. Coral development: from classical embryology to molecular control. Int. J. Dev. Biol. 46,
671–678.
Banaszak, A.T., Lesser, M.P., Kuffner, I.B., Ondrusek, M., 1998. Relationship between ultraviolet (UV) radiation
and mycosporine-like amino acids (MAAs) in marine organisms. Bull. Mar. Sci. 63, 617–628.
Black, N.A., Voellmy, R., Szmant, A.M., 1995. Heat shock protein induction in Montastraea faveolata and
Aiptasia pallida to elevated temperatures. Biol. Bull. 188, 234–240.
Blank, R.J., Trench, R.K., 1985. Speciation and symbiotic dinoflagellates. Science 229, 656–658.
Bridge, D., Cunningham, C.W., DeSalle, R., Buss, L.W., 1995. Class-level relationships in the Phylum Cnidaria:
molecular and morphological evidence. Mol. Biol. Evol. 12, 679–684.
Brown, B.E., 1997. Coral bleaching: causes and consequences. Coral Reefs 16, S129–S138 (Suppl.).
Brown, B.E., Ambarsari, I., Warner, M.E., Fitt, W.K., Dunne, R.P., Gibb, S.W., Cummings, D.G., 1999. Diurnal
changes in photochemical efficiency and xanthophylls concentrations in shallow water reef corals: evidence
for photoinhibition and photoprotection. Coral Reefs 18, 99–105.
Brown, B.E., Downs, C.A., Dunne, R.P., Gibbs, S.W., 2002. Exploring the basis of thermotolerance in the reef
coral Goniastrea aspera. Mar. Ecol. Prog. Ser. 242, 119–129.
Buddemeier, R.W., Fautin, D.G., 1993. Coral bleaching as an adaptive mechanism. Bioscience 43, 320–326.
Bythell, J.C., Barer, M.R., Cooney, R.P., Guest, J.R., O’Donell, A.G., Pantos, O., Le Tissier, M.D.A., 2002.
Histopathological methods for the investigation of microbial communities associated with disease lesions in
reef coral. Lett. Appl. Microbiol. 34, 359–364.
Carpenter, R.C., 1988. Mass mortality of a Caribbean sea urchin: immediate effects on community metabolism
and other herbivores. Proc. Natl. Acad. Sci. 85, 511–514.
Coles, S.L., Brown, B.E., 2003. Coral bleaching-capacity for acclimatization and adaptation. Adv. Mar. Biol. 46,
183–223.
Connell, J.H., 1978. Diversity of tropical rain forests and coral reefs. Science 199, 1081–1085.
Connell, J.H., 1997. Disturbance and recovery of coral assemblages. Coral Reefs 16, S101–S113 (Suppl.).

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252 245
Connell, J.H., Hughes, T.P., Wallace, C.C., 1997. A 30-year study of coral abundance, recruitment, and distur-
bance at several scales in space and time. Ecol. Monogr. 67, 461–488.
Crowley, T.J., 2000. Causes of climate change over the past 1000 years. Science 289, 270–277.
Cushman, J.H., Beattie, A.J., 1991. Mutualisms: assessing the benefits to hosts and visitors. Trends Ecol. Evol. 6,
193–195.
Cutchis, P., 1982. A formula for comparing annual damaging ultraviolet (DUV) radiation doses at tropical and
mid-latitude sites. In: Calkins, J. (Ed.), The Role of Solar Ultraviolet Radiation in Marine Ecosystems.
Plenum, New York, pp. 213–228.
Diekmann, O.E., Bak, R.P.M., Stam, W.T., Olsen, J.L., 2001. Molecular genetic evidence for the probable
reticulate speciation in the coral genus Madracis from a Caribbean fringing reef slope. Mar. Biol. 139,
221–233.
Done, T.J., 1995. Coral zonation: its nature and significance. In: Barnes, D.J. (Ed.), Perspectives on Coral Reefs
AIMS. Brian Clouston Publishers, Townsville, Australia, pp. 107–148.
Done, T.J., Ogden, J.C., Wiebe, W.J., Rosen, B.R., 1996. Biodiversity and ecosystem function of coral reefs.
In: Mooney, H.A., Cushman, J.H., Medine, E., Sala, O.E., Schulze, E.-D. (Eds.), Functional Roles of Biodi-
versity: A Global Perspective. Wiley.
Dove, S.G., Hoegh-Guldberg, O., Ranganathan, S., 2001. Major colour patterns of reef-building corals are due to
a family of GFP-like proteins. Coral Reefs 19, 197–204.
Downs, C.A., Mueller, E., Phillips, S., Fauth, J.E., Woodley, C.M., 2000. A molecular biomarker system
for assessing the health of coral (Montastraea faveolata) during heat stress. Mar. Biotechnol. 2,
533–544.
Downs, C.A., Fauth, J.E., Halas, J.C., Dustan, P., Bemiss, J., Woodley, C.M., 2002. Oxidative stress and seasonal
coral bleaching. Free Radic. Biol. Med. 33, 533–543.
Dunlap, W.C., Yamamoto, Y., 1995. Small-molecule antioxidants in marine organisms: antioxidant activity of
mycosporine-glycine. Comp. Biochem. Physiol. 112B, 106–114.
Dunlap, W.C., Chalker, B.E., Oliver, J.K., 1986. Bathymetric adaptations of reefbuilding corals at Davies
Reef, Great Barrier Reef, Australia: III. UV-B absorbing compounds. J. Exp. Mar. Biol. Ecol. 104,
239–248.
Dunn, S.R., Bythell, J.C., Le Tessier, D.A., Burnett, W.J., Thomason, J.C., 2002. Programmed cell death and
necrosis activity during hyperthermic stress-induced bleaching of the symbiotic sea anemone Aiptasia sp.
J. Exp. Mar. Biol. Ecol. 272, 29–53.
Dunne, R.P., Brown, B.E., 2001. The influence of solar radiation on bleaching of shallow water reef corals in the
Andaman Sea, 1993–1998. Coral Reefs 20, 201–210.
Dykens, J.A., 1984. Enzymic defenses against oxygen toxicity in marine cnidarians containing endosymbiotic
algae. Mar. Biol. Lett. 5, 291–301.
Dykens, J.A., Shick, J.M., 1982. Oxygen production by endosymbiotic algae controls superoxide dismutase
activity in their animal host. Nature 297, 579–580.
Dykens, J.A., Shick, J.M., 1984. Photobiology of the symbiotic sea anemone, Anthopleura elegantissima:
defenses against photodynamic effects, and seasonal photoacclimatization. Biol. Bull. 167, 683–697.
Dykens, J.A., Shick, J.M., Benoit, C., Buettner, G.R., Winston, G.W., 1992. Oxygen radical production in the sea
anemone Anthopleura elegantissima: and its symbiotic algae. J. Exp. Biol. 168, 219–241.
Edmunds, P.J., Carpenter, R.C., 2001. Recovery of Diadema antillarum reduces macroalgal cover and increases
abundance of juvenile corals on a Caribbean reef. Proc. Natl. Acad. Sci. 98, 5067–5071.
Etterson, J.R., Shaw, R.G., 2001. Constraint on adaptive evolution in response to global warming. Science 294,
151–154.
Fagoonee, I., Wilson, H.B., Hassell, M.P., Turner, J.R., 1999. The dynamics of zooxanthellae populations: a long-
term study in the field. Science 283, 843–845.
Falkowski, P.G., Dubinsky, Z., Muscatine, L., Porter, J.W., 1984. Light and the bioenergetics of a symbiotic coral.
Bioscience 34, 709–969.
Falkowski, P.G., Jokiel, P.L., Kinzie III, R.A. 1990. Irradiance and corals. In: Dubinsky, Z. (Ed.), Coral Reefs.
Elsevier, Amsterdam, pp. 89–107.
Fang, L., Huang, S., Lin, K., 1997. High temperature induces the synthesis of heat– shock proteins and the
elevation of intracellular calcium in the coral Acropora grandis. Coral Reefs 16, 127–131.

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252246
Fautin, D.G., Lowenstein, J.M., 1992. Phylogenetic relationships among scleractinians, actinians, and coralli-
morpharians (Coelenterata: Anthozoa). Proc. 7th Int. Coral Reefs Symp. 2, 665–670.
Fitt, W.K., Spero, H.J., Halas, J., White, M.W., Porter, J.W., 1993. Recovery of the coral Montastrea annularis in
the Florida Keys after the 1987 Caribbean ‘‘bleaching event’’. Coral Reefs 12, 57–64.
Fitt, W.K., McFarland, F.K., Warner, M.E., Chilcoat, G.C., 2000. Seasonal patterns of tissue biomass and
densities of symbiotic dinoflagellates in reef corals and relation to coral bleaching. Limnol. Oceanogr. 45,
677–685.
Fitt, W.K., Brown, B.E., Warner, M.E., Dunne, R.P., 2001. Coral bleaching: interpretation of thermal tolerance
limits and thermal thresholds in tropical corals. Coral Reefs 20, 51–65.
Frederick, J.E., Snell, H.E., Haywood, E.K., 1989. Solar ultraviolet radiation at the earth’s surface. Photochem.
Photobiol. 50, 443–450.
Fridovich, I., 1986. Biological effects of the superoxide radical. Arch. Biochem. Biophys. 247, 1–11.
Gardner, T.A., Cote, I.M., Gill, J.A., Grant, A., Watkinson, A.R., 2003. Longterm region-wide declines in
Caribbean corals. Science 301, 958–960.
Gates, R.D., Edmunds, P.J., 1999. The physiological mechanisms of acclimatization in tropical reef corals. Am.
Zool. 39, 30–43.
Gates, R.D., Baghdasarian, G., Muscatine, L., 1992. Temperature stress causes host cell detachment in symbiotic
cnidarians: implications for coral bleaching. Biol. Bull. 182, 324–332.
Gibson, G., 2002. Microarrays in ecology and evolution: a preview. Mol. Ecol. 11, 17–24.
Gilmore, A.M., Larkum, A.W.D., Salih, A., Itoh, S., Shibata, Y., Bena, C., Yamasaki, H., Papina, M., Van
Woesik, R., 2003. Simultaneous time resolution of the emission spectra of fluorescent proteins and zoox-
anthellar chlorophyll in reef-building corals. Photochem. Photobiol. 77, 515–523.
Gleason, D.F., Wellington, G.M., 1993. Ultraviolet radiation and coral bleaching. Nature 365, 836–838.
Glynn, P.W., 1973. Ecology of a Caribbean coral reef. The Porites reef-flat biotype: part II. Plankton community
with evidence for depletion. Mar. Biol. 22, 1–21.
Glynn, P.W., 1988. El Nino-Southern Oscillation 1982–1983-nearshore population, community, and ecosystem
responses. Ann. Rev. Ecol. Syst. 19, 309–345.
Glynn, P.W., 1991. Coral reef bleaching in the 1980s and possible connections with global warming. Trends Ecol.
Evol. 6, 175–179.
Glynn, P.W., 1993. Coral reef bleaching: ecological perspectives. Coral Reefs 12, 1–17.
Glynn, P.W., Imai, R., Sakai, K., Nakano, Y., Yamazato, K., 1992. Experimental responses of Okinawan (Ryukyu
Islands, Japan) reef corals to high sea temperature and UV radiation. In: Richmond, R. (Ed.), Proceedings of
the Seventh Coral Reef Symposium. University of Guam, Guam, pp. 27–37.
Glynn, P.W., Tourino, J.M., Baker, A.C., Calderon, M.O., 2001. Coral bleaching and mortality in Panawa and
Ecuador during the 1997–1998 El Nino-Southern Oscillation event: spatial/temporal patterns and compar-
isons with the 1982–1983 event. Bull. Mar. Sci. 69, 79–109.
Gorbunov, M., Falkowski, P.G., Kolber, Z., 2000. Measurement of photosynthetic parameters in benthic organ-
isms in situ using a SCUBA-based fast repetition rate fluorometer. Limnol. Oceanogr. 45, 242–245.
Gorbunov, M., Kolber, Z.S., Lesser, M.P., Falkowski, P.G., 2001. Photosynthesis and photoprotection in sym-
biotic corals. Limnol. Oceanogr. 46, 75–85.
Green, A.E.S., Sawada, T., Shettle, E.P., 1974. The middle ultraviolet reaching the ground. Photochem. Photobiol.
19, 251–259.
Green, E.P., Mumby, P.J., Edwards, A.J., Clark, C.D., 1996. A review of remote sensing for the assessment and
management of tropical coastal resources. Coast. Manage. 24, 1–40.
Harvell, C.D., Mitchell, C.E., Ward, J.R., Altzier, S., Dobson, A.P., Ostfield, R.S., Samuel, M.D., 2002. Climate
warming and disease risks for terrestrial and marine biota. Science 296, 2158–2162.
Hedley, J.D., Mumby, P.J., 2003a. A remote sensing method for resolving depth and subpixel composition of
aquatic benthos. Limnol. Oceanogr. 48, 480–488.
Hedley, J.D., Mumby, P.J., 2003b. Biological and remote sensing perspectives of pigmentation in coral reef
organisms. Adv. Mar. Biol. 43, 277–317.
Hochberg, E.J., Atkinson, M.J., 2000. Spectral discrimination of coral reef benthic communities. Coral Reefs 19,
164–171.
Hochberg, E.J., Andrefouet, S., Tyler, M.R., 2003. Sea surface correction of high spatial resolution Ikonos

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252 247
images to improve bottom mapping in near-shore environments. IEEE Trans. Geosci. Remote Sens. 41,
1724–1729.
Hoegh-Guldberg, O., 1999. Climate change, coral bleaching and the future of the world’s coral reefs. Mar.
Freshw. Res. 50, 839–866.
Hoegh-Guldberg, O., Jones, R.J., 1999. Photoinhibition and photoprotection in symbiotic dinoflagellates from
reef-building corals. Mar. Ecol. Prog. Ser. 183, 73–86.
Hoegh-Guldberg, O., Smith, G.J., 1989. The effect of sudden changes in temperature, light and salinity on the
population density and export of zooxanthellae from the reef corals Stylophora pistillata Esper and Seria-
topora hystrix Dana. J. Exp. Mar. Biol. Ecol. 129, 279–303.
Hoffmann, A.A., Hercus, M.J., 2000. Environmental stress as an evolutionary force. Bioscience 50, 217–225.
Hughes, T., 1994. Catastrophes, phase shifts, and large-scale degradation of a Caribbean coral reef. Science 265,
1547–1551.
Hughes, L., 2000. Biological consequences of global warming: is the signal already apparent. Trends Ecol. Evol.
15, 56–61.
Hughes, T.P., Connell, J.H., 1999. Multiple stressors on coral reefs: a long-term perspective. Limnol. Oceanogr.
44, 932–940.
Hughes, T.P., Ayre, D., Connell, J.H., 1992. The evolutionary ecology of corals. Trends Ecol. Evol. 7, 292–295.
Hughes, T., Szmant, A.M., Steneck, R., Carpenter, R., Miller, S., 1999. Algal blooms on coral reefs: what are the
causes?. Limnol. Oceanogr. 44, 1583–1586.
Huppert, A., Stone, L., 1998. Chaos in the Pacific’s coral reef bleaching cycle. Am. Nat. 152, 447–459.
Huston, M.A., 1985. Patterns of species diversity on coral reefs. Ann. Rev. Ecolog. Syst. 16, 145–177.
Iglesias-Prieto, R., Matta, J.L., Robins, W.A., Trench, R.K., 1992. Photosynthetic response to elevated temper-
ature in the symbiotic dinoflagellate Symbiodinium microadriaticum in culture. Proc. Natl. Acad. Sci. 89,
10302–10305.
Isoun, E., Fletcher, C., Frazer, N., Gradie, J., 2003. Multi-spectral mapping of reef bathmetry and coral cover;
Kailua Bay, Hawaii. Coral Reefs 22, 68–82.
Jackson, J.B.C., Kirby, M.X., Berger, W.H., et al., 2001. Historical overfishing and the recent collapse of coastal
ecosystems. Science 293, 629–638.
Johnson, A.S., Sebens, K.P., 1993. Consequences of a flattened morphology: effects of flow on feeding rates of
the scleractinian coral Meandrina meandritis. Mar. Ecol. Prog. Ser. 99, 99–114.
Jokiel, P.L., Coles, S.L., 1990. Responses of Hawaiian and other Indo-Pacific reef corals to elevated temper-
atures. Coral Reefs 8, 155–162.
Jokiel, P.L., Lesser, M.P., Ondrusek, M.E., 1997. UV-absorbing compounds in the coral Pocillopora damicornis:
interactive effects of UV radiation, photosynthetically active radiation, and water flow. Limnol. Oceanogr. 42,
1468–1473.
Jompa, J., McCook, L.J., 2002. The effects of nutrients and herbivory on competition between a hard coral
(Porites cylindrica) and a brown alga (Lobophora variegata). Limnol. Oceanogr. 47, 527–534.
Jones, R.J., Hoegh-Guldberg, O., Larkum, A.W.D., Schreiber, U., 1998. Temperature-induced bleaching of corals
begins with impairment of the CO2 fixation mechanism in zooxanthellae. Plant Cell Environ. 21, 1219–1230.
Kawaguti, S., 1969. Effect of the green fluorescent pigment on the productivity of the reef corals. Micronesica 5,
313.
Kelmanson, I.V., Matz, M.V., 2003. Molecular basis and evolutionary origins of color diversity in Gret Star Coral
Montastraea cavernosa (Scleractinia: Faviida). Mol. Biol. Evol. 20, 1125–1133.
Kim, C.S., Lim, W.A., Cho, Y.C., 2001. Mycosprine-like amino acids as the UV sunscreen with oxygen radical
scavenging activity. Bull. Natl. Fish. Res. Dev. Inst. Korea 60, 65–71.
Kinzie III, R.A., Takayama, M., Santos, S.R., Coffroth, M.A. 2001. The adaptive bleaching hypothesis: exper-
imental tests of critical assumptions. Biol. Bull. 200, 51–58.
Kirk, J.T.O., 1994. Light and Photosynthesis in Aquatic Ecosystems Cambridge Univ. Press, London.
Kleypas, J.A., Buddemeier, R.R., Archer, D., Gattuso, J.P., Langdon, C., Opdyke, B.N., 1999. Geochemical
consequences of increased atmospheric CO2 on corals and coral reefs. Science 284, 118–120.
Knowlton, N., 2001. The future of coral reefs. Proc. Natl. Acad. Sci., 5419–5425.
Knowlton, N., Weil, E., Weight, L.A., Guzman, H., 1992. Sibling species in Montastraea annularis, coral
bleaching, and the coral climate record. Science 255, 330–333.

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252248
Knowlton, N., Mate, J.L., Guzman, H.M., Rowan, R., Jara, J., 1997. Direct evidence for reproductive isolation
among the three species of the Montastraea annularis complex in Central America (Panama and Honduras).
Mar. Biol. 127, 705–711.
Kortschak, R.D., Samuel, G., Saint, R., Miller, D.J., 2003. EST analysis of the cnidarian Acropora millepora
reveals extensive gene loss and rapid divergence in the model invertebrates. Curr. Biol. 13, 2190–2195.
Kuhl, M., Cohen, Y., Dalsgaard, T., Jørgensen, B.B., Revsbech, N.P., 1995. Microenvironment and photosyn-
thesis of zooxanthellae in scleractinian corals studied with microsensors for O2, pH, and light. Mar. Ecol.,
Prog. Ser. 117, 159–172.
Kushmaro, A., Loya, Y., Fine, M., Rosenberg, E., 1996. Bacterial infection and coral bleaching. Nature 380, 396.
Kuta, K.G., Richardson, L.L., 1996. Abundance and distribution of black band disease on coral reefs in the
northern Florida Keys. Coral Reefs 15, 219–223.
Labas, Y.A., Gurskaya, N.G., Yanushevich, Y.G., Fradkov, A.F., Lukyanov, K.A., Lukyanov, S.A., Matz,
M., 2002. Diveristy and evolution of the green fluorescent protein family. Proc. Natl. Acad. Sci. 99,
4256–4261.
LaJeunesse, T.C., 2001. Investigating biodiversity, ecology, and phylogeny of endosymbiotic dinoflagellates
in the genus Symbiodinium using the ITS region: in search of a ‘‘species’’ level marker. J. Phycol. 37,
866–880.
LaJeuness, T.C., 2002. Diversity and community structure of symbiotic dinoflagellates from Caribbean coral
reefs. Mar. Biol. 141, 387–400.
LaJeuness, T.C., Loh, W.K.W., van Woesik, R., Hoegh-Guldberg, O., Schmidt, G.W., Fitt, W.K., 2003. Low
symbiont diversity in southern Great Barrier Reef corals, relative to those of the Caribbean. Limnol. Oceanogr.
48, 2046–2054.
Lapointe, B., 1997. Nutrient thresholds for bottom-up control of macroalgal blooms on coral reefs in Jamaica and
southeast Florida. Limnol. Oceanogr. 42, 1119–1131.
Lapointe, B., 1999. Simultaneous top-down and bottom-up forces control macroalgal blooms on coral reefs.
Limnol. Oceanogr. 44, 1586–1592.
Lesser, M.P., 1996. Exposure of symbiotic dinoflagellates to elevated temperatures and ultraviolet radiation
causes oxidative stress and inhibits photosynthesis. Limnol. Oceanogr. 41, 271–283.
Lesser, M.P., 1997. Oxidative stress causes coral bleaching during exposure to elevated temperatures. Coral Reefs
16, 187–192.
Lesser, M.P., 2000. Depth-dependent effects of ultraviolet radiation on photosynthesis in the Caribbean coral,
Montastraea faveolata. Mar. Ecol. Prog. Ser. 192, 137–151.
Lesser, M.P., Farrell, J.H., 2004. Exposure to solar radiation increases damage to both host tissues and algal
symbionts of corals during thermal stress. Coral Reefs (in press).
Lesser, M.P., Gorbunov, M.Y., 2001. Diurnal and bathymetric changes in chlorophyll fluorescence yields of reef
corals measured in situ with a fast repetition rate fluorometer. Mar. Ecol. Prog. Ser. 212, 69–77.
Lesser, M.P., Shick, J.M., 1989. Effects of irradiance and ultraviolet radiation on photoadaptation in the zoo-
xanthellae of Aiptasia pallida: primary production, photoinhibition and enzymic defences against oxygen
toxicity. Mar. Biol. 102, 243–255.
Lesser, M.P., Stochaj, W.R., Tapely, D.W., Shick, J.M., 1990. Bleaching in coral reef anthozoans: effects of
irradiance, ultraviolet radiation and temperature, on the activities of protective enzymes against active oxygen.
Coral Reefs 8, 225–232.
Lesser, M.P., Weis, V.M., Patterson, M.R., Jokiel, P.L., 1994. Effects of morphology and water motion on carbon
delivery and productivity in the reef coral, Pocillopora damicornis (Linnaeus): diffusion barriers, inorganic
carbon limitation, and biochemical plasticity. J. Exp. Mar. Biol. Ecol. 178, 153–179.
Lesser, M.P., Mazel, C., Phinney, D., Yentsch, C.S., 2000. Light absorption and utilization by colonies of the
congeneric hermatypic corals, Montastraea faveolata and Montastraea cavernosa. Limnol. Oceanogr. 45,
76–86.
Lewis, S.M., 1986. The role of herbivorous fishes in the organization of a Caribbean reef community. Ecol.
Monogr. 56, 183–200.
Lipp, E.K., Jarrell, J.L., Griffin, D.W., Lukasik, J., Jacukiewicz, J., Rose, J.B., 2002. Preliminary evidence for
human fecal contamination in corals of the Florida Keys, USA. Mar. Pollut. Bull. 44, 666–670.
Littler, M.M., Littler, D.S., 1991. Comparisons of N- and P-limited productivity between high granitic islands

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252 249
versus low carbonate atolls in the Seychelles archipelago: a test of the relative dominance model. Coral Reefs
10, 199–209.
Lopez, J.V., Kersanach, R., Rehner, S.A., Knowlton, N., 1999. Molecular determination of species boundaries in
corals: genetic analysis of the Montastraea annularis complex using amplified fragment length polymor-
phisms and a microsatellite marker. Biol. Bull. 196, 80–93.
Louchard, E.M., Reid, R.P., Stephens, F.C., Davis, C.O., Leathers, R.A., Downes, T.V., 2003. Optical remote
sensing of benthic habitats and bathymetry in coastal environments at lee Stocking Island, Bahamas: a
comparative spectral classification approach. Limnol. Oceanogr. 48, 511–521.
Loya, Y., 1972. Community structure and species diversity of hermatypic corals at Eilat, Red Sea. Mar. Biol. 13,
100–123.
Loya, Y., Sakai, K., Yamazato, K., Nakano, Y., Sembali, H., van Woesik, R., 2001. Coral bleaching: the winners
and the losers. Ecol. Lett. 4, 122–131.
Madronich, S., McKenzie, R.L., Bjorn, L.O., Caldwell, M.M., 1998. Changes in biologically active ultraviolet
radiation reaching the earth’s surface. Photochem. Photobiol. 46, 5–19.
Matz, M.V., Fradkov, A.F., Labas, Y.A., Savitsky, A.P., Zaraisky, A.G., Markelov, M.L., Lukyanov, S.A., 1999.
Fluorescent proteins from nonbiolumninescent Anthozoa species. Nat. Biotechnol. 17, 969–973.
Mazel, C.H., 1995. Spectral measurements of fluorescence emission in Caribbean cnidarians. Mar. Ecol. Prog.
Ser. 120, 185–191.
Mazel, C.M., Fuchs, E., 2003. Contribution of fluorescence to the spectral signature and perceived color of
corals. Limnol. Oceanogr. 48, 390–401.
Mazel, C.M., Lesser, M.P., Gorbunov, M.Y., Barry, T.M., Farrell, J.H., Wyman, K.D., Falkowski, P.G., 2003.
Green-fluorescent proteins in Caribbean corals. Limnol. Oceanogr. 48, 402–411.
McClanahan, T.R., 2002. The near future of coral reefs. Environ. Conserv. 29, 1–24.
McClanahan, T., Maina, J., Pet-Soede, L., 2002. Effects of the 1998 coral mortality event on Kenyan coral reefs
and fisheries. Ambio 31, 543–550.
Medina, M., Weil, E., Szmant, A.M., 1999. Examination of the Montastraea annularis complex (Cnidaria:
Scleractinia) using ITS and COI sequences. Mar. Biotechnol. 1, 89–97.
Miller, M.W., Hay, M.E., Miller, S.L., Malone, D., Soka, E.E., Szmant, A.M., 1999. Effects of nutrients versus
herbivores on reef algae: a new method for manipulating nutrients on coral reefs. Limnol. Oceanogr. 44,
1847–1861.
Moore, T.S., Campbell, J.W., Feng, H., 2001. A fuzzy logic classification scheme for selecting and blending
satellite ocean color algorithms. IEEE Trans. Geosci. Remote Sens. 39, 1764–1776.
Mueller-Parker, G., D’Elia, C.F., 1997. Interactions between corals and their symbiotic algae. In: Birkeland, C.
(Ed.), Life and Death of Coral Reefs. Chapman & Hall, New York, pp. 96–113.
Mumby, P.J., Green, E.P., Edwards, A.J., Clark, C.D., 1997. Coral reef habitat mapping: how much detail can
remote sensing provide? Mar. Biol. 130, 193–202.
Mumby, P.J., Green, E.P., Clark, C.D., Edwards, A.J., 2000. Digital analysis of multispectral airborne imagery of
coral reefs. Coral Reefs 17, 59–69.
Murdoch, T.J.T., Aronson, R.B., 1999. Scale-dependent spatial variability of coral assemblages along the Florida
reef tract. Coral Reefs 18, 341–351.
Muscatine, L., 1990. The role of symbiotic algae in carbon and energy flux in reef corals. In: Dubinsky, Z. (Ed.),
Ecosystems of the World: Coral Reefs. Elsevier, Amsterdam, pp. 75–87.
Muscatine, L., Porter, J.W., 1977. Reef Corals: mutualistic symbioses adapted to nutrient-poor environments.
Bioscience 27, 454–460.
Muscatine, L., Porter, J.W., Kaplan, I.R., 1989. Resource partitioning by reef corals as determined from stable
isotope composition: I. y13C of zooxanthellae and animal tissue vs. depth. Mar. Biol. 100, 185–193.
Neale, P.J., Banaszak, A.T., Jarriel, C.R., 1998. Ultraviolet suncreens in Gymnodinium sanguineum (Dinophy-
ceae): mycosporine-like amino acids protect against inhibition of photosynthesis. J. Phycol. 34, 928–938.
Nii, C.M., Muscatine, L., 1997. Oxidative stress in the symbiotic sea anemone Aiptasia pulchella (Calgren,
1943): contribution of the animal to superoxide ion production at elevated temperature. Biol. Bull. 192,
444–456.
Osmond, C.B., 1981. Photorespiration and photoinhibition, some implications for the energetics of photosyn-
thesis. Biochim. Biophys. Acta 639, 77–98.

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252250
Ostrander, G.K., Armstrong, K.M., Knobbe, E.T., Gerace, D., Scully, E.P., 2000. Rapid transition in the
structure of a coral reef community: the effects of bleaching and physical disturbance. Proc. Natl. Acad.
Sci. 97, 5297–5302.
Pandolphi, J.M., 1999. Response of Pleistocene coral reefs to environmental change over long temporal scales.
Am. Zool. 39, 113–130.
Patterson, K.L., Porter, J.W., Ritchie, K.B., Polson, S.W., Mueller, E., Peters, E.C., Santavy, D.L., Smith, G.W.,
2002. The etiology of white pox, a lethal disease of the Caribbean elkhorn coral, Acropora palmate. Proc.
Natl. Acad. Sci. 99, 8725–8730.
Porter, J., 1976. Autotrophy, heterotrophy and resource partitioning in Caribbean reef-building corals. Am. Nat.,
731–742.
Quinn, R.J., de Almeida Leone, P., Guymer, G., Hooper, J.N.A., 2002. Australian biodiversity via its plants and
marine organisms: a high-throughput screening approach to drug discovery. Pure Appl. Chem. 74, 519–526.
Richardson, L.L., 1998. Coral diseases: what is really known. Trends Ecol. Evol. 13, 438–443.
Richardson, L.L., Goldberg, W.M., Kuta, K.M., Aronson, R.B., Smith, G.W., Ritchie, K.B., Halas, J.C., Fein-
gold, J.S., Miller, S.L., 1998. Florida’s mystery coral-killer identified. Nature 392, 557–558.
Richter, M., Ruhle, W., Wild, A., 1990. Studies on the mechanism of photosystem II photoinhibition: II. The
involvement of toxic oxygen species. Photosynth. Res. 24, 237–243.
Rodriguez-Lanetty, M., Loh, W., Carter, D., Hoegh-Guldberg, O., 2001. Latitudinal variability in symbiont
specificity within the widespread scleractinian coral Plesiastrea versipora. Mar. Biol. 138, 1175–1181.
Romano, S.L., Cairns, S.D., 2000. Molecular phylogenetic hypotheses for the evolution of scleractinian corals.
Bull. Mar. Sci. 67, 1043–1068.
Romano, S.L., Palumbi, S.R., 1996. Evolution of scleractinian corals inferred from molecular systematics.
Science 271, 640–642.
Rosenberg, E., Ben-Haim, Y., 2002. Microbial diseases of corals and global warming. Environ. Microbiol. 4,
318–326.
Rowan, R., Knowlton, N., 1995. Intraspecific diversity and ecological zonation in coral–algal symbiosis. Proc.
Natl. Acad. Sci. 92, 2850–2853.
Rowan, R.N., Powers, D.A., 1991a. A molecular genetic classification of zooxanthellae and the evolution of
animal–algal symbioses. Science 251, 1348–1351.
Rowan, R.N., Powers, D.A., 1991b. Molecular genetic identification of symbiotic dinoflagellates (zooxanthellae).
Mar. Ecol. Prog. Ser. 71, 65–73.
Rowan, R., Knowlton, N., Baker, A., Javier, J., 1997. Landscape ecology of algal symbionts creates variation in
episodes of coral bleaching. Nature 388, 265–269.
Rutzler, K., Santavy, D.L., Antonius, A., 1983. The black band disease of Atlantic reef corals: III. Distribution,
ecology, and development. P.S.Z.N.I. Mar. Ecol. 4, 329–358.
Ryan, J.F., Finnerty, J.R., 2003. CnidBase: the cnidarian evolutionary genomics database. Nucleic Acids Res. 31,
159–163.
Salih, A., Larkum, A., Cox, G., Kuhl, M., Hoegh-Guldberg, O., 2000. Fluorescent pigments in corals are photo-
protective. Nature 408, 850–853.
Santos, S.R., Taylor, D.J., Kinzie III, R.A., Hidaka, M., Sakai, K., Coffroth, M.A. 2002. Molecular phylogeny of
symbiotic dinoflagellates inferred from partial chlolroplast large subunit (23S)-rDNA sequences. Mol. Phy-
logenet. Evol. 23, 97–111.
Savage, A.M., Trapido-Rosenthal, H., Douglas, A.E., 2002. On the functional significance of molecular variation
in Symbiodinium, the symbiotic algae of Cnidaria: photosynthetic response to irradiance. Mar. Ecol. Prog. Ser.
244, 27–37.
Schoneberg, D.A., Trench, R.K., 1980a. Genetic variation in Symbiodinium (=Gymnodinium) Freudenthal, and
specificity in its symbiosis with invertebrates: I. Isoenzyme and soluble protein patterns of axenic cultures of
S. microadriaticum. Proc. R. Soc. Lond., B 207, 405–427.
Schoneberg, D.A., Trench, R.K., 1980b. Genetic variation in Symbiodinium (=Gymnodinium) Freudenthal, and
specificity in its symbiosis with invertebrates: III. Specificity and infectivity of S. microadriaticum. Proc. R.
Soc. Lond., B 207, 445–460.
Schreiber, U., Schliwa, U., Bilger, W., 1986. Continuous recording of photochemical and non-photochemical
chlorophyll fluorescence quenching with a new type of modulation fluorometer. Photosynth. Res. 10, 51–62.

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252 251
Sebens, K.P., Johnson, A.S., 1991. The effects of water movement on prey capture and distribution of reef corals.
Hydrobiologia 226, 91–101.
Sebens, K.P., Vandersall, K.S., Savina, L.A., Graham, K.R., 1996. Zooplankton capture by two scleractinian
corals, Madracis mirabilis and Montastrea cavernosa, in a field enclosure. Mar. Biol. 127, 303–318.
Sebens, K.P., Grace, S.P., Helmuth, B., Maney, E.J., Miles, J.S., 1998. Water flow and prey capture by three
scelractinian corals, Madracis mirabilis, Montastrea cavernosa, and Porites porites in a field enclosure. Mar.
Biol. 131, 347–360.
Sharp, V.A., Brown, B.E., Miller, D., 1997. Heat shock protein (HSP 70) expression in the tropical reef coral
Goniopora djiboutiensis. J. Therm. Biol. 22, 11–19.
Sheppard, C.R.C., 2003. Predicted recurrences of mass coral mortality in the Indian Ocean. Nature 425,
294–297.
Shick, J.M., Dunlap, W.C., 2000. Mycosporine-like amino acids and related gadusols: biosynthesis, accumula-
tion, and UV-protective functions in aquatic organisms. Annu. Rev. Physiol. 64, 223–262.
Shick, J.M., Lesser, M.P., Jokiel, P.J., 1996. Effects of ultraviolet radiation on corals and other coral reef
organisms. Glob. Chang. Biol. 2, 527–545.
Shick, J.M., Romaine-Lioud, S., Ferrier-Pages, C., Gattuso, J.-P., 1999. Ultrviolet-B radiation stimulates
shikimate pathway-dependent accumulation of mycosporine-like amino acids in the coral Stylophora
pistillata despite decreases in its population of symbiotic dinoflagellates. Limnol. Oceanogr. 44,
1667–1682.
Shick, J.M., Dunlap, W.C., Buettner, G.R., 2000. Ultraviolet (UV) protection in marine organisms II.
Biosynthesis, accumulation, and sunscreening function of mycosporine-like amino acids. In: Yoshi-
kawa, T., et al. (Eds.), Free Radicals in Chemistry, Biology, and Medicine, Chap. 22. OICA Interna-
tional, pp. 215–228.
Shinn, E.A., Smith, G.W., Prospero, J.M., Betzer, P., Hayes, M.L., Garrison, V., Barber, R.T., 2000. Geophys.
Res. Lett. 27, 3029–3032.
Smith, R.C., Baker, K.S., 1979. Penetration of UV-B and biologically effective dose-rates in natural waters.
Photochem. Photobiol. 29, 311–323.
Smith, S.V., Buddemeier, R.W., 1992. Global change and coral reef ecosystems. Ann. Rev. Ecolog. Syst. 23,
89–118.
Stanley, G.D., Fautin, D.G., 2001. The origins of modern corals. Science 291, 1913–1914.
Stimson, J., 1997. The annual cycle of density of zooxanthellae in the tissues of field and laboratory-held
Pocillopora damicornis (Linnaeus). J. Exp. Mar. Biol. Ecol.
Stott, P.A., Tett, S.F.B., Jones, G.S., Allen, M.R., Mitchell, J.F.B., Jenkins, G.J., 2000. External control of 20th
century temperature by natural and anthropogenic forcings. Science 290, 2133–2137.
Suh, H., Lee, H., Jung, J., 2003. Mycosporine glycine protects biological systems against photodynamic damage
by quenching singlet oxygen with a high efficiency. Photochem. Photobiol. 78, 109–113.
Szmant, A.M., Weil, E., Miller, M.W., Colon, D.E., 1997. Hybridization within the species complex of the
scleractinian coral Montastraea annularis. Mar. Biol. 129, 561–572.
Toller, W.W., Rowan, R., Knowlton, N., 2001a. Zooxanthellae of the Montastraea annularis species complex:
patterns of distribution of four taxa of Symbiodinium on different reefs and across depths. Biol. Bull. 201,
348–359.
Toller, W.W., Rowan, R., Knowlton, N., 2001b. Repopulation of zooxanthellae in the Caribbean corals Montas-
traea annularis and M. faveolata following experimental and disease-associated bleaching. Biol. Bull. 201,
360–373.
Trench, R.K., 1979. The cell biology of plant animal symbioses. Annu. Rev. Plant Physiol. 30, 485–531.
Trench, R.K., 1987. Dinoflagellates in non-parasitic symbiosis. In: Taylor, F.J.R. (Ed.), Biology of Dinoflagellates.
Blackwell, Oxford, pp. 530–570.
Trench, R.K., Blank, R.J., 1987. Symbiodinium microadriaticum Freudenthatl, S. goreauii sp. nov., S. Kawagutii
sp. nov. and S. pilosum sp. nov.: gymnodinoid dinoflagellate symbionts of marine invertebrates. J. Phycol. 23,
469–481.
Tsein, R.Y., 1998. The green fluorescent protein. Annu. Rev. Biochem. 67, 509–544.
Urban, F.E., Cole, J.E., Overpeck, J.T., 2000. Influence of mean climate change on climate variability from a 15-
year tropical Pacific coral record. Nature 407, 989–993.

M.P. Lesser / J. Exp. Mar. Biol. Ecol. 300 (2004) 217–252252
Valenzeno, D.P., Pooler, J.P., 1987. Photodynamic action. Bioscience 37, 270–276.
Van Oppen, M.J.H., Willis, B.H., van Vugt, H.W.J.A., Miller, D.J., 2000. Examination of species boundaries in
the Acropora cervicornis group (Scleractinia, Cnidaria) using nuclear DNA sequence analysis. Mol. Ecol. 9,
1363–1373.
Veron, J.E.N., 1995. Corals in Space and Time: The Biogeography and Evolution of the Scleractinia. Cornell
Univ. Press, New York.
Veron, J.E.N., 2000. Corals of the World. Australian Institute of Marine Sciences, Townsville, Queensland.
Vollmer, S.V., Palumbi, S.R., 2002. Hybridization and the evolution of reef coral diversity. Science 296,
2023–2025.
Ware, J.R., Fautin, D.G., Buddemeier, R.W., 1996. Patterns of coral bleaching: modeling the adaptive bleaching
hypothesis. Ecol. Model. 84, 199–214.
Warner, M.E., Fitt, W.K., Schmidt, G.W., 1996. The effect of elevated temperature on the photosynthetic
efficiency of zooxanthellae in hospite from four different species of reef coral: a novel approach. Plant Cell
Environ. 19, 291–299.
Warner, M.E., Fitt, W.K., Schmidt, G.W., 1999. Damage to photosystem II in symbiotic dinoflagellates: a
determinant of coral bleaching. Proc. Natl. Acad. Sci. 96, 8007–8012.
Warner, M.E., Chilcoat, G.C., McFarland, F.K., Fitt, W.K., 2002. Seasonal fluctuations in the photosynthetic
capacity of photosystem II in symbiotic dinoflagellates in the Caribbean reef-building coral Montastraea. Mar.
Biol. 141, 31–38.
Weil, E., Knowlton, N., 1994. A multi-character analysis of the Caribbean coralMontastraea annularis (Ellis and
Solander, 1786) and its two sibling species, M. faveolata (Ellis and Solander, 1786) and M. franksi (Gregory,
1895). Bull. Mar. Sci. 55, 151–175.
Wellington, G.M., 1982. An experimental analysis of the effects of light and zooplankton on coral zonation.
Oecologia 52, 311–320.
Wellington, G.M., Glynn, P.W., Strong, A.E., Novarrete, S.A., Wieters, E., Hubbard, D., 2001. Crisis on coral
reefs linked to climate change. EOS, Trans.-Am. Geophys. Union 82, 1–5.
Wilkinson, C.R., 1999. Global and local threats to coral reef functioning and existence: review and predictions.
Mar. Freshw. Res. 50, 867–878.
Wilkinson, C., 2000. Status of Coral Reefs of the World. Global Coral Reef Monitoring Network, Australia.
Wilkinson, C., Linden, O., Cesar, G., Rubens, J., Strong, A.E., 1999. Ecological and sociological impacts of 1998
coral mortality in the Indian Ocean: an ENSO impact and a warning of future change. Ambio 28, 188–196.
Winters, G., Loya, Y., Rottgers, R., Beer, S., 2003. Photoinhibition in shallow-water colonies of the coral
Stylophora pistillata as measured in situ. Limnol. Oceanogr. 48, 1388–1393.
Copyright © 2022 FDOKUMEN




















