
Effects of depth and ultraviolet radiation on coral reef turf algae
12
Effects of depth and ultraviolet radiation on coral reef turf algae Anna Fricke a,b,c, ⁎, Mirta Teichberg b , Maggy M. Nugues d,e,f , Svenja Beilfuss a , Kai Bischof a a University of Bremen, Department of Marine Botany, Leobener Str. NW2, 28359 Bremen, Germany b Leibniz Center for Tropical Marine Ecology (ZMT), Fahrenheitstrasse 6, 28359 Bremen, Germany c EA4228 ECOMERS, Université de Nice-Sophia Antipolis (UNS), Faculté des Sciences, Parc Valrose, 06108 Nice cedex 2, France d CRIOBE-USR 3278, CNRS-EPHE-UPVD, 58 avenue Paul Alduy, 66860 Perpignan Cedex, France e Laboratoire d'Excellence ‘CORAIL’, France f CARMABI Foundation, Piscaderabaai z/n, P.O. Box 2090, Willemstad, Curaçao, Netherlands Antilles abstract article info Article history: Received 13 February 2014 Received in revised form 24 July 2014 Accepted 27 July 2014 Available online xxxx Keywords: Curaçao Coral reef Macroalgae Cyanobacteria Transplantation UV radiation Despite the increasing dominance of turf algae in coral reefs, few studies have investigated their physiological and ecological responses to changes in abiotic factors. We tested the effects of depth and ultraviolet radiation on turf algae at different levels of successional stages using two experiments. Depth-related differences were found for all turf algal communities, characterized by a higher amount of the cyanobacteria taxon Dichothrix and the red filamentous genera Poly-/Herposiphonia in the shallow and the appearance of oscillating cyanobacteria in deeper waters. In the first experiment, cross-depth transplantation of 153 days old communities influenced percentage cover, biomass and taxa composition. Downward transplantation lowered overall biomass and abundance of the foraminifera Sorites, whereas the crustose green alga Pringsheimiella and filamentous cyanobacteria colonized the communities. A nearly reverse pattern was observed in upward transplanted com- munities. Overall we distinguished between sensitive taxa, like Oscillatoria, and taxa able to acclimate to alter- ations in their environment, like Pringsheimiella, Poly/Herposiphonia and Dichothrix. In the second experiment, algae grown for 285 days at 5 m were exposed together with a set of sterile settlement tiles to three UVR regimes at 2 m for 22 days. UVR had no effect on turf algal communities regardless of successional stage. This study high- lights the presence of high light and UV tolerant species. The high UV tolerance of turf communities may confer a competitive advantage over other more sensitive coral reef biota, such as corals. This study demonstrates that turf algae are dynamic communities exhibiting species-specific resistance to environmental changes. © 2014 Elsevier B.V. All rights reserved. 1. Introduction The degradation of coral reefs is commonly associated with a reduc- tion in coral cover and diversity, coupled with an increase in algal biomass and a shift in algal community composition (Diaz-Pulido et al., 2009; McCook, 1999). On pristine healthy reefs, benthic algae are heavily grazed by herbivores and are dominated by crustose coral- line algae, as well as diminutive, mixed assemblages of filamentous algae, juvenile macroalgae and cyanobacteria, typically referred to as turf algae (Steneck and Simon, 2001). As reefs degrade, algal biomass increases. Crustose coralline algae are replaced by dense turf algae and large macrophytes (McCook, 1999). Studies on the factors driving algal communities have typically focused on large macroalgal genera like Sargassum, Lobophora, Dictyota or Halimeda (Engelen et al., 2005; Mumby, 2009; Mumby et al., 2005; Nugues and Bak, 2008; Teichberg et al., 2013). Turf algae however have become one of the most abundant benthic functional groups on coral reef communities worldwide (Haas and Wild, 2010; Haas et al., 2010; McCook, 2001; Steneck and Simon, 2001). Yet, they have received less attention. Turf algae play a crucial role in coral reefs (Birrell et al., 2005; Fricke et al., 2011b; Vermeij et al., 2010). They perform multiple functions due to their rapid turnover of organic matter (Carpenter and Williams, 2007; Copertino et al., 2005; Wanders, 1976), their role as microhabitats for certain invertebrates and microorganisms (Barott et al., 2011; Chapman and Waters, 1992; Hacker and Steneck, 1990; Kramer et al., 2012; Olabarria and Chapman, 2001) and their accumulation potential for sediment and detritus (Purcell and Bellwood, 2001; Wiebe et al., 1975; Williams and Carpenter, 1997). Furthermore, due to the opportu- nistic life histories of many species, they can rapidly colonize dead or Journal of Experimental Marine Biology and Ecology 461 (2014) 73–84 Abbreviations: EC, early community; LC, late community; P, PAR (400–700 nm) treatment; PA, PAR + UV-A (320–700 nm) treatment; PAB, PAR + UVA + UVB (280–700 nm) treatment; PAR, photosynthetically active radiation (400–700 nm); UV-A, ultraviolet-A radiation, considered in the study as (320–400 nm); UV-B, ultraviolet-B radiation, considered in the study as (280–320 nm); UVR, ultraviolet radiation (280–400 nm); 5 m, community grown at 5 m depth; 15 m, community grown at 15 m depth; 5mT, community grown at 15 m and upward transplanted to 5 m depth at T0; 15mT, community grown at 5 m and downward transplanted to 15 m at T0. ⁎ Corresponding author at: EA4228 ECOMERS, Université de Nice-Sophia Antipolis (UNS), Faculté des Sciences, Parc Valrose, 06108 Nice Cedex 2, France. Tel.: +33 492076846; fax: +33 492076849. E-mail address: [email protected] (A. Fricke). http://dx.doi.org/10.1016/j.jembe.2014.07.017 0022-0981/© 2014 Elsevier B.V. All rights reserved. Contents lists available at ScienceDirect Journal of Experimental Marine Biology and Ecology journal homepage: www.elsevier.com/locate/jembe
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Effects of depth and ultraviolet radiation on coral reef turf algae

Journal of Experimental Marine Biology and Ecology 461 (2014) 73–84
Contents lists available at ScienceDirect
Journal of Experimental Marine Biology and Ecology
j ourna l homepage: www.e lsev ie r .com/ locate / jembe
Effects of depth and ultraviolet radiation on coral reef turf algae
Anna Fricke a,b,c,⁎, Mirta Teichberg b, Maggy M. Nugues d,e,f, Svenja Beilfuss a, Kai Bischof a
a University of Bremen, Department of Marine Botany, Leobener Str. NW2, 28359 Bremen, Germanyb Leibniz Center for Tropical Marine Ecology (ZMT), Fahrenheitstrasse 6, 28359 Bremen, Germanyc EA4228 ECOMERS, Université de Nice-Sophia Antipolis (UNS), Faculté des Sciences, Parc Valrose, 06108 Nice cedex 2, Franced CRIOBE-USR 3278, CNRS-EPHE-UPVD, 58 avenue Paul Alduy, 66860 Perpignan Cedex, Francee Laboratoire d'Excellence ‘CORAIL’, Francef CARMABI Foundation, Piscaderabaai z/n, P.O. Box 2090, Willemstad, Curaçao, Netherlands Antilles
Abbreviations: EC, early community; LC, late commtreatment; PA, PAR + UV-A (320–700 nm) treatme(280–700 nm) treatment; PAR, photosynthetically acUV-A, ultraviolet-A radiation, considered in the stuultraviolet-B radiation, considered in the study as (280–320(280–400 nm); 5 m, community grown at 5 m depth; 15depth; 5mT, community grown at 15 m and upward tra15mT, community grown at 5 m and downward transplan⁎ Corresponding author at: EA4228 ECOMERS, Unive
(UNS), Faculté des Sciences, Parc Valrose, 06108 Nic492076846; fax: +33 492076849.
E-mail address: [email protected] (A. Fricke).
http://dx.doi.org/10.1016/j.jembe.2014.07.0170022-0981/© 2014 Elsevier B.V. All rights reserved.
a b s t r a c t
a r t i c l e i n f oArticle history:Received 13 February 2014Received in revised form 24 July 2014Accepted 27 July 2014Available online xxxx
Keywords:CuraçaoCoral reefMacroalgaeCyanobacteriaTransplantationUV radiation
Despite the increasing dominance of turf algae in coral reefs, few studies have investigated their physiologicaland ecological responses to changes in abiotic factors. We tested the effects of depth and ultraviolet radiationon turf algae at different levels of successional stages using two experiments. Depth-related differences werefound for all turf algal communities, characterized by a higher amount of the cyanobacteria taxon Dichothrixand the red filamentous genera Poly-/Herposiphonia in the shallow and the appearance of oscillatingcyanobacteria in deeperwaters. In thefirst experiment, cross-depth transplantation of 153 days old communitiesinfluenced percentage cover, biomass and taxa composition. Downward transplantation lowered overall biomassand abundance of the foraminifera Sorites, whereas the crustose green alga Pringsheimiella and filamentouscyanobacteria colonized the communities. A nearly reverse pattern was observed in upward transplanted com-munities. Overall we distinguished between sensitive taxa, like Oscillatoria, and taxa able to acclimate to alter-ations in their environment, like Pringsheimiella, Poly/Herposiphonia and Dichothrix. In the second experiment,algae grown for 285 days at 5mwere exposed togetherwith a set of sterile settlement tiles to three UVR regimesat 2m for 22 days. UVR had no effect on turf algal communities regardless of successional stage. This study high-lights the presence of high light and UV tolerant species. The high UV tolerance of turf communities may confer acompetitive advantage over othermore sensitive coral reef biota, such as corals. This study demonstrates that turfalgae are dynamic communities exhibiting species-specific resistance to environmental changes.
© 2014 Elsevier B.V. All rights reserved.
1. Introduction
The degradation of coral reefs is commonly associated with a reduc-tion in coral cover and diversity, coupled with an increase in algalbiomass and a shift in algal community composition (Diaz-Pulidoet al., 2009; McCook, 1999). On pristine healthy reefs, benthic algaeare heavily grazed by herbivores and are dominated by crustose coral-line algae, as well as diminutive, mixed assemblages of filamentousalgae, juvenile macroalgae and cyanobacteria, typically referred to as
unity; P, PAR (400–700 nm)nt; PAB, PAR + UVA + UVBtive radiation (400–700 nm);dy as (320–400 nm); UV-B,nm); UVR, ultraviolet radiationm, community grown at 15 mnsplanted to 5 m depth at T0;ted to 15 m at T0.rsité de Nice-Sophia Antipolise Cedex 2, France. Tel.: +33
turf algae (Steneck and Simon, 2001). As reefs degrade, algal biomassincreases. Crustose coralline algae are replaced by dense turf algae andlarge macrophytes (McCook, 1999). Studies on the factors drivingalgal communities have typically focused on large macroalgal generalike Sargassum, Lobophora, Dictyota or Halimeda (Engelen et al., 2005;Mumby, 2009; Mumby et al., 2005; Nugues and Bak, 2008; Teichberget al., 2013). Turf algae however have become one of themost abundantbenthic functional groups on coral reef communities worldwide (Haasand Wild, 2010; Haas et al., 2010; McCook, 2001; Steneck and Simon,2001). Yet, they have received less attention.
Turf algae play a crucial role in coral reefs (Birrell et al., 2005; Frickeet al., 2011b; Vermeij et al., 2010). They performmultiple functions dueto their rapid turnover of organic matter (Carpenter and Williams,2007; Copertino et al., 2005;Wanders, 1976), their role asmicrohabitatsfor certain invertebrates and microorganisms (Barott et al., 2011;Chapman and Waters, 1992; Hacker and Steneck, 1990; Kramer et al.,2012; Olabarria and Chapman, 2001) and their accumulation potentialfor sediment and detritus (Purcell and Bellwood, 2001; Wiebe et al.,1975;Williams and Carpenter, 1997). Furthermore, due to the opportu-nistic life histories of many species, they can rapidly colonize dead or

Fig. 1. Experimental setups and designs. A) Cross-transplantation experiment showingthe up- or down-transplanted (black-filled) and untransplanted (white-filled) tiles at5 and 15 m depths at the different times: T0 = experimental start, March 17, 2008,T1 = April 9, 2008, and T2 = May 6, 2008. B) Experimental UV exposure setup at 2 mwater depth. C) UV exposure experiment showing the arrangement of experimentalunits within different treatments of light: C = full sunlight, PAB = PAR + UVA + UVB(280–700 nm), PA= PAR+ UVA (320–700 nm) and P= PAR (400–700 nm). The differ-ent communities used in the experiment are symbolized by squares: white = earlysuccessional community (EC), striped= late successional stages (LC), randomly distributedalong the treatments.
74 A. Fricke et al. / Journal of Experimental Marine Biology and Ecology 461 (2014) 73–84
damaged coral colonies and out-compete corals by fast growth rates,potentially reinforcing coral–algal phase shifts (Diaz-Pulido and McCook,2002, 2004; Titlyanov et al., 2005; Vermeij et al., 2010).
Despite their importance to coral reefs, little is known about theecology of turf algal communities and their response to changes inabiotic factors. The abundance and distribution of these benthic algaeare influenced by a suite of environmental parameters, including light,temperature, nutrients, salinity, sedimentation, depth and wave expo-sure (Diaz-Pulido et al., 2009; Fricke et al., 2011b, 2011c; Littler et al.,2010; McCook, 1999; Nugues and Roberts, 2003; Vermeij et al., 2010).However, relatively few studies have experimentally tested the directeffect of these physical factors on turf algae (e.g. Copertino et al.,2006; Fricke et al., 2011b; Littler et al., 2010).
As most benthic coral organisms are sessile and photosyntheticallyactive, coral reefs are areas of intense competition for space and light(Benayahu and Loya, 1981; McCook et al., 2001; Titlyanov andTitlyanova, 2008). Light quantity and quality (i.e. spectral composition)are two crucial factors in the usually oligotrophic transparent waters ofcoral reefs. Short atmospheric path length and the thinness of thestratospheric ozone layer contribute to natural high ultraviolet radiation(UVR) in the tropics (Fleischmann, 1989). As a consequence of climatechange, UVR will increase in all latitudes (Karl and Trenberth, 2003;Shindell et al., 1998; Tsay and Stamnes, 1992; Tuck et al., 1992). So faronly small changes are reported for tropical latitudes, with an increaseof DNA-weighted doses of UVR of 2.8+/−2.1% at 15°S and 15°Nbetween 1979 and 1994 (Shick et al., 1996). But even small changesmight be significant to tropical marine organisms experiencing thehighest irradiances of UVR worldwide (Banaszak and Lesser, 2009).
Light and UVR have important roles in mitigating competition be-tween different reef organisms (Benayahu and Loya, 1981; Jompa andMcCook, 2003; Titlyanov et al., 2005). Corals have been shown to besensitive to intense light and UVR (Gleason et al., 2006; Kuffner, 2001).To date, there are few studies that have investigated UV impacts on trop-ical turf algal communities (Dobretsov et al., 2005; Wahl et al., 2004),and only one (Santas et al., 1998) dealt with UV effects on Caribbeanturf algal communities. This single study showed a decrease in turfalgal biomass under artificially enhanced UV-B radiation in a mesocosmexperiment in the “Living Coral Reef” mesocosm at the Natural HistoryMuseum inWashington. Light effects are often studied over depth gradi-ents. In a recent study, turf algal communities were shown to exhibitlarge depth related differences in species composition and standingcrop, suggesting an important effect of light (Fricke et al., 2011b).
Here we investigated the effect of light quantity related to differentwater depths and light qualitymanipulated by different UVR filter treat-ments on turf algal communities naturally grown in a fringing coral reefat the southwest coast of Curaçao, Caribbean. In a first experiment, turfalgae grown over a period of 153 days on ceramic tiles, for examiningnew recruitment and early succession, at 5 and 15 m depths werecross-depth transplanted (i.e. 5 m tiles were brought to 15 m and viceversa) to test for increasing and decreasing light effects. In a secondexperiment, turf algal communities grown over a period of 285 daysat 5 m depth were exposed together with a set of sterile ceramic tilesto different UVR regimes at 2 m to investigate the effects of increasinglight andUVR. The objectives of the studywere to determine: 1)whethera change in underwater light regime or UVR can alter the diversity,biomass, and taxa composition of tropical turf algal communities;2) which taxa were most affected; and 3) whether light and UV effectsvary with community age, i.e. successional stage.
2. Materials and methods
2.1. Study location and site
This study was conducted in Curaçao, former Netherlands Antilles(12°01′–12°23′N; 68°44′–69°10′W), an island in the southern Caribbeanapproximately 70 km north off the coast of Venezuela. The island is
bordered by a fringing reef formed by plateaus of Pleistocene coral lime-stone that form steep cliffs on the seaward site (for further details seeVan den Hoek et al., 1972; Wanders, 1976). The experimental site,Buoy 0 (12°07′N, 69°57′), also called Klein Piscadera, is located on theSW coast of the island, 100 m west from the entrance to Piscadera Bay.For a detailed description of the experimental site and of the turf algalcommunities, see Van den Hoek (1969), Van den Hoek et al. (1972),Van den Hoek et al. (1975) and Van den Hoek et al. (1978).
2.2. Cross-depth transplantation experiment
Twenty-five 9.6 cm× 9.6 cmunglazed ceramic settlement tiles weredispatched at each of twodepths (5 and 15m) inOctober 2007. The tileswere laid horizontally on oblong grey polyvinyl chloride (PVC) panels(75 × 20 cm) fixed on a frame made of concrete reinforcement bars(100 × 200 cm), anchored on cinder blocks. Each PVC panel carriedtwo- to four-settlement panels fixed by PVC clamps (for further detailssee Fricke et al., 2011b). ln March 2008 (after 153 days of succession),ten tiles from each depth were transplanted either upward to 5 m forthe 15 m tiles or downward to 15 m for the 5 m tiles, thus formingfour different treatments (Fig. 1A): 1) 5 m: community grown at 5 mdepth, 2) 15 m: community grown at 15 m depth, 3) 5mT: communitygrown at 15 m and upward transplanted to 5 m depth at T0, and4) 15mT: community grown at 5 m and downward transplanted to

75A. Fricke et al. / Journal of Experimental Marine Biology and Ecology 461 (2014) 73–84
15 m at T0. To minimize handling disturbance during the experiment,whole PVC panels were carefully moved and transplanted. No losses ofloose algae material and sediments were observed during transporta-tion, so procedural controls were not used. To minimize differencesin environmental conditions and particularly exposure to light, thetransplanted panels were haphazardly positioned on the concrete barsnext to, or in between, the undisturbed control panels. Tiles weredestructively sampled after 23 days (T1 = April 9 2008) and 50 days(T2=May 6 2008) (n=5 per time period and treatment). In addition,5 tiles from each depth were sampled prior to transplantation (T0).Sampling of tiles was done haphazardly.
2.3. UV experiment
We constructed four treatments to test for the potential effectof UV radiation. Each treatment consisted of an aligned PVC box(40 cm × 40 cm) (Fig. 1B). Three of the four PVC boxes were coveredby distinct filters consisting of plexiglass sheets (Sunactive GS Farblos2458, Degussa, Röhm, Germany) with attached radiation filter foils of0.5 mm thickness, allowing certain wavelengths to penetrate: 1) PAR(P, 400–700 nm; 400 Ultraphan URUV ‘Farblos’ foils, Digefra GmbH,Munich, Germany), 2) PAR + UV-A (PA, 320–700 nm; 320 Folanormfoils, Folex GmbH, Dreieich, Germany), and 3) PAR + UV-A + UV-B(PAB, 280–700 nm; 295 Ultraphan transparent 140 foils, DigefraGmbH, Munich, Germany) treatments, respectively. A fourth treatment(C) consisted of the fourth PVC box, which was neither covered by aPerspex sheet nor filter foils, and served as control (i.e., for shading).In order to stay consistent with the cut-off wavelengths provided bythe filter foil, we considered in the following UV-A (315–400 nm) as320–400 nm and UV-B (280–315 nm) as 280–320 nm. To avoid anycage effect, the sides of all plots were left open and tiles were placed inthe center of each plot to avoid sideward light penetration due to waveactivities. To examine the influence of successional stages, each boxcontained one sterile tile (early community = EC) and one 285 day oldtile (late community = LC) positioned horizontally with respect towater surface and at a distance of ≥10 cm to each other at the center ofeach box. Treatments were replicated four times and equally distributedonto two rafts (Fig. 1C) made of concrete reinforcement bars. Since apreliminary experience conducted at 5 m depth over a period of54 days in March 2008 demonstrated no UVR effect (Fricke, 2011), eachraft was anchored to the bottom at 5 m depth and lifted to 2 m depthusing four floating buoys to maximize UVR radiation (Fig. 1B). The tworafts were less than 5 m apart and held the plots parallel to the surfaceand parallel with one another. The LC communities were grown betweenMay 2008 and February 2009 at 5 m depth and the experiment was ini-tiated on February 20, 2009. To control for the effect of the upward trans-plantation effect from 5 m to 2 m depth, an additional treatment (5 m)was included. It consisted of four EC tiles and four LC tiles laid on an ob-long PVC panel (15 × 75 cm) placed at 5 m near the rafts and anchoredon cinder blocks 20 cm from the ground. The experimental setup waschecked daily and filters were cleaned every other day using a softsponge. Tiles were sampled all at once after 22 days.
2.4. Sampling procedure
Sampled tileswere placed in an individual plastic bag filledwith am-bient seawater. To avoid damage through excessive solar radiation ortemperature, theywere alsowrapped in an opaque bag and transportedin a cooling box filled with ambient seawater to the laboratory(b15 min). In the laboratory, samples were taken for pigment analyses,community structure and biomass as described below.
2.5. Pigment analyses
An area (9.6 cm × 2 cm) of each tile was scraped using a sterilescalpel, immediately frozen in liquid nitrogen and stored below
−20 °C for further analyses. Prior to the analyses, all samples fromthe UV experiment were additionally freeze-dried and re-weighted.To investigate the chlorophyll and phycobiliprotein concentrations ofthe different turf algal communities, each sample was split and dividedequally in two preweighed eppendorf cups. Splitting was done underliquid nitrogen at low light conditions to avoid photo destruction ofpigments. Samples were stored in liquid N2 until extraction.
2.5.1. Photometric analysesThe phycobiliprotein content for each sample was determined after
Beer and Eshel (1985). The chlorophyll content of each cross-depthexperiment sample was extracted in 1.5 ml 90% acetone and keptin darkness at 4 °C. After 22 h samples were centrifuged for 5 min10.000 rpm at 4 °C (Heraeus, Biofuge fresco). The supernatant wastransferred into quartz glass cuvettes (QS 10 mm, Hellma) and mea-sured spectrophotometrically (UV-2401 PC, Shimadzu, Tokyo, Japan)at wavelengths of 750, 664, 647 and 630 nm. To quantify the amountof chlorophyll a, b and c within the communities, the equation ofJeffrey and Humphrey was used (Arar and Collins, 1997; Jeffrey andHumphrey, 1975).
2.5.2. HPLC analysesChlorophylls and carotenoids of the turf algae from the EC tiles of
the UV experiment were identified via high pressure liquid chromatog-raphy (HPLC). Approximately 0.01 g of freeze-driedmaterial per samplewas diluted into 1ml acetone (90:10), stirred in the vortex and extract-ed in darkness at 4 °C. After 22 h, samples were centrifuged for 5min at13.000 rpm at 4 °C (Heraeus, Biofuge fresco). After filtration (0.2 μmWhatman membrane filters), extracts were immediately injected intoa Merck Hitachi HPLC (La Chrom Elite) with a photodiode array (DADL-2450). Chromatographic separation was carried out using a C18column for reverse phase chromatography (LiChroCart; 12.5 cm long;4 mm in diameter; 5 μm particles; RP-18) and a 30 min solvent systemprogram, following the modified protocol on Spectraphysics systems,with Solvent A: 80:20 methanol, Solvent B: 90:10 acetonitrile andsolvent C: ethyl acetate (HPLC grade), a flow rate of 1 ml min−1 andan injection volume of 50 μl. Pigment detection was routinely conduct-ed at 436 nm for all chlorophylls and carotenoids. Pigments wereidentified and quantified by comparison to pigment standards (DHI,Waters and Environment, Denmark). Concentrations were expressedon an areal basis (μg cm−2) to account for density variation betweensamples.
2.6. Light absorbing compounds
In order to investigate the concentrations of hydrophobic light-absorbing compounds at different wavelengths within the differentcommunities, pigment extractions used for HPLC analyses (see above)were transferred into quartz glass cuvettes (QS 10 mm, Hellma) andmeasured spectrophotometrically (UV-2401 PC, Shimadzu, Tokyo,Japan). Absorbance was recorded from 260 to 750 nm. Data werecorrected for wet weight of communities and summarized accordingto the different cut off wavelengths of the experimental treatments (seeabove): UV (280–400 nm), UV-A (320–400 nm), UV-B (280–320 nm),and PAR (320–400 nm).
2.7. Community analyses and species identification
To determine the composition of the benthic assemblages, the per-centage cover of individual macrobenthic taxa (N0.5 mm) was recordedfrom each tile collected using a dissection microscope (12×, Zeiss StemiSV6) with an ocular grid (10/10 divisions, 1 cm × 1 cm per visual field).Percentage cover of taxa exceeding 5% were grouped and categorized in5% increments (5–10%, 10–15%, 15–20%, etc. to 95–100%). Due to three-dimensionality, certain taxa overlapped, which caused the overall sumof cover to exceed 100%. Each tile was rinsed gently with filtered
76 A. Fricke et al. / Journal of Experimental Marine Biology and Ecology 461 (2014) 73–84
(60 μm) seawater using awash bottle in order to remove grazers and sed-iment. To avoid any edge effect, only the central part of the tile (5 × 5 cm)was used. Within this area, four visual fields (4 cm2) were analyzed. Themost abundant taxa were identified, except for the EC tiles of the UV ex-periment where all individuals were taken into account. Further detailsof the procedure can be found in Fricke et al. (2011b).
Taxa richness (S) and evenness (J′) were determined for eachsampling time and successional stage from the identified taxa.Additionally, total algal biomass was recorded. To determine totalturf algal community biomass, half of the algal material from thecentral part of each tile was removed with a sterilized razorbladeand dried over 3 days at 60 °C to determine dry weight per unitsurface area.
2.8. Environmental parameters
Irradiance was measured using a submersible spectroradiometer(RAMSES ACC UV–vis, 280–700 nm, Trios, Oldenburg, Germany) in airand underwater from 0.5 to 15 m depth on March 20, March 25 andApril 25, 2008. On each date, two replicate profile measurements wereconducted at 0.5 m intervals, and irradiance was averaged for eachdepth. The diffuse vertical attenuation coefficients of downwardirradiance (kd) for the spectral ranges UV-B, UV-A and PAR weredetermined using the formula by Kirk (1994) cited in Hanelt et al.(2001): kd(m−1) = ln[Ed(z2) / Ed(z1)] × (z1 − z2)−1, in whichEd(z1) and Ed(z2) are the respective irradiances at depth z1 and z2.
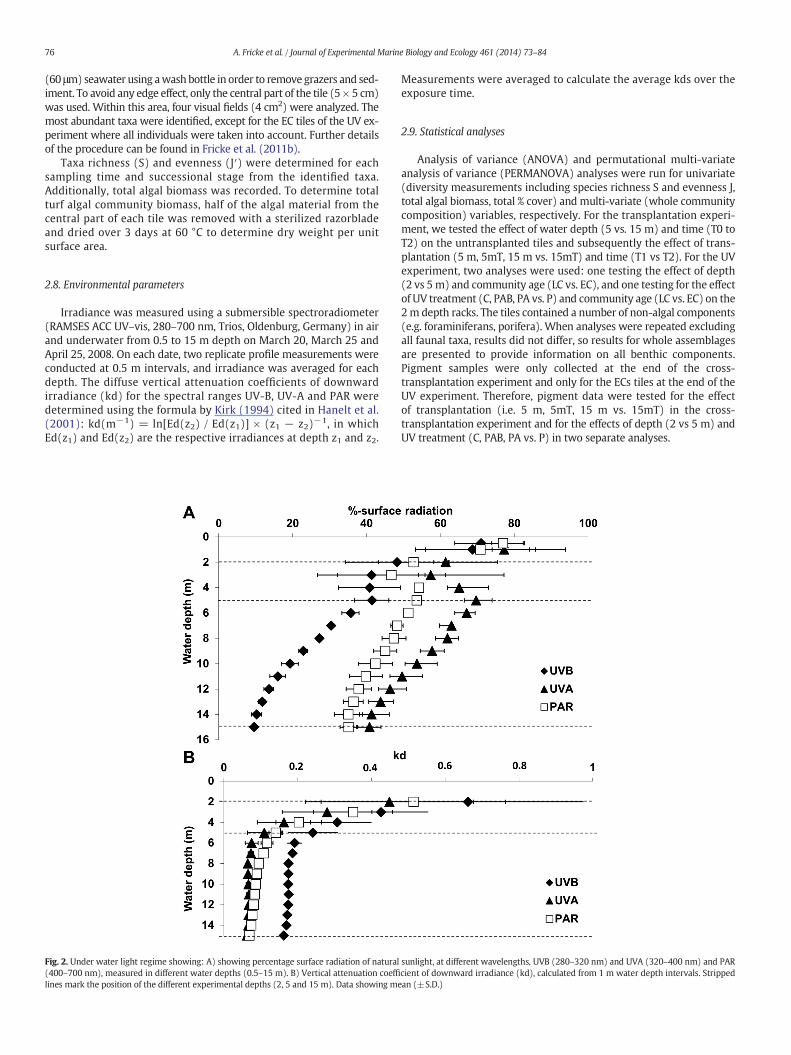
Fig. 2. Under water light regime showing: A) showing percentage surface radiation of natural(400–700 nm), measured in different water depths (0.5–15 m). B) Vertical attenuation coeffilines mark the position of the different experimental depths (2, 5 and 15 m). Data showing me
Measurements were averaged to calculate the average kds over theexposure time.
2.9. Statistical analyses
Analysis of variance (ANOVA) and permutational multi-variateanalysis of variance (PERMANOVA) analyses were run for univariate(diversity measurements including species richness S and evenness J,total algal biomass, total % cover) and multi-variate (whole communitycomposition) variables, respectively. For the transplantation experi-ment, we tested the effect of water depth (5 vs. 15 m) and time (T0 toT2) on the untransplanted tiles and subsequently the effect of trans-plantation (5 m, 5mT, 15 m vs. 15mT) and time (T1 vs T2). For the UVexperiment, two analyses were used: one testing the effect of depth(2 vs 5 m) and community age (LC vs. EC), and one testing for the effectof UV treatment (C, PAB, PA vs. P) and community age (LC vs. EC) on the2m depth racks. The tiles contained a number of non-algal components(e.g. foraminiferans, porifera). When analyses were repeated excludingall faunal taxa, results did not differ, so results for whole assemblagesare presented to provide information on all benthic components.Pigment samples were only collected at the end of the cross-transplantation experiment and only for the ECs tiles at the end of theUV experiment. Therefore, pigment data were tested for the effectof transplantation (i.e. 5 m, 5mT, 15 m vs. 15mT) in the cross-transplantation experiment and for the effects of depth (2 vs 5 m) andUV treatment (C, PAB, PA vs. P) in two separate analyses.
sunlight, at different wavelengths, UVB (280–320 nm) and UVA (320–400 nm) and PARcient of downward irradiance (kd), calculated from 1 m water depth intervals. Strippedan (±S.D.)
77A. Fricke et al. / Journal of Experimental Marine Biology and Ecology 461 (2014) 73–84
Homogeneity of variances of univariate data was tested usingCochran's test, and variances were homogenized by square root orlog transformationwhen necessary. In case no homogeneity was gainedby transformation, datawere analyzed using a non-parametric Kruskal–Wallis test (H). Tukey's Test of Honest Significant Difference (HSD)and multiple comparisons of mean ranks were used for post hoc tests.PERMANOVAs were based on Bray–Curtis similarity indices calculatedfrom percent cover, prior to square root transformations in order toscale-down the importance of the highly abundant taxa. When a factorand/or interaction was identified as significant (P b 0.05), post-hocPERMANOVA pair-wise tests were conducted to detect differenceswithin levels of each relevant factor. Similarity percentages (SIMPER)were used to determine the relative contribution of single taxa respon-sible for differences in specific species composition among treatments.To visualize differences among different communities between the dif-ferent treatments, nonmetric multi-dimensional scaling (MDS) plotswere used based on calculations of Bray–Curtis similarity matrices(Bray and Curtis, 1957).
3. Results
3.1. Environmental parameters
The upper 5 m surface water was highly variable in terms of lightattenuation (Fig. 2). Below 6 m the kd values stabilized, reaching onaverage 0.09 m−1 for PAR, 0.07 m−1 for UV-A and 0.18 m−1 for UV-Bdown to 15 m. The light transparency of the water column was high,
Table 1Results of four ANOVAs and four PERMANOVAs for both experiments, testing the effect of depthand T2), transplantation (DOWN and UP), age (EC and LC) and treatment (C, PAB, PA, PAR) on thare confirmed by pair-wise comparisons within levels of treatments and SIMPER analyses. In or**fourth root transformation, *** log transformation, or analyzed using a nonparametric Kruska
ANOVA
Factors df S J Cover (c) Biomass (b)
F p F/H p F/H p F/H p
a) Cross-depth transplantation experimentEffects of time and depth on turf algal communities
Depth (D) 1 0.53 0.474 0.09 0.759 10.47 K 0.001 K 111.01*** 0.
Time (T) 2 1.11 0.352 1.14 0.343 0.20 0.821 15.24 0.D x T 2 0.61 0.555 0.79 0.469 0.16 0.849 2.04 1.
Res 18Effect of time and transplantation treatments
Transplant (Tr) 3 0.93 0.439 1.62 0.211 4.13** 0.017** 15.32 K 0.
Time (Ti) 1 0.06 0.802 0.43 0.519 0.26 0.614 1.33 0.Tr x Ti 3 0.23 0.871 1.49 0.242 0.11 0.951 1.63 0.
Res 24
b) UV experimentEffect of depth and age
Depth (D) 1 0.01 0.924 2.08 0.175 0.217 0.649 9.06*** 0.Age (A) 1 0.24 0.636 14.35 0.002 23.56 0.000 52.65*** 0.
D x A 1 0.01 0.92 0.69 0.42 0.84 0.378 6.16*** 0.
Res 12Effect of UV treatment and age
Treatment (T) 3 0.62 0.606 0.44 0.73 0.913 0.45 1.112 0.Age (A) 1 0.94 0.343 17.81 K 0.000 K 103.92* 0.000* 117.73*** 0.
T x A 3 0.06 0.98 1.16 0.36 1.17 0.343 1.06 0.Res 24
Bold values are statistically significant.
reaching 35%, 41% and 10% of surface measured PAR, UV-A, and UV-B,respectively at 15 m. From 5 to 15 m, UV-B radiation decreased drasti-cally by 30%, and from 2 to 5 m by 8%, whereas PAR decreased by 20%between 5 and 15 m, but hardly changed between 2 and 5 m.
3.2. General community description
We distinguished 32 different benthic taxa, including six Chloro-phyta (Chl) (1—Pringsheimiella, 2—Derbesia, 3—Ulva, 4—Cladophora,5—Chaetomorpha, 6—Bryopsis), three Phaeophyceae (Ph, 7—Sphacelaria,8—Ectocarpales, 9—Crustous Phaeophycea), and eleven Rhodophyta(Rh, 10—Hydrolithon, 11—Anotrichum, 12—Griffithsia, 13—Rhodophyte,14—Laurencia/Chondria, 15—Jania, 16—Centroceras, 17—Antitrichonella,18—Polysiphonia, 19—Herposiphonia, 20—Ceramium). For theCyanobacteria (Cy), we distinguished six taxa: 21—Dichothrix, 22—Rivularia aff. polyotis, 23—Rivularia spp., 24—Oscillatoria, 25— Calothrix,as well as the taxa 26— filamentous cyanobacteria, which containedmorphologically indistinguishable filamentous forms. Furthermore,we considered one Bacillariophyta (Ba; 27—Tube dwelling diatoms),two foraminifera (Fo; 28—Planorbulina, 29—Sorites) and two taxa ofsessile fauna (Fa; 30—Sedentarier, 31—Porifera). Details to the speciescomposition of these taxa can be found in Fricke et al. (2011b). Forthe transplantation experiment, we grouped taxa 2–6 as filamentousChlorophyta and 18–19 as Poly/Herposiphonia. In addition, we consid-ered the observed cyanobacteria bloom (partly composed of Nostoccommune Vaucher ex Bornet & Flahault, 1888) as separate taxa (32—Cyano-Ecto-mat).
(cross transplantation experiment: 5 and 15 m; UV experiment: 2 and 5 m), time (T0, T1e community composition of different turf algal communities. Conclusions of PERMANOVAder to provide homogeneity, data were transformed prior analyses: * root transformation,l–Wallis test K.
PERMANOVA
Conclusions Community Conclusions
Pseudo-F p
000 ⁎⁎⁎ c : 15 m N 5 mb : 5 m N 15 m
33.68 0.001 5 m ≠ 15 m at all times
000 b : T1 N T0,T2 4.356 0.001 T0 ≠ T1 ≠ T258 4.128 0.001 5 m : T1 ≠ T0, T2 ≠ T3;
Cyano-Ecto bloom causes changein 5 m at T115 m: T0 = T1, T1 = T2;gradually change
002 K c:15 m N 5 m, 15mTb: 5 m N15 m
12.86 0.001 Communities from all treatmentsdiffer from each other at T1 and T2.
260 6.71 0.001 T1 ≠ T2209 2.41 0.003 T1 ≠ T2 in 5 m and 5mT;
T0 = T1 in 15 m and 15mT
01*** b : 5 m N 2 m 3.25 0.003 2 m ≠ 5 m for EC but not for LC00*** b: LC N EC 30.14 0.001 EC ≠ LC in both depths,
Ectocarpales dominated the ECs.03*** b : 5 m N 2 m for EC,
5 m = 2 m for LC1.768 0.126
360 0.596 0.828000*** J : LC N EC
C :LC N ECB : LC N EC
54.868 0.001 EC ≠ LC in both depth;Ectocarpales dominated the ECs.
381 1.149 0.329
78 A. Fricke et al. / Journal of Experimental Marine Biology and Ecology 461 (2014) 73–84
3.3. Cross-depth transplantation experiment
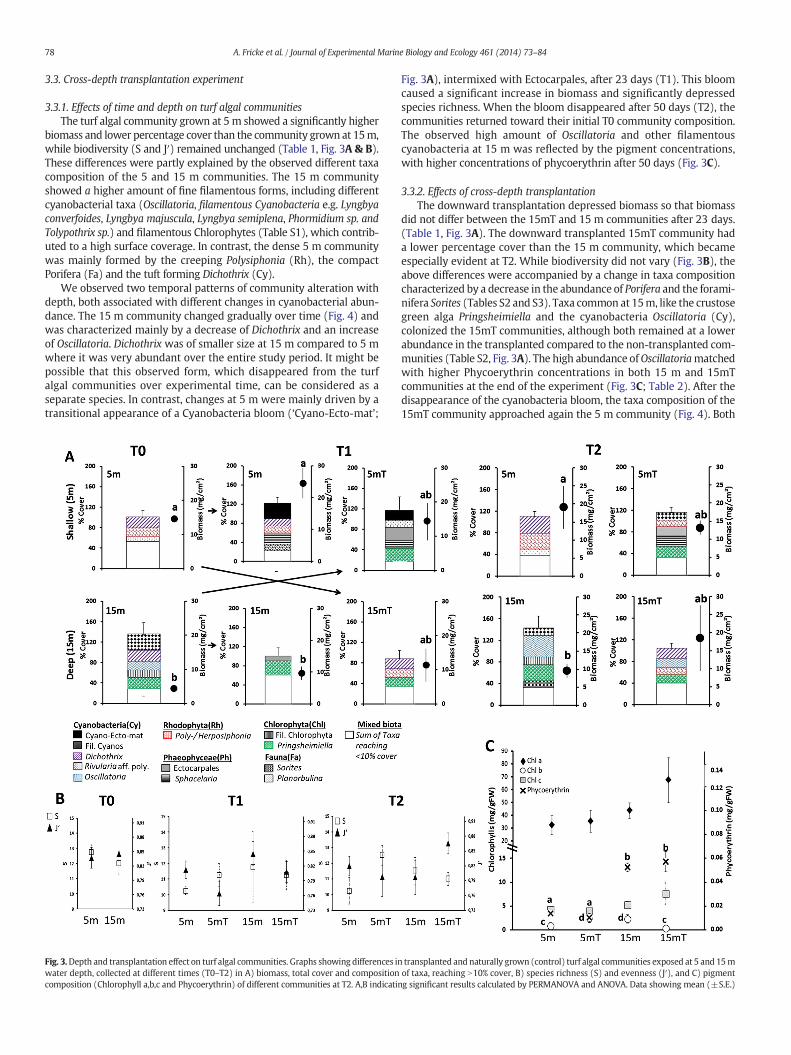
3.3.1. Effects of time and depth on turf algal communitiesThe turf algal community grown at 5m showed a significantly higher
biomass and lower percentage cover than the community grown at 15m,while biodiversity (S and J′) remained unchanged (Table 1, Fig. 3A & B).These differences were partly explained by the observed different taxacomposition of the 5 and 15 m communities. The 15 m communityshowed a higher amount of fine filamentous forms, including differentcyanobacterial taxa (Oscillatoria, filamentous Cyanobacteria e.g. Lyngbyaconverfoides, Lyngbya majuscula, Lyngbya semiplena, Phormidium sp. andTolypothrix sp.) and filamentous Chlorophytes (Table S1), which contrib-uted to a high surface coverage. In contrast, the dense 5 m communitywas mainly formed by the creeping Polysiphonia (Rh), the compactPorifera (Fa) and the tuft forming Dichothrix (Cy).
We observed two temporal patterns of community alteration withdepth, both associated with different changes in cyanobacterial abun-dance. The 15 m community changed gradually over time (Fig. 4) andwas characterized mainly by a decrease of Dichothrix and an increaseof Oscillatoria. Dichothrix was of smaller size at 15 m compared to 5 mwhere it was very abundant over the entire study period. It might bepossible that this observed form, which disappeared from the turfalgal communities over experimental time, can be considered as aseparate species. In contrast, changes at 5 m were mainly driven by atransitional appearance of a Cyanobacteria bloom (‘Cyano-Ecto-mat’;
Fig. 3.Depth and transplantation effect on turf algal communities. Graphs showing differences inwater depth, collected at different times (T0–T2) in A) biomass, total cover and compositioncomposition (Chlorophyll a,b,c and Phycoerythrin) of different communities at T2. A,B indicatin
Fig. 3A), intermixed with Ectocarpales, after 23 days (T1). This bloomcaused a significant increase in biomass and significantly depressedspecies richness. When the bloom disappeared after 50 days (T2), thecommunities returned toward their initial T0 community composition.The observed high amount of Oscillatoria and other filamentouscyanobacteria at 15 m was reflected by the pigment concentrations,with higher concentrations of phycoerythrin after 50 days (Fig. 3C).
3.3.2. Effects of cross-depth transplantationThe downward transplantation depressed biomass so that biomass
did not differ between the 15mT and 15 m communities after 23 days.(Table 1, Fig. 3A). The downward transplanted 15mT community hada lower percentage cover than the 15 m community, which becameespecially evident at T2. While biodiversity did not vary (Fig. 3B), theabove differences were accompanied by a change in taxa compositioncharacterized by a decrease in the abundance of Porifera and the forami-nifera Sorites (Tables S2 and S3). Taxa common at 15m, like the crustosegreen alga Pringsheimiella and the cyanobacteria Oscillatoria (Cy),colonized the 15mT communities, although both remained at a lowerabundance in the transplanted compared to the non-transplanted com-munities (Table S2, Fig. 3A). The high abundance ofOscillatoriamatchedwith higher Phycoerythrin concentrations in both 15 m and 15mTcommunities at the end of the experiment (Fig. 3C; Table 2). After thedisappearance of the cyanobacteria bloom, the taxa composition of the15mT community approached again the 5 m community (Fig. 4). Both
transplanted and naturally grown (control) turf algal communities exposed at 5 and 15mof taxa, reaching N10% cover, B) species richness (S) and evenness (J′), and C) pigmentg significant results calculated by PERMANOVA and ANOVA. Data showing mean (±S.E.)

Fig. 4. Differences between transplanted and non-transplanted communities. Non-metric multidimensional scaling (nMDS) of the changes in taxa composition of transplanted (squares)and none-transplanted (circles) communities exposed 5 and 15 m water depths, investigated at different times (T0–T2). Arrows indicate direction of communities over time, whereasdashed arrows indicate the direction of community after transplantation event. Dashed line encircles communities proven as not significantly different in PERMANOVA. Percentagesimilarity, from pairwise test, is given for every significant pairwise comparison. Data showing averages of communities (n = 4).
79A. Fricke et al. / Journal of Experimental Marine Biology and Ecology 461 (2014) 73–84
the 5m and 15mT communities were characterized by a high content ofthe cyanobacteria Dichothrix and the red algae Poly/Herposiphonia, sug-gesting that these taxa are resistant to increasing depth (i.e. decreasinglight).
In contrast, the upward transplantation caused an increase inbiomass so that biomass did not differ between the 5mT and 5 m com-munities at T1. Changes in taxa composition in the upward transplanta-tion when compared to the original T0 15 m included a decreasein Oscillatoria, filamentous cyanobacteria and Dichothrix (althoughthe latter were abundant in the 5 m communities), an increase inEctocarpales and a temporary (T1 only) increase in Rivularia aff. polyotis(Cy) together with the Cyano-Ecto-mat (Table S3). The transient overalldecrease in filamentous cyanobacteria and Oscillatoria at both 5 mand 5mT communities might be due to the cyanobacterial bloom. Taxatypical to shallow depths such as Poly/Herposiphonia succeeded in colo-nizing the transplanted communities in high numbers, while Dichothrixshowed only sporadic appearance. The crustose Pringsheimiella and cer-tain less abundant filamentous Chlorophyta (Table S2, Fig. 3A) survivedthe upward transplantation, suggesting that these taxa are resistant todecreasing depth (i.e. increasing light). This resulted in a similar highamount of Chlorophycean in the transplanted 5mT and 15m communi-ties at T1 and T2. This pattern was also reflected by higher concentra-tions of Chl b in 5mT and 15 m communities (Fig. 3C).
Table 2Differences in pigment composition of communities exposed to different water depths (Tranconcentrations of Chlorophyll a (Chla) and different ratios of pigments related to Chla: Chloropbetween treatments are identified using Tukey's test of honest significant difference (HSD) anLeven's test.
Chl a Chlb/Chla Chlc/Ch
N° Factor df F p F p F
1 Depth (D) 1 1.93 0.193 0.03 0.864 0.192 Transplant (T) 1 0.42 0.529 0.00 0.987 0.113 D x T 1 1.53 0.703 6.79 0.024 0.18
Res 11
Bold values are statistically significant.
3.4. UV experiment
3.4.1. Effects of community age and water depthStrong differences were found between the ECs and LCs communi-
ties (Table 1, Figs. 5 and 6). At both 2m and 5mdepths, the LCs showedsignificantly higher cover and biomass than the ECs. In terms of biodi-versity, the LCs showed a higher evenness than the ECs, but no differ-ence in species richness. This pattern was caused by a dominance ofEctocarpales on the ECs.
Biodiversity (S, J′) and cover were not affected by depth, regardless ofcommunity age. However, the ECs had a significantly higher biomass at5 m than at 2m,whereas no difference in biomass or community compo-sition between depths was found for the LCs. The biomass difference ofthe ECs correlated with a difference in taxa composition. The 5 m ECsweremainly composed of filamentous forms (e.g. Ectocarpales), whereasthe 2 m ECs contained a higher amount of crustose forms (e.g. crustosePhaeophyceae). Those crusts contained discs of phaeophyceaen cells(Ectocarpales or Sphacelariales).
Furthermore, differenceswere found in pigment contents of the ECs,with increased concentrations of chlorophyll b, lutein and zeaxanthin inthe shallower 2m treatments (Table 3, Fig. 7). Furthermore, the ECs hada significantly higher level in UV-B absorbing compounds at 2m than at5 m, indicating a potential depth dependant UV-B effect.
splant experiment). Overview of analysis of variance (ANOVA, F) testing differences inhyll b (Chlb), Chlorophyll c (Chl c), Phycoerythrin (Ery) and Phycocyanin (Cy). Differencesd are given under each significant difference. Homogeneity of variances was tested using
la Ery/Chla Cy/Chla Conclusions
p F p F p
0.671 10.51 0.008 0.96 0.347 15 m N 5 m0.741 0.01 0.928 1.05 0.3280.683 0.35 0.563 0.11 0.743 15 m, 5mT N 5 m, 15mT
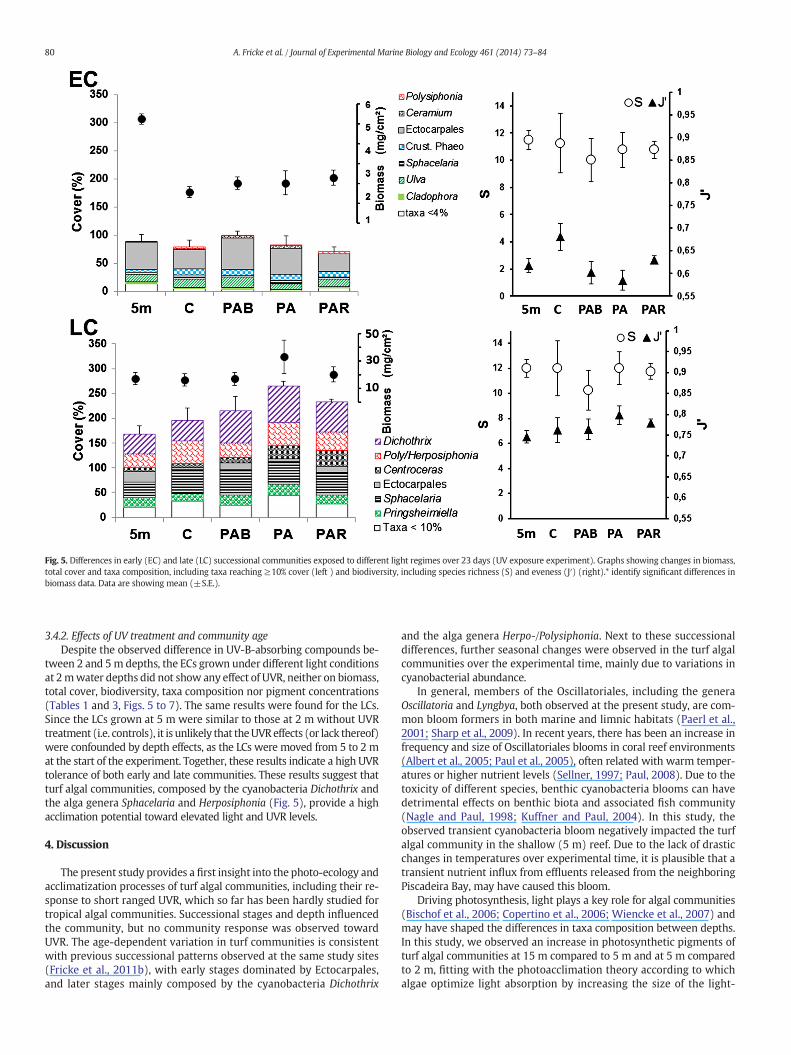
Fig. 5. Differences in early (EC) and late (LC) successional communities exposed to different light regimes over 23 days (UV exposure experiment). Graphs showing changes in biomass,total cover and taxa composition, including taxa reaching ≥10% cover (left ) and biodiversity, including species richness (S) and eveness (J′) (right).* identify significant differences inbiomass data. Data are showing mean (±S.E.).
80 A. Fricke et al. / Journal of Experimental Marine Biology and Ecology 461 (2014) 73–84
3.4.2. Effects of UV treatment and community ageDespite the observed difference in UV-B-absorbing compounds be-
tween 2 and 5m depths, the ECs grown under different light conditionsat 2mwater depths did not show any effect of UVR, neither on biomass,total cover, biodiversity, taxa composition nor pigment concentrations(Tables 1 and 3, Figs. 5 to 7). The same results were found for the LCs.Since the LCs grown at 5 m were similar to those at 2 m without UVRtreatment (i.e. controls), it is unlikely that theUVReffects (or lack thereof)were confounded by depth effects, as the LCs were moved from 5 to 2 mat the start of the experiment. Together, these results indicate a high UVRtolerance of both early and late communities. These results suggest thatturf algal communities, composed by the cyanobacteria Dichothrix andthe alga genera Sphacelaria and Herposiphonia (Fig. 5), provide a highacclimation potential toward elevated light and UVR levels.
4. Discussion
The present study provides a first insight into the photo-ecology andacclimatization processes of turf algal communities, including their re-sponse to short ranged UVR, which so far has been hardly studied fortropical algal communities. Successional stages and depth influencedthe community, but no community response was observed towardUVR. The age-dependent variation in turf communities is consistentwith previous successional patterns observed at the same study sites(Fricke et al., 2011b), with early stages dominated by Ectocarpales,and later stages mainly composed by the cyanobacteria Dichothrix
and the alga genera Herpo-/Polysiphonia. Next to these successionaldifferences, further seasonal changes were observed in the turf algalcommunities over the experimental time, mainly due to variations incyanobacterial abundance.
In general, members of the Oscillatoriales, including the generaOscillatoria and Lyngbya, both observed at the present study, are com-mon bloom formers in both marine and limnic habitats (Paerl et al.,2001; Sharp et al., 2009). In recent years, there has been an increase infrequency and size of Oscillatoriales blooms in coral reef environments(Albert et al., 2005; Paul et al., 2005), often related with warm temper-atures or higher nutrient levels (Sellner, 1997; Paul, 2008). Due to thetoxicity of different species, benthic cyanobacteria blooms can havedetrimental effects on benthic biota and associated fish community(Nagle and Paul, 1998; Kuffner and Paul, 2004). In this study, theobserved transient cyanobacteria bloom negatively impacted the turfalgal community in the shallow (5 m) reef. Due to the lack of drasticchanges in temperatures over experimental time, it is plausible that atransient nutrient influx from effluents released from the neighboringPiscadeira Bay, may have caused this bloom.
Driving photosynthesis, light plays a key role for algal communities(Bischof et al., 2006; Copertino et al., 2006; Wiencke et al., 2007) andmay have shaped the differences in taxa composition between depths.In this study, we observed an increase in photosynthetic pigments ofturf algal communities at 15 m compared to 5 m and at 5 m comparedto 2 m, fitting with the photoacclimation theory according to whichalgae optimize light absorption by increasing the size of the light-

Fig. 6. Non-metric multidimensional scaling (nMDS), showing differences between lateand early successional communities (ECs, LCs). A) Graph showing differences betweencommunities exposed to different water depths (2 and 5 m). B) Graph shows differencesbetween communities exposed to different light levels (open, PAB, PA, P). Data showingaverages of communities (n = 4).
81A. Fricke et al. / Journal of Experimental Marine Biology and Ecology 461 (2014) 73–84
harvesting complex and the ratio of photosynthetic to photoprotectivepigments (Falkowski and Owens, 1978; Sukenik and Shelef, 1984).Due to the high transparency, UV radiation penetrated deeply in thewater column, with 10% of UV-B surface radiation measured at 15 m.To highlight the high penetration depth and potential damaging effectof UVR, Visser et al. (2002) demonstrated that UVR caused considerableDNA damage in bacterioplancton down to 5mwater depth and affectedits production (leucine and thymidine incorporation) to a depth of 10mclose to our study site. In addition, Boelen et al. (2000) found UV-B-induced DNA damage, measured as cyclobutane pyrimidine dimers(CPDs), in phyto-and bacterioplankton sampled from 0 to 15 m waterdepths at several locations in Curaçao. In our study, we found an in-crease in UV-B absorbing compounds from 5 to 2 m depth. Thus, aminor reduction in UV-B radiation (ca. 8%) caused a significant increaseof UV-B absorbing compounds over a depth range of 3 m.
Despite these findings, no UV effect was found on the turf algalcommunities in this study. Already in the pre-experiment done in 5 mwater depth a year before, we did not observe any effect of UV radiation
Table 3Differences in pigment composition of ECs exposed to different light regimes (UV experimenChlorophyll a (Chla) and different ratios of pigments related to Chla: Chlorophyll b (Chlb/Chla), Caswell as differences in light absorbing compounds,measured at differentwavelengths: ultravioradiation (UVA; 320–400 nm), photosynthetically active radiation (PAR; 400–700 nm). HomKruskal Wallis test.
df Chla Chlb/ Chla Chlc/ Chla Fuc/ Chla Lut
a) Effect of depth F p F p F p F p FDepth (D) 1 1.33 0.25 13.97 0.01 4.86 0.07 4.86 0.06 6.6Res 6
b) Effect of UVTreatment (T) 3 0.95 0.45 1.22 0.35 0.93 0.45 0.93 0.46 1.5Res 11
Bold values are statistically significant.
on turf algal community composition or structure over a period of54 days (Fricke, 2011). Conducting a shorter, but more UV exposed, ex-periment of 22 days at the same site and season,we did not observe anyalteration in grown communities (LCs), neither by transplantation, norby further UV exposure. Taken together, this indicates that turf algalcommunities have a high tolerance and acclimatization potential.
However, thisfinding contradicts the results of amesocosm study onCaribbean turf algae (Santas et al., 1998). In their study, the recruitmentof turf algae was investigated under different light conditions (PAR +UV-A + high UV-B, PAR + UV-A + low UV-B and PAR) for a period of42 days, followed by a period of 35 days with reversal treatments tostudy the recovery of established communities. Already after 21 days,this study showed a strong UV-B effect, with a decrease in biomassand change of communities, whereas no effects on established commu-nities (N6 weeks old) were observed.
In order to understand the discrepancy between their and our study,several factors need to be considered. First, the two studies were con-ducted at different depths. Santas' mesocosm study focused on theupper 3 cm (i.e. intertidal zone), while our study was performed onthe shallow coral reef environment and investigated subtidal communi-ties naturally exposed to a lower, but constant irradiance of UVR. Sec-ond, a mesocosm is an artificial environment. The light regimes werespectrally fixed and not based on the environmental light levels whichnaturally vary throughout time as shown in our study. Third, althoughthe algal turf of the mesocosm experiment showed comparable taxacomposition to our field experiment (i.e., predominance of two filamen-tous taxa Ectocarpales and Ulva), different genera were involved in thedifferent studies. As Santas reported mainly the presence of Asteronema(Ectocarpus) rhodochortonoides (Børgesen) D.G.Müller & E.R.Parodi1994, the Ectocarpales in our study comprised mainly species of thegenus Feldmannia, e.g. Feldmannia irregularis (Kützing) G.Hamel 1939,Feldmannia indica (Sonder) Womersley & A.Bailey 1970, Feldmanniamitchelliae (Harvey) H.-S.Kim 2010. Different genera within theEctocarpales (e.g. Asterocladon, Ectocarpus, Pylaiella) have been ob-served to be sensitive against changes in light conditions and UV-B indifferent studies and latitudes (Fricke et al., 2011a; Santas et al., 1998;Wahl et al., 2004). The observed high UV tolerance in the Ectocarpalesat our site could be related to genera specific characteristics.
Genera characteristics may play a key role in turf algal communities,which in addition to light also have to copewith different depth-relatedenvironmental factors, like variations in water currents (Fricke et al.,2011b) or herbivory (De Ruyter van Steveninck and Breeman, 1987).In this study, we observed depth-related differences in taxa and theirresponse to changing depths. These differences could be explainedby differential morphological plasticity in algae (Van Den Hoeket al., 1995). Benthic macroalgae have complex heteromorphic lifecycles (Dethier, 1981; Parente et al., 2003) and a change in life stages(e.g. gametophyte, sporophyte) is a common strategy to adapt tochanging environmental factors, such as seasonality (Dethier, 1981),herbivory (Lubchenco and Cubit, 1980) and light (Zupan and West,
t). Overview of analysis of variance (ANOVA, F) testing differences in concentrations ofhlorophyll c (Chlc/chla), Fucoxanthin (Fuc/chla), Lutein (Lut/chla), Zeaxanthin (Zea/chla),let radiation (UV; 280–400 nm), ultraviolet-B radiation (UVB; 280–320 nm), ultraviolet-Aogeneity of variances was tested using Cochran's test. Underlines indicate analyses with
/ Chla Zea/ Chla UV UVB UVA PAR
p F p F p F p F p F p4 0.04 9.35 0.02 1.35 0.29 20.05 0.00 0.30 0.60 0.37 0.56
0 0.27 0.64 0.61 0.47 0.71 0.41 0.75 0.47 0.71 0.29 0.83

Fig. 7. Differences in chlorophylls (A), xanthophylls (B) and UV absorbing compounds (C) of ECs (UV experiment), exposed to different water depths (2 and 5 m) and light conditions(PAB, PA, P). Chla = Chlorophyll a, Chl b = Chlorophyll b, Chl c = Chlorophyll c, Fuco = Fucoxanthin, Lut = Lutein and Zea = Zeaxanthin; * indicates a significant differences indepth comparison (see Table 3). Dotted lines mark the treatments used for comparisons between 2 (C) and 5 m water depths. Data showing mean (±S.E.).
82 A. Fricke et al. / Journal of Experimental Marine Biology and Ecology 461 (2014) 73–84
1990). In addition, variable phenotypic responses have been describedin benthic algae (Lewis et al., 1987; Parodi and Cáceres, 1991). It is plau-sible that the Ectocarpales, which commonly dominates turf algal com-munities in early successional stages (Fricke et al., 2011b), are capable ofsuch adaptations.
The depth-dependent differences in later successional stages in thisstudy are consistent with a prior study (Fricke et al., 2011b) whereinthe community at 5 m depth was dominated by the tuft formingcyanobacteria Dichothrix and the red algae Poly-/Herposiphonia, whilethe more filamentous community at 15 m was composed of differentfine filamentous cyanobacteria and green algae. Here we distinguishedsensitive taxa, which decreased in abundances and vanished from thecommunities over time; and tolerant taxa, able to acclimate to changing
conditions. For the tolerant taxa, beside Pringsheimiella and Poly-/Herposiphonia, the heterocyst bearing Dichothrix was particularlyremarkable. Dominating turf algal communities at shallow depths, thistaxon showed high resistance during depth transplantation and wasnot affected by the exposure to increased light and UVR levels. Bearingbasal and intercalary heterocysts, this taxon is able to fix nitrogen(Wiebe et al., 1975). Furthermore, its thallus consists of single filaments,trichoms, embedded in an extracellular polysaccharide sheet rich inphotoprotective substances, such as the UV absorbing sheet pigmentscytonemin (Singh et al., 2010; Sinha and Häder, 2008). Its photo-protective potential might also explain the taxa tolerance towardupward transplantation and further UV exposure in the UV experiment.Due to its tuft or mat-like growth form, it can increase its self-shading

83A. Fricke et al. / Journal of Experimental Marine Biology and Ecology 461 (2014) 73–84
capacity and may provide a protective canopy for other adjacent orunderstory species (Reed and Foster, 1984) that might migrate intothe protective structure. Different studies investigating the potentialeffect of UV radiation on the community level at different latitudes indi-cate the importance of community age toward UV sensitivity due to theactivation of morphological and chemical UVR protection (Fricke et al.,2011a; Wahl et al., 2004; Zacher et al., 2007). Thus, corresponding toour observations, UVR resistant taxa, like Dichothrix, might provideshelter to other UV sensitive species thanks to their canopy formingstructure in later successional stages (Molis and Wahl, 2009; Moliset al., 2003).
In contrast to the bottomfixed biota, someof the observed taxawerecapable of active movements, which may allow them to escape fromunsuitable conditions. Thus the filamentous cyanobacteria Oscillatoriais capable of oscillating movements, which allows it to migrate withinthe algal turf (Geitler, 1982). Showing high abundances below 5 mwater depth (Fricke et al., 2011b), it quickly disappeared after the trans-plantation to shallower waters. In fact Nadeau et al. (1999) on a hyper-salinemicrobial mat showed a high sensitivity of Oscillatoria sp. towardelevated light regimes, especially UV radiation, measured in a decreaseof photosynthetic driven incorporation of NAHCO3 and an elevateddownward migration.
The investigation of turf algal communities naturally grownunder different light conditions in the coral reef has revealed twomain conclusions. First, contrary to our assumptions, turf algal com-munities grown and transplanted in the shallow reef (2 m) showedno difference between UV exposed and UV free conditions, provinga strong adaptation potential to high light and UVR levels. The highUV tolerance of the turf algal communities, regardless of their suc-cessional stage might play a crucial role for the future of coral reefsas it may confer a competitive advantage over other more UV sensi-tive coral reef biota, including corals (Banaszak and Lesser, 2009;Baruch et al., 2005; Wellington and Fitt, 2003). As a consequence, thechange and degradation of coral reefs worldwide (Hoegh-Guldberget al., 2007;Mumby et al., 2007) could be intensified under recent prog-nostic climate change scenarios.
Second, turf algae assemblages are highly variable in species compo-sition. The observed natural alterations due to successional and seasonalprocesses (e.g. cyanobacteria bloom), as well as the experimentallyinitiated responses to different environmental conditions (up- anddown-transplantations) prove a high acclimatization potential ofturf algal communities. Turf algae are not simply a diverse assem-blage of phylogenetic groups but dynamic communities, which areshaped by intra-taxon characteristics, inter-taxa relations and environ-mental factors, e.g. light quantity. Turf algae are commonly pooled asa functional group (Barott et al., 2011; Steneck and Simon, 2001;Vermeij et al., 2010). However, to further understand their ecologyand function on coral reefs, we recommend a more specific approach,as turf algae are a complex community on their own.
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.jembe.2014.07.017.
Acknowledgments
For kind assistance with diving and field work we thank DanielleLommen, Julia Kesting, and Daniel Nugues. We are grateful to the staffof the CARMABI foundation: Dr. Mark J.A. Vermeij, Dr Adolphe Debrot,Carlos Winterdaal, Oscar Frans and Sislin Rosalia for friendly support;as well as to the workshop of the University of Bremen for their expertassistance. For help in species identification,we gratefully thank TamaraV. Titlyanova. This study was supported by the Bremen InternationalGraduate School for Marine Sciences (GLOMAR) funded by the GermanResearch Foundation (DFG)within the frame of the Excellence Initiativeby the German federal and state governments to promote science andresearch at German universities. [ST]
References
Albert, S., O'Neil, J.M., Udy, J.W., Ahern, K.S., O'Sullivan, C.M., Dennison, W.C., 2005.Blooms of the cyanobacterium Lyngbya majuscula in coastal Queensland, Australia:disparate sites, common factors. Mar. Pollut. Bull. 51, 428–437.
Arar, E.J., Collins, G.B., 1997. In vitro determination of chlorophylls a, b, c1 + c2 andpheopigments inmarine and freshwater algae by visible spectrophotometry. NationalExposure Research Laboratory, Office of Research and Development, U. S. Environ-mental Protection Agency, Method 446.0, Revision 1.2.
Banaszak, A.T.,Lesser,M.P., 2009. Effects of solar ultraviolet radiation on coral reef organisms.Photochem. Photobiol. Sci. 8 (9), 1276–1294.
Barott, K.L., Rodriguez-Brito, B., Janouskovec, J., Marhaver, K.L., Smith, J.E., Keeling, P.,Rohwer, F.L., 2011. Microbial diversity associated with four functional groups of ben-thic reef algae and the reef-building coral Montastraea annularis. Environ. Microbiol.13 (5), 1192–1204.
Baruch, R.,Avishai, N.,Rabinowitz, C., 2005. UV incites diverse levels of DNA breaks in dif-ferent cellular compartments of a branching coral species. J. Exp. Biol. 208 (Pt 5),843–848.
Beer, S., Eshel, A., 1985. Determining phycoerythrin and phycocyanin concentrations inaqueous crude extracts of red algae. Aust. J. Mar. Freshw. Res. 36, 785–792.
Benayahu, Y.,Loya, Y., 1981. Competition for space among coral-reef sessile organisms atEilat, Red-Sea. Bull. Mar. Sci. 31 (3), 514–522.
Birrell, C.L.,McCook, L.J.,Willis, B.L., 2005. Effects of algal turfs and sediment on coralsettlement. Mar. Pollut. Bull. 51 (1–4), 408–414.
Bischof, K., Gõmez, I.,Molis, M.,Hanelt, D.,Karsten, U., Lüder, U.,Roleda, M.Y., Zacher, K.,Wiencke, C., 2006. Ultraviolet radiation shapes seaweed communities. Rev. Environ.Sci. Biotechnol. 5, 141–166.
Boelen, P., Boer, M.K.d.,Kraay, G.W.,Veldhuis, M.J.W., Buma, A.G.J., 2000. UVBR-inducedDNA damage in natural marine picoplankton assemblages in the tropical AtlanticOcean. Mar. Ecol. Prog. Ser. 193, 1–9.
Bornet, É., Flahault, C., 1888. Revision des Nostocacées hétérocystées contenues dans lesprincipaux herbiers de France (quatrième et dernier fragment). Ann. Sci. Nat., Bot.7, 177–262.
Bray, J.R.,Curtis, J.T., 1957. An ordination of upland forest communities of southern Wis-consin. Ecol. Mono. 27, 325–349.
Carpenter, R.,Williams, S., 2007. Mass transfer limitation of photosynthesis of coral reefalgal turfs. Mar. Biol. 151 (2), 435–450.
Chapman, A.R.O., Waters, D.A., 1992. Epi- and Endobiotic Chlorophytes Algae andSymbioses: Plants, Animals, Fungi, Viruses, Interactions Explored. Biopress Ltd., Bristol,pp. 619–639.
Copertino, M., Connell, S.D., Cheshire, A., 2005. The prevalence and production of turf-forming algae on a temperate subtidal coast. Phycologia 44 (3), 241–248.
Copertino, M.S., Cheshire, A.,Watling, J., 2006. Photoinhibition and photoacclimation ofturf algal communities on a temperate reef, after in situ transplantation experiments.J. Phycol. 42, 580–592.
De Ruyter van Steveninck, E.D.,Breeman, A., 1987. Deep water populations of Lobophoravariegata (Phaeophyceae) on the coral reef of Curagao: influence of grazing anddispersal on distribution patterns. Mar. Ecol. Prog. Ser. 38, 241–250.
Dethier, M., 1981. Heteromorphic algal life histories: the seasonal pattern and response toherbivory of the brown crust, Ralfsia californica. Oecologia 49 (3), 333–339.
Diaz-Pulido, G.,McCook, L.J., 2002. The fate of bleached corals: patterns and dynamics ofalgal recruitment. Mar. Ecol. Prog. Ser. 232, 115–128.
Diaz-Pulido, G.,McCook, L.J., 2004. Effects of live coral, epilithic algal communities andsubstrate type on algal recruitment. Coral Reefs 23 (2), 225–233.
Diaz-Pulido, G.,McCook, L.J.,Dove, S.,Berkelmans, R.,Roff, G.,Kline, D.I.,Weeks, S.,Evans, R.D.,Williamson, D.H.,Hoegh-Guldberg, O., 2009. Doom and boom on a resilient reef:climate change, algal overgrowth and coral recovery. PLoS ONE 4 (4), e5239.
Dobretsov, S.V., Qian, P.Y.,Wahl, M., 2005. Effect of solar ultraviolet radiation on theformation of shallow, early successional biofouling communities in Hong Kong.Mar. Ecol. Prog. Ser. 290, 55–65.
Engelen, A.H.,Breeman, A.M.,Olsen, J.L.,Stam, W.T.,Aberg, P., 2005. Life history flexibilityallows Sargassum polyceratium to persist in different environments subjected tostochastic disturbance events. Coral Reefs 24 (4), 670–680.
Falkowski, P.G.,Owens, T.G., 1978. Effects of Light-Intensity on Photosynthesis and DarkRespiration in 6 Species of Marine-Phytoplankton. Mar. Biol. 45 (4), 289–295.
Fleischmann, E., 1989. The measurement and penetration of ultraviolet-radiation intotropical marine water. Limnol. Oceanogr. 34 (8), 1623–1629.
Fricke, A., 2011. Succession patterns of turf algal communities in a fringing coral reef.(Ph.D. Thesis) University of Bremen, Germany, p. 281.
Fricke, A.,Molis, M.,Wiencke, C.,Valdivia, N.,Chapman, A., 2011a. Effects of UV radiation onthe structure of Arctic macrobenthic communities. Polar Biol. 1–15.
Fricke, A., Teichberg, M., Beilfuss, S., Bischof, K., 2011b. Succession patterns in algal turfvegetation on a Caribbean coral reef. Bot. Mar. 54 (2), 111–126.
Fricke, A., Titlyanova, T.,Nugues, M.M.,Bischof, K., 2011c. Depth-related variation in epi-phytic communities growing on the brown alga Lobophora variegata in a Caribbeancoral reef. Coral Reefs 30 (4), 967–973.
Geitler, L., 1982. On the knowledge of the Oscillatoriacea Pseudanabaena-Galeata(Cyanophyta). Plant Syst. Evol. 141 (2), 169–175.
Gleason, D.,Edmunds, P.,Gates, R., 2006. Ultraviolet radiation effects on the behavior andrecruitment of larvae from the reef coral Porites astreoides. Mar. Biol. 148 (3),503–512.
Haas, A.F.,Naumann, M.S.,Struck, U.,Mayr, C.,el-Zibdah, M.,Wild, C., 2010. Organic matterrelease by coral reef associated benthic algae in the Northern Red Sea. J. Exp. Mar.Biol. Ecol. 389 (1–2), 53–60.
Haas, A.F.,Wild, C., 2010. Composition analysis of organicmatter released by cosmopolitancoral reef-associated green algae. Aquat. Biol. 10 (2), 131–138.

84 A. Fricke et al. / Journal of Experimental Marine Biology and Ecology 461 (2014) 73–84
Hanelt, D.,Tüg, H.,Bischof, K.,Groß, C.,Lippert, H.,Sawall, T.,Wiencke, C., 2001. Light regimein an Arctic fjord: a study related to stratospheric ozone depletion as a basis for deter-mination of UV effects on algal growth. Mar. Biol. 138 (3), 649–658.
Hacker, S.D., Steneck, R.S., 1990. Habitat architecture and the abundance and body-size-dependent habitat selection of a phytal amphipod. Ecology 71 (6), 2269–2285.
Hoegh-Guldberg, O., Mumby, P.J., Hooten, A.J., Steneck, R.S., Greenfield, P., Gomez, E.,Harvell, C.D., Sale, P.F., Edwards, A.J., Caldeira, K., Knowlton, N., Eakin, C.M., Iglesias-Prieto, R., Muthiga, N., Bradbury, R.H., Dubi, A., Hatziolos, M.E., 2007. Coral reefsunder rapid climate change and ocean acidification. Science 318 (5857), 1737–1742.
Jeffrey, S.W.,Humphrey, G.F., 1975. New spectrophotometric equations for determiningchlorophylls a, b, c + c in higher plants, algae and natural phytoplankton. Biochem.Physiol. Pflanz. 167, 191–194.
Jompa, J.,McCook, L.J., 2003. Coral–algal competition:macroalgaewith different propertieshave different effects on corals. Mar. Ecol. Prog. Ser. 258, 87–95.
Karl, T.R., Trenberth, K.E., 2003. Modern global climate change. Science 302 (5651),1719–1723.
Kirk, J.T., 1994. Light and photosynthesis in aquatic ecosystems. Cambridge UniversityPress, New York.
Kramer, M.J.,Bellwood, D.R.,Bellwood, O., 2012. Cryptofauna of the epilithic algal matrixon an inshore coral reef, Great Barrier Reef. Coral Reefs 31 (4), 1007–1015.
Kuffner, I.B., 2001. Effects of ultraviolet radiation and water motion on the reef coralPorites compressa Dana: a flume experiment. Mar. Biol. 138 (3), 467–476.
Kuffner, I.B.,Paul, V.J., 2004. Effects of the benthic cyanobacterium Lyngbya majuscula onlarval recruitment of the reef corals Acropora surculosa and Pocillopora damicornis.Coral Reefs 23, 455–458.
Lewis, S.M.,Norris, J.N., Searles, R.B., 1987. The regulation of morphological plasticity intropical reef algae by herbivory. Ecology 68 (3), 636–641.
Littler, M.M.,Littler, D.S.,Brooks, B.L., 2010. The effects of nitrogen and phosphorus enrich-ment on algal community development: artificial mini-reefs on the Belize BarrierReef sedimentary lagoon. Harmful Algae 9, 255–263.
Lubchenco, J.,Cubit, J., 1980. Heteromorphic life histories of certain marine algae as adap-tations to variations in herbivory. Ecology 61 (3), 676–687.
McCook, L., 2001. Competition between corals and algal turfs along a gradient of ter-restrial influence in the nearshore central Great Barrier Reef. Coral Reefs 19 (4),419–425.
McCook, L.,Jompa, J.,Diaz-Pulido, G., 2001. Competition between corals and algae on coralreefs: a review of evidence and mechanisms. Coral Reefs 19 (4), 400–417.
McCook, L.J., 1999. Macroalgae, nutrients and phase shifts on coral reefs: scientific issuesand management consequences for the Great Barrier Reef. Coral Reefs 18 (4),357–367.
Molis, M., Lenz, M.,Wahl, M., 2003. Radiation effects along a UV-B gradient on speciescomposition and diversity of a shallow-water macrobenthic community in thewestern Baltic. Mar. Ecol. Prog. Ser. 263, 113–125.
Molis, M.,Wahl, M., 2009. Comparison of the impacts of consumers, ambient UV, andfuture UV-B irradiance on mid-latitudinal macroepibenthic assemblages. GlobalChange Biol. 15 (7), 1833–1845.
Mumby, P., 2009. Herbivory versus corallivory: are parrotfish good or bad for Caribbeancoral reefs? Coral Reefs 28 (3), 683–690.
Mumby, P.J.,Foster, N.L.,Fahy, E.A.G., 2005. Patch dynamics of coral reef macroalgae underchronic and acute disturbance. Coral Reefs 24 (4), 681–692.
Mumby, P.J.,Hastings, A.,Edwards, H.J., 2007. Thresholds and the resilience of Caribbeancoral reefs. Nature 450 (7166), 98–101.
Nadeau, T.-L.,Howard-Williams, C.,Castenholz, R.W., 1999. Effects of solar UV and visibleirradiance on photosynthesis and vertical migration of Oscillatoria sp. (Cyanobacteria)in an Antarctic microbial mat. Aquat. Microb. Ecol. 20 (3), 231–243.
Nagle, D.G., Paul, J., 1998. Chemical defense of a marine cyanobacterial bloom. JEMBE 225,29–38.
Nugues, M.M.,Bak, R.P.M., 2008. Long-term dynamics of the brown macroalga Lobophoravariegata on deep reefs in Curaçao. Coral Reefs 27 (2), 389–393.
Nugues, M.M.,Roberts, C.M., 2003. Partial mortality in massive reef corals as an indicatorof sediment stress on coral reefs. Mar. Pollut. Bull. 46 (3), 314–323.
Olabarria, C., Chapman, M.G., 2001. Comparison of patterns of spatial variation ofmicrogastropods between two contrasting intertidal habitats. Mar. Ecol. Prog. Ser.220, 201–211.
Paerl, H.W.,Fulton, R.S.,Moisander, P.H.,Dyble, J., 2001. Harmful freshwater algal blooms,with an emphasis on cyanobacteria. Sci. World J. 1, 76–113.
Parente, M.I., Neto, A.I., Fletcher, R.L., 2003. Morphology and life history of Scytosiphonlomentaria (Scytosiphonaceae, Phaeophyceae) from the Azores. J. Phycol. 39 (2),353–359.
Parodi, E.R.,Cáceres, E.J., 1991. Variation in number of apical ramifications and vegetativecell length in freshwater populations of Cladophora (Ulvophyceae, Chlorophyta).J. Phycol. 27 (5), 628–633.
Paul, V.J., 2008. Global warming and cyanobacterial harmful algal blooms. Cyanobacterialharmful algal blooms: State of the science and research needs. Adv. Exp. Med. Biol.619, 239–257.
Paul, V.,Thacker, R.,Banks, K.,Golubic, S., 2005. Benthic cyanobacterial bloom impacts thereefs of South Florida (Broward County, USA). Coral Reefs 24, 693–697.
Purcell, S.P.,Bellwood, D.B., 2001. Spatial patterns of epilithic algal and detrital resourceson a windward coral reef. Coral Reefs 20 (2), 117–125.
Reed, D.C., Foster, M.S., 1984. The effects of canopy shadings on algal recruitment andgrowth in a giant kelp forest. Ecology 65 (3), 937–948.
Santas, R.,Korda, A., Lianou, C., Santas, P., 1998. Community responses to UV radiation. 1.Enhanced UV-B effects on biomass and community structure of filamentous algalassemblages growing in a coral reef mesocosm. Mar. Biol. 131 (1), 153–162.
Sellner, K.G., 1997. Physiology, ecology, and toxic properties of marine cyanobacteriablooms. Limnol. Oceanogr. 42, 1089–1104.
Sharp, K., Arthur, K.E., Gu, L.,Ross, C.,Harrison, G., Gunasekera, S.P., Meickle, T.,Matthew, S.,Luesch, H., Thacker, R.W., Sherman, D.H., Paul, V.J., 2009. Phylogenetic and chemicaldiversity of three chemotypes of bloom-forming Lyngbya species (Cyanobacteria:Oscillatoriales) from reefs of Southeastern Florida. Appl. Environ. Microbiol. 75,2879–2888.
Shick, J.M.,Lesser, M.P.,Jokiel, P.L., 1996. Effects of ultraviolet radiation on corals and othercoral reef organisms. Global Change Biol. 2 (6), 527–545.
Shindell, D.T.,Rind, D., Lonergan, P., 1998. Increased polar stratospheric ozone losses anddelayed eventual recovery owing to increasing greenhouse-gas concentrations.Nature 392 (6676), 589–592.
Singh, S.P.,Hader, D.P., Sinha, R.P., 2010. Cyanobacteria and ultraviolet radiation (UVR)stress: mitigation strategies. Ageing Res. Rev. 9 (2), 79–90.
Sinha, R.P., Häder, D.-P., 2008. UV-protectants in cyanobacteria. Plant Sci. 174 (3),278–289.
Steneck, R.S.,Simon, A.L., 2001. Functional Groups Encyclopedia of Biodiversity. Elsevier,New York, pp. 121–139.
Sukenik, A., Shelef, G., 1984. Algal autoflocculation - verification and proposed mecha-nism. Biotechnol. Bioeng. 26 (2), 142–147.
Teichberg, M., Fricke, A., Bischof, K., 2013. Increased physiological performance of thecalcifying green macroalga Halimeda opuntia in response to experimental nutrientenrichment on a Caribbean coral reef. Aquat. Bot. 104, 25–33.
Titlyanov, E.,Titlyanova, T., 2008. Coral–algal competition on damaged reefs. Russ. J. Mar.Biol. 34 (4), 199–219.
Titlyanov, E.A.,Titlyanova, T.V.,Yakovleva, I.M.,Nakano, Y.,Bhagooli, R., 2005. Regenerationof artificial injuries on scleractinian corals and coral/algal competition for newlyformed substrate. J. Exp. Mar. Biol. Ecol. 323 (1), 27–42.
Tsay, S., Stamnes, K., 1992. Ultraviolet-radiation in the Arctic: the impact of potentialozone depletions and cloud effects. J. Geophys. Res. - Atmos. 97 (D8), 7829–7840.
Tuck, A., Davies, T., Hovde, S., Nogueralba, M., Fahey, D., Kawa, S., Kelly, K., Murphy, D.,Proffitt, M., Margitan, J., Loewenstein, M., Podolske, J., Strahan, S., Chan, K., 1992.Polar stratospheric cloud processed air and potential vorticity in the northern-hemisphere lower stratosphere at midlatitudes during winter. J. Geophys. Res. -Atmos. 97 (D8), 7883–7904.
Van den Hoek, C., 1969. Algal vegetation-types along the open coasts of Curaçao,Netherlands Antilles. Proceedings of the Koninklijke Nederlandse Akademie vanWetenschappen Series C Biological and Medical Sciences, pp. 537–577.
Van den Hoek, C.,Colijn, F.,Cortel-Breeman, A.M.,Wanders, J.B.W., 1972. Algal vegetation-types along the shores of inner bays and lagoons of Curaçao and of the lagoon Lac(Bonaire), Netherlands Antilles. Verhandelingen Koningklijke Nederlandse Akademievan Wetenschap, Afdeling Natuurkunde. 61, pp. 1–72.
Van den Hoek, C.,Cortel-Breeman, C.A.,Wanders, J.B.W., 1975. Algal zonation in the fring-ing coral reef of Curaçao, Netherlands Antilles, in relation to zonation of corals andgorgonians. Aquat. Bot. 1, 269–308.
Van den Hoek, C.,Breeman, A.M.,Bak, R.P.M.,Van Buurt, G., 1978. The distribution of algae,corals and gorgonians in relation to depth, light attenuation, water movement andgrazing pressure in the fringing coral reef of Curaçao, Netherlands Antilles. Aquat.Bot. 5, 1–46.
Van Den Hoek, C.,Mann, D.G., Jahns, H.M., 1995. Algae—An Introduction to Phycology.Cambridge University Press.
Vermeij, M.J.A.,van Moorselaar, I.,Engelhard, S.,Hörnlein, C.,Vonk, S.M.,Visser, P.M., 2010.The effects of nutrient enrichment and herbivore abundance on the ability of turfalgae to overgrow coral in the Caribbean. PLoS ONE 5 (12), e14312.
Visser, P.M., Poos, J.J., Scheper, B.B., Boelen, P., van Duyl, F.C., 2002. Diurnal variations indepth profiles of UV-induced DNA damage and inhibition of bacterioplankton pro-duction in tropical coastal waters. Mar. Ecol. Prog. Ser. 228, 25–33.
Wahl, M.,Molis, M.,Davis, A.,Dobretsov, S.,Dürr, S.T., Johansson, J., Kinley, J.,Kirugara, D.,Langer, M., Lotze, H.K., Thiel, M., Thomason, J.,Worm, B., Zeevi Ben-Yosef, D., 2004.UVR effects that come and go: a global comparison of marine benthic communitylevel impacts. Global Change Biol. 10 (12), 1962–1972.
Wanders, J.B.W., 1976. The role of benthic algae in the shallow reef of Curaçao (Nether-lands Antilles) I: Primary productivity in the coral reef. Aquat Bot 2, 235–270.
Wellington, G.M.,Fitt, W.K., 2003. Influence of UV radiation on the survival of larvae frombroadcast-spawning reef corals. Mar. Biol. 143 (6), 1185–1192.
Wiebe, W.J., Johannes, R.E.,Webb, K.L., 1975. Nitrogen fixation in a coral reef community.Science 188 (4185), 257–259.
Wiencke, C., Clayton, M.N., Gomez, I., Iken, K., Lüder, U.H., Amsler, C.D., Karsten, U.,Hanelt, D., Bischof, K.,Dunton, K., 2007. Life strategy, ecophysiology and ecology ofseaweeds in polar waters. Rev. Environ. Sci. Biotechnol. 6, 95–126.
Williams, S.L.,Carpenter, R.C., 1997. Grazing effects on nitrogen fixation in coral reef algalturfs. Mar. Biol. 130 (2), 223–231.
Zacher, K.,Hanelt, D.,Wiencke, C.,Wulff, A., 2007. Grazing and UV radiation effects on anAntarctic intertidal microalgal assemblage: a long-term field study. Polar Biol. 30 (9),1203–1212.
Zupan, J.R.,West, J.A., 1990. Photosynthetic responses to light and temperature of theheteromorphic marine alga Mastocarpus papillatus (Rhodophyta). J. Phycol. 26 (2),232–239.




















