
Lipid chain order and dynamics at different bilayer depths in liposomes of several...
12
Chemistry and Physics of LIPIDS ELSEVIER SCIENCE Chemistry and Physics of Lipids IRELAND 71 (1994) 61--72 Lipid chain order and dynamics at different bilayer depths in liposomes of several phosphatidylcholines studied by differential polarized phase fluorescence M.A. Tricerri, H.A. Gardat, R.R. Brennert Instituto de Investigaciones Bioquimicas de la Plata (IN1BIOLP), CONICET-UNLP, Facultad de Ciencias Mddicas, Calles 60 y 120, 1900 La Plata, Argentina (Received 13 October 1993; revision received 27 January 1994; accepted 27 January 1994) Abstract The influence of acyl chain length, double bond number and position on the structural and dynamic properties of phosphatidylcholine bilayers in the liquid crystalline state was studied. The range and rate of the rotation of 1,6- diphenyl-1,3,5-hexatriene and a set of n-(9-anthroyloxy)stearates (n = 2, 7 and 12), which are located at different depths in the lipid bilayers, were measured using differential polarized phase fluorometry in multilamellar liposomes of several unsaturated synthetic phosphatidylcholines. For the anthroyloxystearate probes, two rotational modes, 'in' and 'out' of the plane of the anthroyl aromatic ring, were partially resolved by measuring at different excitation wavelengths. The data obtained here indicate that in lecithins containing the same acyl chain in sn-1 and sn-2, the introduction of double bonds towards the deep interior of the lipid bilayer decreases the order in the external region of the bilayer and the viscous resistance to the rotation of all the probes tested. On the contrary, double bonds located towards the external region of the bilayer increase order and viscous resistance to the rotation of probes located in the external region. Increasing acyl chain length has different effects depending on the probe and on the rotational mode con- sidered. Measurements in mixed chain lecithins indicate that the influence of increasing unsaturation in sn-2 has dif- ferent consequences depending on the acyl group that is present in sn-l. Similarly, increasing acyl chain length in sn-I influences differently the bilayer properties depending on the fatty acid that is present in sn-2. Key words: Differential polarized phase fluorometry; 1,6-diphenyl-l,3,5-hexatriene; n-(9-anthroyloxy)stearic acids; Lipid chain order and dynamics; Phophatidylcholine bilayers 1. Introduction * Corresponding author. $The authors are members of the Carrera del Investigador Cientifico, Consejo National de Investigaciones Cientificas y T6cnicas (CONICET), Argentina. There is a large number ofreports in the litera- ture on the relationship between fatty acid com- position in membrane lipids and physical and 0009-3084/94/$07.00 © 1994 Elsevier Science Ireland Ltd. All rights reserved. SSDI 0009-3084(94)02303-M
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Lipid chain order and dynamics at different bilayer depths in liposomes of several...

Chemistry and Physics of
LIPIDS ELSEVIER SCIENCE Chemistry and Physics of Lipids
IRELAND 71 (1994) 61--72
Lipid chain order and dynamics at different bilayer depths in liposomes of several phosphatidylcholines studied by differential
polarized phase fluorescence
M.A. Tricerri, H.A. Gardat, R.R. B r e n n e r t
Instituto de Investigaciones Bioquimicas de la Plata (IN1BIOLP), CONICET-UNLP, Facultad de Ciencias Mddicas, Calles 60 y 120, 1900 La Plata, Argentina
(Received 13 October 1993; revision received 27 January 1994; accepted 27 January 1994)
Abstract
The influence of acyl chain length, double bond number and position on the structural and dynamic properties of phosphatidylcholine bilayers in the liquid crystalline state was studied. The range and rate of the rotation of 1,6- diphenyl-1,3,5-hexatriene and a set of n-(9-anthroyloxy)stearates (n = 2, 7 and 12), which are located at different depths in the lipid bilayers, were measured using differential polarized phase fluorometry in multilamellar liposomes of several unsaturated synthetic phosphatidylcholines. For the anthroyloxystearate probes, two rotational modes, 'in' and 'out ' of the plane of the anthroyl aromatic ring, were partially resolved by measuring at different excitation wavelengths. The data obtained here indicate that in lecithins containing the same acyl chain in sn-1 and sn-2, the introduction of double bonds towards the deep interior of the lipid bilayer decreases the order in the external region of the bilayer and the viscous resistance to the rotation of all the probes tested. On the contrary, double bonds located towards the external region of the bilayer increase order and viscous resistance to the rotation of probes located in the external region. Increasing acyl chain length has different effects depending on the probe and on the rotational mode con- sidered. Measurements in mixed chain lecithins indicate that the influence of increasing unsaturation in sn-2 has dif- ferent consequences depending on the acyl group that is present in sn-l. Similarly, increasing acyl chain length in sn-I influences differently the bilayer properties depending on the fatty acid that is present in sn-2.
Key words: Differential polarized phase fluorometry; 1,6-diphenyl-l,3,5-hexatriene; n-(9-anthroyloxy)stearic acids; Lipid chain order and dynamics; Phophatidylcholine bilayers
1. Introduction * Corresponding author. $The authors are members of the Carrera del Investigador Cientifico, Consejo National de Investigaciones Cientificas y T6cnicas (CONICET), Argentina.
There is a large number ofreports in the litera- ture on the relationship between fatty acid com- position in membrane lipids and physical and
0009-3084/94/$07.00 © 1994 Elsevier Science Ireland Ltd. All rights reserved. SSDI 0009-3084(94)02303-M

62 M.A. Tricerri et aL / Chem. Phys. Lipids 71 (1994) 61-72
functional membrane properties (reviewed in Ref. 1). Frequently, fatty acid composition is modified by diets or other treatments, but other composi- tional changes, e.g. in phospholipid classes, cho- lesterol or protein content, can take place in addi- tion to fatty acid pattern changes. Fat-free or es- sential fatty acid-deficient diets seem to change fatty acid composition without other composi- tional changes and were used to attempt to relate unsaturation level with membrane physical pro- perties [2-5]. The transversal distribution of dou- ble bonds was considered in some reports and related to acyl chain order and dymamics measured at different bilayer depths [5]. In some membranes it has been observed that in spite of large changes in fatty acid composition, membrane viscotropic properties remain unaltered. 1 A good understanding of the relationship between fatty acid composition and membrane properties requires studies in more simple systems as single lipid membranes.
In this work, the influence of acyl chain length, double bond number and position on the structur- al and dynamic properties of the membrane lipid matrix was studied in single phospholipid systems. The rotational diffusion of probes with different properties and location in the lipid bilayer was measured using differential polarized phase fluorometry in multilamellar liposomes (MLV) of different molecular species of phosphatidylcholine (PC) containing unsaturated acyl chains.
The interpretation of the fluorescence emission anisotropy of a reporter molecule dissolved in a lipid bilayer must include consideration of both, orientational constraint and resistance to the depolarizing rotation of the probe imposed by the surrounding lipid matrix. These two contributions are not resolved in steady state anisotropy measurements but may be distinguished by time- resolved measurements by nanosecond single photon counting after pulse excitation [6,7] or by differential polarized phase fluorometry [8,9]. As reporter probes we have used here 1,6-diphenyl- 1,3,5-hexatriene (DPH), a rigid molecule that senses almost all the lipid matrix and 2-, 7- and 12-
1. H.A. Garda, A.M. Bernasconi and R.R. Brenner, submitted (a).
(9-anthroyloxy) stearic acids (n-AS), whose fluor- escent anthroyl moiety, attached to a flexible acyl chain, is located at different depths from the sur- face to the centre of the lipid bilayer [10], as shown by quenching with KI [11] and recently with brominated and spin labelled lipids [12].
DPH is thought to be inserted into lipid bilayers with its longitudinal axes parallel to the acyl chains. The absorption and emission moments of DPH are almost parallel and aligned to the longi- tudinal axis [13]. Thus, rotation about the longitu- dinal axis does not depolarize the emitted light, and only the wobbling motion about the axis that lies in the plane of the membrane can be observed [6,14]. This wobbling motion of DPH is hindered in lipid bilayers [15]. On the other hand, the n-AS probes would be inserted into the membrane with the stearoyl chain parallel to the phospholipid acyl chain, with the long axis of the anthroyl ring on average almost perpendicular to the plane of the membrane, with little motion about the 9ster link- age [16]. The emission dipole lies in the plane of the anthracene ring perpendicular to its long axis [17]. Absorption moment also lies in the same plane but at an angle from the emission dipole that is dependent on the excitation wavelength [18]. By exciting at a wavelength near 320 nm, the observed fundamental anisotropy is near 0.1, corresponding to an angle of 45 ° between absorption and emis- sion moments, and only the motion 'out of the plane' of the anthracene ring contributes to the depolarization of the emission [18,19]. Two 'out of the plane' rotational modes are possible, i.e. around both the long and the short axis of the anthracene ring. Considering that phase 2 and time-resolved [18,20] measurements indicate that this 'out of the plane' motion of n-AS probes is completely unhindered in protein-free lipid bi- layers, and also that rotation about the short axis of the anthracene ring does not displace the emis- sion dipole, it was proposed that using this excita- tion wavelength only, the rotation about the long axis of the anthroyl moiety or, what is equivalent, about the stearoyl acyl chain would be observed. 3
2. H.A. Garda, A.M. Bernasconi and R.R. Brenner, submitted (b). 3. Ibid.

M.A. Tricerri et al./ Chem. Phys. Lipids 71 (1994) 61-72 63
On the other hand, using higher excitation wavelengths over the red edge of the excitation spectrum, both 'in the plane' and 'out of the plane' modes of rotation produce depolarization of the emitted light. In this case phase 4 and time- resolved [18,20] measurements indicate a residual anisotropy r® which would correspond to a hindered 'in the plane' wobbling of the anthroyl group. Here we have used a 322-nm excitation wavelength to measure the rate of the 'out of the plane' rotation about the stearoyl chain and 388- nm excitation to obtain an evaluation of the range and rate of the 'in the plane' wobbling of the n-AS probes.
2. Materials and methods
2.1. Materials
DPH was obtained from Aldrich Chemical Company (Milwaukee, WI, USA); a stock solution 2.0 mM in tetrahydrofuran was prepared. 2-, 7- and 12-AS were purchased from Molecular Pro- bes, Inc. (Junction City, OR, USA); stock solu- tions 2.0 mM in methanol were prepared. Phosphatidylcholines (dissolved in chloroform) were obtained from Avanti Polar Lipids (Alabaster, AL, USA).
Fatty acid composition of egg and soybean phosphatidylcholines (PC), as analyzed by gas- liquid chromatography were: Egg-PC (16:0: 33.4%; 16:1 (9): 1.2%; 18:0:11.2%; 18:1 (9): 32.1%; 18:2 (9,12): 16.3%; 20:4 (5,8,11,14): 3.1%; and other minor components: 2.7%). Soybean-PC (16:0: 14.9%; 18:0: 3.5%; 18:1 (9): 8.9%; 18:2 (9,12): 66.1%; 18:3 (9,12,15) 5.5%; and other minor com- ponents: 1.1%).
2.2. Multilamellar liposome preparation and la- belling
Multilamellar liposomes (0.25 mM in phospha- tidylcholine) were prepared in 20 mM sodium phosphate pH 7.1 using the procedure described by Kutchai et al. [21]. For labelling, liposome preparations were mixed with the same volume of
2.5 #M probe dispersions in 20 mM sodium phos- phate buffer pH 7.1, agitated and incubated at 40°C for 30 min. All procedures were carried out under an N2 atmosphere.
2.3. Fluorescence measurements
Steady-state fluorescence anisotropy (rs), flu- orescence lifetimes (¢) and differential polarized phase shift (A) were measured at 20°C with an SLM 4800 C phase-modulation spectrofluoro- meter (SLM Instruments, Inc., Urbana, IL, USA) essentially as described by Lakowicz et al. [19,22] with some modifications to apply the background correction previously described.5 Excitation wavelengths were 361 nm for DPH and 322 or 388 nm for 2-, 7- and 12-AS. The emitted light was passed through a sharp cut-off filter (Schott KV 389 far DPH or KV 418 for 2-, 7- and 12-AS).
Lifetime measurements were obtained with the exciting light amplitude-modulated at 18 MHz by a Debye-Sears modulator. After the filter, the emission light was passed through a Glan- Thompson polarizer oriented 55 ° to the vertical to eliminate the effects of Brownian motion [23]. The phase shift and demodulation of the emitted light relative to a reference of known fluorescence lifetime were determined and used to compute the phase lifetime (¢p) and the modulation lifetime (era) of the sample [24]. POPOP (1,4-bis(5- phenyloxazol-2-yl)benzene) in ethanol was used as standard. It has a fluorescent lifetime of 1.35 ns [25,26]. For the n-AS, lifetime was measured only at 388 nm, since we have previously observed that it is independent of the excitation wavelength. 6 For all the samples and probes measured, the dif- ferences between 7p and 7m were within the experi- mental error, indicating homogeneity in the fluorophore population [19,27]. The reported lifetime values ((T)) are the average between Tp and Tm.
The differential polarized phase shift (A) was determined according to Refs. 19 and 22, exciting with light modulated at 18 MHz and vertically polarized and measuring the phase difference be-
5. Ibid. 4. Ibid. 6. Ibid.

64 M.A. Tricerri et al . / Chem. Phys. Lipids 71 (1994)61-72
tween the parallel and perpendicular components of the emitted light. It was corrected for the background contribution. 7
Steady-state anisotropies (rs) were measured with the Debye-Sears modulator turned off. Fun- damental anisotropy values (r0) were previously estimated 8 to be 0.390 for DPH and 0.102 and 0.332 for the n-AS at 322 and 388-nm excitation wavelength, respectively. For r0 = 0.1 the angle between absorption and emission dipoles is 45 ° , and only the rotation 'out of the plane' of the an- thracene ring depolarizes the emitted light, while for r0 = 0.332 the absorption dipole is about 20 ° to the emission dipole and both rotational modes 'in' and 'out' of the anthracene ring plane are observed [18,201.
2.4. Analysis of fluorescence data
When a fluorescent molecule dissolved in an isotropic solvent is excited by a brief pulse of polarized light, the fluorescence anisotropy decays exponentially approaching zero. When the rota- tion of the fluorofore is hindered by the anisotropic environment, the anisotropy decays to a finite value (r**). A modified Perrin equation [28] takes this into account:
r s = r** + ((ro - r**)/(1 + 7/rr)) (1)
where Tr is the correlation time for the probe rotation, which is inversally related to the rate of rotation (R).
rr = 1/6R.
rr and r** are considered as reflecting different limitations on probe rotation. Tr primarily depends on the local viscous resistance to rotation. The infinite time anisotropy (r**) reflects prin- cipally the limitation that the local environment of the fluorophore imposes on the extent or range of probe rotation.
To calculate rr and r** separately we used the
7. Ibid.
8, Ibid.
method described by Lakowicz et al. [19,22], 9 in which R or r r was estimated from measurements of rs, r and the differential polarized phase shift A according to the theory developed by Weber [27]. The r r values obtained in this way were used in Eq. 1, rearranged in order to calculate r**.
Previously reported measurements using differ- ential polarized phase fluorometry l° or time- resolved anisotropy decay [18,20] indicated that the 'out of the plane' motion of the n-AS in protein-free liposomes is totally unhindered, i.e. r** was found to be zero when measured at wavelengths where r0 is about 0.1. Then, correla- tion time for the 'out of the plane motion' of the n-AS was here obtained using the rs values obtained at 322 nm in the unmodified Perrin equation:
rs = ro/(1 + r/rr) (2)
3. Results
3.1. Influence of number and position of double bonds
Fig. 1 shows the lifetimes and the obtained r~. and ¢r parameters for the wobbling motion of DPH in MLV of a series of phosphatidylcholines containing mono- and polyunsaturated acyl chains of 18 and 20 carbons. The lifetime of DPH (Fig. 1 A) has a tendency to diminish with an increase in the unsaturation degree. It is particularly notice- able in di-20:4(5,8,11,14)-PC.
The comparison of r** and rr values obtained in MLV of di- 18: I(6)-PC and dil 8:1 (9)-PC indicates that when the double bonds are in the more exter- nal position 6, DPH rotates a little faster but to a lesser extent than when the double bonds are in position 9. The incorporation of double bonds into the internal region of the lipid bilayer as in di18:2(9,12)-PC results in a strong increase in the extent and also in the rate of DPH wobbling, as indicated by the decrease in the obtained r** and
9. Described in detail in H.A. Garda, A.M. Bernasconi and R.R. Brenner, submitted (b). 10. H.A. Garda, A.M. Bernasconi and R.R. Brenner, submit- ted (b).

M.A. Trieerri et al./ Chem. Phys. Lipids 71 (1994) 61-72 65
~'r values. However, the introduction of a third double bond towards the internal deep region as in di18:3(9,12,15)-PC does not produce an extra- large change in the rotational parameters of DPH. On the other hand, the introduction of double bonds towards the more external region of the lipid bilayer does not have such a strong effect. It is indicated by the comparison between di20:1 (11)- PC and di20:4(5,8,11,14)-PC, in which DPH wob- bling occurs at a similar rate and only a slight decrease in rod is observed in the more unsaturated di-20:4-PC.
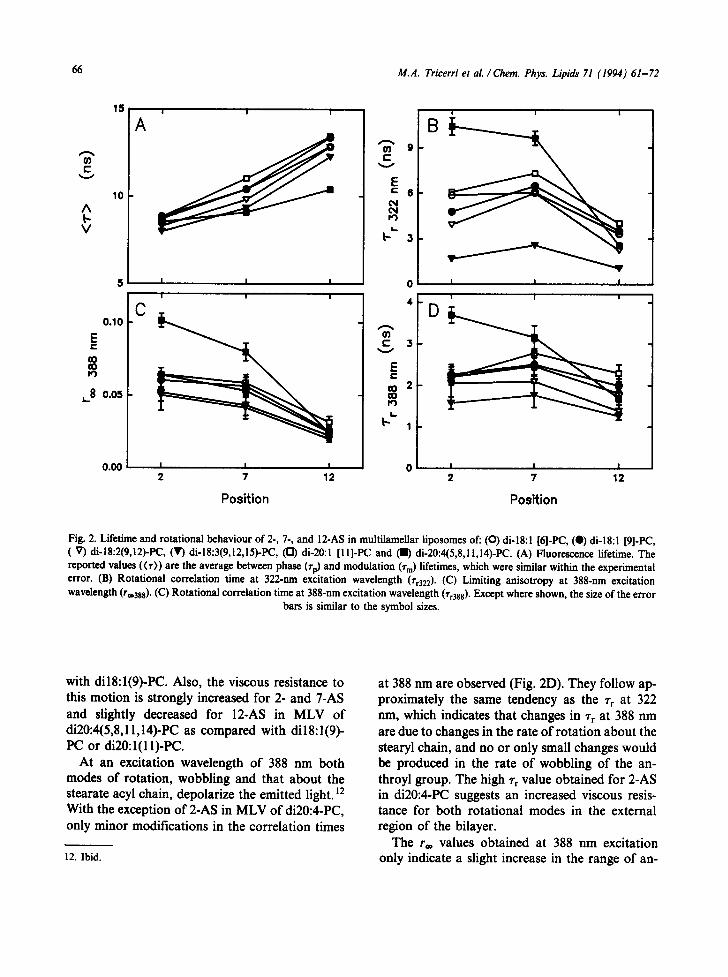
Lifetimes and parameters for the rotation of 2-, 7- and 12-AS obtained for the same series of phos- phatidylcholine MLVs are shown in Fig. 2. As previously observed in model and natural mem- branes [29], lifetime increases in the order 2 < 7 < 12-AS, indicating a decrease in polarity and/or
water content towards the bilayer deep interior. Similarly to DPH, the lifetime of 12-AS in MLV of di20:4(5,8,11,14)-PC is significantly shorter than in MLV of the other PCs (Fig. 2A), suggesting an increased polarity in the microenvironment of these probes in the bilayer of this polyunsaturated lecithin.
As previously discussed, H the correlation times obtained at 322-nm excitation for the n-AS probes would correspond to the unhindered rotation about the stearate acyl chain. The rate of this rota- tional mode is enhanced for all the n-AS tested when the unsaturation degree is increased in the series dilS:l(9)-PC < di18:2(9,12)-PC < di18:3- (9,12,15)-PC (Fig. 2B). The rate of this motion is decreased for 2-AS in dilS:l(6)-PC as compared
10
A
5
0
e- V
A b V
0.04
L_ 8
0.02
0.00
B
i ,,q ,,q
,,q ,,q *I ,,q
,,q
1 I. Ibid.
2
b I"
Fig. 1. Lifetime and rotational behaviour of DPH in multilamellar liposomes of: [] di- ! 8:1 (6)-PC, [] di- 18: i (9)-PC, [] di- 18:2(9, i 2)-PC, [] di-18:3(9,12,1S)-PC, [] di-20:1(11)-PC, and f di-20:4(5,8,11,14)-PC. (A) Fluorescence lifetime. The reported values ((r)) are the
average between phase (¢p) and modulation (¢m) lifetimes, which were similar within the experimental error. (B) Limiting anisotropy (r=). (C) Rotational correlation time (¢r)-
66 M.A. Tricerri et al . / Chem. Phys. Lipids 71 (1994) 61-72
E V
A b V
1 _ q - ~ I ! i
A
I I I i
C . 0.10
E
oo oO F)
L8 0.05
0 . 0 0 i i J 2 7 12
' ~ 9
:e e it')
b ~ 3
!
B
I I I
4 ~ i i i
& 3
0 I 2 7 12
Position Position
Fig. 2. Lifetime and rotational behaviour of 2-, 7-, and 12-AS in multilamellar iiposomes of: ((3) di-18:! [6]-PC, (e) di-18:l [9]-PC, (V) di- 18:2(9,12)-PC, (V) di- 18:3(9,12,15)-PC, ([!) di-20:1 [ 11 ]-PC and (11) di-20:4(5,8,11,14)-PC. (A) Fluorescence lifetime. The reported values (<~)) are the average between phase (Tp) and modulation (~m) lifetimes, which were similar within the experimental error. (B) Rotational correlation time at 322-nm excitation wavelength ('rr322). (C) Limiting anisotropy at 388-nm excitation wavelength (r~.3s8). (C) Rotational correlation time at 388-nm excitation wavelength (~r388)' Except where shown, the size of the error
bars is similar to the symbol sizes.
with dil8:l(9)-PC. Also, the viscous resistance to this motion is strongly increased for 2- and 7-AS and slightly decreased for 12-AS in MLV of di20:4(5,8,11,14)-PC as compared with dil8:l(9)- PC or di20:l(11)-PC.
At an excitation wavelength of 388 nm both modes of rotation, wobbling and that about the stearate acyl chain, depolarize the emitted light. ]2 With the exception of 2-AS in MLV of di20:4-PC, only minor modifications in the correlation times
12. Ibid.
at 388 nm are observed (Fig. 2D). They follow ap- proximately the same tendency as the rr at 322 nm, which indicates that changes in 1" r at 388 nm are due to changes in the rate of rotation about the stearyl chain, and no or only small changes would be produced in the rate of wobbling of the an- throyl group. The high rr value obtained for 2-AS in di20:4-PC suggests an increased viscous resis- tance for both rotational modes in the external region of the bilayer.
The ro. values obtained at 388 nm excitation only indicate a slight increase in the range of an-
M.A. Tricerri et al. / Chem~ Phys. Lipids 71 (1994) 61-72 67
A L B C
%'3
L8
V
0.1
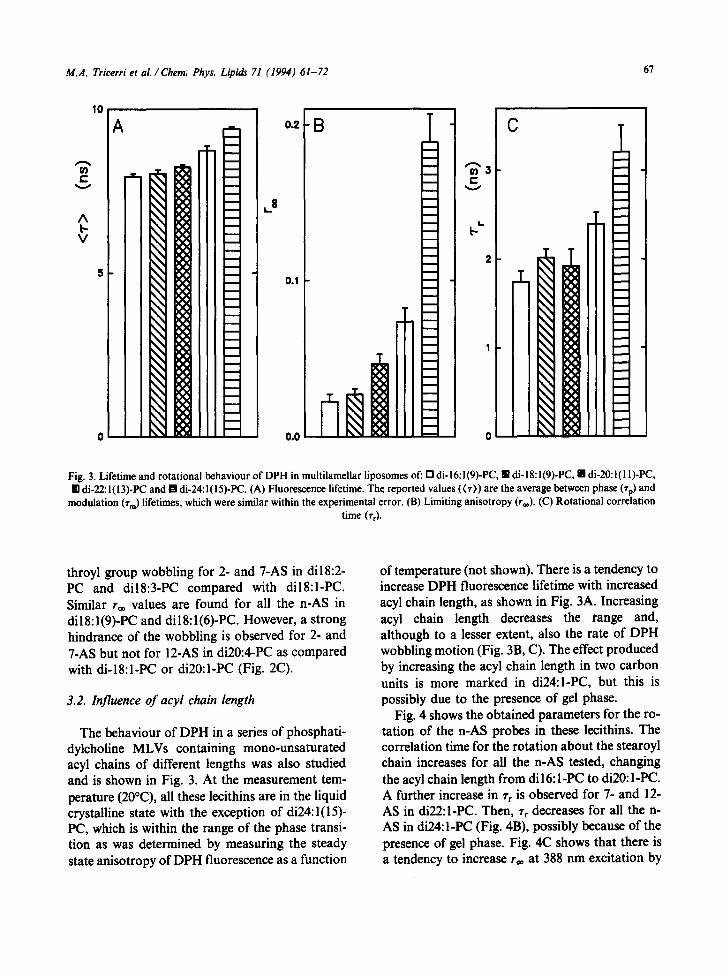
0.0 ~']~~ 0 / Fig. 3. Lifetime and rotational behaviour of DPH in multilamellar l iposomes of" [] di- 16:1 (9)-PC, m di- 18:1 (9)-PC, l1 di-20:1 (1 I)-PC, In di-22: l(13)-PC and a di-24: l(15)-PC. (A) Fluorescence lifetime. The reported values ((r)) are the average between phase (rp) and
modulation (rm) lifetimes, which were similar within the experimental error. (B) Limiting anisotropy (r**). (C) Rotational correlation time (rr)-
throyl group wobbling for 2- and 7-AS in dil8:2- PC and dilS:3-PC compared with dilS:l-PC. Similar r® values are found for all the n-AS in dil 8:1 (9)-PC and dil 8:1 (6)-PC. However, a strong hindrance of the wobbling is observed for 2- and 7-AS but not for 12-AS in di20:4-PC as compared with di-18:l-PC or di20:l-PC (Fig. 2C).
3.2. Influence of acyl chain length
The behaviour of DPH in a series of phosphati- dylcholine MLVs containing mono-unsaturated acyl chains of different lengths was also studied and is shown in Fig. 3. At the measurement tem- perature (20°C), all these lecithins are in the liquid crystalline state with the exception of di24:1 (15)- PC, which is within the range of the phase transi- tion as was determined by measuring the steady state anisotropy of DPH fluorescence as a function
of temperature (not shown). There is a tendency to increase DPH fluorescence lifetime with increased acyl chain length, as shown in Fig. 3A. Increasing acyl chain length decreases the range and, although to a lesser extent, also the rate of DPH wobbling motion (Fig. 3B, C). The effect produced by increasing the acyl chain length in two carbon units is more marked in di24:l-PC, but this is possibly due to the presence of gel phase.
Fig. 4 shows the obtained parameters for the ro- tation of the n-AS probes in these lccithins. The correlation time for the rotation about the stearoyl chain increases for all the n-AS tested, changing the acyl chain length from dil 6: I-PC to di20: I-PC. A further increase in % is observed for 7- and 12- AS in di22:l-PC. Then, Tr decreases for all the n- AS in di24:l-PC (Fig. 4B), possibly because of the presence of gel phase. Fig. 4(2 shows that there is a tendency to increase r~ at 388 nm excitation by

68 M.A. Tricerri et al./ Chem. Phys. Lipids 71 (1994) 61-72
15
lO
V
5
0 .10
i i i i i i
B lO
E_5
o
3
2 e .
I I I I I I
• i i i i
C D
0 .00 I J J 0 I I I 2 7 12 2 7 12
Position Position
Fig. 4. Lifetime and rotational behaviour of 2-, 7- and 12-AS in multilamellar iiposomes of: (O) di-16:l(9)-PC, (0) di-18:l(9)-PC, (V) di-20:l(l D-PC, ( • ) di-22:1(13)-PC and ([2} di-24:1 (15)-PC. (A) Fluorescence lifetime. The reported values ((T)) are the average between phase (rp) and modulation (rm) lifetimes, which were similar within the experimental error. (B) Rotational correlation time at 322-nm excitation wavelength (rr322). (C) Limiting anisotropy at 388-nm excitation wavelength (r**388). (C) Rotational correlation
time at 388-nm excitation wavelength (rr38s). Except where shown, the size of the error bars is similar to the symbol sizes.
increasing tho acyl chain length from di 16:1-PC to di22: I-PC, and a strong hindrance of the extent of 7- and 12-AS wobbling is observed for di24:1-PC. On the other hand, the rr at 388 nm excitation in- creases mainly for 12-AS by changing the acyl chain length from di lb : l -PC to di22:l-PC, and a high increase in the wobbling rate is observed for 2- and 7-AS in di24: I-PC (Fig. 4D). The different behaviour of di24:l-PC is possibly due to the presence of gel phase.
3.3. Influence of unsaturation and chain length in mixed-chain lecithins
Phospholipids which are components of natural membranes are mainly mixed-chain, containing a saturated chain in sn-1 and an unsaturated chain in sn-2. The behaviour of DPH in several mixed- chain lecithins is shown in Fig. 5. An increase in the saturated acyl chain length in sn-1 strongly decreases the range without altering the rate of

M.A. Tricerri et al./ Chem. Phys. Lipids 71 (1994) 61-72 69
10
0~ i-
v
A b V
B T 0.o8 21 C
~.8 b L-
I
0.04 1
Fig. 5. Lifetime and rotational behaviour of DPH in multilamellar liposomes of" r'l 16:0/18:1(9)-PC. [] 18:0/18:1(9)-PC, i 16:0/20:4(5,8,11,14)-PC, [] 18:0/20:4(5,8,11,14)-PC, [] egg-PC and ! soybean-PC. (A) Fluorescence lifetime. The reported values ((r)) are the average between phase (¢p) and modulation (Tm) lifetimes, which were similar within the experimental error. (B) Limiting ani-
sotropy (r®). (C) Rotational correlation time (~'r)'
DPH wobbling if the unsaturated acyl in an-2 is 18:1, but it has a minor influence if the unsaturated acyl in sn-2 is 20:4. It is also possible to see in Fig. 5B and 5C that by changing the unsaturated acyl chain in sn-2 from 18:1 to 20:4 the extent and rate of DPH wobbling are increased. Fig. 5 also in- cludes the data obtained in complex mixtures such as egg and soybean lecithins. Egg-PC is rich in sat- urated fatty acids and contains 20:4, while soybean-PC is rich in 18:2 and contains 18:3. DPH wobbles in a larger range although with a similar rate in soybean-PC compared with egg-PC.
The behaviour of n-AS probes in this series of lecithins is shown in Fig. 6. The rotational rate about the stearoyl chain (Fig. 6B) decreases for the three n-AS by increasing the an-1 saturated acyl chain length if the sn-2 unsaturated acyl chain is
18:1, but the rate of this rotational mode is decreased only for the external 2- and 7-AS probes if 20:4 is present in sn-2. In this case, a faster rota- tion is observed for 12-AS in 18:0/20:4-PC than in 16:0/20:4-PC. It is also interesting to compare the influence when changing the unsaturated chain in sn-2. When the sn-1 acyl group is 16:0, the substi- tution of 20:4 for 18:1 in sn-2 produces a similar increase in the rotational rate about the stearoyl chain for all the n-AS tested. However, when the sn-1 acyl is 18:0, the substitution of 20:4 for 18:1 in sn-2 produces a very large increase in the rate of this rotational mode for 12-AS but in minor pro- portion for 2- and 7-AS probes. The rotation about the stearoyl chain is faster in soybean-PC when compared with egg-PC for all the n-AS tested.

70 M.A. Tricerri et al. / Chem. Phys. Lipids 71 (1994) 61-72
1 5 i i i i i i
&
^ 1Ol- =E
, , 5 ~ 2
E
0 o 0 0 I I I 0 I I I 2 7 12 2 7 12
Position Position
Fig. 6. Lifetime and rotational behaviour of 2-, 7- and 12-AS in muitilamellar liposomes of: (O) 16:0/18:1(9)-PC, (0) 18:0/18:1(9)-PC, (V) 16:0/20:4(5,8,11,14)-PC, (V) 18:0/20:4(5,8,11,14)-PC, (12) egg-PC and ( I ) soybean-PC. (A) Fluorescence lifetime. The reported values ((r)) are the average between phase (rp) and modulation (rm) lifetimes, which were similar within the experimental error. (B) Rotational correlation time at 322-nm excitation wavelength (rr322). (C) Limiting anisotropy at 388-nm excitation wavelength (r®3ss). (C) Rotational correlation time at 388-nm excitation wavelength (rr388). Except where shown, the size of the error bars is similar to
the symbol sizes.
The rr values obtained at 388 nm excitation are similar in this series of lecithins (Fig. 6D). Only the Tr 388 nm for 12-AS in 18:0/20:4-PC is significant- ly smaller than in the other lecithins, indicating an increased wobbling rate. The influence of unsat- uration in sn-2 and acyl chain length in sn-I is more important on the range of the n-AS wobbling (Fig. 6C). By changing the acyl chain length in sn-1 we obtain a different effect, which depends on the unsaturated acyl chain in sn-2. The substitution of 18:0 for 16:0 in sn-1 does not influence the wobbl- ing range of the n-AS when the unsaturated acyl chain is 18:1, but the same change decreases the range of 2- and 7-AS wobbling when the sn-2 acyl
chain is 20:4. Also, it can be observed in Fig. 6C that the influence of substituting 20:4 for 18:1 in sn-2 increases the range of wobbling of all the n- AS tested when 16:0 is the sn-1 fatty acid, but the same change increases the wobbling range only for 12-AS when 18:0 is the sn-I acyl group. A very similar wobbling range is observed for the three n- AS tested in egg-PC or soybean-PC.
4. Discuss ion
The relationship between fatty acid unsatura- tion and physical properties of membrane lipid phase has been and still is a controversial subject.

M.A. Tricerri et aL / Chem. Phys. Lipida 71 (1994) 61-72 71
The introduction of a cis double bond into the lipid matrix affects the physical organization of the lipid bilayer. It disrupts the co-operativity of the tightly packed saturated acyl chains, increasing the cross-sectional area per acyl chain, as indicated by monolayer studies [30-32] (reviewed in Refs. 1, 33). It results in an increase in both range and rate of the wobbling motion of general probes as DPH [15]. However, no simple relationship exists between double bond number and lipid matrix properties. The effect of the first introduced dou- ble bond is more marked than that of the subse- quent ones [1,15,32,33]. Less attention was paid to the double bond position, although in some cases it may have a more important influence than the actual double bond number [1]. Melting temperatures of 18:1 and 18:2 acids [34,35] and phase transition temperatures of dil8:l-phospha- tidylcholines [36] indicate a maximal double bond effect when it is positioned at the centre of the acyl chain.
Data reported in this paper show that a complex relationship exists between double bond number and position and chain length With the bilayer pro- perties in the liquid crystalline state. It depends not only on the type and localization of the probe but also on the particular mode of rotation con- sidered.
The influence of unsaturation on the bilayer structural and dynamic properties is strongly dependent on the position of double bonds. At least in lecithins containing the same acyl chain in sn-I and sn-2, double bonds located in the internal deep region of the bilayer as in dil8:2(9,12)-PC or di18:3(9,12,15)-PC increase the range of wobbling of the externally located 2- and 7-AS probes without largely modifying the order in the internal region where 12-AS is located. On the contrary, as suggested by the results obtained in di20:4(5,8, l l,14)-PC, the externally located double bonds increase the order in the external region where 2- and 7-AS are located, but they increase the range of 12-AS wobbling.
From the comparison with saturated lecithins using 2H-NMR [37,38] and steady state aniso- tropy of n-AS [39], it has been suggested that a cis double bond would produce a separation between acyl chains, thus allowing an increase of mobility
in regions far away from where the double bond is located. However, in the region where the double bond is located a relatively diminished mobility would result, because of the diminished conforma- tional freedom in the non-rotating double bond or a steric hindrance to the probe motion produced by the acyl chain bend. These considerations would explain well the above-mentioned effects on the range of n-AS wobbling, but they cannot ex- plain why internally located double bonds as in dil 8:2(9,12)-PC or di 18:3(9,12,15)-PC decrease the viscous resistance to the motion about the stearoyl chain for all tested n-AS probes, even the internal- ly located 12-AS.
These lecithins containing two polyunsaturated acyl chains are not present in natural membranes. Biomembrane lecithins are generally mixed-chain with a saturated acyl in sn-1 and an unsaturated one in sn-2. The results shown here indicate that the influence of the unsaturation in sn-2 depends on the acyl chain length in sn-1, i.e. by changing the unsaturated sn-2 chain from 18:1 to 20:4 a gen- eral mobility enhancement at all the tested depths is produced if the saturated acyl chain in sn-1 is 16:0, but the same change produces a mobility increase exclusively localized in the region probed by 12-AS when the sn-1 fatty acid is 18:0. The same data but focused from another point of view show that the influence of chain length in sn-1 is depen- dent on the fatty acid that is present in sn-2.
Thus there is a complex relationship between unsaturation or acyl chain length and viscotropic properties of the lipid bilayer even in systems con- taining single molecular species of phospholipids. In systems consisting of mixtures of different mo- lecular components as biological membranes, the interaction between them should also be con- sidered, and further efforts will be required to fully understand the influence of fatty acid composition on the viscotropic properties of the lipid matrix of the membranes.
The influence of double bond position can be relevant in the biological membranes, as suggested by the double bond distribution of plant compared with mammal phospholipids. The fatty acids pre- sent in plant phospholipids are rich in double bonds near the terminal methyl group, i.e. in the internal deeper region of the membrane bilayer.

72 M.A. Tricerri et al . / Chem. Phys. Lipids 71 (1994) 61-72
Mammals take up these fatty acids and modify them by introducing extra double bonds towards the carboxyl group, i.e. towards the external region of the membrane bilayer. The biochemical basis for this is well known [40], but nothing is known about its importance regarding the physical properties and function of animal cellular mem- branes.
5. Acknowledgements
This work was supported by a grant from the Consejo Nacional de Investigaciones Cientificas y T6cnicas (CONICET), Argentina, and by Efamol Research Inc., Canada.
6. References
1 C.D. Stubbs and A.D. Smith (1984) Biochim. Biophys Acta 779, 89-137.
2 D.A. York, P.A. Hyslop and R. French (1982) Biochem. Biophys. Res. Commun. 106, 1478-1483.
3 C.E. Castuma and R.R. Brenner (1983) Biochim. Biophys. Acta 729, 9-16.
4 R.R. Brenner, H.A. Garda, I.N.T. de Gomez Durum and H. Pezzano 0982) Prog. Lipid Res. 20, 315-321.
5 C.L. L~ger, D. Daveloose, R. Christon and J. Viret (1990) Biochemistry 29, 7269-7275.
6 K.K.inosita, S. Kawato and A. Ikegami (1977) Biophys. J. 20, 289-305.
7 G. Lipari and A. Szahn (1980) Biophys. J. 30, 489-506. 8 Weber, G. (1977) J. Chem. Phys. 66, 4081-4091 9 W.W. Mantulin and G. Weber (1977) J. Chem. Phys. 66,
4O92-4O99. 10 A.S. Waggoner and L. Stryer (1970) Proc. Natl. Acad Sci.
USA. 65, 579-589. 11 D.B. Chaipin and A.M. Kleinfeld (1983) Biochim.
Biophys. Acta 731,465-474. 12 F.S. Abrams, Chattopadhyay A. and London E. (1992)
Biochemistry 31, 5322-5327. 13 M.P. Andrich and J.M. Vanderkooi (1976) Biochemistry
15. 1257-1261. 14 S. Kawato, K. Kinosita and A. lkegami (1977)
Biochemistry 16, 2319-2324.
15 C.D. Stubbs, T. Kouyama, K. Kinosita and A. Ikegami (1981) Biochemistry 20, 4257--4262.
16 S.R. Johns, R.I. Willing, K.R. Thulhnrn and W.H. Sawyer (1979) Chem. Phys. Lipids 24, 11-16.
17 R.A. Badley, W.G. Martin and H. Schneider (1973) Biochemistry 12, 268-275.
18 M. Vincent, B. De Foresta, J. Gallay and A. Aifsen (1982) Biochemistry 21,708-716.
19 J.R. Lakowicz (1983) Principles of Fluorescence Spec- troscopy. Plenum Press, New York.
20 M. Vincent, B. De Foresta, J. Gailay and A. Alfsen (1982) Biochem. Biophys. Res. Commun. 107, 914-921.
21 H. Kutchai, L.H. Chandler and G.B. Zavoico (1983) Biochim. Biophys. Acta 736, 137-149.
22 J.R. Lakowicz, F.G. Prendergast and D. Hogen (1979) Biochemistry 18, 508-519.
23 R.D. Spencer and G. Weber (1970) J. Chem. Phys. 52, 1654-1664
24 J.R. Lakowicz and H. Cherek (1980) J. Biol. Chem. 255, 831-834.
25 J.R. Lakowicz, H. Cherek and D.R. Bevan (1980) J. Biol. Chem. 255, 4403-4406.
26 F.G. Prendergast, R.P. Haugland and P.J. CaUahan (1981) Biochemistry 20, 7333-7338.
27 G. Weber (1977) Acta Phys. Pol. A 54, 173-179. 28 G. Weber (1981) J. Phys. Chem. 85, 949-953. 29 K.R. Thulborn and W.H. Sawyer (1978) Biochim.
Biophys. Acta 511, 125-140. 30 R.A. Demel, W.S.M. Geurts van Kessel and L.L.M van
Deenen (1972) Biochim. Biophys. Acta 266, 26-40. 31 D. Ghosh, M.A. Williams and J. Tinoco (1973) Biochim.
Biophys. Acta 291,351-362. 32 R.W. Evans and J. Tinoco (1978) Chem. Phys. Lipids 22,
207-220. 33 R.R. Brenner (1984) Prog. Lipid Res. 23, 69-96. 34 F.D. Gunstone and I.A. Ismail (1967) Chem. Phys. Lipids
1,264-269. 35 W.W. Christie and R.T. Hohnan (1967) Chem. Phys. Lip-
ids I, 407-423. 36 P.G. Barton and F.D. Gunstone (1975) J. Biol. Chem.
250, 4470-4476. 37 A. Seelig and J. Scelig (1977) Biochemistry 16, 45-50. 38 J. Seelig and N. Waespe-S~tr6evic (1978) Biochemistry 17,
3310-3315. 39 K.R. Thulborn, F.E. Treloar and W.H. Sawyer (1978)
Biochem. Biophys. Res. Commun. 81, 42-49. 40 R.R. Brenner (1977) Drug Metab. Rev. 6, 155-212.




















