
Light Directed Gene Transfer by Photochemical Internalisation
24
Current Gene Therapy, 2003, 3, 000-000 1 1566-5232/03 $41.00+.00 © 2003 Bentham Science Publishers Ltd. Light Directed Gene Transfer by Photochemical Internalisation A. Høgset 1, 2 *, L. Prasmickaite 2 , B.Ø. Engesæter 2, 3 , M. Hellum 2 , P.K. Selbo 2 , V.M. Olsen 2 , G.M. Mælandsmo 3 and K. Berg 2 1 PCI Biotech AS, Hoffsvn.48, N-0377 Oslo, Norway; 2 Department of Biophysics, The Norwegian Radium Hospital, Montebello, N-0310 Oslo, Norway and 3 Department of Tumour Biology The Norwegian Radium Hospital, Montebello, N-0310 Oslo, Norway Abstract: Numerous gene therapy vectors, both viral and non-viral, are taken into the cell by endocytosis, and for efficient gene delivery the therapeutic genes carried by such vectors have to escape from endocytic vesicles so that the genes can further be translocated to the nucleus. Since endosomal escape is often an inefficient process, release of the transgene from endosomes represents one of the most important barriers for gene transfer by many such vectors. To improve endosomal escape we have developed a new technology, named photochemical internalisation (PCI). In this technology photochemical reactions are initiated by photosensitising compounds localised in endocytic vesicles, inducing rupture of these vesicles upon light exposure. The technology constitutes an efficient light-inducible gene transfer method in vitro, where light-induced increases in transfection or viral transduction of more than 100 and 30 times can be observed, respectively. The method can potentially be developed into a site- specific method for gene delivery in vivo. This article will review the background for the PCI technology, and several aspects of PCI induced gene delivery with synthetic and viral vectors will be discussed. Among these are: (i) The efficiency of the technology with different gene therapy vectors; (ii) use of PCI with targeted vectors; (iii) the timing of DNA delivery relative to the photochemical treatment. The prospects of using the technology for site-specific gene delivery in vivo will be thoroughly discussed, with special emphasis on the possibilities for clinical use. In this context our in vivo experience with the PCI technology as well as the clinical experience with photodynamic therapy will be treated, as this is highly relevant for the clinical use of PCI-mediated gene delivery. The use of photochemical treatments as a tool for understanding the more general mechanisms of transfection will also be discussed. Keywords: Photochemical internalisation, Endosomes, Site-specific, Gene delivery, Gene therapy, Light-directed 1. BACKGROUND Despite continuous improvement in vector technology, development of methods for efficient and specific delivery of therapeutic gene to the target cells still remains a main issue in the clinical development of gene therapy [Verma and Somia, 1997,Anderson, 1998,Pfeifer and Verma, 2001,Kay et al., 2001,Liu and Huang, 2002]. In vivo gene delivery faces many substantial obstacles, both extracellular and intracellular. Among the extracellular barriers can be mentioned enzymatic degradation of the DNA, inefficient extravascularisation, slow transport through the target tissue, extensive uptake in non-target tissues and inefficient binding to and uptake into the target cells [Nishikawa and Huang, 2001]. Intracellularly also there are many obstacles for the functional delivery of transgenes. Nuclear transport and uptake of the transfecting DNA represent one important such barrier [Zabner et al., 1995,Wilke et al., 1996,Pouton, 1998,Luo and Saltzman, 2000], however, several other *Address correspondence to this author at the PCI Biotech AS, Hoffsvn.48, N-0377 Oslo, Norway; Tel: +47 2325 4003; Fax: +47 2325 4001; E-mail: [email protected] intracellular obstacles can also be identified, e.g.dissociation of DNA from the complexing agent, degradation of the DNA, and sequestration of the DNA in endocytic vesicles [Zabner et al., 1995,Ogris et al., 1998,Pouton et al., 1998,De Smedt et al., 2000,Pouton and Seymour, 2001]. Finally transcription and translation of the transgene also represent possible barriers, especially in cases where it is desirable with long-term expression of the transgene. The importance of these intracellular barriers probably varies considerably between different vector systems. While for some viral vectors, at least in some cell lines, the cellular uptake of just a few virus particles per cell may be sufficient to obtain transduction [Greber et al., 1993,Leopold et al., 1998], with synthetic vectors cells may not be transfected even though they have taken up ample amounts of exogenous DNA. For example it has been estimated that the cells on average contain 2-3 x 10 5 plasmids per cell after delivery by cationic lipids [Zabner et al., 1995,Tseng et al., 1997]. Likewise our own studies showed that for polylysine- mediated delivery cells could contain on average about 6 x 10 4 plasmids, with still only 3 % of the cells expressing the transgene [Prasmickaite et al., 2000]. Therefore, at least for
-
Upload
rr-research -
Category
Documents
-
view
1 -
download
0
Transcript of Light Directed Gene Transfer by Photochemical Internalisation

Current Gene Therapy, 2003, 3, 000-000 1
1566-5232/03 $41.00+.00 © 2003 Bentham Science Publishers Ltd.
Light Directed Gene Transfer by Photochemical Internalisation
A. Høgset1,2*, L. Prasmickaite2, B.Ø. Engesæter2,3, M. Hellum2, P.K. Selbo2, V.M. Olsen2,G.M. Mælandsmo3 and K. Berg2
1PCI Biotech AS, Hoffsvn.48, N-0377 Oslo, Norway; 2Department of Biophysics, The Norwegian Radium Hospital,Montebello, N-0310 Oslo, Norway and 3Department of Tumour Biology The Norwegian Radium Hospital, Montebello,N-0310 Oslo, Norway
Abstract: Numerous gene therapy vectors, both viral and non-viral, are taken into the cell by endocytosis, and forefficient gene delivery the therapeutic genes carried by such vectors have to escape from endocytic vesicles so thatthe genes can further be translocated to the nucleus. Since endosomal escape is often an inefficient process, releaseof the transgene from endosomes represents one of the most important barriers for gene transfer by many suchvectors. To improve endosomal escape we have developed a new technology, named photochemical internalisation(PCI). In this technology photochemical reactions are initiated by photosensitising compounds localised inendocytic vesicles, inducing rupture of these vesicles upon light exposure. The technology constitutes an efficientlight-inducible gene transfer method in vitro, where light-induced increases in transfection or viral transduction ofmore than 100 and 30 times can be observed, respectively. The method can potentially be developed into a site-specific method for gene delivery in vivo.
This article will review the background for the PCI technology, and several aspects of PCI induced gene deliverywith synthetic and viral vectors will be discussed. Among these are: (i) The efficiency of the technology withdifferent gene therapy vectors; (ii) use of PCI with targeted vectors; (iii) the timing of DNA delivery relative to thephotochemical treatment.
The prospects of using the technology for site-specific gene delivery in vivo will be thoroughly discussed, withspecial emphasis on the possibilities for clinical use. In this context our in vivo experience with the PCI technologyas well as the clinical experience with photodynamic therapy will be treated, as this is highly relevant for theclinical use of PCI-mediated gene delivery.
The use of photochemical treatments as a tool for understanding the more general mechanisms of transfection willalso be discussed.
Keywords: Photochemical internalisation, Endosomes, Site-specific, Gene delivery, Gene therapy, Light-directed
1. BACKGROUND
Despite continuous improvement in vector technology,development of methods for efficient and specific delivery oftherapeutic gene to the target cells still remains a main issuein the clinical development of gene therapy [Verma andSomia, 1997,Anderson, 1998,Pfeifer and Verma, 2001,Kayet al., 2001,Liu and Huang, 2002]. In vivo gene deliveryfaces many substantial obstacles, both extracellular andintracellular. Among the extracellular barriers can bementioned enzymatic degradation of the DNA, inefficientextravascularisation, slow transport through the target tissue,extensive uptake in non-target tissues and inefficient bindingto and uptake into the target cells [Nishikawa and Huang,2001]. Intracellularly also there are many obstacles for thefunctional delivery of transgenes. Nuclear transport anduptake of the transfecting DNA represent one important suchbarrier [Zabner et al., 1995,Wilke et al., 1996,Pouton,1998,Luo and Saltzman, 2000], however, several other
*Address correspondence to this author at the PCI Biotech AS, Hoffsvn.48,N-0377 Oslo, Norway; Tel: +47 2325 4003; Fax: +47 2325 4001; E-mail:[email protected]
intracellular obstacles can also be identified, e.g.dissociationof DNA from the complexing agent, degradation of theDNA, and sequestration of the DNA in endocytic vesicles[Zabner et al., 1995,Ogris et al., 1998,Pouton et al.,1998,De Smedt et al., 2000,Pouton and Seymour, 2001].Finally transcription and translation of the transgene alsorepresent possible barriers, especially in cases where it isdesirable with long-term expression of the transgene.
The importance of these intracellular barriers probablyvaries considerably between different vector systems. Whilefor some viral vectors, at least in some cell lines, the cellularuptake of just a few virus particles per cell may be sufficientto obtain transduction [Greber et al., 1993,Leopold et al.,1998], with synthetic vectors cells may not be transfectedeven though they have taken up ample amounts ofexogenous DNA. For example it has been estimated that thecells on average contain 2-3 x 105 plasmids per cell afterdelivery by cationic lipids [Zabner et al., 1995,Tseng et al.,1997]. Likewise our own studies showed that for polylysine-mediated delivery cells could contain on average about 6 x104 plasmids, with still only 3 % of the cells expressing thetransgene [Prasmickaite et al., 2000]. Therefore, at least for
Wasim

2 Current Gene Therapy, 2003, Vol. 3, No. 2 Høgset et al.
synthetic vectors the intracellular barriers are of primeimportance as a target for improving gene delivery.
We have recently described a technology, namedphotochemical internalisation (PCI), where photosensitisingcompounds and light are employed to rupture membranes ofendocytic vesicles, resulting in the release of functionalmacromolecules from these vesicles into the cell cytosol[Berg et al., 1999]. In this article we review the backgroundof this technology, its employment as a gene deliverymethod in combination with different vector systems, and itspossible use as a system for site-specific gene delivery invivo.
1.1. The Importance of Endosomal Escape for GeneDelivery
With most gene delivery vectors, especially non-viral,but also many viral, the major mechanism of uptaking ofgene complexes is through the endocytic pathway [Wickhamet al., 1994,Duan et al., 1999,Douar et al., 2001,van Loo etal., 2001]. Thus, in order for the exogenous DNA to betranslocated to the nucleus it has to be released from theendocytic vesicles before being degraded by lysosomalenzymes. While many viruses have evolved quite efficientsystems for endosomal release [Greber et al., 1993,Leopoldet al., 1998], the situation is different for non-viral vectors,where in many cases endosomal escape is a major obstaclefor efficient gene transfer. This has been indicated by severallines of evidence, including: (i) Transfection by severalsynthetic transfection agents can substantially be improvedby ”lysosomotropic agents” such as chloroquine [Cotten etal., 1990,Wagner et al., 1992,Midoux et al., 1993,Wolfertand Seymour, 1998]. Although the mechanism behind thiseffect is not fully clear [Erbacher et al., 1996], this findingindicates that the transfecting DNA is subjected to adetrimental stay in endocytic vesicles. (ii) Inactivated virusesor viral proteins with “endosomolytic” properties canconsiderably enhance transfection when added to e.g.polylysine-DNA complexes, most probably due to“endosomolytic” properties of the virus proteins [Curiel etal., 1991,Wagner et al., 1992,Cristiano et al.,1993,Harbottle et al., 1998]. (iii) Several “fusogenic”peptides have the same effect [Midoux et al., 1993,Ogris etal., 1998]. (iv) Transfection can be enhanced by glycerol,probably through a glycerol-induced increase in endosomalpermeability [Zauner et al., 1996,Zauner et al., 1997]. (v)The inclusion of “fusogenic” lipids in lipoplexes oftenstimulates transfection [Felgner et al., 1994,Farhood et al.,1995,Wrobel and Collins, 1995].
The importance of endosomal escape as a barrier to genetransfer is believed to vary extensively between different non-viral vectors. Some polycationic vectors, such as polylysine,probably have no inherent mechanism for endosomal escape,while it is thought that e.g. polyethylenimine (PEI) anddendrimers can effect endosomal escape by acting as “protonsponges” inducing swelling and permeabilisation of theendosomes [Boussif et al., 1995,De Smedt et al.,2000,Kirchler et al., 2001]. For cationic lipids, a model hasbeen suggested where the lipids in the
transfection complex induce a “flip-flop” of phospholipids inthe endosomal membranes leading to release of thetransfecting DNA both from the lipid complex and from theendosomes [Xu and Ellington, 1996,Zelphati and Szoka,1996]. The synthetic vector mechanisms for endosomalescape are, however, often probably far less efficient thancorresponding viral mechanisms, implying that moreefficient methods for endosomal release could still lead tosubstantial improvement in transfection with synthetictransfection agents.
In contrast to synthetic vectors, adenovirus is known tobe an efficient vector both for in vitro and in vivo genedelivery [Anderson, 1998,Vorburger and Hunt, 2002]. Theprocess of adenovirus uptake into the cell and the subsequenttrafficking to the cell nucleus is not fully understood, but itis known that adenovirus binds to receptors on the cellsurface, is taken up by endocytosis, escapes from endosomesand is transported to the nucleus via microtubuli[Suomalainen et al., 1999,Nemerow and Stewart,1999,Leopold et al., 2000]. Adenovirus escape fromendosomes is believed to be an efficient process [Greber etal., 1993,Leopold et al., 1998], however, this has only beeninvestigated in a few cell lines, and in many cell systemsendosomal escape may still be an important barrier for genedelivery also by adenoviral vectors. Furthermore,intracellular barriers for gene delivery have been described forother virus vectors [Hansen et al., 2000,Douar et al., 2001],indicating that enhanced endosomal escape could havebeneficial effects also on virus-mediated gene delivery.
A lot of work is going on to develop both viral and non-viral systems for targeted delivery of gene therapy agents tospecific cell types in the body [Wickham, 2000,Haviv andCuriel, 2001,Galanis et al., 2001]. The great majority ofsuch targeting strategies involve binding of the gene therapyvectors to specific molecules (receptors) present on thesurface of the target cells, followed by endocytic uptake ofthe vector/receptor complex. Thus, all such strategies in theend rely on an ability of the endocytosed transgene complexto escape from the endocytic vesicles. Although sometargeted gene therapy vectors may have some built-in featuresfor effecting such escape, for most of the vectors the lack ofefficient endosomal escape may represent a severe limitationfor the efficiency of gene delivery.
1.2. Photochemical Internalisation for ImprovingEndosomal Release
Photochemical internalisation is a novel technologywhere photosensitising compounds (photosensitisers) areused for improving endosomal release of variousendocytosed molecules. Although before the PCI treatmentthe molecules are already taken up by the cell intoendosomes, from the cell membrane view they are still kept‘outside’ separated from the cytosol by the vesiclemembrane, presenting a very significant barrier. The termphotochemical internalisation thus emphasizes the intra-cytosol delivery mediated by light, whereby the endocytosedmolecules are relocalised from what can be viewed as aremnant of the extracellular space into the cell interior.

Light Directed Gene Transfer Current Gene Therapy, 2003, Vol. 3, No. 2 3
1.2.1. Photosensitisers and Photochemical Reactions
1.2.1.1. General Principles
Photosensitisers are compounds that make cellsextraordinary sensitive to illumination with visible light.During exposure to light of wavelengths overlapping theabsorbance spectrum of the photosensitiser in the groundstate (PS), the PS absorbs the energy of a photon (hν) and isconverted into an excited singlet state, 1PS*. Then 1PS* isquickly converted into an excited triplet state, 3PS*. Thistriplet state can initiate further photochemical reactions bytransferring the absorbed energy to other molecules. Withphotosensitisers like porphyrins and structurally relatedcompounds the most important of these reactions proceedsvia singlet oxygen (1O2), a highly reactive form of oxygengenerated after interaction between an excited photosensitisermolecule and triplet ground-state molecular oxygen (3O2)[Moan and Sommer, 1985; Weishaupt et al., 1976;Henderson and Dougherty, 1992] Schematically:
3PS* is thus converted back to PS, which is then readyfor further cycles of excitation and generation of singletoxygen [Kochevar and Redmond, 2000; Moan and Berg,1991]. Thus, three factors: photosensitiser, light and oxygenare absolutely necessary to accomplish the photochemicalreactions.
Singlet oxygen is a very powerful oxidizing agent andcan oxidize many different biomolecules, such as unsaturatedfatty acids, certain amino acids (His, Trp, Cys, Met and Tyr)and the nucleic acid base guanine [Jori and Spikes, 1984].By such oxidations 1O2 can induce damage into variouscellular structures. However, 1O2 has a very short lifetime (<0.1 ms) and accordingly a short range of action (10-20 nm)inside the cell [Moan and Berg, 1991]. This means that onlytargets very close to the excited photosensitiser will besubject to 1O2 damage upon light exposure. Distantmolecules will largely be left intact.
1.2.1.2. Photochemically Induced Cytotoxicity andPhotodynamic Therapy
In general, the oxidative reactions induced by thephotochemical treatment induces cytotoxic effects, since vitalintracellular structures may be damaged [Moan and Berg,1992,Henderson and Dougherty, 1992]. These cytotoxiceffects have been successfully exploited for cancer treatmentin photodynamic therapy (PDT), a treatment modality wherelight exposure leads to photosensitiser-induced killing ofcancer cells [Henderson and Dougherty, 1992,Pass,1993,Dougherty et al., 1998]. PDT is a very specifictreatment, mainly for two reasons. Firstly, the cytotoxiceffects will be generated only in illuminated areas of thebody; in non-illuminated areas the cytotoxicity of thephotosensitisers will generally be very low [Moan and Berg,1992]. Secondly, some photosensitisers show a preferential
accumulation in tumour tissue as compared to adjacentnormal tissue [Pass, 1993]. Thus, ratios in photosensitiserconcentration of tumour to normal surrounding tissues of 3:1up to > 20:1 have been observed [Chopp et al., 1996].Hence, in these cases the light treatment will have a largereffect on the diseased tissue than on the normal adjacenttissue. The reason for this selective photosensitiseraccumulation is unknown, but it seems to be related tocertain properties of tumour tissue, such as low pH, leakyvasculature, poor lymphatic drainage, or the presence oftumour-infiltrating macrophages taking up aggregatedphotosensitisers [Henderson and Dougherty, 1992,Pass,1993].
PDT may seem best suited for superficial cancers, such asbasal cell carcinoma, but, e.g. by the use of fibre opticdevices, the treatment can also be used for a variety ofinternal cancers. Thus, it has been approved for the treatmentof cancers in the lung, head and neck, esophagus, cervix andbladder, and is under investigation e.g. for mesothelioma,ovarian, pancreatic, prostate and colorectal cancer [Doughertyet al., 1998].
In addition to cancer treatment PDT is also suited to non-oncologic conditions, and is being explored in experimentaltreatment of vascular diseases [Neave et al., 1988,Ortu et al.,1992,Gonschior et al., 1996,Yamaguchi et al.,2000,LaMuraglia et al., 2000,Rockson et al., 2000b] andviral infections [Matthews et al., 1988,Ben-Hur et al.,1996,van Moor et al., 1999,Schagen et al., 1999], as well asin immunological disorders [Simkin et al., 2000] such asrheumatoid arthritis [Ratkay et al., 1994,Trauner and Hasan,1996,Ratkay et al., 1998] and psoriasis [Robinson et al.,1999,Boehncke et al., 2000]. However, the most successfulnon-cancer PDT clinical application so far has been thetherapy of patients with visual loss due to age-relatedmacular degeneration with choroidal neovascularisationdisease [Wu et al., 1999,Donati et al., 1999,Regillo,2000,Schmidt-Erfurth and Hasan, 2000] where the treatmentis called photothrombosis.
In contrast to what is the case for other cancer therapieslike chemotherapy and ionising irradiation there is noevidence of PDT-induced carcinogenesis in humans whenutilising clinically relevant photosensitisers, which do notlocalise to the nucleus and do not bind to DNA [Moan etal., 1990]. Thus, unlike ionising irradiation, repeatedinjections and treatments can theoretically be madeindefinitely.
1.2.1.3. Intracellular localisation of the Photosensitisers
Depending on their physicochemical properties,photosensitisers are taken up by the cell either byendocytosis or by active or passive transport through theplasma membrane [Berg et al., 1990a,Rosenkranz, 2000].Depending among other things on the mode of uptake,different photosensitisers localise at different sites inside thecell [Moan et al., 1989,Berg et al., 1990a,Berg et al.,1990b,Rosenkranz, 2000,Prasmickaite et al., 2001a]. Thus,photosensitisers that aggregate and hydrophilicphotosensitisers such as, for example, sulfonated tetraphenylporphines [Berg et al., 1990b,Berg and Moan, 1994] and
PS 1PS* 3PS* PS + 1O2hv O2

4 Current Gene Therapy, 2003, Vol. 3, No. 2 Høgset et al.
sulfonated aluminium phthalocyanines [Moan et al.,1989,Moan et al., 1992,Peng et al., 1996] enter the cells viaendocytosis, and tend to accumulate in endocytic vesicles.As can be seen in Fig. (1) TPPS2a (meso-tetraphenylporphine with two sulfonate groups on adjacentphenyl rings) and AlPcS2a (aluminium phthalocyanine with2 sulfonate groups on adjacent rings) have two sulfonategroups on adjacent phenyl/phthalate rings. This makes thesephotosensitiser molecules amphiphilic, leading to a primarylocalisation in the membranes of endocytic vesicles [Bergand Moan, 1994], with the hydrophobic part of thephotosensitisers inserted into the vesicle membrane [Mamanet al., 1999].
1.2.2. The Principle of Photochemical Internalisation
The development of the photochemical internalisationtechnology was based on the observation that light exposureof cells containing photosensitisers in their endocyticvesicles leads to a permeabilisation of the vesicles andrelease of the photosensitiser into the cytosol [Berg andMoan, 1994, Moan et al., 1994]. In the same studies it wasfound that also substantial amounts of lysosomal enzymeactivities could be found in the cytosol after light treatment.Thus, these studies indicated that in addition to thephotosensitiser also unrelated molecules located inside thelysosomes could be released into the cytosol. Moreover, thisphotochemically induced release could occur withoutinducing extensive cell death, an observation that has beenattributed to a high sensitivity of cathepsins to inactivationby the photochemical treatment [Berg and Moan, 1994].Cathepsins are lysosomal proteases of crucial importance forthe cytotoxic effect of lysosomal rupture [Miller et al.,1983,Wilson et al., 1987], and their inactivation would beexpected to lower the cytotoxic effect of lysosome rupture.
Even though cathepsins seem to be destroyed by thephotochemical treatment, most other vesicular moleculesseem to maintain their biological activity even after the
treatment. A probable reason for this is the short range ofaction of the photochemically generated 1O2, meaning thatonly molecules very close to the excited photosensitiser willbe damaged. Thus, if the photosensitiser mainly localises inthe vesicle membranes, molecules in the vesicle matrixshould not be very liable to photochemical damage. Takentogether it implies that the photochemical treatment may beused to release a host of endocytosed molecules in abiologically active form from endocytic vesicles, a principlewe have named photochemical internalisation (PCI) (Fig.(2)). The introduction of molecules into the cytosol isachieved by first exposing the cells or tissues to aphotosensitising dye and the molecule which one wants todeliver, both of which should preferentially localise inendosomes and/or lysosomes [Berg et al., 1999]. Secondly,the cells or tissues are exposed to light of wavelengthsinducing a photochemical reaction. This reaction will lead todisruption of lysosomal and/or endosomal membranes andthe contents of the endocytic vesicles will be released intothe cytosol. Detailed descriptions of practical aspects of thetechnology can be found in Berg et al. (1996) [Berg et al.,1996] and Prasmickaite et al. (2001) [Prasmickaite et al.,2001b].
In vitro, PCI has been shown to induce endosomalrelease of a variety of molecules, such as plant toxins [Berget al., 1999,Selbo et al., 2000a], immunotoxins [Selbo etal., 2000b], peptides [Berg et al., 1999], ribozymes andoligodeoxynucleotides [Høgset et al., 2000,Prasmickaite etal., 2001a]. In addition, PCI has been shown tosubstantially increase transfection with several differentsynthetic transfection agents [Berg et al., 1999,Høgset et al.,2000,Prasmickaite et al., 2000,Prasmickaite et al.,2002,Høgset et al., 2002b] and also gene transfer byadenoviral vectors [Høgset et al., 2002a] as will be discussedbelow in detail.
Recent experiments on PCI with a protein toxin showthat photochemical internalisation works very well also in
Fig. (1). Chemical structures of the photosensitisers AlPcS2a and TPPS2a.
NH N
HNN
N N
N
N
N
N
N Al
AIPcS2a TPPS2a
-O3 S SO3-
SO3-
SO3-

Light Directed Gene Transfer Current Gene Therapy, 2003, Vol. 3, No. 2 5
vivo [Selbo et al., 2001] (see 4.3.1). These results indicatethat the PCI technology has a clear clinical potential, bothfor the light-directed delivery of drugs and, as will bediscussed below, for the light-induced, site-specific deliveryof genes in in vivo gene therapy.
2. PHOTOCHEMICAL INTERNALISATION AS AGENE DELIVERY TECHNOLOGY
Photochemical internalisation has been studied as a genedelivery technology both with several non-viral [Berg et al.,1999,Høgset et al., 2000, Prasmickaite et al., 2000,Prasmickaite et al., 2001a, Prasmickaite et al., 2002,Høgsetet al., 2002b] and with adenoviral vectors [Høgset et al.,
2002a], mainly by using reporter genes such as genesencoding EGFP (enhanced green fluorescent protein) or β-galactosidase. However, PCI has also been shown to inducethe delivery of therapeutic genes, such as the genes encodingHSV-tk (Herpes Simplex Virus thymidine kinase)(Prasmickaite, unpublished) and IL-12 (interleukin-12)(Høgset, unpublished).
The effect of PCI on gene delivery can be illustrated byexperiments with polylysine-mediated transfection ofAlPcS2a-treated HCT 116 human colon carcinoma cells,where a part of the culture dish was covered by aluminiumfoil, while the rest of the dish was illuminated. As can beseen from Fig. (3), after PCI many of the illuminated HCT116 cells exhibited visible EGFP-fluorescence, in contrast to
Fig. (2). Light-induced cytosolic delivery by PCI.A. A schematic presentation of the induction of the photochemical reactions involved in PCI; (S) photosensitiser; S* excitedphotosensitiser.B. The principle of PCI. I. The cells are incubated with the photosensitiser (S) and the molecule to be delivered (G). II. S and G aretaken up in the cell by endocytosis, localising in the endosome membrane and matrix, respectively. III. Illumination inducesphotochemical damage to the endosome membrane releasing G into the cytosol.C. PCI-induced relocalisation of an oligodeoxynucleotide. THX cells were incubated for 18 h with 20 µg/ml AlPcS2a. After 3 h washin culture medium and 4 h incubation with a polylysine complex of a fluorescein-labelled oligodeoxynucleotide the same cells werephotographed before and after the exposure to 10 s microscopy light. See [Høgset et al., 2000] for details.

6 Current Gene Therapy, 2003, Vol. 3, No. 2 Høgset et al.
what was the case for the non-irradiated cells. Thus, the lighttreatment strongly induces transfection in HCT 116 cells.This experiment also indicates the high degree of site-specificity that can be obtained, reflected by the cleardifference in transfection between the illuminated and thenon-illuminated parts of the culture dish.
2.1. Light Dose Dependence
The PCI effects on transfection have been studied in moredetail by flow cytometry analysis of the transfected cells.From the data presented in Fig. (4A), it can be seen that thelight treatment induced an increase in transfection efficiencyfrom about 1 % EGFP-positive cells at 0 min of light toabout 50 % positives after 5 min illumination for THXmelanoma cells, thus representing a light -inducedenhancement of transfection efficiency of about 50 times.The induction of transfection exhibits a clear light doseresponse, however, the photochemical enhancement oftransfection is effective over a quite large range of light doses(Fig. (4B)), a fact that may have important implications forthe potential clinical use of the technology (see 4.3.3.1).Thus, although the maximal effect of PCI on transfection isusually obtained at light doses killing about 50 % of thecells [Høgset et al., 2000], very substantial enhancement oftransfection can also be obtained at light doses givingconsiderably less cytotoxicity.
In some, but not all, cell lines the transfection efficiencytends to go down at a very highest light dose. There areseveral possible explanations for this. Firstly, it may be dueto a more general cytotoxic effect of the photochemical
treatment at high light doses, e.g. destroying the cell’sability to transcribe or translate the foreign DNA. Secondly,it may be caused by photochemically induced oxidativedamage to the transfecting DNA in the endocytic vesicles,e.g. generating inactivating mutations. The optimal lightdose will vary between different cells lines, dependingamong other things on the efficiency in the uptake of thephotosensitiser in a given cells line.
2.2. PCI Transfection with Different Cell Lines
It is well known that most transfection methods aredependent on the cell line, and the same was found to be thecase for the transfection induced by PCI. It can be seen(Table 1), that after light treatment the absolute level oftransfection achieved varies between different cell lines.However, although the absolute level of transfection varies,for polylysine-mediated transfection PCI has been found tosubstantially increase the transfection level in most cell linestested (Table 1).
With the exception of the THX cells [Høgset et al.,2000] the conditions for transfection has generally not yetbeen optimised for the tested cell lines (except for findingthe optimal light dose). Thus, given the many differentparameters involved in the PCI treatment, there seems to beample opportunities for finding conditions where thetransfection efficiency in these cells can be substantiallyincreased as compared to what has been achieved so far.
2.3. Effects of PCI with Different Photosensitisers
It is inherent in our hypothesis for the mechanism behindphotochemically induced transfection that onlyphotosensitisers localising in endocytic vesicles shouldimprove transfection, and to test this assumption differentphotosensitisers with different intracellular localisation (see1.2.1.3) were tested for their ability to increase transfectionefficiency with polylysine-mediated transfection. Inaccordance with our assumptions light-activation of the non-lysosomally localised lipophilic dye 3THPP (tetra(3-hydroxyphenyl)porphyrin) and 5-aminolevulinic acid (5-ALA)-induced protoporphyrin IX did not have a significanteffect on polylysine-mediated transfection [Prasmickaite etal., 2001a]. Comparison of the photosensitisers localising inendocytic vesicles showed that AlPcS2a and TPPS2a (Fig.(1)) had the strongest stimulating effect on transfection.These photosensitisers have two sulfonate groups on adjacentphthalate/phenyl rings, making the photosensitiser moleculesamphiphilic. In contrast, the effect of the more hydrophilictetrasulfonated dye TPPS4 was lower, whereas the cationichydrophilic dye p-TMPyPH2 and the lysosomotropic weakbase Nile blue A had a very low or no effect, althoughlocalised in endocytic vesicles [Prasmickaite et al., 2001a].Thus, also among photosensitisers localised in endocyticvesicles differences in the transfection enhancing effectscould be observed, indicating that amphiphilicphotosensitisers expecting to localise mainly in themembranes of the endocytic vesicles (see 1.2.3) give the besteffect [Prasmickaite et al., 2001a]. Thus, localisation of
Fig. (3). PCI-induced transfection of HCT 116 cells studied byfluorescence microscopy.Human HCT 116 colon carcinoma cells were incubated for 18 hwith 20 µg/ml AlPcS2a, washed 3 times with culture medium andtreated with a pEGFP-N1/polylysine complex for 6 h. The cellswere washed once with culture medium, and the culture dish waspartly covered by aluminium foil (“no PCI” region) before beingsubjected to 7 min light treatment. 48 h later the cells wereanalysed by fluorescence microscopy for EGFP fluorescence (see[Høgset et al., 2000] for details).

Light Directed Gene Transfer Current Gene Therapy, 2003, Vol. 3, No. 2 7
4A
4B
Fig. (4). Light-induced transfection studied by flow cytometry.A. Human THX melanoma cells were treated with 20 µg/ml AlPcS2a for 18 h, washed in photosensitiser-free medium, and incubatedwith a pEGFP-N1/polylysine complex (5 µg/ml DNA) for 4 h. The cells were exposed to light (illumination times indicated on thefigure), and red (AlPcS2a) and green (EGFP) fluorescence were analysed by flow cytometry as described earlier [Høgset et al., 2000].The cells in the upper right quadrant were taken as positive for GFP-expression, since in control experiments (cells that received noplasmid) there were no cells in this quadrant (not shown). The figure is from Høgset et al. (2000). (Copyright Hum. Gene Ther. (MaryAnn Liebert, Inc)).
B. Light dose response for photochemical transfection. HCT 116 cell were treated with AlPcS2a over night, washed, and incubated witha pEGFP-N1/polylysine complex for 4 h. The cells were then illuminated with red light, and 48 h later EGFP-expressing cells werescored by flow cytometry. Cell survival was measured by the MTT assay as described earlier [Høgset et al., 2000]. n EGFP-positivecells; ¢ cell survival. Error bars are SEM of 3 experiments.

8 Current Gene Therapy, 2003, Vol. 3, No. 2 Høgset et al.
the photosensitiser in the membranes of endocytic vesiclesseems to be important for optimal photochemicalinternalisation of intact DNA molecules.
2.4. PCI with Different Gene Delivery Agents
2.4.1. Non-Viral Vectors
Whereas PCI in general has a positive effect ontransfection with polycations such as polylysine andpolyethylenimine, the effect on transfection with cationiclipids is much more variable [Prasmickaite et al.,2000,Prasmickaite et al., 2001a]. While in some cell linesPCI seems to reduce cationic lipid mediated transfection, inother cell lines PCI can have positive effects (Hellum et al.,submitted). It also seems that the effect of PCI dependsstrongly on the type of lipid composition used fortransfection. E.g. in HCT 116 cells PCI can enhancetransfection mediated by βAE-DMRIE/DOPE, while βAE-DMRIE-mediated transfection is not affected (Hellumet al., submitted) (βAE-DMRIE: β-aminoethyl-dimyristoylRosen-thal inhibitor ether; DOPE: dioleoylphosphatidy-lethanolamine).
The observed differences in the PCI response between thedifferent transfection agents raise several interestingquestions. For transfection agents where transfection is notstimulated by PCI, it may seem that endosomal release isnot a limiting factor for “normal” transfection. The negativeeffects of PCI seen with some lipidic transfection agentsmight have several explanations. Firstly, they might be dueto a photochemically induced cytotoxicity (see 1.2.1.2)having some generally inhibitory effect on transfection. Ifsuch effects play a role they may also affect transfection bypolycationic agents, and may be reflected in the decrease intransfection by such agents that in some cases is observed athigher light doses (see 2.1). PCI-induced transfection couldthen be viewed as a balance between these general negativeeffects and the positive effects caused by increasedendosomal release. Following this reasoning, for transfectionagents that are very ineffective in endosomal release, such as
polylysine, the positive effects would dominate, while foragents very efficient in such release the negative effectswould dominate.
There are, however, also several other possibleexplanations for the negative effect of PCI observed withsome lipoplex transfection agents. One possibility is that themode of endosomal release induced by PCI has a negativeeffect that is specific for lipoplex transfection agents. Forcationic lipid transfection agents, a model proposed bySzoka and co-workers [Xu and Ellington, 1996,Zelphati andSzoka, 1996] suggests that the necessary dissociation of thelipid from the DNA is intimately coupled with the processof endosomal release. It is possible that PCI will release theintact DNA/lipid complex, thereby bypassing an essentialstep of dissociation of the complex taking place during“normal” lipoplex transfection. It is also possible that DNAin lipid complexes are more prone to photochemicallyinduced damage than DNA in polycationic complexes, eitherbecause the lipids are not effective in protecting the DNAagainst oxidative damage, or because lipid complexes arelocated closer to the photosensitiser and are therefore moreliable to damage. A finding supporting this view is that ifPCI is performed by giving the light before the transfectioncomplex (see 2.6.1), it can stimulate transfection with somelipoplex agents where an inhibitory effect is seen if light isgiven after the complex (Engesæter et al., unpublished). Thepossibilities for photochemical damage to the DNA would,of course, seem larger if the DNA is present duringillumination than if it is introduced after illumination.
The observation that transfection by polyethyleniminecomplexes can be positively affected by PCI [Prasmickaite etal., 2000] gives an indication that even if endosomal escapeis induced by PEI, this process probably is not always veryeffective. This may e.g. be related to the size of thePEI/DNA complexes employed, as pointed out by Ogris etal. [Ogris et al., 1998]. With the proposed mechanism ofPEI enhancing endosomal release by acting as a “protonsponge” (see 1.1) it would also be necessary with a certainminimum amount of PEI inside each endocytic vesicle inorder to induce endosomal swelling and lysis. The
Table 1. PCI-Induced Transfection of Different Cell Lines
Cell line Cell type Maximum transfection achieved.(% EGFP-positive cells)
Light-induced fold increasein transfection
THX Human melanoma 70 15
BHK Baby hamster kidney 25 20
HCT 116 Human colon carcinoma 50 150
WiDr Human colon carcinoma 5 10
HeLa Human cervix cancer 50 57
Cos-1 African Green Monkey kidney 35 25
3T3 Mouse fibroblast 6 65
DU145 Human prostate cancer 30 30
Different cell lines were transfected by pEGFP-N1/poly-L-lysine complexes, essentially as described in Høgset et al. (2000). % EGFP-positive cells and the fold increase intransfection achieved by the light treatment was scored.
Light Directed Gene Transfer Current Gene Therapy, 2003, Vol. 3, No. 2 9
observation that PCI shows greater enhancement of PEI-mediated transfection at low doses of DNA/PEI complex(Høgset, manuscript in preparation) is in accordance withthis proposed mechanism, since the positive effects of PCIshould be expected to increase with the decrease in theability of the PEI to effect endosomal release. Thisobservation could also have important implications for theuse of PEI for in vivo gene therapy. With in vivo delivery ofDNA/PEI complexes the amount of complex reaching thetarget cells will often be very limited, and probably oftenwell below the threshold where PEI is able to induceefficient endosomal escape. In these cases PCI could be avery valuable tool for increasing gene delivery specificallywithin the target area for the therapy.
2.4.2. PCI with Adenoviral Vectors
Adenoviral vectors are known to be taken into the cell byendocytosis. Although adenoviral endosomal release isusually regarded as a very efficient process [Greber et al.,1993,Leopold et al., 1998], there are still many cell types inwhich adenoviral gene delivery is rather inefficient. We have,
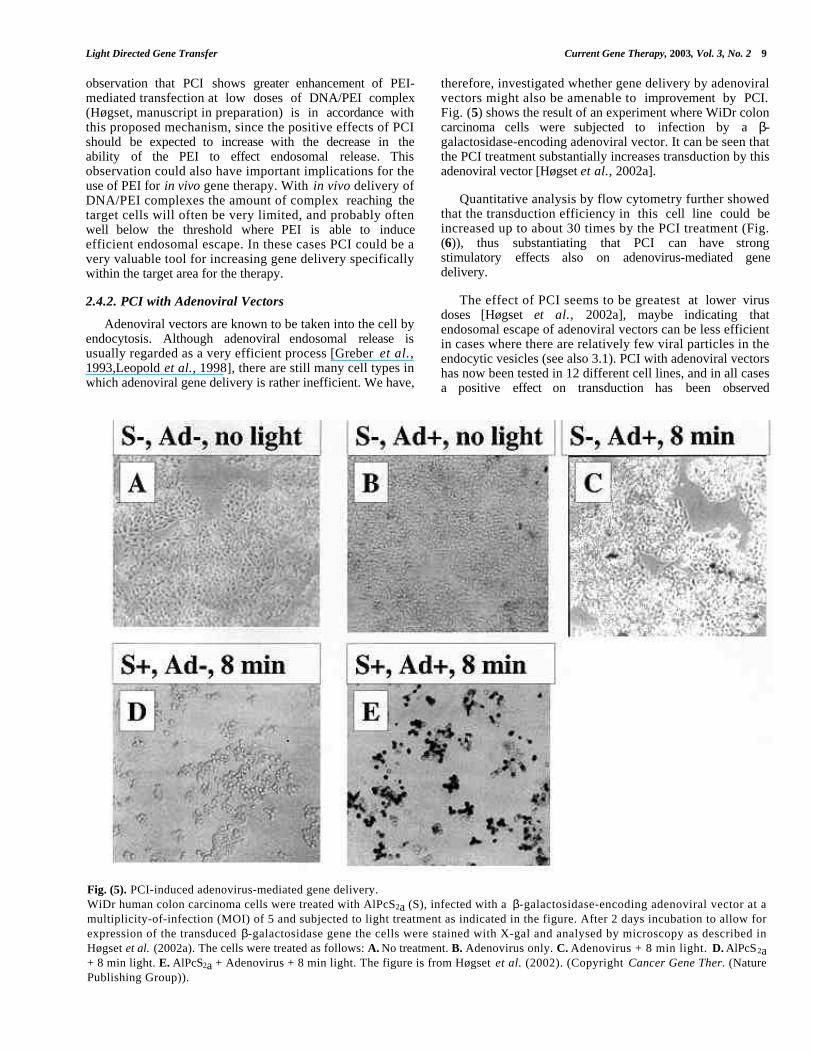
therefore, investigated whether gene delivery by adenoviralvectors might also be amenable to improvement by PCI.Fig. (5) shows the result of an experiment where WiDr coloncarcinoma cells were subjected to infection by a β-galactosidase-encoding adenoviral vector. It can be seen thatthe PCI treatment substantially increases transduction by thisadenoviral vector [Høgset et al., 2002a].
Quantitative analysis by flow cytometry further showedthat the transduction efficiency in this cell line could beincreased up to about 30 times by the PCI treatment (Fig.(6)), thus substantiating that PCI can have strongstimulatory effects also on adenovirus-mediated genedelivery.
The effect of PCI seems to be greatest at lower virusdoses [Høgset et al., 2002a], maybe indicating thatendosomal escape of adenoviral vectors can be less efficientin cases where there are relatively few viral particles in theendocytic vesicles (see also 3.1). PCI with adenoviral vectorshas now been tested in 12 different cell lines, and in all casesa positive effect on transduction has been observed
Fig. (5). PCI-induced adenovirus-mediated gene delivery.WiDr human colon carcinoma cells were treated with AlPcS2a (S), infected with a β-galactosidase-encoding adenoviral vector at amultiplicity-of-infection (MOI) of 5 and subjected to light treatment as indicated in the figure. After 2 days incubation to allow forexpression of the transduced β-galactosidase gene the cells were stained with X-gal and analysed by microscopy as described inHøgset et al. (2002a). The cells were treated as follows: A. No treatment. B. Adenovirus only. C. Adenovirus + 8 min light. D. AlPcS2a+ 8 min light. E. AlPcS2a + Adenovirus + 8 min light. The figure is from Høgset et al. (2002). (Copyright Cancer Gene Ther. (NaturePublishing Group)).

10 Current Gene Therapy, 2003, Vol. 3, No. 2 Høgset et al.
(Engesæter, in preparation). The magnitude of the effectseems, however, to vary quite substantially between differentcell lines, maybe because of different efficiency or uptake andendosomal release in the different cell lines. The uptakemechanism [Fasbender et al., 1997,Miyazawa et al.,1999,Davison et al., 2001,McDonald et al., 2001] and thefollowing intracellular routing of the viral particles areprobably also of importance for the effect of the PCItreatment (Engesæter, in preparation).
2.4.3. PCI with Targeted Vectors
To examine whether PCI would also have positive effectson transfection when the DNA is delivered by receptor-mediated endocytosis, we studied transfection withtransferrin-polylysine. In HCT 116 cells PCI had an evenbetter effect on transfection mediated by transferrin-polylysine than on transfection with unmodified polylysine[Prasmickaite et al., 2000]. The same was true for WiDrcolon carcinoma cells (Olsen, unpublished results). However,in THX melanoma cells no transfection with transferrin-polylysine could be observed, neither with, nor without PCI.Thus, PCI can work very well also with receptor-mediatedtransfection agents, but substantial cell line differences exist,maybe related to the level of receptor expression in the targetcells. The fact that PCI did not induce transfection in theTHX-cells that were resistant to transferrin-mediatedtransfection without PCI, indicates that the specificityobtainable with targeted vectors will not be altered by thePCI treatment.
2.5. The Requirement for Endocytosis
Our hypothesis for the mechanism behind PCI-inducedtransfection dictates that endocytosis should be obligatoryfor the photochemical enhancement of transfection. We have,therefore, performed a number of experiments to investigatethe role of endocytosis and endocytic transport inphotochemical transfection.
An alternative explanation for the enhanced transfectionmight have been that the PCI-treatment increases the overallcellular uptake of the transfecting plasmid, e.g. by inducinga general increase in the plasma membrane permeability.Thus, we studied the intracellular level of transfectingplasmid by Southern-blotting experiments [Prasmickaite etal., 2000]. These experiments clearly indicated that plasmiduptake was not increased by PCI, a conclusion alsosupported by flow cytometry analysis of the uptake of afluorescently labelled oligonucleotide [Prasmickaite et al.,2001a]. Taken together, these studies indicated that PCI doesnot act by increasing the general level of uptake over theplasma membrane.
Nevertheless, it would still be possible that PCI inducesthe transfer of a small subpopulation of plasmid moleculesdirectly through the plasma membrane and that it is thispopulation that gives rise to the observed PCI-inducedtransfection. There are, however, several lines of evidencearguing against this possibility. Firstly, experiments at lowtemperatures where endocytosis is inactive, but where directdiffusion over the plasma membrane can still occur, showedthat endocytosis was necessary for the PCI-inducedstimulation of transfection [Prasmickaite et al., 2000].Secondly, experiments with the inhibitor bafilomycin Aindicated that the PCI effect requires endocytosis, sincebafilomycin A completely blocked the photochemicalstimulation of transfection, while the basal level oftransfection was maintained [Prasmickaite et al., 2000].Bafilomycin A is supposed to block endosomal acidificationand endocytic transport, but apparently does not affect uptakeover the plasma membrane [Clague et al., 1994,van Deurs etal., 1996].
2.6. Importance of Timing of the Treatment
The connection between the PCI response and theendocytic process implies that the timing of the differentcomponents of the treatment could be essential. Thus, thelength of the transfection period and the timing of the lighttreatment relative to the incubation with the transfectioncomplex should be very important. Our experiments indicatethat generally it is best to irradiate the cells directly afterincubation with the DNA. If a chase period is introducedbetween incubation of the cells with DNA and the momentof irradiation, the effect of the light treatment decreases[Prasmickaite et al., 2000] (Hellum et al., submitted),indicating that PCI works best when the plasmid is in earlyendocytic vesicles. This view is further substantiated byresults showing that while the transfection efficiencyincreases sharply with time for transfection times below 1 h,only modest increases can be observed by increasing the
Fig. (6). Light dose dependence of PCI-induced adenoviral genetransfer.
WiDr cells were treated with AlPcS2a, (S) infected with a β-galactosidase-encoding adenoviral vector (Ad) and subjected tolight treatment as described in the legend to Fig. 5. % positivecells (defined as described in Fig. 4A) were scored. (s) S+, Ad-;(¨) S-, Ad+; (l) S+, Ad+). The bars represent SEM of 2 to 5different experiments. The figure is from Høgset et al. (2002).(Copyright Cancer Gene Ther. (Nature Publishing Group)).

Light Directed Gene Transfer Current Gene Therapy, 2003, Vol. 3, No. 2 11
transfection time further [Høgset et al., 2000]. A probablereason for these results is that if illumination is given longafter the uptake of the DNA a large fraction of the DNA willhave reached later endocytic vesicles where it has beendegraded by endosomal or lysosomal nucleases so that it isno longer transfection competent. Thus, a substantial fractionof the DNA would be degraded both with long incubationtimes with the transfection complex, and with introducing achase period between DNA delivery and illumination.
Another possible explanation for these results is that thephotochemical release per se is more effective from early thatfrom late vesicles, but we have as yet no indications that thisshould be the case.
2.6.1. Light Treatment Before Gene Delivery
With regard to the supposed mechanism for the PCIeffect, it was a very surprising finding that an increase ingene transfer efficiency can also be observed when theillumination is performed before the uptake of the gene intothe cell [Prasmickaite et al., 2002]. This mode of PCI-treatment can enhance gene delivery both by non-viral and byviral vectors, and can have important advantages for theemployment of PCI in in vivo gene therapy (see 5.1.1). Themechanisms behind this surprising effect are not yetclarified, a more thorough discussion of this issue can befound in Prasmickaite et al. (2002) [Prasmickaite et al.,2002].
3. MECHANISM OF THE PCI-INDUCED GENEDELIVERY
The main hypothesis for the stimulating effect of PCI ongene delivery is that the photochemical treatment enhancesrelease of the exogenous DNA from endosomes, and asdiscussed above several lines of evidence indicate that theprocess of endocytosis is intimately involved in themechanism of PCI-induced gene delivery. In this chapter this“endosomal release hypothesis” is discussed in relation tothe results obtained with different gene transfer vectors, andalternative possible mechanisms are also discussed in somedetail.
3.1. Endosomal Release as the Mechanism Behind thePCI Effects on Gene Delivery?
For polycationic vectors like polylysine and e.g. forpolyethylenimine our results seem to fit quite well withwhat is generally thought about the mechanism oftransfection with such agents. Thus, PCI gives best effectswith polylysine, a vector that is thought not to be able toeffect endosomal release. Results from experiments withdifferent amounts of PEI and SuperFect (Prasmickaite,unpublished) seem to fit nicely with the “protonsponge”/endosomal swelling hypothesis for the action of thistype of transfection agents [Boussif et al., 1995,De Smedt etal., 2000], in that PCI has a larger effect under conditions(low amounts of complex) where limited endosomalswelling could be expected to be limiting for transfection.Thus, for polycationic vectors the transfection-enhancing
effect of PCI can easily be explained by increased endosomalescape.
However, in contrast to what is the case for polyplexcomplexes, the escape of adenovirus from endosomes isbelieved to be a very efficient process, where most of thevirus particles that bind to the cell seem to reach the cellnucleus [Greber et al., 1993,Leopold et al., 1998]. How PCIcan increase adenovirus-mediated gene transduction >20times is, therefore, at present more unclear. One possibilityis that endosomal release of adenovirus in normal infectionis less efficient if there are few viral particles per endosome.This would be the case at the low MOIs where PCI has thebest effect [Høgset et al., 2002a]. The efficiency ofadenoviral endosomal escape has generally been studiedunder conditions where the amount of virus taken up by thecell has been very large [Greber et al., 1993,Leopold et al.,1998], and it is fully possible that this efficiency maysubstantially lower in cells infected with much smalleramounts of virus.
The efficiency of adenovirus gene transduction also varieswidely between cell lines. This has been attributed todifferences in amounts of adenovirus receptors in differentcells [Walter et al., 1997,Pickles et al., 1998,Li et al.,1999], but relatively little is known about the relationshipbetween the receptor content, receptor use and internalisationroutes and endosomal escape in different cell lines. It isknown that different cell types can use different routes foruptake and intracellular transport of adenoviral particles[Nemerow and Stewart, 1999,Von Seggern et al., 1999,Li etal., 2001], but little is known about the efficiency ofendosomal escape for different such routes [Wang et al.,2000]. It is thus possible that PCI can have a large positiveeffect on transduction for viral particles following some ofthese routes, while the effect with other routes is lesspronounced. Studies to clarify this issue are ongoing in ourlaboratories.
3.2. Other Possible Mechanisms Involved in PCI-Induced Gene Transfer
Although it should be evident that generally endocytosisis necessary for obtaining the PCI-induced stimulation ofgene transfer, it cannot be ruled out that also other factorsmay play a role. One aspect that deserves furtherconsideration is the possibility that the photochemicaltreatment can stimulate transcription from thecytomegalovirus (CMV) promoter employed. This isespecially relevant since it is well established thatphotochemical treatments induce cellular stress responses[Ryter and Gomer, 1993,Luna et al., 1994], and since it hasbeen reported that at least some kind of cellular stress canstimulate transcription from the CMV promoter [Boom etal., 1988,Tang et al., 1997,Bruening et al., 1998,Francisand Rainbow, 2000]. There are, however, several lines ofevidence indicating that stress induction of the CMVpromoter is not a major mechanism behind the stimulationof transfection by PCI: (i) Under conditions inducingequivalent levels of cellular stress, light-induced stimulationof transfection was seen with photosensitisers localising inendocytic vesicles, but not with non-lysosomally localised

12 Current Gene Therapy, 2003, Vol. 3, No. 2 Høgset et al.
sensitisers [Prasmickaite et al., 2001a]. If cellular stress wasthe major determinant for the photochemical effect ontransgene expression, one would have expected to see thesame effects with both classes of photosensitisers. (ii) Thestimulation is dependent on the type of transfection agentemployed. It seems unlikely that the CMV-promoter in thetransgene should respond differently to stress-inductiondepending on the kind of transfection agent used fordelivering the transgene. (iii) Illumination 24 h after deliveryof the DNA stimulates neither polylysine nor DOTAP (1,2-dioleoyl-3-trimethylammonium- propane)- mediated trans-fection in our system [Prasmickaite et al., 2000]. At thistime point one should expect the transgene to be located inthe nucleus where transcription could readily be induced bystress. In accordance with this view, Bruening et al.[Bruening et al., 1998] reported that stress applied up till atleast 40 h after transfection stimulated transcription from theCMV-promoter in transfected plasmids. The same timingshould be expected after PCI-induced transfection if stress-induced stimulation was the main mechanism behind theeffect. (iv) In experiments on cells containing achromosomally integrated GFP-gene controlled by theCMV-promoter no increase in the level of GFP-expressioncould be observed when the cells received a photochemicaltreatment inducing the PCI effect (Prasmickaite,unpublished). This argues against a photochemically inducedstimulation of transcription from the CMV-promoter underthe conditions employed for photochemically inducedtransfection.
There are also other possible mechanisms that couldcontribute to the observed effect of PCI on gene transfer.Although there is ample evidence against it (see 2.5), aminor effect on plasma membrane permeability can still notbe completely excluded. It is also a possibility that PCI mayin some way positively affect cytoplasmic transport towardsthe nucleus or nuclear membrane permeability, althoughthere are at present no specific data indicating that suchmechanisms are of importance.
Finally, there are ample possibilities for additive orcounteracting effects where the overall PCI-effects on genetransfer could be a combination of effects both on endosomalrelease and e.g. on transcription, and where the determinantsleading to the observed net effects may be different betweendifferent combinations of cells, photosensitisers andtransfection agents.
4. USES OF PCI FOR GENE DELIVERY
Probably the most intriguing use of PCI is to utilize thetechnology as a method for site-directed enhancement of genedelivery in in vivo gene therapy. The technology should,however, also be useful as an in vitro gene transfer methodboth with non-viral and with viral vectors.
4.1. PCI for In Vitro Gene Transfer
As discussed above, PCI can induce transfection in avariety of different cell lines, although the absolute level oftransfection varies. At least in some cases transfection
efficiencies greater than what can be obtained by othercommonly used transfection methods can be achieved (M.Hellum, unpublished observations). Thus, PCI may be auseful method for in vitro transfection, adding to thearmament of laboratory gene transfer methods. It will beespecially interesting to see if PCI can stimulate transfectionin cell types that are “notoriously” difficult to transfect withexisting methods. Although the light-induced cytotoxiceffect (see 4.3.3.2) is a drawback for employing PCI as atransfection method, toxicity is a well-known problem alsowith most other efficient transfection methods. Thus, thetoxic effects associated with PCI might in the end well bebalanced by the possibility of using lower amounts of toxictransfection agents, due to the increased transfectionefficiency achievable by PCI. Moreover, PCI as atransfection method is still under development, and it isreasonable to think that further optimisation can lead toconditions where the toxicity will be reduced, whilemaintaining the stimulating effects on transfection.
A quite unique property of the PCI technology is itsability to enhance gene transfer also by adenoviral vectors.Even though adenovirus in many cases is a very efficientgene transfer vector, there are still many cell types that arevery difficult to transduce, or where prohibitively high virusdoses are required to get proper transduction. PCI, oftengiving very good effects in cells that are hard to infect[Høgset et al., 2002a] (Engesæter, in preparation), could be avery valuable tool for in vitro adenovirus gene transfer,making it possible to efficiently transduce “difficult” celltypes, and allowing substantial reductions in the amounts ofvirus needed to get proper transduction.
4.2. PCI as a Tool for Investigating CellularMechanisms of Transfection
PCI can also be used as a method for analysing themolecular mechanisms of transfection, e.g. the importance ofvarious cellular events and compartments in the process oftransfection. Thus, PCI could be used to examine theimportance of endosomal escape as a barrier for transfection,endocytic and cytoplasmic stability of the transgene andnuclear uptake for various transfection agents. A specialfeature of PCI is the instant effect of the light treatment.This should make PCI a particularly valuable tool fortracking down the timing of several critical events in theprocess of transfection.
4.3. Use of PCI for In Vivo Gene Delivery
A very important goal in many forms of in vivo genetherapy is to be able to efficiently deliver a therapeutic geneso that it will be efficiently expressed at a desired location inthe body, but not at other sites (see e.g. [Verma and Somia,1997,Curiel, 1999]). The light-directed enhancement of genedelivery obtainable by the PCI technology could giveimportant contributions to reaching this goal. As discussedabove in vitro PCI has given increases in gene transfer of>100 times with polyplex transfection agents, while 5-30fold increases can be observed for adenovirus mediated genetransfer to a variety of cell types [Høgset et al., 2002a]. It is,

Light Directed Gene Transfer Current Gene Therapy, 2003, Vol. 3, No. 2 13
of course, a crucial question whether the effects seen in vitrowould also be present in an in vivo situation. In this chapterwe will first discuss the results obtain so far with the PCItechnology in vivo, continuing with a discussion of thepotential advantages and disadvantages of using PCI as adelivery technology in in vivo gene therapy, and acomparison of PCI to other technologies for site-specificgene delivery.
4.3.1. PCI In Vivo
Although as yet no conclusive studies have been done onthe effect of PCI on in vivo gene transfer, experiments wherePCI has been used to internalise a protein toxin clearly showthat the PCI technology works very well also in vivo [Selboet al., 2001]. In these studies we used the plant toxingelonin, which is a type I ribosome inactivating toxin, beingan extremely potent inhibitor of protein synthesis in cell-freesystems [Stirpe et al., 1980]. This toxin does not bind to
specific receptors on the cell surface, but is taken up mainlyby fluid phase endocytosis. Since gelonin has no ability toescape from endocytic vesicles it will normally be routed tolysosomes and will be degraded without exerting a toxiceffect [Lambert et al., 1985]. Thus, normally this toxin hasvery low toxicity for intact cells and for animals [Barbieriand Stirpe, 1982,Scott et al., 1987], hence it is very wellsuited for proving the effect of PCI in in vivo experiments. Amouse tumour model for studying the effect of thecombination of gelonin and PCI on tumour growth wasemployed. The results showed (Fig. (7)) that while theinjection of gelonin alone had no effect on tumour growth,the combination with PCI made about 70 % of the treatedmice tumour-free for the rest of their lives [Selbo et al.,2001]. Thus, it can be concluded that the PCI deliverytechnology also works well in vivo. Similar experimentscombining PCI with different gene delivery vectors are inprogress in our laboratory.
Fig. (7). PCI in vivo.
Kaplan-Meier type plot of the response to treatment with the combination of PCI and the protein toxin gelonin of WiDradenocarcinomas subcutaneously growing in athymic mice. The end-point is the time after light exposure when the individualtumours have reach a tumour volume of > 1000 mm3. AlPcS2a PS on the figure was injected i.p. at 10 mg/kg in mice with 100 mm3
tumors on the right hip. After 48 h the tumors were exposed to 135 J/cm2 from a 150 W halogen lamp (Xenophot, HLX64640) filteredthrough a 580-nm long-pass and a 700-nm short-pass filter, emitting 150 mW/cm2. The animals were covered with aluminium foilexcept above the tumor area. The treatment combinations are described in the figure, further experimental details can be found inSelbo et al. (2001).

14 Current Gene Therapy, 2003, Vol. 3, No. 2 Høgset et al.
4.3.2. Potential Benefits of PCI for In Vivo Gene Transfer
4.3.2.1. Gene Delivery to Regions Receiving LimitedAmounts of Gene Therapy agent
One of the main obstacles for efficient gene delivery invivo is that gene complexes are distributed very poorlythrough the target tissue, so that large areas of the targetregion may receive too low concentrations of gene complexto be efficiently transfected. In such a setting the enhancedtransfection efficiency obtainable by PCI may be quitevaluable, possibly allowing also areas of poor complexpenetration to be efficiently transfected. For both adenoviralvectors [Høgset et al., 2002a] and some polycations (Høgset,unpublished) PCI seems to give the largest enhancement ofgene transfer under conditions where a limited amount ofvector is taken up by the target cells, and this indicates thatPCI could especially be well suited for gene delivery toregions with limited vector penetration.
4.3.2.2. Specificity in Photosensitiser Localisation
Probably the most important advantage of PCI is that,because of the light-directed nature of the technology,increases in gene delivery to the target area can be obtainedwithout increasing transfection also in non-target areas, thusavoiding undesirable side effects. In addition to thespecificity achieved by the pure illumination, at least forcancer and some cardiovascular diseases there is also anotherpossible element of specificity inherent in the PCItechnology. Thus, photosensitisers in many cases tend toaccumulate preferentially in areas of disease (see 1.2.1.2).Since both the cytotoxicity and the efficiency ofphotochemical gene delivery are quite dependent on theconcentration of the photosensitiser in the illuminated cells[Høgset et al., 2000], this surrounding normal tissue may toa large extent be spared from the effects of the light treatmentdue to lower photosensitiser concentrations in the normal ascompared to the diseased tissue. For example in braintumours this effect could be very important since, e.g. inglioblastomas, tumour to normal tissue ratios in thephotosensitiser concentration of >20:1 can be observed[Chopp et al., 1996].
There might in the future also be possible to target thephotosensitiser to be more specifically taken up only into thedesired target cells. To this end work has already been donewith coupling photosensitisers to antibodies binding toantigens expressed specifically on various target cells [Goffet al., 1994,Goff et al., 1996,Del Governatore et al.,2000,Soukos et al., 2001].
4.3.2.3. Cytokine Induction
Photochemical treatments are known to induce localinflammatory responses. This often includes the induction ofthe production of various immunostimulatory cytokines suchas IL-6 and TNF-α, as well as different heat-shock proteins[Evans et al., 1990,Kick et al., 1995,Gomer et al., 1996,deVree et al., 1997,Dougherty et al., 1998]. For manyapplications of PCI (e.g. cancer treatment, DNA vaccination,see below) such responses can act synergistically with thetherapeutic gene in generating therapeutic responses. Thus,both cytokines [Cao et al., 1998,Walther and Stein, 1999]and heat-shock proteins [Todryk et al., 1999,Todryk et al.,
2000] are known to be able to attract and alert immune cellsto attack e.g. cancer cells that would otherwise usually nothave been recognized by the immune system.
4.3.2.4. Combination of PCI with Other Technologies forTargeted Gene Therapy
The specificity of PCI-mediated gene transfer could alsobe increased by combining PCI treatment with other meansfor obtaining specific delivery and/or expression of thetherapeutic gene. Thus, most methods for targeting thedelivery of therapeutic genes [Dachs et al., 1997,Zauner etal., 1998,Godbey et al., 1999,Sudimack and Lee, 2000]could probably be combined with PCI. Indeed, theemployment of PCI with the targeted transfection agenttransferrin-polylysine shows that PCI works very well alsowith a transfection agent taken up by receptor-mediatedendocytosis [Prasmickaite et al., 2000]. Also transcriptionaltargeting [Miller and Whelan, 1997,Nettelbeck et al., 2000]should be very well suited for combination with PCI. Inaddition to using promoters that will confer tissue- ordisease-specific expression, a very interesting approachwould be to use promoters inducible by the photochemicaltreatment [Luna et al., 2000] to add further to the specificityachievable with the light treatment.
It is also an advantage with PCI that the same deliverysystem can be used to deliver many different types ofmolecules, such as proteins [Berg et al., 1999,Selbo et al.,2000a,Selbo et al., 2000b], peptides [Berg et al., 1999],oligonucleotides and low molecular weight drugs (Høgset,unpublished). Thus site-specific gene delivery could inprinciple be combined with the site-specific delivery of ahost of other classes of molecules.
4.3.2.5. PCI and the Dose of Gene Therapeutic Agents
The gain in efficiency obtainable by PCI may make itpossible to use lower doses of a gene therapeutic agent, bothwith local and with systemic administration. This shouldlead to a decrease in possible adverse effects, both due to thegene delivery vehicle, and due to the expression of atherapeutic gene in non-target cells. Furthermore, thepossibilities of dose reduction should also decrease problemswith immunogenicity of the gene therapeutic agent, aproblem especially encountered with viral vectors.
4.3.3. Potential Drawbacks of PCI for In Vivo GeneDelivery
4.3.3.1. Limited Light Penetration into the Tissues
A potential drawback for using PCI in in vivo genetherapy is the limited penetration of light into the tissues.Although this limited light penetration is an advantage inthe sense that it makes it possible to confine the effect of thetreatment quite precisely to the desired area, it obviouslymay limit the kind of lesions where PCI could be employed.In photodynamic therapy the range of the treatment usuallyis limited to about 1 cm from the tissue surface, dependingamong other things on the photosensitiser used. For PCI thereach would be expected to be greater, since transfection canbe substantially induced also at relatively non-toxic lightdoses, meaning that effective light doses can be obtainedfurther into the tissue. Thus, our in vivo experiments with

Light Directed Gene Transfer Current Gene Therapy, 2003, Vol. 3, No. 2 15
gelonin (see 4.3.1) indicate that tumours up to a thickness ofat least 8 mm may totally be eradicated by PCI treatment.Experience with photodynamic therapy and theoreticalconsiderations seem to indicate that under optimalconditions the PCI technology should be able to induceefficient gene transfer up to 1-2 cm into the tissues. Inaddition to the external illumination many areas of diseasecan also be illuminated from inside the body e.g. by meansof fibre optic devices (see e.g. 5.1.2 and 5.1.3), thussubstantially expanding the kinds of lesions that can betreated.
Although the effect of PCI is clearly light dosedependent, PCI-induced gene delivery is effective over aquite large range of light doses (Fig. 4B), a fact that mayhave important implications for the clinical use of thetechnology. Firstly, it means that gene delivery can beefficiently induced in the target tissue at different distancesfrom the light source, even though the light dose deliveredto the cells will decrease with this distance. Secondly, itmakes it possible to adjust the balance between the efficiencyof gene delivery and the photochemically induced cytotoxiceffects (see 4.3.3.2).
4.3.3.2. Cytotoxicity
Light induced cytotoxic effects is an obvious drawbackfor some, but by no means all, in vivo applications. For genetherapy approaches where it is essential that the target cellsare not harmed, these effects generally cannot be tolerated,however, in many clinical situations (e.g. in cancer therapy)a certain amount of cell killing may not be harmful, or mayeven be beneficial. Also for cardiovascular applications thecytotoxicity associated with the photochemical treatmentseems to be well tolerated and might have beneficial effects.Thus, studies of photodynamic therapy of restenosis andatherosclerosis indicate that the photochemical treatment initself may have substantial clinical benefit without causingserious adverse effects on the treated blood vessels[Yamaguchi et al., 2000,Rockson et al., 2000a,Rockson etal., 2000b] (see also 5.1.2).
A very important point is that although thephotochemical treatment induces cytotoxic effects, these aregenerally restricted to the illuminated areas of the body[Moan and Berg, 1992], the photosensitisers in themselvesusually have very little systemic toxicity, in contrast to otheragents (e.g. chloroquine) effective in increasing endosomalescape. Thus, damage to vital organs could generally beavoided. Also, there is substantial clinical experience withrelevant photosensitisers, showing that they can be usedsafely in humans [Dougherty et al., 1998].
4.3.4. Comparison with other Technologies for Site-Specific Gene Delivery
In comparison to other physical methods for site-specificgene delivery such as particle bombardment [Rakhmilevichet al., 1996, Muramatsu et al., 1998, Lin et al., 2000, Mir,2001], electroporation [Muramatsu et al., 1998,Mir, 2001],ultrasound [Unger et al., 2001, Porter and Xie, 2001,Newman et al., 2001] or magnetically targeted gene delivery[Scherer et al., 2002] PCI has both advantages anddisadvantages.
Firstly, an advantage of PCI as opposed to all physicalgene delivery methods that works by helping the DNA tocross the plasma membrane directly (e.g. particlebombardment, electroporation and possibly ultrasound), thePCI technology involves endocytosis and therefore is verywell suited for combination with other targeting strategies,e.g. surface targeting via specific ligand-receptor interaction(see 4.3.2.4). Conversely, these other technologies would, ofcourse, be advantageous to PCI in cases where the endocyticuptake of the gene therapy vector is inefficient.
In comparison to ultrasound-mediated gene delivery[Unger et al., 2001,Porter and Xie, 2001,Newman et al.,2001] PCI has the disadvantage that the light penetratesthrough the tissues to a smaller extent than ultrasound, a factthat limits the reach of PCI as compared to ultrasound-mediated delivery. However, as compared to particlebombardment or electroporation the reach of PCI wouldprobably be better. Regarding magnetically targeted genedelivery technologies, these work mainly by generating ahigh concentration of the DNA at the desired location in thebody and at the surface of the target cells, they would stillrely on endocytosis for getting the DNA into the target cells.Thus these technologies might constitute a very interestingcomplement to PCI, where PCI could substantially increasethe gene transfer efficiency of the magnetically targetedDNA.
Recently there have also been a few reports on two novelinteresting technologies for light-induced virus oroligonucleotide transfer; both these technologies are based onprinciples different from PCI. Pandori and co-workers havedescribed a technology where viral vectors can be renderednon-infective by treatment with a photocleavable chemicalagent. Exposure of such viral particles to UV-light willcleave off the agent and restore the infectivity of the viralparticles [Pandori and Sano, 2000,Pandori et al., 2002]. Forin vivo use this technology has the disadvantage that theillumination is done by UV light. UV light has only a verylimited penetration into the tissues, and gene delivery bythis technology would therefore be limited to the verysurface of the target tissue. Normand et al. [Normand et al.,2001] have developed a technology where complexes ofoligonucleotides and the herpes simplex virus VP22 proteincan be disrupted intracellularly by illumination, leading todelivery of the oligonucleotides to the cell nucleus. Thistechnology has not yet been shown to work for gene deliveryalthough it works well for oligonucleotide delivery. Themechanism behind this technology is still unclear, and it isas yet difficult to say how this technology will be comparedto PCI.
5. POTENTIAL CLINICAL USES OF PCI-MEDIATED GENE DELIVERY
PCI could in principle be used as a gene deliverytechnology in all kinds of gene therapy where local genedelivery is necessary or sufficient. The only limitation is thatthe area of gene delivery could in some way be illuminated.Obviously PCI could be used on superficial lesions or skindisorders. However, by means of optical fibres or otherdevices many internal sites can also be reached, such as thelungs, the pancreas, the gastrointestinal tract, the urogenital

16 Current Gene Therapy, 2003, Vol. 3, No. 2 Høgset et al.
system, joints and blood vessels [Pass, 1993,Jenkins et al.,1998,Trauner et al., 1998,Rockson et al., 2000a]. Inaddition PCI-mediated gene delivery could easily be used asa complement to primary surgical treatment and as anadjuvant treatment to systemic therapy. In this chapter thepotential employment of PCI in different clinical situationsis discussed.
5.1. Diseases that Could be Treated with PCI-MediatedGene Therapy
The target diseases for PCI-mediated gene therapy wouldgenerally of course in some way be localised, since localillumination is an essential part of the technology. Althoughthis is an obvious limitation, to date most cancer genetherapy trials have been performed with local delivery of thetherapeutic gene, and PCI could in principle be used in allthese cases. Further, although PCI per se is a local deliverytechnology, systemic responses can also be envisioned, e.g.by exploiting the immune system, such as in immunogenetherapy of cancer. PCI could also be used as a deliverysystem for more traditional DNA vaccine approaches.
5.1.1. PCI-Mediated Gene Therapy of Cancer
Cancer is a natural target for PCI-mediated gene therapy,with skin cancers being one obvious target. Althoughmetastatic spread and systemic disease, of course, is a majorproblem with cancer, there are also many cases wherepatients may die of local disease or where control of localdisease is important for palliative reasons, e.g. brain, headand neck, ovarian and pancreatic cancer. There are also anincreasing amount of cases where cancers are diagnosed at astage where it is still a local disease, e.g. in prostate cancer.Improved diagnostic procedures will in future probably leadto even more cancers being detected at an early stage,implying an increased importance for specific localtreatments. In all such cases local PCI-mediated gene therapycould constitute an attractive specific local treatmentmodality.
PCI-mediated gene therapy could also be combined withsurgery. For many localised cancers surgery will, of course,be the first option for treatment. However, in many cases,e.g. in brain, head and neck and ovarian cancer, localrecurrence often represents a serious problem. In such caseslocal gene therapy killing remaining undetected tumour cellsor inhibiting the proliferation of tumour cells in the treatedarea could give substantial therapeutic benefit. Of course,when PCI is combined with surgery, illumination of thetarget area should represent no problem because of the easyaccessibility to the lesions. Furthermore, employing the“light first” mode of PCI (see 2.6.1) the whole treatmentcould be done in one operation where illumination is donedirectly after surgery, followed immediately by delivery ofthe gene therapeutic agent.
As compared to cancer treatment by PDT, PCI haveseveral potential advantages. Firstly, since PCI is effective atlight doses that in themselves are not lethal PCI will make itpossible to get a therapeutic effect further into the tissue thatwhat is achievable by PDT. Secondly, while PDT kills cellsby a general mechanism (in principle killing all cells that
contain the photosensitiser) a PCI based gene therapeutictreatment can be made much more specific. Thus, in contrastto what is the case for PDT both specific gene deliverysystems, target specific promoters and target specific actionof the therapeutic gene product can be employed to increasethe specificity of the treatment. This can be especiallyimportant e.g. if using PCI for killing off remaining tumorcells after surgery, where the tumor cells may often beinterspersed with normal cells that may have a vital functionin the tissue.
The fact that PCI is a local gene delivery method doesnot, however, rule out the possibility of treating systemicdisease. Many animal studies [Huang et al., 1996,Oshikawaet al., 2001,Maron et al., 2001,Warren et al., 2002] and alsosome patient studies [Rochlitz et al., 1999,Stopeck et al.,2001] indicate that distant metastases may be cured afterlocal administration of a therapeutic gene, probably viaimmunological reactions. In such a setting gene deliverymediated by PCI might be especially beneficial, since thephotochemical treatment is known to haveimmunostimulating effects (see above 4.3.2.3) and therebycan help in generating a systemic immune response to thetumour cells, as a “beneficial side effect” of the deliverymethod.
5.1.2. PCI in Gene Therapy of Cardiovascular Diseases
Gene therapy is under investigation for various kinds ofcardiovascular diseases [Meyerson, 1999, Morishita et al.,2001,Kozarsky, 2001,McKay, 2001,Nikol, 2001,Isner,2002], using many different therapeutic genes [Tio et al.,1998,Nakamura, 2002]. For several prevalent diseaseconditions, e.g. restenosis after angioplasty, in graft veinsand in coronary stents [Quarck, 2001,Klugherz, 2002], it iscrucial to obtain efficient local gene delivery into definedregions of blood vessels. PCI will be very well suited forthis, since light can precisely be delivered both through fibreoptic catheters [Rockson et al., 2000a] and through standardclinical balloon catheters [Jenkins et al., 1998]. Furthermore,photodynamic treatment on its own has already shownpromising results in treating such conditions, both in animalmodels and in humans [Rockson et al., 2000b]. Eventhough the photochemical treatment damages cells in theilluminated area, it seems to be well tolerated with nopermanent damage to the treated vessels. Thus, employingPCI for gene delivery in restenosis could have a doublebeneficial effect. Firstly, the photochemical treatment couldablate the lesions, and secondly PCI would ensure theefficient local delivery of therapeutic genes that could e.g.inhibit the proliferation of smooth muscle cells, therebypreventing recurrence of the lesions.
PCI could also be used more generally for the efficientlocal delivery of therapeutic genes to segments of bloodvessels, without introducing persistent damage to thevessels. This is even more so since the light doses used withPCI would generally be lower than those that have alreadybeen found to be well tolerated in photodynamic therapy.
5.1.3. PCI in Autoimmune Diseases
PCI-mediated gene therapy could also be very useful inthe treatment of autoimmune diseases such as rheumatoid

Light Directed Gene Transfer Current Gene Therapy, 2003, Vol. 3, No. 2 17
arthritis [Tarner and Fathman, 2001,Burstein, 2001]. Inanimal studies the local delivery of genes encoding anti-inflammatory proteins such as IL-10 or IL-1 receptorantagonist has given very promising results [Evans,2001,Ghivizzani, 2001,van de Loo, 2002], and also clinicalgene therapy studies are underway [Evans et al., 2000,Tarnerand Fathman, 2001].
As is the case for cancer and cardiovascular diseasesphotodynamic treatments has also been investigated forrheumatoid arthritis, and results from animal studies showpromising results, employing both fibre-mediatedillumination and external irradiation of small joints[Hendrich, 1997,Ratkay et al., 1998,Trauner et al., 1998].The results show that effective illumination can be performedwith minimally invasive procedures, and that thephotochemical treatment can destroy inflamed synovial cellswithout damaging normal surrounding tissue. Thus, againPCI-mediated gene therapy might have a “double benefit”both in destroying the inflamed cells in a joint in the firsthand, and then by delivering a therapeutic gene whoseexpression could inhibit further inflammatory processes. Anespecially interesting option could be to combine PCI-mediated gene delivery with an “inflammation regulatable”gene therapy system as recently described by Miagkov et al.[Miagkov, 2002].
5.1.4. PCI for Cutaneous Gene Delivery
Obviously, dermatological conditions should be a naturaltarget for PCI-mediated gene therapy. In addition to skincancers, also benign conditions such as psoriasis could betargeted. Thus, PCI would be nearly ideally suitable for thelocal delivery of genes e.g. encoding immunosuppressingproteins to psoriatic lesions [Asadullah et al., 2000,Rao etal., 2002]. PCI could also be used for delivery of genes toskin for treating non-dermatological diseases [Khavari et al.,2002] and for DNA vaccination purposes [Badiavas andFalanga, 2001,Spirito et al., 2001].
5.1.5. PCI as a Delivery System for DNA Vaccines
The development of DNA vaccines has created muchattention during the last years [Berzofsky et al.,2001,Sharma and Khuller, 2001,Gregersen, 2001,McKenzieet al., 2001,Mor and Eliza, 2001,Ulmer, 2001a,Ulmer,2001b], and both prophylactic and therapeutic DNA vaccinesare under clinical development for a host of diseases, such ascancer [Leitner et al., 2001,Haupt et al., 2002] andinfectious diseases [Clark and Johnson, 2001,Michel andLoirat, 2001]. In order to maximize the chances of obtainingthe desired immunological responses, an efficient deliverysystem for the DNA encoding the immunogen is crucial.PCI should be very well suited for this purpose. Firstly, thephotochemical treatment in itself is known to induce theproduction of immunostimulatory factors (4.3.2.3 seeabove), thus potentially acting as an adjuvant. Secondly,DNA for vaccination purposes will typically be deliverednear the surface of the body, at locations very well suited forphotochemical treatments. Studies on the use of PCI forvaccination purposes are in progress in our laboratories.
6. CONCLUDING REMARKS
Photochemical internalisation can be used for enhancinggene delivery with a variety of gene therapy vectors, bothsynthetic and viral, and also including targeted gene deliveryvectors. The light-induction inherent in the PCI technologyshould make it possible to direct this enhancement tospecific regions of the body, e.g. to tumours or to sites ofvascular disease. Thus, PCI can be used to enhance thedelivery of therapeutic genes specifically in areas of disease,avoiding possible adverse effects resulting from theexpression of genes encoding e.g. cytotoxic proteins innormal tissues. PCI is based on the use of photosensitisingcompounds that can be used safely in humans, and for manyof which there is considerable clinical experience. Thus, PCIshould be a very valuable addition to the arsenal of genedelivery methods for in vivo gene therapy.
ABBREVIATIONS
5-ALA = 5-Aminolevulinic acid
AlPcS2a = Aluminium phthalocyanine with twosulfonate groups on adjacent phthalaterings
βAE-DMRIE = β-Aminoethyl-dimyristoyl Rosenthalinhibitor ether
CMV = Cytomegalovirus
DOTAP = 1,2-Dioleoyl-3-trimethylammonium-propane
EGFP = Enhanced green fluorescent protein
MTT = 3-[4,5-Dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide
PCI = Photochemical internalisation
PDT = Photodynamic therapy
pEGFP-N1 = Plasmid encoding EGFP
PEI = Polyethylenimine
PpIX = Protoporphyrin IX
SEM = Standard error of the mean
3THPP = Tetra(3-hydroxyphenyl)porphyrin
TPPS2a = Meso-tetraphenylporphine with twosulfonate groups on adjacent phenylrings
HSV-tk = Herpes Simplex Virus thymidine kinase
IL-12 = Interleukin-12

18 Current Gene Therapy, 2003, Vol. 3, No. 2 Høgset et al.
REFERENCES
Anderson, W.F. (1998). Human gene therapy. Nature 392: 25-30.
Asadullah, K., Docke, W.D., Sabat, R.V., Volk, H.D. and Sterry, W.(2000). The treatment of psoriasis with IL-10: rationale andreview of the first clinical trials. Expert Opin. Investig. Drugs 9,95-102.
Badiavas, E.V. and Falanga, V. (2001). Gene therapy. J. Dermatol.28 : 175-192.
Barbieri, L. and Stirpe, F. (1982). Ribosome-inacting proteinsfrom plants: Properties and possible uses. Cancer Surv. 1: 489-502.
Ben-Hur, E., Moor, A.C., Margolis-Nunno, H., Gottlieb, P., Zuk,M.M., Lustigman, S., Horowitz, B., Brand, A., Van Steveninck, J.and Dubbelman, T.M. (1996). The photodecontamination ofcellular blood components: mechanisms and use ofphotosensitisation in transfusion medicine. Transfus. Med. Rev.10 : 15-22.
Berg, K., Bommer, J.C., Winkelman, J.W. and Moan, J. (1990a).Cellular uptake and relative efficiency in cell inactivation byphotoactivated sulfonated meso-tetraphenylporphines.Photochem. Photobiol. 52 : 775-781.
Berg, K., Western, A., Bommer, J.C. and Moan, J. (1990b).Intracellular localisation of sulfonated meso-tetraphenylporphines in a human carcinoma cell line.Photochem. Photobiol. 52 : 481-487.
Berg, K. and Moan, J. (1994). Lysosomes as photochemicaltargets. Int. J. Cancer 59 : 814-822.
Berg, K., Sandvig, K. and Moan, J. (1996). Transfer of moleculesinto the cytosol of cells. Patent PCT/NO95/00149.
Berg, K., Selbo, P.K., Prasmickaite, L., Tjelle, T.E., Sandvig, K.,Moan, J., Gaudernack, G., Fodstad, Ø., Kjølsrud, S., Anholt, H.,Rodal, G.H., Rodal, S.K. and Høgset, A. (1999). Photochemicalinternalisation: A novel technology for delivery ofmacromolecules into cytosol. Cancer Res. 59 : 1180-1183.
Berzofsky, J.A., Ahlers, J.D. and Belyakov, I.M. (2001).Strategies for designing and optimising new generationvaccines. Nature Rev. Immunol. 1: 209-219.
Boehncke, W.H., Elshorst-Schmidt, T. and Kaufmann, R. (2000).Systemic photodynamic therapy is a safe and effective treatmentfor psoriasis. Arch. Dermatol. 136: 271-272.
Boom, R., Sol, C.J., Minnaar, R.P., Geelen, J.L., Raap, A.K. andvan der Noordaa, J. (1988). Induction of gene expression underhuman cytomegalovirus immediate early enhancer-promotercontrol by inhibition of protein synthesis is cell cycle-dependent. J. Gen. Virol. 69 : 1179-1193.
Boussif, O., Lezoualc'h, F., Zanta, M.A., Mergny, M.D., Scherman,D., Demeneix, B. and Behr, J.P. (1995). A versatile vector forgene and oligonucleotide transfer into cells in culture and invivo: polyethylenimine. Proc. Natl. Acad. Sci. USA 92 : 7297-7301.
Bruening, W., Giasson, B., Mushynski, W. and Durham, H.D.(1998). Activation of stress-activated MAP protein kinases up-regulates expression of transgenes driven by thecytomegalovirus immediate/early promoter. Nucleic Acids Res.26 : 486-489.
Burstein, H. (2001). Gene therapy for rheumatoid arthritis. Curr.Opin. Mol. Ther. 3: 362-374.
Cao, L., Kulmburg, P., Veelken, H., Mackensen, A., Mezes, B.,Lindemann, A., Mertelsmann, R. and Rosenthal, F.M. (1998).Cytokine gene transfer in cancer therapy. Stem Cells 16 Suppl 1:251-260.
Chopp, M., Madigan, L., Dereski, M., Jiang, F. and Li, Y. (1996).Photodynamic therapy of human glioma (U87) in the nude rat.Photochem. Photobiol. 64 : 707-711.
Clague, M.J., Urbé, S., Aniento, F. and Gruenberg, J. (1994).Vacuolar ATPase activity is required for endosomal carriervesicle formation. J. Biol. Chem. 269: 21-24.
Clark, K.R. and Johnson, P.R. (2001). Gene delivery of vaccinesfor infectious disease. Curr. Opin. Mol. Ther. 3: 375-384.
Cotten, M., Langle-Rouault, F., Kirlappos, H., Wagner, E.,Mechtler, K., Zenke, M., Beug, H. and Birnstiel, M.L. (1990).Transferrin-polycation-mediated introduction of DNA intohuman leukemic cells: stimulation by agents that affect thesurvival of transfected DNA or modulate transferrin receptorlevels. Proc. Natl. Acad. Sci. USA 87 : 4033-4037.
Cristiano, R.J., Smith, L.C. and Woo, S.L. (1993). Hepatic genetherapy: adenovirus enhancement of receptor- mediated genedelivery and expression in primary hepatocytes. Proc. Natl.Acad. Sci. USA 90 : 2122-2126.
Curiel, D.T., Agarwal, S., Wagner, E. and Cotten, M. (1991).Adenovirus enhancement of transferrin-polylysine-mediatedgene delivery. Proc. Natl. Acad. Sci. USA 88 : 8850-8854.
Curiel, D.T. (1999). Strategies to adapt adenoviral vectors fortargeted delivery. Ann. N.Y. Acad. Sci. 886: 158-171.
Dachs, G.U., Dougherty, G.J., Stratford, I.J. and Chaplin, D.J.(1997). Targeting gene therapy to cancer: a review. OncologyResearch 9: 313-325.
Davison, E., Kirby, I., Whitehouse, J., Hart, I., Marshall, J.F. andSantis, G. (2001). Adenovirus type 5 uptake by lungadenocarcinoma cells in culture correlates with Ad5 fibrebinding is mediated by alpha(v)β1 integrin and can bemodulated by changes in β1 integrin function. J. Gene Med. 3:550-559.
De Smedt, S.C., Demeester, J. and Hennink, W.E. (2000). Cationicpolymer based gene delivery systems. Pharm. Res. 17 : 113-126.
de Vree, W.J., Essers, M.C., Koster, J.F. and Sluiter, W. (1997).Role of interleukin 1 and granulocyte colony-stimulating factorin photofrin-based photodynamic therapy of ratrhabdomyosarcoma tumors. Cancer Res. 57 : 2555-2558.
Del Governatore, M., Hamblin, M.R., Piccinini, E.E., Ugolini, G.and Hasan, T. (2000). Targeted photodestruction of human coloncancer cells using charged 17.1A chlorine immunoconjugates.Br. J. Cancer 82 : 56-64.
Donati, G., Kapetanios, A.D. and Pournaras, C.J. (1999).Principles of treatment of choroidal neovascularization withphotodynamic therpy in age-related macular degeneration.Semin. Thromb. Hemost. 14 : 2-10.
Douar, A.M., Poulard, K., Stockholm, D. and Danos, O. (2001).Intracellular trafficking of adeno-associated virus vectors:routing to the late endosomal compartment and proteasomedegradation. J. Virol. 75 : 1824-1833.

Light Directed Gene Transfer Current Gene Therapy, 2003, Vol. 3, No. 2 19
Dougherty, T.J., Gomer, C.J., Henderson, B.W., Jori, G., Kessel, D.,Korbelik, M., Moan, J. and Peng, Q. (1998). Photodynamictherapy. J. Natl. Cancer Inst. 90 : 889-905.
Duan, D., Li, Q., Kao, A.W., Yue, Y., Pessin, J.E. and Engelhardt,J.F. (1999). Dynamin is required for recombinant adeno-associated virus type 2 infection. J. Virol. 73 : 10371-10376.
Erbacher, P., Roche, A.C., Monsigny, M. and Midoux, P. (1996).Putative role of chloroquine in gene transfer into a humanhepatoma cell line by DNA/lactosylated polylysine complexes.Exp. Cell Res. 225: 186-194.
Evans, S., Matthews, W., Perry, R., Fraker, D., Norton, J. and Pass,H.I. (1990). Effect of photodynamic therapy on tumor necrosisfactor production by murine macrophages. J. Natl. Cancer Inst.82 : 34-39.
Evans, C.H., Ghivizzani, S.C., Herndon, J.H., Wasko, M.C.,Reinecke, J., Wehling, P. and Robbins, P.D. (2000). Clinicaltrials in the gene therapy of arthritis. Clin. Orthop. 379 Suppl:S300-S307.
Evans, C.H. (2001). Expert Opin. Biol. Ther. 1: 971-978.
Farhood, H., Serbina, N. and Huang, L. (1995). The role ofdioleoyl phosphatidylethanolamine in cationic liposomemediated gene transfer. Biochim. Biophys. Acta 1235: 289-95.
Fasbender, A., Zabner, J., Chillòn, M., Moninger, T.O., Puga, A.P.,Davidson, B.L. and Welsh, M.J. (1997). Complexes ofadenovirus with polycationic polymers and cationic lipidsincrease the efficiency of gene transfer in vitro and in vivo. J.Biol. Chem. 272: 6479-6489.
Felgner, J.H., Kumar, R., Sridhar, C.N., Wheeler, C.J., Tsai, Y.J.,Border, R., Ramsey, P., Martin, M. and Felgner, P.L. (1994).Enhanced gene delivery and mechanism studies with a novelseries of cationic lipid formulations. J. Biol. Chem. 269: 2550-2561.
Francis, M.A. and Rainbow, A.J. (2000). UV-enhancedexpression of a reporter gene is induced at lower UV fluences intranscription-coupled repair deficient compared to normalhuman fibroblasts, and is absent in SV40-transformedcounterparts. Photochem. Photobiol. 72 : 554-561.
Galanis, E., Vile, R. and Russell, S.J. (2001). Delivery systemsintended for in vivo gene therapy of cancer: targeting andreplication competent viral vectors. Crit. Rev. Oncol. Hematol.38 : 177-192.
Ghivizzani, S.C. (2001). Drug Discov. Today 6: 259-267.
Godbey, W.T., Wu, K.K. and Mikos, A.G. (1999).Poly(ethylenimine) and its role in gene delivery. J. Control. Rel.60 : 149-160.
Goff, B.A., Hermanto, U., Rumbaugh, J., Blake, J., Bamberg, M.and Hasan, T. (1994). Photoimmunotherapy and biodistributionwith an OC125-chlorin immunoconjugate in an in vivo murineovarian cancer model. Br. J. Cancer 70 : 474-480.
Goff, B.A., Blake, J., Bamberg, M.P. and Hasan, T. (1996).Treatment of ovarian cancer with photodynamic therapy andimmunoconjugates in a murine ovarian cancer model. Br. J.Cancer 74 : 1194-1198.
Gomer, C.J., Ryter, S.W., Ferrario, A., Rucker, N., Wong, S. andFisher, A.M. (1996). Photodynamic therapy-mediated oxidativestress can induce expression of heat shock proteins. Cancer Res.56 : 2355-2360.
Gonschior, P., Gerheuser, F., Fleuchaus, M., Huehns, T.Y., Goetz,A.E., Welsch, U., Sroka, R., Dellian, M., Lehr, H.A. and Hofling, B.(1996). Local photodynamic therapy reduces tissue hyperplasiain an experimental restenosis model. Photochem. Photobiol. 64 :758-763.
Greber, U.F., Willetts, M., Webster, P. and Helenius, A. (1993).Stepwise dismantling of adenovirus 2 during entry into cells.Cell 75 : 477-486.
Hansen, J., Qing, K., Kwon, H.J., Mah, C. and Srivastava, A.(2000). Impaired intracellular trafficking of adeno-associatedvirus type 2 vectors limits efficient transduction of murinefibroblasts. J. Virol. 74 : 992-996.
Harbottle, R.P., Cooper, R.G., Hart, S.L., Ladhoff, A., McKay, T.,Knight, A.M., Wagner, E., Miller, A.D. and Coutelle, C. (1998).An RGD-oligolysine peptide: a prototype construct for integrin-mediated gene delivery. Hum. Gene Ther. 9: 1037-1047.
Haupt, K., Roggendorf, M. and Mann, K. (2002). The potential ofDNA vaccination against tumor-associated antigens forantitumor therapy. Exp. Biol. Med. (Maywood) 227: 227-237.
Haviv, Y.S. and Curiel, D.T. (2001). Conditional gene targetingfor cancer gene therapy. Adv. Drug Deliv. Rev. 53 : 135-154.
Henderson, B. and Dougherty, T.J. (1992). How doesphotodynamic therapy work? Photochem. Photobiol. 55 : 145-157.
Hendrich, C. (1997). Knee Surg. Sports Traumatol. Arthrosc. 5:58-63.
Huang, H., Chen, S.H., Kosai, K., Finegold, M.J. and Woo, S.L.(1996). Gene therapy for hepatocellular carcinoma: long-termremission of primary and metastatic tumors in mice byinterleukin-2 gene therapy in vivo. Gene Ther. 3: 980-987.
Høgset, A., Prasmickaite, L., Tjelle, T.E. and Berg, K. (2000).Photochemical transfection: A new technology for light-induced, site-directed gene delivery. Hum. Gene Ther. 11 : 869-880.
Høgset, A., Engesæter, B.Ø., Prasmickaite, L., Berg, K., Fodstad,Ø. and Mælandsmo, G.M. (2002a). Light-induced adenovirusgene transfer, an efficient and specific gene delivery technologyfor cancer gene therapy. Cancer Gene Ther. 9; 365-371.
Høgset, A., Prasmickaite, L., Hellum, M., Engesæter, B.Ø., Olsen,V.M., Tjelle, T.E., Wheeler, C.J. and Berg, K. (2002b).Photochemical transfection; a technology for efficient lightdirected gene delivery. In Synthetic DNA Delivery Systems,W.M.Saltzman and D.Luo, eds. (Austin: Landes BiosciencePublishers). In Press.
Isner, J.M. (2002). Myocardial gene therapy. Nature 10 : 234.
Jenkins, M.P., Buonaccorsi, G., MacRobert, A., Bishop, C.C.,Bown, S.G. and McEwan, J.R. (1998). Intra-arterialphotodynamic therapy using 5-ALA in a swine model. Eur. J.Vasc. Endovasc. Surg. 16 : 284-291.
Jori, G. and Spikes, J.D. (1984). Photobiochemistry ofporphyrins. In Topics in photomedicine, K.C. Smith, ed. (NewYork: Plenum Press), pp. 183-318.
Kay, M.A., Glorioso, J.C. and Namavari, M. (2001). Viral vectorsfor gene therapy: the art of turning infectious agents intovehiclles of therapeutics. Nature Med. 7: 33-40.

20 Current Gene Therapy, 2003, Vol. 3, No. 2 Høgset et al.
Khavari, P.A., Rollman, O. and Vahlquist, A. (2002). Cutaneousgene transfer for skin and systemic diseases. J. Intern. Med. 252:1-10.
Kick, G., Messer, G., Goetz, A., Plewig, G. and Kind, P. (1995).Photodynamic therapy induces expression of interleukin 6 byactivation of AP-1 but not NF-kappa B DNA binding. CancerRes. 55 : 2373-2379.
Kichler, A., Leborgne, C., Coeytaux, E. and Danos, O. (2001).Polyethylenimine-mediated gene delivery: a mechanistic study.J. Gene Med. 3: 135-144.
Klugherz, B.D. Song, C., DeFelice, S., Cui, X., Lu, Z., Connolly, J.,Hinson, J.T., Wilensky, R.L. and Levy, R.J. (2002). Gene deliveryto pig coronary arteries from stents carrying antibody-tetheredadenovirus. Hum. Gene Ther. 13 : 443.
Kochevar, I.E. and Redmond, R.W. (2000). Photosensitizedproduction of singlet oxygen. Meth. Enzymol. 319: 20-28.
Kozarsky, K.F. (2001). Gene therapy for cardiovascular disease.Curr. Opin. Pharmacol. 1: 197-202.
Lambert, J.M., Senter, P.D., Yau-Young, A., Blattler, W.A. andGoldmacher, V.S. (1985). Purified immunotoxins that arereactive with human lymphoid cells. Monoclonal antibodiesconjugated to the ribosome-inactivating proteins gelonin andthe pokeweed antiviral proteins. J. Biol. Chem. 260: 12035-12041.
LaMuraglia, G.M., Schiereck, J., Heckenkamp, J., Nigri, G.,Waterman, P., Leszczynski, D. and Kossodo, S. (2000).Photodynamic therapy induces apoptosis in intimalhyperplastic arteries. Am. J. Pathol. 157: 867-875.
Leitner, W.W., Hammerl, P. and Thalhammer, J. (2001). Nucleicacid for the treatment of cancer: genetic vaccines and DNAadjuvants. Curr. Pharm. Des. 7: 1641-1667.
Leopold, P.L., Ferris, B., Grinberg, I., Worgall, S., Hackett, N.R.and Crystal, R.G. (1998). Fluorescent virions: dynamic trackingof the pathway of adenoviral gene transfer vectors in livingcells. Hum. Gene Ther. 9: 367-378.
Leopold, P.L., Kreitzer, G., Miyazawa, N., Rempel, S., Pfister, K.K.,Rodriguez-Boulan, E. and Crystal, R.G. (2000). Dynein- andmicrotubule-mediated translocation of adenovirus serotype 5occurs after endosomal lysis. Hum. Gene Ther. 11 : 151-165.
Li, Y.M., Pong, R.C., Bergelson, J.M., Hall, M.C., Sagalowsky,A.I., Tseng, C.P., Wang, Z. and Hsieh, J.T. (1999). Loss ofadenoviral receptor expression in human bladder cancer cells: Apotential impact on the efficacy of gene therapy. Cancer Res. 59 :325-330.
Li, E., Brown, S.L., Stupack, D.G., Puente, X.S., Cheresh, D.A. andNemerow, G.R. (2001). Integrin alpha(v)beta1 is an adenoviruscoreceptor. J. Virol. 75 : 5405-5409.
Lin, M.T., Pulkkinen, L., Uitto, J. and Yoon, K. (2000). The genegun: current applications in cutaneous gene therapy. Int. J.Dermatol. 39 : 161-170.
Liu, F. and Huang, L. (2002). Development of non-viral vectorsfor systemic gene delivery. J. Control.Rel. 78 : 259-266.
Luna, M.C., Wong, S. and Gomer, C.J. (1994). Photodynamictherapy mediated induction of early response genes. Cancer Res.54 : 1374-1380.
Luna, M.C., Ferrario, A., Wong, S., Fisher, A.M.R. and Gomer, C.J.(2000). Photodynamic therapy-mediated oxidative stress as amolecular switch for the temporal expression of genes ligated tothe human heat shock promoter. Cancer Res. 60 : 1637-1644.
Luo, D. and Saltzman, W.M. (2000). Synthetic DNA deliverysystems. Nature Biotechnol. 18 : 33-37.
Maman, N., Dhami, S., Phillips, D. and Brault, D. (1999). Kineticand equilibrium studies of incorporation of di- sulfonatedaluminum phthalocyanine into unilamellar vesicles. Biochim.Biophys. Acta 1420: 168-178.
Maron, D.J., Choi, E.A. and Spitz, F.R. (2001). Gene therapy ofmetastatic disease: progress and prospects. Surg. Onco. Clin. N.Am. 10 : 449-460.
Matthews, J.L., Newman, J.T., Sogandares-Bernal, F., Judy, M.M.,Skiles, H., Leveson, J.E., Marengo-Rowe, A.J. and Chanh, T.C.(1988). Photodynamic therapy of viral contaminants withpotential for bloo banking applications. Transfusion 28 : 81-83.
McDonald, D., Stockwin, L., Matzow, T., Blair Zajdel, M.E. andBlair, G.E. (2001). Coxackie and adenovirus receptor (CAR)-dependent and major histocompatibility complex (MHC) classI-independent uptake of recombinant adenoviruses into humantumour cells. Gene Ther. 6: 1512-1519.
McKay, M.J.and Gaballa, M.A. (2001). Gene transfer therapy invascular diseases. Cardiovasc. Drug Rev. 19 : 245-262.
McKenzie, B.S., Corbett, A.J., Brady, J.L., Dyer, C.M., Strugnell,R.A., Kent, S.J., Kramer, D.R., Boyle, J.S. and Lew, A.M. (2001).Nucleic acid vaccines: tasks and tactics. Immunol. Res. 24 : 225-244.
Meyerson, S.L. and Schwartz, L.B. (1999). Gene therapy as atherapeutic intervention for vascular disease. J. Cardiovasc.Nurs. 13 : 91.
Miagkov, A.V., Varley, A.W., Munford, R.S. and Makarov, S.S.(2002). Endogenous regulation of a therapeutic transgenerestores homeostasis in arthritic joints. J. Clin. Invest. 109:1223-1229.
Michel, M.L. and Loirat, D. (2001). DNA vaccines forprophylactic or therapeutic immunization against hepatitis B.Intervirology 44 : 78-87.
Midoux, P., Mendes, C., Legrand, A., Raimond, J., Mayer, R.,Monsigny, M. and Roche, A.C. (1993). Specific gene transfermediated by lactosylated poly-L-lysine into hepatoma cells.Nucleic Acids Res. 21 : 871-878.
Miller, D.K., Griffiths, E., Lenard, J. and Firestone, R.A. (1983).Cell killing by lysosomotropic detergents. J. Cell Biol. 97 :1841-1851.
Miller, N. and Whelan, J. (1997). Progress in transcriptionallytargeted and regulatable vectors for genetic therapy. [Review][108 refs]. Hum. Gene Ther. 8: 803-815.
Mir, L.M. (2001). Therapeutic perspectives of in vivo cellelectropermeabilization. Bioelectrochemistry 53 : 1-10.
Miyazawa, N., Leopold, P.L., Hackett, N.R., Ferris, B., Worgall, S.,Falck-Pedersen, E. and Crystal, R.G. (1999). Fiber swap betweenadenovirus subgroups B and C alters intracellular trafficking ofadenovirus gene transfer vectors. J. Virol. 73 : 6056-6065.

Light Directed Gene Transfer Current Gene Therapy, 2003, Vol. 3, No. 2 21
Moan, J. and Sommer, S. (1985). Oxygen dependence of thephotosensitizing effect of hematoporphyrin derivative in NHIK3025 cells. Cancer Res. 45 : 1608-1610.
Moan, J., Berg, K., Kvam, E., Western, A., Malik, Z., Ruck, A. andSchneckenburger, H. (1989). Intracellular localization ofphotosensitizers. CIBA Foundation Symp. 146: 95-107.
Moan, J., Berg, K. and Kvam, E. (1990). Effects of photodynamictreatment on DNA and RNA-related cell functions. InPhototdynamic Therapy of Neoplastic Disease, D. Kessel, ed.(Boca Raton: CRC Press), pp. 197-209.
Moan, J. and Berg, K. (1991). The photodegradation ofporphyrins in cells can be used to estimate the lifetime ofsinglet oxygen. Photochem. Photobiol. 53 : 549-553.
Moan, J. and Berg, K. (1992). Photochemotherapy of cancer:experimental research. Photochem. Photobiol. 55 : 931-948.
Moan, J., Berg, K., Bommer, J.C. and Western, A. (1992). Actionspectra of phthalocyanines with respect to photosensitization ofcells. Photochem. Photobiol. 56 : 171-5.
Moan, J., Berg, K., Anholt, H. and Madslien, K. (1994).Sulfonated aluminium phthalocyanines as sensitizers forphotochemotherapy. Effects of small light doses onlocalization, dye fluorescence and photosensitivity in V79cells. Int. J. Cancer 58 : 865-870.
Mor, G. and Eliza, M. (2001). Plasmid DNA vaccines.Immunology, tolerance, and autoimmunity. Mol. Biotechnol. 19 :245-250.
Morishita, R., Aoki, M., Kaneda, Y. and Ogihara, T. (2001). Genetherapy in vascular medicine: recent advances and futureperspectives. Pharmacol. Ther. 91 : 105-114.
Muramatsu, T., Nakamura, A. and Park, H.M. (1998). In vivoelectroporation: a powerful and convenient means of nonviralgene transfer to tissues of living animals. Int. J. Mol. Med. 1: 55-62.
Nakamura, T., Morishita, R., Asai, T., Tsuboniwa, N., Aoki,M.,Sakonjo, H.,Yamasaki, K., Hashiya, N., Kaneda, Y. andOgihara, T. (2002). Molecular strategy using cis-element 'decoy'of E2F binding site inhibits neointimal formation in porcineballoon-injured coronary artery model.Gene Ther. 9: 488-494.
Neave, V., Gianotta, S., Hyman, S. and Schneider, J. (1988).Hematoporphyrin uptake in atherosclerotic plaques: therapeuticpotentials. Neurosurgery 23 : 307-312.
Nemerow, G.R. and Stewart, P.L. (1999). Role of alpha(v)integrins in adenovirus cell entry and gene delivery. Microbiol.Molec. Biol. Rev. 63 : 725-734.
Nettelbeck, D.M., Jérôme, V. and Müller, R. (2000). Gene therapy- designer promoters for tumour targeting. Trends Genet. 16 :174-181.
Newman, C.M., Lawrie, A., Brisken, A.F. and Cumberland, D.C.(2001). Ultrasound gene therapy: on the road from concept toreality. Echocardiography 18 : 339-347.
Nikol, S. and Huehns, T.Y. (2001). Preclinical and clinicalexperience in vascular gene therapy: advantages overconservative/standard therapy. J. Invasive Cardiol. 13 : 333-338.
Nishikawa, M. and Huang, L. (2001). Nonviral vectors in the newmillennium: delivery barriers in gene transfer. Hum. Gene Ther.12 : 861-870.
Normand, N., van Leeuwen, H. and O'Hare, P. (2001). Particleformation by a conserved domain of the herpes simplex virusprotein VP22 facilitating protein and nucleic acid delivery. J.Biol. Chem. 276: 15042-15050.
Ogris, M., Steinlein, P., Kursa, M., Mechtler, K., Kircheis, R. andWagner, E. (1998). The size of DNA/transferrin-PEI complexes isan important factor for gene expression in cultured cells. GeneTher. 5: 1425-1433.
Ortu, P., LaMuraglia, G.M., Roberts, W.G., Flotte, T.J. and Hasan,T. (1992). Photodynamic therapy of arteries. A novel approachfor treatment of experimental intimal hyperplasia. Circulation85 : 1189-1196.
Oshikawa, K., Rakhmilevich, A.L., Shi, F., Sondel, P.M., Yang, N.and Mahvi, D.M. (2001). Interleukin 12 gene transfer into skindistant from the tumor site elicits antimetastatic effectsequivalent to local gene transfer. Hum. Gene Ther. 12 : 149-160.
Pandori, M.W. and Sano,T. (2000). Photoacivatable retroviralvectors: a strategy for targeted gene delivery. Gene Ther. 7:1999-2006.
Pandori, M.W., Hobson, D.A., Olejnik, J., Krzymanska-Olejnik,E., Rothschild, K.J., Palmer, A.A., Phillips, T.J. and Sano, T.(2002). Photochemical control of the infectivity of adenoviralvectors using a novel photocleavable biotinylation reagent.Chem. Biol. 9: 567-573.
Pass, H.I. (1993). Photodynamic therapy in oncology:Mechanisms and clinical use. J. Natl. Cancer Inst. 85 : 443-456.
Peng, Q., Nesland, J.M., Madslien, K., Danielsen, H.E. and Moan,J. (1996). Subcellular photodynamic action sites of sulfonatedaluminium phthalocyanines in a human melanoma cell line.SPIE 2628: 102-113.
Pfeifer, A. and Verma, I.M. (2001). Gene therapy: promises andproblems. Ann. Rev. Genomics Hum. Genet. 2: 177-211.
Pickles, R.J., McCarty, D., Matsui, H., Hart, P.J., Randell, S.H. andBoucher, R.C. (1998). Limited entry of adenovirus vectors intowell-differentiated airway epithelium is responsible forinefficient gene transfer. J. Virol. 72 : 6014-6023.
Porter, T.R. and Xie, F. (2001). Therapeutic ultrasound for genedelivery. Echocardiography 18 : 349-353.
Pouton, C.W. (1998). Nuclear import of polypeptides,polynucleotides and supramolecular complexes. Adv. DrugDeliv. Rev. 34 : 51-64.
Pouton, C.W., Lucas, P., Thomas, B.J., Uduehi, A.N., Milroy, D.A.and Moss, S.H. (1998). Polycation-DNA complexes for genedelivery: a comparison of the biopharmaceutical properties ofcationic polypeptides and cationic lipids. J. Control Rel. 53 :289-299.
Pouton, C.W. and Seymour, L.W. (2001). Key issues in non-viralgene delivery. Adv. Drug Deliv. Rev. 46 : 187-203.
Prasmickaite, L., Høgset, A., Tjelle, T.E., Olsen, V.M. and Berg, K.(2000). The role of endosomes in gene transfection mediated byphotochemical internalisation. J. Gene Med. 2: 477-488.

22 Current Gene Therapy, 2003, Vol. 3, No. 2 Høgset et al.
Prasmickaite, L., Høgset, A. and Berg, K. Evaluation of differentphotosensitizers for use in photochemical gene transfection(2001a). Photochem. Photobiol. 73 : 388-395.
Prasmickaite, L., Høgset, A. and Berg, K. (2001b). Methods forphotochemical transfection: Light-induced, site-directed genedelivery. In: Methods in Molecular Medicine, Vol. 69 : GeneTherapy Protocols, 2
nd ed.Ed.Morgan, J.R. Humana Press
Inc.,Totowa. pp. 123-135.
Prasmickaite, L., Høgset, A., Selbo, P.K., Engesæter, B.Ø., Hellum,M. and Berg, K. (2002). Photochemical disruption of endocyticvesicles before delivery of drugs: a new strategy for cancertherapy. Br. J. Cancer 86 : 652-657.
Quarck, R. and Holvoet, P. (2001). Restenosis and gene therapy.Expert Opin. Biol. Ther. 1: 79.
Rakhmilevich, A.L., Turner, J., Ford, M.J., McCabe, D., Sun, W.H.,Sondel, P.M., Grota, K. and Yang, N.S. (1996). Gene gun-mediated skin transfection with interleukin 12 gene results inregression of established primary and metastatic murine tumors.Proc. Nat.l Acad. Sci. USA 93 : 6291-6296.
Rao, K.V., He, Y.X. and Ramaswamy, K. (2002). Suppression ofcutaneous inflammation by intradermal gene delivery.GeneTher. 9: 38-45.
Ratkay, L.G., Chowdhary, R.K., Neyndorff, H.C., Tonzetich, J.,Waterfield, J.D. and Levy, J.G. (1994). Photodynamic therapy: acomparison with other immunomodulatory treatments ofadjuvant-enhanced arthritis in MRL-lpr mice. Clin. Exp.Immunol. 95 : 373-377.
Ratkay, L.G., Chowdhary, R.K., Iamaroon, A., Richter, A.M.,Neyndorff, H.C., Keystone, E.C., Waterfield, J.D. and Levy, J.G.(1998). Amelioration of antigen-induced arthritis in rabbits byinduction of apoptosis of inflammatory cells with localapplication of transdermal photodynamic therapy. ArthritisRheum. 41 : 525-534.
Regillo, C.D. (2000). Update on photodynamic therapy. Curr.Opin. Ophthalmol. 11 : 166-170.
Robinson, D.J., Collins, P., Stringer, M.R., Vernon, D.I., Stables,G.I., Brown, S.B. and Sheehan-Dare, R.A. (1999). Improvedresponse of plaque psoriasis after multiple treatments withtopical 5-aminolaevulinic acid photodynamic therapy. ActaDerm. Venereol. 79 : 451-455.
Rochlitz, C., Jantscheff, P., Bongartz, G., Dietrich, P.Y.,Quiquerez, A.L., Schatz, C., Mehtali, M., Courtney, M., Tartour, E.,Dorval, T., Fridman, W.H. and Herrmann, R. (1999). Gene therapystudy of cytokine-transfected xenogeneic cells (Vero-interleukin-2) in patients with metastatic solid tumors. CancerGene Ther. 6: 271-281.
Rockson, S.G., Kramer, P., Razavi, M., Szuba, A., Filardo, S.,Fitzgerald, P., Cooke, J.P., Yousuf, S., DeVault, A.R., Renschler,M.F. and Adelman, D.C. (2000a). Photoangioplasty for humanperipheral atherosclerosis. Results of a phase I trial ofphotodynamic therapy with motexafin lutetium (Antrin).Circulation 102: 2322-2324.
Rockson, S.G., Lorenz, D.P., Cheong, W.-F. and Woodburn, K.W.(2000b). An emerging clinical cardiovascular role forphotodynamic therapy. Circulation 102: 591-596.
Rosenkranz, A.A., Jans, D.A. and Sobolev, A.S. (2000). Targetedintracellular delivery of photosensitizers to enhancephotodynamic efficiency. Immunol. Cell Biol. 78 : 452-464.
Ryter, S.W. and Gomer, C.J. (1993). Nuclear factor kappa Bbinding activity in mouse L1210 cells following photofrin II-mediated photosensitization. Photochem. Photobiol. 58 : 753-756.
Schagen, F.H., Moor, A.C., Cheong, S.C., Cramer, S.J., VanOrmondt, H., Van der Eb, A.J., Dubbelman, T.M. and Hoeben, R.C.(1999). Photodynamic treatment of adenoviral vectors withvisible light: an easy and convenient method for viralinactivation. Gene Ther. 6: 873-881.
Scherer, F., Anton, M., Schillinger, U., Henke, J., Bergemann, C.,Kruger, A., Gansbacher, B. and Plank, C. (2002). Magnetofection:enhancing and targeting gene delivery by magnetic force invitro and in vivo. Gene Ther. 9: 102-109.
Schmidt-Erfurth, U. and Hasan, T. (2000). Mechanism of actionof photodynamic therapy with Verteporfin for the treatment ofage-related macular degeneration. Surv. Ophthalmol. 45 : 195-214.
Scott, C.F.Jr., Lambert, J.M., Goldmacher, V.S., Blattler, W.A.,Sobel, R., Schlossman, S.F. and Benacerraf, B. (1987). Thepharmacokinetics and toxicity of murine monoclonalantibodies and gelonin conjugates of these antibodies. Int. J.Immunopharmacol. 9: 211-225.
Selbo, P.K., Sandvig, K., Kirveliene, V. and Berg, K. (2000a).Release of gelonin from endosomes and lysosomes to cytosolby photochemical internalisation. Biochim. Biophys. Acta 1475:307-313.
Selbo, P.K., Sivam, G., Fodstad, Ø., Sandvig, K. and Berg, K.(2000b). Photochemical internalisation increases the cytotoxiceffect of the immunotoxin MOC31-gelonin. Int. J. Cancer 87 :853-859.
Selbo, P.K., Sivam, G., Fodstad, Ø., Sandvig, K. and Berg, K.(2001). In vivo documentation of photochemical internalisation,a novel approach to site specific cancer therapy. Int. J. Cancer.92 : 761-766.
Sharma, A.K. and Khuller, G.K. (2001). DNA vaccines: futurestrategies and relevance to intracellular pathogens. Immunol.Cell Biol. 79 : 537-546.
Simkin, G.O., Tao, J.S., Levy, J.G. and Hunt, D.W.C. (2000). IL-10contributes to the inhibition of contact hypersensitivity in micetreated with photodynamic therapy. J. Immunol. 164: 2457-2462.
Soukos, N.S., Hamblin, M.R., Keel, S., Fabian, R.L., Deutsch, T.F.and Hasan, T. (2001). Epidermal growth factor receptor-targetedimmunophotodiagnosis and photoimmunotherapy of oralprecancer in vivo. Cancer Res. 61 : 4490-4496.
Spirito, F., Meneguzzi, G., Danos, O. and Mezzina, M. (2001).Cutaneous gene transfer and therapy: the present and the future.J. Gene Med. 3: 21-31.
Stirpe, F., Olsnes, S. and Pihl, A. (1980). Gelonin, a new inhibitorof protein synthesis, nontoxic to intact cells. Isolation,characterization, and preparation of cytotoxic complexes withcancanavalin A. J. Biol. Chem. 255: 6947-6953.
Stopeck, A.T., Jones, A., Hersh, E.M., Thompson, J.A., Finucane,D.M., Gutheil, J.C. and Gonzalez, R. (2001). Phase II study ofdirect intralesional gene transfer of allovectin-7, an HLA-B7/beta2-microglobulin DNA-liposome complex, in patientswith metastatic melanoma. Clin. Cancer Res. 7: 2285-2291.

Light Directed Gene Transfer Current Gene Therapy, 2003, Vol. 3, No. 2 23
Sudimack, J. and Lee, R.J. (2000). Targeted drug delivery via thefolate receptor. Adv. Drug Deliv. Rev. 41 : 147-162.
Suomalainen, M., Nakano, M.Y., Keller, S., Boucke, K., Stidwill,R.P. and Greber, U.F. (1999). Microtubule-dependent plus- andminus end-directed motilities ar competing processes fornuclear targeting of adenovirus. J. Cell Biol. 144: 657-672.
Tang, D.-C., Jennelle, R.S., Shi, Z., Garver, R.I., Jr., Carbone, D.P.,Loya, F., Chang, C.-H. and Curiel, D.T. (1997). Overexpression ofadenovirus-encoded transgenes from the cytomegalovirusimmediate early promoter in irradiated tumor cells. Hum. GeneTher. 8: 2117-2124.
Tarner, I.H. and Fathman, C.G. (2001). Gene therapy inautoimmune disease. Curr. Opin. Immunol. 13 : 676-682.
Tio, R.A., Isner, J.M. and Walsh, K. (1998). Gene therapy toprevent restenosis, the Boston experience. Sem. InterventionalCardiol. 3: 205-210.
Todryk, S., Melcher, A.A., Hardwick, N., Linardakis, E., Bateman,A., Colombo, M.P., Stoppacciaro, A. and Vile, R.G. (1999). Heatshock protein 70 induced during tumor cell killing induces Th1cytokines and targets immature dendritic cell precursors toenhance antigen uptake. J. Immunol. 163: 1398-1408.
Todryk, S.M., Melcher, A.A., Dalgleish, A.G. and Vile, R. (2000).Heat shock proteins refine the danger theory. Immunology 99 :334-337.
Trauner, K.B. and Hasan, T. (1996). Photodynamic treatment ofrheumatoid and inflammatory arthritis. Photochem. Photobiol.64 : 740-750.
Trauner, K., Gandour-Edwards, R., Bamberg, M., Nishioka, N.S.,Flotte, T., Autry, S. and Hasan, T. (1998). Influence of lightdelivery on photodynamic synovectomy in an antigen-inducedarthritis model for rheumatoid arthritis. Lasers in Surgery &Medicine 22 : 147-156.
Tseng, W.C., Haselton, F.R. and Giorgio, T.D. (1997).Transfection by cationic liposomes using simultaneous singlecell measurements of plasmid delivery and transgeneexpression. J. Biol. Chem. 272: 25641-25647.
Ulmer, J.B. (2001a). An update on the state of the art of DNAvaccines. Curr. Opin. Drug Discov. Devel. 4: 192-197.
Ulmer, J.B. (2001b). Tuberculosis DNA vaccines. Scand. J.Infect. Dis. 33 : 246-248.
Unger, E.C., Hersh, E., Vannan, M. and McCreery, T. (2001). Genedelivery using ultrasound contrast agents. Echocardiography18 : 355-361.
van de Loo, F.A. and van den Berg, W.B., (2002). Gene therapyfor rheumatoid arthritis. Lessons from animal models, includingstudies on interleukin-4, interleukin-10, and interleukin-1receptor antagonist as potential disease modulators. Rheum. Dis.Clin. North Am. 28 : 127-149.
van Deurs, B., Holm, P.K. and Sandvig, K. (1996). Inhibition ofthe vacuolar H(+)-ATPase with Bafilomycin reduces delivery ofinternalized molecules from mature multivesicular endosomesto lysosomes in HEp-2 cells. Eur. J. Cell Biol. 69 : 343-350.
van Loo, N.D., Fortunati, E., Ehlert, E., Rabelink, M., Grosveld, F.and Scholte, B.J. (2001). Baculovirus infection of nondividingmammaliam cells: mechanims of entry and nuclear transport ofcapsids. J. Virol. 75 : 961-970.
van Moor, A.C., Dubbelman, T.M., Van Steveninck, J. and Brand,A. (1999). Photodynamic sterilization of red cells and its effecton contaminating white cells: viability and mechanism of celldeath. Transfusion 39 : 599-607.
Verma, I.M. and Somia, N. (1997). Gene therapy - promises,problems and prospects. Nature 389: 239-242.
Von Seggern, D.J., Chiu, C.Y., Fleck, S.K., Stewart, P.L. andNemerow, G.R. (1999). A helper-independent adenovirus vectorwith E1, E3, and fiber deleted: structure and infectivity offiberless particles. J. Virol. 73 : 1601-1608.
Vorburger, S.A. and Hunt, K.K. (2002). Adenoviral gene therapy.Oncologist 7: 46-57.
Wagner, E., Zatloukal, K., Cotten, M., Kirlappos, H., Mechtler, K.,Curiel, D.T. and Birnstiel, M.L. (1992). Coupling of adenoviruseto transferrin-polylysine/DNA complexes greatly enhancesreceptor-mediated gene delivery and expression of transfectedgenes. Proc. Natl. Acad. Sci. USA 89 : 6099-6103.
Walter, E., Croyle, M.A., Roessler, B.J. and Amidon, G.L. (1997).The absence of accessible vitronectin receptors in differentiatedtissue hinders adenoviral-mediated gene transfer to theintestinal epithelium in vitro. Pharm. Res. 14 : 1216-1222.
Walther, W. and Stein,U. (1999). Therapeutic genes for cancertherapy. Mol. Biotechnol. 13 : 21-28.
Wang, K., Guan, T., Cheresh, D.A. and Nemerow, G.R. (2000).Regulation of adenovirus membrane penetration by thecytoplasmic tail of integrin beta5. J. Virol. 74 : 2731-2739.
Warren, P., Song, W., Holle, E., Holmes, L., Wei, Y., Li, J., Wagner,T. and Yu, X. (2002). Combined HSV-TK/GCV and secondarylymphoid tissue chemokine gene therapy inhibits tumor growthand elicits potent antitumor CTL response in tumor-bearingmice. Anticancer Res. 22 : 599-604.
Weishaupt, K.R., Gomer, C.J. and Dougherty, T.J. (1976).Identification of singlet oxygen as the cytotoxic agent inphotoinactivation of a murine tumor. Cancer Res. 36 : 2326-2329.
Wickham, T.J., Filardo, E.J., Cheresh, D.A. and Nemerow, G.R.(1994). Integrin alfav-beta5 selectively promotes adenovirusmediated cell membrane permeabilization. J. Cell Biol. 127:257-264.
Wickham, T.J. (2000). Targeting adenovirus. Gene Ther. 7: 110-114.
Wilke, M., Fortunati, E., van den, B.M., Hoogeveen, A.T. andScholte, B.J. (1996). Efficacy of a peptide-based gene deliverysystem depends on mitotic activity. Gene Ther. 3: 1133-1142.
Wilson, P.D., Firestone, R.A. and Lenard, J. (1987). The role oflysosomal enzymes in killing of mammalian cells by thelysosomotropic detergent N-dodecylimidazole. J. Cell Biol.104: 1223-1229.
Wolfert, M.A. and Seymour, L.W. (1998). Chloroquine andamphipathic peptide helices show synergistic transfection invitro. Gene Ther. 5: 409-414.
Wrobel, I. and Collins, D. (1995). Fusion of cationic liposomeswith mammalian cells occurs after endocytosis. Biochim.Biophys. Acta 1235: 296-304.
Wu, C.Y., Maeda, H., Contursi, C., Ozato, K. and Seder, R.A.(1999). Differential requirement of IFN consensus sequence

24 Current Gene Therapy, 2003, Vol. 3, No. 2 Høgset et al.
binding protein for the production of IL-12 and induction ofTh1-type cells in response to IFN-gamma. J. Immunol. 162: 807-812.
Xu, W. and Ellington, A.D. (1996). Anti-peptide aptamersrecognize amino acid sequence and bind a protein epitope. Proc.Natl. Acad. Sci. USA 93 : 7475-80.
Yamaguchi, A., Woodburn, K.W., Hayase, M. and Robbins, R.C.(2000). Reduction of vein graft disease using photodynamictherapy with motexafin lutetium in a rodent isograft model.Circulation 102 (suppl III): III-275-III-280.
Zabner, J., Fasbender, A.J., Moninger, T., Poellinger, K.A. andWelsh, M.J. (1995). Cellular and molecular barriers to genetransfer by a cationic lipid. J. Biol. Chem. 270: 18997-19007.
Zauner, W., Kichler, A., Schmidt, W., Sinski, A. and Wagner, E.(1996). Glycerol enhancement of ligand-polylysine/DNAtransfection. BioTechniques 20 : 905-913.
Zauner, W., Kichler, A., Schmidt, W., Mechtler, K. and Wagner, E.(1997). Glycerol and polylysine synergize in their ability torupture vesicular membranes: a mechanism for increasedtransferrin- polylysine-mediated gene transfer. Exp. Cell Res.232: 137-145.
Zauner, W., Ogris, M. and Wagner, E. (1998). Polylysine-basedtransfection systems utilizing receptor- mediated delivery. Adv.Drug Deliv. Rev. 30 : 97-113.
Zelphati, O. and Szoka, F.C.J. (1996). Mechanism ofoligonucleotide release from cationic liposomes. Proc. Natl.Acad. Sci. USA 93 : 11493-11498.




















