
Allergen-induced IL-9 directly stimulates mucin transcription in respiratory epithelial cells

1
Leucine Stimulates Protein Synthesis in Skeletal Muscle of Neonatal Pigs by Enhancing
mTORC1 Activation
Agus Suryawan, Asumthia S. Jeyapalan, Renan A. Orellana, Fiona A. Wilson, Hanh V. Nguyen,
and Teresa A. Davis
United States Department Agriculture/Agriculture Research Service, Children’s Nutrition
Research Center, Department of Pediatrics, Baylor College of Medicine, Houston, Texas 77030
Running Title: Leucine-Induced mTOR Signaling in Neonatal Muscle
Correspondence: Teresa A. Davis, Ph.D., USDA/ARS Children’s Nutrition Research Center,
Baylor College of Medicine, 1100 Bates St., Houston, TX 77030 (phone: 713-798-7169, fax:
713-798-7171, email: [email protected]).
Articles in PresS. Am J Physiol Endocrinol Metab (August 5, 2008). doi:10.1152/ajpendo.90314.2008
Copyright © 2008 by the American Physiological Society.

2
ABSTRACT
Skeletal muscle in the neonate grows at a rapid rate due in part to an enhanced sensitivity to the
postprandial rise in amino acids, particularly leucine. To elucidate the molecular mechanism by
which leucine stimulates protein synthesis in neonatal muscle, overnight fasted 7-day-old piglets
were treated with rapamycin (an inhibitor of mammalian target of rapamycin complex
1/mTORC1) for 1 h and then infused with leucine for 1 h. Fractional rates of protein synthesis
and activation of signaling components that lead to mRNA translation were determined in
skeletal muscle. Rapamycin completely blocked leucine-induced muscle protein synthesis.
Rapamycin markedly reduced raptor-mTOR association, an indicator of mTORC1 activation.
Rapamycin blocked the leucine-induced phosphorylation of mTOR, S6K1, and 4EBP1 and
formation of the eIF4E•eIF4G complex and increased eIF4E•4EBP1 complex abundance.
Rapamycin had no effect on the association of mTOR with rictor, a crucial component for
mTORC2 activation, or GβL, a component of mTORC1 and mTORC2 complexes. Neither
leucine nor rapamycin affected the phosphorylation of AMPK, PKB, or TCS2, signaling
components that reside upstream of mTOR. eEF2 phosphorylation was not affected by leucine
or rapamycin, although current dogma indicates that eEF2 phosphorylation is mTOR-dependent.
Taken together, these in vivo data suggest that leucine stimulates muscle protein synthesis in
neonates by enhancing mTORC1 activation and its downstream effectors.
Keywords: mRNA translation; eukaryotic initiation factor 4G; AMP-activated protein kinase;
raptor; rictor

3
INTRODUCTION
The neonatal period is characterized by rapid growth that is supported by a high rate of
protein synthesis (3). However, most infants that are born of low birth weight are discharged
weighing less than the tenth percentile for age despite improvements in their nutritional
management (9). Some remain small to adulthood and exhibit adverse long-term developmental
outcomes including reduced work capacity (27). To improve strategies for the nutritional
management of low birth weight infants, we have used the neonatal pig as a model of the human
neonate so as to identify the mechanisms that regulate protein deposition in neonates. Our
studies have shown that the profound accretion of skeletal muscle protein is in part due to the
ability of neonatal muscle to markedly increase protein synthesis in response to feeding, a
response that declines rapidly with development (5). In skeletal muscle, the feeding-induced
stimulation of protein synthesis is independently modulated by the rise in insulin and amino acids
(6; 24). Recently, we demonstrated that the postprandial rise in leucine alone, but not isoleucine
or valine alone, stimulates skeletal muscle protein synthesis in neonatal pigs and this response
decreases with age (10-12).
Signaling through the mammalian target of rapamycin (mTOR) plays a significant role in
cell growth regulation including protein synthesis (4). Recent studies have revealed that mTOR
exists in two protein complexes (Fig. 1): mTOR complex 1 (mTORC1) and mTOR complex 2
(mTORC2) (4). mTORC1 is rapamycin-sensitive and consists of mTOR, raptor, and G-protein β
subunit-like protein (GβL). This complex is activated by amino acids (especially leucine),
hormones/growth factors, and energy signals (17). mTORC1 regulates mRNA translation by
phosphorylating two of its effectors, ribosomal protein S6 kinase 1 (S6K1) and eIF4E-binding
protein-1 (4E-BP1) (4). Phosphorylated 4E-BP1 releases eIF4E from the inactive eIF4E•4EBP1

4
complex, allowing the formation of the active eIF4G•eIF4E complex (29). mTORC1 has also
been implicated in the regulation of elongation factor 2 (eEF2) activation (38). The second
protein complex, mTORC2, is rapamycin insensitive and consists of mTOR, rictor, and GβL (4).
Recent biochemical studies suggest that one major function of mTORC2 is to activate the growth
factor-regulated protein kinase B (PKB) (28).
The molecular mechanisms by which amino acids or leucine modulate the activation of
mTORC1 in vivo is unknown. Recent studies suggest that amino acids or leucine stimulate
mTORC1 independent of PKB, AMPK and TSC1/TSC2 activation (4). Furthermore, studies
conducted in cell culture suggest that the activation of mTORC1 is partly regulated by the
protein-protein interaction among members of mTORC1, in particular, the interaction of mTOR
with raptor (16). However, we (31) and others (37) have found that feeding stimulates the
activation of mTOR in vivo without affecting the interaction of mTOR with raptor.
Until recently (1; 11; 35), information regarding the molecular mechanism by which
leucine regulates the activation of mTOR leading to the stimulation of muscle protein synthesis
were obtained from cell culture studies. We demonstrated that a physiological rise in plasma
leucine stimulates muscle protein synthesis in neonatal pigs by enhancing translation initiation
factor activation (11). However, detailed study of the effect of physiological levels of leucine on
the activation of mTORC1 leading to the stimulation of protein synthesis in skeletal muscle of
the neonate has not been conducted previously. Therefore, our objective was to determine the
molecular mechanism by which leucine modulates mTORC1 activation in vivo using rapamycin,
a potent inhibitor of mTORC1. Previously we demonstrated that the feeding-induced stimulation
of muscle protein synthesis in neonatal pigs was only partially inhibited by rapamycin (21).

5
MATERIALS AND METHODS
Animals and housing. Two crossbred (Landrace x Yorkshire x Duroc x Hampshire)
pregnant sows (Agriculture Headquarters, Texas Department of Criminal
Justice, Huntsville, TX)
were housed in lactation crates in individual, environmentally controlled rooms 2 wk before
farrowing. Sows were fed a commercial diet (no. 5084; PMI Feeds, Richmond,
IN) and provided
water ad libitum. After farrowing, piglets remained with the sow but were not allowed access to
the sow's diet. A total of 23 piglets from three litters, weighing 2 kg, were studied at 7 days of
age, respectively. Three days prior to the experiment, piglets were anesthetized for sterile
catheter insertion into a jugular vein and carotid artery. Piglets were then returned to the sow
and allowed to suckle freely until studied. The protocol was approved by the Animal Care and
Use Committee of Baylor College of Medicine and was conducted in accordance with the
National Research Council's Guide for the Care and Use of Laboratory Animals.
Treatments and infusion. Piglets were fasted for 12–14 h before infusion and placed in a
sling restraint system. The carotid catheter was used to infuse saline, leucine, rapamycin, and L-
[4-3H]phenylalanine,
whereas the jugular catheter was used for repeated blood sample
collection.
Pigs were randomly assigned to one of four treatment groups: 1) saline (control), 2) saline +
rapamycin, 3) leucine, and 4) leucine + rapamycin. Piglets assigned to the rapamycin groups
were injected with a rapamycin solution (0.75 mg/kg in 5% dimethyl sulfoxide) 1 hour prior to
the initiation of the leucine infusion; other pigs were injected with diluent. Leucine infusion was
initiated with a primed dose (148 µmol/kg) for 10 min, followed by a constant infusion of
leucine at 400 µmol·kg–1
·h–1
for 1 hour. Previous studies (11) have shown that a 2-3 fold
elevation in plasma leucine concentration, similar to that observed with feeding, is achieved by

6
this rate of leucine infusion. During the priming and constant infusion period, saline-infused
pigs received an equal volume of saline as those receiving leucine.
Tissue protein synthesis in vivo. Fractional rates of protein synthesis were measured
using a modification of the flooding dose method (14). At 30 min before the end of the infusion,
pigs were injected with 10 ml/kg body weight of a flooding dose of phenylalanine (Amersham
Biosciences, Piscataway, NJ) which provided 1.5 mM phenylalanine/kg body weight and 0.5
mCi of L-[4-3H]phenylalanine/kg body weight. Samples of whole blood were taken 5, 15, and
30 min after the injection for measurement of the specific radioactivity of the extracellular free
pool of phenylalanine. Pigs were killed at 60 min, and longissimus dorsi muscle samples were
collected and immediately frozen in liquid nitrogen and stored at –70°C until analyzed, as
previously described (8).
Protein synthesis (Ks expressed as % protein synthesized in a day) was calculated
as: Ks
(%/day) = [(Sb/Sa) x (1,440/t)] x 100, where Sb is the specific radioactivity of the protein-bound
phenylalanine; Sa is the specific radioactivity of the tissue free phenylalanine
for the labeling
period, determined from the value of the animal at the time of tissue collection, corrected by the
linear regression of the blood specific radioactivity of the animal against time;
and t is the time of
labeling in minutes. Previous studies have demonstrated that after a flooding dose of L-[4-
3H]phenylalanine is administrated, the specific radioactivity of tissue free phenylalanine is in
equilibrium with the aminoacyl tRNA specific radioactivity, and therefore the tissue free
phenylalanine is a valid measure of the tissue precursor pool specific radioactivity (7).
Tissue extraction and immunoblot analysis. Freshly collected longissimus dorsi muscle
tissue samples were homogenized, centrifuged at 10,000 g for 10 min
at 4°C. Supernatants were
diluted in sample buffer, frozen in liquid nitrogen, and stored at -70oC until analysis. Equal

7
amounts of protein samples were electrophoretically separated in polyacrylamide gels and
transferred to a polyvinylidene difluoride (PVDF) membrane (Bio-Rad, Hercules, CA), which
was incubated with appropriate primary antibodies, washed, and exposed to an appropriate
secondary antibody as previously described (32).
For normalization, immunoblotting performed with antiphospho-specific antibodies were
stripped in stripping buffer (Pierce Biotechnology, Rockford, IL) and reprobed with the
corresponding nonphospho-specific antibodies. Blots were developed
using an enhanced
chemiluminescence kit (Amersham, Piscataway, NJ), visualized,
and analyzed using a
ChemiDoc-It Imaging System (UVP, Upland, CA). Primary antibodies that were used in the
immunoblotting were: PKB (total and Ser 473, Cell Signaling Technology, Inc., Beverly, MA),
AMPKα (total and Thr 172, Cell Signaling), TSC2 (total and Thr 1462, Cell Signaling), mTOR
(total, Ser 2448 and Ser 2481, Cell Signaling), S6K1 (total and Thr 398, Cell Signaling), 4E-BP1
(total, Bethyl Laboratories, Montgomery, TX, and Thr 70, Cell Signaling), and eEF2 (total and
Thr 56, Cell Signaling).
Quantification of eIF4E·4EBP1 and eIF4E·eIF4G complexes. These complexes were
immunoprecipitated using an anti-eIF4E monoclonal antibody (gift of Dr. Leonard Jefferson,
Penn State University College of Medicine, Hershey, PA) from aliquots of fresh tissue
homogenates. Briefly, samples were homogenized in seven volumes of buffer (in mM: 20
HEPES, 2 EGTA, 50 NaF, 100 KCl, and 0.2 EDTA, pH 7.4) containing Sigma P3840 Protease
Inhibitor Cocktail (Sigma Chemical, St. Louis, MO) and centrifuged at 10,000 g for 10
min at
4°C. Supernatants were incubated overnight at 4°C with constant rocking with anti-eIF4E
antibody. Immunoprecipitates
were recovered with goat anti-rabbit IgG magnetic beads
(Polysciences,
Warrington, PA), washed and resuspended in sample buffer as
described

8
elsewhere (8), and immediately subjected to protein immunoblot analysis using rabbit anti-4E-
BP1 (Cell Signaling) antibody or rabbit anti-eIF4G (Bethyl Laboratories). Amounts of 4E-BP1
and eIF4G were corrected by the eIF4E recovered from the immunoprecipitate.
Analysis of mTORC1 and mTORC2 complexes. To determine the association between
mTOR and its partners (with raptor and GβL for the mTORC1 complex, or with rictor and GβL
for the mTORC2 complex), muscle samples were homogenized in CHAPS buffer according to
Williamson et al (39). The CHAPS buffer consisted of 40 mM HEPES, pH 7.5, 120 mM NaCl, 1
mM EDTA, 10 mM pyrophosphate, 10 mM β-glycerolphosphate, 40 mM NaF, 1.5 mM sodium
vanadate, 0.3% CHAPS, 0.1 mM PMSF, 1 mM benzamidine, and 1 mM DTT. The homogenate
was mixed on a platform rocker for 30 min at 4°C and then centrifuged at 1,000 g for 3 min
(4°C). The supernatant containing 500 µg of protein was combined with 2 µl of anti-mTOR
antibody (Cell Signaling) and mixed on a platform rocker overnight at 4°C. Following the
incubation, the immune complexes were isolated with a goat anti-mouse
BioMag IgG
(PerSeptive Diagnostics, Cambridge, MA) bead slurry. The magnetic bead complexes were
collected using a magnetic stand, washed twice with CHAPS buffer, and once in CHAPS buffer
containing 200 mM instead of 120 mM NaCl and 60 mM instead of 40 mM HEPES.
The
precipitates were rinsed with 100 µl of 1X SDS sample buffer and then boiled for 5 min and
centrifuged to collect the supernatant. The samples were subjected to SDS-PAGE followed by
immunoblotting with anti-raptor antibody (Cell Signaling), anti-rictor antibody (Cell Signaling)
and anti-GβL antibody (Cell Signaling). The mTOR-protein complexes were normalized by the
amount of total mTOR in the precipitates.
Statistics. All data were analyzed using one-way analysis of variance (ANOVA) with
saline-treated animals as the independent variable. When a significant overall effect was

9
observed, differences among individual means were assessed by the Tukey-Kramer comparisons
test. Probability values of P < 0.05 were considered statistically significant. Data are presented
as means ± SE. Data are presented as means ± SE.
RESULTS
Fractional protein synthesis rates. Several laboratories have demonstrated that leucine
administration increases in vivo skeletal muscle protein synthesis (1; 2). Recently, we also
provided evidence that leucine administration at physiological levels stimulates protein synthesis
in skeletal muscle of neonatal pigs (11). In this study, we examined whether treatment with
rapamycin could suppress the leucine-induced stimulation of protein synthesis in skeletal muscle
of neonatal pigs. As shown in Fig. 2, leucine infusion, compared to saline infusion, increased the
fractional rates of protein synthesis in skeletal muscle (P < 0.05). Administration of rapamycin
completely blocked the stimulatory effect of leucine on skeletal muscle protein synthesis.
Because it is important to take into account any possible effect of rapamycin on the basal fasting
rate of protein synthesis, rapamycin was also administrated to saline-infused pigs. We found that
rapamycin administration to saline-infused pigs did not reduce the fractional rate of skeletal
muscle protein synthesis below the basal fasting level.
mTORC1 and mTORC2 activation. We previously showed that administration of leucine
to neonatal pigs increases skeletal muscle protein synthesis through the activation of translation
initiation factors (11). However, the mechanism underlying the effect of leucine on mTORC1
was not investigated. Therefore, in this study we wished to determine whether physiological
changes in circulating leucine levels modulates the activation of signaling components upstream
of mTOR (AMPK, PKB, and TSC2) and the protein-protein interactions between members of

10
the mTORC1 and mTORC2 complexes and whether the response is affected by rapamycin
treatment. We found that neither leucine infusion nor rapamycin treatment affected the
phosphorylation of AMPK, PKB, or TSC2 (Fig. 3).
In cell culture (17), rapamycin’s potency for dissociating the interaction of raptor with
mTOR is increased under leucine-rich conditions and this is correlated with a decrease in
mTORC1 kinase activity. Therefore, we hypothesized that rapamycin’s ability to disturb the
raptor-mTOR interaction would be greater in skeletal muscle of leucine-infused piglets than in
saline-infused piglets. As shown in Fig. 4A, rapamycin severely destabilized the raptor-mTOR
complex. Although, there was no significant difference between the presence or absence of
leucine in the potency of rapamycin in perturbing the interaction of raptor with mTOR, there was
a tendency (P=0.20) for the rapamycin-induced dissociation of the raptor-mTOR complex to be
greater in the leucine-treated group compared to the control group. Furthermore, rapamycin did
not affect the GβL-mTOR complex (Fig. 4B), as well as the rictor-mTOR complex (mTORC2)
(Fig. 4C).
Activation of mTOR and its downstream signaling components. Next, we wished to
investigate the effect of rapamycin on the leucine-induced activation of mTOR and its
downstream components leading to the stimulation of skeletal muscle protein synthesis in
neonatal pigs. In this study we determined mTOR activation by measuring mTOR
phosphorylation at Ser 2448 and Ser 2481. Leucine enhanced mTOR phosphorylation at both
sites (P < 0.05) and this effect was abolished by rapamycin (Fig. 5).
mTOR regulates mRNA translation in response to nutrients/leucine and growth factors
via activation of its downstream substrates, S6K1 and 4E-BP1 (4). To determine the effects of
leucine and rapamycin on the activation of these important effectors of mTOR, the

11
phosphorylation state of S6K1 at Thr 398 and 4E-BP1 at Thr 70 were analyzed using
immunoblot analysis. Leucine significantly enhanced the phosphorylation of S6K1 and 4E-BP1
in skeletal muscle (P < 0.05) (Figs. 6A and 6B, respectively) and its effects on the
phosphorylation of these mTOR effectors were completely abolished by rapamycin.
One of the major factors that regulate the formation of an active eIF4E∙eIF4G complex is
4E-BP1 (29). In the hypo-phosphorylated state, this protein binds eIF4E to form an inactive
complex. Conversely, in the hyper-phosphorylated state, 4E-BP1 detaches from eIF4E, allowing
eIF4G to assemble an active eIF4E∙eIF4G complex and initiate mRNA translation. To determine
the effects of leucine and rapamycin on the assembly of an active eIF4E∙eIF4G complex and an
inactive eIF4E∙4EBP1 complex, immunoprecipitation and immunoblot analysis were performed.
As shown in Fig. 7A and 7B, leucine increased the formation of the active eIF4E∙eIF4G complex
(P < 0.05) and inhibited the formation of an inactive eIF4E∙4EBP1 complex (P < 0.05). The
leucine-induce formation of the active eIF4E∙eIF4G complex was completely prevented by
rapamycin (Fig. 7A). Likewise, the inhibitory effect of leucine on the formation of the inactive
eIF4E∙4EBP1 complex was eliminated by rapamycin (Fig. 7B).
Translation elongation in mammalian cells requires two eEFs (eukaryotic elongation
factors), eEF1 and eEF2. eEF2 mediates the translocation of the ribosome by three nucleotides
along the mRNA after the addition of each new amino acid. Phosphorylation of eEF2 on Thr 56
impairs its ability to bind ribosomes, thus inactivating this signaling protein and inhibiting
mRNA translation (18). In this study, we determined the effects of leucine on the
phosphorylation of eEF2 at Thr 56. As shown in Fig. 8, neither leucine nor rapamycin altered
the phosphorylation of eEF2 in skeletal muscle.

12
Effect of rapamycin on the protein abundance of the signaling components. Treatment
with rapamycin, which down regulates mTORC1 activation, has been associated with increased
autophagy, one of the intracellular protein degradation pathways (4). To determine whether
short-term exposure to rapamycin alters the degradation of several signaling proteins, we
measured their protein abundance. We found that the protein abundance of all signaling
components measured in the current study was not affected by short-term exposure to rapamycin
(data not shown)
DISCUSSION
Amino acids, especially the branched-chain amino acid, leucine, serve not only as
precursors for protein synthesis but also as signals that activate protein synthesis via activation of
intracellular signal transduction pathways that regulate mRNA translation (19). However,
despite tremendous efforts, the molecular mechanisms by which amino acids regulate mRNA
translation remain largely unknown. Studies using different cell culture systems have reported
conflicting results. Early data suggest that TSC1 and TSC2 are required for amino acid-induced
signaling through mTOR (34). Other studies indicate that rheb activation, but not TSC1/2
activation, is required for amino acid control of mTOR activation (30). A recent study suggested
that amino acid input into mTOR signaling was not mediated by the TSC1/2 complex or rheb but
by a class 3 phosphatidylinositol 3OH-kinase (PI3K) or human vacuolar protein sorting 34
(hVps34) (23). Our recent work demonstrated that a physiological rise in amino acids in vivo
had no effect on the phosphorylation of AMPK, PKB and TSC2 in skeletal muscle of neonatal
pigs, suggesting that amino acids transmit their signal downstream of the TSC1/2 complex (33).

13
Our in vivo studies have revealed that leucine is effective in stimulating protein synthesis
in skeletal muscle of the neonate although the response to leucine is less than that to a complete
amino acid mixture (11). The leucine-induced stimulation of skeletal muscle protein synthesis
involves the activation of signaling components downstream of mTOR leading to mRNA
translation. We further demonstrated that the acute (1 hr) leucine-induced stimulation of protein
synthesis is not maintained (2 hr), despite continued activation of the mTOR signaling pathway,
because circulating concentrations of essential amino acids fall to less than fasting levels as they
are utilized as substrates for protein synthesis (10). However, when circulating amino acid levels
are maintained at baseline levels for 2 hr, the leucine-induced stimulation of muscle protein
synthesis is maintained. In addition, we demonstrated that the stimulation of the mTOR
signaling pathway and protein synthesis in muscle by leucine, like that by feeding, insulin and
balanced amino acid mixtures, decreases with age (8; 10; 33). In this study, we wished to further
characterize the acute action of leucine by utilizing rapamycin, a potent inhibitor of mTOR.
Amino acids, especially leucine, regulate signaling through mTOR and thereby control a
number of components of the protein synthetic machinery, including initiation and elongation
factors (26). Rapamycin, a well-characterized mTOR inhibitor, has been widely used to study
mTOR functions including protein synthesis (21). Previously, we showed that rapamycin
completely blocked the feeding-induced stimulation of protein synthesis in liver, but only
attenuated 60% of the feeding-induced stimulation of skeletal muscle protein synthesis in
neonatal pigs (21). However, in the current study, infusion of leucine to raise circulating levels
of leucine to fed levels, similar to that achieved in previous feeding studies (5; 13), stimulated
muscle protein synthesis and this was completely blocked by rapamycin.

14
Whilst there is evidence of the involvement of the TSC1/2 complex on the amino acid-
induced activation of mTORC1 (34), in the current in vivo study we did not find any effect of
physiological levels of leucine on the phosphorylation of TSC2 and its kinases, PKB, and
AMPK. This is consistent with our previous finding that fed amino acid levels have no effect on
AMPK, PKB, or TSC2 phosphorylation, although raising insulin to fed levels increased PKB and
TSC2 phosphorylation but not AMPK (33). To the best of our knowledge, this is the first in vivo
study to examine the effect of leucine on the activation of TSC1/TSC2 complex.
Mechanistic studies using cell cultures showed that amino acids regulate the interaction
of raptor with mTOR, resulting in the activation of mTORC1 and its downstream effectors,
S6K1 and 4E-BP1 (4). A recent cell culture study indicates that under nutrient or leucine-rich
conditions, the mTOR-raptor association is destabilized resulting in the activation of mTOR (16).
However, our in vivo studies in skeletal muscle (31; 33) and other studies in heart (37) showed
that although both feeding and amino acid infusion increase S6K1 and 4E-BP1 activation, no
change in the interaction of mTOR with raptor is detected. In this study, leucine infusion also
did not have any effect on the interaction of raptor with mTOR. Thus, further study of the
association of mTOR and raptor is required.
Rapamycin, like nutrient deprivation, inhibits the activation of downstream effectors of
mTOR, but exactly how the drug perturbs mTOR function is unknown (4). A recent study
showed that one of the actions of rapamycin is to interfere with the ability of raptor to effectively
present substrates (such as S6K1 and 4E-BP1) to mTOR (4). In support to this model, we found
that rapamycin significantly reduced the interaction of raptor with mTOR in both control and
leucine-treated groups. In cell culture conditions, additional of leucine to nutrient-poor media
enhanced the destabilizing effect of rapamycin on the raptor-mTOR complex (17). Interestingly,

15
we found that there was a tendency (P=0.20) for the rapamycin-induced dissociation of the
raptor-mTOR complex to be greater in the leucine-treated group compared to the control group.
GβL is a crucial member of mTORC1 because its binding to the kinase domain of mTOR
stabilizes the interaction of raptor with mTOR (4). Furthermore, mTOR, GβL and rictor are
members of mTORC2, a rapamycin insensitive complex (4). While we were able to detect both
GβL and rictor in mTOR-immunoprecipitants, their interactions were not affected by rapamycin
treatment in vivo. To the best of our knowledge, this is the first in vivo study to examine the
effect of rapamycin on leucine regulation of the integrity of the mTORC1 and mTORC2
complexes.
Several lines of evidence suggest that leucine induces the activation of mTOR and its
downstream substrates (S6K1 and 4E-BP1) that control in part the step in translation initiation
involving the binding of mRNA to 40S ribosomal subunit (20). In this study, we found that in
fasting pigs, rapamycin did not reduce the phosphorylation of mTOR, S6K1, and 4E-BP1 below
the basal level. In addition, rapamycin completely blocked the leucine-induced phosphorylation
of mTOR, S6K1 and 4E-BP1 in skeletal muscle. Furthermore, rapamycin completely blunted
the eIF4G association with eIF4E and restored the association between 4E-BP1 and eIF4E. Most
of these observations are in agreement with a study from Vary et al (36).
Recent studies suggest that one of the key roles of mTOR is to regulate the activation of
elongation factors (25). Previously we found that neither amino acids nor leucine alter eEF2
phosphorylation (12; 33). Interestingly, rapamycin did not have any effect on the
phosphorylation of eEF2. Since eEF2 is thought to be one of the major substrates of mTOR
(25), the lack of effect of rapamycin on eEF2 phosphorylation is surprising but it could be due
to the physiological dose of leucine used in this study.

16
In summary, in the present study, we showed that the leucine-induced stimulation of
protein synthesis and the activation of signaling components leading to mRNA translation in
skeletal muscle of the neonate are sensitive to rapamycin. This contrasts with a study published
while we were preparing this manuscript which showed that the feeding-induced activation of 5’
mRNA translation is resistant to rapamycin in fetal liver (hepatocytes) but not adult liver (15).
In the current study, rapamycin blocked leucine-induced phosphorylation of mTOR, S6K1, and
4EBP1 and the formation of active eIF4E•eIF4G complex in neonatal muscle, in agreement with
data from cell culture studies (17; 22). However, study of the effect of leucine on the protein-
protein interaction of members of the mTORC1 complex is more challenging as we were unable
to detect an effect of leucine on raptor-mTOR association in vivo although we did show a
tendency toward leucine-induced loosening of the raptor-mTOR association, a condition that has
been documented in cell culture studies (17). Nevertheless, it is clear that our understanding of
the molecular mechanisms by which leucine regulates the signaling function of mTOC1 in vivo
is incomplete. Considering the potential significance of leucine’s action as a nutrient signal to
stimulate muscle protein synthesis in vivo, further studies to elucidate its mechanistic functions
are warranted.

17
ACKNOWLEDGMENTS
We thank D. E. Miller and J. C. Stubblefield for care of animals and L. F. Weiser for
secretarial assistance.
GRANTS
This work is publication of the United States Department of Agriculture/Agricultural
Research Service (USDA/ARS) Children’s Nutrition Research Center, Department of Pediatrics,
Baylor College of Medicine. This project has been funded in part by National Institutes of
Health (NIH) grant AR-44474 (T. A, Davis), and by the USDA/ARS under Cooperative
Agreement no. 6250510000-33 (T. A. Davis). The contents of this publication do not necessarily
reflect the views or policies of U.S. Department of Agriculture, nor does mention of trade names,
commercial products, or organizations imply endorsement by the U.S. Government.

18
REFERENCES
1. Anthony JC, Yoshizawa F, Anthony TG, Vary TC, Jefferson LS and Kimball SR.
Leucine stimulates translation initiation in skeletal muscle of postabsorptive rats via a
rapamycin-sensitive pathway. J Nutr 130: 2413-2419, 2000.
2. Bolster DR, Vary TC, Kimball SR and Jefferson LS. Leucine regulates translation
initiation in rat skeletal muscle via enhanced eIF4G phosphorylation. J Nutr 134: 1704-
1710, 2004.
3. Chien PF, Smith K, Watt PW, Scrimgeour CM, Taylor DJ and Rennie MJ. Protein
turnover in the human fetus studied at term using stable isotope tracer amino acids. Am J
Physiol 265: E31-E35, 1993.
4. Corradetti MN and Guan KL. Upstream of the mammalian target of rapamycin: do all
roads pass through mTOR? Oncogene 25: 6347-6360, 2006.
5. Davis TA, Burrin DG, Fiorotto ML and Nguyen HV. Protein synthesis in skeletal
muscle and jejunum is more responsive to feeding in 7-than in 26-day-old pigs. Am J
Physiol 270: E802-E809, 1996.
6. Davis TA, Fiorotto ML, Burrin DG, Reeds PJ, Nguyen HV, Beckett PR, Vann RC and
O'Connor PM. Stimulation of protein synthesis by both insulin and amino acids is unique
to skeletal muscle in neonatal pigs. Am J Physiol Endocrinol Metab 282: E880-E890, 2002.

19
7. Davis TA, Fiorotto ML, Nguyen HV and Burrin DG. Aminoacyl-tRNA and tissue free
amino acid pools are equilibrated after a flooding dose of phenylalanine. Am J Physiol 277:
E103-E109, 1999.
8. Davis TA, Nguyen HV, Suryawan A, Bush JA, Jefferson LS and Kimball SR.
Developmental changes in the feeding-induced stimulation of translation initiation in
muscle of neonatal pigs. Am J Physiol Endocrinol Metab 279: E1226-E1234, 2000.
9. Ehrenkranz RA. Early, aggressive nutritional management for very low birth weight
infants: what is the evidence? Semin Perinatol 31: 48-55, 2007.
10. Escobar J, Frank JW, Suryawan A, Nguyen HV and Davis TA. Amino acid
Availability and Age Affect the Leucine Stimulation of Protein Synthesis and eIF4F
Formation in Muscle. Am J Physiol Endocrinol Metab 2007.
11. Escobar J, Frank JW, Suryawan A, Nguyen HV, Kimball SR, Jefferson LS and Davis
TA. Physiological rise in plasma leucine stimulates muscle protein synthesis in neonatal
pigs by enhancing translation initiation factor activation. American Journal of Physiology-
Endocrinology and Metabolism 288: E914-E921, 2005.
12. Escobar J, Frank JW, Suryawan A, Nguyen HV, Kimball SR, Jefferson LS and Davis
TA. Regulation of cardiac and skeletal muscle protein synthesis by individual branched-
chain amino acids in neonatal pigs. American Journal of Physiology-Endocrinology and
Metabolism 290: E612-E621, 2006.

20
13. Frank JW, Escobar J, Suryawan A, Nguyen HV, Kimball SR, Jefferson LS and Davis
TA. Dietary protein and lactose increase translation initiation factor activation and tissue
protein synthesis in neonatal pigs. American Journal of Physiology-Endocrinology and
Metabolism 290: E225-E233, 2006.
14. Garlick PJ, McNurlan MA and Preedy VR. A rapid and convenient technique for
measuring the rate of protein synthesis in tissues by injection of [3H]phenylalanine.
Biochem J 192: 719-723, 1980.
15. Gruppuso PA, Tsai SW, Boylan JM and Sanders JA. Hepatic translation control in the
late gestation fetal rat. Am J Physiol Regul Integr Comp Physiol In Print, 2008.
16. Kim DH and Sabatini DM. Raptor and mTOR: subunits of a nutrient-sensitive complex.
Curr Top Microbiol Immunol 279: 259-270, 2004.
17. Kim DH, Sarbassov DD, Ali SM, King JE, Latek RR, Erdjument-Bromage H, Tempst
P and Sabatini DM. mTOR interacts with raptor to form a nutrient-sensitive complex that
signals to the cell growth machinery. Cell 110: 163-175, 2002.
18. Kimball SR. Regulation of translation initiation by amino acids in eukaryotic cells. Prog
Mol Subcell Biol 26: 155-184, 2001.
19. Kimball SR and Jefferson LS. Regulation of protein synthesis by branched-chain amino
acids. Current Opinion in Clinical Nutrition and Metabolic Care 4: 39-43, 2001.

21
20. Kimball SR and Jefferson LS. Role of amino acids in the translational control of protein
synthesis in mammals. Semin Cell Dev Biol 16: 21-27, 2005.
21. Kimball SR, Jefferson LS, Nguyen HV, Suryawan A, Bush JA and Davis TA. Feeding
stimulates protein synthesis in muscle and liver of neonatal pigs through an mTOR-
dependent process. Am J Physiol Endocrinol Metab 279: E1080-E1087, 2000.
22. Kimball SR, Shantz LM, Horetsky RL and Jefferson LS. Leucine regulates translation
of specific mRNAs in L6 myoblasts through mTOR-mediated changes in availability of
eIF4E and phosphorylation of ribosomal protein S6. J Biol Chem 274: 11647-11652, 1999.
23. Nobukuni T, Joaquin M, Roccio M, Dann SG, Kim SY, Gulati P, Byfield MP, Backer
JM, Natt F, Bos JL, Zwartkruis FJ and Thomas G. Amino acids mediate mTOR/raptor
signaling through activation of class 3 phosphatidylinositol 3OH-kinase. Proc Natl Acad
Sci U S A 102: 14238-14243, 2005.
24. O'Connor PM, Bush JA, Suryawan A, Nguyen HV and Davis TA. Insulin and amino
acids independently stimulate skeletal muscle protein synthesis in neonatal pigs. Am J
Physiol Endocrinol Metab 284: E110-E119, 2003.
25. Proud CG. Role of mTOR signalling in the control of translation initiation and elongation
by nutrients. Curr Top Microbiol Immunol 279: 215-244, 2004.

22
26. Proud CG. Amino acids and mTOR signalling in anabolic function. Biochem Soc Trans
35: 1187-1190, 2007.
27. Saigal S, Stoskopf BL, Streiner DL and Burrows E. Physical growth and current health
status of infants who were of extremely low birth weight and controls at adolescence.
Pediatrics 108: 407-415, 2001.
28. Sarbassov DD, Ali SM, Sengupta S, Sheen JH, Hsu PP, Bagley AF, Markhard AL and
Sabatini DM. Prolonged rapamycin treatment inhibits mTORC2 assembly and Akt/PKB.
Mol Cell 22: 159-168, 2006.
29. Shah OJ, Anthony JC, Kimball SR and Jefferson LS. 4E-BP1 and S6K1: translational
integration sites for nutritional and hormonal information in muscle. Am J Physiol
Endocrinol Metab 279: E715-E729, 2000.
30. Smith EM, Finn SG, Tee AR, Browne GJ and Proud CG. The tuberous sclerosis protein
TSC2 is not required for the regulation of the mammalian target of rapamycin by amino
acids and certain cellular stresses. J Biol Chem 280: 18717-18727, 2005.
31. Suryawan A, Escobar J, Frank JW, Nguyen HV and Davis TA. Developmental
regulation of the activation of signaling components leading to translation initiation in
skeletal muscle of neonatal pigs. Am J Physiol Endocrinol Metab 291: E849-E859, 2006.

23
32. Suryawan A, Nguyen HV, Bush JA and Davis TA. Developmental changes in the
feeding-induced activation of the insulin-signaling pathway in neonatal pigs. Am J Physiol
Endocrinol Metab 281: E908-E915, 2001.
33. Suryawan A, Orellana RA, Nguyen HV, Jeyapalan AS, Fleming JR and Davis TA.
Activation by Insulin and Amino Acids of Signaling Components Leading to Translation
Initiation in Skeletal Muscle of Neonatal Pigs Is Developmentally Regulated. Am J Physiol
Endocrinol Metab 2007.
34. Tee AR, Fingar DC, Manning BD, Kwiatkowski DJ, Cantley LC and Blenis J.
Tuberous sclerosis complex-1 and -2 gene products function together to inhibit mammalian
target of rapamycin (mTOR)-mediated downstream signaling. Proc Natl Acad Sci U S A
99: 13571-13576, 2002.
35. Vary TC. Acute oral leucine administration stimulates protein synthesis during chronic
sepsis through enhanced association of eukaryotic initiation factor 4G with eukaryotic
initiation factor 4E in rats. J Nutr 137: 2074-2079, 2007.
36. Vary TC, Anthony JC, Jefferson LS, Kimball SR and Lynch CJ. Rapamycin blunts
nutrient stimulation of eIF4G, but not PKCepsilon phosphorylation, in skeletal muscle. Am
J Physiol Endocrinol Metab 293: E188-E196, 2007.

24
37. Vary TC, Deiter G and Lynch CJ. Rapamycin limits formation of active eukaryotic
initiation factor 4F complex following meal feeding in rat hearts. J Nutr 137: 1857-1862,
2007.
38. Wang X and Proud CG. Methods for studying signal-dependent regulation of translation
factor activity. Methods Enzymol 431: 113-142, 2007.
39. Williamson DL, Bolster DR, Kimball SR and Jefferson LS. Time course changes in
signaling pathways and protein synthesis in C2C12 myotubes following AMPK activation
by AICAR. Am J Physiol Endocrinol Metab 291: E80-E89, 2006.

25
FIGURE LEGENDS
Figure 1. Current concepts of the amino acid signaling pathway leading to protein
synthesis.
Figure 2. Fractional rates of protein synthesis in skeletal muscle of 7-day-old-pigs after
60 min of infusion of saline (Sal), saline with rapamycin (Sal+Rap), 400 µmol·kg–1
·h–1
of
leucine without rapamycin (Leu), and 400 µmol·kg–1
·h–1
of leucine with rapamycin (Leu+Rap).
Values are means ± pooled SEM; n = 5-7 per treatment. a,b
Values with different superscripts
differ significantly (P < 0.05).
Figure 3. Phosphorylation of protein kinase B (PKB) at Ser473 (A), AMP-activated
protein kinase (AMPK) at Thr172 (B), and tuberous sclerosis complex-2 (TSC2) at Thr1462 (C)
in skeletal muscle of 7-day-old-pigs after 60 min of infusion of saline (Sal), saline with
rapamycin (Sal+Rap), 400 µmol·kg–1
·h–1
of leucine without rapamycin (Leu), and 400 µmol·kg–
1·h
–1 of leucine with rapamycin (Leu+Rap). Phosphorylation of PKB, AMPK, and TSC2 were
corrected by total PKB, AMPK, and TSC2, respectively. Values are means ± pooled SEM; n =
5-7 per treatment.
Figure 4. Association between mTOR and raptor (A), mTOR and GβL (B), and mTOR
and rictor (C) in skeletal muscle of 7-day-old-pigs after 60 min of infusion saline (Sal), saline
with rapamycin (Sal+Rap), 400 µmol·kg–1
·h–1
of leucine without rapamycin (Leu), and 400
µmol·kg–1
·h–1
of leucine with rapamycin (Leu+Rap). Association between mTOR and its
partners was corrected by total mTOR in the immunoprecipitant. Values are means ± pooled
SEM; n = 5-7 per treatment. a,b
Values with different superscripts differ significantly (P < 0.05).
Figure 5. Phosphorylation of mammalian target of rapamycin (mTOR) at Ser2481 (A),
at Ser2448 (B) in skeletal muscle of 7-day-old-pigs after 60 min of infusion of saline (Sal), saline

26
with rapamycin (Sal+Rap), 400 µmol·kg–1
·h–1
of leucine without rapamycin (Leu), and 400
µmol·kg–1
·h–1
of leucine with rapamycin (Leu+Rap). Phosphorylation of mTOR were corrected
by total mTOR. Values are means ± pooled SEM; n = 5-7 per treatment. a,b
Values with different
superscripts differ significantly (P < 0.05)
Figure 6. Phosphorylation of the 70-kDa ribosomal protein S6 kinase (S6K1) at Thr389
(A), and eukaryotic initiation factor (eIF) binding protein-1 (4E-BP1) at Thr70 (B) in skeletal
muscle of 7-day-old-pigs after 60 min of infusion of saline (Sal), saline with rapamycin
(Sal+Rap), 400 µmol·kg–1
·h–1
of leucine without rapamycin (Leu), and 400 µmol·kg–1
·h–1
of
leucine with rapamycin (Leu+Rap). Phosphorylation of S6K1 and 4E-BP1 were corrected by
total S6K1 and 4E-BP1, respectively. Values are means ± pooled SEM; n = 5-7 per treatment.
a,bValues with different superscripts differ significantly (P < 0.05).
Figure 7. Association of eukaryotic initiation factor (eIF) 4G with eIF4E (A) and 4E-
BP1 with eIF4E (B) in skeletal muscle of 7-day-old-pigs after 60 min of infusion of saline (Sal),
saline with rapamycin (Sal+Rap), 400 µmol·kg–1
·h–1
of leucine without rapamycin (Leu), and 400
µmol·kg–1
·h–1
of leucine with rapamycin (Leu+Rap). The formation of eIF4G•eIF4E and 4E-
BP1•eIF4E complexes were corrected by total eIF4E in the immunoprecipitant. Values are
means ± pooled SEM; n = 5-7 per treatment. a,b
Values with different superscripts differ
significantly (P < 0.05).
Figure 8. Phosphorylation of eukaryotic elongation factor 2 (eEF2) at Thr56 in skeletal
muscle of 7-day-old-pigs after 60 min of infusion of saline (Sal), saline with rapamycin
(Sal+Rap), 400 µmol·kg–1
·h–1
of leucine without rapamycin (Leu), and 400 µmol·kg–1
·h–1
of
leucine with rapamycin (Leu+Rap). Phosphorylation of eEF2 was corrected by total eEF2.
Values are means ± pooled SEM; n = 5-7 per treatment.

Figure 1
Growth Factors(Insulin)
IRS 1/2P
PKBP
AMPK
Amino Acids(Leucine)
Rheb
TSC2TSC1P
mTORGβL Rictor mTORC2mTORC1 mTOR
GβL
Raptor
Rapamycin
P
S6K14EBP1
eIF4E eIF4G
Raptor
4EBP1eIF4E
P
P
P
P
eEF2K
eEF2 P
AMPK
S6eIF4E eIF4GeEF2
Initiation Elongation
Protein Synthesis
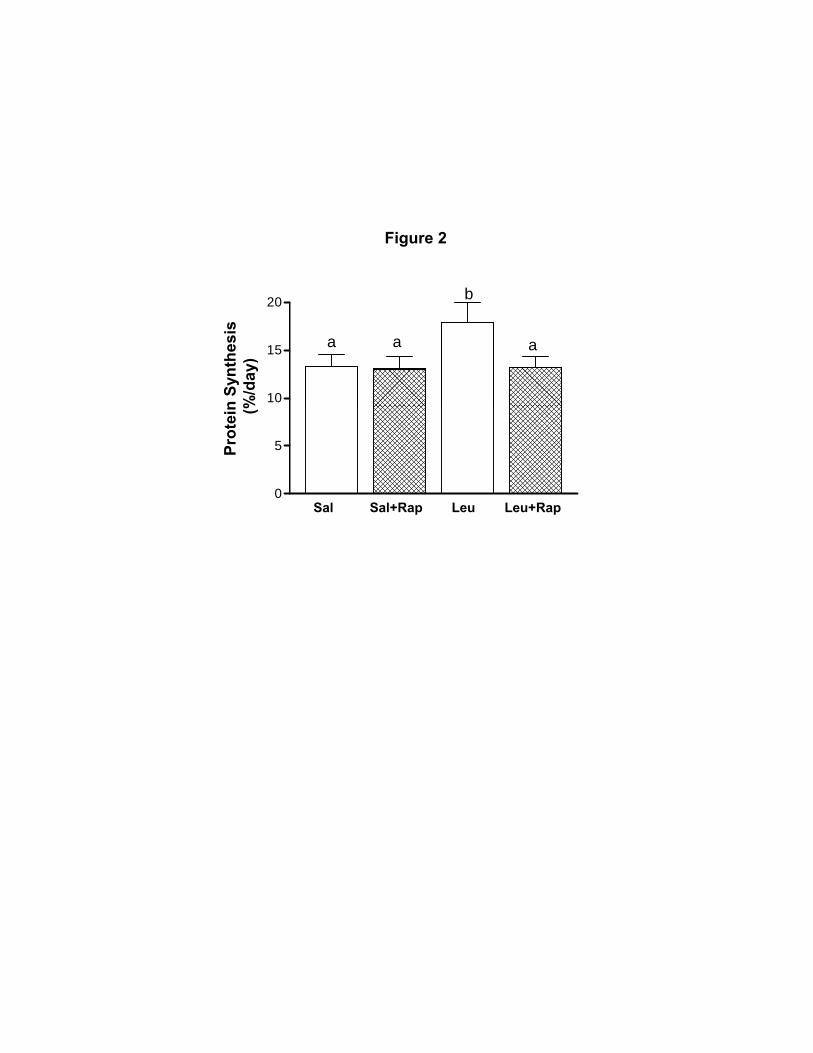
Figure 2n
Synt
hesi
s %
/day
)
10
15
20
a aa
b
Sal Sal+Rap Leu Leu+Rap
Prot
ein (%
0
5

A1.5
Figure 3
PKB
PKBSer 473
P-PK
B (
AU
)
0.5
1.0P
Sal Sal+Rap Leu Leu+Rap0.0
AMPKThr 172
K (A
U)
B
0.75
1.00
AMPK
AMPK
P-A
MPK
0.25
0.50
C
Sal Sal+Rap Leu Leu+Rap0.00
TSC2
TSC2Thr 1462
0.3
0.4
0.5
0.6
C2
(AU
)
0.0
0.1
0.2
P-TS
Sal Sal+Rap Leu Leu+Rap

plex
Figure 4
2.0A
mTOR
Raptor
or -
mTO
R c
omp
(AU
)
0 5
1.0
1.5 aa
Sal Sal+Rap Leu Leu+Rap
Rap
to
0.0
0.5b
b
GβL
ompl
ex
3
4
5B mTOR
β
GβL
-m
TOR
co
(AU
)
0
1
2
3
Sal Sal+Rap Leu Leu+Rap0
7.5C mTOR
Rictor
mTO
R c
ompl
ex
(AU
)
2 5
5.0
7.5
Ric
tor -
m
Sal Sal+Rap Leu Leu+Rap0.0
2.5

Figure 5
Ser 2448
A4
b
mTOR
mTORSer 2448
mTO
R (
AU
)
2
3
a aa
b
P-m
Sal Sal+Rap Leu Leu+Rap0
1
mTOR
mTORSer 2481
0.75
1.00
b
(AU
)
B
0.00
0.25
0.50
a aaP-m
TOR
Sal Sal+Rap Leu Leu+Rap

Figure 6
A1 5
S6K1
S6K1Thr 389
P-S6
K1
(AU
)
1.0
1.5
b
P
Sal Sal+Rap Leu Leu+Rap0.0
0.5
a aa
4EBP1
4EBP1Thr 70
B
P1 (A
U)
0.50
0.75
b
P-4E
BP
Sal Sal+Rap Leu Leu+Rap0.00
0.25a aa
Sal Sal Rap Leu Leu Rap

Figure 7
AeIF4E
eIF4G
F4E
com
plex
(A
U) 5.0
7.5
b
eIF4
G-e
IF
Sal Sal+Rap Leu Leu+Rap0.0
2.5
a aa
IF4E
4EBP1
ompl
ex
B
0.50
0.75a
aa
eIF4E
4EB
P1-e
IF4E
co
(AU
)
0 00
0.25
b0.00
Sal Sal+Rap Leu Leu+Rap

Figure 8
eEF2Thr 56
15
U)
eEF2
5
10
P-eE
F2 (A
U
0Sal Sal+Rap Leu Leu+Rap
Copyright © 2022 FDOKUMEN




















