
Characterization of wheat homeodomain-leucine zipper family ...
Upload
khangminh22Category
view
1download
0
University of Pennsylvania University of Pennsylvania
ScholarlyCommons ScholarlyCommons
Publicly Accessible Penn Dissertations
2011
Extracellular Leucine-Rich Repeat Protein Let-4 is Required to Extracellular Leucine-Rich Repeat Protein Let-4 is Required to
Organize the Extracellular Matrix and Maintain Junctions in C. Organize the Extracellular Matrix and Maintain Junctions in C.
Elegans Epithelia Elegans Epithelia
Vincent Pasquale Mancuso University of Pennsylvania, [email protected]
Follow this and additional works at: https://repository.upenn.edu/edissertations
Part of the Cell Biology Commons, and the Developmental Biology Commons
Recommended Citation Recommended Citation Mancuso, Vincent Pasquale, "Extracellular Leucine-Rich Repeat Protein Let-4 is Required to Organize the Extracellular Matrix and Maintain Junctions in C. Elegans Epithelia" (2011). Publicly Accessible Penn Dissertations. 542. https://repository.upenn.edu/edissertations/542
This paper is posted at ScholarlyCommons. https://repository.upenn.edu/edissertations/542 For more information, please contact [email protected].

Extracellular Leucine-Rich Repeat Protein Let-4 is Required to Organize the Extracellular Leucine-Rich Repeat Protein Let-4 is Required to Organize the Extracellular Matrix and Maintain Junctions in C. Elegans Epithelia Extracellular Matrix and Maintain Junctions in C. Elegans Epithelia
Abstract Abstract Epithelial cells line the interior of many organs, therefore a better understanding of how these cells are maintained could offer insights into many human diseases. This work focuses on two aspects of epithelia: cell junctions and the apical Extracellular Matrix (ECM). Epithelial cell junctions consist of conserved junction proteins that connect cells to each other, serve as a barrier, and separate the apical and basal domains of the cells. The specialized apical ECM of epithelial cells serves to protect the cells and interact with the outside environment. The apical ECM is present in many epithelia, but is poorly studied.
I have characterized the apical domains of epithelial cells and their junctions in the Caenorhabditis elegans excretory system in order to develop the organ as a model for epithelial development and maintenance. With my colleagues, I studied the development of the epithelial cells of the excretory system, and described roles for Ras and Notch in a biased competition model of cell fate determination. I have characterized the localization of conserved proteins of the polarity PAR complex, and the junctional cadherin-catenin complex using molecular markers and immunohistochemistry. This has established that the excretory system shares common features with other epithelia, which supports the C. elegans excretory system as a good epithelial model system.
I have also shown that extracellular leucine-rich repeat protein LET-4 has a role in organization of the apical ECM, and junction maintenance in both the excretory system and epidermis of C. elegans. Characterization of the LET-4 protein has indicated that it localizes apically in epithelial cells, but is not enriched at junctions. I characterized the let-4 loss of function phenotype with molecular markers, electron microscopy, and genetic interaction experiments, and have detected no defect in initial formation of junctions. These data suggest that it is not initial junction formation, but junction integrity that is affected in let-4 mutants. The let-4 analysis suggests that epithelial junction maintenance requires an intact apical ECM. The continuation of this work will involve further characterization of the molecular components of the apical ECM and identification of LET-4 interacting proteins.
Degree Type Degree Type Dissertation
Degree Name Degree Name Doctor of Philosophy (PhD)
Graduate Group Graduate Group Cell & Molecular Biology
First Advisor First Advisor Meera V. Sundaram
Keywords Keywords C. elegans, Epithelial cells, Extracellular matrix
Subject Categories Subject Categories Cell Biology | Developmental Biology
This dissertation is available at ScholarlyCommons: https://repository.upenn.edu/edissertations/542

EXTRACELLULAR LEUCINE-RICH REPEAT PROTEIN LET-4 IS REQUIRED TO
ORGANIZE THE EXTRACELLULAR MATRIX AND MAINTAIN JUNCTIONS IN
C. ELEGANS EPITHELIA
Vincent Pasquale Mancuso
A DISSERTATION
in
Cell and Molecular Biology
Presented to the Faculties of the University of Pennsylvania
in
Partial Fulfillment of the Requirements for the
Degree of Doctor of Philosophy
2011
Supervisor of Dissertation
________________________ Meera V. Sundaram. Ph.D., Associate Professor of Genetics
Graduate Group Chairperson
__________________________ Daniel S. Kessler, Ph.D., Associate Professor of Cell and Developmental Biology
Dissertation Committee:
Douglas J. Epstein, Ph.D., Associate Professor of Genetics
Stephen DiNardo, Ph.D., Professor of Cell and Developmental Biology
Amin S. Ghabrial, Ph.D., Assistant Professor of Cell and Developmental Biology
S. Todd Lamitina, Ph.D., Assistant Professor of Physiology

ii
Acknowledgements
Thank you to my thesis advisor Meera Sundaram, for welcoming me into your
lab, and patiently guiding me through my graduate career and research. I’ve always been
impressed with your intellectual rigor, enthusiasm for research, and your willingness to
give students your time. Thanks for being so generous with your time and talent to help
me grow as a scientist, speaker, teacher, and writer.
Thank you to the members of my thesis committee, Doug Epstein, Steve
DiNardo, Amin Ghabrial and Todd Lamitina, for your time and valuable guidance.
Thank you to the Worm Group for your helpful suggestions and insights over the years.
Thank you to my friends here at Penn for your support and insights, and particularly
Kelly Howell, Balpreet Bhogal and Jean Parry for helping me have at least a little fun in
lab every day.
Thank you to my whole family, particularly my parents, sister, brother and Uncle
Tom for your ceaseless encouragement as I pursued my goal of becoming a scientist.
Finally, I’m especially thankful to my wife and daughter; your patience, support and love
will always inspire me.

iii
ABSTRACT
EXTRACELLULAR LEUCINE-RICH REPEAT PROTEIN LET-4 IS REQUIRED
TO ORGANIZE THE EXTRACELLULAR MATRIX AND MAINTAIN JUNCTIONS IN
C. ELEGANS EPITHELIA
Vincent Pasquale Mancuso
Meera V. Sundaram
Epithelial cells line the interior of many organs, therefore a better understanding
of how these cells are maintained could offer insights into many human diseases. This
work focuses on two aspects of epithelia: cell junctions and the apical Extracellular
Matrix (ECM). Epithelial cell junctions consist of conserved junction proteins that
connect cells to each other, serve as a barrier, and separate the apical and basal domains
of the cells. The specialized apical ECM of epithelial cells serves to protect the cells and
interact with the outside environment. The apical ECM is present in many epithelia, but
is poorly studied.
I have characterized the apical domains of epithelial cells and their junctions in
the Caenorhabditis elegans excretory system in order to develop the organ as a model for
epithelial development and maintenance. With my colleagues, I studied the development
of the epithelial cells of the excretory system, and described roles for Ras and Notch in a
biased competition model of cell fate determination. I have characterized the localization
of conserved proteins of the polarity PAR complex, and the junctional cadherin-catenin

iv
complex using molecular markers and immunohistochemistry. This has established that
the excretory system shares common features with other epithelia, which supports the C.
elegans excretory system as a good epithelial model system.
I have also shown that extracellular leucine-rich repeat protein LET-4 has a role
in organization of the apical ECM, and junction maintenance in both the excretory system
and epidermis of C. elegans. Characterization of the LET-4 protein has indicated that it
localizes apically in epithelial cells, but is not enriched at junctions. I characterized the
let-4 loss of function phenotype with molecular markers, electron microscopy, and
genetic interaction experiments, and have detected no defect in initial formation of
junctions. These data suggest that it is not initial junction formation, but junction
integrity that is affected in let-4 mutants. The let-4 analysis suggests that epithelial
junction maintenance requires an intact apical ECM. The continuation of this work will
involve further characterization of the molecular components of the apical ECM and
identification of LET-4 interacting proteins.

v
Table of Contents
Title Page……………………………………………………………………..………i
Acknowledgements……………………………………………………..……...…….ii
Abstract…………………………………………………………………………..….iii
Table of Contents………………………………………………………………….....v
List of Tables………………………………………….…………………………....vii
List of Illustrations………………………………………………………………...viii
Chapter 1: General Introduction: Epithelial cells, their junctions,
and the apical extracellular matrix ………………………………………………….1
Chapter 2: Extracellular leucine-rich repeat proteins are required to organize
the apical extracellular matrix and maintain epithelial junction integrity
in C. elegans. . . …………….....................................................................................16
Chapter 3: Notch and Ras promote sequential steps of excretory tube development
in C.elegans………………………………………………………………….…..….61
Chapter 4: Discussion: The role of LET-4 in the apical ECM and junctions,
and approaches to identify LET-4 interacting partners………………………….....104
Appendix 1: Characterization of apical and cytoskeletal markers
in the C. elegans excretory system…………………………………………...…….131
Appendix 2: let-4 expression in the excretory system……………….………..…....154
Appendix 3: TEM analysis of the C. elegans excretory system…………………....161

vi
Appendix 4: Detailed protocols…………………………………….………….……198
TEM section staining protocol………………………………….....….……..199
Larvae immunostaing protocol…………………………..……..…….….….201
Freeze-crack immunostaining protocol…………………..……….…....……204
Laser ablation protocol……………………………………..……...…..…….205
Bibliography……………………………………………..…………………….…….206

vii
List of Tables
Table 3.1 Physical or genetic removal of the canal cell reduces but does not prevent
duct fate specification……………….………………………...……………..…90
Table 4.1 let-4; lpr-1 double mutant phenotype……………………………………….125
Table 4.2 RNAi knockdown of worm lipocalins does not result in larval lethality ..…126
Table 4.3 let-4 and let-653 RNAi ……………………………………………………..127
Table Ap1.1 Excretory System markers appear normal in let-4 mutants…….……..…142
Table Ap3.1 Summary of 3-fold embryos analyzed by TEM …………………………170

viii
List of Illustrations
Figure 1.1 Epithelial cells have apical and basal sides; both can be surrounded by an
ECM………………………………………………………………………….15
Figure 2.1 The apical ECM differs between excretory tube types…………….……..53
Figure 2.2 let-4 and egg-6 encode related transmembrane proteins with extracellular
leucine-rich repeats…………………………………………………………..54
Figure 2.3 LET-4::GFP and EGG-6::GFP localize to the apical domains of the excretory
duct, pore and epidermal cells…………………………………………….….55
Figure 2.4 let-4 and egg-6 are each required to maintain junction integrity between the
excretory duct and pore………………………………………………..……..56
Figure 2.5 let-4 and sym-1 are redundantly required to maintain junction integrity in the
epidermis……………………………………………………………………..57
Figure 2.6 The cytoplasmic and transmembrane domains of LET-4 are dispensable
for function……………………………………………………..……….……58
Figure 2.7 let-4 and egg-6 are required for proper apical ECM organization….…….59
Figure 2.8. LET-4::GFP and EGG-6::GFP localize apically in epithelia………...…..60
Figure 3.1 Timeline of excretory system development………………………...…….96
Figure 3.2 let-60/Ras promotes the duct vs. G1 pore fate…………………….….…..97
Figure 3.3 lin-3/EGF, let-23/EGFR and lin-12/Notch reporter expression
in the excretory system…………………………………………..…….……..98
Figure 3.4 let-60/Ras hypomorphs reveal defects in cell competition and stacking....99
Figure 3.5 The canal cell is required for duct and G1 pore stacking and
tubulogenesis………………………………………………………………....100
Figure 3.6 sos-1 temperature shift experiments reveal continued requirements
during duct morphogenesis and differentiation………….……………..….…101

ix
Figure 3.7 G1 withdrawal and G2 entry can occur independently………..………….102
Figure 3.8 Transmission Electron Micrographs of ventral enclosure stage embryo….103
Figure 4.1 Three models for destabilization of the apical ECM leading to junction
breakage …………………………………………………………………....…129
Figure 4.2 Summary of let-4 genetic interaction data…………………………….…..130
Figure Ap1.1 The PAR complex proteins are present in the cells
of the excretory system…………………………………………..…...…...…..148
Figure Ap1.2 The C. elegans E-Cadherin homolog HMR-1 and associated protein
p120 catenin (JAC-1) are localized dynamically in the duct cell…….…..…...149
Figure Ap1.3 F-actin (Phalloidin) Staining in epidermal cells and the excretory
system…………………………………………………………………...….....150
Figure Ap1.4 Chitin and WGA epitope staining…………………..……………….....151
Figure Ap1.5 AJM-1 and IF staining in let-4 RNAi-treated embryos…….…..........…152
Figure Ap1.6 Further characterization of the let-4 phenotype……………….....……..153
Figure Ap2.1 Expression of let-4 in the excretory system……………………...……. 160
Figure Ap3.1 TEM images of the excretory system and detached
inner eggshell layer in 1.5-fold let-4 embryos……………………..….….…. .176
Figure Ap3.2 TEM images of the excretory system in wild-type 3-fold
embryos………………………………………………………………..…..…..178
Figure Ap3.3 TEM images of the excretory system in let-4 3-fold
embryos………………………………………………………………....….….180

Chapter 1
General Introduction: Epithelial cells, their junctions, and the apical extracellular
matrix
1

Defects in epithelial cells are responsible for a variety of human diseases
The interior of organs and the exterior surfaces of organisms are exposed to a
variety of environments, and therefore require specialized polarized epithelial cells. Two
major formations of epithelia are planar and tubular arrangements. In planar epithelia,
cells line up side by side, and the apical face of the epithelium faces the outside of the
organism. Tubular epithelia are joined together to form a tube shape to produce an apical
surface facing the interior lumen of internal tubular organs. Epithelial cells are
characterized by conserved junctions, which separate apical and basal sides of the cells.
Epithelial cells are surrounded by a specialized extracellular matrix (ECM), consisting of
proteins, lipids and polysaccharides (Fig. 1.1).
Because of the wide variety of epithelia, defects in epithelial development or
maintenance contribute to many human diseases. Maintenance of epithelial polarity is
required for adsorption and secretion in the kidney, and as a result, epithelial polarity
defects are associated a number of diseases, including Polycystic Kidney Disease
(Wilson, 2011). Polarity defects in skin cells results in blistering diseases (Niessen et al.,
2010). Epithelial barrier defects in the small intestine, which are associated with diseases
such as Crohn’s disease and ulcerative colitis, can be the result of failure of localization
of apical junctional proteins. (Marchiando et al., 2010). A deeper understanding of
epithelial maintenance could offer insight into treatment or prevention of many diseases.
Epithelial cell maintenance also has a key role in prevention of cancer. Epithelial-
to-mesenchymal transition (EMT), a required step for metastasis, is characterized by loss
of epithelial characteristics, such as cell-cell junctions and polarity, and acquisition of
2

motility and invasiveness. In EMT, epithelial cells lose their junctions with neighboring
cells, and break through the extra-cellular matrix (ECM), in order to colonize sites of
metastasis (Kalluri and Weinberg, 2009; Polyak and Weinberg, 2009). Given their role
in cancer progression, it is important to develop a better understanding of the mechanisms
by which epithelial junctions and the extracellular matrix are formed.
Epithelial junction proteins are conserved
Epithelial cells are characterized by specialized junctions which provide tissue
structure, form a permeability barrier, and separate the apical and basolateral surfaces of
epithelial cells (Knust and Bossinger, 2002; Shin et al., 2006; Giepmans and van
Ijzendoorn, 2009). Although epithelial cells are in a variety of tissue types in so many
organisms, the features of epithelial junctions are evolutionarily conserved. Cadherin-
based adherens junctions mediate cell-cell adhesion, and anchor actin to the membrane,
while claudin based junctions provide a permeability barrier, and separate the apical and
basal domains of the epithelial cells. Although these junctional domains are conserved,
the relative position of the junctions differs among organisms. In mammals, adherens
junctions are located basally to claudin-based tight junctions. In contrast, Drosophila
adherens junctions are apical to claudin-based septate junctions. In Caenorhabditis
elegans, adherens junctions and septate-junction-like domains are localized together
apically (Lynch and Hardin, 2009). In these diverse organisms, proper localization of
these junctional proteins requires conserved apical polarity regulators such as PAR
complex, Crumbs, and Scribble complex (Goldstein and Macara, 2007). PAR-6, PKC-3
and Crumbs localize to the most apical domain, PAR-3 localizes sub-apically to epithelial
3

junctions, and Scribble localizes basolaterally. As development proceeds, epithelial
junctions are disassembled or re-formed as tissues mature (Acloque et al., 2009; Baum
and Georgiou, 2011; St Johnston and Sanson, 2011). Although the precise mechanisms
are still being elucidated, interactions among polarity proteins, extracellular cues, the
protein trafficking machinery and the cytoskeleton all contribute to the initial
establishment of apico-basal polarity and appropriate placement of epithelial junctions.
The apical ECM is a dynamic structure with roles in pathogen protection, cell shape
determination, and signaling
Epithelial cells are surrounded by an extracellular matrix (ECM) with roles in
signaling, transmission of mechanical feedback to cells, and maintenance of cell structure
(Hynes et al, 2009). Epithelial cells have both basal and apical ECMs. Basal domains of
epithelia cells contain integrins, which link the cytoskeleton to the underlying ECM.
The basal ECM has a role in signaling, by binding and interacting with growth factors.
For example, the basal ECM regulates TGFβ-signaling (Hynes, 2009). TGFβ is bound in
inactive forms by basal ECM proteins, and is activated by the actions of other proteins in
the ECM. Also, structural abnormalities in basal ECM have been associated with a
number of disorders, including Polycystic Kidney Disease (Wilson et al., 2011).
Additionally, the basal ECM has been known to have roles in cell adhesion and migration
(Berrier and Yamada, 2007), but the understudied apical ECM may also have important
roles in these processes.
4

Because epithelial cells are exposed to a variety of environments as the border to
the outside of an organism, or the interior of an organ lumen, they require specialized
apical ECMs. Apical ECM is present in both planar and tubular epithelial cells. The
apical ECM has been demonstrated to have roles in epithelial cell shape, tube structure,
signaling, and pathogen defense. The function and molecular components of the apical
ECM varies widely in different tissues, but some features are common. Apical ECMs
contain both transmembrane and secreted proteins. They also commonly contain
collagen, glycoproteins, and polysaccharides. Apical ECMs are commonly modified
after components are placed, and are commonly re-formed during development. Below, I
will describe some model systems, and what is known about epithelial cells, apical ECM,
and junctions.
The apical ECM in Mammals
Mammals have apical ECM in a variety of tissues, such as the egg, eye, ear and
intestine. Surrounding the mouse oocyte, the zona pellucida consists of a meshwork of
three glycosylated proteins, ZP1, ZP2 and ZP3. These proteins are synthesized and
secreted by growing oocytes. In this context, the ECM serves to allow fertilization by
just one sperm (Wassarman and Litscher, 2009). The ZP domain, a common domain in
apical ECM proteins gets its name from the ZP proteins. In the mammalian inner ear,
apical ECM structure is called the tectoral membrane. It includes ZP domain
glycoproteins called tectorins, and aids in the transmission of sensory input (Goodyear
5

and Richardson, 2002). The apical ECM of the mammalian ocular surface contains
glycosylated mucins (Govindarajan and Gipson, 2010), which serve to protect the eye.
Endothelial blood vessels contain a glycocalax, a membrane associated matrix of protein
and sugars (Pries et al 2000). In vivo, capillaries contain a fibrillar material early in
development, which is later cleared from the lumen (Folkman,1980).
The mammalian gastrointestinal tract provides an example of an apical ECM with
a known role in pathogen protections. The apical ECM protects from pathogens by both
signaling and forming of a physical barrier between the lumen of the intestine and the
epithelial cells. The apical ECM of the gastrointestinal tract includes glycosylated
mucins, which block underlying epithelial cells from proteases, and obstruct colonization
by bacteria in the gastrointestinal tract. The apical ECM in the gastrointestinal tract also
prevents pathogen invasion through signaling. The apical ECM includes Toll-like
receptors (TRLs), Leucine-Rich Repeat domain proteins which detect pathogens and
trigger the innate immune response (Moncada et al., 2003). This apical ECM, therefore,
has pathogen protection and signaling roles.
In mammals, a link between ECM and epithelial junctions has been suggested.
MUC1 is a transmembrane mucin glycoprotein, associated with pancreatic ductal
carcinoma. In humans, overexpression or mislocalization of MUC1 or MUC4 are
associated with poor prognosis in breast, lung and pancreatic cancers (Kufe et al., 2009).
In mouse models, overexpression of MUC1 triggers the molecular steps of EMT,
including the repression of epithelial junction protein E-cadherin (Roy et al., 2001).
6

Correct regulation and localization of mucins seems to be required to maintain polarity
and junctional connections in epithelial cells (Kufe et al., 2009).
The apical ECM in Drosophila
Drosophila is a powerful model organism, and has therefore been a useful system
to study the apical ECM. The apical ECM initially surrounding the Drosophila embryo is
the eggshell, which functions to protect the egg from the outside environment. The
Drosophila eggshell components are secreted apically over the oocyte by the somatic
epithelial follicle cells. The eggshell consists of predicted chitin binding and mucin-like
proteins (Fakouri et al., 2006) in addition to other chorion proteins (Trougakos and
Margaritis, 1998). Genes that contribute to the eggshell are tightly regulated temporally,
and are localized to specific portions of the embryo. So called early, middle and late
chorion genes are expressed at distinct times between stages 9-14, producing a final
eggshell with five distinct layers (Margaritis et al., 1980). This tight regulation indicates
a carefully coordinated deposition of eggshell layers in the developing ECM (Cavalieri et
al., 2008; Tootle et al., 2011). Later in development, Drosophila produces a cuticle. The
Drosophila cuticle contains ZP (Uv and Moussian, 2010) proteins, chitin, and lipids
(Payre, 2004). The outer layer, called the epicuticle, contains lipids and glycoproteins.
Cuticle is secreted apically by epidermal cells. After initial deposition, the cuticle must
be modified, as evidenced by the requirement for modifying protein matrix
7

metalloprotease (Glasheen et al., 2010). Thus, the apical ECM surrounding the
Drosophila epidermis is re-formed and modified several times during development.
A link between ECM and cell shape has been noted in Drosophila epithelia. The
apical ECM affects the shape of epithelial cells that form trichomes (Fernandes et al.,
2010). Loss of ZP domain proteins results in the detachment of epidermal cells from the
cuticle (Fernandes et al., 2010). The proteins are required for epidermal adhesion to the
cuticle (Bokel et al., 2005), and have also been demonstrated to have a role in controlling
epithelial cell shape (Plaza et al., 2010).
A particularly well-studied apical ECM in an epithelial tube system is the
Drosophila trachea. The trachea is a complex structure consisting of both multicellular
and unicellular tubes (Lubarski and Krasnow, 2003; Schottenfeld et al., 2010).
Interactions between the cytoskeleton and basal ECM are necessary in the Drosophila
trachea. Mutations in talin and integrin, two proteins involved in linking the cytoskeleton
to the ECM, result in the lumen of branch cells becoming convoluted and breaking up in
larval stages (Levi et al., 2006). On the apical side of the trachea, chitin fibers are
secreted into the apical lumen in the early development of the trachea, and form a chitin
cable, which is required for proper expansion of the dorsal trunk lumen diameter (Devine
et al., 2005; Tonning et al., 2005). Once deposited, proper maintenance of this chitin
apical ECM is necessary; mutations in chitin modifying enzymes Serp and Verm result in
elongated lumen (Wang et al., 2006; Luschnig et al., 2006).
8

Links between ECM and junctional maintenance have been suggested in the
Drosophila trachea. Secretion of Serp and Verm into the lumen requires intact septate
junctions (Wang et al., 2006). Also, apically localized ZP proteins Piopio and Dumpy
contain transmembrane domains and are part of the apical ECM. In the absence of Piopio
and Dumpy, epithelial junctions are not maintained during the switch from intercellular
to autocellular junctions in tracheal cells (Jazwinska et al., 2003), suggesting a
connection between the apical ECM and junction stability. Despite these data, we still
have a limited understanding of how the apical ECM contributes to epithelial morphology
and junction dynamics. New models to study the connection between apical ECM and
junctions could shed light on these interactions.
Epithelial cells and apical ECM in Caenorhabditis elegans
Several convenient features of C. elegans can be exploited to study the conserved
junctions and the apical ECM of epithelial cells. The worm has a short generation time
and large brood size, and it therefore a powerful genetic system. Additionally, C. elegans
is amenable to transgenic arrays and RNAi, allowing genetic manipulation. The worm is
transparent, and many molecular markers are available, or can be generated, allowing
visualization of cells and proteins in the living organism. C. elegans is therefore a
potentially informative system to study epithelial junction and apical ECM formation and
maintenance. I will now review epithelial tissues in the worm, and what is known about
their junctions and apical ECMs.
9

The epidermis
The C. elegans epidermis consists of planar epithelial cells that surround the
worm. The basal side of the epithelial cells faces muscle or pseudoceolom; the apical
side faces the outside of the worm. The cells of the epidermis have conserved epithelial
junctions (Bossinger et al., 2001) and apically localized conserved PAR complex proteins
(Praitis et al., 2005). Also, ERM-1 and SMA-1/βH-spectrin cytoskeletal linkers, which
organize actin, are localized apically in these cells (van Furden et al., 2004; Gobel et al.,
2004; Praitis et al., 2005). Throughout the life of the worm, the apical ECM surrounding
the epidermis changes several times. At the 1.5-fold stage, the embryonic epidermis is
lined by a thin apical ECM, the embryonic sheath (Priess and Hirsh, 1986; Costa et al.,
1997). The sheath is visible by transmission electron microscopy (TEM), but the
molecular components of the sheath are unknown. Beyond the sheath, four additional
layers of ECM constitute the eggshell (Benenati et al., 2009; Rappleye et al., 1999). The
ECM layer closest to the sheath is the inner-eggshell layer, a sac-like structure that
surrounds the entire embryo. The inner-eggshell layer hugs the embryo closely, and is
therefore close to the sheath in most regions, but separates from the sheath where the
embryo bends inward. The components of the sheath, and the mechanism of the
connection between the sheath and inner eggshell layer are unknown.
In the 3-fold stage of development, epidermal cells produce a new apical ECM,
the cuticle. The cuticle serves to protect the worm, help the worm move, and control the
shape of the worm. The cuticle contains collagen and also is made of cuticlins, ZP
domain proteins that have a role in cuticle patterning (Sapio et al., 2005; Page and
10

Johnstone, 2007; Plaza et al., 2010). The cuticle is an apical ECM that is replaced four
times in the worm’s lifetime; a new cuticle is secreted with each larval molt. Mutations
in the cuticle proteins have revealed a role for this ECM in permeability, locomotion, and
worm shape (Page and Johnstone, 2007).
The alimentary tract
Several C. elegans organs contain tubular epithelia. The largest tubular organ
system in the worm is the alimentary tract, consisting of the pharynx, intestine, and rectal
cells. The intestinal epithelial cells, enterocytes, have actin protein ACT-5, ERM-1, and
SMA-1 (Gobel et al., 2004; Segbert et al., 2004; Praitis et al., 2005) localized apically,
and the intestine has conserved apical junction proteins like discs large complex and
cadherin/catenin complex, as well as PAR complex proteins (Bossinger et al., 2001). The
pharynx has an apical chitinous matrix, but the intestine is not lined by cuticle (Zhang et
al., 2005). Although many junctional and apical components are the same in the intestine
and epidermal cells, the striking difference between these tissues is in the extracellular
matrix. The epidermal apical ECM contains cuticle, and the intestine does not,
suggesting very different ECM components between these similar tissues.
11

The excretory system
Despite the progress made in multiple models, the relationship between conserved
epithelial junctions and apical ECM formation and maintenance needs to be better
understood in a simple system. The C. elegans excretory system is emerging as a useful
organ to study the basic properties of epithelial tube cells. This ‘primitive renal system’
forms a continuous lumen through three tandem unicellular tubes: the canal cell, the duct
cell, and the pore cell (Nelson et al., 1983; Nelson and Riddle, 1984). The advantage of
the excretory system is that it is a very simple system with multiple types of unicellular
epithelial tubes in adjacent cells. Two different processes of tube formation occur in the
excretory system. The duct and pore cells form by a wrapping around mechanism in
which the cells form autocellular junctions. In the duct cell, the cell fuses, and the
autocellular junctions dissolve (Stone et al., 2009). The canal cell lumen forms without
the formation of autocellular junctions (Buechner, 2002; Berry et al., 2003). At the 1.5-
fold stage of embryonic development, the unicellular tubes of the excretory system have
formed a continuous lumen connects through all three cells. The cells are tandemly
connected, and epithelial junctions are visible by TEM and with molecular markers
(Abdus-Saboor et al., 2011). After this initial stage of tube formation and junction
formation, the excretory system undergoes a process of elongation. Over the next several
hours, the cells undergo morphological changes, and need to maintain junctions as the
cells elongate until the 3-fold stage of embryonic development. Through these changes
the lumen diameter remains the same, but the lumen length increases. The taxing process
12

of elongation requires junctional maintenance and growth of the apical ECM, making the
excretory system an exciting system to study junctional and apical ECM dynamics.
The apical ECM of the excretory system is still mysterious, and varies among the
cells of the excretory system. At the 1.5-fold stage, a wispy apical ECM is visible by
TEM. Between the 1.5-fold and 3-fold stage, cuticle is deposited in the lumen of the duct
and pore. The canal cell, in contrast, does not have luminal cuticle. Proteins known to
have a role in luminal maintenance in the canal cell include ion channels (Berry et al.,
2003) and components of the cytoskeleton such as ERM-1 and SMA-1/β-H-spectrin
(Gobel et al., 2004; Buechner, 2002). Like the intestine, the canal cell is a tubular
epithelial structure with SMA-1 and ERM-1 apically localized, but the canal cell lumen is
formed by a single cell. The C. elegans excretory system has the potential to be a model
to study epithelial junctions and different types of apical ECM in a simple organ. For the
excretory system to be established as a model requires a better understanding of the duct
and pore cell, and many important questions need to be addressed. What proteins
contribute to the ECM? What is its role in luminal growth/maintenance? What is the
relationship of the ECM to junction dynamics? A better understanding of the role of the
ECM in the formation and maintenance of epithelial tubes in these tiny cells could
provide insight into the junctional dynamics and requirement of apical ECM in epithelial
cells.
13

Figure Legend
Figure 1.1. Epithelial cells have apical and basal sides; both can be surrounded by
an ECM. Planar (A) and tubular (B) epithelia are depicted. Cells are held together by
apical junctional proteins. The apical ECM consists of collagen, mucins and ZP proteins.
The basal ECM includes collagen, laminins, and intergrins.
14
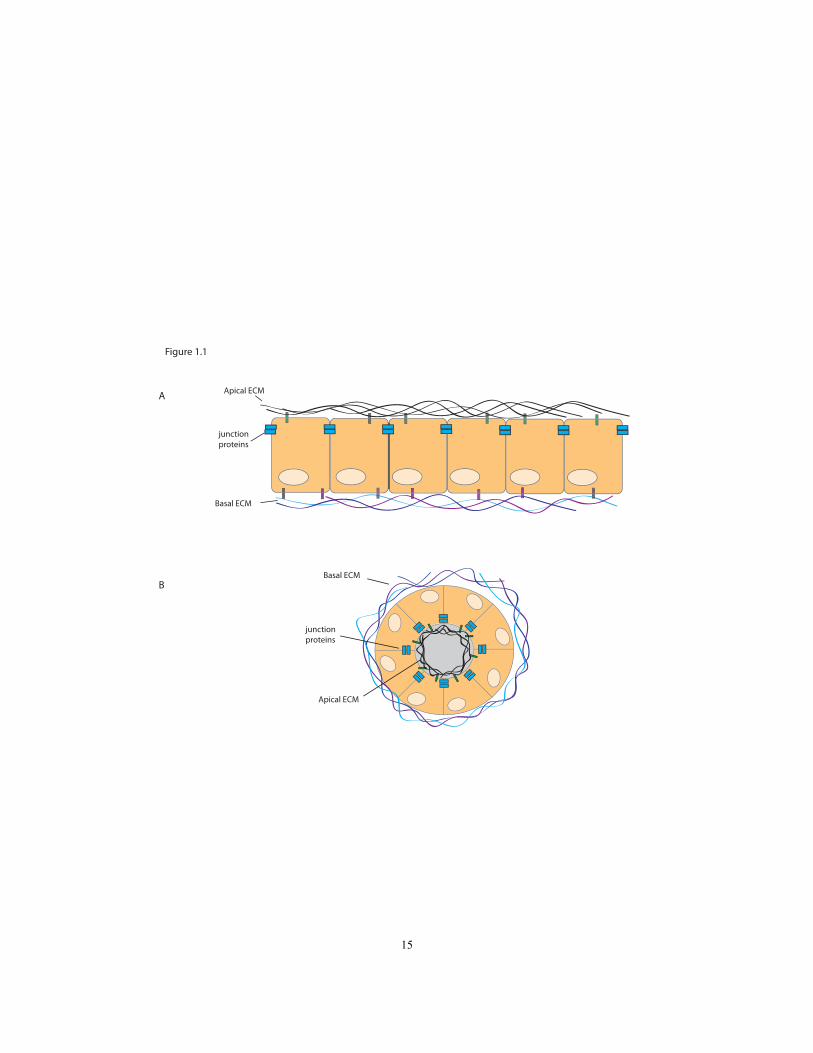
Figure 1.1
junction proteins
Apical ECM
Basal ECM
Basal ECM
Apical ECM
junction proteins
A
B
15

Chapter 2
Extracellular leucine-rich repeat proteins are required to organize the apical
extracellular matrix and maintain epithelial junction integrity in C. elegans
This chapter has been submitted as a manuscript:
Mancuso, V.P., Parry, J.M., Storer, L., Poggioli, C., Nguyen, K.C.Q., Hall, D.H. and
Sundaram, M.V. Extracellular leucine-rich repeat proteins are required to organize
the apical extracellular matrix and maintain epithelial junction integrity in C.
elegans
16

Roles of Authors
Chapter 2, as presented here, with some modifications, is a submitted manuscript.
The bulk of the work of the chapter is my let-4 analysis. In parallel to my work on let-4,
other members of the lab characterized the gene egg-6. The allele egg-6(cs67) was
identified in a screen for rod-like larval lethality, a phenotype indicative of an excretory
system defect (Craig Stone Thesis, 2008). Analysis of egg-6 revealed that the mutant had
similar defects to let-4, and was also in the eLLRon protein family. In order to present
my let-4 work in the larger context of eLLRon proteins, we incorporated our lab‟s work
on egg-6 into this chapter.
I performed the genetic analysis, fluorescent marker analysis, transmission
electron microscopy, deletion construct analysis, RNAi, permeability assay and
immunostaining for the let-4 analysis. Meera Sundaram positionally cloned let-
4(mn105), and performed some of the let-4 RNAi and TEM analysis. Ken Nguyen and
David Hall performed the sectioning and assisted in the TEM analysis. Luke Storer and
Corey Poggioli helped to generate the LET-4 deletion constructs and performed some
RNAi experiments. Jean Parry, with the assistance of Luke Storer, performed the
cloning, phenotypic characterization, genetic analysis, and marker analysis of EGG-6.
The work in the Appendices 1-3 is my work which supplements the main body of
Chapter 2.
17

Abstract
The specialized junctions that hold epithelial cells together are essential for organ
integrity yet often must be remodeled or disassembled during normal morphogenesis and
tissue turnover. Loss of epithelial junction integrity during epithelial-to-mesenchymal
transition (EMT) is a key feature of tumor metastasis, the major cause of cancer
morbidity. The factors that control junction stability and dynamics are poorly
understood, but include both cell-intrinsic and environmental cues. We identified a set of
extracellular leucine-rich repeat only (eLRRon) proteins in C. elegans (LET-4 and EGG-
6) that are expressed on the apical surfaces of epidermal cells and some tubular epithelia,
including the excretory duct and pore. A previously characterized paralog, SYM-1, is
also expressed in epidermal cells and secreted into the apical extracellular matrix (ECM).
Mutants lacking one or more of these eLRRon proteins show multiple defects in apical
ECM organization. Furthermore, epithelial junctions initially form in the correct
locations but then break at the time of collagen matrix secretion and remodeling of cell-
matrix interactions. This work identifies eLRRon proteins as important components and
organizers of the pre-cuticular and cuticular apical ECM, and adds to the small but
growing body of evidence linking the apical ECM to epithelial junction stability. We
propose that eLRRon-dependent apical ECM organization contributes to cell-cell
adhesion and may modulate epithelial junction dynamics in both normal and disease
situations.
18

Introduction
Polarized epithelial cells organize together to form many of the surfaces in our
bodies, including the outer epidermis and the lining of many internal tubular organs such
as the kidney, lung and gastrointestinal tract. Consequently, defects in epithelial
development or maintenance underlie a variety of human diseases (Marchiando et al.,
2010; Chamcheu et al., 2011; Wilson, 2011). Loss of epithelial character during
epithelial-to-mesenchymal transition (EMT) is a key feature of tumor metastasis, the
major cause of cancer morbidity (Kalluri and Weinberg, 2009; Polyak and Weinberg,
2009). Thus, it is important to understand how epithelial structures are formed and
maintained.
Epithelial cells are linked by specialized junctions that hold the tissue together,
create a paracellular barrier, and separate the cells' apical and basolateral surfaces (Shin et
al., 2006; Giepmans and van Ijzendoorn, 2009). Many junction components are
evolutionarily conserved, although junction organization differs somewhat among
organisms. In mammals, cadherin-based adherens junctions, which mediate cell-cell
adhesion, are located basally to claudin-based tight junctions, which form the paracellular
barrier and demarcate the apical and basolateral membrane surfaces. In Drosophila,
adherens junctions are located apically to claudin-based septate junctions. In
Caenorhabditis elegans, a single electron-dense structure, termed the “apical junction”,
contains adjacent adherens junction and septate-junction-like domains (Lynch and
Hardin, 2009). Initial junction assembly depends on conserved polarity regulators such as
the PAR, Crumbs and Scribble complexes (Goldstein and Macara, 2007). Once
19

assembled, epithelial junctions are dynamic structures that must be frequently
disassembled or remodeled during morphogenesis and tissue turnover (Acloque et al.,
2009; Baum and Georgiou, 2011; St Johnston and Sanson, 2011). The mechanisms that
control junction stability and dynamics are still poorly understood.
The basal and apical surfaces of epithelia contain different types of proteins and
lipids, and each surface secretes and interacts with different factors in the extracellular
matrix (ECM). Basal surfaces face towards the basement membrane and neighboring
tissues. In simple planar epithelia, apical surfaces face towards the outside of the body,
and in tubular epithelia apical surfaces face towards the lumen. Basal domains typically
contain integrins, which link the actin cytoskeleton to basement membrane components
such as laminins and collagens (Hynes, 2009). Apical domains contain other types of
transmembrane proteins, such as zona-pellucida (ZP)-domain proteins and mucins that
interact with or contribute to the apical ECM (Bafna et al., 2010; Plaza et al., 2010). It
has long been appreciated that the basal ECM influences epithelial cell polarity, cell
shape and cell motility (Berrier and Yamada, 2007). The apical ECM, in contrast, has
generally been viewed as a more passive protective barrier against pathogens and other
environmental toxins. However, there is increasing evidence that the apical ECM also
helps to shape epithelial cell morphology and can influence junction dynamics. For
example, in the Drosophila trachea, a temporary chitinous apical ECM controls tube
length (Devine et al., 2005; Tonning et al., 2005) and the ZP-domain proteins Piopio and
Dumpy influence junction remodeling (Jazwinska et al., 2003). In humans,
overexpression of the mucin MUC1 is observed in >90% of metastatic pancreatic ductal
adenocarcinoma, and MUC1 can influence EMT in mouse models (Kufe, 2009; Roy et
20

al., 2011). However, we still have a limited understanding of how the apical ECM
contributes to epithelial morphology and junction dynamics.
We use the C. elegans excretory (renal-like) system as a simple model for
epithelial tube development. The excretory system consists of three tandem unicellular
tubes: the large canal cell, which extends along the entire length of the body, and the
smaller duct and pore tube cells, which connect the canal cell to the outside environment
to allow for fluid waste excretion (Nelson et al., 1983; Nelson and Riddle, 1984) (Fig.
2.1A). Each unicellular tube has an intracellular apical or lumenal domain and an
extracellular basal domain, and the three tubes are connected in tandem via apico-lateral
junctions. All three tubes develop from initially non-epithelial precursors, but they are
morphologically distinct and form lumens via different processes. The canal cell forms a
lumen intracellularly at the site of the duct-canal cell junction, presumably through a
vesicular trafficking mechanism (Buechner, 2002; Berry et al., 2003). The duct and pore
tubes form by a wrapping mechanism in which the cells form autocellular junctions and
create an internal lumen from a previously external surface. The duct cell then auto-fuses
to dissolve its autocellular junction and become a seamless toroid, while the pore cell
retains its autocellular junction (Stone et al., 2009). After these initial steps of
tubulogenesis, all three tube cells elongate and undergo morphological changes to adopt
their unique sizes and shapes. Later in larval development, the original pore cell (G1)
withdraws from the organ to become a neuroblast, and is replaced by a second pore cell
(G2), which also forms a tube via wrapping (Sulston et al., 1983; Stone et al., 2009;
Abdus-Saboor et al., 2011). Thus, the excretory system is a simple model for studying
21

lumen development and maintenance and the dynamic control of epithelial junctions.
When searching for mutants that affect excretory duct and pore morphology, we
identified two leucine-rich repeat transmembrane proteins, LET-4 and EGG-6, that
localize to the apical domains of the duct, pore and epidermis. Here we show that LET-4,
EGG-6 and a paralog SYM-1 are important both to organize the apical ECM and to
maintain epithelial junction integrity.
22

Results
The apical ECM of the excretory duct and pore is contiguous with that of the
epidermis
The apical ECM and cytoskeleton differ significantly between different tube types
in the excretory system. The excretory canal cell resembles the C. elegans gut in that it is
not lined by cuticle, but has a specialized apical cytoskeleton containing the FERM
domain protein ERM-1 (Gobel et al., 2004; van Furden et al., 2004) (Fig. 2.1 B,C). It
also requires a set of specialized "exc" gene products for its lumenal maintenance
(Buechner et al, 1999, Buechner 2002). In contrast, the duct and pore do not appear to
express ERM-1 or most exc genes, but the mature duct and pore lumens are lined by a
collagenous cuticle that is contiguous with that of the epidermis (Nelson et al., 1983)
(Fig. 2.1A-E). However, the bulk of cuticle secretion does not occur until the latter part
of embryogenesis (Costa et al., 1997; Johnstone and Barry, 1996), after the duct and pore
have taken their mature shapes.
To examine the pre-cuticular duct and pore ECM, we analyzed existing
transmission electron micrographs (TEMs) of 1.5-fold embryos (Fig. 2.1F-J). At this
stage, the embryonic epidermis is lined by a thin apical ECM termed the "embryonic
sheath" (Priess and Hirsh, 1986), which later becomes an outer layer of the L1 cuticle
(Costa et al., 1997). Outside of the sheath, four additional ECM layers were visible that
together constitute the eggshell and are secreted by the embryo soon after fertilization
(Benenati et al., 2009; Rappleye et al., 1999). The inner-most of these layers was a sac-
23

like structure that encased the entire embryo; it was closely apposed to the sheath in most
regions, but separated from the sheath at points where the embryo bends inward,
including at the excretory pore opening (Fig. 2.1I). Within the nascent excretory pore
and duct lumen, a very thin lining of gray material was visible that may correspond to a
sheath-like ECM. The remainder of the duct and pore lumenal space, as well as the lumen
of the canal cell, was filled with fibrous electron-dense material (Fig. 2.1I, J). This
fibrous ECM material disappeared from the duct and pore by 5-6 hours later, at which
time the cuticle lining of the duct and pore had been secreted (Fig. 2.1D,E). In summary,
TEM analysis revealed the presence of two apical ECM layers within the duct and pore
prior to cuticle secretion; both of these layers are morphologically distinct from the
innermost layers of the epidermal ECM with which they are in contact.
let-4 and egg-6 encode transmembrane proteins with extracellular leucine-rich
repeats.
To identify genes important for excretory duct and pore development or
maintenance, we searched for mutants with defects in these epithelial tubes. let-
4(mn105) mutants previously were reported to have a rod-like lethal phenotype indicative
of excretory system defects (Meneely and Herman, 1979; Buechner et al., 1999), making
let-4 a candidate of interest. We isolated egg-6(cs67) in an EMS mutagenesis screen for
rod-like lethal mutants (Materials and Methods); a second allele, egg-6(ok1506), was
obtained from the C. elegans gene knockout consortium (Moerman and Barstead, 2008).
In both let-4 and egg-6 mutants, excretory junction morphology appeared initially
24

normal, but the pore autocellular junction disappeared shortly before or after hatching,
implicating these genes in the maintenance of junction integrity (see below).
let-4(mn105) is a recessive, loss-of-function mutation and caused highly, but not
completely, penetrant lethality (Fig. 2.2A). The majority of mutants died as early L1
larvae with excretory defects. A smaller percentage of mutants died as embryos; this
embryonic lethal phenotype was temperature sensitive. Approximately 2% of mutants
were "escapers" that survived to adulthood and were fertile, but exhibited defects in
locomotion and egg-laying. The progeny of these escaper homozygotes had the same rate
of lethality as progeny from heterozygous mothers, indicating that there was no maternal
effect on lethality (Fig. 2.2A).
egg-6(cs67) and egg-6(ok1506) are also recessive loss-of-function mutations and
both caused recessive, fully penetrant L1 lethality due to excretory defects (Fig. 2.2A).
Animals rescued for this zygotic lethality by an egg-6(+) transgene (see below) gave
100% dead embryos in the next generation, revealing a maternal egg-6 requirement.
Embryos lacking maternal egg-6 arrested at the ~40 cell stage and had fragile eggshells,
as also observed after egg-6 RNAi (Sonnichsen et al., 2005).
We positionally cloned let-4 and found that it corresponded to the gene sym-
5/C44H4.2, which encodes a predicted type I transmembrane protein with 14
extracellular leucine-rich repeat (LRR) domains and a short cytoplasmic tail (Fig.
2.2B,C). let-4(mn105) mutants had a C to T nucleotide change in the fourth exon of
C44H4.2, introducing a stop codon into the 11th LRR. A 5.3kb genomic fragment
encompassing C44H4.2 and no other genes rescued mn105 lethality. RNAi against
25

C44H4.2 also recapitulated some aspects of the let-4 phenotype (see below). Although
C44H4.2 has been previously called sym-5 (synthetic lethal with mec-8) based on genetic
interactions with the mec-8 splicing factor observed in RNAi experiments (Davies et al.,
1999), the let-4 gene name pre-dates those studies. Therefore, we refer to C44H4.2 as
LET-4.
We positionally cloned cs67 and found that it corresponded to the gene egg-
6/K07A12.2, which also encodes a predicted type I transmembrane protein with 14
extracellular LRR domains and a short cytoplasmic tail (Fig. 2.2B,C). egg-6 was
independently identified and named based on its eggshell-defective RNAi phenotype
(Andrew Singson and Karen Oegema, personal communication). cs67 mutants had a C
to T nucleotide change in the eighth exon of egg-6/K07A12.2, introducing a stop codon
into the extracellular domain. cs67 failed to complement egg-6(ok1506), which deletes
1678 bp of the coding region, completely eliminating the LRR domain. A 10.5kb
genomic fragment encompassing K07A12.2 and no other genes rescued cs67 and ok1506
zygotic lethality. Thus, we conclude that cs67 is an allele of egg-6, and that let-4 and
egg-6 encode related transmembrane proteins.
LET-4 and EGG-6 belong to the large family of extracellular LRR (eLRR)
proteins, which includes many proteins involved in cell adhesion, ECM interactions and
signaling. LET-4 and EGG-6 specifically belong to the "eLRR only" or "eLRRon"
subgroup (Dolan et al., 2007), since they contain no other recognizable domains. Mice
have 52 eLRRon proteins, including LRRTM1-3, which are involved in synaptic junction
formation or stabilization (Brose, 2009; de Wit et al., 2009; Ko et al., 2009; Linhoff et al.,
2009; Siddiqui et al., 2010) and the small leucine-rich proteoglycans (SLRPs), which
26

modulate collagen matrix assembly (Kalamajski and Oldberg, 2010); many others remain
uncharacterized. In addition to LET-4 and EGG-6, C. elegans has 15 other members of
the eLRRon family, including SYM-1, a secreted epithelial eLRRon protein that
functions redundantly with LET-4 in the epidermis (Davies et al., 1999). The LRR
domain of LET-4 is more similar to that of SYM-1 (53%) than to that of EGG-6 (49%) or
any other eLRRon protein.
LET-4::GFP and EGG-6::GFP localize to the apical (luminal) side of the duct, pore
and other external epithelia
To visualize the localization of LET-4 and EGG-6, we generated fusion proteins
by inserting GFP at the LET-4 or EGG-6 C-terminus within our genomic rescue
fragments. Both the LET-4::GFP and EGG-6::GFP fusion proteins rescued lethality of
the corresponding mutants, indicating that all required regulatory elements were included
in the transgenes and that the tagged proteins were functional (Fig. 2.2A,B).
LET-4::GFP and EGG-6::GFP were expressed in a subset of epithelial cells,
including epidermal, vulval and rectal cells and the excretory duct and pore (Figs. 2.3,
2.8). EGG-6::GFP was also observed in some neurons (Fig. 2.8). Expression began
around the ventral enclosure stage of embryogenesis and continued through larval
development, but then decreased in adulthood. Expression was notably absent from other
internal epithelial tissues such as the gut and pharyngeal tubes (Fig. 2.3C-F). LET-
4::GFP was transiently expressed in the excretory canal cell at the 1.5-fold stage (Fig.
2.3C), but no longer visible in this cell by hatch. Notably, with the exception of the canal
27

cell, the epithelia that expressed LET-4 and EGG-6 were those that would eventually
become cuticle-lined.
In almost all epithelia where they were expressed, LET-4::GFP and EGG-6::GFP
appeared strongly apically enriched (Figs. 2.3, 2.8). In the excretory duct and pore, LET-
4::GFP and EGG-6::GFP lined the luminal membrane (Fig 2.3A-B). In the epidermis,
both fusions were distributed across the apical surfaces of most dorsal and ventral
epidermal cells but were observed more weakly or variably in the lateral (seam)
epidermis (Fig. 2.3G-H). Neither fusion was strongly enriched at apical junctions based
on co-visualization with DLG-1/Discs Large::mcherry (Fig. 2.3 B,D,F,G,H). Both fusions
partially overlapped with but did not strongly co-localize with transepidermal
intermediate filaments (IFs) at hemidesmosomes, based on co-staining with the IF
antibody MH4 (Fig. 2.8 and data not shown). Both fusions were present in many large
puncta, potentially representing a vesicular compartment trafficking to or from the
membrane. In summary, LET-4 and EGG-6 topology and apical localization suggest a
configuration in which the LRR domains extend into the apical ECM, but localization is
not limited to known sites of epidermal-apical ECM attachments.
Interestingly, the one exception to the apical localization of LET-4::GFP was the
excretory canal cell. At the 1.5-fold stage, when LET-4::GFP was transiently expressed
in the canal cell, LET-4::GFP localized uniformly around the plasma membrane and not
to the developing internal lumen (Fig. 2.3 C,C‟). While the significance of this
expression is unclear, we speculate that the unique localization pattern reflects molecular
28

differences in the apical domain of the canal cell vs. the apical domains of other LET-
4::GFP expressing cells.
let-4 and egg-6 are required to maintain junction and lumen integrity in the
excretory duct and pore
The majority of let-4(mn105) mutants and all egg-6(cs67) or egg-6(ok1506)
mutants arrested as L1 larvae with excretory defects (Fig. 2.2A). The overall morphology
and junctional pattern of the excretory system appeared initially normal in mutant 3-fold
embryos, but became detectably abnormal shortly prior to hatch (Fig. 2.4). The first
detectable abnormality was a swelling of the canal cell lumen in the region proximal to
the canal-duct junction (Fig. 2.4C-E). Subsequently, the duct and pore cells separated
from each other and the pore autocellular junction disappeared (Fig. 2.4F-H). Remants of
junction material sometimes remained at the separation points, suggesting junction
breakage. The duct-canal junction always remained intact, and the duct lumen often
swelled considerably. Canal lumen swelling was a secondary consequence of defects in
the duct and pore, since the excretory phenotype was rescued by lpr-1p-driven LET-4(+)
or EGG-6(+) transgenes expressed specifically in the duct, pore and epidermal cells but
not in the canal cell (Fig. 2.2A). Neither let-4 nor egg-6 was rescued by dpy-7p-driven
transgenes expressed in the pore and epidermal cells. Thus, let-4 and egg-6 are required
in the excretory duct and likely also in the pore, but not in the canal cell, a requirement
that fits with the expression patterns described above. Within the duct and pore, let-4 and
egg-6 are required for both luminal and junction maintenance.
29

To confirm these interpretations for let-4 mutants, and to visualize the narrow
lumen of the canal cell, duct cell and pore cell directly, we performed transmission
electron microscopy (TEM) of successive serial thin sections. We analyzed five let-4
embryos at the 1.5-fold stage, just after initial tubulogenesis, and confirmed that the
lumen had a generally normal shape and was continuous through all three cells (Fig. 2.4I
and Fig Ap3.1). Fibrous ECM material was visible in the lumen as in wild-type (Fig.
2.4I). We analyzed three wild-type and nine let-4 mutant embryos at the late 3-fold stage,
surrounding the window when the duct and pore have taken their mature shape and
defects first become visible by light microscopy (For details of these results, see
Appendix 3, Figures Ap3.2 and Ap3.3). In 5/9 let-4 3-fold embryos, all three tube cells
were still properly connected and the lumen was continuous as in wild-type, with no
apparent distortions. Intercellular apical junctions appeared normal, as did the cuticular
lining of the duct and pore (Fig. 2.4J). Because 97% of let-4 embryos eventually display
excretory defects, we infer that these embryos would have displayed defects shortly
thereafter, had they been allowed to mature. The absence of any detectable junction or
luminal defect in these embryos indicates that initial steps of junction formation, lumen
growth and cuticle secretion are fairly normal in let-4 mutants
In 4/9 let-4 3-fold embryos, the canal and duct tubes remained connected but the
duct and pore appeared to have separated, as we had also observed by confocal
microscopy. In one of these embryos, the existing duct lumen and the canal lumen
appeared normal. In another, the duct lumen appeared normal, but the canal lumen was
greatly enlarged. In the remaining two embryos, the duct lumen diameter was enlarged
30

proximal to the duct cell body, and the apical membrane in this region had separated from
the cuticle lining (Fig. 2.4K). Because the cuticle ring had a normal diameter in these
cases, we infer that lumen distortion occurred subsequent to cuticle secretion. In two
embryos, we were able to trace the duct lumen to its premature termination within the
duct process (Fig. 2.4L). We were unable to recognize the excretory pore cell in three of
these embryos, suggesting that the pore lacked its characteristic autocellular junction and
lumen. Our interpretation is that duct and pore separation leads to pore collapse and
lumen retraction, and that duct and canal cell lumen swelling behind the break is a
secondary consequence of excretory fluid backup. Thus the primary junctional defect in
let-4 mutants appears to be a failure to maintain the duct-pore intercellular junction.
Paralogs let-4 and sym-1 function redundantly to maintain epidermal junction
integrity during embryonic elongation
A small proportion of let-4 mutant or let-4 RNAi embryos ruptured during
embryonic elongation and failed to hatch (Figs. 2.2A, 2.5A). This phenotype reflected a
semi-redundant role of let-4 and its closest paralog, sym-1. Like LET-4, SYM-1 also is
expressed in epidermal cells, but unlike LET-4, SYM-1 lacks a transmembrane domain
and is secreted into the apical ECM (Davies et al., 1999). Whereas essentially all sym-1
embryos developed normally and hatched, ~100% of sym-1; let-4(RNAi) embryos
ruptured (Fig. 2.5A). The rare embryos that did not rupture swelled abnormally as they
approached hatch, suggesting a defect in osmotic integrity (Fig. 2.5B). Similar osmotic
defects were seen in mec-8; let-4(RNAi) embryos (Fig. 2.5A), the basis for the alternative
let-4 name sym-5 (Davies et al., 1999).
31

Epidermal rupture during embryonic elongation can be caused by excessive actin-
myosin contractile activity, which provides the force for elongation (Wissmann et al.,
1997; Wissmann et al., 1999; Piekny et al., 2000; Piekny et al., 2003), or by defects in
structural components of the epidermal junctions (Costa et al., 1998; Totong et al., 2007).
In sym-1; let-4(RNAi) embryos, AJM-1::GFP and HMR-1/cadherin::GFP both appeared
normally localized prior to significant elongation and rupture (Fig. 2.5E,K), suggesting
that junction organization had been established properly. Junctions did not appear
distorted during the early steps of elongation as in known mutants with increased
contractile activity (Diogon et al., 2007). Furthermore, in temporal analyses, most
embryos managed to elongate to the 3-fold stage before rupturing and retracting to a
shorter length (Fig. 2.5F-N). Thus, as for the excretory system, we found no evidence for
defects in junction establishment. Rather, LET-4 and SYM-1 are required to prevent
epidermal junction breaks during the latter part of embryogenesis. Notably, this is the
time frame when cuticle secretion begins and epidermal-ECM interactions must be
remodeled (Costa et al., 1997).
The LET-4 transmembrane and cytoplasmic domains are dispensable for function
Some eLRRon proteins, including SYM-1, lack a transmembrane domain, and are
secreted into the apical ECM (Davies et al., 1999). Furthermore, although EGG-6 has a
predicted transmembrane domain, some proportion of EGG-6::GFP was still secreted , as
it accumulated between the embryo and eggshell (Fig. 2.8). To ask if LET-4 must be
32

tethered to the membrane and to identify domains important for its function, we deleted
the transmembrane (TM), cytoplasmic (Cterm) or extracellular LRR domains in the
context of an lpr-1p::LET-4 transgene construct tagged with GFP to visualize localization
within the excretory duct (Fig. 2.6). LET-4(ΔLRR) failed to rescue let-4 lethality;
furthermore, the fusion protein was not enriched at the apical face of the excretory duct
and other epithelial cells (Fig. 2.6 B,D). In contrast, LET-4(ΔCterm) efficiently rescued
let-4 lethality and appeared properly localized (Fig. 2.6 C,D). LET-4(ΔTM) appeared
toxic to embryos and we were able to obtain only a few transgenic lines with very low,
undetectable levels of expression. LET-4(ΔTM) transgenes were apparently expressed,
however, since they partially rescued let-4 larval lethality (Fig. 2.6D). We conclude that
the LRR domains are required for proper LET-4 function and localization, whereas the
cytoplasmic domain is dispensable, and that tethering of LET-4 to the membrane is not
absolutely required for function.
let-4 and egg-6 are required for proper apical ECM organization
The above studies suggested that LET-4, SYM-1 and EGG-6 all might function
extracellularly as part of the apical ECM. Although the excretory duct and pore lumen
ECM appeared morphologically indistinguishable from wild-type in let-4 mutants,
several abnormalities in the epidermal apical ECM were observed in let-4 and egg-6
mutants. First, TEM analysis revealed that, although all apical ECM layers were present,
the inner eggshell layer was more widely separated from the epidermal embryonic sheath
33

layer in 5/5 let-4 embryos at the 1.5-fold stage (Fig. 2.7A), suggesting a problem within
one or both of these layers. Second, in most let-4 and egg-6 mutant 3-fold embryos,
many globular structures accumulated between the embryo and the eggshell (Fig. 2.7B-
D). These globules contained cytoplasm, since they were marked by GFP in transgenic
embryos expressing cytoplasmic GFP reporters (Fig. 2.7B). In let-4 TEMs, these
cytoplasts appeared to be membrane-bound and were positioned between the nascent
cuticle and the inner eggshell layer (Fig. 2.7D), indicating that cell fragments had been
shed prior to cuticle secretion. This again suggest a defect in the embryonic sheath.
Third, most let-4 and egg-6 mutant L1 larvae showed abnormal permeability to dye (Fig.
2.7E-J), indicating a defect in larval cuticle organization. This defect was also observed
in sym-1 mutants (Fig. 2.7I, J). A requirement for eLRRon proteins in apical ECM
organization is further supported by the eggshell defects observed after depletion of
maternal egg-6 (Sonnichsen et al., 2005) (A. Singson and K. Oegema, personal
communication). Together, these observations suggest that let-4, egg-6 and sym-1 are
required to organize the apical ECM. We propose that defects in apical ECM lead
secondarily to defects in epithelial junction maintenance.
34

Discussion
It has long been recognized that extracellular cues from contact with neighboring
cells and the ECM influence epithelial polarity, cell shape, and motility (Bryant and
Mostov, 2008). Epithelial cells also secrete their own ECM factors. While most studies
have focused on the importance of basal ECM factors, the work presented here suggests a
link between the apical ECM and maintenance of epithelial junction integrity in C.
elegans. This work also identifies extracellular LRR proteins as important components
and organizers of the pre-cuticular and cuticular apical ECM.
eLRRon proteins and the extracellular matrix
The eLRRon family of proteins includes 52 members in mice, 35 in Drosophila
and 17 in C. elegans (Dolan et al., 2007) several of which are involved in ECM
organization. In mammals, decorin and other secreted SLRPs bind directly to collagen
and modulate collagen fibril assembly (Kalamajski and Oldberg, 2010). Although many
SLRPs appear confined to stromal tissues, several are expressed in the kidney or other
epithelia (Ross et al., 2003; Shimizu-Hirota et al., 2004). SLRP knockout mice have
disorganized collagen fibrils and various tissue fragility phenotypes, and mutations in
certain SLRPs are associated with similar syndromic conditions in humans (Ameye and
Young, 2002; Schaefer and Iozzo, 2008). In Drosophila, the eLRRon protein Convoluted
is required for proper tracheal ECM organization and tube length (Swanson et al., 2009).
Several transmembrane eLRRon proteins, including mammalian LRRTM1-3 and
35

Drosophila Capricious and Tartan, are involved in neuronal cell adhesion and synaptic
junction formation or maintenance (Shishido et al., 1998; Taniguchi et al., 2000; Shinza-
Kameda et al., 2006; Kurusu et al., 2008); these junction phenotypes have not (thus far)
been linked to any additional ECM defect. We have shown here that transmembrane
eLRRon proteins LET-4 and EGG-6 and their secreted paralog SYM-1 are required for
both apical ECM organization and epithelial junction stability in the C. elegans epidermis
and excretory duct and pore tubes.
Like many other invertebrates, C. elegans has a tough outer exoskeleton or cuticle
that lines the epidermis and other exposed epithelia including the excretory duct and pore.
The mature cuticle consists primarily of collagens and ZP-domain proteins termed
cuticulins, and is coated by a lipid-rich epicuticle and a glycoprotein-rich surface coat
(Page and Johnstone, 2007). The mature cuticle forms relatively late in embryogenesis;
prior to that, the lipid- and glycoprotein-rich outer layers appear to comprise the early
embryonic sheath ECM and are in direct contact with the epidermis at microfilament-
based attachment sites (Priess and Hirsch, 1986; Costa et al., 1997). When the inner
cuticle layers are secreted, the earlier sheath layers detach from the epidermis and are
pushed outward, so the epidermis is subjected to a changing matrix environment. The
cuticle is subsequently shed and re-synthesized at each larval molt, so membrane-matrix
attachments must be constantly remodeled during development.
eLRRon proteins LET-4, SYM-1 and EGG-6 all localize to the apical domains of
epithelia that are or will eventually become cuticle-lined, and they are important for the
proper organization of both the pre-cuticular and cuticular apical ECM. eLRRon proteins
36

could play several possible roles in ECM organization. They could be functioning as
structural components of the ECM that contribute to the strength and impermeability of
the matrix. The ability of both LET-4 and SYM-1 to function in the absence of a
transmembrane domain is consistent with this possibility. eLRRon proteins could also
modify or modulate associations among other matrix components such as collagens, as
proposed for the mammalian SLRPs (Kalamajski and Oldberg 2010). Alternatively, or in
addition, eLRRon proteins could affect protein trafficking mechanisms that deliver other
ECM components to the apical surface. Although some pre-cuticular ECM material and
cuticle are still secreted in let-4 mutants as seen by TEM, we have limited knowledge of
the molecular constituents of the ECM and cannot exclude the possibility that some
specific constituents are missing.
Apical ECM organization and epithelial junction stability
A link between apical ECM organization and apical junction stability is suggested
by the concomitant presence of both types of defects in eLRRon mutants. eLRRon
proteins could play independent roles in both processes, or defects in one process may
lead secondarily to defects in the other. For example, in Drosophila, defects in septate
junctions prevent apical secretion of chitin modifying enzymes, disrupting apical ECM
organization (Wang et al., 2006; Luschnig et al., 2006). However, the initially normal
morphology of junctions in eLRRon mutants, the broad apical localization patterns of the
proteins, and the ability of LET-4 and SYM-1 to function in the absence of
37

transmembrane domains are difficult to reconcile with direct roles in junction
organization. Furthermore, egg-6 eggshell defects arise prior to formation of epithelial
junctions, and most let-4 and egg-6 single mutants have defects in epidermal ECM
organization that are not accompanied by defects in epidermal junction maintenance.
Finally, junction breaks in the epidermis (in sym-1 let-4 RNAi mutants) and in the
excretory system (in let-4 or egg-6 single mutants) occur relatively late, after ECM
defects are already apparent. Therefore, we favor a direct role for eLRRon proteins in
ECM organization, with secondary effects on junction integrity.
Mutations in other specific apical ECM components generally do not cause
excretory or epidermal junction phenotypes such as those described here, suggesting a
relatively specific role for eLRRon proteins in junction integrity. Instead, mutations in
individual cuticle collagen or cuticulin genes cause cuticle blistering or defects in body
shape or cuticle patterning (Johnstone, 2000; Page and Johnstone, 2007). Mutations in
glycosyltransferases that perturb the outer cuticle layer alter the susceptibility of larvae to
bacterial infection and only in some cases increase permeability (Partridge et al., 2008).
Nevertheless, there have been prior indications that the apical ECM influences junctional
size and stability. Embryos mutant for the cuticle collagen sqt-3 elongate initially and
then retract, revealing a requirement for cuticle to stabilize epidermal cell shape (Priess
and Hirsh, 1986). Several mutations that perturb molting also affect epidermal integrity
(Moribe et al., 2004; Fritz and Behm, 2009). Finally, sec-23 mutations that impair cuticle
secretion (and presumbably secretion of eLRRon proteins as well) cause embryonic
38

rupturing at the 2-3-fold stage as described here for sym-1; let-4(RNAi) embryos (Roberts
et al., 2003).
There are several mechanisms by which apical ECM organization might affect
junction integrity. Junction breakage in eLRRon mutants could result from increased
forces placed on those junctions. The interconnected nature of the apical ECM may help
bind together epithelial cells that share that ECM, reducing stress on individual junctions.
Indeed, the embryonic sheath originally was proposed to distribute circumferential actin-
myosin contractile forces across the embryo (Priess and Hirsh, 1986). Thus,
abnormalities in apical ECM organization may lead to uneven distribution of those
pulling forces, leading to junction breaks. Alternatively, junction breakage could reflect
inherent weaknesses in epithelial junctions. The interconnected nature of the apical ECM
may help bind together epithelial cells that share that ECM. The apical ECM also is in a
good position to interact with cadherins or with transmembrane apical polarity proteins
such as Crumbs, and could potentially influence polarity and junction maintenance
through such interactions. Finally, many ECM components, including SLRPs, affect
signaling pathways that could alter gene expression and/or cytoskeletal organization to
influence polarity and junction maintenance (Bulow and Hobert, 2006; Schaefer and
Iozzo, 2008). In the C. elegans excretory system, EGF-Ras signaling promotes multiple
aspects of duct development, including lumen and junction maintenance (Abdus-Saboor
et al., 2011); it will be interesting to test if LET-4 or EGG-6 influence EGF signaling.
This study adds to the growing body of evidence for links between eLRRon
proteins and ECM organization and for links between the apical ECM and epithelial
39

junction stability. We hypothesize that cell-type specific modulation of eLRRon
expression or activity could be a general strategy for junction remodeling during
development. The programmed EMT-like withdrawal of the excretory pore provides an
attractive model system for testing this idea.
40

Materials and Methods
Strains and alleles
Strains were grown at 20˚ C and maintained under standard conditions (Brenner,
1974) unless otherwise noted in this chapter, and all work in this thesis. Bristol strain N2
was used as wild-type. Alleles used were: I: mec-8(u218) (Chalfie and Au, 1989). X:
let-4(mn105) (Meneely and Herman, 1979), unc-3(e151) (Hodgkin, 1997), sym-1(mn601)
(Davies et al., 1999). egg-6(cs67) was obtained after standard EMS mutagenesis of N2
(Brenner, 1974). Transgenes used were: csEx146 (lin-48p:mCherry) (Abdus-Saboor et
al., 2011), fgEx11 (ERM-1::GFP) (Gobel et al, 2004), jcIs1 (AJM-1::GFP) (Koppen et
al., 2001), mcIs46 (DLG-1::RFP) (Diogon et al., 2007), qnEx59 (dct-5p::mCherry)
(Abdus-Saboor et al., 2011), saIs14 (lin-48p::GFP) (Johnson et al., 2001), xnIs96 (HMR-
1::GFP) (Achilleos et al., 2010), vha-1p::GFP (Oka et al., 1997).
Molecular analysis
let-4 had been previously mapped to the right arm of the X chromosome
(Meneely and Herman, 1979). cs67 was mapped to chromosome I by linkage analysis
and deficiency mapping (see Wormbase). Both genes were subsequently identified via
transgenic rescue experiments. Gene structures were confirmed by sequencing
41

C44H4.2/let-4 cDNA clones yk8g5, yk134h6, yk1661a04, yk1708a10 and
K07A12.2/egg-6 cDNA clones yk117f12 and yk4a1.
pVM3(LET-4 genomic region) was generated by inserting the let-4 genomic
region [NsiI-XbaI (chrX:14575767-14581099)] from fosmid WRM0620cC02 into a
pBlueScript vector.
To generate pVM4(LET-4::GFP), GFP obtained from pPD103.87 (Addgene) with
a BamHI digest was inserted into the native BamHI site in the final exon of LET-4 in the
pVM3 vector.
To generate pVM7(let-4p::GFP) (described in Appendix 2) required an
intermediate step to generate a fragment with the desired restriction sites. First, the 2.2kb
let-4 promoter region was cut out of a genomic fragment with HindIII and AccI. This
fragment contained the upstream region, and the first 6 amino acids of LET-4. This
fragment was then ligated into the HindIII and AccI sites of pBlueScript. In the
BlueScript vector, an XhoI site was immediately following the AccI site. The new vector
(called pVM6) was digested with HindIII and XhoI. This fragment contained the LET-4
upstream region, the first 6 amino acids of LET-4, including the AccI site, followed by a
cut XhoI site. The fragment was then ligated in-frame between the HindIII and SalI sites
of pPD121.89, which contains GFP. (Addgene)
To generate pVM9(dpy-7p::let-4cDNA), let-4 cDNA was cut out of pVM9 with
NheI and KpnI, and ligated into pKH11, which contains 216bp of the dpy-7 promoter.
42

dpy-7p drives expression in the pore cell and epidermis (Gilleard et al., 1997; Stone et al.,
2009).
To generate pVM16(lpr-1p::let-4cDNA), let-4 cDNA was cut out of pVM8
(which contains let-4 coding region from yk134h6) with NheI and KpnI, and ligated into
pBG12. lpr-1p drives expression in the duct cell, pore cell, and epidermis (Stone et al.,
2009).
To generate pVM20(lpr-1p::ssGFP::LET-4(ΔLRR)), a truncated form of let-
4cDNA was amplified off of pVM16 using primers which amplify the region of LET-4
after the LRR domain (amino acids 398-773). This fragment was then inserted into
pVM16 between the NheI and KpnI sites (This plasmid was called pVM19). PCR was
then performed to amplify ssGFP from pPD95.85 and insert NheI sites on either side.
This GFP fragment was then inserted into the NheI site of pVM19.
To generate pVM21(lpr-1p::LET-4(ΔC-term)::GFP), a truncated form of let-4
cDNA was amplified off of pVM16 using primers which amplify the region of LET-4
from the Start codon to 2 amino acids past the transmembrane domain (amino acids 1-
713). This fragment was then inserted into pVM16 between the NheI and KpnI sites
(This plasmid was called pVM18). PCR was then performed to amplify GFP from
pPD95.75 (Addgene) and insert KpnI sites on either side. The GFP fragment was then
inserted into the KpnI site of pVM18.
To generate pVM25(lpr-1p::LET-4(ΔTM)::GFP), a truncated form of let-4 cDNA
was amplififed off of pVM16 using primers which amplify the region of LET-4 from the
43

Start codon to just before the transmembrane domain (amino acids 1-688). This fragment
was then inserted into pVM16 between the NheI and KpnI sites (This plasmid was called
pVM17). PCR was then performed to amplify GFP from pPD95.75 (Addgene) and insert
KpnI sites on either side. The GFP fragment was then inserted nito the KpnI site of
pVM17.
The egg-6 genomic rescue fragment was cloned from fosmid WRM0617dE11.
GFP obtained from pPD103.87 (Addgene) was inserted into an engineered Nhe1 site to
generate EGG-6::GFP reporter pMS204. Transgenic lines were generated by co-
injecting each construct (2ng/ul) with pRF4 (98ng/uL).
For egg-6 tissue-specific promoter constructs, cDNAs obtained from yk117f12
was cloned into vectors pBG12 (lpr-1p), pKH11 (dpy-7p) or pHS4 (lin-48p), which are
all derivatives of pPD49.26 (Addgene). lin-48p drives expression in the duct cell
(Johnson et al., 2001). Transgenic lines were obtained by coinjecting each construct at 2-
10 ng/ul with either pHS4 (lin-48p::mCherry) at 2.5 ng/ul or pIM175 (unc-119::GFP) at
90ng/uL.
44

RNAi.
let-4 double stranded RNA (dsRNA) was synthesized using the Megascript RNAi Kit
(Ambion), using as template a fragment of the let-4 cDNA corresponding to exons 8-10
(Primers: oMS199
5‟GTAATACGACTCACTATAGGGCAGTCGTGAAGATGAGATTCGC3‟ and
oMS200
5‟GTAATACGACTCACTATAGGGCGCAATAACTGGATCCAGGATTG3‟). dsRNA
was injected into gravid hermaphrodites and embryos laid >16 hrs later were assessed for
phenotypes.
Immunostaining
L4 larvae were collected and fixed described in Appendix 4. Primary antibody
concentrations used were: MH4 (1:50) (Francis and Waterston, 1991); goat anti-GFP
(1:50) (Rockland Immunochemicals). Secondary antibodies: Cy3 donkey anti-mouse
(1:200); FITC donkey anti-goat (1:20) (Jackson Immunoresearch).
Microscopy
Images were captured by differential interference contract (DIC) and
epifluorescence using a Zeiss Axioskop (Jena, Germany) microscope with a Hamamatsu
Chilled CCD camera (Hamamatsu City, Japan). Confocal Mucroscopy was performed
45

with a Leica TCS CP (Wetzlar, Germany). All confocal images were analyzed with Leica
Confocal Software and ImageJ Software.
For TEM, 1.5-fold or late 3-fold embryos from wild-type or let-4(mn105) mothers
were fixed by high pressure freezing followed by freeze substitution (Weimer, 2006),
embedded in Eponate resin and cut into serial thin sections between 50 and 100 nm each.
Sections were observed on a Philips CM10 transmission electron microscope
(Amsterdam, The Netherlands) or a FEI-Tecnai T12. Serial section TEM images of a
similarly processed 1.5-fold embryo, N2E6B, were provided to the Center of C. elegans
Anatomy by Shai Shaham (Rockefeller Univ.).
Hoechst permeability assay
L1 larvae of each genotype were collected in M9 and incubated in 2μg/mL Hoescht dye
33258 (Sigma) for 15 minutes at room temperature, then washed twice with M9.
46

Acknowledgements
We thank the C. elegans knockout consortium, the Caenorhabditis Genetics Center
(University of Minnesota), Verena Gobel, Jeff Hardin, Michel Labouesse and Jeremy
Nance for providing strains, Shai Shaham for providing archival TEM images, Juan
Jimenez and Leslie Gunther for help in conducting HPF fixations, Amin Ghabrial and
Ishmail Abdus-Saboor for helpful comments on the manuscript, and Andrew Singson and
Karen Oegema for sharing results prior to publication. Electron microscopy was
performed in the EMRL at the University of Pennsylvania, supported in part by NIH
grant 5 P30 CA 016520. MH4 antibody was obtained from the Developmental Studies
Hybridoma Bank (University of Iowa) developed under the auspices of the NICHD. This
research was supported by NIHGM58540 to M.V.S. and NIHRR12596 to D.H.H.
V.P.M. was supported in part by T32 GM008216 and J.M.P. was supported in part by
T32 HD007516.
47

Figure Legends
Figure 2.1. The apical ECM differs between excretory tube types
(A,B) Schematics of the late 3-fold or early L1 excretory system. (A) Lateral view.
(B) Cross sections. Cuticle lines the duct and pore lumen. In all schematics, canal cell is
red, duct cell is yellow, and pore cell is blue. Green indicates the embryonic sheath,
which at this stage makes up the outer layer of the cuticle. Dark lines and circles indicate
apical junctions; arrowhead, duct-canal junction; bracket, duct cell body; arrow, pore
autocellular junction. (C) Excretory system of early L1 larva. ERM-1::GFP lines the
lumen of the canal cell (c), and is absent from the pore (p) and duct (d) cells (labeled with
dct-5p::mCherry). Anterior is to the left and ventral down in all figures unless otherwise
noted. (D-E) TEM of 3-fold embryo showing the duct cell lumen (lu) (D) and pore cell
lumen (E). In (E) the cuticle reaches from the outside of the worm into the lumen of the
pore cell. (F-H) Schematics of the 1.5-fold excretory system. (F) Whole embryo and
eggshell layers. (G) Lateral view. (H) Cross sections. The inner eggshell layer (orange)
surrounds the embryo. A thin sheath-like layer (green) lines the pore and duct lumen.
Cuticle has not yet been secreted, and the lumen of all three tube cells contains a fibrous
electron-dense material (ECM).
(I-J) TEM of 1.75-fold embryo, showing the inner eggshell layer (orange arrows) and
embryonic sheath layer (green arrows). Fibrous electron dense material (lines) is visible
in the duct and pore cells (Iiii) as well as the canal cell (J). Embryo “N2E6B” TEM (I-J)
kindly provided by Shai Shaham (Rockefeller U.).
48

Figure 2.2. let-4 and egg-6 encode related transmembrane proteins with
extracellular leucine-rich repeats.
(A) Genetic analyses and transgenic rescue experiments. For transgene rescue
experiments, progeny were collected from let-4(mn105) homozygotes carrying csEx173
(C44H4 cosmid +sur-5p::GFP) and a separately marked transgene of interest, or from
egg-6(ok1506)/hT2[qIs48] heterozygotes carrying a marked transgene of interest.
Rescue was determined by scoring each hatched worm for L1 lethality. (B) let-4 and egg-
6 genomic regions and rescue fragments. Note that paralog sym-1 is immediately
adjacent to let-4. (C) Predicted protein structures and mutant lesions. The LET-4 and
EGG-6 LRR domains share 49% similarity. Data contributed by VM, JP, MS and LS.
Figure 2.3. LET-4::GFP and EGG-6::GFP localize to the apical domains of the
excretory duct, pore and epidermal cells
(A,B) In L1 larvae, LET-4::GFP and EGG-6::GFP localize apically within the excretory
duct and pore. Symbols as in Fig. 1. (A) LET-4::GFP. (A‟) lin-48p::mCherry labels the
duct cell cytoplasm. (A'') Overlay. (B) EGG-6::GFP. (B‟) DLG-1::mCherry marks apical
junctions of the duct and pore cells. (B") Overlay. (C-H) In 1.5-fold (C,E,G,H) and 3-fold
(D,F) stage embryos, LET-4::GFP (C,D,G) and EGG-6::GFP (E,F,H) localize apically
within the epidermis. Apical junctions are marked with DLG-1::mCherry. (C-F) Mid-
plane. Note LET-4::GFP expression on the outer surface of the canal cell (asterisk in C).
(G-H) Surface view. Scale bars: 5μm. Data contributed by V.M. and J.P.
49

Figure 2.4. let-4 and egg-6 are each required to maintain junction integrity between
the excretory duct and pore
(A-H) Late 3-fold embryos (C,D) or early L1 larvae (A,F,G) expressing apical junction
marker AJM-1::GFP. L1 larvae also express duct and pore cell marker dct-5p::mCherry.
Arrowhead indicates canal-duct junction; arrow indicates pore autojunction. Dotted line
indicates pharynx, which is also labeled by AJM-1::GFP. B, E and H are schematic
interpretations. Junction morphology initially appears normal in let-4(mn105) (C) and
egg-6(ok1506) (D) mutants, but the canal cell lumen is beginning to swell (asterisk). In
older let-4 (F) and egg-6 (G) mutants, the duct and pore cells have separated, but contain
junction remnants (caret) and the pore autocellular junction has disappeared. Swelling of
the canal cell lumen is more pronounced (asterisk). (I-M) TEM images of the excretory
system. Canal cell is colored red; duct cell, yellow; pore cell, blue; lu indicates lumen;
cut, cuticle. At 1.5-fold (I), let-4 lumen morphology and lumen ECM (line) were
indistinguishable from wild-type (compare to Fig. 2.1I, J). At 3-fold (J-L), 5/9 let-4
embryos also appeared normal (J) (compare to wild-type, Fig 2.1D). In some let-4
embryos that lacked duct-pore connectivity, the duct lumen behind the break was swollen
and the apical membrane had separated from the cuticle lining (K). Panels L-L„‟‟ show
adjacent serial sections documenting duct lumen termination. A bolus of cuticle-like
material is present near the termination point (L). Scale bars in A-H:5μm; scale bars in I-
J:1μm. Data contributed by V.M. and J.P.
Figure 2.5. let-4 and sym-1 are redundantly required to maintain junction integrity
in the epidermis
50

(A) Quantification of embryonic lethality after let-4 RNAi. (B) Rare sym-1; let-4(RNAi)
embryos that do not rupture swell as they approach hatch. (C-H) AJM-1::GFP and (I-N)
HMR-1::GFP apical junction markers appear normal in sym-1 control embryos (C-D, I-J)
and sym-1; let-4(RNAi) embryos (E-F, K-L) at the 1.5 fold stage (C,E,I, K) and at the 3-
fold stage prior to rupture (D,F,J, L). (G,M) Most sym-1; let-4(RNAi) embryos begin to
extrude epidermal cells (asterisk) and rupture after elongating to the 3-fold stage. (H, N)
Terminal arrest phenotype. Scale bars: 5μm. Data contributed by V.M., M.S. and C.P.
Figure 2.6. The cytoplasmic and transmembrane domains of LET-4 are dispensable
for function
(A,B,C) L4 larvae expressing lpr-1p::LET-4 deletion constructs tagged with GFP. lin-
48p::mCherry labels the entire cytoplasm of the duct cell. (A) LET-4::GFP and (C) LET-
4(ΔC-term)::GFP localize along the lumen of the duct cell. (B) signalseqGFP::LET-
4(ΔLRR) is dispersed thoughout the duct cell cytoplasm. (D) let-4(mn105) rescue data.
At least two independent transgenic lines were scored per construct, and non-transgenic
siblings (grey) were scored as negative controls. low, injected at 2.5ng/uL; high, injected
at 5ng/uL. Scale bar: 5μm. LET-4(ΔLRR) contains LET-4 amino acids 398-773; LET-
4(ΔC-term) 1-713; LET-4(ΔTM) 1-688.
51

Figure. 2.7. let-4 and egg-6 are required for proper apical ECM organization
(A) TEM image of 1.5-fold let-4 mutant, adjacent to the pore opening. The distance
between the inner eggshell layer (orange arrows) and the embryonic sheath layer (green
arrows) is increased compared to wild-type, (Fig. 2.1I). (B-C) DIC image of extra-
embryonic cytoplasts in let-4(mn105)(B,B‟) and egg-6(ok1506) 3-fold embryos (C).
Cytoplasts are labeled with vha-1p::GFP in let-4 (B‟). (D) TEM image of extra-
embryonic cytoplasts in let-4 3-fold embryo. (E-J) Hoescht dye permeability assay. DIC
(E-I) and fluorescence (E‟-I‟) images of L1 larvae treated with Hoechst dye, 0.7 seconds
exposure time. (J) Quantification of dye permeability. Scale bars: 5μm. Data contributed
by V.M. and J.P.
Figure. 2.8. LET-4::GFP and EGG-6::GFP localize apically in epithelia
(A,C) LET-4::GFP and (B,D) EGG-6::GFP localize to the apical face of epithelial rectal
cells (A,B) and vulval cells (C,D) in L4 larvae. lin-48p::mCherry labels the cytoplasm of
rectal cells in A‟. (E) EGG-6::GFP is also visible in some neurons (arrows). (F) EGG-
6::GFP is secreted into the extra-embryonic space inside the eggshell in 3-fold embryos.
(G) LET-4::GFP is not enriched at intermediate filaments (G‟) at hemidesmosomes.
Overlay (G‟‟). L4 larva, ventral view. Bracket marks the region of the ventral epidermis
that does not contact body muscle and lacks hemidesmosomes. Scale bars: 5μm. Data
contributed by V.M. and J.P.
52

duct
cuticle
lumen
Figure 2.1
duct cell canal cell
G1pore D
A
ERM-1
canal
lumen
F
B G1 pore
autojunction
G
A
duct
ECM
lumen
ERM-1
canal
lumen
H G1 pore
autojunction
ii
iii
I
G1 pore
ductcell
canalcell
duct
sheath
inner eggshell
ECM
canal cell
G1 pore
cuticle
Jii iii
A
VD
A
cd
p
i
lECM
cuticle
G1 pore
luEC D lu
duct
cuticle
sheath
5μm 1μm 1μm
1μm 1μm1μm5μm
53

NsiI BamHI XbaI
LET-4::GFP rescuing fragment
LG X:
let-4/sym-5 (C44H4.2)C44H4.1 sym-1B
Figure 2.2
GFP
0 20 40 60 80 100egg-6(ok1506); lpr-1p::EGG-6(+)
egg-6(ok1506); dpy-7p::EGG-6(+)
egg-6(ok1506); ExEGG-6::GFP
egg-6(ok1506)/egg-6(cs67)
egg-6(ok1506)
egg-6(cs67)
let-4; lpr-1p::LET-4(+)
let-4; dpy-7p::LET-4(+)
let-4; ExLET-4::GFP
let-4
let-4 unc-3; ExC44H4.2
let-4 unc-3
egg-6(ok1506); ExEGG-6::GFP
+/egg-6(ok1506)
+/egg-6(cs67)
let-4 15°C
let-4 25°C
let-4
+/let-4
Zygotic Genotype
Maternal Genotype (n)
(n)
A
% embryonic lethal % L1 lethal
(242)(208)(72)
(124)(127)
(329)(many)
(84)
(53)(57)
(30)
(many)(many)
(many)(23)
(4)
(18)
(55)(30)
1kb
mn105: Q -> STOP
cs67: Q -> STOP
Signalseq Leucine-Rich Repeats
LET-4 (C44H4.2)
EGG-6 (K07A12.2)
ok1506 (-1678bp)
TM
C
XbaI XhoI
egg-6 (K07A12.2) K07A12.6K07A12.8
LG I:
GFP1kb
EGG-6::GFP rescuing fragment
54

A’A
C’
A’’
C
* *
Figure 2.3
B’B B’’
E E’
D
F
Overlay
Overlay
Overlay
LET-4
LET-4 LET-4
EGG-6
EGG-6
DLG-1
DLG-1
DLG-1
lin-48
EGG-6
G
H
G’
H’
G’’
H’’Overlay
Overlay
LET-4
EGG-6
DLG-1
DLG-1
LET-4
EGG-6
55

Figure 2.4wild-type
d
p
cB
*
**
egg-6let-4
DC
GF
E
H
duct cellcanal cellG1
pore DA
BA
let-4
I
**
*V
V
V
K
J
let-4
lu
dcut
cut
d lu
let-4
lu
d d dcut
lulu
let-4 let-4 let-4
L L’ L’’
5μm
1μm
56
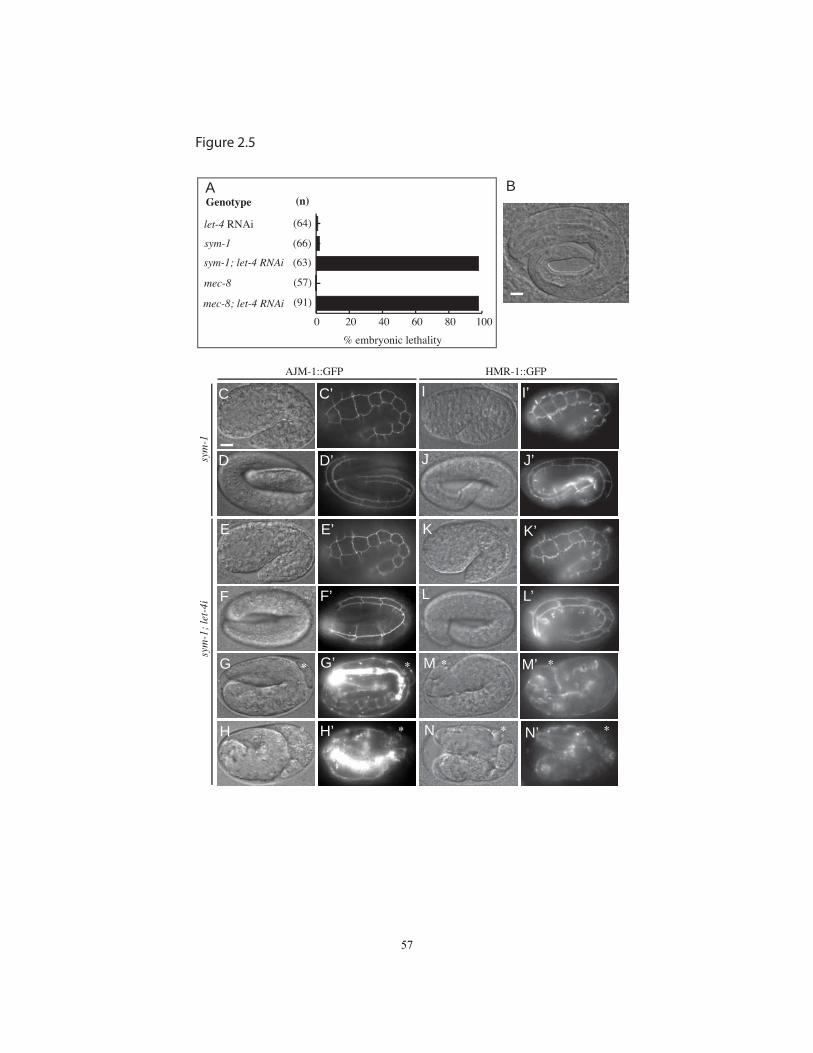
% embryonic lethality
(n)GenotypeA
Figure 2.5
0 20 40 60 80 100
mec-8; let-4 RNAi
mec-8
sym-1; let-4 RNAi
sym-1
let-4 RNAi (64)
(66)
(57)
(91)
(63)
sym
-1;
let-
4isy
m-1
AJM-1::GFP HMR-1::GFP
B
C C’
D D’
E
F
E’
F’
G
H
G’
H’
I
K
I
J
L
M
N
K’
I’
J’
L’
M’
N’
* *
*
* *
* * *
57

A’A
LET-4 lin-48
A’’
Overlay
C’C C’’
B’B B’’
Figure 2.6
transgeneD
(n) (8)
(9)
(40) (19)
(57) (89)
(115)(35)
(35)(14)
0 20 40 60 80 100
(33)(21)
(28) (35)
LET-4(∆LRR) lin-48 Overlay
LET-4(∆C-term) lin-48 Overlay
LET-4(∆LRR)
LET-4(∆LRR)
LET-4(∆C-term)
LET-4(∆C-term)
LET-4(∆TM) (low)
LET-4(∆TM) (high)
LET-4(∆TM) (high)
%lethal0 20 40 60 80 100
transgenic non-transgenic
58
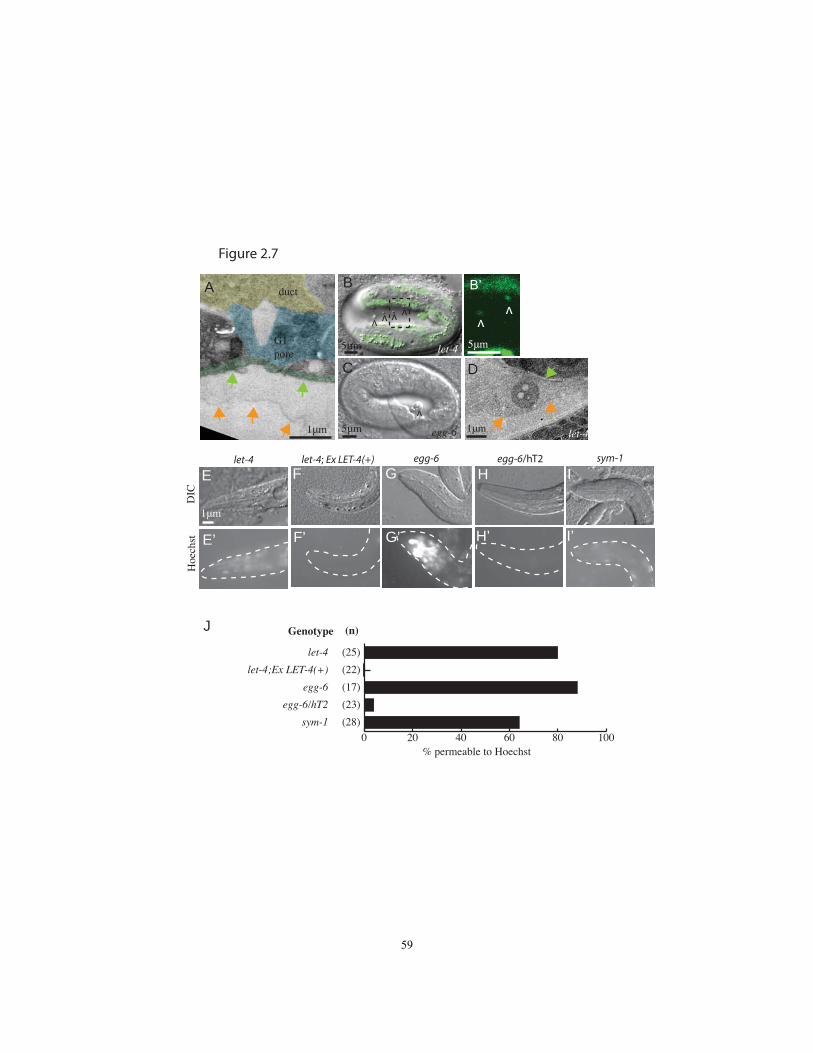
Figure 2.7
B
vv vv
let-4
egg-6
C
v
vv
DIC
egg-6 egg-6/hT2let-4 let-4; Ex LET-4(+)
Hoe
chst
B’
E
E’
F
F’
G
G’
H
H’
D
J
0 20 40 60 80 100
sym-1 (28)
egg-6/hT2 (23)
egg-6 (17)
let-4;Ex LET-4(+) (22)
let-4 (25)
(n)Genotype
% permeable to Hoechst
I
I’
sym-1
let-41μm5μm
1μm
5μm5μmG1 pore
ductA
1μm
59
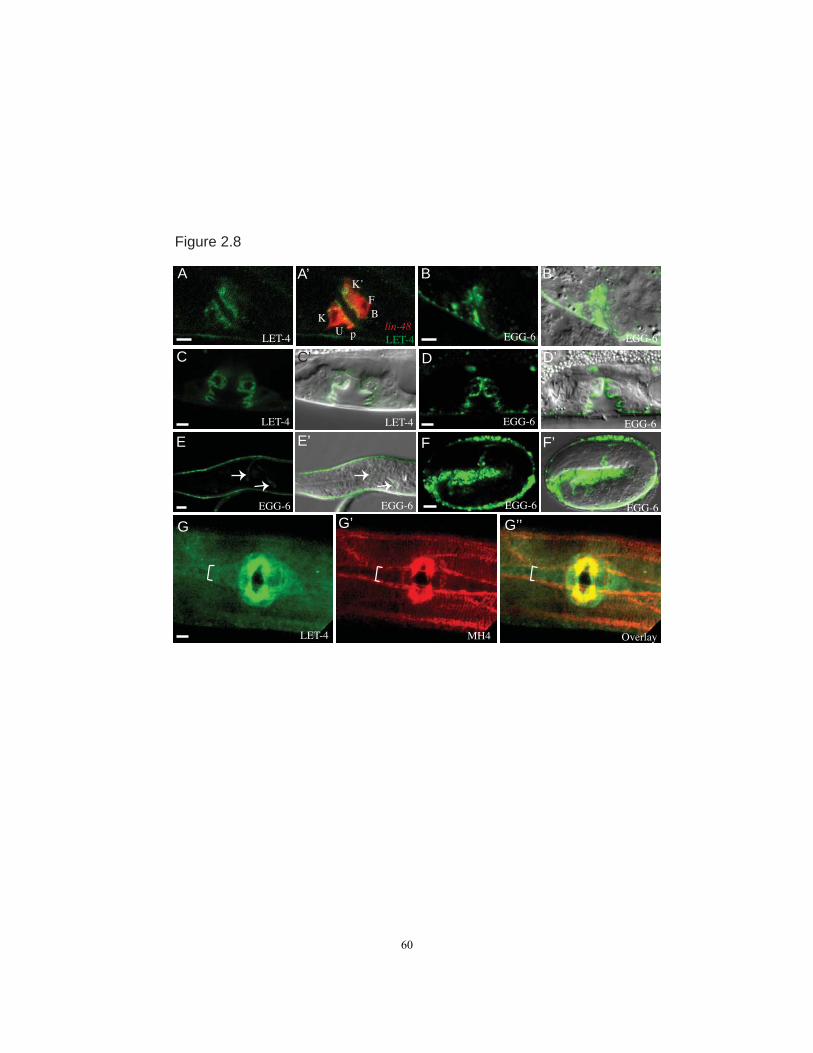
LET-4
K’
FBK
U p
A A’
Figure 2.8
LET-4 LET-4
C C’
EGG-6 EGG-6
D D’
B B’
LET-4lin-48
EGG-6 EGG-6
EGG-6 EGG-6
E E’
EGG-6
F’
EGG-6
LET-4 MH4 Overlay
G G’ G’’
F
60

Chapter 3
Notch and Ras promote sequential steps of excretory tube development in C. elegans
This chapter was previously published:
Abdus-Saboor, I.*, Mancuso, V.P.*, Murray, J.I., Palozola, K., Norris, C., Hall,
D.H., Howell, K., Huang, K., and Sundaram, M.V. (2011) Notch and Ras promote
sequential steps of excretory tube development in C. elegans Development 138, 3545-
3555
*These authors contributed equally
61

Role of Authors
Chapter 3 has been published as presented here. This chapter was a collaborative
effort involving multiple authors, as documented below and in each individual Figure
legend. My work in this manuscript began as an investigation to explore the possibility
of cooperative interaction of Ras and Notch to specify the fate of the cells of the
excretory system.
Ras and Notch are two common signaling pathways, but, in different contexts,
they work sequentially, cooperatively or antagonistically (Sundaram, 2005). For
example, they antagonize each other in the C. elegans vulva. In this system, the anchor
cell produces the Ras pathway activating EGF ligand, which promotes the primary vulval
fate in a vulval precursor cell (Sternberg, 2005). The primary vulval fate is suppressed in
neighboring cells via activation of the Notch pathway. In contrast, in the Drosophila eye,
the Ras and Notch pathways work cooperatively to promote the R7 photoreceptor fate
(Voas and Rebay, 2004). In order to take the R7 fate, the cell must receive both EGF and
DSL, the Notch activating ligand. Ras and Notch are much more commonly in
antagonistic relationships, and further exploration of the two pathways working
cooperatively would be valuable.
Ras and Notch mutants each lack a duct (Lambie and Kimble, 1991; Yochem et
al., 1997), which suggests that Ras and Notch could be working cooperatively to promote
the duct cell fate in the C. elegans excretory system. I decided to explore the possibility
of Ras and Notch working cooperatively in this system, and began by asking basic
questions about the loss of Notch signaling, and removal of the canal cell.
62

I used molecular markers to assay the duct and canal cell fate in a variety of
Notch pathway mutants, and confirmed that the canonical Notch signaling pathway was
involved in duct fate specification. I also found that the duct cell phenotype was
variably and incompletely penetrant in Notch loss of function mutations and RNAi
experiments, suggesting perdurance, maternal effects, or the involvement of other
pathways on the phenotype.
I attempted to determine the source of the Ras pathway inductive signal required
for duct fate specification. In Notch mutants, in addition to the duct being absent, the
canal cell was also absent. We proposed that the duct cell’s absence could be explained
by the canal cell’s absence, because the canal cell expresses LIN-3/EGF. I tested this
model, and demonstrated that the duct cell took its fate even in the absence of the canal
cell through two lines of experimentation. First, in genetic experiments, Notch mutants
lacked a canal cell, but a significant percentage still expressed cell fate marker lin-
48p::GFP in the canal cell. Second, after laser ablating both the canal cell itself, and the
canal cell mother, the presumptive duct cell expressed duct fate marker lin-48::GFP.
The endurance of the duct cell even in the absence of the canal cell indicated that the
canal cell was not the sole source of the inductive signal, and LIN-3 signal from other
sources was sufficient to induce the duct cell fate.
Given the above experiments, evidence seemed to point to an indirect role for
Notch in the specification of the duct cell fate, and multiple sources of ligand for Ras
pathway activation in the duct. I then decided to focus my efforts on investigating let-4
(Chapter 2). We extended my Ras and Notch work, and incorporated new experiments
exploring the roles of these pathways in the development of the excretory system into the
63

manuscript presented here, which includes data from other members of the lab, and other
collaborators.
In the following Chapter, I performed Notch pathway experiments, laser ablation
experiments (Table 3.1, Fig. 3.5), and some LIN-12::GFP characterization (Fig. 3.3).
Ishmail Abdus-Saboor performed Ras pathway mutant phenotyping (Fig. 3.2), sos-1
experiments (Fig. 3.6) and immunostaining (Fig. 3.3). Meera Sundaram performed Ras
and Notch pathway mutant phenotyping (Fig. 3.4, Fig. 3.5, Fig.3.7). Carolyn Norris and
David Hall performed TEM analysis (Fig. 3.1 and Fig. 3.8). John Murray performed 4D
imaging. Katherine Palozola, Kelly Howell, Kai Huang performed Ras pathway loss-of-
function marker analysis (Fig. 3.2).
64

Abstract
Receptor Tyrosine Kinases and Notch are critical for tube formation and
branching morphogenesis in many systems, but the specific cellular processes that require
signaling are poorly understood. Here we describe sequential roles for Notch and
Epidermal Growth Factor (EGF)-Ras-ERK signaling in the development of epithelial tube
cells in the C. elegans excretory (renal-like) organ. This simple organ consists of three
tandemly connected unicellular tubes, the excretory canal cell, duct and G1 pore. lin-12
and glp-1/Notch are required to generate the canal cell, which is a source of LIN-3/EGF
ligand and physically attaches to the duct during de novo epithelialization and
tubulogenesis. Canal cell asymmetry and let-60/Ras signaling influence which of two
equivalent precursors will attach to the canal cell. Ras then specifies duct identity,
inducing auto-fusion and a permanent epithelial character; the remaining precursor
becomes the G1 pore, which eventually loses epithelial character and withdraws from the
organ to become a neuroblast. Ras continues to promote subsequent aspects of duct
morphogenesis and differentiation, and acts primarily through Raf-ERK and the
transcriptional effectors LIN-1/Ets and EOR-1. These results reveal multiple genetically-
separable roles for Ras signaling in tube development, as well as similarities to Ras-
mediated control of branching morphogenesis in more complex organs, including the
mammalian kidney. The relative simplicity of the excretory system makes it an attractive
model for addressing basic questions about how cells gain or lose epithelial character and
organize into tubular networks.
65

Introduction
Many organs, such as the mammalian kidney and the vasculature, consist of
complex networks of tubules that develop from clusters of initially unpolarized
mesenchymal cells (Hogan and Kolodziej, 2002; Lubarsky and Krasnow, 2003; Dressler,
2009). The processes by which these cells polarize, form epithelial or endothelial
junctions, and then organize into complex tubular shapes are only beginning to be
elucidated. In many cases, signaling pathways involving Receptor Tyrosine Kinases
(RTKs) and Ras are critical for formation and patterning of the tubular network. For
example, during branching morphogenesis of the ureteric bud in the kidney, signaling by
the Ret RTK promotes tip cell identity and specifies the location of new branches
(Shakya et al., 2005; Chi et al., 2009). Similarly, during sprouting angiogenesis,
signaling by vascular endothelial growth factor receptors promotes tip cell identity
(Gerhardt, 2008). Absence of RTK signaling results in renal or vascular agenesis.
Although the importance of RTK pathways in controlling tube development is clear, the
specific cellular behaviors that require signaling, and the downstream mechanisms that
control them, are not well understood.
Tubulogenesis can be reversible, as cells can withdraw from an existing tube and
give rise to different cell types. For example, venous endothelial cells in the mouse de-
differentiate and divide to give rise to new coronary arteries, capillaries and veins as part
of their normal developmental program (Red-Horse et al., 2010). Epithelial-to-
mesenchymal transition (EMT) or endothelial-to-mesenchymal transition (EndMT) are
central features of injury-induced fibrosis in the kidney and heart (Kalluri and Neilson,
66

2003; Zeisberg et al., 2007) and underlie the metastatic properties of many tumor cells
(Kalluri and Weinberg, 2009). Tubes that form by de novo polarization may be
particularly prone to EMT, but the mechanisms that promote or restrain such behaviors
remain poorly understood.
The C. elegans excretory system is a simple example of an epithelial tube
network. The excretory system is the worm’s renal-like system and is required for fluid
waste expulsion (Nelson et al., 1983; Nelson and Riddle, 1984; Buechner, 2002). It
consists of three tandemly arranged unicellular tubes: the large canal cell (which runs the
length of the body and appears to collect waste fluid), and the smaller duct and pore cells
(which connect the canal cell to the outside environment) (Fig. 3.1). While the canal cell
and duct tubes are permanent throughout the life of the animal, the G1 pore eventually
withdraws from the excretory system to become a neuroblast, at which time a
neighboring epidermal cell (G2) replaces G1 as the excretory pore tube (Sulston et al.,
1983; Stone et al., 2009). Thus the excretory system provides a simple, genetically
tractable system for studying the dynamic control of epithelial junctions, cell shape and
cell identity.
The progenitors of the excretory duct and G1 pore tubes are left/right lineal
homologs that appear to compete for the duct fate (Sulston et al., 1983). In wild-type
animals, the left cell always becomes the duct and adopts the most canal cell-proximal
position, while the right cell always becomes G1 and adopts a more distal position.
However, ablation of the mother of the presumptive duct causes the presumptive G1 to
adopt a duct-like position and morphology, showing that both cells have the capacity to
become a duct and suggesting some lateral inhibitory mechanism that prevents both from
67

doing so. Both let-60/Ras and lin-12 glp-1/Notch mutants lack an excretory duct,
implicating Ras and Notch in the duct vs. G1 pore fate decision or some other aspect of
duct development (Lambie and Kimble, 1991; Yochem et al., 1997).
Here we show that Notch and Epidermal Growth Factor (EGF)-Ras-ERK act
sequentially during excretory tube development. We identify multiple, genetically
separable requirements for signaling in controlling tube cell position, identity, shape and
function. Finally, we establish the excretory duct and G1 pore system as a model for
investigating many basic cell biological processes associated with tube development and
EMT-coupled cell fate plasticity.
68

Results
Excretory tube development involves de novo formation and remodeling of epithelial
junctions
Excretory tube development occurs in three broad phases
(migration/tubulogenesis, morphogenesis/differentiation, and G1 withdrawal/remodeling)
(Fig. 3.1A) (Sulston et al., 1983; Buechner, 2002; Berry et al., 2003; Stone et al., 2009).
To visualize the excretory duct and G1 pore during these phases, we used lineage-specific
markers in combination with epithelial apical junction markers AJM-1 (Koppen et al.,
2001) and DLG-1/Discs Large (Bossinger et al., 2001)(Fig. 3.1). GFP::MLS-2 marks all
ABpl/rpaaa descendants (including the duct and G1) plus additional lineages during
ventral enclosure (Yoshimura et al., 2008) (J.I.M., unpublished data) (Fig. 3.1B-C,E),
while dct-5p::mCherry marks the duct, G1 and some other epithelial cells during L1 (this
work, Fig. 3.1J-K). We also analyzed a ventral enclosure embryo by transmission
electron microscopy (TEM) of serial sections (Figs 3.1D, 3.8) and traced the canal, duct,
and G1 pore lineages through ventral enclosure from 3D confocal movies of eight
histone::mCherry-expressing embryos (Materials and Methods).
The canal cell, duct and G1 pore progenitors are born in disparate locations of the
embryo. During ventral enclosure (Fig. 3.1B-E), the duct (left) and G1 pore (right)
progenitors migrate toward the canal cell, which is located slightly left of the ventral
midline. The duct progenitor has a shorter distance to migrate, and it appears to reach the
69

canal cell first (Fig. 3.1C). TEM of an embryo at ventral enclosure shows the duct
progenitor and the canal cell closely apposed, while the G1 progenitor is excluded from
the canal cell by the duct and other intervening cell bodies (Fig. 3.1D). In 6/8 3D
confocal movies, the duct nucleus arrives adjacent to the canal cell about 5-10 minutes
before G1. At this time, 1-2 nuclei still separate the canal from G1; the most consistent
of these is RIS, a right-derived neuron whose left homolog is related to the canal cell and
undergoes programmed cell death. In 2/8 movies, the duct and G1 nuclei arrive near the
canal cell at approximately the same time, so it is not possible to determine which cell
contacts the canal cell first in the absence of a membrane or cytoplasmic label. After the
duct reaches the canal cell, they begin to ingress, while G1 moves to a ventral position
between the G2 and W epidermal cells (Fig. 3.1E). Together, these observations suggest
that asymmetry of the canal cell and its lineal relatives might contribute to asymmetry in
duct and G1 pore behavior.
The cells initially lack epithelial junctions (Fig. 3.1B, D), but after they contact
each other, they form epithelial junctions and undergo tubulogenesis (Fig. 3.1F). As
described previously (Stone et al., 2009), the duct and G1 pore cells wrap up into tube
shapes and form autocellular junctions. G1 retains this autocellular junction, but the duct
cell rapidly auto-fuses, becoming a seamless toroid. The canal cell forms lumen
intracellularly at the site of the duct-canal cell intercellular junction. By the 1.5 fold
stage, the canal cell, duct and pore form a simple block-like stack of tandemly connected
unicellular tubes with a continuous lumen and prominent epithelial junctions (Fig.
3.1A,F).
70

Further morphogenesis occurs during the latter part of embryogenesis, such that
by the first larval stage, the excretory duct (Fig. 3.1G-J) and canal cell (Fig. 3.1L) have
distinctive elongated shapes. The cells also begin expressing unique differentiation
markers, such as the lin-48/Ovo transcription factor in the duct (Fig. 3.1I) (Johnson et al.,
2001).
G1 withdrawal and G2 entry occur in the first larval stage, after the excretory
system has already begun to function. At this time, G1 migrates dorsally and loses its
epithelial junctions while a neighboring epidermal cell, G2, forms an autocellular
junction and replaces it as the pore (Fig. 3.1K) (Sulston et al., 1983; Stone et al., 2009).
G2 subsequently divides in L2 to generate a neuronal daughter (G2.a) and an epithelial
daughter (G2.p) that replaces it as the permanent pore tube (Sulston and Horvitz, 1977).
Throughout this time the duct process must remodel its ventral junction to connect to its
new partners.
let-60/Ras is both necessary and sufficient for duct vs. G1 pore fate specification
let-60/Ras is required cell autonomously within the excretory duct cell for proper
excretory system function and organismal viability and was previously proposed to
promote the duct vs. G1 pore fate (Yochem et al., 1997). To test this model, we used
AJM-1::GFP and lin-48p::GFP markers to examine let-60 ras mutants (Fig. 3.2).
Most let-60(sy101sy127lf) null mutants, obtained from heterozygous mothers,
have two pore-shaped cells with autocellular junctions and no lin-48p::GFP (Fig. 3.2B-
C,K-L), consistent with a duct-to-G1 pore cell fate transformation. Although the mutants
lack a duct-like cell, the overall arrangement of the excretory system resembles that of
71

wild-type animals: two cells are arranged in tandem, with one contacting the canal cell
and the other contacting the ventral epidermis. Thus, initial migration, stacking and
tubulogenesis appear normal, but auto-fusion does not occur and duct-specific
differentiation markers are not expressed.
let-60(sy101sy127lf) mutants die as late L1 larvae with a rigid, fluid-filled
appearance termed “rod-like lethality”. Fluid first accumulates near or within the two
pore-like tubes (Fig. 3.2C'). Notably, the timing of fluid accumulation in mid-L1
coincides with the normal timing of pore remodeling and is often associated with large
junctional rings or discontinuities (Fig. 3.2C). Since withdrawal from the excretory
system is a normal feature of G1 pore identity, withdrawal of both pore-like cells may
explain the inviability.
let-60(n1046gf) hypermorphic mutants have two duct-like nuclei expressing lin-
48p::GFP and no autocellular junctions (Fig. 3.2D,K-L), consistent with a G1 pore-to-
duct cell fate transformation. The two duct cells fuse to form a binucleate cell, as would
be predicted for two adjacent cells expressing the fusogen aff-1, which is required for
duct auto-fusion (Stone et al., 2009) and generally sufficient for fusion of adjacent cells
(Sapir et al., 2007). Removal of aff-1 in a let-60(gf) background restored both
intercellular and autocellular junctions (data not shown). The binucleate duct cell attaches
to the ventral epidermis, allowing for fluid excretion, and is permanent throughout the
life of the animal.
We conclude that Ras signaling is both necessary and sufficient to promote duct
vs. G1 pore identity, and that identity can be uncoupled from cell position. Notably,
however, some let-60 null mutants still possess a cell with at least partial duct-like
72

character (Fig. 3.2K-L). Evidence below suggests that this is due to maternally-provided
let-60 activity; maternal activity cannot be completely removed in our experiments
because of the requirements for let-60 and other pathway components in germline
development (Church et al., 1995).
let-60/Ras functions within the canonical EGF-Ras-ERK pathway to promote the
duct fate
Animals mutant for lin-3/EGF or various other components of the canonical EGF-
Ras-ERK pathway all display a rod-like lethal phenotype associated with excretory
system failure (Ferguson and Horvitz, 1985; Sundaram, 2006). We examined mutants for
lin-3/EGF and lin-1/Ets, which lie at the beginning and end of this pathway, respectively
(Hill and Sternberg, 1992; Beitel et al., 1995). lin-3(lf) and lin-1(gf) mutants appear
similar to let-60(lf) mutants, and lin-3 overexpression and lin-1(lf) mutants appear similar
to let-60(gf) mutants (Fig. 3.2E-H,K-L). Furthermore, a variety of other Ras pathway
mutants examined (including hypomorphic alleles of let-23/EGFR and lin-45/Raf) also
show evidence of duct-to-pore fate transformations (Fig. 3.2K-L). Finally, eor-1 and sur-
2 are nuclear factors that act redundantly downstream of MPK-1/ERK (Singh and Han,
1995; Tuck and Greenwald, 1995; Howard and Sundaram, 2002); eor-1 also appears to
act redundantly with a cryptic positive function of lin-1/Ets (Howard and Sundaram,
2002; Tiensuu et al., 2005). We found that eor-1; sur-2(RNAi) and lin-1 eor-1 double
mutants frequently have two pore-like cells (Fig. 3.2I-L). These data are consistent with
the entire canonical pathway promoting duct vs. G1 pore identity.
73

During this analysis, we noted that some mutants with reduced signaling had
paradoxical "0 G1"-like junction patterns, without concomitant duct fate duplication, or
had excretory failure despite apparently normal junction patterns and fates (Fig. 3.2K-L).
These observations suggested that Ras signaling plays roles beyond cell fate specification
(see below).
The excretory canal cell expresses lin-3/EGF
Since the mutant analyses above suggest that signaling by LIN-3/EGF through
LET-23/EGFR is responsible for LET-60/Ras activation in the duct, we asked where
these proteins are expressed. Consistent with the fact that both cells can respond to LIN-
3 to adopt the duct fate, a functional LET-23::GFP reporter, gaIs27 (Simske et al., 1996),
is expressed in both presumptive duct and G1 pore cells during ventral enclosure (Fig.
3.3A). To examine lin-3 expression, we used a lin-3 promoter::GFP reporter (syIs107)
that contains ~2.5 kb of upstream regulatory sequence as well as the first lin-3 intron
(Hwang and Sternberg, 2004). Most notably, lin-3p::GFP is strongly expressed in the
excretory canal cell, beginning soon after canal cell birth and continuing into early larval
development (Fig. 3.3B, D). lin-3p::GFP is also expressed in a variety of other cells that
are further away from the presumptive duct and G1 pore. These data suggested that the
canal cell might be a relevant source of the duct-inducing signal, a model that fits with
the observation that the left member of the equivalence group, which is closest to the
canal cell, is the cell that normally adopts the duct fate.
Ras signaling promotes cell stacking and a canal cell-proximal position
74

EGF-Ras signaling could promote duct vs. G1 pore cell fate specification
independently of cell stacking and tubulogenesis, or signaling could also control initial
cell positioning. The latter possibility was suggested by results of a prior mosaic
analysis, in which a let-60(+) presumptive G1 cell could outcompete a let-60(-)
presumptive duct cell for the more dorsal, canal cell-proximal position and take on the
duct fate (Yochem et al., 1997).
Consistent with a model in which both cells compete for the canal cell-proximal
position, animals homozygous for a partial loss-of-function allele, let-60(n2021), display
a variable phenotype in which the presumptive duct and G1 cells often adopt adjacent
positions rather than stacking on top of each other (Fig. 3.4C-D). In many of these cases,
a single duct-like cell reaches from the ventral epidermis to the canal cell, while the
second cell is mispositioned to the side and appears non-tubular, giving a "1 duct, 0 G1"
phenotype (Fig. 3.4G-H). In other cases, a single pore-like (un-induced) cell reaches from
the ventral epidermis to the canal cell, giving a “0 duct, 1 G1” phenotype (Fig. 3.4I-J).
Similar defects are seen in other hypomorphic mutants and in lin-1; eor-1 double mutants
(Fig. 3.2K and data not shown). We conclude that Ras signaling influences duct and G1
pore stacking.
Notably, the adjacent phenotype is observed only occasionally in let-60 or lin-3
null mutants obtained from heterozygous mothers (Fig. 3.2K, Fig. 3.4K). This is due in
part to maternal rescue, since most let-60(n2021rf) mutants obtained from heterozygous
mothers also have normal cell stacking, in contrast to those obtained from homozygous
mutant mothers (Fig. 4K). Nevertheless, progeny from let-60(sy101sy127lf)/let-
60(n2021rf) mothers have a lower frequency of adjacent cells than those from let-
75

60(n2021rf) mothers (Fig. 3.4K). Therefore, the adjacent phenotype may reflect
problems in resolving cell competition under circumstances where Ras signaling is sub-
optimal but not absent (see Discussion).
The canal cell is required for stacking and tubulogenesis of the duct and G1 pore
To test if LIN-3/EGF expression by the canal cell is required for duct fate
specification or cell stacking, we first removed the canal cell (or its mother) physically by
laser ablation. In the absence of the canal cell, most animals still had a lin-48p::GFP+
cell (Table 3.1), indicating that other sources of LIN-3 are sufficient to induce at least
some features of duct identity. However, duct morphology was abnormal and the G1
pore autocellular junction was missing (Fig. 3.5L-M), suggesting that stacking had been
disrupted.
We next examined the effects of removing the canal cell genetically using Notch
mutants. Mutants lacking both C. elegans Notch receptors, LIN-12 and GLP-1, the DSL
ligand LAG-2 or the CSL transcription factor LAG-1 have a constellation of defects
referred to as the “Lag” (lin-12 and glp-1) phenotype (Lambie and Kimble, 1991). lag
mutants lack an excretory canal cell due to a lineage transformation affecting the canal
cell’s great-grandmother ABplpapp (Lambie and Kimble, 1991; Moskowitz and
Rothman, 1996). As in canal cell-ablated animals, lag mutants often possess a
morphologically abnormal lin-48p::GFP+ cell, and lack a G1 pore autocellular junction
(Table 1, Fig. 3.5). A ventral perspective revealed that the presumptive duct and G1 pore
cells adopt adjacent ventral positions in the epidermis (Fig. 3.5B,C). Thus lag mutants
resemble let-60/Ras partial loss-of-function mutants.
76

Several lines of evidence suggest that the lag duct and G1 pore stacking defects
are a secondary consequence of canal cell absence. First, canal cell ablation in wild-type
embryos can phenocopy lag mutants. Second, a functional LIN-12::GFP reporter
(arIs41) is not detectably expressed in the duct or G1 pore progenitors during ventral
enclosure (Fig. 3.3E), nor is a LIN-12- and GLP-1-responsive reporter, ref-1p::GFP (data
not shown); thus Notch signaling is unlikely to impact directly on duct and G1 pore fate
specification or tubulogenesis. Third, lin-12 hypermorphic mutants, which have a single
canal cell, have normal duct and G1 pore morphology (Table 3.1, Fig. 3.5M). Finally,
examination of lin-12 null mutants or lag-2(q420) hypomorphic mutants, in which
absence of the canal cell is variable, revealed a strong correlation between absence of the
canal cell and failure of the duct and G1 pore to stack and undergo tubulogenesis (Fig.
3.5 D-E,G-H,M).
Together, the ablation and Notch mutant data support a model in which the canal
cell facilitates duct and G1 pore stacking and tubulogenesis. The canal cell likely
provides physical support to the duct and G1 pore as it adheres to the duct during these
processes. Stacking and tubulogenesis appear independent of canal cell-expressed lin-
3/EGF, since these processes are intact in lin-3 zygotic null mutants, despite defects in
cell fate specification (Fig. 3.2K-L). Duct fate specification also appears partially
independent of canal cell-expressed lin-3/EGF, stacking and tubulogenesis, since it is
only mildly affected by canal cell absence (Table 3.1). Nevertheless, since the canal cell
does express lin-3/EGF, and partial reduction of let-60/Ras can mimic canal cell absence,
localized LIN-3/EGF expression by the canal cell may help orient relative duct and G1
77

pore positions during stacking and bias which cell ultimately adopts the duct fate (see
Discussion).
Continued signaling through SOS-1 and Ras is required for duct morphogenesis
and differentiation
To test the temporal requirements for Ras signaling, we conducted temperature-
shift experiments with a sos-1 (Ras guanine nucleotide exchange factor) temperature
sensitive allele (Fig. 3.6). sos-1(cs41) mutants appear essentially normal at 20˚ C but
arrest with excretory system abnormalities when raised at 25˚ C (Rocheleau et al., 2002).
The cs41 lesion affects the CDC25-related Ras GEF domain of SOS-1, and importantly,
sos-1(cs41ts) lethality is almost completely suppressed by let-60(n1046gf) (Rocheleau et
al., 2002) or lin-1(e1275lf) mutations (Fig. 3.6A) , indicating that lethal defects are
caused by a failure in Ras-ERK-mediated signaling. Since let-60 ras is required only in
the duct cell (and not in the G1 or G2 pore or canal cell) for proper excretory function
and viability (Yochem et al., 1997), we further infer that any excretory abnormalities of
sos-1(ts) animals reflect requirements for Ras-ERK signaling in the developing duct cell.
In sos-1(ts) upshift experiments, upshifts prior to the 1.5-fold stage of
embryogenesis could recapitulate the let-60 ras zygotic null phenotype, in which the two
cells stacked properly but failed to undergo auto-fusion or to express the marker lin-
48p::GFP, suggesting both had adopted G1 pore-like fates (Fig. 3.6B,C). The earliest
maternal upshifts could occasionally generate adjacent cells as seen in let-60(n2021)
hypomorphs (data not shown). In sos-1(ts) downshift experiments, most animals were
normal for excretory morphology as long as they were moved to permissive temperature
78

by the bean stage of embryogenesis (Fig. 3.6C). These results are consistent with the
model that Ras signaling and cell fate specification occur as the presumptive duct and G1
pore cells approach the canal cell and undergo tubulogenesis.
Unexpectedly, the sos-1(ts) temperature-sensitive period for lethal excretory
defects extended from the bean stage into the L1 larval stage (Fig. 3.6D), revealing
continued requirements for signaling beyond initial cell fate specification. At least 70%
of animals upshifted at the 1.5-fold, 2-fold, 3-fold or early L1 stages (n>20 each)
accumulated fluid either within the excretory tubules or near the canal-duct junction,
despite an initially normal junction and lin-48p::GFP marker pattern (Fig. 3.6E, F),
suggesting other defects in organ architecture. While additional studies will be needed to
understand the cellular basis of these later defects, we conclude that SOS-1 and Ras, and
most likely the entire EGF-Ras-ERK pathway, play additional roles in duct
morphogenesis and differentiation.
G1 pore withdrawal can still occur in the absence of G2
When the G1 pore withdraws from the excretory system during L1, a neighboring
epidermal cell, G2, moves in to replace it as the pore (Sulston et al., 1983; Stone et al.,
2009) (Fig. 3.1J; Fig. 3.7A-C,M). By examining Ras and Notch pathway mutants, we
were able to address a basic question about the G1-G2 remodeling event: is
communication between G1 and G2 important to trigger G1’s withdrawal and/or G2’s
entry into the excretory system?
As described above, let-60(n1046gf) mutants invariably lack a G1 pore and have a
binucleate duct cell attached directly to the ventral epidermis. In 16% (11/67) of such
79

mutants, G2 still moves in and gives rise to a morphologically normal larval pore cell; in
the remainder, G2 (or G2p) wraps around the base of the duct but does not form a pore of
normal height (Fig. 3.7 D-F,M). Thus, G2 entry does not require a “come here” signal
from the G1 pore, but its morphogenesis and ability to insert between the duct and
epidermis may be facilitated by the act of G1 withdrawal.
To test the requirements for G2, we used lin-12/Notch single mutants, which
affect the G2 vs. W neuroblast cell fates (Greenwald et al., 1983). lin-12(d) hypermorphic
mutants have two G2 cells, and one of these forms a normal larval pore while the other
wraps around its ventral base (Fig. 3.7J-L,M). Conversely, lin-12(0) loss-of-function
mutants lack a G2 cell. In such mutants, G1 still withdraws from the excretory system
during mid-L1, and the duct then attaches directly to the ventral epidermis (Fig. 3.7G-
I,M). Thus, G1 withdrawal does not require a “go away” signal from G2.
80

Discussion
We have shown that Notch signaling and Ras signaling function sequentially to
control tube development in the C. elegans excretory system. Notch signaling is required
to generate the canal cell, which is a central organizer of duct and G1 pore development,
serving both as a source of LIN-3/EGF ligand (which contributes to Ras activation) and as
a physical attachment site for the duct (which is important for cell stacking and
tubulogenesis). Ras signaling influences cell positions, specifies duct vs. G1 pore
identity, and promotes subsequent aspects of duct morphogenesis and differentiation.
Below we propose a model for duct and G1 pore development and discuss similarities and
differences between development of the excretory system and development of more
complex tube networks.
A biased competition model for excretory duct vs. G1 pore fate specification
All excretory tubes are examples of left-right asymmetries in what is a mostly
bilaterally symmetric embryo (Sulston et al., 1983; Pohl and Bao, 2010). Notch signaling
on the left side of the embryo is required for the earliest of these asymmetries, generation
of the excretory canal cell (Lambie and Kimble, 1991; Moskowitz and Rothman, 1996).
We propose that Notch-dependent asymmetry of the canal cell leads to the Ras-dependent
asymmetry of the excretory duct and G1 pore.
According to this biased competition model, the presumptive duct and G1 pore
cells are initially equivalent. As these cells migrate toward the canal cell during ventral
enclosure, the left cell has an advantage due to the left-biased asymmetric position of the
81

canal cell; this bias may be strengthened by the presence of cells on the right side whose
left relatives undergo cell death. The left cell therefore reaches and adheres to the canal
cell first, and also receives earlier or quantitatively more LIN-3/EGF signal. LIN-3/EGF
signaling stimulates LET-60/Ras to promote duct identity and strengthen adhesion with
the canal cell. Signaling may also trigger production of an unknown lateral inhibitory
signal that prevents the presumptive G1 from also responding to LIN-3/EGF. Steric
hindrance or differences in relative Ras vs. inhibitory signaling levels cause the
presumptive G1 to take a more ventral position. Polarization and initial tubulogenesis
appear independent of Ras signaling; however, after both cells wrap up into tube shapes,
continued LIN-3/EGF signaling from the canal cell (and elsewhere) promotes duct vs. G1
pore identity and later aspects of duct morphogenesis and differentiation into a functional
tube.
Two aspects of this model can explain the stacking defects of let-60/Ras
hypomorphs, in which depletion of both maternal and zygotic let-60/Ras compromises
(but does not eliminate) the earliest steps of signaling. First, the presumptive duct, upon
reaching the canal cell, may not adhere to it strongly. Second, the presumptive duct may
not express the proposed inhibitory signal in a timely manner. Under conditions where
Ras signaling is reduced but not absent, this would allow the presumptive G1 pore to
respond to LIN-3/EGF and compete for a canal-cell proximal position. Failure of either
cell to adhere to the canal cell (as in lin-12 glp-1/Notch mutants), or failure to resolve
competition between the two cells such that both adhere, could lead to the observed
adjacent positions.
82

Lateral inhibition is a central feature of RTK-mediated branching morphogenesis
in several tubular organs (Ghabrial and Krasnow, 2006; Chi et al., 2009) and lateral
inhibition of Ras-dependent processes is frequently mediated by Notch signaling
(Sundaram, 2005). However, we find no evidence that Notch signaling directly influences
excretory duct vs. G1 pore cell fates. Differences in cell adhesion and steric hindrance
may be sufficient to explain the stacking process, but they are unlikely to explain how
only a single duct-like (lin-48p::GFP+) cell is specified from the two adjacent precursors
in a lin-12 glp-1/Notch mutant (Table 3.1). Therefore, an unknown signaling pathway
may be used to mediate lateral inhibition of the duct fate.
Downstream consequences of EGF-Ras-ERK signaling in the excretory duct
In addition to influencing cell positions, EGF-Ras-ERK signaling is both
necessary and sufficient for several aspects of duct vs. G1 pore identity, including duct-
specific patterns of gene expression, auto-fusion, and a permanent epithelial identity. This
latter difference in duct epithelial permanence vs. G1 withdrawal may ultimately explain
the lethality of the duct-to-G1 pore fate change in let-60 ras null mutants. G1 withdrawal
does not depend on cues from the replacement cell G2, but instead appears to be an
intrinsically programmed characteristic of the duct and G1 progenitors that is repressed
by Ras signaling. Ras may inhibit withdrawal in part by stimulating aff-1-dependent auto-
fusion to permanently remove the duct autocellular junction and prevent its later
unwrapping; however, Ras must have additional effects since the duct cell still remains
permanent in most aff-1 mutants despite a failure of auto-fusion (Stone et al., 2009).
83

sos-1(ts) temperature-shift experiments suggest that Ras signaling continues to be
required after initial fate specification for development of a fully functional duct tube.
After its auto-fusion to form a toroid, the duct elongates, changes shape, and elaborates a
complex lumen (Stone et al., 2009). The junctions between the duct and its neighboring
tubes must be maintained and may undergo further maturation to establish barrier
functions and prevent excretory fluid leakage. Finally, the duct-pore junction must be
remodeled as G1 withdraws and G2 enters. The continued requirement for sos-1 as these
events are occurring suggests that Ras signaling may directly promote such
morphogenetic and differentiation processes.
Most or all of the responses to Ras signaling in the excretory duct appear to be
transcriptionally-mediated. sos-1(ts) defects can be rescued by loss of the LIN-1/Ets
transcription factor, which is regulated by MPK-1 ERK phosphorylation (Jacobs et al.,
1998) and acts as a repressor of the duct fate. sos-1(ts) defects also can be mimicked by
combinatorial loss of LIN-1 and another downstream transcription factor, EOR-1 (a
BTB-zinc finger protein) (Howard and Sundaram, 2002; Howell et al., 2010), revealing a
second (but redundant) activity of LIN-1/Ets in promoting the duct fate. A challenge for
future work will be to connect these transcriptional effectors to downstream targets that
control the various cell biological processes of duct auto-fusion, morphogenesis, and
epithelial maintenance.
Similarities and differences between the excretory system and more complex tube
networks
84

C. elegans excretory tubes are topologically quite different from epithelial tubes
in other renal systems in that they are each only one cell in diameter. However, similar
unicellular tubes have been described in other organ systems, including the Drosophila
trachea (Ghabrial et al., 2003) and the mammalian microvasculature (Bar et al., 1984).
Furthermore, in vitro studies suggest that unicellular tubes may be developmental
precursors to some larger bore tubes in the vasculature (Folkman and Haudenschild, 1980;
Iruela-Arispe and Davis, 2009).
Despite their topological differences, C. elegans excretory tubes and larger
multicellular tubes must undergo many similar cell biological processes. For example,
initially unpolarized cells must transition to an epithelial state, define an appropriate apical
domain, form new junctions, and build a lumen; the difference is that excretory tubes
define an intracellular rather than an extracellular lumen. Furthermore, distinct tube types
must join to form a continuous conduit. The maturing tubes must be structurally strong to
withstand internal pressure from their contents, yet flexible enough to elongate and grow
as organismal size or physiological demands increase. Finally, some epithelial tube cells,
like the G1 pore, retain the developmental potential to adopt different fates (Jarriault et al.,
2008; Mani et al., 2008; Weaver and Krasnow, 2008; Kalluri and Weinberg, 2009; Red-
Horse et al., 2010). Given the simplicity of the C. elegans excretory system and its
amenability to genetic manipulations, further studies in this system should give insights
into basic cellular mechanisms involved in these common steps of tubular organ
development
85

Materials and Methods
Strains and alleles
N2 var. Bristol was the wild-type strain. Unless otherwise indicated, all strains were
grown at 20˚C under standard conditions (Brenner, 1974) and all mutant alleles are
described in (Riddle et al., 1997). I: lag-1(q385), lag-2(q411), lag-2(q420). II: let-
23(sy97). III: lin-12(n137), lin-12(n137n720), lin-12 (n941), glp-1(q46), glp-1(q231).
IV: eor-1(cs28) (Rocheleau et al., 2002), let-60(sy101sy127), let-60(n1046), let-
60(n2021), lin-1(e1275), lin-1(n304) (Beitel et al., 1995), lin-1(n1761) (Jacobs et al.,
1998), lin-3(n1059), lin-45(n2018). V: sos-1(cs41) (Rocheleau et al., 2002). X: lin-
15(n765), sem-5(n2019). Transgenes used are: arIs12 (lin-12 intra) (Struhl et al., 1993),
arIs41 (LIN-12::GFP) (Levitan and Greenwald, 1998), gaIs27 (LET-23::GFP) (Simske et
al., 1996), jcIs1 (AJM-1::GFP) (Koppen et al., 2001), saIs14 (lin-48p::GFP) (Johnson et
al., 2001), syIs107 (lin-3p::GFP) (Hwang and Sternberg, 2004), wIs78 (AJM-1::GFP)
(Koh and Rothman, 2001), xnIs17 (DLG-1::GFP) (Totong et al., 2007), vha-1p::GFP
(Oka et al., 1997), zuIs143 (ref-1p::GFP) (Neves and Priess, 2005). qnEx59 (dct-
5p::mcherry) was provided by Julia and David Raizen and contains 845 bp of the dct-5 5'
region. csIs55 (GFP::MLS-2) was generated from a pYJ59-containing array (Jiang et al.,
2005) by gamma-irradiation-induced integration. csEx146 (lin-48p::mcherry) contains
4.8 kb of the lin-48 5’ region and mcherry in vector pPD49.26 (Fire et al., 1990). lin-3
overexpression was achieved with an integrated lin-3p::LIN-3::GFP transgene provided
by Min Han.
86

Marker Analysis and Imaging
Images were captured by differential interference contrast (DIC) and epi-fluorescence
microscopy using a Zeiss Axioskop and Hamamatsu C5985 camera, or by confocal
microscopy using a Leica SP5. Images were processed for brightness and contrast using
Photoshop or ImageJ. Some AJM-1::GFP images were inverted for clarity.
For electron microscopy, embryos were mounted on an agarose pad and observed
under light microscopy to identify timepoints for fixation. A laser was used to place 3-4
holes in the eggshell, allowing the embryo to be aldehyde fixed while on the pad (see
more details at www.wormatlas.org/laserhole.htm). The fixed embryo was postfixed with
osmium tetroxide, potassium ferrocyanide, and tannic acid, and then post-stained with
uranyl acetate before embedding in plastic resin. Transverse serial thin sections were
collected on slot grids and photographed on a Philips CM10 electron microscope. The G1
and duct cells were identified within a series of 600 serial thin sections on the basis of
their positions relative to the canal cell and to the G2 and W epidermal cells (Fig. 3.8)
and by comparison to known nuclear positions in time-lapse confocal movies.
To visualize the duct and pore progenitor migration paths and timing, we
generated 3D confocal movies of strains UP2051 (pie-1::mCherry::HIS-58::pie-1utr; his-
72pro::HIS-24::mCherry::let-858utr; GFP::MLS-2) and RW10890 (pie-1::mCherry::HIS-
58::pie-1utr; his-72pro::HIS-24::mCherry::let-858utr; PAL-1::GFP) as previously
described (Murray et al., 2006) on a Leica TCS SP5 resonance-scanning confocal
microscope with 0.5 micron z slice spacing and 1.5 minute time point spacing.
Temperature was 22.5˚C. We used a hybrid blob-slice model and StarryNite (Bao et al.,
87

2006; Santella et al., 2010) for automated lineage tracing and curated the duct, pore and
canal lineages (ABplpaa and ABprpaa) through ventral enclosure (approx. 275 minutes)
with AceTree (Boyle et al., 2006).
Ablations
Laser ablations were performed with a Micropoint Laser Ablation system (Photonic
Instruments, St. Charles, IL) mounted to a Leica DM5500B or Zeiss Axiophot
microscope. The canal cell mother (ABplpappaa) was identified using zuIs143 (ref-
1p::GFP) (Neves and Priess, 2005). Successful ablation was confirmed by the absence
of the canal cell as assessed by DIC and either vha-1p::GFP or AJM-1::GFP patterns.
For more detailed description of ablation technique, see Appendix 4.
Immunostaining
Embryos were permeabilized by freeze-cracking and fixed in methanol as described in
Appendix 4 and incubated with primary antibodies overnight at 4C and with secondary
antibodies for 2hrs at room temperature. The following antibodies were used:
preadsorbed rat anti-MLS-2 (CUMCR6; 1:400) (Jiang et al., 2005) goat polyclonal anti-
GFP (Rockland; 1:50), rabbit polyclonal anti-DLG-1 (1:50 to 1:100) (Segbert et al.,
2004). All secondary antibodies were from Jackson ImmunoResearch Laboratories and
were used at a dilution of 1:50 to 1:200.
.
88

Acknowledgements
We thank Matthew Buechner, Helen Chamberlin, Iva Greenwald, Min Han, Jeff Hardin,
Jun Liu, Jim Priess, David Raizen, Paul Sternberg and the Caenorhabditis Genetics
Center (U. Minnesota) for providing strains, Craig Stone and Helin Shiah for generating
reporters, and Iva Greenwald and John Yochem for helpful discussion and comments on
the manuscript. TEM of embryo morphogenesis was originally done in collaboration with
Ed Hedgecock. This work was funded by NIH GM58540 to M.S. I.A. and V.M. were
supported in part by T32 GM008216.
89
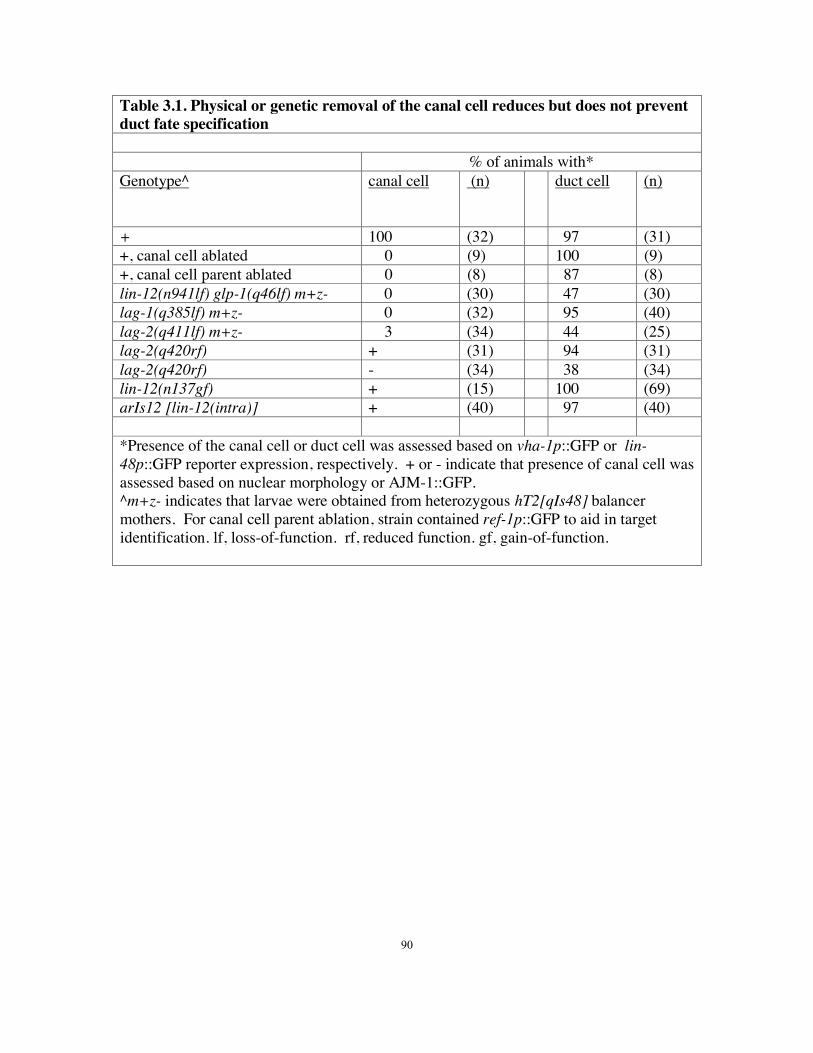
Table 3.1. Physical or genetic removal of the canal cell reduces but does not prevent duct fate specification
*htiw slamina fo %
Genotype^ llec lanac (n)
duct cell (n)
+ 100 (32) 97 (31) +, canal cell ablated 0 (9) 100 (9) +, canal cell parent ablated 0 (8) 87 (8) lin-12(n941lf) glp-1(q46lf) m+z- 0 (30) 47 (30) lag-1(q385lf) m+z- 0 (32) 95 (40) lag-2(q411lf) m+z- 3 (34) 44 (25) lag-2(q420rf) + (31) 94 (31) lag-2(q420rf) - (34) 38 (34) lin-12(n137gf) + (15) 100 (69) arIs12 [lin-12(intra)] + (40) 97 (40)
*Presence of the canal cell or duct cell was assessed based on vha-1p::GFP or lin-48p::GFP reporter expression, respectively. + or - indicate that presence of canal cell was assessed based on nuclear morphology or AJM-1::GFP. ^m+z- indicates that larvae were obtained from heterozygous hT2[qIs48] balancer mothers. For canal cell parent ablation, strain contained ref-1p::GFP to aid in target identification. lf, loss-of-function. rf, reduced function. gf, gain-of-function.
90

Figure Legends
Figure 3.1: Timeline of excretory system development
(A) Schematics of excretory canal cell (red, ABplpappaap), duct (yellow, ABplpaaaapa),
G1 (blue, ABprpaaaapa), G2 (green, ABplapaapa) and W (green, ABprapaapa) at
different developmental stages, based on Sulston et al. (1983), prior electron microscopy
(Stone et al., 2009), and this work. Dark black lines indicate apical junctions. Dotted
line, duct auto-fusion. Arrow, pore autocellular junction. Arrowhead, duct-canal cell
intercellular junction. Bracket, duct cell body. Not shown are the non-essential excretory
gland cells, which also connect to the duct-canal junction (Nelson et al., 1983; Nelson
and Riddle, 1984). (B-E) Progressively older ventral enclosure stage embryos. (B-C, E)
Ventral views. GFP::MLS-2 marks the presumptive duct and G1 pore nuclei. DLG-
1::GFP marks epidermal cell junctions in B and E, which are confocal projections. (B)
The presumptive duct and G1 initially lack junctions. (C) The presumptive duct is closer
to the canal cell than is the presumptive G1. (D) Transmission electron micrograph of a
wild-type embryo at similar stage as C, with cells pseudo-colored as in A. Transverse
anterior view. The presumptive duct and G1 have met at the ventral midline. The duct
makes extensive contact with the canal cell, while G1 is excluded. No epithelial junctions
or lumen are detectable. (E) G1 moves ventrally. Asterisk indicates site of future G1
pore opening between G2 and W epidermal cells (see also Fig. S1). (F-L) Left lateral
views. (F) 1.5-fold stage embryo immunostained for DLG-1, showing newly formed
autocellular junctions (inset) just prior to duct auto-fusion. (G-J) L1 larvae. Box in G'
indicates region magnified in H. AJM-1::GFP marks junctions. The duct no longer has
91

an autocellular junction. (I) lin-48p::mcherry marks the duct. (J) dct-5p::mcherry marks
the duct and G1 pore in early L1 and (K) the duct and G1 in late L1 after G1 withdrawal
and G2 entry. (L) Adult canal cell marked with vha-1p::GFP. Note that canal cell
elongates extensively. Data contributed by all authors.
Figure 3.2: let-60/Ras promotes the duct vs. G1 pore fate
(A-J') AJM-1::GFP (left column) and lin-48p::GFP or (C') dct-5p::mCherry (middle
column) expression in L1 larvae of the indicated genotypes. Lateral views, with
schematic interpretations (right column) and symbols as in Fig. 1. Colors represent lineal
identity, not fate.
Mutants with reduced signaling usually have 2 pore-like cells with autocellular junctions,
but as fluid (carat) accumulates during L1 (C), large junctional rings (asterisk) are
common. In (C'), white arrows indicate two stacked pore-like cells. Mutants with
increased signaling have a seamless binucleate duct that connects to the ventral
epidermis. Scale bar 2 m. (K, L) Quantification of marker phenotypes. Note that some
mutants with “0 G1” have defects in cell stacking and tubulogenesis rather than in cell
fate specification (see Fig. 4). Data contributed by M.S., K.P., K Howell, K. Huang, and
I.A.
Figure 3.3. lin-3/EGF, let-23/EGFR and lin-12/Notch reporter expression in the
excretory system
(A, C) LET-23::GFP is expressed in the presumptive duct and G1 pore at ventral
enclosure (A) and in the duct (bracket) at 3-fold (C). (B, D) lin-3p::GFP is expressed in
92

the canal cell from ventral enclosure (B) through L1 (D). (E) LIN-12::GFP is expressed
in the presumptive G2 and W but not in the presumptive duct or pore at ventral enclosure.
In A, C and E, embryos were co-stained with anti-GFP and either anti-MLS-2 or anti-
AJM-1 to mark the duct and G1 pore (n > 10 each). Scale bars, 5 m. Data contributed
by V.M. and I.A.
Figure 3.4. let-60/Ras hypomorphs reveal defects in cell competition and stacking
(A-B, E-F) Wild-type. (C-D, G-L) let-60(n2021rf). (A, C) AJM-1::GFP in 3-fold
embryos, ventral view. Lines indicate ventral junctions between the presumptive G1 or
duct and the epidermis. (E, G, I, K) AJM-1::GFP and dct-5p::mCherry in early L1s,
lateral view. Asterisks indicate ventral cells with neither duct-like nor pore-like
morphology. In let-60(n2021rf) mutants, the presumptive duct and G1 adopt adjacent
positions in the epidermis (C-D) and one cell usually reaches from the ventral epidermis
to the canal cell (G-J). The lineal identity of this cell is unknown and may be variable.
(K) Quantification of adjacent defects in early L1 larvae from heterozygous vs.
homozygous mutant mothers. Animals with no G1 pore autocellular junction or with a
single autocellular junction that stretched from the ventral epidermis to the canal junction
were scored as "adjacent". Data contributed by M.S. and I.A.
Figure 3.5: The canal cell is required for duct and G1 pore stacking and
tubulogenesis
93

(A-I, L) AJM-1::GFP. D, G, H also contain lin-48p::GFP. Scale bars, 2 m. (J, K)
Schematic diagrams. (A-C) early 3-fold embryos, ventral view. (D-F) Early L1s, ventral
view. (G-L) Early L1s, lateral view. In wild-type (A), the G1 pore contacts G2 and W in
the ventral epidermis. In lag-1(RNAi) (B) or lin-12(n941) glp-1(q46) double mutants (C,
F, I), the presumptive duct and G1 pore (lines) both contact the epidermis and lack
autocellular junctions. In lag-2(q420rf) mutants (D, E, G, H), presence of a canal cell (D,
G) correlates with normal duct and G1 pore morphology. aff-1(tm2214) (E) has no
impact on the lag-2(q420rf) phenotype. (L) Ablation of the canal cell mother eliminates
the G1 pore autocellular junction. (M) Quantification of junction phenotypes in early L1
larvae. Animals with no G1 pore autocellular junction were scored as "adjacent". Data
contributed by V.M. and M.S.
Figure 3.6. sos-1 temperature shift experiments reveal continued requirements
during duct morphogenesis and differentiation.
(A) sos-1(cs41ts) lethality at 25˚ C is rescued by let-60(n1046gf) or lin-1(e1275lf). n>50
for each genotype. (B-D) sos-1(ts) animals bearing AJM-1::GFP or lin-48p::GFP markers
were up-shifted or down-shifted at the stages indicated. n>20 for each time point. sos-1
is required prior to the 1.5 fold stage to promote lin-48p::GFP duct marker expression (B)
or duct auto-fusion (C). (D) The sos-1(ts) temperature-sensitive period (TSP) for lethality
extends from the bean stage of embryogenesis to L2. The majority of animals upshifted
prior to L2 arrested with excretory abnormalities (see C,E,F). Animals upshifted during
L2 displayed a scrawny phenotype similar to that reported for egl-15/FGFR mutants
(DeVore et al., 1995; Roubin et al., 1999). (E',F') Fluid (carats) accumulated in or near
94

the duct in 3-fold upshifted sos-1(ts) animals, while AJM-1::GFP (E) and lin-48p::GFP
(F) patterns were unaffected in these same animals. Scale bar, 2 m. Data contributed by
I.A.
Figure 3.7: G1 withdrawal and G2 entry can occur independently
AJM-1::GFP in L4 larvae. (A, D, G, J) lateral views. (B, E, H, K) ventral views. (C, F, I,
L) Schematic diagrams. In wild-type (A-C), G2p forms the pore. In let-60(n1046gf)
mutants (D-F), G2p usually wraps around the base of the duct. In lin-12(n941lf) mutants
(G-I), the duct attaches directly to the ventral epidermis after G1 withdrawal. In lin-
12(n137gf) mutants (J-L), the extra G2p cell wraps around the ventral base of the pore.
Lines indicate ventral junctions with the epidermis. Scale bars, 2 m. (M) Quantification
of junction phenotypes. Data contributed by M.S.
Figure 3.8: Transmission Electron Micrographs of ventral enclosure stage embryo
Thin sections eighteen (A) and thirty-eight (B) sections posterior of the section shown in
Fig. 3.1D. The canal cell, duct, G1, G2 and W are pseudo-colored as in Fig. 3.1A. hyp7
cells ABpl/raapppa have already met at the ventral midline and fused, but G2 and W have
not yet met at the midline, leaving an open ventral cleft through which parts of the canal
cell, duct and G1 are exposed on the outside of the embryo. The canal cell and duct
remain in contact over more than forty sections in this embryo, but the canal cell and G1
never touch. Data contributed by D.H. and C.N.
95
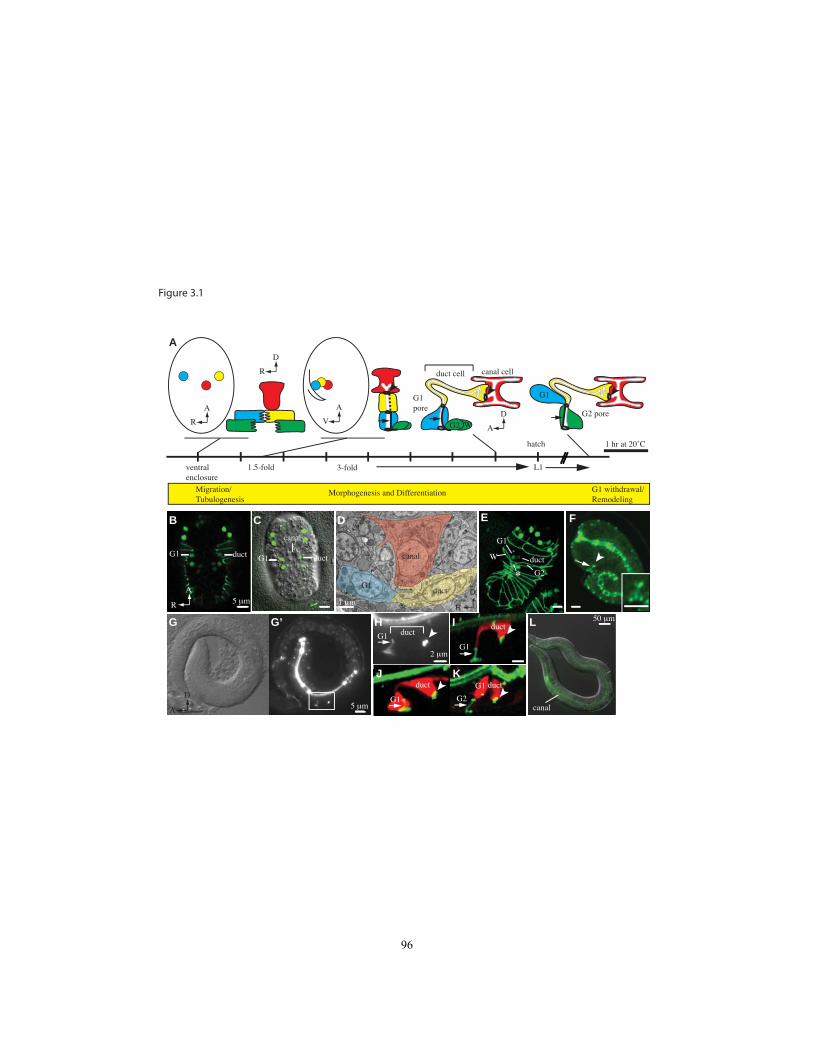
ventralenclosure
1.5-fold 3-fold L1
hatch
A
Migration/Tubulogenesis
Morphogenesis and Differentiation G1 withdrawal/Remodeling
A
R
A
V
duct cell
G1
canal cell
G1pore
G2 poreD
AG2
H I ductG1
F
F F
canalcell
G12 μm
5 μm
B
5 μm
G G’
ductG1
duct FF
canacanananaanaallllllcellcelllcanal
L
JG1
duct
G2
JJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJJG1G11GGG1G1G11G1G111111G111111G11111G
ducdducducdddducducductductductducucuctdududucdddududucduucduducducduducdducducduucuctduddduudududucdduduuudududuuudduucucudduuuuddduuccddddddduuucdddducccddddudd ccdddduc
GGGGGGG2G22GGGGGGGGGGGGGGGGGGGGGGGGGGGGGG2G2G2G2G22GG
KductG1
G2
IIIIIIIIIIIIIIIIIIJ
G1
duct
C
G1 duct
canal
50 μm
A
R
ductG1
canal
E
D
R
D
W
1 hr at 20˚C
1 μm
D
A
D
R
G1
duct
G2
W
*
Figure 3.1
96
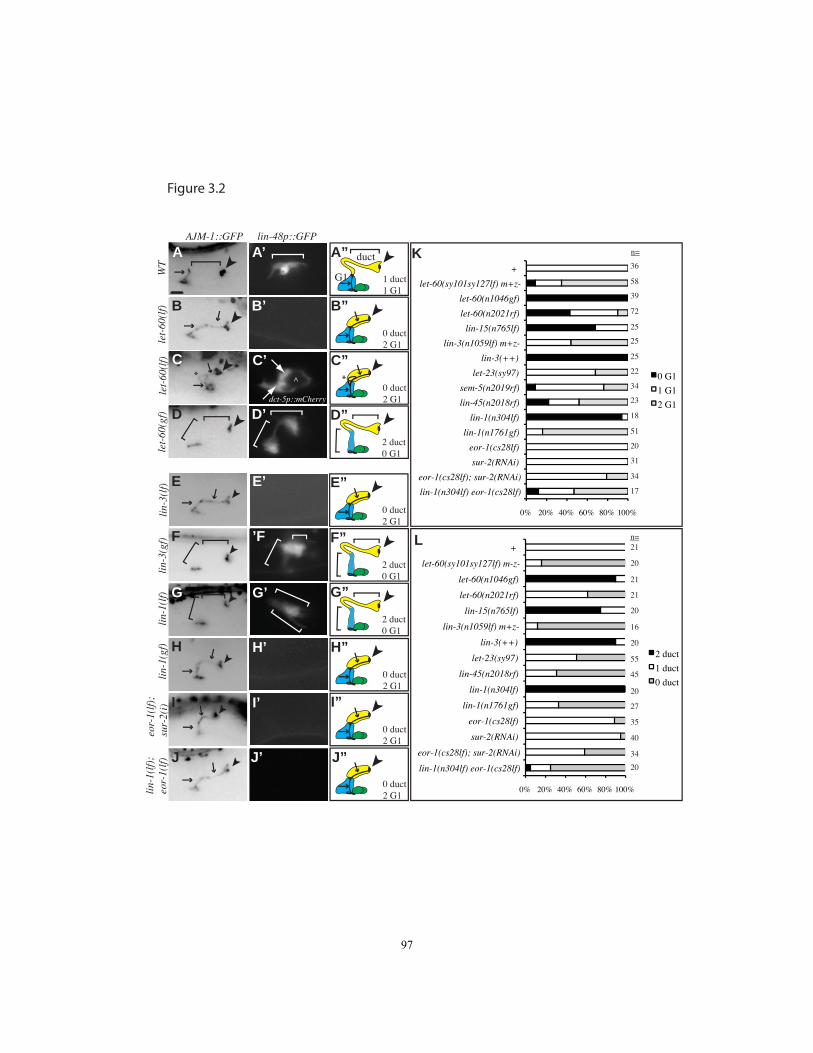
0% 20% 40% 60% 80% 100%
+
let-60(sy101sy127lf) m+z-
let-60(n1046gf)
let-60(n2021rf)
lin-15(n765lf)
lin-3(n1059lf) m+z-
lin-3(++)
let-23(sy97)
sem-5(n2019rf)
lin-45(n2018rf)
lin-1(n304lf)
lin-1(n1761gf)
eor-1(cs28lf)
sur-2(RNAi)
eor-1(cs28lf); sur-2(RNAi)
lin-1(n304lf) eor-1(cs28lf)
0 G1
1 G1
2 G1
0% 20% 40% 60% 80% 100%
+
let-60(sy101sy127lf) m-z-
let-60(n1046gf)
let-60(n2021rf)
lin-15(n765lf)
lin-3(n1059lf) m+z-
lin-3(++)
let-23(sy97)
lin-45(n2018rf)
lin-1(n304lf)
lin-1(n1761gf)
eor-1(cs28lf)
sur-2(RNAi)
eor-1(cs28lf); sur-2(RNAi)
lin-1(n304lf) eor-1(cs28lf)
2 duct
1 duct
0 duct
A A’
D”
B B’
D D’
WT
let-
60(l
f)le
t-60
(gf)
let-
60(l
f) C C’
2 duct0 G1
*
AJM-1::GFP
dct-5p::mCherry
^
A”
1 duct1 G1
G1
duct
B”
0 duct2 G1
C”
0 duct2 G1
*
lin-48p::GFP
lin-
3(lf
)
G
E’
lin-
1(lf
)li
n-1(
gf)
lin-
3(gf
)eo
r-1(
lf);
sur-
2(i)
E
F
H
I
’F
G’
H’
I’
lin-
1(lf
);eo
r-1(
lf) J
F”
2 duct0 G1
G”
2 duct0 G1
E”
0 duct2 G1
H”
0 duct2 G1
I”
0 duct2 G1
J”
0 duct2 G1
K
L n=
n=
58
36
39
72
25
25
25
34
23
18
51
20
31
34
17
22
20
21
21
21
20
16
20
45
20
27
35
40
34
55
J’20
Figure 3.2
97
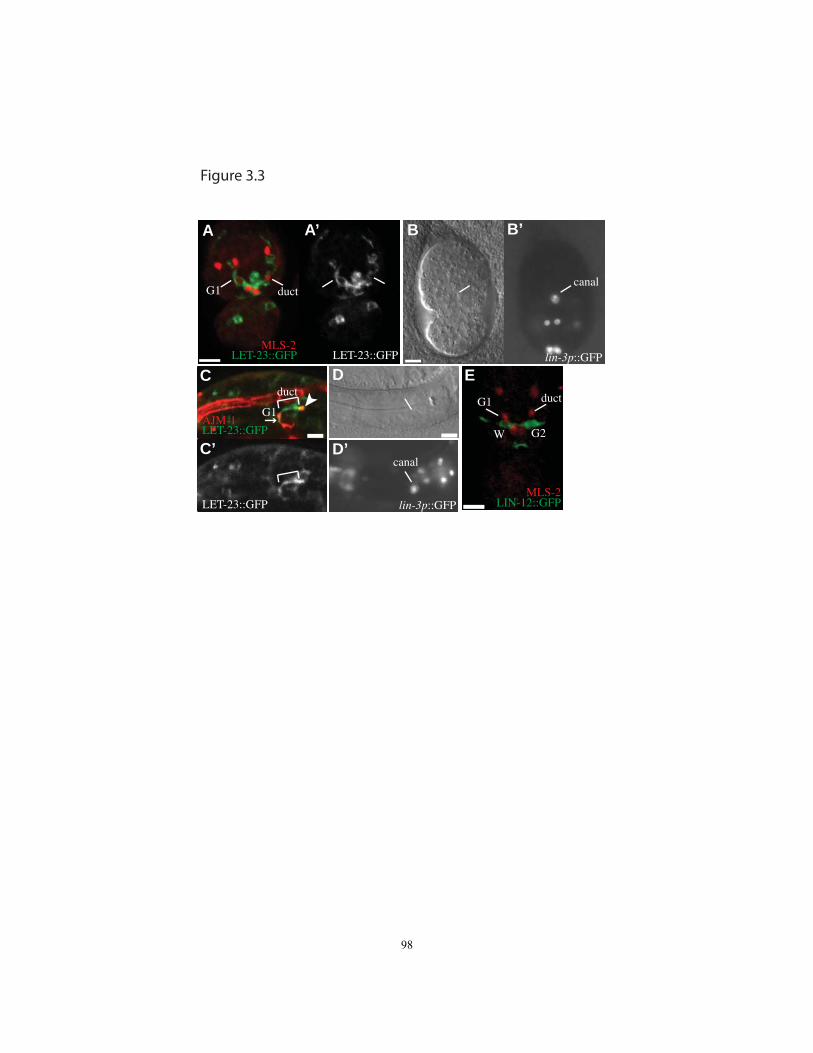
A A’
C
B B’
LET-23::GFP LET-23::GFP lin-3p::GFPMLS-2
C’
D
D’
E
MLS-2LET-23::GFP LIN-12::GFPlin-3p::GFP
LET-23::GFPAJM-1
G2W
G1 ductcanal
G1
duct ductG1
canal
Figure 3.3
98
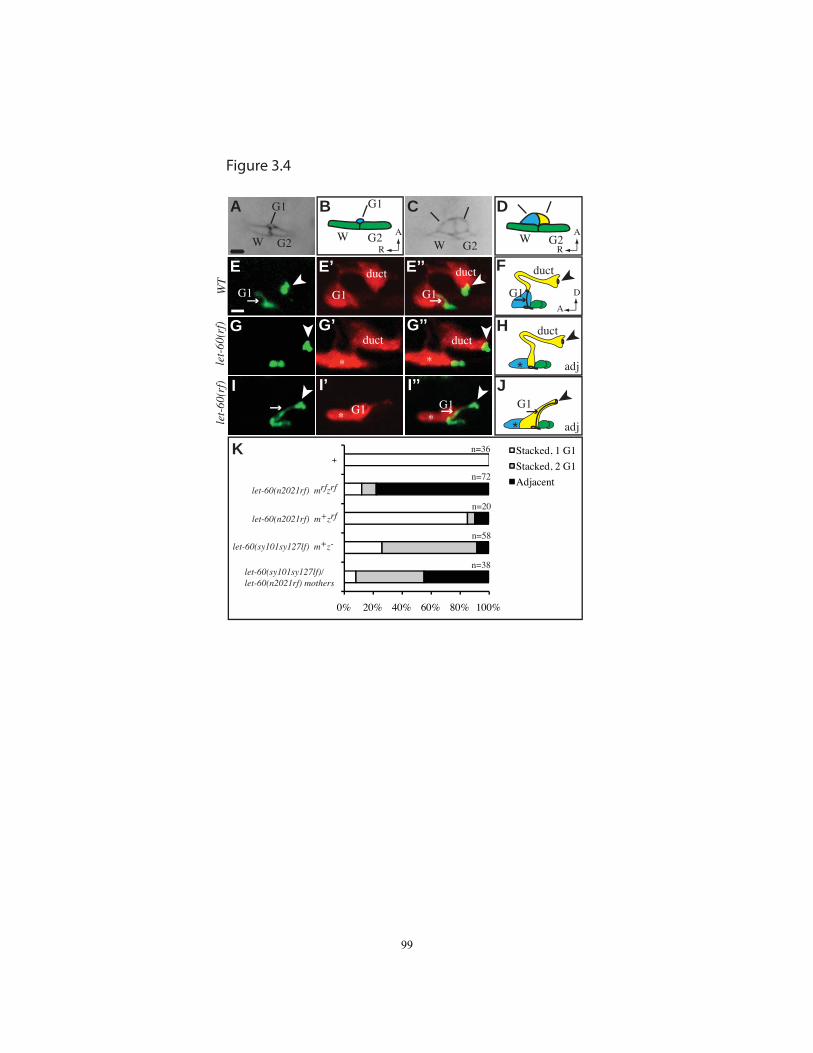
Stacked, 1 G1
Stacked, 2 G1
Adjacent
0% 20% 40% 60% 80% 100%
G1
W G2
AA
R
CB D
W G2
G1
G2WW G2
E E’ E’’G1
duct duct
G1G1
F
G G’ G’’ H
J
D
A
I’ I’’IG1
duct duct
G1
A
R
K
*
* *
*
G1
duct
*
duct
G1
*n=36
n=72
n=20
n=58
n=38
WT
let-
60(r
f)le
t-60
(rf)
let-60(n2021rf) mrfzrf
let-60(n2021rf) m+zrf
let-60(sy101sy127lf) m+z-
let-60(sy101sy127lf)/let-60(n2021rf) mothers
+
adj
adj
Figure 3.4
99

0% 20% 40% 60% 80% 100%
++
lin-12(lf) glp-1(lf) m+z-lag-1(RNAi)
lag-2(rf)lag-2(rf)
aff-1; lag-2(rf)lin-12(lf) m+z-lin-12(lf) m+z-glp-1(lf) m+z-
lin-12(gf)
adjacent stacked
G1
W G2
A B
G H
D
K
E I
I
F
J
C
L
WT lag
WT, canal-
G1
duct
D
A
A
R
D
A
A
R
M3652453134212182015
n
canal
+
+
+
-
+
-
-
-
-
+
-
canal+ canal- canal-
vent
ral
vent
ral
late
ral
late
ral
Figure 3.5
100
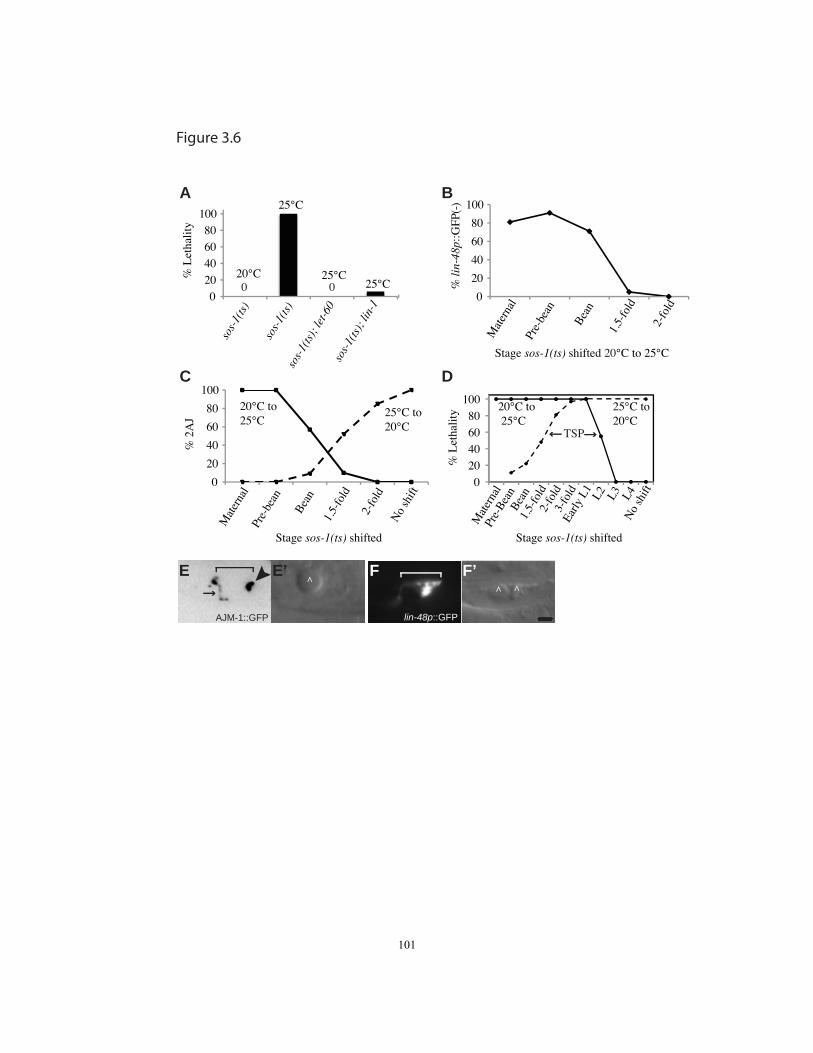
0
20
40
60
80
100
Mat
erna
l Pr
e-be
an
Bea
n 1.
5-fo
ld
2-fo
ld
No
shift
% 2
AJ
Stage sos-1(ts) shifted
20°C to 25°C
25°C to 20°C
C
0
20
40
60
80
100
Mat
erna
l Pr
e-be
an
Bea
n
1.5-
fold
2-fo
ld
% li
n-48
p::G
FP(-
)
B
Stage sos-1(ts) shifted 20°C to 25°C
0 20 40 60 80
100 so
s-1(
ts)
sos-
1(ts
) so
s-1(
ts);
let-6
0so
s-1(
ts);
lin-
1
% L
etha
lity
A
0 0
25°C
25°C25°C
20°C
0 20 40 60 80
100
Mat
erna
l Pr
e-B
ean
Bea
n 1.
5-fo
ld
2-fo
ld
3-fo
ld
Early
L1
L2
L3
L4
No
shift
% L
etha
lity
D
TSP
25°C to 20°C
20°C to 25°C
lin-48p::GFPlin-48p::GFP
F’E E’ F
AJM-1::GFP
^ ^^
Stage sos-1(ts) shifted
Figure 3.6
101
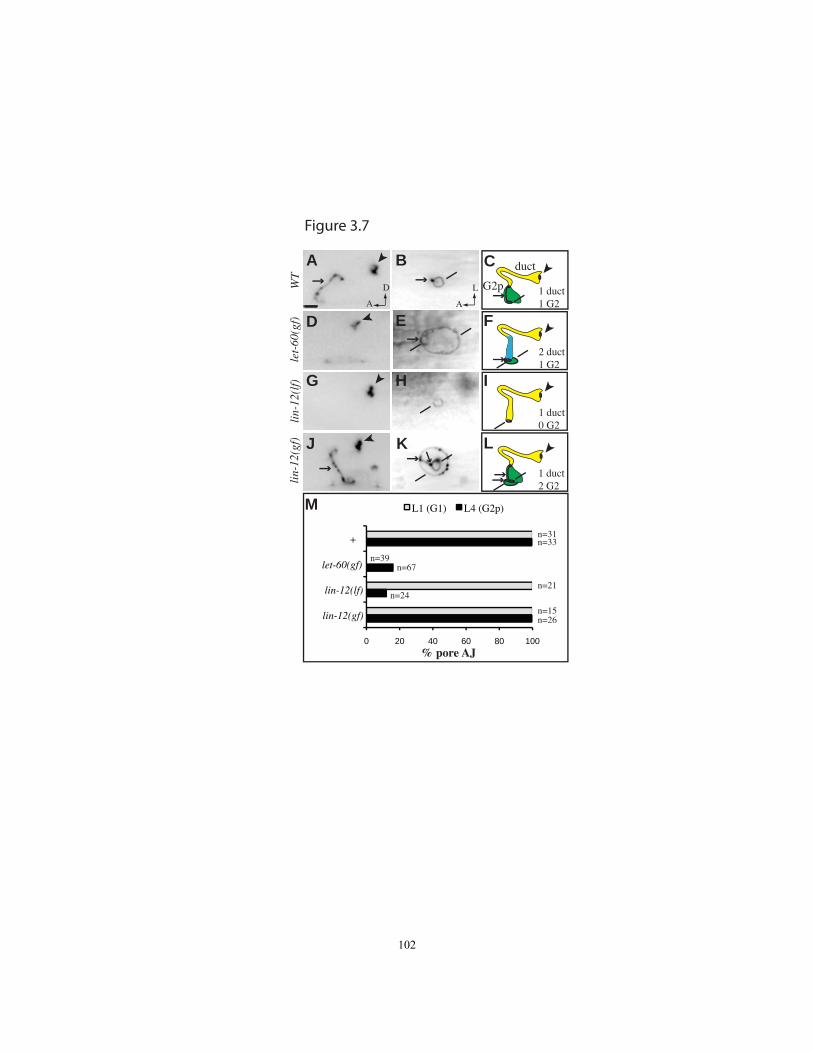
0 20 40 60 80 100
+
let-60(gf)
lin-12(lf)
lin-12(gf)
L1 (G1) L4 (G2p)
% pore AJ
n=31n=33
n=21
n=15n=26
n=39n=67
n=24
let-
60(g
f)li
n-12
(lf)
WT
lin-
12(g
f)1 duct1 G2
2 duct1 G2
1 duct0 G2
A B
D E
G H
J K
F
I
D
A
L
A
C
1 duct2 G2
L
duct
G2p
M
Figure 3.7
102

canal
ductG1
G2
W
canal
ductG1W
G2
hyp7
hyp7ventralcleft
B
A
1 μm
1 μm
D
R
Figure 3.8
103

Chapter 4
Discussion: The role of LET-4 in the apical ECM and junctions, and approaches to
identify LET-4 interacting partners.
104

Introduction
This work helps establish the excretory system as a new model to ask important
questions about epithelial tubes. With my colleagues, I have explored the role of Notch
and Ras in the excretory system, and proposed a biased competition model for duct cell
fate. I have characterized the pattern of conserved junctional proteins in the excretory
system, such as the PAR complex proteins and cadherin-catenin complex. This work has
demonstrated the presence of conserved proteins with roles common to many epithelia,
and therefore supports the excretory system as a good model for epithelial development,
luminal maintenance, and apical ECM formation and maintenance. I have also
demonstrated that loss of apical transmembrane protein LET-4 results in defects in both
apical ECM organization and junction integrity, suggesting an unexpected link in the
maintenance of these two epithelial cell structures.
Loss of LET-4 results in junctional and apical ECM defects
My let-4 analysis has characterized two classes of phenotypes: ECM phenotypes and
junctional phenotypes. The ECM defects are: (1) detached inner eggshell layer in 1.5-
fold embryos, (2) extra-embryonic cytoplasts in 3-fold let-4 embryos, (3) in the duct cell,
the cuticle becomes detached from the apical face of the cell in late in 3-fold, and (4)
increased L1 cuticle permeability. There are two epithelial junction phenotypes: (1) the
pore autojunction, and duct/pore connection break late in 3-fold, and (2) in let-4 RNAi-
105

treated sym-1 worms, epidermal junctions break late in 3-fold. My analysis has found no
evidence for defects in initial apical specification or junction formation in these cells.
Considering these phenotypes, I propose that LET-4 is required for proper apical ECM
organization in the excretory system and epithelial cells. In the absence of LET-4 in the
apical ECM, epithelial junctions cannot be maintained as a secondary consequence of
apical ECM disorganization. Based on this proposal, I have generated 3 models that
could explain LET-4‟s indirect role in jucntional maintenance (Fig. 4.1). First, a
disorganized apical ECM could result in the apical ECM failing to absorb force, causing
the junctions themselves to be exposed to more force (Fig 4.1A). Second, the apical
ECM may be playing a role in signaling pathway required for junctinal maintenance (Fig.
4.1B). Third, a still unknown component of the apical ECM could directly stabilize
junctional proteins (Fig. 4.1C).
While I cannot rule out the possibility that loss of LET-4 is affecting the junctions
independent of the extracellular matrix, we favor the model that the ECM is the primary
defect, and the junctions are affected because of ECM defects. The junction breakage
may be due to extra force sustained by the junctions in the absence of a wild-type apical
ECM. We favor this model for several reasons. First, LET-4::GFP reporter transgenes
are not enriched at the junctions like other junctional components are. Second, ECM
defects are more penetrant than junctional defects. The defects in the apical ECM such as
inner eggshell layer detachment, presence of extra-embryonic cytoplasts, and cuticle
permeability are highly penetrant, and junctional defects like embryonic rupture are rarer,
except in the sym-1 background. In the excretory system, though, detachment of the
106

cuticle and pore autojunction breakdown are both highly penetrant. Third, the defect in
the sheath occurs at 1.5-fold, before elongation, and before any junctional phentoypes are
apparent.
I have identified a system with a correlation between apical ECM components
and junctional defects, adding to the growing field of apical ECM studies. This is
different from the previously described connections related to MUC1 in mammals and
Piopio and Dpy in Drosophila (Discussed in Chapter 1). MUC1 is an apical ECM
protein, and mislocalization of it results in polarity defects, leading to EMT (Roy et al.,
2001; Kufe et al., 2009). The let-4 defect is distinct from this because in let-4, I‟ve
detected no defect in apical polarity. Also, MUC1 has been demonstrated to signal
through the cytoplasmic domain (Kufe et al., 2009), and I‟ve found that the LET-4
cytoplasmic domain is dispensable for function. The authors of the Piopio and Dumpy
study propose that the apical ECM serves as a structure to guide migrating cells as they
intercalate and form new autocellular junctions, and the absence of the normal ECM
results in failure of proper junction formation (Jazwinska et al., 2003). In let-4 mutants,
cells are not migrating relative to each other when I observe junctional phenotypes, and
no new junctions are forming. The tubes of the excretory system have already formed
and elongated by the time the defect is apparent. The junctions of this system may
require maintenance which does not occur due to the absence of LET-4 in the apical
ECM.
The let-4 excretory system defect, consisting of duct cell lumen collapse, duct
detachment from the pore, and loss of pore autojunction occurs late in embryogenesis,
107

after deposition of the cuticle (Chapter 2). The excretory system does not have a defect
prior to 3-fold, or at later larval molts, indicating that LET-4 acts at a very specific time
in the excretory system. Similarly, I have found no evidence for any defect in the
excretory system apical ECM until late in 3-fold stage. This suggests the maintenance of
the apical ECM is a regulated process, with requirements for specific proteins at specific
times. Because these excretory system defects are not apparent until 3-fold stage, this
could mean that LET-4 has a role in apical ECM at the cuticle stage.
In the excretory system, I see the specific junctions being affected, which argues
against let-4 mutants having a general junctional defect. If there were a general defect in
junctional integrity in let-4 mutants, I would expect to see detachment between the pore
and the outside, or the duct and canal cell. The duct/pore junction always becomes
disconnected, while the duct/canal cell junction stays intact. The data suggest that
specific junctional structures are sensitive to an abnormal ECM.
The epidermal rupture phenotype observed in let-4 RNAi treated sym-1 embryos
has revealed a different type of epidermal rupture phenotype than has been previously
described. The let-4 embryonic phenotype is reminiscent of other mutant phenotypes,
such as par-6 (Totong et al., 2007), hmr-1/e-cadherin (Costa et al., 1998), ajm-1 (Koppen
et al., 2001), ifb-1 (Woo et al., 2004) or spektraplakin locus genes vab-10A and vab-10B
(Bosher et al., 2003). These other mutants, however, have mutated junctions or
cytoskeletal components, and rupture early in elongation. I‟ve found that let-4 mutants
have an epidermal defect that is very different because the junctions and cytoskeletal
components appear to be patterned normal by TEM, markers, and
108

immunohistochemistry, until the junction breakage at 3-fold stage (Chapter 2, Appendix
2a).
Why does the excretory system phenotype present itself when it does? Perhaps
the junctions of the excretory system undergo a remodeling at the time of cuticle
secretion in 3-fold stage, making them vulnerable to breakage due to an incomplete ECM
at this stage. Although junctional proteins are known to be dynamic (Acloque et
al.,2009; Baum and Georgiou, 2011; St Johnston and Sanson, 2011), the replenishing of
junctions in the excretory system has not been described.
Many questions remain about the role of LET-4 and its relationship with the
apical extracellular matrix. The components of the embryonic sheath in the embryo and
the extracellular matrix in the lumen of the duct and pore cell still remains mysterious. A
deeper understanding of the molecular partners of let -4 will involve a closer
characterization of the extra-cellular matrix and identification of let-4 interacting
proteins. Below, I list more outstanding questions, describe experiments performed to
address these questions, and discuss my conclusions.
Why are there so many eLLRons?
The extra-cellular Leucine-Rich Repeat only (eLLRon) family is extensive.
There are 17 members in C. elegans, 35 in Drosophila, 52 in Mouse and 57 in human
(Dolan et al., 2007). This large number of members in such a specific protein category
suggests redundancy among these proteins. Redundancy is not unprecedented for
components of the C. elegans extracellular matrix.
109

There are about 170 collagens in the C. elegans genome (Page and Johnstone,
2007), but only a handful have been found to have mutant phenotypes. Often, those
phenotypes are only mild, such as missing alae, roller locomotion defects, or a shortened
body length, suggesting that worms can form a sufficient cuticle even without the most
important cuticular collagens.
A family of extracellular Leucine-Rich Repeat proteins which may provide
insight into how LET-4 may function in an ECM is the Small Leucine-Rich
Proteoglycans (SLRPs). SLRPs consist of LRR motifs in a protein core, and covalent
linkage to glycosaminoglycan side chains. SLRPs modulate collagen matrix assembly
(Kalamajski and Oldberg, 2010), and bind collagen with their LRR domains (Shimizu-
Hirota et al., 2004). SLRPs are ECM proteins that are required for collagen matrix
assembly. They are hypothesized to work as a type of scaffold in the apical ECM, to
regulate the formation of collagen structures. (Kalamajski and Oldberg, 2010). SLRPs,
therefore are not affecting the production of the ECM components, but the proper
ordering of the components. Perhaps LET-4 is performing a similar role, and affecting
the organization of some collagens in the cuticle.
Other eLLRons have been shown to have a role in cell-cell adhesion in neurons.
Although the let-4 defects described here are in epithelial cell junctions, it is intriguing
that eLLRons have been demonstrated to have roles at synaptic junctions. In mammals,
LRRTM1-3 are required for excitatory synapse formation, and bind neurexins (deWit et
al., 2009; Ko et al., 2009; Siddiqui et al., 2010). The Drosophila protein Capricious is
well characterized eLLRon known to have a role in neurons. Caps is required for
110

targeting of embryonic motor neurons to specific muscles, and for proper synaptic
formation; some of these roles require the cytoplasmic domain (Taniguchi et al., 2000,
Shinza-Kameda et al., 2006, Mao et al., 2008, Kohsaka and Nose 2009, Shishido et al.,
2009). Notably, Caps is also required in the trachea for branch outgrowth (Krause et al.,
2006). Although other eLLRons have roles in neurons, I‟ve found no evidence for
transcription or translation of let-4 in neurons.
Why are both LET-4 and EGG-6 required in the excretory system?
A better comparison of the phenotype of let-4 and egg-6 requires further
investigation of the egg-6 phenotype. Molecular characterization of additional junctional
components of the excretory system will be useful. Also, TEM of the egg-6 mutant
phenotype will reveal the state of the duct lumen and inner eggshell layer. The AJM-
1::GFP junctional phenotype and duct detachment phenotypes of let-4 and egg-6 are
similar, so the duct luminal phenotype and inner eggshell layer phenotype could be very
similar as well.
LET-4 and EGG-6 are similar proteins with similar phenotypes; are they
interchangeable? To investigate this, ongoing experiments are being performed to test for
rescue of the egg-6 phenotype with let-4 overexpression, and vice versa. A transgene
containing let-4cDNA driven by the lpr-1 promoter, which rescues the let-4 defect, failed
to rescue egg-6 larval lethality (J. Parry, personal communication). This suggests that the
two proteins, though similar, are serving non-redundant functions in the excretory
111

system. These data highlight the diversity of functions of the eLLRons, even in the same
tissue.
What ECM components could LET-4 be interacting with?
An initial candidate for a let-4-interacting protein was LPR-1, a secreted lipocalin.
Many aspects of the phenotype of lpr-1 are very similar to let-4. In addition to a phasmid
dye-filling defect, both mutants have a highly, but incompletely, penetrant L1 larval
lethal phenotype which results from a duct cell lumen connectivity defect (Stone et al.,
2009). The lumen connectivity defect in lpr-1 was observed with a TEM analysis
performed on the 3-fold stage lpr-1 mutants, therefore it is currently unknown if the
lumen is initially normal like in let-4 at the 1.5-fold stage. In the lpr-1 3-fold TEM
analysis, though the duct cell was still in contact with the pore cell, in contrast to the let-4
mutant, in which the two cells were no longer connected. As a secreted protein, lpr-1
may play a role in the secretory pathway. Alternatively, LPR-1 could be secreted, and
interact with the ECM of the duct cell. LPR-1 could even be a ligand of LET-4.
Leucine-rich repeats interact with a variety of targets, including secreted ligands (Kobe
and Deisenhofer, 1994, Bella et al., 2008, Kawai et al., 2011). To look for evidence of
let-4 and lpr-1 interaction, I analyzed the timing of fluid accumulation in lpr-1(cs73) I;
let-4(mn105) X double mutants. If let-4 and lpr-1 were involved in the same pathway, an
earlier defect may have occurred in these double mutants, but I found no enhancement of
the onset of the first visible distortion in the excretory system (Table 4.1). This was not
112

an exhaustive analysis, but I have determined that the lumen defects don‟t result in an
earlier phenotype. Given the common appearance of the first fluid distortion visible it in
the late 3-fold stage, it may be that this is the time at which the excretory system starts to
function, and thus the first time fluid passes through the excretory system. A better
comparison between the luminal defects will require further TEM of lpr-1 mutant
embryos.
Although the above experiments did not detect an interaction, lipocalins remain a
tempting candidate for LET-4 ligands. To further explore the potential of lipocalins to
have a role in excretory system development, RNAi knockdown of other C. elegans
lipocalin proteins was performed in wild-type and lpr-1 background. We used the
temperature sensitive allele of lpr-1(h276) for this analysis. We performed a feeding
RNAi experiment with lpr-2, lpr-3, lpr-4, lpr-5, lpr-6 and lpr-7. None of the lipocalins
produced significant embryonic or larval lethality in wild-type RNAi-treated worms, and
none of the lipocalins strongly enhanced the lpr-1 phenotype. (Table 4.2) This data is not
conclusive, however, because RNAi is known to have variable levels of efficacy, and
seems to be particularly weak in the excretory system based on experiments performed in
our lab.
To identify proteins and pathways interacting with let-4, I tested several genes for
a genetic interaction with let-4. The embryonic defect which results from let-4 RNAi in a
sym-1 background is similar to reported embryonic phenotypes for junctional and
cytoskeletal mutants (Costa et al., 1998, Koppen et al., 2001, Bosher et al., 2003, Woo et
al., 2004, Totong et al., 2007), therefore, I chose candidates for the genetic interactions
113

that were junctional and cytoskeletal components. Many of the alleles tested produce a
homozygous lethal phenotype. For this analysis, trans-heterozygotes of let-4 and a
candidate allele were generated, and the rate of embryonic plus larval lethality in the next
generation was measured. The expectation is if let-4 and the candidate had a strong
genetic interaction, the rate of lethality would be greater than the additive lethality of two
homozygous genes. If both genes were involved in the same pathway, decreasing the
levels of both genes in the trans-heterozygote could lead to increased lethality. For all the
candidates tested, no enhancement of let-4 embryonic and larval lethality was observed
(Fig. 4.2). However, no increased lethality was observed in sym-1 or mec-8 in the trans-
heterozygote experiment, even though it has been established that RNAi produces an
interaction between let-4 and these two genes (Davies et al., 1999). This suggests that the
trans-heterozygote genetic interaction is not a sensitive assay, or let-4 is not sensitive to
genetic reduction. Below, I list in more detail of the reasoning for choosing specific
candidates for genetic interaction experiments.
mec-8 and sym-1 were chosen because they were previously shown to have
genetic interactions with let-4 (Davies et al,1999, this work). Also, sym-1 is an eLLRon
protein. egg-6 was chosen because it, too is an eLLRon, and because of the similar
phenotype to let-4. We chose hmr-1, the worm homolog of E-cadherin because this
protein is a widely expressed epithelial junctional protein, and has a role in maintaining
epidermal integrity during embryonic elongation (Costa et al., 1998).
The worm‟s protective outer aECM, the cuticle, is made of collagen, and
therefore let-4 could be affecting the ECM by binding collagen. Choosing candidate
114

collagens is a difficult task, because there are about 170 predicted cuticle collagens in C.
elegans, and many are not well-studied (Page and Johnstone, 2007). For this analysis, I
choose to look for a genetic interaction between let-4 and cuticle collagen sqt-3. Of the
characterized collagens, many are known to only be expressed in later larval stages and
adulthood. SQT-3 is the collagen with the earliest reported expression, and earliest
reported phenotype: 2-fold arrest (Novelli et al., 2006).
Defects in the epidermal cells could be a result of a weakened cytoskeleton due to
actin disorganization. To explore genetic interactions between let-4 and actin
components, I performed genetic interaction experiments with act-5, rho-1 and arx-5. C.
elegans has five actin proteins; act-5::GFP has been found to be expressed in the
excretory system and the intestine. In the intestine, it has been reported to be apically
localized (MacQueen et al., 2005), therefore I chose it as a candidate. I chose arx-5
because it produces a necessary subunit of the Arp2/3 complex, which nucleates actin
branching. Depletion of some Arp-2/3 components by RNAi results in enlarged luminal
width, apical accumulation of junctional proteins, and decreased F-actin levels in the
intestine (Bernadskaya et al., 2011). Another gene chosen for this analysis, rho-1, is a
GTPase. It is ubiquitiously expressed, regulates Arp-2/3, and has been associated with
polarity protein localization and actin contractility defects (Schonegg and Hyman 2006;
Motegi and Sugimoto 2006).
I looked for a genetic interaction in two genes coding for proteins that anchor the
cytoskeleton to the apical face of epithelial cells: sma-1 and erm-1. sma-1 is the C.
elegans ortholog of Drosophila βHspectrin. βHspectrin is required for actin organization
115

and cell adhesion in Drosophila. SMA-1 localizes to the apical face of elongating
epithelial cells, and is responsible for actin organization. It is expressed in the canal cell
(McKeown et al.., 1998) and other epithelial cells (Praitis et al., 2005). In sma-1 mutants,
actin is not associated with the apical cell membrane. ERM-1 is an apical protein
affecting junction remodeling in epithelial cells, and is expressed in the canal cell. Ezrin-
radixin-moesin protiens are membrane-cytoskeletal linkers. Perturbation of erm-1 results
in disordered F-actin and then mislocalization of junctions (van Furden et al., 2004;
Gobel et al., 2004).
Epithelial cells are anchored to the basal ECM through transmembrane integrins,
which are anchored to laminins in the basement membrane (Hynes et al., 2009). The
LRR of PKD1 has been shown to interact with ECM components such as collagen,
fibronectin, and laminin in vitro (Malhas et al., 2002). Therefore, we chose intergins and
a laminin as candidates for genetic interaction. C. elegans genes pat-2 and pat-3 are
integrins. (Williams and Waterston, 1994) Depletion of pat-2 pat-3 and talin results in
muscle actin to be disorganized. (Gettner et al., 1995, Lee et al., 2001). The single
laminin-β gene is lam-1 (Kao et al., 2006). Each laminin molecule consists of a β-
subunit, therefore loss of LAM-1 should affect all laminin molecules. RNAi against lam-
1 results in some early embryonic arrest, some post-elongation arrest, and a weak loss of
function allele gives wild-type worms with „unc‟ movement defects.
Because of its roles in junctional maintenance, we chose to test the C. elegans
spektraplakin locus, VAB-10. VAB-10 codes for two isoforms: VAB-10A and VAB-
116

10B; loss of either results in elongation defects. VAB-10A localizes to the
hemidesmosomes, and is associated with intermediate filaments, while VAB-10B
associates with actin (Bosher et al., 2003). This was an enticing candidate for a LET-4-
interacting protein because it has roles in junctional maintenance, and a homolog has a
role in the lumen of unicellular tubes. The Drosophila spektraplakin, short stop, is
required for the proper formation of the lumen of the unicellular terminal cells in the
Drosophila trachea (B. Levi Thesis, 2006). Therefore, I pursued the possibility of an
interaction further, and injected let-4 RNAi into vab-10 mutants (Figure 4.2). However,
like the other genes listed above, there was no genetic interaction detected.
A gene that became interesting to our lab because of its similar phenotype to let- 4
is let-653. let-653 mutants have fluid accumulation in the excretory system, and die as
L1 larvae (Jones and Baille 1995, Buechner et al., 1999). LET-653 is a secreted mucin.
Mucins are glycosylated proteins that contribute to the apical extracellular matrix. LET-
653 is also a ZP-domain containing protein. In Drosophila ZP domain mutants, both
apical ECM attachment and microtubule organization are affected. (Bokel et al., 2005).
The let-653 loss of function phenotype, expression pattern and predicted localization
make it a candidate for a LET-4 interacting protein. To explore this possibility, we tested
LET-4 and LET-653 for a genetic interaction. We treated wild-type worms with let-653
and let-4 RNAi. We detected no enhancement of embryonic or larval lethality (Table
4.3). Surprisingly, we saw an egg-laying defect. The majority of let-653 and let-4
RNAi-treated worms that grow to adulthood die with the „bag-of-worms‟ defect. let-653
or let-4 RNAi treatment alone produced no bagged worms. This phenotype is similar to
117

the phenotype observed in let-4 mutants that survived to adulthood (discussed further
below). This result supports an interaction between these two proteins, although it was
not detected in the excretory system. Experiments in our lab have found repeatedly that
the excretory system may be resistant to RNAi, therefore this experiment hasn‟t ruled out
an interaction between these two proteins in the excretory system. The excretory system
defect and the fact that mucins are part of the apical ECM make let-653 a tempting
candidate for a let-4-interacting protein. The role of let-653 in the excretory system, its
localization, and its potential to interact with eLLRons is being investigated in our lab.
How is the ECM affected in let-4 mutants?
What is the molecular reason for the inner eggshell layer being detached? In let-4
mutants, I observed a detached inner eggshell at the 1.5-fold stage, and a detached cuticle
in the duct cell in the worms with a distorted lumen. Could these phenotypes be related?
In both cases, the ECM is too far removed from the apical surface of epithelial cells
which normally express LET-4. LET-4 may play a role in anchoring the ECM to the
apical face of epithelial cells, or LET-4 could provide stability to the structure of the
ECM. My structure/function analysis of LET-4 (Chapter 2) has determined that a form
of the protein without a transmembrane domain can partially rescue the let-4 defect,
favoring the model that LET-4 provides stability, and is not directly anchoring the ECM
to the membrane. Although these phenotypes are similar, the result of each is very
different. The excretory system phenotype is associated with a catastrophic failure of the
excretory system, while the detached inner eggshell layer phenotype does not result in
118

failure to elongate. In order to further explore this possibility, the actual role of LET-4 in
the ECM needs to be investigated by identifying proteins interacting with LET-4.
What is the significance of the let-4 escaper phenotypes?
A small percentage of let-4(mn105) mutants survive the embryonic and L1 larval
stage, and are referred to as „escapers.‟ These individuals were found to have a variety of
non-lethal phenotypes, including phasmid dye-filling defect, an egg-laying defect, and
mildly uncoordinated movement.
The phasmid is a sensory structure consisting of neurons and associated tubular
glial cells. A dye-filling defect suggests a problem with epithelial glial cells associated
with the neurons, the sheath and socket cells (Perens and Shaham, 2005; Inglis et al.,
2006; Stone et al., 2009). Intriguingly, the socket cells are unicellular tubes, like the
tubes of the excretory system. The nature of apical ECM in these cells is uninvestigated,
but is it possible that LET-4 is required for interaction between the socket and or sheath
cells and their apical extracellular matrix.
The let-4 egg laying defect results in adult worms which are unable to lay eggs
before they hatch. As a result, the eggs hatch while still in the mother. A classic cause of
this „bag-of-worms‟ phenotype is vulval defects (Horvitz and Sulston, 1980). The vulva
is a tubular epithelial structure, in which LET-4 and EGG-6 localize apically. To
investigate the bag-of-worms defect, I performed a closer analysis of the vulva in let-4
escaper worms. The general morphology of the vulva appeared normal through the “X-
mas tree stage” of the L4 larva. This analysis was performed with DIC microscopy. To
investigate the junctions of the vulva in let-4 mutants, I looked at AJM-1::GFP, and it
119

appeared normal (data not shown). The vulva, therefore, does not have major polarity or
junctional defects. This phenotype could also be the result of muscle defects, although
the LET-4::GFP localization was enriched where epithelial cells were not in contact with
muscles. The vulva is lined by cuticle; therefore LET-4 may have a role in the
organization of the epithelial cells of the vulva and the ECM, though no obvious defects
were observed. The cause of the egg-laying defect needs further investigation to be fully
understood.
Intriguingly, the let-4 escaper phenotypes described here could be the result of
neuronal defects. Phasmid dye-filling could be the result of phasmid neuron defects. An
egg-laying defect could be the result of defects in muscle defects, or neuronal defects.
Egg-laying is ceased with the ablation of some vulval muscles and egg-laying is
influenced by environmental cues, which are mediated by neurons (Schafer, 2006).
Uncoordinated movement, another escaper phenotype, is also often the result of neuronal
defects (Bastiani and Mendel, 2006). I have presented no evidence for expression or
function of LET-4 in C. elegans neurons, however, the fact that there are roles for other
eLLRon proteins in neurons (described above) and the indirect evidence in the let-4
escaper phenotypes hints at a role in neurons.
Why is the apical domain of the canal cell so different from the duct and pore cells?
There are several differences between the apical domain of the canal cell and the
duct and pore. The canal cell has different cytoskeletal components than the duct and
120

pore. Second, the cuticle lines the duct and pore, not the canal. Third, the duct and pore
form by wrapping around and forming an autojunction, while the canal cell has never
been observed to form autojunctions. Fourth, PKC-3 and PAR-6 line the entire lumen of
the duct and pore, but are more restricted in the canal cell. The canal cell localization of
PAR-6::GFP and PKC-3 is particularly interesting. At the 1.5-fold stage, when the lumen
of the canal cell is localized in the main cell body, both proteins were localized apically.
As the lumen of the canal cell extended as the worm progressed through 2-fold and into
3-fold, the area of PAR protein localization did not change (Appendix 1). The result is
that both proteins are localized apically, but only in the part of the canal cell lumen
immediately adjacent to the secretory junction. The PAR complex proteins may be
required to “seed” the apical surface of the canal cell, but are not required along the
apical luminal face as it expands during the maturation of the canal cell. Fifth, LET-
4::GFP lines the apical lumen of the duct and pore, but not the canal cell. LET-4::GFP
lines the basal side of the canal cell in 1.5-fold stage (Chapter 2 and Appendix 2). Sixth,
HMR-1::GFP/E-cadherin reporter is present in the duct and pore, not the canal (Appendix
1). Why are these two types of lumen so different? They are known to have a different
mechanism of formation; they also likely have different functions. The canal cell is a
specialized cell which collects fluid, as plays a role in the worm‟s osmoregulation
(Nelson and Riddle 1984, Buechner, 1999). The duct and pore may serve to simply
transport the fluid produced by the canal cell, and not have the same secretory functions
as the canal cell, so a very different extracellular matrix may be required in the duct and
pore. Regardless of the reason for the differences in apical surfaces and ECMs, the
121

differences in these types of lumen only enhances the value of careful dissection of the
steps of formation and maintenance of the excretory system.
In summary, identification of more components of the apical ECM will allow us
to better understand the role of LET-4. Closer investigation of LET-4 and the other
eLLRons will offer insights into the role of this poorly understood gene family.
122

Materials and Methods
Alleles used were:
I: mec-8(u218) (Chalfie and Au, 1989), egg-6(ok1506) (Moerman and Barstead, 2008),
hmr-1(zu248) (Costa et al, 1998), vab-10(ok817) (Moerman and Barstead, 2008), vab-
10A(e698) (Hodgkin, 1983), vab-10B(mc44) (Bosher et al, 2003), erm-1(tm677) (Gobel
et al 2004), lpr-1(cs73) (Stone et al, 2009).
III: act-5(ok1397) (Moerman and Barstead, 2008), arx-5(ok1990) (Moerman and
Barstead, 2008), pat-2(ok2148)(Moerman and Barstead, 2008), pat-3(st564) (Williams
and Waterston, 1994).
IV: rho-1(ok2418) (Moerman and Barstead, 2008), let-653(s1733) (Clark and Baille
1992), lam-1(ok3139) (Moerman and Barstead, 2008),
X: let-4(mn105) (Meneely and Herman, 1979), sym-1(mn601) (Davies, et al 1999)
V: sqt-3(sc63) (Cox et al, 1980), sma-1(e30) (Brenner, 1974)
Transgenes used were:
jcIs1 (AJM-1::GFP) (Koppen et al., 2001)
Microscopy
Microscopy for analysis of let-4 vulvas was performed as described in Chapter 2
Materials and Methods.
123

RNAi
For RNAi injection experiments, let-4 dsRNA was generated and injected into adults as
described in Chapter 2 Materials and Methods. F1 pulse lays were performed, and
lethality was quantitated. For feeding RNAi experiments, L4 adults were exposed to
RNAi-expressing clones. Pulse lays were performed on the first day of adulthood.
Percent survival was defined as the proportion of embryos that reached L4 stage. Percent
embryonic lethal was the proportion of embryos that never hatched. Percent larval
lethality was the proportion of embryos that died as L1s.
124

Table 4.1 – let-4; lpr-1 double mutant phenotype
% of embryos with fluid in canal
cell body
Genotype
comma +
4hrs (n)
comma +
5hrs (n)
comma
+6hrs (n)
lpr-1(cs73) I; let-4(mn105);unc-3(e151)
X 0% (9) 18% (11) 64% (11)
lpr-1(cs73) I; let-4(mn105); unc-3(e151)
X; C44H4 0% (23) 8% (12) 86% (7)
125

Table 4.2- RNAi knockdown of worm lipocalins does not result in larval lethality
Genotype RNAi
eggs
(n) embryonic lethal L1 lethal survival
N2 GFP 155 1% 0% 99%
N2 T12A7.5/lpr-2 59 0% 0% 100%
N2 W04G3.8/lpr-3 101 0% 0% 100%
N2 W04G3.3/lpr-4 86 3% 0% 97%
N2 W04G3.2/lpr-5 227 0% 0% 100%
N2 W04G3.1/lpr-6 92 1% 0% 99%
N2 T19D7.3/lpr-7 216 0% 0% 100%
lpr-1(h276) GFP 132 10% 64% 27%
lpr-1(h276) T12A7.5/lpr-2 82 0% 68% 32%
lpr-1(h276) W04G3.8/lpr-3 141 19% 43% 38%
lpr-1(h276) W04G3.3/lpr-4 144 24% 66% 10%
lpr-1(h276) W04G3.2/lpr-5 111 4% 72% 24%
lpr-1(h276) W04G3.1/ lpr-6 96 0% 76% 24%
lpr-1(h276) T19D7.3/lpr-7 219 10% 56% 35%
126

Table 4.3 – let-4 and let-653 RNAi
Gentoype RNAi* n
embryonic
lethal
L1
lethal
Survival
(L4)
% of adults
with bag-of-
worms
phenotype
+ let-653 169 5% 0% 95 0%
+ let-4 82 12% 0% 88 0%
+ let-653 + let-4 116 18% 1% 81 59%
*Worms were exposed to let-653 dsRNA by feeding, and let-4 dsRNA by injection
127

Figure Legends
Figure 4.1. Three models for destabilization of the apical ECM leading to junction
breakage. (A) In the absence of LET-4, the apical ECM is destabilized, and the junction
proteins (blue) are subjected to increased force (red arrows). (B) In the absence of LET-
4, the apical ECM does not function to assist in ligand binding for an unknown signaling
pathway, which weakens junctions. (C) In the absence of LET-4, components of the
apical ECM fail to localize correctly and stabilize junctional proteins.
Figure 4.2. Summary of let-4 genetic interaction data. For each gene, single
heterozygous worms and worms also heterozygous for let-4 were generated, and their
progeny were phenotyped. Combined embryonic and larval lethality in the F1 generation
is depicted. Worms heterozygous for let-4 alone gave 22% lethality in the F1s. No genes
had a strong interaction with let-4 in this experiment. let-4 dsRNA was injected into
wild-type, sym-1, mec-8 and vab-10A homozygotes, and vab-10B heterozygotes. RNAi
experiments were not performed in the other mutants. Red line indicates 44%, which is
expected lethality in the case of purely additive lethality.
128

A
B
C
wild-type let-4
wild-type let-4
wild-type let-4
Figure 4.1
129

0
20
40
60
80
100
let-4 RNAi
let-4 trans-heterozygote
heterozygote
vab-10/spectra
plakin
sma-1/β
-H sp
ectrin
erm-1/cyto
skeletal li
nker
hmr-1/ca
dherin
lam-1/laminin
pat-3/β
-integrin
pat-2/α
-integrin
arx-5/Arp
2/3 su
bunit
rho-1/R
ho GTPase
act-5/actin
sqt-3
/colla
gen
egg-6/LRR protein
let-653/m
ucin
vab-10B/spectra
plakin
vab-10A/spectra
plakin
mec-8/R
RM
sym-1/LRR pro
teinlet-4
wild-ty
pe
let-4 genetic interactions
%le
thal
ity
Figure 4.2
130

Appendix 1
Characterization of apical and cytoskeletal markers in the
C. elegans excretory system.
131

Introduction
Simple genetically tractable model organisms such as C. elegans and Drosophila
have been instrumental in defining factors important for apical vs. basal specification and
proper development and maintenance of epithelial tubes. We aim to exploit the C.
elegans excretory system to continue to investigate these processes.
The C. elegans excretory system is a simple tubular organ that consists of just
three tandem unicellular tubes: the canal cell, the duct cell, and the pore cell. These
three tubes are morphologically distinct and form lumen via different processes. The
duct and pore cells form by a wrapping around mechanism in which the cells form
autocellular junctions, and the inside face of the cell adopt apical characteristics. The
duct cell then auto-fuses to dissolve its autocellular junction, while in the pore cell, the
autocellular junction remains. The canal cell forms lumen intracellularly at the site of the
duct-canal cell junction, apparently through a vesicular trafficking mechanism, and does
not possess an autocellular junction. These initial steps of tubulogenesis are complete by
the 1.5-fold stage of embryogenesis. During the latter part of embryogenesis, all three
cells elongate and undergo morphological changes to adopt their unique sizes and shapes.
The excretory system, therefore, is a simple system to study lumen formation, growth and
maintenance.
As described in Chapter 1, many epithelia are characterized by apical localization
of PAR complex proteins and the cadherin/catenin complex. Thus far, little is known
about the molecular components required for the formation and maintenance of the
excretory system. In this appendix, I describe the characterization of some conserved
132

apical factors in the excretory system. After the initial characterization of these markers,
I studied many of these factors in let-4 mutants, to look for defects.
Results
PAR protein localization in the excretory system
PAR-3, PAR-6 and atypical Protein Kinase C (PKC) are key regulators of
epithelial polarity and junction assembly, and they typically localize to the most apical
regions of epithelial cells. I investigated the localization of these proteins in the 1.5-fold
and 3-fold excretory system (Fig. Ap1.1). In the excretory system, I found that PAR-3 co-
localized with the apical junction markers AJM-1 and DLG-1/Discs Large at intercellular
junctions between the canal, duct and pore cells and along the pore autocellular junction
(Fig. Ap1.1A,F). PKC-3 lined the entire lumen of the duct and pore, and extended only a
short distance into the canal cell body. At the 1.5-fold stage, PKC-3 appeared to be in the
lumen of the canal cell, but as seen in 3-fold stage, the canals of the canal cell were not
lined with PKC-3 (Fig Ap1.1 C,H). PAR-6::GFP showed a localization pattern similar
to PKC-3 (Fig. Ap1.1E,J) PKC-3 and PAR-6 were not enriched at the junctions between
the cells of the excretory system. Thus, the apical domains of the canal cell vs. the duct
and pore have different molecular constituents, likely reflecting different mechanisms of
lumen and apical ECM formation and maintenance.
In let-4(mn105) mutants, the localization of PAR-3 (Fig. Ap1.1B,G) and PKC-
3(Fig. Ap1.1D,I) did not change. PAR-6::GFP localization was not studied in the let-4
background.
133

E-Cadherin and p120 catenin localization in the excretory system
To characterize the localization of the C. elegans E-cadherin homolog in the
excretory system, we used HMR-1::GFP and JAC-1::GFP reporters. HMR-1 is the worm
homolog of E-cadherin and JAC-1 is the worm homolog of cadherin/catenin complex
regulator p120 catenin (Pettit et al., 2003). Both these reporters displayed a strong signal
in the duct in early 3-fold, which degrades over the course of 3-fold. (Fig. Ap1.2) To
quantify the fading signal, individuals at different developmental stages were phenotyped
and placed into one of three categories describing duct cell expression of the GFP fusion
protein: ‘Strong,’ ‘Faint,’ or ‘Absent.’ (Fig. Ap1.2) For both reporters, the largest
category was Strong at the beginning of 3-fold (1.5-fold + 4 hrs). At the end of 3-fold
stage (1.5-fold + 8hrs), most embryos were Faint, and a large proportion were Absent.
Almost all worms fell into the Absent category in late larval stages.
HMR-1::GFP expression categories are as follows: in Strong individuals, the
expression is even along the length of the pore and duct lumen. Based on the shape of
the GFP signal, the whole duct cell cytoplasm was not labeled, only the apical face of the
cell, lining the lumen. In the Faint category, the signal in the duct contained gaps.
Finally, in individuals labeled Absent, the signal in the pore and secretory (duct-canal)
junction was still strong, but the signal was absent from the duct.
For JAC-1::GFP expression, ‘Strong’ individuals had the GFP signal in the duct
cell, and the signal was enriched at junctions. ‘Faint’ individuals had much more faint
expression in the duct, and ‘Absent’ individuals had no GFP in the duct. JAC-1::GFP
134

expression was not significantly affected in let-4(mn105) individuals. (In let-4, 30%
strong, 70% faint, n=20)
Cytoskeletal and Extracellular Matrix markers in the excretory system
I performed some preliminary studies of F-actin in the excretory system in 1.5-
fold and 3-fold wild-type embryos (Fig. Ap1.3E-H). In 4/4 1.5-fold embryos analyzed,
F-actin was visible in the canal cell and pore. In the duct cell, the F-actin was strong in
2/4 embryos, in one it was very faint in the duct, and in one it appeared broken up and
faint. Based on the pattern, the lumen of the canal cell had F-actin at the 1.5-fold stage.
Later in development, F-actin staining only appeared at junctions in the excretory system
in 3-fold and L1 worms (Fig. Ap1.3I,J). These data may suggest that F-actin is present at
the initial formation of the lumen, but is absent by the time the excretory system has
matured. Alternatively, phalloidin staining may not be efficient in these cells at the 3-
fold stage. To better analyze the timing of the F-actin pattern in the excretory system,
further experiments should be performed.
Proper Drosophila trachea development requires a temporary chitin scaffold
(Devine et al., 2005; Moussian et al., 2006). Could the excretory system have a similar
structure? To investigate this possibility, we stained for chitin, which had previously
been reported to be in the eggshell and adult pharynx (Zhang et al., 2005). To visualize
chitin, we stained embryos with Chitin Binding Protein (CBP) and MH27 (Fig. Ap1.4.i).
MH27 (anti-AJM-1) was used to mark the junctions of the excretory system. We
135

observed chitin staining in the eggshell, but no staining anywhere else in the embryo,
including the excretory system, in both 1.5-fold and 3-fold embryos (Fig. Ap1.4.i).
RNAi knockdown of a chitin sythaase results in L1 arrest, suggesting that chitin is
important in the pharynx early in development (Zhang et al., 2005). Because chitin was
not detected, either chitin is not present until after 3-fold, or it is, and this reagent is not
staining chitin in the pharynx at this early stage.
After finding no CBP staining in the excretory system, we tried a less stringent
reagent, Wheat Germ Agglutinin (WGA). WGA is a carbohydrate-binding protein that
binds some sugar residues found on cell membranes. In C. elegans, it has been reported
to stain the canal cell in adults (Hedgecock et al., 1990). We stained 1.5-fold and 3-fold
stage embryos to look for WGA-epitope staining in the cells of the excretory system. To
visualize the cells of the excretory system, we co-stained with MH27 to label the
junctions of the excretory system. In both 1.5-fold, and 3-fold embryos, WGA stained
the pharynx, but did not appear enriched at the junctions of the excretory system, or in the
duct and pore lumens, although non-specific staining was observed throughout the
embryo (Fig Ap1.4.ii).
Apical junction, intermediate filament (IF) and F-actin organization in the
epidermis
To further analyze the junctions of the epithelial cells in let-4 RNAi-treated sym-
1(mn601) embryos, MH27 (anti-AJM-1) and MH4 (anti-IF) staining were performed
136

(Fig. Ap1.5). At the 1.5-fold stage, MH27 and MH4 patterns are not affected in the
hypodermis in let-4 mutants, indicating that the elongation defect is not the results of the
failure of localization of these junctional components. (Fig. Ap1.5.i A,B and Fig. Ap1.5.ii
A,B). At the 3-fold stage, MH27 staining looked normal, but the intermediate filaments
are mildly disorganized, and the region to where IFs are localized is expanded (Fig.
Ap1.5.i A,B and Fig. Ap1.5.ii A,B). Because these embryos are ruptured in late 3-fold,
the IF effects may be a secondary consequence of the rupture. Also, the MH4 antibody
recognizes only a subset of intermediate filaments (Francis and Waterston, 1991),
therefore some IFs may be affected in these mutant embryos.
let-4 RNAi treatment of sym-1(mn601) embryos produced embryonic elongation
defects, as described in Chapter 2 (Fig. 2.5). To further investigate this phenotype, we
used phalloidin to stain for F-actin in the epidermal cells (Fig. Ap1.3A-D). The pattern
doesn’t appear to change in the 1.5-fold epidermal cells in the treated v. untreated sym-
1(mn601) embryos. In 3-fold epidermal cells, there are gaps in the circumferential
filament bundles (CFB) in let-4 RNAi-treated embryos (Fig. Ap1.3 D). These gaps
appear after rupture of these embryos, so the gaps may be a secondary consequence of the
rupture.
Cell type specific markers in the excretory system
To confirm correct fate specification and positioning of the cells of the excretory
system, several molecular markers were analyzed. Canal, duct, and pore markers appear
137

wild-type in let-4 mutants (Fig. Ap1.6, Table Ap1.1). vha-1::GFP, which labels the canal
cell is expressed normally in let-4 mutants (Fig. Ap1.6B-C), and allowed visualization of
the excretory canals, which revealed that the canals extend normally before massive fluid
accumulation distorts the canals. HMP-1/α-catenin::GFP, which labels the duct and pore
cells, were both unaffected in let-4(mn105) mutants, indicating that the cells were the
correct shape, and positioned correctly. (Fig. Ap1.6D-E).
let-4 mutants have a phasmid dye-filling defect
Because of the defects observed in the duct cell of let-4 mutants, we investigated
the possibility of defects in other tubular epithelia. Another duct lumen connectivity
mutant, lpr-1, has been demonstrated to have a phasmid dye-filling defect (Stone et al,
2009). Phasmid dye-filling defect could indicate defects in the tubular socket or sheath
cells surrounding the phasmid neurons (Perens and Shaham, 2005). To investigate
phasmid dye uptake in let-4 mutants, rare L4 let-4(mn105) escaper worms were exposed
to DiO. let-4 mutants were found to have a dye-filling defect, suggesting a defect in the
phasmid neurons themselves, or in the tubular glial cells surrounding the neurons (Fig.
Ap1.6 A).
Discussion
The tubular cells of the excretory system share apical characteristics of other
epithelial cells. These data have illustrated that the cells of the excretory system contain
138

the PAR proteins, indicative of conserved apical character. For further discussion of the
apical character of the duct, pore, and canal cell, see Chapter 4.
These experiments have demonstrated that in let-4 mutants, these apical markers
and junctions are unaffected at the resolution of this analysis. Thus, let-4 is not
responsible for fate specification or apical specification of the epithelial cells of the
excretory system or hypodermis.
Materials and Methods
Alleles used were: X: let-4(mn105), sym-1(mn601).
Transgenes used were: csEx146 (lin-48p:mCherry) (Abdus-Saboor et al., 2011), fgEx11
(ERM-1::GFP) (Gobel et al, 2004), jcIs1 (AJM-1::GFP) (Koppen et al., 2001), mcIs46
(DLG-1::RFP) (Diogon et al., 2007), qnEx59 (dct-5p::mCherry) (Abdus-Saboor et al.,
2011), saIs14 (lin-48p::GFP) (Johnson et al., 2001), xnIs96 (HMR-1::GFP) (Achilleos et
al., 2010), vha-1p::GFP (Oka et al., 1997), jcIs25(jac-1::GFP) Pettitt et al 2003, HMP-
1::GFP (Hardin Lab), xnIs3[par-6p::PAR-6::GFP; unc-119(+)] (Anderson et al, 2008)
Microscopy
Microscopy was performed as described in Chapter 2 Materials and Methods
Phasmid dye uptake assay
139

L4 let-4(mn105) worms were placed in 2mg/mL DiO (1:200 in M9) for 2 hours, then
allowed to recover on NGM plates for one hour before being mounted to a slide for
microscopy.
Immunostaining
Embryos were collected by bleaching gravid adult worms. The recovered
embryos were then allowed to incubate at room temperature to allow the eggs to reach
the desired developmental stage. The embryos were permeabilized by freeze-cracking
and fixed in methanol as described in Leung et al 1999 and Appendix 4. Primary
antibody concentrations used were: P4A1 (mouse anti-PAR-3) (1:25) (Obtained from the
DSHB); goat anti-GFP (1:50) (Rockland Immunochemicals), rat anti-PKC-3 (1:150)
(Tabuse et al 1998), mouse anti-IF (MH4) (1:50) (Francis and Waterston, 1991, Obtained
from the DSHB), mouse anti-AJM-1 (MH27) (1:100) (Obtained from the DSHB).
Secondary antibodies: Cy3 donkey anti-mouse (1:200); FITC donkey anti-goat (1:50)
(Jackson Immunoresearch) Cy3 donkey anti-rabbit (Jackson Immunoresearch) Cy3
donkey anti-rat (PKC-3) (1:200), FITC goat a-mouse (1:20). For Chitin staining, NEB
Chitin Binding Protein (NEB P5211S) (1:500 dilution) was used, and for WGA staining
WGA Texas-Red-conj (Molecular Probes W21405) (100mg/µL). No secondary antibody
was required for CBP and WGA staining.
140

Phalloidin Staining
Gravid adults were bleached, the embryos were collected and incubated in M9 for 5 hrs
to enrich for 3-fold stage. Freeze crack permeabilization was performed as described in
Appendix 4. Embryos were fixed [75%methanol, 3.7% paraformaldehyde, 1:200
phalloidin stock solution (1mg/mL of AlexaFluor 488 or 546 (Molecular Probes)in PBS)
for 30 minutes at -20°C, followed by 2x10min PBS-T(PBS + 0.1% Tween 20) wash, then
phalloidin stain (1:200) 1hr at room temperature, another 2x10min PBS-T wash, then
mounted on slides for visualization. (Protocol modified from van Furden, et al 2004.)
Acknowledgements
Thank you to Bossinger Lab for providing rabbit anti-DLG-1, the Kemphues Lab for
providing rat anti-PKC-3. The P4A1(PAR-3), MH27 and MH4 supernatants developed
by Francis and Waterston and were obtained from the Developmental Studies Hybridoma
Bank developed under the auspices of the NICHD and maintained by The University of
Iowa, Department of Biology, Iowa City, IA 52242.
141
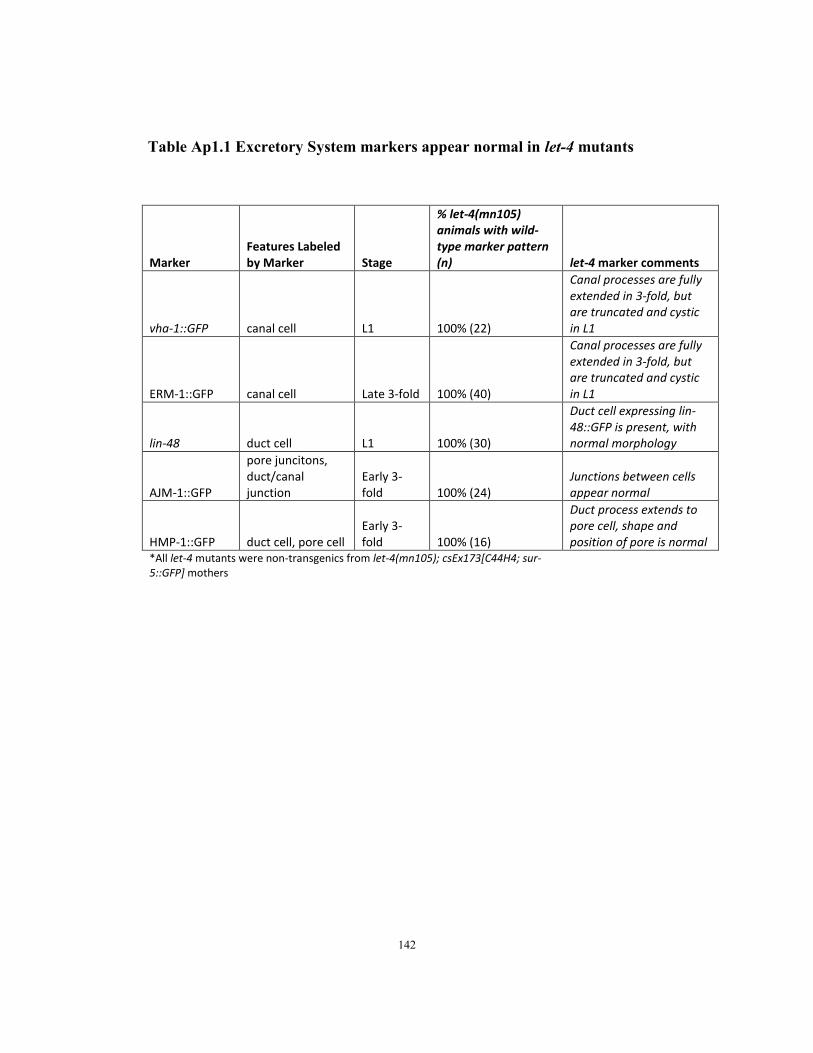
Table Ap1.1 Excretory System markers appear normal in let-4 mutants
Marker Features Labeled by Marker Stage
% let-4(mn105) animals with wild-type marker pattern (n) let-4 marker comments
vha-1::GFP canal cell L1 100% (22)
Canal processes are fully extended in 3-fold, but are truncated and cystic in L1
ERM-1::GFP canal cell Late 3-fold 100% (40)
Canal processes are fully extended in 3-fold, but are truncated and cystic in L1
lin-48 duct cell L1 100% (30)
Duct cell expressing lin-48::GFP is present, with normal morphology
AJM-1::GFP
pore juncitons, duct/canal junction
Early 3-fold 100% (24)
Junctions between cells appear normal
HMP-1::GFP duct cell, pore cell Early 3-fold 100% (16)
Duct process extends to pore cell, shape and position of pore is normal
*All let-4 mutants were non-transgenics from let-4(mn105); csEx173[C44H4; sur-5::GFP] mothers
142

Figure Legends
Figure Ap1.1. The PAR complex proteins are present in the cells of the excretory
system.
PAR-3 overlaps with DLG-1 staining in wild type at 1.5-fold (A) and 3-fold (F); pattern
is unaffected in let-4(mn105) mutants at both 1.5-fold(B) and 3-fold(G).
PKC-3 staining extends through the pore, duct, and into the canal cell lumen in both the
1.5-fold (C) and 3-fold (H) excretory system; the pattern is unaffected in let-4 mutants at
1.5-fold (D) and 3-fold (I) stage. n numbers = ( number of embryos with the illustrated
pattern)/(number of embryos analyzed).
PAR-6::GFP is present in the cells of the excretory system. Overlay (E,J) and GFP
(E’,J’) of 1.5-fold (E) and 3-fold (J) wild-type embryos expressing PAR-6::GFP. Regions
in dotted boxes in (E’, J’) are magnified in (E’’J’’). Arrow indicates estimated position
of the secretory junction, bracket indicated the duct, and arrow marks the pore cell. PAR-
6::GFP localization was not measured in let-4 mutants. n numbers indicate numbers of
embryos with the described pattern/number of embryos analyzed. Scale bars: 5µm.
Figure Ap1.2. The C. elegans E-Cadherin homolog HMR-1 and associated protein
p120 catenin (JAC-1) are localized dynamically in the duct cell.
Confocal images of GFP (A-H) and overlay with DIC (A’-H’) of the excretory system in
wild-type 3-fold embryos expressing HMR-1::GFP (A,D,G) or JAC-1::GFP (B,E,H) and
143

let-4(mn105) embryos expressing JAC-1::GFP (C,F). Dotted lines indicate canal cell
body. Scale bar: 5µm.
Throughout 3-fold, expression levels remained the same at apical junctions, while
expression of both reporters along the duct lumen was reduced (I,J). Expression of
HMR-1::GFP and JAC-1::GFP was categorized into three phenotypes: Strong, Faint, or
Absent in the duct.
HMR-1::GFP expression categories: Strong- expression is even throughout the duct
lumen (A). Faint- expression is uneven with breaks (D). Absent- GFP is visible at
apical junctions, but not along the duct cell (G).
JAC-1::GFP expression categories: Strong- expression is enriched at the junctions, but
even along the length of the duct (B). Faint- expression is faint in the duct (E). Absent-
GFP is visible in pore and secretory junction, but not the duct cell (H).
(C,F) JAC-1::GFP expression patterns were unaffected in let-4(mn105) mutants.
(I) Quantification of HMR-1::GFP in the duct cell during development
(J) Quantification of JAC-1::GFP in the duct cell during development.
Figure Ap1.3. F-actin (Phalloidin) Staining in epidermal cells and the excretory
system.
144

(A-D) F-actin is initially ordered, but later mildly disordered in the epidermis of let-
4 RNAi treated sym-1(mn601) worms. Phalloidin stains epidermal junctions, muscle
bands, and circumferential filament bundles (CFBs) of actin. Region in dotted box in
A,B,C and D are magnified in A’, B’, C’ and D’, respectively. In both untreated sym-1
(A, A’), and let-4 RNAi treated sym-1 (B, B’) pre- elongation embryos, CFBs are evenly
spaced. In B, most of the embryo is out of the plane of focus. Later, CFBs are evenly
spaced in untreated embryos (C,C’), but contain gaps in the let-4 RNAi-treated animals
(D,D’) Asterisk marks a gap between filaments.
(E-H) F-actin is present in the cells of the excretory system in wild-type 1.5-fold
embryos. E’F’G’H’ are magnified insets of E,F,G, H. Arrowhead marks estimated
position of secretory junction; bracket, duct cell; arrow, pore cell. Of the 4 embryos
stained for this analysis, phalloidin staining is strong and even in the duct in 2 (F,G) and
broken up or faint in 2 (E,H).
(I-J) F-actin is visible only in the junctions of the excretory system in wild-type 3-
fold and L1 embryos. F-actin (I,J) and AJM-1::GFP (I’,J’) of the excretory system in
the 3-fold embryo (I) and L1 larva (J). Arrowhead indicates secretory junction; bracket,
duct; arrow, pore autojunction. F-actin is stained with phalloidin, AJM-1::GFP is
visualized with anti-goat GFP. In the excretory system, phalloidin staining overlaps with
AJM-1::GFP. Scale bar: 5µm
Figure Ap1.4. Chitin and WGA epitope staining.
145

i.Chitin is only present in the eggshell at 1.5-fold and 3-fold stages. Chitin Binding
Protein (A,D) and MH27 immunostaining (B,E) in 1.5-fold (A-C) and 3-fold (D-F).
Regions in dotted boxes in (A-F) are magnified in (A’-F’). Arrowhead indicates
secretory junction, bracket duct, arrow pore autojunction. Scale bar 5µM.
ii.WGA is not enriched in the excretory system. WGA staining (A) and MH27
immunostaining (B) in 1.5-fold embryos (A-C) and L1 larvae (D-F). Regions in dotted
box in A-C are magnified in A’-C.’ Arrowhead indicates secretory junction; bracket,
duct; arrow, pore autojunction. Scale bar 5µM.
Figure AP1.5. AJM-1 and IF staining in let-4 RNAi-treated embryos
i. AJM-1, a discs large complex component, is unaffected in let-4 RNAi treated sym-
1(mn601) worms. MH27 (anti-AJM-1, green) stains a double band at the junctions of
epithelial cells. The double band results from a single band on the apical face of each of
two neighboring cells. Region in dotted box in A,B,C and D are magnified in A’, B’, C’
and D’, respectively. In both untreated (A,A’), and let-4 RNAi treated (B,B’) pre-
elongation embryos, AJM-1 is present at the apical face of adjacent cells, and localized
tightly at the apical face. AJM-1 is localized correctly at the apical face of adjacent cells
in later embryos (C,C’), also in let-4 RNAi treated embryos with epidermal lesions
(D,D’). Notice also in D, the epidermal cells outlined by AJM-1 are misshapen, not
regular and rectangular as wild-type. Scale bar =10µM.
146

ii. Intermediate Filaments (IFs) are initially ordered, but later mildly disordered in
let-4 RNAi treated sym-1(mn601) worms. MH4 (anti-IF, red in A, green in B-D) stains
epidermal cells where muscle will attach. The staining pattern is the same in both
untreated (A), and let-4 RNAi treated (B) pre- elongation embryos. Arrows indicate
staining of epithelial cells in A and B. Later, CFBs are tightly packed and evenly spaced
in wild-type (C) but are wider (brackets) and contain gaps (arrowheads) in the let-4
RNAi-treated animals (D). Scale bar =10µM
Figure Ap1.6. Further characterization of the let-4 phenotype.
(A) let-4(mn105) mutants have a dye-filling defect. Worms were exposed to DiO, and
assayed for dye uptake into phasmids. 100% of wild-type worms uptake dye into all 4
phasmids. let-4(mn105) worms show a reduced ability to uptake dye. Rate of defect is
similar to level seen for another duct cell lumen mutant, lpr-1(cs73) (lpr-1 data from
Craig Stone).
(B-E) The cells of the excretory system are present and positioned correctly in let-
4(mn105) mutants. (B-C) vha-1::GFP marks the canal cell, which initially extends canals
normally, although the cell becomes distorted in let-4 mutants (C). (D-E) HMP-1::GFP
labels the duct and pore cells. Arrow indicates estimated position of the secretory
junction, bracket indicates the duct, and arrow marks the pore cell. Pattern is unaffected
in let-4 mutants (E). Scale bar: 5µm.
147

DLG-1
DLG-1
PAR-3
PKC-3
1.5
fold
3-fo
ld
E
J
E’
J’
E’’
J’’
n=36/36
n=11/12*
C
PAR-6::GFP
PAR-6::GFP
PKC-3 DLG-1
PAR-3 DLG-1
wild-type let-4
DLG-1PAR-3
DLG-1PKC-3
PKC-3 DLG-1
PAR-3 DLG-1
A A’ A’’ B B’ B’’
C C’ C’’ D D’ D’’
G G’ G’’F F’ F’’
H H’ H’’ I I’ I’’
n=15/16 n=6/7
n=10/11 n=12/12
n=5/5
n=2/2n=12/12
n=13/13
Figure Ap1.1
148
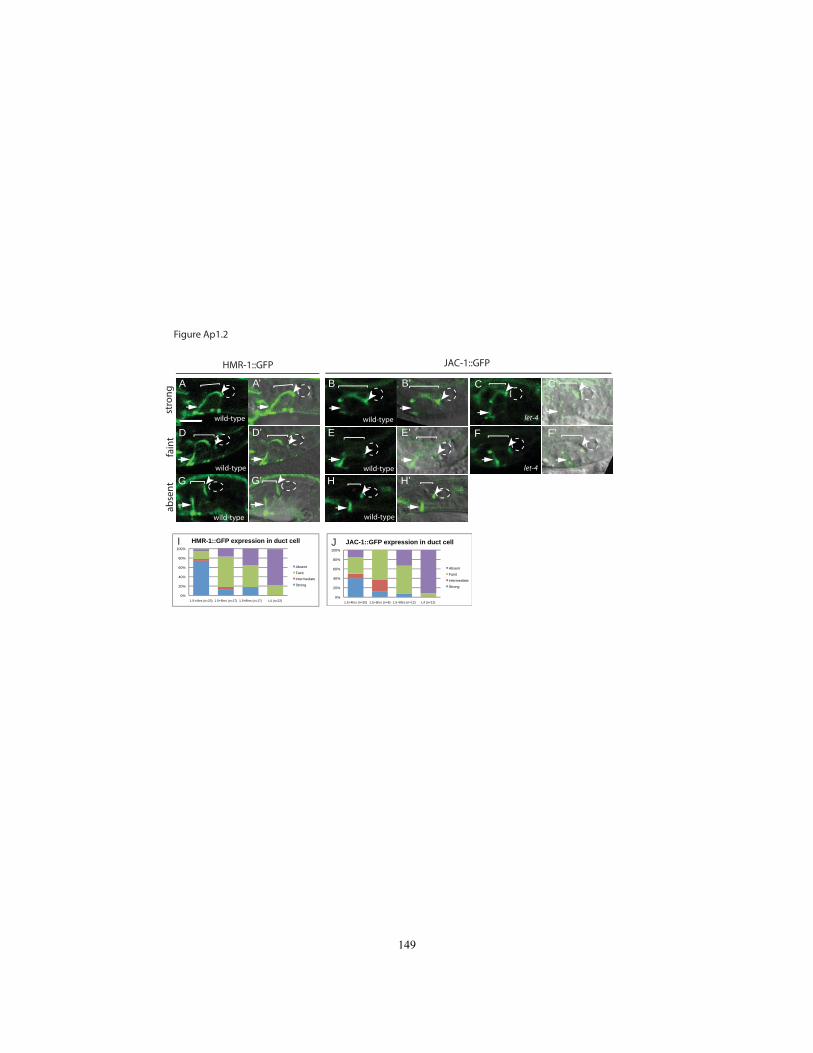
0%
20%
40%
60%
80%
100%
1.5+4hrs (n=23) 1.5+6hrs (n=17) 1.5+8hrs (n=17) L4 (n=22)
HMR-1::GFP expression in duct cell
Absent
Faint
intermediate
Strong
0%
20%
40%
60%
80%
100%
1.5+4hrs (n=20) 1.5+6hrs (n=8) 1.5+8hrs (n=12) L4 (n=12)
JAC-1::GFP expression in duct cell
Absent
Faint
intermediate
Strong
AJ
stro
ngfa
int
abse
nt
HMR-1::GFP JAC-1::GFP
Figure Ap1.2
wild-type wild-type
wild-type
wild-typewild-type
wild-type
let-4
let-4
I
A B C
D E F
G H
A’ B’ C’
D’ E’ F’
G’ H’
149
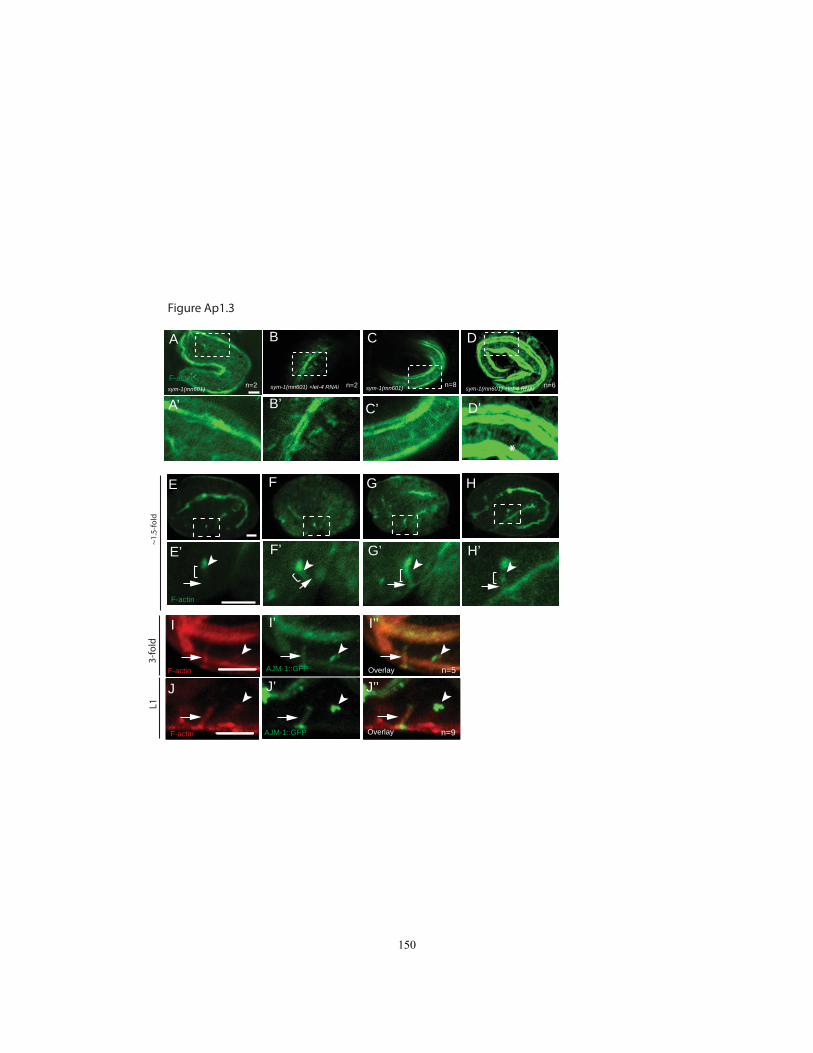
AJM-1::GFP
AJM-1::GFP
F-actin
F-actin
Overlay
Overlay
n=5
n=9
3-fo
ldL1
~1.5
-fold
A
A’
B
B’
C
C’
D
D’sym-1(mn601) sym-1(mn601)sym-1(mn601) +let-4 RNAi sym-1(mn601) +let-4 RNAin=2 n=2 n=8 n=6
E F G H
E’ F’ G’ H’
I I’ I’’
J J’ J’’
*
F-actin
F-actin
Figure Ap1.3
150

L1
~1.5
-fold
B
B’
A
A’
C
C’
ED F
MH27
MH27
MH27WGA
WGA
WGA Overlay
Overlay
Overlay
i.
A
A’
B
B’
C
C’
D
D’
E
E’
F
F’
ii.
Overlay
Overlay
Overlay
Overlay
Chitin MH27
Chitin MH27
Chitin MH27
Chitin MH27
3-fo
ld~1
.5-fo
ld n=2
n=9 n=20
n=2
Figure Ap1.4
151
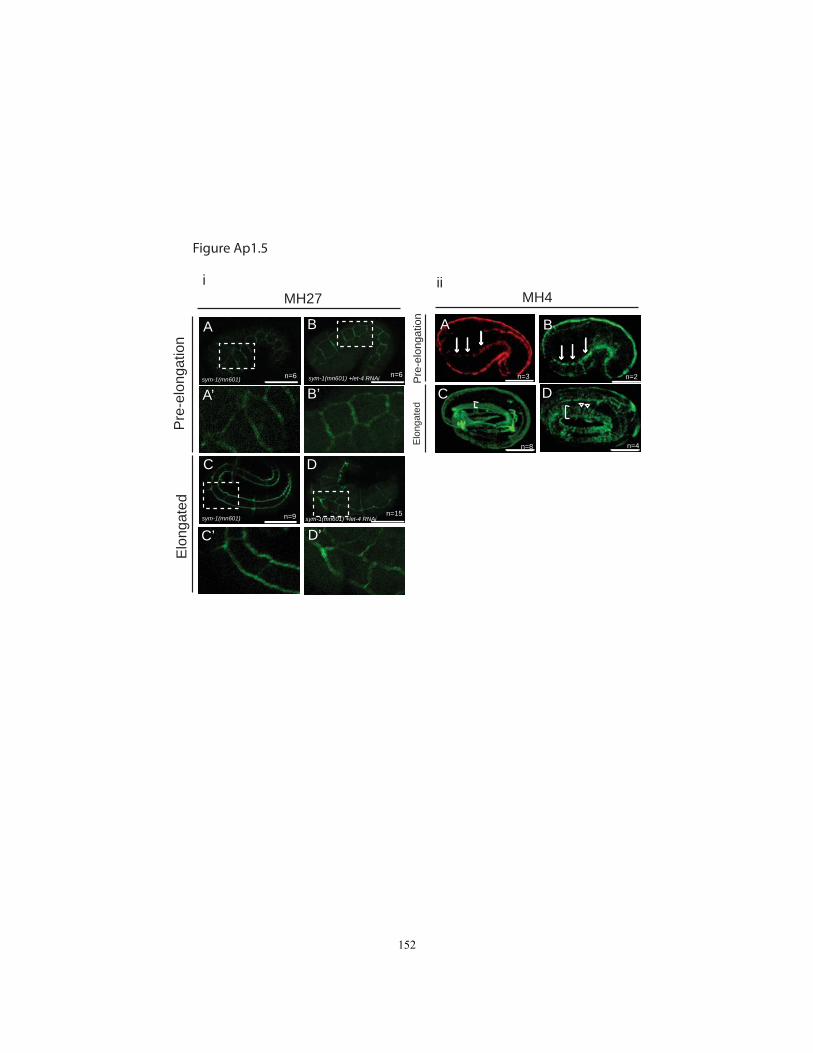
A
A’
B
B’
C
C’
D
D’
Pre
-elo
ngat
ion
Elo
ngat
ed
i
n=6 n=6
n=9 n=15
sym-1(mn601)
sym-1(mn601)
sym-1(mn601) +let-4 RNAi
sym-1(mn601) +let-4 RNAi
MH27 MH4
A B
C D
ii
n=3 n=2
n=8 n=4
Pre
-elo
ngat
ion
Elo
ngat
ed
Figure Ap1.5
152
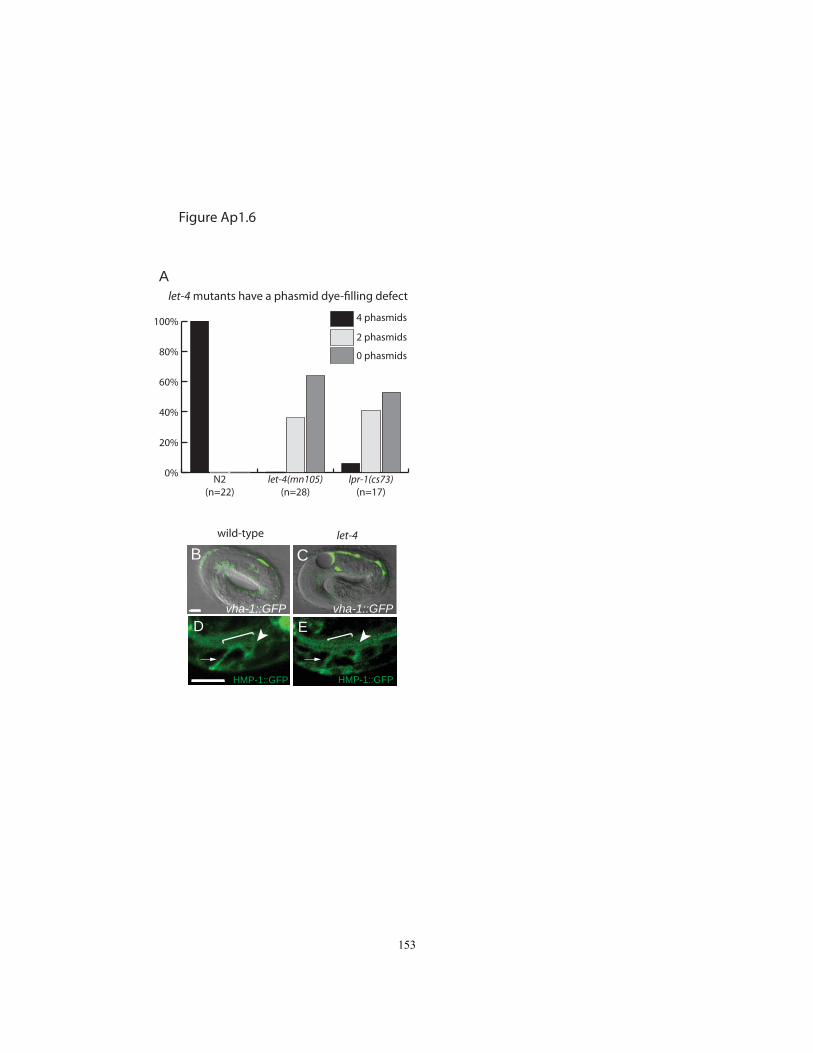
0%
20%
40%
60%
80%
100%
0 phasmids
2 phasmids
4 phasmids
lpr-1(cs73)(n=17)
let-4(mn105)(n=28)
N2(n=22)
let-4 mutants have a phasmid dye-filling defect
Figure Ap1.6
A
HMP-1::GFP HMP-1::GFP
D E
wild-type let-4
vha-1::GFP vha-1::GFP
B C
153

Appendix 2
let-4 expression in the excretory system
154

Introduction
To understand the expression and localization of LET-4, I generated
transcriptional and translational reporters, summarized in Chapter 2. The let-4p::GFP
and LET-4::GFP transgenes revealed dynamic expression patterns in the excretory
system. This appendix includes a detailed analysis of the expression pattern of the
transcriptional and translational let-4 reporters.
Results
let-4 is transcribed dynamically in the excretory system. (Fig. Ap2.1) At the 1.5-
fold stage, when the lumen has first formed in all three cells of the excretory system, let-
4p::GFP was expressed in all 3 cells of the excretory system. Over the course of 3-fold
stage, expression was reduced in the canal cell, but GFP expression was still present in
the duct and pore cell (Fig. Ap2.1 B). This expression pattern is mirrored by the
expression of LET-4::GFP in the excretory system (Fig. Ap2.1 C). LET-4::GFP was
present in the duct, pore and canal cell at 1.5-fold stage. The GFP signal remained in the
duct and pore through the L4 larval stage, while expression in the canal cell was not
visible by hatch.
To determine the relative localization of LET-4::GFP, we co-immunostained GFP
and DLG-1. The junctional DLG-1 protein localized to the pore connection to the
outside, pore autojunction, and duct-pore connection (Fig. Ap2.1F-H). At the 1.5-fold
155

stage, in the excretory system, even with DLG-1 labelled, it is difficult to resolve the
localization of LET-4 because the duct and pore cell themselves are so narrow. A ventral
view of the pore opening in a 1.5-fold embryo shows LET-4::GFP in the pore cell, but it
is difficult to resolve LET-4::GFP is enriched apical to DLG-1 (Fig. Ap2.1G). By the 3-
fold stage, when the cells are larger and have taken their distinct shapes (Refer to Fig. 2.4
A,B), LET-4::GFP was clearly localized to the lumen of the duct and pore cells, and did
not fill the entire cytoplasm of the cells (FigAp2.1H). Localization of LET-4::GFP is not
exclusively to the apical face of cells; it is dispersed in the cytoplasm and enriched at the
apical face.
Discussion
The let-4 promoter drives expression in the duct and pore beginning at 1.5-fold,
which may indicate a role for let-4 at this early stage. However by TEM and marker
analysis (Chapter 2, Appendix 3), the duct and pore lumen and junctions appear wild type
during 1.5-fold, and well into 3-fold. It is possible that LET-4 may first be serving a role
in 1.5-fold, and the deficiency doesn’t cause a phenotype until late in 3-fold, when the
excretory system begins to function. To determine the timing of let-4 requirement, a
transgene was generated driving LET-4 under the control of a heat-shock inducible
promoter, but this construct was too toxic to be useful (data not shown). Most rescue
experiments have indicated that worms are very sensitive to overexpression of LET-4.
156

Why is the let-4 promoter driving expression earlier in the canal cell than the duct
and pore? One possibility could be an early role for let-4 in canal cell lumen
development. However, based on the TEM analysis (see Appendix 3) the canal cell
lumen appears unaffected in let-4 mutants. Also, expression of let-4 in the duct and pore,
but not the canal cell, is sufficient to rescue let-4(mn105) (Fig. 2.2). Therefore any role
of LET-4 in the canal cell is dispensable. Given the redundancy of LET-4 and SYM-1 in
the epidermal elongation, perhaps LET-4 functions redundantly with other eLLRons in
the canal cell.
157

Materials and Methods
Transgenes used were:
csEx203 [10ng/uL pVM7(let-4p::GFP) + 100ng/uL pRF4[rol-6(su1006)] and csEx210
[2ng/µLpVM4(LET-4::GFP) +98ng/µLpRF4(rol-6(su1006)]. For further description of
these transgenes, see Chapter 2 Materials and Methods.
Microscopy
Microscopy was performed as described in Chapter 2 Materials and Methods
Immunostaining
Freeze-crack of LET-4::GFP embryos was performed as described in Appendix 4.
Primary antibodies: goat anti-GFP (1:50) (Rockland Immunochemicals); rabbit anti-
DLG-1 (1:400) (Segbert et al 2004) Secondary antibodies: FITC donkey anti-goat (1:20)
and Cy3 anti-rabbit (1:200) (Jackson Immunoresearch)
Acknowledgements
I thank Olaf Bossinger for kindly providing the rabbit anti-DLG-1 antibody.
158

Figure Legend
Figure Ap2.1. Expression of let-4 in the excretory system.
(A-B) let-4p::GFP is dynamically expressed in the excretory system. The boxed regions
in A and B are magnified in A’and B’. At 1.5-fold stage (A), let-4p::GFP is expressed in
the canal cell (c) duct cell(d) and pore cell(p) nucleus; in the canal cell, expression
decreases during 3-fold (B,C). The individual shown in (B) is also carrying AJM-1::GFP
for easy identification of the cells of the excretory system. (C) Quantification of presence
of let-4p::GFP in the canal cell from 1.5-fold to late 3-fold. (Hatch normally occurs ~1.5-
fold + 8hrs.) (D-E) LET-4::GFP is dynamically expressed in the excretory system. LET-
4::GFP is localized to the basal membrane of the canal cell and to the lumen of duct and
pore cell in 1.5-fold (D) and 3-fold (E). (F-H) Immunostaining of GFP (F,G,H) and
DLG-1(F’,G’,H’) in LET-4::GFP. DLG-1 labels the pore connection to the outside, pore
autojunction and the duct-pore connection. (F) 1.5-fold excretory system. (G) Ventral
view of the 1.5-fold excretory system. (H) 3-fold excretory system. LET-4::GFP is
localized to the basal membrane of the canal cell and to the lumen of duct and pore cell in
1.5-fold (F) and 3-fold (H). Arrow indicates estimated position of duct-canal junction;
bracket indicates duct, and arrow marks pore cell. Scale bars: 5µm.
159
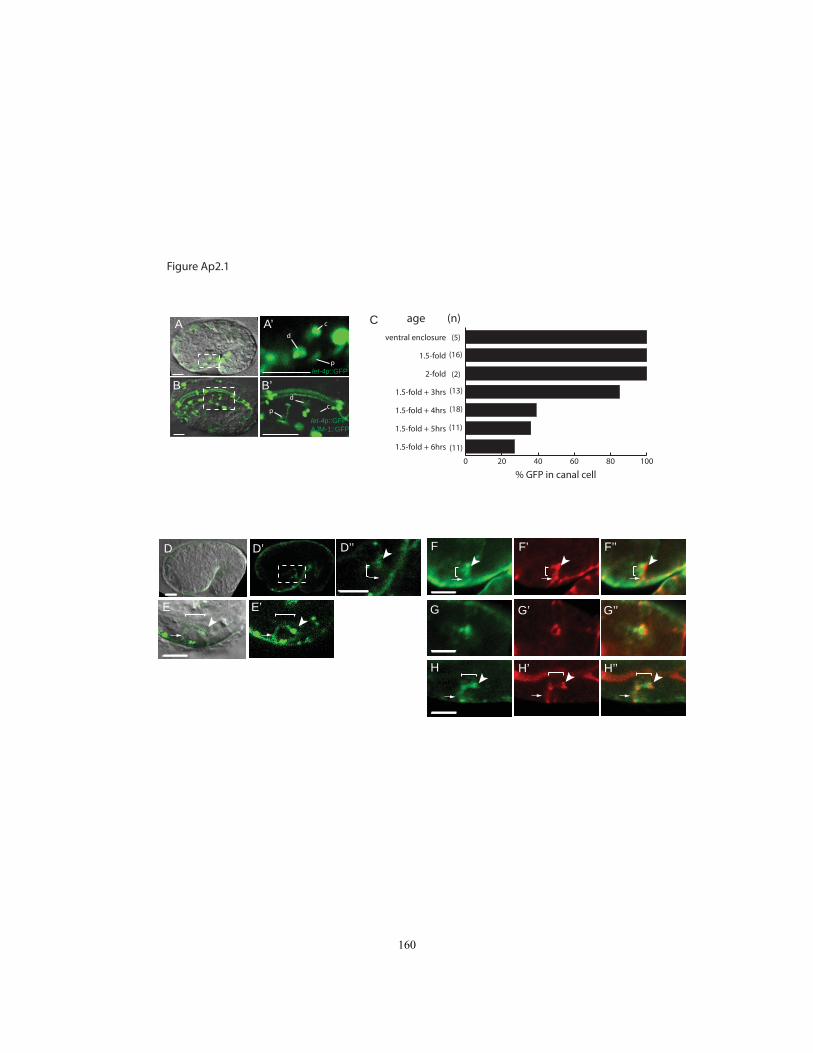
Figure Ap2.1
0 20 40 60 80 100
1.5-fold + 6hrs
1.5-fold + 5hrs
1.5-fold + 4hrs
1.5-fold + 3hrs
2-fold
1.5-fold
ventral enclosure
age
% GFP in canal cell
(n)
(5)
(16)
(2)
(13)
(18)
(11)
(11)
d
p
c
cp
d
A A’
B B’
D D’
E E’
C
let-4p::GFPAJM-1::GFP
let-4p::GFP
D’’ F
G
H
F’
G’
H’
F’’
G’’
H’’
160

Appendix 3
TEM analysis of the C. elegans excretory system
161

Introduction
The C. elegans excretory system is a tiny structure; the lumen of the duct, pore
and canal cells can be more narrow than 1 µm, making visualization of the lumen
difficult with molecular markers and confocal microscopy. For a more detailed imaging
of the lumen on junctions in both wild-type and let-4 mutants, we performed
Transmission Electron Microscopy (TEM). TEM has been successful before in
visualizing excretory system luminal defects (Stone et al, 2009). This analysis provides
serial sections, allowing us to trace the entire lumen through the three cells of the
excretory system. Additionally, TEM allows closer examination of the apical extra-
cellular matrix (ECM) in the excretory system, and outside the embryo. The resolution of
TEM analysis allows us to address some specific questions that could not be addressed
with confocal microscopy: Does the lumen form normally? Is the lumen maintained
through late stages of embryonic development? Are there visible junctional defects in
let-4 mutants? Are there apical ECM defects in let-4 mutants?
We chose to perform the analysis on both 1.5-fold and 3-fold stage let-4(mn105)
mutants. 1.5-fold stage is just after the initial formation of the junctions and lumen of the
excretory system. By investigating this stage, we could determine if the junctions and
lumen initially form correctly in let-4 mutants. After initial formation in 1.5-fold stage,
the excretory system matures until 3-fold stage. We chose to perform the 3-fold stage
analysis as late as possible in development before the excretory system phenotype was
observed with confocal microscopy. This time point was chosen to visualize the matured
162

excretory system before the system was damaged by secondary effects caused by fluid
accumulation. A brief summary of the TEM results, as well as my conclusions were
presented in Chapter 2. Here, I present more detailed description of the analysis. At the
end of this appendix, I’ve included a summary datasheet for each embryo analyzed.
Results
TEM analysis revealed no excretory system defect in 1.5-fold stage let-4 mutants.
(Fig. Ap3.1). We sectioned through the excretory system of five embryos. The lumen
does not have any obvious lumen enlargement relative to available images of archived
wild-type embryos (Archived in the Hall Lab). The lumen was clearly continuous in 4/5
embryos. In a fifth embryo, let-4-1, a key serial section through the lumen was missing,
however, there is no indication of a luminal defect.
Although no excretory system defect was observed, TEM analysis revealed an
inner eggshell layer defect in 1.5-fold let-4 mutants. In available embryos fixed by HPF,
the inner eggshell layer is close to the embryo, and reaches across the pore cell opening
(See Chapter 2). In all five of the 1.5-fold let-4 mutants analyzed here, the inner eggshell
layer is pulled away from the surface of the embryo (Fig. Ap3.1), suggesting
compromised apical ECM integrity.
We sectioned through three wild-type 3-fold embryos to serve as a control for the
let-4 3-fold analysis. Key images from these embryos highlighting features of the
excretory system are presented in Figure Ap3.2.
163

We sectioned through nine let-4 3-fold embryos (Fig. Ap3.3). For a summary of
the 3-fold TEM analysis, see Table Ap3.1. Four of the nine had luminal defects. In two
of these embryos, let-4-8-1 (Fig. Ap3.3A) and let-4-9-2 (Fig. Ap3.3B), the duct cell
lumen connected to the canal cell, but terminated in the duct cell.
Of the four let-4 3-fold embryos with luminal defects, I did not observe duct/pore
cell connectivity in any of them. In let-4-8-1, let-4-9-2 and let-4-7-1 (Fig. Ap3.3C), the
pore cell could not be identified. In the TEM analysis pore cell features such as the
autojunction and lumen are the distinguishing characteristic. These features are lost in
let-4 mutants (as observed with Confocal microscopy), prohibiting pore identification. In
let-4-7-2, the pore cell was observed to still have an autojunction, and connect to the
outside of the worm, but connectivity to the duct could not be assessed, due to section
damage (Fig. Ap3.3D). This embryo is the most ambiguous of the 3-fold analyzed. The
lumen may be distorted in some sections, and the pore may hae no cuticle lining,
although the lumen can’t be followed all the way through, because sections are missing.
In the embryos with an enlarged lumen (Fig. Ap3.3.C, D, J), the cuticle inside the duct
lumen was observed to be pulled away from the surface of the duct lumen.
Based on confocal and TEM analysis, the duct/pore connection is lost late in 3-
fold stage in let-4 mutants (Fig. 2.4, Ap3.3). Therefore, it could be possible there are
defects in this junction before the phenotype is apparent. However, in the 5 let-4 3-fold
mutants that did not yet have duct lumen defects, the junction between the duct and pore
was not observed to be different than wild-type (Fig. 3.3E-I). Based on TEM analysis,
the dark, electron dense junctions were present, and appeared wild-type in these let-4 3-
164

fold mutants. Sections highlighting these junctions are presented in Figure Ap3.3.
(Compare to wild-type duct/pore junctions, Fig. Ap3.2). Therefore, at the resolution of
this analysis, the duct/pore junction, which destabilizes late in 3-fold, is normal into the
3-fold stage.
In addition to the nine let-4 3-fold embryos described above, partial analysis was
performed on two other embryos. In these two let-4 3-fold embryos, many sections were
lost and shuffled during the sectioning process. Thus the full analysis of the excretory
system could not be performed, although some data could be collected from these
embryos. In the embryo let-4-10-1, sections near the secretory junction were recovered
and analyzed (Fig. Ap3.3J). These sections revealed an enlarged duct cell lumen, a
detached cuticle inside the duct cell, and a mildly enlarged canal cell lumen. In let-4-10-
6, sections posterior to the terminal bulb of the pharynx contained what appears to be
accumulation of fluid (Fig. Ap3.3K). Although the origin of this fluid could not be traced
due to damage to more anterior sections, the accumulation looks similar to the fluid
visible in the embryo let-4-8-1, which was distorted significantly.
Discussion
In let-4 mutants, which phenotype occurs first: enlarged lumen, or detached
lumen? In all embryos with a luminal phenotype except one, the lumens are both
enlarged and detached. In let-4-9-2, the duct cell lumen is truncated, but the lumen of
neither the duct nor canal cell is enlarged. This suggests that the lumen detachment
occurs before the lumen becomes enlarged, leading to the following model: let-4 mutants
165

develop normally until late 3-fold, at which time the junction between the duct and pore
destabilizes due to the absence of a destabilized apical extracellular matrix. The loss of
junctional integrity then results in several events. The duct cell becomes detached from
the pore cell, the duct cell lumen collapses, and the pore cell autojunction falls apart. As
fluid is collected by the excretory system with a collapsed duct lumen, it backs up into
the duct and canal cell lumens leading to swelling. These events occur rapidly, and with
the current data the order of the junction destabilization cannot be determined. To
evaluate this model, and determine the order of the events, more TEM analysis is
required.
Based on confocal and TEM analysis, let-4 mutants lose the pore autojunction and
the junctions between the duct and the pore. However, the connection between the canal
cell and duct cell, the secretory junction, was unobstructed and looked normal in all let-4
1.5-fold and 3-fold mutants (Except for let-4-2-5, which was damaged during HPF).
Why isn’t this junction affected? Based on tissue-specific rescue (Fig. 2.2), let-4 is not
required in the canal cell. The apical character of the canal cell and its ECM is different
than the duct and pore, and perhaps other proteins at the secretory junction maintain the
junction even in the absence of LET-4.
The cuticle, as visualized by TEM, was pulled away from the apical face of the
duct cells in let-4 mutants with enlarged lumen. However, despite the swelling of the
lumen in some let-4 mutants, the cuticle inside the duct cell stays intact. In all embryos,
the cuticle was continuous through all sections containing a lumen, and maintained a
normal diameter even as the lumen of the duct cell was enlarged (Fig. Ap3.3D.i-iii,
166

Fig.Ap3.3J.i,ii). This indicates that LET-4 is not required for the cuticle to maintain its
shape, but LET-4 may be required to anchor the cuticle to the apical surface of the duct
cell. The most significant change observed in the cuticle structure was in let-4-7-1,
where cuticle appears distorted in the duct cell body (Fig. Ap3.3C.ii). (Also, the cuticle
terminated in a bolus when the lumen terminated in let-4-8.) The apical ECM, inside the
duct cell, which includes the cuticle, likely contains components that are not visible using
the fixation and staining procedures used here, and loss of LET-4 likely has an effect on a
part of the luminal ECM which is not preserved during this fixation.
Detachment of inner eggshell layer in 1.5-fold let-4 mutants could be the result of
destabilized interactions of the embryonic sheath with the eggshell layer. This de-
stabilization is not catastrophic for the embryo, though, because this defect was observed
in 5/5 embryos, and most let-4 embryos progress beyond the 1.5-fold stage and elongate
normally. It is only in the sym-1 let-4 double loss-of-function embryos that embryonic
elongation defects are observed at a high rate (Fig. 2.5). The connection of a de-
stabilized inner-eggshell layer and the elongation defect is still unknown, and little is
known about the molecular components of the inner eggshell layer. For further
exploration, see Chapter 4.
167

Materials and Methods
Alleles used were: let-4(mn105) X, progeny from let-4(mn105) mothers.
For TEM analysis, non-green let-4 escaper worms were picked from the strain UP1787
[let-4(mn105); C44H4] and maintained as a stock. 1.5-fold embryos from this escaper
line were picked for 1.5-fold stage analysis. For the 3-fold stage analysis, embryos from
this let-4 escaper line and the wild-type N2 line were picked as 1.5-fold embryos, and
incubated for 6 hours at 20°C before performing HPF. During sectioning, TEM sections
were laid out on grids. The position of the sections on the grid was recorded by the
sectioning technician. Occasionally, sections could fall on the grid on top of each other,
or sections could curl up, leading to individual sections being lost, damaged, or out of
order.
For TEM fixation and staining procedures, see Chapter 2 materials and methods and
Appendix 4. Images were processed and colored with Adobe Photoshop and ImageJ.
Acknowledgements
I thank David Hall, Ken Nguyen, Juan Jimenez and Leslie Gunther for help in conducting
HPF fixations. Ken Nguyen sectioned the fixed embryos. David Hall provided expertise
in analysis. I thank Shai Shaham, John Suslton, Jonathan Hodgkin, Richard Fetter and
Cornelia Bargman for providing archival TEM images. I performed the electron
168

microscopy in the EMRL at the University of Pennsylvania, supported in part by NIH
grant 5 P30 CA 016520.
169
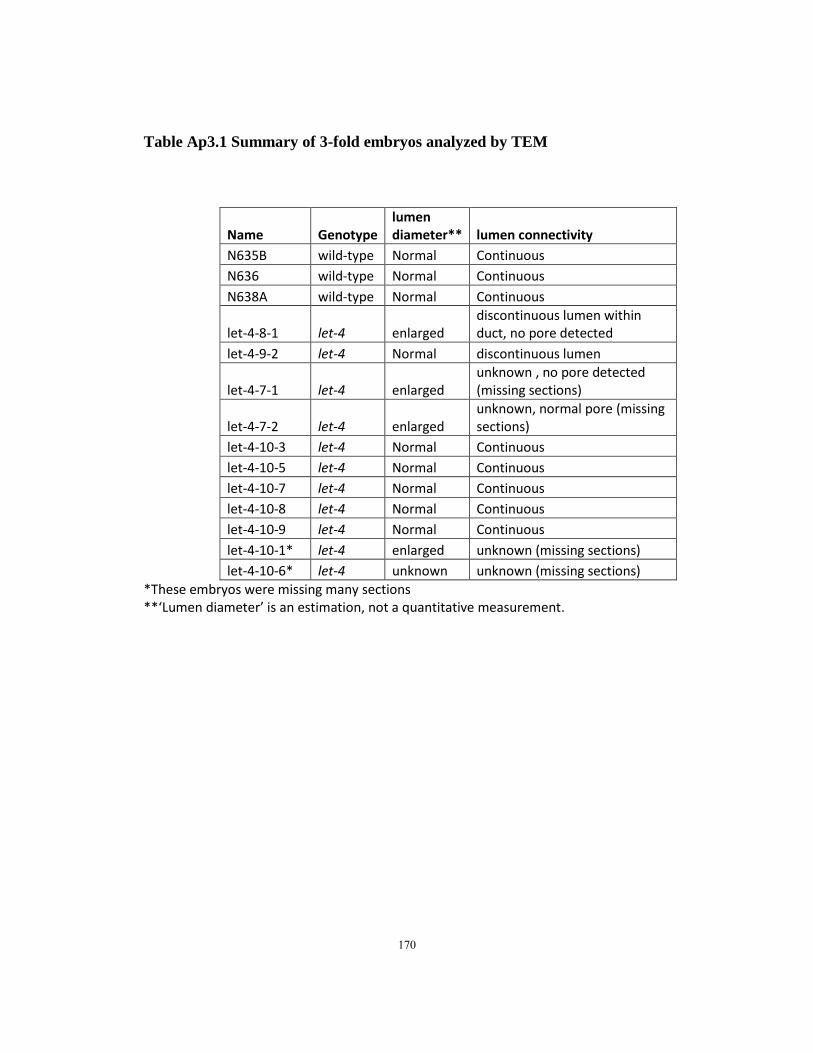
Table Ap3.1 Summary of 3-fold embryos analyzed by TEM
Name Genotype lumen diameter** lumen connectivity
N635B wild-type Normal Continuous
N636 wild-type Normal Continuous
N638A wild-type Normal Continuous
let-4-8-1 let-4 enlarged discontinuous lumen within duct, no pore detected
let-4-9-2 let-4 Normal discontinuous lumen
let-4-7-1 let-4 enlarged unknown , no pore detected (missing sections)
let-4-7-2 let-4 enlarged unknown, normal pore (missing sections)
let-4-10-3 let-4 Normal Continuous
let-4-10-5 let-4 Normal Continuous
let-4-10-7 let-4 Normal Continuous
let-4-10-8 let-4 Normal Continuous
let-4-10-9 let-4 Normal Continuous
let-4-10-1* let-4 enlarged unknown (missing sections)
let-4-10-6* let-4 unknown unknown (missing sections)
*These embryos were missing many sections **‘Lumen diameter’ is an estimation, not a quantitative measurement.
170

Figure Legends
Figure Ap3.1. TEM images of the excretory system and detached inner eggshell
layer in 1.5-fold let-4 embryos. In all panels, canal cell (c) is labeled red; duct cell (d),
yellow; pore cell (p), blue and gland cell, purple.
A.let-4-1. Orientation is nearly transverse, sections go posteriorly as numbers increase
(i-iii)TEM images of the excretory system. (i) lumen is visible in canal call, duct and
pore. Part of the duct cell lumen is in other sections. (ii) whispy EC material is visible in
canal cell. (iv) TEM image illustrating detached inner eggshell layer(orange arrows).
Luminal ECM (line) is visible in (ii). Green arrows indicate embryonic sheath. This
embryo was missing a key section such that lumen connectivity could not be assessed.
B. let-4-2-3. Orientation is longitudinal, sections go ventral as numbers increase.
(i-vi) TEM of the excretory system. (i) canal cell lumen, excretory gland cell is visible.
(ii) duct cell luminal ECM (lines) is visible in (ii-iii). (iii) pore cell; ECM is visible. (iv)
pore cell opening. (v) TEM image illustrating detached inner eggshell layer (orange
arrow). Green arrows mark embryonic sheath.
C. let-4-2-5. Orientation is transverse, but tilted posterior. (Dorsal part of the section is
more posterior than the ventral part). Sections go posterior as numbers increase. (i-iv)
TEM of the excretory system. (i,ii) canal cell and pore cell lumen. (iii) duct cell lumen.
(iv) pore cell lumen and opening to outside of worm. Excretory gland cell is visible in
(iii-iv). (v) TEM image illustrating detached inner eggshell layer (orange arrow). This
embryo was slightly damaged during high pressure freezing.
171

D. let-4-3. Orientation is nearly transverse; sections go posterior as numbers increase. (i-
v)TEM images of excretory system. (i) canal cell lumen. (ii) duct cell lumen and
autojunction is visible. (iii,iv) duct cell lumen. (v) duct and pore lumen. Part of the
excretory gland cell is visible in (i). ECM (lines) is visible in (iii-v). (vi) TEM of
detached inner eggshell layer (orange arrows).
E. let-4-4. Orientation is nearly transverse; sections go anterior as numbers increase. (i-
iii)TEM images of the excretory system. Pore opening is visible in (i). ECM (lines) is
visible in lumen in (ii) and (iii). (iv) TEM image illustrating detached inner eggshell layer
(orange arrow).
Figure Ap3.2. TEM images of the excretory system in wild-type 3-fold embryos. In
all panels, canal cell (c) is colored red; duct cell (d), yellow; pore cell (p) is colored blue.
A. N635B. Wild-type. The embryo is oriented so that generally, the dorsal side of the
worm is facing the top of the image. As section numbers increase, they move posterior.
(i) Secretory junction (ii) duct cell extension (iii,iv)pore cell lumen, (v) pore cell opening.
(vi-ix) Serial sections showing the duct/pore connection. Arrows indicate electron dense
junctions between the duct and pore cells.
B. N636. Wild-type. The embryo is oriented so the dorsal is facing the top of the image.
A section numbers increase, they move posterior. (i) duct cell body (ii) pore cell lumen,
and connection to the outside (iii) duct cell extension and pore cell lumen. (iv-viii) Serial
172

sections showing the duct/pore junction. Arrows indicate electron dense junctions
between the duct and pore cells.
C. N638A. Wild-type. The embryo is oriented so dorsal is facing the top of the image.
As section numbers increase, they move posterior. (i) duct cell body (ii-iii) secretory
junction (iv) canal cell body. (v-x) Serial sections showing the duct.pore connection.
Arrows indicate electron dense junctions between the duct and pore cells.
Figure Ap3.3. TEM images of the excretory system in let-4 3-fold embryos. In all
panels, canal cell (c) is colored red; duct cell (d), yellow; pore cell (p) blue.
A. let-4-8. Embryo with discontinuous duct lumen and swollen canal lumen. The embryo
is oriented so the dorsal side is generally facing the top of the image. As the sections
numbers increase, the sections are more posterior. (i) The canal cell is greatly distorted.
(ii) The lumen of the duct cell is continuous with the canal cell lumen. (iv-vii) Serial
sections showing the terminating duct cell lumen. The cuticle terminates in a bolus (iv).
B. let-4-9-2. Embryo with discontinuous duct lumen, no lumen swelling. The embryo is
oriented so the segment of the worm containing the excretory system is toward the top of
the image. In this segment, the dorsal portion is generally toward the top of the image.
As section numbers increase, the sections are more posterior. (i,ii) The secretory junction
and duct cell body. (iii,iv) The duct cell extension. Canals of the canal cell are also
visible.
173

C. let-4-7-1. Embryo with unknown connectivity, swollen duct and canal lumen. The
sections are oriented so that dorsal is generally facing up, as section numbers increase,
the sections move posterior. (i) Duct cell extension (ii) The secretory junction. The
lumen of the duct and canal cells is enlarged. (iii)The canal cell lumen is enlarged.
D. let-4-7-2. Embryo with unknown connectivity, swollen duct and canal lumen. Sections
are oriented so that the dorsal side of the worm is facing the top of the image. As section
numbers increase, the sections are more posterior. (i,ii,iii) duct cell lumen is enlarged;
pore lumen, autojunction, and pore opening (ii) is visible. Canals are also visible. (iv)
Canal cell lumen is enlarged. (v-vii) Serial sections showing the secretory junction.
From this orientation, sections are moving straight down the lumen from the duct cell
into the canal cell, making it difficult to know which section is the last duct section, and
which is the first canal cell section. Arrows indicate electron dense junctions.
E. let-4-10-3. Embryo with normal connectivity and lumen. The embryo is oriented so
the dorsal side is generally toward the top of the image. As section numbers increase, the
sections are more anterior. (i) Canal cell and duct cell bodies. (ii) secretory junction. (iii)
pore cell lumen (iv) pore cell opening. (v-viii) Sections adjacent to the duct.pore
connection. Sections 2131-2136, which contain part of the connection between the duct
and pore, were damaged.
F. let-4-10-9. Embryo with normal connectivity and lumen. The embryo is oriented
awkwardly because the cells of the excretory system are close to the end of the egg. As
section numbers increase, the sections are more ventral. (i,ii) Duct cell body and
174

extension (c) secretory junction. (iv) pore cell. (v-xii) Sections showing the duct cell-pore
cell connection. Arrows indicate electron dense junctions between the cells.
G. let-4-10-5. Embryo with normal connectivity and lumen. The embryo is oriented so
the dorsal side is facing the top of the image. As section numbers increase, the sections
are more anterior. (i) canal cell and duct cell bodies. (ii) secretory junction. (iii) duct cell
extension and pore cell. (iv) pore cell opening. (v) duct/pore cell connection. (vi-xii)
Serial sections showing the pore/duct connection.
H. let-4-10-7. Embryo with normal connectivity and lumen. The embryo is oriented so
the dorsal portion is toward the top of the image. As section numbers increase, the
sections are more anterior. (i) Secretory junction. (ii) Canal and duct cell bodies. (iii)
Duct cell extensions and pore cell opening. (iv-viii) Serial sections showing the
duct/pore connection.
I. let-4-10-8. Embryo with normal connectivity and lumen. The embryo is oriented
awkwardly because the cells of the excretory system are close to a fold in the embryo.
As section numbers increase, the sections are generally more posterior. (i-vi) Secretory
junction and pore cell lumen. (vii-xiii) Serial sections showing the pore/duct cell
connection. Arrows indicate electron dense junctions between the cells.
J. let-4-10-1 (i-ii) Duct cell lumen is enlarged, canal cell lumen is slightly enlarged.
Section numbers are unknown.
K. let-4-10-6 (i,ii) Excess fluid (orange asterisks) is present in the area of the terminal
pharyngeal bulb; more anterior sections are damaged.
175

c
d p
c
d p
c
dp
c d
p p
c
d
p
c
d
p
c
d
p
c
dp
let-
4-1
iv
2μm
d
p
i ii iii
sec280 sec284 sec285 sec280
sec740 sec719
sec706 sec703
sec1223 sec1224 sec1228 sec1232
i ii
iii iv
i ii iii iv
2μm
2μm
2μmsec1216
c
d
p
v
2μm
let-
4-2-
5
sec709
let-
4-2-
3
2μm
v
A.
B.
C.
Figure Ap3.1
176

sec396
c
d
p
c
d
p
c
d
p
cd
p
c
dp
cd
p
c
dp
c
d
p
sec388 sec396 sec398
sec400 sec401
sec453 sec454 sec455
let-
4-4
i ii iii
iv
i ii iii
v
2μm
2μm
sec4432μm
iv
vi
sec363
let-
4-3
c
dp
D.
E.
2μm
Figure Ap3.1, continued
177

N635B
sec465 sec466 sec468sec467p
d
2μm
i ii
iii iv v
vi vii viii ix
A.
2μm
Figure Ap3.2
178
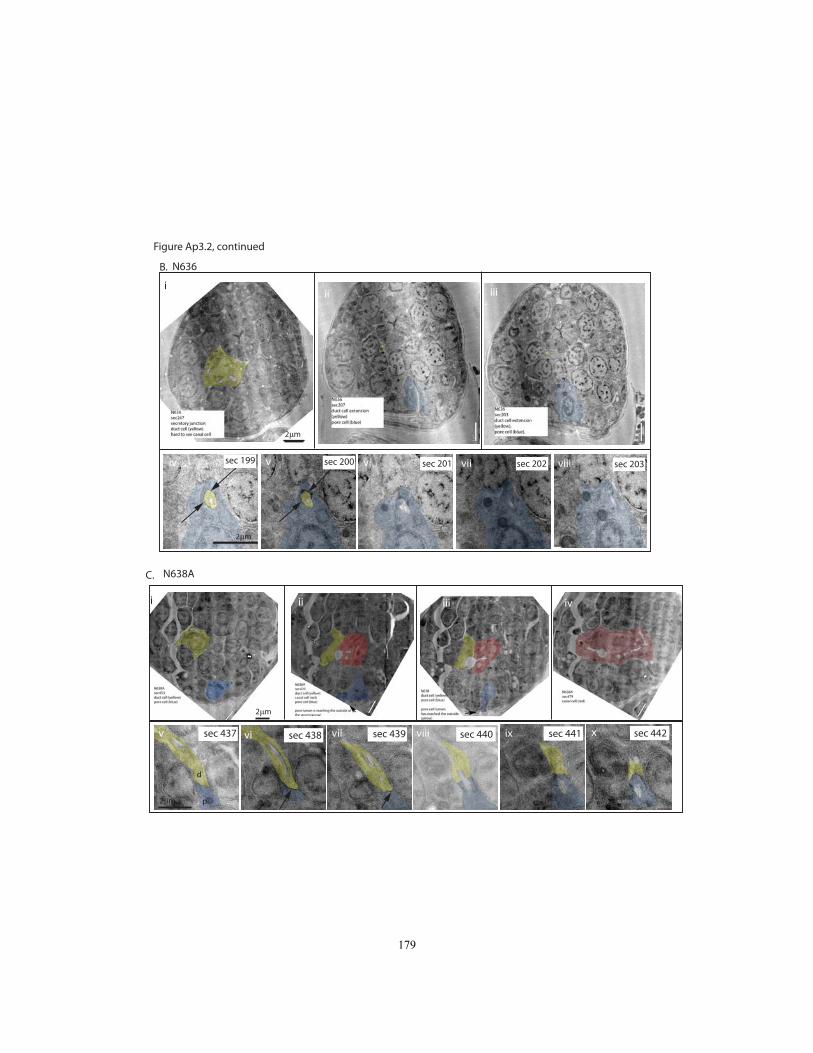
N636
sec 199 sec 200 sec 201 sec 202 sec 203
2μm
ii iii
iv v vi vii viii
i
B.
2μm
N638A
sec 437 sec 438 sec 439 sec 442sec 441sec 440
2μm
p
d
i ii iii iv
v vi vii viii ix x
C.
2μm
Figure Ap3.2, continued
179
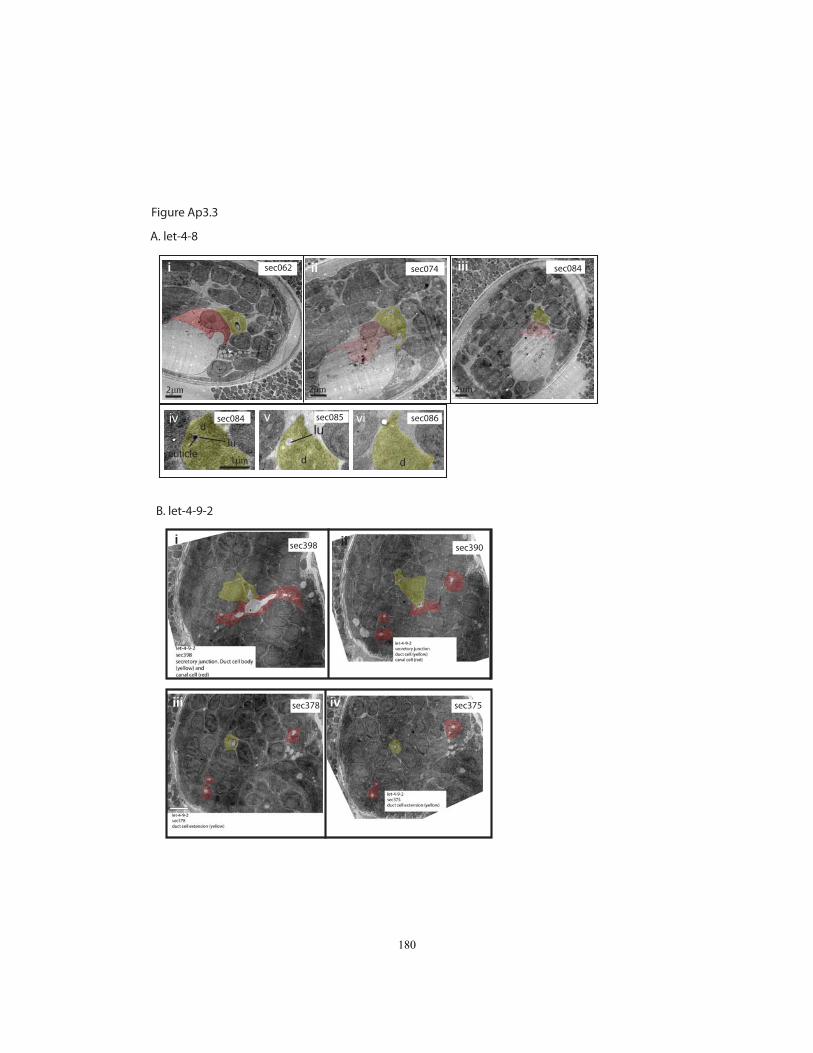
A. let-4-8
d
d dcuticle
lulu
sec062 sec084sec074
sec086sec085sec084
i ii iii
iv v vi
1μm
B. let-4-9-2
sec398
sec378 sec375
sec390i ii
iii iv
2μm
2μm 2μm 2μm
Figure Ap3.3
180

C. let-4-7-1
sec462sec489 sec506
2μm
i ii iii ivsec476
2um
D. let-4-7-2
sec496 sec497 sec498v vi vii
2μm
sec489sec480sec476
sec506ii ivi iii
2μm
Figure Ap3.3, continued
181

E. let-4-10-3
sec 2139 sec 2140sec 2138sec 2137
v vi vii viii
sec2155sec2140
sec2086sec2078i ii iii iv
2um
2um
F. let-4-10-9
sec2755 sec2756 sec2757 sec2758 sec2759 sec2761 sec2763 sec2765
v vi vii viii ix x xi xii
2um
i ii iii ivsec2746
sec2758 sec2755 sec2793
2um
Figure Ap3.3, continued
182
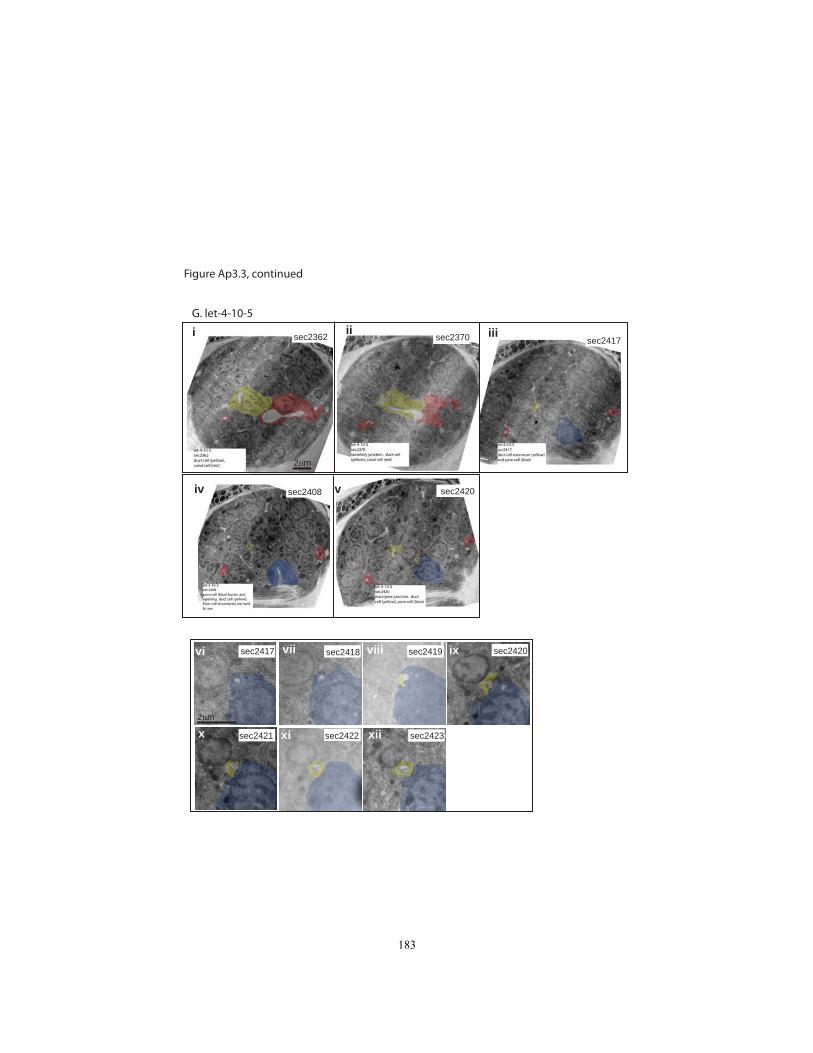
G. let-4-10-5
i ii iii
iv v
vi vii viii ix
x xi xii
sec2417 sec2418 sec2419 sec2420
sec2421 sec2422 sec2423
sec2362 sec2370 sec2417
sec2408 sec2420
2μm
2μm
Figure Ap3.3, continued
183
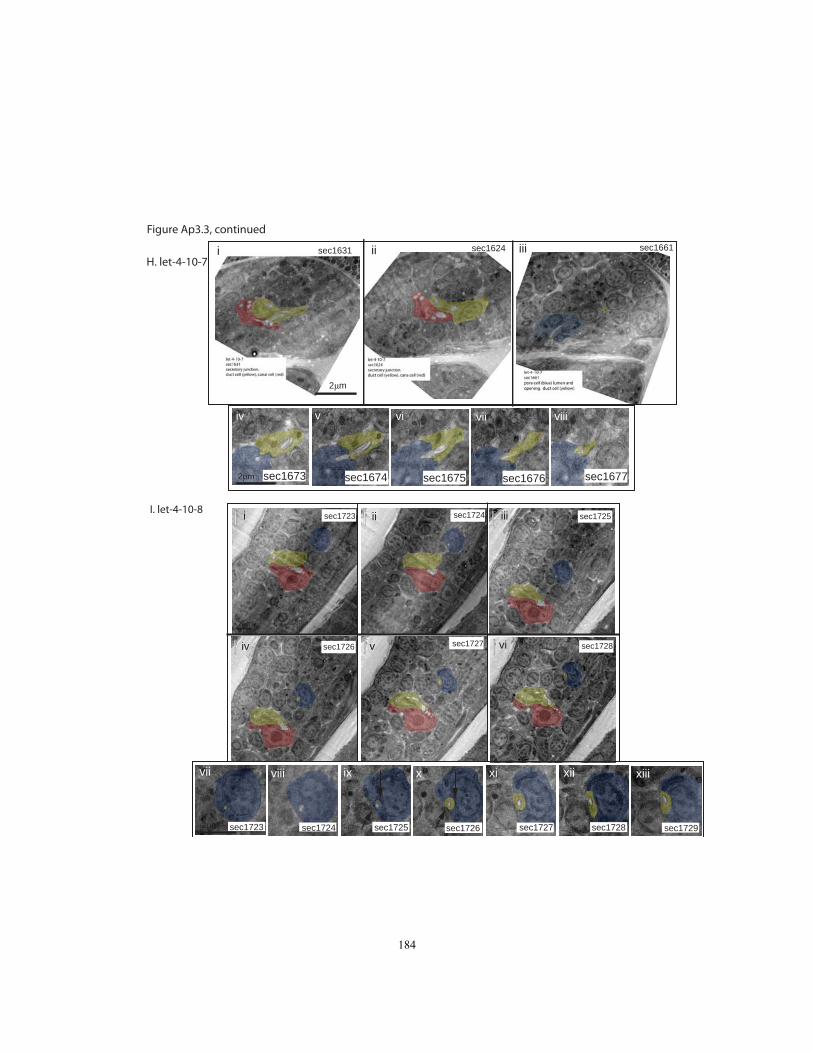
H. let-4-10-7
iv v vi vii viii
sec1673 sec1674 sec1675 sec1676 sec1677
i ii iiisec1631 sec1624 sec1661
2μm
2μm
2μm
i ii iii
iv v vi
vii viii ix x xi xii xiii
sec1723 sec1724 sec1725
sec1726 sec1727 sec1728
sec1723 sec1724 sec1725 sec1726 sec1727 sec1728 sec17292μm
I. let-4-10-8
Figure Ap3.3, continued
184
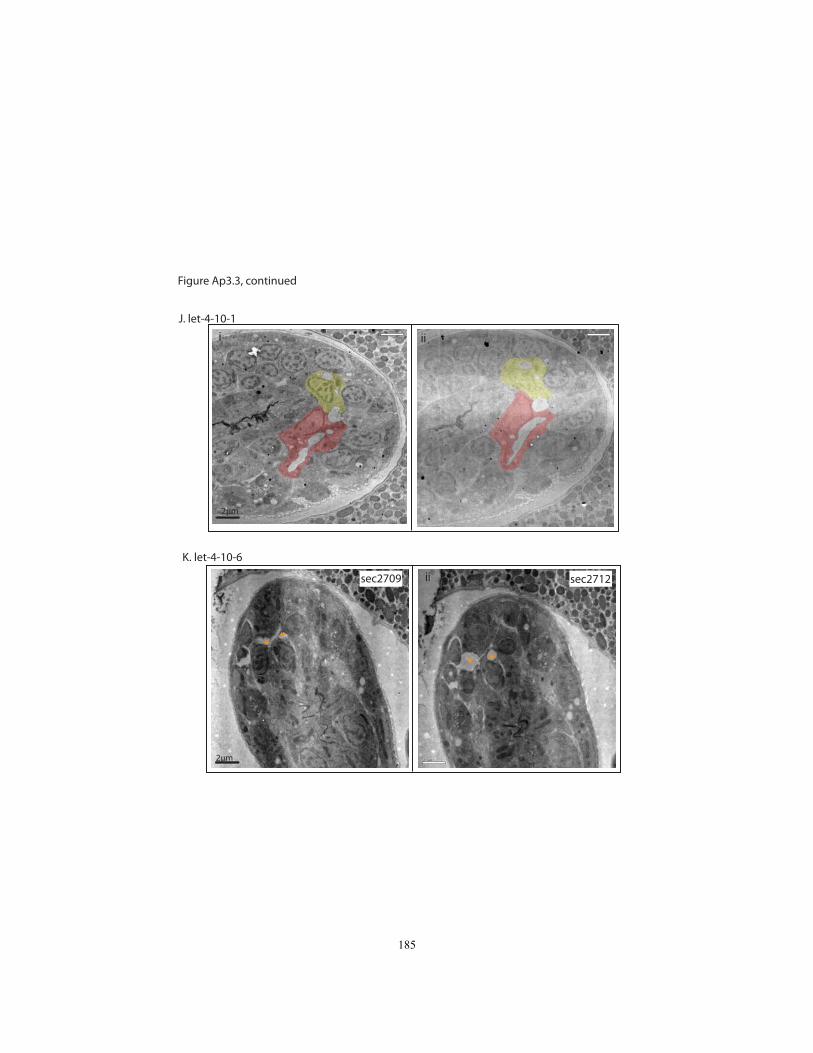
J. let-4-10-1
K. let-4-10-6
2μm
2μm
i ii
sec2709 sec2712i ii
** *
*
Figure Ap3.3, continued
185

N635B – wild type
N2 Age: 1.5-fold + 6hrs HPF: 12/1/09 Sectioned: 1/20/10 Scoped: 6/10
The embryo is oriented so that generally, the dorsal side of the worm is facing the top of the
image. As section numbers increase, they move posterior.
Key Sections:
Section#466 - pore/duct junction (pretty hard to make out) Sections #467-489- pore lumen Section# 489 – pore opening Sections#467-521 – duct cell 467-489- duct cell extension; cuticle visible in lumen in many sections 496-521- duct cell body 503- three passes of lumen through section. Sections# 503-511 – secretory junction Section#500-521- canal cell body lumen Sections#497-521- canal cell body, nucleus Cuticle visible in lumen in: Duct cell body: sec518 (also in most of the cell body) Duct cell extension: sec465-467 Pore cell lumen: sec468-478 Closest to sec junc: sec500
Updated 7/12/10
186

N636 – wild type
N2 Age: 1.5-fold + 6hrs HPF: 12/1/09 Sectioned: 1/24/10 Scoped: 6/10
As of 7/10/10: I don’t have the canal cell lumen. I’ve scoped Excretory System sections #199-
248
The embryo is oriented so the portion containing the ExSys is toward the top of the image.
Dorsal is facing the top of the image. A section numbers increase, they move posterior.
Key Sections: Sections #199-204- pore cell nucleus Sections #199-207- pore cell lumen Sections #204-207- pore cell opening Sections # 207-213- pore cell autojunction Sections #199-215- duct cell lumen, with cuticle, is visible throughout these sections Sections #190-230 – duct cell extension Sections #230 -248*- duct cell body Sections #199-201- duct/pore junction Sections #244-248* – secretory junction *Section#248 is the highest section number I’ve looked at.
Missing pieces: The rest of the secretory junction, and the canal cell are in higher section
numbers
Cuticle visible in lumen in:
Duct cell body: sec239-247 (scaffold not visible in 248) Duct cell extension, along the entire length of the extension. Pore lumen: sec200,201,207 Cuticle closest to sec. junc: sec247
187

N638A – wild type
N2 Age: 1.5-fold + 6hrs HPF: 12/1/09 Sectioned: 1/28/10 Scoped: 6/10
The embryo is oriented so the portion containing the ExSys is toward the top of the image.
Dorsal is facing the top of the image. As section numbers increase, they move posterior.
Key Sections: Section #450- end of pore cell nucleus Section #470- pore cell opening Sections #440-470- pore cell lumen Sections #437-439- pore/duct junction Sections #448-464- duct cell nucleus Sections# 432-442 –duct cell extension Sections #432- ~468- duct cell lumen
#450-454- duct cell lumen cuticle is visible Sections #466-475- secretory junction Sections #472-493- canal cell nucleus Sections#469-480- canal cell body lumen Cuticle visible inside lumen in: Duct cell body: sec# 451-454 Duct cell extension: sec# 438-439 Pore cell lumen: sec# 463-470 Closest to sec. junc: sec461
188

let-4-8 – duct lumen ends without connecting to pore; severe
fluid damage
UP1787(without GFP-marked rescuing transgene): let-4(mn105) Age: 1.5-fold + 6hrs HPF : 12/1/09 Sectioned: 1/3/10
Scoped: mostly 4/10 Result: lumen does not extend all the way through the duct. This embryo has a lot of fluid accumulation. The fluid can be traced back to the EC (excretory canal cell) lumen, which connects to the duct lumen, which extends into the duct, but doesn’t go all the way through the duct. I can’t identify the pore cell due to all the damage done to the embryo by the fluid accumulation. The worm is in one piece in the sections containing the canal cell and duct cells. These sections begin near the very beginning of the embryo. The embryo is oriented so the dorsal side is generally facing the top of the image. As the sections numbers increase, the sections are more posterior.
Key Sections: Section #035 Canal cell lumen is already in view, part of the canal cell lumen is in view in all the sections I’ve looked at. Section #050 Excretory canal cell (red), appears to contain the pocket of fluid. The cell just below the canal cell in this image is the cell that will form junctions with the canal cell, likely the gland cell. Sections #056-070 I see the series of bubbles which look like they’re in a cell, but the membrane containing them doesn’t seem to connect to any of the cells I’m interested in, or a nucleus anywhere. Sections #059-074 Secretory junction.
Section#062 Junctions are visible between the excretory canal cell (red) and another cell, probably the gland cell (purple). The duct cell (yellow) is next to the excretory cell, and its lumen is visible. Section#074 is the section with the best view of what’s left of the damaged duct/canal cell junction. The lumen of the duct cell (yellow), the lumen of the excretory cell (red), and the pocket of fluid are continuous. The duct cell and gland cell come in contact, although dark junctional material isn’t apparent.
Sections #058-089 - duct cell body Sections #060-076 Duct cell lumen In following these sections, it’s apparent that the duct cell lumen is continuous with the canal cell lumen which is continuous with the large pocket of accumulated fluid. Sections #064-085 Duct cell nucleus. Cuticle visible in lumen in: Duct cell body: sec63-85, (only sec062 looks like it’s not detached) Duct cell extension: N/A Pore cell: N/A Detachment closest to sec. junc:sec75 (it’s difficult to tell, with all the damage)
189

let-4-9-2 – duct lumen ends without connecting to pore; no fluid accumulation
UP1787(without GFP-marked rescuing transgene): let-4(mn105)
Age: 1.5-fold + 6hrs HPF : 12/1/09 Sectioned: 1/4/10 Scoped: mostly 5/10
Result: The duct lumen just seems to stop in grid C3. The excretory canal cell (EC) looks normal;
the secretory junction is present; I can’t identify the pore, even though I’ve gone up to section
#262, ~90 sections anterior of where the duct cell lumen ends.
The embryo is oriented so the segment of the worm containing the excretory system is toward the top of the image. In this segment, the dorsal portion is generally toward the top of the image. As section numbers increase, the sections are more posterior.
I’m missing sections #371-373. I think the thing I was calling lumen before #371 may just be a
random vacuole, and I’m losing track of the lumen in this tiny gap.
Key Sections: Sections#354-417 duct cell lumen
#354- this is the first bit of duct lumen I can find; I believe it’s the anterior end of the duct cell lumen. When I trace the duct lumen from the opposite direction, this is as far as I can follow it. #374-387 duct cell nucleus (below the lumen) #375-386 cuticle is obvious #417- last bit of duct lumen in this direction, it winds back toward the anterior end #376-417 duct cell body #354-374 duct cell extension (it’s hard to tell exactly where the body/extension
boundary is in this orientation, so I made my best guess) Sections# 391-396 secretory junction, #391 is the best view of the sec. junc. Sections# 392-403 canal cell lumen Section# 420- canal cell nucleus first becomes visible
Cuticle visible in lumen in:
Duct cell body: in all lumen sec376-417 Duct cell extension: cuticle isn’t visible in the duct in sections lower than sec#375 Pore cell: N/A Cuticle closest to sec. junc: sec398 190

let-4-7-1 – Fluid in duct and canal cell; gaps/missing
sections
UP1787(without GFP-marked rescuing transgene): let-4(mn105)
Age: 1.5-fold + 6hrs HPF : 12/1/09 Sectioned: 12/22/09 Scoped: mostly 3/10 Result: The duct lumen is connected to the EC lumen, but I can’t find it ever reaching a pore. I
lose the lumen in the gap between sections 454 and 458; I am missing some crucial sections
around this area that are damaged and folded over; the pore is likely located in these sections.
Conclusions I can make from this embryo:
The duct cell is present, has a lumen, and the lumen connects with the EC lumen
EC is present, with a lumen, and a few bubbles of fluid accumulation.
The sections are oriented to that the dorsal part of the segment of the worm containing the
excretory system is generally facing up. The embryo is in two large segments in the lower
sections numbers, and the sections continue to the end of the embryo.
Key images: Sections #462-463- Duct cell extension Sections #475-485- duct cell body Sections #458-489- Duct cell lumen
Section#463 Narrow part of duct lumen; I don’t ever get to see where this lumen leads to, due to damaged sections. From the opposite direction, I can follow the lumen from the cell body (with some gaps) but I don’t know what happens at the anterior end of the lumen. Notice dark ring of cuticle inside duct cell lumen Section #476 Lumen is continuous through duct (yellow) and EC(red), some of the secretory junction is visible; duct nucleus is visible. The duct lumen and EC lumen each pass through this section twice. (I don’t have the sections where the two pieces of duct lumen connect, but I am missing #464-474 and #477-479.) Cuticle is still present in the duct cell lumen.
Sections #475-480 –secretory junction Sections #475-509- Canal Cell body lumen
#481-496- There are clearly a few bubbles of fluid accumulation in the EC lumen. Why are these bubbles right next to each other; what gives them their shape? Also, notice the dark shading inside the EC lumen.
Cuticle is visible in the lumen in: Duct cell body: sec475,476; but not in sec480-496. Duct cell extension: cuticle is visible in all the sections I have: sec458,461,462,463 Pore cell: N/A; Cuticle closest to sec. junc: sec475 (there is a gap next to this section) 191

let-4-7-2 - fluid in duct and canal cell; gaps/missing sections UP1787(without GFP-marked rescuing transgene): let-4(mn105)
Age: 1.5-fold + 6hrs ;HPF : 12/1/09; Sectioned: 12/22/09; Scoped: mostly 3/10 Result: Grids C1 and C2, which contain crucial sections, had a lot of folded and damaged
sections, so I can’t be sure of ExSys lumen continuity. What I can be sure of is:
Pore cell is present with autocellular junctions, a lumen, and a pore opening.
Duct lumen is connected to EC lumen, and at least partially present.
EC is present, with a lumen and fluid accumulation
Sections are oriented so that the portion of the worm which contains the excretory system
is the top segment, and the dorsal side of this segment is toward the top of the image. The
pharynx is visible throughout these sections, and the cells of the excretory system appear
ventral (below) the pharynx. Section numbers increase as we move posterior.
Key sections:
Sections #475-483 Pore lumen; autocellular junction. Section#s 469-474 and 477-479 are torn/damaged, so I can’t see if the pore lumen connects to the duct lumen, but the pore autojunction is still intact, so it’s probably attached to the duct cell. In the colorized image of section #480, the duct cell is yellow, and the pore cell is blue. 475-476; 480-482: pore lumen
480-482: pore opening Sections #457-490 Duct cell lumen. I can follow it to the secretory junction in the posterior direction, but I can’t see where it ends, or where it connects in the anterior direction. I lose track of the lumen in the gap between #454 and 457. I’m also missing 445-453. 458-496: duct cell body Sections #496-498 Secretory junction; we’re transitioning from the duct cell part of the lumen to the excretory cell. The lumen is swollen on both sides of the junction, and at the junction. The excretory cell nucleus is coming in to view Sections #501-519 Canal Cell main body lumen (canal lumen is visible in all these sections) (Dark shading is visible inside lumen in #501-507) Are the two large pockets of fluid ventral to the EC nucleus continuous? Section #506 Excretory cell nucleus and two pockets of fluid. This embryo has two adjacent pockets of fluid; what is keeping them separate? Section# 507 This is the one section where the two adjacent pockets of fluid seem to connect, but it almost looks like the tissue between the two pockets of fluid was damaged, maybe during freezing. Cuticle visible in the lumen in: Duct cell body: visible in the whole lumen from sec458-496 Duct cell extension: N/A Pore cell: cuticle is visible in sec480,481,482 (not visible in sec475,476) Cuticle closest to sec junc:490 (there is a gap between this section and the sec junc.)
192

let-4-10-3 – no fluid accumulation; lumen is continuous
UP1787(without GFP-marked rescuing transgene): let-4(mn105)
Age: 1.5-fold + 6hrs HPF : 12/1/09 Sectioned: 1/5/10 Scoped: mostly 6/10 Result: The lumen is present, and appears continuous through the excretory canal cell (EC),
duct, and pore (although it’s very hard to follow the duct-pore connection around section
#2109-2127
The embryo is oriented so the segment of the worm containing the excretory system is toward the top of the image. In this segment, the dorsal portion is generally toward the top of the image. As section numbers increase, the sections are more anterior. The excretory system is cut through in cross sections. As Section #s increase, we move from the EC nucleus and lumen, to the duct lumen, then the pore lumen. Key Sections: Sections#2059-2086: canal cell nucleus; canal cell body lumen is visible from 2056-2122. Sections#2064-2098- canal cell body lumen Sections #2086-2095: secretory junction, and lumen on both sides of the junction Sections #2065-2138: duct lumen- contains cuticle 2064-2096- duct cell body
2098-2138- duct cell extension 2068- duct lumen passes through section once 2069 duct lumen passes through section twice 2086-2091 duct lumen is connected to canal cell lumen 2100-2126 sections are damaged, but duct lumen is still there. 2127-2138 duct lumen reaches across the plane of the section, toward the pore, the lumen is really tiny here. 2132-2136 are missing. I think this was a section numbering error. Based on the sections, I’m not missing much between #2131 and #2137, and there weren’t any more sections on the grids. Section#2137- duct/pore junction Sections# 2139- 2156 pore lumen; pore auto junction visible; cuticle present in pore lumen (is pore lumen narrow?) 2137-2160- pore nucleus
2139-2157- pore lumen 2157 pore opening
Cuticle visible in lumen in: Duct cell body: anywhere from sec2064-2096 Duct cell extension: clearest in sec2182 Pore: anywhere from 2139-2157 Cuticle closest to sec junc: sec2086
193

let-4-10-9 – no fluid accumulation; lumen is continuous
UP1787(without GFP-marked rescuing transgene): let-4(mn105)
Age: 1.5-fold + 6hrs HPF : 12/1/09 Sectioned: 1/7/10 Scoped: 7/10 Result: The lumen is present, and appears continuous through the canal cell, duct, and pore;
there is no excess fluid accumulation in the lumen or canals.
The embryo is oriented awkwardly because the cells of the excretory system are close to the end of the egg. As section numbers increase, the sections are more ventral.
Sections lower than 2738 may have a little duct cell cytoplasm, but I can’t visualize them
because the grid was damaged.
The duct cell lumen begins at the secretory junction in sections #2757-2772, then winds through
the body, then moves into the duct cell extension, moving toward the top of the image, as
section numbers increase, then the lumen connects to the pore cell, and continues through
lower section numbers. The duct/pore junction is at 2758, then the pore cell is narrow for some
sections, before the pore cell nucleus comes into view at about section #2781.
Key Sections:
Sections #2759-2772: canal cell lumen Sections #2757-2772: secretory junction 2759, 2761: lumen on both sides of junction Sections #2738-2758: duct cell lumen
2738-2765- duct cell body 2740-2758- duct cell extension
Sections #2758 pore/duct junction Sections #2758-2798: pore cell 2758-2798: pore autojunction/lumen 2781-2798: pore cell nucleus 2810: as far as I can go toward the pore opening Cuticle is visible inside the lumen in: Duct cell body : sec# 2757-2755 Duct cell extension: sec#2743,2744 Pore cell lumen: sec# 2761-2765; 2775 Closest to secretory junction: sec# 2757 194

let-4-10-5 – no fluid accumulation; lumen is continuous
UP1787(without GFP-marked rescuing transgene): let-4(mn105)
Age: 1.5-fold + 6hrs HPF : 12/1/09 Sectioned: 1/7/10 Scoped: 6/10
Result: The lumen is present, and appears continuous through the canal cell, duct, and pore;
there is only a little excess fluid accumulation in the canal cell, and maybe some swelling in the
duct.
The embryo is oriented so the segment of the worm containing the excretory system is toward the top of the image. In this segment, the dorsal portion is generally toward the top of the image. As section numbers increase, the sections are more anterior. The duct lumen is a little confusing; the duct cell process is up against the pharynx and goes in the anterior direction, anterior of the pore, up to section 2433, where it bends ventrally, then moves posterior to connect with the pore at section 2421
Key Sections: Sections #2349-2372 excretory canal cell lumen (posterior part of lumen is in lower section numbers) #2349-2367 canal cell nucleus Sections #2367-2373 Secretory junction 2367-2371: lumen on both sides of the junction Sections #2351 -2433 Duct cell 2355-2373: duct cell nucleus 2349-2382: duct cell body 2383-2433, then winding back to 2420: duct cell extension (with lumen) Section #2420 – duct/pore junction Sections #2404-2433 Pore cell #2404-2406 pore autojunction #2406-2408 pore opening #2409-2420 pore lumen
#2412-2433- pore cell nucleus Cuticle visible in lumen in: Duct cell body: basically visible in all lumen, especially from sec2360-2373 Duct cell extension: all the way through the extension: sec2383-2433 Pore cell: sec2417,2418,2419; sec2406,2407,2408 Cuticle closest to sec junc: sec 2370
195

let-4-10-7 – no fluid accumulation; lumen is continuous
UP1787(without GFP-marked rescuing transgene): let-4(mn105)
Age: 1.5-fold + 6hrs HPF : 12/1/09 Sectioned: 1/5/10 Scoped: 6/10
Result: The lumen is present, and appears continuous through the canal cell, duct, and pore;
there is no excess fluid accumulation in the lumen or canals.
The embryo is oriented so the segment of the worm containing the excretory system is toward the top of the image. In this segment, the dorsal portion is generally toward the top of the image. As section numbers increase, the sections are more anterior.
Sections 1587-1595 are damaged. These sections contain the canal cell nucleus, and some of
the canal cell lumen.
Key Sections: Sections #1596-1624: canal cell lumen #1603-1624- canal cell lumen is unusually narrow #1627-1630- lumen connects to duct cell lumen Sections #1624-1630: secretory junction Sections #1618-1675: duct cell lumen
1610-1639- duct cell body 1641-1676- duct cell extension
Sections #1673-1677 pore/duct junction 1675: pore/duct junction, with lumen on both sides Sections #1659-1677: pore cell 1659-1661: pore autojunction 1660-1664: pore opening 1660-1675: pore lumen- some parts are hard to follow, use autojunction as a guide
Cuticle in lumen visible in:
Duct cell body: visible in all the cell body lumen, especially 1621-1631 Duct cell extension: all the way through, sec1641-1676 Pore cell: the whole lumen: sec1660-1664 Cuticle closest to sec junc: sec1633 196

let-4-10-8 – no fluid accumulation; lumen is continuous
UP1787(without GFP-marked rescuing transgene): let-4(mn105)
Age: 1.5-fold + 6hrs HPF : 12/1/09 Sectioned: 1/6/10 Scoped: mostly 8/10
Result: The lumen is present, and appears continuous through the canal cell, duct, and pore;
there is no excess fluid accumulation in the lumen or canals. It’s very difficult to see the cuticle
inside the lumen in this animal.
The embryo is oriented awkwardly because the cells of the excretory system are close to a fold in the embryo. As section numbers increase, the sections are generally more posterior.
Key Sections: Sections #1720-1733: canal cell lumen Sections #1724-1729: secretory junction 1725,1726: lumen on both sides of junction Sections #1724->1744->1726: duct cell lumen
1723-1742- duct cell body 1738->1744->1726 - duct cell extension (it reaches toward higher section numbers, then turns back at section #1744)
Sections #1725-1726: pore/duct junction Sections #: pore cell (I’m missing a few sections in the pore lumen) 1700-1725: pore autojunction/lumen 1718-1732: pore cell nucleus 1700: pore cell opening Cuticle inside lumen visible in: Duct cell body: sec1738 Closest to sec. junction: 1727 Pore cell: sec1701
197

Appendix 4
Detailed Protocols
198

TEM Grid Staining Protocol – From the Hall Lab and Electron Microscopy
Sciences Protocol.
I. Filter solutions:
1) Attach filter to syringe, pull plunger all the way out
2) Decant solution into syringe
3) Press plunger in until *snap*
4) Pull plunger back
II. Stain Grids:
1) Create two staining chambers with plates or petri dishes with a piece of filter
paper in each. Label one UrAc and the other lead citrate.
2) Fill 4 small beakers with CO2-free H2O.
3) Apply 1M NaOH to filter paper in each chamber, just enough for the paper to
be saturated.
4) Place parafilm on top of wet filter paper. Place several NaOH pellets in the
lead citrate petri dish.
5) Using the filtered syringe, place one drop of each staining solution on the
parafilm. You will need a different drop of solution for each grid to be
stained.
6) Using forceps, take out the grid to be stained. Place the grid face down on the
drop of UrAc, and leave it for 4 minutes. (In the Hall lab, grid boxes are
prepared so the top face of the grid is facing to the left.)
7) Take the grid off the UrAc, and dip it into the first beaker of H2O 5 times,
submerging it for 10 seconds each time. Then, dip the grid into the next
beaker of H2O the same way 5 times.
8) Place the grid face down in the drop of lead citrate, and leave it for 2 minutes.
9) Take the grid off the lead citrate, then dip it into the 3rd beaker 5 times, then
4th
beaker 5 times.
10) Allow the grid to air dry in the forceps, then return the grid to the grid box.
Solutions:
I. CO2-free H2O
Boil mQH2O for 1 minute, and store it immediately in tightly-capped containers.
Let the water cool completely before using it for staining. Open the containers
immediately before use; there should be a hissing sound when the cap is opened.
199

II. 2% Uranyl Acetate in H2O. – UrAc will go into solution best if H2O is 60°C.
III. Lead Staining Stock Solution (Reynold’s)
1) In a 50 ml flask add: Lead Nitrate Pb(NO3)2 1.33 g Sodium Citrate
Na3(C6H5O7).2H2O 1.76 g Boiled, cooled, CO2 free dist. water 30 mL
2) Cover the flask, shake vigorously for at least one minute.
3) Add 8 ml of 1N NaOH (in CO2 free H2O)
4) Mix until clear.
5) Check the pH. It should be 12.0 ±0.1. If the pH is low add more NaOH to the
clear solution. If the pH is above 12.1 start over this time adding a smaller
amount of NaOH.
6) Add CO2-free water to bring the solution to a final volume of 50 mL.
7) Let stand several hours before use.
8) Filter stock solution, then store it (protected from light) at 4°C for 2 days
before use.
9) Use a 1:100 dilution of this stock to stain grids.
200

Antibody Staining Protocol –Modified from Finney and Ruvkin 1990 and Anne
Marie McKnight
I.Fixation:
1) Wash worms from ~4 crowded plates with M9 into 15mL conical tube, spun
down 1.5krpms, for 1 min.
2) Wash worms with M9 3 times, spinning down and removing supernatant after
each wash.
3) Wash worms with mQH2O
4) Resuspend pellet in mQH2O to a total of 900µL
5) In hood- add 1mL 2XRFB and 110uL Formaldehyde (37%)
6) Mix gently, parafilm tube
7) Freeze in dry ice/ethanol mix (still in 15mL conical), then store the frozen worms
at -80°C (Worms can stay like this overnight)
8) Remove tube from -80°C. Thaw under warm water,; place on ice/water bath for 3
hrs, gently shaking every half hour.
9) Spin at 1krpm for 1 min.
10) Wash 2x with 2mL TTE
11) After second wash, add 2 mLs of TTE +1% β-mercaptoethanol.
12) Incubate worms in 37°C water bath for 3 hrs, mixing gently every half hour.
13) Spin 1000rpm for 1 min.
14) Remove supernatant (in hood)
15) Wash with 2mLs 1xBO3 buffer +10mM DTT. Remove Super.
16) Add 2mLs 1xBO3 Buffer
17) Incubate for 15mins at 37°C
18) Spin, remove super, wash with 2mLs 1XBO3.
19) Add 2mL 1xBO3 + 0.3% H2O2. Incubate for 15mins at 37°C.
20) Spin down and remove super. Wash with 2mL 1xBO3, remove super.
21) Add 3mL PTC. Rock at RT for 30min.
22) Spin down and remove super. Add 2 mL PTB.
23) Store worms at 4°C.
II.Staining:
1) Transfer 40uL packed fixed worms to a microcentrifuge tube.
2) Add primary antibodies and PTB, up to 200uL
3) Rock overnight at 4°C.
4) Spin down 1500RPM for 1 min.
201

5) Remove super and wash with 1mL PTC
6) Spin, remove super, resuspend in 1mL PTC
7) Shake/rock 1 hr.
8) Repeat 2 more times
9) Final wash with PTB
10) After last wash, add PTB to 40uL.
11) Add secondary antibodies in PTB up to 200uL
12) Rock for 2.5hrs at room temperature.
13) Immediate and 1 hr PTB wash
14) Resuspend in 200uL PTC, and rock overnight.
15) Quick wash and 1 hr wash with PTC.
16) Store at 4°C in 200uL PTC. Worms are ready to mount.
Solutions for antibody staining:
1) 5xPBS pH 7.0-7.2
7.31g NaCl
2.36g Na2HPO4
1.31g NaH2PO4*2H2O
Add dH2O up to 1L
2) 2XMRWB (2XRFB)
5mL 0.2M EGTS
5mL 0.3M PIPES
8mL 1.0 M KCl
2mL 1.0M KCl
0.5mL 1.0M spermadine
pH to 7.4 with HCl
add dH2O to 25mLs (~4.5mL)
add 25mLs Methanol
3) TTB (TTE)
10mL 1M Tris *Cl pH7.4
200uL 0.5M EDTA
1mL Triton X-100
Add dH2O to 100mL (~89mL)
4) 100xBO3 (Borate) Buffer
1M H3BO3, pH9.2
5) PTB ( aka AbA)
48.72mL 1xPBS
0.5g BSA
100uL 0.5M EDTA
202

0.781mL 1M NaN3
250µL Triton X-100
6) PTC ( aka AbB)
194.9 mL 1xPBS
0.2g BSA
1.0 mL 1.5M EDTA
3.1 mL 1M NaN3
1.0mL Triton X-100
203

Freeze-Crack Staining Protocol
Adapted from the Priess lab (Leung et al 1999).
Freeze embryos by placing them in a drop of M9 on a poly-L-lysine coated slide (from
4degC incubator) and dropping a coverslip on top, leaving some overhang, for easy
removal of the coverslip. Use as small a drop of M9 as possible, so the embryos don’t
get washed away when you drop on the coverslip. Place the slide on a metal plate on dry
ice. (The metal plate should be pre-cooled on the dry ice for at least an hour before
placing the slide on it)
Crack coverslip off of slide after at least 1 hr on cold metal plate, then immediately
proceeded with the following steps:
15 min in -20degC Methanol – after methanol wash, let the slide air dry for 30 sec-1min.
30 min PBS-T wash (PBS + 0.1% Tween-20) x2
30 min Block step (PBS + 1% BSA)
Overnight room temperature incubation of Primary antibody
30 min PBS-T wash (PBS + 0.1% Tween-20) x2
4hr room temperature incubation with Secondary antibody
Rinse with PBS.
Mount embryos with ~50% PBS and 50% mounting media.
204

General C. elegans embryo ablation protocol
I. Prepare laser:
1) Place a slide with a coverslip (and no crucial samples) onto the microscope.
Focus on coverslip of the slide.
2) Beginning with the attenuator on a very low setting, fire at the coverslip. Step
up attenuator (and therefore laser power) one notch at a time, until one shot of
the laser can crack the coverslip.
3) Shift the attenuator back down one step, and use this attenuator setting for
ablations.
II. Mount embryos:
The key is minimizing the amount of time the embryo spends on a slide.
1) Pick a few embryos at a time onto a slide with a 4% Noble Agar pad.
2) If the embryo is not the desired stage, fire at the surface off the eggshell to
destroy the embryo so it will not be recovered from the slide.
3) Focus on the nucleus of the target cell, and fire. Record the number of
“clicks” of the laser. The number of clicks required, as well as the best
attenuator setting will depend on the target cell, and the age of the dye in the
laser. Perform the ablations quickly, then remove the slide from the scope.
III. Recover embryos:
1) Pop off the coverslip. To generate a useful tool for this, use a razor to sharpen
one edge of a pipette tip, then wedge the tip between the glass slide and the
overhang of the coverslip. Use the modified pipette like a lever to pop off the
coverslip.
2) Recover the ablated embryos (off of the agar pad, or the coverslip) using a
pick covered with a lot of bacteria.
205

Bibliography
Abdus-Saboor, I., Mancuso, V. P., Murray, J. I., Palozola, K., Norris, C., Hall, D. H.,
Howell, K., Huang, K., Sundaram, M. V., 2011. Notch and Ras promote
sequential steps of excretory tube development in C. elegans. Development. 138,
3545-55.
Achilleos, A., Wehman, A. M., Nance, J., 2010. PAR-3 mediates the initial clustering and
apical localization of junction and polarity proteins during C. elegans intestinal
epithelial cell polarization. Development. 137, 1833-42.
Acloque, H., Adams, M. S., Fishwick, K., Bronner-Fraser, M., Nieto, M. A., 2009.
Epithelial-mesenchymal transitions: the importance of changing cell state in
development and disease. J Clin Invest. 119, 1438-49.
Ameye, L., Young, M. F., 2002. Mice deficient in small leucine-rich proteoglycans:
novel in vivo models for osteoporosis, osteoarthritis, Ehlers-Danlos syndrome,
muscular dystrophy, and corneal diseases. Glycobiology. 12, 107R-16R.
Anderson, D. C., Gill, J. S., Cinalli, R. M., Nance, J., 2008. Polarization of the C. elegans
embryo by RhoGAP-mediated exclusion of PAR-6 from cell contacts. Science.
320, 1771-4.
Bafna, S., Kaur, S., Batra, S. K., 2010. Membrane-bound mucins: the mechanistic basis
for alterations in the growth and survival of cancer cells. Oncogene. 29, 2893-904.
Bao, Z., Murray, J. I., Boyle, T., Ooi, S. L., Sandel, M. J., Waterston, R. H., 2006.
Automated cell lineage tracing in Caenorhabditis elegans. Proc Natl Acad Sci U S
A. 103, 2707-12.
Bar, T., Guldner, F. H., Wolff, J. R., 1984. "Seamless" endothelial cells of blood
capillaries. Cell Tissue Res. 235, 99-106.
Bastiani, C., Mendel, J., 2006. Heterotrimeric G proteins in C. elegans. WormBook. 1-
25.
Baum, B., Georgiou, M., 2011. Dynamics of adherens junctions in epithelial
establishment, maintenance, and remodeling. J Cell Biol. 192, 907-17.
Beitel, G. J., Tuck, S., Greenwald, I., Horvitz, H. R., 1995. The Caenorhabditis elegans
gene lin-1 encodes an ETS-domain protein and defines a branch of the vulval
induction pathway. Genes Dev. 9, 3149-62.
Benenati, G., Penkov, S., Muller-Reichert, T., Entchev, E. V., Kurzchalia, T. V., 2009.
Two cytochrome P450s in Caenorhabditis elegans are essential for the
organization of eggshell, correct execution of meiosis and the polarization of
embryo. Mech Dev. 126, 382-93.
Bernadskaya, Y. Y., Patel, F. B., Hsu, H. T., Soto, M. C., 2011. Arp2/3 promotes junction
formation and maintenance in the Caenorhabditis elegans intestine by regulating
membrane association of apical proteins. Mol Biol Cell. 22, 2886-99.
Bernadskaya, Y. Y., Patel, F. B., Hsu, H. T., Soto, M. C., 2011. Arp2/3 promotes junction
formation and maintenance in the Caenorhabditis elegans intestine by regulating
membrane association of apical proteins. Mol Biol Cell. 22, 2886-99.
Berrier, A. L., Yamada, K. M., 2007. Cell-matrix adhesion. J Cell Physiol. 213, 565-73.
Berry, K. L., Bulow, H. E., Hall, D. H., Hobert, O., 2003. A C. elegans CLIC-like protein
206

required for intracellular tube formation and maintenance. Science. 302, 2134-7.
Bokel, C., Prokop, A., Brown, N. H., 2005. Papillote and Piopio: Drosophila ZP-domain
proteins required for cell adhesion to the apical extracellular matrix and
microtubule organization. J Cell Sci. 118, 633-42.
Bosher, J. M., Hahn, B. S., Legouis, R., Sookhareea, S., Weimer, R. M., Gansmuller, A.,
Chisholm, A. D., Rose, A. M., Bessereau, J. L., Labouesse, M., 2003. The
Caenorhabditis elegans vab-10 spectraplakin isoforms protect the epidermis
against internal and external forces. J Cell Biol. 161, 757-68.
Bossinger, O., Fukushige, T., Claeys, M., Borgonie, G., McGhee, J. D., 2004. The apical
disposition of the Caenorhabditis elegans intestinal terminal web is maintained by
LET-413. Dev Biol. 268, 448-56.
Bossinger, O., Klebes, A., Segbert, C., Theres, C., Knust, E., 2001. Zonula adherens
formation in Caenorhabditis elegans requires dlg-1, the homologue of the
Drosophila gene discs large. Dev Biol. 230, 29-42.
Boyle, T. J., Bao, Z., Murray, J. I., Araya, C. L., Waterston, R. H., 2006. AceTree: a tool
for visual analysis of Caenorhabditis elegans embryogenesis. BMC
Bioinformatics. 7, 275.
Brenner, S., 1974. The genetics of Caenorhabditis elegans. Genetics. 77, 71-94.
Broadie, K., Baumgartner, S., Prokop, A., 2011. Extracellular matrix and its receptors in
Drosophila neural development. Dev Neurobiol.
Brose, N., 2009. Synaptogenic proteins and synaptic organizers: "many hands make light
work". Neuron. 61, 650-2.
Bryant, D. M., Mostov, K. E., 2008. From cells to organs: building polarized tissue. Nat
Rev Mol Cell Biol. 9, 887-901.
Buechner, M., 2002. Tubes and the single C. elegans excretory cell. Trends Cell Biol. 12,
479-84.
Buechner, M., Hall, D. H., Bhatt, H., Hedgecock, E. M., 1999. Cystic canal mutants in
Caenorhabditis elegans are defective in the apical membrane domain of the renal
(excretory) cell. Dev Biol. 214, 227-41.
Bulow, H. E., Hobert, O., 2006. The molecular diversity of glycosaminoglycans shapes
animal development. Annu Rev Cell Dev Biol. 22, 375-407.
Cavaliere, V., Bernardi, F., Romani, P., Duchi, S., Gargiulo, G., 2008. Building up the
Drosophila eggshell: first of all the eggshell genes must be transcribed. Dev Dyn.
237, 2061-72.
Chalfie, M., Au, M., 1989. Genetic control of differentiation of the Caenorhabditis
elegans touch receptor neurons. Science. 243, 1027-33.
Chalfie, M., Thomson, J. N., Sulston, J. E., 1983. Induction of neuronal branching in
Caenorhabditis elegans. Science. 221, 61-3.
Chamcheu, J. C., Siddiqui, I. A., Syed, D. N., Adhami, V. M., Liovic, M., Mukhtar, H.,
2011. Keratin gene mutations in disorders of human skin and its appendages.
Arch Biochem Biophys. 508, 123-37.
Chi, X., Michos, O., Shakya, R., Riccio, P., Enomoto, H., Licht, J. D., Asai, N.,
Takahashi, M., Ohgami, N., Kato, M., Mendelsohn, C., Costantini, F., 2009. Ret-
dependent cell rearrangements in the Wolffian duct epithelium initiate ureteric
bud morphogenesis. Dev Cell. 17, 199-209.
207

Church, D. L., Guan, K. L., Lambie, E. J., 1995. Three genes of the MAP kinase cascade,
mek-2, mpk-1/sur-1 and let-60 ras, are required for meiotic cell cycle progression
in Caenorhabditis elegans. Development. 121, 2525-35.
Clark, D. V., Baillie, D. L., 1992. Genetic analysis and complementation by germ-line
transformation of lethal mutations in the unc-22 IV region of Caenorhabditis
elegans. Mol Gen Genet. 232, 97-105.
Costa, M., Draper, B. W., Priess, J. R., 1997. The role of actin filaments in patterning the
Caenorhabditis elegans cuticle. Dev Biol. 184, 373-84.
Costa, M., Raich, W., Agbunag, C., Leung, B., Hardin, J., Priess, J. R., 1998. A putative
catenin-cadherin system mediates morphogenesis of the Caenorhabditis elegans
embryo. J Cell Biol. 141, 297-308.
Cox, E. A., Hardin, J., 2004. Sticky worms: adhesion complexes in C. elegans. J Cell Sci.
117, 1885-97.
Cox, G. N., Laufer, J. S., Kusch, M., Edgar, R. S., 1980. Genetic and Phenotypic
Characterization of Roller Mutants of CAENORHABDITIS ELEGANS.
Genetics. 95, 317-39.
Davies, A. G., Spike, C. A., Shaw, J. E., Herman, R. K., 1999. Functional overlap
between the mec-8 gene and five sym genes in Caenorhabditis elegans. Genetics.
153, 117-34.
de Wit, J., Sylwestrak, E., O'Sullivan, M. L., Otto, S., Tiglio, K., Savas, J. N., Yates, J.
R., 3rd, Comoletti, D., Taylor, P., Ghosh, A., 2009. LRRTM2 interacts with
Neurexin1 and regulates excitatory synapse formation. Neuron. 64, 799-806.
Devine, W. P., Lubarsky, B., Shaw, K., Luschnig, S., Messina, L., Krasnow, M. A., 2005.
Requirement for chitin biosynthesis in epithelial tube morphogenesis. Proc Natl
Acad Sci U S A. 102, 17014-9.
DeVore, D. L., Horvitz, H. R., Stern, M. J., 1995. An FGF receptor signaling pathway is
required for the normal cell migrations of the sex myoblasts in C. elegans
hermaphrodites. Cell. 83, 611-20.
Ding, M., Woo, W. M., Chisholm, A. D., 2004. The cytoskeleton and epidermal
morphogenesis in C. elegans. Exp Cell Res. 301, 84-90.
Diogon, M., Wissler, F., Quintin, S., Nagamatsu, Y., Sookhareea, S., Landmann, F.,
Hutter, H., Vitale, N., Labouesse, M., 2007. The RhoGAP RGA-2 and LET-
502/ROCK achieve a balance of actomyosin-dependent forces in C. elegans
epidermis to control morphogenesis. Development. 134, 2469-79.
Dityatev, A., Seidenbecher, C. I., Schachner, M., 2010. Compartmentalization from the
outside: the extracellular matrix and functional microdomains in the brain. Trends
Neurosci. 33, 503-12.
Dolan, J., Walshe, K., Alsbury, S., Hokamp, K., O'Keeffe, S., Okafuji, T., Miller, S. F.,
Tear, G., Mitchell, K. J., 2007. The extracellular leucine-rich repeat superfamily;
a comparative survey and analysis of evolutionary relationships and expression
patterns. BMC Genomics. 8, 320.
Dressler, G. R., 2009. Advances in early kidney specification, development and
patterning. Development. 136, 3863-74.
Duerr, J. S., Frisby, D. L., Gaskin, J., Duke, A., Asermely, K., Huddleston, D., Eiden, L.
E., Rand, J. B., 1999. The cat-1 gene of Caenorhabditis elegans encodes a
208

vesicular monoamine transporter required for specific monoamine-dependent
behaviors. J Neurosci. 19, 72-84.
Fakhouri, M., Elalayli, M., Sherling, D., Hall, J. D., Miller, E., Sun, X., Wells, L.,
LeMosy, E. K., 2006. Minor proteins and enzymes of the Drosophila eggshell
matrix. Dev Biol. 293, 127-41.
Ferguson, E. L., Horvitz, H. R., 1985. Identification and characterization of 22 genes that
affect the vulval cell lineages of the nematode Caenorhabditis elegans. Genetics.
110, 17-72.
Fernandes, I., Chanut-Delalande, H., Ferrer, P., Latapie, Y., Waltzer, L., Affolter, M.,
Payre, F., Plaza, S., 2010. Zona pellucida domain proteins remodel the apical
compartment for localized cell shape changes. Dev Cell. 18, 64-76.
Finney, M., Ruvkun, G., 1990. The unc-86 gene product couples cell lineage and cell
identity in C. elegans. Cell. 63, 895-905.
Fire, A., Kondo, K., Waterston, R., 1990. Vectors for low copy transformation of C.
elegans. Nucleic Acids Res. 18, 4269-70.
Folkman, J., Haudenschild, C., 1980. Angiogenesis in vitro. Nature. 288, 551-6.
Francis, R., Waterston, R. H., 1991. Muscle cell attachment in Caenorhabditis elegans. J
Cell Biol. 114, 465-79.
Fritz, J. A., Behm, C. A., 2009. CUTI-1: A novel tetraspan protein involved in C. elegans
CUTicle formation and epithelial integrity. PLoS One. 4, e5117.
Gendler, S. J., Spicer, A. P., 1995. Epithelial mucin genes. Annu Rev Physiol. 57, 607-
34.
Gerhardt, H., 2008. VEGF and endothelial guidance in angiogenic sprouting.
Organogenesis. 4, 241-6.
Gettner, S. N., Kenyon, C., Reichardt, L. F., 1995. Characterization of beta pat-3
heterodimers, a family of essential integrin receptors in C. elegans. J Cell Biol.
129, 1127-41.
Ghabrial, A., Luschnig, S., Metzstein, M. M., Krasnow, M. A., 2003. Branching
morphogenesis of the Drosophila tracheal system. Annu Rev Cell Dev Biol. 19,
623-47.
Ghabrial, A. S., Krasnow, M. A., 2006. Social interactions among epithelial cells during
tracheal branching morphogenesis. Nature. 441, 746-9.
Giepmans, B. N., van Ijzendoorn, S. C., 2009. Epithelial cell-cell junctions and plasma
membrane domains. Biochim Biophys Acta. 1788, 820-31.
Gilleard, J. S., Barry, J. D., Johnstone, I. L., 1997. cis regulatory requirements for
hypodermal cell-specific expression of the Caenorhabditis elegans cuticle
collagen gene dpy-7. Mol Cell Biol. 17, 2301-11.
Glasheen, B. M., Robbins, R. M., Piette, C., Beitel, G. J., Page-McCaw, A., 2010. A
matrix metalloproteinase mediates airway remodeling in Drosophila. Dev Biol.
344, 772-83.
Gobel, V., Barrett, P. L., Hall, D. H., Fleming, J. T., 2004. Lumen morphogenesis in C.
elegans requires the membrane-cytoskeleton linker erm-1. Dev Cell. 6, 865-73.
Goldstein, B., Macara, I. G., 2007. The PAR proteins: fundamental players in animal cell
polarization. Dev Cell. 13, 609-22.
Goodyear, R. J., Richardson, G. P., 2002. Extracellular matrices associated with the
209

apical surfaces of sensory epithelia in the inner ear: molecular and structural
diversity. J Neurobiol. 53, 212-27.
Govindarajan, B., Gipson, I. K., 2010. Membrane-tethered mucins have multiple
functions on the ocular surface. Exp Eye Res. 90, 655-63.
Greenwald, I. S., Sternberg, P. W., Horvitz, H. R., 1983. The lin-12 locus specifies cell
fates in Caenorhabditis elegans. Cell. 34, 435-44.
Hedgecock, E. M., Culotti, J. G., Hall, D. H., 1990. The unc-5, unc-6, and unc-40 genes
guide circumferential migrations of pioneer axons and mesodermal cells on the
epidermis in C. elegans. Neuron. 4, 61-85.
Hedgecock, E. M., Sulston, J. E., Thomson, J. N., 1983. Mutations affecting programmed
cell deaths in the nematode Caenorhabditis elegans. Science. 220, 1277-9.
Herman, R. K., 1978. Crossover suppressors and balanced recessive lethals in
Caenorhabditis elegans. Genetics. 88, 49-65.
Hill, R. J., Sternberg, P. W., 1992. The gene lin-3 encodes an inductive signal for vulval
development in C. elegans. Nature. 358, 470-6.
Hodgkin, J., 1983. Male Phenotypes and Mating Efficiency in CAENORHABDITIS
ELEGANS. Genetics. 103, 43-64.
Hodgkin, J., Genetics. In: T. B. D.L. Riddle, B.J. Meyer, and J.R. Priess, (Ed.), C.
elegans II. Cold Spring Harbor Laboratory Press, Plainview, N.Y., 1997, pp. 881-
1047.
Hogan, B. L., Kolodziej, P. A., 2002. Organogenesis: molecular mechanisms of
tubulogenesis. Nat Rev Genet. 3, 513-23.
Hollingsworth, M. A., Swanson, B. J., 2004. Mucins in cancer: protection and control of
the cell surface. Nat Rev Cancer. 4, 45-60.
Horvitz, H. R., Sulston, J. E., 1980. Isolation and genetic characterization of cell-lineage
mutants of the nematode Caenorhabditis elegans. Genetics. 96, 435-54.
Howard, R. M., Sundaram, M. V., 2002. C. elegans EOR-1/PLZF and EOR-2 positively
regulate Ras and Wnt signaling and function redundantly with LIN-25 and the
SUR-2 Mediator component. Genes Dev. 16, 1815-27.
Howell, K., Arur, S., Schedl, T., Sundaram, M. V., 2010. EOR-2 is an obligate binding
partner of the BTB-zinc finger protein EOR-1 in Caenorhabditis elegans.
Genetics. 184, 899-913.
Hudson, M. J., Stamp, G. W., Chaudhary, K. S., Hewitt, R., Stubbs, A. P., Abel, P. D.,
Lalani, E. N., 2001. Human MUC1 mucin: a potent glandular morphogen. J
Pathol. 194, 373-83.
Hwang, B. J., Sternberg, P. W., 2004. A cell-specific enhancer that specifies lin-3
expression in the C. elegans anchor cell for vulval development. Development.
131, 143-51.
Hynes, R. O., 2009. The extracellular matrix: not just pretty fibrils. Science. 326, 1216-9.
Inglis, P. N., Boroevich, K. A., Leroux, M. R., 2006. Piecing together a ciliome. Trends
Genet. 22, 491-500.
Iruela-Arispe, M. L., Davis, G. E., 2009. Cellular and molecular mechanisms of vascular
lumen formation. Dev Cell. 16, 222-31.
Jacobs, D., Beitel, G. J., Clark, S. G., Horvitz, H. R., Kornfeld, K., 1998. Gain-of-
function mutations in the Caenorhabditis elegans lin-1 ETS gene identify a C-
210

terminal regulatory domain phosphorylated by ERK MAP kinase. Genetics. 149,
1809-22.
Jarriault, S., Schwab, Y., Greenwald, I., 2008. A Caenorhabditis elegans model for
epithelial-neuronal transdifferentiation. Proc Natl Acad Sci U S A. 105, 3790-5.
Jazwinska, A., Affolter, M., 2004. A family of genes encoding zona pellucida (ZP)
domain proteins is expressed in various epithelial tissues during Drosophila
embryogenesis. Gene Expr Patterns. 4, 413-21.
Jazwinska, A., Ribeiro, C., Affolter, M., 2003. Epithelial tube morphogenesis during
Drosophila tracheal development requires Piopio, a luminal ZP protein. Nat Cell
Biol. 5, 895-901.
Jiang, Y., Horner, V., Liu, J., 2005. The HMX homeodomain protein MLS-2 regulates
cleavage orientation, cell proliferation and cell fate specification in the C. elegans
postembryonic mesoderm. Development. 132, 4119-30.
Johnson, A. D., Fitzsimmons, D., Hagman, J., Chamberlin, H. M., 2001. EGL-38 Pax
regulates the ovo-related gene lin-48 during Caenorhabditis elegans organ
development. Development. 128, 2857-65.
Johnson, C. D., Duckett, J. G., Culotti, J. G., Herman, R. K., Meneely, P. M., Russell, R.
L., 1981. An acetylcholinesterase-deficient mutant of the nematode
Caenorhabditis elegans. Genetics. 97, 261-79.
Johnstone, I. L., 2000. Cuticle collagen genes. Expression in Caenorhabditis elegans.
Trends Genet. 16, 21-7.
Johnstone, I. L., Barry, J. D., 1996. Temporal reiteration of a precise gene expression
pattern during nematode development. EMBO J. 15, 3633-9.
Jones, S. J., Baillie, D. L., 1995. Characterization of the let-653 gene in Caenorhabditis
elegans. Mol Gen Genet. 248, 719-26.
Kalamajski, S., Oldberg, A., 2010. The role of small leucine-rich proteoglycans in
collagen fibrillogenesis. Matrix Biol. 29, 248-53.
Kalluri, R., Neilson, E. G., 2003. Epithelial-mesenchymal transition and its implications
for fibrosis. J Clin Invest. 112, 1776-84.
Kalluri, R., Weinberg, R. A., 2009. The basics of epithelial-mesenchymal transition. J
Clin Invest. 119, 1420-8.
Kao, G., Huang, C. C., Hedgecock, E. M., Hall, D. H., Wadsworth, W. G., 2006. The role
of the laminin beta subunit in laminin heterotrimer assembly and basement
membrane function and development in C. elegans. Dev Biol. 290, 211-9.
Kawai, T., Akira, S., 2010. The role of pattern-recognition receptors in innate immunity:
update on Toll-like receptors. Nat Immunol. 11, 373-84.
Knust, E., Bossinger, O., 2002. Composition and formation of intercellular junctions in
epithelial cells. Science. 298, 1955-9.
Ko, J., Fuccillo, M. V., Malenka, R. C., Sudhof, T. C., 2009. LRRTM2 functions as a
neurexin ligand in promoting excitatory synapse formation. Neuron. 64, 791-8.
Kobe, B., Deisenhofer, J., 1994. The leucine-rich repeat: a versatile binding motif. Trends
Biochem Sci. 19, 415-21.
Koh, K., Rothman, J. H., 2001. ELT-5 and ELT-6 are required continuously to regulate
epidermal seam cell differentiation and cell fusion in C. elegans. Development.
128, 2867-80.
211

Kohsaka, H., Nose, A., 2009. Target recognition at the tips of postsynaptic filopodia:
accumulation and function of Capricious. Development. 136, 1127-35.
Kondo, M., Kanemoto, N., Taniguchi, Y., Iwanaga, T., Arita, N., Nose, M., Tanigami,
A., 2000. Atypical hyperplasia of choledocho-pancreatic duct epithelium in an
Otsuka Long Evans Tokushima Fatty strain of rats. Pathol Int. 50, 126-35.
Koppen, M., Simske, J. S., Sims, P. A., Firestein, B. L., Hall, D. H., Radice, A. D.,
Rongo, C., Hardin, J. D., 2001. Cooperative regulation of AJM-1 controls
junctional integrity in Caenorhabditis elegans epithelia. Nat Cell Biol. 3, 983-91.
Kramer, J. M., Extracellular Matrix. In: T. B. D.L. Riddle, B.J. Meyer, and J.R. Priess,
(Ed.), C. elegans II. Cold Spring Harbor Laboratory Press, Plainview, N.Y., 1997,
pp. 471-500.
Krause, C., Wolf, C., Hemphala, J., Samakovlis, C., Schuh, R., 2006. Distinct functions
of the leucine-rich repeat transmembrane proteins capricious and tartan in the
Drosophila tracheal morphogenesis. Dev Biol. 296, 253-64.
Kufe, D. W., 2009. Mucins in cancer: function, prognosis and therapy. Nat Rev Cancer.
9, 874-85.
Kurusu, M., Cording, A., Taniguchi, M., Menon, K., Suzuki, E., Zinn, K., 2008. A screen
of cell-surface molecules identifies leucine-rich repeat proteins as key mediators
of synaptic target selection. Neuron. 59, 972-85.
Labouesse, M., 2006. Epithelial junctions and attachments. WormBook. 1-21.
Lambie, E. J., Kimble, J., 1991. Two homologous regulatory genes, lin-12 and glp-1,
have overlapping functions. Development. 112, 231-40.
Lee, M., Cram, E. J., Shen, B., Schwarzbauer, J. E., 2001. Roles for beta(pat-3) integrins
in development and function of Caenorhabditis elegans muscles and gonads. J
Biol Chem. 276, 36404-10.
Legouis, R., Gansmuller, A., Sookhareea, S., Bosher, J. M., Baillie, D. L., Labouesse, M.,
2000. LET-413 is a basolateral protein required for the assembly of adherens
junctions in Caenorhabditis elegans. Nat Cell Biol. 2, 415-22.
Leung, B., Hermann, G. J., Priess, J. R., 1999. Organogenesis of the Caenorhabditis
elegans intestine. Dev Biol. 216, 114-34.
Levi, B. P., Genetic control of tube formation and maintenance in Drosophila treacheal
terminal cells. Stanford University, 2006.
Levi, B. P., Ghabrial, A. S., Krasnow, M. A., 2006. Drosophila talin and integrin genes
are required for maintenance of tracheal terminal branches and luminal
organization. Development. 133, 2383-93.
Levitan, D., Greenwald, I., 1998. LIN-12 protein expression and localization during
vulval development in C. elegans. Development. 125, 3101-9.
Linhoff, M. W., Lauren, J., Cassidy, R. M., Dobie, F. A., Takahashi, H., Nygaard, H. B.,
Airaksinen, M. S., Strittmatter, S. M., Craig, A. M., 2009. An unbiased expression
screen for synaptogenic proteins identifies the LRRTM protein family as synaptic
organizers. Neuron. 61, 734-49.
Litscher, E. S., Williams, Z., Wassarman, P. M., 2009. Zona pellucida glycoprotein ZP3
and fertilization in mammals. Mol Reprod Dev. 76, 933-41.
Lubarsky, B., Krasnow, M. A., 2003. Tube morphogenesis: making and shaping
biological tubes. Cell. 112, 19-28.
212

Luschnig, S., Batz, T., Armbruster, K., Krasnow, M. A., 2006. serpentine and vermiform
encode matrix proteins with chitin binding and deacetylation domains that limit
tracheal tube length in Drosophila. Curr Biol. 16, 186-94.
Lynch, A. M., Hardin, J., 2009. The assembly and maintenance of epithelial junctions in
C. elegans. Front Biosci. 14, 1414-32.
MacQueen, A. J., Baggett, J. J., Perumov, N., Bauer, R. A., Januszewski, T., Schriefer,
L., Waddle, J. A., 2005. ACT-5 is an essential Caenorhabditis elegans actin
required for intestinal microvilli formation. Mol Biol Cell. 16, 3247-59.
Malhas, A. N., Abuknesha, R. A., Price, R. G., 2002. Interaction of the leucine-rich
repeats of polycystin-1 with extracellular matrix proteins: possible role in cell
proliferation. J Am Soc Nephrol. 13, 19-26.
Mani, S. A., Guo, W., Liao, M. J., Eaton, E. N., Ayyanan, A., Zhou, A. Y., Brooks, M.,
Reinhard, F., Zhang, C. C., Shipitsin, M., Campbell, L. L., Polyak, K., Brisken,
C., Yang, J., Weinberg, R. A., 2008. The epithelial-mesenchymal transition
generates cells with properties of stem cells. Cell. 133, 704-15.
Mao, Y., Kerr, M., Freeman, M., 2008. Modulation of Drosophila retinal epithelial
integrity by the adhesion proteins capricious and tartan. PLoS One. 3, e1827.
Marchiando, A. M., Graham, W. V., Turner, J. R., 2010. Epithelial barriers in
homeostasis and disease. Annu Rev Pathol. 5, 119-44.
Margaritis, L. H., Kafatos, F. C., Petri, W. H., 1980. The eggshell of Drosophila
melanogaster. I. Fine structure of the layers and regions of the wild-type eggshell.
J Cell Sci. 43, 1-35.
McCaffrey, L. M., Macara, I. G., 2011. Epithelial organization, cell polarity, and
tumorigenesis. Trends Cell Biol.
McKeown, C., Praitis, V., Austin, J., 1998. sma-1 encodes a betaH-spectrin homolog
required for Caenorhabditis elegans morphogenesis. Development. 125, 2087-98.
McMahon, L., Legouis, R., Vonesch, J. L., Labouesse, M., 2001. Assembly of C. elegans
apical junctions involves positioning and compaction by LET-413 and protein
aggregation by the MAGUK protein DLG-1. J Cell Sci. 114, 2265-77.
Meneely, P. M., Herman, R. K., 1979. Lethals, steriles and deficiencies in a region of the
X chromosome of Caenorhabditis elegans. Genetics. 92, 99-115.
Meneely, P. M., Herman, R. K., 1981. Suppression and function of X-linked lethal and
sterile mutations in Caenorhabditis elegans. Genetics. 97, 65-84.
Moerman, D. G., Barstead, R. J., 2008. Towards a mutation in every gene in
Caenorhabditis elegans. Brief Funct Genomic Proteomic. 7, 195-204.
Moncada, D. M., Kammanadiminti, S. J., Chadee, K., 2003. Mucin and Toll-like
receptors in host defense against intestinal parasites. Trends Parasitol. 19, 305-11.
Moribe, H., Yochem, J., Yamada, H., Tabuse, Y., Fujimoto, T., Mekada, E., 2004.
Tetraspanin protein (TSP-15) is required for epidermal integrity in Caenorhabditis
elegans. J Cell Sci. 117, 5209-20.
Moskowitz, I. P., Rothman, J. H., 1996. lin-12 and glp-1 are required zygotically for
early embryonic cellular interactions and are regulated by maternal GLP-1
signaling in Caenorhabditis elegans. Development. 122, 4105-17.
Motegi, F., Sugimoto, A., 2006. Sequential functioning of the ECT-2 RhoGEF, RHO-1
and CDC-42 establishes cell polarity in Caenorhabditis elegans embryos. Nat Cell
213

Biol. 8, 978-85.
Moussian, B., Tang, E., Tonning, A., Helms, S., Schwarz, H., Nusslein-Volhard, C., Uv,
A. E., 2006. Drosophila Knickkopf and Retroactive are needed for epithelial tube
growth and cuticle differentiation through their specific requirement for chitin
filament organization. Development. 133, 163-71.
Murray, J. I., Bao, Z., Boyle, T. J., Waterston, R. H., 2006. The lineaging of
fluorescently-labeled Caenorhabditis elegans embryos with StarryNite and
AceTree. Nat Protoc. 1, 1468-76.
Nelson, C. M., Bissell, M. J., 2006. Of extracellular matrix, scaffolds, and signaling:
tissue architecture regulates development, homeostasis, and cancer. Annu Rev
Cell Dev Biol. 22, 287-309.
Nelson, F. K., Albert, P. S., Riddle, D. L., 1983. Fine structure of the Caenorhabditis
elegans secretory-excretory system. J Ultrastruct Res. 82, 156-71.
Nelson, F. K., Riddle, D. L., 1984. Functional study of the Caenorhabditis elegans
secretory-excretory system using laser microsurgery. J Exp Zool. 231, 45-56.
Neves, A., Priess, J. R., 2005. The REF-1 family of bHLH transcription factors pattern C.
elegans embryos through Notch-dependent and Notch-independent pathways. Dev
Cell. 8, 867-79.
Novelli, J., Page, A. P., Hodgkin, J., 2006. The C terminus of collagen SQT-3 has
complex and essential functions in nematode collagen assembly. Genetics. 172,
2253-67.
Oka, T., Yamamoto, R., Futai, M., 1997. Three vha genes encode proteolipids of
Caenorhabditis elegans vacuolar-type ATPase. Gene structures and preferential
expression in an H-shaped excretory cell and rectal cells. J Biol Chem. 272,
24387-92.
Page, A. P., Johnstone, I. L., 2007. The cuticle. WormBook. 1-15.
Partridge, F. A., Tearle, A. W., Gravato-Nobre, M. J., Schafer, W. R., Hodgkin, J., 2008.
The C. elegans glycosyltransferase BUS-8 has two distinct and essential roles in
epidermal morphogenesis. Dev Biol. 317, 549-59.
Payre, F., 2004. Genetic control of epidermis differentiation in Drosophila. Int J Dev
Biol. 48, 207-15.
Perens, E. A., Shaham, S., 2005. C. elegans daf-6 encodes a patched-related protein
required for lumen formation. Dev Cell. 8, 893-906.
Pettitt, J., Cox, E. A., Broadbent, I. D., Flett, A., Hardin, J., 2003. The Caenorhabditis
elegans p120 catenin homologue, JAC-1, modulates cadherin-catenin function
during epidermal morphogenesis. J Cell Biol. 162, 15-22.
Piekny, A. J., Johnson, J. L., Cham, G. D., Mains, P. E., 2003. The Caenorhabditis
elegans nonmuscle myosin genes nmy-1 and nmy-2 function as redundant
components of the let-502/Rho-binding kinase and mel-11/myosin phosphatase
pathway during embryonic morphogenesis. Development. 130, 5695-704.
Piekny, A. J., Wissmann, A., Mains, P. E., 2000. Embryonic morphogenesis in
Caenorhabditis elegans integrates the activity of LET-502 Rho-binding kinase,
MEL-11 myosin phosphatase, DAF-2 insulin receptor and FEM-2 PP2c
phosphatase. Genetics. 156, 1671-89.
Plaza, S., Chanut-Delalande, H., Fernandes, I., Wassarman, P. M., Payre, F., 2010. From
214

A to Z: apical structures and zona pellucida-domain proteins. Trends Cell Biol.
20, 524-32.
Pohl, C., Bao, Z., 2010. Chiral forces organize left-right patterning in C. elegans by
uncoupling midline and anteroposterior axis. Dev Cell. 19, 402-12.
Polyak, K., Weinberg, R. A., 2009. Transitions between epithelial and mesenchymal
states: acquisition of malignant and stem cell traits. Nat Rev Cancer. 9, 265-73.
Praitis, V., Ciccone, E., Austin, J., 2005. SMA-1 spectrin has essential roles in epithelial
cell sheet morphogenesis in C. elegans. Dev Biol. 283, 157-70.
Pries, A. R., Secomb, T. W., Gaehtgens, P., 2000. The endothelial surface layer. Pflugers
Arch. 440, 653-66.
Priess, J. R., Hirsh, D. I., 1986. Caenorhabditis elegans morphogenesis: the role of the
cytoskeleton in elongation of the embryo. Dev Biol. 117, 156-73.
Rappleye, C. A., Paredez, A. R., Smith, C. W., McDonald, K. L., Aroian, R. V., 1999.
The coronin-like protein POD-1 is required for anterior-posterior axis formation
and cellular architecture in the nematode caenorhabditis elegans. Genes Dev. 13,
2838-51.
Red-Horse, K., Ueno, H., Weissman, I. L., Krasnow, M. A., 2010. Coronary arteries form
by developmental reprogramming of venous cells. Nature. 464, 549-53.
Riddle, D. L., Blumenthal, T., Meyer, B. J., Priess, J. R., 1997. Introduction to C.
elegans.
Roberts, B., Clucas, C., Johnstone, I. L., 2003. Loss of SEC-23 in Caenorhabditis elegans
causes defects in oogenesis, morphogenesis, and extracellular matrix secretion.
Mol Biol Cell. 14, 4414-26.
Rocheleau, C. E., Howard, R. M., Goldman, A. P., Volk, M. L., Girard, L. J., Sundaram,
M. V., 2002. A lin-45 raf enhancer screen identifies eor-1, eor-2 and unusual
alleles of Ras pathway genes in Caenorhabditis elegans. Genetics. 161, 121-31.
Ross, M. D., Bruggeman, L. A., Hanss, B., Sunamoto, M., Marras, D., Klotman, M. E.,
Klotman, P. E., 2003. Podocan, a novel small leucine-rich repeat protein
expressed in the sclerotic glomerular lesion of experimental HIV-associated
nephropathy. J Biol Chem. 278, 33248-55.
Roubin, R., Naert, K., Popovici, C., Vatcher, G., Coulier, F., Thierry-Mieg, J., Pontarotti,
P., Birnbaum, D., Baillie, D., Thierry-Mieg, D., 1999. let-756, a C. elegans fgf
essential for worm development. Oncogene. 18, 6741-7.
Roy, L. D., Sahraei, M., Subramani, D. B., Besmer, D., Nath, S., Tinder, T. L., Bajaj, E.,
Shanmugam, K., Lee, Y. Y., Hwang, S. I., Gendler, S. J., Mukherjee, P., 2011.
MUC1 enhances invasiveness of pancreatic cancer cells by inducing epithelial to
mesenchymal transition. Oncogene. 30, 1449-59.
Santella, A., Du, Z., Nowotschin, S., Hadjantonakis, A. K., Bao, Z., 2010. A hybrid blob-
slice model for accurate and efficient detection of fluorescence labeled nuclei in
3D. BMC Bioinformatics. 11, 580.
Sapio, M. R., Hilliard, M. A., Cermola, M., Favre, R., Bazzicalupo, P., 2005. The Zona
Pellucida domain containing proteins, CUT-1, CUT-3 and CUT-5, play essential
roles in the development of the larval alae in Caenorhabditis elegans. Dev Biol.
282, 231-45.
Sapir, A., Choi, J., Leikina, E., Avinoam, O., Valansi, C., Chernomordik, L. V.,
215

Newman, A. P., Podbilewicz, B., 2007. AFF-1, a FOS-1-regulated fusogen,
mediates fusion of the anchor cell in C. elegans. Dev Cell. 12, 683-98.
Schaefer, L., 2011. Small leucine-rich proteoglycans in kidney disease. J Am Soc
Nephrol. 22, 1200-7.
Schaefer, L., Iozzo, R. V., 2008. Biological functions of the small leucine-rich
proteoglycans: from genetics to signal transduction. J Biol Chem. 283, 21305-9.
Schafer, W. F., 2006. Genetics of egg-laying in worms. Annu Rev Genet. 40, 487-509.
Schonegg, S., Hyman, A. A., 2006. CDC-42 and RHO-1 coordinate acto-myosin
contractility and PAR protein localization during polarity establishment in C.
elegans embryos. Development. 133, 3507-16.
Schottenfeld, J., Song, Y., Ghabrial, A. S., 2010. Tube continued: morphogenesis of the
Drosophila tracheal system. Curr Opin Cell Biol. 22, 633-9.
Segbert, C., Johnson, K., Theres, C., van Furden, D., Bossinger, O., 2004. Molecular and
functional analysis of apical junction formation in the gut epithelium of
Caenorhabditis elegans. Dev Biol. 266, 17-26.
Shakya, R., Watanabe, T., Costantini, F., 2005. The role of GDNF/Ret signaling in
ureteric bud cell fate and branching morphogenesis. Dev Cell. 8, 65-74.
Shimizu-Hirota, R., Sasamura, H., Kuroda, M., Kobayashi, E., Saruta, T., 2004.
Functional characterization of podocan, a member of a new class in the small
leucine-rich repeat protein family. FEBS Lett. 563, 69-74.
Shin, K., Fogg, V. C., Margolis, B., 2006. Tight junctions and cell polarity. Annu Rev
Cell Dev Biol. 22, 207-35.
Shinza-Kameda, M., Takasu, E., Sakurai, K., Hayashi, S., Nose, A., 2006. Regulation of
layer-specific targeting by reciprocal expression of a cell adhesion molecule,
capricious. Neuron. 49, 205-13.
Shishido, E., Takeichi, M., Nose, A., 1998. Drosophila synapse formation: regulation by
transmembrane protein with Leu-rich repeats, CAPRICIOUS. Science. 280, 2118-
21.
Siddiqui, T. J., Pancaroglu, R., Kang, Y., Rooyakkers, A., Craig, A. M., 2010. LRRTMs
and neuroligins bind neurexins with a differential code to cooperate in glutamate
synapse development. J Neurosci. 30, 7495-506.
Simske, J. S., Hardin, J., 2011. Claudin family proteins in Caenorhabditis elegans.
Methods Mol Biol. 762, 147-69.
Simske, J. S., Kaech, S. M., Harp, S. A., Kim, S. K., 1996. LET-23 receptor localization
by the cell junction protein LIN-7 during C. elegans vulval induction. Cell. 85,
195-204.
Singh, N., Han, M., 1995. sur-2, a novel gene, functions late in the let-60 ras-mediated
signaling pathway during Caenorhabditis elegans vulval induction. Genes Dev. 9,
2251-65.
Sonnichsen, B., Koski, L. B., Walsh, A., Marschall, P., Neumann, B., Brehm, M.,
Alleaume, A. M., Artelt, J., Bettencourt, P., Cassin, E., Hewitson, M., Holz, C.,
Khan, M., Lazik, S., Martin, C., Nitzsche, B., Ruer, M., Stamford, J., Winzi, M.,
Heinkel, R., Roder, M., Finell, J., Hantsch, H., Jones, S. J., Jones, M., Piano, F.,
Gunsalus, K. C., Oegema, K., Gonczy, P., Coulson, A., Hyman, A. A., Echeverri,
C. J., 2005. Full-genome RNAi profiling of early embryogenesis in
216

Caenorhabditis elegans. Nature. 434, 462-9.
St Johnston, D., Sanson, B., 2011. Epithelial polarity and morphogenesis. Curr Opin Cell
Biol.
Sternberg, P. W., 2005. Vulval development. WormBook. 1-28.
Stone, C. E., Hall, D. H., Sundaram, M. V., 2009. Lipocalin signaling controls unicellular
tube development in the Caenorhabditis elegans excretory system. Dev Biol. 329,
201-11.
Struhl, G., Fitzgerald, K., Greenwald, I., 1993. Intrinsic activity of the Lin-12 and Notch
intracellular domains in vivo. Cell. 74, 331-45.
Sulston, J. E., Horvitz, H. R., 1977. Post-embryonic cell lineages of the nematode,
Caenorhabditis elegans. Dev Biol. 56, 110-56.
Sulston, J. E., Schierenberg, E., White, J. G., Thomson, J. N., 1983. The embryonic cell
lineage of the nematode Caenorhabditis elegans. Dev Biol. 100, 64-119.
Sundaram, M. V., 2005. The love-hate relationship between Ras and Notch. Genes Dev.
19, 1825-39.
Sundaram, M. V., 2006. RTK/Ras/MAPK signaling. WormBook. 1-19.
Swanson, L. E., Yu, M., Nelson, K. S., Laprise, P., Tepass, U., Beitel, G. J., 2009.
Drosophila convoluted/dALS is an essential gene required for tracheal tube
morphogenesis and apical matrix organization. Genetics. 181, 1281-90.
Tabuse, Y., Izumi, Y., Piano, F., Kemphues, K. J., Miwa, J., Ohno, S., 1998. Atypical
protein kinase C cooperates with PAR-3 to establish embryonic polarity in
Caenorhabditis elegans. Development. 125, 3607-14.
Taniguchi, H., Shishido, E., Takeichi, M., Nose, A., 2000. Functional dissection of
drosophila capricious: its novel roles in neuronal pathfinding and selective
synapse formation. J Neurobiol. 42, 104-16.
Tervonen, T. A., Partanen, J. I., Saarikoski, S. T., Myllynen, M., Marques, E., Paasonen,
K., Moilanen, A., Wohlfahrt, G., Kovanen, P. E., Klefstrom, J., 2011. Faulty
epithelial polarity genes and cancer. Adv Cancer Res. 111, 97-161.
Thornton, D. J., Rousseau, K., McGuckin, M. A., 2008. Structure and function of the
polymeric mucins in airways mucus. Annu Rev Physiol. 70, 459-86.
Tiensuu, T., Larsen, M. K., Vernersson, E., Tuck, S., 2005. lin-1 has both positive and
negative functions in specifying multiple cell fates induced by Ras/MAP kinase
signaling in C. elegans. Dev Biol. 286, 338-51.
Tonning, A., Hemphala, J., Tang, E., Nannmark, U., Samakovlis, C., Uv, A., 2005. A
transient luminal chitinous matrix is required to model epithelial tube diameter in
the Drosophila trachea. Dev Cell. 9, 423-30.
Tootle, T. L., Williams, D., Hubb, A., Frederick, R., Spradling, A., 2011. Drosophila
eggshell production: identification of new genes and coordination by Pxt. PLoS
One. 6, e19943.
Totong, R., Achilleos, A., Nance, J., 2007. PAR-6 is required for junction formation but
not apicobasal polarization in C. elegans embryonic epithelial cells. Development.
134, 1259-68.
Trougakos, I. P., Margaritis, L. H., 1998. Immunolocalization of the temporally "early"
secreted major structural chorion proteins, Dvs38 and Dvs36, in the eggshell
layers and regions of Drosophila virilis. J Struct Biol. 123, 111-23.
217

Tsarouhas, V., Senti, K. A., Jayaram, S. A., Tiklova, K., Hemphala, J., Adler, J.,
Samakovlis, C., 2007. Sequential pulses of apical epithelial secretion and
endocytosis drive airway maturation in Drosophila. Dev Cell. 13, 214-25.
Tuck, S., Greenwald, I., 1995. lin-25, a gene required for vulval induction in
Caenorhabditis elegans. Genes Dev. 9, 341-57.
Uv, A., Moussian, B., 2010. The apical plasma membrane of Drosophila embryonic
epithelia. Eur J Cell Biol. 89, 208-11.
Van Furden, D., Johnson, K., Segbert, C., Bossinger, O., 2004. The C. elegans ezrin-
radixin-moesin protein ERM-1 is necessary for apical junction remodelling and
tubulogenesis in the intestine. Dev Biol. 272, 262-76.
Voas, M. G., Rebay, I., 2004. Signal integration during development: insights from the
Drosophila eye. Dev Dyn. 229, 162-75.
Wang, S., Jayaram, S. A., Hemphala, J., Senti, K. A., Tsarouhas, V., Jin, H., Samakovlis,
C., 2006. Septate-junction-dependent luminal deposition of chitin deacetylases
restricts tube elongation in the Drosophila trachea. Curr Biol. 16, 180-5.
Wassarman, P. M., Litscher, E. S., 2009. The multifunctional zona pellucida and
mammalian fertilization. J Reprod Immunol. 83, 45-9.
Weaver, M., Krasnow, M. A., 2008. Dual origin of tissue-specific progenitor cells in
Drosophila tracheal remodeling. Science. 321, 1496-9.
Weimer, R. M., 2006. Preservation of C. elegans tissue via high-pressure freezing and
freeze-substitution for ultrastructural analysis and immunocytochemistry.
Methods Mol Biol. 351, 203-21.
Williams, B. D., Waterston, R. H., 1994. Genes critical for muscle development and
function in Caenorhabditis elegans identified through lethal mutations. J Cell
Biol. 124, 475-90.
Wilson, P. D., 2011. Apico-basal polarity in polycystic kidney disease epithelia. Biochim
Biophys Acta. 1812, 1239-48.
Wissmann, A., Ingles, J., Mains, P. E., 1999. The Caenorhabditis elegans mel-11 myosin
phosphatase regulatory subunit affects tissue contraction in the somatic gonad and
the embryonic epidermis and genetically interacts with the Rac signaling
pathway. Dev Biol. 209, 111-27.
Wissmann, A., Ingles, J., McGhee, J. D., Mains, P. E., 1997. Caenorhabditis elegans
LET-502 is related to Rho-binding kinases and human myotonic dystrophy kinase
and interacts genetically with a homolog of the regulatory subunit of smooth
muscle myosin phosphatase to affect cell shape. Genes Dev. 11, 409-22.
Woo, W. M., Goncharov, A., Jin, Y., Chisholm, A. D., 2004. Intermediate filaments are
required for C. elegans epidermal elongation. Dev Biol. 267, 216-29.
Yochem, J., Sundaram, M., Han, M., 1997. Ras is required for a limited number of cell
fates and not for general proliferation in Caenorhabditis elegans. Mol Cell Biol.
17, 2716-22.
Yoshimura, S., Murray, J. I., Lu, Y., Waterston, R. H., Shaham, S., 2008. mls-2 and vab-
3 Control glia development, hlh-17/Olig expression and glia-dependent neurite
extension in C. elegans. Development. 135, 2263-75.
Young, M. F., Bi, Y., Ameye, L., Chen, X. D., 2002. Biglycan knockout mice: new
models for musculoskeletal diseases. Glycoconj J. 19, 257-62.
218

Zeisberg, E. M., Tarnavski, O., Zeisberg, M., Dorfman, A. L., McMullen, J. R.,
Gustafsson, E., Chandraker, A., Yuan, X., Pu, W. T., Roberts, A. B., Neilson, E.
G., Sayegh, M. H., Izumo, S., Kalluri, R., 2007. Endothelial-to-mesenchymal
transition contributes to cardiac fibrosis. Nat Med. 13, 952-61.
Zhang, Y., Foster, J. M., Nelson, L. S., Ma, D., Carlow, C. K., 2005. The chitin synthase
genes chs-1 and chs-2 are essential for C. elegans development and responsible
for chitin deposition in the eggshell and pharynx, respectively. Dev Biol. 285,
330-9.
219
Copyright © 2022 FDOKUMEN




















