
Laser Interstitial Thermal Therapy in Grade 2/3 IDH1/2 Mutant ...
Upload
independentCategory
view
1download
0
Fax +41 61 306 12 34E-Mail [email protected]
Original Paper
Cells Tissues Organs 2012;196:175–188 DOI: 10.1159/000332381
Late Embryonic and Postnatal Development of Interstitial Cells of Cajal in Mouse Esophagus: Distribution, Proliferation and Kit Dependence
Xiao He a, b Wen-cui Yang a, b Xiang-yin Wen a, b Di Tang a, b Lan Xiao a
Juan Han c Bin Yu a Wei Zhang d Feng Mei a
Departments of a Histology and Embryology, and Development Biology, and b 9th Team of the Cader Brigade, Third Military Medical University, c Department of Emergency, People’s Liberation Army Hospital 324, and d Department of Cardiothoracic Surgery, First Affiliated Hospital, Third Military Medical University, Chongqing , China
birth through adulthood (P28) with a gradually decreasing cell number. At 24 h, Kit+ ICC were dramatically decreased and almost missing 48 h after administration of imatinib (a Kit inhibitor). Our results indicate that ICC proliferation is age dependent and persists throughout the postnatal period. There is a dramatic decrease in the ICC number from P0 to adult life. The Kit signal is essential for the postnatal develop-ment of ICC in the esophagus.
Copyright © 2012 S. Karger AG, Basel
Key Words
Bromodeoxyuridine � Esophageal model � Interstitial cells of Cajal � Ki67 � Kit � Postnatal development � Proliferation
Abstract
This paper investigates alterations in interstitial cells of Cajal (ICC) in the esophagus of mice from embryonic day 13.5 (E13.5) to 36 days postpartum (P0–P36) using immunohisto-chemistry. At E13.5, Kit+ cells presented in clusters and dif-ferentiated into spindle-like cells with biopolar processes within the outer (longitudinal) and inner (circular) muscle layers at E17.5. These Kit+ ICC with long processes were also Ano1+ and prominent at birth. The density of ICC gradually decreased, and at P36 it became about one twentieth of that at birth. Kit ligand (stem cell factor) expression is lower in stri-ated muscle cells than that in smooth muscle cells. The ICC number was higher in the distal (close to the cardia) than in the proximal esophagus (close to the pharynx). Some Kit+/Ki67+ and Kit+/bromodeoxyuridine+ cells were observed within the muscle layers, and proliferation persisted from
Accepted after revision: August 29, 2011 Published online: January 20, 2012
Dr. Feng Mei Departments of Histology and Embryology, and Development Biology Third Military Medical UniversityChongqing 400038 (China) Tel. +86 23 6877 5221, E-Mail meifeng.mail @ 163.com
Co-corresponding author: Dr. Wei Zhang Department of Cardiothoracic SurgeryThe First Affiliated Hospital of the Third Military Medical UniversityChongqing 400038 (China)Tel. +86 23 6877 3084, E-Mail zhangw1966 @ 126.com
© 2012 S. Karger AG, Basel1422–6405/12/1962–0175$38.00/0
Accessible online at:www.karger.com/cto
Abbreviations used in this paper
BrdU bromodeoxyuridineGI gastrointestinalICC interstitial cells of CajalICC-IM intramuscular ICCSCF stem cell factorSMC smooth muscle cells
X.H. and W.Y. contributed equally to this work.

He /Yang /Wen /Tang /Xiao /Han /Yu /Zhang /Mei
Cells Tissues Organs 2012;196:175–188176
Introduction
Interstitial cells of Cajal (ICC) are mesenchymal-de-rived cells distributed within the entire gastrointestinal (GI) tract wall [Lecoin et al., 1996; Young et al., 1996; Kluppel et al., 1998; Young, 1999; Wu et al., 2000]. ICC act as pacemaker cells in the small intestine, stomach and colon [Ward, et al., 1994; Liu and Huizinga, 1994; Hui-zinga et al., 1995; Rumessen and Thuneberg, 1996; Sand-ers, 1996; Horiguchi et al., 2001; Ward and Sanders, 2001]. ICC are also closely connected to nerve and muscle cells in the esophagus [Rumessen et al., 2001]. These cells form synapse-like structures with nerve fibers, and also gap junctions to smooth muscle cells (SMC) in opossums, dogs and humans [Daniel and Posey-Daniel, 1984; Wong et al., 1990; Berezin et al., 1994]. It has been suggested that ICC in the esophagus probably function as mediators of neural inputs [Ward et al., 1998] and mechanosensors [Rumessen et al., 2001; Fox et al., 2002].
ICC and SMC of the GI tract are derived from com-mon mesenchymal precursors [Lecoin et al., 1996; Young et al., 1996; Kluppel et al., 1998; Young, 1999]. SMC pro-vide an adaptive microenvironment for the development of ICC [Horvath et al., 2006; Lin et al., 2010]. Distinct from other segments of the GI tract, the microenviron-ment in the esophageal wall undergoes a dramatic change during late embryonic and postnatal life [Stratton et al., 2000]. Smooth muscle precursors are predominant in the muscle layers of the murine esophagus during the embry-onic period [Patapoutian et al., 1995; Sang and Young, 1997], and striated muscle cells gradually appear and re-place SMC before adulthood. Previous studies have indi-cated that ICC increased gradually within the small in-testine and colon from the neonatal stage to adulthood, and the proliferation of ICC is involved in the postnatal development of gut [Mei et al., 2009c; Han et al., 2010a; Tharayil et al., 2010]. With respect to the dramatic chang-es observed in muscle cells, the postnatal development of ICC in the esophagus may differ from other segments of the GI tract, and is still not well understood yet.
ICC progenitors begin to express Kit protein, a mark-er for ICC, during the embryonic period in the foregut and hindgut of mice on embryonic days 12 (E12) and E14, respectively [Wu et al., 2000]. Kit ligand, stem cell factor (SCF), is expressed by SMC and neurons. Kit signaling is essential for the genesis and development of ICC in the stomach, small intestine and colon [Torihashi et al., 1999b; Shimojima et al., 2005; Beckett et al., 2007; Han et al., 2010b]. Inactivation of Kit with neutralizing antibod-ies or Kit inhibitor in fetal and neonatal animals, or non-
lethal mutations of Kit or SCF in rats and mice, leads to abnormal ICC development in the GI tract [Ward et al., 1995; Mikkelsen et al., 1998; Nakama et al., 1998; Naka-gawa et al., 2005; Sanders and Ward, 2007]. However, the effects of Kit signaling on ICC in the esophagus during postnatal development are still unclear.
To address these issues, we have utilized immunofluo-rescent staining to investigate the alterations in the den-sity, morphology and proliferation of ICC in the murine esophagus from late embryonic (E13.5) to adult life [post-natal day 36 (P36)], and the response of ICC to Kit signal-ing blockade using imatinib, a known Kit inhibitor.
Materials and Methods
Animals BALB/C mice were purchased from the Animal Center of the
Third Military Medical University (Chongqing, China) and paired to produce offspring; 86 mice were used in our experi-ments. Animals were divided into 3 groups: (1) 45 mice were killed at E13.5, E17.5, P0, P2, P7, P12, P18, P28 and P36 and used for whole-mount preparations, and mice aged from P2 to P36 were injected with 1 dose of bromodeoxyuridine (BrdU; 0.1 mg/g i.p.) 24 h before being sacrificed; (2) 21 mice were sacrificed at P0, P2, P7, P12, P18, P28 and P150 and used for sectioning; (3) 10 mice at P8 were treated intragastrically with imatinib mesylate (Glivec � ; Novartis) at a dosage of 0.5 mg/g per day, and sacrificed 24 (n = 5) and 48 h (n = 5) later, and distilled water was used to treat 10 mice as the control group. All experiments were performed in accor-dance with our University Health Guide for the Care and Use of Laboratory Animals.
Immunofluorescence To obtain frozen sections, the esophagus was placed in the op-
timal cutting temperature compound and quickly frozen in liquid nitrogen. Transection sections (6–8 � m thick) were made with a cryostat (Leica 1850) and fixed with 100% acetone for 15 min (at 4 ° C). For whole-mount preparations, the esophagus was fixed with 100% cold acetone and divided into two parts (proximal and distal segments) with identical lengths, then opened along the lu-men, and the mucosa and submucosa were removed by sharp dis-section using a dissection microscope (except for E13.5 mice). Immunostaining procedures were as described herein. Whole-mount preparations or frozen sections were incubated with rat anti-Kit (Ack2, 1: 100; eBioscience), rabbit anti-Ki67 (1: 100; Neo-marker), rabbit anti-SCF (Santa Cruz, 1: 200), rabbit anti-Ano1 (Abcam, 1: 300) or mouse anti-desmin (1: 100; Santa Cruz) anti-body overnight at 4 ° C. Secondary antibodies were utilized as fol-lows: Cy3-conjugated anti-rat IgG (1: 100; Zymed), FITC-conju-gated anti-mouse IgG (1: 100; DAKO) and Alexa488-conjugated anti-rabbit IgG (1: 100; Invitrogen). To identify BrdU incorpora-tion, whole-mount preparations were processed as follows: the specimens were additionally fixed in 4% paraformaldehyde in 0.1 M PBS at pH 7.2 for 10 min. After rinsing with 0.01 M PBS (pH 7.2), the specimens were treated with 2 M HCl for 30 min at 37 ° C. To reveal BrdU labeling, the specimens were incubated with a mouse

ICC Development in Mouse Esophagus Cells Tissues Organs 2012;196:175–188 177
monoclonal antibody raised against BrdU (1: 100; DAKO) over-night at 4 ° C. Negative controls were performed by omission of primary antibodies. The stained results were detected using a Nikon 90i fluorescence microscope (Nikon) or Olympus confocal laser scanning microscope (FV1000; Olympus) with an excitation wavelength appropriate for FITC (488 nm) and Cy3 (552 nm).
Measurement and Statistical Analysis After immunofluorescent staining, photomicrographs of Kit+
cells were taken in 9 random fields (magnification ! 200, 0.3969 mm 2 ) per whole-mount preparation with a laser confocal scan-ning microscope (PV100; Olympus), and both muscle layers were scanned, stepping at 2 � m in order to capture all of the ICC con-tained within the muscle layers. The number of Kit+ cells was counted with Image-Pro Plus 5.0 (Media Cybernetics). Data are expressed as means 8 SEM. Numbers (n) reported in the text re-fer to the number of animals used. Differences in the data were evaluated by Student’s t test, and p ! 0.05 was considered statisti-cally significant.
Results
Alterations in ICC Distribution To characterize the ICC distribution in the esophageal
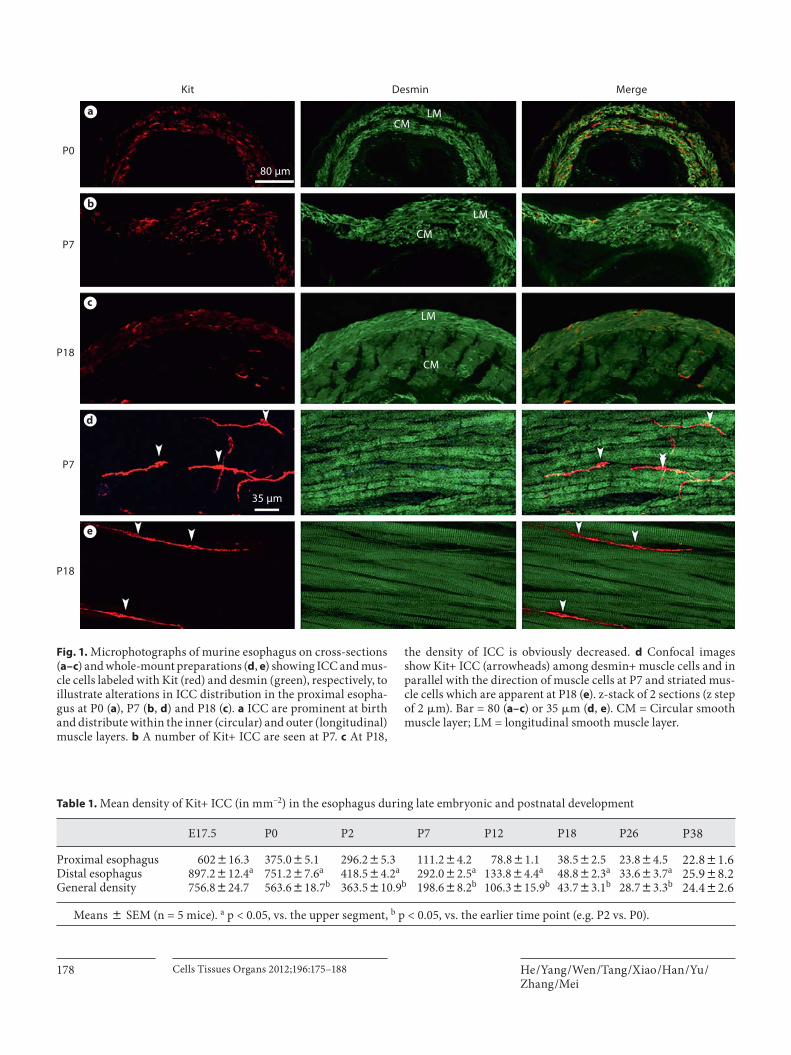
wall from birth (P0) to adulthood (P36), Kit (ICC marker, red) and desmin (a marker for both types of muscle cells, green) double labeling was performed. These Kit+ ICC were distributed within the outer (longitudinal) and in-ner (circular) muscle layers (desmin+) on frozen cross-sections, and within muscle layers (ICC-IM) they were prominent at P0 in the esophagus ( fig. 1 a). The density of Kit+ cells appeared unchanged during the thickening of smooth muscle layers by P2 and P7 ( fig. 1 b), but cell den-sity subsequently appeared much lower at P18 ( fig. 1 c) and P28. These Kit+ ICC were located between muscle cells and in parallel with the orientation of muscle cells on whole-mount preparations, as revealed by confocal scan-ning, and closely in contact with adjacent muscle cells by extended processes at P2 ( fig. 1 d) and P18 ( fig. 1 e); stri-ated muscle was apparent at P18 ( fig. 1 e).
ICC Changes in Morphology and Density To investigate the changes in the morphology and den-
sity of ICC during embryonic (E13.5 and E17.5) and post-natal periods (P0–P36), Kit immunofluorescent staining was carried out on whole-mount esophageal preparations. In the esophagus, Kit+ cells were present in clusters on E13.5 with round cell bodies and slender processes (E14.5; fig. 2 a, a’). Kit+ ICC with oval-shaped cell bodies and 2 or 3 processes appeared by E17.5. These cells were in parallel with muscle fibers and formed a mesh-like network con-necting to other ICC ( fig. 2 b). The density of ICC was
about 750 per mm –2 . Kit+ ICC had prolonged processes and a tightly organized network at birth (P0; fig. 2 c), and density was decreased (to about 563 mm –2 ) compared to that at E17.5 ( table 1 ), with longer processes but a lower density on P2 ( fig. 2 d). The density of Kit+ ICC was obvi-ously decreased by P12 (to about 106 mm –2 ; fig. 2 e), and the value progressively decreased from P18, P28 to P36 ( fig. 2 f), with a decreased density at P36 (about one twen-tieth of that observed at birth; table 1 ). Ano1, another mar-ker of ICC, was used to identify ICC in the esophagus be-sides Kit, and Kit/Ano1 double labeling indicated that al-most all Kit+ cells were also DOG1 positive ( fig. 2 g–i). These results indicate that Kit+ cells are ICC in the esoph-agus and similar to those in the stomach and intestine.
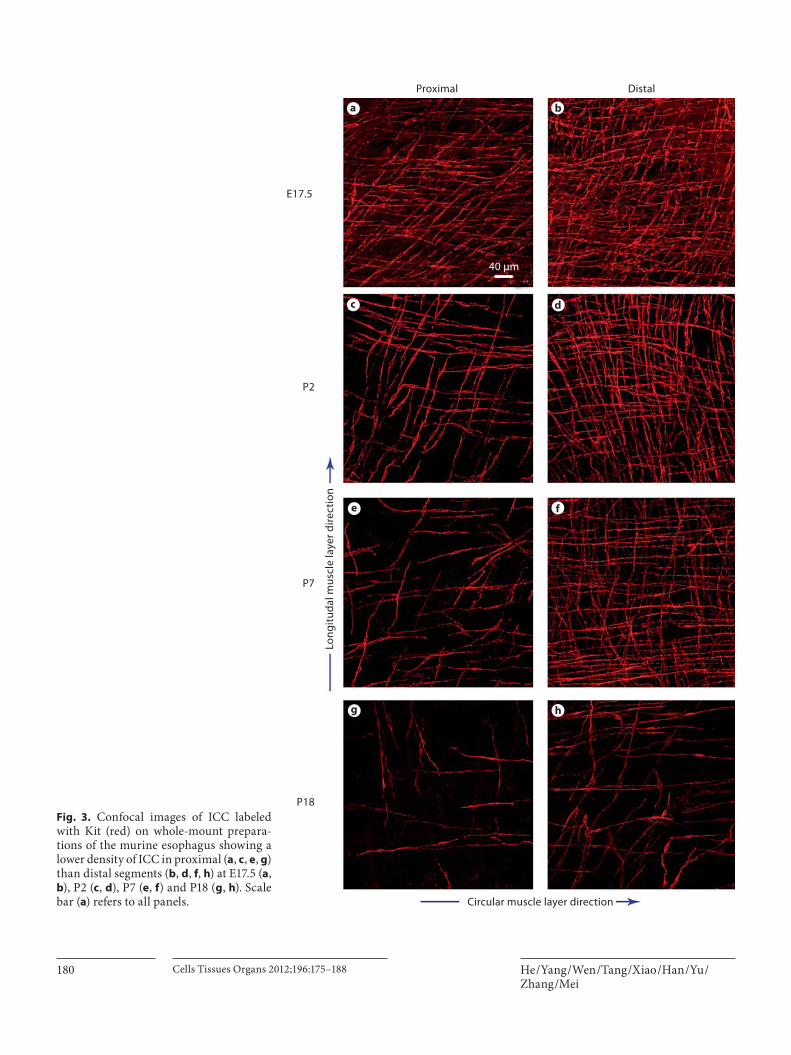
We compared the distribution of ICC in the proximal (close to the pharynx) and distal (close to the cardia) seg-ments of the esophagus, where a distribution difference was observed between these two segments. At E17.5, more Kit+ ICC were present in the proximal esophagus ( fig. 3 a) than in the distal portion ( fig. 3 b). This distribution dif-ference was more apparent at P2 ( fig. 3 c, d), and counting indicated that the density in the distal segment was about 751 mm –2 ( table 1 ). The value was significantly higher than that for the proximal portion, which was about 375 mm –2 ( table 1 ). The tendency was still present at P7 ( fig. 3 e, f) and P18 ( fig. 3 g, h), and persisted throughout the postnatal development of the esophagus ( table 1 ).
Proliferation of ICC With respect to the persistent decrease in cell density,
we aimed to determine whether or not proliferation of ICC is also involved in the development of the esophagus ( table 1 ). BrdU was injected 24 h before sacrifice, and the results indicated that Kit/BrdU double-labeled cells could be seen at P2 ( fig. 4 a–c), and were still present at P7 ( fig. 4 d–f) and P12 ( fig. 4 g–i), although the number of proliferative ICC gradually decreased during this period. It was surprising that the Kit+/BrdU+ cells could be de-tected at P18 ( fig. 4 j–l) and P28 ( fig. 4 m–o), despite the dramatically decreasing ICC density at these time points.
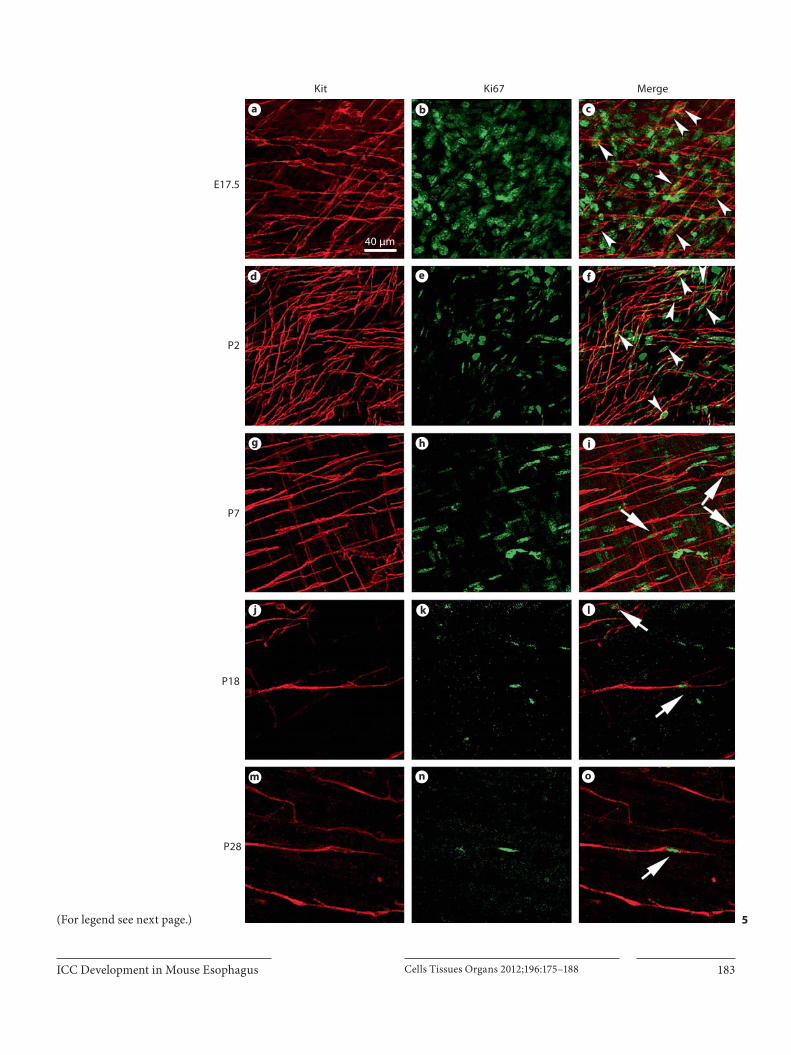
To further confirm the proliferative events of ICC, anti-Ki67 antibody was used to detect the presence of prolifera-tive ICC. Numerous Kit+/Ki67+-labeled cells were scat-tered at E17.5 ( fig. 5 a–c). Such cells were still present at P0 and P2 ( fig. 5 d–f) at a high density. At P7, a few Kit+/Ki67+ double-labeled cells were observed ( fig. 5 g–i). Similar to the results using a BrdU incorporation assay, Kit+/Ki67+ cells were still present at either P18 ( fig. 5 j–l) or P28 ( fig.5 m–o), implying that the proliferation of ICC would persist from late embryonic stage to adulthood in the esophagus.
He /Yang /Wen /Tang /Xiao /Han /Yu /Zhang /Mei
Cells Tissues Organs 2012;196:175–188178
Table 1. Mean density of Kit+ ICC (in mm–2) in the esophagus during late embryonic and postnatal development
E17.5 P0 P2 P7 P12 P18 P26 P38
Proximal esophagus 602816.3 375.085.1 296.285.3 111.284.2 78.881.1 38.582.5 23.884.5 22.881.6Distal esophagus 897.2812.4a 751.287.6a 418.584.2a 292.082.5a 133.884.4a 48.882.3a 33.683.7a 25.988.2General density 756.8824.7 563.6818.7b 363.5810.9b 198.688.2b 106.3815.9b 43.783.1b 28.783.3b 24.482.6
M eans 8 SEM (n = 5 mice). a p < 0.05, vs. the upper segment, b p < 0.05, vs. the earlier time point (e.g. P2 vs. P0).
Kit Desmin Merge
P0
P7
P18
P7
P18
80 μm
35 μm
LM
LM
LM
CM
CM
CM
a
b
c
d
e
Fig. 1. Microphotographs of murine esophagus on cross-sections ( a–c ) and whole-mount preparations ( d , e ) showing ICC and mus-cle cells labeled with Kit (red) and desmin (green), respectively, to illustrate alterations in ICC distribution in the proximal esopha-gus at P0 ( a ), P7 ( b , d ) and P18 ( c ). a ICC are prominent at birth and distribute within the inner (circular) and outer (longitudinal) muscle layers. b A number of Kit+ ICC are seen at P7. c At P18,
the density of ICC is obviously decreased. d Confocal images show Kit+ ICC (arrowheads) among desmin+ muscle cells and in parallel with the direction of muscle cells at P7 and striated mus-cle cells which are apparent at P18 ( e ). z-stack of 2 sections (z step of 2 � m). Bar = 80 ( a–c ) or 35 � m ( d , e ). CM = Circular smooth muscle layer; LM = longitudinal smooth muscle layer.

ICC Development in Mouse Esophagus Cells Tissues Organs 2012;196:175–188 179
E13.5 E17.5 P0
P2 P12 P36
Kit ANO1 Merge
20 μm
80 μm
50 μm
P2
Lon
git
udin
al m
uscl
e la
yer d
irec
tion
Circular muscle layer direction
a’a b
g h i
c
d e f
Fig. 2. Confocal microphotographs showing the Kit+ (red) and Ano1+ (green) ICC in the proximal segment of the esophagus on whole-mount preparations. a Kit+ ICC (arrowheads) clustered at E13.5 without processes. a’ ICC (arrow) at higher magnification. b These cells develop a spindle-like structure with bipolar pro-
cesses at E17.5. c ICC connecting with adjacent cells form a mesh-like network at birth. d Those Kit+ cells have long processes and present at lower density at P2; the density further decreased at P12 ( e ) and P36 ( f ). g –i These Kit+ cells are also labeled with Ano1+ in the muscle layers. Bar = 20 ( a’ ), 80 ( a – f ) or 50 � m ( g–i ).
He /Yang /Wen /Tang /Xiao /Han /Yu /Zhang /Mei
Cells Tissues Organs 2012;196:175–188180
Proximal Distal
E17.5
P2
P7
P18
Lon
git
udal
mus
cle
laye
r dir
ecti
on
Circular muscle layer direction
40 μm
ba
c d
e f
g h
Fig. 3. Confocal images of ICC labeled with Kit (red) on whole-mount prepara-tions of the murine esophagus showing a lower density of ICC in proximal ( a , c , e , g ) than distal segments ( b , d , f , h ) at E17.5 ( a , b ), P2 ( c , d ), P7 ( e , f ) and P18 ( g , h ). Scale bar ( a ) refers to all panels.

ICC Development in Mouse Esophagus Cells Tissues Organs 2012;196:175–188 181
Kit BrdU Merge
P2
P7
P12
P18
P28
40 μm
a b c
d e f
g h i
j k l
m n o
4 (For legend see next page.)

He /Yang /Wen /Tang /Xiao /Han /Yu /Zhang /Mei
Cells Tissues Organs 2012;196:175–188182
Response to the Kit Signaling Blockade To investigate the effects of Kit signaling on ICC de-
velopment in the esophagus during postnatal life, SCF expression was also studied. Our results indicated that either SMC or striated muscle cells expressed SCF at P2 ( fig. 6 b) and P21 ( fig. 6 f), respectively, but it should be noted that ICC density was decreased at P21 ( fig. 6 d) ver-sus P2 ( fig. 6 a); meanwhile SCF expression was down-regulated in striated muscle cells at P18 ( fig. 6 g) and ICC were absent around those cells (arrows in fig. 6 f).
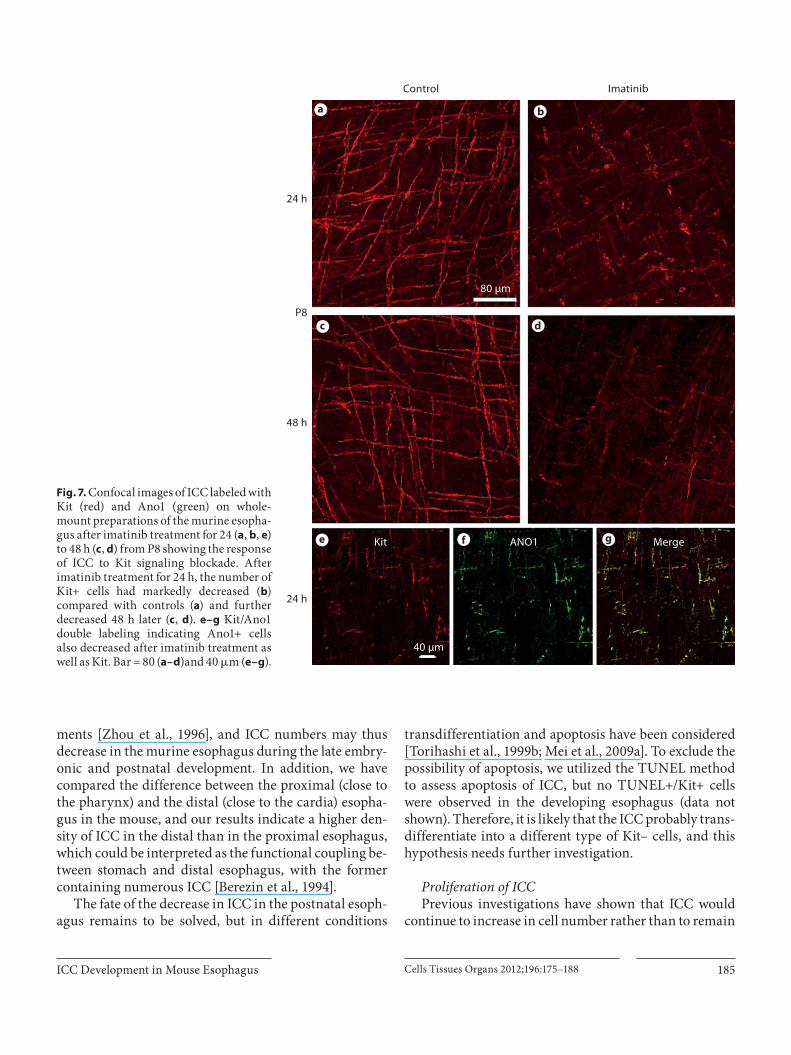
Furthermore, imatinib, a Kit signaling inhibitor, was used to treat 8-day-old mice for either 24 or 48 h. Our re-sults indicated that after 24-hour administration of ima-tinib, there was a dramatic decrease in ICC numbers, and some of them still had Kit+ cell bodies, but processes were evidently shortened compared with the control ( fig. 7 a, b). Moreover, there were very few Kit+ cells at 48 h ( fig. 7 c, d). To further confirm whether there is a real reduction in ICC, Ano1/Kit double labeling was carried out, and the results indicated that imatinib treatment also decreased Ano1+ cells, and these two markers were well co-local-ized ( fig. 7 e–g). These outcomes indicate that Kit signal-ing is essential for the postnatal development of ICC and maintenance of the phenotype in the esophagus.
Discussion
The present study demonstrates that in the murine esophagus: (1) an age-dependent decrease and a distal-to-proximal gradient of ICC numbers is evident in the post-natal development of the murine esophagus; (2) ICC pro-liferation persists from embryonic stage to adulthood, and (3) Kit signaling is essential for the postnatal develop-ment of ICC.
Fig. 4. Confocal images of ICC labeled with Kit (red) and BrdU (green) on whole-mount preparations of the murine esophagus at P2 ( a–c ), P7 ( d–f ), P12 ( g–i ), P18 ( j–l ) and P28 ( m–o ) showing the proliferation of ICC. a–c At P2, a large number of Kit/BrdU dou-ble-labeled cells (arrowheads) are observed. d–f At P7, prolifera-tive ICC present spindle like (arrowheads); the number of these cells is lower than at P2. g–i At P12, a small number of Kit+/BrdU+ (arrowheads) cells also can be observed in a lower density (ar-rows). Such cells (arrows) were still present at P18 ( j–l ) and P28 ( m–o ). Scale bar ( a ) refers to all panels.
ICC in the Esophagus during Late Embryonic and Postnatal Periods During the embryonic period, ICC progenitors begin
to express Kit protein, a marker of ICC, in the foregut on E12 and hindgut of mice on E14 [Wu et al., 2000], respec-tively. Our results indicated that Kit+ cells were present in the esophagus at E13.5, and these cells have round cell bodies with slender and short processes which may be-long to affected ICC progenitors [Huizinga and White, 2008; Lorincz et al., 2008]. Besides Kit, we have utilized another marker of ICC, Ano1, a Ca 2+ -activated Cl – chan-nel, which is also named DOG1 or TMEM16A to label ICC [Gomez-Pinilla et al., 2009; Han et al., 2010b]. The double labeling indicates that almost all Kit+ cells also express Ano1, i.e. they could be used to label ICC in the esophagus. These Kit+ cells gradually develop into spin-dle-like cells at E17.5, and form mesh-like networks with adjacent cells with their processes. A similarly organized ICC network has been observed in the canine distal esophagus [Berezin et al., 1994]. ICC in the esophagus are similar to those located within the muscular layers (ICC-IM) of the stomach, intestines and cecum [Serio et al., 1991; Burns et al., 1997; Horisawa et al., 1998; Ordog et al., 1999; Han et al., 2010a]. Previous studies have shown that ICC-IM play an important role in NO-dependent neurotransmission in the lower esophageal and pyloric sphincter, and loss of these cells may lead to dysfunction of relaxation and normal motility in the sphincters [Ward et al., 1998].
In the esophagus, those ICC distributed within the outer (longitudinal) and inner (circular) muscle layers and occurred in parallel with muscle cells, consistent with previous observations of ICC in the esophagus of cats [Huizinga et al., 2008], humans [Torihashi et al., 1999a; Radenkovic et al., 2010], rats [Farre et al., 2007], guinea pigs [Burns et al., 1997] and adult mice [Rumessen et al., 2001]. ICC density further decreased throughout the late embryonic and postnatal periods, but ICC pro-cesses gradually elongated and were closely in contact with adjacent muscle cells, which may contribute to their functions. The exact function of ICC in the esophagus remains poorly defined; previous studies have provided evidence in support of the hypothesis that esophageal ICC may act as mediators of neural inputs [Daniel and Posey-Daniel, 1984; Faussone-Pellegrini and Cortesini, 1985; Sang and Young, 1997; Rumessen et al., 2001] and also mechanosensors [Thuneberg, 1999; Rumessen et al., 2001]. However, it should be noted that some of the ob-servations have been carried out in SMC and ICC, such as ICC and lower esophageal sphincter [Daniel and Posey-
ICC Development in Mouse Esophagus Cells Tissues Organs 2012;196:175–188 183
Kit Ki67 Merge
40 μm
E17.5
P2
P7
P18
P28
a b c
d e f
g h i
j k l
m n o
5(For legend see next page.)

He /Yang /Wen /Tang /Xiao /Han /Yu /Zhang /Mei
Cells Tissues Organs 2012;196:175–188184
Daniel, 1984], and there is still lack of substantial evi-dence in support of ICC as mediators of neural inputs, especially ICC presenting in low density within the esophagus which has lost SMC. ICC seem unlikely to act as a pacemaker in the murine esophagus due to the ab-sence of slow waves in striated muscles.
ICC Density Changes in the Esophagus We have shown that the density of ICC in the murine
colon and small intestine did not change considerably from birth to adulthood [Torihashi et al., 1997; Mei et al., 2009c; Han et al., 2010a]; however, in the esophagus, ICC density gradually decreased from E17.5 to adulthood (P36), and the value at P36 was about one twentieth of that observed at P0. Moreover, it should be noted that the ICC in the intestines gradually form an intact and com-plex network after birth, but the close connection be-
tween those cells in the esophagus could be found only from E17.5 to P7; thereafter, these cells were distributed separately along with a decrease in density. Our hypoth-esis is that the changes in ICC in the esophagus may be related to differences in the musculature. Distinct from other segments of the murine GI tract, the esophageal musculature initially develops as smooth muscle, and striated muscles begin to appear at E14.5; eventually the esophageal muscularis propria becomes completely stri-ated at P14, although it is subject to controversy that the smooth-to-striated muscle transition occurs through transdifferentiation [Zhou et al., 1996; Stratton et al., 2000; Rumessen et al., 2001; Rishniw et al., 2009] or stri-ated myogenesis in parallel to SMC apoptosis [Rishniw et al., 2003; Rishniw et al., 2007; Katori et al., 2010]. Differ-ent from that of mice, the esophageal wall of humans is composed of striated muscles in the proximal segment, and smooth muscle in the distal portion, and a mixture in the middle portion [Kallmunzer et al., 2008]. The Kit+ ICC are significantly increased in density in the lower third segment compared with the other two segments, with the number being the lowest in the upper third [Radenkovic et al., 2010]; furthermore this difference be-comes apparent along with the emergence of striated muscles from 9 to 10 weeks of gestation [Katori et al., 2010; Radenkovic et al., 2010]. Therefore, it seems that ICC development differs between SMC and striated mus-cle cells, respectively, in these distinct microenviron-
g
P2
Mea
n in
ten
sity
of S
CF
P18
*
4
0
8
12
Kit SCF Merge
P2
P18
50 μm
a b c
d e f
Fig. 5. Confocal images of ICC labeled with Kit (red) and Ki67 (green) on whole-mount preparations of the murine esophagus at E17.5 ( a–c ), P2 ( d–f ), P7 ( g–i ), P18 ( j–l ) and P28 ( m–o ) showing the proliferation of ICC. a–c At E17.5, a number of Kit/Ki67 dou-ble-labeled cells was observed (arrowheads). d–f At P2, these cells presented with spindle-like structure (arrowheads). g–i At P7, those Kit+/Ki67+ cells are decreased in density (arrows) but still present at P18 ( j–l ) and P28 ( m–o ). Scale bar ( a ) refers to all panels.
Fig. 6. Microphotographs of the murine proximal esophagus on cross-sections showing Kit+ ICC (red) and SCF (green) at P2 ( a–c ) and P18 ( d–f ). a–c ICC are prominent at P2 and SCF is expressed by muscle cells. d–f At P18, the density of ICC is obviously de-
creased and SCF expression in muscle cells is down-regulated (ar-rows). g SCF intensity is significantly lower at P18 than that at P2 (n = 5, p ! 0.05). Scale bar ( a ) refers to all panels.
ICC Development in Mouse Esophagus Cells Tissues Organs 2012;196:175–188 185
Control Imatinib
24 h
48 h
24 h
P8
80 μm
40 μm
Kit ANO1 Merge
a b
c d
e f g
ments [Zhou et al., 1996], and ICC numbers may thus decrease in the murine esophagus during the late embry-onic and postnatal development. In addition, we have compared the difference between the proximal (close to the pharynx) and the distal (close to the cardia) esopha-gus in the mouse, and our results indicate a higher den-sity of ICC in the distal than in the proximal esophagus, which could be interpreted as the functional coupling be-tween stomach and distal esophagus, with the former containing numerous ICC [Berezin et al., 1994].
The fate of the decrease in ICC in the postnatal esoph-agus remains to be solved, but in different conditions
transdifferentiation and apoptosis have been considered [Torihashi et al., 1999b; Mei et al., 2009a]. To exclude the possibility of apoptosis, we utilized the TUNEL method to assess apoptosis of ICC, but no TUNEL+/Kit+ cells were observed in the developing esophagus (data not shown). Therefore, it is likely that the ICC probably trans-differentiate into a different type of Kit– cells, and this hypothesis needs further investigation.
Proliferation of ICC Previous investigations have shown that ICC would
continue to increase in cell number rather than to remain
Fig. 7. Confocal images of ICC labeled with Kit (red) and Ano1 (green) on whole-mount preparations of the murine esopha-gus after imatinib treatment for 24 ( a , b , e ) to 48 h ( c , d ) from P8 showing the response of ICC to Kit signaling blockade. After imatinib treatment for 24 h, the number of Kit+ cells had markedly decreased ( b ) compared with controls ( a ) and further decreased 48 h later ( c , d ). e–g Kit/Ano1 double labeling indicating Ano1+ cells also decreased after imatinib treatment as well as Kit. Bar = 80 ( a–d )and 40 � m ( e–g ).

He /Yang /Wen /Tang /Xiao /Han /Yu /Zhang /Mei
Cells Tissues Organs 2012;196:175–188186
constant during the postnatal period in both small intes-tine and colon, and an age-dependent proliferation was involved along with the dramatic increase in intestinal diameter and length [Mei et al., 2009c; Han et al., 2010a]. In this case, we wanted to know whether or not prolif-eration is also involved in the postnatal development of ICC in the esophagus, since there is a similar increase in length and diameter. Therefore, BrdU incorporation was used to assess the proliferative events. BrdU injection was carried out 24 h before sacrifice, and our results indicated that numerous Kit+/BrdU+ cells were found in the esoph-ageal wall from P2 to P7. It is surprising that such cells were still present in the esophagus from P12 to adulthood (P28), although ICC density dramatically decreased dur-ing that period. Those Kit+/BrdU+ cells may originate from the proliferation of Kit+ ICC and/or the prolifera-tion of Kit– ICC progenitors, and then differentiate into Kit+ cells [Huizinga and White, 2008; Lorincz et al., 2008].
To further confirm the proliferation of Kit+ cells, Ki67, a nuclear protein that is expressed in proliferating cells, was detected [Ohta and Ichimura, 2000], and our results indicated a number of Kit+ ICC expressing Ki67 from E17.5 to P7, and, similarly, some Kit+ cells were still Ki67+ from P12 to adulthood (P36). These results are consistent with previous reports that have shown the proliferation of ICC would persist until adulthood in the small intes-tine and colon during postnatal development [Mei et al., 2009c; Han et al., 2010a; Tharayil et al., 2010].
The proliferative ICC Kit+ ICC in the stomach and small intestine may originate from ICC progenitors which are characterized by the co-expression of insulin-like growth factor 1 receptor (IGF-1R)/insulin receptor/CD34 and Kit [Lorincz et al., 2008; Mei et al., 2009a]. In this case, we also have tried to investigate whether or not those Kit+/IGF-1R+ ICC progenitors may give rise toproliferative ICC, but unfortunately we did not observe any IGF-1R/Kit double-labeled cells (online suppl. fig.1, www.karger.com/doi/10.1159/000332381). So it seems likely those proliferative ICC may originate from mature ICC rather than ICC progenitors, or alternatively ICC progenitors in the esophagus may be different from those in the small intestine and stomach. All these possibilities need further investigation, particularly the ultrastruc-ture and biologic characteristics of ICC in the esophagus.
Kit Signaling Effects on ICC Development It seems difficult to understand the fact that ICC con-
tinue to proliferate, while the actual cell number keeps falling in the murine esophagus from birth to adulthood.
One explanation is the dramatic change in the microen-vironment [Faussone-Pellegrini et al., 1990; Sperelakis and Daniel, 2004; Horvath et al., 2006]. Kit ligand, SCF, is produced by muscle cells and neurons, and binds with Kit and activates Kit signaling. It has been well docu-mented that Kit signaling is essential for the genesis of ICC in the stomach, small intestine and colon in mice and rats [Maeda et al., 1992; Ward et al., 1994; Huizinga et al., 1995; Torihashi et al., 1997; Nakama et al., 1998; Tori-hashi et al., 1999b; Beckett et al., 2007; Mei et al., 2009b; Han et al., 2010b]. Although SCF mRNA has been de-tected in the striated muscles of the esophagus using RT-PCR [Rumessen et al., 2001], SCF expression levels in muscle cells at different ages have not been determined. Therefore, we studied the expression level of SCF after immunofluorescent staining of esophageal sections. The results indicated that both the SMC and the striated mus-cle cells express SCF, but the SCF level was much lower in striated muscle cells, and ICC are absent around these low-SCF-expressing striated muscle cells suggesting the down-regulation of SCF may be related to the decreased ICC density.
It has been shown that ICC are present in the fetal esophagus but absent in adult ICC-deficient Kit w-lacz /Kit Wv mice [Rumessen et al., 2001], which implies absence of the Kit signal would not affect the genesis of ICC, but would lead to aplasia of ICC in adults. In this case, we tried to understand whether Kit signaling is required for the phenotype of ICC to remain unchanged. Therefore, imatinib, a Kit signal inhibitor, was used, and our results indicated that imatinib treatment leads to a dramatic de-crease in ICC density, which was confirmed by Kit and Ano1 staining, suggesting Kit signaling is essential to maintain the phenotype of ICC in the esophagus.
In summary, this paper has shown an age- and Kit-dependent development of ICC in the murine esophagus during late embryonic and postnatal stages, and these outcomes will increase our understanding of the ICC de-velopment in a microenvironment in transition from SMC into striated muscles. This field of research would benefit from further studies on the possible role for ICC in the control of motility in the esophagus.
Acknowledgment
This work was supported in part by the High Education Re-form Key Project of Chongqing (09-2-006).

ICC Development in Mouse Esophagus Cells Tissues Organs 2012;196:175–188 187
References
Beckett, E.A., S. Ro, Y. Bayguinov, K.M. Sanders, S.M. Ward (2007) Kit signaling is essential for development and maintenance of inter-stitial cells of Cajal and electrical rhythmic-ity in the embryonic gastrointestinal tract. Dev Dyn 236: 60–72.
Berezin, I., E.E. Daniel, J.D. Huizinga (1994) Ul-trastructure of interstitial cells of Cajal in the canine distal esophagus. Can J Physiol Phar-macol 72: 1049–1059.
Burns, A.J., T.M. Herbert, S.M. Ward, K.M. Sanders (1997) Interstitial cells of Cajal in the guinea-pig gastrointestinal tract as re-vealed by c-Kit immunohistochemistry. Cell Tissue Res 290: 11–20.
Daniel, E.E., V. Posey-Daniel (1984) Neuromus-cular structures in opossum esophagus: role of interstitial cells of Cajal. Am J Physiol 246: G305–G315.
Farre, R., X.Y. Wang, E. Vidal, A. Domenech, M. Pumarola, P. Clave, J.D. Huizinga, M. Ji-menez (2007) Interstitial cells of Cajal and neuromuscular transmission in the rat lower oesophageal sphincter. Neurogastroenterol Motil 19: 484–496.
Faussone-Pellegrini, M.S., C. Cortesini (1985) Ultrastructural features and localization of the interstitial cells of Cajal in the smooth muscle coat of human esophagus. J Submi-crosc Cytol 17: 187–197.
Faussone-Pellegrini, M.S., D. Pantalone, C. Cor-tesini (1990) Smooth muscle cells, interstitial cells of Cajal and myenteric plexus interrela-tionships in the human colon. Acta Anat (Basel) 139: 31–44.
Fox, E.A., R.J. Phillips, M.S. Byerly, E.A. Baro-nowsky, M.M. Chi, T.L. Powley (2002) Selec-tive loss of vagal intramuscular mechanore-ceptors in mice mutant for steel factor, the c-Kit receptor ligand. Anat Embryol (Berl) 205: 325–342.
Gomez-Pinilla, P.J., S.J. Gibbons, M.R. Bardsley, A. Lorincz, M.J. Pozo, P.J. Pasricha (2009) Ano1 is a selective marker of interstitial cells of Cajal in the human and mouse gastroin-testinal tract. Am J Physiol Gastrointest Liv-er Physiol 296: G1370–G1381.
Han, J., W.H. Shen, Y.Z. Jiang, B. Yu, Y.T. He, N. Li, F. Mei (2010a) Distribution, development and proliferation of interstitial cells of Cajal in murine colon: an immunohistochemical study from neonatal to adult life. Histochem Cell Biol 133: 163–175.
Han, J., Y.P. Zhou, Y.Z. Jiang, Y.T. He, F. Mei (2010b) Postnatal development of interstitial cells of Cajal in mouse colon in response to Kit signal blockade with imatinib (Glivec � ). Acta Histochem 112: 215–221.
Horiguchi, K., G.S. Semple, K.M. Sanders, S.M. Ward (2001) Distribution of pacemaker function through the tunica muscularis of the canine gastric antrum. J Physiol 537: 237–250.
Horisawa, M., Y. Watanabe, S. Torihashi (1998) Distribution of c-Kit immunopositivecells in normal human colon and in Hirsch–sprung’s disease. J Pediatr Surg 33: 1209–1214.
Horvath, V.J., H. Vittal, A. Lorincz, H. Chen,G. Meida-Porada, D. Redelman, T. Ordog (2006) Reduced stem cell factor links smooth myopathy and loss of interstitial cells of Cajal in murine diabetic gastroparesis. Gastroen-terology 130: 759–770.
Huizinga, J.D., D.E. Reed, I. Berezin, X.Y. Wang, D.T. Valdez, L.W. Liu, N.E. Diamant (2008) Survival dependency of intramuscular ICC on vagal afferent nerves in the cat esophagus. Am J Physiol Regul Integr Comp Physiol 294: R302–R310.
Huizinga, J.D., L. Thuneberg, M. Kluppel, J. Malysz, H.B. Mikkelsen, A. Bernstein (1995) W/Kit gene required for interstitial cells of Cajal and for intestinal pacemaker activity. Nature 373: 347–349.
Huizinga, J.D., E.J. White (2008) Progenitor cells of interstitial cells of Cajal: on the road to tis-sue repair. Gastroenterology 134: 1252–1254.
Kallmunzer, B., B. Sorensen, W.L. Neuhuber, J. Worl (2008) Enteric co-innervation of stri-ated muscle fibres in human oesophagus. Neurogastroenterol.Motil 20: 597–610.
Katori, Y., B.H. Cho, C.H. Song, M. Fujimiya, G. Murakami, T. Kawase (2010) Smooth-to-striated muscle transition in human esopha-gus: an immunohistochemical study using fetal and adult materials. Ann Anat 192 : 33–41
Kluppel, M., J.D. Huizinga, J. Malysz, A. Bern-stein (1998) Developmental origin and Kit-dependent development of the interstitial cells of Cajal in the mammalian small intes-tine. Dev Dyn 211: 60–71.
Lecoin, L., G. Gabella, N. Le Douarin (1996) Or-igin of the c-kit-positive interstitial cells in the avian bowel. Development 122: 725–733.
Lin, L., L.M. Xu, W. Zhang, Y.B. Ge, Y.R. Tang, H.J. Zhang, X.L. Li, J.D. Chen (2010) Roles of stem cell factor on the depletion of intersti-tial cells of Cajal in the colon of diabetic mice. Am J Physiol Gastrointest Liver Physi-ol 298 : G241–G247.
Liu, L.W., J.D. Huizinga (1994) Canine colonic circular muscle generates action potentials without the pacemaker component. Can J Physiol Pharmaco l 72 : 70–81.
Lorincz, A., D. Redelman, V.J. Horvath, M.R. Bardsley, H. Chen, T. Ordog (2008) Progeni-tors of interstitial cells of Cajal in the postna-tal murine stomach. Gastroenterology 134: 1083–1093.
Maeda, H., A. Yamagata, S. Nishikawa, K. Yoshi-naga, S. Kobayashi, K. Nishi, S. Nishikawa (1992) Requirement of c-kit for development of intestinal pacemaker system. Develop-ment 116: 369–375.
Mei, F., S. Guo, Y.T. He, J. Zhu, D.S. Zhou, J.Q. Niu, H.Z. Wang, Y.P. Tian (2009a) Apoptosis of interstitial cells of Cajal, smooth muscle cells, and enteric neurons induced by intes-tinal ischemia and reperfusion injury in adult guinea pigs. Virchows Arch 454: 401–409.
Mei, F., J. Han, Y. Huang, Z.Y. Jiang, C.J. Xiong, D.S. Zhou (2009b) Plasticity of interstitial cells of Cajal: a study in the small intestine of adult guinea pigs. Anat Rec (Hoboken) 292: 985–993.
Mei, F., J. Zhu, S. Guo, D.S. Zhou, J. Han, B. Yu, S.F. Li, Z.Y. Jiang, C.J. Xiong (2009c) An age-dependent proliferation is involved in the postnatal development of interstitial cells of Cajal in the small intestine of mice. Histo-chem Cell Biol 131: 43–53.
Mikkelsen, H.B., J. Malysz, J.D. Huizinga, L. Thuneberg (1998) Action potential genera-tion, Kit receptor immunohistochemistry and morphology of steel-Dickie (Sl/Sld) mu-tant mouse small intestine. Neurogastroen-terol Motil 10: 11–26.
Nakagawa, T., H. Misawa, Y. Nakajima, M. Taka-ki (2005) Absence of peristalsis in the ileum of W/W(V) mutant mice that are selectively deficient in myenteric interstitial cells of Cajal. J Smooth Muscle Res 41: 141–151.
Nakama, A., S. Hirota, T. Okazaki, K. Nagano, S. Kawano, M. Hori, Y. Kitamura (1998) Dis-turbed pyloric motility in Ws/Ws mutant rats due to deficiency of c-kit-expressing in-terstitial cells of Cajal. Pathol Int 48 : 843–849.
Ohta, Y., K. Ichimura (2000) Proliferation mark-ers, proliferating cell nuclear antigen, Ki67, 5-bromo-2 � -deoxyuridine, and cyclin D1 in mouse olfactory epithelium. Ann Otol Rhi-nol Laryngol 109: 1046–1048.
Ordog, T., S.M. Ward, K.M. Sanders (1999) In-terstitial cells of Cajal generate electrical slow waves in the murine stomach. J Physiol 518: 257–269.
Patapoutian, A., B.J. Wold, R.A. Wagner (1995) Evidence for developmentally programmed transdifferentiation in mouse esophageal muscle. Science 270 : 1818–1821.
Radenkovic, G., I. Ilic, D. Zivanovic, S. Vlajko-vic, V. Petrovic, O. Mitrovic (2010) C-kit-im-munopositive interstitial cells of Cajal in hu-man embryonal and fetal oesophagus. Cell Tissue Res 340: 427–436.
Rishniw, M., P.J. Fisher, R.M. Doran, S.P. Bliss, M.I. Kotlikoff (2009) Striated myogenesis and peristalsis in the fetal murine esophagus occur without cell migration or interstitial cells of Cajal. Cells Tissues Organs 189: 410–419.
Rishniw, M., P.W. Fisher, R.M. Doran, E. Mead-ows, W.H. Klein, M.I. Kotlikoff (2007) Smooth muscle persists in the muscularis ex-terna of developing and adult mouse esopha-gus. J Muscle Res Cell Motil 28: 153–165.

He /Yang /Wen /Tang /Xiao /Han /Yu /Zhang /Mei
Cells Tissues Organs 2012;196:175–188188
Rishniw, M., H.B. Xin, K.Y. Deng, M.I. Kotlikoff (2003) Skeletal myogenesis in the mouse esophagus does not occur through transdif-ferentiation. Genesis 36: 81–82.
Rumessen, J.J., A. Kerchove, S. Mignon, F. Ber-nex, J.P. Timmermans, S.N. Schiffmann, J.J. Panthier, J.M. Vanderwinden (2001) Inter-stitial cells of Cajal in the striated muscula-ture of the mouse esophagus. Cell Tissue Res 306: 1–14.
Rumessen, J.J., L. Thuneberg (1996) Pacemaker cells in the gastrointestinal tract: interstitial cells of Cajal. Scand J Gastroenterol 216(sup-pl): 82–94.
Sanders, K.M. (1996) A case for interstitial cells of Cajal as pacemakers and mediators of neu-rotransmission in the gastrointestinal tract. Gastroenterology 111: 492–515.
Sanders, K.M., S.M. Ward (2007) Kit mutants and gastrointestinal physiology. J Physiol 578: 33–42.
Sang, Q., H.M. Young (1997) Development of nicotinic receptor clusters and innervation accompanying the change in muscle pheno-type in the mouse esophagus. J Comp Neurol 386: 119–136.
Serio, R., C. Barajas-Lopez, E.E. Daniel, I. Be-rezin, J.D. Huizinga (1991) Slow-wave activ-ity in colon: role of network of submucosal interstitial cells of Cajal. Am J Physiol 260: G636–G645.
Shimojima, N., T. Nakaki, Y. Morikawa, K. Hoshino, M. Kitajima (2005) Imatinib blocks spontaneous mechanical activities in the adult mouse small intestine: possible in-hibition of c-Kit signaling. Pharmacology 74: 95–99.
Sperelakis, N., E.E. Daniel (2004) Activation of intestinal smooth muscle cells by interstitial cells of Cajal in simulation studies. Am J Physiol Gastrointest Liver Physiol 286 : G234–G243.
Stratton, C.J., Y. Bayguinov, K.M. Sanders, S.M. Ward (2000) Ultrastructural analysis of the transdifferentiation of smooth muscle to skeletal muscle in the murine esophagus. Cell Tissue Res 301: 283–298.
Tharayil, V.S., M.M. Wouters, J.E. Stanich, et al. (2010) Lack of serotonin 5-HT2B receptor al-ters proliferation and network volume of in-terstitial cells of Cajal in vivo. Neurogastro-enterol Motil 22: 462–410.
Thuneberg, L. (1999) One hundred years of in-terstitial cells of Cajal. Microsc Res Tech 47: 223–238.
Torihashi, S., M. Horisawa, Y. Watanabe (1999a) c-Kit immunoreactive interstitial cells in the human gastrointestinal tract. J Auton Nerv Syst 75: 38–50.
Torihashi, S., K. Nishi, Y. Tokutomi, T. Nishi, S. Ward, K.M. Sanders (1999b) Blockade of kit signaling induces transdifferentiation of in-terstitial cells of Cajal to a smooth muscle phenotype. Gastroenterology 117: 140–148.
Torihashi, S., S.M. Ward, K.M. Sanders (1997) Development of c-Kit-positive cells and the onset of electrical rhythmicity in murine small intestine. Gastroenterology 112: 144–155.
Ward, S.M., A.J. Burns, S. Torihashi, S.C. Har-ney, K.M. Sanders (1995) Impaired develop-ment of interstitial cells and intestinal elec-trical rhythmicity in steel mutants. Am J Physiol 269: C1577–C1585.
Ward, S.M., A.J. Burns, S. Torihashi, K.M. Sand-ers (1994) Mutation of the proto-oncogene c-kit blocks development of interstitial cells and electrical rhythmicity in murine intes-tine. J Physiol 480: 91–97.
Ward, S.M., G. Morris, L. Reese, X.Y. Wang, K.M. Sanders (1998) Interstitial cells of Cajal mediate enteric inhibitory neurotransmis-sion in the lower esophageal and pyloric sphincters. Gastroenterology 115: 314–329.
Ward, S.M., K.M. Sanders (2001) Physiology and pathophysiology of the interstitial cell of Cajal: from bench to bedside. I. Functional development and plasticity of interstitial cells of Cajal networks. Am J Physiol Gastro-intest Liver Physiol 281: G602–G611.
Wong, W.C., S.H. Tan, T.Y. Yick, E.A. Ling (1990) Ultrastructure of interstitial cells of Cajal at the gastro-oesophageal junction of the monkey (Macaca fascicularis) . Acta Anat (Basel) 138: 318–326.
Wu, J.J., T.P. Rothman, M.D. Gershon (2000) De-velopment of the interstitial cell of Cajal: or-igin, kit dependence and neuronal and non-neuronal sources of kit ligand. J Neurosci Res 59: 384–401.
Young, H.M. (1999) Embryological origin of in-terstitial cells of Cajal. Microsc Res Tech 47: 303–308.
Young, H.M., D. Ciampoli, B.R. Southwell, D.F. Newgreen (1996) Origin of interstitial cells of Cajal in the mouse intestine. Dev Biol 180: 97–107.
Zhou, D.S., J. Desaki, T. Komuro (1996) Neuro-muscular junctions of longitudinal and cir-cular muscle fibers of the guinea-pig esopha-gus and their relation to myenteric plexus. J Auton Nerv Syst 58: 63–68.
Copyright © 2022 FDOKUMEN




















