
LAPORAN PENELITIAN KARIOTIPE CACING NIPAH ...
19
LAPORAN PENELITIAN KARIOTIPE CACING NIPAH Namalycastis rhodochorde (POLYCHAETA : NEREIDIDAE) Oleh : SITI IFADATIN, S.Si., M.Si JUNARDI, S.Si., M.Si FIRMAN SAPUTRA, S.Si DIBIAYAI OLEH DANA DIPA FMIPA UNTAN TAHUN 2009 SURAT PERJANJIAN PELAKSANAAN PENELITIAN NOMOR : 1486/H22.8/PL/2009 TANGGAL 29 JULI 2009 JURUSAN BIOLOGI FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM UNIVERSITAS TANJUNGPURA PONTIANAK 2009
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of LAPORAN PENELITIAN KARIOTIPE CACING NIPAH ...

LAPORAN PENELITIAN
KARIOTIPE CACING NIPAH Namalycastis rhodochorde
(POLYCHAETA : NEREIDIDAE)
Oleh :
SITI IFADATIN, S.Si., M.Si
JUNARDI, S.Si., M.Si
FIRMAN SAPUTRA, S.Si
DIBIAYAI OLEH DANA DIPA FMIPA UNTAN TAHUN 2009
SURAT PERJANJIAN PELAKSANAAN PENELITIAN
NOMOR : 1486/H22.8/PL/2009 TANGGAL 29 JULI 2009
JURUSAN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN
ALAM
UNIVERSITAS TANJUNGPURA
PONTIANAK
2009

1
BAB I
PENDAHULUAN
1.1. Latar Belakang
Cacing Nipah (Namalycastis rhodochorde.) merupakan salah satu cacing
subfamili Namanereididae famili Nereididae dari kelas polychaeta (Glasby, 1999).
Namalycastis termasuk cacing yang hidup di daerah tropis dan subtropik dengan
ukuran panjang antara 100-200 cm (Glasby, et al., 2003). Cacing ini memiliki
peran sangat penting sebagai pendaur ulang nutrien (Beesley et al., 2000) dan
sumber protein (Batista et al., 2003). Pemberian cacing nipah sebagai pakan dapat
meningkatkan kandungan asam lemak tidak jenuh pada ikan dan udang (Olive,
1999).
Cacing nipah termasuk salah satu spesies polychaeta yang banyak dijumpai
di estuari Sei Kakap Kalimantan Barat. Cacing Namalycastis telah menjadi
komoditas yang diekspor antarnegara sebagai bahan baku pakan ikan dan udang
(Thomson & Alam, 2005). Potensi komersial cacing nipah juga dapat
dikembangkan di Kalimantan Barat. Saat ini pemanfaatan cacing nipah di
Kalimantan Barat umumnya hanya untuk umpan pancing. Cacing ini dijual di
pasar-pasar tradisional dengan harga relatif tinggi. Satu ekor cacing nipah hidup
dihargai Rp. 6.000-10.000 per ekor dengan berat antara 5-10 gram.
Budidaya adalah upaya yang tepat untuk komersialisasi dan mencegah
pengambilan berlebih (over fishing) cacing nipah di alam yang cenderung merusak
habitat. Informasi awal yang harus diketahui dalam biologi cacing nipah adalah
penentuan jenis kelamin. Determinasi kelamin cacing nipah selama ini hanya
diketahui berdasarkan karakter morfologi dan gamet yang dapat dibedakan antara
jantan dan betina pada individu dewasa. Pada individu pradewasa, jenis kelamin
dapat diketahui dengan melihat karakter kromosom.
1.2. Perumusan Masalah
Cacing nipah bersifat dioseus, dapat dibedakan antara individu jantan dan
betina namun hanya pada individu dewasa yang berbeda morfologi dan tingkat
kematangan gamet. Pada individu pradewasa jenis kelamin belum dapat dibedakan.

2
Determinasi kelamin pada cacing nipah merupakan faktor penting yang
berpengaruh untuk mengestimasi populasinya di alam. Pada budidaya, determinasi
kelamin juga akan berpengaruh pada keberhasilan reproduksi artifisial yang selama
ini belum berhasil dilakukan pada cacing nipah. Teknik pemeriksaan dengan
kromosom akan membantu untuk mengetahui lebih awal kenis kelamin cacing
nipah pada individu pradewasa. Penelitian ini akan mencoba mengetahui tipe dan
susunan kromosom (kariotipe) cacing nipah
1.3. Tujuan Penelitian
1. Mengetahui tipe dan susunan kromosom (kariotipe) cacing nipah
2. Mengetahui perbedaan kromosom cacing nipah jantan dan betina
1.4. Manfaat Penelitian
Data hasil penelitian ini dapat digunakan untuk menentukan jenis kelamin
cacing nipah yang dapat mendukung teknik fertilisasi artifisial dalam reproduksi
cacing nipah. Keberhasilan dalam reproduksi cacing ini akan membantu proses dan
metode budidaya dan produksi massal cacing nipah.

3
BAB II
TINJAUAN PUSTAKA
2.1. Cacing Nipah (Namalycastis rhodochorde)
Cacing nipah (Namalycastis rhodochorde) merupakan salah satu cacing
subfamili Namanereididae famili Nereididae dari kelas polychaeta (Glasby, 1999).
Polychaeta telah ditemukan sebanyak 20.000 spesies yang dimasukkan ke dalam
namanereidinae, Namanereis dan Namalycastis. Perbedaan antara kedua genus
tersebut antara lain pada ukuran antenna, bentuk pygidium, ada tidaknya notoseta,
banyak sedikitnya telur matang dan bentuk telur matang (Glasby, 1999).
Cacing nipah termasuk cacing yang hidup di daerah tropis dan subtropik
dengan ukuran panjang antara 100-200 cm (Glasby, et.al., 2003). Di Kalimantan
Barat, cacing nipah banyak dijumpai di estuari Desa Sungai Kakap. Desa Sungai
Kakap merupakan desa pesisir yang memiliki estuaria mangrove yang didominasi
tumbuhan nipah (Nypa sp.) yang merupakan habitat alami cacing nipah.
2.2. Determinasi Kelamin Cacing Nipah
Tipe reproduksi polychaeta umumnya dapat dibedakan menjadi tiga,
hermaprodit simultan, hermaprodit sekuensial dan gonokhoristik (Sella, 2006).
Pada Nereididae, umumnya reproduksinya gonokhoristik, cacing memiliki jenis
kelamin yang terpisah, jantan dan betina. Penentuan jenis kelamin cacing nipah
dilakukan melalui pendekatan morfologi dan anatomi. Pendekatan morfologi
ditentukan berdasarkan panjang, bobot dan warna tubuh serta jumlah segmen.
Perbedaan morfologi jantan dan betina hanya dapat dilihat jika telah
memasuki fase dewasa. Pada fase pradewasa, jantan dan betina hanya dapat dilihat
berdasarkan ada tidaknya sperma dan ovum dalam selom. Individu dewasa cacing
nipah memiliki panjang dan robot tubuh lebih kecil serta jumlah segmen yang lebih
sedikit dibandingkan individu pradewasa. Perubahan morfologi melalui
pengurangan panjang, bobot dan jumlah segmen terjadi pada saat ekdisis. Setelah
ekdisis individu immature berubah menjadi mature, tidak ada data mengenai
lamanya waktu ekdisis.

4
Cacing yang akan memasuki fase remaja (submature) baik jantan maupun
betina memiliki dua pasang mata yang jelas yang akan digunakan untuk berenang
bebas saat memijah (Glasby et al., 2007). Penelitian ini hanya mendapatkan dua
jantan dan dua betina dewasa, oleh sebab itu tidak cukup data untuk mencari
perbedaan yang signifikan berdasarkan bobot dan panjang tubuh serta jumlah
segmen.
Cacing nipah jantan umumnya lebih pendek, bobot tubuh ringan dan jumlah
segmen yang sedikit dibandingkan betina. Perbedaan warna tubuh hanya tampak
pada individu dewasa, sedangkan pada jantan pradewasa sangat mirip betina.
Bagian ventral tubuh posterior cacing jantan dewasa berwarna kehijauan. Warna
tubuh N. rhodochorde jantan yang berwarna kehijauan juga didapatkan oleh M.
Lopuszanski Tahun 2002 di Delta Sungai Mekong (Glasby et al., 2007).
Betina dewasa memiliki panjang, bobot dan jumlah segmen yang lebih
banyak dibandingkan jantan dewasa. Perbedaan warna tubuh betina dan jantan
dewasa dapat dilihat pada bagian ventral tubuh. Betina dewasa memiliki warna
ventral tubuh putih pucat. Betina pradewasa mulai dapat dikenali dari adanya telur
yang berukuran 30µm dan bentuk yang bulat. Ukuran dan bentuk telur <30µm
umumnya amorf dan belum dapat dibedakan dengan eleosit.
2.3. Kromosom dan Kariotipe
Bahan genetik utama jasad eukariot terletak dalam inti sel (nukleus) dan
dikemas sedemikian rupa membentuk struktur yang disebut kromosom. Jumlah
kromosom pada kelompok jasad eukariot sangat bervariasi. Selain kromosom,
beberapa sel eukariot juga mempunyai DNA di luar kromosom yaitu DNA pada
mitokondria dan pada kloroplas (pada sel tumbuhan hijau).
DNA utama pada sel eukariot dikemas dengan sistem yang sangat efisien
dan kompak (Gambar 2.1). DNA dikemas menggunakan protein histon yang terdiri
dari lima macam yaitu H1, H2A, H2B, H3, dan H4. Protein histon digunakan untuk
menggulung molekul DNA sekitar 200 bp membentuk struktur disebut nukleosom.
Pembentukan struktur nukleosom menyebabkan kondensasi atau pengemasan DNA
menjadi enam kali lebih kompak. Rangkaian nukleosom membentuk struktur yang
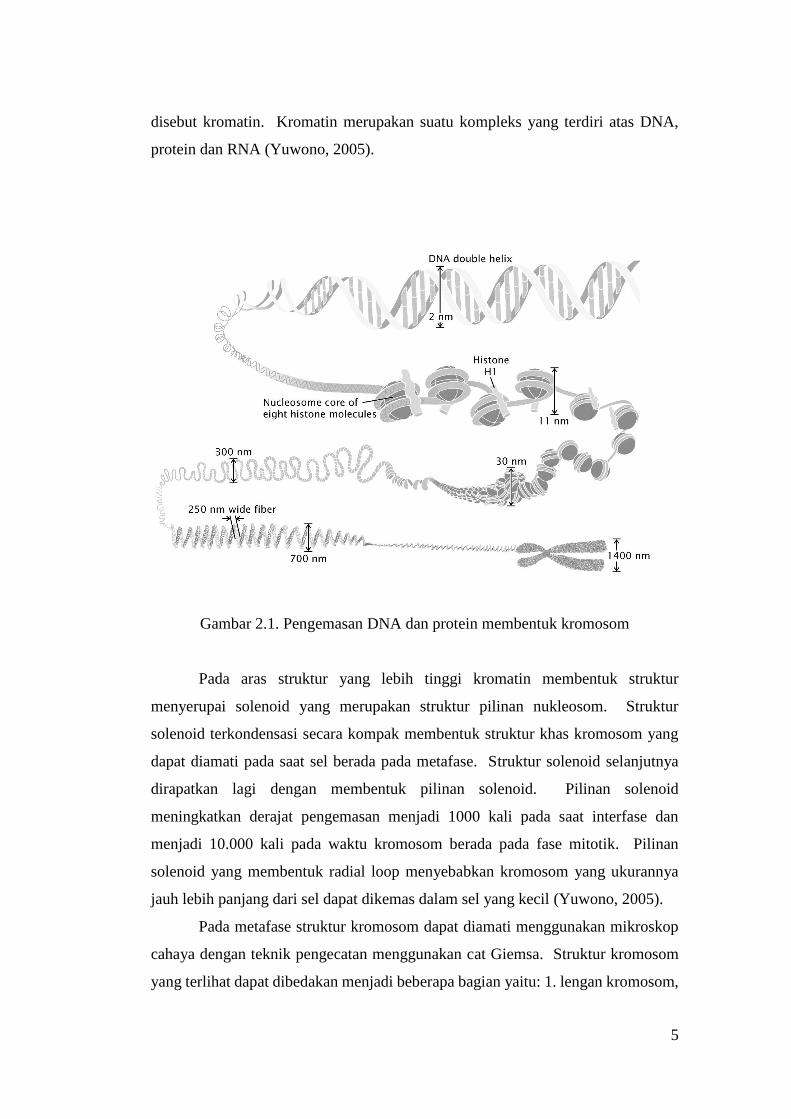
5
disebut kromatin. Kromatin merupakan suatu kompleks yang terdiri atas DNA,
protein dan RNA (Yuwono, 2005).
Gambar 2.1. Pengemasan DNA dan protein membentuk kromosom
Pada aras struktur yang lebih tinggi kromatin membentuk struktur
menyerupai solenoid yang merupakan struktur pilinan nukleosom. Struktur
solenoid terkondensasi secara kompak membentuk struktur khas kromosom yang
dapat diamati pada saat sel berada pada metafase. Struktur solenoid selanjutnya
dirapatkan lagi dengan membentuk pilinan solenoid. Pilinan solenoid
meningkatkan derajat pengemasan menjadi 1000 kali pada saat interfase dan
menjadi 10.000 kali pada waktu kromosom berada pada fase mitotik. Pilinan
solenoid yang membentuk radial loop menyebabkan kromosom yang ukurannya
jauh lebih panjang dari sel dapat dikemas dalam sel yang kecil (Yuwono, 2005).
Pada metafase struktur kromosom dapat diamati menggunakan mikroskop
cahaya dengan teknik pengecatan menggunakan cat Giemsa. Struktur kromosom
yang terlihat dapat dibedakan menjadi beberapa bagian yaitu: 1. lengan kromosom,

6
2. sentromer, dan 3. telomer (Gambar 2.2). Lengan kromosom adalah bagian
kromosom yang mengandung rangkaian gen, sentromer adalah bagian tengah
kromosom yang berfungsi dalam proses distribusi kromosom pada waktu terjadi
pembelahan sel, sedangkan telomer adalah bagian ujung kromosom (Yuwono,
2005).
Gambar 2.2. Struktur kromosom
Pengamatan kromosom menggunakan cat Giemsa dapat diamati adanya
perbedaan dalam pengikatannya terhadap cat tersebut pada bagian-bagian tertentu.
Bagian kromosom yang tidak terlalu rapat pengemasannya mengikat lebih sedikit
cat Giemsa, bagian ini disebut eukromatin. Bagian kromosom yang lebih rapat
mengikat cat Giemsa lebih banyak, bagian ini disebut heterokromatin. Pada jasad
tingkat tinggi kromosom dibedakan atas kromosom non-seks yang disebut autosom
dan kromosom seks yaitu kromosom X dan Y (Yuwono, 2005). Perbedaan dalam
pengikatan cat Giemsa dapat dilihat pada gambar kariotipe manusia (laki-laki)
dibawah ini.

7
Gambar 2.3. Kariotipe manusia (laki-laki)
Kariotipe adalah gambar fotografi yang menunjukkan jumlah dan bentuk
seluruh kromosom yang ada di dalam sel. Dalam teknik kariotipe kromosom
ditampilkan dalam urutan yang menunjukkan perpasangan kromosom homolog.
Kromosom homolog ditetapkan berdasarkan pasangan kromosom yang mempunyai
panjang, posisi sentromer dan pola pewarnaan yang sama (Gambar 2.3).
Teknik kariotipe telah digunakan pada banyak organisme untuk berbagai
tujuan. Tujuan utama adalah mengetahui jumlah kromosom setiap spesies sebagai
salah satu penciri spesies tersebut. Selain itu kariotipe digunakan untuk sistematika
(McBee et.al 1985; Cereali et.al 2008; Fukui 1996: Noleto et.al 2006), membedakan
jantan dan betina (Tosuji et.al., 2004), analisis genom (Yulianty et.al, 2006) dan
melihat pengaruh senyawa kimia terhadap aberasi kromosom (Dixon et.al, 1999).

8
BAB III
BAHAN DAN METODE
3.1. Tempat dan Waktu Penelitian
Sampel diambil dari Muara Sungai Kapuas, Pelaksana penelitian ini akan
dilaksanakan di Laboratorium Hewan Uji FMIPA Untan mulai bulan Agustus
sampai Nopember 2009
3.2. Alat dan Bahan
Alat yang digunakan pada penelitian ini adalah mikroskop, dissecting set,
alat-alat gelas, dan kamera, sedangkan bahan yang digunakan meliputi cacing
nipah, kolkisin, metanol, asam asetat, alkohol 70%, air laut, sodium sitrat, larutan
Giemsa, buffer fosfat 6,8.
3.3. Metode Penelitian
Pembuatan preparat kromosom
Untuk mendapatkan kromosom pada fase metafase digunakan ekor cacing
nipah yang mengalami regenerasi menggunakan metode Ikeda dan Sato (1991).
Cacing nipah yang digunakan untuk percobaan dipilih cacing yang sedang
mengalami regenerasi pada ujung posterior. Cacing kemudian dibius menggunakan
alkohol 5% dan dipotong lebih kurang 1-2 mm.
Potongan ekor dimasukkan dalam kolkisin yang dilarutkan dalam air laut pada
konsentrasi 0,04-0,05%, diinkubasi selama 15-24 jam pada suhu 17-20ºC. Ekor
dipindahkan ke dalam larutan hipotonik sodium sitrat 1% selama 30-40 menit pada
suhu ruang. Ekor difiksasi dengan larutan metanol:asam asetat (3:1) selama 40-60
menit pada suhu ruang. Spesimen yang telah difiksasi diletakkan pada gelas objek
dan dimaserasi menggunakan jarum bedah.
Preparat dikeringkan menggunakan alkohol dan diwarnai dengan larutan
Giemsa 2% yang dilarutkan dalam buffer fosfat pH 6,8. Preparat kemudian diamati
dengan mikroskop cahaya dan difoto. Pengukuran panjang lengan kromosom
menggunakan mikrometer okuler.

9
Pembuatan Kariotipe
Foto kromosom dianalisis dengan menggunakan program Adobe Photoshop
cs2 versi 9. Hasil foto kromosom dicetak hitam putih kemudian setiap kromosom
digunting dan dipasang-pasangkan untuk mengetahui pasangan kromosom
homolog berdasarkan ukuran, bentuk, dan pola pewarnaan.

10
BAB IV
HASIL DAN PEMBAHASAN
4.1. Sel cacing nipah
Berdasarkan prosedur pembuatan kariotipe cacing nipah diperoleh
gambaran sel cacing nipah (Gambar 4.1). Sel cacing nipah tampak jelas tersusun
rapat berukuran sekitar 100 µm. Sel pada fase interfase tampak jelas inti selnya
berbentuk bulat terletak di tengah sel, terwarnai ungu oleh Giemsa, berukuran 30-
50 µm Sitoplasma sel tampak lebih terang dengan pewarnaan Giemsa.
Gambar 4.1. Sel cacing nipah
Keterangan: 1. inti sel; 2. sitoplasma
4.2. Kariotipe cacing nipah
Kariotipe merupakan salah satu cara untuk menentukan jumlah dan tipe
kromosom setiap organisme. Setiap organisme memiliki jumlah dan tipe kromosom
yang bervariasi sehingga kromosom juga merupakan salah satu penciri spesies.
Selain itu dengan teknik kariotipe dapat diketahui jenis kelamin, dan perubahan
dalam jumlah dan struktur kromosom (aberasi kromosom)
Berdasarkan teknik pembuatan kariotipe Ikeda dan Sato (1991) dibuat
kariotipe untuk cacing nipah (Namalycastis rhodochorde). Sampel diambil dari

11
bagian posterior cacing yang sedang mengalami pertumbuhan yang dapat dilihat
dari ukuran yang lebih kecil dibandingkan bagian lainnya. Bagian posterior cacing
dipotong 2 cm kemudian dimasukkan dalam larutan kolkisin. Kolkisin merupakan
senyawa kimia yang dapat menghambat pembentukan apparatus mitotik sehingga
menghambat terjadinya pembelahan sel. Dengan pemberian kolkisin, sel yang
sedang membelah, tahapan pembelahan selnya akan terhenti. Selanjutnya perlakuan
dengan larutan hipotonis (sodium sitrat 1%) untuk menjaga bentuk sel agar lebih
mudah diamati. Bagian posterior tersebut kemudian dimasukkan ke dalam larutan
fiksatif dan diwarnai dengan pewarna Giemsa 2 %.
Berdasarkan teknik pembuatan kariotipe tersebut dapat diperoleh gambaran
seperti kromosom, yaitu bentuk batang panjang dengan bagian tengah lebih sempit
(Gambar 4.2). Panjang kromosom berkisar antara 20 – 50 µm. Kromosom
terwarnai ungu oleh pewarna Giemsa dan terlihat menyebar. Dari hasil pengamatan
diperoleh jumlah kromosom yang sangat banyak dan menyebar tak beraturan,
sehingga tidak dapat dilihat kromosom per selnya. Kondisi ini kemungkinan
disebabkan membran sel rusak sehingga kromosom antar sel bercampur satu
dengan lainnya. Hal ini menyulitkan dalam menghitung jumlah kromosom untuk
setiap sel. Demikian juga dalam menentukan pasangan kromosom homolog dan tipe
kromosomnya. Membran sel yang rusak diduga disebabkan oleh perendaman dalam
larutan hipotonik yang terlalu lama.
Gambar 4.2. Kromosom cacing nipah (Namalycastis rhodochorde)

12
BAB V
SIMPULAN DAN SARAN
Ukuran sel cacing nipah dengan pewarnaan Giemsa sekitar 100 µm. Inti sel
berukuran 30 – 50 µm terwarnai ungu dan sitoplasma terpulas lebih terang.
Kromosom terwarnai ungu oleh Giemsa dengan ukuran 20 – 50 µm. Jumlah
kromosom setiap sel belum dapat ditentukan.
Saran
Teknik kariotipe pada cacing nipah perlu diperbaiki lagi dengan optimasi
pada setiap tahap perlakuan untuk mendapatkan jumlah kromosom setiap sel dan
perbedaan jenis kelamin. Objek untuk pembuatan kariotipe menggunakan cacing
muda yang sedang regenerasi untuk memperoleh sel-sel yang aktif membelah.

13
DAFTAR PUSTAKA
Andries, JC. 2001. Endocrine and environmental control of reproduction in
polychaeta. Canadian Journal Zoology 79:254-270.
Batista.FM, PF. Costa, A. Ramos, AM. Passos, PP. Ferreira, LC. Fonseca. 2003.
Production of ragworm Nereis diversicolor (O.F. Muller, 1776), fed with a
diet for gilthead seabream Sparus auratus L. 1758: survival, growth, feed
utilization and oogenesis. Bol.Inst. Oceanogr. 19:447-451.
Beesley, P.L, GJB. Ross. CJ. Glasby. 2000. Polychaetes and allies: The Southern
synthesis. Fauna of Australia vol. 4A. Polychaeta, Myzostomida,
Pogonophora, Echiura, Sipuncula. Melbourne: SCIRO.
Cereali, SS, Pomini, E., Rosa, R., Zawadzki, LH., Froehlich, O., Caetano, LG.
2008. Genetics and Molecular Research. 7(3) : 583-591.
Costa, PF. 1999. Reproduction and growth in captivity of the polychaete Nereis
diversicolor O.F Muller, 1776, using two different kinds of sediment;
preliminary assays. Bol. Inst. Oceanogr. 15:352-355.
Dixon, DR, Wilson, JT., Pascoe, PL., Parry, JM. 1999. Anaphase Aberration in The
Embryos of The Marine Tubeworm Pomatoceros lamarckii (Polychaeta :
Serpulidae) : A New in vivo test assay for detecting Aneugenus and
Clastogenus in The Marine Environment. Mutagenesis vol. 14 no. 4 pp. 375-
383
Fukui, Y. 1996. Karyotype of The Sea Anemone Aiptasiomorpha sp. (Anthozoa,
Actiniaria) from Japan BiolBell 190: 6-7.
Glasby, CJ. 1999. The Namanereidinae (Polychaeta:Nereididae). Part 1. Taxonomy
and phylogeny. Rec. Aus. Mus. Supp.. 25:1-129.
Glasby, CJ, M. Mogi, K. Takahashi, 2003. Occurance of the polychaete
Namalycastis hawaiiensis Johnson, 1903 (Nereididae: Namanereidinae) in
Pandanus leaf axils on Palau West Pacific. The Beagle. Records of the
Museum and Art Galleries of the Northern Territory. 19: 97-99.
Noleto, RB., Kantek, DLZ., Swarca, AC., Dias, AL., Fenocchio, AS., Cestari, MM.
2006. Karyotype Characterization of Hydromedusa tectifera (Testudines,
Pleurodia) from The Upper Iguacu River in The Brazilian State of Parana.
Genetics and Molecular Biology. 29:2; 263-266.
Olive, PJW. 1999. Polychaete aquaculture and polychaete science: a mutual
synergism. Hydrobiologia 402: 175-183.

14
Thomson, JA, SK Alam. 2005. Analysis of customs trade data to characterize
importation of live bait. Fisheries 30:36-39.
Tosuji, H., Miyamoto, J., Hayata, Y., Sato, M. 2004. Karyotyping of Female and
Male Hediste japonica (Polychaeta, Annelids) in comparison with those of
two closely related species, H. diadore and H. atoka. Zoological Science 21 :
147-152.
Yulianty, M., Pujawati, ED., Badasaufari. 2006. Analisis Kariotipe pisang Mauli.
Bioscientiae vol. 3 no. 2 : 103-109.
Yuwono, T. 2005. Biologi Molekular. Penerbit Erlangga. Hal: 80-83

15
LAMPIRAN
BIODATA
1. Ketua Peneliti
Nama Lengkap dan gelar : Siti Ifadatin, S.Si., M.Si
NIP : 197103272000032001
Jabatan Fungsional : Lektor
Jenis Kelamin : Perempuan
Tempat Tanggal Lahir : Kebumen, 27 Maret 1971
Alamat institusi : Program Studi Biologi FMIPA Untan
Bidang Keahlian : Genetika
Pendidikan
a. Magister Sains Biologi, PPs Institut Pertanian Bogor, 2002
b. Sarjana Biologi, Universitas Gadjah Mada, 1997
Penelitian
2007 : Keanekaragaman genetik genus Garcinia asal Kalimantan
Barat berdasarkan penanda isozim (DIKTI)
2006 : Keanekaragaman genetik padi lokal Kalimantan Barat
berdasarkan penanda isozim (DIKTI)
2004 : Tinjauan taksonomi Brugmansia Pers berdasarkan analisis
isozim (Forum HEDS)
2002 : Kemiripan genetik enam populasi kelapa Dalam asal
Kalimantan Barat berdasarkan penanda RAPD (Random
Amplified Polymorphic DNA) (Tesis)
Pelatihan
2006 : Pelatihan penulisan artikel ilmiah terpusat, 16-18 November
2006, Surabaya. (Peserta)
2005 : Pelatihan teknik analisis isozim, Agustus 2005, Bogor
Publikasi
1. Siti Ifadatin dan E. R. P. Wardoyo. 2007. Keanekaragaman genetik padi
lokal Kalimantan Barat berdasarkan penanda isozim. Floribunda
Pontianak, Desember 2009
Siti Ifadatin, S.Si., M.Si.
NIP. 197103272000032001

16
2. Anggota Peneliti
Nama Lengkap dan gelar : Junardi, S.Si., M.Si
NIP : 132257938
Jabatan Fungsional : Lektor Kepala
Jenis Kelamin : Laki-laki
Tempat Tanggal Lahir : Lampung, 13 Juni 1972
Alamat institusi : Program Studi Biologi FMIPA Untan
Bidang Keahlian : Biologi Laut (Polychaeta)
Pendidikan:
c. Magister Sains Biologi, PPs Institut Pertanian Bogor, 2001
d. Sarjana Biologi, Universitas Lampung, 1997
Penelitian:
2008 : Aspek reproduksi dan Perkembangan Cacing Nipah Namalycastis
rhodochorde (Polychaeta: Nereididae) (Hibah Pekerti Tahun I,
DIKTI)
2007 : Struktur Populasi dan Preferensi Habitat Cacing Nipah
(Namalycastis sp.) di Perairan Sei Kakap Kalimantan Barat (DIKTI)
2004 : Keanekaragaman Cacing Laut (Polychaeta) dan Dekomposisi
Serasah Hutan Mangrove Peniti Besar Kalimantan Barat (DIKTI)
2004: Peneliti Struktur Komunitas Cacing Laut (Polychaeta) di Perairan
Mangrove Peniti Kalimantan Barat (PPD Forum HEDS)
2001 : Keanekaragaman, Pola Penyebaran dan Ciri-ciri Substrat
Polychaeta (Annelida) di Perairan Pantai Timur Lampung Selatan
(Tesis PPs IPB)
Pelatihan dan Workshop
2007 : Manajemen dan Teknik Identifikasi Polychaeta. Pelatihan
Manajeman Laboratorium 18 November 2007, Pontianak,
Kalimantan Barat (Pemateri).
2007 : Keanekaragaman Polychaeta. Pendidikan dan Pelatihan Dasar
(DIKLATSAR) Mahasiswa Biologi FMIPA Untan, 15 Desember
2007, Pontianak, Kalimantan Barat (Pemateri).
2007 : Pesky Polychaetes: A Workshop on Marine Polychaete Taxonomy,
Museum of Art and Gallery of The Northern Territory, Darwin,
October,1-5, AUSTRALIA (Peserta)
2005 : Pelatihan Identifikasi Biota Laut (Zooplankton), 5-15 September
2005, P2O-LIPI, Jakarta (Peserta)
2005 : International Field Biology Course IBOY-DIWPA III, Research
Center for Biology, Indonesia Institute of Science, December 12-
21, 2005, Cibinong (Peserta).
Publikasi

17
1. Junardi. 2009. Karakteristik Morfologi dan Habitat Cacing Nipah
Namalycastis rhodochorde (Polychaeta: Nereididae: Namanereididae) di
Kawasan Hutan Mangrove Estuaria Sei Kakap Kalimantan Barat Jurnal Sains
MIPA Vol 93:
2. Junardi dan E.R.P Wardoyo. 2008. Struktur Komunitas dan Karakteristik
Habitat Cacing Laut (Polychaeta) di Perairan Mangrove Peniti Kalimantan
Barat. Biodiversitas Vol 9 (3): 213-216.
3. Glasby, C.J., T. Miura. E. Nishi, and Junardi. 2007. A new species of
Namalycastis (Polychaeta: Nereididae: Namanereidinae) from the shores of
South-east Asia. The Beagle, Records of the Museum and Art Galleries of the
Northern Territory 23:21-27.
4. Junardi dan E.R.P. Wardoyo. 2006 Keanekaragaman Cacing Laut
(Polychaeta) dan Dekomposisi Serasah Hutan Mangrove Peniti Kalimantan
Barat. Pemberdayaan Masyarakat.
5. Junardi dan Nofrita. 2004. Keanekaragaman Makrozoobentos Sebagai
Indikator Kualitas Perairan Sungai Jawi. Suara Almamater. Vol. 19 (2):37-46.
6. Junardi dan S. Murwani. 2004. Keanekaragaman dan Pola Distribusi Genus
Polychaeta di Perairan Pantai Timur Lampung Selatan. Jurnal Sains dan
Teknologi 10 (1): 58-64.
Pontianak, Desember 2009
Junardi, S.Si, M.Si
NIP : 132257938
2. Anggota Peneliti
Nama Lengkap dan gelar : Firman Saputra, S.Si.

18
NIP : 198302112008121003
Jabatan Fungsional :
Jenis Kelamin : Laki-laki
Tempat Tanggal Lahir : Pontianak, 11 Februari 1983
Alamat institusi : Program Studi Biologi FMIPA Untan
Bidang Keahlian : Parasitologi
Pendidikan:
a. Sarjana Biologi, Universitas Gadjah Mada, 2007
Penelitian:
2007 : Pengaruh Ekstrak Metanol Spons (Familia Tetilliade) terhadap
Toxoplasma gondii pada Mencit (Mus musculus)
2005 : Pengaruh Suplemen Spirulina terhadap Kadar Eritrosit Tikus
(Rattus norvegicus) yang Terdedah Radiasi
Pontianak, Desember 2009
Firman Saputra, S.Si.
NIP : 198302112008121003




















