
La caractérisation des dépôts animaux de La Tène ancienne à Geispolsheim « Schwobenfeld »...
44
—25— Épreuves d’auteur La caractérisation des dépôts animaux de La Tène ancienne à Geispolsheim « Schwobenfeld » (Bas-Rhin) Olivier Putelat 1 et Michaël Landolt 2 1 Pôle d’Archéologie Interdépartemental Rhénan. Sélestat & École Doctorale d’Archéologie ED 112 - Université de Paris 1 Panthéon-Sorbonne. Maison de l’Archéologie et de l’Ethnologie - UMR 7041 ArScAn, Équipe Archéologies environnementales. [email protected] 2 Pôle d’Archéologie Interdépartemental Rhénan. Sélestat & Archéologie et Histoire ancienne : Méditerranée-Europe - UMR 7044 Archimède. [email protected] Introduction L’occupation de La Tène ancienne à Geispolsheim « Schwobenfeld » (Bas-Rhin) a fait l’objet de plusieurs diagnostics et fouilles préventives entre 2006 et 2009 (Landolt, Fleischer 2011 ; Landolt et al. 2007 ; 2008 ; 2013). Le site est localisé à une dizaine de kilomètres au sud-est de Strasbourg le long de la rive sud d’un ancien bras de la Bruche en activité durant la Protohistoire (zone 5) (fig. 1). D’un point de vue chronologique, il succède à un habitat, attribué à la transition Hallstatt D3/La Tène A1, situé à Entzheim « In der Klamm » à 600 mètres au nord de l’autre côté de la rivière (zone 1 : 119 structures). Un habitat beaucoup plus restreint de la même période est également à noter à Geispolsheim « Schwobenfeld », en rive sud de l’ancien bras de la Bruche, à une centaine de mètres au nord-est de l’occupation laténienne (zone 4 : 14 structures). En raison de l’importante surface fouillée (environ 20 000 m 2 ), du nombre de structures et de l’abondance du mobilier, l’étude du site laténien de Geispolsheim améliore la connaissance de l’organisation des sites ouverts de La Tène ancienne dans la plaine du Rhin supé- rieur. Régionalement, de par ses 290 structures fouillées, cette occupation se démarque nettement des autres sites de plaine, moins étendus, déjà reconnus pour La Tène ancienne. Signalons, par exemple, que les sites bas-rhinois de Rosheim « Mittelweg » et de Souffelweyersheim « Les Sept Arpents » ne comportent respectivement que 20 et 38 structures (Jeunesse et al. 1993 ; Lefranc et al. 2008). Seul le site récemment fouillé de Colmar « Jardin des Aubépines » (Haut-Rhin) se distingue avec ses 191 struc- tures (Roth-Zehner et al. 2008). À Geispolsheim « Schwobenfeld », 205 silos, 79 fosses et 6 fonds de cabanes excavées ont été attribués à La Tène A2-B1a (fouilles 2006 à 2008). Malgré la relative bonne conservation des structures (notamment des silos parfois très bien conservés jusqu’au goulot) et l’abondance des matériaux de construction en terre crue (environ 250 kg), aucun bâtiment n’a pu être identifié. Contrairement aux 0 300 km 0 50 km N Scheer Ill RHIN RHIN Bruche Meurthe Moder Zorn STRASBOURG SELESTAT MOLSHEIM SAVERNE HAGUENAU WISSEMBOURG Entzheim- Geispolsheim Fig. 1 : Localisation des communes d’Entzheim et de Geispolsheim (Pair).
-
Upload
culturecommunication-fr -
Category
Documents
-
view
1 -
download
0
Transcript of La caractérisation des dépôts animaux de La Tène ancienne à Geispolsheim « Schwobenfeld »...

—25—
Épreuves d’auteur
La caractérisation des dépôts animaux de La Tène ancienne à Geispolsheim « Schwobenfeld » (Bas-Rhin)
Olivier Putelat1 et Michaël Landolt2
1 Pôle d’Archéologie Interdépartemental Rhénan. Sélestat &École Doctorale d’Archéologie ED 112 - Université de Paris 1 Panthéon-Sorbonne.Maison de l’Archéologie et de l’Ethnologie - UMR 7041 ArScAn, Équipe Archéologies [email protected] Pôle d’Archéologie Interdépartemental Rhénan. Sélestat &Archéologie et Histoire ancienne : Méditerranée-Europe - UMR 7044 Archimè[email protected]
IntroductionL’occupation de La Tène ancienne à Geispolsheim
« Schwobenfeld » (Bas-Rhin) a fait l’objet de plusieurs diagnostics et fouilles préventives entre 2006 et 2009 (Landolt, Fleischer 2011 ; Landolt et al. 2007 ; 2008 ; 2013). Le site est localisé à une dizaine de kilomètres au sud-est de Strasbourg le long de la rive sud d’un ancien bras de la Bruche en activité durant la Protohistoire (zone 5) (fig. 1). D’un point de vue chronologique, il succède à un habitat, attribué à la transition Hallstatt D3/La Tène A1, situé à Entzheim « In der Klamm » à 600 mètres au nord de l’autre côté de la rivière (zone 1 : 119 structures). Un habitat beaucoup plus restreint de la même période est également à noter à Geispolsheim « Schwobenfeld », en rive sud de l’ancien bras de la Bruche, à une centaine de mètres au nord-est de l’occupation laténienne (zone 4 : 14 structures).
En raison de l’importante surface fouillée (environ 20 000 m2), du nombre de structures et de l’abondance du mobilier, l’étude du site laténien de Geispolsheim améliore la connaissance de l’organisation des sites ouverts de La Tène ancienne dans la plaine du Rhin supé-rieur. Régionalement, de par ses 290 structures fouillées, cette occupation se démarque nettement des autres sites de plaine, moins étendus, déjà reconnus pour La Tène ancienne. Signalons, par exemple, que les sites bas-rhinois de Rosheim « Mittelweg » et de Souffelweyersheim « Les Sept Arpents » ne comportent respectivement que 20 et 38 structures (Jeunesse et al. 1993 ; Lefranc et al. 2008). Seul le site récemment fouillé de Colmar « Jardin des Aubépines » (Haut-Rhin) se distingue avec ses 191 struc-tures (Roth-Zehner et al. 2008).
À Geispolsheim « Schwobenfeld », 205 silos, 79 fosses et 6 fonds de cabanes excavées ont été attribués à La Tène A2-B1a (fouilles 2006 à 2008). Malgré la relative bonne conservation des structures (notamment des silos parfois très bien conservés jusqu’au goulot) et l’abondance des matériaux de construction en terre crue (environ 250 kg), aucun bâtiment n’a pu être identifié. Contrairement aux
0 300 km
0 50 km
N
Sche
er
Ill
RHIN
RH
IN
Bruche
Meurthe
Moder
Zorn
STRASBOURG
SELESTAT
MOLSHEIM
SAVERNE
HAGUENAU
WISSEMBOURG
Entzheim-Geispolsheim
Fig. 1 : Localisation des communes d’Entzheim et de Geispolsheim (Pair).

Les dépôts d’ossements d’animaux en France, de la fouille à l’interprétation
—26—
Épreuves d’auteur
silos et aux fosses qui se répartissent sur toute la zone fouillée, les fonds de cabanes excavées se concentrent dans la partie sud de la fouille (fig. 2). De plus, la réparti-tion spatiale des rejets montre que le cœur du site se trouve au sud du décapage et doit se prolonger au-delà. Les struc-tures d’habitat pourraient donc se trouver immédiatement au sud des surfaces décapées.
L’important corpus céramique (2833 NMI), associé à des objets métalliques datés, apporte un nouveau regard sur les ensembles du début de La Tène, situés au sud de la plaine du Rhin supérieur. La détermination de fibules attri-buées à La Tène A2-B1 a permis d’identifier cette phase chronologique avec certitude et d’établir une première ébauche typo-chronologique (Landolt et al. 2012). La céramique tournée permet d’aborder les problématiques de son origine et de sa diffusion. Enfin, certains récipients en céramique modelée témoignent d’échanges à plus ou moins grande distance : une situle en pâte graphitée est d’origine orientale (sud-ouest de l’Allemagne, Bohême ou Autriche) ; une coupe à profil sinueux ornée de barbotine est très probablement de provenance occidentale (culture Aisne-Marne).
Les activités domestiques et artisanales sont nombreu-
ses et diversifiées. Le filage et le tissage sont bien attestés. Les meules et molettes en rhyolithe nous renseignent sur les circuits d’approvisionnements en matériel de mouture à moyenne distance (La Salle, Vosges). La découverte de plusieurs fragments de barres en céramique, identiques à celles du briquetage, est à mettre en relation avec la production de sel à l’âge du Fer dans la région de Marsal (Moselle). Les traces d’activités métallurgiques sont nombreuses : forge (culots, battitures, fibule en cours de fabrication), artisanat du bronze (creusets et lingotières) et orfèvrerie (lingotière). Ces indices sont généralement retrouvés en association ce qui permet de confirmer que ces différentes activités étaient encore étroitement liées à cette période. Généralement mises au jour dans ou à proximité des cabanes excavées, ces découvertes viennent confirmer l’hypothèse de la dévolution de ces structures aux pratiques artisanales. Parmi les vestiges d’activités métallurgiques, une fibule en fer appartenant à un type hybride apparenté à la famille des Certosa a pu être recon-nue. L’objet, en cours de fabrication, apporte de nouveaux éléments à l’étude des phénomènes d’hybridation des fibules, mis en évidence au nord des Alpes, depuis le Hallstatt final et pendant La Tène ancienne, et résultant de contacts entre le sud des Alpes et l’Europe moyenne. Les rares objets en os ne permettent pas de conclure quant au travail des matières dures animales sur le site.
La flore, révélée par les analyses carpologiques, est composée de plantes ancrées dans les traditions de l’âge du Bronze et du Premier âge du Fer. Les blés, l’orge et le millet constituent la base de l’alimentation végétale. Hormis l’absence de l’épeautre, cette flore s’insère bien dans les données interrégionales. Aucune nouvelle plante n’a été mise en évidence dans les assemblages botaniques. De par ses 205 silos, le site de Geispolsheim apparaît indiscutablement comme étant un site de production agricole. Il s’agit en effet d’une aire d’ensilage intensive, à proximité de laquelle diverses activités artisanales et domestiques étaient pratiquées.
Entre le Hallstatt final et La Tène ancienne, l’existence de ces grandes aires d’ensilages, parfois destinées à appro-visionner en nourriture d’autres sites, a pu être mise en évidence ailleurs. C’est par exemple le cas à Soupir « Le Champ Grand Jacques » (Aisne) (Gransar et al. 2002) ou à Gondreville/Fontenoy-sur-Moselle (Meurthe-et-Moselle) (Deffressigne et al. 2002). Une des caractéristiques du site de Geispolsheim est d’avoir livré de très nombreux rejets domestiques, ce qui le distingue d’autres sites d’en-silages intensifs généralement très pauvres en mobilier. Témoignent-ils de la très grande proximité de l’habitat ?
À Geispolsheim « Schwobenfeld », plusieurs types de dépôts humains, animaux, et probablement métalliques, forment un ensemble dont la signification nous échappe encore. Ils s’inscrivent dans un phénomène bien docu-menté en Alsace et dans une partie de l’Europe, principa-lement entre le Ve et le IVe s. avant J.-C. (Landolt et al. 2011, p. 220-226). Six inhumations ont été mises au jour dans des structures d’ensilage. Elles se concentrent dans deux parties du site, en limite sud de la fouille et dans
1079
1204
Limites communales 100 m0
N
Occupation de La Tène A2-B1a
Occupations du Hallstatt D3/La Tène A1
Zone 1
Zone 6Zone 7
Zone 4
Zone 8-10
Lingolsheim
Entzheim « In der Klamm »
Geispolsheim « Schwobenfeld »
Zone 9
Zone 11
Zone 5
Chenal de la Bruche(lit majeur)
Fig. 2 : Plan général des occupations du Hallstatt D3/La Tène A1 et de La Tène A2-B1a d’Entzheim « In der Klamm » et de Geispolsheim « Schwobenfeld » (M. Landolt).

O. Putelat et M. Landolt, La caractérisation des dépôts animaux de La Tène ancienne à Geispolsheim
—27—
Épreuves d’auteur
la moitié nord de la fouille. Les dépôts humains en silo sont exclusivement individuels et souvent primaires. La position et l’orientation des corps sont variées. Le recrute-ment des sujets n’est pas lié au sexe ni à l’âge des défunts puisque l’échantillon rassemble des sujets des deux sexes appartenant à toutes les classes d’âges (enfant, adulte jeune, mature et âgé).
Plusieurs objets métalliques complets pourraient eux aussi avoir fait l’objet de dépôts. On notera, par exemple, un couteau en fer dans la même structure qu’un squelette de lièvre et un fer de lance plié volontairement dans une autre.
Les dépôts animaux, qui font l’objet de cette étude, sont situés à proximité des inhumations humaines. Ils concernent 27 structures et mettent en jeu une quarantaine d’individus, appartenant à différents taxons, domestiques pour la plupart. Ces dépôts concernent aussi bien des squelettes que des parties de squelette, en connexion, manipulés, voire partiellement consommés. Leur présen-tation détaillée est précédée d’un rapide aperçu de l’apport de l’archéozoologie à la connaissance de la part carnée de l’alimentation des habitants du site.
1. Le corpus faunique et les rejets alimentaires
Pour les périodes protohistoriques et historiques, les fouilles de 2008 à Geispolsheim « Schwobenfeld » ont livré environ 17000 restes animaux (microfaune non comprise), pour une masse totale d’environ 208 kg. Leur répartition chronologique est étagée du Hallstatt C à l’épo-que contemporaine. Ils viennent s’ajouter aux 6161 restes protohistoriques et historiques issus de la fouille de 2006 à Geispolsheim « Schwobenfeld » et Entzheim « In der Klamm », intégralement étudiés par O. Putelat (Landolt et al. 2007).
Pour La Tène A2-B1a, la fouille 2008 à Geispolsheim « Schwobenfeld » a livré 12200 restes (pour une masse totale de 152 kg), mis au jour dans 207 structures, dont 153 silos. N’ayant pas eu la possibilité matérielle d’étu-dier l’ensemble de ce matériel, cependant intégralement examiné, nous en avons privilégié l’étude des dépôts animaux, qui constituent à notre sens la spécificité archéo-zoologique du site.
Les dépôts animaux (squelettes, parties de squelettes, assemblages fauniques atypiques) concernent au total 27 structures, dont 25 silos1. Ces dépôts sont souvent mêlés à des déchets. Un peu plus de 2000 restes ainsi étudiés sont des rejets alimentaires usuels qui constituent un échantillon quantitativement significatif mais biaisé. En effet, cet échantillon est issu de structures sélectionnées en
1 Soit 4 silos fouillés en 2006, 21 silos et 2 structures en 2008, soit 11 % des structures et 15 % des silos mis au jour. Les 4790 restes de macrofaune livrés par les 23 structures de 2008 ont été intégralement étudiés (soit environ 39 % du corpus osseux laténien de cette fouille).
fonction de leurs spécificités (dépôts animaux et assem-blages atypiques), et non d’un choix aléatoire. Ces déchets osseux permettent cependant une première approche des pratiques alimentaires à Geispolsheim « Schwobenfeld » (fouilles 2008) pendant La Tène A2-B1a2.
Les ossements qui composent les rejets sont dans l’ensemble bien conservés. Leurs surfaces sont lisses, la masse moyenne satisfaisante (8,9 g) et le nombre de restes non strictement dentaires est très élevé (98 %). Le taux d’ossements brûlés (carbonisés et calcinés en proportions sensiblement comparables) est relativement important (27 % du NR), mais est extrêmement variable d’une structure à l’autre. Le spectre de faune laténien de Geispolsheim « Schwobenfeld » (fouilles 2008), majori-tairement domestique, est présenté ci-après (tab. 1). Il est
2 Cette approche est complétée par quelques données issues du pré-examen des 6000 restes laténiens non étudiés à ce jour (fouilles 2008), par l’étude de la faune mise au jour en 2006, ainsi que par les connaissances acquises pour le Hallstatt D3/La Tène A1 sur le site voisin d’Entzheim « In der Klamm ».
Espèces N.R. % N R NMIc Masse (g)
% masse
Bœuf : Bos taurus L. 189 14,6% 3 5993,0 32,3%Mouton : Ovis aries L. 125 6Chèvre : Capra hircus L. 10 1Caprinés 671 7Porc : Sus domesticus E. et suinés 126 9,8% 5 1146,9 6,2%Equidés : Equus sp 67 5,2% 2 3164,3 17,1%Chien : Canis familiaris L. 30 2,3% 3 365,6 2,0%Poule : Gallus domesticus 20 1,5% 2 11,3 0,1%Total des animaux domestiques 1238 95,9% 29 16135,1 87,0%Lièvre : Lepus capensis P. 1 0,1% 1 1,4 0,0%Renard : Vulpes vulpes L. 1 0,1% 1 4,0 0,0%Loutre d'Europe : Lutra lutra L. 1 0,1% 1 1,0 0,0%Aurochs-Bison 4 0,3% 1 231,6 1,2%Sanglier : Sus scrofa L. 4 0,3% 1 152,1 0,8%Cerf élaphe : Cervus elaphus L. 4 0,3% 1 386,1 2,1%Chevreuil : Capreolus capreolus L. 2 0,2% 1 19,2 0,1%Canard : Anas sp 1 0,1% 1 0,5 0,0%Bécasse des bois : Scolopax rusticola 2 0,2% 1 1,2 0,0%Total des animaux sauvages 20 1,5% 9 797,1 4,3%Oie : Anser sp 1 0,1% 1 2,0 0,0%Oiseaux non différenciés 1 0,1% 1,2 0,0%Brochet: Esox lucius 4 0,3% 1 1,0 0,0%Cyprinidés 5 0,4% 1 0,6 0,0%Poisson indéterminé 22 1,7% 0 0,1 0,0%Autres groupes 33 2,6% 3 4,9 0,0%Nombre de restes déterminés 1291 100,0% 41 16937,1 91,3%Grands mammifères 11 675,3 3,6%Mammifères de taille moyenne 574 576,1 3,1%Esquilles indéterminées 213 353,2 1,9%Total 2089 41 18541,7 100,0%Bois de cervidés : NR / PR 1 18,0Grand hamster : Cricetus cricetus L. 7 8,6Micromammifères 57 19,0Crapaud : Bufo sp 8 2,4Grenouille : Rana sp 4 0,2Amphibiens indéterminés 190 15,1Cyprinidés (écailles) 1257 45,0Escargots : Cepaea sp 2 1,9Moules d'eau douce: unionidés 2 10,2
29,4%62,4% 5454,0
Tab. 1 : Geispolsheim « Schwobenfeld » (fouilles 2008). Spectre de faune du matériel osseux laténien étudié hors dépôts (O. Putelat).

Les dépôts d’ossements d’animaux en France, de la fouille à l’interprétation
—28—
Épreuves d’auteur
complété par quelques observations relevées lors du pré-examen des ossements non étudiés (tab. 2).
Les caprinés, mouton et chèvre, prévalent en nombres de restes et d’individus. Ils égalent presque le bœuf en masse des restes, plus ou moins corrélée à la masse de viande, et participent donc de façon très conséquente à l’apport carné.
Toutefois, le matériel osseux des fouilles de 2008 dif-fère sur ce point par rapport aux fouilles de 2006 (fig. 3 et tab. 3). Certes, d’une manière générale pour l’Alsace, la comparaison des proportions de restes de caprinés vis-à-vis du nombre de restes des principales espèces mammaliennes domestiques (bœuf, caprinés, porc, équi-dés, chien) montre leur assez forte représentation à La Tène ancienne : 32,4 % à Geispolsheim « Schwobenfeld » (fouilles 2006), 42,6 % à Rosheim « Mittelweg » et « Sandgrube » (Méniel 1993), 51,7 % à Mittelhausen « Liesbuhl Gimbretter Weg » (Broc 2011). Il n’en reste pas moins que, pour ce critère, les données de Geispolsheim
« Schwobenfeld » (fouilles 2008), où les caprinés repré-sentent 71,9 % des espèces domestiques, sont très élevées par rapport aux autres données alsaciennes. Il en est de même lorsque l’on compare cette proportion aux travaux de synthèse pour La Tène ancienne dans la moitié nord de la Gaule (Méniel et al. 2009). Une telle surreprésentation des caprinés à Geispolsheim « Schwobenfeld » (fouilles 2008) paraît donc difficile à valider en l’état actuel de la recherche. Notre sentiment est que ce taux très élevé est surévalué, résultant probablement d’un biais, lié à au fait que l’étude archéozoologique n’a concerné que les silos livrant des dépôts animaux. En effet, les caprinés étant très bien représentés parmi ces dépôts, leur surreprésen-tation dans les déchets osseux peut être due à un « effet de traîne » des dépôts animaux et assemblages fauniques atypiques (cf § 2). Ce point serait sans doute relativisé par l’étude exhaustive de la faune laténienne de Geispolsheim « Schwobenfeld » (fouilles 2008).
À Geispolsheim « Schwobenfeld » (fouilles 2006 et 2008), la proportion de faune sauvage est faible, mais la ressource cynégétique est diversifiée, mêlant très grands et petits gibiers (tab. 1, 2, 3). Quelques espèces sont carac-téristiques de la proximité de milieux humides : la loutre, le castor, le héron, le canard, la grenouille, le brochet, un cyprinidé (au moins), des mollusques dulçaquicoles. Le sanglier, le cerf, la bécasse des bois sont plutôt à mettre en rapport avec la proximité de forêts. La présence du lapin et du hamster est sans doute le fait d’une intrusion ulté-rieure de ces fouisseurs. En comparant les données sur un plan diachronique (fig. 4), les résultats de Geispolsheim « Schwobenfeld » confirment les observations réalisées pour le tout début de La Tène ancienne à Entzheim « In der Klamm ». Sur ce territoire, la pratique cynégétique régresse considérablement entre le Hallstatt D3, Hallstatt D3/La Tène A1 et La Tène A2 (gibier en noir sur la fig. 4). Cette régression, qui ne peut être fortuite et dont les
Ossements échantillonnés (167 structures) NRHumain 4Lapin de garenne : Oryctolagus cuniculus L. 7Grand hamster : Cricetus cricetus L. 3Castor : Castor fiber L. 2Loup : Canis lupus L. 1Chat forestier : Felis sylvestris Schreber 1Aurochs-Bison 6Sanglier : Sus scrofa L. 5Cerf élaphe : Cervus elaphus L. 8Héron cendré : Ardea cinerea 1Canard colvert : Anas platyrhynchos 1Pigeon ramier : Columba palumbus 1Poisson indéterminé 1Poisson indéterminé (écailles) <10Escargots : Cepaea sp 25Escargots : Helix pomatia 1Moules d'eau douce : unionidés 17
Tab. 2 : Geispolsheim « Schwobenfeld » (fouilles 2008). Taxons supplémentaires mis en évidence dans le matériel osseux laténien pré-examiné (non étudié) (O. Putelat).
Bœuf
Caprinés
Suinés
0
20
40
60
80
100
Entz. Ha D3 (184)
Entz. Ha D3-LT A
(330)
Geisp. 2006 LT A-B
(879)
Geisp. 2008 LT A-B (1121)
% du nombre de restes
Fig. 3 : Diagramme ternaire diachronique de répartition des taxons de la triade domestique, exprimés en nombre de restes pour les sites d’Entzheim (Entz.) « In der Klamm » (Hallstatt D3/La Tène A1) et de Geispolsheim (Geisp.) « Schwobenfeld » (La Tène A2-B1a) (fouilles 2006 et 2008). Le NR de la triade figure entre parenthèses (O. Putelat).
Espèces N.R. % NR NMIc Masse (g) % masse Bœuf : Bos taurus L. 313 31,6% 5 12470,1 45,8%Mouton : Ovis aries L. 17 5Chèvre : Capra hircus L. 7 2Caprinés 265 3Porc : Sus domesticus E. et suinés 277 28,0% 16 4439,7 16,3%Equidé: Equus sp 37 3,7% 4 4458,4 16,4%Chien : Canis familiaris L. 60 6,1% 5 746,6 2,7%Poule : Gallus domesticus 5 0,5% 1 3,4 0,0%Total des animaux domestiques 981 99,2% 41 24734,4 90,8%Chevreuil 1 0,1% 1 3,9 0,0%Sanglier cf 1 0,1% 1 74,7 0,3%Lièvre 5 0,5% 1 10,7 0,0%Castor 1 0,1% 1 4,7 0,0%Total des animaux sauvages 8 0,8% 4 94 0,3%Nombre de restes déterminés 989 100,0% 45 24828,4 91,2%Grands mammifères 90 711,6 2,6%Mammifères de taille moyenne 424 1094,5 4,0%Esquilles indéterminées 174 594,7 2,2%Total 1677 45 27229,2 100,0%Bois de cervidés : NR / MR 2 61,0Humain 1 3,7Grand hamster : Cricetus cricetus L. 2 0,7Micromammifères 3 1,0Bufo sp. 11 0,9Rana sp, 2 0,1Amphibiens indéterminés 1 0,0
29,2% 2616,2 9,6%
Tab. 3 : Spectre de la faune laténienne de Geispolsheim « Schwobenfeld », fouille 2006, hors dépôts (O. Putelat).
O. Putelat et M. Landolt, La caractérisation des dépôts animaux de La Tène ancienne à Geispolsheim
—29—
Épreuves d’auteur
causes restent à comprendre, concerne principalement la chasse au cerf élaphe, qui représente 11,9 % des restes déterminés au Hallstatt D3 et 11,5 % au Hallstatt D3/La Tène A1, mais qui ne représente plus que 0,3 % des restes déterminés à Geispolsheim « Schwobenfeld » à La Tène A2-B1a (fouilles 2008) (tab. 1) et n’apparaît pas dans la faune mise au jour en 2006 (tab. 3).
2. Les dépôts animaux
2.1. Matériel et méthodeLa faune laténienne de Geispolsheim « Schwobenfeld »
comprend, outre les déchets alimentaires présentés précé-demment, divers ensembles anatomiques composés de squelettes complets ou incomplets, de segments anatomi-ques connectés, de têtes osseuses, d’assemblages atypi-ques, mis au jour dans des fosses et silos. Ces découvertes, livrées en 2008 par 23 structures différentes, s’inscrivent dans la continuité des fouilles de ce site en 2006, où quatre structures avaient livré en tout quatre squelettes de chien et deux de lièvre, attribués eux aussi à la même période (Landolt et al. 2007). Les dépôts animaux des fouilles de 2006 et de 2008 sont réunis ici (tab. 4). Les espèces impli-quées sont majoritairement domestiques : bœuf, caprinés (mouton et/ou chèvre), porc, cheval, chien, auxquelles s’ajoutent une espèce mammalienne sauvage, le lièvre, et éventuellement des poissons.
Grâce à l’expérience acquise lors des fouilles de 2006, il a été porté une attention particulière au prélèvement de ces différents ensembles. À chaque découverte d’un sque-lette, ou d’un assemblage osseux évoquant la présence d’un squelette, le décapage mécanique était interrompu, les os éventuellement endommagés lors du décapage étant récoltés et conditionnés de manière séparée. Une fouille fine s’ensuivait, complétée par une documentation photographique de qualité (vues d’ensemble et détail des
connexions anatomiques). La majorité des prélèvements a été effectuée par l’archéozoologue3, le plus souvent assisté par un archéologue4.
Un des buts de cette recherche, initiée dès la fouille de 2006 (Landolt et al. 2011), est d’affiner les modalités de mise en place de ces divers assemblages fauniques. Les squelettes sont-ils complets ? Quel fut leur mode d’en-fouissement ? Distingue-t-on des remaniements squeletti-ques post-mortem et de quelle nature sont-ils ? Certaines espèces, certaines attributions sexuelles et classes d’âges sont-elles plus fréquentes que d’autres ? Quelles signifi-cations pouvons-nous attribuer à ces dépôts, à la présence de tombes animales multiples ? S’agit-il d’une simple élimination de cadavres ou résultent-ils de pratiques com-
3 Olivier Putelat.
4 L’étude des ossements en laboratoire a été confiée à Ludivine Paleau, dans le cadre d’une thèse réalisée à l’Université de Strasbourg sous la tutelle de Rose-Marie Arbogast et Patrice Méniel : « aspects cultuels et économiques des animaux à l’Âge du Fer en Alsace ». Ce travail de thèse est définitivement interrompu depuis le mois d’avril 2011. Cette étude a fait l’objet d’un rapport inédit (Paleau 2011), normalisé par Olivier Putelat et repris de façon collégiale dans le rapport de fouilles de Geispolsheim « Schwobenfeld » (Landolt et al. 2013).
0
20
40
60
80
100
Entz. Ha D3 (233)
Entz. Ha D3-LT A
(414)
Geisp. 2006 LT A-B (989)
Geisp. 2008 LT A-B
(1258)
Triade Divers domestiques Sauvage% Nombre de restes
Fig. 4 : Diagramme diachronique de répartition des grandes catégories de restes déterminés (triade domestique, divers domestiques, sauvage), exprimées en nombre de restes pour les sites d’Entzheim (Entz.) « In der Klamm » et de Geispolsheim (Geisp.) « Schwobenfeld ». Le NRD figure entre parenthèses (O. Putelat).
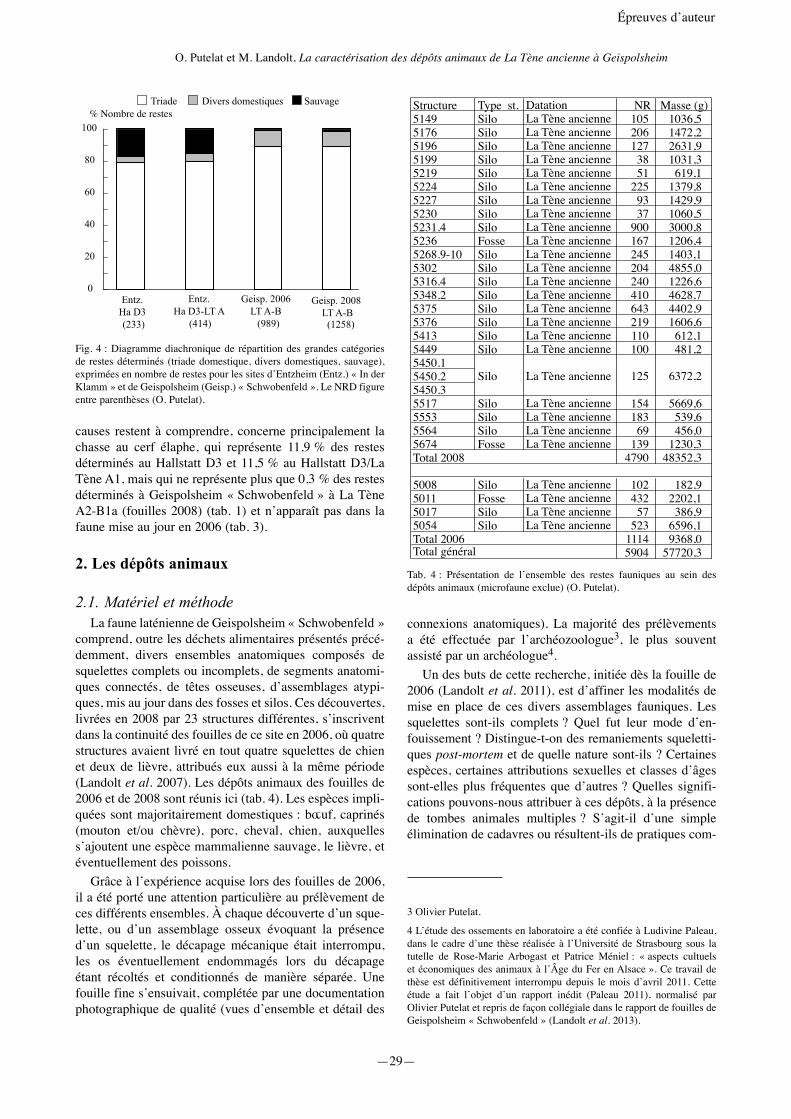
Structure Type st. Datation NR Masse (g)5149 Silo La Tène ancienne 105 1036,55176 Silo La Tène ancienne 206 1472,25196 Silo La Tène ancienne 127 2631,95199 Silo La Tène ancienne 38 1031,35219 Silo La Tène ancienne 51 619,15224 Silo La Tène ancienne 225 1379,85227 Silo La Tène ancienne 93 1429,95230 Silo La Tène ancienne 37 1060,55231.4 Silo La Tène ancienne 900 3000,85236 Fosse La Tène ancienne 167 1206,45268.9-10 Silo La Tène ancienne 245 1403,15302 Silo La Tène ancienne 204 4855,05316.4 Silo La Tène ancienne 240 1226,65348.2 Silo La Tène ancienne 410 4628,75375 Silo La Tène ancienne 643 4402,95376 Silo La Tène ancienne 219 1606,65413 Silo La Tène ancienne 110 612,15449 Silo La Tène ancienne 100 481,25450.15450.25450.35517 Silo La Tène ancienne 154 5669,65553 Silo La Tène ancienne 183 539,65564 Silo La Tène ancienne 69 456,05674 Fosse La Tène ancienne 139 1230,3Total 2008 4790 48352,3
5008 Silo La Tène ancienne 102 182,95011 Fosse La Tène ancienne 432 2202,15017 Silo La Tène ancienne 57 386,95054 Silo La Tène ancienne 523 6596,1Total 2006 1114 9368,0Total général 5904 57720,3
Silo La Tène ancienne 125 6372,2
Tab. 4 : Présentation de l’ensemble des restes fauniques au sein des dépôts animaux (microfaune exclue) (O. Putelat).

Les dépôts d’ossements d’animaux en France, de la fouille à l’interprétation
—30—
Épreuves d’auteur
plexes, dont la compréhension nous échappe en partie ? En présence d’un ensemble anatomique connecté, sur quel argumentaire peut-on se fonder pour différencier ce qui résulterait d’une certaine attention portée au dépôt, de ce qui résulterait du rejet de déchets ? Existe-t-il des parallèles à ces pratiques à La Tène ancienne, en Alsace, ou ailleurs ?
Le champ de la recherche est vaste et les interpréta-tions données aux découvertes ne sont pas exemptes de subjectivité. Ainsi, notre vision actuelle des assemblages osseux connectés porte sur des vestiges osseux nettoyés, débarrassés de leurs enveloppes de peaux, de chairs et de tendons. Ces vestiges fauniques assemblés nous apparais-sent sous forme de « tableaux ». Pour chaque occurrence on a ainsi pu évoquer de manière débonnaire au sein de l’équipe de fouilles le « cheval baleine », le « mouton papillon ou araignée », les « trois petits cochons ». Il va de soi que cette vision des choses ne peut en aucun cas être appliquée à celle des Laténiens de Geispolsheim, puisque la mise en place, puis la vision des restes animaux, concer-nait non pas des squelettes, ou des parties de squelettes, mais bien des cadavres, des carcasses, ou des parties de carcasses, dans des états de conservation variables, encore garnis des matières périssables.
L’étude de ces ensembles particuliers nécessite donc une présentation claire des dépôts, préalablement à toute tentative d’interprétation. Pour cela, nous les avons clas-sés selon leur composition anatomique, puis taxonomique. Nous évoquons tout d’abord ici les dépôts de têtes osseu-ses, les dépôts d’ensembles anatomiques issus du squelette appendiculaires, puis les dépôts de squelettes, complets ou presque complets. Par ailleurs, lors du pré examen de la faune laténienne, certains faits archéologiques nous ont paru livrer des rejets alimentaires atypiques, car volumi-neux, parfois monospécifiques, souvent carbonisés pour tout ou partie. Au vu de ces caractéristiques, il nous a paru nécessaire de les dissocier de l’étude des rejets domesti-ques et de les présenter aussi dans le cadre des dépôts. Nous les avons regroupés sous le terme de restes de « repas collectifs ». Après la présentation des résultats par type de dépôt et par structure, ceux-ci sont ensuite discutés à l’échelon du site, puis mis succinctement en perspective avec un état de la question régional.
2.2 Les résultats
2.2.1. Les têtes osseuses
Le silo bitronconique 5149Le silo 5149 livre 105 restes dont 31 déterminés, pour
une masse totale d’environ 1 kg. Il s’agit d’au moins 90 déchets alimentaires auxquels s’ajoutent deux têtes bovine et équine, et un assemblage d’ossements canins.
Une tête de bœuf fractionnée est partiellement repré-sentée par sa partie basio-occipitale et par la moitié droite du crâne et de la face. Une tête d’équidé très fragmentée
est représentée par les principaux fragments hétérolaté-raux de la région du crâne et de la face. L’absence totale de dents jugales pour ces deux taxons semble indiquer une exposition prolongée à l’air libre avant enfouisse-ment. Un squelette de chien (âgé de plus de 18 mois), aux connexions non documentées, est mis en évidence par la moitié gauche de sa tête, deux vertèbres tranchées cauda-lement, un os coxal montrant des traces de désossement, un radius et une ulna, un os du carpe et un métacarpe, une fibula.
Ces deux têtes de bœuf et d’équidé semblent avoir séjourné à l’air libre. Les restes canins portent des traces de découpe. En l’absence d’attribution stratigraphique pour ces vestiges et ne pouvant les différencier de rejets de boucherie, nous prenons le parti d’interpréter ces restes comme étant des déchets alimentaires.
Le silo tronconique 5196Le silo 5196 livre 127 restes de macrofaune, dont 76
déterminés, pour une masse totale d’environ 2,6 kg. Il s’agit de 126 ossements correspondant à des déchets ali-mentaires, auxquels s’ajoute la tête de porc sur laquelle nous nous interrogeons (fig. 5). Cette tête provient de la dernière US de comblement du silo. Elle est presque complète, en connexion étroite avec ses deux mandibules. L’écaille occipitale est enfoncée, suite à l’assommage de ce porc, âgé de 8-10 mois. Les os hyoïdes manquent, non conservés ou arrachés avec la langue.
Les déchets de ce silo sont caractérisés par une bonne diversité taxonomique et une surreprésentation de la région de la tête. Le bœuf est représenté par deux man-dibules hétérolatérales et une côte. Les caprinés sont déterminés au vu de trois restes crâniens, quatre mandibu-les correspondant à quatre individus différents et par six restes du squelette appendiculaire. La surreprésentation pondérale de la tête par rapport à des squelettes de réfé-
Fig. 5 : Tête de porc dans l’US 2 du silo 5196 (M. Landolt).
O. Putelat et M. Landolt, La caractérisation des dépôts animaux de La Tène ancienne à Geispolsheim
—31—
Épreuves d’auteur
rence, non figurée ici, et le NMI élevé indiqué par les res-tes dentaires, évoquent la présence de rejets primaires de boucherie. Au porc sont attribués six autres fragments de tête(s) osseuse(s) qui montrent la présence d’un individu surnuméraire par rapport à la tête déposée. La région de la tête est également surreprésentée parmi la faune équine (41 fragments). Trente deux de ces restes ont subi une ustion à des degrés variables : carbonisés à calcinés. Le chien est décelé au vu d’une dent, d’un métacarpe, d’une côte. Le cerf est décelé par un bois de massacre : la moitié du frontal et de l’occipital droits sont présents, le merrain est brisé (et manque) immédiatement au dessus de l’an-douiller basilaire. La bécasse des bois est déterminée par un humérus et un carpométacarpe.
Le fait que les vestiges fauniques de ce silo corres-pondent majoritairement à la région de la tête nous fait envisager un comblement constitué de rejets primaires de boucherie. Remarquons d’ailleurs que la tête de cerf est privée de ses bois, ce qui accrédite l’idée d’un déchet alimentaire, plutôt que du dépôt d’un crâne de cerf. Dans ce contexte, la présence d’une tête de porc complète se banalise, après remise en contexte au sein des rejets de boucherie (dix-huit restes dans l’US 2). Il n’est donc pas possible de prouver que cette tête ne provient pas d’une activité culinaire. Pour cette raison, nous la considérons par défaut comme un déchet alimentaire.
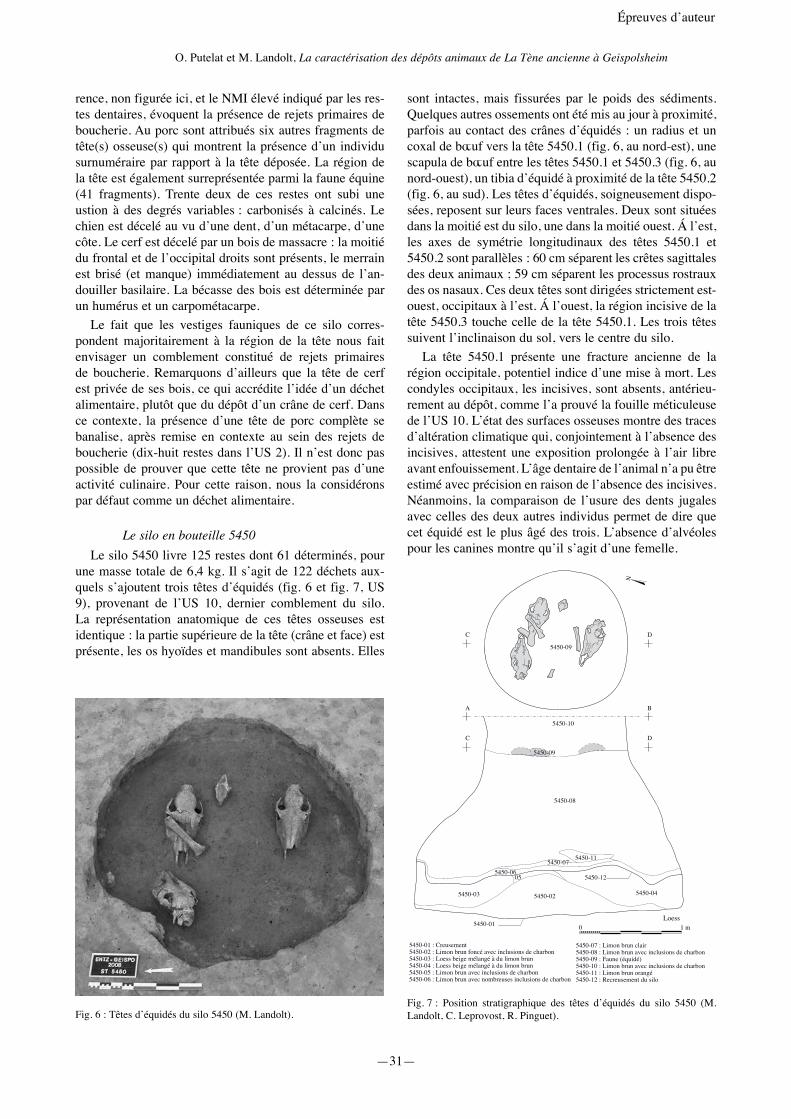
Le silo en bouteille 5450Le silo 5450 livre 125 restes dont 61 déterminés, pour
une masse totale de 6,4 kg. Il s’agit de 122 déchets aux-quels s’ajoutent trois têtes d’équidés (fig. 6 et fig. 7, US 9), provenant de l’US 10, dernier comblement du silo. La représentation anatomique de ces têtes osseuses est identique : la partie supérieure de la tête (crâne et face) est présente, les os hyoïdes et mandibules sont absents. Elles
sont intactes, mais fissurées par le poids des sédiments. Quelques autres ossements ont été mis au jour à proximité, parfois au contact des crânes d’équidés : un radius et un coxal de bœuf vers la tête 5450.1 (fig. 6, au nord-est), une scapula de bœuf entre les têtes 5450.1 et 5450.3 (fig. 6, au nord-ouest), un tibia d’équidé à proximité de la tête 5450.2 (fig. 6, au sud). Les têtes d’équidés, soigneusement dispo-sées, reposent sur leurs faces ventrales. Deux sont situées dans la moitié est du silo, une dans la moitié ouest. Á l’est, les axes de symétrie longitudinaux des têtes 5450.1 et 5450.2 sont parallèles : 60 cm séparent les crêtes sagittales des deux animaux ; 59 cm séparent les processus rostraux des os nasaux. Ces deux têtes sont dirigées strictement est-ouest, occipitaux à l’est. Á l’ouest, la région incisive de la tête 5450.3 touche celle de la tête 5450.1. Les trois têtes suivent l’inclinaison du sol, vers le centre du silo.
La tête 5450.1 présente une fracture ancienne de la région occipitale, potentiel indice d’une mise à mort. Les condyles occipitaux, les incisives, sont absents, antérieu-rement au dépôt, comme l’a prouvé la fouille méticuleuse de l’US 10. L’état des surfaces osseuses montre des traces d’altération climatique qui, conjointement à l’absence des incisives, attestent une exposition prolongée à l’air libre avant enfouissement. L’âge dentaire de l’animal n’a pu être estimé avec précision en raison de l’absence des incisives. Néanmoins, la comparaison de l’usure des dents jugales avec celles des deux autres individus permet de dire que cet équidé est le plus âgé des trois. L’absence d’alvéoles pour les canines montre qu’il s’agit d’une femelle.
Fig. 6 : Têtes d’équidés du silo 5450 (M. Landolt).
C D
A B
5450-01
C D
5450-01 : Creusement5450-02 : Limon brun foncé avec inclusions de charbon5450-03 : Loess beige mélangé à du limon brun5450-04 : Loess beige mélangé à du limon brun5450-05 : Limon brun avec inclusions de charbon5450-06 : Limon brun avec nombreuses inclusions de charbon
5450-07 : Limon brun clair5450-08 : Limon brun avec inclusions de charbon5450-09 : Faune (équidé) 5450-10 : Limon brun avec inclusions de charbon5450-11 : Limon brun orangé5450-12 : Recreusement du silo
Loess
5450-10
5450-08
5450-09
5450-115450-07
05
5450-02 5450-045450-03
5450-065450-12
5450-09
N
0 1 m
Fig. 7 : Position stratigraphique des têtes d’équidés du silo 5450 (M. Landolt, C. Leprovost, R. Pinguet).

Les dépôts d’ossements d’animaux en France, de la fouille à l’interprétation
—32—
Épreuves d’auteur
La tête 5450.2 est complète. L’état des surfaces osseu-ses montre un enfouissement rapide. L’âge dentaire de l’animal a été estimé par l’examen des incisives à environ 12 à 15 ans. L’absence d’alvéoles canines montre qu’il s’agit d’une femelle.
La tête 5450.3 est complète. L’état des surfaces osseu-ses montre un enfouissement lent. L’âge dentaire de l’ani-mal a été estimé par l’examen des incisives à environ 6 ans. La présence de canines bien développées montre qu’il s’agit très probablement d’un mâle.
En résumé, le dernier comblement du silo 5450 a livré trois têtes d’équidés adultes, un mâle et deux femelles, déposées et agencées avec soin. Deux de ces têtes ont séjourné un certain temps à l’air libre avant leur enfouis-sement. La disposition des têtes et le soin apporté au dépôt ne nous invitent pas à considérer cet assemblage osseux comme un rejet de détritus.
2.2.2 Les ensembles anatomiques issus du squelette appendiculaire
2.2.2.1 Le bœuf
Le silo tronconique 5227Le silo 5227 livre 93 restes dont 76 déterminés, pour
une masse totale d’environ 1,4 kg. Il s’agit pour moitié de nombreux déchets, mais aussi de trois ensembles anatomi-ques bovins composés d’un autopode antérieur et de deux autopodes postérieurs presque complets.
Les rejets bovins portent de nombreuses traces de découpe et appellent quelques commentaires. En effet, la fracturation anthropique de deux fragments de métatarses les distingue des métatarses intacts présents dans ce silo. Ces fragments brisés sont surnuméraires et proviennent d’un bovin différent de celui qui livre les métatarses intacts. Ce constat est conforté par la découverte d’os carpiens de taille incompatible avec l’autopode antérieur complet. Il en est de même pour une phalange proximale antérieure. Enfin, divers sésamoïdes bovins proviennent de l’échantillon mis au jour au décapage. Il est probable qu’une partie d’entre eux soit à mettre au compte des seg-ments anatomiques connectés des bovins (infra), mais en l’absence de certitudes, ce matériel est considéré comme appartenant aux rejets.
Les ensembles anatomiques bovins connectés ont été mis au jour dans les US 2 et 3 (premiers comblements limono-argileux du fond du silo) (fig. 8 et 9). Il s’agit de deux autopodes hétérolatéraux postérieurs (fig. 9), aux connexions partiellement conservées (documentées en fouilles), mais aussi d’un autopode antérieur droit, recons-titué parmi le mobilier prélevé au décapage.
L’autopode antérieur droit, remonté lors de l’étude archéozoologique, est complet à l’exception de deux os de la rangée proximale du carpe (pisiforme et pyramidal) et de l’ensemble des sésamoïdes, probablement intégrés au corpus des déchets (supra).
Les autopodes postérieurs droit et gauche sont com-plets, ou quasi complets, à l’exception de leurs rangées tarsales proximales (talus, calcanéus). Diverses incisions anthropiques anciennes, fines et profondes, marquent les bords médiaux et latéraux des gorges de la surface proximale des naviculo-cuboïdes. Il s’agit ici de traces de dépouille, ou d’une désarticulation, effectuée par sépara-tion des deux rangées du tarse. Le prélèvement des cuirs ou des tendons est matérialisé par des incisions fines et profondes qui affectent systématiquement les faces exter-nes des parties distales des phalanges proximales des trois autopodes. Les os de l’autopode postérieur droit (fig. 9 au nord) sont proches les uns des autres, mais le plus sou-
N
A B
A B
C D
C D
St. 5227
5227-01
5227-01 : Creusement5227-02 : Limon argileux brun avec inclusions de charbon 5227-03 : Limon argileux brun clair avec inclusions de charbon 5227-04 : Limon brun foncé noir avec très nombreuses inclusions de charbon5227-05 : Limon beige5227-06 : Limon brun clair mélangé à du loess beige5227-07 : Faune (bœuf)5227-08 : Faune (bœuf)
Loess
5227-025227-03
5227-03
5227-04
5227-055227-06
5227-04 5227-02
5227-07 5227-08
5227-075227-08
0 1 m
Fig. 8 : Position stratigraphique des autopodes bovins connectés du silo 5227 (M. Landolt, M. Leprovost, R. Pinguet).

O. Putelat et M. Landolt, La caractérisation des dépôts animaux de La Tène ancienne à Geispolsheim
—33—
Épreuves d’auteur
vent déconnectés, en raison d’un pendage d’une dizaine de centimètres. La connexion tarse-métatarse est lâche. La connexion métatarse-phalanges est inexistante alors que les connexions inter-phalangiennes sont simplement déplacées, du fait que les phalanges reposent sur le plat du comblement de la structure. Il en est de même pour l’auto-pode postérieur gauche (fig. 9, au sud) mais les phalanges proximales, absentes sur le terrain lors du prélèvement, ont été retrouvées parmi le mobilier prélevé au décapage.
L’homogénéité ostéologique et ostéométrique de ces trois autopodes nous les font attribuer à un même animal, dont l’âge est évalué à plus de deux ans.
En résumé, ce dépôt est constitué de trois autopodes quasiment complets, attribués à un même bovin adulte, déposés dans les deux premières couches de comblement du silo. Les connexions anatomiques sont lâches et/ou déplacées, en raison d’une première phase de décomposi-tion à l’air libre et du pendage de la structure.
La nature de cette découverte est ambigüe. Il s’agit de toute façon du rejet de pieds bovins, non consommés et détachés du squelette durant son dépeçage. Cependant, les métapodes bovins entiers ne sont pas prévalents au sein des restes fauniques5. La rareté des découvertes d’auto-podes quasiment complets, avec leurs métapodes intacts, semblerait donc conférer une certaine spécificité aux trois autopodes bovins du silo 5227. Il ne nous est toutefois pas possible de différencier ici ce qui pourrait relever du dépôt intentionnel de trois autopodes, ou du dépôt intentionnel d’une peau bovine, encore munie de ses extrémités (voir par exemple Méniel 2008, p. 76), ou encore du rejet de trois autopodes non consommés.
5 Parmi les restes échantillonnés dans le matériel osseux laténien des fouilles 2008, où au moins quinze métapodes bovins ont été déterminés, seuls cinq sont entiers. Il en est de même pour les fouilles de 2006, où parmi onze métapodes bovins déterminés, seuls cinq sont entiers. De plus, la découverte simultanée de métapodes entiers et des os courts des basipodes et / ou acropodes au sein d’un même fait archéologique, est rare (2 occurrences sur 14 pour les fouilles de 2006 ; 3 occurrences sur 13 pour les fouilles de 2008).
2.2.2.2 Les caprinés
Le silo cylindrique 5219Le silo 5219 livre 51 restes dont 47 sont déterminés,
pour une masse totale de 0,6 kg. Il s’agit de quelques déchets mais surtout d’un ensemble constitué d’une tête osseuse et de plusieurs autopodes ovins (46 restes), pro-venant du fond de la structure (fig. 10 et 11).
L’hémi-crâne droit et sa mandibule, tous deux en connexion étroite, apparaissent en vue latérale et reposent à plat sur leurs faces médiales. La mandibule est encore connectée à l’os temporal et les arcs dentaires jugaux supérieurs et inférieurs sont au contact l’un de l’autre. L’hémi-crâne gauche est complètement désolidarisé de sa mandibule, qui repose sur sa face médiale, symétrique-ment à la mandibule droite. L’hémi-crâne gauche a été brisé, antérieurement au dépôt : le rang jugal M1M2M3 est plaqué sous la mandibule gauche, tandis que les par-
Fig. 9 : Autopodes bovins connectés du silo 5227 (M. Landolt).
N
A B
A B
C D
St. 5219
5219-01
5219-01 : Creusement5219-02 : Limon brun5219-03 : Faune (ovin)
Loess
5219-03
C D
St. 05213
5219-02
5219-03
0 1 m
Fig. 10 : Position stratigraphique du dépôt ovin du silo 5219 (M. Landolt, C. Leprovost, G. Pinguet).

Les dépôts d’ossements d’animaux en France, de la fouille à l’interprétation
—34—
Épreuves d’auteur
ties frontale, occipitale et une partie du rang jugal (P3P4) recouvrent cette mandibule. L’examen des surfaces osseu-ses prouve la fente sagittale de cette tête, à l’aide d’un cou-teau lourd, de la face à la région pariétale. L’os occipital est fendu manuellement, par écartement des deux moitiés de crâne précédemment dégagées. Les os hyoïdes sont absents, bien qu’ils aient été recherchés lors de la fouille. L’atlas découvert au décapage s’adapte parfaitement aux condyles occipitaux de cette tête. Il est tranché transversa-lement, du crânio-dorsal vers le caudo-ventral.
Quatre autopodes antérieurs et postérieurs hétérolaté-raux sont présents, avec des connexions variables, étroites (tarse-métatarse), lâches (métapodes-phalanges) et dépla-cées (phalanges). Ils sont parallèles les uns aux autres, à proximité immédiate et au nord de l’hémi-crâne droit. On reconnait, de droite à gauche (fig. 11), l’autopode antérieur gauche en vue palmaire, proximum à l’est ; les autopodes postérieur (en vue latérale) et antérieur (en vue palmaire) droits, parties proximales à l’ouest ; l’autopode postérieur gauche en vue médiale, proximum à l’est.
Les données ostéométriques très homogènes confir-ment l’observation macroscopique. Il est très vraisembla-ble que ces autopodes proviennent des quatre membres d’un même individu. L’essentiel des vestiges attendus est présent, à l’exception de toutes les rangées proxima-les des carpes et des tarses. Ces absences sont à mettre en relation avec les désarticulations effectuées entre les rangées proximales et distales. Cette pratique est avérée pour le tarse, puisque nous relevons sur les faces médiale et crâniale du naviculo-cuboïde droit un ensemble d’une dizaine de très fines incisions parallèles et profondes, causées par la dépouille de l’animal et la désarticulation du pied. D’autres absences (quelques sésamoïdes, deux
phalanges distales) sont probablement liées à des causes taphonomiques.
L’état de conservation des surfaces osseuses et le maintien de la plus grande partie des connexions plaident en faveur d’un enfouissement rapide des vestiges. Ainsi qu’en témoigne une certaine dislocation des acropodes (ensemble des phalanges), leur recouvrement n’a toutefois pas suivi immédiatement le dépôt.
La forme et les dimensions des chevilles osseuses et des autopodes attestent la présence d’un ovin mâle. Les données odonto-ostéologiques permettent d’estimer son âge aux environs de deux ans.
La fente sagittale de la tête osseuse et l’absence des os hyoïdes sont à mettre au compte du prélèvement de la cervelle et de la langue. Les traces de dépouille et de désarticulation observées sur les autopodes n’impliquent pas la consommation de ces parties du squelette, mais le rejet de parties peu charnues. Si ce n’est l’absence de traces d’ustion, la fente sagittale de la tête et l’intégrité anatomique des métapodes ne diffèrent pas des obser-vations effectuées dans certains rejets alimentaires (silos 5231, 5268 et 5316).
Le dépôt symétrique, en miroir, des deux moitiés de tête et un effet d’alignement des autopodes sont remar-quables. Faut-il pour autant y voir l’effet d’un agencement intentionnel des rejets ? Une autre hypothèse pourrait être avancée, où l’on envisagerait, dans un premier temps, le dépôt dans un récipient de la tête fendue, suivi du retour-nement de l’ensemble au moment du rejet. Une reconsti-tution expérimentale du geste supposé a été tentée. Elle montre que pour obtenir la disposition des deux moitiés de tête observée sur le terrain, il est obligatoire dans un premier temps, après la fente sagittale de la tête et son ouverture par basculement latéral des deux moitiés (pour prélever langue et cervelle), que les deux moitiés, crâne et mandibules, reposent à plat, leurs faces latérales sur le fond du récipient. Le « démoulage » du contenu pour-rait alors effectivement reproduire l’effet observé sur le terrain. Cependant, le parallélisme observé sur le terrain entre les divers autopodes resterait à expliquer : s’agit-il d’un agencement délibéré, d’un effet d’alignement ou de l’éventuel dépôt d’une peau, encore munie d’une partie des extrémités du corps ?
2.2.2.3 Les équidés
Le silo tronconique 5199Le silo 5199 livre 38 restes de macrofaune, dont 26
déterminés, pour une masse totale d’environ un kg. Les vestiges sont en majorité des déchets alimentaires et por-tent de nombreuses traces de découpe. On trouve aussi un ensemble anatomique constitué d’un membre postérieur droit d’équidé. Cet ensemble, partiellement représenté, provient de l’US 4, première couche de comblement du silo (fig. 12 et 13). Il comprend six os : le tibia, le calcanéus (prélevé au décapage), le talus, les métatarses II, III et IV. Le tibia et les métatarses reposent sur leurs
Fig. 11 : Détail de la tête et des autopodes ovins du silo 5219 (M. Landolt).

O. Putelat et M. Landolt, La caractérisation des dépôts animaux de La Tène ancienne à Geispolsheim
—35—
Épreuves d’auteur
faces médiales et forment un angle aigu. La connexion tibia-talus est lâche, la connexion tarse-talus est déplacée. Quatre des six os du tarse manquent, soit l’intégralité des deux étages de sa rangée distale. L’épiphysation des os indique qu’il s’agit d’un animal âgé de plus de trois ans et demi. On constate l’absence de traces de découpes et la présence d’altérations climatiques sur les surfaces osseu-ses, moins bien conservées que celles des déchets alimen-taires. Il est probable que ce lot a séjourné quelque temps à l’air libre. Il ne nous est pas possible de différencier ici ce qui résulterait du dépôt intentionnel d’un ensemble ana-tomique connecté (tibia-tarse-métatarse) de ce qui pourrait être mis au compte du rejet d’une pièce de viande non consommable (viande à évolution anormale ou avariée).
Le silo tronconique 5230Le silo 5230 livre 37 restes, dont 29 déterminés, pour
une masse totale d’environ 1 kg. Il s’agit pour moitié de déchets, mais aussi de deux autopodes d’équidé (16 restes). Ces deux autopodes antérieur et postérieur droits incomplets, aux connexions partiellement conservées, ont été mis au jour à une cinquantaine de centimètres l’un
de l’autre, au sommet de l’US 2 (premier comblement peu épais du fond du silo (fig. 14 et 15). L’autopode antérieur droit (fig. 15, à gauche) est composé des méta-carpes II-III-IV, des trois phalanges et d’un sésamoïde. La connexion métacarpe III-phalange proximale est déplacée. Les connexions phalangiennes sont étroites. L’autopode postérieur droit (fig. 15, à droite) est composé de la rangée distale du tarse, des métatarses II-III-IV, de trois phalanges et de deux sésamoïdes. Les connexions anatomiques sont strictes. L’homogénéité macroscopique et ostéométrique de ces deux autopodes nous les font attribuer à un même animal. Les phalanges proximales antérieure et postérieu-re livrent toutes deux des traces de dépouille, matérialisées par la présence de séries d’incisions fines, profondes, et parallèles les unes aux autres, localisées sur la face dor-sale de la diaphyse de la phalange antérieure, ou sur les bords latéral et médial de la partie distale de la phalange postérieure. L’âge de cet individu est évalué à plus de 15 mois, mais des remaniements osseux plaident en faveur de la présence d’un adulte. Le métatarse III de cet équidé est légèrement plus petit, mais surtout plus gracile que celui du silo 5199.
En résumé, ces deux autopodes incomplets, prove-nant probablement d’un même équidé adulte différent de celui du silo 5199, ont été déposés au fond du silo, sur la première couche de comblement. Les connexions anatomiques sont relativement bien conservées en raison du comblement rapide de la structure. Les phalanges mon-trent des traces de dépouille. Il ne nous est pas possible de différencier ici ce qui résulterait du dépôt intentionnel d’ensembles anatomiques connectés, ou du dépôt d’une peau encore munie d’une partie des extrémités du corps, ou encore ce qui pourrait être mis au compte du rejet de parties du squelette peu appréciées.
N
A B
A B
St. 5199
5199-01 Loess
5199-02
5199-01 : Creusement5199-02 : Limon argileux brun 5199-03 : Limon argileux brun clair 5199-04 : Limon brun foncé5199-05 : Limon argileux brun mélangé à du loess beige 5199-06 : Limon argileux brun mélangé à du loess beige 5199-07 : Limon sableux brun5199-08 : Faune (équidé)
5199-03
5199-04
06
055199-07
C D
C D
5199-08
5199-08
0 1 m
Fig. 12 : Position stratigraphique du membre d’équidé du silo 5199 (M. Landolt, C. Leprovost, R. Pinguet).
Fig. 13 : Détail du membre postérieur d’équidé du silo 5199 (M. Landolt).

Les dépôts d’ossements d’animaux en France, de la fouille à l’interprétation
—36—
Épreuves d’auteur
2.2.3 Les squelettes
2.2.3.1 Le bœuf
Le silo tronconique 5302Le silo 5302 livre 204 restes dont 164 déterminés,
pour une masse totale de 4,8 kg. Il s’agit de 67 déchets auxquels s’ajoute un squelette bovin mis au jour sur le fond du silo.
Les ossements du bovin, recouverts par les limons argileux de l’US 10, sont très bien conservés, parfois soli-darisés par des concrétions minérales (fig. 16 et 17). Le squelette est incomplet. Les lacunes les plus importantes affectent le membre antérieur et le coxal droits ainsi que le membre postérieur gauche (fig. 18). Les données odonto-ostéologiques permettent d’estimer l’âge de ce bovin aux environs de 20 à 24 mois au moment de son décès. La forme de l’os pubien montre la présence d’un mâle.
N
A B
A B
C D
C D
St. 5230
5230-01
5230-01 : Creusement5230-02 : Limon argileux brun 5230-03 : Dépôt de faune (équidé)5230-04 : Limon argilo-sableux brun gris
Loess5230-02
5230-03
5230-04
5230-05
5230-03
5230-05
5230-02
5230-02
0 1 m
Fig. 14 : Position stratigraphique des autopodes équins du silo 5230 (M. Landolt, C. Leprovost, R. Pinguet).
Fig. 15 : Autopodes équins du silo 5230 (M. Landolt).
N
A B
A B
St. 5302
5302-01
5302-01 : Creusement5302-02 : Limon argileux brun foncé 5302-03 : Limon brun clair mélangé à du loess beige5302-04 : Limon sableux brun mélangé à du loess beige 5302-05 : Limon brun clair mélangé à du loess beige5302-06 : Limon brun mélangé à du loess brun clair5302-07 : Limon argileux brun mélangé à du loess beige 5302-08 : Limon sableux brun mélangé à du loess beige 5302-09 : Limon sabelux brun foncé 5302-10 : Limon argileux brun foncé 5302-11 : Loess beige5302-12 : Faune (bœuf)
Loess
5302-07
5302-02
5302-06
5302-04
5302-10
05
5302-0911
5302-12
5302-08
5302-03
C D
C D
0 1 m
Fig. 16 : Position stratigraphique du bovin du silo 5302 (M. Landolt, C. Leprovost, R. Pinguet).
O. Putelat et M. Landolt, La caractérisation des dépôts animaux de La Tène ancienne à Geispolsheim
—37—
Épreuves d’auteur
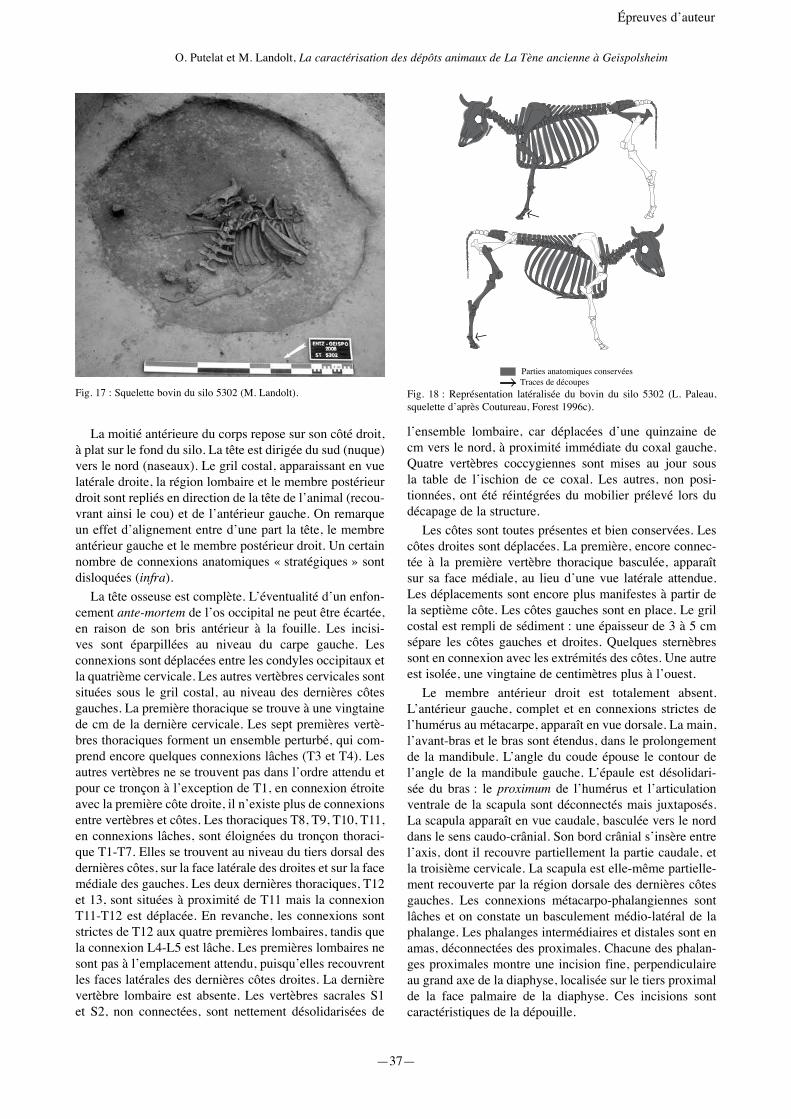
La moitié antérieure du corps repose sur son côté droit, à plat sur le fond du silo. La tête est dirigée du sud (nuque) vers le nord (naseaux). Le gril costal, apparaissant en vue latérale droite, la région lombaire et le membre postérieur droit sont repliés en direction de la tête de l’animal (recou-vrant ainsi le cou) et de l’antérieur gauche. On remarque un effet d’alignement entre d’une part la tête, le membre antérieur gauche et le membre postérieur droit. Un certain nombre de connexions anatomiques « stratégiques » sont disloquées (infra).
La tête osseuse est complète. L’éventualité d’un enfon-cement ante-mortem de l’os occipital ne peut être écartée, en raison de son bris antérieur à la fouille. Les incisi-ves sont éparpillées au niveau du carpe gauche. Les connexions sont déplacées entre les condyles occipitaux et la quatrième cervicale. Les autres vertèbres cervicales sont situées sous le gril costal, au niveau des dernières côtes gauches. La première thoracique se trouve à une vingtaine de cm de la dernière cervicale. Les sept premières vertè-bres thoraciques forment un ensemble perturbé, qui com-prend encore quelques connexions lâches (T3 et T4). Les autres vertèbres ne se trouvent pas dans l’ordre attendu et pour ce tronçon à l’exception de T1, en connexion étroite avec la première côte droite, il n’existe plus de connexions entre vertèbres et côtes. Les thoraciques T8, T9, T10, T11, en connexions lâches, sont éloignées du tronçon thoraci-que T1-T7. Elles se trouvent au niveau du tiers dorsal des dernières côtes, sur la face latérale des droites et sur la face médiale des gauches. Les deux dernières thoraciques, T12 et 13, sont situées à proximité de T11 mais la connexion T11-T12 est déplacée. En revanche, les connexions sont strictes de T12 aux quatre premières lombaires, tandis que la connexion L4-L5 est lâche. Les premières lombaires ne sont pas à l’emplacement attendu, puisqu’elles recouvrent les faces latérales des dernières côtes droites. La dernière vertèbre lombaire est absente. Les vertèbres sacrales S1 et S2, non connectées, sont nettement désolidarisées de
l’ensemble lombaire, car déplacées d’une quinzaine de cm vers le nord, à proximité immédiate du coxal gauche. Quatre vertèbres coccygiennes sont mises au jour sous la table de l’ischion de ce coxal. Les autres, non posi-tionnées, ont été réintégrées du mobilier prélevé lors du décapage de la structure.
Les côtes sont toutes présentes et bien conservées. Les côtes droites sont déplacées. La première, encore connec-tée à la première vertèbre thoracique basculée, apparaît sur sa face médiale, au lieu d’une vue latérale attendue. Les déplacements sont encore plus manifestes à partir de la septième côte. Les côtes gauches sont en place. Le gril costal est rempli de sédiment : une épaisseur de 3 à 5 cm sépare les côtes gauches et droites. Quelques sternèbres sont en connexion avec les extrémités des côtes. Une autre est isolée, une vingtaine de centimètres plus à l’ouest.
Le membre antérieur droit est totalement absent. L’antérieur gauche, complet et en connexions strictes de l’humérus au métacarpe, apparaît en vue dorsale. La main, l’avant-bras et le bras sont étendus, dans le prolongement de la mandibule. L’angle du coude épouse le contour de l’angle de la mandibule gauche. L’épaule est désolidari-sée du bras : le proximum de l’humérus et l’articulation ventrale de la scapula sont déconnectés mais juxtaposés. La scapula apparaît en vue caudale, basculée vers le nord dans le sens caudo-crânial. Son bord crânial s’insère entre l’axis, dont il recouvre partiellement la partie caudale, et la troisième cervicale. La scapula est elle-même partielle-ment recouverte par la région dorsale des dernières côtes gauches. Les connexions métacarpo-phalangiennes sont lâches et on constate un basculement médio-latéral de la phalange. Les phalanges intermédiaires et distales sont en amas, déconnectées des proximales. Chacune des phalan-ges proximales montre une incision fine, perpendiculaire au grand axe de la diaphyse, localisée sur le tiers proximal de la face palmaire de la diaphyse. Ces incisions sont caractéristiques de la dépouille.
Fig. 17 : Squelette bovin du silo 5302 (M. Landolt).
Parties anatomiques conservéesTraces de découpes
Fig. 18 : Représentation latéralisée du bovin du silo 5302 (L. Paleau, squelette d’après Coutureau, Forest 1996c).

Les dépôts d’ossements d’animaux en France, de la fouille à l’interprétation
—38—
Épreuves d’auteur
Le coxal droit est absent. Le membre postérieur droit apparaît en vue latérale, en extension dans le sens sud-nord (la jambe au nord), strictement connecté du fémur à la rangée proximale du tarse (talus, calcanéus). Des concrétions minérales solidarisent la patella au fémur, la malléole et la rangée proximale du tarse au tibia. Le métatarse droit est mis au jour sous le gril costal, dans le prolongement du tarse dont il est déconnecté. Il forme un angle d’environ 90° avec l’axe du tibia. La rangée distale du tarse, naviculo-cuboïde et grand cunéiforme est décon-nectée et découverte entre le talus et le métatarse. Les phalanges et leurs sésamoïdes sont encore en connexion stricte au distum du métatarse. Chacune des phalanges proximales montre trois rangées d’incisions parallèles, au milieu des faces dorsale et latérale de la diaphyse, elles aussi caractéristiques de la dépouille. Le coxal gauche, très bien conservé, apparaît en vue dorsale. Le membre postérieur gauche est absent.
La position des ossements permet d’envisager leur stra-tification définitive au fond du silo selon l’ordre suivant : tête osseuse avec les sept vertèbres cervicales, en cours de déconnexion ; membre antérieur gauche (étendu dans le prolongement de la mandibule, l’angle du coude épousant le contour de l’angle de la mandibule gauche et la scapula recouvrant partiellement la partie caudale de l’axis en s’insérant entre l’axis et la troisième cervicale) ; mem-bre pelvien droit ; thorax ; régions lombaire et sacrale, coxal gauche. La complexité de l’ordonnancement de cet assemblage osseux nous amène tout d’abord à formaliser quelques observations :
- les seules traces anthropiques décelées, sur les pha-langes proximales, ne concernent que l’écorchement de l’animal ;
- alors que les surfaces osseuses sont dans l’ensemble très lisibles, aucun indice tracéologique ne documente le dépeçage partiel du bovin, et encore moins le désossement des viandes ;
- un dépeçage partiel, ou une réduction de la carcasse, est irréfutable en raison de l’absence des membres anté-rieur droit et postérieur gauche, qui ne peut en aucun cas être mise au compte de phénomènes taphonomiques consécutifs à la fouille ;
- le membre antérieur, extrêmement facile à détacher du corps, pourrait éventuellement avoir été prélevé sans laisser de traces visibles avant le dépôt/rejet du bovin dans le silo ;
- il est en revanche improbable que l’ablation du coxal droit et du membre postérieur gauche ait pu être effectuée sur une carcasse fraîche sans laisser de traces.
Bien que les connexions anatomiques soient nombreu-ses et souvent strictes, il apparaît que ce bovin a été dis-loqué durant sa décomposition, et avant un enfouissement définitif relativement rapide, comme l’atteste l’excellent état de conservation des surfaces osseuses. Le bon ordon-nancement général des différentes régions du squelette fait écarter l’hypothèse d’un rejet « tête la première » dans le silo, car une décomposition de ce type aurait entraîné des
effets de torsion, une dispersion des côtes, etc. La compa-raison des connexions anatomiques de ce bovin avec un état théorique attendu dans des conditions normales/humi-des de décomposition (Méniel 2008, p. 147 ; Morris 2008, p. 48), montre des divergences. Des connexions très labi-les sont conservées (crâne-mandibule, fémur/patella, pha-langes postérieures), tandis que les connexions résistantes de la colonne vertébrale sont déplacées. L’hypothèse d’une décomposition normale du squelette bovin est donc écartée. Le fait que les déplacements les plus nombreux concernent les restes vertébraux évoque les modifications du processus de décomposition causées par des phases de dessiccation du squelette en cours de putréfaction. En effet, dans certaines conditions, le corps peut subir une momification naturelle, où la peau joue le rôle d’une enveloppe protégeant la tête et le squelette appendicu-laire (Méniel 2008, p. 56-57). Au retour de l’humidité la décomposition concerne en priorité le tronc (Ibid.). Cette hypothèse n’est toutefois pas entièrement satisfaisante car nous avons démontré le prélèvement de la peau. Il nous semble que la mise en place de l’assemblage osseux est complexe et fait conjointement appel à des agents naturels et des actions anthropiques. Nous proposons donc l’hypo-thèse suivante :
- mise à mort éventuelle par assommage ou mort natu-relle ;
- écorchement partiel du bovin ;- dépôt dans un bref laps de temps du bovin dans
le silo en décubitus latéral droit (prélèvement possible dès ce stade du membre antérieur droit par levée « en raquette ») ;
- décomposition naturelle au fond du silo ;- intervention anthropique avec prélèvement du posté-
rieur gauche, prélèvement de l’antérieur droit avec bascu-lement du squelette vers le nord entraînant le déplacement des vertèbres cervicales et thoraciques, prélèvement du coxal droit.
C’est donc en fait le prélèvement anthropique, durant le processus de décomposition, qui aurait causé les décon-nexions et les déplacements les plus conséquents, entraî-nant un effet de pliage du squelette, et un alignement du squelette appendiculaire non prélevé (membre antérieur gauche et postérieur droit).
2.2.3.2 Les caprinésLe silo tronconique 5376Le silo 5376 livre 219 restes dont 188 déterminés, pour
une masse de 1,6 kg. Il s’agit de 75 déchets auxquels s’ajoute le squelette d’un ovin (US 4). Ce squelette, qua-siment complet, mais remanié, a été mis au jour sur l’US 3, deuxième US de comblement, à proximité de sa paroi nord-ouest (fig. 19, 20, 21). La morphologie du pelvis indique une brebis, et l’attrition des dents jugales permet d’estimer l’âge entre 3 et 4 ans.
L’assemblage osseux se compose d’un noyau central comprenant la tête (complète), le cou (connecté à la région

O. Putelat et M. Landolt, La caractérisation des dépôts animaux de La Tène ancienne à Geispolsheim
—39—
Épreuves d’auteur
occipitale), le squelette axial (du thorax au sacrum) et la ceinture pelvienne (fig. 20). Deux « appendices » latéraux se déploient de part et d’autre du noyau central : au nord-est, les membres postérieurs gauche et droit complets ; au sud, les membres antérieurs partiellement représentés.
Dans le noyau central, la tête, entière, est déformée. La fracture ancienne du pariétal se manifeste par trois fissu-res à gauche et une cassure franche à droite. La bouche est close. L’atlas est connecté à l’occipital et les cervica-les sont connectées de C1 à C5. La tête et les vertèbres cervicales remplissent la cavité abdominale. La face est recouverte par les dernières thoraciques, tandis que les cervicales de rang 3, 4 et 5 sont recouvertes par les trois dernières vertèbres lombaires.
Au nord-est, les membres postérieurs gauche et droit sont complets, en extension selon un axe nord-est/sud-ouest (parties fémorales au sud-ouest), et se croisent par-tiellement. Les connexions du postérieur droit sont strictes entre le fémur et le coxal, lâche à déplacée entre le fémur et le tibia6, lâche entre le tibia et le tarse, strictes pour l’autopode. Celles du postérieur gauche sont strictes entre le fémur et le coxal, lâche à déplacée entre le fémur et le tibia, strictes du tibia aux phalanges distales.
Au sud, les membres antérieurs gauche et droit sont partiellement représentés. Pour l’antérieur droit, la face médiale de la scapula est au contact des quatre dernières vertèbres lombaires et recouvre également quelques côtes déplacées. La connexion scapulo-humorale est lâche : il existe un écart d’un demi centimètre entre l’articulation ventrale de la scapula et la partie proximale de l’humérus. La connexion entre l’humérus et le radius est stricte, tandis que l’absence de l’ulna reste inexpliquée. Le métacarpe
6 Il existe pour les genoux gauche et droit des écarts respectifs de deux et trois centimètres entre les articulations distales des fémurs et les articulations proximales des tibias.
B
B
A
A
C D
St. 5376
5376-01
5376-01 : Creusement5376-02 : Limon loessique brun5376-03 : Limon loessique gris à noir charbonneux5376-04 : Faune (ovin)5376-05 : Limon loessique brun5376-06 : Limon loessique brun très charbonneux avec inclusions de charbon5376-07 : Limon loessique brun clair5376-08 : Limon loessique brun charbonneux avec inclusions de charbon5376-09 : Loess beige clair
Loess5376-02
N
5376-03
5376-0506 5376-07
5376-08 09
C D
5376-04
5376-04
0 1 m
Fig. 19 : Position stratigraphique du squelette ovin du silo 5376 (M. Landolt, C. Leprovost, R. Pinguet).
Fig. 20 : Vue de détail du squelette ovin du silo 5376 (M. Landolt).
Parties anatomiques conservéesFig. 21 : Représentation latéralisée du squelette ovin du silo 5376 (L. Paleau, squelette d’après Coutureau, Forest 1996a).

Les dépôts d’ossements d’animaux en France, de la fouille à l’interprétation
—40—
Épreuves d’auteur
est à l’emplacement attendu : son proximum étant distant de la partie distale du radius d’environ 1,5 cm. Toutes les connexions de l’autopode sont strictes, du métacarpe aux phalanges, mais une phalange distale manque. La position du membre antérieur gauche est perturbée. La scapula est mise au jour au fond du silo, sous les cervicales C3, C4, C5. L’humérus et le carpe gauches ont été prélevés durant le décapage de l’US. Le radius gauche est recouvert par le radius droit. La connexion entre la partie distale du radius (et peut-être le carpe) et le métacarpe est de toute façon déplacée, puisque le proximum du métacarpe droit est éloigné d’une dizaine de centimètres du distum du radius. Les phalanges de cette main gauche sont déplacées.
Les modalités de mise en place du dépôt sont relative-ment complexes. Les traces anthropiques sont rares et non avérées : le bris ancien de l’os pariétal pourrait indiquer un assommage ; un enfoncement ancien concerne la partie dorsale de la scapula droite. Aucune autre trace n’indique de prélèvement. Les remaniements stratigraphiques de la vertèbre cervicale 7, d’une vertèbre coccygienne et de quelques os de l’antérieur gauche sont mis au compte du décapage, ou de bioturbations attestées. De fait, les prin-cipales absences affectent les premières vertèbres thoraci-ques, de T2 à T6 ou 7 et l’ulna droite, non détectées parmi le matériel osseux prélevé au décapage. Les déplacements osseux les plus notables concernent d’une part le déplace-ment de la tête, connectée aux cinq premières cervicales, puis sa mise en place dans l’abdomen de la brebis, et d’autre part le décollement des deux membres antérieurs, repositionnés ensuite dans l’alignement des postérieurs. Le déplacement de la tête et du cou n’a pu intervenir que par arrachement du cou, au cours de la décomposition de l’animal, in situ ou à l’extérieur du silo, puisqu’aucune trace de découpe n’affecte les vertèbres C5 (connectée au cou), C6 (mise au jour avec le gril costal), C7 (pro-venant du décapage), T1 (mise au jour dans un ensemble de côtes perturbées situées sous la scapula droite). Il est très probable que la disparition des thoraciques (supra) et l’éparpillement de quelques côtes ait eu lieu à cette occasion. Le décollement des membres antérieurs a pu avoir lieu immédiatement après le décès de l’animal, sans laisser de traces, puisque l’ablation du membre thoracique s’effectue extrêmement facilement chez les herbivores du fait de l’absence de clavicule. Cette opération, dite « levée en raquette » (Lignereux, Peters 1996), se résume à la section des muscles du cou et du thorax attachés à la scapula et ne nécessite pas d’entrer au contact des sur-fasses osseuses. Plus vraisemblablement, elle a du être effectuée en même temps que le repositionnement de la tête, la connexion scapula-gril costal étant extrêmement labile (Morris 2008). L’assemblage osseux a été ensuite enseveli dans un laps de temps relativement court, comme en attestent le maintien d’une bonne partie des connexions anatomiques et la conservation des surfaces osseuses. La finalité de ces manipulations anthropiques intentionnelles restant inconnue, nous nous limiterons à constater une évi-dente volonté de mise en scène, caractérisée par un ordon-nancement soigneux des diverses parties du squelette.
2.2.3.3 Les suinés
Le silo tronconique 5176Le silo 5176 livre 206 restes dont 192 déterminés, pour
une masse totale d’environ 1,5 kg. Il s’agit de quelques rejets, mais surtout d’un squelette de porc (177 restes). Les ossements prélevés lors du décapage mécanique ont été réintégrés au corpus, sans certitude d’exhaustivité. La forme des canines supérieures indique la présence d’une femelle, dont l’âge est estimé entre 16 et 18 mois.
La carcasse, rejetée parmi la première couche de com-blement du silo (US 3, fig. 22), montre un pendage d’en-viron 20 cm entre sa région coxale et sa région mandibu-laire.L’animal est partiellement représenté, en connexions partielles (fig. 23 et 24). Sa moitié antérieure repose en décubitus latéral gauche. La moitié postérieure, partielle-ment représentée, a été soumise à un effet de torsion, au moment du dépôt, puisque la région lombaire apparaît en vue ventrale. La tête, quasi intacte, apparaît en vue latérale droite, reposant à plat sur son côté gauche. La colonne vertébrale est vrillée. Les vertèbres cervicales et lombaires
N
A B
A B
5176-01
5176-01 : Creusement5176-02 : Limon sableux brun gris 5176-03 : Limon sableux gris sombre 5176-04 : Loess beige mélangé à du limon sableux brun5176-05 : Limon sableux brun5176-06 : Loess beige mélangé à du limon sableux brun5176-07 : Limon sableux brun foncé 5176-08 : Loess beige mélangé à du limon sableux brun5176-09 : Loess beige5176-10 : Loess beige mélangé à du limon sableux brun5176-11 : Faune (porc)
Loess
5176-025176-115176-03
5176-09
5176-045176-05
5176-08
5176-075176-10
5176-06
C D
C D
5176-11
0 1 m
St. 5176
Fig. 22 : Position stratigraphique du squelette de porc du silo 5176 (M. Landolt, C. Leprovost, R. Pinguet).

O. Putelat et M. Landolt, La caractérisation des dépôts animaux de La Tène ancienne à Geispolsheim
—41—
Épreuves d’auteur
sont pour la plupart en connexions étroites. L’absence de la vertèbre thoracique 2 est constatée lors de l’étude en laboratoire. Le sacrum a été endommagé lors du décapage. Les vertèbres coccygiennes sont absentes. Le gril costal, affaissé, rempli de terre, suit les mouvements de torsion de la colonne vertébrale. Les côtes droites sont entrecroisées sous la scapula droite. Les côtes gauches sont situées sous l’animal. Une côte cassée en cours de consolidation a pu être observée.
Le membre antérieur droit est en flexion contrainte vers l’extérieur du corps (à l’ouest). Son autopode, par-tiellement engagé sous le corps, se montre en vue médio-palmaire. Le membre antérieur gauche est complet, en connexion étroite et en flexion forcée sous l’animal. Son autopode apparaît en vue palmaire, légèrement vrillé en direction de l’intérieur du corps.
Le membre postérieur droit est presque complet. Environ 10 cm en contrebas du sacrum, et à l’aplomb de celui-ci, se trouvent l’acétabulum du coxal et le grand trochanter du fémur. La patella et la fibula sont absentes. L’autopode est découvert à gauche du corps, dirigé de l’est vers l’ouest, et le pied apparaît en vue plantaire. La distance entre la région coxale et le tarse est compatible avec l’hypothèse d’une connexion anatomique totale du postérieur au coxal, bien que l’emplacement du fémur et du tibia, prélevés au décapage, ne soit pas documenté. La plupart des os du tarse sont en connexion lâche, mais le calcanéus fait exception, puisqu’il est déplacé à l’aplomb de la main droite. Les métapodes et les phalanges sont en connexion étroite.
Le membre postérieur gauche et sa ceinture pelvienne sont totalement absents.
Aucune trace anthropique n’a été relevée qui puisse traduire la mise à mort ou la récupération de parties car-nées. L’os occipital et la région frontale étant intacts, nous ne relevons pas d’indices avérés de l’assommage du porc. Aucun indice d’égorgement ne marque les faces ventrales
des vertèbres cervicales. La palette de la scapula gauche est trop endommagée pour statuer quant à une éventuelle saignée portée directement au cœur.
En sus de l’absence du membre postérieur gauche (supra), divers manques anatomiques sont décelés. L’absence de quelques os courts est probablement liée au décapage mécanique (vertèbres coccygiennes, os des carpes, phalanges). Il est plus difficile de statuer sur l’ab-sence totale du postérieur gauche, puisqu’aucun vestige ne permet d’en supposer la présence au moment du dépôt du cadavre. De même les absences de la seconde thoracique, très probablement présente lors du prélèvement, et de la fibula droite restent incomprises.
En résumé, cette jeune truie a été déposée dans une des premières couches de comblement du silo, avec un léger pendage, la tête au point le plus bas. Il n’a pas été apporté un soin particulier à la mise en place du cadavre, mais le bon état de conservation des surfaces osseuses et les faibles déplacements des os dans le silo indiquent un enfouissement rapide. Le corps, vrillé, reposait à plat sur sa face antérieure gauche, tandis que la moitié posté-rieure du corps se présentait sur sa face ventrale, la cuisse droite marquant un angle très ouvert par rapport au corps. L’absence du membre postérieur gauche reste inexpli-quée : est-elle antérieure au dépôt ? Est-elle consécutive aux façons culturales ultérieures ou à la fouille ?
Le silo cylindrique 5375Le silo 5375 livre 643 restes dont 614 déterminés,
pour une masse totale de 4,4 kg. Il s’agit de 121 déchets osseux, auxquels s’ajoutent trois squelettes de porcs dépo-sés sur l’US 2, premier comblement du silo (fig. 25). Le prélèvement des porcs 1 et 2 (US 3 et 4) (fig. 26) a révélé la présence d’un troisième porcelet, immédiatement sous-jacent (US 5) (fig. 27). La conservation des vestiges est excellente. Les données odonto-ostéologiques montrent la présence d’une jeune truie, âgée de 14 à 18 mois (porc 1,
Fig. 23 : Squelette de porc du silo 5176 (M. Landolt).
Parties anatomiques conservées
Fig. 24 : Représentation latéralisée du squelette de porc du silo 5176 (L. Paleau, squelette d’après Coutureau et al. 2003).
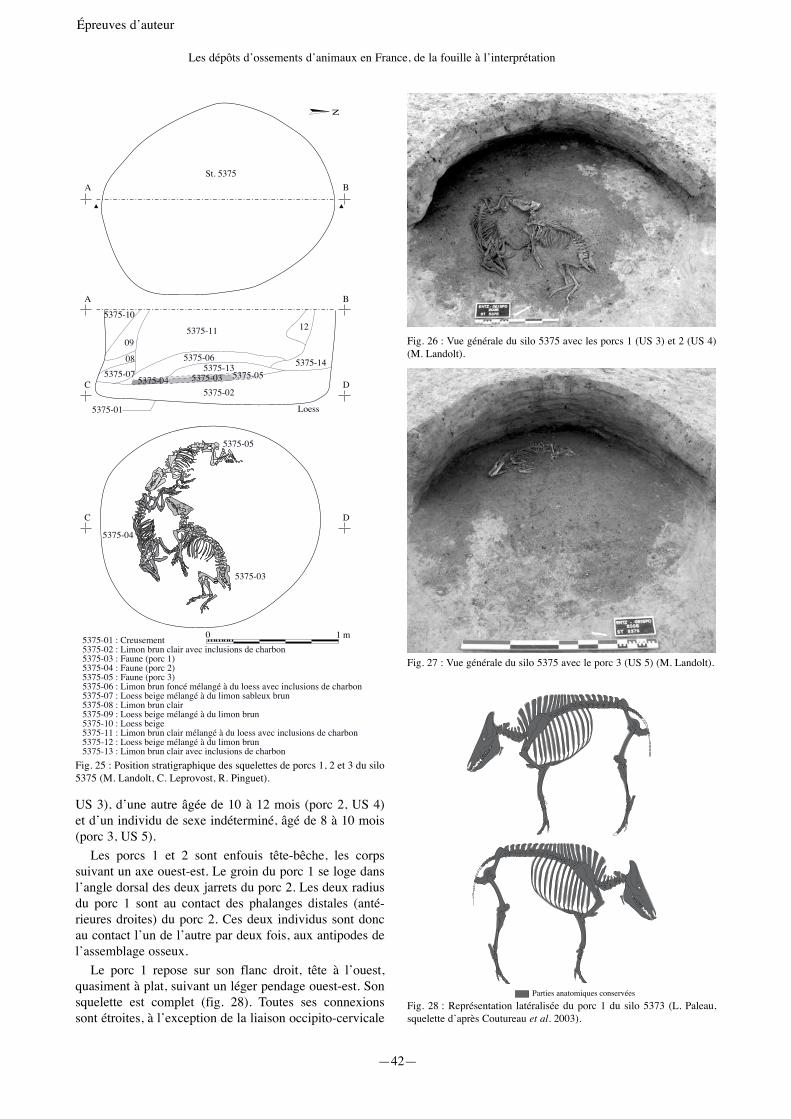
Les dépôts d’ossements d’animaux en France, de la fouille à l’interprétation
—42—
Épreuves d’auteur
US 3), d’une autre âgée de 10 à 12 mois (porc 2, US 4) et d’un individu de sexe indéterminé, âgé de 8 à 10 mois (porc 3, US 5).
Les porcs 1 et 2 sont enfouis tête-bêche, les corps suivant un axe ouest-est. Le groin du porc 1 se loge dans l’angle dorsal des deux jarrets du porc 2. Les deux radius du porc 1 sont au contact des phalanges distales (anté-rieures droites) du porc 2. Ces deux individus sont donc au contact l’un de l’autre par deux fois, aux antipodes de l’assemblage osseux.
Le porc 1 repose sur son flanc droit, tête à l’ouest, quasiment à plat, suivant un léger pendage ouest-est. Son squelette est complet (fig. 28). Toutes ses connexions sont étroites, à l’exception de la liaison occipito-cervicale
N
A B
A B
C D
C D
St. 5375
5375-01 Loess
5375-06
5375-01 : Creusement5375-02 : Limon brun clair avec inclusions de charbon 5375-03 : Faune (porc 1)5375-04 : Faune (porc 2) 5375-05 : Faune (porc 3) 5375-06 : Limon brun foncé mélangé à du loess avec inclusions de charbon 5375-07 : Loess beige mélangé à du limon sableux brun 5375-08 : Limon brun clair5375-09 : Loess beige mélangé à du limon brun 5375-10 : Loess beige5375-11 : Limon brun clair mélangé à du loess avec inclusions de charbon 5375-12 : Loess beige mélangé à du limon brun5375-13 : Limon brun clair avec inclusions de charbon
5375-11
5375-07
5375-10
08
09
5375-13 5375-14
12
5375-02
5375-04
5375-03
0 1 m
5375-05
5375-035375-04 5375-05
Fig. 25 : Position stratigraphique des squelettes de porcs 1, 2 et 3 du silo 5375 (M. Landolt, C. Leprovost, R. Pinguet).
Fig. 26 : Vue générale du silo 5375 avec les porcs 1 (US 3) et 2 (US 4) (M. Landolt).
Fig. 27 : Vue générale du silo 5375 avec le porc 3 (US 5) (M. Landolt).
Fig. 28 : Représentation latéralisée du porc 1 du silo 5373 (L. Paleau, squelette d’après Coutureau et al. 2003).
Parties anatomiques conservées

O. Putelat et M. Landolt, La caractérisation des dépôts animaux de La Tène ancienne à Geispolsheim
—43—
Épreuves d’auteur
(l’atlas, légèrement basculé vers la gauche, est partiel-lement déconnecté des condyles occipitaux). La tête est intacte, mandibules closes. Le dos est légèrement voûté. Le gril costal est effondré. Les pattes antérieures sont entrecroisées. La patte droite, au contact du sol par sa face latérale est sensiblement perpendiculaire à l’axe vertébral. L’antérieure gauche est repliée en direction de l’arrière du corps et forme un angle d’environ 45° avec l’axe verté-bral. Les pattes postérieures sont parallèles l’une à l’autre, légèrement fléchies, sensiblement perpendiculaires à l’axe vertébral. Les patellas sont en place.
Le porc 2 repose quasiment à plat, tête à l’est, en décu-bitus dorso-latéral droit. Le squelette est complet (fig. 29). Toutes les connexions sont étroites. La tête est intacte, gueule close. Les mandibules, apparaissant en vue latéro-ventrale gauche sont basculées vers la gauche. L’axe ver-tébral est légèrement arqué, sans être voûté. Le gril costal est effondré. Les pattes antérieures se situent à l’aplomb des mandibules. L’antérieure gauche est en flexion forcée, la scapula outrepasse l’axe vertébral. Les pattes postérieu-res sont fléchies, à l’aplomb de la région pelvienne.
La tête du porc 3 est apparue lors du prélèvement des os coxaux du porc 2, et le contact de la tête du porc 3 avec la région pelvienne et/ou fémorale du porc 2 est avéré. L’animal repose en décubitus latéral droit, à plat, selon un axe nord-ouest/sud-est, tête au sud-est. Le squelette est complet (fig. 30). Toutes les connexions sont étroites, à l’exception de la connexion scapulo-humérale gauche, plus lâche. La tête est intacte, la gueule est close. L’arc vertébral est légèrement incurvé et suit les contours du silo, distants d’une vingtaine de centimètres. Le gril cos-tal est effondré en place. Les antérieurs sont parallèles, à
l’aplomb du garrot. La scapula gauche, perforée de façon ancienne, pourrait indiquer une mise à mort de l’animal, par perforation du cœur. Les membres postérieurs se superposent. En semi-flexion, ils sont parallèles, mais les autopodes sont légèrement décalés l’un par rapport à l’autre.
En résumé, les trois porcs présentent chacun un sque-lette complet. Aucune trace de découpe, aucune absence remarquable (à l’exception d’un petit déficit d’os courts imputable à la fouille) n’ont été observés. L’état de conser-vation des squelettes, les excellentes représentations anatomiques, le maintien des connexions, indiquent un enfouissement relativement rapide, suivi d’une décompo-sition en espace colmaté. Les âges de ces trois porcs, deux femelles et un animal de sexe indéterminé, sont étagés entre 8 et 18 mois, ce qui signifie qu’ils n’appartiennent pas à la même portée. Sans mise en évidence de traces anthropiques, la mise à mort des porcs 1 et 2 n’est pas avé-rée. La scapula gauche du porc 3 montre une hypothétique trace de perforation du cœur.
Le dépôt semble s’être déroulé dans l’ordre suivant : dépôt et recouvrement du porc 3 par un peu de sédiment dans un premier temps, suivi très rapidement du dépôt simultané tête-bêche des porcs 1 et 2. La disposition des corps, reposant tous étendus sur leurs côtés droits et non emmêlés, laisse envisager la recherche d’un certain ordon-nancement. En particulier, le fait que les porcs ne reposent pas sur le cône d’effondrement du silo, mais sur sa pre-mière couche de comblement, et que le porc 3 soit situé à proximité de la paroi, et non à l’aplomb de la gueule du silo, nous autorise à réfuter l’hypothèse d’un positionne-ment aléatoire des cadavres, jetés depuis la surface.
Parties anatomiques conservéesFig. 29 : Représentation latéralisée du porc 2 du silo 5375. (L. Paleau, squelette d’après Coutureau et al. 2003).
Parties anatomiques conservées
Fig. 30 : Représentation latéralisée du porc 3 du silo 5375 (L. Paleau, squelette d’après Coutureau et al. 2003).

Les dépôts d’ossements d’animaux en France, de la fouille à l’interprétation
—44—
Épreuves d’auteur
Le silo cylindrique 5564Le silo 5564 livre 69 restes, tous déterminés, pour une
masse totale de 0,5 kg. L’essentiel de l’assemblage osseux est constitué par le squelette d’un porc (68 restes), mis au jour au sein de l’US 2, premier comblement argilo-limo-neux du silo (fig. 31). Une datation radiocarbone engagée sur un de ses os permet une attribution chronologique comprise entre 751 et 401 avant J.-C. à 95,5 % près, c’est à dire entre le Hallstatt C et la fin de La Tène A2 (Poz-32052 : 2425 ± 35 BP). L’hypothèse d’une attribution chronologique à La Tène ancienne est liée au contexte archéologique (les dépôts de faune sont attribués à l’oc-cupation laténienne). Cependant, une attribution chrono-logique au Hallstatt C pourrait également être proposée, puisqu’une occupation de cette époque est attestée en zone 5.
Les données odonto-ostéologiques montrent la pré-sence d’un animal âgé de 6 à 8 mois, de sexe indéterminé. Les ossements de l’animal sont souvent solidarisés par des concrétions calcaires. Les documents de terrain ne permettent d’observer correctement que la connexion conservée entre la tête et les mandibules. Les vertè-bres cervicales sont proches de la localisation attendue, mais mal déterminables. Les os longs, relativement bien conservés, ne semblent pas avoir été connectés au moment de l’enfouissement du squelette. On note l’écrasement de la quasi-totalité des vertèbres, du gril costal, et l’absence de la totalité des autopodes (fig. 32 et 33). Aucune trace de découpe ni de cuisson n’est observée sur les surfaces osseuses.
A B
A B
St. 5564
5564-01
5564-02
Loess
St. 05565
5564-01 : Creusement5564-02 : Limon argileux brun clair mélangé à du loess beige 5564-03 : Faune (porc)5564-04 : Limon argileux brun clair mélangé à du loesss beige
5564-03
5564-03
5564-02
C D
C D
0 1 m
N
Fig. 31 : Position stratigraphique du squelette de porc du silo 5564 (M. Landolt, C. Leprovost, R. Pinguet).
Fig. 32 : Vue générale du silo 5564 avec le squelette de porc (M. Landolt).
Fig. 33 : Représentation latéralisée du squelette de porc du silo 5564 (L. Paleau, squelette d’après Coutureau et al. 2003).
Parties anatomiques conservées

O. Putelat et M. Landolt, La caractérisation des dépôts animaux de La Tène ancienne à Geispolsheim
—45—
Épreuves d’auteur
La concentration de faune 5674La concentration de faune 5674 livre 139 restes, dont
125 déterminés, pour une masse totale d’environ 1,2 kg. Tous les restes déterminés proviennent d’un même squelette de porc, mis au jour dans ce qui semblerait être une fosse très arasée dont les contours n’ont pas pu être observés.
La datation radiocarbone d’un os de l’animal permet une attribution chronologique comprise entre 770 et 416 avant J.-C. à 95,5 % près, c’est-à-dire entre le Hallstatt C et la fin de La Tène A2 (Poz-32053 : 2480 ± 35 BP). Ici encore, en raison des difficultés de datation liées au palier du Premier âge du Fer, l’attribution chronologique à La Tène ancienne est fonction du contexte archéologique. Cependant, comme pour le silo 5564, une attribution chro-nologique à l’occupation du Hallstatt C attestée en zone 5 pourrait également être proposée.
L’épiphysation des os indique que cet individu, de sexe indéterminé, est âgé d’environ 42 à 48 mois.
Les observations de terrain montrent que l’animal, de bonne taille, a été déposé sur le dos, thorax au nord, pelvis au sud-est (fig. 34). Le squelette a été endommagé lors du décapage mécanique, ce qui explique au moins partielle-ment les lacunes anatomiques constatées parmi la colonne vertébrale et les autopodes (fig. 35). Aucun indice de la présence de la tête n’a toutefois été détecté parmi les restes collectés en vrac.
Les vertèbres thoraciques, lombaires et sacrales appa-raissent en vue ventrale. L’ensemble, en connexions étroites, forme un arc de cercle, légèrement vrillé vers la gauche dans la région du garrot. Les côtes droites sont pla-quées vers la gauche dans la région du garrot ; affaissées pour les vertèbres suivantes ; ouverte vers les dernières thoraciques. Les côtes gauches sont plaquées contre le fond de la fosse. Les sternèbres en connexions lâches avec quelques ligaments intercostaux, ont basculé vers la face médiale des côtes gauches.
L’antérieur droit est déplié, affaissé au contact de la face ventrale du gril costal. La scapula apparaît en vue
caudale, basculée d’un quart de tour au nord, en direction de la tête du porc. Sa connexion avec l’humérus n’est pas observable, car détruite par le décapage. L’humérus est encore connecté strictement au radius et à l’ulna et l’en-semble apparaît en vue dorsale. Le carpe est absent. Les métacarpes II et III, la phalange proximale du doigt III sont représentés sur les documents de terrain, à proximité du distum du radius droit et du proximum de tibia droit. Cette remarque tend à indiquer que l’absence du carpe et de la plupart des phalanges de ce membre est consécutive à la mise au jour du squelette. On trouve, mêlé aux pre-mières thoraciques conservées, un fragment de scapula gauche, en vue caudale. C’est le seul témoin du membre antérieur gauche non perturbé par le décapage. Nous constatons par ailleurs (fig. 36) qu’un déficit anatomique affecte l’antérieur gauche par rapport au droit (radius, autopode), y compris après réintégration au corpus des os ramassés en vrac.
Les os coxaux apparaissent en vue ventrale et les ischions ont disparu au décapage. Le fémur, intact, a été prélevé au décapage de la fosse. Le tibia est proche de la position attendue, mais outrepasse l’axe vertébral, vers le nord-est. Sa connexion avec le talus est étroite et ce der-nier est en connexion avec le calcanéus. Le fémur gauche est connecté au coxal et apparaît en vue caudo-latérale ; sa patella, qui a migré durant la décomposition des chairs, est mise au jour contre les corps des vertèbres thoraciques et sous les côtes gauches.
La position des vestiges montre donc un dépôt de l’ani-mal sur le dos, légèrement vrillé vers la gauche dans la région thoracique. Le membre antérieur droit est étendu dans l’axe du corps en direction caudale. Les deux mem-
Fig. 34 : Squelette de porc 5674 (M. Landolt).
Parties anatomiques conservées
Fig. 35 : Représentation latéralisée du squelette de porc 5674 (L. Paleau, squelette d’après Coutureau et al. 2003).

Les dépôts d’ossements d’animaux en France, de la fouille à l’interprétation
—46—
Épreuves d’auteur
bres postérieurs sont dépliés, vers l’avant et en travers du corps. Les données de terrain sont trop incomplètes pour qu’il nous soit possible de trancher entre deux hypothè-ses : enfouissement en décubitus dorsal d’un individu mort en décubitus ventral, membres engagés sous le corps, ou enfouissement en décubitus dorsal, avant rigidité cadavérique, et pratique d’une certaine contrainte sur les membres pour les adapter à la hauteur de la fosse ?
Des remaniements post-mortem, amplifiés par le décu-bitus dorsal du suiné, sont à imputer aux phénomènes de décomposition : la déconnexion de l’autopode antérieur droit, la migration de la patella gauche. Ces remaniements n’impliquent pas nécessairement la pratique d’un enfouis-sement tardif. Il en effet possible que la peau du porc ait pu résister quelques temps aux gaz de décomposition, occasionnant ainsi un espace creux dans le dépôt colmaté, et des remaniements ponctuels ultérieurs.
L’absence de quelques os, principalement celle des autopodes, peut être mise au compte du décapage méca-nique, et probablement aussi des pratiques aratoires, le décubitus dorsal facilitant l’arasement des extrémités corporelles. En revanche, l’absence de la tête ne peut être expliquée avec certitude : est-elle antérieure à l’enfouisse-ment ou résulte-t-elle d’événements ultérieurs ?
2.2.3.4 Les équidés
Le silo tronconique 5517Le silo 5517 livre 154 restes de macrofaune dont 139
déterminés, pour une masse totale de 5,7 kg. Il s’agit de 32 déchets auxquels s’ajoute un squelette d’équidé (US 14, fig. 36) mis au jour sur le fond du silo (fig. 36 et 37). La tête et l’os coxal gauche ont été endommagés par le décapage mécanique, tandis que les côtes l’ont été lors de leur prélèvement. La présence de quatre grosses canines montre qu’il s’agit d’un mâle, et l’usure de ses incisives inférieures indique un âge au décès d’environ 13 ans.
Le squelette axial et la ceinture pelvienne sont seuls représentés, le squelette appendiculaire est totalement absent (fig. 37 et 38). L’animal repose en décubitus latéral droit, dos légèrement arqué, la région coccygienne au nord-ouest, le thorax au sud-est. La tête et le cou sont en position doublement inversée, puisque d’une part ils reposent sur leurs côtés gauches, alors qu’il devrait s’agir du droit (comme le reste du squelette), et que d’autre part la région occipitale se situe au débouché du gril costal, à l’emplacement que devraient occuper les dernières vertè-bres cervicales, tandis que ces dernières sont positionnées dans le prolongement du garrot.
La tête repose sur son côté gauche. La bouche est légèrement entrouverte, les deux os hyoïdes sont présents. L’atlas est connecté aux condyles occipitaux et les sept vertèbres cervicales sont connectées. Le cou est rectili-gne et l’angle qu’il compose avec la branche montante de la mandibule semble indiquer une flexion forcée. La déconnexion de cet ensemble cervical est anthropique, comme le prouvent la découverte d’une forte incision
A B
A B
St. 5517
5517-01
5517-01 : Creusement5517-02 : Limon argileux brun mêlé à du loess5517-03 : Limon noir5517-04 : Limon noir5517-05 : Limon brun5517-06 : Limon argileux brun clair et loess5517-07 : Limon beige foncé5517-08 : Limon argilo sableux et inclusions de charbon
5517-09 : Limon brun avec inclusions de charbon5517-10 : Limon brun avec inclusions de charbon5517-11 : Limon beige foncé5517-12 : Limon beige foncé5517-13 : Limon beige5517-14 : Faune (équidé)
Loess
5517-02
N
5517-14
5517-14
5517-08
5517-05
5517-07
5517-12 5517-10
5517-13
5517-04
5517-11
03 06C D
C D
0 1 m
Fig. 36 : Position stratigraphique de l’équidé du silo 5517 (M. Landolt, C. Leprovost, R. Pinguet).
Fig. 37 : Vue générale du squelette d’équidé du silo 5517 (M. Landolt).

O. Putelat et M. Landolt, La caractérisation des dépôts animaux de La Tène ancienne à Geispolsheim
—47—
Épreuves d’auteur
latéro-médiale sur la face dorsale du processus articulaire caudal droit de C5 et le fait que la C7 soit tranchée dans sa partie caudale. Les autres vertèbres sont en connexions étroites, des premières thoraciques aux dernières sacrales. De nombreuses lésions les affectent, de C3 à T4, de T10 à T18 et de L4 à L5. Elles sont d’origine inflammatoire et diversement handicapantes (ostéophytes, « becs de perro-quet », ankylose). Quelques coccygiennes, déconnectées, sont présentes. Le gril costal est aplati et ordonné. Le sternum et les ligaments intercostaux sont présents et interconnectés. Les côtes gauches sont plaquées sur un comblement terreux. L’os coxal droit est en place, plaqué sous l’animal. Les débris du coxal gauche, endommagé au décapage, sont à l’emplacement attendu. Cette partie du corps a fait l’objet d’une exposition à la chaleur prolon-gée. La face latérale de l’aile de l’ilion gauche est carboni-sée en profondeur. L’épine iliaque ventro-crâniale (l’angle de la hanche) est brûlée à cœur, carbonisée sur la face latérale, calcinée sur la face médiale. La zone située entre la crête iliaque et l’épine iliaque dorso-crâniale (angle de la croupe) est intacte.
En résumé, l’équidé du silo 5517 était un adulte âgé d’environ 13 ans souffrant de pathologies vertébrales de gravités diverses. Les seules traces anthropiques attestées concernent la section du cou. En effet, bien que la structu-re ait été assez vaste pour pouvoir recevoir l’animal entier, la tête et le cou ont été prélevés en bloc très rapidement
après sa mort, comme en atteste le maintien de connexions labiles, telles les liaisons crâne-mandibule et occipito-cervicale (Morris 2008, p. 48-50). Dans un second temps, cet ensemble a été replacé auprès de l’animal dans une position différente : la tête est dans l’alignement du reste du corps mais déposée sur le côté gauche, alors que les vertèbres cervicales sont placées perpendiculairement au corps, dans l’alignement du garrot, et reposent sur leur côté gauche.
Aucun indice de présence des membres n’a été retrouvé au sein de la structure. La simple absence de traces de découpe au niveau des coxaux ne permet pas de savoir si les membres ont été découpés ou s’ils ont été prélevés après décomposition. Toutefois, le maintien de toutes les connexions cervicales (alors que certaines sont plus labiles que celles du squelette appendiculaire) et le tho-rax aplati indiquent un enfouissement rapide de l’équidé et sa décomposition en espace colmaté. Il est donc très vraisemblable que le dépeçage de l’animal a eu lieu sur une carcasse relativement fraîche. L’ustion de l’os coxal gauche reste inexpliquée à ce stade de l’étude. Les causes du décès de cet animal encore dans la force de l’âge sont inconnues. Il est au demeurant possible que la pathologie observée sur la région du garrot en soit la cause indirecte, entraînant la réforme de ce cheval handicapé.
2.2.3.5 Les chiensLa fosse 5011 livre 432 restes, tous déterminés, pour
une masse totale de 2,2 kg. Il s’agit de trois squelettes de chiens (US 6, 7, 8) mis au jour dans le comblement de la structure (fig. 39 et 40). La chienne 5011-06 (fig. 39 au
Parties anatomiques conservéesFig. 38 : Représentation latéralisée de l’équidé 5517 (L. Paleau, squelette d’après Barone 1986).
Loess
5011-01 : Creusement5011-02 : Limon beige clair5011-03 : Limon beige clair5011-04 : Limon brun foncé
5011-05 : Limon beige clair5011-06 : Faune (chien 2)5011-07 : Faune (chien 3)5011-08 : Faune (chien 1)
0 1 m
St. 55011
5011-01
5011-07
5011-08
5011-06
5011-07
5011-025011-03 5011-03
5011-045011-045011-05
5011-08 5011-06
N
A B
A B
Fig. 39 : Position stratigraphique des squelettes de chiens de la fosse 5011 (M. Landolt).
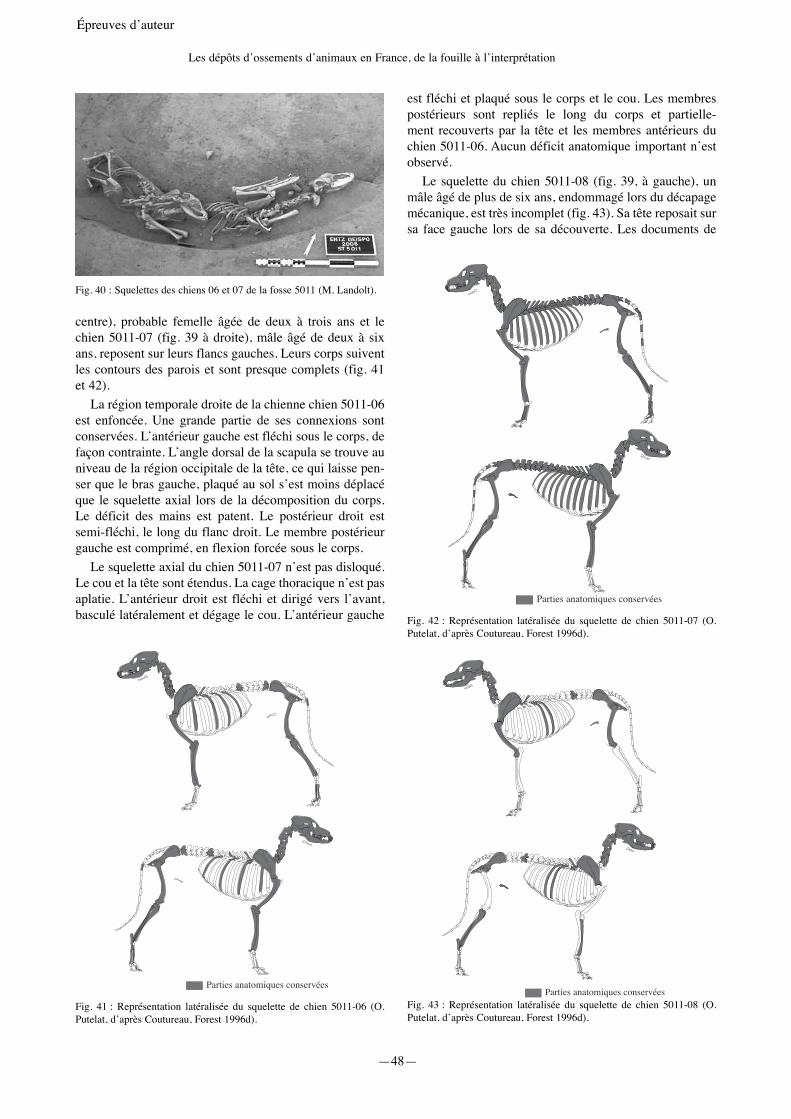
Les dépôts d’ossements d’animaux en France, de la fouille à l’interprétation
—48—
Épreuves d’auteur
centre), probable femelle âgée de deux à trois ans et le chien 5011-07 (fig. 39 à droite), mâle âgé de deux à six ans, reposent sur leurs flancs gauches. Leurs corps suivent les contours des parois et sont presque complets (fig. 41 et 42).
La région temporale droite de la chienne chien 5011-06 est enfoncée. Une grande partie de ses connexions sont conservées. L’antérieur gauche est fléchi sous le corps, de façon contrainte. L’angle dorsal de la scapula se trouve au niveau de la région occipitale de la tête, ce qui laisse pen-ser que le bras gauche, plaqué au sol s’est moins déplacé que le squelette axial lors de la décomposition du corps. Le déficit des mains est patent. Le postérieur droit est semi-fléchi, le long du flanc droit. Le membre postérieur gauche est comprimé, en flexion forcée sous le corps.
Le squelette axial du chien 5011-07 n’est pas disloqué. Le cou et la tête sont étendus. La cage thoracique n’est pas aplatie. L’antérieur droit est fléchi et dirigé vers l’avant, basculé latéralement et dégage le cou. L’antérieur gauche
est fléchi et plaqué sous le corps et le cou. Les membres postérieurs sont repliés le long du corps et partielle-ment recouverts par la tête et les membres antérieurs du chien 5011-06. Aucun déficit anatomique important n’est observé.
Le squelette du chien 5011-08 (fig. 39, à gauche), un mâle âgé de plus de six ans, endommagé lors du décapage mécanique, est très incomplet (fig. 43). Sa tête reposait sur sa face gauche lors de sa découverte. Les documents de
Fig. 40 : Squelettes des chiens 06 et 07 de la fosse 5011 (M. Landolt).
Parties anatomiques conservées
Fig. 41 : Représentation latéralisée du squelette de chien 5011-06 (O. Putelat, d’après Coutureau, Forest 1996d).
Parties anatomiques conservées
Fig. 42 : Représentation latéralisée du squelette de chien 5011-07 (O. Putelat, d’après Coutureau, Forest 1996d).
Parties anatomiques conservéesFig. 43 : Représentation latéralisée du squelette de chien 5011-08 (O. Putelat, d’après Coutureau, Forest 1996d).

O. Putelat et M. Landolt, La caractérisation des dépôts animaux de La Tène ancienne à Geispolsheim
—49—
Épreuves d’auteur
terrain montrent que les ossements fouillés manuellement, non perturbés par le décapage mécanique, sont éparpillés. Le squelette a donc probablement été, au moins partielle-ment, disloqué avant son enfouissement.
En résumé, le mâle 5011-07 a été déposé le premier, puis recouvert partiellement par la femelle 5011-06. Le mâle 5011-08 a été déposé en dernier et son squelette n’a probablement pas été recouvert très rapidement, puisqu’il est éparpillé. Aucune trace anthropique n’a été relevée qui puisse indiquer les causes du décès de ces animaux mais la relative simultanéité des dépôts peut faire envisager que ces décès soient dus à la main de l’homme. L’état des crânes pourrait indiquer l’assommage des chiens, mais ce point reste délicat, les parois des têtes ayant pu s’enfoncer naturellement sous le poids des sédiments. La coupe de la fosse (fig. 39) montre que les chiens ont été déposés sur le cône de sédiment provenant de l’effondrement du bord supérieur. La disposition des corps le long de la paroi, les compressions des membres, sont donc la traduction de contraintes spatiales et altimétriques. Un certain ordon-nancement a présidé au dépôt, puisque les corps ne sont pas enchevêtrés et qu’ils respectent une même direction et un même mode de dépôt en décubitus latéral gauche.
Le silo en bouteille 5054Le silo 5054 livre 523 restes, dont 160 déterminés,
pour une masse totale de 6,6 kg. Il s’agit de 371 déchets osseux auxquels s’ajoute un squelette de chiot, âgé de un à quatre mois (152 restes).
La position stratigraphique et les connexions anatomi-ques ne sont pas documentées. Aucune trace anthropique n’a été observée sur les ossements de ce squelette bien représenté (fig. 44).
Le silo tronconique 5236 Le silo 5236 livre 167 restes, dont 160 déterminés,
pour une masse totale de 1,2 kg. Il s’agit de quelques déchets auxquels s’ajoute un squelette de chien (US 8) mis au jour dans le comblement du silo (fig. 45 et 46). Une couche de fragments d’argile rubéfiée (US 6) est plaquée directement sur le cadavre.
Il s’agit d’un squelette complet (fig. 47), endommagé lors de la fouille de la moitié nord de la structure. Une partie des ossements exhumés à cette occasion a pu être récoltée en vrac et conservée. La morphologie de l’os pubien indique qu’il s’agit d’une femelle. Les données odonto-ostéologiques permettent d’estimer l’âge de l’ani-mal à plus de quatre ans.
L’animal repose en décubitus latéral gauche, tête au sud-est, bassin au nord-ouest. Le thorax est aplati. Les connexions des quatre membres sont étroites. Les pattes antérieures sont parallèles, légèrement fléchies et perpen-diculaires à l’axe du corps. La main gauche repose sur la
Parties anatomiques conservées
Fig. 44 : Représentation latéralisée du squelette de chien 5054 (O. Putelat, d’après Coutureau, Forest 1996d).
N
A B
A B
C D
St. 5236
5236-01 Loess
5236-02
5236-01 : Creusement5236-02 : Limon sableux brun5236-03 : Loess brun5236-04 : Limon sableux brun mélangé à du loess 5236-05 : Limon sableux brun5236-06 : Torchis fragmentés en position de rejet recouvrant le chien5236-07 : Limon sableux brun5236-08 : Faune (chien)
C D
5236-045236-05
5236-03
5236-065236-07
5236-08
5236-08
0 1 m
Fig. 45 : Position stratigraphique du squelette de chien du silo 5236 (M. Landolt, C. Leprovost, R. Pinguet).

Les dépôts d’ossements d’animaux en France, de la fouille à l’interprétation
—50—
Épreuves d’auteur
main droite ; les métapodes et les phalanges sont présents. Les pattes postérieures sont perpendiculaires à l’axe vertébral. La postérieure gauche est légèrement repliée vers l’abdomen, vers l’est. Le membre postérieur droit est étendu. Il n’a pas été observé de traces anthropiques anciennes. Quelques petits os du carpe, du tarse, ainsi qu’environ la moitié des phalanges manquent suite à la fouille. Ce chien s’est décomposé en espace colmaté et a été enfoui rapidement après son dépôt, comme l’indiquent la conservation des surfaces osseuses et la prévalence des connexions strictes.
Le silo tronconique 5413Le silo 5413 livre 110 restes dont 101 sont déterminés,
pour une masse totale de 0,6 kg. Il s’agit de onze déchets auxquels s’ajoute un squelette de chien quasi complet, mis au jour sur le fond du silo (fig. 48). Ce squelette, mal conservé, partiellement disloqué (fig. 49) et légèrement
Fig. 46 : Squelette de chien du silo 5236 (M. Landolt).
Parties anatomiques conservées
Fig. 47 : Représentation latéralisée du squelette de chien du silo 5236 (L. Paleau, squelette d’après Coutureau, Forest 1996d).
A B
A B
St. 5413
5413-01
5413-01 : Creusement5413-02 : Limon argileux brun 5413-03 : Limon brun clair 5413-04 : Limon brun clair5413-05 : Limon argileux brun avec inclusions de charbon 5413-06 : Faune (chien)
Loess
5413-02
N
5413-05
5413-045413-03
5413-06
C D
5413-06
C D
0 1 m
Fig. 48 : Position stratigraphique du squelette de chien du silo 5413 (M. Landolt, C. Leprovost, R. Pinguet).
Fig. 49 : Vue générale du silo 5413 avec le squelette de chien (M. Landolt).

O. Putelat et M. Landolt, La caractérisation des dépôts animaux de La Tène ancienne à Geispolsheim
—51—
Épreuves d’auteur
incomplet (fig. 50), provient d’un chiot âgé de trois à cinq mois. L’animal repose en décubitus latéral droit. Le ventre et les pattes, repliées sous le corps, sont plaqués contre la paroi orientale de la fosse. La plupart des connexions observables sont très lâches. Le crâne est bien conservé, ce qui tend à exclure l’hypothèse d’un assommage. Les mandibules sont déplacées par les phénomènes de bas-culement qui ont affecté la région de la tête. La colonne vertébrale est en connexion, de l’atlas aux dernières lom-baires, mais l’agencement de quelques vertèbres cervica-les et thoraciques est perturbé et des vertèbres manquent à l’étude. Les côtes sont repoussées en direction caudale. Le remplissage du gril costal est terreux. Les membres antérieurs sont plaqués sous le thorax. Le fémur droit est en place, mais le reste du membre postérieur droit est déplacé d’au moins 10 cm. Le membre postérieur gauche est allongé sous le ventre du chien, en direction caudo-crâniale, plaqué contre le bord du silo. Les autopodes, disloqués et très proches les un des autres, voire superpo-sés, sont mélangés.
Les remaniements anatomiques concernent la tête, les parties cervicales et thoraciques du squelette axial, les parties les plus proximales des deux membres antérieurs et le postérieur droit (tibia, fibula et autopode). Ces modi-fications spatiales peuvent résulter de déplacements de la tête, du cou et du garrot. La vue en coupe de la structure (fig. 48) montre d’ailleurs que l’US 5 qui scelle le sque-lette à la forme d’un cône d’effondrement. Il est donc très envisageable que ces diverses anomalies de position-nement soient consécutives à une décomposition à l’air libre, suivie de l’effondrement des parois du silo.
2.2.3.6 Les lièvresLes silos tronconique 5008, ovoïde 5017 et bitronco-
nique 5224Chacun des silos 5008, 5017 et 5224 a livré un sque-
lette de lièvre. Ces animaux sont jeunes : l’absence de synostose coxale indique qu’il s’agit de levrauts âgés de moins de six mois (Habermehl 1985). Bien qu’aucun des ossements ne soit épiphysé, la détermination du lièvre est confirmée au vu des critères de C. Callou (1997), appli-qués aux mandibules, humérus, radius, ulnas.
Le silo 5008 a livré 104 restes animaux, dont 71 restes de lièvre (US 5), pour une masse totale de 0,2 kg. Le silo 5017 a livré 68 restes animaux, dont 11 restes d’un même bufonidé, et 41 restes de levraut (US 2), pour une masse totale de 0,4 kg. Dans les deux cas, les squelettes de lièvres se présentent sous la forme d’un amoncellement épars d’ossements au fond du silo (fig. 51-52 et 53-54). Quelques connexions sont encore visibles, de façon plus
Parties anatomiques conservées
Fig. 50 : Représentation latéralisée du squelette de chien du silo 5413 (L. Paleau, squelette d’après Coutureau, Forest 1996d).
St. 5008
Loess
5008-02
5008-04
5008-03
5008-05
5008-01
0 1 m
A B
A B
C D
C D
5008-05
N
5008-01 : Creusement5008-02 : Limon sableux brun5008-03 : Limon jaune à brun5008-04 : Limon sableux brun5008-05 : Faune (lièvre)
Fig. 51 : Position stratigraphique du squelette de lièvre du silo 5008 (M. Landolt).

Les dépôts d’ossements d’animaux en France, de la fouille à l’interprétation
—52—
Épreuves d’auteur
marquée dans le silo 5008 que dans la structure 5017. Ces individus se sont décomposés sur place à l’air libre, en espace protégé, avant l’effondrement des parois. Dans le silo 5017, l’US 9 qui contient le squelette de lièvre livre aussi des éléments d’un squelette de bufonidé, probable-ment victime d’un piégeage naturel. Les levrauts sont relativement complets eu égard à la petite taille de leurs ossements (fig. 55 et 56), mais la tête du lièvre 5008 est absente.
Fig. 52 : Lièvre déposé au fond du silo 5008 (M. Landolt).
St. 5017
Loess
5017-08
5017-09
5017-065017-07
5017-05
5017-04
5017-11
5017-03
5017-10
5017-02
5017-02
5017-01
0 1 m
N
A B
C D
C D
A B
5017-01 : Creusement5017-02 : Faune (lièvre)5017-03 : Limon brun clair5017-04 : Loess jaune5017-05 : Loess jaune5017-06 : Loess jaune
5017-07 : Argile et sable brun à jaune5017-08 : Argile, sable et loess brun à jaune5017-09 : Argile, sable et cendre noire5017-10 : Argile et sable brun clair5017-11 : Argile et sable et loess brun à jaune
Fig. 53 : Position stratigraphique du squelette de lièvre du silo 5017 (M. Landolt).
Fig. 54 : Lièvre déposé au fond du silo 5017 (M. Landolt).
Parties anatomiques conservées
Fig. 55 : Représentation latéralisée du squelette de lièvre du silo 5008 (O. Putelat, squelette d’après Coutureau, Forest 1996b, squelette de lapin).
O. Putelat et M. Landolt, La caractérisation des dépôts animaux de La Tène ancienne à Geispolsheim
—53—
Épreuves d’auteur
Le squelette de lièvre 5224 (93 restes) a été mis au jour dans l’US 02, première US de comblement du silo. Les connexions anatomiques ne sont pas documentées, l’assemblage osseux n’ayant pas été identifié en tant que squelette à la fouille. Nous supposons qu’il se présentait sous la forme d’un amoncellement épars d’ossements, comme les levrauts 5008 et 5017. Sa représentation ana-tomique montre la présence d’un squelette quasi complet (fig. 57), où les manques sont imputables aux phénomènes taphonomiques, dont la fouille.
Le silo cylindrique 5553Le silo 5553 livre un squelette humain et 183 restes de
macrofaune, dont 178 déterminés, pour une masse totale de 540 g. Il s’agit de quelques déchets, mais aussi d’un squelette de lièvre (164 restes) déposé à l’emplacement initial de la jambe et du pied gauches du défunt, le long de la bordure est du silo (Pélissier in Landolt et al. 2013). A cette macrofaune s’ajoutent 157 restes de microfaune7.
L’animal (US 9) provient de l’US 2, cône de comble-ment du silo, où il occupe l’emplacement initial de la jambe et du pied gauches du défunt (fig. 58, 59, 60).
7 La microfaune est exclusivement composée d’ossements d’amphibiens, prélevés en amas. Ces vestiges n’ayant pas, à ce jour, fait l’objet d’une étude détaillée, nous limiterons à observer que toutes les grandes régions du squelette sont représentées, provenant d’au moins trois individus. Il est possible que ces amphibiens aient été piégés dans le silo, resté ouvert durant la première phase de décomposition du squelette humain.
Parties anatomiques conservées
Fig. 56 : Représentation latéralisée du squelette de lièvre du silo 5017 (O. Putelat, squelette d’après Coutureau, Forest 1996b, squelette de lapin).
Parties anatomiques conservées
Fig. 57 : Représentation latéralisée du squelette de lièvre du silo 5224 (L. Paleau, squelette d’après Coutureau, Forest 1996b, squelette de lapin).
N
A B
A B
C D
C D
St. 5553
5553-01 Loess
5553-02
5553-01 : Creusement5553-02 : Limon brun foncé5553-03 : Limon brun mêlé à du loess5553-04 : Limon brun5553-05 : Limon brun clair
5553-06 : Limon brun05223-07 : Limon brun mêlé à du loess05223-08 : Squelette humain05223-09 : Faune (lièvre)
5553-035553-04
5553-07
5553-05 5553-06
5553-08
5553-08
5553-09
0 1 m
Fig. 58 : Position stratigraphique du squelette de lièvre, avec le défunt du silo 5553 (M. Landolt, C. Leprovost, R. Pinguet).
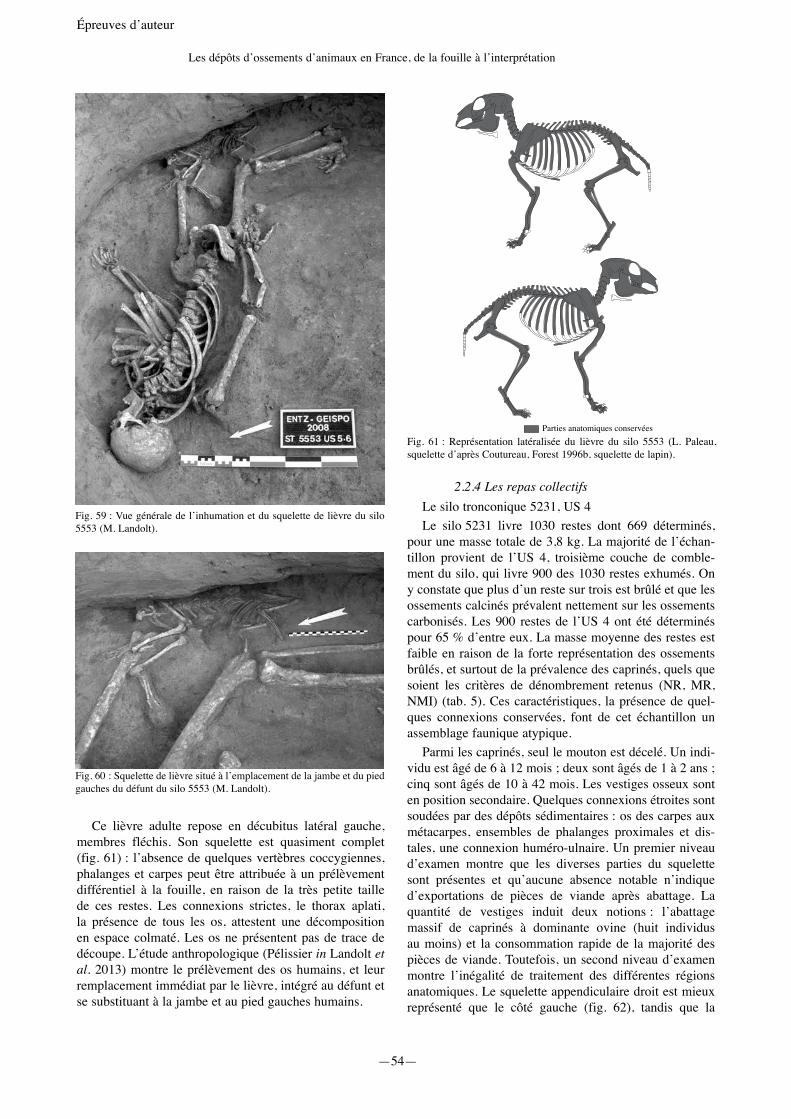
Les dépôts d’ossements d’animaux en France, de la fouille à l’interprétation
—54—
Épreuves d’auteur
Ce lièvre adulte repose en décubitus latéral gauche, membres fléchis. Son squelette est quasiment complet (fig. 61) : l’absence de quelques vertèbres coccygiennes, phalanges et carpes peut être attribuée à un prélèvement différentiel à la fouille, en raison de la très petite taille de ces restes. Les connexions strictes, le thorax aplati, la présence de tous les os, attestent une décomposition en espace colmaté. Les os ne présentent pas de trace de découpe. L’étude anthropologique (Pélissier in Landolt et al. 2013) montre le prélèvement des os humains, et leur remplacement immédiat par le lièvre, intégré au défunt et se substituant à la jambe et au pied gauches humains.
2.2.4 Les repas collectifsLe silo tronconique 5231, US 4Le silo 5231 livre 1030 restes dont 669 déterminés,
pour une masse totale de 3,8 kg. La majorité de l’échan-tillon provient de l’US 4, troisième couche de comble-ment du silo, qui livre 900 des 1030 restes exhumés. On y constate que plus d’un reste sur trois est brûlé et que les ossements calcinés prévalent nettement sur les ossements carbonisés. Les 900 restes de l’US 4 ont été déterminés pour 65 % d’entre eux. La masse moyenne des restes est faible en raison de la forte représentation des ossements brûlés, et surtout de la prévalence des caprinés, quels que soient les critères de dénombrement retenus (NR, MR, NMI) (tab. 5). Ces caractéristiques, la présence de quel-ques connexions conservées, font de cet échantillon un assemblage faunique atypique.
Parmi les caprinés, seul le mouton est décelé. Un indi-vidu est âgé de 6 à 12 mois ; deux sont âgés de 1 à 2 ans ; cinq sont âgés de 10 à 42 mois. Les vestiges osseux sont en position secondaire. Quelques connexions étroites sont soudées par des dépôts sédimentaires : os des carpes aux métacarpes, ensembles de phalanges proximales et dis-tales, une connexion huméro-ulnaire. Un premier niveau d’examen montre que les diverses parties du squelette sont présentes et qu’aucune absence notable n’indique d’exportations de pièces de viande après abattage. La quantité de vestiges induit deux notions : l’abattage massif de caprinés à dominante ovine (huit individus au moins) et la consommation rapide de la majorité des pièces de viande. Toutefois, un second niveau d’examen montre l’inégalité de traitement des différentes régions anatomiques. Le squelette appendiculaire droit est mieux représenté que le côté gauche (fig. 62), tandis que la
Fig. 59 : Vue générale de l’inhumation et du squelette de lièvre du silo 5553 (M. Landolt).
Fig. 60 : Squelette de lièvre situé à l’emplacement de la jambe et du pied gauches du défunt du silo 5553 (M. Landolt).
Parties anatomiques conservéesFig. 61 : Représentation latéralisée du lièvre du silo 5553 (L. Paleau, squelette d’après Coutureau, Forest 1996b, squelette de lapin).

O. Putelat et M. Landolt, La caractérisation des dépôts animaux de La Tène ancienne à Geispolsheim
—55—
Épreuves d’auteur
ceinture pelvienne et les membres postérieurs sont plus touchés par les phénomènes d’ustion que les membres antérieurs. Les traces anthropiques sont de deux sortes. Des traces de découpe concernent essentiellement les crânes (fente sagittale de trois crânes) et les vertèbres (tronçonnées transversalement, parfois fendues sagitta-lement). Des traces de désossement affectent plus rare-ment des vertèbres cervicales et lombaires et des côtes. S’il est indéniable que les concrétions sédimentaires et l’ustion des vestiges occultent une partie du processus de consommation des viandes, il n’en reste pas moins que la bonne représentation d’os longs non fragmentés et non brûlés du squelette appendiculaire antérieur (par exemple six scapulas, six humérus, cinq radius) contraste avec le manque d’observations quant aux traces de découpe sur ces mêmes segments anatomiques.
En résumé, l’échantillon faunique de l’US 4 peut être défini comme étant une concentration très dense d’osse-ments de caprinés, très bien conservés, rejetés au contact les uns des autres, résultant de l’abattage massif et de l’utilisation rapide d’au moins huit caprinés. L’hypothèse d’un repas collectif, au centre duquel le mouton tiendrait la place principale, doit être évoquée. Cependant, l’iné-galité de traitement des diverses parties des squelettes, la rareté des informations quant à la consommation des vian-des, à l’exception du squelette axial où elle est attestée sur des vertèbres et des côtes, nous font émettre l’hypothèse que puissent se côtoyer ici rejets alimentaires et dépôts de pièces de viande non consommées.
Le silo en bouteille 5268, US 9 et 10Le silo 5268 livre 334 restes de macrofaune (dont 280
déterminés), pour une masse totale de 2,3 kg. Des US 9 et 10, situées à la base du silo (fig. 63), provient la plus grande partie des vestiges (245 restes). Il s’agit là de rejets culinaires, auxquels s’ajoute un petit dépôt constitué de mandibules et de scapulas ovines.
Le matériel osseux des US 9-10 se présente sous la forme d’un amas allongé, de couleur brun sombre, aux contours irréguliers mais bien délimités. L’excellente conservation des restes, déterminés pour 82 % d’entre eux, indique leur recouvrement relativement rapide par
Espèces N.R. % NR NMIc Masse (g) % masse Bœuf : Bos taurus L. 5 0,8% 1 254,3 8,5%Mouton : Ovis aries L. 99 6Caprinés 481 2Porc : Sus domesticus E. et suinés 2 0,3% 1 28,8 1,0%Total des animaux domestiques 587 99,7% 10 2729,4 91,0%Oiseau non différencié 1 0,2% 1 0,9 0,0%Total des animaux sauvages 1 0,2% 1 0,9 0,0%Poisson 1 0,2% 1 0,1 0,0%Nombre de restes déterminés 589 100,0% 12 2730,4 91,0%Mammifères de taille moyenne 309 268,6 9,0%Esquilles indéterminées 2 1,8 0,1%Total 900 12 3000,8 100,0%
2446,3 81,5%98,5%
Tab. 5 : Spectre faunique du silo 5231, US 4 (O. Putelat).
6 5 4 3 2 1 0 1 2 3 4 5 6 7
MétatarseMétacarpe
TarseTibia FémurCoxalCarpe
Radius-UlnaHumérusScapula
CervicalesMandibuleMaxillaire
Nombre minimal de segments
Fig. 62 : Silo 5231, US 4. Mise en évidence du NMI de fréquence des caprinés à partir de la représentation latéralisée des nombres minimum d’éléments anatomiques les plus caractéristiques (O. Putelat).
N
A B
A B
St. 5268
5268-01
5268-02
5268-01 : Creusement5268-02 : Limon brun foncé à noir 5268-03 : Loess mélangé à du limon beige5268-04 : Loess mélangé à du limon beige5268-05 : Limon brun5268-06 : Limon cendreux gris foncé à noir5268-07 : Limon brun5268-08 : Limon cendreux gris foncé5268-09 : Limon argileux brun5268-10 : Faune (ovin)
Loess
5268-05
5268-06
5268-07
5268-085268-09
5268-0304
C D
C D
5268-10
0 1 m
Ovin sans cornesBrebis cornue
5268-10
Fig. 63 : Position stratigraphique des ossements animaux du silo 5268 (M. Landolt, C. Leprovost, R. Pinguet).

Les dépôts d’ossements d’animaux en France, de la fouille à l’interprétation
—56—
Épreuves d’auteur
un sédiment lœssique fin, physiquement et chimiquement non agressif. Il s’agit de rejets alimentaires, comme en attestent de nombreuses traces de découpe et des brûlures de cuisson (infra). De rares connexions sont conservées et ne concernent que quelques vertèbres. Le halo brun-orga-nique très conséquent qui entoure les vestiges fauniques ne peut donc pas être imputé à la décomposition de sque-lettes animaux. Le spectre faunique est caractérisé par la prévalence des caprinés, quels que soient les critères de dénombrement (tab. 6).
Chez les caprinés, où seul le mouton est différencié, les parties du squelette sont représentées de façon assez homogène (fig. 64) et il n’a pas été observé de représenta-tion latéralisée préférentielle. Deux têtes osseuses ovines, fendues sagittalement, mais quasiment complètes, sont mises en évidence par juxtaposition des surfaces osseuses conservées. Il s’agit d’une brebis cornue et d’une brebis motte (acère). L’examen des dents jugales indique des âges d’abattage semblables pour ces deux individus : 4 à 6 ans. Deux brebis sont également sexuées au vu de la forme de leurs os pubiens. L’homogénéité macroscopique de l’échantillon, le fait que les âges osseux confirment systématiquement les âges dentaires des deux ovins, rend très vraisemblable que ces deux brebis soient à l’origine de
la plus grande partie de l’assemblage osseux, bien qu’un fragment métatarsien surnuméraire indique la présence discrète d’un troisième capriné. Les traces de découpe et de cuisson sur les os de caprinés sont nombreuses et variées. Les têtes osseuses sont fendues, et probablement grillées comme en attestent les surfaces osseuses des frontaux. Les diverses parties du squelette montrent des traces de désarticulation, de désossement, de dépeçage. Il faut remarquer que, dans ce contexte culinaire, le bris des métapodes n’est pas systématique, puisque quatre des sept métapodes présents sont intacts.
Lors du prélèvement du matériel osseux de l’US 10, au désordre des ossements (fig. 65), s’opposait l’ordre de deux ensembles osseux, que nous attribuons à un agence-ment anthropique. Au nord-est du dépôt, deux scapulas provenant d’un même ovin sont disposées « tête-bêche » (fig. 65, ellipse blanche et fig. 66). La gauche (fig. 65, à gauche dans l’ellipse blanche), repose sur sa face médiale, angle dorsal à l’ouest. La droite (fig. 65, à droite dans l’ellipse blanche) repose sur sa face latérale, angle dorsal à l’est. Ces deux scapulas cachent la mandibule gauche de la brebis cornue déjà mentionnée.
Espèces N.R. % NR NMIc Masse (g) % masseBœuf : Bos taurus L. 10 5,0% 1 81,2 5,8%Mouton : Ovis aries L. 41 2Caprinés 149 1Chien : Canis familiaris L. 1 0,5% 1 12,0 0,9%Total des animaux domestiques 201 100,0% 5 1339,6 95,5%Total des animaux sauvages 0 0,0% 0 0,0 0,0%Nombre de restes déterminés 201 100,0% 5 1339,6 95,5%Grands mammifères 1 14,4 1,0%Mammifères de taille moyenne 40 45,3 3,2%Esquilles indéterminées 3 3,8 0,3%Total 245 5 1403,1 100,0%Amphibiens 10 2 2,4
1246,4 88,8%94,5%
Tab. 6 : Spectre de faune des US 9-10 du silo 5268 (O. Putelat).
NME droits, gauches et non latéralisés
0 1 2 3 4 5
Métatarse
Métacarpe
Tarse
Tibia
Fémur
Coxal
Carpe
Radius-Ulna
Humérus
Scapula
Cervicales
Mandibule
Crâne
Nombre minimal de segments
5268 US 9-10
Fig. 64 : Silo 5268, US 9-10. Mise en évidence du NMI de fréquence des caprinés à partir des nombres minimum d’éléments anatomiques les plus caractéristiques (O. Putelat).
Fig. 65 : Matériel osseux de l’US 10 dans la partie nord-ouest du silo 5268 (M. Landolt).
Fig. 66 : Silo 5268, US 10. Vue de détail de la mandibule gauche de la brebis cornue, recouverte par deux scapulas hétérolatérales (M. Landolt).

O. Putelat et M. Landolt, La caractérisation des dépôts animaux de La Tène ancienne à Geispolsheim
—57—
Épreuves d’auteur
Au centre du dépôt, deux mandibules hétérolatérales, strictement parallèles et distantes l’une de l’autre d’une quinzaine de centimètres, suivent un axe nord-est/sud-ouest, parties incisives vers le sud-ouest (fig. 65, ellipse blanche). A l’est, l’une (fig. 65, en haut dans l’ellipse blanche) repose sur sa face latérale. Il s’agit de la mandi-bule gauche de la brebis motte déjà citée. L’autre (fig. 65, en bas dans l’ellipse blanche) repose sur sa face médiale. Il s’agit de la mandibule droite de la brebis cornue. Sans que la finalité du geste nous apparaisse clairement, et sans en exagérer la portée, il nous semble qu’a eu lieu un agencement anthropique de quelques détritus culinaires au moment de leur rejet ou après. Trois hémi-mandibu-les et deux scapulas ont été dissociées des déchets, puis positionnées assez soigneusement. Le positionnement des scapulas tête-bêche, cachant parfaitement une mandibule, dont la présence renvoie elle-même au positionnement de deux autres mandibules parallèles au centre du dépôt (livrées par deux ovins différents) nous semble bien diffi-cile à imputer aux seuls effets du hasard.
Le silo tronconique 5316 Le silo 5316 livre 359 restes de macrofaune, dont 228
déterminés, pour une masse totale de 1,9 kg. Les espèces sont domestiques et les caprinés prévalent. La majorité de l’échantillon provient de l’US 4, première couche de comblement du silo, qui livre 240 des 359 restes. On constate que dans cette US près des trois-quarts des ossements sont carbonisés et que les caprinés sont le seul taxon déterminé. Ces deux observations, la présence de connexions conservées, font de l’US 4 un assemblage faunique atypique.
Le mouton et la chèvre sont tous deux mis en évidence (tab. 7). Les données odonto-ostéologiques montrent la présence de deux caprinés (dont un ovin) âgés respective-ment de 2 à 4 ans et 4 à 6 ans. La chèvre (stricto sensu) est âgée de plus de 3 ans. Les vestiges osseux sont en position secondaire. Quelques connexions étroites sont conservées par des dépôts sédimentaires : deux vertèbres thoraciques, deux vertèbres lombaires, un autopode ovin (carpe, métacarpe, phalanges proximales). La proportion de restes carbonisés est très importante. La majorité des brûlures résultent du dépôt/rejet au feu des ossements. Destructrices, elles masquent traces de découpe et brûlu-res de cuisson. Nous n’avons pas observé de différence de traitement dans l’exposition au feu entre les membres antérieurs et postérieurs. Les diverses parties du sque-lette sont mises en évidence, mais un déficit affecte les scapulas et les autopodes (fig. 67). Les régions du corps
les mieux représentées sont la tête (toujours fendue sagittalement) et les parties charnues des membres. Nous n’avons pas constaté de dissymétrie dans la représentation anatomique, à l’exception des radius-ulnas (un droit, trois gauches). L’exclusivité taxonomique des vestiges, leur répartition anatomique, montrent l’abattage et la consom-mation de la majorité des pièces de viande de plusieurs caprinés, dans un bref laps de temps. L’hypothèse que l’assemblage osseux résulte de la pratique d’un repas col-lectif est envisageable.
Le silo tronconique 5348Le silo 5348 livre au total 542 restes osseux, pour une
masse de 5,0 kg. Plus de la moitié des 347 restes déter-minés, exhumés pour l’essentiel au diagnostic de 2007 (structure 186) (fig. 68), mais aussi lors des fouilles de 2008, proviennent d’un même bovin. La fouille de 2008 en a précisé le rattachement stratigraphique à l’US 2 (première couche de comblement du silo). Le spectre de faune de ce silo est présenté tab. 8. Le bœuf y prévaut, mais les ossements de caprinés sont néanmoins très bien représentés.
Aucune connexion anatomique ne lie les ossements du bovin 5348. Cependant, l’homogénéité morphologique des restes, la concordance des estimations d’âges réalisées sur différentes parties du squelette, montrent que l’échan-
Espèces N.R. % NR NMI Masse (g) % masse Mouton : Ovis aries L. 17 1Chèvre : Capra hircus L. 2 1Caprinés 124 1Nombre de restes déterminés 143 100,0% 3 1122,9 91,5%Mammifères de taille moyenne 90 85,6 7,0%Esquilles indéterminées 7 18,1 1,5%Total 240 3 1226,6 100,0%
91,5%1122,9100,0%
Tab. 7 : Spectre de faune du silo 5316, US 4 (O. Putelat).
NME droits, gauches et non latéralisés
0 1 2 3 4 5
MétatarseMétacarpe
TarseTibia FémurCoxalCarpe
Radius-UlnaHumérusScapula
MandibuleMaxillaire
Nombre minimal de segments
Fig. 67 : Silo 5316, US 4. Mise en évidence du NMI de fréquence des caprinés à partir des nombres minimum d’éléments anatomiques les plus caractéristiques (O. Putelat).
Espèces N.R. % NR NMIc Masse (g) % masseSquelette bovin 5348 190 65,3% 1 3631,3 78,5%Bœuf : Bos taurus L. 7 2,4% 1 466,0 10,1%Mouton : Ovis aries L. 31 2Chèvre : Capra hircus L. 2 1Caprinés 60Porc : Sus domesticus E. et suinés 1 0,3% 1 0,8 0,0%Total des animaux domestiques 291 100,0% 6 4547,9 98,3%Total des animaux sauvages 0 0,0% 0 0,0 0,0%Nombre de restes déterminés 291 100,0% 6 4547,9 98,3%Grands mammifères 2 13,7 0,3%Mammifères de taille moyenne 73 36,1 0,8%Esquilles indéterminées 44 31,0 0,7%Total 410 6 4628,7 100,0%Mollusque 1,0 1 0,8
449,8 9,7%32,0%
Tab. 8 : Spectre de faune du silo 5348 : faune du diagnostic et US 2 de la fouille (O. Putelat).

Les dépôts d’ossements d’animaux en France, de la fouille à l’interprétation
—58—
Épreuves d’auteur
tillon provient d’un même mâle, âgé d’environ 18 à 24 mois. Le bovin a été consommé (comme en attestent de multiples traces anthropiques et des brûlures d’extrémi-tés), puis dispersé sur le fond du silo 5348. Son squelette est incomplet : les lacunes les plus importantes concernent la colonne vertébrale, le gril costal, les autopodes anté-rieur gauche et postérieur droit (fig. 69).
Parmi les autres déchets, plusieurs restes proviennent d’un second bovin, d’un gabarit beaucoup plus important que le bovin 5348.
Les ossements de caprinés sont nombreux et méritent d’être exposés en détail, pour les mêmes raisons que ceux du bovin 5348, puisqu’ils participent du rejet simultané d’une grande quantité de restes culinaires. Le mouton prévaut, mais la chèvre est mise en évidence. Deux ovins, au moins, sont présents. L’un est âgé de 6 à 12 mois et l’autre de plus de 30 mois. L’âge du caprin n’a pu être établi. Les ossements proviennent de toutes les régions du squelette. Les traces de découpe sont rares, souvent occultées par les concrétions minérales. La fragmentation des vestiges, les nombreuses brûlures d’extrémité sont toutefois caractéristiques de rejets alimentaires. Quelques connexions étroites sont documentées par la présence de segments anatomiques concrétionnés.
En résumé, le fait que la moitié de l’assemblage osseux provienne d’un même bovin prouve la simultanéité de rejets issus de l’ensemble des régions du squelette. Les parties les plus riches sont bien représentées, tant chez le bovin 5348 que parmi les deux moutons et le caprin abat-tus. Le statut alimentaire de l’échantillon est confirmé par de nombreuses traces de découpes et de cuisson. L’abattage et la consommation quasi simultanés de ces animaux sont
Parties anatomiques conservées
Fig. 69 : Représentation latéralisée du bovin du silo 5348 (L. Paleau, squelette d’après Coutureau, Forest 1996c).
Fig. 68 : Ossements du bovin et ossements divers dans le comblement du silo 5348 (F. Reutenauer).

O. Putelat et M. Landolt, La caractérisation des dépôts animaux de La Tène ancienne à Geispolsheim
—59—
Épreuves d’auteur
très probables. En l’absence de tout indice d’exportation de parties charnues du squelette, ils induisent les notions de disponibilité d’une quantité conséquente de viande et de sa consommation in situ. L’hypothèse qu’il s’agisse des reliefs d’un repas collectif d’une certaine ampleur nous paraît très vraisemblable.
2.2.5. Divers
Le silo tronconique 5449Le silo 5449 livre 100 restes dont 74 déterminés, pour
une masse totale de 0,5 kg. A cet échantillon s’ajoutent des restes d’ichtyofaune8 (fig. 70), livrés principalement par l’US 4, sixième couche de comblement du silo. De l’US 4 proviennent un reste dentaire et un os paras-phénoïde de brochet ; un os pharyngien et une vertèbre de cyprinidé, une côte partiellement carbonisée et 682 écailles de cyprinidé(s). De nombreuses écailles sont également présentes dans le sédiment prélevé (non étudié à ce jour). Le statut de ces restes qui mettent en œuvre deux taxons différents, le brochet et un cyprinidé, n’est pas établi à ce jour. S’agit-il d’un rejet culinaire ou d’un autre type de dépôt ?
8 L’ichtyofaune a été gracieusement déterminée par notre collègue Benoît Clavel (MNHN, Paris). Nous l’en remercions vivement.
3. SynthèseL’analyse archéozoologique montre la variabilité de
composition des dépôts étudiés. Elle permet aussi d’en proposer un classement. Nous avons distingué des dépôts de têtes osseuses, d’ensembles anatomiques issus du sque-lette appendiculaire, de squelettes et des dépôts issus de repas collectifs. Nous constatons que :
- à deux exceptions près, les fosses 5236 et 5674, la totalité des dépôts de têtes, de parties de squelettes ou de squelettes provient de silos ;
- ces dépôts sont le plus souvent associés à des rejets domestiques ;
- même si la faune sauvage est relativement fréquente en très petite quantité dans les silos, les dépôts sont le plus souvent composés d’espèces domestiques ;
- le seul animal sauvage représenté les dépôts de sque-lettes est le lièvre. C’est aussi le seul animal mis au jour en contexte funéraire sur le site (silo 5553) ;
- les traces anthropiques pouvant être mises en relation avec la mise à mort des animaux présents dans les dépôts sont rares ou inexistantes ;
- des cadavres ont subi des manipulations intentionnel-les et non alimentaires indiscutables ;
- d’autres cadavres peuvent résulter d’une mortalité naturelle des animaux domestiques ;
0 1 cm
1
2
3
4
A B
Fig. 70 : A gauche, amas d’écailles de poisson du silo 5449 (M. Landolt). A droite, écaille de cyprinidé (1) ; os pharyngien de cyprinidé (2) ; vertèbre de cyprinidé (3) ; côtes de cyprinidé (4) (I. Dechanez-Clerc).

Les dépôts d’ossements d’animaux en France, de la fouille à l’interprétation
—60—
Épreuves d’auteur
- certains dépôts de têtes osseuses ou d’ensembles ana-tomiques issus du squelette appendiculaire peuvent être des « dépôts particuliers », c’est-à-dire des restes témoi-gnant de pratiques rituelles domestiques (d’après Auxiette et al. 2010, p. 193) ;
- d’autres peuvent aussi bien être des rejets alimentaires (silos 5149 et 5196) ou des rejets non consommables.
Ces assemblages osseux sont regroupés et synthétisés (tab. 9). En excluant les dépôts douteux des silos 5149 et 5196, et en envisageant la possibilité que les parties de squelette appendiculaire puissent correspondre à l’un des quatre chevaux révélés par leurs têtes, les dépôts de têtes osseuses, d’ensembles anatomiques issus du squelette appendiculaire et de squelettes mettent en jeu au moins vingt-quatre individus différents, tandis que les restes de repas collectifs correspondent à au moins quatorze indivi-dus consommés (tab. 9). Sur un plan spatial, ils se regrou-pent en trois grandes zones au sein du site (fig. 71).
3.1. Les têtes osseusesLa tête de porc complète du silo 5196 constituerait un
« dépôt particulier » très convaincant au vu de l’icono-graphie (fig. 5). Cependant, la présence d’autres restes alimentaires de porc et de fragments de têtes d’autres mammifères laisse penser qu’il s’agit d’un « effet de fouille » qui ne peut être attribué avec certitude à un dépôt particulier.
Le silo 5149 a livré les restes des têtes d’un bœuf et d’un équidé, associées à des ossements de chien marqués de traces de boucherie. L’état de conservation de ces têtes montre qu’elles sont probablement restées à l’air libre avant leur enfouissement. L’hétérogénéité de l’assembla-ge osseux, son éloignement des trois aires riches en dépôts (fig. 71) et sa proximité d’une zone non fouillée rendent trop incertaine son attribution à un dépôt particulier.
Au contraire, les trois têtes d’équidés du silo 5450, soigneusement ordonnancées, et dont deux semblent avoir été exposées quelques temps à l’air libre, s’inscrivent dans la logique des dépôts particuliers. En Alsace, à La Tène B à Colmar « Jardin des Aubépines » une tête de cheval a été retrouvée dans le silo 339, associée à l’inhumation d’un humain périnatal9 (Roth-Zehner et al. 2008, p. 116). Sur le même site, un dépôt d’équidé acéphale est mentionné dans le silo 235 (Ibid., p. 130).
3.2. Les ensembles anatomiques issus du squelette appendiculaire
Les caractéristiques des différents dépôts d’ensembles anatomiques issus du squelette appendiculaire (accompa-gnés de la tête dans le cas de l’ovin du silo 5219) ont été discutées lors de leurs présentations. Il reste difficile de
9 Pour ce silo, les archéologues ne concluent pas à un dépôt « rituel » en raison du contexte détritique du dépôt.
différencier pour ces ensembles ce qui ressort du « dépôt particulier », du rejet de détritus, ou encore du dépôt de peaux dont la finalité nous échappe. L’interprétation de cette catégorie de dépôts reste subjective. On voit par exemple que dans le Haut-Rhin, au Hallstatt D3 à Wolfgantzen « Niederfeld » (fig. 72), une patte de cheval en connexion est interprétée comme un rejet de découpe (Kuhnle et al. 2000, p. 151). A Geispolsheim « Schwobenfeld », les principaux arguments pour voir des dépôts particuliers dans ce type d’ensembles anatomiques seraient l’ordonnancement éventuel de certains lots, la présence d’ossements entiers beaucoup plus rares dans les rejets que dans les dépôts supposés, mais surtout l’inser-tion de ces assemblages dans l’une des trois aires riches en dépôts (fig. 71). Le membre postérieur d’équidé du silo 5199 est toutefois extérieur à ces aires.
3.3. Les squelettesLe bovin du silo 5302 s’est décomposé à l’air libre au
fond du silo, puis a été remanié par le prélèvement d’os-sements. La finalité de l’effet d’ordonnancement observé reste inconnue : est-elle volontaire ou n’est-elle qu’un effet secondaire du prélèvement des os ?
Pour La Tène ancienne en Alsace, en l’état actuel de la recherche, et dans l’attente de la publication du site de Colmar « Jardin des Aubépines », ce squelette reste la seule occurrence connue de dépôt bovin.
La brebis du silo 5376 s’est décomposée à l’air libre sur le second comblement du silo. Son cadavre a été remanié dans un deuxième temps, en faisant l’objet d’une disposition intentionnelle des ossements, que l’on pourrait qualifier de « tableau ».
Dans le Haut-Rhin, d’autres squelettes de caprinés sont attestés pour La Tène ancienne (fig. 72). Le silo 235 de Colmar « Jardin des Aubépines » a livré plu-sieurs squelettes de caprinés dans un contexte attribué à La Tène B (Roth-Zehner et al. 2008, p. 119). A Illfurth « Naegelberg », le squelette d’un capriné en connexion a été identifié sur une occupation de La Tène B-C (Roth-Zehner et al. 2004, p. 20 et 26).
Concernant les porcs, la disposition des squelettes révèle d’une part des individus déposés sans soin particu-lier : le squelette « vrillé » avec pendage du silo 5176, le squelette en décubitus dorsal de la présumée fosse 5674. Le mauvais état de conservation du porc 5564 ne permet pas d’observations satisfaisantes. On observe d’autre part trois individus déposés plus soigneusement, sur le pre-mier comblement du silo 5375, sans simultanéité stricte du dépôt (le porc 3 a été recouvert d’un peu de sédiment, avant dépôt, dans un bref laps de temps, des porcs 1 et 2). L’absence de traces de mise à mort, ou l’incertitude quant aux traces observées, l’étagement des âges des décès des porcins exhumés, la simultanéité des décès dans le cas du silo 5375, pourraient constituer les indices d’une mortalité naturelle du troupeau, d’autant que quelques individus périnataux ont également été décelés lors du pré-examen des vestiges (silos 5270 et 5308). Cependant, l’absence
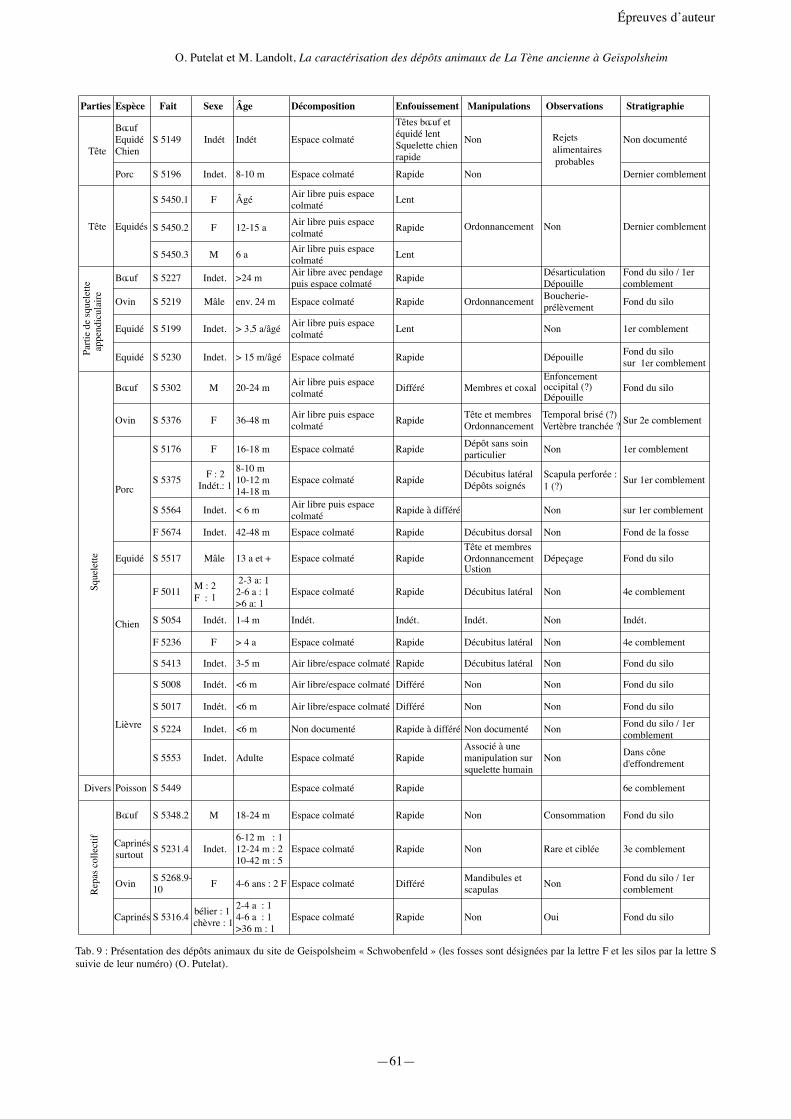
O. Putelat et M. Landolt, La caractérisation des dépôts animaux de La Tène ancienne à Geispolsheim
—61—
Épreuves d’auteur
Parties Espèce Fait Sexe Âge Décomposition Enfouissement Manipulations Observations Stratigraphie
Bœuf Equidé Chien
S 5149 Indét Indét Espace colmaté
Têtes bœuf et équidé lent Squelette chien rapide
Non Non documenté
Porc S 5196 Indet. 8-10 m Espace colmaté Rapide Non Dernier comblement
S 5450.1 F Âgé Air libre puis espace colmaté Lent
S 5450.2 F 12-15 a Air libre puis espace colmaté Rapide
S 5450.3 M 6 a Air libre puis espace colmaté Lent
Bœuf S 5227 Indet. >24 m Air libre avec pendagepuis espace colmaté Rapide Désarticulation
DépouilleFond du silo / 1er comblement
Ovin S 5219 Mâle env. 24 m Espace colmaté Rapide Ordonnancement Boucherie-prélèvement Fond du silo
Equidé S 5199 Indet. > 3,5 a/âgé Air libre puis espace colmaté Lent Non 1er comblement
Equidé S 5230 Indet. > 15 m/âgé Espace colmaté Rapide Dépouille Fond du silo sur 1er comblement
Bœuf S 5302 M 20-24 m Air libre puis espace colmaté Différé Membres et coxal
Enfoncement occipital (?) Dépouille
Fond du silo
Ovin S 5376 F 36-48 m Air libre puis espace colmaté Rapide Tête et membres
OrdonnancementTemporal brisé (?) Vertèbre tranchée ? Sur 2e comblement
S 5176 F 16-18 m Espace colmaté Rapide Dépôt sans soin particulier Non 1er comblement
S 5375 F : 2 Indét.: 1
8-10 m 10-12 m 14-18 m
Espace colmaté Rapide Décubitus latéral Dépôts soignés
Scapula perforée :1 (?) Sur 1er comblement
S 5564 Indet. < 6 m Air libre puis espace colmaté Rapide à différé Non sur 1er comblement
F 5674 Indet. 42-48 m Espace colmaté Rapide Décubitus dorsal Non Fond de la fosse
Equidé S 5517 Mâle 13 a et + Espace colmaté RapideTête et membres Ordonnancement Ustion
Dépeçage Fond du silo
F 5011 M : 2 F : 1
2-3 a: 1 2-6 a : 1 >6 a: 1
Espace colmaté Rapide Décubitus latéral Non 4e comblement
S 5054 Indét. 1-4 m Indét. Indét. Indét. Non Indét.
F 5236 F > 4 a Espace colmaté Rapide Décubitus latéral Non 4e comblement
S 5413 Indet. 3-5 m Air libre/espace colmaté Rapide Décubitus latéral Non Fond du silo
S 5008 Indét. <6 m Air libre/espace colmaté Différé Non Non Fond du silo
S 5017 Indét. <6 m Air libre/espace colmaté Différé Non Non Fond du silo
S 5224 Indet. <6 m Non documenté Rapide à différé Non documenté Non Fond du silo / 1er comblement
S 5553 Indet. Adulte Espace colmaté RapideAssocié à une manipulation sur squelette humain
Non Dans cône d'effondrement
Bœuf S 5348.2 M 18-24 m Espace colmaté Rapide Non Consommation Fond du silo
Caprinés surtout S 5231.4 Indet.
6-12 m : 1 12-24 m : 2 10-42 m : 5
Espace colmaté Rapide Non Rare et ciblée 3e comblement
Ovin S 5268.9-10 F 4-6 ans : 2 F Espace colmaté Différé Mandibules et
scapulas Non Fond du silo / 1er comblement
Caprinés S 5316.4 bélier : 1 chèvre : 1
2-4 a : 1 4-6 a : 1 >36 m : 1
Espace colmaté Rapide Non Oui Fond du silo
Rep
as c
olle
ctif
Chien
Porc
Lièvre
Divers
Tête
Tête Equidés
Sque
lette
Parti
e de
sque
lette
ap
pend
icul
aire
Rejets alimentaires probables
S 5449Poisson Espace colmaté Rapide 6e comblement
Ordonnancement Non Dernier comblement
Tab. 9 : Présentation des dépôts animaux du site de Geispolsheim « Schwobenfeld » (les fosses sont désignées par la lettre F et les silos par la lettre S suivie de leur numéro) (O. Putelat).

Les dépôts d’ossements d’animaux en France, de la fouille à l’interprétation
—62—
Épreuves d’auteur
de traces anthropiques ne suffit pas à exclure une mise à mort, puisqu’il existe des techniques traditionnelles non sanglantes d’abattage (Vigne, Marinval-Vigne 1991, p. 73-75). De plus, la mise au jour de ces porcs au sein des trois aires riches en dépôts plaide, au moins pour ceux qui y sont situés, en la faveur de dépôts particuliers, et peut être en la défaveur de l’enfouissement des victimes d’un épisode pathologique, l’un n’empêchant toutefois pas l’autre. Les individus des silos 5375, 5564 et de la fosse 5674 pourraient donc éventuellement être assimilés à des dépôts particuliers, tandis qu’il resterait impossible de statuer pour le porc du silo 5176, situé à proximité d’une zone non fouillée.
D’autres sites régionaux ont livré des squelettes de porcs (fig. 72). Dans le Bas-Rhin, les sites de Pfulgriesheim « Langgarten-Buetzel », daté de La Tène A1 (Balzer,
Meunier 2005) et de Littenheim « Auf den Sand », daté de La Tène A-B (inédit, diagnostic Christophe Croutsch, Pair ; Putelat 2008) ont livré tous deux un squelette de porcelet déposé dans un silo. Dans le Haut-Rhin, le dépôt multispécifique du silo 235 de Colmar « Jardin des Aubépines » a lui aussi livré un squelette de por-celet (supra). Outre-Rhin, à proximité dans le Bade-Wurtemberg, le lien entre un dépôt de porc et une inhu-mation humaine a pu être démontré à March-Neuerhausen « Lindenacker » (La Tène) où trois inhumations humaines étaient associées dans la même structure à un squelette de porc (Jeunesse, Ehretsmann 1988).
L’aspect du squelette équin 5517, relativement specta-culaire, évoque encore une fois l’hypothèse de la compo-sition d’un « tableau ». Cependant, tout en défendant cette hypothèse, et par là même celle d’un dépôt particulier,
Zone 5(2006)
Zone 5(2008)
0 50 m
N
paléochenal de la Bruche
Zone détruite
Zone détruite
5054
5017
50115008
5413
5564
5149
5176
519651995219
5224522752305236
5674
52685302
5348
53755450
5517
5553
5316
5231
5376
5449
5142
5275
5683
5229
5015
5010
5252
5444
5197
BœufChien
Poisson
ChevalLièvreOvin
Porc
Dépôt d’objet métallique (?)
SquelettesTêtes
Repas collectifs
Parties de squelette appendiculaire
Os humain erratiqueInhumation humaine
Faune
Métal
Humain
Fig. 71 : Répartition spatiale des dépôts de faune, humain et métallique de Geispolsheim « Schwobenfeld » (M. Landolt).

O. Putelat et M. Landolt, La caractérisation des dépôts animaux de La Tène ancienne à Geispolsheim
—63—
Épreuves d’auteur
en cohérence avec l’appartenance de l’équidé à l’une des trois aires riches en dépôts (fig. 71), nous constatons que l’interprétation des données reste ambigüe. Les quatre membres de ce cheval ont-ils été consommés après son éventuelle mise à mort ? Pour quelles raisons son os coxal gauche est-il partiellement carbonisé ? La plaie de garrot décelée était-elle de nature à dissuader de la consommation du squelette axial ? En d’autres termes, la non-consommation du squelette axial fut-elle délibérée ou fut-elle la conséquence d’écoulements purulents persis-tants dans la région du garrot ?
Des exemples régionaux montrent que le cheval peut être inhumé seul, comme par exemple à Emmendigen-Forchheim « Heckle » (Bade-Wurtemberg) dans un silo daté du début de La Tène (Maise 1996), ou avec d’autres espèces animales, comme à Colmar « Jardin des Aubépines » dans le silo 235 (supra).
L’association humain/cheval est aussi reconnue. Dans le Haut-Rhin, à La Tène A, à Wettolsheim « Ricoh » (fig. 72), le squelette d’un cheval acéphale a été mis au jour au fond d’un silo (Jeunesse, Ehretsmann 1988 ; Lambach 1988 ; Méniel 1988). Il s’agit d’un mâle adulte, déposé enroulé sur lui-même en épousant la forme de la struc-ture. Son crâne a été prélevé durant la décomposition du cadavre. Dans un second temps, après remblaiement du dépôt, une femme a été inhumée en position fléchie. Pour le Bade-Wurtemberg citons le dépôt d’Ihringen « Burghalde », constitué d’un individu et d’un cheval (Kraft, Unser 1947), probablement daté de La Tène A-B.
L’étude archéozoologique des squelettes de chiens issus des fouilles de 2006 avait conclu au simple rejet de cadavre au sujet du chiot du silo 5054 et restait dans l’ex-pectative pour les trois chiens de la fosse 5011 (Landolt et al. 2007, p. 305). Les fouilles de 2008 ont livré deux chiens supplémentaires dans le silo 5413 et la fosse 5236. A l’instar des porcs, l’absence de traces de mise à mort, l’étagement des âges des décès, confirmé par la décou-verte d’ossements de chiens périnataux dans les silos 5279 et 5308, pourraient indiquer que ces squelettes de chiens résultent d’une mortalité naturelle. Au vu de la place tenue par l’élevage des caprinés à Geispolsheim « Schwobenfeld », on pourrait aussi évoquer pour les chiens subadultes et adultes l’élimination de chiens « tueurs de mouton » (Putelat inédit-a) car, ici encore, l’absence de traces anthropiques ne suffit pas à exclure une mise à mort. Nous avons par exemple recueilli, dans le cas précis des « tueurs de moutons », des témoignages récents d’éleveurs qui pensaient que l’élimination de ces chiens au comportement déviant ne justifiait pas de gâcher une cartouche de fusil, et préféraient opter pour une mort par pendaison à une échelle, à un arbre flexible, etc.
Cependant, le soin (relatif) apporté à la disposition des trois chiens de la fosse 5011 et à ceux des silos 5236 et 5413, leur appartenance aux trois aires riches en dépôts animaux, voisinant ou non des inhumations en silos (fig. 71), plaident pour considérer ces cinq individus comme étant des dépôts particuliers.
Dans le Haut-Rhin, les deux dépôts, mis en évi-dence dans de « petites alvéoles » aménagées du site de Mulhouse « Moenchsberg » (Haut-Rhin) sont pro-bablement des chiens10 (Schweitzer 1977) (fig. 72). Dans le Bas-Rhin, les occupations de Geispolsheim « Bruechel » datée du Hallstatt D (Pétry 1984, p. 256) et de Mittelhausen « Liesbuehl Gimbretter Weg », datée de La Tène A (Broc, Putelat 2012), ont toutes deux livré un dépôt de chien en silo (fig. 72).
Une certaine proximité des dépôts canins avec les sque-lettes humains est notée à Geispolsheim « Schwobenfeld ». C’est aussi le cas à Colmar « Jardin des Aubépines » où le dépôt multispécifique du silo 235 qui livre plusieurs chiens se trouve à proximité de dépôts humains en silos (Roth-Zehner et al. 2008).
10 En l’absence d’étude archéozoologique, cette identification reste incertaine.
0 25 75 100 km50
Dépôts de faune
Hallstatt C-D1
1 2-5 ›5Hallstatt D2 à La Tène C
Inhumations humaines
1 2-5 ›5Nombre d'individus :
Nombre d'individus :
Dachstein
Morschwiller-le-Bas
Entzheim
Souffelweyersheim
Strasbourg
Bischoffsheim
Mulhouse
Eckbolsheim
Colmar
Didenheim
Geispolsheim
Holtzheim
Illfurth
Littenheim
Marlenheim
Meistratzheim
Mittelhausen
Pfulgriesheim
Rosheim
Steinbourg
Wettolsheim
Ettendorf
GoxwillerObernai
Bergheim
Gundolsheim
Wolfgantzen
Fig. 72 : Carte de répartition des inhumations humaines et animales en silo du Hallstatt D à La Tène C en Alsace (M. Landolt).

Les dépôts d’ossements d’animaux en France, de la fouille à l’interprétation
—64—
Épreuves d’auteur
A Geispolsheim « Schwobenfeld », l’étude archéozoo-logique des deux squelettes de levrauts issus des fouilles de 2006 (silos 5008 et 5017) avait conclu à la présence d’animaux tombés et décomposés au fond de ces pièges naturels que constituent les silos (Landolt et al. 2007, p. 310). Les fouilles de 2008 ont de nouveau livré un sque-lette de levraut, probablement partiellement disloqué (silo 5224), et un squelette de lièvre adulte, physiquement asso-cié à un squelette humain (silo 5553). La nature de cette association est complexe. L’hypothèse d’un ensevelisse-ment du cadavre humain en plusieurs étapes est attestée par la présence de manipulations post-mortem pendant la décomposition. L’anthropologie a mis en évidence un déplacement du bas de la jambe droite pour accéder à la gauche pendant les premiers stades de la décomposition de l’individu, dans le but d’y prélever le tibia gauche. Les autres éléments osseux du bas de la jambe ont été ensuite réduits le long de la paroi pour y déposer immédiatement après la dépouille d’un lièvre. L’absence de dépôt sédi-mentaire entre les os du défunt et ceux de l’animal prouve la simultanéité des dépôts dans un espace-temps réduit (le corps de l’individu n’était pas encore complètement décharné). Il apparaitrait ainsi que ce dépôt de lièvre soit une sorte de « compensation » symbolique pour l’individu après le prélèvement de son tibia gauche (Pélissier in Landolt et al. 2013, p. 317). Le rôle joué par ce lièvre qui remplace la jambe du défunt, et qui est de surcroît le seul animal sauvage à être partie prenante des dépôts animaux sur ce site, nous amène à revoir l’hypothèse émise en 2007 d’un piégeage naturel des levrauts dans les silos. La localisation des squelettes de levrauts à proximité d’inhu-mations en silos (5008 et 5017), ou simplement dans une des trois aires du site riches en dépôts animaux (silo 5224) (fig. 71) nous pousse vers une réinterprétation des décou-vertes, du piégeage naturel au dépôt particulier.
Un certain nombre de découvertes de squelettes de lièvres en silos sont connues de la Préhistoire à la Protohistoire (Arbogast 2006, p. 41). Dans le Bas-Rhin signalons les silos 134, 143 et 183 de Rosheim « Leimen » (Ibid.), datés probablement du Néolithique pour le silo 183 et du Hallstatt pour les silos 134 et 14311. A Mittelhausen « Liesbuehl Gimbretter Weg », le silo 85 daté de La Tène A livre lui aussi un squelette incom-plet de levraut (Broc 2011, p. 28). Dans le Haut-Rhin, à Didenheim « Kahlberg » un squelette de lièvre provient du silo 59, daté du Bronze final ou du Hallstatt, et un squelette de léporidé immature (lièvre ou lapin intrusif) a été découvert dans la partie supérieure du comblement de l’inhumation double du silo 13, daté de La Tène ancienne (Denaire et al. 2011). Sur un plan régional étendu, il faut aussi souligner la singularité du site de Möhlin (Suisse, Argovie), daté du Hallstatt D3-La Tène A1, où des dépôts en silos de squelettes entiers ou partiels de cerfs, de lièvres
11 Nous remercions vivement Ph. Lefranc (Inrap), responsable de la fouille, pour ces renseignements. .
et de hérissons ont été identifiés (Brogli et al. 2003). Malgré cela, il nous reste une réticence à assimiler avec
certitude à des dépôts particuliers toutes les découvertes de squelettes de lièvres en silos. Nous constatons qu’il s’agit très souvent d’animaux immatures. Prenant par exemple les trois levrauts mis au jour dans les silos 134, 143 et 183 de Rosheim « Leimen » (supra), le vaste inter-valle chronologique qui sépare le silo 183, probablement néolithique, des silos hallstattiens 134 et 143 suggère autant les effets de piégeages naturels mêlés à des dépôts anthropiques, que la perpétuation « immémoriale » de la pratique de dépôts de léporidés. Par ailleurs, plus tardive-ment, c’est sans ambigüité qu’à Sierentz « Landstrasse » (Haut-Rhin), un squelette de jeune lièvre quasiment com-plet (puits antique FR 31) a été attribué à un piégeage naturel (Vigne et al. 1994, p. 75).
3.4. Les restes de « repas collectifs »La plus grande partie des assemblages fauniques liés
aux rejets de « repas collectifs » fut mise en évidence dès la phase de terrain, en raison de la quantité d’ossements qui les caractérisent. Le pré-examen de la faune laténien-ne de Geispolsheim « Schwobenfeld » a permis d’établir la spécificité du matériel faunique de certaines structures, mésestimée sur le terrain. L’étude archéozoologique par US permet d’approcher avec précision le spectre fauni-que et la répartition anatomique des taxons consommés. Les principales caractéristiques de ces rejets de « repas collectifs », mettant en jeu au moins un bovin et dix-sept caprinés, sont assez constantes (tab. 9). L’essentiel de la ressource carnée peut être fourni par un individu dont la plus grande partie du squelette est présente et dont la consommation est attestée par des traces de découpe (bovin du silo 5348). Il peut aussi s’agir de plusieurs individus dont les têtes (fendues, consommées, parfois grillées) et les parties charnues du squelette sont les mieux représentées (huit caprinés de l’US 5231.4, trois caprinés des US 5268.9-10, trois caprinés de l’US 5316.4, trois caprinés du silo 5348). Des pratiques spécifiques à chaque silo sont souvent perceptibles. On constate la rareté des traces de désossage au regard des traces de désarticulation (US 5231.4), la consommation préférentielle du côté droit et une différence dans l’exposition au feu entre les mem-bres antérieurs et postérieurs (US 5231.4), le rejet au feu de plus des trois-quarts des ossements (US 5316.4), des manipulations d’ossements in situ, peut être postérieure-ment à leurs rejets dans le silo (mandibules et scapulas du silo 5268). Enfin, il s’agit d’animaux jeunes (probable-ment sous-représentés dans l’US 5316.4, en raison de la forte ustion des vestiges) et subadultes.
3.5. DiversL’amas ichtyofaunique du silo 5449, composé de plus
d’un millier d’écailles et de quelques os appartient à l’une d’une des trois aires riches en dépôts (fig. 71), mais nous n’avons pu déterminer s’il s’agissait d’un rejet d’écaillage ou d’un dépôt particulier. Son statut reste donc incertain.

O. Putelat et M. Landolt, La caractérisation des dépôts animaux de La Tène ancienne à Geispolsheim
—65—
Épreuves d’auteur
ConclusionLe passage des dénominations neutres de dépôt, ou d’as-
semblages fauniques, au qualificatif de « dépôt particulier » peut être subjectif. À Geispolsheim « Schwobenfeld » c’est l’organisation spatiale des dépôts, la présence de dépôts particuliers avérés comme le bovin du silo 5302, l’ovin du silo 5376, les manipulations de mandibules et scapulas du silo 5268, les têtes équines du silo 5450, l’in-tégration du squelette de lièvre au squelette humain du silo 5553, qui forgent le cadre du raisonnement et nous amè-nent à identifier comme étant des dépôts particuliers des squelettes animaux pour lesquels nous aurions sans doute envisagé une interprétation différente en d’autres contex-tes. La mise en évidence de manipulations post-mortem, entièrement tributaire de la qualité de la documentation de terrain, fournit donc des indices indispensables à la compréhension des modalités d’assemblage des dépôts animaux.
Parmi les 27 dépôts étudiés, deux ont été considérés comme étant des rejets de déchets (silos 5149 et 5196) et un est indéterminé (ichtyofaune du silo 5449). Vingt-qua-tre dépôts seulement sont donc retenus. Les squelettes y sont les mieux représentés, suivis par les ensembles anato-miques issus du squelette appendiculaire, puis par les têtes osseuses. Ces squelettes proviennent de quinze structures et représentent six taxons pour dix-neuf individus, dont un en contexte funéraire (tab. 9). Les ensembles anatomiques issus du squelette appendiculaire proviennent de quatre structures et représentent trois taxons pour quatre indivi-dus, étant entendu qu’il n’est pas possible d’exclure l’hy-pothèse que les segments anatomiques appendiculaires équins des silos 5199 et 5230 proviennent d’un des quatre équidés déterminés par leurs têtes osseuses (silos 5450 et 5517). Le seul dépôt de tête revêtant un caractère particu-lier est le triple dépôt de têtes d’équidés du silo 5450.
Les équidés (six occurrences) et les chiens (six occur-rences) prévalent au sein des dépôts, toutes catégories confondues. Les équidés figurent une fois parmi les squelettes et prévalent au sein des deux groupes consti-tués par les têtes osseuses (une occurrence de trois têtes osseuses) et segments connectés (deux occurrences). Les chiens n’apparaissent qu’au sein du groupe constitué par les squelettes (six individus). Le porc est représenté quatre fois au sein de la catégorie des squelettes (six individus). Les caprinés, peu représentés parmi les squelettes et par-ties de squelettes, prévalent en revanche au sein des rejets de repas collectifs.
Pour l’Alsace, les études récentes ont montré l’im-portante du phénomène des dépôts animaux au cours du Néolithique moyen-récent (Lefranc et al. 2010). Après quelques attestations encore rares pour l’Age du Bronze ancien et moyen, le phénomène connaît un important regain entre le Hallstatt C et La Tène B où dix-neuf occu-pations ont livré des vestiges de ce type (fig. 72).
Pour le Bas-Rhin, onmentionnera les sites d’Eck-bolsheim « Zénith » (fouille inédite, Gérard Bataille, Inrap), Pfulgriesheim « Langgarten/Buetzel » (Balzer, Meunier 2005), de Geispolsheim « Bruechel/Kirstenfeld » (Pétry 1984) et « Schwobenfeld », d’Holtzheim « Am Schuesselberg » (Kuhnle, Tesnier-Hermetey 2005), de Littenheim « Auf den Sand » (Putelat 2008), de Marlenheim « Le Domaine de la Couronne d’Or » (Thomas et al. 2010; Putelat, ce volume), de Meistratzheim « Foegel » (Véber et al. 2008) et « Pfett » (Murer et al. 2010), de Mittelhausen « Liesbuehl Gimbretter Weg » (Broc 2011), de Rosheim « Leimen » (Lefranc et al. 2006) et « Lotissement Saint-Odile » (Jeunesse, Sainty 1985) et de Steinbourg « Altenberg/Ramsberg » (Putelat en cours).
Dans le Haut-Rhin, on signalera les occupations de Colmar « Jardin des Aubépines » (Roth-Zehner et al. 2008), de Didenheim « Kahlberg » (Denaire et al. 2011), d’Illfurth « Naegelberg » (Roth-Zehner et al. 2004), de Mulhouse « Moenchsberg » (Schweitzer 1977), de Wettolsheim « Ricoh » (Méniel 1988) et de Wolfgantzgen « Niederfeld » (Kuhnle et al. 2000).
Pour la Protohistoire, les dépôts d’animaux complets, ou de parties de leurs corps, existent également en Europe, du Hallstatt D à La Tène C (voir, par exemple, Auxiette 2000, Auxiette et al. 2010, Gransar et al. 2002, Landolt et al. 2011, Méniel 2008). De la même façon, en gardant à l’esprit qu’elle ne correspond très probablement qu’à des évènements ponctuels dans la vie des Laténiens de Geispolsheim « Schwobenfeld », nous constatons que la pratique de repas collectifs s’insère bien dans l’état bibliographique de la question pour l’âge du Fer (voir, par exemple, Le Bihan et Méniel 2002 ou Deffressigne et al. 2007).
A Geispolsheim « Schwobenfeld »la relation entre les inhumations humaines en silos, d’éventuels dépôts d’objets métalliques et les dépôts animaux est clairement établie sur un plan spatial, bien que l’on ne puisse pas réel-lement parler de dépôts funéraires (fig. 71). Cette relation est poussée à l’extrême dans le cas du silo 5553, où le liè-vre vient remplacer la jambe prélevée sur le défunt. Pour le Hallstatt final et La Tène ancienne, des associations spatiales entre inhumations humaines en silos et dépôts animaux sont elles aussi connues régionalement (fig. 72) (voir aussi Landolt et al. 2011).
Sur un vaste plan régional le site de Geispolsheim « Schwobenfeld » se distingue donc à La Tène ancienne par le nombre et la variété de ses dépôts animaux. On y trouve regroupées sur un même site diverses occurrences taxonomiques qui n’apparaissent le plus souvent qu’au cas par cas sur d’autres sites en Alsace, des manipulations de squelettes, la pratique de repas collectifs, des associations spatiales entre inhumations humaines et dépôts animaux. On peut sans risque parler pour ce site d’une réelle spéci-ficité, caractérisée par le caractère cumulatif de dépôts de différentes catégories.

Les dépôts d’ossements d’animaux en France, de la fouille à l’interprétation
—66—
Épreuves d’auteur
Bibliographie
Arbogast 2006 : R.-M. Arbogast, La faune. In : Ph. Lefranc, E. Boës avec la coll. de R.-M. Arbogast, Rosheim « Leimen » (Bas-Rhin) : Occupations du Néolithique récent, du début du Bronze moyen et du Premier Âge du Fer. Document final de synthèse de fouille d’archéologie préventive, dactylographié, Inrap, Strasbourg, 2006, 38-41.
Auxiette 2000 : G. Auxiette, Les rejets non domestiques des établissements ruraux du Hallstatt final à La Tène finale dans la vallée de l’Aisne et de la Vesle. In : S. Marion, G. Blancquaert (dir.), Les installations agrico-les à l’Age du Fer en France septentrionale, Actes du colloque de Paris, 29 et 30 novembre 1997. Éditions Rue d’Ulm, ENS, Études d’Histoire et d’Archéologie 6, 2000, 169-180.
Auxiette et al. 2011 : G. Auxiette, A. Baudry, P. Méniel, Une histoire de l’élevage dans l’ouest de la Normandie : les sites de Mondeville, Ifs, Fleury, Creully (Calvados) et les autres… In : Ph. Barral, B. Dedet, F. Delrieu, P. Giraud, I. Le Goff, S. Marion, A. Villard-le-Tiec (dir.), L’âge du Fer en Basse-Normandie, Actes du XXXIIIe colloque de l’AFEAF de Caen, 20-24 mai 2009. Presses Universitaires de Franche-Comté, Annales Littéraires de l’Université de Franche-Comté, Série environnement, sociétés et archéologie, 14, vol. 2, 2011, 185-202.
Balzer, Meunier 2005 : I. Balzer, K. Meunier, Un site d’habitat du Hallstatt final et de La Tène ancienne à Pfulgriesheim (Bas-Rhin). Eine späthallstatt- und frühlatènezeitliche Siedlung in Pfulgriesheim (Bas-Rhin). In : A.-M. Adam (dir.), Recherches de protohis-toire alsacienne. La céramique d’habitat du Bronze final III à La Tène ancienne. Société Archéologique de l’Est, supp. 23, Dijon, 2005, 257-273.
Barone 1986 : R. Barone, Anatomie comparée des mam-mifères domestiques. Volume 1 : Ostéologie. Vigot frè-res, 3e édition, Paris 1986.
Broc 2011 : E. Broc, Initiation aux méthodes de l’archéo-zoologie : Étude de deux sites du Bas-Rhin, Mittelhausen et Meistratzheim. Mémoire de Master, dactylographié, Université de Bourgogne, Dijon 2011.
Broc, Putelat 2012 : E. Broc, O. Putelat, La faune. In : Ch. Croutsch (dir), Mittelhausen, Bas Rhin, « Liesbuehl, Gimbretter Weg », construction de la LGV Est Européenne Tronçon H, Site 10-1. Structures domesti-ques et funéraires du Michelsberg et de La Tène. Rapport de fouille préventive, dactylographié, Pôle d’Archéolo-gie Interdépartemental Rhénan, Sélestat, 2012, 152-168.
Brogli et al. 2003 : W. Brogli, P. Jud, J. Schibler, Frühlatènezeitliche Gruben mit Tierdeponierungen aus Möhlin (AG). In : S. Plouin, P. Jud (éd.), Habitats, mobiliers et groupes régionaux à l’Âge du Fer, actes du XXe colloque A.F.E.A.F, Colmar-Mittelwihr, 16-19 mai 1996. Revue Archéologique de l’Est, supp. 20, Dijon, 2003, 137-146.
Callou 1997 : C. Callou, Diagnose différentielle des principaux éléments squelettiques du lapin (genre Oryctolagus) et du lièvre (genre Lepus) en Europe occi-dentale. Fiches d’Ostéologie Animale pour l’Archéolo-gie, Série B : Mammifères, 8, CRA du CNRS, Valbonne-Sophia Antipolis 1997.
Coutureau et al. 2003 : M. Coutureau, C. Beauval, J.-G. Ferrié, J.-H. Yvinec, Corpus de squelettes de mammifè-res. Le porc gaulois (Sus domesticus). [En ligne] www.arkzoo.archeozoologie-archaeozoology.org/sommaire.
Coutureau, Forest 1996a : M. Coutureau, V. Forest, Corpus de squelettes de mammifères. Le mouton (Ovis aries). [En ligne] www.arkzoo.archeozoologie-archaeozoology.org/sommaire.
Coutureau, Forest 1996b : M. Coutureau, V. Forest, Corpus de squelettes de mammifères. Le lapin (Oryctolagus cuni-culus). [En ligne] www.arkzoo.archeozoologie-archaeo-zoology.org/sommaire.
Coutureau, Forest 1996c : M. Coutureau, V. Forest, Corpus de squelettes de mammifères. La vache (Bos taurus). [En ligne] www.arkzoo.archeozoologie-archaeozoology.org/sommaire.
Coutureau, Forest 1996d : M. Coutureau, V. Forest, Corpus de squelettes de mammifères. Le chien (Canis familiaris). [En ligne] www.arkzoo.archeozoologie-archaeozoology.org/sommaire.
Deffressigne et al. 2002 : S. Deffressigne, N. Tikonoff, K. Boulanger-Boucher, Ch. Chaussé, C. Tesnier-Hermetey avec la coll. de G. Allenet, B. Chaume, A. De Hingh, J.-P. Legendre, M. Leroy, Ch. Leroyer, P. Merluzzo, Les gisements d’habitat de la fin du Premier Âge du Fer à Gondreville-Fontenoy-sur-Moselle (54). Le stockage intensif et ses conséquences économiques et sociales. Archaeologia Mosellana, 4, 2002, 81-184.
Deffressigne et al. 2007 : S. Deffressigne, N. Tikonoff, G. Auxiette, C. Tesnier-Hermetey, Rejets particuliers de la fin de l’âge du Bronze à travers quelques exemples de l’Est de la France. In : Ph. Barral, A. Daubigney, C. Dunning, G. Kaenel, M.-J. Roulière-Lambert (éd.), L’âge du Fer dans l’arc jurassien et ses marges. Dépôts, lieux sacrés et territorialité à l’âge du Fer, Actes du XXIXe colloque international de l’AFEAF, Bienne, 5-8 mai 2005. Presses universitaires de Franche-Comté, Besançon, vol. 2, 2007, 621-628.
Denaire et al. 2011 : A. Denaire, (L. Boury, E. Cartier-Mamie, S. Goepfert, Didenheim « ZAC Espace-Kahlberg ». Un nouveau site du Néolithique récent, de l’Âge du Bronze (RSFO) et du premier Âge du Fer. Rapport d’archéologie préventive, dactylographié, Antea-Archéologie, Habsheim 2011.

O. Putelat et M. Landolt, La caractérisation des dépôts animaux de La Tène ancienne à Geispolsheim
—67—
Épreuves d’auteur
Gransar et al. 2002 : F. Gransar avec la coll. de S. Desenne, M. Gransar, La batterie de silos de Soupir « Le Champ Grand Jacques » (Aisne) : contribution à l’identification d’une centralisation du stockage à la Tène B dans le nord de la France. In : P. Méniel, B. Lambot, Découvertes récentes de l’âge du Fer dans le massif des Ardennes et ses marges. Repas des vivants et nourriture pour les morts en Gaule, actes du XXVe colloque de l’AFEAF, Charleville-Mézières, 24-27 mai 2001. Mémoire de la Société Archéologique Champenoise, 16, Reims, 2002, 67-80.
Habermehl 1985 : K.-H. Habermehl, Alterbestimmung bei Wild- und Pelztieren. Paul Parey Verlag, Berlin, Hamburg 1985.
Jeunesse et al. 1993 : Ch. Jeunesse, P. Méniel, B. Röder, L’habitat La Tène ancienne de Rosheim « Mittelweg » (Bas-Rhin). Cahiers de l’Association pour la promotion de la Recherche Archéologique en Alsace, Supp. 1, Strasbourg 1993.
Jeunesse, Ehretsmann 1988 : C. Jeunesse, M. Ehretsmann, La jeune femme, le cheval et le silo, une tombe de La Tène ancienne sur le site de Wettolsheim « Ricoh » (Haut-Rhin). Cahiers Alsaciens d’Art, d’Archéologie et d’Histoire, 31, 1988, 45-54.
Jeunesse, Sainty 1985 : Ch. Jeunesse, J. Sainty, Paysans de l’Âge de pierre : un village préhistorique il y a 6000 ans à Rosheim, Catalogue d’exposition, 11-19 mai 1985, Rosheim. Direction des Antiquités Préhistoriques d’Al-sace, Strasbourg 1985.
Kraft, Unser 1947 : G. Kraft, S. Unser, Ihringen, 1941-1947. Badische Fundberichte, 25, 1947, 311-312.
Kuhnle et al. 2000 : G. Kuhnle, C. Tesnier-Hermetey, S. Plouin Ch. Thurnheer, Ch. Maise, L’habitat hallstattien D2/D3 de Wolfgantzen (Bas-Rhin). Revue Archéologie de l’Est, 49, 2000, 135-181.
Kuhnle, Tesnier-Hermetey 2005 : G. Kuhnle, C. Tesnier-Hermetey, Holtzheim Am Schluesselberg (Bas-Rhin) : un habitat hallstattien de bord de terrasse loessique dans la plaine du Rhin supérieur. In : A.-M. Adam (dir.), Recherches de protohistoire alsacienne. La céramique d’habitat du Bronze final III à La Tène ancienne. Société Archéologique de l’Est, supp. 23, Dijon, 2005, 179-209.
Lambach 1988 : F. Lambach, Étude anthropologique de la sépulture 201 de Wettolsheim « Ricoh ». Cahiers Alsaciens d’Archéologie, d’Art et d’Histoire, 31, 1988, 55-70.
Landolt et al. 2007 : M. Landolt, M. Van Es, O. Putelat, D. Bouquin, C. Schaal, E. Boës avec la coll. de E. Baccharetti, D. Bevilacqua, I. Dechanez-Cler, Entzheim-Geispolsheim (Alsace, Bas-Rhin), Aéroparc (Lidl-CUS), volume 3 : Les occupations protohistoriques. Rapport final d’opération de fouille, Pôle Archéologique Interrégional Rhénan, Sélestat 2007.
Landolt et al. 2008 : M. Landolt, D. Bouquin, M. van Es, O. Putelat, C. Schaal, N. Sneider Les occupations halls-tattiennes et de La Tène ancienne d’Entzheim « In der Klamm » et de Geispolsheim « Schwobenfeld » (Bas-Rhin). Bulletin de l’Association Française de l’Étude de l’Age du Fer, 23, 2008, 33-36.
Landolt et al. 2011 : M. Landolt, E. Millet, M. Roth-Zehner avec la coll. de H. Barrand, E. Cartier, A. Mauduit, O. Putelat, Pratiques funéraires en Alsace du Ve au Ier siè-cle avant J.-C. In : Ph. Barral, B. Dedet, F. Delrieu, P. Giraud, I. Le Goff, S. Marion, A. Villard-le-Tiec)(dir.), Gestes funéraires en Gaule au second âge du Fer, Actes du XXXIIIe colloque de l’AFEAF de Caen, 20-24 mai 2009. Annales Littéraires de l’Université de Franche-Comté, Série environnement, sociétés et archéologie, 14, Presses Universitaires de Franche-Comté, 2011, vol. 2, 207-230.
Landolt et al. 2012 : M. Landolt, M. Roth-Zehner, F. Fleischer avec la coll. de E. Millet, M. Van Es, La chro-nologie de la céramique de la fin du Hallstatt D3 à La Tène B2 dans le sud de la plaine du Rhin supérieur : l’apport des nouveaux sites alsaciens d’Entzheim (Bas-Rhin), de Geispolsheim (Bas-Rhin) et de Colmar (Haut-Rhin). In : M. Schönfelder, S. Sievers (dir.), L’âge du Fer entre la Champagne et la vallée du Rhin, Actes du XXXIVe colloque de l’AFEAF d’Aschaffenburg, 13-16 mai 2010. RGZM Tagungen, 4, Verlag des Römisch-Germanischen Zentralmuseums, Mainz, 2012, 471-502.
Landolt et al. 2013 : M. Landolt (dir.), A.-M. Adam, G. Alix, A. Charrie, A. Ferrier, F. Fleischer, S. Francisco, B. Gratuze, C. Leprovost, E. Millet, L. Paleau, A. Pelissier, O. Putelat, M. Roth-Zhener, C. Schaal, Entzheim « In der Klamm » et Geispolsheim « Schwobenfeld » (Alsace, Bas-Rhin) : les occupations protohistoriques. Rapport final d’opération de fouille, dactylographié, Pôle d’Ar-chéologie Interdépartemental Rhénan, Sélestat 2013.
Landolt, Fleischer 2011 : M. Landolt, F. Fleischer, Les occupations d’Entzheim « In der Klamm » et de Geispolsheim « Schwobenfeld » (Bas-Rhin) du Hallstatt C à La Tène B (fouilles 2006 à 2009). Bulletin de l’As-sociation Française pour l’Étude de l’Age du Fer, 29, 2011, 13-17.
Le Bihan, Méniel 2002 : J.-P. Le Bihan, P. Méniel, Un dépôt d’ossements du premier âge du Fer sur l’Île d’Oues-sant : déchets alimentaires ou restes de banquets ? In : P. Méniel, B. Lambot (édit.), Découvertes récentes de l’âge du Fer dans le massif des Ardennes et ses marges. Repas des vivants et nourriture pour les morts en Gaule, actes du XXVe colloque de l’A.F.E.A.F., Charleville-Mézières, 24-27 mai 2001. Mémoire de la Société Archéologique Champenoise, 16, Reims, 2002, 303-316.
Lefranc et al. 2008 : Ph. Lefranc, E. Boës, C. Véber, Un habitat de La Tène ancienne à Souffelweyersheim « Les Sept Arpents » (Bas-Rhin) ». Revue Archéologique de l’Est, 57, 2008, 41-74.

Les dépôts d’ossements d’animaux en France, de la fouille à l’interprétation
—68—
Épreuves d’auteur
Lignereux, Peters 1996 : Y. Lignereux, J. Peters, Techniques de boucherie et rejets osseux en Gaule romaine. Anthropozoologica, 24, 1996, 45-98.
Maise 1996 : C. Maise, Eine Pferdebestattung der Frühlatènezeit und hallstattzeitliche Siedlungsreste in Forchheim, Kreis Emmendingen. Archäologische Ausgrabungen in Baden-Württemberg 1995, 1996, 110-112.
Méniel 1988 : P. Méniel, Un cheval sous la sépulture gauloise de Wettolsheim « Ricoh » (Haut-Rhin). Cahiers Alsaciens d’Archéologie, d’Art et d’Histoire, 31, 1988, 55-70.
Méniel 1993 : P. Méniel, La faune. In : Ch. Jeunesse, P. Méniel, B. Röder, L’habitat La Tène ancienne de Rosheim « Mittelweg » (Bas-Rhin). Cahiers de l’Asso-ciation pour la promotion de la Recherche Archéologique en Alsace, supp. 1, 1993, 132-149.
Méniel 2008 : P. Méniel, Manuel d’archéozoologie funé-raire et sacrificielle. Âge du Fer. Infolio, Gollion 2008.
Méniel et al. 2009 : P. Méniel, G. Auxiette, D. Germinet, A. Baudry, M.-P. Horard-Herbin, Une base de données sur les études de faunes des établissements ruraux en Gaule. In : I. Bertrand, A. Duval, J. Gomez de Soto, P. Maguer (dir.), Habitats et paysages ruraux en Gaule et regards sur d’autres régions du monde celtique, Actes du XXXIe colloque international de l’Association fran-çaise pour l’Étude de l’Âge du Fer, 17-20 mai 2007, Chauvigny. Association des Publications Chauvinoises, 35, Chauvigny, vol. 2, 2009, 417-446.
Morris 2008 : J. Morris, Re-examining Associated Bone Groups from Southern England and Yorkshire, c. 4000 BC to AD 1550, Mémoire de thèse, Bournemouth University, Bournemouth 2008 [En ligne] http://www.animalbones.org/publications.html.
Murer et al. 2010 : A. Murer, H. Barrand, B. Commercon, S. Goepfert, A. Langlois, A. Le Martret, A. Mauduit, B. Perrin, X. Perrin, L. Schott-Toullec, M. Roth-Zehner, Meistratzheim. Station d’épuration intercommuna-le (Alsace, Bas-Rhin, 67). Rapport final d’opération de fouille préventive, dactylographié, 2 vol, Antea-Archéologie, Habsheim 2010.
Paleau 2011 : L. Paleau, Les ossements animaux de Geispolsheim « Schwobenfeld »(Bas-Rhin), site 5046. Rapport d’étude archéozoologique, dactylographié, Pôle d’Archéologie Interdépartemental Rhénan, Sélestat 2011.
Pétry 1984 : F. Pétry, Geispolsheim. Gallia, 40-1982, 1984, 256-257.
Putelat 2002 : O. Putelat, Quelques réflexions sur la mor-talité en élevage ovin. In : I. Chenal-Velarde (dir.), La faune du site néolithique de Sion-Avenue Ritz (Valais, Suisse). Histoire d’un élevage villageois il y a 5000 ans. BAR International Series, 1081, Oxford, 2002, 105-115.
Putelat 2008 : O. Putelat, Les ossements animaux de Littenheim-Lupstein (Bas-Rhin). Rapport d’étude archéolozoologique, dactylographié, Pôle d’Archéologie Interdépartemental Rhénan, Sélestat 2008.
Roth-Zehner et al. 2004 : M. Roth-Zehner, avec la coll. de B. Bakaj, J.-B. Gaillard, J. Pellissier, Illfurth « Naegelberg », Lotissement du Chemin du buis (Alsace–Haut-Rhin). Rapport final d’archéologie préventive, dac-tylographié, Antea-Archéologie, Sierentz 2004.
Roth-Zehner et al. 2008 : M. Roth-Zehner, avec la col-laboration de A. Boyer, E. Cartier, S. Coubel, A. Le Martret, A. Mauduit, A. Richard, V. Rougier, Colmar : Jardin des Aubépines : Alsace, Haut-Rhin. Rapport final d’opération de fouille préventive, 4 vol., Antea-Archéologie, Habsheim 2008.
Schweitzer 1977 : J. Schweitzer, Riedisheim à l’épo-que celtique, Bulletin de la Société des Amis du Vieux Riedisheim, 5, 1977, 20-32.
Thomas et al. 2010 : Y. Thomas (dir.), A. Bandelli, P. Méniel, M. van Es, F. Jodry, N. Schneider, J. Wiethold, Marlenheim. Lotissement « Le Domaine de la Couronne d’Or » (Bas-Rhin), vestiges d’habitat du premier âge du Fer. Rapport final d’opération, dactylographié, Inrap, Strasbourg 2010.
Véber et al. 2008 : C. Véber, G. Bataille, P. Girard, F. Bergantz, V. Bourson, B. Bonaventure, H. Cicutta, F. Jodry, D. Lalaï, F. Latron, Ph. Lefranc, M. Leroy, Ch. Leyenberger, F. Schneikert, N. Schneider, W. Tegel, M. Van Es, J. Wiethold, Meistratzheim. Lotissement Foegel (Bas-Rhin, Alsace). Habitats du Néolithique, âge du Bronze, Hallstatt et La Tène finale. Sépulture Hallstatt. Rapport final d’opération de fouille préventive, dactylo-graphié, 2 vol., Inrap, Strasbourg 2008.
Vigne et al. 1994 : J.-D. Vigne, S. Bailon, C. Vallet, Les microvertébrés terrestres de Sierentz-Landstrasse (Haut-Rhin). Cahiers de l’Association pour la Promotion de la Recherche Archéologique en Alsace, 10, 1994, 68-96.
Vigne, Marinval-Vigne 1991 : J.-D. Vigne, M.-C. Marinval-Vigne, A propos de l’abattage sans effusion de sang : le cas du porc en Corse du Sud. Anthropozoologica, 14-15, 1991, 73-75.




















