
KATP channel subunits in rat dorsal root ganglia: alterations by painful axotomy
Upload
independentCategory
view
0download
0
Catherine M. Fuller and Dale J. BenosQadri, Zsuzsanna Bebok, James K. Bubien, Niren Kapoor, Rafal Bartoszewski, Yawar J. MigrationGlioblastoma Whole Cell Current and CellSodium Channel Subunits Inhibits Knockdown of ASIC1 and EpithelialDevelopmental Biology:Molecular Basis of Cell and
doi: 10.1074/jbc.M109.037390 originally published online June 26, 20092009, 284:24526-24541.J. Biol. Chem.
10.1074/jbc.M109.037390Access the most updated version of this article at doi:
.JBC Affinity SitesFind articles, minireviews, Reflections and Classics on similar topics on the
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2009/06/25/M109.037390.DC1.html
http://www.jbc.org/content/284/36/24526.full.html#ref-list-1
This article cites 58 references, 34 of which can be accessed free at
by guest on June 1, 2013http://www.jbc.org/Downloaded from

Knockdown of ASIC1 and Epithelial Sodium Channel SubunitsInhibits Glioblastoma Whole Cell Current and Cell Migration*□S
Received for publication, June 24, 2009 Published, JBC Papers in Press, June 26, 2009, DOI 10.1074/jbc.M109.037390
Niren Kapoor‡, Rafal Bartoszewski§, Yawar J. Qadri¶, Zsuzsanna Bebok§, James K. Bubien¶, Catherine M. Fuller¶,and Dale J. Benos‡§¶1
From the Departments of ‡Neurobiology, §Cell Biology, and ¶Physiology and Biophysics, University of Alabama at Birmingham,Birmingham, Alabama 35294
High grade gliomas such as glioblastoma multiformeexpress multiple members of the epithelial sodium channel(ENaC)/Degenerin family, characteristically displaying abasally active amiloride-sensitive cation current not seen innormal human astrocytes or lower grade gliomas. Usingquantitative real time PCR, we have shown higher expressionof ASIC1, �ENaC, and �ENaC in D54-MG human glioblas-toma multiforme cells compared with primary human astro-cytes. We hypothesize that this glioma current is mediated bya hybrid channel composed of a mixture of ENaC and acid-sensing ion channel (ASIC) subunits. To test this hypothesiswe made dominant negative cDNAs for ASIC1, �ENaC,�ENaC, and �ENaC. D54-MG cells transfected with the dom-inant negative constructs for ASIC1, �ENaC, or �ENaCshowed reduced protein expression and a significant reduc-tion in the amiloride-sensitive whole cell current as com-pared with untransfected D54-MG cells. Knocking down�ENaC or �ENaC also abolished the high PK
�/PNa� of
D54-MG cells. Knocking down �ENaC in D54-MG cellsreduced �ENaC protein expression but had no effect oneither the whole cell current or K� permeability. Using co-immunoprecipitation we show interactions between ASIC1,�ENaC, and �ENaC, consistent with these subunits interact-ing with each other to form an ion channel in glioma cells.Wealso found a significant inhibition of D54-MG cell migrationafter ASIC1, �ENaC, or �ENaC knockdown, consistent withthe hypothesis that ENaC/Degenerin subunits play an impor-tant role in glioma cell biology.
Gliomas are the most common primary tumors of the cen-tral nervous system. These tumors arise either from astro-cytes or their progenitor cells (1). Gliomas are divided intofour grades based on the degree of malignancy. Glioblastomamultiforme (GBM),2 Grade IV, is the most frequently occur-ring, most invasive, and has the worst prognostic outcome
with a median survival of approximately one year from diag-nosis (2).We have previously reported the presence of an amiloride-
sensitive current in glioblastoma cells that is not seen innormal astrocytes or low grade gliomas (3). Amiloride is apotassium sparing diuretic that inhibits sodium channelscomposed of subunits from the epithelial sodium channel(ENaC)/Degenerin (Deg) family. Amiloride-sensitive Na�
channels are essential for the regulation of Na� transportinto cells and tissues throughout the body. These channelsare found in all body tissues; from epithelia, endothelia,osteoblasts, keratinocytes, taste cells, lymphocytes, andbrain (4). Apart from the ENaCs, the ENaC/Deg family alsoincludes acid-sensing ion channels (ASICs) which have beenfound predominantly in neurons (4–6). Primary malfunc-tions of ENaC/Deg family members underlie or are involvedin the pathophysiology of several human diseases such assalt-sensitive hypertension (7, 8), pseudohypoaldosteronismtype I (7), cystic fibrosis (9), chronic airway diseases (10, 11),and flu (12).The ENaC/Deg family subunits share the same structural
topology. They all have short intracellular N and C termini,two transmembrane spanning domains, and a large extracel-lular cysteine-rich loop (4, 5). There are five ENaC subunitstermed �, �, �, �, and �. Functional ion channels arise from amultimeric assembly of these subunits. The prototypicalENaC channel of the collecting duct principal cell is thoughtto be ���ENaC (13, 14). The �-ENaC subunit appears to bethe core conducting element, whereas the �- and �-ENaCsubunits are associated with trafficking and insertion of thechannel in the cell membrane (13, 15, 16). ASICs are homol-ogous to ENaCs and are most prevalently expressed in thebrain and nervous system (17–19), although they are alsofound in the retina (20–22), testes (23), pituitary gland (24),lung epithelia (22), and bone and cartilage (25). Four ASICgenes have been identified so far, ASIC1–4. Of these,ASIC1–3 has multiple splice variants (19, 22). The crystalstructure of chicken ASIC1 has revealed it to be a homotri-mer (26). ASICs differ from their ENaC counterparts in thatthey are transiently activated by extracellular acid (19) andare much less sensitive to inhibition by amiloride (27, 28).Also ASIC1 is inhibited with high affinity by psalmotoxin 1(PcTX-1), a 40-amino acid peptide found in the venom of the
* This work was supported, in whole or in part, by National Institutes of HealthGrant CA101952 and DK37206.
□S The on-line version of this article (available at http://www.jbc.org) containssupplemental Fig. 1.
1 To whom correspondence should be addressed: University of Alabama atBirmingham, 1918 University Blvd., Birmingham, AL 35294. Tel.: 205-934-6220; Fax: 205-934-2377; E-mail: [email protected].
2 The abbreviations used are: GBM, glioblastoma multiforme; ENaC, epithelialsodium channel; Deg, Degenerin; ASIC, acid-sensing ion channel; GFP,green fluorescent protein; eGFP, enhanced GFP; YFP, yellow fluorescentprotein; eYFP, enhanced YFP; CFP, cerulean fluorescent protein; DN, dom-
inant negative; CHAPS, 3-[(3-cholamidopropyl)dimethylammonio]-1-pro-panesulfonic acid.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 284, NO. 36, pp. 24526 –24541, September 4, 2009© 2009 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
24526 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 284 • NUMBER 36 • SEPTEMBER 4, 2009 by guest on June 1, 2013http://www.jbc.org/Downloaded from

West Indies tarantula, Psalmopoeus Cambridgei (29). ASICs,because they are activated by acidic pH, have been suggestedto play a role in chemical pain associated with increasedtissue acidification as occurs in ischemia (30, 31). They havealso been implicated in touch sensation (32), taste (33), fear-conditioning (6), and learning and memory (34).Our laboratory has proposed that ENaC/Deg channels
underlie the basally activated cation current measured inhigh grade glioma cells (3).We hypothesize that the channelsforming this current pathway are composed of a mixture ofASIC and ENaC subunits. RNA profiling of a large number ofGBM-derived cell lines and freshly resected tumors haverevealed the presence of a myriad of ASIC/ENaC compo-nents (3). The basally active current seen in GBM cells can besignificantly reduced by amiloride or benzamil (a higheraffinity amiloride analog), both of which are inhibitors of theENaC/Deg family of ion channels (3). PcTX1, a selectiveASIC1 blocker, also effectively abolishes the basally activeGBM current (35).We have previously shown that ENaC andASIC subunits can form cross-clade interactions in a heter-ologous expression system (36). This study aims to probe thecomposition of the novel ENaC/Deg heteromer in a gliomacell line, D54-MG. Our study postulates that a change inGBM cell electrophysiological properties after subunitknockdown would be indicative of that subunit being a partof the GBM channel. We have sequentially knocked downdifferent ENaC/Deg subunits from the D54-MG glioma cellsand measured amiloride-sensitive whole cell current usingpatch clamp. We found that knocking down various ENaC/Deg subunits significantly reduced the whole cell patchclamp current in glioma cells and changed the restingNa�/K� permeability of the these cells. After subunit knock-down, glioma cells showed a reduced cell migration as com-pared with control cells, consistent with our hypothesis thatENaC/Deg subunits play an important role in glioma cellpathophysiology.
EXPERIMENTAL PROCEDURES
Cell Culture—Experiments were performed on a glioma-derived cell line D54-MG (GBM, derived from a WorldHealth Organization grade IV tumor; a kind gift of Dr. D.Bigner, Duke University, Durham, NC), primary humanastrocytes (a kind gift of Dr. Yancey Gillipsie, University ofAlabama at Birmingham, Birmingham, AL), and on CHO-K1cells. Cells were maintained in continuous cell culture in50:50 Dulbecco’s modified Eagle’s medium/F-12 medium(Invitrogen) supplemented with 10% fetal bovine serum(Hyclone, Logan, UT) and 1% penicillin/streptomycin(Invitrogen). For electrophysiological recording, cells weresplit 48 h before recording onto 35-mm dishes containingflame-sterilized coverslips. In the case of transfection, cellswere split 24 h before the transfection experiment and thenallowed to grow for 24–48 h before use.Isolation of Cellular RNA—Total cellular RNA was isolated
from D54-MG cells and primary human astrocytes usingRNeasy (Qiagen), according to the manufacturer’s recommen-dations. RNA isolated 48 h post-transfection from CHO-K1cells transfected with cDNAs for human ASIC1, �ENaC,
�ENaC, and �ENaC was used as a positive control. RNA con-centration was calculated based on the absorbance at 260 nm.RNA samples were stored at �20 °C.Measurement of Relative mRNA Levels Using Real Time PCR—
Real time PCR to measure ASIC1 (Hs00241630_m1), �ENaC(Hs00168906_m1), �ENaC (Hs00168918_m1), and �ENaC(Hs00161595_m1) mRNA was performed using TaqMan�One-Step RT-PCR Master Mix Reagents (Applied Biosys-tems, catalog no. 4309169) using the manufacturer’s hand-book as a reference (Relative Quantification; Applied Biosys-tems 7300/7500 Real Time PCR system; 2004). 18 S rRNA(Hs99999901_m1) was amplified as an internal control andused as a reference.Generation of Dominant Negative cDNAs—Site-directed
mutagenesis was performed using QuikChange XL IImutagenesis kit (Stratagene) to engineer premature stopcodons in human ASIC1, human �ENaC, human �ENaC,and human �ENaC cDNA. Briefly, wild type constructs ofeach subunit (previously ligated at the N terminus to theenhanced green fluorescent protein (eGFP) or to enhancedyellow fluorescent protein (eYFP) at the C-terminal expres-sion vector and shown to form functional ion channels (36),were subjected to PCRwith sense and antisense primers con-taining the specific base change to result in the desiredcodonmutations. For ASIC1, a stop codon was introduced atTyr-67, for �ENaC at Ser-155, for �ENaC at Glu-34, and for�ENaC at Ser-35. The reaction mixture was subjected to thecycling parameters according to the manufacturer’s hand-book; 1 cycle at 95 °C for 1 min followed by 18 cycles sequen-tially at 95 °C for 50 s, 60 °C for 50 s, and 68 °C for 1 min/kilobase of plasmid length and finally followed by 1 cycle at68 °C for 7 min. PCR products were transformed, and Esch-erichia coli were plated on Luria Bertani (LB) plates withkanamycin (30 �g/ml) and incubated overnight at 37 °C. Sin-gle colonies were grown on LB medium with 30 �g/ml kana-mycin and incubated overnight at 37 °C with constant shak-ing. DNA was isolated using the Qiagen miniprep kit(Qiagen), and the presence of themutation was confirmed bysequencing (Heflin Genetics Center, University of Alabamaat Birmingham). The colonies were grown in 250 ml of LBmedium with antibiotics, and DNA was isolated using theQiagen Maxiprep kit (Qiagen).Cell Transfections—For transfecting dominant negative
cDNAs, electroporation using the Bio-Rad Gene PulserX-cell electroporator (Bio-Rad) was used. Cells are detachedfrom the flask with 0.1 mM EDTA in phosphate-bufferedsaline and suspended at 1.3 � 107 cells/ml in RPMI 1640 �10 mM dextrose and 0.1 mM dithiothreitol in a 4-mm sterilecuvette (0.3 ml/cuvette). 5–10 �g of cDNA was added to thecuvette. The cuvette was then pulsed using a time lapse pro-tocol of a single square wave pulse of 300 V for 35 ms and 960microfarads as per the manufacturer’s protocol, and the cellswere returned to 60-mm dish in the incubator for 24–48 hbefore use.Stable ASIC1-GFP Cell Line—Post-transfection with ASIC1
ligated to GFP as described above, D54-MG cells were treatedwith 500 �g/ml G418 to select for transfected cells. After anti-biotic selection, GFP-positive cells were sterile-sorted by the
ENaC/Deg Subunits and Glioblastoma
SEPTEMBER 4, 2009 • VOLUME 284 • NUMBER 36 JOURNAL OF BIOLOGICAL CHEMISTRY 24527 by guest on June 1, 2013http://www.jbc.org/Downloaded from

Center for AIDS Research Fluorescent Activated Cell Sorting(FACS) facility at University of Alabama at Birmingham.Antibodies—�ENaCand�ENaC rabbit polyclonal antibodies
were generated against synthetic peptide in collaboration withDr. Mark Knepper (National Institutes of Health) as described(37). Briefly, antigenic peptide with the sequences human�ENaC, LMKGNKREEQGLGPEPAAPQQPTC, and human�ENaC, CNTLRLDSAFSSQLTDTQLTNEF, were synthesizedby Lofstrand Laboratories (Gaithersburg, MD). All peptideswere high performance liquid chromatography-purified.ASIC1 antibody was obtained from Alomone Labs (product#ASC-014), �ENaC antibody was obtained from Santa CruzBiotechnology (catalog #H-230: sc-21015), and GFP antibodywas obtained from Abgent (catalog #AM1009a).Total Membrane and Plasma Membrane Isolation—
D54-MG cells stably transfected with ASIC1-GFP weregrown on 100-mm tissue culture dishes. Cells were allowedto grow until 90–100% confluent and then scraped and col-lected by centrifugation at 600 � g for 5 min at 4 °C. Totalmembranes and plasma membranes were isolated using aplasmamembrane protein extraction kit (BioVision) accord-ing to the manufacturer’s protocol. Total membrane andplasma membrane were dissolved in lysis buffer containing20 mM Tris HCl, pH 8, 137 mM NaCl, 10% glycerol, 2 mM
EDTA, 1% w/v CHAPS, and protease inhibitor (Complete,Roche Applied Science). Total membrane and plasma mem-brane protein concentration was measured using the BCAprotein assay kit (Pierce).Immunoblotting and Co-immunoprecipitation—For im-
munoblotting, cell lysates were assayed for protein concen-tration (BCA, Pierce), and then 50 �g of protein was loadedin 2� SDS sample buffer containing 5% �-mercaptoethanolinto the wells of a 6–8% SDS-PAGE gel (4% stacking gel).The gels were transferred onto polyvinylidene difluoridemembranes and blocked with 5% nonfat dry milk in TBSTbuffer (100 mM Tris, pH 7.5, 150 mM NaCl, 0.1% Tween 20)for 1 h and then incubated overnight at 4 °C with primaryantibody. The blots were developed with horseradish perox-idase-conjugated secondary antibodies and enhancedchemiluminescence (SuperSignal West Pico, Pierce) andautoradiography. Band densities were assessed using Scana-lytic software.In co-immunoprecipitation experiments total membrane
and plasma membrane fractions of D54-MG stably trans-fected with ASIC1-GFP was used. For control experimentsCHO-K1 cells were transiently transfected with cDNA forASIC1-GFP, and CHO-K1 and D54-MG cells were tran-siently transfected with CFP-CLC1. Cells were lysed usinglysis buffer with 1% CHAPS and protease inhibitors (Com-plete, Roche Applied Science). 2 �g of primary antibody wasadded to 500 �g of cell lysate and incubated overnight at4 °C. 100 �l of protein A-agarose beads were added andallowed to incubate for 4 h. The beads were then pelleted andwashed 3–4 times with lysis buffer. To elute the immunecomplex, 50 �l of elution buffer (0.1 M glycine-HCl, pH 2.5–3.0) was added to the beads and allowed to incubate on icefor 5 min and centrifuged at 2500 � g for 1–3 min at 4 °C.Supernatant was collected, and the pH was adjusted to phys-
iological pH by adding 10 �l of 1 M Tris. 10–25 �l of loadingbuffer was added to the eluate and loaded onto a 6% SDS gel.Immunoblotting was carried out as described above usingthe appropriate primary and secondary antibodies. All buff-ers used for preparing cell lysates, membranes, and subcel-lular fractions included protease inhibitors (Complete,Roche Applied Science).Whole Cell Patch Clamp Recordings—Amiloride-sensitive
whole cell currents were recorded in transfected and untrans-fected glioma cells. Cells were cultured on flame-sterilized cov-erslips and transfected 48 h before patch-clamping. Patchpipettes were made of thin-walled borosilicate glass using aP-97 Flaming/Browning Micropipette puller (Sutter Instru-ments) and typically had a resistance of 5–10 megaohms.Pipettes were filled with an electrolyte solution containing 100mM potassium gluconate, 30 mM KCl, 10 mM NaCl, 20 mM
HEPES, 0.5 mM EGTA, 4 mM ATP, pH 7.2. Current recordingswere low pass-filtered at 2 kHz and digitized online at 10 kHzwith a Digidata 1200 (Axon Instruments). pClamp 9.0 (AxonInstruments) was used to acquire and store data. Our standardbath solution consisted of 125 mM NaCl, 5 mM KCl, 1.2 mM
MgSO4, 1mMCaCl2, 1.6 mMNa2HPO4, 0.4 mMNaH2PO4, 10.5mM glucose, and 32.5 mM HEPES acid; pH was adjusted to 7.4with NaOH. The whole cell patch configuration was estab-lished as described previously (35). Currents were recordedby holding the cell at �60 mV and clamped sequentially tomembrane potentials between �100 mV and �80 mV in20-mV increments.To characterize the ion selectivity of the whole cell currents
and calculate the relative permeability for Na� and K�, bathsolutionswere used as described previously (35), i.e. solutionsAand B that contained Na� and K�, respectively, as the majorcation. Bath solution A contained 140 mM NaCl, 5 mM KCl, 1mM CaCl2, 1 mM MgCl2, 10 mM HEPES, and 5 mM dextrose.Bath solution B contained 140 mM KCl, 1 mM CaCl2, 1 mM
MgCl2, 10 mMHEPES, and 5 mM dextrose. The pH of the solu-tions was adjusted with NaOH or KOH to 7.5. The solution ofthe entire chamber was perfused, whereas the cell was still inwhole cell patch clamp mode.Migration Assay—D54-MG cells were transfected with
YFP-tagged dominant negative cDNAs for ASIC1, �ENaC,�ENaC, or with eYFP for the control group using electropo-ration as described above. Cells were transfected for 48 hbefore use and then detached (phosphate-buffered saline� 1mM EDTA) and counted using a hemocytometer. 50,000YFP-labeled cells transfected with either test or controlcDNAs were replated onto high throughput screening Flu-oroblokTM (BD Bioscience) inserts coated on the undersidewith 5 �g/ml vitronectin. High throughput screening Flu-oroblok inserts have a special polyethylene terephthalatetrack-etched polymer coating on the membrane whichblocks the passage of light through the membrane so onlyfluorescent cells that have migrated through the filter pores(8-�m diameter) were visualized. Cells were allowed tomigrate for 24 h and fixed with 4% paraformaldehyde andvisualized using fluorescence. Because GFP and YFP fluoro-phores have overlapping emission spectra, YFP-tagged cellscan be visualized using a fluorescein isothiocyanate filter.
ENaC/Deg Subunits and Glioblastoma
24528 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 284 • NUMBER 36 • SEPTEMBER 4, 2009 by guest on June 1, 2013http://www.jbc.org/Downloaded from
YFP-expressing cells that had migrated through the pores tothe other side of the filter were counted in five random fieldsper filter using a Nikon TE200V microscope equipped forepifluorescence. Three filters equal an n of 1.Statistics—All experiments were repeated aminimum of three
times. Differences between groups were assessed using standardstatistical tests, primarily Student’s t test. Data are presented asthe means � 1 S.D. Significance was set at p � 0.05.
RESULTS
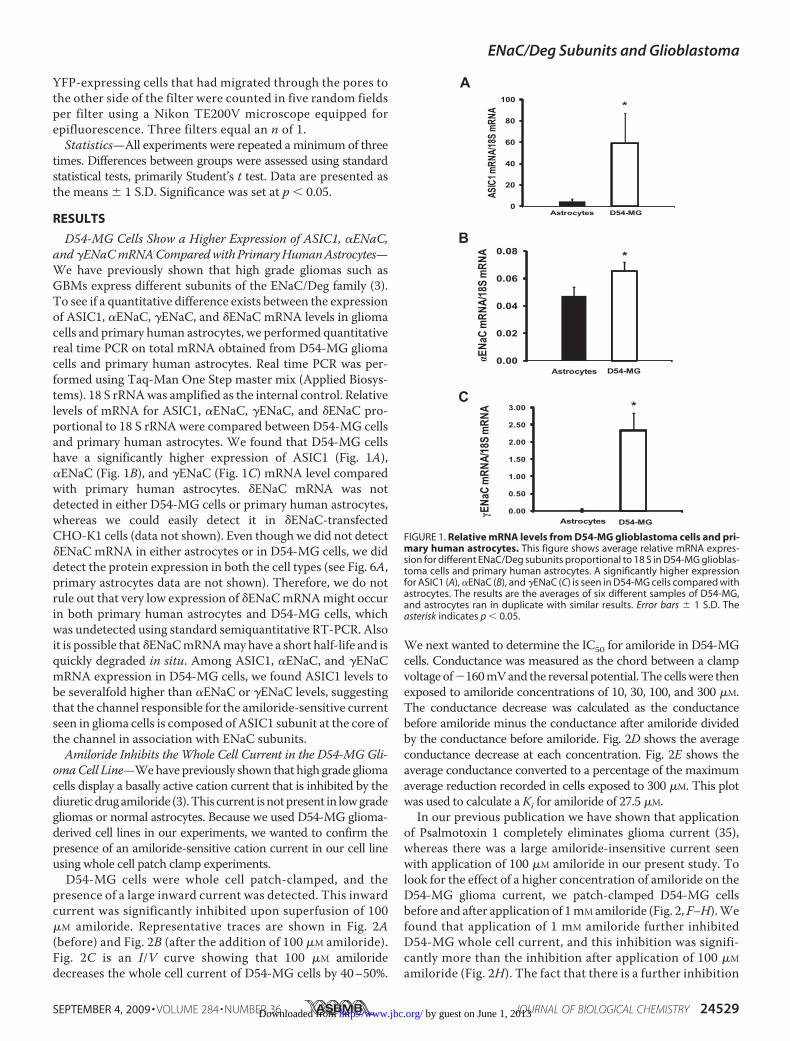
D54-MG Cells Show a Higher Expression of ASIC1, �ENaC,and�ENaCmRNAComparedwithPrimaryHumanAstrocytes—We have previously shown that high grade gliomas such asGBMs express different subunits of the ENaC/Deg family (3).To see if a quantitative difference exists between the expressionof ASIC1, �ENaC, �ENaC, and �ENaC mRNA levels in gliomacells and primary human astrocytes, we performed quantitativereal time PCR on total mRNA obtained from D54-MG gliomacells and primary human astrocytes. Real time PCR was per-formed using Taq-Man One Step master mix (Applied Biosys-tems). 18 S rRNAwas amplified as the internal control. Relativelevels of mRNA for ASIC1, �ENaC, �ENaC, and �ENaC pro-portional to 18 S rRNA were compared between D54-MG cellsand primary human astrocytes. We found that D54-MG cellshave a significantly higher expression of ASIC1 (Fig. 1A),�ENaC (Fig. 1B), and �ENaC (Fig. 1C) mRNA level comparedwith primary human astrocytes. �ENaC mRNA was notdetected in either D54-MG cells or primary human astrocytes,whereas we could easily detect it in �ENaC-transfectedCHO-K1 cells (data not shown). Even though we did not detect�ENaC mRNA in either astrocytes or in D54-MG cells, we diddetect the protein expression in both the cell types (see Fig. 6A,primary astrocytes data are not shown). Therefore, we do notrule out that very low expression of �ENaCmRNAmight occurin both primary human astrocytes and D54-MG cells, whichwas undetected using standard semiquantitative RT-PCR. Alsoit is possible that �ENaCmRNAmayhave a short half-life and isquickly degraded in situ. Among ASIC1, �ENaC, and �ENaCmRNA expression in D54-MG cells, we found ASIC1 levels tobe severalfold higher than �ENaC or �ENaC levels, suggestingthat the channel responsible for the amiloride-sensitive currentseen in glioma cells is composed of ASIC1 subunit at the core ofthe channel in association with ENaC subunits.Amiloride Inhibits the Whole Cell Current in the D54-MG Gli-
omaCell Line—Wehave previously shown that high grade gliomacells display a basally active cation current that is inhibited by thediureticdrugamiloride (3).This current isnotpresent in lowgradegliomas or normal astrocytes. Because we used D54-MG glioma-derived cell lines in our experiments, we wanted to confirm thepresence of an amiloride-sensitive cation current in our cell lineusing whole cell patch clamp experiments.D54-MG cells were whole cell patch-clamped, and the
presence of a large inward current was detected. This inwardcurrent was significantly inhibited upon superfusion of 100�M amiloride. Representative traces are shown in Fig. 2A(before) and Fig. 2B (after the addition of 100 �M amiloride).Fig. 2C is an I/V curve showing that 100 �M amiloridedecreases the whole cell current of D54-MG cells by 40–50%.
We next wanted to determine the IC50 for amiloride in D54-MGcells. Conductance was measured as the chord between a clampvoltageof�160mVand the reversal potential. The cellswere thenexposed to amiloride concentrations of 10, 30, 100, and 300 �M.The conductance decrease was calculated as the conductancebefore amiloride minus the conductance after amiloride dividedby the conductance before amiloride. Fig. 2D shows the averageconductance decrease at each concentration. Fig. 2E shows theaverage conductance converted to a percentage of the maximumaverage reduction recorded in cells exposed to 300 �M. This plotwas used to calculate aKi for amiloride of 27.5 �M.
In our previous publication we have shown that applicationof Psalmotoxin 1 completely eliminates glioma current (35),whereas there was a large amiloride-insensitive current seenwith application of 100 �M amiloride in our present study. Tolook for the effect of a higher concentration of amiloride on theD54-MG glioma current, we patch-clamped D54-MG cellsbefore and after application of 1mMamiloride (Fig. 2, F–H).Wefound that application of 1 mM amiloride further inhibitedD54-MG whole cell current, and this inhibition was signifi-cantly more than the inhibition after application of 100 �M
amiloride (Fig. 2H). The fact that there is a further inhibition
FIGURE 1. Relative mRNA levels from D54-MG glioblastoma cells and pri-mary human astrocytes. This figure shows average relative mRNA expres-sion for different ENaC/Deg subunits proportional to 18 S in D54-MG glioblas-toma cells and primary human astrocytes. A significantly higher expressionfor ASIC1 (A), �ENaC (B), and �ENaC (C) is seen in D54-MG cells compared withastrocytes. The results are the averages of six different samples of D54-MG,and astrocytes ran in duplicate with similar results. Error bars � 1 S.D. Theasterisk indicates p � 0.05.
ENaC/Deg Subunits and Glioblastoma
SEPTEMBER 4, 2009 • VOLUME 284 • NUMBER 36 JOURNAL OF BIOLOGICAL CHEMISTRY 24529 by guest on June 1, 2013http://www.jbc.org/Downloaded from
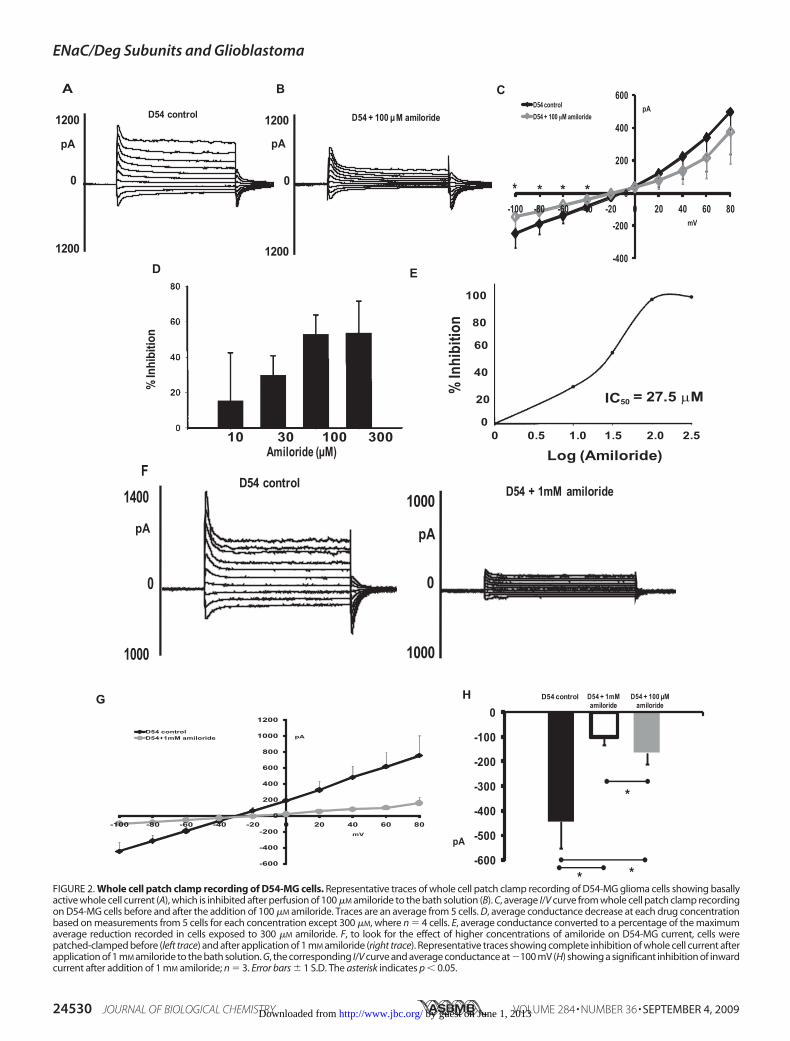
FIGURE 2. Whole cell patch clamp recording of D54-MG cells. Representative traces of whole cell patch clamp recording of D54-MG glioma cells showing basallyactive whole cell current (A), which is inhibited after perfusion of 100 �M amiloride to the bath solution (B). C, average I/V curve from whole cell patch clamp recordingon D54-MG cells before and after the addition of 100 �M amiloride. Traces are an average from 5 cells. D, average conductance decrease at each drug concentrationbased on measurements from 5 cells for each concentration except 300 �M, where n � 4 cells. E, average conductance converted to a percentage of the maximumaverage reduction recorded in cells exposed to 300 �M amiloride. F, to look for the effect of higher concentrations of amiloride on D54-MG current, cells werepatched-clamped before (left trace) and after application of 1 mM amiloride (right trace). Representative traces showing complete inhibition of whole cell current afterapplication of 1 mM amiloride to the bath solution. G, the corresponding I/V curve and average conductance at �100 mV (H) showing a significant inhibition of inwardcurrent after addition of 1 mM amiloride; n � 3. Error bars � 1 S.D. The asterisk indicates p � 0.05.
ENaC/Deg Subunits and Glioblastoma
24530 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 284 • NUMBER 36 • SEPTEMBER 4, 2009 by guest on June 1, 2013http://www.jbc.org/Downloaded from
of glioma cell whole cell current upon application of 1 mM
amiloride after initial saturation of the inhibition at 100 �M
suggests that the glioma channel shows a bimodal inhibitorycurve to amiloride.To confirm that the current seen in D54-MG cells was sen-
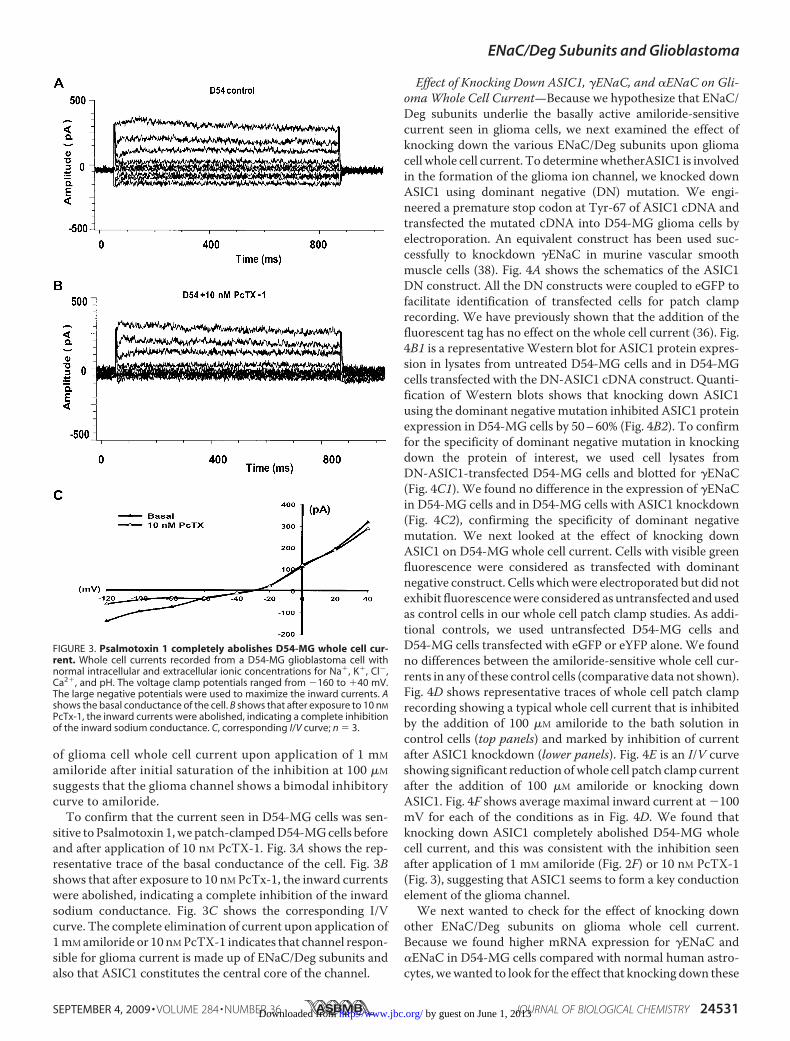
sitive to Psalmotoxin 1,we patch-clampedD54-MGcells beforeand after application of 10 nM PcTX-1. Fig. 3A shows the rep-resentative trace of the basal conductance of the cell. Fig. 3Bshows that after exposure to 10 nM PcTx-1, the inward currentswere abolished, indicating a complete inhibition of the inwardsodium conductance. Fig. 3C shows the corresponding I/Vcurve. The complete elimination of current upon application of1mMamiloride or 10 nMPcTX-1 indicates that channel respon-sible for glioma current is made up of ENaC/Deg subunits andalso that ASIC1 constitutes the central core of the channel.
Effect of Knocking Down ASIC1, �ENaC, and �ENaC on Gli-omaWhole Cell Current—Because we hypothesize that ENaC/Deg subunits underlie the basally active amiloride-sensitivecurrent seen in glioma cells, we next examined the effect ofknocking down the various ENaC/Deg subunits upon gliomacell whole cell current. To determinewhetherASIC1 is involvedin the formation of the glioma ion channel, we knocked downASIC1 using dominant negative (DN) mutation. We engi-neered a premature stop codon at Tyr-67 of ASIC1 cDNA andtransfected the mutated cDNA into D54-MG glioma cells byelectroporation. An equivalent construct has been used suc-cessfully to knockdown �ENaC in murine vascular smoothmuscle cells (38). Fig. 4A shows the schematics of the ASIC1DN construct. All the DN constructs were coupled to eGFP tofacilitate identification of transfected cells for patch clamprecording. We have previously shown that the addition of thefluorescent tag has no effect on the whole cell current (36). Fig.4B1 is a representativeWestern blot for ASIC1 protein expres-sion in lysates from untreated D54-MG cells and in D54-MGcells transfected with the DN-ASIC1 cDNA construct. Quanti-fication of Western blots shows that knocking down ASIC1using the dominant negativemutation inhibited ASIC1 proteinexpression in D54-MG cells by 50–60% (Fig. 4B2). To confirmfor the specificity of dominant negative mutation in knockingdown the protein of interest, we used cell lysates fromDN-ASIC1-transfected D54-MG cells and blotted for �ENaC(Fig. 4C1). We found no difference in the expression of �ENaCin D54-MG cells and in D54-MG cells with ASIC1 knockdown(Fig. 4C2), confirming the specificity of dominant negativemutation. We next looked at the effect of knocking downASIC1 on D54-MG whole cell current. Cells with visible greenfluorescence were considered as transfected with dominantnegative construct. Cells whichwere electroporated but did notexhibit fluorescencewere considered as untransfected andusedas control cells in our whole cell patch clamp studies. As addi-tional controls, we used untransfected D54-MG cells andD54-MG cells transfected with eGFP or eYFP alone. We foundno differences between the amiloride-sensitive whole cell cur-rents in any of these control cells (comparative data not shown).Fig. 4D shows representative traces of whole cell patch clamprecording showing a typical whole cell current that is inhibitedby the addition of 100 �M amiloride to the bath solution incontrol cells (top panels) and marked by inhibition of currentafter ASIC1 knockdown (lower panels). Fig. 4E is an I/V curveshowing significant reduction ofwhole cell patch clamp currentafter the addition of 100 �M amiloride or knocking downASIC1. Fig. 4F shows average maximal inward current at �100mV for each of the conditions as in Fig. 4D. We found thatknocking down ASIC1 completely abolished D54-MG wholecell current, and this was consistent with the inhibition seenafter application of 1 mM amiloride (Fig. 2F) or 10 nM PcTX-1(Fig. 3), suggesting that ASIC1 seems to form a key conductionelement of the glioma channel.We next wanted to check for the effect of knocking down
other ENaC/Deg subunits on glioma whole cell current.Because we found higher mRNA expression for �ENaC and�ENaC in D54-MG cells compared with normal human astro-cytes, wewanted to look for the effect that knocking down these
FIGURE 3. Psalmotoxin 1 completely abolishes D54-MG whole cell cur-rent. Whole cell currents recorded from a D54-MG glioblastoma cell withnormal intracellular and extracellular ionic concentrations for Na�, K�, Cl�,Ca2�, and pH. The voltage clamp potentials ranged from �160 to �40 mV.The large negative potentials were used to maximize the inward currents. Ashows the basal conductance of the cell. B shows that after exposure to 10 nM
PcTx-1, the inward currents were abolished, indicating a complete inhibitionof the inward sodium conductance. C, corresponding I/V curve; n � 3.
ENaC/Deg Subunits and Glioblastoma
SEPTEMBER 4, 2009 • VOLUME 284 • NUMBER 36 JOURNAL OF BIOLOGICAL CHEMISTRY 24531 by guest on June 1, 2013http://www.jbc.org/Downloaded from
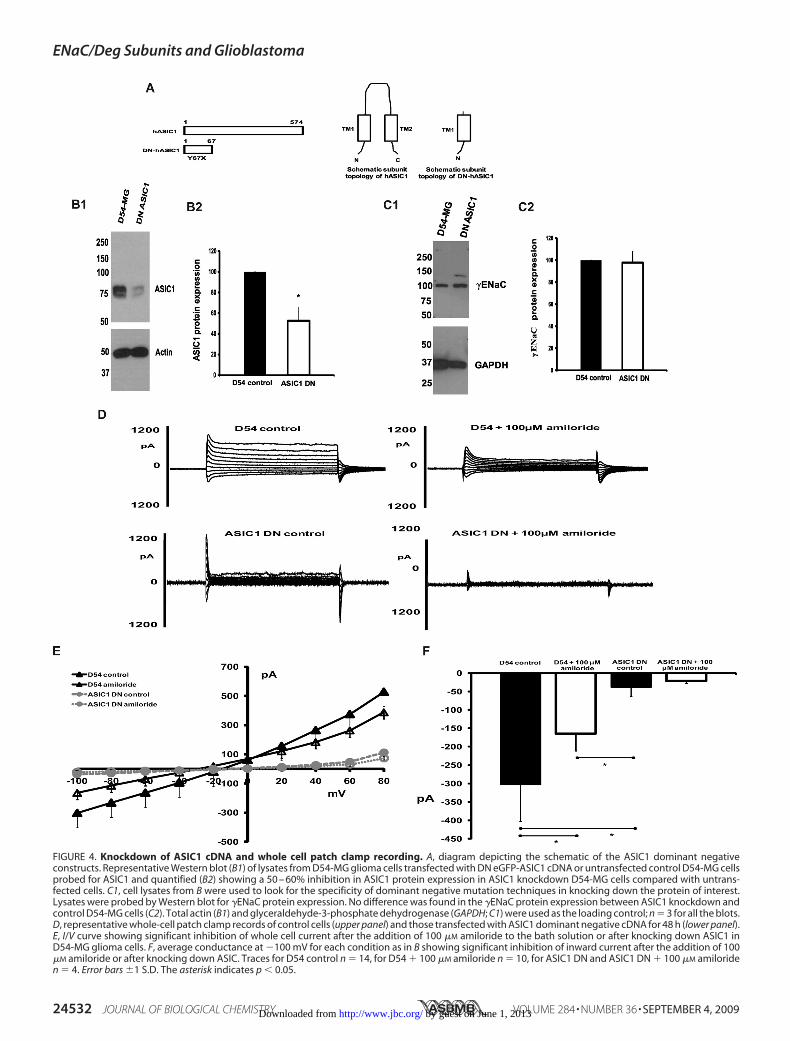
FIGURE 4. Knockdown of ASIC1 cDNA and whole cell patch clamp recording. A, diagram depicting the schematic of the ASIC1 dominant negativeconstructs. Representative Western blot (B1) of lysates from D54-MG glioma cells transfected with DN eGFP-ASIC1 cDNA or untransfected control D54-MG cellsprobed for ASIC1 and quantified (B2) showing a 50 – 60% inhibition in ASIC1 protein expression in ASIC1 knockdown D54-MG cells compared with untrans-fected cells. C1, cell lysates from B were used to look for the specificity of dominant negative mutation techniques in knocking down the protein of interest.Lysates were probed by Western blot for �ENaC protein expression. No difference was found in the �ENaC protein expression between ASIC1 knockdown andcontrol D54-MG cells (C2). Total actin (B1) and glyceraldehyde-3-phosphate dehydrogenase (GAPDH; C1) were used as the loading control; n � 3 for all the blots.D, representative whole-cell patch clamp records of control cells (upper panel) and those transfected with ASIC1 dominant negative cDNA for 48 h (lower panel).E, I/V curve showing significant inhibition of whole cell current after the addition of 100 �M amiloride to the bath solution or after knocking down ASIC1 inD54-MG glioma cells. F, average conductance at �100 mV for each condition as in B showing significant inhibition of inward current after the addition of 100�M amiloride or after knocking down ASIC. Traces for D54 control n � 14, for D54 � 100 �M amiloride n � 10, for ASIC1 DN and ASIC1 DN � 100 �M amiloriden � 4. Error bars �1 S.D. The asterisk indicates p � 0.05.
ENaC/Deg Subunits and Glioblastoma
24532 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 284 • NUMBER 36 • SEPTEMBER 4, 2009 by guest on June 1, 2013http://www.jbc.org/Downloaded from

two subunits would have on glioma cell current. We knockeddown �ENaC by constructing a dominant negative cDNA for�ENaC (eYFP-S155X) similar to the DN-ASIC1 construct andtransfecting the mutated cDNA in D54-MG cells (Fig. 5A).Western blot for�ENaC showed a 60–70% reduction in proteinexpression in D54-MG cells transfected with DN-�ENaC com-pared with untransfected D54-MG cells (Fig. 5, B1 and B2). Tocheck for the specificity of the dominant negative mutation inknocking down only �ENaC, we probed for the expression ofASIC1 (Fig. 5C1) and �ENaC (Fig. 5D1) protein levels in lysatesfrom untransfected and transfected D54-MG cells and foundno difference in the expression level of either ASIC1 (Fig. 5C2)or �ENaC (Fig. 5D2).
To study the effect of knocking down �ENaC on the gli-oma cell current, we patch-clamped control and �ENaCknocked-down D54-MG cells. D54-MG control cells exhib-ited a whole cell current that was inhibited by 100 �M amilo-ride (Fig. 5E, representative traces, top panel). The amilo-ride-sensitive current seen in D54-MG cells was significantlyinhibited after knocking down �ENaC in transfected cells ascompared with control cells (representative traces, lowerpanel). Fig. 5F shows the corresponding I/V curve. Fig. 5Gshows the average maximal inward current at �100 mV foreach of the conditions illustrated in Fig. 5E.We next knocked down �ENaC in D54-MG cells using a dom-
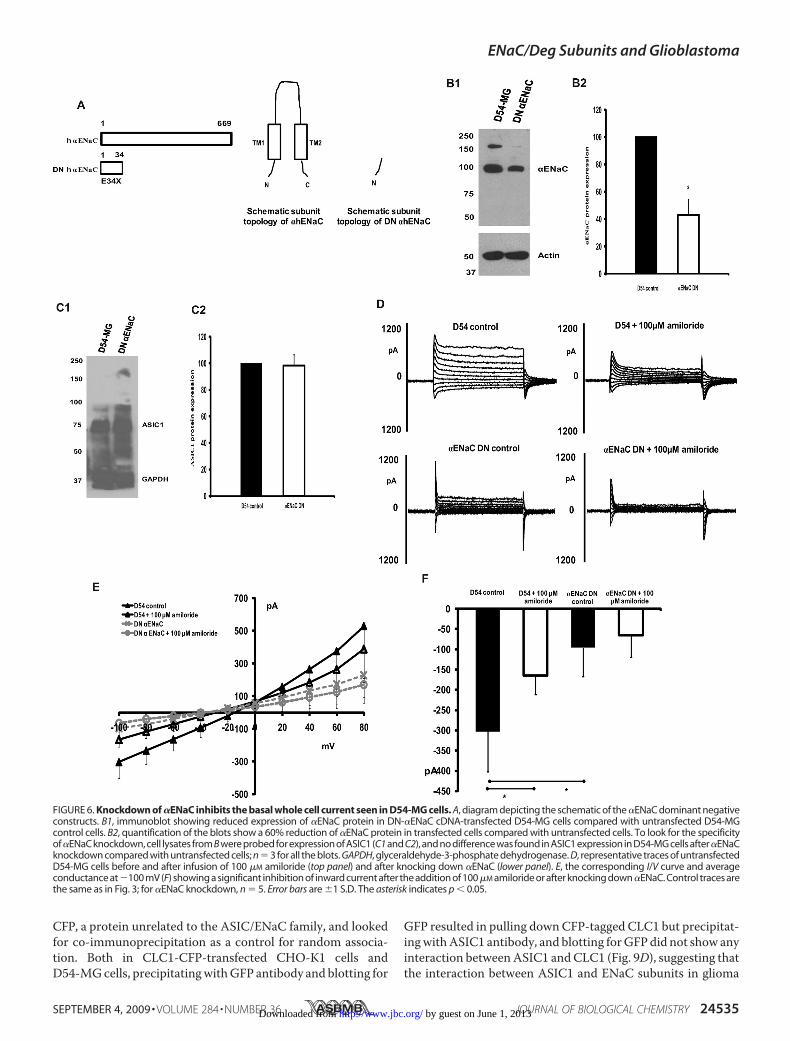
inantnegative cDNAfor�ENaC(eYFP-E34X) (Fig. 6A) and founda60%reductionof�ENaCprotein levels as comparedwithcontrolD54-MG cells, as shown by Western blot (Fig. 6, B1 and B2).Knocking down �ENaC did not change the expression of ASIC1protein level (Fig. 6,C1 andC2), again confirming the specificity ofthe dominant negative mutation in knocking down the protein ofinterest. Similar to what we saw for �ENaC, knocking down�ENaC significantly inhibited the amiloride-sensitive whole cellcurrent seen in D54-MG glioma cells (Fig. 6, D–F). Knockingdowneither�ENaCor�ENaCreduced thewhole cell patchclampcurrent seen in D54-MG cells to a larger extent than after theaddition of 100 �M amiloride. Because knocking down either�ENaC or �ENaC significantly reduced the glioma cell conduct-ance, both �ENaC and�ENaC seem to formkey elements of theglioma channel.Knocking Down �ENaCDoes Not Change Glioma Cell Whole
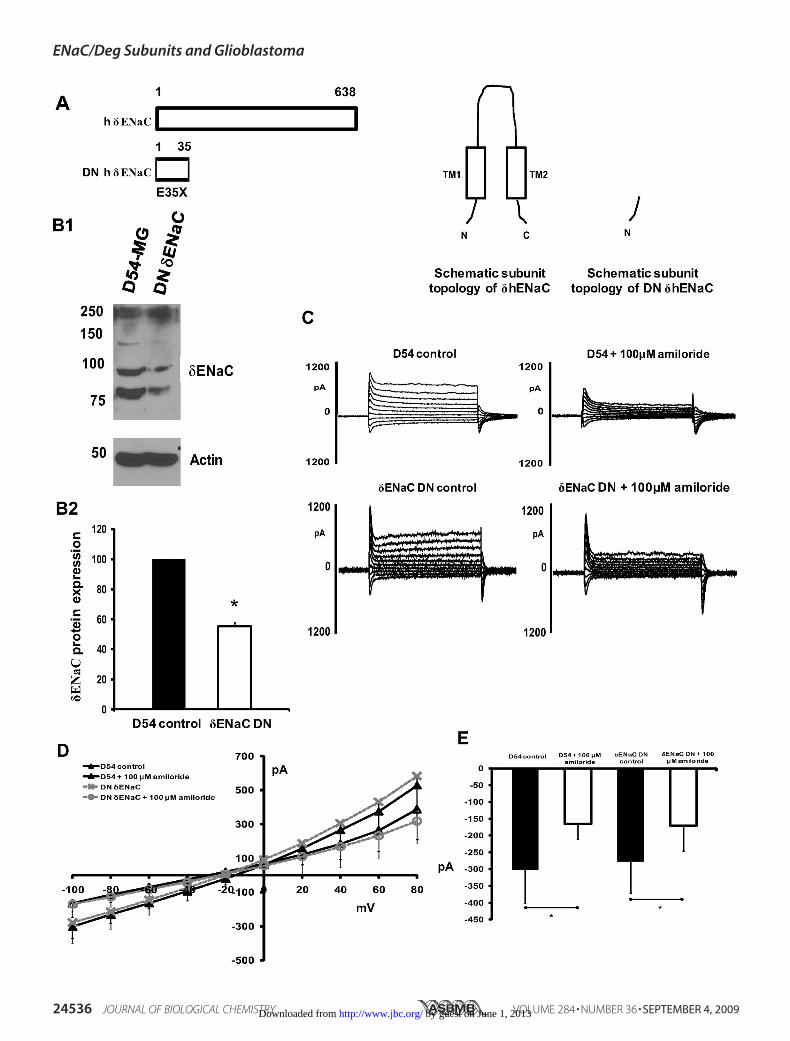
Cell Current—Although we did not detect �ENaC mRNAexpression levels in GBM cells or in normal astrocytes as theprotein was present, we wanted to test whether knockingdown �ENaC would have an effect upon GBM whole cellcurrents. eYFP-DN-�ENaC (S35X) cDNA was transfectedinto D54-MG glioma cells (Fig. 7A shows the schematics ofDN-�ENaC construct), and cells were whole cell patch-clamped. We saw a 50% reduction in protein expression for�ENaC in cells transfectedwith theDNconstruct comparedwithuntransfected control cells as shown by Western blot (Fig. 7, B1and B2). Even though �ENaC protein expression was reduced incells transfected with DN �ENaC cDNA, no change in whole cellpatch clamp currents between control cells and cells with �ENaCknockdown was observed. Fig. 7C shows representative tracesbefore (top panel) and after (lower panel) knocking down�ENaC in D54-MG cells. Fig. 7,D and E, show the correspond-ing I/V curve and average inward current at �100 mV, respec-
tively, for each of the conditions shown in Fig. 7C. Becauseknocking down �ENaC causes no significant change in GBMwhole cell current, we conclude that even though �ENaC isexpressed in glioma cells, �ENaC is not a part of the gliomachannel complex.Glioma Cells Have a Higher K� Selectivity over Na� That Is
Abolished after Knocking Down �ENaC and �ENaC—In a pre-vious publicationwe showed that high grade glioma cells have ahigher ionic selectivity for K� over Na� (35). In that study cellsfrom three different high grade glioma cell lines (SK-MG,U251-MG, and U87-MG) were whole cell patch-clamped and,whereas in the patch clampconfiguration, the bath solutionwaschanged sequentially by perfusing the entire chamber withsolutions containing Li�, Ca2�, or K� as the major cationinstead of Na�. In every case when Na� was substituted withK� as the major cation, a large increase in amiloride-sensitiveinward current was observed. Using similar experimental con-ditions, we found a significant large increase in inward currentin D54-MG cells when Na� was substituted with K� (Fig. 8).This increase in inward current was abolished in D54-MG cellsin which �ENaC or �ENaC were knocked down. Cells with�ENaC knockdown showed a significant increase in inwardcurrent upon substituting K� as the major cation similar to theeffect seen in control cells. These data further suggest thatknocking down �ENaC or �ENaC changes important electro-physiological properties of glioma cells in addition to reducingthe whole cell current, whereas cells with �ENaC knockeddown behave similarly to the control cells.Co-immunoprecipitation of ASIC1 and ENaC Subunits in
Glioma Cells—Our hypothesis is that different ENaC/Deg sub-units interact with each other to form a channel complex inglioma cells, giving these cells their unique properties. To probefor this interactionweusedco-immunoprecipitation to look for aninteractionbetweenASIC1,�ENaC, and�ENaC inD54-MGcells.Endogenous glioma cells have a relatively low level of ENaC/Degproteinexpression.Toovercomethis limitation,weoverexpressedoneof the putative subunits,ASIC1, and epitope-tagged it to allowfor more sensitive immunoprecipitation and immunodetection.Total cellmembrane andplasmamembrane fractions ofD54-MGcells stably transfected with ASIC1-GFP were obtained using themethods detailed above. Lysate from CHO-K1 cells transfectedwith ASIC1-GFP served as positive controls. Both in CHO-K1(Fig. 9A) cells and in total membrane (Fig. 9B) and plasmamembrane (Fig. 9C), isolates of D54-MG cells immunoprecipi-tating with GFP or ASIC1 antibodies and blotting with GFPpulled down GFP-ASIC1 at 100 kDa, consistent with theexpected size of GFP tagged ASIC1. Furthermore, in both themembrane fractions of ASIC1-GFP-overexpressing D54-MGcells, immunoprecipitating with �ENaC or �ENaC antibodiesand blotting with GFP also resulted in pulling down GFP-ASIC1 at 100 kDa, suggesting an interaction between ASIC1and�ENaCandbetweenASIC1 and�ENaC inD54-MGgliomacells (Fig. 9, B and C).One of the inherent limitations of overexpression studies is the
occurrenceof randomornonspecificassociations.Toruleout ran-dom association resulting from overexpression as a cause ofASIC1/ENaC interaction, we transfected D54-MG cells andCHO-K1 cells with cDNA for sarcolemmal Cl� channel, CLC1-
ENaC/Deg Subunits and Glioblastoma
SEPTEMBER 4, 2009 • VOLUME 284 • NUMBER 36 JOURNAL OF BIOLOGICAL CHEMISTRY 24533 by guest on June 1, 2013http://www.jbc.org/Downloaded from
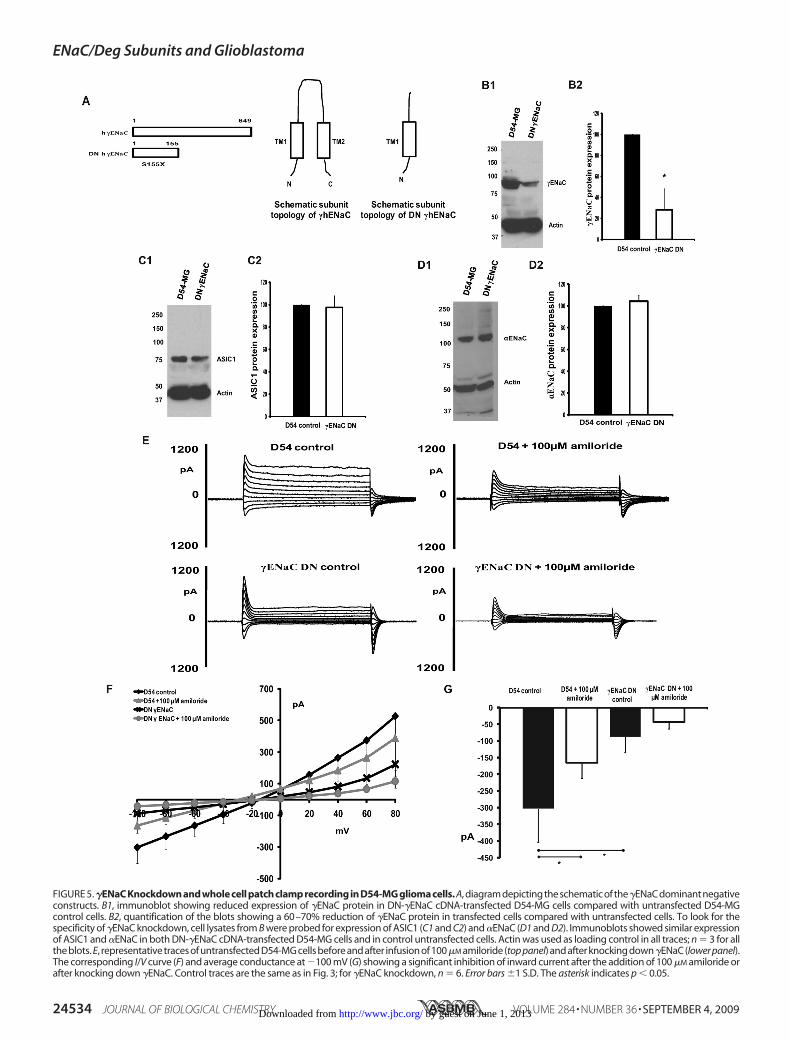
FIGURE 5. �ENaC Knockdown and whole cell patch clamp recording in D54-MG glioma cells. A, diagram depicting the schematic of the�ENaC dominant negativeconstructs. B1, immunoblot showing reduced expression of �ENaC protein in DN-�ENaC cDNA-transfected D54-MG cells compared with untransfected D54-MGcontrol cells. B2, quantification of the blots showing a 60–70% reduction of �ENaC protein in transfected cells compared with untransfected cells. To look for thespecificity of �ENaC knockdown, cell lysates from B were probed for expression of ASIC1 (C1 and C2) and �ENaC (D1 and D2). Immunoblots showed similar expressionof ASIC1 and �ENaC in both DN-�ENaC cDNA-transfected D54-MG cells and in control untransfected cells. Actin was used as loading control in all traces; n � 3 for allthe blots. E, representative traces of untransfected D54-MG cells before and after infusion of 100�M amiloride (top panel) and after knocking down�ENaC (lower panel).The corresponding I/V curve (F) and average conductance at �100 mV (G) showing a significant inhibition of inward current after the addition of 100 �M amiloride orafter knocking down �ENaC. Control traces are the same as in Fig. 3; for �ENaC knockdown, n � 6. Error bars �1 S.D. The asterisk indicates p � 0.05.
ENaC/Deg Subunits and Glioblastoma
24534 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 284 • NUMBER 36 • SEPTEMBER 4, 2009 by guest on June 1, 2013http://www.jbc.org/Downloaded from
CFP, a protein unrelated to the ASIC/ENaC family, and lookedfor co-immunoprecipitation as a control for random associa-tion. Both in CLC1-CFP-transfected CHO-K1 cells andD54-MGcells, precipitatingwithGFP antibody and blotting for
GFP resulted in pulling downCFP-tagged CLC1 but precipitat-ingwithASIC1 antibody, and blotting forGFPdid not show anyinteraction betweenASIC1 andCLC1 (Fig. 9D), suggesting thatthe interaction between ASIC1 and ENaC subunits in glioma
FIGURE 6. Knockdown of �ENaC inhibits the basal whole cell current seen in D54-MG cells. A, diagram depicting the schematic of the �ENaC dominant negativeconstructs. B1, immunoblot showing reduced expression of �ENaC protein in DN-�ENaC cDNA-transfected D54-MG cells compared with untransfected D54-MGcontrol cells. B2, quantification of the blots show a 60% reduction of �ENaC protein in transfected cells compared with untransfected cells. To look for the specificityof�ENaC knockdown, cell lysates from B were probed for expression of ASIC1 (C1 and C2), and no difference was found in ASIC1 expression in D54-MG cells after�ENaCknockdown compared with untransfected cells; n � 3 for all the blots. GAPDH, glyceraldehyde-3-phosphate dehydrogenase. D, representative traces of untransfectedD54-MG cells before and after infusion of 100 �M amiloride (top panel) and after knocking down �ENaC (lower panel). E, the corresponding I/V curve and averageconductance at�100 mV (F) showing a significant inhibition of inward current after the addition of 100 �M amiloride or after knocking down �ENaC. Control traces arethe same as in Fig. 3; for �ENaC knockdown, n � 5. Error bars are �1 S.D. The asterisk indicates p � 0.05.
ENaC/Deg Subunits and Glioblastoma
SEPTEMBER 4, 2009 • VOLUME 284 • NUMBER 36 JOURNAL OF BIOLOGICAL CHEMISTRY 24535 by guest on June 1, 2013http://www.jbc.org/Downloaded from
ENaC/Deg Subunits and Glioblastoma
24536 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 284 • NUMBER 36 • SEPTEMBER 4, 2009 by guest on June 1, 2013http://www.jbc.org/Downloaded from
cells was not because of a random association resulting fromoverexpression.We found that even though we had not transfected D54-MG
cells with either �ENaC or �ENaC, both these subunits immu-noprecipitated with ASIC1 in transfected D54-MG cells. Thisresult combined with the electrophysiological data showingthat knocking down these three subunits significantly reducesglioma cell whole cell current strongly suggests that these sub-units come together to form a functional ion channel in gliomacells and that the active channel in the glioma cell contains (atthe minimum) ASIC1, �ENaC, and �ENaC.Knocking Down ASIC1, �ENaC, or �ENaC Inhibits GBM Cell
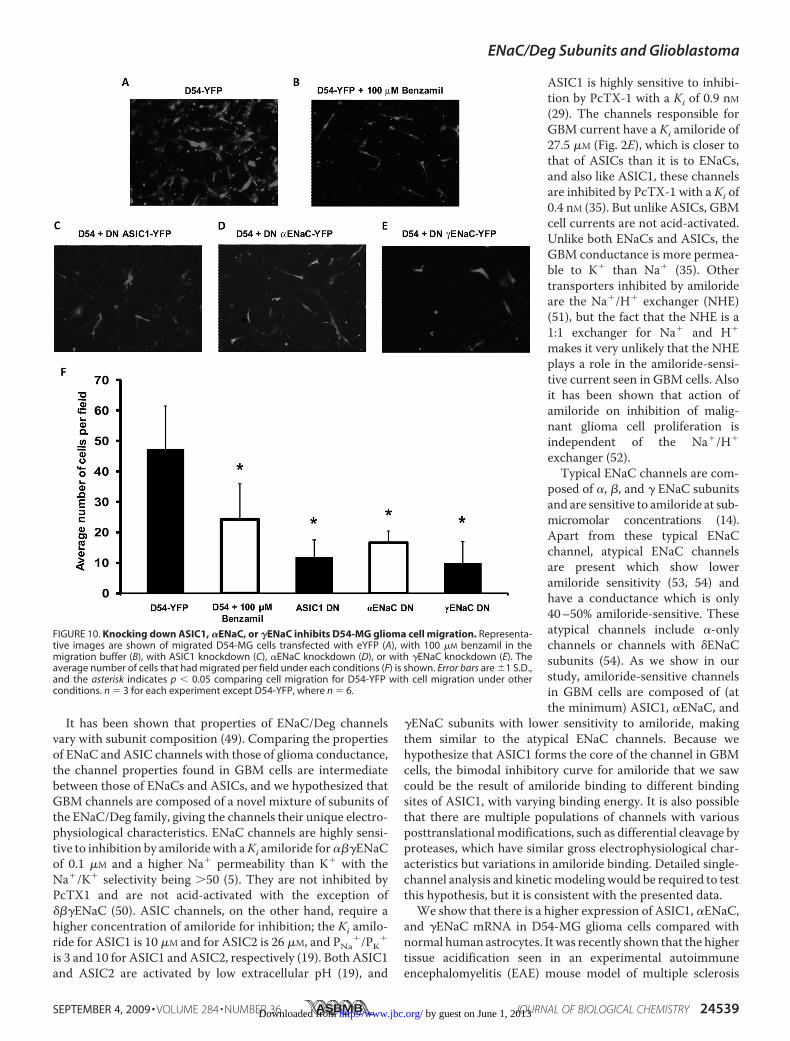
Migration—Because ENaC/Deg family members are hypothe-sized to play an important role in glioma cell biology, we predictthat knocking down the different ENaC/Deg subunits wouldhave an inhibitory effect upon glioma cell migration. To studythemigration properties of glioma cells before and after knock-ing down the different subunits, we used a Transwell migrationassay using high throughput screening Fluoroblok inserts withan 8-�mpore size (BD Bioscience). To allow for sufficient timefor cell migration, we performed a 24-h migration assay.Because we found 24 h application of 100 �M amiloride was
toxic to cells, we substituted 100 �M benzamil (an analogue ofamiloride which inhibits D54-MGwhole cell currents similarlyto amiloride; supplemental Fig. 1) instead of amiloride in themigration assay study. Fig. 10 shows representative images ofmigratedD54-MGcells transfectedwith eYFP (Fig. 10A), trans-fected D54-MG cells with 100 �M benzamil in the migrationbuffer (Fig. 10B), and migrated D54-MG cells with ASIC1knockdown (Fig. 10C), �ENaC knockdown (Fig. 10D), or�ENaC knockdown (Fig. 10E). Fig. 10F shows the average num-ber of cells counted per field for each of the conditions. Wefound that 100 �M benzamil or knocking downASIC1, �ENaC,or �ENaC had a significant inhibitory effect upon D54-MG cellmigration, suggesting that ASIC1 and ENaC subunits play arole in glioma cell migration.
DISCUSSION
In our recent publication we showed that ASIC and ENaCsubunits are capable of forming cross-clade heteromers (36).We found that expression of combinations of ASIC1 withENaC subunits exhibited novel electrophysiological charac-teristics as compared with ASIC1 alone. That study sug-gested that heteromeric complexes of ASIC and ENaC sub-units may contribute to the diversity of amiloride-sensitivecation conductance observed in a wide variety of tissues andcell types where co-expression of ASIC and ENaC subunitshas been observed (36) in addition to differences due to post-translational modification, e.g. phosphorylation (39–43),carboxymethylation (44, 45), and protease clipping (46, 47).In this study we tested the hypothesis that the glioma cationcurrent is comprised of a unique mixture of subunits fromthe ENaC/Degenerin superfamily of ion channels. Our aimwas to assess the role of these different ENaC/Deg subunitsin glioma biology and to probe the composition of thesenovel ENaC/Deg heteromers.We used a dominant negativemutation strategy to knockdown
ASIC1 and ENaC subunits, similar to the approach used by Jerni-gan and Drummond (38). A similar approach had been used tomakeN-terminal-truncated rat�-,�-, and�ENaCsubunits, and itwas shown to reduce INa when co-expressedwithwild type rat�-,�-, and �ENaC (4). Although the mechanism of action of dom-inant negative mutation is not fully understood, some possibil-ities are that the mutated subunit when overexpressed in cellsdisplaces thewild type subunit from ion channel complexes andforms a non-functional ion channel (4, 38). They also causedegradation of wild type subunits, as a reduction of the pro-tein level is seen in dominant negative transfected cells. Theloss of channel function can be explained by the fact that themutated subunits are not full-length proteins and most likelynever get folded into a functional subunit, are unstable, andtherefore, are degraded (38, 48).
FIGURE 7. Knockdown of �ENaC does not change the D54-MG glioma cell whole cell current. A, schematic diagram of �ENaC dominant negative construct.B1, representative Western blot of DN �ENaC and D54-MG cell lysate. B2, quantification of Western blots showing a 50% reduction of �ENaC protein in cellstransfected with dominant negative �ENaC cDNA compared with untransfected D54-MG cells; n � 3. C shows representative traces of untransfected D54-MGcells before and after infusion of 100 �M amiloride (top panel) and after knocking down �ENaC (lower panel). D shows an I/V curve from whole-cell patch clamprecording of DN �ENaC-transfected D54-MG cells showing no difference in whole cell current before or after knocking down �ENaC. E, average conductance at�100 mV for each condition as in Fig. 6B, showing similar inward current for both D54-MG control cells and in D54-MG cells with �ENaC knockdown. Amiloridesignificantly inhibited the inward current under both the conditions. Control traces are the same as in Fig. 3; for �ENaC knockdown, traces n � 5. Error bars are�1 S.D. The asterisk indicates p � 0.05.
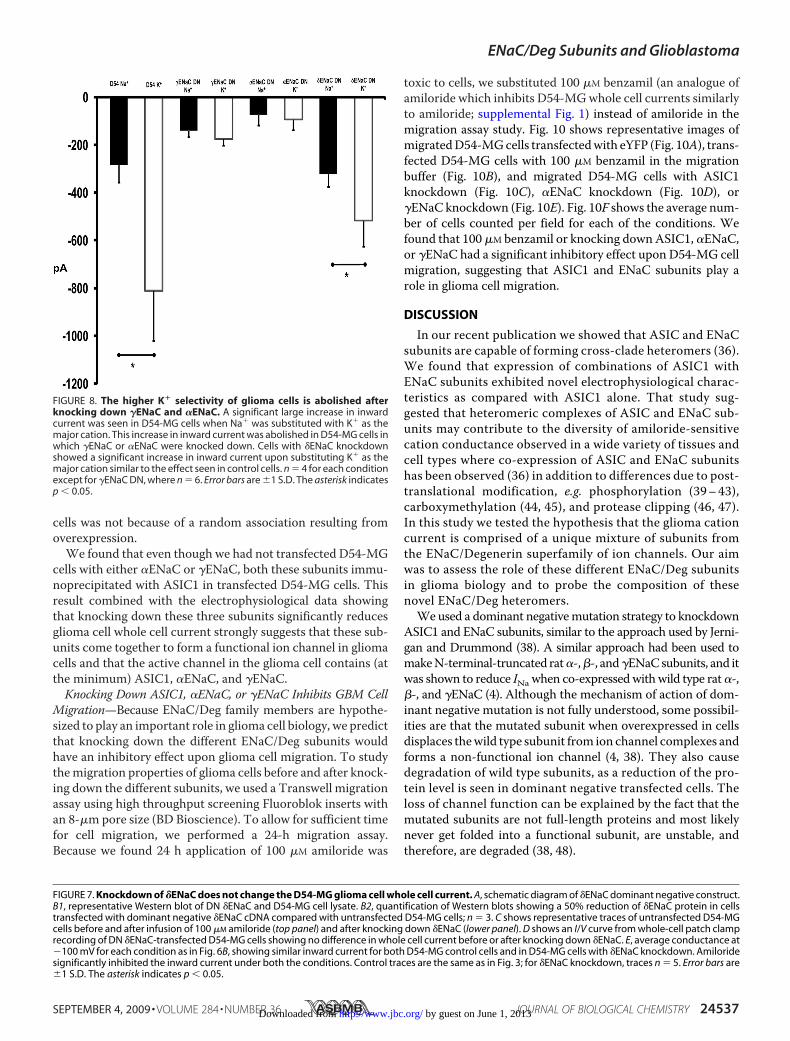
FIGURE 8. The higher K� selectivity of glioma cells is abolished afterknocking down �ENaC and �ENaC. A significant large increase in inwardcurrent was seen in D54-MG cells when Na� was substituted with K� as themajor cation. This increase in inward current was abolished in D54-MG cells inwhich �ENaC or �ENaC were knocked down. Cells with �ENaC knockdownshowed a significant increase in inward current upon substituting K� as themajor cation similar to the effect seen in control cells. n � 4 for each conditionexcept for �ENaC DN, where n � 6. Error bars are �1 S.D. The asterisk indicatesp � 0.05.
ENaC/Deg Subunits and Glioblastoma
SEPTEMBER 4, 2009 • VOLUME 284 • NUMBER 36 JOURNAL OF BIOLOGICAL CHEMISTRY 24537 by guest on June 1, 2013http://www.jbc.org/Downloaded from
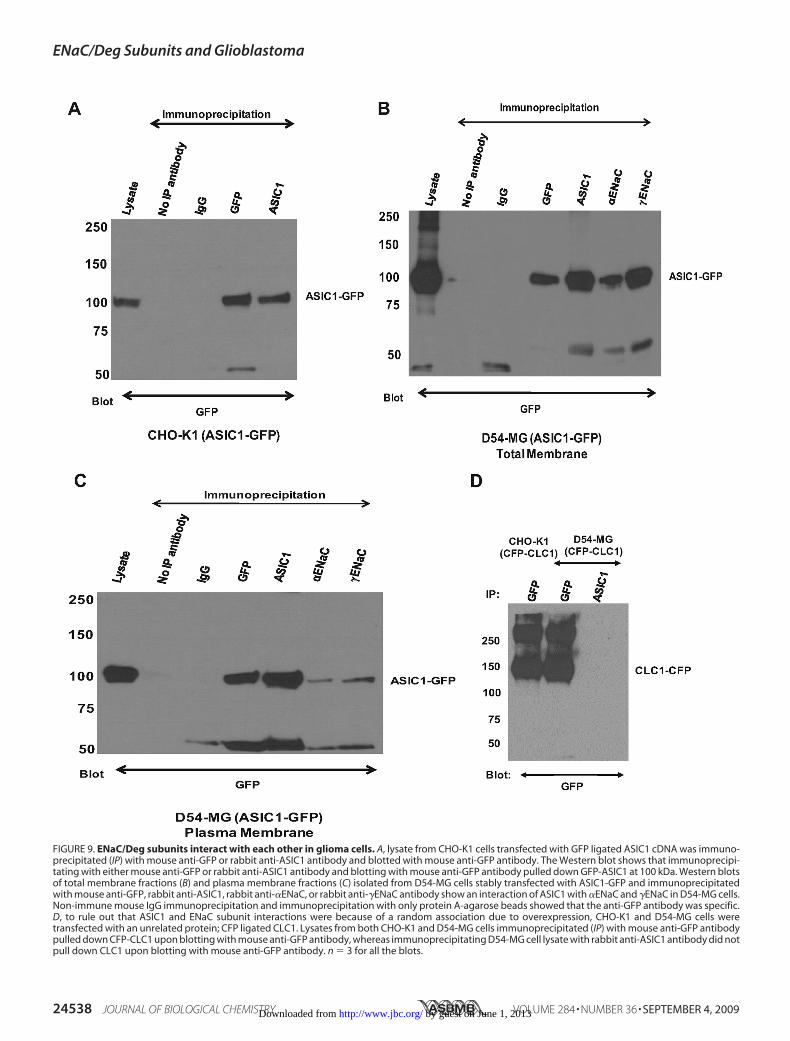
FIGURE 9. ENaC/Deg subunits interact with each other in glioma cells. A, lysate from CHO-K1 cells transfected with GFP ligated ASIC1 cDNA was immuno-precipitated (IP) with mouse anti-GFP or rabbit anti-ASIC1 antibody and blotted with mouse anti-GFP antibody. The Western blot shows that immunoprecipi-tating with either mouse anti-GFP or rabbit anti-ASIC1 antibody and blotting with mouse anti-GFP antibody pulled down GFP-ASIC1 at 100 kDa. Western blotsof total membrane fractions (B) and plasma membrane fractions (C) isolated from D54-MG cells stably transfected with ASIC1-GFP and immunoprecipitatedwith mouse anti-GFP, rabbit anti-ASIC1, rabbit anti-�ENaC, or rabbit anti-�ENaC antibody show an interaction of ASIC1 with �ENaC and �ENaC in D54-MG cells.Non-immune mouse IgG immunoprecipitation and immunoprecipitation with only protein A-agarose beads showed that the anti-GFP antibody was specific.D, to rule out that ASIC1 and ENaC subunit interactions were because of a random association due to overexpression, CHO-K1 and D54-MG cells weretransfected with an unrelated protein; CFP ligated CLC1. Lysates from both CHO-K1 and D54-MG cells immunoprecipitated (IP) with mouse anti-GFP antibodypulled down CFP-CLC1 upon blotting with mouse anti-GFP antibody, whereas immunoprecipitating D54-MG cell lysate with rabbit anti-ASIC1 antibody did notpull down CLC1 upon blotting with mouse anti-GFP antibody. n � 3 for all the blots.
ENaC/Deg Subunits and Glioblastoma
24538 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 284 • NUMBER 36 • SEPTEMBER 4, 2009 by guest on June 1, 2013http://www.jbc.org/Downloaded from
It has been shown that properties of ENaC/Deg channelsvary with subunit composition (49). Comparing the propertiesof ENaC and ASIC channels with those of glioma conductance,the channel properties found in GBM cells are intermediatebetween those of ENaCs and ASICs, and we hypothesized thatGBM channels are composed of a novel mixture of subunits ofthe ENaC/Deg family, giving the channels their unique electro-physiological characteristics. ENaC channels are highly sensi-tive to inhibition by amiloridewith aKi amiloride for���ENaCof 0.1 �M and a higher Na� permeability than K� with theNa�/K� selectivity being �50 (5). They are not inhibited byPcTX1 and are not acid-activated with the exception of���ENaC (50). ASIC channels, on the other hand, require ahigher concentration of amiloride for inhibition; the Ki amilo-ride for ASIC1 is 10 �M and for ASIC2 is 26 �M, and PNa
�/PK�
is 3 and 10 for ASIC1 and ASIC2, respectively (19). Both ASIC1and ASIC2 are activated by low extracellular pH (19), and
ASIC1 is highly sensitive to inhibi-tion by PcTX-1 with a Ki of 0.9 nM(29). The channels responsible forGBM current have a Ki amiloride of27.5 �M (Fig. 2E), which is closer tothat of ASICs than it is to ENaCs,and also like ASIC1, these channelsare inhibited by PcTX-1 with a Ki of0.4 nM (35). But unlike ASICs, GBMcell currents are not acid-activated.Unlike both ENaCs and ASICs, theGBM conductance is more permea-ble to K� than Na� (35). Othertransporters inhibited by amilorideare the Na�/H� exchanger (NHE)(51), but the fact that the NHE is a1:1 exchanger for Na� and H�
makes it very unlikely that the NHEplays a role in the amiloride-sensi-tive current seen in GBM cells. Alsoit has been shown that action ofamiloride on inhibition of malig-nant glioma cell proliferation isindependent of the Na�/H�
exchanger (52).Typical ENaC channels are com-
posed of �, �, and � ENaC subunitsand are sensitive to amiloride at sub-micromolar concentrations (14).Apart from these typical ENaCchannel, atypical ENaC channelsare present which show loweramiloride sensitivity (53, 54) andhave a conductance which is only40–50% amiloride-sensitive. Theseatypical channels include �-onlychannels or channels with �ENaCsubunits (54). As we show in ourstudy, amiloride-sensitive channelsin GBM cells are composed of (atthe minimum) ASIC1, �ENaC, and
�ENaC subunits with lower sensitivity to amiloride, makingthem similar to the atypical ENaC channels. Because wehypothesize that ASIC1 forms the core of the channel in GBMcells, the bimodal inhibitory curve for amiloride that we sawcould be the result of amiloride binding to different bindingsites of ASIC1, with varying binding energy. It is also possiblethat there are multiple populations of channels with variousposttranslational modifications, such as differential cleavage byproteases, which have similar gross electrophysiological char-acteristics but variations in amiloride binding. Detailed single-channel analysis and kineticmodelingwould be required to testthis hypothesis, but it is consistent with the presented data.We show that there is a higher expression of ASIC1, �ENaC,
and �ENaC mRNA in D54-MG glioma cells compared withnormal human astrocytes. It was recently shown that the highertissue acidification seen in an experimental autoimmuneencephalomyelitis (EAE) mouse model of multiple sclerosis
FIGURE 10. Knocking down ASIC1, �ENaC, or �ENaC inhibits D54-MG glioma cell migration. Representa-tive images are shown of migrated D54-MG cells transfected with eYFP (A), with 100 �M benzamil in themigration buffer (B), with ASIC1 knockdown (C), �ENaC knockdown (D), or with �ENaC knockdown (E). Theaverage number of cells that had migrated per field under each conditions (F) is shown. Error bars are �1 S.D.,and the asterisk indicates p � 0.05 comparing cell migration for D54-YFP with cell migration under otherconditions. n � 3 for each experiment except D54-YFP, where n � 6.
ENaC/Deg Subunits and Glioblastoma
SEPTEMBER 4, 2009 • VOLUME 284 • NUMBER 36 JOURNAL OF BIOLOGICAL CHEMISTRY 24539 by guest on June 1, 2013http://www.jbc.org/Downloaded from

increased the expression and function of ASIC1 in EAE micebrains (55). Although we did not test for tissue acidification inGBMs, we predict that glioma cells growing at a rapid ratewould lead to tissue acidosis and a higher expression of ASIC1similar to what was seen in experimental autoimmune enceph-alomyelitis. We also show a higher expression of �ENaC and�ENaC in GBM cells. Proteolytic cleavage of both these sub-units is required to maximally activate ENaC channels (56).Even though we showed that knocking down ASIC1 signifi-cantly inhibited GBM whole cell current, glioma cells do notexhibit the typical acid induced ASIC1 current. One potentialreason for the lack of acid activation may be that the channel isalready maximally activated, and therefore, a further change in[H�] does not exert an additional stimulus. Another possibilityis that the proton-sensitive region is masked by the presence ofadditional ENaC subunits.One of the important prognostic factors in treatment of
glioblastoma is secondary metastases, as these tumors areknown to infiltrate widely throughout the brain. It has beenshown that for cell migration to occur, the leading edge ofthe cell or lamellipodium needs to expand; movement ofNa� and H2O to the cell interior plays an important role inthis process (57).We have shown previously that both amilo-ride and PcTX1 inhibits glioma cell swelling (58).We have alsoshown that amiloride inhibits glioma cell migration. In thisstudy we show that knocking down ASIC1, �ENaC, or �ENaCinhibits glioma cell migration, possibly because of the inhibitoryeffect upon Na� influx after channel knockdown and the corre-sponding inhibition of cell swelling required for lamellipodiumexpansion.Althoughmuchwork is needed to fully understand theENaC/ASICcross-clade interactions ingliomacells andhowthesechannels relate to each other on a biochemical and molecularlevel, this study represents the first evidence to show the pres-ence of an endogenous functional amiloride-sensitive Na�
channel that is a ENaC/ASIC heteromer.
Acknowledgments—We thankMelissaMcCarthy for taking care of allthe tissue culture needs. We also thank Dr. Wanda Vila-Carriles,Edlira Bashari, Dr. Bakhrom Berdiev, and Dr. Robert Meltzer forhelpful discussions.
REFERENCES1. Maher, E. A., Furnari, F. B., Bachoo, R. M., Rowitch, D. H., Louis, D. N.,
Cavenee, W. K., and DePinho, R. A. (2001) Genes Dev. 15, 1311–13332. Holland, E. C. (2000) Proc. Natl. Acad. Sci. U.S.A. 97, 6242–62443. Berdiev, B. K., Xia, J., McLean, L. A., Markert, J. M., Gillespie, G. Y., Map-
stone, T. B., Naren, A. P., Jovov, B., Bubien, J. K., Ji, H. L., Fuller, C.M., Kirk,K. L., and Benos, D. J. (2003) J. Biol. Chem. 278, 15023–15034
4. Benos, D. J., and Stanton, B. A. (1999) J. Physiol. 520, 631–6445. Kellenberger, S., and Schild, L. (2002) Physiol. Rev. 82, 735–7676. Wemmie, J. A., Askwith, C. C., Lamani, E., Cassell, M. D., Freeman, J. H.,
Jr., and Welsh, M. J. (2003) J. Neurosci. 23, 5496–55027. Snyder, P. M. (2002) Endocr. Rev. 23, 258–2758. Swift, P. A., and MacGregor, G. A. (2004) Am. J. Pharmacogenomics 4,
161–1689. Berdiev, B. K., Qadri, Y. J., and Benos, D. J. (2009)Mol. Biosyst. 5, 123–12710. Mall, M. A. (2008) J. Aerosol Med. Pulm. Drug Deliv. 21, 13–2411. Matalon, S., Lazrak, A., Jain, L., and Eaton, D. C. (2002) J. Appl. Physiol. 93,
1852–185912. Guggino,W. B., and Guggino, S. E. (2000) Proc. Natl. Acad. Sci. U.S.A. 97,
9827–982913. Canessa, C. M., Merillat, A. M., and Rossier, B. C. (1994) Am. J. Physiol.
267, C1682–C169014. Canessa, C.M., Schild, L., Buell, G., Thorens, B., Gautschi, I., Horisberger,
J. D., and Rossier, B. C. (1994) Nature 367, 463–46715. Staruschenko, A., Adams, E., Booth, R. E., and Stockand, J. D. (2005) Bio-
phys. J. 88, 3966–397516. Firsov, D., Gautschi, I., Merillat, A. M., Rossier, B. C., and Schild, L. (1998)
EMBO J. 17, 344–35217. García-Anoveros, J., Derfler, B., Neville-Golden, J., Hyman, B. T., and
Corey, D. P. (1997) Proc. Natl. Acad. Sci. U.S.A. 94, 1459–146418. Wemmie, J. A., Price, M. P., and Welsh, M. J. (2006) Trends Neurosci. 29,
578–58619. Krishtal, O. (2003) Trends Neurosci. 26, 477–48320. Brockway, L. M., Zhou, Z. H., Bubien, J. K., Jovov, B., Benos, D. J., and
Keyser, K. T. (2002) Am. J. Physiol. Cell Physiol. 283, C126–C13421. Ettaiche, M., Guy, N., Hofman, P., Lazdunski, M., and Waldmann, R.
(2004) J. Neurosci. 24, 1005–101222. Lingueglia, E. (2007) J. Biol. Chem. 282, 17325–1732923. Ishibashi, K., and Marumo, F. (1998) Biochem. Biophys. Res. Commun.
245, 589–59324. Grunder, S., Geissler, H. S., Bassler, E. L., and Ruppersberg, J. P. (2000)
Neuroreport 11, 1607–161125. Jahr, H., vanDriel,M., vanOsch, G. J.,Weinans, H., and van Leeuwen, J. P.
(2005) Biochem. Biophys. Res. Commun. 337, 349–35426. Jasti, J., Furukawa, H., Gonzales, E. B., and Gouaux, E. (2007)Nature 449,
316–32327. Waldmann, R., Champigny, G., Bassilana, F., Heurteaux, C., and Lazdun-
ski, M. (1997) Nature 386, 173–17728. Diochot, S., Salinas, M., Baron, A., Escoubas, P., and Lazdunski, M. (2007)
Toxicon 49, 271–28429. Escoubas, P., De Weille, J. R., Lecoq, A., Diochot, S., Waldmann, R.,
Champigny, G., Moinier, D., Menez, A., and Lazdunski, M. (2000) J. Biol.Chem. 275, 25116–25121
30. Naves, L. A., and McCleskey, E. W. (2005) Braz. J. Med. Biol. Res. 38,1561–1569
31. Immke, D. C., and McCleskey, E. W. (2001) ScientificWorldJournal 1,510–512
32. Price, M. P., Lewin, G. R., McIlwrath, S. L., Cheng, C., Xie, J., Heppenstall,P. A., Stucky, C. L., Mannsfeldt, A. G., Brennan, T. J., Drummond, H. A.,Qiao, J., Benson, C. J., Tarr, D. E., Hrstka, R. F., Yang, B.,Williamson, R. A.,and Welsh, M. J. (2000) Nature 407, 1007–1011
33. Lin, W., Ogura, T., and Kinnamon, S. C. (2002) J. Neurophysiol. 88,133–141
34. Wemmie, J. A., Chen, J., Askwith, C. C., Hruska-Hageman, A. M., Price,M. P., Nolan, B. C., Yoder, P. G., Lamani, E., Hoshi, T., Freeman, J. H., Jr.,and Welsh, M. J. (2002) Neuron 34, 463–477
35. Bubien, J. K., Ji, H. L., Gillespie, G. Y., Fuller, C. M., Markert, J. M., Map-stone, T. B., and Benos, D. J. (2004) Am. J. Physiol. Cell Physiol 287,C1282–C1291
36. Meltzer, R. H., Kapoor, N., Qadri, Y. J., Anderson, S. J., Fuller, C. M., andBenos, D. J. (2007) J. Biol. Chem. 282, 25548–25559
37. Berdiev, B. K., Cormet-Boyaka, E., Tousson, A., Qadri, Y. J., Oosterveld-Hut, H. M., Hong, J. S., Gonzales, P. A., Fuller, C. M., Sorscher, E. J.,Lukacs, G. L., and Benos, D. J. (2007) J. Biol. Chem. 282, 36481–36488
38. Jernigan, N. L., andDrummond, H. A. (2006)Am. J. Physiol. Renal Physiol.291, F1184–F1191
39. Chigaev, A., Lu,G., Shi,H., Asher, C., Xu, R., Latter,H., Seger, R., Garty,H.,and Reuveny, E. (2001) Am. J. Physiol. Renal Physiol. 280, F1030–F1036
40. Shi, H., Asher, C., Yung, Y., Kligman, L., Reuveny, E., Seger, R., and Garty,H. (2002) Eur. J. Biochem. 269, 4551–4558
41. Shimkets, R. A., Lifton, R., and Canessa, C.M. (1998) Proc. Natl. Acad. Sci.U.S.A. 95, 3301–3305
42. Yang, L. M., Rinke, R., and Korbmacher, C. (2006) J. Biol. Chem. 281,9859–9868
43. Bashari, E., Qadri, Y. J., Zhou, Z. H., Kapoor, N., Anderson, S. J., Meltzer,R. H., Fuller, C.M., and Benos, D. J. (2009)Am. J. Physiol. Cell Physiol. 296,C372–C384
ENaC/Deg Subunits and Glioblastoma
24540 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 284 • NUMBER 36 • SEPTEMBER 4, 2009 by guest on June 1, 2013http://www.jbc.org/Downloaded from

44. Ismailov, II,McDuffie, J. H., Sariban-Sohraby, S., Johnson, J. P., and Benos,D. J. (1994) J. Biol. Chem. 269, 22193–22197
45. Edinger, R. S., Yospin, J., Perry, C., Kleyman, T. R., and Johnson, J. P. (2006)J. Biol. Chem. 281, 9110–9117
46. Kleyman, T. R.,Myerburg,M.M., andHughey, R. P. (2006)Kidney Int. 70,1391–1392
47. Rossier, B. C. (2004) Proc. Am. Thorac. Soc. 1, 4–948. Adams, C. M., Snyder, P. M., and Welsh, M. J. (1997) J. Biol. Chem. 272,
27295–2730049. Fyfe, G. K., and Canessa, C. M. (1998) J. Gen. Physiol. 112, 423–43250. Ji, H. L., and Benos, D. J. (2004) J. Biol. Chem. 279, 26939–2694751. Masereel, B., Pochet, L., and Laeckmann, D. (2003) Eur. J. Med. Chem. 38,
547–554
52. Hegde, M., Roscoe, J., Cala, P., and Gorin, F. (2004) J. Pharmacol. Exp.Ther. 310, 67–74
53. O’Brodovich, H., Yang, P., Gandhi, S., and Otulakowski, G. (2008) Am. J.Physiol. Lung Cell. Mol. Physiol. 294, L401–L408
54. Folkesson, H. G. (2008) Am. J. Physiol. Lung Cell. Mol. Physiol. 294,L399–L400
55. Friese, M. A., Craner, M. J., Etzensperger, R., Vergo, S., Wemmie, J. A.,Welsh, M. J., Vincent, A., and Fugger, L. (2007) Nat. Med. 13, 1483–1489
56. Hughey, R. P., Carattino, M. D., and Kleyman, T. R. (2007) Curr. Opin.Nephrol. Hypertens. 16, 444–450
57. Schwab, A. (2001) News Physiol. Sci. 16, 29–3358. Ross, S. B., Fuller, C.M., Bubien, J. K., andBenos, D. J. (2007)Am. J. Physiol.
Cell Physiol. 293, C1181–C1185
ENaC/Deg Subunits and Glioblastoma
SEPTEMBER 4, 2009 • VOLUME 284 • NUMBER 36 JOURNAL OF BIOLOGICAL CHEMISTRY 24541 by guest on June 1, 2013http://www.jbc.org/Downloaded from

Citations http://www.jbc.org/content/284/36/24526#otherarticles
This article has been cited by 13 HighWire-hosted articles:
by guest on June 1, 2013http://www.jbc.org/Downloaded from
Copyright © 2022 FDOKUMEN




















