
Kinematic analysis of thalamic versus subthalamic neurostimulation in postural and intention tremor
18
Kinematic analysis of thalamic versus subthalamic neurostimulation in postural and intention tremor Jan Herzog, 1 Wolfgang Hamel, 2,4 Roland Wenzelburger, 1 Monika Po« tter, 1 Marcus O. Pinsker, 2 Jan Bartussek, 3 Andre Morsnowski, 3 Frank Steigerwald, 1 Gu« nther Deuschl 1 and Jens Volkmann 1 1 Department of Neurology, Christian Albrechts University Kiel, Germany, 2 Department of Neurosurgery, Christian Albrechts University Kiel, Germany and 3 Institute of Experimental and Applied Physics, Christian-Albrechts-Universita« t Kiel, Germany 4 Present address: Department of Neurosurgery, University Hospital Hamburg, Germany Correspondence to: Prof. Dr. Jens Volkmann, MD, Christian Albrechts University Kiel, Germany E-mail: [email protected] Deep brain stimulation of the thalamus (thalamic DBS) is an established therapy for medically intractable essential tremor and tremor caused by multiple sclerosis. In both disorders, motor disability results from complex interaction between kinetic tremor and accompanying ataxia with voluntary movements. In clinical studies, the efficacy of thalamic DBS has been thoroughly assessed. However, the optimal anatomical target structure for neurostimulation is still debated and has never been analysed in conjunction with objective measurements of the different aspects of motor impairment. In 10 essential tremor and 11 multiple sclerosis patients, we analysed the effect of thalamic DBS through each contact of the quadripolar electrode on the contralateral tremor rating scale, accelerometry and kinematic measures of reach-to-grasp-movements. These measures were correlated with the anatomical position of the stimulating electrode in stereotactic space and in relation to nuclear boundaries derived from intraoperative microrecording. We found a significant impact of the stereotactic z -coordinate of stimulation contacts on theTRS, accelerometry total power and spatial deviation in the deceleration and target period of reach-to-grasp-movements. Most effective contacts clustered within the subthalamic area (STA) covering the posterior Zona incerta and prelemniscal radiation. Stimulation within this region led to a mean reduction of the lateralized tremor rating scale by 15.8 points which was significantly superior to stimulation within the thalamus (P50.05, student’s t -test). STA stimulation resulted in reduction of the accelerometry total power by 99%, whereas stimulation at the ventral thalamic border (68%) or within the thalamus proper (2.5%) was significantly less effective (P50.01). Concomitantly, STA stimulation led to a significantly higher increase of tremor frequency and decrease in EMG synchronization compared to stimulation within the thalamus proper (P50.001). In reach-to-grasp movements, STA stimulation reduced the spatial variability of the movement path in the deceleration period by 28.9% and in the target period by 58.4%, whereas stimulation within the thalamus was again significantly less effective (P50.05), with a reduction in the deceleration period between 6.5 and 21.8% and in the target period between 1.2 and 11.3%. An analysis of the nuclear boundaries from intraoperative microrecording confirmed the anatomical impression that most effective electrodes were located within the STA. Our data demonstrate a profound effect of deep brain stimulation of the thalamic region on tremor and ataxia in essential tremor and tremor caused by multiple sclerosis. The better efficacy of stimulation within the STA compared to thalamus proper favours the concept of a modulation of cerebello^thalamic projections underlying the improvement of these symptoms. Keywords: deep brain stimulation; tremor; ataxia; thalamus; subthalamic white matter; kinematic analysis Abbreviations: DBS ¼ deep brain stimulation; ET ¼ essential tremor; MS ¼ multiple sclerosis; STA ¼ subthalamic area; TRS ¼ tremor rating scale; VIM ¼ ventral intermedius nucleus Received November 8, 2006. Revised January 17 , 2007 . Accepted March 15, 2007 . Advance Access publication April 17 , 2007 doi:10.1093/brain/awm077 Brain (2007), 130, 1608 ^1625 ß The Author (2007). Published by Oxford University Press on behalf of the Guarantors of Brain. All rights reserved. For Permissions, please email: [email protected] by guest on December 11, 2013 http://brain.oxfordjournals.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Kinematic analysis of thalamic versus subthalamic neurostimulation in postural and intention tremor

Kinematic analysis of thalamic versus subthalamicneurostimulation in postural and intention tremorJan Herzog,1Wolfgang Hamel,2,4 Roland Wenzelburger,1Monika Po« tter,1Marcus O. Pinsker,2
Jan Bartussek,3 Andre Morsnowski,3 Frank Steigerwald,1Gu« nther Deuschl1 and Jens Volkmann1
1Department of Neurology, Christian Albrechts University Kiel, Germany, 2Department of Neurosurgery,Christian Albrechts University Kiel, Germany and 3Institute of Experimental and Applied Physics,Christian-Albrechts-Universita« t Kiel, Germany4Present address: Department of Neurosurgery, University Hospital Hamburg,Germany
Correspondence to: Prof. Dr. Jens Volkmann, MD, Christian Albrechts University Kiel, GermanyE-mail: [email protected]
Deep brain stimulation of the thalamus (thalamic DBS) is an established therapy for medically intractableessential tremor and tremor caused by multiple sclerosis. In both disorders, motor disability results fromcomplex interaction between kinetic tremor and accompanying ataxia with voluntary movements. In clinicalstudies, the efficacy of thalamic DBS has been thoroughly assessed. However, the optimal anatomical targetstructure for neurostimulation is still debated and has never been analysed in conjunction with objectivemeasurements of the different aspects of motor impairment. In 10 essential tremor and 11 multiple sclerosispatients, we analysed the effect of thalamic DBS through each contact of the quadripolar electrode on thecontralateral tremor rating scale, accelerometry and kinematic measures of reach-to-grasp-movements.These measures were correlated with the anatomical position of the stimulating electrode in stereotacticspace and in relation to nuclear boundaries derived from intraoperative microrecording.We found a significant impact of the stereotactic z-coordinate of stimulation contacts on theTRS, accelerometrytotal power and spatial deviation in the deceleration and target period of reach-to-grasp-movements.Most effective contacts clustered within the subthalamic area (STA) covering the posterior Zona incerta andprelemniscal radiation. Stimulation within this region led to a mean reduction of the lateralized tremor ratingscale by15.8 points whichwas significantly superior to stimulationwithin the thalamus (P50.05, student’s t-test).STA stimulation resulted in reduction of the accelerometry total power by 99%, whereas stimulation at theventral thalamic border (68%) or within the thalamus proper (2.5%) was significantly less effective (P50.01).Concomitantly, STA stimulation led to a significantly higher increase of tremor frequency and decreasein EMG synchronization compared to stimulation within the thalamus proper (P50.001). In reach-to-graspmovements, STA stimulation reduced the spatial variability of the movement path in the deceleration periodby 28.9% and in the target period by 58.4%, whereas stimulation within the thalamus was again significantlyless effective (P50.05), with a reduction in the deceleration period between 6.5 and 21.8% and in the targetperiod between 1.2 and 11.3%. An analysis of the nuclear boundaries from intraoperative microrecordingconfirmed the anatomical impression that most effective electrodes were located within the STA.Our data demonstrate a profound effect of deep brain stimulation of the thalamic region on tremor and ataxiain essential tremor and tremor caused by multiple sclerosis.The better efficacy of stimulation within the STAcompared to thalamus proper favours the concept of a modulation of cerebello^thalamic projections underlyingthe improvement of these symptoms.
Keywords: deep brain stimulation; tremor; ataxia; thalamus; subthalamic white matter; kinematic analysis
Abbreviations: DBS¼ deep brain stimulation; ET¼ essential tremor; MS¼multiple sclerosis; STA¼ subthalamic area;TRS¼ tremor rating scale; VIM¼ ventral intermedius nucleus
Received November 8, 2006. Revised January17, 2007. Accepted March 15, 2007. Advance Access publication April 17, 2007
doi:10.1093/brain/awm077 Brain (2007), 130, 1608^1625
� The Author (2007). Published by Oxford University Press on behalf of the Guarantors of Brain. All rights reserved. For Permissions, please email: [email protected]
by guest on Decem
ber 11, 2013http://brain.oxfordjournals.org/
Dow
nloaded from

IntroductionDeep brain stimulation of the ventrolateral thalamus(thalamic DBS) is used to treat a wide range of medicallyintractable tremor syndromes, nowadays mostly severeessential tremor (ET) and tremor caused by multiplesclerosis (MS). In both diseases, motor disability resultsfrom the complex interaction of kinetic (postural,action and intention) tremor and ataxia with voluntarymovements. The efficacy of thalamic DBS in ET is provenin randomized and controlled trials (Dick, 2003). For MStremor, no controlled studies are available and the currentdata is therefore insufficient to support clinical efficacy onan evidence-based level. Nevertheless, results from caseseries support the clinical impression of a significantsymptomatic benefit from thalamic DBS in at least asubpopulation of MS tremor patients (Wishart et al., 2003).Whereas clinical effects of thalamic DBS have beenthoroughly assessed, crucial aspects of its neuralmechanisms still await further characterization: first, theoptimal anatomical target structure for DBS in tremor isstill debated and has never been analysed in conjunctionwith objective measures of different aspects ofmotor disability in ET and MS patients. Second, thephysiological influence of thalamic DBS on the differentcomponents of kinetic tremor as well as abnormalitiesattributed to cerebellar dysfunction in ET and MS have notbeen completely characterized.The standard stereotactic coordinates for thalamic DBS
are located at the border between the ventral intermediusnucleus (VIM) and the subthalamic white matter (Kracket al., 2002). It is therefore debatable whether tremorreduction is caused by neural elements within the thalamusproper or the subthalamic area. Furthermore, the basicphysiological mechanisms of DBS as well as the neuralelements influenced by neurostimulation have not yetbeen conclusively defined. From electrophysiological stud-ies, there is evidence for impact of DBS both on fibre tracts(Ranck, 1975; Holsheimer et al., 2000; Kiss et al., 2003a;Anderson et al., 2004, 2006) and neuronal somata (Beurrieret al., 2001; Kiss et al., 2002; Magarinos-Ascone et al.,2002). It has been proposed that stimulation within thethalamus proper predominantly influences pathologicalneuronal activity of thalamocortical projection neurons(McIntyre et al., 2004). Alternatively, stimulation of thesubthalamic area would primarily impact afferent cerebello–thalamic fibres (Anderson et al., 2006). Recent case serieshave challenged the concept of tremor abolition byneurostimulation of the thalamus proper and located themost effective stimulation site within the subthalamic fibrearea (Kitagawa et al., 2000; Murata et al., 2003; Plaha et al.,2004).Postural and intentional components are essential
features of advanced ET (Louis, 2005) and MStremor (Alusi et al., 2001b). In ET, postural tremor is themost and disease defining symptom (Deuschl et al., 1998).
Thalamic DBS modulates the amplitude, frequency,regularity and tremor-EMG coherence in ET posturaltremor (Vaillancourt et al., 2003). In MS, postural tremoris the principal constituent of the broad clinical spectrumof ‘jerky’ movements with a prevalence of 92%and correlates highly with the tremor disability score(Alusi et al., 2001b). The influence of thalamic DBS onelectrophysiological properties of MS postural tremor hasnot yet been investigated.
Intention tremor has a lower prevalence than posturaltremor in both diseases. Approximately 15% of ET patientssuffer from intention tremor (Biary and Koller, 1987),whereas the proportion among MS tremor patients seemsto be even lower (Alusi et al., 2001b). Nevertheless,intention tremor is regarded the predominant source ofdisability (Tranchant et al., 1995; Liu et al., 1997) and animportant factor in considering functional neurosurgery.Clinically clearly defined as a tremor with increasingamplitude during movements towards a target (Deuschlet al., 1998), the electrophysiological assessment ofintention tremor is difficult (Frost, 1978; Jankovic andFrost, 1981). Intention tremor is part of atactic (literally:‘disordered’) movements in cerebellar dysfunction and assuch difficult to separate from other features like proximalpostural instability or serial dysmetria. One option isthe kinematic analysis of visually guided reach-to-graspmovements which allows to measure abnormalities of themovement trajectories during different phases of the motortask. While serial dysmetria is characterized by earlycorrectional movements during the reach, ataxia andintention tremor are most prominent in the decelerationphase of patients with cerebellar disorders (Flament andHore, 1986; Hallett et al., 1991; Hore et al., 1991; Topkaet al., 1998). MS tremor patients with lesions of thecerebellum and infratentorial structures typically shareaspects of ataxia with patients suffering from cerebellardeficit of other origin (Feys et al., 2005a). In ET,interestingly, both EMG and kinematic analysis foundsimilar abnormalities in the deceleration period leading tothe suggestion of cerebellar dysfunction in a subgroup ofET patients (Britton et al., 1994; Deuschl et al., 2000; Kosteret al., 2002). It has been proposed that serial dysmetria inearly periods of targeting movements causes delayedcompensatory moves which facilitate the occurrence ofintention tremor (Elble, 1998). It is therefore important toquantify the effect of thalamic DBS on the different phasesof reach-to-grasp movements to elucidate the underlyingpathophysiological mechanisms.
The present study aimed at defining fundamentalelectrophysiological effects of thalamic DBS in severepostural and intention tremor. We investigated theinfluence of each stimulation contact on the contralateralclinical tremor rating scale, accelerometry and kinematicfeatures of grasp-to-reach movements. We then correlatedthese changes to the anatomical site of stimulation in orderto define the optimal target for tremor suppression.
Thalamic or subthalamic DBS in tremor Brain (2007), 130, 1608^1625 1609
by guest on Decem
ber 11, 2013http://brain.oxfordjournals.org/
Dow
nloaded from
Subjects and methodsSubjectsTwenty-one patients (9 females and 12 males) who have beentreated with uni- (5) or bilateral (16) thalamic DBS at our centrefor a medically intractable tremor according to the diagnosticcriteria of the Tremor Investigation Group and the Consensusstatement of the Movement Disorder Society Group (Deuschlet al., 1998) caused by MS (11) or ET (10) participated in thisstudy after providing informed consent. Entry criteria for thisstudy were a stable clinical response to thalamic DBS for at least 4months and a score of at least 2/4 on the postural and intentiontremor items of the Fahn–Tolosa–Marin tremor rating scale (TRS)(Fahn et al., 1993). No patient was taking anti-tremor medicationat the time of assessment. Clinical details of the patientsare summarized in Table 1. In MS patients (patients no. 11–21),clinical signs in the arms other than tremor and ataxia, symptomsin other parts of the body due to MS as well as the ExpandedDisability Status Scale (EDSS) (Kurtzke, 1983) are listed inTable 2.
Surgical procedure, microrecordingMounting of the stereotactic ring (Zamoran–Dujovny openceramic version, Stryker–Leibinger, Freiburg, Germany) wasfollowed by a stereotactic MRI (Magnetom Vision 1.5 tesla,Siemens, Erlangen, Germany) with axial gadolinium-enhancedT1-sequences under general anaesthesia. Target coordinatesfor thalamic DBS (x, y, z coordinates relative to the midpointof the anterior and posterior commissure [midACPC]
in mm: 13, �5.5, 0) were defined on the T1-images according
to the atlas coordinates of the VIM. If the third ventricle exceeded
a width of 14mm, the laterality of the target was adjusted.
The trajectories were planed on the gadolinium-enhanced
T1-sequences to avoid vessels, sulci and ventricles. For intrao-
perative microrecording, we used five microelectrodes in a
‘Ben-gun’ configuration (MEAS, Stryker–Leibinger) with one
central electrode surrounded by four electrodes situated 2.3mm
anterior, posterior, medial and lateral. Electrodes were simulta-
neously advanced from 6mm above the target to 3–4mm below
the target at steps of 0.5mm. Intraoperatively, we defined the
optimal stimulation site by evaluating beneficial and adverse
effects of 130Hz microstimulation at different levels of the five
trajectories and then implanted the permanent macroelectrode
(model 3387 or 3389, Medtronic, Minneapolis, MN, USA). 43%
of all electrodes (16/37) were implanted into the central
trajectory, 38% (14/37) into the medial, 8% (3/37) into the
lateral, 5.5% (2/37) into the anterior and 5.5% (2/37) into the
posterior trajectory.
Intraoperative microrecordings were monitored online
(Leadpoint-system, Medtronic) and stored for offline analysis
(CED 1401 system, sampling rate of 25 kHz; Cambridge Electronic
Design, Camebridge, UK) with the Spike2 version 5.05 CED
software. We defined the neuronal discharge characteristics at
each electrode position according to discharge frequency, response
to intraoperative cutaneous somatosensory stimulation and
rapid active or passive joint movements, as well as the
configuration of single unit activity (SUA). Recordings were
classified as intrathalamic if SUA had a discharge frequency of at
Table 1 Patients’ characteristics and stimulation parameters used for assessment of stimulation effects
Patientno.
Gender Diagnosis Age(years)
Diseaseduration(years)
Preoperativetotal TRS(max 144)
Postoperativetotal TRS(max 144)
Side ofstimulation
Durationaftersurgery(months)
Stimulation-parametersright electrode(V, ms, Hz)
Stimulation-parametersleft electrode(V, ms, Hz)
1 M ET 69 31 71 12 Bilateral 6 1.2, 60, 130 1.5, 60, 1302 M ET 43 13 48 9 Bilateral 6 2.2, 60, 180 3.0, 60, 1803 F ET 78 14 47 10 Bilateral 31 2.3, 60, 130 1.5, 60, 1304 M ET 74 33 73 30 Bilateral 22 1.8, 60; 130 2.2, 60, 1305 F ET 69 52 60 29 Bilateral 24 2.5, 60, 130 1.5, 60, 1306 M ET 68 39 48 23 Bilateral 41 1.5, 60, 130 2.0, 60, 1307 M ET 66 20 53 14 Bilateral 6 2.8, 60, 130 2.5, 60, 1308 M ET 74 22 58 21 Bilateral 6 2.8, 60, 130 2.4, 60, 1309 F ET 74 41 60 31 Bilateral 24 3.0, 60, 130 2.5, 60, 13010 F ET 69 16 79 39 Bilateral 6 2.5, 60, 130 2.8, 60, 13011 F MS 57 4 88 35 Bilateral 10 3.5, 90, 180 2.8, 90, 18012 M MS 42 5 79 34 Bilateral 9 3.5, 60, 160 3.5, 60, 16013 M MS 30 7 30 13 Right 9 3.7, 90, 180 ^14 F MS 34 15 60 27 Left 12 ^ 2.8, 90, 15015 M MS 57 13 50 29 Left 19 ^ 3.5, 90, 15016 F MS 43 18 56 30 Left 13 ^ 2.0, 90, 13017 F MS 28 7 58 26 Left 6 ^ 3.5, 90, 15018 M MS 42 6 98 58 Bilateral 25 1.5, 60, 130 2.0, 60, 13019 F MS 54 15 77 48 Bilateral 10 2.0, 90, 130 2.0, 90, 13020 M MS 46 15 129 78 Bilateral 6 2.5, 90, 150 3.2, 90, 15021 M MS 23 6 93 28 Bilateral 6 1.5, 60, 130 2.8, 90, 130
Mean � SME ET 68.4�3.1 28.1�4.2 59.7� 3.6 21.8� 3.3MS 41.5�3.5 10.1�1.5 74.4� 8.2 36.9�5.4total 54.3�3.8 18.7� 2.9 67.4� 4.8 29.7� 3.6
1610 Brain (2007), 130, 1608^1625 J. Herzog et al.
by guest on Decem
ber 11, 2013http://brain.oxfordjournals.org/
Dow
nloaded from

least 15Hz and/or responded to somatosensory stimulation
or joint movements (Kiss et al., 2003b). Recordings from thesubthalamic area were defined in the absence of SUA and
low background activity or spare SUA below 5Hz withoutresponse to somatosensory stimuli or joint movements (Velasco
and Molina-Negro, 1973; Velasco et al., 1976; Eaton and Moss,
1989). If the electrode location could not be definitelyclassified according to these criteria, the respective trajectory
was excluded from further analysis. In 19 finally implantedelectrodes, microrecordings allowed to determine the ventral
thalamic border (thalamic border) as indicated by a transition
from intrathalamic recordings to recordings of thesubthalamic area.
Evaluation of electrode position instereotactic spaceThe stereotactic coordinates of each contact of the quadripolar
macroelectrode related to midACPC were calculated based onpostoperative X-ray radiographies in anteroposterior and lateral
projections (Stereoplan plus v.2.3 software, Stryker–Leibinger). Ifpostoperative stereotactic X-rays were unavailable, the coordinates
were determined by fusion of the preoperative stereotactic and
postoperative MRI.
If microrecordings allowed to define the ventral thalamic
border, as described earlier, we calculated the distance of eachcontact of the macroelectrode to the thalamic border by
reconstructing a plane within the commissure-based coordinatesystem connecting the points of all microelectrodes displaying
transition from thalamus to the subthalamic area. The Euclidean
distance between the centre of each contact and the reconstructedplane was measured (Fig. 1) using self-written routines in Matlab
(MathWorks, Natick, NJ, USA).
Evaluation of neurostimulation effectsAll patients underwent a clinical and kinematic assessment of the
neurostimulation effects through each contact of the quadripolar
stimulation electrode. For standardized comparison, we stimulated
each contact with the same parameters used for chronic
stimulation in this electrode (Table 1).
Clinical evaluationTremor severity was evaluated using the lateralized TRS, which
was calculated from items 5 or 6 (tremor right or left upper
extremity), 11 (drawing large Archimedes spiral), 12 (drawing
small Archimedes spiral), 13 (drawing straight lines) and 14
(pouring a glass of water) (maximum 28 with higher scores
indicating more severe tremor).
AccelerometryPatients were comfortably seated on an armchair while holding
both arms elevated at the shoulder level in a wing-beating position
with the elbows flexed at 90� and wrists extended in line with the
elbows at the level of the patient’s nose. The postural tremor was
recorded by a piezoelectric accelerometer attached to the dorsum
of each hand 9 cm distal to the processus styloideus ulnae on the
distal part of the third metacarpal bone. Simultaneously, surface
electromyographic activity of the M. flexor carpi ulnaris and
M. extensor carpi ulnaris was recorded with electrodes positioned
2 cm apart, close to the motor point of the respective muscle. The
accelerometer signals and the EMG channels were sampled over
32 s at a rate of 800Hz. Tremor activity in DBS OFF as well as in
DBS ON with subsequent stimulation of all four contacts was
analysed for accelerometer total power [log (mg2)/(mV2)] and
peak frequency. From the rectified EMG, we determined the
Table 2 Clinical symptoms other than ataxia and tremor as well as Expanded Disability Status Scale (EDSS) of MS tremorpatients
Patient no. Clinical symptoms of arms otherthan tremor and ataxia
Additional clinical symptoms EDSS
11 ^ Paraparesis (mostly wheelchair-bound),sensory dysfunction
7.0
12 ^ Paraparesis (wheelchair-bound), nystagmus, dysarthria,urinary incontinence
8.0
13 Mild paresis left arm Dysarthria 4.514 ^ Paraparesis, nystagmus, dysarthria,
slight visual disturbance6.5
15 ^ Paraparesis (wheelchair-bound), nystagnus, dysarthria,slight visual disturbance
7.5
16 ^ Paraparesis, dysarthria, sensory dysfunction,urinary frequency
7.0
17 ^ Paraparesis (mostly wheelchair-bound), nystagmus,dysarthria
7.0
18 Sensory dysfunction both arms Paraparesis (wheelchair-bound), nystagmus, oculomotoric paresis,dysarthria, urinary incontinence
8.0
19 ^ Paraparesis, nystagmus, oculomotoric paresis,dysarthria
6.5
20 Sensory dysfunction both arms Paraparesis (wheelchair-bound) nystagmus, dysarthria,urinay incontinence
8.0
21 Mild paresis right arm Paraparesis (wheelchair-bound), nystagmus, oculomotoric paresis,dysarthria, urinary dysfunction
8.0
Thalamic or subthalamic DBS in tremor Brain (2007), 130, 1608^1625 1611
by guest on Decem
ber 11, 2013http://brain.oxfordjournals.org/
Dow
nloaded from
synchronization between mechanical oscillations and muscle
activity accelerometry tremor and EMG (Raethjen et al., 2000).
Kinematic analysis of reach-to-grasp movementsPatients were seated on a comfortable chair with their back
supported and the active hand resting with the solum on a small
platform located in front. The shoulder was in a neutral position
and the elbow flexed at 90�. The target consisted of a cylinder
(1 cm diameter, 6 cm length) which was fixed to a heavy support
34 cm above the table at a distance of �50 cm from the body. An
acoustic signal instructed the subjects to reach out and grasp the
cylinder with their thumb and index. After each movement the
hand was returned to the starting position. All patients performed
training trials until they were familiar with the task. Twenty trials
were sampled in each condition.
The movements were recorded using a magnetic six-degree-
of-freedom measurement system, (3SPACE FASTRAK, Polhemus
Inc, Colchester, VT, USA). In that system, a low-frequency
magnetic field is emanated from a stationary transmitting antenna
and sensed with movable receiving antenna at a sampling rate of
40Hz using LabView-software (National instruments Cooperation,
Austin, TX, USA). Markers at a size of 1� 1� 1 cm3 containing
the receiving antennas were placed on the tip of the thumb and
index finger as well as the epicondylus of the wrist. This allowed
recording of movement paths of thumb, index and wrist within a
3D-Cartesian coordinate system. Data were analysed by Matlab-
software.
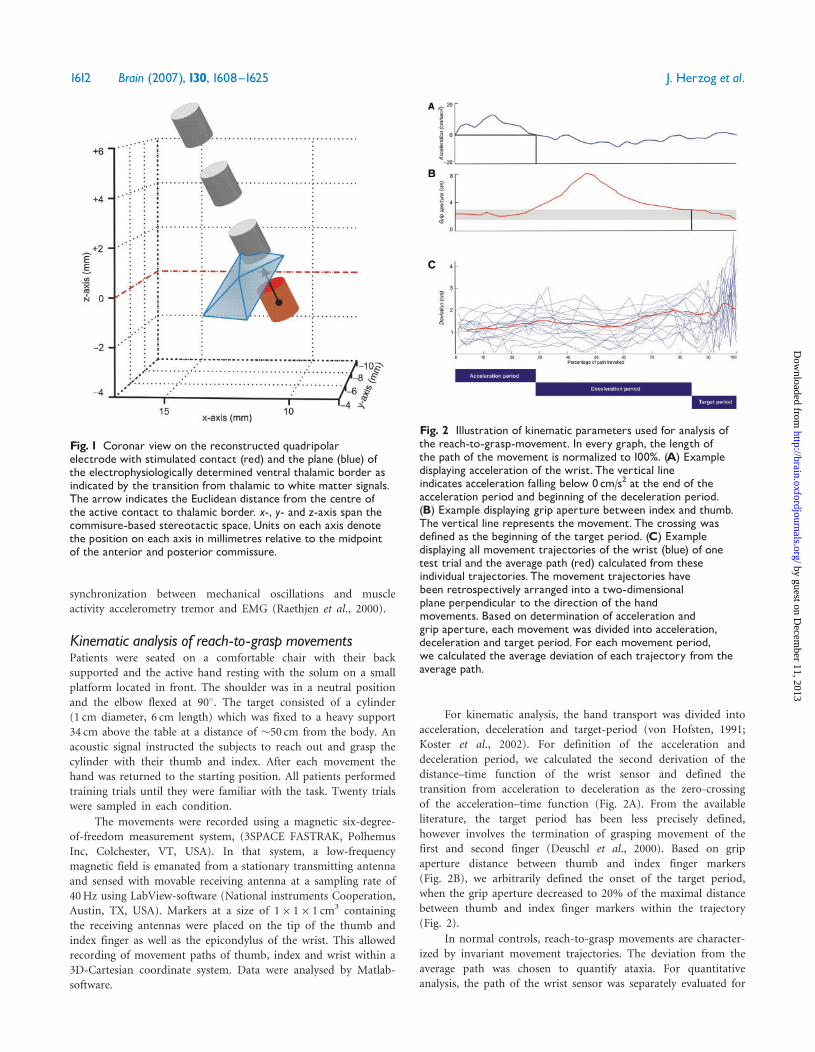
For kinematic analysis, the hand transport was divided into
acceleration, deceleration and target-period (von Hofsten, 1991;
Koster et al., 2002). For definition of the acceleration and
deceleration period, we calculated the second derivation of the
distance–time function of the wrist sensor and defined the
transition from acceleration to deceleration as the zero-crossing
of the acceleration–time function (Fig. 2A). From the available
literature, the target period has been less precisely defined,
however involves the termination of grasping movement of the
first and second finger (Deuschl et al., 2000). Based on grip
aperture distance between thumb and index finger markers
(Fig. 2B), we arbitrarily defined the onset of the target period,
when the grip aperture decreased to 20% of the maximal distance
between thumb and index finger markers within the trajectory
(Fig. 2).
In normal controls, reach-to-grasp movements are character-
ized by invariant movement trajectories. The deviation from the
average path was chosen to quantify ataxia. For quantitative
analysis, the path of the wrist sensor was separately evaluated for
Fig. 2 Illustration of kinematic parameters used for analysis ofthe reach-to-grasp-movement. In every graph, the length ofthe path of the movement is normalized to 100%. (A) Exampledisplaying acceleration of the wrist. The vertical lineindicates acceleration falling below 0cm/s2 at the end of theacceleration period and beginning of the deceleration period.(B) Example displaying grip aperture between index and thumb.The vertical line represents the movement. The crossing wasdefined as the beginning of the target period. (C) Exampledisplaying all movement trajectories of the wrist (blue) of onetest trial and the average path (red) calculated from theseindividual trajectories. The movement trajectories havebeen retrospectively arranged into a two-dimensionalplane perpendicular to the direction of the handmovements. Based on determination of acceleration andgrip aperture, each movement was divided into acceleration,deceleration and target period. For each movement period,we calculated the average deviation of each trajectory from theaverage path.
Fig. 1 Coronar view on the reconstructed quadripolarelectrode with stimulated contact (red) and the plane (blue) ofthe electrophysiologically determined ventral thalamic border asindicated by the transition from thalamic to white matter signals.The arrow indicates the Euclidean distance from the centre ofthe active contact to thalamic border. x-, y- and z-axis span thecommisure-based stereotactic space.Units on each axis denotethe position on each axis in millimetres relative to the midpointof the anterior and posterior commissure.
1612 Brain (2007), 130, 1608^1625 J. Herzog et al.
by guest on Decem
ber 11, 2013http://brain.oxfordjournals.org/
Dow
nloaded from

the acceleration, deceleration and target period to assign spatialabnormalities to the respective period of the movement. For thatpurpose, we defined the average path of all available grasping trialsin one condition and determined the root mean square (RMS incm) of each movement trajectory to the average path in a two-dimensional plane perpendicular to the direction of the handmovement (Fig. 2C). Subsequently, we calculated the mean of theRMS of all trajectories from the average path for each of threemovement periods. Kinematic analysis was performed in DBS OFFand for each of four contacts with the identical stimulationparameters used in assessment of lateralized TRS and accelero-metry. The absolute values for mean deviation from the averagepath showed a high degree of interindividual variability due todifferences in symptom severity. Therefore, the relative changefrom baseline (DBS OFF) was used to assess the impact ofstimulation through the four contacts on the different movementperiods.
Determination of charge of stimulationcontactWe calculated the charge (mC) of each stimulation contactaccording to the following formula:
Amplitude ½V� � pulse width ½�s�=impedance ½��
Statistical analysisTo describe the relative position of each stimulation contact to thestandard target (x¼ 13, y¼�5.5, z¼ 0), we calculated thedifference (in mm) along each axis of the Cartesian stereotacticcoordinate system (called ‘coordinate difference’). Based on thiscalculated difference, each contact was assigned to one of the threecategories according to its position on the x, y and z axis,respectively. Along the x-axis, a difference 5�0.5mm wasconsidered ‘medially’ to the standard target, between �0.5 andþ0.5mm ‘at standard coordinates’ and 4þ0.5mm ‘laterally’ tothe standard target. Along the y-axis, a difference5�0.5mm wasconsidered ‘posterior’ to the standard target, between �0.5 andþ0.5mm ‘at standard coordinates’ and 4þ0.5mm ‘anterior’ tothe standard target. Along the z-axis, a difference 5�0.5mmindicated a position ‘below (ventral)’ to standard target, between�0.5 and þ0.5mm ‘at standard coordinates’ and 4þ0.5mm‘above (dorsal)’ to standard target.
We used repeated measures of covariance (ANCOVA) withthe following dependent variables: change in lateralized TRS,accelerometry total power, tremor frequency and change of spatialdeviation in the deceleration and target period and the followingcovariates: Disease (ET versus MS), stimulation charge andcoordinate difference (x, y, z). The ANCOVA results directedmeasures of variance (ANOVA) and student’s t-test for individualcomparisons.
To describe the position of the stimulation contact inrelation to the electrophysiologically determined ventral thalamicborder, we calculated the distance of each stimulation contact tothat border. In contacts located below the electrophysiologicallydetermined thalamic border, the mean distance to the ventralborder was 1.5� 0.2mm (mean� standard error of the mean). Incontacts located above, the mean distance was 3.3� 0.3mm. Weused ANCOVA analysis to describe the effect of disease,stimulation charge and distance to ventral thalamic border onchange in lateralized TRS, accelerometry total power, change in
tremor frequency and change of spatial deviation within thedeceleration and target period. To compare the effect ofstimulation contacts below and above the thalamic border weused student’s t-test.
All tests were two-sided with a P50.05.
ResultsClinical effect of neurostimulationThalamic DBS using the postoperatively determinedchronic stimulation parameters was highly effective inall patients (Table 1). The average reduction of thepreoperative TRS in all patients was 55.9� 4.4%(mean� standard error of the mean), in ET patients64.2� 4.4%, and in MS patients 50.4� 3.1%.
Neurostimulation effect on lateralized TRS
Correlation of lateralized TRS with stereotactic coordinatesIn DBS OFF, the mean lateralized TRS was 22.3� 1.0 for allpatients. ANCOVA analysis revealed that the magnitude ofchange in lateralized TRS in DBS ON depended on thez-coordinate difference of the stimulation contact[F(1,148)¼ 32.1, P50.001]. No other covariate orinteraction term was associated with a significant difference(Table 3). One-way ANOVA showed a significant differencein change of lateralized TRS between contacts locatedbelow, at level of standard target and above[F(2,145)¼ 68.0, P50.001]. The largest change inlateralized TRS (15.8� 1.0) was found for contacts belowthe standard target, whereas stimulation contacts at level ofthe standard target or above were significantly less effective(12.1� 1.3 and 4.7� 0.5) (Fig. 3A).
Correlation of lateralized TRS with distance ofstimulation site to the ventral thalamic borderIn 19 electrodes (76 contacts in 13 patients), electrophy-siological recordings allowed to determine the ventralthalamic border. In these cases, the mean lateralized TRSin DBS OFF was 21.5� 1.6. In the ANCOVA analysis,only distance to the electrophysiologically determinedventral thalamic border had a significant relation tochange in lateralized TRS [F(1,76)¼ 63.2, P50.001)(Table 4). Stimulation of contacts below the thalamicborder led to a significant larger change of lateralized TRS(15.4� 1.3) than contacts above (5.2� 0.6, P50.001, t-test)(Fig. 3B).
Neurostimulation effect on accelerometry inpostural tremor
Correlation of accelerometry with stereotactic coordinatesIn 33 electrodes (132 contacts in 21 patients), we evaluatedthe effect of thalamic DBS on accelerometry total
Thalamic or subthalamic DBS in tremor Brain (2007), 130, 1608^1625 1613
by guest on Decem
ber 11, 2013http://brain.oxfordjournals.org/
Dow
nloaded from
power, change in tremor frequency and EMG synchroni-zation. For the remaining electrodes, accelerometry wasnot tolerated by the patient due to severe tremor inDBS OFF.The total power in DBS OFF was 7.9� 102� 2.2� 102
[log (mg2)/(mV2)]. ANCOVA analysis showed that thetotal power depended on the z-coordinate difference ofstimulation contacts [F(1,132)¼ 5.1, P50.025], whereasother covariates or interactions did not reveal a significanteffect (Table 3). One-way ANOVA showed a significantdifference in total power for stimulation contacts below,at standard level and above [F(2,145)¼ 6.8, P50.002].Lowest accelerometry total power was found for contactslocated below standard z-coordinate (7.7� 100� 5.2� 100),whereas stimulation at level of standard coordinates(7.8� 101� 5.3� 101) and above (2.5� 102� 4.8� 101)was significantly less effective (Fig. 3C).In DBS OFF, tremor frequency differed between ET
(3.9� 0.1Hz) and MS patients (3.2� 0.2Hz) (P50.001,t-test). ANCOVA analysis revealed significant impact onchange of tremor frequency by DBS for disease[F(1,132)¼ 4.8, P50.031], x-coordinate difference[F(1,132)¼ 24.6, P50.001], z-coordinate difference[F(1,132)¼ 8.6, P50.003] as well as the interaction terms[disease x x-coordinate difference] [F(1,132)¼ 4.0,P50.047] and [x-coordinate difference x z-coordinatedifference] [F(1,132)¼ 4.3, P50.041] (Table 3). In ET,stimulation contacts below the standard z-coordinate led toan increase of tremor frequency by 0.78� 0.12Hz, at levelof standard z-coordinate by 0.60� 0.19Hz and above by0.13� 0.05Hz. In MS, effect on tremor frequency was lesspronounced with an increase by 0.39� 0.13Hz below
standard z-coordinate, 0.03� 0.03Hz at standard coordi-nate and 0.01� 0.04Hz above (Table 5). With respect tothe x-coordinate, in ET patients, stimulation contactslocated medially to the standard x-coordinate led to anincrease by 0.95� 0.09Hz, at standard x-coordinate by0.43� 0.15Hz and laterally by 0.12� 0.06Hz. In MS, therewas no significant difference in change of the tremorfrequency in relation to the x-coordinate difference(Table 5).
In DBS OFF, we found synchronization between EMGand accelerometry in 92% of contacts. Synchronization wassignificantly less frequent (P50.001, chi square test) withcontacts below the standard z-coordinate (33%) comparedto the standard target and above (70.5 and 96%).
Correlation of accelerometry with distance to theventral thalamic borderIn 17 electrodes (68 contacts from 12 patients), electro-physiological recordings allowed to determine the ventralthalamic border. In these cases, the mean total power was5.5� 102� 3.2� 102 in DBS OFF. The change in totalpower was significantly related to the distance of thecontact from the ventral thalamic border [ANCOVA,F(1,68)¼ 5.7, P50.020] (Table 4). Stimulation belowthe thalamic border reduced total power to1.6� 100� 5.6� 10�1 but stimulation above only to2.2� 102� 6.1� 101 (P50.01, t-test) (Fig. 3D).
In DBS OFF, tremor frequency differed significantlybetween ET (3.94� 0.11Hz) and MS patients(3.1� 0.07Hz) (P50.001, t-test). ANCOVA analysisshowed a significant influence on the change of tremor
Table 3 Effects of disease, stimulation charge and position of stimulation contacts within the stereotactic space on ETandMS tremor
Change inlateralized TRS
Accelerometrytotal power
Change in tremorfrequency
Percentage change ofspatial deviation(deceleration period)
Percentage change ofspatial deviation(target period)
F (1,148) P F (1,132) P F (1,132) P F (1,96) P F (1,96) P
1: Disease 0.002 0.965 0.058 0.811 4.765 0.031 0.631 0.429 0.071 0.7902: Stimulation charge 3.625 0.059 1.753 0.188 0.263 0.609 0.792 0.376 0.133 0.7163: x-coordinate difference 2.439 0.121 0.234 0.629 24.613 0.001 0.401 0.528 3.292 0.0734: y-coordinate difference 0.831 0.364 0.108 0.743 0.364 0.547 0.899 0.346 0.681 0.4125: z-coordinate difference 32.071 0.001 5.136 0.025 8.647 0.003 6.233 0.015 13.352 0.001Interaction term [1�2] 0.560 0.455 0.402 0.527 0.004 0.951 2.457 0.121 0.243 0.623Interaction term [1�3] 3.217 0.070 3.340 0.070 4.031 0.047 0.025 0.876 0.005 0.943Interaction term [1�4] 0.000 0.983 0.460 0.499 1.137 0.288 0.578 0.449 0.016 0.899Interaction term [1�5] 0.800 0.373 1.247 0.266 2.129 0.147 0.129 0.721 0.199 0.657Interaction term [3� 4] 0.150 0.699 2.578 0.111 0.544 0.462 0.372 0.544 0.525 0.471Interaction term [3�5] 0.251 0.617 0.319 0.573 4.288 0.041 0.029 0.865 0.119 0.731Interaction term [4� 5] 0.052 0.820 0.548 0.461 1.999 0.160 0.356 0.552 1.932 0.168Interaction term [3� 4� 5] 3.297 0.72 0.464 0.497 0.170 0.681 0.044 0.834 2.262 0.136
Note: Measures of covariance (ANCOVA) with following dependent variables: change in lateralized TRS, accelerometry total power, changein tremor frequency and percentage deviation in the deceleration and target period of reach-to-grasp movements. Disease, stimulationcharge, x-/y-/z-coordinates of stimulation contacts and respective interaction terms represent covariates. Significant impact of covariatesand interaction terms are bold type printed.
1614 Brain (2007), 130, 1608^1625 J. Herzog et al.
by guest on Decem
ber 11, 2013http://brain.oxfordjournals.org/
Dow
nloaded from

frequency for disease [F(1,68)¼ 16.1, P50.001] anddistance of stimulation contacts to the thalamic border[F(1,68)¼ 9.6, P50.003] (Table 4). In ET, stimulationof contacts below thalamic border led to a significantlygreater increase of tremor (0.71� 0.12Hz) comparedto stimulation above (0.21� 0.07Hz) (P50.001, t-test).
In MS, there was no significant increase of tremorfrequency (Table 6).
Stimulation below the thalamic border reduced theoccurrence of EMG synchronization from 90 (DBS OFF)to 20%, whereas stimulation above did not (93%)(P50.001, chi square test).
Fig. 3 Mean change of lateralized TRS and accelerometry total power. (A) Mean change of lateralized TRS in relation to the difference ofz-coordinate of the stimulated contact to the standard target z-value. According to the calculated difference, contacts have been dividedinto three groups: below (5̂ 0.5mm) to the standard z-value, at level of standard z-value (within an interval of �0.5mm) and above(40.5mm) to standard z-value. (B) Mean change of lateralized TRS in relation to the electrophysiologically determined ventral thalamicborder. Contacts have been divided into two groups: below ventral thalamic border and above. (C) Mean accelerometry total power inrelation to the difference of z-coordinates of the stimulated contact to the standard target z-value: as described earlier, according tothe calculated difference, contacts have been divided into three groups: below, at level of standard z-value and above to standard z-value.(D) Mean accelerometry total power in relation to the electrophysiologically determined ventral thalamic border. Contacts have beendivided into two groups: below ventral thalamic border and above. Student’s t-test has been performed comparing effect of contactslocated below to contacts located at level of standard coordinate and above. ���P50.001, ��P50.01, �P50.05.
Thalamic or subthalamic DBS in tremor Brain (2007), 130, 1608^1625 1615
by guest on Decem
ber 11, 2013http://brain.oxfordjournals.org/
Dow
nloaded from

Neurostimulation effect on reach-to-graspmovements
Correlation of change in kinematic parameters withstereotactic coordinatesKinematic analysis was available for 24 electrodes (96contacts in 17 patients). In the other cases tremor in DBSOFF was too severe to complete the task.Thalamic DBS had no consistent influence on the
acceleration period of reach-to-grasp-movements.However, we found significant improvements in thevariability of the movement trajectories duringthe deceleration and target period in ET (Fig. 4A–C) andMS patients (Fig. 4D–F). Z-coordinate difference from thestandard target was the only significant predictor ofimprovement in the deceleration period [ANCOVA,F(1,96)¼ 6.2, P50.015]. The highest percentage improve-ment was found for contacts located below standardz-coordinate (29.9� 4.5%), whereas contacts located atstandard z-coordinates (6.5� 7.5%) and above (1.3� 4.4%)were significantly less effective (P50.05, t-test) (Fig. 5A).Likewise, improvement in the target period was sig-
nificantly related only to the z-coordinate difference[ANCOVA, F(1,96)¼ 13.4, P50.001] (Table 3). The
relative improvement in the target period was greatest forcontacts below standard z-coordinate (58.4� 3.3%),whereas contacts located at level of standard z-coordinates(21.7� 10.1%) and above (11.3� 3.2%) were significantlyless effective [ANOVA, F(2,95)¼ 8.4, P50.01] (Fig. 5B).
Table 4 Effect of disease, stimulation charge and position of stimulation contact in relation to the ventral thalamic border onETand MS tremor
Change inlateralized TRS
Accelerometrytotal power
Change in tremorfrequency
Percentage change ofspatial deviation(deceleration period)
Percentage change ofspatial deviation(target period)
F (1,76) P F (1,68) P F (1,68) P F (1,52) P F (1,52) P
1: Disease 3.747 0.57 0.126 0.724 16.11 0.001 0.756 0.389 0.404 0.5282: Stimulation charge 0.090 0.765 1.297 0.259 2.580 0.113 0.163 0.688 2.028 0.1613: Distance to VIM border 63.182 0.001 5.732 0.020 9.682 0.003 10.383 0.002 25.596 0.001Interaction term [1�2] 3,821 0.054 0.022 0.884 2.455 0.109 0.989 0.325 0.001 0.978Interaction term [1�3] 0.031 0.861 0.039 0.844 2.465 0.122 0.021 0.884 0.978 0.328
Note: Measures of covariance (ANCOVA) with following dependent variables: change in lateralized TRS, accelerometry total power,change in tremor frequency and percentage deviation in the deceleration and target period of reach-to-grasp movements.Disease, stimulation charge, position of stimulation contacts relative to the ventral thalamic border and respective interactionterms represent covariates. Significant impact of covariates and interaction terms are bold type printed.
Table 5 Effect of position of stimulation contact within the stereotactic space on increase of tremor frequency
z-coordinate-relative to standard value x-coordinate relative to standard value
Below Standard Above Medial Standard Lateral
ET 0.78� 0.12 0.60� 0.19 0.13� 0.05 0.95� 0.09 0.43� 0.15 0.12� 0.06(P¼ 0.466) (P50.001) (P¼ 0.006) (P40.001)
MS 0.39� 0.13 0.03� 0.03 0.01� 0.04 0.19� 0.07 0.16� 0.04 0.12� 0.09(P¼ 0.015) (P¼ 0.001) (P¼ 0.455) (P¼ 0.130)
Note: Mean change of tremor frequency (Hz) (�standard error of the mean) in DBS ON compared to DBS OFF. Since ANCOVA revealed asignificant influence of disease, analysis has been performed separately for ETand MS. Stimulation contacts have been divided into threegroups according to the difference of coordinate of the stimulated contact to the standard y-value and x-value, respectively. Student’s t-testwas used for comparison of values at standard coordinates and above/laterally with values located below/medially to standard values.Significant differences are bold type printed.
Table 6 Effect of position of stimulation contact in relationto the ventral thalamic border on tremor frequency
Position relative to electrophysiologicallydetermined ventral thalamic border
Below Above
ET 0.71�0.12 0.21�0.07(P50.001)
MS 0.14� 0.09 0.04� 0.06(P¼ 0.118)
Note: Mean change of tremor frequency (Hz) (�standard errorof the mean) compared to DBS OFF. Since ANCOVA revealeda significant influence of disease, analysis has been performedseparately for ETand MS. Stimulation contacts have beendivided according to their position below and above theelectrophysiologically determined thalamic border. Student’s t-testwas used for comparison of values above with values below ventralthalamic border. Significant changes are bold type printed.
1616 Brain (2007), 130, 1608^1625 J. Herzog et al.
by guest on Decem
ber 11, 2013http://brain.oxfordjournals.org/
Dow
nloaded from

The stereotactic coordinates of the most effective contactsin reducing the spatial variability in the target periodprojected onto the prelemniscal radiation and posterior partof the Zona incerta in the Schaltenbrand–Wahren atlas(Fig. 6).
Correlation of change in kinematic parameters withdistance to the ventral thalamic borderIn 14 electrodes (56 contacts in 9 patients), electrophysio-logical recordings allowed to determine the thalamic borderand to relate the electrode location to the performance inreach-to-grasp movements. In both, the deceleration andthe target period, the vertical distance from the thalamic
border had a significant bearing on the relative improve-ment of spatial variability [ANCOVA, F(1,52)¼ 10.3,P50.002] and [F(1,52)¼ 25.6, P50.001] (Table 4).Stimulation of contacts below the thalamic border wassignificantly more effective than above as displayed inFig. 5C and D.
DiscussionWe observed pronounced effects of thalamic DBS onfundamental electrophysiological features of MS andessential tremor which strongly depended on the anatomi-cal site of stimulation. Thalamic DBS significantly reducedpathological features of postural tremor and also improved
Fig. 4 Examples of reach-to-grasp movements in ET patients (A^C) and MS patients (D^F) in DBS OFF condition and with stimulation ofeach contact of the quadripolar electrode. In each graph, outermost left trajectory display DBSOFF. Subsequently, the effect of stimulationcontacts 0 to 3 are demonstrated from left to right.Grey boxes under the trajectories show the target phase of each reach-to-graspmovement drawn at larger scale.
Thalamic or subthalamic DBS in tremor Brain (2007), 130, 1608^1625 1617
by guest on Decem
ber 11, 2013http://brain.oxfordjournals.org/
Dow
nloaded from

unsteadiness in the terminal phase of goal-directed move-ments, which is a typical feature of atactic movements incerebellar dysfunction. These effects were markedly strongerfor electrode contacts below the AC–PC line as comparedto stimulation at the standard VIM target within theventrolateral thalamus. The ventrodorsal location ofstimulation (stereotactic z-axis) was the only significantcovariate of the change in tremor and movementparameters, with the exception of the change in tremor
frequency, which was also related to the location of theelectrode along the x-axis.
Methodical considerationsThe level of the AC–PC line defines the ventral thalamicborder in most stereotactic reference systems (Guiot et al.,1976; Schaltenbrand and Wahren, 1977). We thereforeargue that the most effective contacts were probably located
Fig. 5 Mean percentage reduction of the spatial deviation from the average path in the deceleration and target period of reach-to-graspmovements. (A) Mean percentage reduction in the deceleration period. Stimulation contacts have been divided into three groups accordingto the difference of z-coordinate of the stimulated contact to the standard target z-value: below (5̂ 0.5mm) to the standard z-value,at level of standard z-value (within an interval of �0.5mm) and above (40.5mm) to standard z-value. (B) Mean percentage reduction inthe target period. As described earlier, according to the calculated difference, contacts have been divided into three groups: below, atlevel of standard z-value and above to standard z-value. (C) Mean percentage reduction in the deceleration period in relation to theelectrophysiologically determined ventral thalamic border.Contacts have been divided into two groups: below thalamic border and above.(D) Mean percentage reduction in the target period in relation to the ventral thalamic border.Contacts have been divided into two groups:below ventral thalamic border and above. Student’s t-test has been performed comparing effect of contacts located below to the standardcoordinates to contacts located at level of standard coordinate and above. ���P50.001, ��P50.01, �P50.05.
1618 Brain (2007), 130, 1608^1625 J. Herzog et al.
by guest on Decem
ber 11, 2013http://brain.oxfordjournals.org/
Dow
nloaded from

within the subthalamic white matter and not withinthalamic nuclei. Before interpreting this data, somepotentially confounding factors have to be considered:our anatomical interpretation is based on superimpositionof landmark-based stereotactic coordinates on a standardbrain atlas. Especially in MS patients, due to atrophyof grey and white matter, the thalamic region mayshow a significant atrophy in MRI-based quantification ofparenchyma volume (Wylezinska et al., 2003; Carone et al.,2006) and therefore not match the standard anatomicalatlas. We did not correlate the postoperative location of theelectrode on MRI to the ventral border of the thalamus,which may be difficult to identify. Based on microelectroderecordings, which enables discrimination between grey andwhite matter signals, we confirmed our observation of abetter stimulation efficacy below the ventral thalamicborder in a subset of patients. However, accuracy of theposition of the macroelectrode position relative to theelectrophysiologically determined thalamic boundary maybe impaired by different factors. First, there may be a
mismatch between the position of the trajectory used formicrorecording and the finally implanted macroelectrode.According to the available literature, the difference betweenthe calculated target and the implanted electrode for thez-axis is in the range of 0.8mm (Dormont et al., 1997;Ferroli et al., 2004). In our analysis of contacts consideredto be located within the subthalamic area, the meandistance to the electrophysiologically determined ventralthalamic border was 1.5mm. Therefore, we cannot excludea potential imprecision for few contacts located in closevicinity to ventral thalamic border as determined bymicrorecording. In the majority of contacts, however, thevertical spacing between contact and electrophysiologicallydetermined thalamic border was clearly larger. Second,intraoperative brain shift due to leakage of cerebrospinalfluid and re-shifting during the postoperative periodhave to be considered. Although there is no systematicquantitative study available on the magnitude of brainshifting following DBS, it may introduce uncertainties whenpostoperative imaging is used for electrode location.
Fig. 6 Projection of stimulated contacts on horizontal sections of the Schaltenbrand^Wahren atlas (negative values indicate sectionsbelow AC^PC line, positive values indicate sections above AC^PC line). The relative changes of spatial deviations in the target period ofreach-to-grasp movements are colour-coded with positive values (green) indicating improvement and negative values (red) indicatingdeterioration.
Thalamic or subthalamic DBS in tremor Brain (2007), 130, 1608^1625 1619
by guest on Decem
ber 11, 2013http://brain.oxfordjournals.org/
Dow
nloaded from

The effect of brain shift in the target area, however, is likelyto be small given the very accurate match betweenstereotactic planning and actual electrode location onpostoperative stereotactic imaging in most cases. Third,simultaneously advancing of five microelectrodes as in oursurgical approach may push forward the adhering braintissue. This may lead to the impression that the recordedactivity is deeper than it actually is when the brain tissuehas resettled around the chronic macroelectrode. Thislocation error is probably small, because the microelec-trodes in our setup are sharp tipped and have a diameter ofonly 125�m, which enables them to penetrate the tissuewith little resistance.
Influence of thalamic neurostimulationon postural tremorWe analysed accelerometry total power, EMG synchroniza-tion and tremor frequency during a postural task and foundthat stimulating the subthalamic area reduced the totalpower of tremor by nearly three dimension scalesapproaching the range of physiological tremor (Deuschlet al., 2001). In contrast, stimulating the thalamus properreduced postural tremor only within the range ofpathological values. The synchronization between EMGand mechanical tremor oscillations—reflecting the centraldriving oscillator—was disrupted by stimulation throughmost of the contacts within the subthalamic area, but wasstill present with stimulation within ventrolateral thalamus.Moreover, stimulation of contacts in the subthalamic areasignificantly increased tremor frequency, which also reflectsa reduced contribution of a pathological central tremoroscillator (Deuschl and Bergman, 2002). This effect wasstronger in ET than in MS tremor, but still present in MSpatients. The similar effects of neurostimulation in the twopatient groups, underline the symptomatic efficacy ofthalamic DBS in postural tremor in general, irrespectiveof the underlying disease.A previous study has characterized the neurophysiologi-
cal consequences of thalamic DBS on postural tremor in ET(Vaillancourt et al., 2003). The main finding was a markedreduction of tremor amplitude comparable to values ofphysiological tremor in control subjects. Additionally,effective thalamic DBS resulted in reduced tremor-EMGcoherence and an increase of tremor frequency by �1Hz.In general, these observations are in good agreement withour results. Some differences in analysing tremor data,patient characteristics and study design, however, have tobe considered in comparing both studies. In contrast to ourapproach, the former study investigated the root meansquare displacement from the dominant tremor frequencyand the degree of entrainment for determination of tremorstrength and EMG-tremor coherence, respectively. Basedon our qualitative method of analysing EMG-tremorcoherence, we cannot completely exclude a shift of EMGactivity to higher frequencies with a lower and broader
coherence peak. Despite these differences, both studiessupport the view that effective thalamic DBS suppressescentral tremor oscillations and allow mechanical-reflexfactors, represented by a desynchronization and anincrease of tremor frequency, to play a larger role. Basiccharacteristics of ET patients were different in both studies,most notably for tremor frequency in DBS OFF. Althoughdisease duration in the former study was not stated,the markedly lower frequency of 3.8Hz in our studysuggests that our patients probably had been operatedin a more advanced stage of disease. As a major difference,the previous study did not correlate the clinical andelectrophysiological effects of thalamic DBS to theanatomical site of stimulation. It was therefore not possibleto conclude which anatomical part of the central tremoroscillator was affected by thalamic DBS.
ET is thought to arise from abnormal oscillatory activitywithin the olivo–cerebello–thalamic circuit. In the animalmodel of harmaline-induced tremor (Lamarre and Mercier,1971; Elble, 1998), resembling ET in some aspects,the cerebellar cortex and the deep cerebellar nucleisynchronously show similar oscillations (Wilms et al.,1999). Cerebellar information is relayed from the deepcerebellar nuclei to thalamic nuclei and consecutively tomotor cortical areas (Steriade et al., 1997). Along thispathophysiological model, thalamic DBS may act byinterrupting the transfer of pathologic oscillations in theolivo–cerebellar circuit to the motor cortex (Benabid et al.,1996; Kiss et al., 2002; Anderson et al., 2003). Based onelectrophysiological findings with approximately 40 timeslower thresholds for fibres of passage and axonal terminalsthan for cell somata (Holsheimer et al., 2000; Kiss et al.,2003a; Anderson et al., 2004, 2006), it has been suggestedthat the targeted neuronal elements in thalamic DBSare predominantly axonal. However, despite a higherexcitability for fibre tracts (Ranck, 1975), it cannot beexcluded that DBS also exerts some somatodendritic effectswithin the volume of current spread, which may contributeto the clinical benefit. Based on PET studies demonstratinga stimulation-associated increase of regional blood flow inthe primary motor cortex (Ceballos-Baumann et al., 2001)and supplementary motor area (Perlmutter et al., 2002),it has been hypothesized that effective thalamic DBSexcites thalamocortical projections. By high-frequencyexcitation, thalamic DBS may mask the abnormalinput of synchronous oscillations to the motor cortex andtherefore cancel the rhythmic entrainment of spinalmotoneurons and muscles (Elble, 1996; Hua et al., 1998).However, magnetoencephalographic experiments havechallenged the idea of a primary role of the thalamocorticalcircuit in development of ET (Halliday et al., 2000).Our finding with most effective stimulation in thesubthalamic area is neither consistent with the concept ofmodulating the thalamocortical efferent outflow for tremorsuppression. Instead it favours the idea that presynapticstimulation of cerebellar input to the thalamus may be
1620 Brain (2007), 130, 1608^1625 J. Herzog et al.
by guest on Decem
ber 11, 2013http://brain.oxfordjournals.org/
Dow
nloaded from

more appropriate to disrupt oscillatory activity. Recentelectrophysiological studies in brain slices and intraopera-tive microrecordings in ET patients (Anderson et al., 2006)support that elimination of oscillatory input to thalamictremor cells might be most accountable for stoppingtremor propagation. Nevertheless, more effective stimula-tion by affecting cerebello–thalamic projections doesnot necessarily conflict the findings of the aforementionedPET studies. Since cerebello–thalamic projections areexcitatory by nature, high-frequency stimulation withinthe subthalamic area could drive downstream thalamicneurons resulting in activation of cortical areas inPET imaging. A problem in interpreting the present PETdata is the lack of information on the stereotacticlocalization of the effective electrode contacts. Furtherimaging studies focusing on the differential effect of thestimulation site may therefore improve our understandingof whether stimulation within the subthalamic area leads toa different activation pattern compared to stimulationwithin thalamus proper.To our knowledge, no study has assessed the impact
of thalamic DBS on electrophysiological parameters ofpostural tremor in MS. Similar to ET, stimulation ofsubthalamic white matter was most effective in reducingtremor power and EMG-tremor coherence. The lesspronounced change in tremor frequency may reflectadditional cerebellar symptoms of MS such as proximalpostural instability which may contribute to low-frequencymechanical oscillations and prevent the dominance ofhigher-frequency mechanical-reflex oscillations.Despite the widespread lesions in MS, action tremor
seems to be specifically related to damage of the cerebellumor its connections (Liu et al., 1999). There is supportingevidence from animal experiments showing that sufficientmyelination of the olivocerebellar connection is crucial forgenerating synchronous and exactly timed complex spikeactivity in the cerebellum (Lang and Rosenbluth, 2003). Inmyelin-deficient rats, disturbance of myelination in theolivocerebellar system results in abnormal cerebellar activityand is associated with the appearance of tremor. In MSpatients, CNS myelin deficiency of the olivocerebellarsystem may similarly contribute to tremor activity whichis conveyed to the thalamus and the thalamocortical system.Similar to ET, presynaptic blocking of the abnormalafferent input to thalamic neurons appears to be mosteffective in reducing postural tremor.
Influence of thalamic neurostimulationon reach-to-grasp movementsThalamic DBS significantly reduced the spatial variabilityin the target and deceleration period of visually guidedreach-to-grasp movements. The improvement of these twokinematic parameters was only observed with stimulation ofthe subthalamic area.
So far no study had assessed the impact of thalamic DBSon kinematic parameters of visually guided reach-to-graspmovements in ET. However, increasing experimentalevidence suggests the evolution of cerebellar dysfunctionin advanced stages of the disease, which is typically reflectedby imprecise targeting during goal-directed movements.In a tracking paradigm moving a horizontal manipulandumeither with the wrist (Britton et al., 1994) or elbow (Kosteret al., 2002), the second agonist burst was significantlydelayed in ET resulting in an abnormal deceleration profile,movement overshot and intention tremor (Berardelli et al.,1996). One study (Zackowski et al., 2002) studied the effectof thalamic DBS on the agonist-antagonist-pattern ina sudden stretch load, but did not find a change inthe timing relationship between the initial agonist andantagonist burst. Since, however, the load was applied inthe hold position of the wrist, the experimental settingsdoes not meet the criteria of a goal-directed movement butrather of a postural task. The inaccuracy of goal-directedhand movements was related to intention tremor andhypermetria in the target period and slowing during thedeceleration period in a three-dimensional kinematicanalysis of ET patients (Deuschl et al., 2000).
The aetiology of cerebellar dysfunction in ET is presentlyunder discussion (Louis, 2005). It is unlikely to resultfrom structural cerebellar damage, but may rather reflectinterference with the feed-forward function of the cerebel-lum caused by abnormal timing signals from a dysfunc-tional olivocerebellar circuitry. In animal studies, complexspike activities of olivocerebellar climbing fibres have beencorrelated with tracking direction, amplitude and speed(Ebner et al., 2002). In combination with simple spikeactivity carried by mossy fibres from the spinal cord,brainstem and cerebral cortex, they probably provide thebasis for a ‘velocity command signal’ essential for thecontrol of limb movement (van Kan et al., 1993; Ebner,1998). In the harmaline model of ET, there is enhancedrhythmic activity of complex spikes and simultaneouslyelimination of simple spikes (Llinas and Volkind, 1973;De Montigny and Lamarre, 1975), most likely causingdisintegration of sensory and olivary input on the level ofPurkinje neurons. This misarranged activation may perturbthe feed-forward function of the cerebellum (Allen andTsukahara, 1974) leading to disturbances in the decelerationand target period resembling ataxia in primary cerebellardisease. Support for this hypothesis comes from theeffect of ethanol which decreases complex spikes andincrease of single-spike activity in the animal model(Sinclair et al., 1982) and significantly improves cerebel-lar-like gait ataxia in ET (Klebe et al., 2005). However,experimental evidence for the beneficial effect of ethanoland subsequent normalisation of the olivocerebellarpathway on limb ataxia is lacking. A possible alternativeto a primary cerebellar dysfunction underlying imprecisetargeting in ET could be the massive interference of tremoritself with the proprioceptive feedback signal necessary to
Thalamic or subthalamic DBS in tremor Brain (2007), 130, 1608^1625 1621
by guest on Decem
ber 11, 2013http://brain.oxfordjournals.org/
Dow
nloaded from

control the reaching movement. Presently, it is not knownwhich pathophysiological mechanism is improved bythalamic DBS. However, modulating the cerebello–thalamicfibre system appears to be most effective in improvingataxia. This again favours a direct impact of thalamicneurostimulation on the propagation of pathologicalcerebellar outflow signals and not the interruption ofa reverberating tremor oscillation within the thalamo-cortical loop.In MS patients, reach-to-grasp movements are severely
compromised by overshot in the deceleration period as wellas intention tremor in the target period (Deuschl et al.,2000). Also abnormalities in the acceleration period ofelbow flexion movements have been described (Hallettet al., 1991). In MS patients suffering from cerebellar ataxia,infratentorial lesions in the brainstem and cerebellum arecommonly found on MRI (Nakashima et al., 1999).Intention tremor, in particular, is more frequentlyassociated with lesions in the cerebellar peduncles and thepons than in the cerebellar hemispheres (Feys et al., 2005b).Lesions in these areas would mainly affect the afferentpathways of the cerebellum, the rubroolivary and thecorticoolivary pathway, which could impair cerebellaractivation. Neurostimulation of the cerebello–thalamicprojections may disrupt pathological efferent neural activitywithin the cerebello–thalamo–cortical loop and therebyimprove ataxia. MS lesions, however, could also disturbascending projections within the cerebello–thalamicpathways. Additionally, disabled coordination in MSpatients has been correlated with supratentorial lesions inthe commissural fibres tracts (Charil et al., 2003). ThalamicDBS is unlikely to restore deficient efferent cerebellaractivity or to alleviate ataxia of cortical origin. This mightexplain the lower efficacy on ataxia in MS patientscompared to ET patients.Thalamic neurostimulation reduced the spatial variability
already in the deceleration period of reach-to-graspmovements in both MS and ET patients. This is probablyrelated to a reduced movement overshot which has beendescribed in previous studies of ET and MS tremor. In ourstudy, we did not compare kinematic parameters ofcontrols and patients. However, previous studies innormal controls demonstrated the regularity of movementtrajectories with only minimal deviations from the averagewrist path (Bastian et al., 1996). The reduced deviation inthe deceleration period by subthalamic area stimulationprobably reflects improvement of dysmetria, another featureof ataxia. The extent of improvement in the decelerationperiod is not as high as in the target period. However, pooraccuracy already in the deceleration period may lead tosubsequent agonist bursts as a compensatory mechanismwith the end result of intention tremor (Berardelli et al.,1996). Therefore, even moderate improvement in thedeceleration period may be causative for the much higherimprovement in the target period.
Clinical effect of thalamic DBSOn the clinical TRS our ET patients improved by 64% andour MS patients by 50%, which is in good accordance withprevious clinical studies. For ET, the average improvementof extremity tremor was between 50 and 80% and of headand voice tremor between 55 and 95% (Ondo et al., 1998;Limousin et al., 1999; Obwegeser et al., 2001; Sydow et al.,2003). In MS, most studies found a qualitative post-operative improvement in 50 to 100% of patients (Benabidet al., 1996; Geny et al., 1996; Montgomery et al., 1999;Berk et al., 2002; Hooper et al., 2002; Schulder et al., 2003).In the few studies providing quantitative assessment,thalamic DBS reduced the total TRS in a range between36 and 70% (Berk et al., 2002; Schulder et al., 2003; Bittaret al., 2005).
The lateralized TRS, which was used in our study tospecifically assess extremity tremor, was associated withslightly higher reduction in the range of 70 and 80% for ETpatients and 62 to 76% in MS patients. This experimentalparameter is difficult to compare to previous clinical trials,which typically evaluate tremor after prolonged periods ofstimulation, not immediately after a change in stimulationparameters.
In keeping with the objective experimental measures,clinical tremor ratings in ET and MS were markedlybetter for stimulation within the subthalamic area thanwithin ventrolateral thalamus. This emphasizes that ourexperimental design captured relevant features of symptomseverity.
Characterization of the subthalamic areaWhich could be the neural target elements of subthalamicarea stimulation in ET and MS tremor? After projectiononto the Schaltenbrand–Wahren atlas, the most effectiveelectrode contacts localized onto the prelemniscal radiationwhich is the posterior extension of field H of Forel. Fromlabelling and degeneration studies in animals, the course offibres from deep cerebellar nuclei to the cerebellar thalamushas been accurately defined (Mehler and Nauta, 1974;Stanton, 1980; Ilinsky and Kultas-Ilinsky, 1984). Afterleaving the dentate nucleus and crossing to the contralateralside, fibres form a distinct bundle which ascends rostrallyand passes through and around the lateral portion ofthe red nucleus. The bulk of fibres continuing in therostro-lateral direction passes through the field H of Foreland enters the thalamus, notably the VIM, through itsventral aspects. Stimulation within the subthalamic area,therefore, probably takes advantage of an anatomical ‘bottleneck’ where a large proportion of cerebellothalamicafferents can be affected by a small volume of currentspread, before the fibres spread out to innervate the entireVIM nucleus (Sakai et al., 1996). Three previous clinicalcase series of DBS in ET also described a more effectivetremor reduction by stimulation of subthalamic targets
1622 Brain (2007), 130, 1608^1625 J. Herzog et al.
by guest on Decem
ber 11, 2013http://brain.oxfordjournals.org/
Dow
nloaded from

including the field of Forel, prelemniscal radiation andZona incerta (Kitagawa et al., 2000; Murata et al., 2003;Plaha et al., 2004). There is no data so far on the clinicallyoptimal stimulation site in MS tremor, but in a series of 13MS tremor patients treated by thalamotomy the best resultswere obtained in patients where the lesion was confined tothe subthalamic area (Alusi et al., 2001a). This observationcorroborates the concepts of ‘subthalamotomy’ or ‘cam-potomy’ which has been established early in the era oflesional tremor surgery (Spiegel et al., 1963; Story et al.,1965; Mundinger, 1969; Velasco et al., 1972). A directcomparison of our findings to the results of lesionalsurgery, however, may be difficult for two reasons: first,localizations of lesions have not always been exactlydetermined due to lack of modern imaging techniquesin historical studies. Second, in both procedures theunderlying neural mechanism leading to tremor reductionis clearly distinct. In contrast to lesioning, DBS does notsilence neuronal activity but rather acts by modulatingpathological information transfer without the need fordestroying brain tissue.
ConclusionThalamic neurostimulation does not only reduce clinicaland electrophysiological measures of postural tremor inET or MS. It is also highly effective in decreasing the spatialvariability in the deceleration and target period ofgoal-directed movements, which are kinematic features ofcerebellar ataxia. This effect on target-oriented movementsmay underlie the clinically impressive improvement ofintention tremor in both conditions. The more pronouncedtremor reduction when stimulating the subthalamic areafavours the concept of modulating cerebello–thalamicprojections.
ReferencesAllen GI, Tsukahara N. Cerebrocerebellar communication systems.
Physiol Rev 1974; 54: 957–1006.
Alusi SH, Aziz TZ, Glickman S, Jahanshahi M, Stein JF, Bain PG.
Stereotactic lesional surgery for the treatment of tremor in multiple
sclerosis: a prospective case-controlled study. Brain 2001a; 124: 1576–89.
Alusi SH, Worthington J, Glickman S, Bain PG. A study of tremor in
multiple sclerosis. Brain 2001b; 124: 720–30.
Anderson ME, Postupna N, Ruffo M. Effects of high-frequency stimulation
in the internal globus pallidus on the activity of thalamic neurons in the
awake monkey. J Neurophysiol 2003; 89: 1150–60.
Anderson T, Hu B, Pittman Q, Kiss ZH. Mechanisms of deep brain
stimulation: an intracellular study in rat thalamus. J Physiol 2004; 559:
301–13.
Anderson TR, Hu B, Iremonger K, Kiss ZH. Selective attenuation of
afferent synaptic transmission as a mechanism of thalamic deep brain
stimulation-induced tremor arrest. J Neurosci 2006; 26: 841–50.
Bastian AJ, Martin TA, Keating JG, Thach WT. Cerebellar ataxia: abnormal
control of interaction torques across multiple joints. J Neurophysiol
1996; 76: 492–509.
Benabid AL, Pollak P, Gao D, Hoffmann D, Limousin P, Gay E, et al.
Chronic electrical stimulation of the ventralis intermedius nucleus of the
thalamus as a treatment of movement disorders. J Neurosurg 1996; 84:
203–14.
Berardelli A, Hallett M, Rothwell JC, Agostino R, Manfredi M,
Thompson PD, et al. Single-joint rapid arm movements in normal
subjects and in patients with motor disorders. Brain 1996; 119 (Pt 2):
661–74.
Berk C, Carr J, Sinden M, Martzke J, Honey CR. Thalamic deep brain
stimulation for the treatment of tremor due to multiple sclerosis: a
prospective study of tremor and quality of life. J Neurosurg 2002; 97:
815–20.
Beurrier C, Bioulac B, Audin J, Hammond C. High-frequency stimulation
produces a transient blockade of voltage-gated currents in subthalamic
neurons. J Neurophysiol 2001; 85: 1351–6.
Biary N, Koller W. Kinetic predominant essential tremor: successful
treatment with clonazepam. Neurology 1987; 37: 471–4.
Bittar RG, Hyam J, Nandi D, Wang S, Liu X, Joint C, et al.
Thalamotomy versus thalamic stimulation for multiple sclerosis
tremor. J Clin Neurosci 2005; 12: 638–42.
Britton TC, Thompson PD, Day BL, Rothwell JC, Findley LJ, Marsden CD.
Rapid wrist movements in patients with essential tremor. The critical
role of the second agonist burst. Brain 1994; 117 (Pt 1): 39–47.
Carone DA, Benedict RH, Dwyer MG, Cookfair DL, Srinivasaraghavan B,
Tjoa CW, et al. Semi-automatic brain region extraction (SABRE) reveals
superior cortical and deep gray matter atrophy in MS. Neuroimage
2006; 29: 505–14.
Ceballos-Baumann AO, Boecker H, Fogel W, Alesch F, Bartenstein P,
Conrad B, et al. Thalamic stimulation for essential tremor activates
motor and deactivates vestibular cortex. Neurology 2001; 56: 1347–54.
Charil A, Zijdenbos AP, Taylor J, Boelman C, Worsley KJ, Evans AC, et al.
Statistical mapping analysis of lesion location and neurological disability
in multiple sclerosis: application to 452 patient data sets. Neuroimage
2003; 19: 532–44.
De Montigny C, Lamarre Y. Effects produced by local applications of
harmaline in the inferior olive. Can J Physiol Pharmacol 1975; 53:
845–9.
Deuschl G, Bain P, Brin M. Consensus statement of the Movement
Disorder Society on Tremor. Ad Hoc Scientific Committee. Mov Disord
1998; 13 (Suppl 3): 2–23.
Deuschl G, Bergman H. Pathophysiology of nonparkinsonian tremors.
Mov Disord 2002; 17 (Suppl 3): S41–8.
Deuschl G, Raethjen J, Lindemann M, Krack P. The pathophysiology of
tremor. Muscle Nerve 2001; 24: 716–35.
Deuschl G, Wenzelburger R, Loffler K, Raethjen J, Stolze H. Essential
tremor and cerebellar dysfunction clinical and kinematic analysis of
intention tremor. Brain 2000; 123 (Pt 8): 1568–80.
Dick JP. Multicentre European study of thalamic stimulation in essential
tremor. J Neurol Neurosurg Psychiatry 2003; 74: 1362–3.
Dormont D, Cornu P, Pidoux B, Bonnet AM, Biondi A, Oppenheim C,
et al. Chronic thalamic stimulation with three-dimensional MR
stereotactic guidance. Ajnr: Am J Neuroradiol 1997; 18: 1093–107.
Eaton MJ, Moss RL. Electrophysiology and pharmacology of neurons of
the medial zona incerta: an in vitro slice study. Brain Res 1989; 502:
117–26.
Ebner TJ. A role for the cerebellum in the control of limb movement
velocity. Curr Opin Neurobiol 1998; 8: 762–9.
Ebner TJ, Johnson MT, Roitman A, Fu Q. What do complex spikes signal
about limb movements? Ann N Y Acad Sci 2002; 978: 205–18.
Elble RJ. Central mechanisms of tremor. J Clin Neurophysiol 1996; 13:
133–44.
Elble RJ. Animal models of action tremor. Mov Disord 1998; 13 (Suppl 3):
35–9.
Fahn S, Tolosa E, Marin C. Clinical rating scale for tremor. In:
Jankovic J, Tolosa E, editors. Parkinson’s disease and movement
disorders. Baltimore: Williams and Wilkins; 1993. p. 271–80.
Ferroli P, Franzini A, Marras C, Maccagnano E, D’Incerti L, Broggi G. A
simple method to assess accuracy of deep brain stimulation electrode
placement: pre-operative stereotactic CT þ postoperative MR image
fusion. Stereotactic Functional Neurosurg 2004; 82: 14–9.
Thalamic or subthalamic DBS in tremor Brain (2007), 130, 1608^1625 1623
by guest on Decem
ber 11, 2013http://brain.oxfordjournals.org/
Dow
nloaded from

Feys P, Helsen WF, Liu X, Nuttin B, Lavrysen A, Swinnen SP, et al.
Interaction between eye and hand movements in multiple sclerosis
patients with intention tremor. Mov Disord 2005a; 20: 705–13.
Feys P, Maes F, Nuttin B, Helsen W, Malfait V, Nagels G, et al.
Relationship between multiple sclerosis intention tremor severity and
lesion load in the brainstem. Neuroreport 2005b; 16: 1379–82.
Flament D, Hore J. Movement and electromyographic disorders associated
with cerebellar dysmetria. J Neurophysiol 1986; 55: 1221–33.
Frost JD, Jr. Triaxial vector accelerometry: a method for quantifying
tremor and ataxia. IEEE Trans Biomed Eng 1978; 25: 17–27.
Geny C, Nguyen JP, Pollin B, Feve A, Ricolfi F, Cesaro P, et al.
Improvement of severe postural cerebellar tremor in multiple sclerosis
by chronic thalamic stimulation. Mov Disord 1996; 11: 489–94.
Guiot G, Derome P, Robert G. Stereotaxic surgery of tremors.
Neurochirurgie 1976; 22: 532–5.
Hallett M, Berardelli A, Matheson J, Rothwell J, Marsden CD.
Physiological analysis of simple rapid movements in patients with
cerebellar deficits. J Neurol Neurosurg Psychiatry 1991; 54: 124–33.
Halliday DM, Conway BA, Farmer SF, Shahani U, Russell AJ,
Rosenberg JR. Coherence between low-frequency activation of the
motor cortex and tremor in patients with essential tremor. Lancet 2000;
355: 1149–53.
Holsheimer J, Demeulemeester H, Nuttin B, de Sutter P. Identification
of the target neuronal elements in electrical deep brain stimulation.
Eur J Neurosci 2000; 12: 4573–7.
Hooper J, Taylor R, Pentland B, Whittle IR. A prospective study of
thalamic deep brain stimulation for the treatment of movement
disorders in multiple sclerosis. Br J Neurosurg 2002; 16: 102–9.
Hore J, Wild B, Diener HC. Cerebellar dysmetria at the elbow, wrist, and
fingers. J Neurophysiol 1991; 65: 563–71.
Hua SE, Lenz FA, Zirh TA, Reich SG, Dougherty PM. Thalamic neuronal
activity correlated with essential tremor. J Neurol Neurosurg Psychiatry
1998; 64: 273–6.
Ilinsky IA, Kultas-Ilinsky K. An autoradiographic study of topographical
relationships between pallidal and cerebellar projections to the cat
thalamus. Exp Brain Res 1984; 54: 95–106.
Jankovic J, Frost JD, Jr. Quantitative assessment of parkinsonian and
essential tremor: clinical application of triaxial accelerometry. Neurology
1981; 31: 1235–40.
Kiss ZH, Anderson T, Hansen T, Kirstein D, Suchowersky O, Hu B.
Neural substrates of microstimulation-evoked tingling: a chronaxie
study in human somatosensory thalamus. Eur J Neurosci 2003a; 18:
728–32.
Kiss ZH, Davis KD, Tasker RR, Lozano AM, Hu B, Dostrovsky JO.
Kinaesthetic neurons in thalamus of humans with and without tremor.
Exp Brain Res 2003b; 150: 85–94.
Kiss ZH, Mooney DM, Renaud L, Hu B. Neuronal response to local
electrical stimulation in rat thalamus: physiological implications
for mechanisms of deep brain stimulation. Neuroscience 2002; 113:
137–43.
Kitagawa M, Murata J, Kikuchi S, Sawamura Y, Saito H, Sasaki H, et al.
Deep brain stimulation of subthalamic area for severe proximal tremor.
Neurology 2000; 55: 114–6.
Klebe S, Stolze H, Grensing K, Volkmann J, Wenzelburger R, Deuschl G.
Influence of alcohol on gait in patients with essential tremor. Neurology
2005; 65: 96–101.
Koster B, Deuschl G, Lauk M, Timmer J, Guschlbauer B, Lucking CH.
Essential tremor and cerebellar dysfunction: abnormal ballistic move-
ments. J Neurol Neurosurg Psychiatry 2002; 73: 400–5.
Krack P, Dostrovsky J, Ilinsky I, Kultas-Ilinsky K, Lenz F, Lozano A, et al.
Surgery of the motor thalamus: problems with the present nomencla-
tures. Mov Disord 2002; 17 (Suppl 3): S2–8.
Kurtzke JF. Rating neurologic impairment in multiple sclerosis: an
expanded disability status scale (EDSS). Neurology 1983; 33: 1444–52.
Lamarre Y, Mercier LA. Neurophysiological studies of harmaline-induced
tremor in the cat. Can J Physiol Pharmacol 1971; 49: 1049–58.
Lang EJ, Rosenbluth J. Role of myelination in the development of a
uniform olivocerebellar conduction time. J Neurophysiol 2003; 89:
2259–70.
Limousin P, Speelman JD, Gielen F, Janssens M. Multicentre European
study of thalamic stimulation in parkinsonian and essential tremor.
J Neurol Neurosurg Psychiatry 1999; 66: 289–96.
Liu X, Ingram HA, Palace JA, Miall RC. Dissociation of ‘on-line’ and
‘off-line’ visuomotor control of the arm by focal lesions in the
cerebellum and brainstem. Neurosci Lett 1999; 264: 121–4.
Liu X, Miall C, Aziz TZ, Palace JA, Haggard PN, Stein JF. Analysis of
action tremor and impaired control of movement velocity in multiple
sclerosis during visually guided wrist-tracking tasks. Mov Disord 1997;
12: 992–9.
Llinas R, Volkind RA. The olivo-cerebellar system: functional properties as
revealed by harmaline-induced tremor. Exp Brain Res 1973; 18: 69–87.
Louis ED. Essential tremor. Lancet Neurol 2005; 4: 100–10.
Magarinos-Ascone C, Pazo JH, Macadar O, Buno W. High-frequency
stimulation of the subthalamic nucleus silences subthalamic neurons: a
possible cellular mechanism in Parkinson’s disease. Neuroscience 2002;
115: 1109–17.
McIntyre CC, Grill WM, Sherman DL, Thakor NV. Cellular effects of deep
brain stimulation: model-based analysis of activation and inhibition.
J Neurophysiol 2004; 91: 1457–69.
Mehler WR, Nauta WJ. Connections of the basal ganglia and of the
cerebellum. Confin Neurol 1974; 36: 205–22.
Montgomery EB, Jr, Baker KB, Kinkel RP, Barnett G. Chronic thalamic
stimulation for the tremor of multiple sclerosis. Neurology 1999; 53:
625–8.
Mundinger F. Results of 500 subthalamotomies in the region of the zona
incerta. Third Symposium on Parkinson’s Disease. Edinburgh and
London: E&S Livingstone; 1969. p. 261–5.
Murata J, Kitagawa M, Uesugi H, Saito H, Iwasaki Y, Kikuchi S, et al.
Electrical stimulation of the posterior subthalamic area for the treatment
of intractable proximal tremor. J Neurosurg 2003; 99: 708–15.
Nakashima I, Fujihara K, Okita N, Takase S, Itoyama Y. Clinical and
MRI study of brain stem and cerebellar involvement in Japanese
patients with multiple sclerosis. J Neurol Neurosurg Psychiatry 1999; 67:
153–7.
Obwegeser AA, Uitti RJ, Witte RJ, Lucas JA, Turk MF, Wharen RE, Jr.
Quantitative and qualitative outcome measures after thalamic deep
brain stimulation to treat disabling tremors. Neurosurgery 2001; 48:
274–81; discussion 281–4.
Ondo W, Jankovic J, Schwartz K, Almaguer M, Simpson RK. Unilateral
thalamic deep brain stimulation for refractory essential tremor and
Parkinson’s disease tremor. Neurology 1998; 51: 1063–9.
Perlmutter JS, Mink JW, Bastian AJ, Zackowski K, Hershey T, Miyawaki E,
et al. Blood flow responses to deep brain stimulation of thalamus.
Neurology 2002; 58: 1388–94.
Plaha P, Patel NK, Gill SS. Stimulation of the subthalamic region for
essential tremor. J Neurosurg 2004; 101: 48–54.
Raethjen J, Pawlas F, Lindemann M, Wenzelburger R, Deuschl G.
Determinants of physiologic tremor in a large normal population.
Clin Neurophysiol 2000; 111: 1825–37.
Ranck JB, Jr. Which elements are excited in electrical stimulation of
mammalian central nervous system: a review. Brain Res 1975; 98:
417–40.
Sakai ST, Inase M, Tanji J. Comparison of cerebellothalamic and
pallidothalamic projections in the monkey (Macaca fuscata): a double
anterograde labeling study. J Comp Neurol 1996; 368: 215–28.
Schaltenbrand G, Wahren W. Atlas for stereotaxy of the human brain.
Stuttgart: Thieme; 1977.
Schulder M, Sernas TJ, Karimi R. Thalamic stimulation in patients with
multiple sclerosis: long-term follow-up. Stereotact Funct Neurosurg
2003; 80: 48–55.
Sinclair JG, Lo GF, Harris DP. Ethanol effects on the olivocerebellar
system. Can J Physiol Pharmacol 1982; 60: 610–4.
1624 Brain (2007), 130, 1608^1625 J. Herzog et al.
by guest on Decem
ber 11, 2013http://brain.oxfordjournals.org/
Dow
nloaded from

Spiegel EA, Wycis HT, Szekely EG, Adams DJ, Flanagan M, Baird HW 3rd.
Campotomy in various extrapyramidal disorders. J Neurosurg 1963; 20:
871–84.
Stanton GB. Topographical organization of ascending cerebellar
projections from the dentate and interposed nuclei in Macaca mulatta:
an anterograde degeneration study. J Comp Neurol 1980; 190: 699–731.
Steriade M, Jones E, McCormick D. Thalamus. New York: Elsvier; 1997.
Story JL, French LA, Chou SN, Meier MJ. Experiences with subthalamic
lesions in patients with movement disorders. Confin Neurol 1965; 26:
218–21.
Sydow O, Thobois S, Alesch F, Speelman JD. Multicentre European study
of thalamic stimulation in essential tremor: a six year follow up.
J Neurol Neurosurg Psychiatry 2003; 74: 1387–91.
Topka H, Konczak J, Dichgans J. Coordination of multi-joint arm
movements in cerebellar ataxia: analysis of hand and angular kinematics.
Exp Brain Res 1998; 119: 483–92.
Tranchant C, Bhatia KP, Marsden CD. Movement disorders in multiple
sclerosis. Mov Disord 1995; 10: 418–23.
Vaillancourt DE, SturmanMM, Verhagen Metman L, Bakay RA, Corcos DM.
Deep brain stimulation of the VIM thalamic nucleus modifies several
features of essential tremor. Neurology 2003; 61: 919–25.
van Kan PL, Houk JC, Gibson AR. Output organization of intermediate
cerebellum of the monkey. J Neurophysiol 1993; 69: 57–73.
Velasco F, Molina-Negro P. Electrophysiological topography of the human
diencephalon. J Neurosurg 1973; 38: 204–14.
Velasco F, Velasco M, Maldonado H. Identification and lesion
of prelemniscal radiation in the surgical treatment of tremor.
Arch Invest Med (Mex) 1976; 7: 29–42.
Velasco FC, Molina-Negro P, Bertrand C, Hardy J. Further definition of
the subthalamic target for arrest of tremor. J Neurosurg 1972; 36:
184–91.
von Hofsten C. Strucuturing in early reaching movements: a longitudinal
study. J Mot Behav 1991; 23: 280–92.
Wilms H, Sievers J, Deuschl G. Animal models of tremor. Mov Disord
1999; 14: 557–71.
Wishart HA, Roberts DW, Roth RM, McDonald BC, Coffey DJ,
Mamourian AC, et al. Chronic deep brain stimulation for the
treatment of tremor in multiple sclerosis: review and case reports.
J Neurol Neurosurg Psychiatry 2003; 74: 1392–7.
Wylezinska M, Cifelli A, Jezzard P, Palace J, Alecci M, Matthews PM.
Thalamic neurodegeneration in relapsing-remitting multiple sclerosis.
Neurology 2003; 60: 1949–54.
Zackowski KM, Bastian AJ, Hakimian S, Mink JW, Perlmutter JS,
Koller WC, et al. Thalamic stimulation reduces essential tremor
but not the delayed antagonist muscle timing. Neurology 2002; 58:
402–10.
Thalamic or subthalamic DBS in tremor Brain (2007), 130, 1608^1625 1625
by guest on Decem
ber 11, 2013http://brain.oxfordjournals.org/
Dow
nloaded from




















