
Hairy Baskets" Associated With Degenerative Purkinje Cell Changes in Essential Tremor
20
“Hairy Baskets” Associated with Degenerative Purkinje Cell Changes in Essential Tremor Cordelia Erickson-Davis, BA 1 , Phyllis L. Faust, MD, PhD 2 , Jean-Paul G. Vonsattel, MD 2,3 , Sachin Gupta, MD 1 , Lawrence S. Honig, MD, PhD 1,3,4 , and Elan D. Louis, MD, MSc 1,3,4,5 1 GH Sergievsky Center, College of Physicians and Surgeons, Columbia University, New York, NY 2 Department of Pathology and Cell Biology, College of Physicians and Surgeons, Columbia University, New York, NY 3 Taub Institute for Research on Alzheimer’s Disease and the Aging Brain, College of Physicians and Surgeons, Columbia University, New York, NY 4 Department of Neurology, College of Physicians and Surgeons, Columbia University, New York, NY 5 Department of Epidemiology, Mailman School of Public Health, Columbia University, New York, NY Abstract Essential tremor (ET) is one of the most common neurological diseases. Increased numbers of torpedoes and Purkinje cell loss have been documented in the brains of patients with ET. We recently observed a dense and tangled appearance (“hairiness”) of the basket cell axonal plexuses that surround Purkinje cell soma in Bielschowsky preparations of cerebellar cortex in ET brains. Here, we assessed basket cell “hairiness” in 37 ET (32 cerebellar ET; 5 Lewy body variant ET [LBVET]), 21 non-disease control, and 48 disease control brains using a semiquantitative scale. In 8 cerebellar ET cases (25%) there were high basket scores (rating = 3), whereas no LBVET, 1 non-disease control (4.8%) and 2 diseased controls (4.2%) had high basket scores (p = 0.001). The hairy basket scores correlated with numbers of torpedoes (p < 0.001) and inversely with numbers of Purkinje cells (p = 0.06). Axonal plexus density obtained by image analysis of basket cell processes traced from digitized images was higher in ET than in non-diseased control cases (p = 0.016). Closely spaced sites of synaptic contact between basket cell processes and Purkinje cells were identified by electron microscopy in ET cases. These data indicate that structural changes are not restricted to Purkinje cells in ET and that other neurons within their functional network may be involved in its pathogenesis. Keywords Basket cells; Cerebellum; Essential tremor; Neurodegenerative; Pathology; Pathophysiology; Purkinje cells INTRODUCTION Essential tremor (ET) is a chronic brain disease, the most recognizable feature of which is a 4 to 12 Hz action tremor (1,2). It is one of the most common neurological disorders present in Correspondence and reprint requests to: Dr. Elan Louis, Unit 198, Neurological Institute, 710 West 168th Street, New York, NY, 10032. Tel: (212) 305 – 9194; Fax: (212) 305-1304; [email protected]. NIH Public Access Author Manuscript J Neuropathol Exp Neurol. Author manuscript; available in PMC 2011 March 1. Published in final edited form as: J Neuropathol Exp Neurol. 2010 March ; 69(3): 262–271. doi:10.1097/NEN.0b013e3181d1ad04. NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
Transcript of Hairy Baskets" Associated With Degenerative Purkinje Cell Changes in Essential Tremor

“Hairy Baskets” Associated with Degenerative Purkinje CellChanges in Essential Tremor
Cordelia Erickson-Davis, BA1, Phyllis L. Faust, MD, PhD2, Jean-Paul G. Vonsattel, MD2,3,Sachin Gupta, MD1, Lawrence S. Honig, MD, PhD1,3,4, and Elan D. Louis, MD, MSc1,3,4,51 GH Sergievsky Center, College of Physicians and Surgeons, Columbia University, New York, NY2 Department of Pathology and Cell Biology, College of Physicians and Surgeons, ColumbiaUniversity, New York, NY3 Taub Institute for Research on Alzheimer’s Disease and the Aging Brain, College of Physiciansand Surgeons, Columbia University, New York, NY4 Department of Neurology, College of Physicians and Surgeons, Columbia University, New York,NY5 Department of Epidemiology, Mailman School of Public Health, Columbia University, New York,NY
AbstractEssential tremor (ET) is one of the most common neurological diseases. Increased numbers oftorpedoes and Purkinje cell loss have been documented in the brains of patients with ET. We recentlyobserved a dense and tangled appearance (“hairiness”) of the basket cell axonal plexuses thatsurround Purkinje cell soma in Bielschowsky preparations of cerebellar cortex in ET brains. Here,we assessed basket cell “hairiness” in 37 ET (32 cerebellar ET; 5 Lewy body variant ET [LBVET]),21 non-disease control, and 48 disease control brains using a semiquantitative scale. In 8 cerebellarET cases (25%) there were high basket scores (rating = 3), whereas no LBVET, 1 non-disease control(4.8%) and 2 diseased controls (4.2%) had high basket scores (p = 0.001). The hairy basket scorescorrelated with numbers of torpedoes (p < 0.001) and inversely with numbers of Purkinje cells (p =0.06). Axonal plexus density obtained by image analysis of basket cell processes traced from digitizedimages was higher in ET than in non-diseased control cases (p = 0.016). Closely spaced sites ofsynaptic contact between basket cell processes and Purkinje cells were identified by electronmicroscopy in ET cases. These data indicate that structural changes are not restricted to Purkinjecells in ET and that other neurons within their functional network may be involved in its pathogenesis.
KeywordsBasket cells; Cerebellum; Essential tremor; Neurodegenerative; Pathology; Pathophysiology;Purkinje cells
INTRODUCTIONEssential tremor (ET) is a chronic brain disease, the most recognizable feature of which is a 4to 12 Hz action tremor (1,2). It is one of the most common neurological disorders present in
Correspondence and reprint requests to: Dr. Elan Louis, Unit 198, Neurological Institute, 710 West 168th Street, New York, NY, 10032.Tel: (212) 305 – 9194; Fax: (212) 305-1304; [email protected].
NIH Public AccessAuthor ManuscriptJ Neuropathol Exp Neurol. Author manuscript; available in PMC 2011 March 1.
Published in final edited form as:J Neuropathol Exp Neurol. 2010 March ; 69(3): 262–271. doi:10.1097/NEN.0b013e3181d1ad04.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

4.0% of individuals ≥40 years of age (3), and in as many as 21.7% of individuals aged 95 yearsand older (4). The tremor is usually progressive (5) and produces functional disabilities (6).Recent evidence also suggests an associated increased risk of mortality (7).
A wealth of clinical data suggests that ET is a disorder of cerebellar dysfunction. Intention (i.e.“cerebellar”) tremor of the hands occurs in 44% of ET cases (8). Abnormalities in tandem gaitand balance have been repeatedly described in ET patients (9–11), and ET patients withintention tremor may also have other cerebellar signs such as dysdiadochokinesia (12). Eyemovement abnormalities that indicate cerebellar dysfunction have also been described in ET(13). Unilateral cerebellar stroke has been reported to abolish ipsilateral arm tremor in ET(14) and cerebellar outflow pathways are the targets of deep brain stimulation surgery, whichis effective in treating ET (15,16). Numerous neuroimaging studies employing functionalmagnetic resonance imaging (17), positron emission tomography (18,19), [1H] magneticresonance spectroscopic imaging (20,21), diffusion-tensor imaging (22), and voxel-basedmorphometry (23) have provided evidence that cerebellar structure and function are abnormalin ET.
Until recently, there have been few postmortem studies of ET (24). Studies that explore thepathological anatomy and improve our understanding of the pathophysiology of ET arecritically important since there is no cure for ET; the 2 first-line medications are estimated tobe ineffective in as many as 50% of patients (25). In each of 2 recent large postmortem series,the presence of degenerative changes in the cerebellum has become evident (24,26–28). Todate, these changes consist of an increased number of torpedoes (5- to 6-fold greater than seenin age-matched control brains) and a mild, approximately 40%, loss of Purkinje cells (PCs)(27,29). Whether the structural changes in ET are restricted to PCs or whether they involveother neurons in their functional network have not been investigated.
Recently, we observed an unusual dense and tangled (we now term, “hairy”) appearance of thebasket cell axonal plexus surrounding PC soma in Bielschowsky preparations of cerebellarcortical sections in ET cases. Basket cells are γ-aminobutyric acid (GABA)-ergic inhibitoryinterneurons found in the molecular layer; they send out axonal collaterals to form a pericellularbasket around PC perikarya. To our knowledge, there are only 2 prior reports describing adense and tangled appearance of basket cell axonal plexus profiles occurring with cerebellardegeneration: a single case of Creutzfeldt-Jakob disease (30) and an early study examiningbrains of individuals diagnosed with “senile dementia” (31). Because this pathological findingmay provide important mechanistic insights into the pathogenesis of ET, we assessed basketcell changes in ET, other neurodegenerative disease and non-diseased control cases.
MATERIALS AND METHODSBrain Repository
This study was conducted at the Essential Tremor Centralized Brain Repository (ETCBR) atthe New York Brain Bank, Columbia University (32). The 106 brains included 37 from ETcases; 31 (83.8%) of these were collected prospectively beginning in 2003 and 6 were archival.All 37 had been diagnosed by their treating neurologist and all diagnoses were confirmed usingETCBR criteria (27). ET cases were divided into 2 groups based on the presence or absenceof brainstem Lewy bodies on postmortem examination (27,29,33). ET brains without Lewybodies have an array of cerebellar abnormalities that includes increased numbers of torpedoesand PC loss (27,29). The Lewy body-free brains have been referred to as cerebellar ET andbrains with Lewy bodies as Lewy body variant of ET (LBVET) in previous publications (24,32). Demographic and clinical information were collected on each brain; the severity of tremoron Archimedes spiral drawings were rated using a validated rating scale (0–3) by a neurologist(EDL) who was blinded to postmortem data (35).
Erickson-Davis et al. Page 2
J Neuropathol Exp Neurol. Author manuscript; available in PMC 2011 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Diseased control brains (n = 49) included individuals diagnosed with AD (n = 20), PD or DLBD(n = 15) that were selected for matching to the age distribution of available cerebellar ET brains.AD brains were from individuals who had been enrolled as AD cases in the Alzheimer DiseaseResearch Center or the Washington Heights Inwood Columbia Aging Project, where they werefollowed prospectively with serial neurological assessments, including assessment of actiontremor, and were free of ET, PD, DLBD or PSP. PD or DLBD brains at the New York BrainBank had been patients at the Movement Disorder Division, Neurological Institute of NewYork or were outside referrals to the brain bank. We also used the brains of 12 PSP cases andof 1 case of clinically diagnosed adult-onset focal dystonia (torticollis) even if they wereyounger than the cerebellar ET cases since tissue availability was limited.
Non-diseased control brains (n = 21) were matched to the age distribution of cerebellar ETbrains and were from individuals who had been enrolled as normal elderly control subjects inthe Alzheimer’s Disease Research Center or the Washington Heights Inwood Columbia AgingProject; they were followed prospectively with serial neurological assessments and were freeof the diagnosis of AD, ET, PD, DLBD or PSP, and had normal neuropathologicalexaminations.
Tissue ProcessingAll brains underwent complete neuropathological assessment at the New York Brain Bank(26,27,32,33,36). Brains received ratings of neurofibrillary tangles using Braak and Braakstaging (37,38) and Consortium to Establish a Registry for Alzheimer’s Disease (CERAD)ratings (39) for neuritic plaques. National Institute on Aging - Reagan Institute (NIA - RI)criteria for AD (40) were also assigned. Postmortem interval (PMI) was the number of hoursbetween death and placement of the brain in a cold room or on ice.
A standard 3 × 20 × 25 mm parasagittal tissue block from the neocerebellum was harvestedfrom each brain as described (26,27,29,33). Seven-μm-thick paraffin sections, impregnatedusing a modified Bielschowsky silver technique, were used for semiquantitative andquantitative analyses of basket cell processes and quantification of torpedoes. Sections werealso stained with Luxol fast blue-hematoxylin and eosin (LH&E) for quantification oftorpedoes and PCs (26,27,33). For immunohistochemical analyses, sections were incubatedwith mouse monoclonal SMI-31 (Covance, Princeton, NJ), as previously described (41). Ineach brain, torpedoes in an entire LH&E section and an entire Bielschowsky preparation werecounted, and PCs in five 100× LH&E fields were counted and averaged, as previouslydescribed (27).
Semiquantitative Rating of Hairy BasketsA semiquantitative rating of the appearance of the basket cell plexus surrounding PC bodiesthroughout Bielschowsky preparations was carried out by a senior neuropathologist (P.L.F.)who was blinded to all clinical information. The following scale was used: 0 (few, or nodiscernible processes); 1 (sparse number of processes); 2 (moderate number of processes); and3 (dense tangle of processes). In some instances, the rater used intermediate values (0.5, 1.5,and 2.5). Hence, the hairy basket rating ranged included the values 0, 0.5, 1, 1.5, 2, 2.5 and 3.
Quantitative Analysis of Basket Cell PlexusQuantification of the density of the basket cell axonal plexus formations was performed on asub-sample of 10 brains (including 5 randomly-selected cerebellar ET brains and 5 randomlyselected non-diseased control brains) by a single trained physician who was blinded to clinicalinformation. Digital images of the Bielschowsky neocerebellum preparations were obtainedusing a Zeiss Axioplan 2 microscope fit with an Axiocam HR digital camera (20× objectivelens). For image acquisition, slides were oriented with the dentate nucleus situated on the right
Erickson-Davis et al. Page 3
J Neuropathol Exp Neurol. Author manuscript; available in PMC 2011 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

and the folia branches extending leftward. Folia were labeled numerically and the 2 folialocated most centrally in the slide were selected. Within each of these folia, 3 smaller sub-regions consisting of 1 to 2 distal peaks (gyri) and 1 to 2 distal troughs (sulci) were chosensuch that there were 6 sub-regions photographed per brain. In each photograph, the basket cellprocesses surrounding each visible PC perikaryon were traced using Adobe Photoshop 5.0(approximately 40 PCs/brain). Images of the basket cell tracings were then inverted andimported into Image J (NIH), where the integrated density was assessed (i.e. a function of theprogram whereby each pixel of the basket tracing was assigned a gray value using a variablegray scale, and then summed).
Ultrastructural AnalysisFlat Eponate 12 (Ted Pella, Redding, CA) embedded vibratome sections of cerebellar corticaltissue were dissected into 3 × 4 mm sections and re-embedded in the same resin. Semithinsections were then stained with toluidine blue and examined by light microscopy for PCs. OncePCs were identified on semithin section, the block was further trimmed around the PCs to thesize required for ultrathin sectioning. Ultrathin sections were stained with uranyl acetate andlead citrate and then examined by transmission electron microscope (JEOL 1200EXII,Peabody, MA), as described (41).
Statistical AnalysesAll analyses were performed in SPSS (Version 16.0). As noted above, a priori, the focus ofour interest was cerebellar ET cases rather than LBVET cases. Demographic and clinicalfeatures of diagnostic groups (e.g. cerebellar ET vs. controls) were compared using Chi-squaretests and analysis of variance (ANOVA) with Tukey’s post-hoc comparisons (Table 1).
The hairy basket rating included the values 0, 0.5, 1, 1.5, 2, 2.5 and 3. In non-diseased controls,the correlates of the hairy basket rating were assessed using Pearson’s correlation coefficients(e.g., hairy basket rating by age). To facilitate further analyses, hairy basket ratings were thencollapsed into a smaller number of rating categories as follows: low (0 or 1, and also includingany ratings of 0.5 and 1.5); intermediate (2 and also including any ratings of 2.5); and high(3). Diagnostic groups were compared with regards to hairy basket rating category (Mantel-Haenszel Chi-square test, Table 2). For several analyses, diagnostic groups were combined(AD + PD/DLBD + PSP + dystonia = diseased controls; diseased controls + non-diseasedcontrols = all controls, Table 2). Due to the modest sample size, basket cell plexus density wascompared across diagnostic groups using the Mann Whitney test.
In an unadjusted logistic regression model, diagnosis (cerebellar ET vs. all controls) was theoutcome variable. In this model, the hairy basket rating category was the independentcategorical variable (high, intermediate, and low, as described above). In adjusted regressionmodels, covariates that were associated with either the outcome or independent variable or forwhich a priori evidence was considerable that the variable may be a confounder were included.
The correlates of the hairy basket rating in cerebellar ET cases and non-diseased controls andin cerebellar ET cases alone were determined using Pearson’s correlation coefficient whenboth variables were normally distributed; otherwise, Spearman’s correlation coefficient wasused. The degeneration index was defined as the number of torpedoes on Bielschowsky sectiondivided by the mean number of PCs per 100× LH&E-stained high-power field. High valuesindicated large numbers of torpedoes and/or low PC counts (i.e. PC loss) (42).
Erickson-Davis et al. Page 4
J Neuropathol Exp Neurol. Author manuscript; available in PMC 2011 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

RESULTSET, Non-Diseased Controls and Diseased Controls
Cerebellar ET cases were similar to LBVET, AD, and PD/DLBLD cases and non-diseasedcontrols with respect to age and PMI. However, when diseased controls (AD + PD/DLBD +PSP + dystonia) were combined and all controls were combined (diseased controls + non-diseased controls), cerebellar ET cases were older by an average ~5 years and mean PMI wasshorter (Table 1). All groups were similar with respect to gender and brain weight (Table 1).CERAD scores and Braak AD scores were higher in AD cases than cerebellar ET cases (Table1). Braak AD scores were higher in cerebellar ET cases than non-diseased controls (Table 1).
Basket Cell Plexus Alterations in Cerebellar ETBielschowsky sections of cerebellar neocortex revealed variable appearances of basket cellaxon collaterals that surround the PC perikarya. Figure 1 illustrates examples of thesemiquantitative scores of the basket cell plexus morphologies. “Hairy baskets” (rating = 3,the severe end of the spectrum) were defined as a dense and tangled axonal plexus (Fig. 1D).These hairy baskets generally surrounded viable-appearing PCs rather than PCs that hadpreviously died (i.e. “empty baskets”). In each of the 8 ET cases with hairy basket scores of 3,20 hairy baskets were assessed (160 hairy baskets). PC somata were visible in 141 of these(88.1%). Immunohistochemistry highlighted the neurofilament-rich basket cell axonalprocesses that comprise the hairy baskets and PC axonal torpedoes (Fig. 2).
We also assessed whether the hairy basket rating was similar in different neocerebellar regionswithin the same brain. For this analysis, 1 Bielschowsky section was examined from 3 separateneocerebellar blocks in 7 brains with hairy basket ratings ranging from 0 to 3. The agreementbetween ratings was high (Block 1 vs. Block 2, Pearson’s r = 0.91, p = 0.005; Block 1 vs. Block3, Pearson’s r = 0.95, p = 0.001; and Block 2 vs. Block 3, Pearson’s r = 0.96, p < 0.001). Thisindicated that the data, from a single block were likely to be broadly representative of the entireneocerebellum.
The hairy basket rating was not correlated with age (Pearson’s r = 0.24, p = 0.31), PMI(Pearson’s r = −0.04, p = 0.87), brain weight (Pearson’s r = −0.20, p = 0.41), CERAD score(Pearson’s r = −0.09, p = 0.71) or Braak AD score (Pearson’s r = 0.05, p = 0.83). It also didnot differ by gender in non-diseased controls (1.5 ± 0.9 in males vs. 1.1 ± 0.8 in females, p =0.34).
Hairy basket rating category was higher in cerebellar ET cases than in AD, PSP, dystonia, PD/DLBD (marginal difference) and non-diseased controls (Table 2). When control groups werecombined, the differences with cerebellar ET were more marked. In 8 out of 32 ET cases(25.0%) there was a hairy basket rating of 3, whereas only 4.2% of diseased controls and 4.8%of non-diseased controls had a score of 3 (Table 2). In an unadjusted logistic regression modelin which diagnosis (cerebellar ET vs. all controls) was the outcome variable, brains in the high(rating = 3) hairy basket rating category were 10 times more likely to be cerebellar ET thancontrols (odds ratio [OR] = 10.05, 95% confidence interval [CI] = 2.33–43.32, p = 0.002).Brains in the intermediate (rating = 2) basket rating category were 2 to 3 times more likely tobe cerebellar ET than controls (OR = 2.44, 95% CI = 0.92 – 6.46, p = 0.07). In a logisticregression model that adjusted for age, gender, PMI, brain weight, CERAD score, and BraakAD score, these values were as follows: OR = 18.95 (95% CI = 1.88–191.56, p = 0.01) andOR = 3.52 (95% CI = 0.82–15.02, p = 0.089).
To provide a quantitative measure of these basket cell plexus alterations, the axonal plexusessurrounding extant PCs in 5 cerebellar ET cases and 5 non-diseased controls were traced fromBielschowsky preparation images (Fig. 3A–D). When the densities of the tracings were
Erickson-Davis et al. Page 5
J Neuropathol Exp Neurol. Author manuscript; available in PMC 2011 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

determined, the average basket cell plexus in cerebellar ET cases was approximately twice thedensity of the average plexus in non-diseased controls (mean = 13.97 ± 3.56 [median = 15.10]vs. 6.98 ± 2.20 [median = 6.68], Mann- Whitney z = 2.40, p = 0.016) (Figure 3E). Basket cellplexus density was highly correlated with the hairy basket rating (Pearson’s r = 0.73, p = 0.018).Basket cell plexus density was also strongly associated with the number of torpedoes(Spearman’s r = 0.79, p = 0.007), marginally inversely with the number of PCs (Spearman’s r= −0.25, p = 0.49), and with the degeneration index (Spearman’s r = 0.76, p = 0.01).
Ultrastructural Analysis of the Basket Cell PlexusWe performed ultrastructural studies to compare basket cell processes in ET and control cases.In an ET case with a hairy basket rating of 3, there were several layers of neurofilament-richbasket cell processes adjacent to the PC soma, which intertwined at various angles (Fig. 4A,C–E). In contrast, in a non-diseased control with a hairy basket rating of 0, only 1 or 2 basketcell processes having a somewhat thinner diameter were found adjacent to the PC soma (Fig.4B).
The identity of these neurofilament-rich processes as basket cell fibers was further confirmedin the ET case by the identification of synapses with the PC soma and its initial axon segment.In several instances, contacts with the PC somata were marked by multiple, closely spacedsynaptic sites within 1 basket cell process (Fig. 5A–C, arrows). A basket fiber axon was alsoidentified making contact with the axon hillock of the Purkinje cell. In the basket axon, therewere multiple synaptic sites marked by vesicle accumulations beneath the axonal membrane(Figure 5D, d′). These findings further corroborate and extend our light microscopic findingsof basket cell plexus morphology.
Correlates of the Hairy Basket RatingThe hairy basket rating was robustly correlated with number of torpedoes (Spearman’s r =0.49, p < 0.001) (Fig. 6A), inversely with the number of PCs (Spearman’s r = −0.27, p = 0.06)(Fig. 6B), and inversely with the degeneration index (Spearman’s r = 0.48, p = 0.001) (Fig.6C), indicating that these basket cell changes were associated with torpedo formation and PCloss. Within cerebellar ET cases, the hairy basket rating was associated with age (Pearson’s r= 0.35, p = 0.48) and marginally with disease duration (disease duration was known in 19 casesand Pearson’s r = 0.35, p = 0.19). Hairy basket rating was associated with number of torpedoes(Spearman’s r = 0.49, p = 0.005) but not with number of PCs (Pearson’s r = 0.01, p = 0.98);however, it was associated with the degeneration index (Spearman’s r = 0.45, p = 0.01). Hairybasket rating did not differ by family history (presence vs. absence of ≥1 first-degree relativewith ET), age of tremor onset (Pearson’s r = −0.21, p = 0.44), severity (0–3) of tremor onArchimedes spiral (Pearson’s r = −0.08, p = 0.79) or presence versus absence of head tremor(data not shown).
DISCUSSIONRecent postmortem studies have documented degenerative changes in the cerebellum of ETcases, as manifested by increased numbers of torpedoes and loss of PCs (24,27–29,42). Thesepostmortem data complement a rich history of clinical, physiological and imaging studies thathave long implicated cerebellar and cerebellothalamacortical circuitry abnormalities in ET(8–15,19–21,43). While prior pathologic studies focused on degenerative changes in PCs, therelationship between ET and cerebellar interneurons has remained unexplored. We examinedarchival and prospectively acquired cerebellar tissue of ET cases and both diseased and non-diseased controls and found that the axonal plexuses of basket cells surrounding PCs incerebellar ET brains are significantly “hairier,” as compared to controls.
Erickson-Davis et al. Page 6
J Neuropathol Exp Neurol. Author manuscript; available in PMC 2011 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Basket cells are GABA-ergic inhibitory interneurons that receive input from parallel fibers andto a limited extent from climbing and mossy fibers. In humans, up to 50 axon collaterals fromneighboring basket cells descend from the molecular layer and combine to form a complexbasket structure around the PC soma, to which the basket cells’ entire axonal output is devoted(44). Reductions in cerebellar basket cell axonal plexuses have been reported in several diseasestates, including hypothyroidism (44), ataxia telangiectasia (45), and Menkes kinky hairdisease (46). Preservation of basket cell plexuses in the setting of PC death (i.e. “emptybaskets”) has been reported in Creutzfeldt-Jakob disease (47) and spinocerebellar ataxia type1 (48), but a dense and tangled appearance of the axonal plexus profile has not to our knowledgebeen documented in the ataxia literature. Curiously, an early 20th century study, describing PCabnormalities in 16 brains of individuals diagnosed with “senile dementia”, noted increasedbasket cell fiber “tangles and masses” alongside PC soma showing signs of degeneration in 3cases (31). Another more recent study qualitatively examined basket cells in Creutzfeldt-Jakobdisease and noted empty baskets with abnormally dense phosphorylated neurofilamentpositivity in a single case (31). Here, we documented that individuals with a diagnosis ofcerebellar ET were more than 10 times as likely as controls (both diseased and non-diseased)to have a hairy basket rating of 3 (the marked or severe end of the spectrum). Also, the medianaxonal plexus density in cerebellar ET was approximately 2 to 3 times higher than in controls.“Hairiness” of basket cell axonal plexuses in these independent semiquantitative andquantitative measures was positively associated with number of torpedoes and the degenerationindex, and inversely with number of PCs, suggesting that hairy baskets and torpedoes may beconcomitant features of cerebellar degeneration in ET.
The mechanism by which this increased “hairiness” occurs is unknown. One possibleexplanation is that the increased profiles observed in our study represent an accumulation ofconverging basket cell processes recruited from neighboring PCs that have been damaged ordied. Although there has been little investigation of such a phenomenon in the humancerebellum, selective preservation and reorganization of basket cell axonal processes has beendemonstrated in basket cells in the CA1 and CA3 region of the hippocampus (49,50). Thesecells form baskets around hippocampal pyramidal cells and function as local circuit inhibitoryGABA-ergic interneurons, analogous to the relationship between cerebellar basket cells andPCs. These disease-resistant hippocampal basket cells undergo extensive reorganization in thesetting of pyramidal cell death (51). In a few cases, basket cell plexuses displayed increaseddensity adjacent to areas with severe pyramidal cell loss and no basket formations, suggestingthat these basket cell processes might be converging on and reorganizing around remainingpyramidal cells (51).
The molecular mechanisms that underlie the degenerative changes observed in the ETcerebellum are not clear. The array of cellular changes within the complex neuronal networkin cerebellar cortex have yet to be fully catalogued in ET; thus, further work with humanpostmortem tissue is needed. In addition, genes have not been identified for ET and notransgenic mice currently exist that provide an animal model for ET. The most widely usedanimal model for ET is based on the administration of neurotoxins (e.g. harmaline andibogaine) to rodents and other mammals. These toxins induce synchronous firing of inferiorolivary neurons at an 8 to 12 Hz frequency and sustained glutamatergic stimulation of PCs viatheir climbing fiber afferents (52–54), triggering a massive increase in intracellular calciumand an excitotoxic-mediated PC degeneration and cell death (55–57). However, this toxin-based animal model produces an acute, reversible action tremor. Most animals shake for amatter of hours and there is more severe destruction of cerebellar cortex in readily identifiablelinear bands. While these latter clinical and pathologic features clearly differ from those in ET,this model demonstrates the functional significance of the olivocerebellar system and apotential role for excitotoxic-mediated PC death in the generation of this type of tremor. It isintriguing that harmane (a tremor-producing, indole-alkaloid structurally similar to harmaline)
Erickson-Davis et al. Page 7
J Neuropathol Exp Neurol. Author manuscript; available in PMC 2011 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

has been found in the blood of some ET patients at levels up to 50% higher than age-matchedcontrols (58). While additional studies are needed to establish whether this exposure is ofetiologic importance in ET, we postulate that the PC loss documented in ET (whetherexcitotoxic or not) leads to a reorganization of the cerebellar interneuron network, withsprouting and/or accumulation of neighboring basket cell processes on surviving PC bodies.
Ultrastructural analysis revealed the presence of multiple, closely spaced sites of synapticcontact between the dense, multi-layered basket cell processes and PC soma in ET. Theneurophysiological abnormality in ET is not clear, but is thought to be the result of decreasedcerebellar PC inhibitory output. Our observations raise the possibility that the increased densityof basket cell processes identified in a significant number of ET cases contribute to thisinhibitory effect on PC output. Recent studies have demonstrated that climbing fiber responsesin PCs are modulated through cerebellar interneuron networks that include the basket cell(59). Basket cell output is also inhibited by the axon collaterals of PCs and excited by bothparallel fibers and collaterals of climbing fibers (60). Cerebellar interneurons are also coupledby dendro-somatic, dendro-dendritic and somato-somatic gap junctions, which may lead tosynchronization of firing in groups of organized neural networks (61). Specialized axo-axonicseptate junctions electrically couple basket fiber processes in the axon hillock region (62).Thus, the disease process in ET may cause an imbalance in the interactions between cerebellarafferents, PCs and their axon collaterals and the cerebellar interneuron network that includesbasket cells. Additional studies on other cerebellar neurons and their interactions in ET areclearly needed to understand the basis for tremor generation in this disorder.
Despite being the largest series of ET brains reported to date, the sample size was modest.Despite this limitation, we were able to detect significant case-control differences, indicatingthat the sample was sufficient. This study had several strengths. This was the first investigationof a relatively novel pathological finding in a large series of ET brains. We examined basketcell profiles both semiquantitatively and quantitatively and, furthermore, compared ET brainsagainst both non-diseased control brains and a group of diseased controls with a variety ofneurodegenerative pathologies.
In summary, the degenerative changes in PCs that characterize cerebellar ET are accompanied,in some cases, by a dense and tangled, or “hairy,” appearance of basket cell axonal plexusformations. This relatively novel finding was not observed to any degree in either diseasedcontrols or normal control brains. This finding provides initial evidence that structural changesin ET are not restricted to the PC but also involve their functional network. The presence ofhairy baskets is a heretofore uninvestigated phenomenon and further exploration is warrantedin order to understand its relationship to PC degeneration and its role in the pathogenesis ofET.
AcknowledgmentsFunding: R01 NS42859, P50 AG08702, and P01 AG07232 from the National Institutes of Health (Bethesda, MD);the Parkinson’s Disease Foundation (New York, NY); and the Arlene Bronstein Essential Tremor Research Fund(Columbia University).
We would like to thank Hong Yi (Emory University) for assistance with preparation of tissue for ultrastructural studies.
References1. Benito-Leon J, Louis ED. Essential tremor: Emerging views of a common disorder. Nat Clin Pract
Neurol 2006;2:666–78. quiz 662p following 691. [PubMed: 17117170]2. Louis ED. Essential tremor. Lancet Neurol 2005;4:100–10. [PubMed: 15664542]
Erickson-Davis et al. Page 8
J Neuropathol Exp Neurol. Author manuscript; available in PMC 2011 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

3. Dogu O, Sevim S, Camdeviren H, et al. Prevalence of essential tremor: Door-to-door neurologic examsin Mersin Province, Turkey. Neurology 2003;61:1804–6. [PubMed: 14694055]
4. Louis ED, Thawani SP, Andrews HF. Prevalence of essential tremor in a multiethnic, community-based study in northern Manhattan, New York, NY. Neuroepidemiology 2009;32:208–14. [PubMed:19169043]
5. Critchley M. Observations on essential (heredofamilial) tremor. Brain 1949;72:113–39. [PubMed:18136705]
6. Busenbark KL, Nash J, Nash S, et al. Is essential tremor benign? Neurology 1991;41:1982–3. [PubMed:1745359]
7. Louis ED, Benito-Leon J, Ottman R, et al. A population-based study of mortality in essential tremor.Neurology 2007;69:1982–9. [PubMed: 18025392]
8. Louis ED, Frucht SJ, Rios E. Intention tremor in essential tremor: Prevalence and association withdisease duration. Mov Disord 2009;24:626–7. [PubMed: 19185016]
9. Singer C, Sanchez-Ramos J, Weiner WJ. Gait abnormality in essential tremor. Mov Disord1994;9:193–6. [PubMed: 8196682]
10. Stolze H, Petersen G, Raethjen J, et al. The gait disorder of advanced essential tremor. Brain2001;124:2278–86. [PubMed: 11673328]
11. Parisi SL, Heroux ME, Culham EG, et al. Functional mobility and postural control in essential tremor.Arch Phys Med Rehabil 2006;87:1357–64. [PubMed: 17023246]
12. Koster B, Deuschl G, Lauk M, et al. Essential tremor and cerebellar dysfunction: Abnormal ballisticmovements. J Neurol Neurosurg Psychiatry 2002;73:400–405. [PubMed: 12235308]
13. Helmchen C, Hagenow A, Miesner J, et al. Eye movement abnormalities in essential tremor mayindicate cerebellar dysfunction. Brain 2003;126:1319–32. [PubMed: 12764054]
14. Dupuis MJ, Delwaide PJ, Boucquey D, et al. Homolateral disappearance of essential tremor aftercerebellar stroke. Mov Disord 1989;4:183–7. [PubMed: 2733709]
15. Schuurman PR, Bosch DA, Bossuyt PM, et al. A comparison of continuous thalamic stimulation andthalamotomy for suppression of severe tremor. N Engl J Med 2000;342:461–8. [PubMed: 10675426]
16. Benabid AL, Pollak P, Seigneuret E, et al. Chronic VIM thalamic stimulation in Parkinson’s disease,essential tremor and extra-pyramidal dyskinesias. Acta Neurochir Suppl (Wien) 1993;58:39–44.[PubMed: 8109299]
17. Bucher SF, Seelos KC, Dodel RC, et al. Activation mapping in essential tremor with functionalmagnetic resonance imaging. Ann Neurol 1997;41:32–40. [PubMed: 9005863]
18. Colebatch JG, Findley LJ, Frackowiak RS, et al. Preliminary report: Activation of the cerebellum inessential tremor. Lancet 1990;336:1028–30. [PubMed: 1977019]
19. Jenkins IH, Bain PG, Colebatch JG, et al. A positron emission tomography study of essential tremor:evidence for overactivity of cerebellar connections. Ann Neurol 1993;34:82–90. [PubMed: 8517685]
20. Louis ED, Shungu DC, Chan S, et al. Metabolic abnormality in the cerebellum in patients withessential tremor: A proton magnetic resonance spectroscopic imaging study. Neurosci Lett2002;333:17–20. [PubMed: 12401550]
21. Pagan FL, Butman JA, Dambrosia JM, et al. Evaluation of essential tremor with multi-voxel magneticresonance spectroscopy. Neurology 2003;60:1344–7. [PubMed: 12707440]
22. Shin DH, Han BS, Kim HS, et al. Diffusion tensor imaging in patients with essential tremor. AJNRAm J Neuroradiol 2008;29:151–3. [PubMed: 17921227]
23. Cerasa A, Messina D, Nicoletti G, et al. Cerebellar atrophy in essential tremor using an automatedsegmentation method. AJNR Am J Neuroradiol 2009;30:1240–3. [PubMed: 19342539]
24. Louis ED, Vonsattel JP. The emerging neuropathology of essential tremor. Mov Disord 2007;23:174–82. [PubMed: 17999421]
25. Zesiewicz TA, Elble R, Louis ED, et al. Practice parameter: therapies for essential tremor: Report ofthe Quality Standards Subcommittee of the American Academy of Neurology. Neurology2005;64:2008–20. [PubMed: 15972843]
26. Louis ED, Vonsattel JP, Honig LS, et al. Essential tremor associated with pathologic changes in thecerebellum. Arch Neurol 2006;63:1189–93. [PubMed: 16908751]
Erickson-Davis et al. Page 9
J Neuropathol Exp Neurol. Author manuscript; available in PMC 2011 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

27. Louis ED, Faust PL, Vonsattel JP, et al. Neuropathological changes in essential tremor: 33 casescompared with 21 controls. Brain 2007;130:3297–3307. [PubMed: 18025031]
28. Shill HA, Adler CH, Sabbagh MN, et al. Pathologic findings in prospectively ascertained essentialtremor subjects. Neurology 2008;70:1452–5. [PubMed: 18413570]
29. Axelrad JE, Louis ED, Honig LS, et al. Reduced purkinje cell number in essential tremor: Apostmortem study. Arch Neurol 2008;65:101–107. [PubMed: 18195146]
30. Yokoo H, Isoda K, Yamanouchi H, et al. Cerebellar basket cells of Creutzfeldt-Jakob disease:immunohistochemical and ultrastructural study. Pathol Int 2000;50:291–6. [PubMed: 10849314]
31. Uyematsu S. A study of some peculiar changes found in the oxons and dendrites of the Purkinje cells.Arch Neurol Psych 1921;5:231–69.
32. Louis ED, Borden S, Moskowitz CB. Essential tremor centralized brain repository: Diagnostic validityand clinical characteristics of a highly selected group of essential tremor cases. Mov Disord2005;20:1361–5. [PubMed: 16001407]
33. Louis ED, Vonsattel JP, Honig LS, et al. Neuropathologic findings in essential tremor. Neurology2006;66:1756–9. [PubMed: 16769958]
34. Louis ED, Faust PL, Vonsattel JP, et al. Torpedoes in Parkinson’s disease, Alzheimer’s disease,essential tremor, and control brains. Mov Disord 2009;24:1600–5. [PubMed: 19526585]
35. Louis ED, Ottman R, Ford B, et al. The Washington Heights-Inwood Genetic Study of EssentialTremor: Methodologic issues in essential-tremor research. Neuroepidemiology 1997;16:124–33.[PubMed: 9159767]
36. Louis ED, Honig LS, Vonsattel JP, et al. Essential tremor associated with focal nonnigral Lewybodies: a clinicopathologic study. Arch Neurol 2005;62:1004–7. [PubMed: 15956174]
37. Braak H, Braak E. Diagnostic criteria for neuropathologic assessment of Alzheimer’s disease.Neurobiol Aging 1997;18:S85–8. [PubMed: 9330992]
38. Braak H, Alafuzoff I, Arzberger T, et al. Staging of Alzheimer disease-associated neurofibrillarypathology using paraffin sections and immunocytochemistry. Acta Neuropathol 2006;112:389–404.[PubMed: 16906426]
39. Mirra SS. The CERAD neuropathology protocol and consensus recommendations for the postmortemdiagnosis of Alzheimer’s disease: a commentary. Neurobiol Aging 1997;18:S91–4. [PubMed:9330994]
40. The National Institute on Aging and Reagan Institute Working Group on Diagnostic Criteria for theNeuropathological Assessment of Alzheimer’s Disease 11: Consensus Recommendations for thePostmortem Diagnosis of Alzheimer’s disease. Neurobiol Aging 1997;18(S4):S1–2. [PubMed:9330978]
41. Louis ED, Yi H, Erickson-Davis C, et al. Structural study of Purkinje cell axonal torpedoes in essentialtremor. Neurosci Lett 2009;450:287–91. [PubMed: 19047012]
42. Louis ED, Vonsattel JPG, Honig LS, et al. Older onset essential tremor: More rapid progression andmore degenerative pathology. Mov Disord. 2009 (In Press).
43. Deuschl G, Wenzelburger R, Loffler K, et al. Essential tremor and cerebellar dysfunction clinical andkinematic analysis of intention tremor. Brain 2000;123 ( Pt 8):1568–80. [PubMed: 10908187]
44. Leclerc N, Gravel C, Plioplys A, et al. Basket cell development in the normal and hypothyroid ratcerebellar cortex studied with a monoclonal anti-neurofilament antibody. Can J Biochem Cell Biol1985;63:564–76. [PubMed: 2412672]
45. Perry TL, Kish SJ, Hinton D, et al. Neurochemical abnormalities in a patient with ataxia-telangiectasia. Neurology 1984;34:187–91. [PubMed: 6320055]
46. Iwata M, Hirano A, French JH. Degeneration of the cerebellar system in X-chromosome-linked coppermalabsorption. Ann Neurol 1979;5:542–9. [PubMed: 475349]
47. Tiller-Borcich JK, Urich H. Abnormal arborizations of Purkinje cell dendrites in Creutzfeldt-Jakobdisease: A manifestation of neuronal plasticity? J Neurol Neurosurg Psychiatry 1986;49:581–4.[PubMed: 3519873]
48. Robitaille Y, Schut L, Kish SJ. Structural and immunocytochemical features of olivopontocerebellaratrophy caused by the spinocerebellar ataxia type 1 (SCA-1) mutation define a unique phenotype.Acta Neuropathol 1995;90:572–81. [PubMed: 8615077]
Erickson-Davis et al. Page 10
J Neuropathol Exp Neurol. Author manuscript; available in PMC 2011 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

49. de Lanerolle NC, Kim JH, Robbins RJ, et al. Hippocampal interneuron loss and plasticity in humantemporal lobe epilepsy. Brain Res 1989;495:387–95. [PubMed: 2569920]
50. Buckmaster PS, Jongen-Relo AL. Highly specific neuron loss preserves lateral inhibitory circuits inthe dentate gyrus of kainite-induced epileptic rats. J Neurosci 1999;19:9519–29. [PubMed:10531454]
51. Arellano JI, Munoz A, Ballesteros-Yanez I, et al. Histopathology and reorganization of chandeliercells in the human epileptic sclerotic hippocampus. Brain 2004;127(1):45–64. [PubMed: 14534159]
52. De Montigny C, Lamarre Y. Rhythmic Activity Induced by harmaline in the olivocerebello-bulbarsystem of the cat. Brain Res 1973;53:81–95. [PubMed: 4697252]
53. Llinás R, Volkind RA. The olivo-cerebellar system: Functional properties as revealed by bulbarsystem of the cat. Brain Res 1973;63:430–4. [PubMed: 4764311]
54. Llinás R, Mühlethaler M. Electrophysiology of guinea-pig cerebellar nuclear cells in the in vitro brainstem-cerebellar preparation. J Physiol 1988;404:241–58. [PubMed: 2855348]
55. Trump BF, Berezesky IK. Calcium-mediated cell injury and cell death. FASEB J 1995;9:219–28.[PubMed: 7781924]
56. Slemmer JE, De Zeeuw CI, Weber JT. Don’t get too excited: Mechanisms of glutamate-mediatedPurkinje cell death. Prog Brain Res 2005;148:367–90. [PubMed: 15661204]
57. Weber JT, De Zeeuw CI, Linden DJ, et al. Long-term depression of climbing fiber-evoked calciumtransients in Purkinje cell dendrites. Proc Natl Acad Sci USA 2003;100:2878–83. [PubMed:12601151]
58. Louis ED, Jiang W, Pellegrino KM, et al. Elevated blood harmane (1-methyl-9H-pyrido[3,4-b]indole)concentrations in essential tremor. Neurotoxicology 2008;29:294–300. [PubMed: 18242711]
59. Barmack NH, Yakhnitsa V. Functions of interneurons in mouse cerebellum. J Neurosci2008;28:1140–52. [PubMed: 18234892]
60. O’Donoghue DL, King JS, Bishop GA. Physiological and anatomical studies of the interactionsbetween Purkinje cells and basket cells in the cat’s cerebellar cortex: Evidence for a unitaryrelationship. J Neurosci 1989;9:2141–50. [PubMed: 2723768]
61. Sotelo C, Llinás R. Specialized membrane junctions between neurons in the vertebrate cerebellarcortex. J Cell Biol 1972;53:271–89. [PubMed: 4537207]
62. Gobel S. Axo-axonic septate junctions in the basket formations of the cat cerebellar cortex. J CellBiol 1971;51:328–33. [PubMed: 5165176]
Erickson-Davis et al. Page 11
J Neuropathol Exp Neurol. Author manuscript; available in PMC 2011 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 1.Semiquantitative rating of basket cell processes. (A–D) A rating scale (from 0 to 3) of theappearance of the basket cell plexus surrounding extant Purkinje cell bodies was determinedin a blinded analysis of Bielschowsky preparations of neocerebellum sections. (A) Score = 0(few, or no discernible processes); (B) Score = 1 (sparse number of processes); (C) score = 2(moderate number of processes); (D) score = 3 (dense tangle of processes, i.e. hairy basket).
Erickson-Davis et al. Page 12
J Neuropathol Exp Neurol. Author manuscript; available in PMC 2011 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 2.Immunohistochemical analysis of basket cell processes. (A, B) Vibratome sections (100-μm-thick) immunostained with monoclonal antibody SMI-31 demonstrate prominence ofphosphorylated neurofilaments in basket cell processes (long arrows). (A) A dense and tangled,or “hairy,” appearance of basket cell axonal plexus formation =in a section from a cerebellaressential tremor case with a hairy basket rating of 3. (B) A control case with a rating of 0.Torpedoes (short arrows) are also seen.
Erickson-Davis et al. Page 13
J Neuropathol Exp Neurol. Author manuscript; available in PMC 2011 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
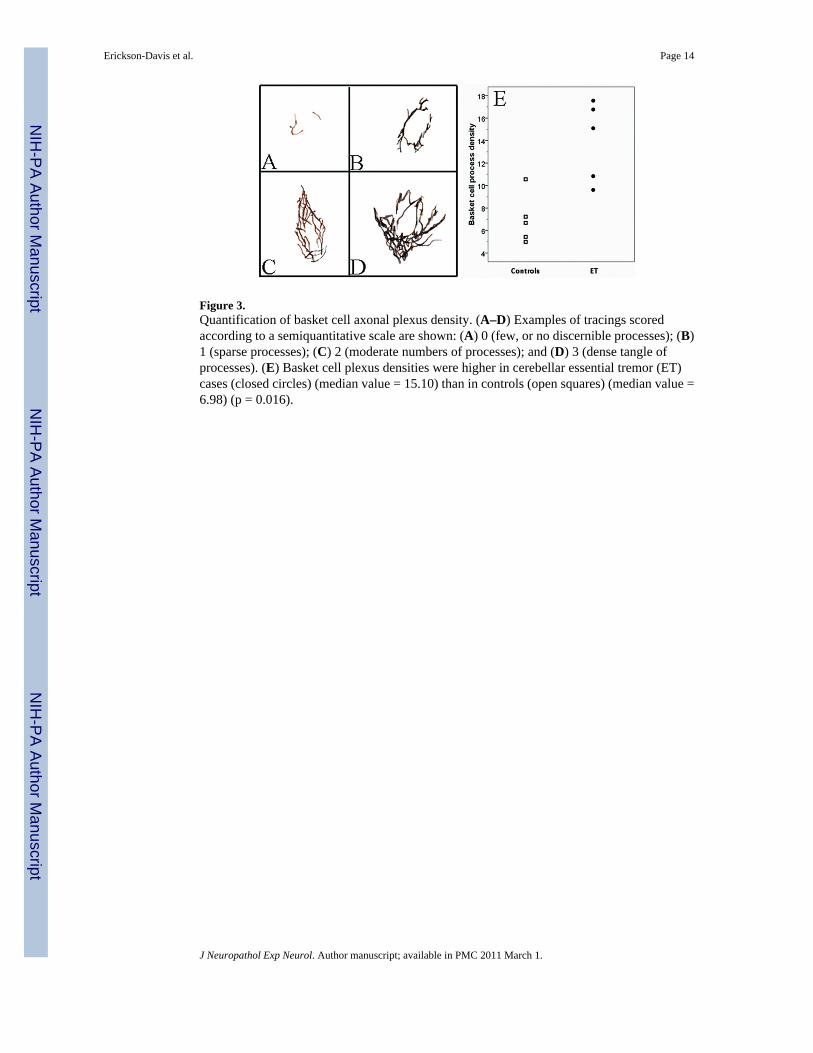
Figure 3.Quantification of basket cell axonal plexus density. (A–D) Examples of tracings scoredaccording to a semiquantitative scale are shown: (A) 0 (few, or no discernible processes); (B)1 (sparse processes); (C) 2 (moderate numbers of processes); and (D) 3 (dense tangle ofprocesses). (E) Basket cell plexus densities were higher in cerebellar essential tremor (ET)cases (closed circles) (median value = 15.10) than in controls (open squares) (median value =6.98) (p = 0.016).
Erickson-Davis et al. Page 14
J Neuropathol Exp Neurol. Author manuscript; available in PMC 2011 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 4.Ultrastructure of neurofilament-rich basket cell processes in cerebellar essential tremor (ET)and control cases. (A) The dense plexus of basket cell processes (arrows) adjacent to thePurkinje cell soma (P) in a cerebellar ET case contains neurofilaments and mitochondria. (B)Only 1 to 2 layer(s) of basket cell processes (arrows) are seen adjacent to a Purkinje cell soma(P) in a control case. (C) Higher magnification of basket cell axonal plexus in (A) shows manylayers of the basket cell formation (arrows). (D) A dense nest of basket cell processes (arrows)surrounds another Purkinje cell in an ET case. (E) Intertwining of basket cell processes aroundone another in an ET case. Some processes are longitudinal to the plane of section (long arrows);
Erickson-Davis et al. Page 15
J Neuropathol Exp Neurol. Author manuscript; available in PMC 2011 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

others have been cut transversely (short arrows). Magnifications: A, B, 10,000×; C, 20,000×;D, 25,000×; E, 30,000×.
Erickson-Davis et al. Page 16
J Neuropathol Exp Neurol. Author manuscript; available in PMC 2011 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 5.Ultrastructural morphology of basket cell-Purkinje cell (P) contacts in essential tremor cases.(A–C) Multiple, closely spaced sites of synaptic contact (arrows) between a neurofilament-rich basket fiber and a Purkinje cell soma. Post-synaptic densities are apparent in (A) and (B)(arrows). The basket fiber in (C) shows several vesicle-lined contacts with the Purkinje cell(arrows). (D) Numerous processes converging on the initial segment (axon hillock) of aPurkinje cell (P-axh); at least one of these is a neurofilament-rich basket cell axon (large arrow).(d′) In a higher magnification of (D), the Purkinje cell contact shows 2 distinct clusters ofvesicles (arrows). Magnifications: A, B, d’, 40,000×; C, 50,000×; D, 20,000×.
Erickson-Davis et al. Page 17
J Neuropathol Exp Neurol. Author manuscript; available in PMC 2011 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 6.Correlations of hairy basket scores in cerebellar essential tremor (ET) and non-diseased controlcases. (A–C) Hairy basket score correlated with number of torpedoes (A) (Spearman’s r = 0.49,p < 0.001), inversely with the number of Purkinje cells (B) (Spearman’s r = −0.27, p = 0.06),and degeneration index (C) (Spearman’s r = 0.48, p = 0.001).
Erickson-Davis et al. Page 18
J Neuropathol Exp Neurol. Author manuscript; available in PMC 2011 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Erickson-Davis et al. Page 19
Tabl
e 1
Dem
ogra
phic
and
Clin
ical
Cha
ract
eris
tics a
nd D
iagn
oses
of C
ases
NC
ereb
ella
r E
TL
BV
ET
AD
PD/D
LB
DPS
PD
ysto
nia
Non
- dis
ease
d co
ntro
lsD
isea
sed
cont
rols
¥A
ll C
ontr
ols¥
325
2015
121
2148
69
Age
(yea
rs)
83.6
± 7
.686
.8 ±
4.7
82.9
± 5
.278
.1 ±
6.7
73.4
± 7
.2**
6877
.3 ±
13.
678
.7 ±
7.3
**78
.3 ±
9.6
**
Fem
ale
Gen
der
20 (6
2.5%
)2
(40.
0%)
12 (6
0.0%
)6
(40.
0%)
5 (4
1.7%
)1
(100
%)
8 (3
8.1%
)24
(50.
0%)
32 (4
6.4%
)
PMI (
hour
s)4.
7 ±
4.1
7.7
± 2.
87.
7 ±
7.3
7.7
± 4.
88.
0 ±
7.3
7.1
8.8
± 8.
47.
8 ±
6.5*
8.1
± 7.
0**
Bra
in w
eigh
t (gm
)12
11 ±
138
1184
± 1
3110
97 ±
119
1128
±272
1164
± 1
1016
3212
84 ±
182
1135
± 1
9211
80 ±
200
CER
AD
scor
e0.
9 ±
1.1
1.2
± 1.
33.
0 ±
0.0*
**1.
5 ±
1.1
0.4
± 0.
70
0.2
± 0.
41.
8 ±
1.3*
*1.
3 ±
1.3
Bra
ak A
D sc
ore
2.0
± 1.
22.
2 ±
0.8
5.8
± 0.
4***
1.3
± 1.
01.
2 ±
0.9
10.
6 ±
0.9*
**3.
2 ±
2.4*
*2.
4 ±
0.4
ET =
inte
ntio
n tre
mor
; LB
VET
= L
ewy
body
var
iant
of E
T; A
D =
Alz
heim
er d
isea
se; P
D =
Par
kins
on d
isea
se; D
LBD
= d
iffus
e Le
wy
body
dis
ease
; PSP
= p
rogr
essi
ve su
pran
ucle
ar p
alsy
.
¥ Dis
ease
d co
ntro
ls =
AD
+ P
D/D
LBD
+ P
SP +
dys
toni
a. A
ll co
ntro
ls =
dis
ease
d co
ntro
ls a
nd n
on-d
isea
sed
cont
rols
.
PMI =
pos
tmor
tem
inte
rval
; Val
ues a
re e
ither
mea
n ±
SD o
r num
ber (
perc
enta
ge).
* p <
0.05
,
**p<
0.0
1,
*** p<
0.00
1 co
mpa
red
to c
ereb
ella
r ET
(AN
OV
A w
ith T
ukey
’s p
ost-h
oc c
ompa
rison
).
J Neuropathol Exp Neurol. Author manuscript; available in PMC 2011 March 1.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Erickson-Davis et al. Page 20
Tabl
e 2
Hai
ry B
aske
t Cat
egor
ies b
y D
iagn
osis
Cer
ebel
lar
ET
(n =
32)
LB
VE
T(n
= 5
)A
D(n
= 2
0)PD
/DL
BD
(n =
15)
PSP
(n =
12)
Dys
toni
a(n
= 1)
Dis
ease
d co
ntro
ls(n
= 4
8)N
on- d
isea
sed
Con
trol
s(n
= 2
1)A
ll C
ontr
ols
(69)
Hai
ry B
aske
t Cat
egor
y
1 (lo
w)
13 (4
0.6%
)2
(40.
0%)
15 (7
5.0%
)10
(66.
7%)
9 (7
5.0%
)1
(100
%)
35 (7
2.9%
)14
(66.
7%)
49 (7
1.0%
)
2 (I
nter
med
iate
)11
(34.
4%)
3 (6
0.0%
)4
(20.
0%)
4 (2
6.7%
)3
(25.
0%)
0 (0
.0%
)11
(22.
9%)
6 (2
8.6%
)17
(24.
6%)
3 (h
igh)
8 (2
5.0%
)0
(0.0
%)
1 (5
.0%
)1
(6.7
%)
0 (0
.0%
)0
(0.0
%)
2 (4
.2%
)1
(4.8
%)
3 (4
.3%
)
P va
lue*
Not
app
licab
le0.
510.
010.
068
0.02
^0.
001
0.03
0.00
1
ET =
ess
entia
l tre
mor
; LB
VET
= L
ewy
body
var
iant
of E
T; A
D =
Alz
heim
er d
isea
se; P
D =
Par
kins
on d
isea
se, D
LBD
= d
iffus
e Le
wy
body
dis
ease
, PSP
= p
rogr
essi
ve su
pran
ucle
ar p
alsy
.
Hai
ry B
aske
t Cat
egor
y: 1
(low
, hai
ry b
aske
t rat
ing
= 0
– 1.
5), 2
(int
erm
edia
te, h
airy
bas
ket r
atin
g =
2 –
2.5)
, 3 (h
igh,
hai
ry b
aske
t rat
ing
= 3)
. Val
ues a
re n
umbe
r of c
ases
(per
cent
age)
.
* com
pare
d to
cer
ebel
lar E
T (M
ante
l-Hae
nsze
l chi
-squ
are
test
).
^ Onl
y on
e ca
se; p
val
ue n
ot c
ompu
ted.
J Neuropathol Exp Neurol. Author manuscript; available in PMC 2011 March 1.




















