
Isozyme relative mobility (R m ) changes related to leaf position; apparently smoothR m trends and...
14
Biochemical Genetics, Vol. 23, Nos. 9/10, 1985 Isozyme Relative Mobility (Rm) Changes Related to Leaf Position; Apparently Smooth R m Trends and Some Implications Hugh Tyson, ~ Mary Ann Fieldes, 1 Cindy Cheung, I and Jean Starobin I Received 14 Sept. 1984--FinalH Mar. 1985 Relative mobilities (Rm's) of peroxidase and acid phosphatase isozymes were examined in leaves of flax (Linum usitatissimum L.). The leaves were sampled from four equidistantly spaced positions from main stem base to apex in various genotypes. Rm's for the two slowest-migrating isozymes of each enzyme system changed in a simple, coherent fashion in leaves from stem bases toward apices. The R m trends up the stem seen in two highly branched flax types were somewhat different from those in two sparsely branched types. The coherent Rm trends in the four types, suggesting a smooth continuum and a potentially large number of slightly different forms of these isozymes, are discussed in relation to other data for such R m trends. In the study reported here, both enzyme systems behaved similarly. This fact and the simple Mendelian genetical system with no codominance controlling Rm's in flax peroxidases and acid phosphatases suggest posttranscriptional or posttranslational modifications as plausible mechanisms underlying the numerous, presumably small molecular changes generating the small, con- sistent changes in Rm'S. KEY WORDS: isozyme R,,; leaf position and Rm. INTRODUCTION Electrophoretic variation in isozymes of peroxidase, acid phosphatase, and esterase has been examined in genotypes and genotrophs of flax (Linum The financial assistance of the Natural Sciences and Engineering Council of Canada is acknowledged with thanks. Biology Department, McGill University,Montreal, Quebec, Canada H3A 1B1. 641 0006 2928/85/100~0641504.50/0 © 1985 Plenum Publishing Corporation
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Isozyme relative mobility (R m ) changes related to leaf position; apparently smoothR m trends and...

Biochemical Genetics, Vol. 23, Nos. 9/10, 1985
Isozyme Relative Mobility (Rm) Changes Related to Leaf Position; Apparently Smooth R m Trends and Some Implications
Hugh Tyson, ~ Mary Ann Fieldes, 1 Cindy Cheung, I and Jean Starobin I
Received 14 Sept. 1984--Final H Mar. 1985
Relative mobilities (Rm's) o f peroxidase and acid phosphatase isozymes were examined in leaves of flax (Linum usitatissimum L.). The leaves were sampled from four equidistantly spaced positions from main stem base to apex in various genotypes. Rm's for the two slowest-migrating isozymes of each enzyme system changed in a simple, coherent fashion in leaves from stem bases toward apices. The R m trends up the stem seen in two highly branched flax types were somewhat different from those in two sparsely branched types. The coherent Rm trends in the four types, suggesting a smooth continuum and a potentially large number of slightly different forms of these isozymes, are discussed in relation to other data for such R m trends. In the study reported here, both enzyme systems behaved similarly. This fact and the simple Mendelian genetical system with no codominance controlling Rm's in flax peroxidases and acid phosphatases suggest posttranscriptional or posttranslational modifications as plausible mechanisms underlying the numerous, presumably small molecular changes generating the small, con- sistent changes in Rm'S.
KEY WORDS: isozyme R,,; leaf position and Rm.
INTRODUCTION
Electrophoretic variation in isozymes of peroxidase, acid phosphatase, and esterase has been examined in genotypes and genotrophs of flax (Linum
The financial assistance of the Natural Sciences and Engineering Council of Canada is acknowledged with thanks.
Biology Department, McGill University, Montreal, Quebec, Canada H3A 1B 1.
641 0006 2928/85/100~0641504.50/0 © 1985 Plenum Publishing Corporation

642 Tyson, Fieldes, Cheung, and Starobin
usi tat iss imum L.). This species is a complete inbreeder. In these previous studies (Fieldes, and Tyson, 1972, 1984), samples for assay of a number of enzyme systems were prepared from main stem tissue, using the complete stem (minus leaves) from base to apex. Comparisons were made between different genotypes to assess the number of peroxidase, acid phosphatase, esterase, etc, isozymes present and measure their individual relative mobilities (R,,) from electrophoretic separations. Four anionic isozyme bands have been observed for peroxidases and for acid phosphatases in the flax genotypes so far examined. Both peroxidases and acid phosphatases are asparagine-N-acetyl- glucosamine-type glycoproteins (Welinder, 1976; Hemmings et al, 1981). Flax peroxidases are certainly monomeric.
Differences between the R~'s of purified peroxidase isozymes have been established in comparisons of genotypes and "genotrophs" of flax (Fieldes et al., 1976a, 1977). Flax genotrophs (Durrant, 1971), it should be explained briefly, display heritable changes induced within a single completely inbreed- ing genotype, i.e., a group of genetically identical plants, by a single generation's growth in contrasting environmental conditions, specifically high fertiliser levels of NPK versus NK. Progenies of the NPK and NK treatment subgroups which are then grown in uniform environmental conditions reflect induced changes due to NPK or NK, for many subsequent generations. The changes are not explainable through selection, and the flax results have been paralleled by similar findings with tobacco Nicotiana rustica (see e,g., Perkins et al., 1971). The origin of genotrophs thus contrasts with that of genotypes (cultivars) produced by artificial selection.
In comparing two flax genotrophs (genotrophs L and S, Durrant, 1971) for peroxidase R,,'s it has been shown that (a) the four anionic isozymes present in the main stem tissue of each genotroph have distinctly increasing Rm'S from the slowest (isozyme 1) to the fastest migrating (isozyme 4). If one compares, for example, the slowest-migrating stem isozyme in L with that in S, there is a smaller, but still consistent, clear-cut R,~ difference. It has been shown that (b) this small difference, in the same direction, is present for each of the other three peroxidase isozymes; genotroph (L) has a set of four isozymes, all of which run slightly faster than the corresponding four from the other (S) genotroph (Fieldes and Tyson, 1972). The R m shift remains after isozyme purification (Fieldes et al., 1977).
The "correspondence" between peroxidase isozymes has been confirmed by crosses between different parental genotypes or between the two geno- trophs. In the F1 hybrids between the L and the S genotrophs, the slowest- moving isozyme 1, for example, has a n R m which is identical to that of the faster version of isozyme 1 found in L. The L × S F1 hybrid thus has an isozyme 1 displaying complete dominance for the faster (L) version, with no codominance. The single F1 band for isozyme 1 lies, therefore, on the upper R,, boundary of the range displayed by L and S for the slowest-migrating

Isozyme Rm Dependence on Leaf Position 643
peroxidase. This same complete dominance in the L direction occurs for each of the other three peroxidase bands, so that the F1 hybrid contains four isozymes, each having the same Rm as the faster (L) parental version. Control of this Rm difference for at least the first two isozymes is through a simple Mendelian system with two alleles at one locus; the allele for the faster- migrating (L) version is dominant to the S allele, and the expected 3:1 segregation of fast (L) to slow (S) is found in the F 2 generation from the L x S cross for isozymes 1 and 2 (Tyson et al., 1978). Correspondence of two isozymes, each the slowest migrating in the respective sample extracts, implies then that the Rm variation between these two is clearly less than that between either of them and the next slowest bands in the samples. If the samples represent genotypes, crosses between them can confirm correspondence, whether codominance is present or absent.
For acid phosphatases and esterases, the same undirectional Rm shift (faster Rm in L) for all isozymes is observed. In a total of 10 isozyme bands from the three enzyme systems, the L versions always run slightly faster than the corresponding S forms. This undirectional Rm shift raises the possibility of a single mechanism affecting R m (and hence molecular weight and/or net charge) in several enzyme systems.
If peroxidase Rm were examined across a range of genotypes, variation in R,, would be expected for one or more isozymes. The exact Rm of, say, the slowest-moving isozyme (1) may differ from one genotype to the next. The correspondence between all slowest (isozyme 1) bands can again be confirmed by crosses; this process can be extended to the next slowest isozyme bands, and so on. Correspondence for specific isozyme pairs can then be detailed for a specific pair of genotypes.
An important proviso for comparisons between genotypes is that such variation between Rm's of corresponding isozymes should not be confused (i.e., confounded) with other sources of Rm variation. Different developmental stages and/or different tissues (collectively "differences within plants") or, alternatively, the effects of external environmental influences, are factors which have to be assessed and controlled, to avoid spurious Rm differences.
Studies to disentangle differences between plants of different genotypes from differences within plants and from the effects of other factors affecting R m have been carried out in flax (Fieldes et al., 1976b). It has been shown that there are small, but discernible, Rm changes from the base of the main stem to its apex for a given isozyme. There are also small Rm changes which can be detected over the period of vegetative growth (Fieldes and Tyson, 1973). External environmental influences such as temperature and light conditions can also affect R m in flax (Tyson, Fieldes, and Cheung, unpublished). Elsewhere, developmental effects o n Rm's have been detected in Petunia (Van den Berg, 1983), and temperature (or greenhouse) conditions have been shown to affect R m data from populations of Si lene pratens is (Mastonbroek et

644 Tyson, Fieldes, Cheung, and Starobin
al., 1984). Thus, isozyme comparisons using a range of genotypes require careful planning to eliminate possible confusion from factors such as these.
Although such factors influencing Rm's clearly have to be checked when looking at the genetic control of Rm variation, the fact that R m appears to vary with stage of development and environment can supply other information and can be turned to advantage. A given isozyme's polypeptide moiety is the product of at least one specific structural gene. In the context of an array of four isozymes detectable in flax main stem tissue, one would argue for four responsible structural genes. If one flax genotype differred from another in that the R m of each of these four isozymes was slightly faster, the assumption would be that it possessed four slightly different structural genes. This reasoning has been extended to cope with R m differences from one develop- mental stage or one tissue to another, for example. It does, however, lead to an eventual dilemma, namely, that for every developmental/tissue variant an underlying structural gene variant is required.
The R m changes found for a given peroxidase isozyme (e.g., isozyme 1) from the base of the stem compared to the apex are initially explainable through "switching on" different peroxidase genes between the base and the apex. If, however, data from intervening zones of the stem suggest the emergence of a smooth trend in Rm values, one may speculate that continued finer subdivision of the stem length, coupled with R,~ measurement at each point, could produce Ru values changing in an essentially continuous fashion over the stem length. The argument that every change in Rm represents the output of a new structural gene and the shutdown of all others now becomes difficult to sustain, since it leads to the implausible requirement for a virtually infinite number of responsible structural genes.
A hint that just such smooth R m trends were occurring along the main stem tissue from base to apex of the flax main stem has been referred to above. The aim of the work described here was to see whether, in leaves sampled at four points along the main stem length, a smooth trend could be detected for peroxidases and acid phosphatases in different flax genotypes and genotrophs grown at two contrasting temperatures. Samples of a few leaves from a plant (flax has numerous small leaves) are nondestructive, and extraction is simpler and easier than for stem tissue. If smooth trends, as in stem tissue, were detectable, they would again lead to considerations of the repertoire of genes responsible and the mechanisms which might underlie a seemingly continuous range of small modifications in the enzyme molecule.
MATERIALS AND METHODS
Plants of two flax genotypes and two flax genotrophs were all grown at two different temperatures, namely, 15 and 30°C. All plants were grown in two

lsozyme Rm Dependence on Leaf Position 645
growth chambers; the 15°C environment was maintained in one growth chamber, while the second chamber was set to 30°C. There were duplicates of all plants within the 15°C chamber and within the 30°C chamber. Lighting duration (18-hr day length) and intensity were identical in both chambers; all plants were raised in vermiculite to which nutrient solution (modified from Murashige and Skoog, 1962) was added.
The two flax genotypes (cultivars R = Royal and M = Mandarin) were, respectively, linseed and fiber types, produced by conventional selection procedures and used as commercial cultivars. Linseed types are typically highly branched, especially from the base of the main stem, whereas fiber types are typically unbranched but tall. The other two were genotrophs generated within the commercial cultivar Stormont cirrus by Durrant 1971. In this case, the two genotrophs (L = large and S = small) displayed the induced heritable effects of one generation of soil-supplied inorganic nutrient treatments. This phenomenon has been extensively documented by Durrant 1971 and others elsewhere. Essentially it involves induced heritable changes in a wide range of characters, including isozyme R,,, resulting from growing a genetically homogeneous seed stock in high levels of NPK on the one hand compared to NK on the other. Selection was absent and does not explain the results. Progenies from random sampling of the treatment generation were grown in completely uniform environmental conditions, displaying heritable changes due to NPK or NK. In terms of morphological appearance, L resembled R (pronounced branching and side shooting), while S mimicked M (sparse branching and side shooting). R, M, L, and S are referred to jointly in what follows as "types."
At 47 days after germination leaves were sampled from four equidis- tantly spaced positions up the main stems. Simple leaf squashes supplied measureable quantities of isozymes of both enzyme systems. The leaf positions are referred to as bottom, mid-2, mid-l, and top. Leaf samples were frozen at - 17°C. Extracts were prepared by grinding the samples in 0.1 M, pH 8.0, phosphate buffer. Two peroxidase and four acid phosphatase isozymes were detected in all leaf samples.
Electrophoresis was carried out exactly as detailed by Fieldes and Tyson (1984). Electrophoresis buffer at 0.37 M ensured adequate control of pH throughout the procedure. Extract/isozyme concentration has no effect on R,, (Fieldes and Tyson, 1972). The visualization of peroxidases and acid phospha- tases was as outlined by Fieldes et at. (1984b), so that isozymes from both systems could be scanned from the same gels. Electrophoresis gels were scanned spectrophotometrically. Recorded optical density (OD) scans were measured to obtain the migration distances for each of the bands. These were converted to relative mobilities (Rm's) in relation to the bromophenol blue boundary distance for the particular gel. The slowest two isozymes (1 and 2)

646 Tyson, Fieldes, Cheung, and Starobin
in each enzyme system were examined in detail. Duplication of the electropho- resis separations means that, in all, there were four values for each type, temperature, isozyme, and leaf position combination, for a total of 512 values. These were examined through analyses of variance (Sokal and Rohlf, 1981). The error term, against which all main factors and their interactions were tested, was generated by summing the 128 within-combination sums of squares. The possibility of interactions between the main effects, particularly between the genotypes and the leaf positions and/or the temperatures was of primary concern. The single degree of freedom plant-type comparisons used were (a) the difference between morphological types--highly branched types R and L compared to sparsely branched M and S; (b) the difference between genetic backgrounds--selected types R and M compared to induced types L and S; and (c) the interaction between morphological types and genetic background. The position component of the analysis looked for linear, quadratic, and cubic changes over the leaf positions, and the isozyme component examined the differences between the two isozymes within each enzyme system plus the (arbitrary) difference between the peroxidase and the acid phosphatase isozyme means.
RESULTS
The Rm data are summarized in Table I. Analyses (summarized in Table II) show that there are significant differences among types, positions, and isozymes and between the two temperatures used. Each of these four factors, of course, contains Rm values averaged over all the other factors. Overall conclusions are, therefore, dependent on interactions between them. Differ- ences between isozyme 1 and isozyme 2 within both enzyme systems are significant, and the acid phosphatases have a faster mean Rm than the peroxidases. The trend in average Rm from base to apex is not linear.
For genotypes R and M, the mean R m across all isozymes is faster (8.9%) in the sparsely branched M compared to the highly branched R. For the L and S genotrophs, the highly branched L has the faster (6.2%) mean Rm. More importantly, there are differences for the mean Rm values among the four leaf positions sampled. From base to apex the principal Rm difference is between the mean of the bottom two and the mean of the top two positions. There is, however, a significant interaction between leaf positions and types. For R and M the Rm difference is largest in leaves at the stem base; for L and S it is largest along the midportion of the stem, while at the apex a reversal in R,~ direction toward S occurs. There is also a temperature effect averaged over all isozymes, positions, and types; the average Rm at 30°C is significantly faster than at 15°C, but two of the three first-order interactions with temperature are significant.

Table I. Mean R,, Values for Types, Isozymes, and Positions °
(A)
R M
PI P2 AP1 AP2 P1 P2 AP1 AP2
Top 0.0859 0.2130 0.1573 0.3718 0.0837 0 .2231 0.1775 0.4002 Mid-1 0.0859 0.2176 0.1467 0.3619 0.0812 0.2345 0.1688 0.3921 Mid-2 0.0893 0.2304 0.1619 0.3787 0.0950 0 .2391 0.1746 0.4030 Bottom 0.0718 0.2234 0.1520 0 .3801 0.1045 0.2504 0.1769 0.4194
(B)
L S
P1 P2 AP1 AP2 P1 P2 AP1 AP2
Top 0.0714 0.1965 0.1465 0.3757 0.0845 0.2112 0 .1581 0.3760 Mid-1 0.0758 0 .2131 0.1729 0 .4001 0 .0751 0.1922 0.1337 0.3602 Mid-2 0.0836 0.2305 0.1669 0.3979 0.0744 0.2093 0.1496 0.3656 Bottom 0.0844 0.2267 0.1627 0 .3911 0.0700 0.2022 0.1543 0.3809
(c)
Type
R M L S
Top 0.2070 0 .2211 0.1975 0.2074 Mid-I 0.2030 0 .2191 0.2155 0.1909 Mid-2 0.2150 0.2279 0.2197 0.1997 Bottom 0.2068 0.2378 0.2162 0.2019
(D)
Position 15°C 30°C
Top 0.2015 0.2150 Mid-1 0.2017 0.2123 Mid-2 0.2086 0.2226 Bottom 0.2025 0.2288
(E)
Isozyme 15°C 30°C
P1 0.0782 0.0864 P2 0.2112 0.2279 AP1 0.1551 0.1649 AP2 0.3698 0.3995
aR, Royal; M, Mandarin; L, large; S, small; PI, peroxidase isozyme 1; P2, peroxidase isozyme 2; AP1, acid phosphatase isozyme 1; AP2, acid phosphatase isozyme 2.

648
Source
Tyson, Fieldes, Cheung, and Starobin
Table II. Analysis of Variance of Rm Data
dF Mean squares F
Total 511 Types (G) 3 0;.01595881 28.65**
R - M + L - S (a) 1 0.00120873 2.17 R + M - L - S (b) 1 0.01601906 28.76** R - M - L + S (c) 1 0.03064862 55.03**
Positions (P) 3 0.00276612 4.97** Linear (x) 1 0.006079;31 10.91"* Quadratic (y) 1 0.00006032 < 1 Cubic (z) 1 0.00215873 3.88**
Isozymes (I) 3 2.10764545 3784.10"* Temperatures (T) 1 0.03313736 59.50** First-order interactions
G x P 9 0.00171122 3.07* (a) x (y) 1 0.00644288 11.57"* (c) x (x) 1 0.00530164 9.52** (All other G x P interaction components not significant)
G x T 3 0.00105151 1.89 G x I 9 0.00104072 1.87 P x T 3 0.00154245 2.77* P x I 9 0.00039952 <1 T x I 3 0.00306336 5.50*
Higher-order interactions 81 0.00053477 < 1
Error 384 0.00055698
* Significant at probability 0.05 ** Significant at probability 0.01.
The detailed breakdown (Table [I) of the type by position interaction shows two significant components. The first concerns the comparison between morphological types (highly versus sparsely branched) and the shape of the RF~ changes with the different leaf positions. In terms of Rm shifts, the main stem organization in the highly branched types (R and L) appears distinc- tively different from that in the sparsely branched types (M and S). These differences are shown in Fig. 1. The second interaction component reflects the reversal in mean R,, between L and S apices compared to the consistent R and M difference.
The two significant first-order interactions involving temperature show that the size of the temperature effect varies (a) with position, being much larger at the base than any other position, and (b) with isozyme, being much larger for acid phosphatase 2 than for acid phosphatase 1. This can be seen from the data summary in Table [, D and E.
The second- and higher-order interactions among the main factors here (accounting for a mean square of 0.00053477 with 81 degrees of freedom)

Isozyme R., Dependence on Leaf Position 649
~2400
G2300
w
N O
G2200
O
< G2100
w > 0 E
Z < ~2000
G1900
G1800
M
i "ilSL
BOTTOM MID2 MID1 TOP
LEAF POSITION ON MAIN STEM
Fig. 1. Relative mobilities for leaf samples from stem base to apex. Solid lines represent R,~ changes for the individual plant types. Dashed lines represent R,, behavior in highly branched (R and L) versus sparsely branched (M and S) types,
have not been dissected in detail since the mean square for these remaining interactions is not significant and is extremely small in relation to those for main effects and their first-order interactions, as can be seen from Table I. A check of the second-order type x position x isozyme interaction shows that it also is not significant; there is thus reasonable agreement between the behavior of peroxidase and the behavior of acid phosphatase isozymes.

650 Tyson, Fieldes, Cheung, and Starobin
In summary, these results indicate that significant R,, trends occur over the four leaf positions sampled up the main stem. There is a distinct difference between the trends for leaf position effects for highly branched types and those for sparsely branched types, with higher R,~'s occurring in the middle stem zones for R and L, in contrast to higher R,,'s at the base and apex for M and S in both enzyme systems. Temperature also produces R,, shifts, although it must be borne in mind that with the markedly different temperatures used here throughout the 47-day growing period prior to sampling, the correspond- ing rates of development were very different. Thus, the temperature effect on R,, may well be a developmental difference.
DISCUSSION
The leaf position, temperature, and genotype/genotroph effects on the Rm'S of peroxidases and acid phosphatases which are reported here interact with one another. Nevertheless, the results shown in Fig. 1 strongly suggest that Rm changes from one point on the stem to the next are not random but represent real and coherent trends. These trends are not merely linear but display at least one turning point in R,, change between base and apex, with the Rm reaching a maximum or a minimum between base and apex. The shape of these trends also appears to be related to the characteristic branching phenotype of the genotype/genotroph; pronounced branching, at least in this small sample, appears to be associated with a Rm maximum just above the stern base, whereas the contrasting less branched phenotypes display a R m
minimum just below the apex. The stem to apex, within-type variability here of from 6 to 11% is in the same range as the 6 to 9% variability among the means of the four types.
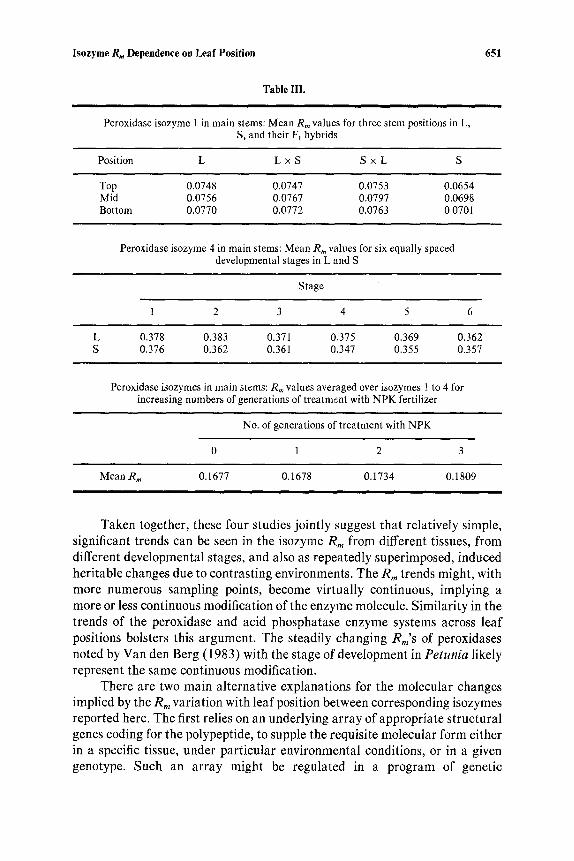
Data indicating significant Rm trends have been obtained from three previous studies using flax and peroxidase isozymes; R,~ values from these are summarized in Table III. They show that R m shifts for isozyme 1 are also detectable in stem tissue sampled at three positions from the main stem base to the apex (Fieldes et al., 1976b). For isozyme 4 in main stems over a 20-day development period during the vegetative phase, there appears to be a steady, linear decrease in Rm from day 16 to day 36 in both L and S (Fieldes and Tyson, 1973). Finally, in looking at progenies from plants repeatedly treated in either NPK or NK fertilizers, there is a discernible R m increase superim- posed for each successive generation of treatment with NPK relative to NK (Fieldes and Tyson, 1972). The Rm'S were assayed in stem tissues of plants which had been grown in uniform conditions for two further generations after the final fertilizer treatment; the Rm shifts detected represented induced heritable changes of the type documented at length by Durrant (1971). This R,, increase, dependent on the number of generations of NPK treatment, is essentially linear, using the means of the four peroxidase isozymes.
Isozyme R~, Dependence on Leaf Position
Table III.
Peroxidase isozyme 1 in main stems: Mean R,~ values for three stem positions in L, S, and their FI hybrids
651
Position L L x S S x L S
Top 0.0748 0.0747 0.0753 0.0654 Mid 0.0756 0.0767 0.0797 0.0698 Bottom 0.0770 0.0772 0.0763 0.0701
Peroxidase isozyme 4 in main stems: Mean R,, values for six equally spaced developmental stages in L and S
Stage
1 2 3 4 5 6
L 0.378 0.383 0.371 0.375 0.369 0.362 S 0.376 0.362 0.361 0.347 0.355 0.357
Peroxidase isozymes in main stems: Rm values averaged over isozymes 1 to 4 for increasing numbers of generations of treatm~.nt with NPK fertilizer
No. of generations of treatment with NPK
0 1 2 3
Mean Rm 0.1677 0.1678 0.1734 0.1809
Taken together, these four studies jointly suggest that relatively simple, significant trends can be seen in the isozyme R,, from different tissues, from different developmental stages, and also as repeatedly superimposed, induced heritable changes due to contrasting environments. The Rm trends might, with more numerous sampling points, become virtually continuous, implying a more or less continuous modification of the enzyme molecule. Similarity in the trends of the peroxidase and acid phosphatase enzyme systems across leaf positions bolsters this argument. The steadily changing Rm's of peroxidases noted by Van den Berg (1983) with the stage of development in Petunia likely represent the same continuous modification.
There are two main alternative explanations for the molecular changes implied by the Rm variation with leaf position between corresponding isozymes reported here. The first relies on an underlying array of appropriate structural genes coding for the polypeptide, to supple the requisite molecular form either in a specific tissue, under particular environmental conditions, or in a given genotype. Such an array might be regulated in a program of genetic

652 Tyson, Fieldes, Cheung, and Starobin
expression determined by some master gene activity, by feedback control of the transcription of the genes in the array, or by controlled degradation of mRNA or protein variants. The second explanation concerns posttranscrip- tional or posttranslational modifications, for example, alterations in mRNA, processing of the translated polypeptide, or changes due to the system's (de)glycosylating (glyco)proteins.
For isozymes which correspond, as defined in the Introduction, the more variation of Rm in different tissues, developmental stages, or genotypes/ genotrophs, the less the likelihood that this molecular variation arises from an increasingly large repertoire of structural genes coding for the polypeptides. Moreover, where isozymes of two or more enzyme systems display the same R,, trends, a multiplicity of underlying structural genes accounting for the trends is even less likely in view of the complexity of the regulatory systems needed. There must also be some limit to the number which can be generated of closely related, workable polypeptide variants for a given enzyme function. If the number of structural loci coding for the proteins were minimized by physical reshuffling within each structural DNA sequence responsible for each enzyme (Williams et al., 1979), the control mechanisms needed for more than one enzyme system to produce such parallel R,, changes would still be complex. Finally, even single codon changes in a given structural gene would be expected to generate discrete stepwise R,, changes for the resultant product, as opposed to a virtual continuum. As a result of these basic considerations, it would be more plausible to look at posttranscriptional and/or posttranslational modification.
Peroxidases in plants are monomeric glycoproteins, and monomerism likely holds for acid phosphatases also. Rm differences between corresponding peroxidase and acid phosphatase isozymes in the L and S genotrophs reflect molecular weight changes (Fieldes and Tyson, 1983). It is possible that the R m
differences related to the leaf position for corresponding isozymes also reflect molecular weight changes. This situation with monomeric isozymes cannot be explained on the basis of association changes in the type and number of enzyme subunits, as, for example, in the case of LDH variation in different mammalian tissues (Maclean, 1976).
Modification in the enzyme glycoprotein carbohydrate (CHO) moiety as development, etc., proceeds is thus a reasonable alternative mechanism, given the relatively large proportion of CHO (up to 20%) in these plant peroxidases and the small molecular changes suggested by the Rm data. CHO modification also fits with the complete absence of codominance in hybrids, an absence implying masking of other molecular differences by CHO. The higher molecular weights of individual S-genotroph peroxidases compared to corre- sponding L's might thus reflect more attached CHO. Genetic systems elsewhere in which posttranslational modifications in enzyme molecules may

Isozyme Rm Dependence on Leaf Position 653
be the cause of alterations in isozyme R m have been described in Drosophila by Finnerty and Johnson (1979) and in Calocedrus by Harry (1983). Intracellular control of CHO modifications in a glycoprotein has been examined by Williams and Lennarz (1984); processing of attached CHO appears to be a factor in protecting specific glycoproteins from intracellular proteolytic digestion. This appears to depend on both the conformation of the target glycoprotein and the CHO processing enzymes available within the cell.
The last case leads into the question of the adaptive significance, if any, of the isozyme modifications seen in the data from flax genotypes and genotrophs. One aspect concerns the adaptation of cells in different tissues within the organism; another relates to differences between individuals in populations. The isozymes of leucine aminopeptidase in Mytilus edulis possess a spectrum of altered characteristics, and Koehn (1978) has demon- strated the possible adaptive significance of such variation for Mytilus populations. In flax, in vitro thermal stability differences have been detected between corresponding L and S peroxidases. This finding needs, of course, to be coupled with evidence of differential fitness due to the alternative L and S forms.
One last point from the data here concerns the overlapping at the top of the stem of the curves for three of the four types (fig. 1). This suggests that the three can all make the same versions of the enzymes, yet in the more differentiated tissue towards the stem base they clearly produce distinctly different versions, as can be seen by the non-overlap of all four curves in fig. 1. Taken together with the fact that the curves are different for the two main morphological types, regulatory control by a genome with a considerable repertoire/potential is suggested. The genetical analyses of R m control, and the R,, variability within the genotypes, all seem to point to this potential, with the Rm changes stemming from post-transcriptional or post-translational modifications.
REFERENCES
Durrant, A. (1971). The environmental induction of heritable change in Linum. Heredity 17:27. Fieldes, M. A., and Tyson, H. (1972). Activity and relative mobility of peroxidase isozymes in
genotrophs and genotypes of flax (Linum usitatissimum L.). Can. J. Genet. Cytol. 14:625. Fieldes, M. A., and Tyson, H. (1973). Activity and relative mobility of peroxidase and esterase
isozymes of flax (Linum usitatissimum) genotrophs. I. Developing main stems. Can. J. Genet. Cytol. 15:731.
Fieldes, M. A., and Tyson, H. (1983). Molecular weight differences in acid phosphatases of stem homogenates from L and S flax genotrophs. Biochem. Gen. 21:391.
Fieldes, M. A., and Tyson, H. (1984). Possible post-translational modification, and its genetic control, in flax genotroph isozymes. Biochem. Genet. 22:99.
Fieldes, M. A., Bashour, N., Deal, C. L., and Tyson, H. (1976a). Isolation of peroxidase isozymes from two flax genotypes by column chromatography. Can. J. Bot. 54:1180.

L
654 Tyson, Fieldes, Cheung, and Starobin
Fieldes, M. A., Tyson, H., and Bashour, N. (1976b). Relative mobility shifts in anionic peroxidase isozymes between stem base and apex of flax genotrophs. Phytochemistry 15:247.
Fieldes, M. A., Deal, C. L., and Tyson, H. (1977). Preliminary characterisation of peroxidase isozymes isolated from two flax genotrophs. Can. J. Bot. 55:1465.
Fieldes, M. A., Starobin, J., and Tyson, H. (1984). Visualisation of both peroxidase and acid phosphatase isozymes from flax (Linum) on the same gels. Anal. Biochem. 137:146.
Finnerty, V., and Johnson, G. (1979). Post-translational modification as a potential explanation of high levels of enzyme polymorphism: Xanthine dehydrogenase and alcohol oxidase in Drosophila melanogaster. Genetics 91:695.
Harry, D. (1983). Identification of a locus modifying the electrophoretic mobility of malate dehydrogenase isozymes in incense cedar (Calocedrus decurrens), and its implications for population studies. Biochem. Genet. 21:417.
Hemmings, B. A., Zubenko, G. S., Haslik, A., and Jones, E. W. (1981). Mutant defective in processing of an enzyme located in the lysosome-like vacuole of Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 78:435.
Koehn, R. K. (1978). Physiology and biochemistry of enzyme variation: The interface of ecology and population genetics. In Brussard, P. F. (ed.), Ecological Genetics: The Interface, Springer-Verlag, New York, pp. 51-72.
Maclean, N. (1976). Control of Gene Expression, Academic Press, New York, p. 46. Mastonbroek, O., Hogeweg, P., Heringa, J., Niemann, G. J., Van Nigtevecht, G., and Van
Brederode, J. (1984). Isozyme variation in Silene pratensis: A response to different environments. Biochem. System. Ecol. 12:29.
Murashige, T., and Skoog, F. (1962). A revised medium for rapid growth and bio-assays with tobacco tissue. Physiol. Plant. 15:473.
Perkins, J. M., Eglinton, E. G., and Jinks, J. L. (1971). The nature of the inheritance of the permanently induced changes in Nicotiana rustica. Heredity 27:441.
Sokal, R. R., and Rohlf, F. J. (1981). Biometry, Freeman, San Francisco, p. 179. Tyson, H., Taylor, S. A., and Fieldes, M. A. (1978). Segregation of the environmentally induced
relative mobility shifts in flax genotroph peroxidase isozymes. Heredity 40:281. Van den Berg, B. M., Bianchi, F., and Wijsman, H. J. W. (1983). Genetics of the peroxidase
isoenzymes in Petunia. 5. Differential temporal expression of prxA alleles. Theoret. Appl. Genet. 65:1.
Welinder, K. G. (1976). Covalent structure of the glycoprotein horseradish peroxidase (EC 1.11.1.7). FEBS Lett. 72:19.
Williams, D. B., and Lennarz, W. J. (1984). Control of asparagine-linked oligosaccharide chain processing: Studies on bovine pancreatic ribonuclease B--an in vitro system for the processing of exogenous glycoproteins. J. Biol. Chem. 259:5105.
Williams, R. O., Young, J. R., and Majiwa, P. A. O. (1979). Genomic rearrangements correlated with antigenic variation in Trypanosoma brucei. Nature (Lond.) 282:847.














![[Hepatic steatosis, visceral fat and metabolic alterations in apparently healthy overweight/obese individuals]](https://static.fdokumen.com/doc/165x107/6324f8237fd2bfd0cb03375f/hepatic-steatosis-visceral-fat-and-metabolic-alterations-in-apparently-healthy.jpg)





