Isolation, sequencing, and functional analysis of the TATA-less human ATPase II promoter
13
Promoter paper Isolation, sequencing, and functional analysis of the TATA-less human ATPase II promoter $ Tomasz Sobocki a , Farah Jayman a , Malgorzata B. Sobocka b , Ruth Duchatellier a , Probal Banerjee a, * a Department of Chemistry and the CSI/IBR Center for Developmental Neuroscience, The City University of New York at The College of Staten Island, Staten Island, NY 10314, United States b Department of Medicine, SUNY Downstate Medical Center, Brooklyn, NY 10203, United States Received 17 December 2004; received in revised form 10 February 2005; accepted 21 February 2005 Available online 24 March 2005 Abstract Multiple lines of evidence indicate that the P-type Mg 2+ -ATPase, termed ATPase II, could play an important role in apoptosis. With the long-term objective of studying the regulation of this protein during apoptosis, we delineated the exon – intron organization of the human ATPase II gene (within chromosome 4). Subsequently, we used RNA ligase-mediated rapid amplification of cDNA ends to identify a major transcription start site at position 143 with respect to the translation start site. Luciferase reporter analysis of a 1.2-kb 5V -flanking sequence (1222 to +94 with respect to the transcription start site) revealed strong promoter activity in three human cell lines, human oligodendroglioma (HOG), SHSY5Y (hybrid neuroblastoma), and EA.hy926 (endothelial cell line). Serial deletions from the 5V end of this sequence up to nucleotide 291 yielded some decrease in activity only in the EA.hy926 cells. Further deletion to 217 caused a drastic decrease in activity in all three cell lines, but a 148 fragment showed preferential reduction in activity in the EA.hy926 cells. The promoter activity was nearly equal in two sequence variants of the promoter, one of which (designated as Variant 2) contained a 15-bp direct repeat within a GC-rich region. Additionally, there were several single base-pair changes from the sequence reported by the human genome project. Despite the presence of enhancer/repressor elements, such as Sp1 and NFnB, relatively small differences in promoter activity were observed in the three cell lines. However, it is likely that such sequence elements could cause major regulation of promoter activity in cells subjected to conditions that trigger apoptosis. The ATPase II promoter sequence will provide valuable clues to the regulation and role of the ATPase II protein. D 2005 Elsevier B.V. All rights reserved. Keywords: P-type ATPase; Promoter; Human ATPase II; Apoptosis; Aminophospholipid translocase 1. Introduction In live cells, the aminophosphospholipid, phosphatidyl- serine (PS), is normally sequestered to the inner leaflet of the plasma membrane [1–7]. This is caused by an enzymatic activity, termed aminophospholipid translocase (APTL), which uses energy derived from ATP hydrolysis to drive translocation of PS from the outer to the inner leaflet of the plasma membrane [4]. The lipid translocase activity is Mg 2+ -dependent, and is inhibited by vanadate, high intracellular calcium, and the sulfhydryl-modifying agent N-ethylmaleimide [4,8]. The mammalian enzyme ATPase II that belongs to a family of P-type ATPases is also inhibited by the same inhibitors described above for APTL [6,7]. Both activities, i.e. the PS-translocating activity of APTL as well as the ATPase activity of ATPase II, are inhibited in the presence of increased intracellular 0167-4781/$ - see front matter D 2005 Elsevier B.V. All rights reserved. doi:10.1016/j.bbaexp.2005.02.007 Abbreviations: HGP, Human Genome Project; TF, transcription factor; RLM-RACE, RNA ligase-mediated rapid amplification of cDNA ends; APTL, aminophospholipid translocase $ The sequence data presented in this article have been entered into the EMBL/GenBank database under accession numbers AY775564 and AY775565. * Corresponding author. Tel.: +1 718 982 3938; fax: +1 718 982 3944. E-mail address: [email protected] (P. Banerjee). Biochimica et Biophysica Acta 1728 (2005) 186 – 198 http://www.elsevier.com/locate/bba
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Isolation, sequencing, and functional analysis of the TATA-less human ATPase II promoter

http://www.elsevier.com/locate/bba
Biochimica et Biophysica Act
Promoter paper
Isolation, sequencing, and functional analysis of the TATA-less human
ATPase II promoter$
Tomasz Sobockia, Farah Jaymana, Malgorzata B. Sobockab, Ruth Duchatelliera,
Probal Banerjeea,*
aDepartment of Chemistry and the CSI/IBR Center for Developmental Neuroscience, The City University of New York at The College of Staten Island,
Staten Island, NY 10314, United StatesbDepartment of Medicine, SUNY Downstate Medical Center, Brooklyn, NY 10203, United States
Received 17 December 2004; received in revised form 10 February 2005; accepted 21 February 2005
Available online 24 March 2005
Abstract
Multiple lines of evidence indicate that the P-type Mg2+-ATPase, termed ATPase II, could play an important role in apoptosis. With the
long-term objective of studying the regulation of this protein during apoptosis, we delineated the exon–intron organization of the human
ATPase II gene (within chromosome 4). Subsequently, we used RNA ligase-mediated rapid amplification of cDNA ends to identify a major
transcription start site at position �143 with respect to the translation start site. Luciferase reporter analysis of a 1.2-kb 5V-flanking sequence
(�1222 to +94 with respect to the transcription start site) revealed strong promoter activity in three human cell lines, human
oligodendroglioma (HOG), SHSY5Y (hybrid neuroblastoma), and EA.hy926 (endothelial cell line). Serial deletions from the 5V end of this
sequence up to nucleotide �291 yielded some decrease in activity only in the EA.hy926 cells. Further deletion to �217 caused a drastic
decrease in activity in all three cell lines, but a �148 fragment showed preferential reduction in activity in the EA.hy926 cells. The promoter
activity was nearly equal in two sequence variants of the promoter, one of which (designated as Variant 2) contained a 15-bp direct repeat
within a GC-rich region. Additionally, there were several single base-pair changes from the sequence reported by the human genome project.
Despite the presence of enhancer/repressor elements, such as Sp1 and NFnB, relatively small differences in promoter activity were observed
in the three cell lines. However, it is likely that such sequence elements could cause major regulation of promoter activity in cells subjected to
conditions that trigger apoptosis. The ATPase II promoter sequence will provide valuable clues to the regulation and role of the ATPase II
protein.
D 2005 Elsevier B.V. All rights reserved.
Keywords: P-type ATPase; Promoter; Human ATPase II; Apoptosis; Aminophospholipid translocase
1. Introduction
In live cells, the aminophosphospholipid, phosphatidyl-
serine (PS), is normally sequestered to the inner leaflet of
0167-4781/$ - see front matter D 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.bbaexp.2005.02.007
Abbreviations: HGP, Human Genome Project; TF, transcription factor;
RLM-RACE, RNA ligase-mediated rapid amplification of cDNA ends;
APTL, aminophospholipid translocase$ The sequence data presented in this article have been entered into the
EMBL/GenBank database under accession numbers AY775564 and
AY775565.
* Corresponding author. Tel.: +1 718 982 3938; fax: +1 718 982 3944.
E-mail address: [email protected] (P. Banerjee).
the plasma membrane [1–7]. This is caused by an
enzymatic activity, termed aminophospholipid translocase
(APTL), which uses energy derived from ATP hydrolysis
to drive translocation of PS from the outer to the inner
leaflet of the plasma membrane [4]. The lipid translocase
activity is Mg2+-dependent, and is inhibited by vanadate,
high intracellular calcium, and the sulfhydryl-modifying
agent N-ethylmaleimide [4,8]. The mammalian enzyme
ATPase II that belongs to a family of P-type ATPases is
also inhibited by the same inhibitors described above for
APTL [6,7]. Both activities, i.e. the PS-translocating
activity of APTL as well as the ATPase activity of ATPase
II, are inhibited in the presence of increased intracellular
a 1728 (2005) 186 – 198

T. Sobocki et al. / Biochimica et Biophysica Acta 1728 (2005) 186–198 187
Ca2+ concentration (>0.2 MM) and selective depletion of
intracellular Ca2+, which causes an inhibition of the Ca2+-
ATPase, leaves the enzymes completely active [9,10].
Furthermore, a yeast strain with a mutant gene (Drs2),
which is homologous to the mammalian ATPase II gene,
showed inhibited translocation of PS across the plasma
membrane [7]. Finally, our earlier studies have demon-
strated that the overexpression of the mouse ATPase II
cDNA in the mouse hippocampus-derived, hybrid neuro-
blastoma (HN2) causes an increase in PS translocation and
expression of voltage-gated calcium currents (which
regulate apoptosis) in this otherwise calcium channel-
deficient cell line [6,11–13]. Thus, besides its activity as
an ATPase, ATPase II could play a major role in the
maintenance of lipid asymmetry of the plasma membrane
of a cell.
The P-type ATPases are so named because they harbor
a DKTG motif in which the Asp (D) residue accepts the
;-phosphate (P) from ATP during the catalytic cycle,
forming a covalent acylphosphate intermediate, which is a
signature property of these transporters [14]. For the
completion of the catalytic cycle of the P-type ATPases, it
is essential that this phosphate group is eventually cleaved
[15,16]. Remarkably, such dephosphorylation of ATPase
II proceeds only in the presence of phosphatidylserine
(PS) [17]. The large family of more than 150 P-type
ATPases includes many proteins that are believed to
perform important physiological functions such as the
transport of metal ions like Mg2+, Cu2+ or Ca2+.
Mutations in certain P-type ATPases cause a copper
transport defect named Wilson’s disease and also two
forms of hereditary Cholestasis [14,18]. As an Mg2+-
ATPase, ATPase II does not transport Na+ or K+ ions and
is insensitive to the concentrations of these ions in the
medium.
During apoptosis, PS is externalized to serve as a
marker, which is recognized by scavenger cells that engulf
and degrade apoptotic cells [1,3,19]. Since APTL inhib-
ition might be central to this PS-externalization, the
expression of ATPase II could be critical in this process
and thus may be tightly regulated so as to control
phagocytic clearance of specific cell types in many parts
of an organism. The tissue specificity of the expression of
the ATPase II mRNA has been reported earlier [6,12], but
no information has been available on the sequence of the
ATPase II promoter. Surprisingly, among the 150 or more
P-type ATPases known so far, the promoter of only one P-
type ATPase gene (the Wilson Disease gene) has been
identified [20]. Our study reports the isolation of the
ATPase II promoter, and analysis of its structure and
function. Further analysis of this promoter in more cell
types, under various conditions of growth factor-mediated
regulation, will reveal its mechanistic role in controlling
ATPase II expression under specific conditions. This will
help build a clearer understanding of the regulation and
role of this protein in apoptosis.
2. Materials and methods
2.1. Organization of the ATPase II gene
In order to elucidate the chromosomal organization of
the ATPaseII gene, we compared the sequences of ATPaseII
cDNAs AB013452, AF067820 and NM_006095 to Human
Genome Sequence using the Human Genome Browser
(http://genome.ucsc.edu/; [21,22]). The data presented in
this paper has been obtained using assembly as of May
2004 [21,22], which is the most resent version. Align-
ments of genomic and cDNA sequences were carried out
using ClustalW ([23]; http://www.ebi.ac.uk/clustalw/) or
BCM Search Launcher: Multiple Sequence Alignments
software package ([24]; http://searchlauncher.bcm.tmc.edu/
multi-align/multi-align.html). Wherever necessary, sequen-
ces were aligned manually using standard word process-
ing software and visually inspected.
2.2. Searching for the potential promoter region and
transcription factor binding sites in the ATPaseII gene
The search was performed on the Internet using
GenomatixSuite collection of software (Genomatix Soft-
ware GmbH, Munich, Germany) available at http://www.
genomatix.de. The PromoterInspector [25] mammalian
promoter prediction software was used to analyze the 5Vend of the ATPaseII gene for the presence of the promoter
and the MatInspector [26] to identify the potential tran-
scription factor binding sites. The parameters of the
MatInspector search were set to the core similarity of 1.00
and the optimized matrix similarity.
2.3. Primer design
In order to select PCR primers of specific length and
temperature of annealing, we used the ‘‘RAWPRIMER’’
program (http://alces.med.umn.edu/rawprimer.html). Pri-
mers were analyzed for the absence of stems or hairpin loops
using the ‘‘Primerdesign’’ utility (http://www.cybergene.se/
primer.html). The forward primers contained CGG GGT
ACC, while the reverse primers harbored CTA GCT AGC
(KpnI or NheI sites respectively, underlined plus three
protecting bases). These restriction sites enabled us to
perform directional cloning of the PCR products into the
reporter vector pGL3Basic (Promega Corporation, Madison,
WI). The custom primers used in this project (Table 1) were
synthesized by Invitrogen (Carlsbad, CA).
2.4. Preparation of human genomic DNA
Genomic DNA from human white blood cells was
prepared using the Wizard\ Genomic DNA Purification
Kit from Promega as specified in the manufacturer’s
protocol. The precipitated genomic DNA was re-hydrated
overnight at 4 -C. The concentration, purity and quality of
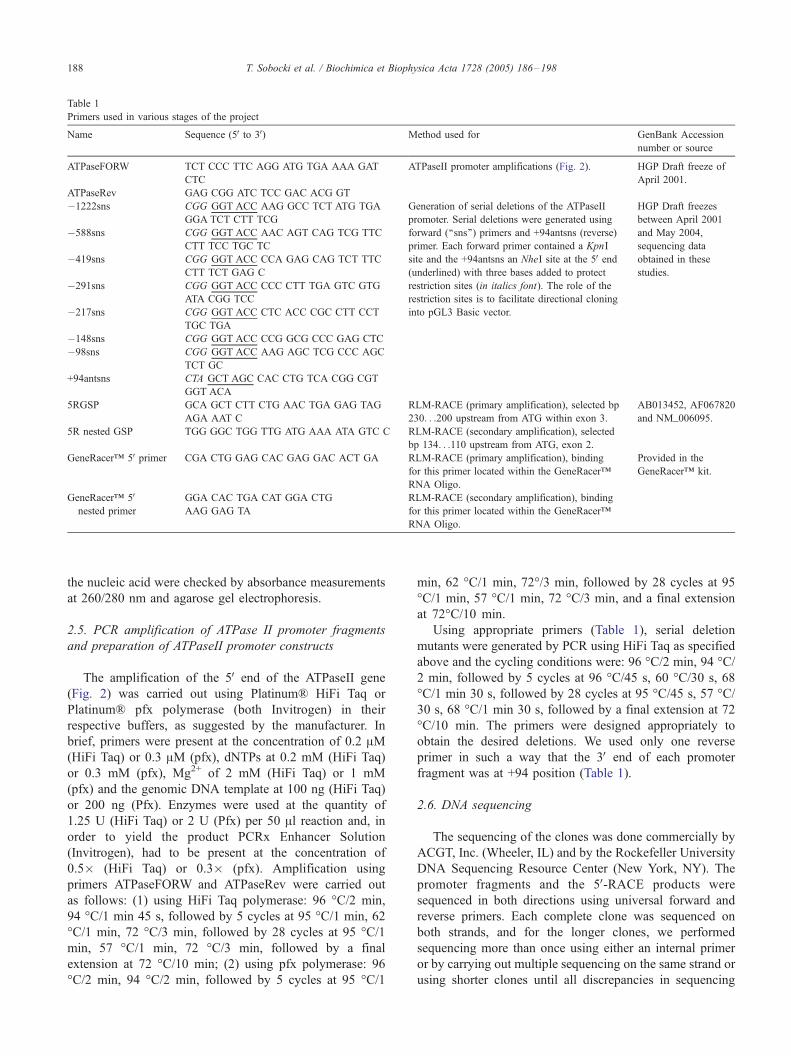
Table 1
Primers used in various stages of the project
Name Sequence (5V to 3V) Method used for GenBank Accession
number or source
ATPaseFORW TCT CCC TTC AGG ATG TGA AAA GAT
CTC
ATPaseII promoter amplifications (Fig. 2). HGP Draft freeze of
April 2001.
ATPaseRev GAG CGG ATC TCC GAC ACG GT
�1222sns CGG GGT ACC AAG GCC TCT ATG TGA
GGA TCT CTT TCG
Generation of serial deletions of the ATPaseII
promoter. Serial deletions were generated using
forward (‘‘sns’’) primers and +94antsns (reverse)
primer. Each forward primer contained a KpnI
site and the +94antsns an NheI site at the 5V end(underlined) with three bases added to protect
restriction sites (in italics font). The role of the
restriction sites is to facilitate directional cloning
into pGL3 Basic vector.
HGP Draft freezes
between April 2001
and May 2004,
sequencing data
obtained in these
studies.
�588sns CGG GGT ACC AAC AGT CAG TCG TTC
CTT TCC TGC TC
�419sns CGG GGT ACC CCA GAG CAG TCT TTC
CTT TCT GAG C
�291sns CGG GGT ACC CCC CTT TGA GTC GTG
ATA CGG TCC
�217sns CGG GGT ACC CTC ACC CGC CTT CCT
TGC TGA
�148sns CGG GGT ACC CCG GCG CCC GAG CTC
�98sns CGG GGT ACC AAG AGC TCG CCC AGC
TCT GC
+94antsns CTA GCT AGC CAC CTG TCA CGG CGT
GGT ACA
5RGSP GCA GCT CTT CTG AAC TGA GAG TAG
AGA AAT C
RLM-RACE (primary amplification), selected bp
230. . .200 upstream from ATG within exon 3.
AB013452, AF067820
and NM_006095.
5R nested GSP TGG GGC TGG TTG ATG AAA ATA GTC C RLM-RACE (secondary amplification), selected
bp 134. . .110 upstream from ATG, exon 2.
GeneRaceri 5V primer CGA CTG GAG CAC GAG GAC ACT GA RLM-RACE (primary amplification), binding
for this primer located within the GeneRaceri
RNA Oligo.
Provided in the
GeneRaceri kit.
GeneRaceri 5Vnested primer
GGA CAC TGA CAT GGA CTG
AAG GAG TA
RLM-RACE (secondary amplification), binding
for this primer located within the GeneRaceri
RNA Oligo.
T. Sobocki et al. / Biochimica et Biophysica Acta 1728 (2005) 186–198188
the nucleic acid were checked by absorbance measurements
at 260/280 nm and agarose gel electrophoresis.
2.5. PCR amplification of ATPase II promoter fragments
and preparation of ATPaseII promoter constructs
The amplification of the 5V end of the ATPaseII gene
(Fig. 2) was carried out using Platinum\ HiFi Taq or
Platinum\ pfx polymerase (both Invitrogen) in their
respective buffers, as suggested by the manufacturer. In
brief, primers were present at the concentration of 0.2 AM(HiFi Taq) or 0.3 AM (pfx), dNTPs at 0.2 mM (HiFi Taq)
or 0.3 mM (pfx), Mg2+ of 2 mM (HiFi Taq) or 1 mM
(pfx) and the genomic DNA template at 100 ng (HiFi Taq)
or 200 ng (Pfx). Enzymes were used at the quantity of
1.25 U (HiFi Taq) or 2 U (Pfx) per 50 Al reaction and, in
order to yield the product PCRx Enhancer Solution
(Invitrogen), had to be present at the concentration of
0.5� (HiFi Taq) or 0.3� (pfx). Amplification using
primers ATPaseFORW and ATPaseRev were carried out
as follows: (1) using HiFi Taq polymerase: 96 -C/2 min,
94 -C/1 min 45 s, followed by 5 cycles at 95 -C/1 min, 62
-C/1 min, 72 -C/3 min, followed by 28 cycles at 95 -C/1min, 57 -C/1 min, 72 -C/3 min, followed by a final
extension at 72 -C/10 min; (2) using pfx polymerase: 96
-C/2 min, 94 -C/2 min, followed by 5 cycles at 95 -C/1
min, 62 -C/1 min, 72-/3 min, followed by 28 cycles at 95
-C/1 min, 57 -C/1 min, 72 -C/3 min, and a final extension
at 72-C/10 min.
Using appropriate primers (Table 1), serial deletion
mutants were generated by PCR using HiFi Taq as specified
above and the cycling conditions were: 96 -C/2 min, 94 -C/2 min, followed by 5 cycles at 96 -C/45 s, 60 -C/30 s, 68
-C/1 min 30 s, followed by 28 cycles at 95 -C/45 s, 57 -C/30 s, 68 -C/1 min 30 s, followed by a final extension at 72
-C/10 min. The primers were designed appropriately to
obtain the desired deletions. We used only one reverse
primer in such a way that the 3V end of each promoter
fragment was at +94 position (Table 1).
2.6. DNA sequencing
The sequencing of the clones was done commercially by
ACGT, Inc. (Wheeler, IL) and by the Rockefeller University
DNA Sequencing Resource Center (New York, NY). The
promoter fragments and the 5V-RACE products were
sequenced in both directions using universal forward and
reverse primers. Each complete clone was sequenced on
both strands, and for the longer clones, we performed
sequencing more than once using either an internal primer
or by carrying out multiple sequencing on the same strand or
using shorter clones until all discrepancies in sequencing

T. Sobocki et al. / Biochimica et Biophysica Acta 1728 (2005) 186–198 189
were resolved. Any isolated sequence that was different
from the corresponding sequence obtained from the human
genome project (for example a 15-bp repeat observed in
human ATPase II promoter) was sequenced several times.
2.7. Transient transfection studies of the ATPase II promoter
Transient transfections with ATPase II promoter con-
structs in pGL3Basic (firefly luciferase) and pRL-TK
(Renilla luciferase) into HOG, SHSY5Y and EA.hy926
cells were carried out using the Effectene reagent (Qiagen,
Valencia, CA) according to the manufacturer’s recommen-
dations. In brief, cotransfection mixtures contained (per well
of cells in 24-well plates) promoter construct pDNA (195 ng
for EA.hy926 and SHSY5Y or 200 ng for HOG), pRL-TK
(5 ng for EA.hy926 and SHSY5Y or 2.5 ng for HOG), EC
buffer (60 Al), Enhancer (1.6 Al), Effectenei (5 Al) and
DMEM (350 Al, 10% FBS, 1% Penicillin–Streptomycin).
The cotransfection mixture was added to cells overlayered
with 350 Al of DMEM (10% FBS, 1% Penicillin–
Streptomycin). The medium plus cotransfection mixes were
subsequently removed after either 4 h (EA.hy926) or
approximately 24 h (SHSY5Y or HOG), and the cells were
overlayered with 1 ml of DMEM (10% FBS, 1% Penicillin–
Streptomycin) per well. Cells were harvested 48 h after
transfection, and luciferase activities were assayed using a
dual luciferase assay kit (Promega) as specified by the
manufacturer. Luminescence from either firefly or Renilla
luciferase was measured using a TD20/20 Luminometer
(Turner Design, Sunnyvale, CA). Results were expressed as
a ratio of Firefly to Renilla luminescence. All results were
normalized to the activity obtained from the pGL3Promoter
vector (Promega).
2.8. Analysis of the 5V-untranslated sequences by RLM-RACE
Total RNA samples (5 Ag each) obtained from human
brain and skeletal muscle (Ambion, Austin, TX) were
processed for RLM-RACE using the GeneRacer kit as
detailed in the manufacturer’s protocol. In brief, the RNA
samples were dephosphorylated in the presence of 10 U of
calf intestinal phosphatase (CIP) at 50 -C for 1 h.
Following the purification of the RNA sample by ethanol
precipitation, the mRNA cap structure was removed by
treating the dephosphorylated RNA with 0.5 U of tobacco
acid pyrophosphatase (TAP). The decapped mRNA was
purified and then ligated in the presence of T4 ligase (5U)
to an adapter RNA oligonucleotide sequence that was
specific for both the GeneRacer 5V Primer and the
GeneRacer 5V Nested Primer. An aliquot of the purified
ligation product, containing approximately 1.2 Ag of
RACE-ready RNA, was reverse transcribed in the presence
of an oligo dT primer using 15 U Thermoscripti RT
(Invitrogen). The product was subsequently treated with 2
U of RNase H at 37 -C for 20 min and then the cDNA
product used immediately for PCR amplification. PCR
amplification was performed first with the GeneRacer 5VPrimer and an ATPase II-specific primer 5RGSP (Table 1).
The conditions of PCR were 94 -C/2 min followed by 5
cycles of 94 -C/30 s, 70 -C/1 min 30 s, followed by 5
cycles of 94 -C/30 s, 68 -C/1 min 30 s, followed by 25
cycles of 94 -C/25s, 63 -C/30 s, 68 -C/1 min, followed by
a final extension at 72 -C/10 min. An aliquot containing 1/
50th of the primary PCR product was then reamplified
using the GeneRacer 5V Nested Primer and a second
ATPase II-specific primer 5RNestedGSP (Table 1). The
conditions of PCR were 94 -C/2 min, followed by 25
cycles of 94 -C/25 s, 63 -C/30 s, 68 -C/1 min 30 s, and a
final extension at 72 -C/10 min.
3. Results
3.1. Organization of the ATPase II gene
The alignment of the ATPaseII cDNAs with sequence
of Human Genome revealed that the ATPaseII gene is
located in chromosome 4p13 and spans the region from
42,255,563 to 42,499,828 with the coding region matching
to the negative strand between bases 42,255,864 and
42,499,818 within chromosome 4. The gene is composed
of 37 exons (sizes: 41 to 396 bp) and 36 introns (size: 92
to 29,715bp). The exon/intron organization is shown in
Table 2 and the schematic structure of the gene is shown
in Fig. 1. Analysis of the 5V end of the gene revealed the
presence of the CpG island of the size equal to 1272
(42,499,207 to 42,500,478) [27]. The percentage of CpG
within the island is 17.3% and the total percentage of
either C or G equals to 62.9%. The parts of the ATPaseII
gene where the CpG island was present included the entire
5VUTR, the entire first exon and first 563 bases of the first
intron. In order to predict a potential promoter within the
5Vend of the ATPaseII gene, we used PromoterInspector to
scan 30-kb of DNA sequence starting 20 kb upstream of
the translation start site and spanning even 10 kb down-
stream of this site. Thus, the sequence scanned included
the entire first exon and the 5V end of the first intron. The
search indicated a promoter region from bp 19,617 to
19,828, which corresponded to bases �250 to �38 in
Fig. 2. Our search also revealed the lack of a TATA box
and a consensus initiator sequence. Thus, the transcription
initiation of the ATPase II gene could occur within a
segment of the core promoter, densely lined with tran-
scription factor binding sites (Fig. 2, Table 2, and Fig. 3).
For our analysis of promoter activity, however, we
considered the complete 1.2-kb 5V-flanking region. Our
objective was to look for possible enhancer/repressor
sequences, the deletion of which would produce a dramatic
change in promoter activity. It should be noted that the
complete sequences presented in this article have been
entered into the EMBL/GenBank database under accession
numbers AY775564 and AY775565.

Table 2
Exon– intron boundaries of the human ATPase II gene
Exon–Intron junctions on the human ATPase II gene
Exon cDNA AB013452 Exon size (bp) Chromosome 4p13 Intron|Exon|Intron
From To From To
1 1 62 �62 42499828 42499770 . . .. . .. . .. . .. . .. . .. . .. . .. . .. . .CCGAAG|GTAAGG
2 63 177 115 42470054 42469940 TTTAAG|GTTATG. . .. . .. . .TGTCAG|GTAAGA
3 178 277 100 42468658 42468559 TTCTAG|CACTGC. . .. . .. . .CTGCAG|GTAAAG
4 278 376 99 42467579 42467481 AAACAG|CAAATA. . .. . .. . .GATATT|GTAAGT
5 377 422 46 42459023 42458978 CTTTAG|AAACGA. . .. . .. . .CGCAAG|GTAGGA
6 423 463 41 42443463 42443423 TTCCAG|TTTTGA. . .. . .. . .GAAAAG|GTATGC
7 464 537 74 42433829 42433756 GTGTAG|GTGGCA. . .. . .. . .CTCAAG|GTTAGA
8 538 607 70 42431274 42431205 CTACAG|TGAGCC. . .. . .. . .AGACAG|GTAAGA
9 608 735 128 42429421 42429294 AAACAG|GGCTTA. . .. . .. . .ACATGG|GTATGT
10 736 847 111 42424677 42424566 TCCTAG|CACCGT. . .. . .. . .ATGCAG|GTGAAA
11 848 1013 166 42422923 42422758 TTACAG|AATTCA. . .. . .. . .TAAACT|GTAAGC
12 1014 1141 128 42421332 42421205 CAACAG|ATGGTG. . .. . .. . .AATTGG|GTAAAT
13 1142 1219 78 42418644 42418567 TTAAAG|GATCTT. . .. . .. . .GGCCAG|GTAAAC
14 1220 1308 89 42417652 42417564 TTTTAG|GTTAAA. . .. . .. . .TTATGG|GTGAGT
15 1309 1353 45 42412150 42412106 CTGAAG|CCATGT. . .. . .. . .TGAATG|GTAAGC
16 1354 1426 73 42398985 42398913 CTTCAG|GCAGAA. . .. . .. . .AATCAT|GTAAGT
17 1427 1532 106 42395555 42395450 TCACAG|CCAACT. . .. . .. . .CTCCAG|GTACAA
18 1533 1615 83 42394225 42394143 AAACAG|ATGAGG. . .. . .. . .GATTCA|GTAAGT
19 1616 1665 50 42392007 42391958 TCTTAG|CTGGGG. . .. . .. . .TACCAG|GTAAAA
20 1666 1735 70 42386931 42386862 TTTCAG|TGCTAG. . .. . .. . .GGAGCT|GTAAGT
21 1736 1820 85 42367792 42367708 TTCCAG|GACACT. . .. . .. . .CAGAAG|GTAAGT
22 1821 1960 140 42365244 42365105 TTTCAG|GGTTAA. . .. . .. . .GAAAAG|GTACGT
23 1961 2099 139 42350099 42349961 TTTCAG|AATCTT. . .. . .. . .ACATCG|GTAATT
24 2100 2164 65 42346459 42346395 ATACAG|GACACT. . .. . .. . .CTTGAT|GTAAGT
25 2165 2337 173 42328613 42328441 TTCCAG|GGAACA. . .. . .. . .CTGTCG|GTAAGT
26 2338 2521 184 42308021 42307838 TTTCAG|GGTTTC. . .. . .. . .GCTCAG|GTAAAG
27 2522 2632 111 42307745 42307635 TTTTAG|TTCAAA. . .. . .. . .ATCGAG|GTAACA
28 2633 2707 75 42298544 42298470 TCCTAG|ATCTGG. . .. . .. . .AACGTG|GTAAGT
29 2708 2830 123 42298364 42298242 CTTTAG|ATGTTT. . .. . .. . .ACCAAG|GTAGGG
30 2831 2909 79 42295004 42294926 CCTCAG|GTTTTC. . .. . .. . .AGTATG|GTAAGT
31 2910 2971 62 42289589 42289528 GTACAG|GTACTG. . .. . .. . .TACACT|GTGAGT
32 2972 3028 57 42287579 42287523 TTTTAG|TTTGTG. . .. . .. . .ACATGG|GTAAGA
33 3029 3136 108 42286617 42286510 CTACAG|TTCAGC. . .. . .. . .GGAGAG|GTAATG
34 3137 3225 89 42266650 42266562 ACACAG|GCAGCC. . .. . .. . .CAAGGT|GTAAGT
35 3226 3318 93 42265844 42265752 TTGCAG|TATCAA. . .. . .. . .AAAAAG|GTAAAA
36 3319 3410 92 42257663 42257572 CTGCAG|CCTGAC. . .. . .. . .TGCTCC|GTGAGT
37 3411 3806 �396 42255958 42255563 CTACAG|ATGGGT. . .. . .. . .GTATTC
Delineation of ATPase II gene structure: Exon– intron boundaries. The table shows consecutive exons, their corresponding position within ATPaseII cDNA
(AB013452) and within chromosome 4p13. Exon size and details of intron/exon/intron junctions are provided for each exon. Please note that according to the
HGP, ATPaseII exons match to the negative strand of the chromosome, which accounts for the fact that the consecutive numbering within the chromosome
decreases as the numbering within the cDNA increases.
T. Sobocki et al. / Biochimica et Biophysica Acta 1728 (2005) 186–198190
3.2. Transcription factor binding sites
The putative promoter sequence harbored many tran-
scription factor-binding sites, which are summarized in
Fig. 2 and Table 3. We have found some deviations from
the genomic sequence reported in the human genome
database (Table 4 and Fig. 3). The region from position
�419 up to position +38, as shown in Fig. 2 and Table 3,
contained a large number of transcription factor binding
sites and also sites selective for the core promoter binding
protein (CPBP), which is often observed in TATA-less
promoters [28]. Other transcription factor binding sequen-
ces were identified upstream of this region of the putative
promoter (Fig. 2 and Table 3).
3.3. Dissimilarities between the isolated ATPase II sequence
variants and the human genome project (HGP)
Two sequence variants were obtained. One of them
contained a 15-bp direct repeat in the core promoter
region (Variant 2, Table 4 and Fig. 3). They were
designated as Variant 1 {without the repeat and (�)}
and Variant 2 {with the repeat and (+)} (Table 4). Both
Variant 1 and Variant 2 sequences showed further
difference from the HGP at single base pairs listed in
Table 4. These differences could be due to single
nucleotide polymorphism. Since this 15-bp sequence
contained a core promoter binding protein (CPBP), an
EGR1, a myc-associated zinc finger protein (MAZ), and a

Fig. 1. Gene structure of human ATPase II. Delineation of the ATPase II gene structure using bioinformatics data obtained from public databases (GenBank/
NCBI, Human Genome Project, etc.). Top panel: schematic illustration of human chromosome 4 as interpreted by the Human Genome Project (HGP). A thin
vertical line and a triangle above indicate the position of the ATPaseII gene. Bottom panel: Structure of the ATPase gene drawn to scale. Exons are indicated by
thin vertical lines.
T. Sobocki et al. / Biochimica et Biophysica Acta 1728 (2005) 186–198 191
myc-associated zinc finger protein related transcription
factor binding sites, the repeat introduced additional
copies of these sites to the core promoter sequence
(Table 5).
Fig. 2. Isolation and characterization of the ATPaseII promoter. Shown is the varian
transcription start site determined by 5VRACE (Fig. 4). The start of transcription as
position where the additional 15-bp direct repeat is present in the Variant 2 of the
different from the HGP (see Table 3). TF binding sites determined by MATINSPEC
factors above the sequence. Primers used to amplify this fragment (ATPaseFoRW a
end of the sequence. The promoter region detected by PROMOTERINSPECTO
transcription factor (TF) binding sites, which could not be incorporated in this f
promoter region detected by PROMOTERINSPECTOR.
3.4. The transcription-start site on the ATPase II promoter
The transcription start site of the ATPase II gene has
not been identified so far. Additionally, the predicted
t I of the promoter (AY775564). The numbering of bases is in relation to the
determined by the 5VRACE is shown as ‘‘�’’. The symbol ‘‘ ’’ indicates the
promoter (Fig. 3). Asterisks (F*_) above the sequence indicate bases that are
TOR are indicated by lines below the sequence with names of transcription
nd ATPaseRev, Table 1) are depicted in gray italics at the beginning and the
R (�250/�38) is shown in bold. For the purpose of clarity, additional
igure, are listed in Table 3. Majority of these TF binding sites are in the

*ATPase(-) -140 CGAGCTCCGCCCCTCGCCGAGCCGCTCCTC---------------CGCGGCTGCAGCAAG -96ATPase+ -155 CGAGCTCCGCCCCTCGCCGAGCCGCCCCTCGCCGAGCCGCCCCTCCGCGGCTGCAGCAAG -96
Fig. 3. ATPase II promoter variant II (AY775565) contains 15-bp direct repeat. Location of the direct repeat is indicated by a horizontal line, beginning and
ending with arrowheads. The asterisk (F*_) above indicates mutation at base �115 (Table 4). This figure shows only the fragment of the two variants of the
ATPase II promoter where the sequences differ. Flanking sequences are included for the purpose of clarity.
T. Sobocki et al. / Biochimica et Biophysica Acta 1728 (2005) 186–198192
ATPase II promoter sequence is devoid of a TATA box or
a consensus eukaryotic initiator sequence, and it is known
that such TATA-less promoters often contain multiple
transcription start sites, which had to be determined. One
important part of our strategy was to design the reverse,
gene-specific primer in such a way that no genomic DNA
contamination was obtained in the 5V-RACE product. This
was achieved by positioning the gene-specific reverse
5RGSP and the nested gene-specific reverse 5RNes-
tedGSP primers (Table 1) on exon 3 and exon 2
respectively (Table 2). Since the 5V-untranslated region
of the ATPase II gene contains many GC-rich regions,
RT-PCR amplification of such templates could have been
difficult. We overcame this difficulty by using the
Thermoscripti RT (Invitrogen) for the reverse tran-
scription step.
5V-RACE required two rounds of PCR amplification i.e.
initial amplification of primary RACE products, followed
by subsequent re-amplification as described in Materials
and methods. Secondary products were clearly visible
upon analysis by agarose gel electrophoresis. The
Table 3
Additional transcription factor binding sites in the ATPase II promoter
Transcription factor Strand
OCT1 (�)
EGR1 (+)
(+)
(�)
Myc-associated zinc finger protein related TF (+)
(+)
(�)
(+)
(�)
GC Box/SP1 (+)
(�)
(�)
Myeloid zinc finger protein (MZF1) (�)
Myc associated zinc finger protein (MAZ) (�)
(�)
(�)
(�)
ZBP-89 (�)
(+)
Zinc finger/POZ domain TF (+)
(+)
AP2 (+)
CPBP (�)
(�)
HEN1 (�)
AP4 (�)
expected size of such secondary products is exactly 30
bp larger due to the presence at the 5V ends of the
products an additional stretch of oligodeoxynucleotide
originating from a part of reverse transcribed GeneR-
aceri RNA oligonucleotide sequence. We obtained major
bands of approximately 300-bp in RACE from brain and
skeletal muscle samples. In addition, 5V-RACE of brain
mRNA gave two shorter species of approximately 280 and
230 bp, respectively (Fig. 4A). The products were cloned
into pCR4-TOPO vector and sequenced (total of nine
clones). In the brain, the major band at ¨300 bp
corresponded to products from two closely spaced tran-
scription start sites at �150G and �143C with respect to the
translation initiation codon (Fig. 4A and B). The shorter
species at ¨230 bp corresponded to a transcription start
site at �71C. None of these transcription initiation sites
corresponded to the consensus, eukaryotic initiation
sequence. Nonetheless, such deviations are observed in a
large number of promoters. In the skeletal muscle, there
was one major band immediately below 300 bp, which
corresponded to the same transcription start site at �143C
Sequence Position
TAAATTTTAATTTGC �973/�959
GCGGGGCGGGGCGGGGC �264/�248
GCGGGGCGGGGCGAAGG �259/�243
CGGGTGAGGGGCGGGGA �226/�210
CGGGGCGGGGCGG �263/�251
CGGGGCGGGGCGA �258/�246
AGGGGCGGGGAGA �228/�216
GCGGGAGGGGGAG �190/�178
GGGGGAGGGGCCT �165/�153
GGCGGGGCGGGGCGA �260/�246
TGAGGGGCGGGGAGA �228/�214
CGAGGGGCGGAGCTC �139/�125
GCGGGGA �226/�220
GGGTGAGGGGCGG �223/�211
CGGCGAGGGGCGG �134/�122
CGCGGAGGAGCGG �119/�107
GGAGGAGGAGAAA �46/�34
GGCTCGCGCTCCCCCTCCCGCGG �192/�170
GCAGGCCCCTCCCCCGGCGCCGG �167/�145
GGGAGCGCGAG �182/�172
CTGAGCGCTCT +38/+48
CGCCCGGCGGCGG �14/�3
CGCGCCGCCGCCCACCTAGGGCA +2/+24
GCCTGGGCCGCGCCGCCGCCCAC +10/+32
GAGCGCTCAGCTGCAGCCTGG +27/+47
GCGCTCAGCTGCAGCCT +29/+45
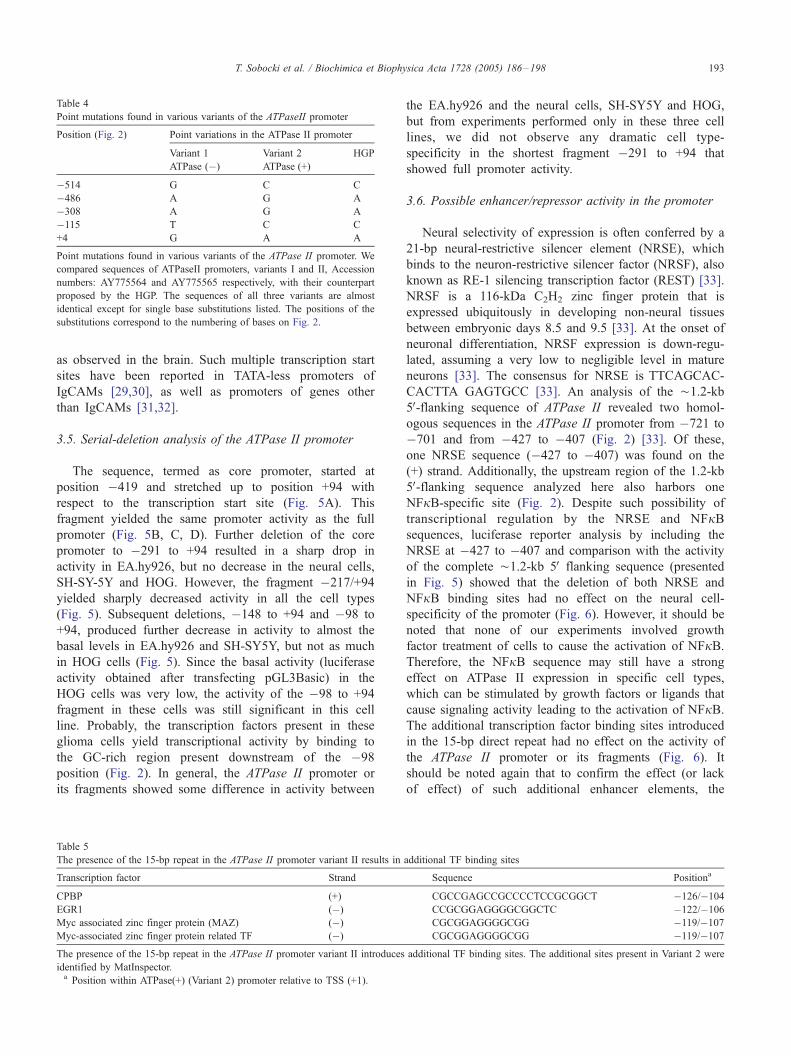
Table 4
Point mutations found in various variants of the ATPaseII promoter
Position (Fig. 2) Point variations in the ATPase II promoter
Variant 1
ATPase (�)
Variant 2
ATPase (+)
HGP
�514 G C C
�486 A G A
�308 A G A
�115 T C C
+4 G A A
Point mutations found in various variants of the ATPase II promoter. We
compared sequences of ATPaseII promoters, variants I and II, Accession
numbers: AY775564 and AY775565 respectively, with their counterpart
proposed by the HGP. The sequences of all three variants are almost
identical except for single base substitutions listed. The positions of the
substitutions correspond to the numbering of bases on Fig. 2.
T. Sobocki et al. / Biochimica et Biophysica Acta 1728 (2005) 186–198 193
as observed in the brain. Such multiple transcription start
sites have been reported in TATA-less promoters of
IgCAMs [29,30], as well as promoters of genes other
than IgCAMs [31,32].
3.5. Serial-deletion analysis of the ATPase II promoter
The sequence, termed as core promoter, started at
position �419 and stretched up to position +94 with
respect to the transcription start site (Fig. 5A). This
fragment yielded the same promoter activity as the full
promoter (Fig. 5B, C, D). Further deletion of the core
promoter to �291 to +94 resulted in a sharp drop in
activity in EA.hy926, but no decrease in the neural cells,
SH-SY-5Y and HOG. However, the fragment �217/+94
yielded sharply decreased activity in all the cell types
(Fig. 5). Subsequent deletions, �148 to +94 and �98 to
+94, produced further decrease in activity to almost the
basal levels in EA.hy926 and SH-SY5Y, but not as much
in HOG cells (Fig. 5). Since the basal activity (luciferase
activity obtained after transfecting pGL3Basic) in the
HOG cells was very low, the activity of the �98 to +94
fragment in these cells was still significant in this cell
line. Probably, the transcription factors present in these
glioma cells yield transcriptional activity by binding to
the GC-rich region present downstream of the �98
position (Fig. 2). In general, the ATPase II promoter or
its fragments showed some difference in activity between
Table 5
The presence of the 15-bp repeat in the ATPase II promoter variant II results in
Transcription factor Strand
CPBP (+)
EGR1 (�)
Myc associated zinc finger protein (MAZ) (�)
Myc-associated zinc finger protein related TF (�)
The presence of the 15-bp repeat in the ATPase II promoter variant II introduces
identified by MatInspector.a Position within ATPase(+) (Variant 2) promoter relative to TSS (+1).
the EA.hy926 and the neural cells, SH-SY5Y and HOG,
but from experiments performed only in these three cell
lines, we did not observe any dramatic cell type-
specificity in the shortest fragment �291 to +94 that
showed full promoter activity.
3.6. Possible enhancer/repressor activity in the promoter
Neural selectivity of expression is often conferred by a
21-bp neural-restrictive silencer element (NRSE), which
binds to the neuron-restrictive silencer factor (NRSF), also
known as RE-1 silencing transcription factor (REST) [33].
NRSF is a 116-kDa C2H2 zinc finger protein that is
expressed ubiquitously in developing non-neural tissues
between embryonic days 8.5 and 9.5 [33]. At the onset of
neuronal differentiation, NRSF expression is down-regu-
lated, assuming a very low to negligible level in mature
neurons [33]. The consensus for NRSE is TTCAGCAC-
CACTTA GAGTGCC [33]. An analysis of the ¨1.2-kb
5V-flanking sequence of ATPase II revealed two homol-
ogous sequences in the ATPase II promoter from �721 to
�701 and from �427 to �407 (Fig. 2) [33]. Of these,
one NRSE sequence (�427 to �407) was found on the
(+) strand. Additionally, the upstream region of the 1.2-kb
5V-flanking sequence analyzed here also harbors one
NFjB-specific site (Fig. 2). Despite such possibility of
transcriptional regulation by the NRSE and NFjBsequences, luciferase reporter analysis by including the
NRSE at �427 to �407 and comparison with the activity
of the complete ¨1.2-kb 5V flanking sequence (presented
in Fig. 5) showed that the deletion of both NRSE and
NFjB binding sites had no effect on the neural cell-
specificity of the promoter (Fig. 6). However, it should be
noted that none of our experiments involved growth
factor treatment of cells to cause the activation of NFjB.Therefore, the NFjB sequence may still have a strong
effect on ATPase II expression in specific cell types,
which can be stimulated by growth factors or ligands that
cause signaling activity leading to the activation of NFjB.The additional transcription factor binding sites introduced
in the 15-bp direct repeat had no effect on the activity of
the ATPase II promoter or its fragments (Fig. 6). It
should be noted again that to confirm the effect (or lack
of effect) of such additional enhancer elements, the
additional TF binding sites
Sequence Positiona
CGCCGAGCCGCCCCTCCGCGGCT �126/�104
CCGCGGAGGGGCGGCTC �122/�106
CGCGGAGGGGCGG �119/�107
CGCGGAGGGGCGG �119/�107
additional TF binding sites. The additional sites present in Variant 2 were

A.
B.+1
PromoVar1: GTGCGGGGGC CGCGCCCGGC GGCGGCTCTG CCCTAGGTGG GCGGCGGCGC GGCCCAGGCT GCAGCTGAGC GCTCTGCGCG GCGCAGCCGG GTCTCCCGCG 1328/+73Clone1: 1GGCGGCTCTG CCCTAGGTGG GCGGCGGCGC GGCCCAGGCT GCAGCTGAGC GCTCTGCGCG GCGCAGCCGG GTCTCCCGCG 80Clones2-5: 1 CTG CCCTAGGTGG GCGGCGGCGC GGCCCAGGCT GCAGCTGAGC GCTCTGCGCG GCGCAGCCGG GTCTCCCGCG 73Clone6: 1 CTG CCCTAGGTGG GCGGCGGCGC GGCCCAGGCT GCAGCTGAGC GCTCTGCGCG GCGCAGCCGG GTCTCCCGCG 73Clone7: 1 G CCCTAGGTGG GCGGCGGCGC GGCCCAGGCT GCAGCTGAGC GCTCTGCGCG GCGCAGCCGG GTCTCCCGCG 71Clones8,9: 1 CTCCCGCG 8
(Coding sequence…)PromoVar1: TGTACCACGC CGTGACAGGT GCAGAGTCCG GGCTGAGGAC CCACCTGCAG CCGCCGCCGC GATGCCCACC ATGCGGAGGA CCGTGTCGGA GATCCGCTC- 1427/+172Clone1: TGTACCACGC CGTGACAGGT GCAGAGTCCG GGCTGAGGAC CCACCTGCAG CCGCCGCCGC GATGCCCACC ATGCGGAGGA CCGTGTCGGA GATCCGCTCG 180Clones2-5: TGTACCACGC CGTGACAGGT GCAGAGTCCG GGCTGAGGAC CCACCTGCAG CCGCCGCCGC GATGCCCACC ATGCGGAGGA CCGTGTCGGA GATCCGCTCG 173Clone6: TGTACCACGC CGTGACAGGT GCAGAGTCCG GGCTGAGGAC CCACCTGCAG CCGCCGCCGC GATGCCCACC ATGCGGAGGA CCGTGTCGGA GATCCGCTCG 173Clone7: TGTACCACGC CGTGACAGGT GCAGAGTCCG GGCTGAGGAC CCACCTGCAG CCGCCGCCGC GATGCCCACC ATGCGGAGGA CCGTGTCGGA GATCCGCTCG 171Clones8,9: TGTACCACGC CGTGACAGGT GCAGAGTCCG GGCTGAGGAC CCACCTGCAG CCGCCGCCGC GATGCCCACC ATGCGGAGGA CCGTGTCGGA GATCCGCTCG 108
350 bp
250 bp
300 bp
50bp
ladd
er
BrainSke
letal
muscle
Fig. 4. RLM-RACE analysis of the ATPase II promoter shows that the major transcription start site in the brain and skeletal muscle is 143-bp 5V of thetranslation initiation site. (A) Secondary 5VRACE products obtained using GeneRaceri 5V Nested Forward and 5RNestedGSP (Table 5) are shown. In the
brain, there was a major band at ¨300 bp and a few bands with lesser molecular weights. The skeletal muscle showed one major band that moves slightly faster
than 300 bp. (B) The major band at ¨300 bp in the brain corresponded to products from two closely spaced transcription start sites at – 150G (clone 1), – 143C
(clone 6) and – 141G (clone 7) with respect to the translation initiation codon. A faster-moving band at ¨230 bp corresponded to a transcription start site at – 71C
(clones 8 and 9). The skeletal muscle samples showed one major band below 300 bp, which corresponded to the same transcription start site at – 143C (clones
2–5) as observed in the brain. The promoter region (Variant 1, AY775564) shown corresponds to bases 1229 to 1427 (�27 to +172 relative to a transcription
start site; Fig. 2). Even though all the clones were obtained with 5RNestedGSP (Table 1) as a reverse primer, only the 5V part of the sequences of the clonescorresponding to the promoter region indicated is shown on the figure. The location of the coding sequence within the ATPase II gene is indicated.
T. Sobocki et al. / Biochimica et Biophysica Acta 1728 (2005) 186–198194
promoter activity has to be tested in many more cell
types.
4. Discussion
Many P-type ATPases have been identified and their
coding sequences identified. However, the promoter
sequence of only one of these genes (the Wilson disease
gene-WD gene) has been identified [20]. Although some of
the ATPase genes are believed to be putative amino-
phospholipid translocases, our previous study showed for
the first time that the overexpression of the cDNA for such a
P-type ATPase (the ATPase II cDNA) causes an increase in
aminophospholipid translocase activity [13]. Therefore,
such studies are likely to shed new light on the identity
and expression of the enigmatic aminophospholipid trans-
locase enzyme. As discussed earlier, further studies will
include testing the effect of signaling pathways that are
known to activate transcription factors specific for number
of the enhancer/repressor elements identified on the ATPase
II promoter. Such studies in a wider variety of cell lines
would reveal the expression profile of the ATPase II gene,
which is likely to mimic its expression in various human cell
types.
Earlier studies on the ATPase II gene reported its cDNA
sequences from bovine, mouse, and human [7,11,12].
However, the intron–exon organization of the gene had
not been delineated in these reports. Our analysis has
yielded the complete organization of the gene at the locus
4p13 on the human genome (Fig. 1 and Table 1). ATPase II
appears to be a large gene with 37 exons (Fig. 1 and Table
1). Interestingly, both the WD gene promoter and the
ATPase II promoter lack a TATA box and contain GC-rich
regions harboring multiple transcription factor binding
sequences. The promoter region on the WD gene is,
however, located further upstream (¨�700 bp) from the
translation-start site than the promoter of the ATPase II gene
(¨�250 bp). Furthermore, unlike the WD gene promoter,
which contains a repressor element, deletion analysis of the
ATPase II promoter indicated the lack of such repressor
elements. However, repressor elements could still exist
either upstream or downstream of the (¨1.2-kb sequence
analyzed in this study).
The ATPase II message levels show dramatic variation in
different cell types [6,12]. Based on such observations, we
planned to investigate the structure and role of the ATPase II
promoter. Although the overall activity of both the full and
core promoters of this gene show similar levels of activity in
three different cell types (Fig. 5), the deletions of the core
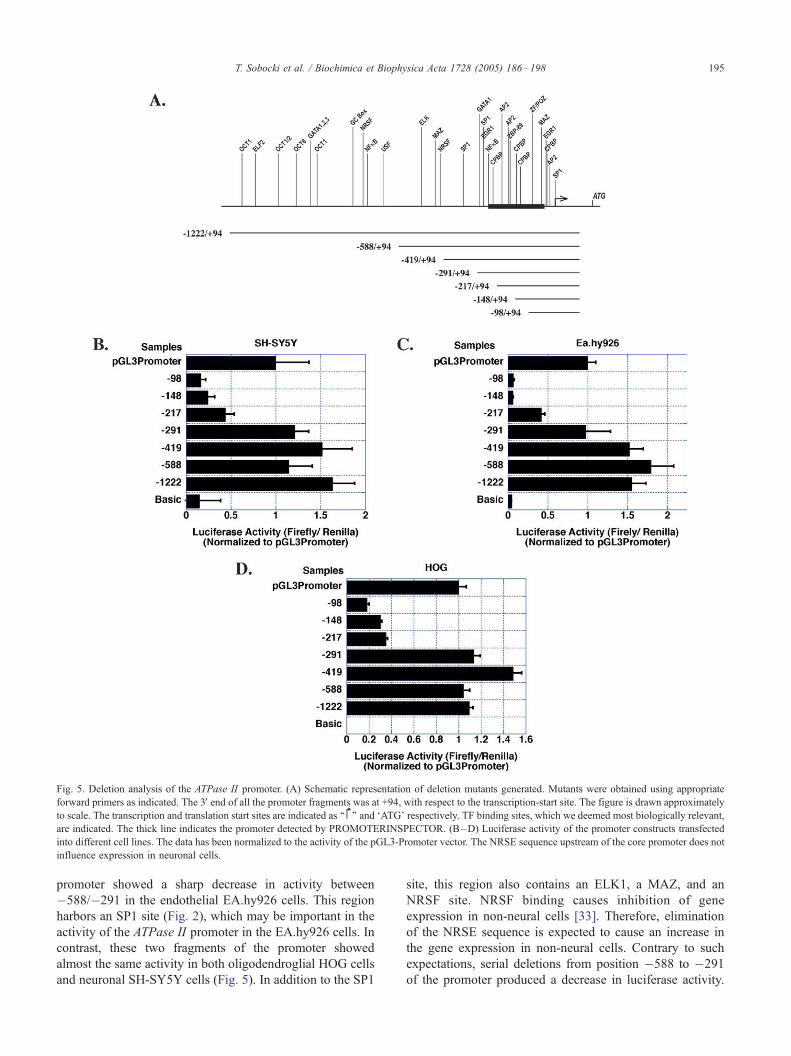
Fig. 5. Deletion analysis of the ATPase II promoter. (A) Schematic representation of deletion mutants generated. Mutants were obtained using appropriate
forward primers as indicated. The 3V end of all the promoter fragments was at +94, with respect to the transcription-start site. The figure is drawn approximately
to scale. The transcription and translation start sites are indicated as ‘‘�’’ and FATG_ respectively. TF binding sites, which we deemed most biologically relevant,
are indicated. The thick line indicates the promoter detected by PROMOTERINSPECTOR. (B–D) Luciferase activity of the promoter constructs transfected
into different cell lines. The data has been normalized to the activity of the pGL3-Promoter vector. The NRSE sequence upstream of the core promoter does not
influence expression in neuronal cells.
T. Sobocki et al. / Biochimica et Biophysica Acta 1728 (2005) 186–198 195
promoter showed a sharp decrease in activity between
�588/�291 in the endothelial EA.hy926 cells. This region
harbors an SP1 site (Fig. 2), which may be important in the
activity of the ATPase II promoter in the EA.hy926 cells. In
contrast, these two fragments of the promoter showed
almost the same activity in both oligodendroglial HOG cells
and neuronal SH-SY5Y cells (Fig. 5). In addition to the SP1
site, this region also contains an ELK1, a MAZ, and an
NRSF site. NRSF binding causes inhibition of gene
expression in non-neural cells [33]. Therefore, elimination
of the NRSE sequence is expected to cause an increase in
the gene expression in non-neural cells. Contrary to such
expectations, serial deletions from position �588 to �291
of the promoter produced a decrease in luciferase activity.

Fig. 6. Deletion analysis of the ATPase II promoter: the 15-bp insert does not alter promoter activity. Mutants were obtained using appropriate forward primers
as indicated. After cloning, the mutants were tested for the presence of the 15-bp repeat. The 3V end of all the promoter fragments was at +94, with respect to the
transcription-start site. Panels A–C: Luciferase activity of the promoter constructs transfected into different cell lines. The data were normalized to the activity
of the pGL3-Promoter vector (Promega, CA). The inclusion of the NRSE sequence in the promoter fragments did not influence expression in neuronal cells.
There was no significant difference in the activity of the promoter fragments with and without the insert.
T. Sobocki et al. / Biochimica et Biophysica Acta 1728 (2005) 186–198196
Therefore, the NRSE site does not play an important role in
the activity of the ATPase II promoter in the EA.hy926
cells. Further deletion beyond �217 (�148 and �98)
caused a suppression of promoter activity to a minimum
in all the three cell types. Intriguingly, the minimum activity
of the deleted promoter was still significantly higher than
that of the empty vector (pGL3Basic) in the HOG cells but
not in the other two cell lines. This could be due to the
presence of an EGR1, a Zinc finger/POZ, and a core
promoter binding protein (CPBP) sites present in this short
fragment of the promoter (Fig. 2 and Table 2). The presence
of three CPBP sites in the region upstream of the
transcription start site is typical for a TATA-less promoter
and strongly supports the results of MatInspector analysis
suggesting the presence of a core promoter between �491
and +94.
One 15-base pair direct repeat and several single base-
pair changes were observed in the clones of the ATPase
II promoter (Figs. 2 and 3). The 15-base pair repeat
occurs in a GC-rich region within the core promoter and
thus contains several additional transcription factor-bind-
ing sites including a CPBP and an EGR1 sites (Fig. 3b
and c). Thus, transcription factors could bind to this
repeat to cause regulation of gene expression. However,
in the three cell lines tested, this repeat sequence did not
produce any change in the activity of the promoter
fragments (Fig. 6). Nonetheless, only by testing such
promoter variations in more cell types, and under
conditions that affect the activity of these transcription
factors, we would be able to understand the effect of such
sequence variations in the actual expression of the
ATPase II gene in human cells.
Like ATPase II, another similar protein, ATPase 1B [12],
is also believed to harbor lipid-translocating activity that
could be crucial in the phagocytic uptake of apoptotic cells.
It has been reported that this gene is highly expressed in the
brain [6,12]. As opposed to our earlier expectation that the
ATPase II promoter is neuron-cell specific, our data shows
that this promoter may not have any dramatic cell type-
specificity. Therefore, among other likely candidates,

T. Sobocki et al. / Biochimica et Biophysica Acta 1728 (2005) 186–198 197
ATPase 1B protein could be the main regulator of
phagocytosis in the brain. In our earlier studies, we had
expressed the ATPase II cDNA in neuronal cells to show an
increase in PS internalization in these cells [13]. Such
studies should also be performed with the ATPase 1B cDNA
and a thorough analysis of the ATPase 1B promoter and its
possible neural-cell specificity are required.
The P-type ATPases have received much attention in
recent years. The involvement of these proteins in multiple
drug resistance, Wilson’s disease, and familial Cholestasis
has been established in many studies. The genes for such
physiologically important proteins often undergo regulated
expression to help maintain appropriate levels of the gene
products. Moreover, during apoptosis, many other genes and
their products are regulated to effect signature changes that
are observed in apoptotic cells. As discussed earlier, the
overexpression of ATPase II in the hybrid neuroblastoma cell
line HN2 causes an increase in APTL activity [13]. Our
earlier studies have also shown that apoptosis is associated
with an inhibition of APTL and externalization of PS [19].
Because of this correlation, it could be expected that ATPase
II could be linked to the expression and regulation of
apoptosis-associated proteins. Thus, there is an acute need
for a thorough analysis of ATPase II expression in various cell
types under different conditions of trophic support or stress.
Results reported here lay the groundwork for such future
experiments, which will help delineate the role of ATPase II.
Acknowledgment
FJ and RD were supported by funds from the New
York Louis Stokes Alliance for Minority Participation
Program, MAGNET/AGEP, and NIH minority supple-
ments. This project was supported by a grant from the
NIH (CA77803-03).
References
[1] V.A. Fadok, D.R. Voelker, P.A. Campbell, J.J. Cohen, D.L. Bratton,
P.M. Hensen, Exposure of phosphatidylserine on the surface of
apoptotic lymphocytes triggers specific recognition and removal by
macrophages, J. Immunol. 148 (1992) 2207–2216.
[2] S.J. Martin, D.M. Finucane, G.P. Amarante-Mendes, G.A. O’Brien,
D.R. Green, Phosphatidylserine externalization during CD95-induced
apoptosis of cells and cytoplasts requires ICE/CED-3 protease activity,
J. Biol. Chem. 271 (1996) 28753–28756.
[3] T. Adayev, R. Estephan, S. Meserole, B. Mazza, E.J. Yurkow, P.
Banerjee, Externalization of phosphatidylserine may not be an early
signal of apoptosis in neuronal cells, but only the phosphatidylserine-
displaying apoptotic cells are phagocytosed by microglia, J. Neuro-
chem. 71 (1998) 1854–1864.
[4] R.F.A. Zwaal, R.A. Schroit, Pathophysiologic implications of
membrane phospholipid asymmetry in blood cells, Blood 89 (1997)
1121–1132.
[5] B. Verhoven, S. Krahling, R.A. Schlegel, P. Williamson, Regulation of
phosphatidylserine exposure and phagocytosis of apoptotic T lym-
phocytes, J. Cell Death Differ. 6 (1999) 262–270.
[6] M.S. Halleck, D. Pradhan, C. Blackman, C. Berkes, P.
Williamson, R.A. Schlegel, Differential expression of putative
transbilayer amphipath transporters, Physiol. Genomics 1 (1999)
139–150.
[7] X. Tang, M.S. Halleck, R.A. Schlegel, P. Williamson, A subfamily of
P-Type ATPases with aminophospholipid transporting activity, Sci-
ence 272 (1996) 1495–1497.
[8] R.F.A. Tilly, J.M.G. Senden, P. Comfurius, E.M. Bevers, R.F.A.
Zwaal, Increased aminophospholipid translocase activity in human
platelets during secretion, Biochim. Biophys. Acta 1029 (1990)
188–190.
[9] D.L. Daleke, W.H. Heustis, Incorporation and translocation of
aminophospholipids in human erythrocytes, Biochemistry 24 (1985)
5406–5416.
[10] M. Bitbol, P. Fellmann, A. Zachowski, P.F. Devaux, Ion regulation of
phosphatidylserine and phosphatidylethanolamine outside-inside
translocation in human erythrocytes, Biochim. Biophys. Acta 904
(1987) 268–282.
[11] I. Mouro, M.S. Halleck, R.A. Schlegel, M.M. Genevieve, P.
Williamson, A. Zachowski, P. Devaux, J. Cartron, Y. Colin, Cloning,
expression, and chromosomal mapping of a human ATPase 2 gene,
member of the third subfamily of P-Type ATPases and Orthologous to
the presumed bovine and murine aminophospholipid translocase, J.
Biol. Chem. 257 (1999) 333–339.
[12] M.S. Halleck, D. Pradhan, C. Blackman, C. Berkes, P. Williamson,
R.A. Schlegel, Multiple members of a third subfamily of P-Type
ATPases identified by genomic sequences and ESTs, Genome Res. 8
(1998) 354–361.
[13] G. Chin, Y. El-Sherif, F. Jayman, R. Estephan, A. Wieraszko, P.
Banerjee, Appearance of voltage-gated calcium channels following
overexpression of ATPase II cDNA in neuronal HN2 cells, Mol. Brain
Res. 117 (2003) 109–115.
[14] R. Tsivkovskii, J.F. Eisses, J.H. Kaplan, S. Lutsenko, Functional
properties of the copper-transporting ATPase ATP7B (the Wilson’s
Disease Protein) expressed in insect cells, J. Biol. Chem. 277 (2002)
976–983.
[15] W. Kuhlbrandt, Biology, structure and mechanism of P-type ATPases,
Nat. Rev. 5 (2004) 282–295.
[16] P.L. Pederson, E. Carafoli, Ion motive ATPases: I. Ubiquity, proper-
ties, and significance to cell function, Trends Biochem. Sci. 12 (1987)
146–150.
[17] J. Ding, Z. Wu, B.P. Crider, Y. Ma, X. Li, C. Slaughter, L. Gong, X.-S.
Xie, Identification and functional expression of four isoforms of
ATPase II, the putative aminophospholipid translocase, J. Biol. Chem.
275 (2000) 23378–23386.
[18] L.N. Bull, M.J.T. van Eijk, L. Pawlikowska, J.A. De Young, J.A.
Juijn, M. Liao, L.W.J. Klomp, N. Lomri, R. Berger, B.F. Scharsch-
midt, A.S. Knisely, R.H.J. Houwen, N.B. Freimer, A gene encoding a
P-type ATPase mutated in two forms of hereditary cholestasis, Nat.
Genet. 18 (1998) 219–224.
[19] P. Das, R. Estephan, P. Banerjee, Apoptosis is associated with an
inhibition of aminophospholipid translocase (APTL) in CNS-derived
HN2-5 and HOG cells and phosphatidylserine is a recognition
molecule in microglial uptake of the apoptotic HN2-5 cells, Life Sci.
72 (2003) 2617–2627.
[20] W.-J. Oh, E.K. Kim, K.D. Park, S.H. Hahn, O.J. Yoo, Cloning and
characterization of the promoter region of the Wilson disease gene,
Biochem. Biophys. Res. Commun. 259 (1999) 206–211.
[21] W.J. Kent, BLAT—The BLAST-like alignment tool, Genome Res. 12
(2002) 656–664.
[22] W.J. Kent, C.W. Sugnet, T.S. Furey, K.M. Roskin, K.M. Roskin, T.H.
Pringle, A.M. Zahler, D. Haussler, The human genome browser at
UCSC, Genome Res. 12 (2002) 996–1006.
[23] J.D. Thompson, D.G. Higgins, T.J. Gibson, CLUSTALW: improving
the sensitivity of progressive multiple sequence alignment through
sequence weighting, position-specific gap penalties and weight matrix
choice, Nucleic Acids Res. 22 (1994) 4673–4680.

T. Sobocki et al. / Biochimica et Biophysica Acta 1728 (2005) 186–198198
[24] R.F. Smith, B.A. Wiese, M.K. Wojzynski, D.B. Davison, K.C. Worley,
BCM Search Launcher—an integrated interface to molecular biology
data base search and analysis services available on the World Wide
Web, Genome Res. 6 (1996) 454–462.
[25] M. Scherf, A. Klingenhoff, T. Werner, Highly specific localization of
promoter regions in large genomic sequences by PromoterInspector: a
novel context analysis approach, J. Mol. Biol. 297 (2000) 599–606.
[26] K. Quandt, K. Frech, H. Karas, E. Wingender, T. Werner, MatInd and
MatInspector: new fast and versatile tools for detection of consensus
matches in nucleotide sequence data, Nucleic Acids Res. 23 (1995)
4878–4884.
[27] M. Gardiner-Garden, M. Frommer, CpG islands in vertebrate
genomes, J. Mol. Biol. 196 (1987) 261–282.
[28] C.D. Novina, A.L. Roy, Core promoters and transcriptional control,
Trends Genet. 12 (1996) 351–355.
[29] R.J. Gumina, N.E. Kirschbaum, K. Piotrowski, P.J. Newman, Charac-
terization of the human platelet/endothelial cell adhesion molecule-1
promoter: identification of a GATA-2 binding element required for
optimal transcriptional activity, Blood 89 (1997) 1260–1269.
[30] P.J. Cowan, D. Tsang, C.M. Pedic, L.R. Abbott, T.A. Shinkel, A.J.
d’Apice, M.J. Pearse, The human ICAM-2 promoter is endothelial
cell-specific in vitro and in vivo and contains critical Sp1 and GATA
binding sites, J. Biol. Chem. 273 (1998) 11737–11744.
[31] J. Lu, W. Lee, C. Jiang, E.B. Keller, Start site selection Sp1 in the
TATA-less human Ha-ras promoter, J. Biol. Chem. 269 (1994)
5391–5402.
[32] C. Muller, R. Yang, L. Beck-von-Peccoz, G. Idos, W. Verbeek, H.P.
Koefffler, Cloning of the cyclin A1 genomic structure and character-
ization of the promoter region. GC boxes are essential for cell cycle-
regulated transcription of the cyclin A1 gene, J. Biol. Chem. 274
(1999) 11220–11228.
[33] C.J. Schoenherr, A.J. Paquette, D.J. Anderson, Identification of
potential target genes for the neuron-restrictive silencer factor, Proc.
Natl. Acad. Sci. 93 (1996) 9881–9886.