
Isolation and gene expression of haematopoietic-cell-free preparations of highly purified murine...
9
Original Full Length Article Isolation and gene expression of haematopoietic-cell-free preparations of highly purified murine osteocytes Ling Yeong Chia, Nicole C. Walsh, T. John Martin, Natalie A. Sims ⁎ St. Vincent's Institute of Medical Research, Fitzroy, Victoria, Australia Department of Medicine at St. Vincent's Hospital, The University of Melbourne, Fitzroy, Victoria, Australia abstract article info Article history: Received 14 August 2014 Revised 14 October 2014 Accepted 11 November 2014 Available online 15 November 2014 Edited by: Mark Johnson Keywords: Osteocyte Osteoclast To define their gene expression and function, osteocytes are commonly isolated and purified by fluorescence- activated cell sorting (FACS) from mice expressing GFP directed by the dentin matrix protein 1 (Dmp1) promoter (DMP1-GFP). These cells express mRNA for osteocyte genes, including sclerostin (Sost) and Dmp1, and genes associated with the osteoclast phenotype: Dcstamp, Oscar, Cathepsin K (Ctsk), tartrate resistant acid phosphatase (TRAP/Acp5) and calcitonin receptor (Calcr). This suggests either that osteoclasts and osteocytes share genes and functions or that DMP1-GFP + preparations contain haematopoietic osteoclasts. To resolve this we stained DMP1-GFP cells for haematopoietic lineage (Lin) surface markers (CD2, CD3e, CD4, CD45, CD5, CD8, CD11b, B220, Gr1, Ter119) and CD31. Lin - CD31 - (Lin - ) and Lin + CD31 + (Lin + ) populations were analysed for GFP, and the four resulting populations assessed by quantitative real-time PCR. Lin - GFP + cells expressed mRNAs for Sost, Dmp1, and Mepe, confirming their osteocyte identity. Dcstamp and Oscar mRNAs were restricted to haematopoietic (Lin + ) cells, but Calcr, Ctsk and Acp5 were readily detected in pu- rified osteocytes (Lin - GFP + ). The capacity of these purified osteocytes to support osteoclastogenesis was assessed: no TRAP+ cells with N 2 nuclei were formed when purified osteocytes were cultured with bone mar- row macrophages and stimulated with 1,25-dihydroxyvitamin-D 3 /prostaglandin E 2 . Lin - GFP + osteocytes also expressed lower levels of Tnfsf11 (RANKL) mRNA than the osteoblast-enriched population (Lin - GFP - ). This demonstrates the importance of haematopoietic depletion in generating highly purified osteocytes and shows that osteocytes express Acp5, Ctsk and Calcr, but not other osteoclast markers, and do not fully support osteoclast formation in vitro. © 2014 Elsevier Inc. All rights reserved. Introduction Osteocytes are terminally differentiated osteoblasts and constitute more than 95% of all bone cells in the adult skeleton [1]. They reside in lacunae, small cavities within the mineralised bone matrix, and possess long, dendrite-like processes. These extend through small tunnels in the bone matrix (canaliculi) thereby connecting osteocytes with each other and with cells on the bone surface, including osteoblasts and bone lining cells [1,2]. This forms a well-organised network suitable for integration of local and systemic signals. In recent years, osteocytes have been re- ported to play significant paracrine and endocrine roles that regulate bone formation [3,4], phosphate metabolism [5–7], and osteoclast dif- ferentiation [8–12]. Despite their abundance, these cells remain poorly understood, mainly due to their position, deeply embedded in mineralised bone, and the difficulty of isolating highly purified primary cells. Several methods to isolate a highly purified osteocyte population have been described. Initial efforts using cells digested from embryonic chick calvaria purified them using a monoclonal antibody to the osteocytic protein, phosphate regulating endopeptidase homolog, X-linked (Phex) [13,14]. Osteocytes were also collected from serial digestion of newborn rat calvariae [15] or outgrowth of cells from previ- ously digested bone [16]. Both methods generated a population of oste- ocytes that showed typical stellate shape in culture, that produced low levels of alkaline phosphatase [15,17] and high levels of extracellular matrix proteins including osteocalcin [15,18], osteonectin [18] and osteopontin [18]. However, these cultures were impure and contami- nating cells, including fibroblasts, proliferated and overgrew the post mitotic osteocytes [15,17]. The more recent identification of several osteocytic gene markers such as Dmp1, Mepe, Phex, podoplanin (Pdpn/E11) and Sost indicated that late digest fractions contain higher levels of osteocyte genes includ- ing Dmp1 and Sost compared to earlier osteoblast-enriched fractions [19], allowing isolation of cells including a high proportion of osteo- cytes. Stern et al. [20] also successfully isolated primary cells displaying several characteristics of osteocytes from mice by combining serial Bone 72 (2015) 34–42 ⁎ Corresponding author at: St. Vincent's Institute of Medical Research, 9 Princes St., Fitzroy, Victoria 3065, Australia. E-mail address: [email protected] (N.A. Sims). http://dx.doi.org/10.1016/j.bone.2014.11.005 8756-3282/© 2014 Elsevier Inc. All rights reserved. Contents lists available at ScienceDirect Bone journal homepage: www.elsevier.com/locate/bone
Transcript of Isolation and gene expression of haematopoietic-cell-free preparations of highly purified murine...

Bone 72 (2015) 34–42
Contents lists available at ScienceDirect
Bone
j ourna l homepage: www.e lsev ie r .com/ locate /bone
Original Full Length Article
Isolation and gene expression of haematopoietic-cell-free preparations ofhighly purified murine osteocytes
Ling Yeong Chia, Nicole C. Walsh, T. John Martin, Natalie A. Sims ⁎St. Vincent's Institute of Medical Research, Fitzroy, Victoria, AustraliaDepartment of Medicine at St. Vincent's Hospital, The University of Melbourne, Fitzroy, Victoria, Australia
⁎ Corresponding author at: St. Vincent's Institute of MFitzroy, Victoria 3065, Australia.
E-mail address: [email protected] (N.A. Sims).
http://dx.doi.org/10.1016/j.bone.2014.11.0058756-3282/© 2014 Elsevier Inc. All rights reserved.
a b s t r a c t
a r t i c l e i n f oArticle history:Received 14 August 2014Revised 14 October 2014Accepted 11 November 2014Available online 15 November 2014
Edited by: Mark Johnson
Keywords:OsteocyteOsteoclast
To define their gene expression and function, osteocytes are commonly isolated and purified by fluorescence-activated cell sorting (FACS) frommice expressing GFP directed by the dentinmatrix protein 1 (Dmp1) promoter(DMP1-GFP). These cells express mRNA for osteocyte genes, including sclerostin (Sost) and Dmp1, and genesassociated with the osteoclast phenotype:Dcstamp, Oscar, Cathepsin K (Ctsk), tartrate resistant acid phosphatase(TRAP/Acp5) and calcitonin receptor (Calcr). This suggests either that osteoclasts and osteocytes share genes andfunctions or that DMP1-GFP+ preparations contain haematopoietic osteoclasts.To resolve this we stained DMP1-GFP cells for haematopoietic lineage (Lin) surface markers (CD2, CD3e, CD4,CD45, CD5, CD8, CD11b, B220, Gr1, Ter119) and CD31. Lin−CD31− (Lin−) and Lin+CD31+ (Lin+) populationswere analysed for GFP, and the four resulting populations assessed by quantitative real-time PCR.Lin−GFP+ cells expressed mRNAs for Sost, Dmp1, and Mepe, confirming their osteocyte identity. Dcstamp andOscarmRNAswere restricted to haematopoietic (Lin+) cells, but Calcr, Ctsk and Acp5were readily detected in pu-rified osteocytes (Lin−GFP+). The capacity of these purified osteocytes to support osteoclastogenesis wasassessed: no TRAP+ cells with N2 nuclei were formed when purified osteocytes were cultured with bone mar-row macrophages and stimulated with 1,25-dihydroxyvitamin-D3/prostaglandin E2. Lin−GFP+ osteocytes alsoexpressed lower levels of Tnfsf11 (RANKL) mRNA than the osteoblast-enriched population (Lin−GFP−).This demonstrates the importance of haematopoietic depletion in generating highly purified osteocytes andshows that osteocytes express Acp5, Ctsk and Calcr, but not other osteoclast markers, and do not fully supportosteoclast formation in vitro.
© 2014 Elsevier Inc. All rights reserved.
Introduction
Osteocytes are terminally differentiated osteoblasts and constitutemore than 95% of all bone cells in the adult skeleton [1]. They reside inlacunae, small cavities within the mineralised bonematrix, and possesslong, dendrite-like processes. These extend through small tunnels in thebonematrix (canaliculi) thereby connecting osteocytes with each otherandwith cells on the bone surface, including osteoblasts and bone liningcells [1,2]. This forms a well-organised network suitable for integrationof local and systemic signals. In recent years, osteocytes have been re-ported to play significant paracrine and endocrine roles that regulatebone formation [3,4], phosphate metabolism [5–7], and osteoclast dif-ferentiation [8–12]. Despite their abundance, these cells remain poorlyunderstood, mainly due to their position, deeply embedded inmineralised bone, and the difficulty of isolating highly purified primarycells.
edical Research, 9 Princes St.,
Several methods to isolate a highly purified osteocyte populationhave been described. Initial efforts using cells digested from embryonicchick calvaria purified them using a monoclonal antibody to theosteocytic protein, phosphate regulating endopeptidase homolog,X-linked (Phex) [13,14]. Osteocytes were also collected from serialdigestion of newborn rat calvariae [15] or outgrowth of cells from previ-ously digested bone [16]. Bothmethods generated a population of oste-ocytes that showed typical stellate shape in culture, that produced lowlevels of alkaline phosphatase [15,17] and high levels of extracellularmatrix proteins including osteocalcin [15,18], osteonectin [18] andosteopontin [18]. However, these cultures were impure and contami-nating cells, including fibroblasts, proliferated and overgrew the postmitotic osteocytes [15,17].
The more recent identification of several osteocytic gene markerssuch as Dmp1, Mepe, Phex, podoplanin (Pdpn/E11) and Sost indicatedthat late digest fractions contain higher levels of osteocyte genes includ-ing Dmp1 and Sost compared to earlier osteoblast-enriched fractions[19], allowing isolation of cells including a high proportion of osteo-cytes. Stern et al. [20] also successfully isolated primary cells displayingseveral characteristics of osteocytes from mice by combining serial

35L.Y. Chia et al. / Bone 72 (2015) 34–42
digestions with tissue homogenisation. In their studies, the late digestfraction expressed several osteocytic genes, including Pdpn, Sost, Mepe,Dmp1 and Phex [20]. The possibility of contaminating proliferating cellssuch as fibroblasts in the cell preparation remained an open question.
Based on the restricted expression of Dmp1 in osteocytes [21,22],Kalajzic et al. [22] used 8 kb of the Dmp1 promoter region to generatea transgenic mouse line with GFP expression directed to osteocytes inosteoid and bone [22]. Subsequently, cells sorted for GFP from calvarialbones of neonatal DMP1-GFP mice have been used to examineosteocytic gene expression compared to osteoblasts [19,23]. An alterna-tive approach to isolate osteocytes fromneonatal calvariae used fluores-cent labelling directed by a 10 kb-promoter region driven DMP1-Cretransgene [8]. However, expression of GFP driven by the 10 kb DMP1-Cre transgene also occurs in osteoblasts [12,24], perhaps providing anexplanation for recent contradicting data obtained when RANKL levelsare compared between osteocytes and osteoblasts prepared by thismethod [8] vs a comparison of early and late digest fractions [25].
We used the 8 kbDMP1-GFP transgenicmice to generate osteocytes.By FACS sorting for GFP only, we observed the presence of osteoclast-specific markers Dcstamp and OscarmRNA in the GFP positive popula-tion, consistent with previous reports of common genes expressed byosteocytes and haematopoietic osteoclasts including Ctsk [26], Acp5[26], Oscar, Dcstamp [23], Calcr [23,27,28] and Sost [29]. We developeda method to further purify this osteocyte population, utilising standardhaematopoietic-lineage and endothelial cell depletion methods [30–33]in order to determine the extent to which osteocyte gene expressionoverlaps with that of osteoclasts.
Materials and methods
Animal experiments
DMP1-GFPmice, that express green fluorescent protein (GFP) underthe control of 8 kb of the DMP1 promoter region plus 4439 bp of thefirstexon, first intron and part of the second exon [22], were obtained fromDr Ivo Kalajzic, University of Connecticut Health Science Center, via thecolony of Dr Hong Zhou, ANZAC Research Institute, Sydney. All animalexperiments were approved by the St. Vincent's Melbourne Health An-imal Ethics Committee.
Osteocyte isolation
To allow comparison with previously published work [8,19,23],calvariae were dissected from 4 to 9 day old DMP1-GFP transgenicmice and subjected to 7 sequential 15-minute digestions in a 2 ml mix-ture containing 4 mg dispase (Gibco, NY, USA) and 2 mg collagenasetype 2 (Worthington, Australia) as previously described [27]. Cell frac-tions 1–7 were collected, pooled, and resuspended in alpha modifiedEagle's medium (αMEM, Gibco) containing 10% FBS, and centrifuged.Pellets were resuspended in erythroid cell lysis buffer (150 mMNH4CI, 10 mM CHKO3, 0.1 mM Na2EDTA, pH 7.2) before cell sorting.
Fluorescence activated cell sorting (FACS) of green-fluorescent protein(GFP) positive osteocytes
Isolated cells were subjected to FACS for GFP expression alone orlineage depletion plus GFP as follows (Fig. 1). Prior to sorting, deadcells and debris were removed based on side scatter (SSC) area andforward scatter (FSC) area, and doublets were excluded based both onSSC width (W) vs SSC height (H) and on FSC-W vs FSC-H. Cells weresorted with excitation 488 nm and 530/30 or 530/40-emission filter forGFP on a BD FACSAria cell sorter (BD Bioscience) or a BD FACS Influxcell sorter (BDBioscience), respectively. For each preparation, calvarial di-gests from C57BL/6 mice were used to determine the GFP negative gate.
For haematopoietic lineage and endothelial cell depletion, cells werestained for 30 min with an antibody cocktail comprised of biotin-
labelled antibodies against mouse CD2, CD3e, CD4, CD45, CD5,CD8, CD11b, B220, Gr1, Ter119 (all eBioscience) to identify thehaematopoietic lineage population [30–33] and CD31 (eBioscience) toidentify endothelial cells [34]. None of these markers have been detect-ed by microarray in any mesenchymal cell (BioGPS: http://biogps.org;accessed 14 October 2014) or in neonatal osteocytes by in situ hybridi-zation (Gene eXpression Database: http://www.informatics.jax.org/mgihome/homepages/expression.shtml; accessed 14 October 2014).In the text that follows, the Lin+ population includes CD31+ cells, andthe Lin− population is also CD31−. Biotinylated antibodies weredetected with Qdot 605 streptavidin conjugate (Invitrogen) as previ-ously described [35]. Cells were then subjected to FACS with excitationat 405 nmand 610/20 emission filter for Qdot 605, and GFP as describedabove.
The following cell populations were isolated: Lin+GFP+ (fluorescenthaematopoietic cells), Lin+GFP− (haematopoietic lineage cells),Lin−GFP+ (highly purified osteocytes) and Lin−GFP− (mesenchymallineage cells, including osteoblasts) (Fig. 1).
cDNA preparation and PCR
cDNA was prepared from RNA using random hexamers (Promega,Australia) and Superscript III (Invitrogen, Australia) according to themanufacturer's protocol. Real timequantitative RT-PCR (qPCR)wasper-formed using SYBR green detection on a Stratagene Mx3000P(Invitrogen, Australia) as previously described [27]. Cycling conditionswere (95 °C for 10′), (95 °C for 15″, 60 °C for 1′) × 40 cycles. Sampleswere analysed using Stratagene softwareMxPro and reported using lin-ear ΔCT values normalised to the geometric average of Bms1 and Pgk1.Primers for genes of interest are as previously published: Calcr, Dmp1,Mepe, Sost and Bms1 [27,28]; Alpl, Pthr1 and Tnfsf11 [36]; Dcstamp andCtsk [37]; Tnfrsf11b [38]; and Bglap [39]; novel primers are listed inTable 1. Calcr transcript was confirmed by sequencing as follows: PCRproducts were cloned into pGEMT™T-A cloning vector (Promega) andtransformed into Stable 3 bacterial cells. Plasmid DNA was preparedusing Qiagen mini-prep kit (Qiagen) and cycle sequencing was per-formed by Applied Genetic Diagnostics (Department of Pathology, TheUniversity of Melbourne).
In vitro osteoclastogenesis
To generate bone marrow macrophages, mouse bone marrow wasisolated from C57BL/6 mice and cultured in L-cell conditioned mediafor 2 days. Non-adherent cells (bone marrow macrophages) (250,000cells/cm2) were used as osteoclast precursor cells, which werecocultured with osteoblasts (50,000 cells/cm2) or osteocytes (50,000cells/cm2) in the presence of 10−7 M 1,25-dihydroxyvitamin D3 and10−7 M prostaglandin E2 for 6–7 days as previously described [8].Cells were then fixed with 4% paraformaldehyde (Sigma-Aldrich) for10 min on ice and stained for tartrate-resistant acid phosphatase(TRAP) enzyme activity [40].
Results
GFP+ population generated by FACS for GFP without haematopoieticdepletion contains osteoclasts
DMP1-GFP+ calvarial cells obtained by FACS for GFP alone from 4 to9 day oldmice expressed the osteocytemarker, Sost at a three-fold higherlevel than GFP− calvarial cells, confirming enrichment of osteocytes inthis population (Fig. 2). Consistent with our previous finding, Calcrwasalso detected in this population [28] (Fig. 2). Testing for the presence ofother osteoclast markers, such as dendritic cell-specific transmembraneprotein (Dcstamp) and osteoclast-associated immunoglobulin-like recep-tor (Oscar), revealed their presence in both GFP+ and GFP− populations(Fig. 2). This indicated either that both osteocytes and osteoclasts express

GFP sort Lineage and GFP sortneage and
DMP1-GFP 4-9d old; n=30 and C57BL/6
7 x 15 min collagenase digestions
Lin+
Lin-Lin-
A B
Calvariae
Lin- GFP+
Lin- GFP-
Lin+ GFP+
Lin+ GFP-
Highly purified osteocytes
Osteoblast-enriched mesenchymal cells
Fluorescent haematopoietic & endothelial cells
Haematopoietic & endothelial cells
pre-sort: debris & doublet removal by 1. SSC-A vs FSC-A2. SSC-W vs SSC-H3. FSC-W vs FSC-H
0
104
103
102
105
<GFP
-A>
FSC-A0 100K 200K
0
104
103
102
105
FSC-A0 100K 200K
<Qdo
t 605
-A:L
in>
Haematopoietic & Endothelial
Mesenchymal
<GFP
-A>
<GFP
-A>
FSC-A
FSC-A
Fig. 1. Schematic of FACS purification of GFP positive osteocytes. Calvariae collected from 4 to 9 day old WT and DMP1-GFP pups were subjected to 7 sequential 15 minute digestions indispase and collagenase type 2. Cells from fractions 1–7 were pooled, and subjected to either (A) GFP sorting alone or (B) lineage depletion and GFP sorting to produce 4 fractions: TheLin+GFP+ fraction contains GFP+ haematopoietic cells, Lin+GFP− fraction contains other haematopoietic cells, Lin−GFP+ fraction contains highly purified osteocytes and Lin−GFP−
fraction contains other mesenchymal cells including osteoblasts.
36 L.Y. Chia et al. / Bone 72 (2015) 34–42
these marker genes, or that osteocyte preparations generated by GFPsorting alone contain contaminating osteoclasts or their precursors.
Haematopoietic lineage depletion of DMP1-GFP osteocyte preparation
Haematopoietic and endothelial cells were removed fromDMP1-GFPosteocyte preparations using lineage depletion followed by sorting forGFP by FACS; this provided 4 subsequent populations: Lin+GFP+ cells
Table 1Novel primer sequences used for PCR analysis of murine transcripts.
Protein Gene name NCBI RefSeq
Acid phosphatase, tartrate resistant Acp5 NM_007388Keratocan Kera NM_008438Matrix extracellular phosphoglycoprotein Mepe NM_053172Osteoclast associated receptor Oscar NM_175632Phosphoglycerate kinase 1 Pgk1 NM_008828Sclerostin Sost NM_024449
(fluorescent haematopoietic cells), Lin+GFP− cells (non-fluorescenthaematopoietic cells), Lin−GFP+ cells (GFP-expressing osteocytes), andLin−GFP− cells (non-fluorescent mesenchymal cells) (Fig. 1); note thatLin− cells are also CD31−. Excluding those cells that fell between the lin-eage andGFP gateswhichwere discarded, the proportion of isolated cellsin each population were Lin+GFP+: 0.33 ± 0.17%, Lin+GFP−: 38.52 ±6.97%, Lin−GFP+: 6.91 ± 1.86%, and Lin−GFP−: 54.20 ± 6.09%(mean ± SEM of 4 independent experiments). At the same time, we
Forward sequence (5′ to 3′) Reverse sequence (5′ to 3′)
GGGGACAATTTCTACTTCACTGG GCAAACGGTAGTAAGGGCTGTCCCCCATCAACTTATTTTAGC GGTTGCCATTACAGCACCTTAGCAAATGCCCAGAGACTAAGCCC TGAGGCCCTCTGGCGTCATTCAGTAACGGATCAGCTCCCCAG CGCGGTACAGTGCAAAACTCCCCGGCATTCTGCACGCTTCAA GGTCGAAAGGCCCGGAGATGACCACAGAGGTCATCCCA GACACATCTTTGGCGTCATAG
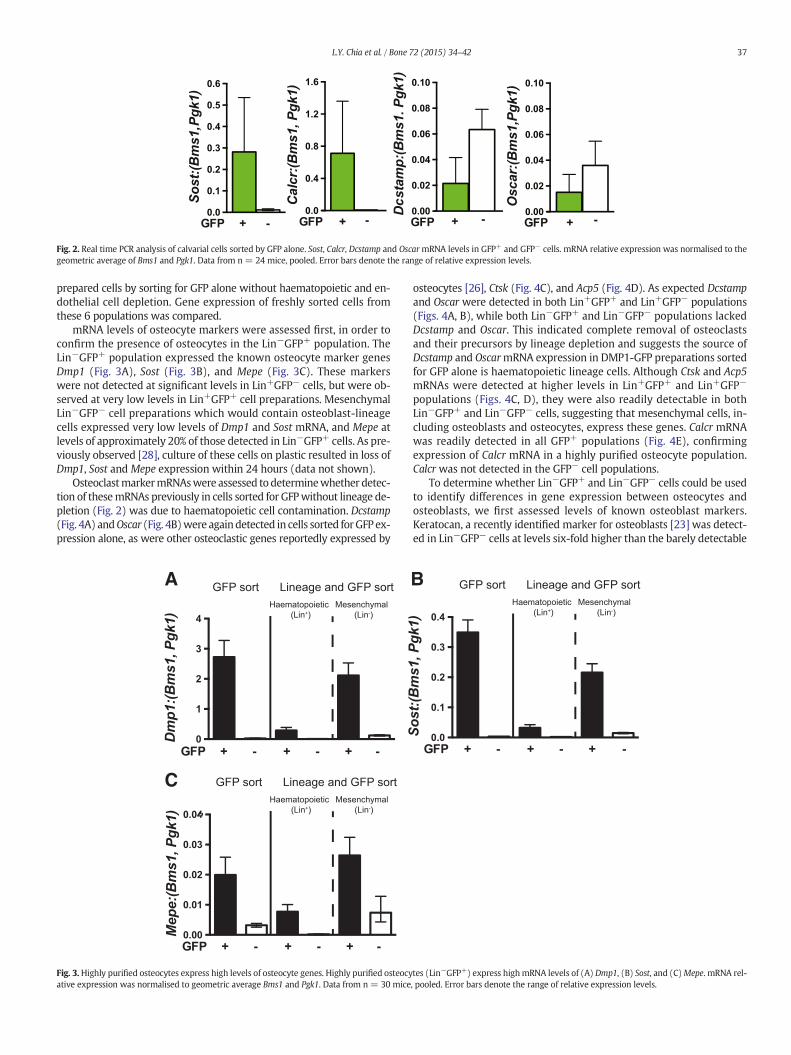
Fig. 2. Real time PCR analysis of calvarial cells sorted by GFP alone. Sost, Calcr, Dcstamp and OscarmRNA levels in GFP+ and GFP− cells. mRNA relative expression was normalised to thegeometric average of Bms1 and Pgk1. Data from n = 24 mice, pooled. Error bars denote the range of relative expression levels.
37L.Y. Chia et al. / Bone 72 (2015) 34–42
prepared cells by sorting for GFP alone without haematopoietic and en-dothelial cell depletion. Gene expression of freshly sorted cells fromthese 6 populations was compared.
mRNA levels of osteocyte markers were assessed first, in order toconfirm the presence of osteocytes in the Lin−GFP+ population. TheLin−GFP+ population expressed the known osteocyte marker genesDmp1 (Fig. 3A), Sost (Fig. 3B), and Mepe (Fig. 3C). These markerswere not detected at significant levels in Lin+GFP− cells, but were ob-served at very low levels in Lin+GFP+ cell preparations. MesenchymalLin−GFP− cell preparations which would contain osteoblast-lineagecells expressed very low levels of Dmp1 and Sost mRNA, and Mepe atlevels of approximately 20% of those detected in Lin−GFP+ cells. As pre-viously observed [28], culture of these cells on plastic resulted in loss ofDmp1, Sost and Mepe expression within 24 hours (data not shown).
OsteoclastmarkermRNAswere assessed todeterminewhether detec-tion of thesemRNAs previously in cells sorted for GFPwithout lineage de-pletion (Fig. 2) was due to haematopoietic cell contamination. Dcstamp(Fig. 4A) andOscar (Fig. 4B)were againdetected in cells sorted forGFP ex-pression alone, as were other osteoclastic genes reportedly expressed by
A
C
Fig. 3.Highly purified osteocytes express high levels of osteocyte genes. Highly purified osteocyative expression was normalised to geometric average Bms1 and Pgk1. Data from n = 30 mice
osteocytes [26], Ctsk (Fig. 4C), and Acp5 (Fig. 4D). As expected Dcstampand Oscar were detected in both Lin+GFP+ and Lin+GFP− populations(Figs. 4A, B), while both Lin−GFP+ and Lin−GFP− populations lackedDcstamp and Oscar. This indicated complete removal of osteoclastsand their precursors by lineage depletion and suggests the source ofDcstamp and OscarmRNA expression in DMP1-GFP preparations sortedfor GFP alone is haematopoietic lineage cells. Although Ctsk and Acp5mRNAs were detected at higher levels in Lin+GFP+ and Lin+GFP−
populations (Figs. 4C, D), they were also readily detectable in bothLin−GFP+ and Lin−GFP− cells, suggesting that mesenchymal cells, in-cluding osteoblasts and osteocytes, express these genes. Calcr mRNAwas readily detected in all GFP+ populations (Fig. 4E), confirmingexpression of Calcr mRNA in a highly purified osteocyte population.Calcr was not detected in the GFP− cell populations.
To determine whether Lin−GFP+ and Lin−GFP− cells could be usedto identify differences in gene expression between osteocytes andosteoblasts, we first assessed levels of known osteoblast markers.Keratocan, a recently identified marker for osteoblasts [23] was detect-ed in Lin−GFP− cells at levels six-fold higher than the barely detectable
B
tes (Lin−GFP+) express highmRNA levels of (A) Dmp1, (B) Sost, and (C)Mepe. mRNA rel-, pooled. Error bars denote the range of relative expression levels.

Fig. 4.Defining the expression of classic osteoclast genes inhaematopoietic and osteocytic populations. (A)High levels ofDcstamp and (B)OscarmRNA in Lin+GFP+but not Lin−GFP+ cellsconfirmed their expression in haematopoietic cells only. (C, D) Detection of mRNA for Ctsk and Acp5 in the Lin−GFP+ population confirms their expression in haematopoietic-depletedosteocytes. (E) Detection of CalcrmRNA in the Lin−GFP+ population confirms their expression in osteocytes. mRNA relative expression was normalised to geometric average of Bms1and Pgk1. Data from n = 30 mice, pooled. Error bars denote the range of relative expression levels.
38 L.Y. Chia et al. / Bone 72 (2015) 34–42
levels in osteocytes (Lin−GFP+) and both Lin+ populations (Fig. 5A),confirming that the Lin−GFP− population is enriched in osteoblasts. An-other osteoblast gene, Alpl was expressed at four-fold higher levels inboth Lin− populations compared to the Lin+ populations (Fig. 5B);Alpl and Pthr1mRNA levelswere equivalent in Lin+GFP+ and Lin+GFP−
populations (Figs. 5B, C). The late osteoblast/early osteocyte marker,Bglapwas expressed at six-fold higher levels in Lin−GFP+ cells comparedto Lin−GFP− cells, was detected at lower levels in Lin+GFP+ cells, butwas not present in Lin+GFP− cells (Fig. 5D). Thus, Lin−GFP+ cells repre-sent a purified osteocyte population while Lin−GFP−mesenchymal cellsexpress higher levels of early-mid differentiation stage osteoblast cellmarkers, indicating enrichment of these cells in this population.
Since the relative expression levels of Tnfsf11 (RANKL) and Tnfrsf11b(OPG) in osteoblasts and osteocytes are controversial we assessed thesemarkers in our highly purified populations. Both Tnfsf11 and Tnfrsf11bwere expressed at levels two-fold lower in purified osteocytes
(Lin−GFP+) compared to the enriched osteoblast population (Lin−
GFP− cells) (Figs. 5E, F).
GFP+ osteocyte preparations do not fully support osteoclast formationin vitro
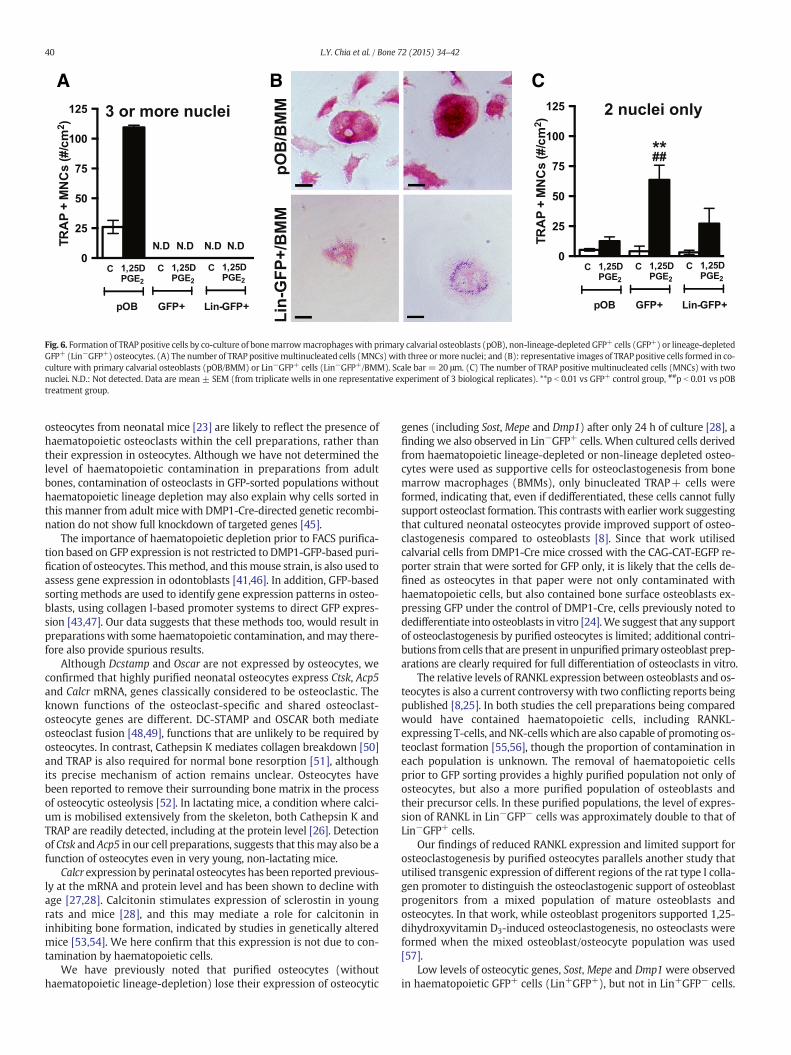
To assess whether purified osteocytes support osteoclast formationin vitro, we co-cultured osteocytes obtained by GFP sorting alone,or Lin−GFP+ cells, with bonemarrowmacrophages (BMMs), in the pres-ence of 1,25-dihydroxyvitamin D3 and prostaglandin E2 for 6–7 days. Pri-mary calvarial osteoblasts were used as a positive control. Althoughtartrate resistant acid phosphatase (TRAP)-positive osteoclasts withthree ormore nuclei were formed in the presence of primary calvarial os-teoblasts, neither osteocyte preparation supported their formation(Fig. 6A). However, we noted the presence of faintly TRAP+ binucleatedcells in the co-cultures of BMM with GFP+ cells (both GFP+ and

Fig. 5. Levels of early and late osteoblast genes and genes regulating osteoclastogenesis in highly purified osteocytes (Lin−GFP+) compared to highly purified osteoblasts (Lin−GFP−).Highly purified osteocytes (Lin−GFP+) expressed low mRNA levels of early osteoblastic gene (A) Kera, but high mRNA levels of late osteoblastic genes (B) Alpl, (C) Pthr1, (D) Bglap,(E) Tnfsf11 and (F) Tnfrsf11b. mRNA relative expression was normalised to geometric average of Bms1 and Pgk1. Data from n = 30 mice, pooled. Error bars denote the range of relativeexpression levels.
39L.Y. Chia et al. / Bone 72 (2015) 34–42
Lin−GFP+) (Fig. 6B). When quantified, we observed that while few ofthese cells were present in the primary calvarial osteoblast co-cultures,significantly more were observed when BMMs were cultured in thepresence of Lin−GFP+ cells with the osteoclastogenic stimulus (Fig. 6C)suggesting that they may be incompletely differentiated osteoclasts.
Discussion
In order to understand the paracrine and endocrine functions of os-teocytes, there is a growing need to isolate highly purified primary oste-ocytes. In thisworkwe showed that haematopoietic and endothelial celldepletion is required to obtain a highly purified preparation of thesecells, even fromDMP1-GFPmice. In doing so,we have confirmed the ex-pression in osteocytes of several genes previously reported to be sharedbetween osteoclasts and osteocytes (Calcr, Acp5 and Ctsk), but not othergenes (Dcstamp andOscar). Thismethod also allowed us to confirm that
osteoblasts express relatively higher levels of Tnfsf11 and Tnfrsf11bmRNA than osteocytes, and finally that these purified osteocytes donot support osteoclast differentiation to the same extent as primarycalvarial osteoblasts. In this study, in order to be thorough, we used afull suite of antibodies against the haematopoietic cell lineages and en-dothelial cells, however a simpler approach using antibodies to CD45,CD11b, Ter119 and CD31 would likely provide similar results.
The use of FACS to purify osteocytes and odontoblasts from DMP1-GFPmice is an increasingly commonmethod in skeletal and dental biol-ogy [19,23,27,28,41–44], but prior depletion of haematopoietic cells hasnot been used. Our finding that purification of cellular digests of DMP1-GFP bones by FACS sorting for GFP alone results in a population of cellscontaining approximately 5% haematopoietic-lineage cells, with signifi-cant expression of osteoclast-specific mRNAs, calls into question someearlier conclusions about osteocytic gene expression. For example,Dcstamp and Oscar mRNAs previously detected in isolated calvarial
A B C
Fig. 6. Formation of TRAP positive cells by co-culture of bonemarrowmacrophageswith primary calvarial osteoblasts (pOB), non-lineage-depleted GFP+ cells (GFP+) or lineage-depletedGFP+ (Lin−GFP+) osteocytes. (A) The number of TRAP positivemultinucleated cells (MNCs) with three ormore nuclei; and (B): representative images of TRAP positive cells formed in co-culture with primary calvarial osteoblasts (pOB/BMM) or Lin−GFP+ cells (Lin−GFP+/BMM). Scale bar = 20 μm. (C) The number of TRAP positive multinucleated cells (MNCs) with twonuclei. N.D.: Not detected. Data are mean ± SEM (from triplicate wells in one representative experiment of 3 biological replicates). **p b 0.01 vs GFP+ control group, ##p b 0.01 vs pOBtreatment group.
40 L.Y. Chia et al. / Bone 72 (2015) 34–42
osteocytes from neonatal mice [23] are likely to reflect the presence ofhaematopoietic osteoclasts within the cell preparations, rather thantheir expression in osteocytes. Although we have not determined thelevel of haematopoietic contamination in preparations from adultbones, contamination of osteoclasts in GFP-sorted populations withouthaematopoietic lineage depletion may also explain why cells sorted inthis manner from adult micewith DMP1-Cre-directed genetic recombi-nation do not show full knockdown of targeted genes [45].
The importance of haematopoietic depletion prior to FACS purifica-tion based on GFP expression is not restricted to DMP1-GFP-based puri-fication of osteocytes. Thismethod, and thismouse strain, is also used toassess gene expression in odontoblasts [41,46]. In addition, GFP-basedsorting methods are used to identify gene expression patterns in osteo-blasts, using collagen I-based promoter systems to direct GFP expres-sion [43,47]. Our data suggests that these methods too, would result inpreparationswith some haematopoietic contamination, andmay there-fore also provide spurious results.
Although Dcstamp and Oscar are not expressed by osteocytes, weconfirmed that highly purified neonatal osteocytes express Ctsk, Acp5and Calcr mRNA, genes classically considered to be osteoclastic. Theknown functions of the osteoclast-specific and shared osteoclast-osteocyte genes are different. DC-STAMP and OSCAR both mediateosteoclast fusion [48,49], functions that are unlikely to be required byosteocytes. In contrast, Cathepsin K mediates collagen breakdown [50]and TRAP is also required for normal bone resorption [51], althoughits precise mechanism of action remains unclear. Osteocytes havebeen reported to remove their surrounding bone matrix in the processof osteocytic osteolysis [52]. In lactating mice, a condition where calci-um is mobilised extensively from the skeleton, both Cathepsin K andTRAP are readily detected, including at the protein level [26]. Detectionof Ctsk and Acp5 in our cell preparations, suggests that thismay also be afunction of osteocytes even in very young, non-lactating mice.
Calcr expression by perinatal osteocytes has been reported previous-ly at the mRNA and protein level and has been shown to decline withage [27,28]. Calcitonin stimulates expression of sclerostin in youngrats and mice [28], and this may mediate a role for calcitonin ininhibiting bone formation, indicated by studies in genetically alteredmice [53,54]. We here confirm that this expression is not due to con-tamination by haematopoietic cells.
We have previously noted that purified osteocytes (withouthaematopoietic lineage-depletion) lose their expression of osteocytic
genes (including Sost,Mepe and Dmp1) after only 24 h of culture [28], afinding we also observed in Lin−GFP+ cells. When cultured cells derivedfrom haematopoietic lineage-depleted or non-lineage depleted osteo-cytes were used as supportive cells for osteoclastogenesis from bonemarrow macrophages (BMMs), only binucleated TRAP+ cells wereformed, indicating that, even if dedifferentiated, these cells cannot fullysupport osteoclast formation. This contrastswith earlier work suggestingthat cultured neonatal osteocytes provide improved support of osteo-clastogenesis compared to osteoblasts [8]. Since that work utilisedcalvarial cells from DMP1-Cre mice crossed with the CAG-CAT-EGFP re-porter strain that were sorted for GFP only, it is likely that the cells de-fined as osteocytes in that paper were not only contaminated withhaematopoietic cells, but also contained bone surface osteoblasts ex-pressing GFP under the control of DMP1-Cre, cells previously noted todedifferentiate into osteoblasts in vitro [24].We suggest that any supportof osteoclastogenesis by purified osteocytes is limited; additional contri-butions fromcells that are present in unpurifiedprimary osteoblast prep-arations are clearly required for full differentiation of osteoclasts in vitro.
The relative levels of RANKL expression between osteoblasts and os-teocytes is also a current controversywith two conflicting reports beingpublished [8,25]. In both studies the cell preparations being comparedwould have contained haematopoietic cells, including RANKL-expressing T-cells, andNK-cells which are also capable of promoting os-teoclast formation [55,56], though the proportion of contamination ineach population is unknown. The removal of haematopoietic cellsprior to GFP sorting provides a highly purified population not only ofosteocytes, but also a more purified population of osteoblasts andtheir precursor cells. In these purified populations, the level of expres-sion of RANKL in Lin−GFP− cells was approximately double to that ofLin−GFP+ cells.
Our findings of reduced RANKL expression and limited support forosteoclastogenesis by purified osteocytes parallels another study thatutilised transgenic expression of different regions of the rat type I colla-gen promoter to distinguish the osteoclastogenic support of osteoblastprogenitors from a mixed population of mature osteoblasts andosteocytes. In that work, while osteoblast progenitors supported 1,25-dihydroxyvitamin D3-induced osteoclastogenesis, no osteoclasts wereformed when the mixed osteoblast/osteocyte population was used[57].
Low levels of osteocytic genes, Sost, Mepe and Dmp1were observedin haematopoietic GFP+ cells (Lin+GFP+), but not in Lin+GFP− cells.

41L.Y. Chia et al. / Bone 72 (2015) 34–42
This is consistentwith prior reports of low levels of sclerostinmRNAandprotein in osteoclasts and endothelial cells during bone development[58] and in conditioned medium from isolated osteoclasts from agedmice [29]. Although sclerostin is generally considered to be osteocyte-specific in adult bone, it is expressed in multiple tissues in the develop-ing embryo, including the kidney, liver and heart [59], and this may ex-plain the detection of sclerostin in non-mesenchymal GFP+ cells in thepresent study of neonatalmice. Similarly, DMP1 andMEPE protein havebeen identified in the kidney and pancreas [60,61], but expression inhaematopoietic lineage cells has not been shown previously.
Unfortunately, haematopoietic lineage depletion coupled with GFPpurification inevitably provides a small amount of material to workwith. For each replicate of this study, we pooled cells from 30 mice,and obtained only sufficient RNA from the Lin−GFP+ population to as-sess expression of 14mRNAs in duplicate PCRs. This means the methodis useful for verifying data obtainedwithout purification, but is not a de-sirable method for large scale analysis or routine laboratory investiga-tions. Such small cellular yields also limits the capability to carry out apost-sort analysis to confirm purity of cell populations, however, sinceLin+GFP+ cells displayed a range of GFP intensities (see FACS plot inFig. 1), it is unlikely that complete removal of haematopoietic and endo-thelial cells can be achieved without lineage depletion by simplyrestricting the GFP gate. In addition, although this method produced ahighly purified population of osteocytes, these cells did not retainosteocytic gene expression when cultured in vitro as previously ob-served [28] and discussed above.Whether the cells remaining in culturecould be considered true osteocytes or have de-differentiated toosteoblasts is not known; in either case, they are unable to fully supportosteoclastogenesis. We suggest that a combination of cell-sorting and3D culture with appropriate matrix may be required for primary osteo-cytes to maintain their phenotype in culture and allow mechanisticstudies that resemble in vivo conditions.
In conclusion, this study demonstrates the importance ofhaematopoietic and endothelial lineage depletion to generate highlypurified osteocytes. With lineage depletion we have confirmed thatneonatal calvarial osteocytes express Acp5, Ctsk and Calcr, but notother osteoclast markers, and that cells purified in this manner do notfully support osteoclast formation in vitro. We suggest that lineagedepletion should be used routinely when assessing osteocyte geneexpression in order to eliminate ambiguous results obtained by thepresence of other cell types.
Acknowledgments
The authors acknowledge the advice of Emma K Baker and Carl RWalkley (St. Vincent's Institute, Melbourne) on appropriate markersfor haematopoietic and endothelial cell depletion, and technical assis-tance from Narelle McGregor. This work was funded by NHMRC ProjectGrant 1002730. St. Vincent's Institute receives support from theVictorian State Government O.I.S. Programme.
References
[1] Bonewald LF. Osteocytes as dynamic multifunctional cells. Ann N Y Acad Sci 2007;1116:281–90.
[2] Schaffler MB, Cheung WY, Majeska R, Kennedy O. Osteocytes: master orchestratorsof bone. Calcif Tissue Int 2014;94:5–24.
[3] Poole KE, van Bezooijen RL, Loveridge N, Hamersma H, Papapoulos SE, Lowik CW,et al. Sclerostin is a delayed secreted product of osteocytes that inhibits bone forma-tion. FASEB J 2005;19:1842–4.
[4] Leupin O, Piters E, Halleux C, Hu S, Kramer I, Morvan F, et al. Bone overgrowth-associated mutations in the LRP4 gene impair sclerostin facilitator function. J BiolChem 2011;286:19489–500.
[5] Feng JQ, Ward LM, Liu S, Lu Y, Xie Y, Yuan B, et al. Loss of DMP1 causes rickets andosteomalacia and identifies a role for osteocytes in mineral metabolism. Nat Genet2006;38:1310–5.
[6] Lorenz-Depiereux B, Bastepe M, Benet-Pages A, Amyere M, Wagenstaller J, Muller-Barth U, et al. DMP1mutations in autosomal recessive hypophosphatemia implicate
a bone matrix protein in the regulation of phosphate homeostasis. Nat Genet 2006;38:1248–50.
[7] Bergwitz C, Juppner H. FGF23 and syndromes of abnormal renal phosphate handling.Adv Exp Med Biol 2012;728:41–64.
[8] Nakashima T, Hayashi M, Fukunaga T, Kurata K, Oh-Hora M, Feng JQ, et al. Evidencefor osteocyte regulation of bone homeostasis through RANKL expression. Nat Med2011;17:1231–4.
[9] Vashishth D, Verborgt O, Divine G, Schaffler MB, Fyhrie DP. Decline in osteocyte la-cunar density in human cortical bone is associated with accumulation ofmicrocracks with age. Bone 2000;26:375–80.
[10] Verborgt O, Gibson GJ, Schaffler MB. Loss of osteocyte integrity in association withmicrodamage and bone remodeling after fatigue in vivo. J Bone Miner Res 2000;15:60–7.
[11] Cardoso L, Herman BC, Verborgt O, Laudier D, Majeska RJ, Schaffler MB. Osteocyteapoptosis controls activation of intracortical resorption in response to bone fatigue.J Bone Miner Res 2009;24:597–605.
[12] Xiong J, Onal M, Jilka RL, Weinstein RS, Manolagas SC, O'Brien CA. Matrix-embeddedcells control osteoclast formation. Nat Med 2011;17:1235–41.
[13] Nijweide PJ, Mulder RJ. Identification of osteocytes in osteoblast-like cell culturesusing amonoclonal antibody specifically directed against osteocytes. Histochemistry1986;84:342–7.
[14] Westbroek I, De Rooij KE, Nijweide PJ. Osteocyte-specific monoclonal antibody MAbOB7.3 is directed against Phex protein. J Bone Miner Res 2002;17:845–53.
[15] Mikuni-Takagaki Y, Kakai Y, SatoyoshiM, Kawano E, Suzuki Y, Kawase T, et al. Matrixmineralization and the differentiation of osteocyte-like cells in culture. J Bone MinerRes 1995;10:231–42.
[16] Kawata A, Mikuni-Takagaki Y. Mechanotransduction in stretched osteocytes—temporal expression of immediate early and other genes. Biochem Biophys ResCommun 1998;246:404–8.
[17] van der Plas A, Aarden EM, Feijen JH, de Boer AH, Wiltink A, Alblas MJ, et al. Charac-teristics and properties of osteocytes in culture. J Bone Miner Res 1994;9:1697–704.
[18] Aarden EM, Wassenaar AM, Alblas MJ, Nijweide PJ. Immunocytochemical demon-stration of extracellular matrix proteins in isolated osteocytes. Histochem Cell Biol1996;106:495–501.
[19] Halleux C, Kramer I, Allard C, Kneissel M. Isolation of mouse osteocytes using cellfractionation for gene expression analysis. Methods Mol Biol 2012;816:55–66.
[20] Stern AR, Stern MM, Van Dyke ME, Jahn K, Prideaux M, Bonewald LF. Isolation andculture of primary osteocytes from the long bones of skeletally mature and agedmice. Biotechniques 2012;52:361–73.
[21] Toyosawa S, Shintani S, Fujiwara T, Ooshima T, Sato A, Ijuhin N, et al. Dentin matrixprotein 1 is predominantly expressed in chicken and rat osteocytes but not inosteoblasts. J Bone Miner Res 2001;16:2017–26.
[22] Kalajzic I, Braut A, Guo D, Jiang X, Kronenberg MS, Mina M, et al. Dentin matrixprotein 1 expression during osteoblastic differentiation, generation of an osteocyteGFP-transgene. Bone 2004;35:74–82.
[23] Paic F, Igwe JC, Nori R, Kronenberg MS, Franceschetti T, Harrington P, et al. Identifi-cation of differentially expressed genes between osteoblasts and osteocytes. Bone2009;45:682–92.
[24] Torreggiani E, Matthews BG, Pejda S, Matic I, Horowitz MC, Grcevic D, et al.Preosteocytes/osteocytes have the potential to dedifferentiate becoming a sourceof osteoblasts. PLoS One 2013;8:e75204.
[25] Fumoto T, Takeshita S, Ito M, Ikeda K. Physiological functions of osteoblast lineageand T cell-derived RANKL in bone homeostasis. J Bone Miner Res 2014;29:830–42.
[26] Qing H, Ardeshirpour L, Pajevic PD, Dusevich V, Jahn K, Kato S, et al. Demonstrationof osteocytic perilacunar/canalicular remodeling in mice during lactation. J BoneMiner Res 2012;27:1018–29.
[27] Gooi JH, Chia LY, Walsh NC, Karsdal MA, Quinn JMW, Martin TJ, et al. Decline in cal-citonin receptor expression in osteocytes with age. J Endocrinol 2014;221:181–91.
[28] Gooi JH, Pompolo S, Karsdal MA, Kulkarni NH, Kalajzic I, McAhren SH, et al. Calcito-nin impairs the anabolic effect of PTH in young rats and stimulates expression ofsclerostin by osteocytes. Bone 2010;46:1486–97.
[29] Ota K, Quint P, Ruan M, Pederson L, Westendorf JJ, Khosla S, et al. Sclerostin isexpressed in osteoclasts from aged mice and reduces osteoclast-mediated stimula-tion of mineralization. J Cell Biochem 2013;114:1901–7.
[30] Lagasse E, Connors H, Al-Dhalimy M, Reitsma M, Dohse M, Osborne L, et al. Purifiedhematopoietic stem cells can differentiate into hepatocytes in vivo. NatMed 2000;6:1229–34.
[31] Semerad CL, Christopher MJ, Liu F, Short B, Simmons PJ, Winkler I, et al. G-CSF po-tently inhibits osteoblast activity and CXCL12mRNA expression in the bonemarrow.Blood 2005;106:3020–7.
[32] Rybtsov S, Sobiesiak M, Taoudi S, Souilhol C, Senserrich J, Liakhovitskaia A, et al. Hi-erarchical organization and early hematopoietic specification of the developing HSClineage in the AGM region. J Exp Med 2011;208:1305–15.
[33] Pearce DJ, Ridler CM, Simpson C, Bonnet D. Multiparameter analysis of murine bonemarrow side population cells. Blood 2004;103:2541–6.
[34] Asahara T, Murohara T, Sullivan A, Silver M, van der Zee R, Li T, et al. Isolation ofputative progenitor endothelial cells for angiogenesis. Science 1997;275:964–7.
[35] Singbrant S, Russell MR, Jovic T, Liddicoat B, Izon DJ, Purton LE, et al. Erythropoietincouples erythropoiesis, B-lymphopoiesis, and bone homeostasis within the bonemarrow microenvironment. Blood 2011;117:5631–42.
[36] Allan EH, Hausler KD, Wei T, Gooi JH, Quinn JM, Crimeen-Irwin B, et al. EphrinB2regulation by PTH and PTHrP revealed by molecular profiling in differentiatingosteoblasts. J Bone Miner Res 2008;23:1170–81.
[37] Takyar FM, Tonna S, Ho PW, Crimeen-Irwin B, Baker EK, Martin TJ, et al. EphrinB2/EphB4 inhibition in the osteoblast lineage modifies the anabolic response toparathyroid hormone. J Bone Miner Res 2013;28:912–25.

42 L.Y. Chia et al. / Bone 72 (2015) 34–42
[38] Le Goff B, Singbrant S, Tonkin BA, Martin TJ, Romas E, Sims NA, et al. Oncostatin Macting via OSMR, augments the actions of IL-1 and TNF in synovial fibroblasts.Cytokine 2014;68:101–9.
[39] Nakamura A, Ly C, Cipetic M, Sims NA, Vieusseux J, Kartsogiannis V, et al. Osteoclastinhibitory lectin (OCIL) inhibits osteoblast differentiation and function in vitro. Bone2007;40:305–15.
[40] Parkinson IH, Fazzalari NL, Durbridge TC, Moore AJ. Simplified approach to enzymat-ic identification of osteoclastic bone resorption. J Histotechnol 1991;14:81–3.
[41] Balic A, MinaM. Identification of secretory odontoblasts using DMP1-GFP transgenicmice. Bone 2011;48:927–37.
[42] Igwe JC, Jiang X, Paic F, Ma L, Adams DJ, Baldock PA, et al. Neuropeptide Y is expressedby osteocytes and can inhibit osteoblastic activity. J Cell Biochem 2009;108:621–30.
[43] Li H, Marijanovic I, Kronenberg MS, Erceg I, Stover ML, Velonis D, et al. Expressionand function of Dlx genes in the osteoblast lineage. Dev Biol 2008;316:458–70.
[44] Kondoh S, Inoue K, Igarashi K, Sugizaki H, Shirode-Fukuda Y, Inoue E, et al. Estrogenreceptor alpha in osteocytes regulates trabecular bone formation in female mice.Bone 2014;60:68–77.
[45] Bivi N, Nelson MT, Faillace ME, Li J, Miller LM, Plotkin LI. Deletion of Cx43 from oste-ocytes results in defective bone material properties but does not decrease extrinsicstrength in cortical bone. Calcif Tissue Int 2012;91:215–24.
[46] Balic A, Aguila HL, Mina M. Identification of cells at early and late stages of polariza-tion during odontoblast differentiation using pOBCol3.6GFP and pOBCol2.3GFPtransgenic mice. Bone 2010;47:948–58.
[47] Powell Jr WF, Barry KJ, Tulum I, Kobayashi T, Harris SE, Bringhurst FR, et al. Targetedablation of the PTH/PTHrP receptor in osteocytes impairs bone structure andhomeostatic calcemic responses. J Endocrinol 2011;209:21–32.
[48] Yagi M, Miyamoto T, Sawatani Y, Iwamoto K, Hosogane N, Fujita N, et al. DC-STAMPis essential for cell–cell fusion in osteoclasts and foreign body giant cells. J Exp Med2005;202:345–51.
[49] Barrow AD, Raynal N, Andersen TL, Slatter DA, Bihan D, Pugh N, et al. OSCAR is a col-lagen receptor that costimulates osteoclastogenesis in DAP12-deficient humans andmice. J Clin Invest 2011;121:3505–16.
[50] Garnero P, Borel O, Byrjalsen I, Ferreras M, Drake FH, McQueney MS, et al. Thecollagenolytic activity of Cathepsin K is unique among mammalian proteinases. JBiol Chem 1998;273:32347–52.
[51] Hayman AR, Jones SJ, Boyde A, Foster D, ColledgeWH, Carlton MB, et al. Mice lackingtartrate-resistant acid phosphatase (Acp 5) have disrupted endochondral ossifica-tion and mild osteopetrosis. Development 1996;122:3151–62.
[52] Belanger LF, Robichon J, Migicovsky BB, Copp DH, Vincent J. Resorption withoutosteoclasts (osteolysis). Mechanisms of hard tissue destruction. Washington, D.C.:American Association for the, Advancement of Science; 1963. p. 531–56.
[53] Hoff AO, Catala-Lehnen P, Thomas PM, Priemel M, Rueger JM, Nasonkin I, et al.Increased bone mass is an unexpected phenotype associated with deletion of thecalcitonin gene. J Clin Invest 2002;110:1849–57.
[54] Dacquin R, Davey RA, Laplace C, Levasseur R, Morris HA, Goldring SR, et al. Amylininhibits bone resorption while the calcitonin receptor controls bone formationin vivo. J Cell Biol 2004;164:509–14.
[55] Horwood NJ, Kartsogiannis V, Quinn JM, Romas E, Martin TJ, Gillespie MT. ActivatedT lymphocytes support osteoclast formation in vitro. Biochem Biophys Res Commun1999;265:144–50.
[56] Soderstrom K, Stein E, Colmenero P, Purath U, Muller-Ladner U, de Matos CT, et al.Natural killer cells trigger osteoclastogenesis and bone destruction in arthritis.Proc Natl Acad Sci U S A 2010;107:13028–33.
[57] Li H, Jiang X, Delaney J, Franceschetti T, Bilic-Curcic I, Kalinovsky J, et al. Immatureosteoblast lineage cells increase osteoclastogenesis in osteogenesis imperfecta mu-rine. Am J Pathol 2010;176:2405–13.
[58] Kusu N, Laurikkala J, Imanishi M, Usui H, Konishi M, Miyake A, et al. Sclerostin is anovel secreted osteoclast-derived bone morphogenetic protein antagonist withunique ligand specificity. J Biol Chem 2003;278:24113–7.
[59] Loots GG, Kneissel M, Keller H, Baptist M, Chang J, Collette NM, et al. Genomic dele-tion of a long-range bone enhancer misregulates sclerostin in Van Buchem disease.Genome Res 2005;15:928–35.
[60] TerasawaM, Shimokawa R, Terashima T, Ohya K, Takagi Y, Shimokawa H. Expressionof dentin matrix protein 1 (DMP1) in nonmineralized tissues. J Bone Miner Metab2004;22:430–8.
[61] Ogbureke KU, Fisher LW. Renal expression of SIBLING proteins and their partnermatrix metalloproteinases (MMPs). Kidney Int 2005;68:155–66.




















