
Identification of new populations of chicken natural killer (NK) cells

ORIGINAL ARTICLE
Interleukin-15 liver gene transfer increasesthe number and function of IKDCs and NK cells
A Arina1,4, O Murillo1,4, J Dubrot1, A Azpilikueta1, I Gabari1, JL Perez-Gracia1,2, C Alfaro1, C Berasain1,
J Prieto1,2, S Ferrini3, S Hervas-Stubbs1,5 and I Melero1,2,5
1Gene Therapy Unit, Centro de Investigacion Medica Aplicada (CIMA), Universidad de Navarra, Pamplona, Spain; 2ClınicaUniversitaria, Universidad de Navarra, Pamplona, Spain and 3Immunological Therapy, Istituto Nazionale per la Ricerca sul Cancro(IST-Genova), Genova, Italy
The surface phenotype CD3�NK1.1+DX5+CD11cintB220+
GR1� has been recently ascribed to a novel subset of mouseleukocytes termed interferon (IFN)-producing killer dendriticcells (IKDCs) that shares functions with natural killer (NK) cellsand DCs. Interleukin-15 (IL-15) is critical for NK cells but itsrelationship with IKDC remained unexplored. An expressioncassette encoding human IL-15 (hIL-15) has been transferredby hydrodynamic injection into the liver of mice, resulting intransient expression of the cytokine that is detectable duringthe first 48 h. hIL-15 hydrodynamic gene transfer resulted in anexpansion of NK cells and IKDCs. Relative expansions ofIKDCs were more dramatic in the IL-15 gene-transferredhepatic tissue than in the spleen. Adoptively transferred DX5+
cells comprising both NK cells and IKDCs proliferated inresponse to hydrodynamic injection of hIL-15, indicating thatquantitative increases are at least in part the result ofproliferation from already differentiated cells. Expansion isaccompanied by enhanced cytolytic activity and increasedexpression of TRAIL and CD137 (4-1BB), without augmentinginterferon-g production. The effects of a single hydrodynamicinjection surpassed those of two intraperitoneal doses of therecombinant protein. The novel functional link betweencirculating IL-15 and IKDCs opens new possibilities to studythe biology and applications of this minority cell subset.Gene Therapy (2008) 15, 473–483; doi:10.1038/gt.2008.4;published online 14 February 2008
Keywords: interleukin-15; gene transfer; natural killer cells; dendritic cells
Introduction
Interferon (IFN)-producing killer dendritic cells (IKDCs),a cell type with intermediate features of naturalkiller (NK) cells and DCs, have been recentlydescribed in mice.1,2 These cells are defined asCD3�NK1.1+DX5+CD11cintB220+GR1� and are keymediators of antigen-independent IFN-g production inresponse to various stimuli, such as tumor cells andcytokines.1,2 Interestingly, this subset has been reportedto mediate both natural cytotoxicity and major histo-compatibility complex class II-mediated antigen presen-tation to T cells.1 IKDCs, through a direct tumoricidalactivity, have been implicated in the innate immuno-therapeutic effects of a combination of interleukin (IL)-2plus imatinib against B16F10-derived melanomas.2
Previous reports had also postulated the existence of apopulation of cells sharing some of the functional andphenotypical features of both NK cells and DCs in mice,3
mainly on the basis of CD11c and NK1.1 coexpression.There is evidence that these so-called NKDCs (that
comprise the IKDC subset) play an important role in thedefense against Listeria primarily by producing IFN-g inresponse to IL-12 and IL-18.4 Recently published experi-ments show that NK and IKDC cell lineages originate inmice from different primitive lymphoid precursors,supporting the notion that these cells represent a distincthematopoietic lineage,5 albeit an homologous cell subsetin human beings still defies identification.6,7 However,several groups have recently questioned the existence ofthis cell subset as a separate lineage from NK cells, withthe putative antigen presentation function of IKDC at thecore of the controversy.8–10 For some of these authors,IKDCs may represent an activated status of conventionalNK cells.8
IL-15 11,12 is an absolutely essential cytokine for thedifferentiation and maintenance of memory CD8+ T,NKT and NK cells.13 IL-15 shows a broad tissuedistribution at the mRNA level, but post-transcriptionaland intracellular trafficking mechanisms confine func-tional production mainly to myeloid cells.14 The effects ofIL-15 on NK cells have been studied in detail.12 IL-15 andIL-15Ra are necessary for the differentiation of NK cellsfrom lymphoid precursors and for their steady-statesurvival.13,15,16 IL-15 is mitogenic for NK cells, enhancessecretion of pro-inflammatory cytokines and upregulatestheir cytolytic machinery.12,17
The multimeric receptor for IL-15 encompasses twosignaling subunits IL-2Rb (CD122) and IL-2Rg (CD132),while the high-affinity specific chain (IL-15Ra) makes
Received 2 August 2007; revised 26 December 2007; accepted 29December 2007; published online 14 February 2008
Correspondence: Dr I Melero or Dr S Hervas-Stubbs, Gene TherapyUnit, Centro de Investigacion Medica Aplicada (CIMA), Universi-dad de Navarra, Avda Pio XII, 55, Pamplona 31008, Spain.E-mails: [email protected] or [email protected] authors contributed equally to this work.5These authors will equally share credit for senior authorship.
Gene Therapy (2008) 15, 473–483& 2008 Nature Publishing Group All rights reserved 0969-7128/08 $30.00
www.nature.com/gt
limited or no contribution to signaling. The main role ofIL-15Ra is trans-presenting its bound cytokine to thesignaling receptor subunits on the membrane of ajuxtaposed cell.18 In this sense, IL-15 complexed to IL-15Ra on the plasma membrane of an antigen-presentingcell acts more as a co-stimulatory molecule rather than asa soluble cytokine.19 Trans-presentation events can bemediated by monocytes and DCs, which are the mainproducers of this cytokine.18,20
Hydrodynamic injection is a relatively simple anduseful technique in rodents for gene delivery in vivo thatrelies in rapidly injecting a solution containing nakedDNA into the tail veins.21 As a result of the hydrostaticpressure and the reflux of the DNA solution from thecava to the hepatic veins, the plasmid extravasates fromliver sinusoids and enters into hepatocytes. The proce-dure gives rise to 2–20% hepatocytes productivelytransfected and has been used to express in vivocytokines, such as sFLT-3L22 and IL-2.23 Several ex vivogene transfer strategies involving IL-15 have beenstudied in mice,12,19,24–29 but in vivo gene transfer toattain circulating systemic levels of IL-15 has not beentried and can be a valuable tool to study the biology ofthis cytokine.
The complete lack of DX5+ and NK1.1+ cells in IL-15�/�
mice13 and the absence of IKDCs in CD122�/� mice1
made us suspect that IL-15 could be critical for thefunction of IKDCs, since they share these surfacemarkers with conventional NK cells. In this study, weshow that gene-transferred liver tissue can be used as an
endogenous source of IL-15, which does increase boththe function and number of IKDCs and NK cells.
Results
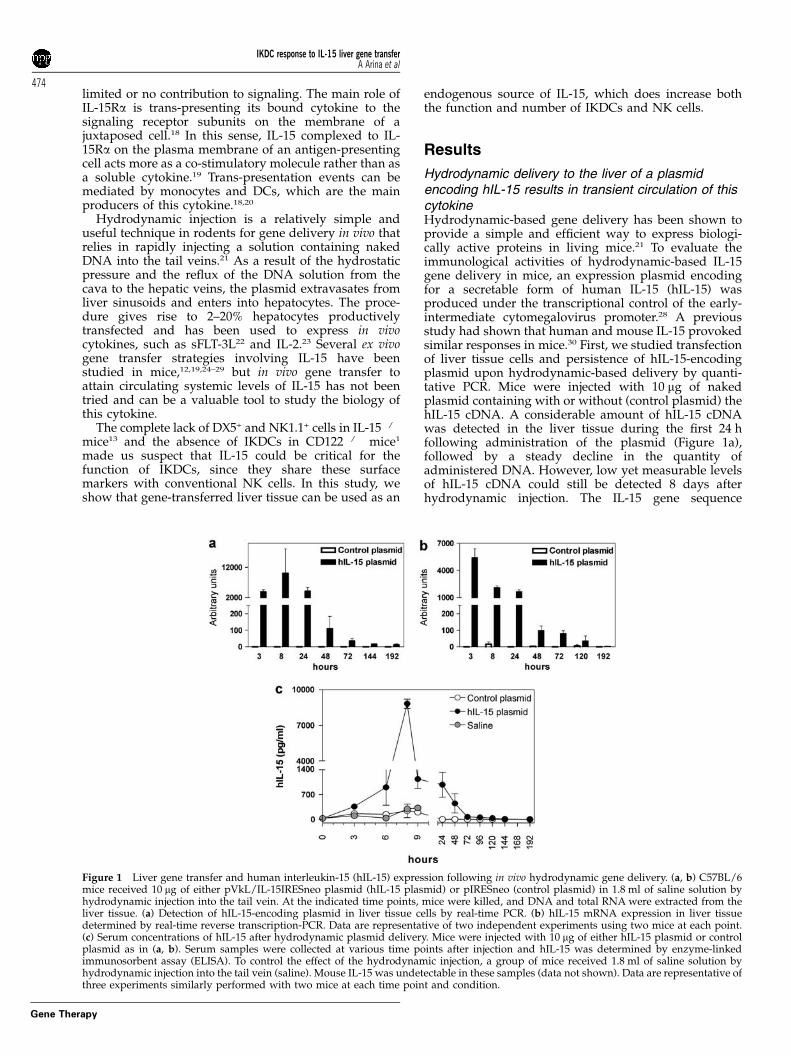
Hydrodynamic delivery to the liver of a plasmidencoding hIL-15 results in transient circulation of thiscytokineHydrodynamic-based gene delivery has been shown toprovide a simple and efficient way to express biologi-cally active proteins in living mice.21 To evaluate theimmunological activities of hydrodynamic-based IL-15gene delivery in mice, an expression plasmid encodingfor a secretable form of human IL-15 (hIL-15) wasproduced under the transcriptional control of the early-intermediate cytomegalovirus promoter.28 A previousstudy had shown that human and mouse IL-15 provokedsimilar responses in mice.30 First, we studied transfectionof liver tissue cells and persistence of hIL-15-encodingplasmid upon hydrodynamic-based delivery by quanti-tative PCR. Mice were injected with 10 mg of nakedplasmid containing with or without (control plasmid) thehIL-15 cDNA. A considerable amount of hIL-15 cDNAwas detected in the liver tissue during the first 24 hfollowing administration of the plasmid (Figure 1a),followed by a steady decline in the quantity ofadministered DNA. However, low yet measurable levelsof hIL-15 cDNA could still be detected 8 days afterhydrodynamic injection. The IL-15 gene sequence
Figure 1 Liver gene transfer and human interleukin-15 (hIL-15) expression following in vivo hydrodynamic gene delivery. (a, b) C57BL/6mice received 10 mg of either pVkL/IL-15IRESneo plasmid (hIL-15 plasmid) or pIRESneo (control plasmid) in 1.8 ml of saline solution byhydrodynamic injection into the tail vein. At the indicated time points, mice were killed, and DNA and total RNA were extracted from theliver tissue. (a) Detection of hIL-15-encoding plasmid in liver tissue cells by real-time PCR. (b) hIL-15 mRNA expression in liver tissuedetermined by real-time reverse transcription-PCR. Data are representative of two independent experiments using two mice at each point.(c) Serum concentrations of hIL-15 after hydrodynamic plasmid delivery. Mice were injected with 10 mg of either hIL-15 plasmid or controlplasmid as in (a, b). Serum samples were collected at various time points after injection and hIL-15 was determined by enzyme-linkedimmunosorbent assay (ELISA). To control the effect of the hydrodynamic injection, a group of mice received 1.8 ml of saline solution byhydrodynamic injection into the tail vein (saline). Mouse IL-15 was undetectable in these samples (data not shown). Data are representative ofthree experiments similarly performed with two mice at each time point and condition.
IKDC response to IL-15 liver gene transferA Arina et al
474
Gene Therapy
detected corresponds to the transgene and not to the hostgene since no IL-15 DNA was amplified in the group ofmice that received the control plasmid (Figure 1a).
Foreign gene expression could be detected by quanti-tative reverse transcription-PCR following hydro-dynamic hIL-15 cDNA delivery (Figure 1b). The kineticsof hIL-15 gene expression closely resembled the kineticsof detection of hIL-15-encoding plasmid in liver tissues,with high levels of hIL-15 mRNA during the first 24 hfollowed by low but measurable levels of gene expres-sion that persisted up to day 5. Administration of hIL-15expression plasmid resulted in a transient expression ofhIL-15 whose concentration peaked in serum 8 h afterinjection (reaching up to 8 ng ml�1) and was detectablethrough 48 h (Figure 1c). The loss of the hIL-15-encodingplasmid in the liver together with the short half-life of IL-15 is conceivably the main causes for the fast decline ofthis cytokine in the serum. No mouse IL-15, IFN-g, tumornecrosis factor (TNF)-a, IL-12(p70) were detected inserial serum samples drawn after hydrodynamic deliv-ery of either hIL-15-encoding or control plasmid (datanot shown). Similarly, we checked the possible inductionof host IL-2, since this cytokine shares with IL-15 tworeceptor subunits and some biological functions. How-ever, very low levels (below 50 pg ml�1) of mouse IL-2could be detected in these serum samples (data notshown). Moreover, such small quantities of mIL-2 weresimilar in the IL-15 and the control plasmid-treated mice,hence excluding a role of IL-2 on the effects of IL-15hydrodynamic gene transfer.
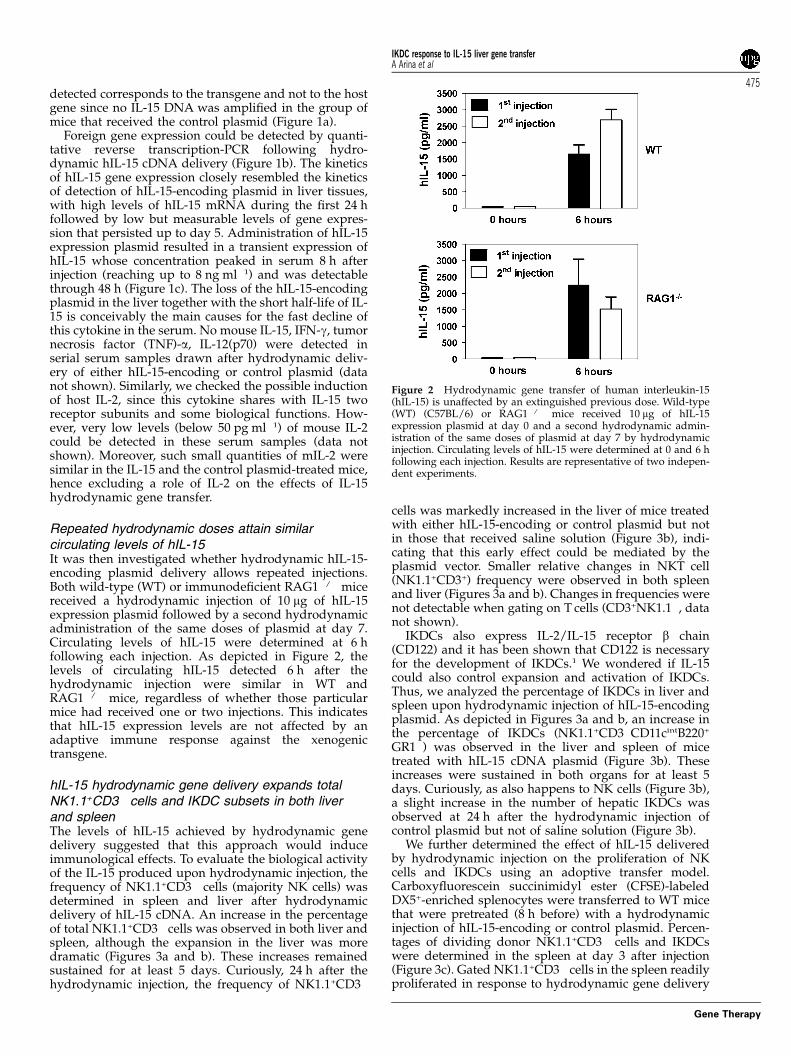
Repeated hydrodynamic doses attain similarcirculating levels of hIL-15It was then investigated whether hydrodynamic hIL-15-encoding plasmid delivery allows repeated injections.Both wild-type (WT) or immunodeficient RAG1�/� micereceived a hydrodynamic injection of 10 mg of hIL-15expression plasmid followed by a second hydrodynamicadministration of the same doses of plasmid at day 7.Circulating levels of hIL-15 were determined at 6 hfollowing each injection. As depicted in Figure 2, thelevels of circulating hIL-15 detected 6 h after thehydrodynamic injection were similar in WT andRAG1�/� mice, regardless of whether those particularmice had received one or two injections. This indicatesthat hIL-15 expression levels are not affected by anadaptive immune response against the xenogenictransgene.
hIL-15 hydrodynamic gene delivery expands totalNK1.1+CD3� cells and IKDC subsets in both liverand spleenThe levels of hIL-15 achieved by hydrodynamic genedelivery suggested that this approach would induceimmunological effects. To evaluate the biological activityof the IL-15 produced upon hydrodynamic injection, thefrequency of NK1.1+CD3� cells (majority NK cells) wasdetermined in spleen and liver after hydrodynamicdelivery of hIL-15 cDNA. An increase in the percentageof total NK1.1+CD3� cells was observed in both liver andspleen, although the expansion in the liver was moredramatic (Figures 3a and b). These increases remainedsustained for at least 5 days. Curiously, 24 h after thehydrodynamic injection, the frequency of NK1.1+CD3�
cells was markedly increased in the liver of mice treatedwith either hIL-15-encoding or control plasmid but notin those that received saline solution (Figure 3b), indi-cating that this early effect could be mediated by theplasmid vector. Smaller relative changes in NKT cell(NK1.1+CD3+) frequency were observed in both spleenand liver (Figures 3a and b). Changes in frequencies werenot detectable when gating on T cells (CD3+NK1.1�, datanot shown).
IKDCs also express IL-2/IL-15 receptor b chain(CD122) and it has been shown that CD122 is necessaryfor the development of IKDCs.1 We wondered if IL-15could also control expansion and activation of IKDCs.Thus, we analyzed the percentage of IKDCs in liver andspleen upon hydrodynamic injection of hIL-15-encodingplasmid. As depicted in Figures 3a and b, an increase inthe percentage of IKDCs (NK1.1+CD3�CD11cintB220+
GR1�) was observed in the liver and spleen of micetreated with hIL-15 cDNA plasmid (Figure 3b). Theseincreases were sustained in both organs for at least 5days. Curiously, as also happens to NK cells (Figure 3b),a slight increase in the number of hepatic IKDCs wasobserved at 24 h after the hydrodynamic injection ofcontrol plasmid but not of saline solution (Figure 3b).
We further determined the effect of hIL-15 deliveredby hydrodynamic injection on the proliferation of NKcells and IKDCs using an adoptive transfer model.Carboxyfluorescein succinimidyl ester (CFSE)-labeledDX5+-enriched splenocytes were transferred to WT micethat were pretreated (8 h before) with a hydrodynamicinjection of hIL-15-encoding or control plasmid. Percen-tages of dividing donor NK1.1+CD3� cells and IKDCswere determined in the spleen at day 3 after injection(Figure 3c). Gated NK1.1+CD3� cells in the spleen readilyproliferated in response to hydrodynamic gene delivery
Figure 2 Hydrodynamic gene transfer of human interleukin-15(hIL-15) is unaffected by an extinguished previous dose. Wild-type(WT) (C57BL/6) or RAG1�/� mice received 10 mg of hIL-15expression plasmid at day 0 and a second hydrodynamic admin-istration of the same doses of plasmid at day 7 by hydrodynamicinjection. Circulating levels of hIL-15 were determined at 0 and 6 hfollowing each injection. Results are representative of two indepen-dent experiments.
IKDC response to IL-15 liver gene transferA Arina et al
475
Gene Therapy
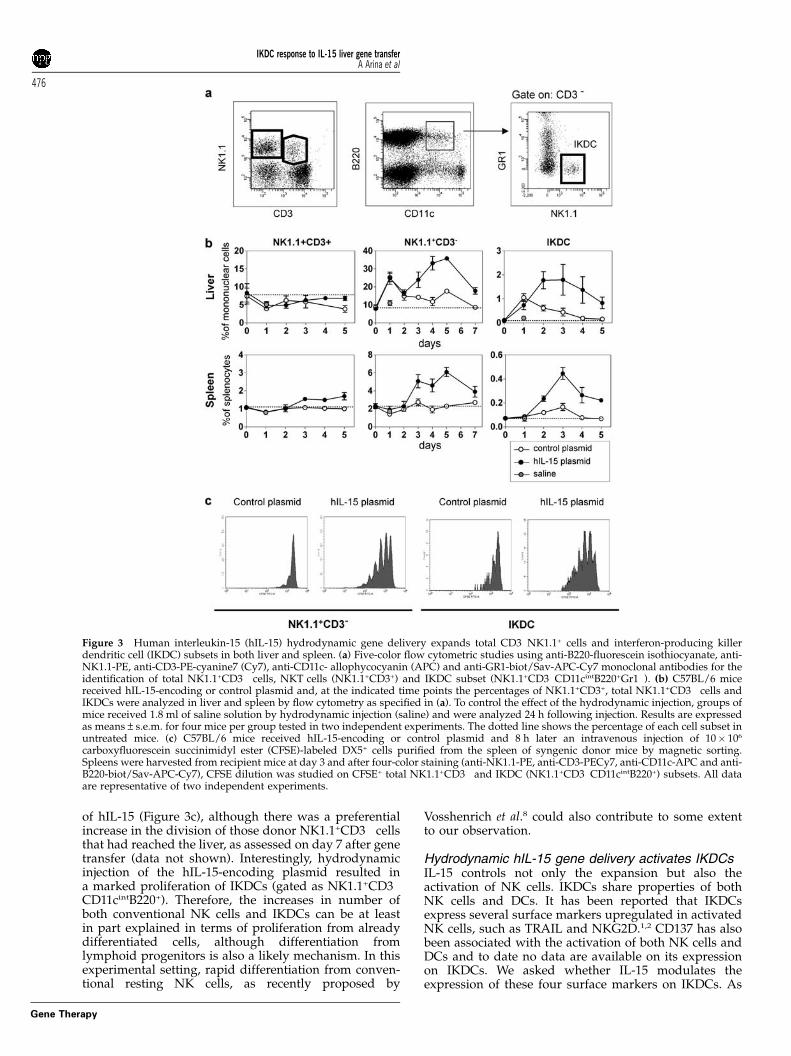
of hIL-15 (Figure 3c), although there was a preferentialincrease in the division of those donor NK1.1+CD3� cellsthat had reached the liver, as assessed on day 7 after genetransfer (data not shown). Interestingly, hydrodynamicinjection of the hIL-15-encoding plasmid resulted ina marked proliferation of IKDCs (gated as NK1.1+CD3�
CD11cintB220+). Therefore, the increases in number ofboth conventional NK cells and IKDCs can be at leastin part explained in terms of proliferation from alreadydifferentiated cells, although differentiation fromlymphoid progenitors is also a likely mechanism. In thisexperimental setting, rapid differentiation from conven-tional resting NK cells, as recently proposed by
Vosshenrich et al.8 could also contribute to some extentto our observation.
Hydrodynamic hIL-15 gene delivery activates IKDCsIL-15 controls not only the expansion but also theactivation of NK cells. IKDCs share properties of bothNK cells and DCs. It has been reported that IKDCsexpress several surface markers upregulated in activatedNK cells, such as TRAIL and NKG2D.1,2 CD137 has alsobeen associated with the activation of both NK cells andDCs and to date no data are available on its expressionon IKDCs. We asked whether IL-15 modulates theexpression of these four surface markers on IKDCs. As
Figure 3 Human interleukin-15 (hIL-15) hydrodynamic gene delivery expands total CD3�NK1.1+ cells and interferon-producing killerdendritic cell (IKDC) subsets in both liver and spleen. (a) Five-color flow cytometric studies using anti-B220-fluorescein isothiocyanate, anti-NK1.1-PE, anti-CD3-PE-cyanine7 (Cy7), anti-CD11c- allophycocyanin (APC) and anti-GR1-biot/Sav-APC-Cy7 monoclonal antibodies for theidentification of total NK1.1+CD3� cells, NKT cells (NK1.1+CD3+) and IKDC subset (NK1.1+CD3�CD11cintB220+Gr1�). (b) C57BL/6 micereceived hIL-15-encoding or control plasmid and, at the indicated time points the percentages of NK1.1+CD3+, total NK1.1+CD3� cells andIKDCs were analyzed in liver and spleen by flow cytometry as specified in (a). To control the effect of the hydrodynamic injection, groups ofmice received 1.8 ml of saline solution by hydrodynamic injection (saline) and were analyzed 24 h following injection. Results are expressedas means±s.e.m. for four mice per group tested in two independent experiments. The dotted line shows the percentage of each cell subset inuntreated mice. (c) C57BL/6 mice received hIL-15-encoding or control plasmid and 8 h later an intravenous injection of 10� 106
carboxyfluorescein succinimidyl ester (CFSE)-labeled DX5+ cells purified from the spleen of syngenic donor mice by magnetic sorting.Spleens were harvested from recipient mice at day 3 and after four-color staining (anti-NK1.1-PE, anti-CD3-PECy7, anti-CD11c-APC and anti-B220-biot/Sav-APC-Cy7), CFSE dilution was studied on CFSE+ total NK1.1+CD3� and IKDC (NK1.1+CD3�CD11cintB220+) subsets. All dataare representative of two independent experiments.
IKDC response to IL-15 liver gene transferA Arina et al
476
Gene Therapy
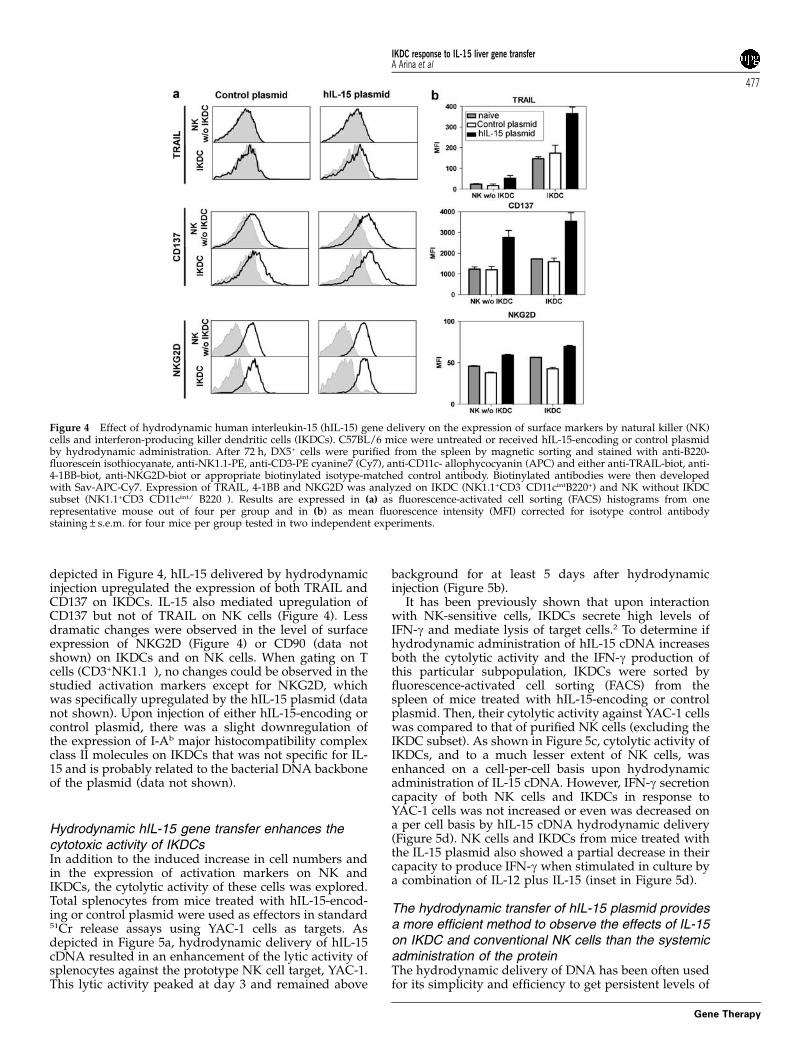
depicted in Figure 4, hIL-15 delivered by hydrodynamicinjection upregulated the expression of both TRAIL andCD137 on IKDCs. IL-15 also mediated upregulation ofCD137 but not of TRAIL on NK cells (Figure 4). Lessdramatic changes were observed in the level of surfaceexpression of NKG2D (Figure 4) or CD90 (data notshown) on IKDCs and on NK cells. When gating on Tcells (CD3+NK1.1�), no changes could be observed in thestudied activation markers except for NKG2D, whichwas specifically upregulated by the hIL-15 plasmid (datanot shown). Upon injection of either hIL-15-encoding orcontrol plasmid, there was a slight downregulation ofthe expression of I-Ab major histocompatibility complexclass II molecules on IKDCs that was not specific for IL-15 and is probably related to the bacterial DNA backboneof the plasmid (data not shown).
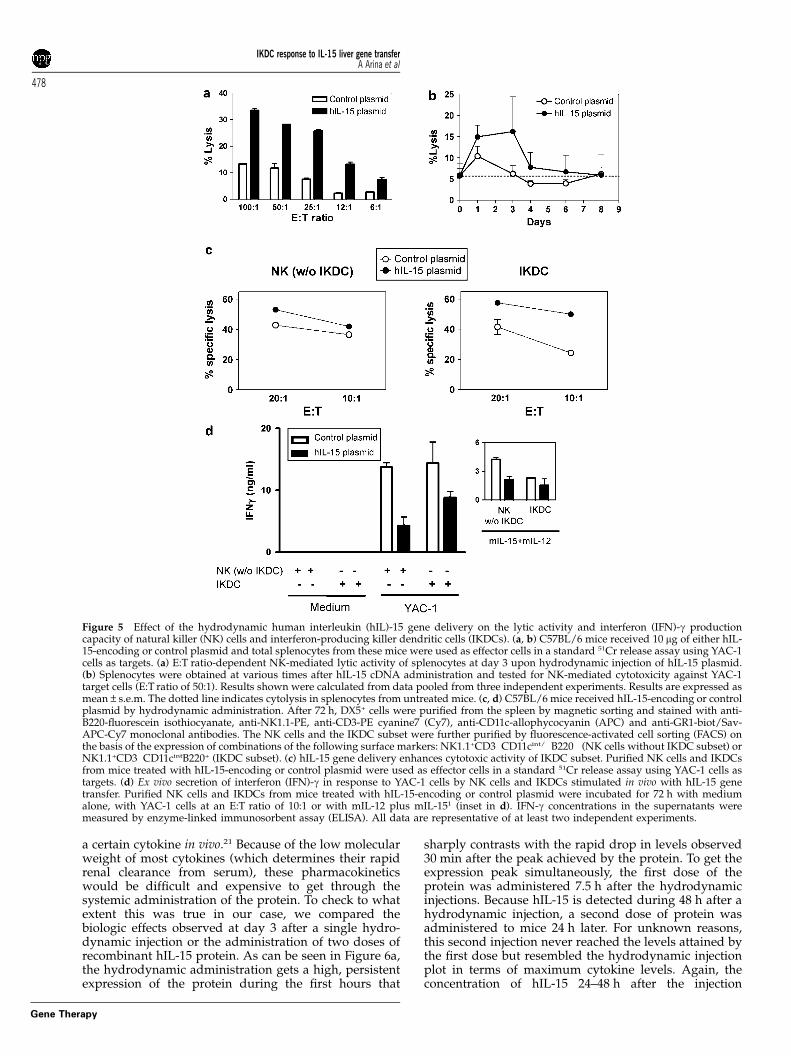
Hydrodynamic hIL-15 gene transfer enhances thecytotoxic activity of IKDCsIn addition to the induced increase in cell numbers andin the expression of activation markers on NK andIKDCs, the cytolytic activity of these cells was explored.Total splenocytes from mice treated with hIL-15-encod-ing or control plasmid were used as effectors in standard51Cr release assays using YAC-1 cells as targets. Asdepicted in Figure 5a, hydrodynamic delivery of hIL-15cDNA resulted in an enhancement of the lytic activity ofsplenocytes against the prototype NK cell target, YAC-1.This lytic activity peaked at day 3 and remained above
background for at least 5 days after hydrodynamicinjection (Figure 5b).
It has been previously shown that upon interactionwith NK-sensitive cells, IKDCs secrete high levels ofIFN-g and mediate lysis of target cells.2 To determine ifhydrodynamic administration of hIL-15 cDNA increasesboth the cytolytic activity and the IFN-g production ofthis particular subpopulation, IKDCs were sorted byfluorescence-activated cell sorting (FACS) from thespleen of mice treated with hIL-15-encoding or controlplasmid. Then, their cytolytic activity against YAC-1 cellswas compared to that of purified NK cells (excluding theIKDC subset). As shown in Figure 5c, cytolytic activity ofIKDCs, and to a much lesser extent of NK cells, wasenhanced on a cell-per-cell basis upon hydrodynamicadministration of IL-15 cDNA. However, IFN-g secretioncapacity of both NK cells and IKDCs in response toYAC-1 cells was not increased or even was decreased ona per cell basis by hIL-15 cDNA hydrodynamic delivery(Figure 5d). NK cells and IKDCs from mice treated withthe IL-15 plasmid also showed a partial decrease in theircapacity to produce IFN-g when stimulated in culture bya combination of IL-12 plus IL-15 (inset in Figure 5d).
The hydrodynamic transfer of hIL-15 plasmid providesa more efficient method to observe the effects of IL-15on IKDC and conventional NK cells than the systemicadministration of the proteinThe hydrodynamic delivery of DNA has been often usedfor its simplicity and efficiency to get persistent levels of
Figure 4 Effect of hydrodynamic human interleukin-15 (hIL-15) gene delivery on the expression of surface markers by natural killer (NK)cells and interferon-producing killer dendritic cells (IKDCs). C57BL/6 mice were untreated or received hIL-15-encoding or control plasmidby hydrodynamic administration. After 72 h, DX5+ cells were purified from the spleen by magnetic sorting and stained with anti-B220-fluorescein isothiocyanate, anti-NK1.1-PE, anti-CD3-PE cyanine7 (Cy7), anti-CD11c- allophycocyanin (APC) and either anti-TRAIL-biot, anti-4-1BB-biot, anti-NKG2D-biot or appropriate biotinylated isotype-matched control antibody. Biotinylated antibodies were then developedwith Sav-APC-Cy7. Expression of TRAIL, 4-1BB and NKG2D was analyzed on IKDC (NK1.1+CD3�CD11cintB220+) and NK without IKDCsubset (NK1.1+CD3�CD11cint/�B220�). Results are expressed in (a) as fluorescence-activated cell sorting (FACS) histograms from onerepresentative mouse out of four per group and in (b) as mean fluorescence intensity (MFI) corrected for isotype control antibodystaining±s.e.m. for four mice per group tested in two independent experiments.
IKDC response to IL-15 liver gene transferA Arina et al
477
Gene Therapy
a certain cytokine in vivo.21 Because of the low molecularweight of most cytokines (which determines their rapidrenal clearance from serum), these pharmacokineticswould be difficult and expensive to get through thesystemic administration of the protein. To check to whatextent this was true in our case, we compared thebiologic effects observed at day 3 after a single hydro-dynamic injection or the administration of two doses ofrecombinant hIL-15 protein. As can be seen in Figure 6a,the hydrodynamic administration gets a high, persistentexpression of the protein during the first hours that
sharply contrasts with the rapid drop in levels observed30 min after the peak achieved by the protein. To get theexpression peak simultaneously, the first dose of theprotein was administered 7.5 h after the hydrodynamicinjections. Because hIL-15 is detected during 48 h after ahydrodynamic injection, a second dose of protein wasadministered to mice 24 h later. For unknown reasons,this second injection never reached the levels attained bythe first dose but resembled the hydrodynamic injectionplot in terms of maximum cytokine levels. Again, theconcentration of hIL-15 24–48 h after the injection
Figure 5 Effect of the hydrodynamic human interleukin (hIL)-15 gene delivery on the lytic activity and interferon (IFN)-g productioncapacity of natural killer (NK) cells and interferon-producing killer dendritic cells (IKDCs). (a, b) C57BL/6 mice received 10 mg of either hIL-15-encoding or control plasmid and total splenocytes from these mice were used as effector cells in a standard 51Cr release assay using YAC-1cells as targets. (a) E:T ratio-dependent NK-mediated lytic activity of splenocytes at day 3 upon hydrodynamic injection of hIL-15 plasmid.(b) Splenocytes were obtained at various times after hIL-15 cDNA administration and tested for NK-mediated cytotoxicity against YAC-1target cells (E:T ratio of 50:1). Results shown were calculated from data pooled from three independent experiments. Results are expressed asmean±s.e.m. The dotted line indicates cytolysis in splenocytes from untreated mice. (c, d) C57BL/6 mice received hIL-15-encoding or controlplasmid by hydrodynamic administration. After 72 h, DX5+ cells were purified from the spleen by magnetic sorting and stained with anti-B220-fluorescein isothiocyanate, anti-NK1.1-PE, anti-CD3-PE cyanine7 (Cy7), anti-CD11c-allophycocyanin (APC) and anti-GR1-biot/Sav-APC-Cy7 monoclonal antibodies. The NK cells and the IKDC subset were further purified by fluorescence-activated cell sorting (FACS) onthe basis of the expression of combinations of the following surface markers: NK1.1+CD3�CD11cint/�B220� (NK cells without IKDC subset) orNK1.1+CD3�CD11cintB220+ (IKDC subset). (c) hIL-15 gene delivery enhances cytotoxic activity of IKDC subset. Purified NK cells and IKDCsfrom mice treated with hIL-15-encoding or control plasmid were used as effector cells in a standard 51Cr release assay using YAC-1 cells astargets. (d) Ex vivo secretion of interferon (IFN)-g in response to YAC-1 cells by NK cells and IKDCs stimulated in vivo with hIL-15 genetransfer. Purified NK cells and IKDCs from mice treated with hIL-15-encoding or control plasmid were incubated for 72 h with mediumalone, with YAC-1 cells at an E:T ratio of 10:1 or with mIL-12 plus mIL-151 (inset in d). IFN-g concentrations in the supernatants weremeasured by enzyme-linked immunosorbent assay (ELISA). All data are representative of at least two independent experiments.
IKDC response to IL-15 liver gene transferA Arina et al
478
Gene Therapy
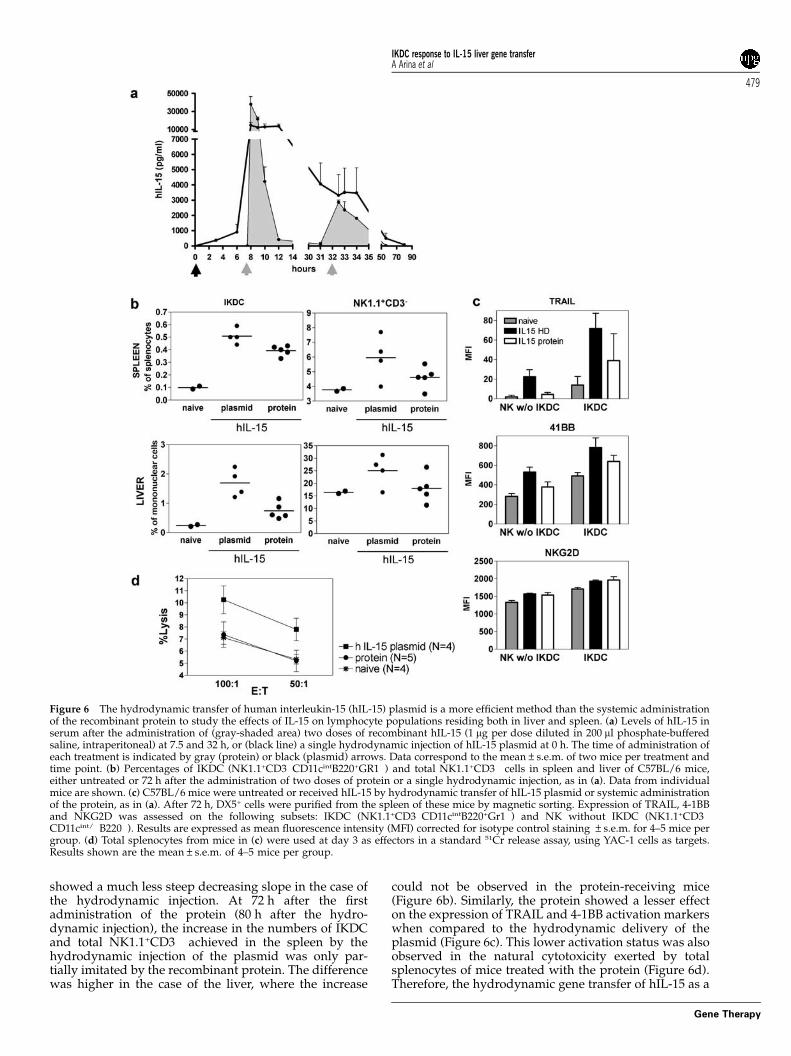
showed a much less steep decreasing slope in the case ofthe hydrodynamic injection. At 72 h after the firstadministration of the protein (80 h after the hydro-dynamic injection), the increase in the numbers of IKDCand total NK1.1+CD3� achieved in the spleen by thehydrodynamic injection of the plasmid was only par-tially imitated by the recombinant protein. The differencewas higher in the case of the liver, where the increase
could not be observed in the protein-receiving mice(Figure 6b). Similarly, the protein showed a lesser effecton the expression of TRAIL and 4-1BB activation markerswhen compared to the hydrodynamic delivery of theplasmid (Figure 6c). This lower activation status was alsoobserved in the natural cytotoxicity exerted by totalsplenocytes of mice treated with the protein (Figure 6d).Therefore, the hydrodynamic gene transfer of hIL-15 as a
Figure 6 The hydrodynamic transfer of human interleukin-15 (hIL-15) plasmid is a more efficient method than the systemic administrationof the recombinant protein to study the effects of IL-15 on lymphocyte populations residing both in liver and spleen. (a) Levels of hIL-15 inserum after the administration of (gray-shaded area) two doses of recombinant hIL-15 (1 mg per dose diluted in 200 ml phosphate-bufferedsaline, intraperitoneal) at 7.5 and 32 h, or (black line) a single hydrodynamic injection of hIL-15 plasmid at 0 h. The time of administration ofeach treatment is indicated by gray (protein) or black (plasmid) arrows. Data correspond to the mean±s.e.m. of two mice per treatment andtime point. (b) Percentages of IKDC (NK1.1+CD3�CD11cintB220+GR1�) and total NK1.1+CD3� cells in spleen and liver of C57BL/6 mice,either untreated or 72 h after the administration of two doses of protein or a single hydrodynamic injection, as in (a). Data from individualmice are shown. (c) C57BL/6 mice were untreated or received hIL-15 by hydrodynamic transfer of hIL-15 plasmid or systemic administrationof the protein, as in (a). After 72 h, DX5+ cells were purified from the spleen of these mice by magnetic sorting. Expression of TRAIL, 4-1BBand NKG2D was assessed on the following subsets: IKDC (NK1.1+CD3�CD11cintB220+Gr1�) and NK without IKDC (NK1.1+CD3�
CD11cint/�B220�). Results are expressed as mean fluorescence intensity (MFI) corrected for isotype control staining ±s.e.m. for 4–5 mice pergroup. (d) Total splenocytes from mice in (c) were used at day 3 as effectors in a standard 51Cr release assay, using YAC-1 cells as targets.Results shown are the mean±s.e.m. of 4–5 mice per group.
IKDC response to IL-15 liver gene transferA Arina et al
479
Gene Therapy

method to enhance innate immunity is more efficientthan the systemic administration of the protein.
Taken together, all these data indicate that hydro-dynamic delivery of IL-15 cDNA provides a usefulexperimental method to study the in vivo effects of IL-15on IKDCs and NK cells.
Discussion
This study proves the concept that liver gene transfer inmice provides a source of IL-15 that thereby impacts thebiology of IKDCs and NK cells upregulating both theirnumbers and their cytolytic function.
The liver seems to be the organ in which the biologicaleffects of this gene transfer approach are more pro-nounced. Local concentrations of IL-15 at the sourcepoint are conceivably higher and the cytokine may lastlonger in this organ where it might remain adsorbed ontoconstitutively expressed IL-15Ra.31 In this study, weidentify small numbers of IKDCs residing in theuntreated liver that increase their number followingtransient gene expression of hIL-15 in this organ. Fromnow on, IKDCs should be considered an additionalcellular player in the incompletely understood functionsof the liver immune system.32
Functional plasticity seems to be a characteristicfeature of IKDCs1 and NKDCs33 because these cellsseem to modulate the expression of particular sets ofreceptors depending on the stimulus provided to them.In our study, we observed an upregulation of activationmarkers typically expressed by cytotoxic effector cells,TRAIL and CD137, following the administration of hIL-15 gene. Interestingly, IL-15 increases surface expressionlevels of TRAIL, a molecule reportedly involved in thetumoricidal effects of IKDCs promoted by IL-2 plusimatinib.2,6 The surface antigen CD137 was constitutivelyfound on these cells and its expression was increased as aresult of IL-15 gene therapy. Hence, it will be interestingto investigate whether the therapeutic cellular immunityenhanced by agonistic anti-CD137 monoclonal antibo-dies is in part mediated via these cells as is the case withNK cells.34 Upregulation of these activation markers byIKDCs correlated with an augmented cytotoxic activity.Nonetheless, under IL-15 in vivo gene transfer there wasan unexpected partial reduction in the capability ofsorted NK cells and IKDCs to produce IFN-g in culture inresponse to tumor cells. This indicates that hydro-dynamic IL-15 gene transfer differentially modulatescytokine production and cytotoxicity as elicited byYAC-1 cells.
In our model, IL-15 expression is transient andcirculating levels are relatively modest. Mechanismsmediating extinction of gene expression encompass lossof the plasmid vector and probably silencing of thecytomegalovirus promoter.35 Higher and more sustainedlevels of circulating IL-15 upon gene transfer couldbe achieved by exogenously regulatable promoterstargeting transcription to the liver and using liver-tropicviral vectors such as those based on ‘gut-less’ adeno-viruses or adeno-associated viruses.36 Alternatively, away to optimize the pharmacokinetics of IL-15 is tocomplex it with an IgFc chimeric protein that encom-passes IL-15Ra, in such a way that renal elimination isretarded and trans-presentation is favored.30,37 Overall,
hydrodynamic gene transfer was by far a better methodto get sustained levels of the cytokine in serum, whencompared to the systemic administration of the protein.Therefore, hydrodynamic gene delivery represents ahighly efficient method to study the effects of IL-15 onIKDC and conventional NK cells, especially in the liver.
Despite the transient nature of gene expression in oursystem, we were able to observe biological effects at timepoints in which no detectable IL-15 could be found asprotein or as mRNA. This could be due to the fact thatcirculating IL-15 can be trapped and trans-presentedadsorbed onto recycling IL-15Ra in cells such as DCs orstromal fibroblastic cells.20 Alternatively, this couldhappen because a short in vivo exposure to IL-15 imprintsa response in the IKDCs and NK cells that is detected atlater time points. Although we readily observe IL-15trans-presentation using DCs pulsed in vitro withrecombinant hIL-15, we could not ascertain this phe-nomenon in DCs isolated from the spleen followinghydrodynamic gene transfer of hIL-15 in vivo (data notshown). Higher levels of circulating cytokine could becritical for this purpose as proposed by Sato et al.38 Itremains to be seen if IKDCs also sense IL-15 viaDC-mediated trans-presentation.
Contrary to IL-2,22 the hydrodynamic delivery of IL-15did not show curative effects in several murine cancermodels (B16F10, RENCA, CT26, RMA-S; data notshown). This difference might be related to the lowerlevels of cytokine achieved by our plasmid, or to anintrinsic inferiority of IL-15 when compared to IL-2 interms of activation of anti-tumor functions. In thisregard, NKDC expanded in IL-15 had to be pretreatedwith IL-12 plus IL-18 to exhibit antitumor activity.39 Inany case, rodent data showing lesser toxicity and a muchmore consistent immunostimulatory profile back thechoice of IL-15 over IL-2 for immunotherapy.40 Indeed,the hydrodynamic hIL-15 gene transfer has clear biologiceffects in our hands, and we believe that it could be usedin combination with other immunotherapy agents toenhance their effectiveness, as has already been docu-mented with the recombinant protein.41,42 Our studyindicates that these beneficial effects could be greatlypromoted by the higher efficiency of the hydrodynamicinjections. Also, it would be interesting to study theeffects of hIL-15 liver gene transfer of on other cellpopulations, such as memory T cells, due to theirreported dependence on IL-15.13 These points arecurrently under exploration.
The scanty numbers of IKDCs make their isolation andstudy very difficult and therefore administration of IL-15as a soluble cytokine or by gene transfer could be avaluable tool to explore IKDC’s biology, much in thesame ways as sFLT-3L hydrodynamic gene transfer wasuseful for the study of different DC populations.22,43 As atool to increase IKDC numbers, hydrodynamic genetransfer of IL-15 can be valuable to address and settle thepolemics on the true nature of IKDCs.7,44 From thisexperimental point of view, hIL-15 can be readministeredto mice in successive hydrodynamic injections of theencoding plasmid, thereby achieving comparable levelsof circulating cytokine.
As a whole, this study (i) demonstrates that the livercan be used as an endogenous source of secretable IL-15following hepatocyte gene transfer and (ii) providesexperimental evidence for a functional link between
IKDC response to IL-15 liver gene transferA Arina et al
480
Gene Therapy

circulating IL-15 and the number and effector function ofIKDCs.
Materials and methods
Mice and cell linesC57BL/6 mice (6–10 weeks old) were purchased fromHarlan Laboratories (Barcelona, Spain). RAG1�/� miceon C57BL/6 background from the Jackson Laboratory(Bar Harbor, ME, USA) were bred in our animal facility.All animal procedures were conducted under institu-tional guidelines (study approval number 003/02) thatcomply with national laws and policies. YAC-1 cells wereobtained from American Type Culture Collection.
Plasmids and hIL-15 proteinThe pVkL/IL-15IRESneo expression plasmid, whichcarries a modified cDNA encoding for hIL-15 pre-proteinbearing the IgK light-chain signal peptide (VkL) underthe transcriptional control of the cytomegalovirus pro-moter/enhancer (hIL-15 plasmid), and the empty pIR-ES1neo (control plasmid) were produced and purified asdescribed.28 hIL-15 recombinant protein purchased fromPeprotech (Rocky Hill, NJ, USA) was intraperitoneallyinjected in 1 mg doses, diluted in 200 ml of phosphate-buffered saline (PBS) per dose.
Hydrodynamic injectionsC57BL/6 or RAG1�/� mice received an intravenousinjection (tail vein) of 10 mg of either hIL-15-encoding orcontrol plasmid in a volume of 100 ml kg�1 using a 27-Gneedle at a rate of 0.4 ml s�1, as previously described.22
Antibodies and flow cytometryCells isolated from liver and spleens were pretreatedwith anti-CD16/32 (eBioscience, San Diego, CA, USA) toreduce nonspecific staining. Monoclonal antibodiesto the following mouse antigens were conjugated tofluorescein isothiocyanate, phycoerythrin (PE), PE-cya-nine7 (PE-Cy7), allophycocyanin (APC) or biotin: CD3(145-2C11), CD11c (HL3), NK1.1 (PK-136), B220 (RA3-6B2), GR1 (RB6-8C5), I-Ab (KH74), CD137 (17B5) andappropriate isotype controls were obtained from BDBiosciences (San Diego, CA, USA); NKG2D (MI6) andTRAIL (N2B2) were purchased from eBioscience. Bioti-nylated antibodies were visualized with streptavidin(Sav)-APC-Cy7 (BD Biosciences). FACSAria and FACS-Calibur cytometers were used for cell acquisition, anddata analysis was performed using FACS DiVa (BDBiosciences) and FlowJo 7.2.1 (Tree Star Inc., Ashland,OR, USA).
Total RNA and DNA purification and real-time PCRTotal RNA was extracted from liver tissue using TRIreagent (Sigma-Aldrich, Barcelona, Spain). We treatedRNA with DNaseI (Gibco-BRL, Gaithersburg, MD, USA)prior to reverse transcription with M-MLV reversetranscriptase (Gibco-BRL) in the presence of RNaseOUT(Gibco-BRL). Real-time PCR was performed using aniCycler (Bio-Rad, Hercules, CA, USA) and the iQ SYBRGreen Supermix (Bio-Rad). hIL-15 cDNA was amplifiedusing primers GCCAACTGGGTGAATGTAATAAG andGAATCAATTGCAATCAAGAAGTG specific for hIL-15.Mouse b-actin cDNA was amplified as control using
primers CGCGTCCACCCGCGAG and CCTGGTGCCTAGGGCG. Alternatively, total DNA was isolated fromliver tissue using the DNeasy tissue Kit (Qiagen, Madrid,Spain). hIL-15 DNA was amplified by real-time PCRusing the same pair of primers described above. Mousegenomic DNA for SOCS3, used as control, was amplifiedusing primers TACCAGCTGGTGGTGAACGC andGCGGCATGTAGTGGTGCACC. To monitor the specifi-city, final PCR products were analyzed by melting curvesand electrophoresis. The amount of hIL-15 was ex-pressed as arbitrary units defined as the n-fold differencerelative to the control gene b-actin (for transcription) orSOCS3 (for genomic DNA) 2Ct�100, where DCt repre-sents the difference in threshold cycle between thecontrol and target genes).
Cell isolationLiver non-parenchymal cells and splenocytes wereisolated from mice injected with hIL-15-encoding orcontrol plasmid at the indicated time points. Briefly,spleens were incubated in collagenase and DNase(Roche, Basel, Switzerland) for 15 min at 37 1C, and thenboth livers and spleens were mechanically disruptedbefore being passed through a sterile 70-mm nylon meshfilter (BD Falcon, San Jose, CA, USA). Hepatocytes wereexcluded from the liver samples by discarding the pelletsafter a single-step Percoll gradient (GE Healthcare,Chalfont St Giles, UK). In either case, erythrocytes werelysed with ACK buffer. In certain experiments, spleno-cytes were enriched in DX5+ cells using immunomag-netic beads according to the manufacturer’s protocol(Miltenyi Biotec, Bergisch Gladbach, Germany). At thisstep, the cell suspension was composed of 50–70% of NKcells, including 3–8% of IKDCs. The NK cells and IKDCswere further purified on a FACSAria sorter on the basisof the expression of NK1.1+CD3�CD11cneg/intB220� (NKcells without IKDC subset) or NK1.1+CD3�CD11cintB220+
(IKDCs). Purity exceeded 90% in every experiment.
CFSE labeling of cells and adoptive transferEnriched DX5+ cells were washed twice with phosphate-buffered saline and resuspended at 107 cells ml�1 in 5 mM
CFSE (Sigma-Aldrich) for 15 min at room temperature.The labeling was stopped by washing twice with coldphosphate-buffered saline 10% fetal bovine serum. Theproliferation of adoptively transferred NK cells(CFSE+NK1.1+CD3�) and IKDCs (CFSE+NK1.1+CD3�
CD11cintB220+) was analyzed by CFSE dilution using aFACSAria cytometer.
Ex vivo analysis of effector functionTo determine IFN-g secretion capacity, FACS-sorted NKcells and IKDCs from mice treated with hIL-15-encodingor control plasmid were incubated for 72 h with mediumalone or with YAC-1 cells (104 targets per well) at a 10:1E:T ratio. In both cases, human IL-2 (Chiron, Emeryville,CA, USA) was present at 100 IU ml�1. When indicatedmIL-15 (R&D, Minneapolis, MN, USA) at 50 ng ml�1 andmIL-12 (Peprotech) at 1 ng ml�1 were added to thecultures.1 Cytotoxic activity of either total splenocytesor sorted NK cells and IKDCs from mice treated withhIL-15-encoding or control plasmid was assessedby standard 5 h 51Cr release assays,34 using 51Cr-labeledYAC-1 cells as targets (4� 104 targets per well). The
IKDC response to IL-15 liver gene transferA Arina et al
481
Gene Therapy

51Cr-release was counted using TopCount NXT (Perkin-Elmer, Waltham, MA, USA).
Enzyme-linked immunosorbent assayhIL-15, mTNF-a, mIFN-g, mIL-12(p70) and mIL-2 fromindividual sera and mouse IFN-g content in the super-natants from the ex vivo cultured cells were quantified bycommercial enzyme-linked immunosorbent assay (ELI-SA) (OptEIA; BD Biosciences). The ELISA pair OptEIAset for hIL-15 shows no cross-reactivity with any of thecytokines tested (for example, mouse IL-1b, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-9, IL-10, IL-12 p70, IL-15, GM-CSF,IFN-g, MCP-1, TCA-3, TNF) according to the manu-facturer. For the quantification of mIL-15, an ELISA setfrom R&D was used.
Abbreviation
IKDC, interferon-producing killer dendritic cell
Acknowledgements
This study was financially supported by Ministerio deEducacion y Ciencia (MEC-SAF2005-03131), Departa-mentos de Educacion y Salud del Gobierno de Navarra,Redes tematicas de investigacion cooperativa and ‘UTEfor project FIMA.’ Associazione Italiana per la Ricerca sulCancro funded SF’s work. OM and AAr are recipients ofscholarships from Ministerio de Educacion y Ciencia andFondo de Investigacion Sanitaria. SH-S is supported byAsociacion Espanola Contra el Cancer.
References
1 Chan CW, Crafton E, Fan HN, Flook J, Yoshimura K, Skarica Met al. Interferon-producing killer dendritic cells provide a linkbetween innate and adaptive immunity. Nat Med 2006; 12:207–213.
2 Taieb J, Chaput N, Menard C, Apetoh L, Ullrich E, Bonmort Met al. A novel dendritic cell subset involved in tumorimmunosurveillance. Nat Med 2006; 12: 214–219.
3 Pillarisetty VG, Katz SC, Bleier JI, Shah AB, Dematteo RP.Natural killer dendritic cells have both antigen presenting andlytic function and in response to CpG produce IFN-gamma viaautocrine IL-12. J Immunol 2005; 174: 2612–2618.
4 Plitas G, Chaudhry UI, Kingham TP, Raab JR, DeMatteo RP. NKdendritic cells are innate immune responders to Listeriamonocytogenes infection. J Immunol 2007; 178: 4411–4416.
5 Welner RS, Pelayo R, Garrett KP, Chen X, Perry SS, Sun XH et al.Interferon-producing killer dendritic cells (IKDC) arise via aunique differentiation pathway from primitive c-kitHiCD62L+
lymphoid progenitors. Blood 2007; 22: 22.6 Ullrich E, Bonmort M, Mignot G, Chaput N, Taieb J, Menard C
et al. Therapy-induced tumor immunosurveillance involves IFN-producing killer dendritic cells. Cancer Res 2007; 67: 851–853.
7 Spits H, Lanier LL. Natural killer or dendritic: what’s in a name?Immunity 2007; 26: 11–16.
8 Vosshenrich CA, Lesjean-Pottier S, Hasan M, Richard-Le Goff O,Corcuff E, Mandelboim O et al. CD11cloB220+ interferon-producing killer dendritic cells are activated natural killer cells.J Exp Med 2007; 204: 2569–2578.
9 Blasius AL, Barchet W, Cella M, Colonna M. Development andfunction of murine B220+CD11c+NK1.1+ cells identify them as asubset of NK cells. J Exp Med 2007; 204: 2561–2568.
10 Caminschi I, Ahmet F, Heger K, Brady J, Nutt SL, Vremec D et al.Putative IKDCs are functionally and developmentally similar tonatural killer cells, but not to dendritic cells. J Exp Med 2007; 204:2579–2590.
11 Budagian V, Bulanova E, Paus R, Bulfone-Paus S. IL-15/IL-15receptor biology: a guided tour through an expanding universe.Cytokine Growth Factor Rev 2006; 17: 259–280.
12 Becknell B, Caligiuri MA. Interleukin-2, interleukin-15, and theirroles in human natural killer cells. Adv Immunol 2005; 86:209–239.
13 Kennedy MK, Glaccum M, Brown SN, Butz EA, Viney JL,Embers M et al. Reversible defects in natural killer and memoryCD8T cell lineages in interleukin 15-deficient mice. J Exp Med2000; 191: 771–780.
14 Fehniger TA, Suzuki K, Ponnappan A, VanDeusen JB, CooperMA, Florea SM et al. Fatal leukemia in interleukin 15 transgenicmice follows early expansions in natural killer and memoryphenotype CD8+ T cells. J Exp Med 2001; 193: 219–231.
15 Lodolce JP, Boone DL, Chai S, Swain RE, Dassopoulos T, Trettin Set al. IL-15 receptor maintains lymphoid homeostasis bysupporting lymphocyte homing and proliferation. Immunity1998; 9: 669–676.
16 Koka R, Burkett PR, Chien M, Chai S, Chan F, Lodolce JP et al.Interleukin (IL)-15R[alpha]-deficient natural killer cells survivein normal but not IL-15R[alpha]-deficient mice. J Exp Med 2003;197: 977–984.
17 Carson WE, Giri JG, Lindemann MJ, Linett ML, Ahdieh M,Paxton R et al. Interleukin (IL) 15 is a novel cytokine thatactivates human natural killer cells via components of the IL-2receptor. J Exp Med 1994; 180: 1395–1403.
18 Dubois S, Mariner J, Waldmann TA, Tagaya Y. IL-15Ralpharecycles and presents IL-15 in trans to neighboring cells.Immunity 2002; 17: 537–547.
19 Waldmann TA. The biology of interleukin-2 and interleukin-15:implications for cancer therapy and vaccine design. Nat RevImmunol 2006; 6: 595–601.
20 Kobayashi H, Dubois S, Sato N, Sabzevari H, Sakai Y, Waldmann TAet al. Role of trans-cellular IL-15 presentation in the activation ofNK cell-mediated killing, which leads to enhanced tumorimmunosurveillance. Blood 2005; 105: 721–727.
21 Niidome T, Huang L. Gene therapy progress and prospects:nonviral vectors. Gene Ther 2002; 9: 1647–1652.
22 He Y, Pimenov AA, Nayak JV, Plowey J, Falo Jr LD, Huang L.Intravenous injection of naked DNA encoding secreted flt3ligand dramatically increases the number of dendritic cells andnatural killer cells in vivo. Hum Gene Ther 2000; 11: 547–554.
23 Ortaldo JR, Winkler-Pickett RT, Bere Jr EW, Watanabe M,Murphy WJ, Wiltrout RH. In vivo hydrodynamic delivery ofcDNA encoding IL-2: rapid, sustained redistribution, activationof mouse NK cells, and therapeutic potential in the absence ofNKT cells. J Immunol 2005; 175: 693–699.
24 Hazama S, Noma T, Wang F, Iizuka N, Ogura Y, Yoshimura Ket al. Tumour cells engineered to secrete interleukin-15 augmentanti-tumour immune responses in vivo. Br J Cancer 1999; 80:1420–1426.
25 Di Carlo E, Comes A, Basso S, De Ambrosis A, Meazza R,Musiani P et al. The combined action of IL-15 and IL-12 genetransfer can induce tumor cell rejection without T and NK cellinvolvement. J Immunol 2000; 165: 3111–3118.
26 Kishida T, Asada H, Itokawa Y, Cui FD, Shin-Ya M, Gojo S et al.Interleukin (IL)-21 and IL-15 genetic transfer synergisticallyaugments therapeutic antitumor immunity and promotesregression of metastatic lymphoma. Mol Ther 2003; 8: 552–558.
27 Croce M, Meazza R, Orengo AM, Radic L, De Giovanni B,Gambini C et al. Sequential immunogene therapy with inter-leukin-12- and interleukin-15-engineered neuroblastoma cellscures metastatic disease in syngeneic mice. Clin Cancer Res 2005;11: 735–742.
IKDC response to IL-15 liver gene transferA Arina et al
482
Gene Therapy

28 Meazza R, Lollini PL, Nanni P, De Giovanni C, Gaggero A,Comes A et al. Gene transfer of a secretable form of IL-15 inmurine adenocarcinoma cells: effects on tumorigenicity,metastatic potential and immune response. Int J Cancer 2000;87: 574–581.
29 Vera M, Razquin N, Prieto J, Melero I, Fortes P,Gonzalez-Aseguinolaza G. Intratumoral injection of dendriticcells transduced by an SV40-based vector expressing inter-leukin-15 induces curative immunity mediated by CD8+ Tlymphocytes and NK cells. Mol Ther 2005; 12: 950–959.
30 Stoklasek TA, Schluns KS, Lefrancois L. Combined IL-15/IL-15Ralpha immunotherapy maximizes IL-15 activity in vivo.J Immunol 2006; 177: 6072–6080.
31 Suzuki A, McCall S, Choi SS, Sicklick JK, Huang J, Qi Y et al.Interleukin-15 increases hepatic regenerative activity. J Hepatol2006; 45: 410–418.
32 Mehal WZ, Azzaroli F, Crispe IN. Immunology of the healthyliver: old questions and new insights. Gastroenterology 2001; 120:250–260.
33 Chen L, Calomeni E, Wen J, Ozato K, Shen R, Gao JX. Naturalkiller dendritic cells are an intermediate of developing dendriticcells. J Leukoc Biol 2007; 1: 1.
34 Melero I, Johnston JV, Shufford WW, Mittler RS, Chen L. NK1.1cells express 4-1BB (CDw137) costimulatory molecule and arerequired for tumor immunity elicited by anti-4-1BB monoclonalantibodies. Cell Immunol 1998; 190: 167–172.
35 Qin L, Ding Y, Pahud DR, Chang E, Imperiale MJ, Bromberg JS.Promoter attenuation in gene therapy: interferon-gammaand tumor necrosis factor-alpha inhibit transgene expression.Hum Gene Ther 1997; 8: 2019–2029.
36 Wang L, Hernandez-Alcoceba R, Shankar V, Zabala M,Kochanek S, Sangro B et al. Prolonged and inducible transgeneexpression in the liver using gutless adenovirus: a potentialtherapy for liver cancer. Gastroenterology 2004; 126: 278–289.
37 Rubinstein MP, Kovar M, Purton JF, Cho JH, Boyman O, Surh CDet al. Converting IL-15 to a superagonist by binding to solubleIL-15R{alpha}. Proc Natl Acad Sci USA 2006; 103: 9166–9171.
38 Sato N, Patel HJ, Waldmann TA, Tagaya Y. The IL-15/IL-15Ralpha on cell surfaces enables sustained IL-15 activity andcontributes to the long survival of CD8 memory T cells. Proc NatlAcad Sci USA 2007; 104: 588–593.
39 Chaudhry UI, Plitas G, Burt BM, Kingham TP, Raab JR,DeMatteo RP. NK dendritic cells expanded in IL-15 exhibitantitumor responses in vivo. J Immunol 2007; 179: 4654–4660.
40 Waldmann TA. Effective cancer therapy through immuno-modulation. Annu Rev Med 2006; 57: 65–81.
41 Klebanoff CA, Finkelstein SE, Surman DR, Lichtman MK,Gattinoni L, Theoret MR et al. IL-15 enhances the in vivoantitumor activity of tumor-reactive CD8+ T cells. Proc Natl AcadSci USA 2004; 101: 1969–1974.
42 Waldmann TA. The biology of interleukin-2 and interleukin-15:implications for cancer therapy and vaccine design. Nat RevImmunol 2006; 6: 595–601.
43 Maraskovsky E, Brasel K, Teepe M, Roux ER, Lyman SD,Shortman K et al. Dramatic increase in the numbers offunctionally mature dendritic cells in Flt3 ligand-treated mice:multiple dendritic cell subpopulations identified. J Exp Med1996; 184: 1953–1962.
44 Shortman K, Villadangos JA. Is it a DC, is it an NK? No, it’s anIKDC. Nat Med 2006; 12: 167–168.
IKDC response to IL-15 liver gene transferA Arina et al
483
Gene Therapy
Copyright © 2022 FDOKUMEN




















