
Higher Intensity Exercise Increases Protein Turnover and ...
116
Higher Intensity Exercise Increases Protein Turnover and Daily Protein Requirements of Male Endurance Athletes During Recovery by Carolyn Adams A thesis submitted in conformity with the requirements for the degree of Master of Science in Exercise Science Faculty of Kinesiology and Physical Education University of Toronto © Copyright by Carolyn Adams 2021
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of Higher Intensity Exercise Increases Protein Turnover and ...

Higher Intensity Exercise Increases Protein Turnover and Daily Protein Requirements of Male Endurance Athletes During Recovery
by
Carolyn Adams
A thesis submitted in conformity with the requirements for the degree of Master of Science in Exercise Science
Faculty of Kinesiology and Physical Education University of Toronto
© Copyright by Carolyn Adams 2021

ii
Higher Intensity Exercise Increases Protein Turnover and Daily Protein Requirements of Male Endurance Athletes During Recovery
Carolyn Adams Master of Science Department of Exercise Science University of Toronto 2021
Abstract
Purpose: Examine the effect of endurance exercise intensity on whole-body protein metabolism
and protein requirements during recovery. Methods: Seven male endurance athletes completed
randomized trials (20-km run) at LOW (~55% V̇O2peak), or HIGH (~72% V̇O2peak), intensity.
Participants consumed 0.6g·kg-1 CHO pre-exercise. During 8 h post-exercise, participants
consumed 8.6g·kg-1·d-1 CHO and hourly meals providing 0.93g ·kg-1·d-1 protein, enriched with
indicator AA [13C] phenylalanine. Breath and urine samples were collected to determine whole-
body phenylalanine flux (PheRa), oxidation (PheOx), protein synthesis (PS), breakdown (PB) and
net balance (NB). Results: Exercising CHO oxidation rate was 60% greater in HIGH vs LOW
(P<0.001). PheOx had a large effect size and tended to be greater in HIGH (d=0.844,
P=0.094). NB had a large effect size and tended to be less positive in HIGH (d=0.844, P=0.094).
PheRa, PS and PB had a moderate effect size. Conclusion: Protein requirements are elevated
following high vs low intensity endurance exercise.

iii
Acknowledgements
I would like to thank my supervisor, Dr. Daniel Moore for his support and commitment to my academic growth over the past two years.
I wish to also acknowledge the support of my committee members, Dr. Jenna Gillen and
Dr. Marius Locke for being great teachers and providing their expertise and support. These were instrumental in the completion of my project.
Thank you to all of my lab mates for their encouragement and friendship over the past
two years. In particular, I want to acknowledge the contributions of Dr. Nathan Hodson, Stephanie Estafanos, Hugo Fung, and Cass Tineline-Goodfellow for supporting me in the completion of this project.
Thank you to the Faculty of Kinesiology and Physical Education and to the University of
Toronto for providing me with a place to grow academically and personally over the past seven years. A special thank you to Dean Ira Jacobs for supporting my early interests in research and believing in me.
Thank you to the University of Toronto Track and Field team for being my home away
from home and to Carl Georgevski for supporting my passions and for many years of invaluable mentorship.
Lastly, thank you to my family and friends for your patience, encouragement, and love. I
have truly appreciated you being there for me along this journey.

iv
Table of Contents
Abstract ........................................................................................................................................... ii
Acknowledgements ........................................................................................................................ iii
List of Abbreviation ....................................................................................................................... vi
List of Tables ................................................................................................................................ vii
List of Figures ............................................................................................................................... vii
List of Appendices ........................................................................................................................ vii
Chapter 1. Review of the Literature .................................................................................................1
1.1 Introduction ..........................................................................................................................1
1.2 Introduction to Protein Metabolism .....................................................................................3
1.3 Fuel use during Endurance Exercise ....................................................................................5
1.4 Protein Metabolism and Endurance Exercise .......................................................................9
1.4.1. Muscle protein synthesis and breakdown ..................................................................9
1.4.2. Whole body protein synthesis and breakdown ........................................................12
1.4.3 Amino acid oxidation ................................................................................................14
1.4.4 The effect of CHO availability on AA oxidation during exercise ............................16
1.4.5 The effect of exercise intensity and duration on AA oxidation and post-exercise protein requirements ..............................................................................................19
1.5 Estimating Dietary Protein Requirements for Endurance Athletes .....................................22
1.5.1 Methods of estimating protein requirements ............................................................23
1.5.2 Current protein intake recommendations for endurance athletes .............................25
1.5.3 Habitual protein intake of endurance athletes ...........................................................26
1.6 Research Gaps and Conclusion ...........................................................................................27
Chapter 2. Study Proposal ..............................................................................................................29
2.1 Introduction .........................................................................................................................29
2.2 Hypothesis ...........................................................................................................................30

v
2.3 Research Phases ..................................................................................................................30
Chapter 3. Methodology ................................................................................................................33
3.1 Introduction .........................................................................................................................33
3.2 Participants and Study Design ............................................................................................33
3.3 Informed Consent and Familiarization ...............................................................................34
3.4 Fitness Assessment and Anthropometric Measurement Collection ....................................35
3.5 Experimental Trials .............................................................................................................36
3.6 Data Collection and Analysis ..............................................................................................38
3.6.1 Statistical Analysis ....................................................................................................41
Chapter 4. Results ..........................................................................................................................42
4.1 Descriptive Characteristics of Exercise ..............................................................................42
4.2 Phenylalanine Oxidation and Tracer Kinetics .....................................................................45
Chapter 5. Discussion ....................................................................................................................48
5.1 Overview of the Findings ....................................................................................................48
5.2 Exercise Outcomes ..............................................................................................................49
5.3 Phenylalanine Oxidation and Tracer Kinetics .....................................................................53
5.4 Estimating Protein Requirements ........................................................................................62
5.5 Methodological Limitations ................................................................................................65
5.6 Research Implications and Practical Recommendations .....................................................67
5.7 Summary and Conclusion ...................................................................................................70
References ......................................................................................................................................72
Appendices .....................................................................................................................................94
A. Consent Form .......................................................................................................................95
B. CSEP ‘Get Active’ Questionnaire ......................................................................................104
C. Training Log and Screening/Information Questionnaire ...................................................107

vi
List of Abbreviation AA - Amino Acid ADP - Adenosine Diphosphate AMPK - 5’ AMP-activated Protein Kinase ATGL - Adipose Tissue Triacylglycerol Lipase ATP - Adenosine Triphosphate APE - Atom Percent Excess BCAAs - Branched-Chain Amino Acids BCOAD - Branched-Chain 2 oxo-acid Dehydrogenase Enzyme CHO - Carbohydrate EAA - Essential Amino Acid EAR - Estimated Average Requirement FFA - Free Fatty Acid FM - Fat Mass FFM - Fat-free Mass FSR - Fractional Synthetic Rate F13CO2 - Breath Enrichment of 13CO2 GLUT-4 - Glucose Transporter Type 4 HR - Heart Rate HSL - Hormone Sensitive Lipase IAA - Indicator Amino Acid IAAO - Indicator Amino Acid Oxidation IMTG - Intramuscular Triglycerides MPS - Muscle Protein Synthesis MPB - Muscle Protein Breakdown MyoPS - Myofibrillar Protein Synthesis MitoPS - Mitochondrial Protein Synthesis N - Nitrogen NB - Net Protein Balance NBAL - Nitrogen Balance NOLD - Non-Oxidative Leucine Disposal PB - Protein Breakdown PS - Protein Synthesis PheOx - Phenylalanine Oxidation PheRa - Phenylalanine Rate of Appearance (flux) Ra - Rate of Appearance REE - Resting Energy Expenditure RDA - Recommended Dietary Allowance RER - Respiratory Exchange Ratio RQ - Respiratory Quotient V̇O2 - Oxygen Consumption V̇O2max - Maximal Oxygen Consumption 3-MH - 3-Methylhistidine

vii
List of Tables
Table 1: Overall Study Protocol Table 2: Participant Characteristics Table 3: Exercise outcomes at 2-km, 8-km, 18-km and overall during the 20-km run Table 4: Substrate utilization during 20-km running performance in LOW and HIGH
List of Figures
Figure 1: Schematic Illustration of Whole-body Protein Metabolism Figure 2: Schematic Illustration of Exercise and Metabolic Trial Sessions Figure 3: RER at 3 time points (2-km, 8-km, 18-km) during 20-km run in HIGH and LOW Figure 4: Percent CHO and fat oxidation during exercise over time (2-km, 8-km, 18-km) Figure 5: F13CO2 following 20-km run in HIGH vs. LOW Figure 6: PheOx, Protein Synthesis, Protein Breakdown following 20-km run in HIGH vs. LOW Figure 7: Net protein balance following 20-km run in HIGH vs. LOW Figure 8: Assumptions of the IAAO model
List of Appendices
Appendix A: Consent Form Appendix B: CSEP ‘Get Active’ Questionnaire Appendix C: Training Log and Screening/Information Questionnaire

1
Chapter 1. Review of the Literature
1.1 Introduction
Endurance exercise training (e.g. running, cycling, triathlon) is a popular activity that is
carried out at a range of intensities from low-moderate to intense for the recreational and elite,
high-performance athletes, respectively. At the elite level, endurance athletes are driven to enhance
exercise adaptations, improve their performance and optimize recovery between training bouts.
Given that endurance training results in elevated metabolic demands (e.g. increased energy
expenditure and endogenous fuel depletion) (Loucks, 2007; Burke, Kiens, & Ivy, 2004), one area
that has received considerable focus in terms of its capacity to support and improve athletic
performance is sports-specific nutrition tailored to the unique needs of endurance athletes (Hawley
and Hopkins 1995; Jeukendrup, 2011).
During endurance exercise, glycogen and free fatty acids (FFA) constitute the main sources
of energy (Lemon & Mullin, 1980; Van Loon, Greenhaff, Constantin-Teodosiu, Saris, &
Wagenmakers, 2001). Furthermore, substrate utilization for fuel during prolonged exercise is a
function of the relative exercise intensity and duration (Van Loon et al., 2001). In general, lipids
constitute a major fuel source during rest and low intensity exercise and are utilized maximally at
~55% V̇O2max (Van Loon et al., 2001). At moderate to high exercise intensities (i.e. >55%
V̇O2max), there is a progressive decrease in absolute lipid utilization and a reciprocal increase in
the contribution of carbohydrates (CHO) to fuel the metabolic demands (Brooks, 1998; Van Loon
et al., 2001; Romijn et al., 1993). In trained individuals, higher intensity (~70-75% V̇O2max) but
not lower intensity (30% V̇O2max) exercise causes glycogen availability to become limited after
~90 min of continuous exercise (Hermansen et al., 1967; Gollnick et al., 1981). Limited glycogen
availability results in greater dependence on alternate fuel sources including amino acids (AA) (i.e.

2
amino acid oxidation) to support the energy demands of exercise (Lemon & Mullin, 1980;
Millward, Bowtell, Pacy, & Rennie, 1994; Tarnopolsky, 2004; Wagenmakers et al., 1991).
Ultimately, greater AA utilization (oxidation) during exercise limits the availability of AA in the
post-exercise period to support muscle and whole-body protein synthesis (Kato, Suzuki, Bannai,
& Moore, 2016; Mazzulla et al., 2017). Thus, post-exercise protein requirements to support
recovery and exercise adaptations may differ depending on the exercise intensity and associated
substrate utilization.
Understanding the relationship between exercise intensity and duration as well as the
relative contributions of different substrates (e.g. glycogen, FFA, AA) to support the energy
demands of exercise is an important consideration for trained endurance athletes. The weekly
training of endurance athletes consists of a range of exercise intensities (from higher intensity
sessions (i.e. ~70-75% V̇O2max) to lower intensity ‘recovery’ sessions (i.e. ~50-55% V̇O2max)
(Stellingwerff, 2012). Typically, an athlete’s periodization schedules are structured around major
competition events with the exercise intensity and total training duration (or volume) modified
based on the temporal proximity to major competitions. Specifically, higher intensity training of
lesser total weekly volume will be performed close to competition dates to try to match the training
demands to the demands of competition (Heikura et al., 2018; Shepley et al., 1992; Fiskerstrand
and Seiler 2004; Stellingwerff, 2012; Stöggl and Sperlich 2015). With this in mind, the substrate
utilization at different times in an athlete’s program (pre-competition vs base training season) or
even between daily workouts could be vastly different. Substrate utilization related to endogenous
CHO depletion and AA oxidation may have implications on post-exercise dietary protein
requirements to support exercise recovery and adaptations. This is of particular importance for the
endurance athlete population as suboptimal protein intakes have been suggested to compromise

3
endurance exercise training quality and performance outcomes (Williamson et al., 2019). Thus,
pre- and post-exercise nutrition strategies could be tailored to fit athletes’ needs depending on the
exercise demands (i.e. exercise intensity and duration) and associated substrate utilization.
This review of the literature will discuss the relevant background information and current
research related to dietary protein requirements for endurance athletes, specifically focusing on the
effects of exercise intensity and duration on primary substrate utilization and exercise-induced AA
oxidation. The review first provides a general introduction to protein metabolism (Section 1.2) and
primary fuel sources for endurance exercise (Section 1.3). This is followed by review of the effects
of endurance exercise on whole-body protein metabolism (Section 1.4). Finally, dietary protein
requirements for endurance athletes (Section 1.5), as well as factors such as exercise intensity and
duration that modulate dietary protein requirements (Section 1.6) are discussed.
1.2 Introduction to Protein Metabolism
Proteins and their constituent AA are the basic building blocks of the human body and
serve a range of essential functions. These functions include; i) structural support, ii) support for
muscular contraction, iii) regulation of hormones and enzymes, and iv) regulation of energy
demands and metabolism (Ott, Amunts, & Brown, 2016; Tarnopolsky, 2004; Tiidus, Tupling, &
Houston, 2012). The basic structure of AA consists of a central carbon skeleton with four bound
groups; i) an amino group (-NH2), ii) a carboxyl group (-COOH), iii) a hydrogen atom and iv) a
variable side chain (R group) (Tiidus et al. 2012). Proteins are comprised of twenty unique AA
that differ in their respective R group side chain. Of the twenty AA that make up proteins, nine are
considered ‘essential AA’ (EAA) or indispensable meaning that they are not produced
endogenously in sufficient quantities to satisfy their metabolic demand and thus, must be obtained
exogenously from dietary sources (Tarnopolsky, 2004; Tiidus et al., 2012). These AA join

4
together through a series of peptide bonds which form polypeptide chains that make up all proteins
in the body (Tiidus et al., 2012; Ott et al., 2016).
Body proteins (including muscle) are regulated by constant ‘turnover’, which is
characterized by the breakdown of old/damaged proteins into their constituent AA and the
reutilization of these AA in combination with exogenous AA to synthesize new proteins (e.g.
mitochondrial proteins and myofibrillar proteins) in their place (Burd, Tang, Moore, & Phillips,
2009; Tiidus et al., 2012; Wagenmakers, 1999). Net protein balance (NB) is a function of the
algebraic difference between overall rate of whole body protein synthesis (PS) and protein
breakdown (PB) (see Figure 1) (Burd et al., 2009; Phillips, Hartman, & Wilkinson, 2005).
Importantly, AA cannot be stored in the human body; when they are provided in excess of their
ability to be incorporated into new body proteins, AA are deaminated and either directly oxidized
or utilized as gluconeogenic precursors (Wolfe, 2006; Tiidus et al., 2012).
The dynamic process of protein turnover is responsive to dietary AA ingestion and
exercise (e.g. heavy resistance and endurance). Dietary AA are needed in sufficient quantities and
ratios of EAA to support protein accretion. For example, a 20g intake of high quality, EAA-
enriched protein (~8-10g of EAA) promotes MPS following resistance exercise (Moore et al.,
2009). Similarly, Churchward-Venne et al. (2020) showed that a 30g intake of high quality protein
was needed to maximize MPS after endurance exercise. Further, it is believed that the EAA are of
particular importance as they are needed to stimulate MPS (Tipton et al., 1999; Børsheim et al.,
2004). For this reason, protein sources containing all EAA in sufficient amounts (e.g. whey and
egg protein) are superior to EAA-deficient sources (e.g. plant-based sources such as rice and soy
protein) in their capacity to promote protein accretion (Wilkinson et al., 2007). Thus, adequate
dietary protein of a high quality, containing all EAA in the correct ratios, is needed to promote

5
optimal protein accretion, supporting lean body mass preservation and growth (Burd et al., 2009;
Wilkinson et al., 2007).
Figure 1: Schematic illustration of whole-body protein metabolism Adapted from (Wagenmakers, 1999). Abbreviations: amino acid (AA), protein synthesis (PS), protein breakdown (PB), excretion (Ex), oxidation (Ox), carbon dioxide (CO2), ammonia (NH3). 1.3 Fuel use during Endurance Exercise
Endurance exercise is associated with elevated energy requirements to fuel the demands of
exercise. During prolonged periods of aerobic-based exercise, energy in the form of adenosine
triphosphate (ATP) is supplied primarily by oxidative metabolism of CHO and lipids (oxidative
phosphorylation) (Blomstrand & Saltin, 1999; Burke, Van Loon, & Hawley, 2017; Cermak & Van
Loon, 2013; Gollnick et al., 1981; Hargreaves, 2000; Hawley & Hopkins, 1995; Holloszy & Coyle,
2016; Jeukendrup et al., 1999; Klein, Coyle, & Wolfe, 1994; Murray & Rosenbloom, 2018;
Tarnopolsky, 2004). At higher-intensity exercise (~70-75% V̇O2max), CHO constitutes the main
source of fuel and the availability of CHO is a major contributor to sustained high-intensity
endurance exercise capacity (Burke et al., 2017). In contrast, during lower exercise intensity

6
(<55% V̇O2max) there is relatively less CHO oxidation and depletion of muscle glycogen and
greater lipid utilization (Van Loon et al., 2001).
Carbohydrates as an Energy Substrate
During exercise, elevations in whole-body glucose turnover occur through enhanced
endogenous glucose production and subsequent skeletal muscle glucose uptake (Jeukendrup et al.,
1999). Muscle glucose uptake is stimulated by exercise contraction, which acts to increase glucose
transporter type 4 (GLUT-4) translocation to the sarcolemma to increase glucose uptake into the
cell (Jeukendrup et al., 1999; Murray & Rosenbloom, 2018). Endogenous glucose production,
which occurs in the liver primarily, is regulated tightly by central nervous system mediated
feedforward mechanisms (glucosensors in the hepatoportal region transmit signals to the
hypothalamus) and feedback mechanisms in response to decreased blood glucose concentration
and energy stress (e.g. increased 5’ AMP-activated protein kinase (AMPK) activity) (Jeukendrup
et al., 1999; Taylor and Shulman, 2011). In addition, during exercise, catecholamines (epinephrine
and norepinephrine) regulate CHO metabolism via beta adrenoceptors which stimulate
glycogenolysis and gluconeogenesis and inhibiting the effect of insulin (Barth et al., 2007).
Endogenous glucose production occurs via; i) glycogenolysis, the breakdown of stored glycogen
particles into individual glucose molecules, and; ii) gluconeogenesis the production of glucose
from non-glucose substrates including AA (Jeukendrup et al., 1999; Murray & Rosenbloom,
2018). Endogenous CHO is present in the form of stored muscle glycogen (400-600g) and liver
glycogen (80-160g). Muscle glycogen is distributed within the muscle cell in three intracellular
‘compartments’; i) intramyofibrillar, ii) intermyofibrillar, and iii) subsarcolemmal glycogen stores
(Murray & Rosenbloom, 2018). During prolonged, continuous endurance events lasting >90-120
minutes, endogenous CHO stores are depleted in an intensity-dependent manner (Hargreaves,

7
2000; Romijn et al., 1993). For example, during an all-out exhaustive exercise bout, glycogen is
degraded and releases glucose molecules at a rate of ~40mmol glucose ∙ kg wet mass-1• min-1
whereas during lower-intensity (~50% V̇O2max) exercise, glycogen breakdown occurs at ~1-
2mmol glucose ∙ kg wet mass-1 •min-1 (Hargreaves et al., 2012). This endogenous CHO depletion
ultimately increases the reliance on non-CHO gluconeogenic substrates (e.g. AA) as a fuel source
(Hermansen et al., 1967; Coyle et al., 1986; Coggan & Coyle, 1987; Romijn et al., 1993;
Jeukendrup et al., 1999; Hargreaves, 2000; Murray & Rosenbloom, 2018).
Given the pivotal role of endogenous CHO availability (e.g. muscle glycogen and blood
glucose) as energy for sustained higher intensity exercise (Burke, 2010; Burke, Van Loon, &
Hawley, 2017; Gollnick et al., 1981; Murray & Rosenbloom, 2018), endurance athletes frequently
attempt to manipulate CHO availability. This is done through different nutrition strategies
including pre-exercise CHO feeding and glycogen loading and depletion techniques. The ingestion
of exogenous CHO before prolonged exercise is a common strategy that reduces the reliance on
endogenous CHO sources (e.g. muscle glycogen) and improves the capacity for sustained high-
intensity exercise (Coyle et al., 1986; Coggan and Coyle 1987; Heikura et al., 2018). Pre-exercise
CHO feeding also maintains the rate of CHO oxidation and blood glucose concentrations into the
later stages of exercise (Jeukendrup et al., 1999; Murray & Rosenbloom, 2018; Riddell et al.,
2003). Manipulating endogenous CHO storage through CHO supercompensation (“CHO
loading”), and low-CHO-availability training (e.g. fasted exercise and/or exercise with low
glycogen content) is also commonly practiced (Burke, 2010; Hargreaves, 2015; Heikura et al.,
2018; Jeukendrup, 2011). CHO loading is associated with superior muscle glycogen storage and
enhanced exercise performance (Coggan & Coyle 1987; Hawley et al., 1997). Low CHO
(glycogen depleted) training has been shown to increase AMPK activity, GLUT-4 activation and

8
fat oxidation during exercise (Hawley et al., 2006; Hawley & Morton, 2014). This enhancement
in intracellular signaling and gene expression regulating metabolic function may enhance exercise
recovery and aerobic adaptations (e.g. increased mitochondrial biogenesis, muscle oxidative
capacity, resting muscle glycogen concentration and citrate synthase activity ) (D'Antona et al.,
2010; Di Donato et al., 2014; Ferguson-Stegall et al., 2011; Hansen et al., 2005; Hawley et al.,
2006; Hulston et al., 2010; Psilander, Frank, Flockhart, & Sahlin, 2013; Rowlands et al., 2011)
and, as such, has become a popular nutrition strategy for training (Burke et al., 2010 & 2017;
Hansen et al., 2005; Hawley et al., 2014; Hearris et al., 2018; Lane et al., 2015; Murray et al.,
2018; Pilegaard et al., 2002). Despite the adaptive changes associated with training with low-CHO
availability, there is some question as to whether these changes translate into superior aerobic
exercise performance (Burke et al., 2010).
Lipids as an Energy Substrate
Lipids in the form of FFA and IMTG constitute a greater fuel source for low-moderate
intensity (~50-55% V̇O2max) compared to higher intensity exercise (~70-75% V̇O2max)
(Horowitz et al., 1997; Kiens et al., 1999; Romijn et al., 1993; Van Loon et al., 2001). Watt &
Spriet, (2004) suggest that lower fat oxidation during high-intensity exercise may be a result of
decreased hormone-sensitive lipase (HSL) activity, a key enzyme involved in lipolysis. Activation
of energy-sensitive AMPK, and phosphorylation of inhibitory sites on HSL, may blunt lipolysis
and consequently limit the rate of fat oxidation during exercise at high intensities (Kiens et al.,
1999; Watt & Spriet, 2004). Moreover, elevations in the flow of metabolites through the glycolytic
pathway (i.e. glycolytic flux) (Teslaa & Teitell, 2014) during higher-intensity exercise, is
associated with an accumulation of acetyl coenzyme A (acetyl-CoA) and is suggested to limit long-
chain fatty acid oxidation during exercise (Dyck et al., 1993; Sidossis et al., 1996).

9
The rate of fat oxidation during endurance exercise is not only dependent on the intensity
and duration of exercise but also on the CHO availability and concentration of circulating FFA
(Gollnick et al., 1981; Horowitz et al., 1997; Van Loon et al., 2001). High CHO availability
through adequate muscle glycogen storage and pre-exercise CHO ingestion have been shown to
augment the rate of CHO oxidation during exercise and exert a potent inhibitory effect on lipolysis
during low-moderate and high-intensity exercise (Coyle et al., 1997; Horowitz et al., 1997;
Sidossis et al., 1996; Spriet, 2014). Horowitz et al., (1997) showed that modest pre-exercise CHO
feeding (fructose: 9uU/ml, glucose: 30uU/ml) was associated with a small rise in insulin
concentration and an insulin-induced suppression in lipolysis, the latter finding being consistent
with several other studies (Campbell et al., 1992; Romijn et al., 1993; Sidossis et al., 1996). The
effect of insulin on lipolysis is suggested to occur through blunting enzymes, adipose tissue
triacylglycerol lipase (ATGL) and HSL, (both key enzymes in IMTG hydrolysis) (Sidossis et al.,
1996). As the rate of fat oxidation can only exceed the rate of lipolysis for a brief period of time,
blunted lipolysis ultimately limits the hydrolysis of triglycerides and the availability of fatty acid
entry into the mitochondria for oxidation (Horowitz et al., 1997; Sidossis et al., 1996). Importantly,
chronic endurance training is associated with a refinement of the metabolic processes favouring a
greater capacity for muscle glycogen storage (Hargreaves, 2015) and greater relative fat utilization
for fuel during exercise (Murray & Rosenbloom, 2018; Tarnopolsky, MacDougall, Atkinson, &
Sutton, 1990). Thus, patterns of substrate utilization during exercise are not only affected by the
intensity and duration of exercise but also by endogenous substrate availability, pre- and during
exercise nutrition strategies (e.g. pre-exercise CHO) as well as individual training status.
1.4 Protein Metabolism and Endurance Exercise 1.4.1. Muscle protein synthesis and breakdown

10
Compared to resistance exercise, the effects of endurance exercise on muscle (and whole-
body) protein metabolism are less well characterized. Following acute endurance exercise, rates
of mixed-muscle protein synthesis are augmented in the fasted (Carraro et al., 1990; Sheffield-
Moore et al., 2004; Harber et al., 2010; Mascher et al., 2011) and fed (Harber et al., 2010) states.
Further, endurance exercise has been shown to augment myofibrillar protein synthesis (MyoPS)
in the fasted state (Di Donato et al. 2014) and mitochondrial protein synthesis (MitoPS) in both
the fasted (Di Donato et al., 2014) and fed (Wilkinson et al., 2008) state. Mascher et al., (2011)
showed that mixed-muscle protein FSR increased gradually in the 3 h recovery period in the fasted
state following a high-intensity single leg cycling protocol (65-70% V̇O2 max). Similarly, Harber
et al., (2010) showed that in the 6 h recovery period following 60 min of high-intensity (72%
V̇O2max) biking exercise, mixed-muscle FSR was elevated above resting conditions in the
postabsorptive and postprandial state. Further, CHO + protein feeding (0.83 g CHO/kg body mass
and 0.37 g protein/kg body mass) immediately after and 1 h after exercise did not further augment
rates of mixed-muscle FSR compared to a postabsorptive condition. Sheffield-Moore et al., (2004)
examined the effect of low-moderate intensity (40% V̇O2max) walking exercise for 45 min on
mixed-muscle FSR in the recovery period. They showed an elevation above rest in mixed-muscle
FSR at 10 and 60 min after exercise; however, this elevation was more transient than what is seen
in some other studies. Interestingly, at a higher exercise intensity (60 min of running at 70%
V̇O2max), Abou Sawan et al., (2018) showed no difference in rates of fed state (18g protein and
60g CHO) MyoPS compared to a resting condition. They speculated that this may be a result of
the trained population being examined who would exhibit a more refined muscle protein synthetic
response to exercise (Abou Sawan et al., 2018).

11
Discrepancies in these results may also be due in part to differences in the exercise
modality. For example, studies using a cycling exercise protocol have shown elevations in muscle
protein FSR in the post-exercise period (Harber et al., 2010; Mascher et al., 2011; Di Donato et
al., 2014); however, cycling protocols may exert a more potent and concentrated stimulus on the
vastus lateralis muscle (Bijker et al., 2002), the site at which muscle biopsies are commonly
obtained to quantify FSR through tracer incorporation (Wagenmakers, 1999). In contrast, running
exerts a more whole-body stimulus (Loftin et al., 2004; Millet et al., 2009). Further, the relative
intensity and total duration of exercise has been shown to influence the magnitude of effect on
rates of MPS. For example, Di Donato et al., (2014) showed that both high (30 min at 60% Wmax)
and low intensity (60 min at 30% Wmax cycling exercise in the fasted state elevated MyoPS in
the early (0.5-4.5 hours) post-exercise recovery period. However, MyoPS remained elevated into
the later stages of recovery (24-28 h) only in the high intensity exercise condition. Moreover,
MitoPS was significantly greater at both early and late stage recovery in the high intensity
compared to low intensity condition. Thus, endurance exercise augments MPS and has been shown
to be influenced by factors including the exercise modality as well as relative intensity and duration
of exercise.
The ingestion of post-exercise protein is important for endurance athlete populations as it
replenishes AA oxidative loss, augments the rate of mixed-muscle (Howarth et al., 2009) and
MyoPS (Abou Sawan et al., 2018; Breen et al., 2011) after exercise, which supports post-exercise
recovery (Harber et al., 2010). Interestingly, increases in myofibrillar but not mitochondrial FSR
following post-exercise with protein feeding have also been reported (Breen et al., 2011; Abou
Sawan et al., 2018), suggesting a diverging response to post-exercise nutrition between protein
subfractions with mitochondrial proteins being less responsive to nutrition following endurance

12
exercise. Churchward-Venne et al., (2020) showed that protein ingestion (30g) increased rates of
MyoPS but not MitoPS. However, dietary protein derived AA incorporation into MitoPS was
responsive to increasing protein intakes in a dose-dependent manner as AA feeding supplies the
necessary AA precursors for the synthesis of new proteins (Churchward-Venne et al., 2020).
Further, the dose-dependent nature of the response to post- exercise protein feeding indicates that
the diverging responses across studies may be due in part to variability in protein feeding protocols
(e.g. timing, dose, whole food vs. intravenous AA infusion) (Abou Sawan et al., 2018).
Endurance exercise stimulates the degradation of muscle proteins into the free AA pool
through protein breakdown in the fasted state (Carraro, Stuart, Hartl, Rosenblatt, & Wolfe, 1990;
Harber et al., 2010; Rennie et al., 1981). For example, Carraro et al., (1990) assessed MPB through
3-methylhistidine excretion (3-MH) and observed that following 4 h of low-intensity treadmill
exercise, there was a ~85% elevation in 3-MH excretion. Harber et al., (2010) estimated MPB
indirectly via skeletal muscle mRNA expression of proteolytic factors involved in the ubiquitin-
proteasome and Ca2+-dependent protein degradation pathways. They showed that following
cycling exercise in the post-absorptive condition, expression of proteolytic factors were elevated
above rest. Importantly, post-exercise feeding of protein and CHO attenuated the expression of
proteolytic factors (MuRF-1 and calpain-2) (Harber et al., 2010) which is consistent with the
previously reported insulin-induced suppression in muscle protein degradation (Gelfand & Barrett,
1987). Thus, post-exercise protein and CHO ingestion supports muscle protein accretion and a
more positive NB (Levenhagen et al., 2002).
1.4.2. Whole body protein synthesis and breakdown
At the whole-body level, endurance exercise induces a negative protein balance during and
after exercise in the postabsorptive condition (Levenhagen et al., 2002). This occurs through a

13
greater diversion of AA in the free AA pool toward energy production by exercise-induced AA
oxidative loss (2-3 fold greater) and lower rates of whole-body protein synthesis during exercise
(Bowtell et al., 2000; Mazzulla et al., 2017; Phillips, Atkinson, Tarnopolsky, & MacDougall,
1993). Phillips et al., (1993) showed that rates of leucine flux, which were presumed to reflect
whole-body protein breakdown, were elevated during 90 min of running exercise at 65% V̇O2max
compared to a resting condition. However, this was not seen by Mazzulla et al., (2017) who showed
that whole-body leucine balance and leucine Ra (protein breakdown) in the postprandial period (5
h) following protein ingestion (18g) were not different between a resting condition and following
endurance exercise (running for 60 min at 70% V̇O2max). However, when exercise-induced
leucine oxidation was accounted for, leucine balance became negative (Mazzulla et al., 2017).
Moreover, during moderate-intensity (60% V̇O2max) prolonged (2 h) endurance exercise, it has
been estimated that leucine is oxidized at a rate of ~8 mg·kg-1·h-1 and overall, ~13g of protein is
catabolized (Moore et al., 2014). Thus, endurance exercise shifts AA metabolism toward energy
production (discussed further in Section 1.4.2) and away from energy consuming processes such
as PS. During the postabsorptive condition or without the provision of adequate amounts of post-
exercise dietary protein, endurance exercise results in a catabolic body environment and negative
whole-body NB (Mazzulla et al., 2017; Rennie et al., 1981). Adequate post-exercise protein
feeding serves to replenish exercise-induced AA oxidative losses and support a positive
postprandial NB.
Bowtell et al. (2000) showed that rates of whole-body protein breakdown were higher
significantly during exercise compared to the post-exercise period following protein alone or CHO
+ protein (0.75 g CHO ·kg-1·h-1 and 1.8 g protein ·kg-1·h-1 ) ingestion. Indeed, protein or CHO +
protein feeding during and after exercise augments whole body protein synthesis (Levenhagen et

14
al., 2002), and attenuates the rise in whole-body protein breakdown (Howarth et al., 2009;
Koopman et al., 2004; Mazzulla et al., 2017) which supports a positive (or less negative) whole-
body NB (Howarth et al., 2009; Koopman et al., 2004; Levenhagen et al., 2002). Thus, whole-
body endurance exercise induces a negative protein balance during and after exercise which can
be offset by post-exercise dietary protein ingestion.
Horwarth et al. (2009) proposed that methodological differences in post-exercise protein
feeding may explain the diverging post-exercise response. Specifically, Levenhagen et al. (2002)
showed providing a single bolus of post-exercise CHO (8g) + protein (10g) augmented whole-
body PS during recovery while Koopman et al. (2004) showed no differences in rates of synthesis
but an attenuated whole-body PB after exercise when exercisers received small doses of protein at
regular intervals (0.25 g protein·kg-1·h-1). Howarth at el., (2009) showed a diverging response to
a continuous hourly CHO (1.2 g CHO·kg-1·h-1) + protein (0.4 g protein·kg-1·h-1) feeding protocol
after exercise. Specifically, feeding induced an increase in post-exercise muscle FSR; however,
this was not seen at the whole-body level in NOLD, which is representative of whole-body PS. As
mixed-muscle FSR accounts for ~25% of whole-body PS (Nair et al., 1988), there may be a
diverging response of different body protein pools to post-exercise CHO + protein feeding.
Moreover, methodological differences in the post-exercise feeding (e.g. dose and timing) may
explain some of these discrepancies.
1.4.3 Amino acid oxidation
Carbohydrates (i.e. glycogen and glucose) and fat (i.e. FFA) typically constitute the major
sources of fuel for oxidative phosphorylation during prolonged endurance exercise with AA
supplying ~2-5% of total fuel (Blomstrand & Saltin, 1999; Lemon & Mullin, 1980; Van Loon et
al., 2001). The use of AA as a respiratory substrate involves the removal of the AA amino group

15
(-NH2) and use of the carbon skeleton to supply carbon atoms as an alternate source of fuel (Lemon
& Mullin, 1980; Tiidus et al., 2012). The carbon skeleton of AA supports energy during exercise
through; i) direct oxidation through the production of tricarboxylic acid cycle
precursors/intermediates via the muscle mitochondria (Jungas, Halperin, & Brosnan, 1992;
Tarnopolsky, 2004), and; ii) glucose formation from non-carbohydrate precursor (e.g. glucogenic
AA, lactate, glycerol) via gluconeogenesis in the liver (Kato et al., 2018; Shimomura et al., 2004;
Tarnopolsky, 2004; Tiidus et al., 2012).The amine group (-NH2) removed through deamination is
either i) converted to ammonia and subsequently converted to urea in the liver which is eventually
excreted in the urine (Tiidus et al., 2012) or ii) transferred to pyruvate to form alanine which is
subsequently converted back to glucose in the liver through the Cori cycle (Berg et al., 2002).
When CHO availability becomes limited, AA are increasingly diverted away from events
supporting protein synthesis and towards events supporting energy production (i.e. AA oxidation)
(Mazzulla et al., 2017). For example, exercise-induced AA oxidation can supply ~5-10% of total
energy during CHO depleted conditions and this is a major contributor to the catabolic nature of
endurance exercise (Lemon & Mullin, 1980; Tarnopolsky, 2004). The branched-chain amino acids
(BCAA) (i.e. leucine, isoleucine and valine) play a regulatory role in protein metabolism (Moore
et al., 2014) and are oxidized preferentially during endurance exercise within skeletal muscle
(Bowtell et al., 2000; Jungas et al., 1992; Kato et al., 2018a; Koopman et al., 2004; Lamont,
McCullough, & Kalhan, 1999; Millward et al., 1994; Shimomura, Murakami, Nakai, Nagasaki, &
Harris, 2004). The extent of exercise-induced AA oxidation, and primarily of the BCAA, is largely
dependent on: i) the energy demands of exercise (i.e. exercise intensity and duration), and; ii) CHO
availability (i.e. muscle glycogen storage) to support energy needs (discussed further in Section
1.4.3 ) (Lemon & Mullin, 1980; Millward et al., 1994; Wagenmakers et al., 1991). Indeed, the

16
activation of the mitochondrial rate-limiting enzyme in BCAA oxidation [branched-chain 2 oxo-
acid dehydrogenase enzyme (BCOAD] occurs with decreases in both the ATP/ADP ratio and
muscle glycogen content (Kasperek & Snider, 1987; McKenzie et al., 2000; Wagenmakers et al.,
1991; Wagenmakers, Brookes, Coakley, Reilly, & Edwards, 1989).
The exercise-induced oxidation of leucine has received considerable attention due to the
fact that it is oxidized preferentially during endurance exercise and supports anabolism through its
unique ability to stimulate PS independently by activation of the mTOR signalling pathway
(Anthony, Kimball, & Jefferson, 2001; Kimball & Jefferson, 2006). Whole-body leucine oxidation
and BCOAD activity has been shown to increase proportionally to relative exercise intensity
(Lamont et al., 1982 & 2001). During higher intensity exercise (60-70% V̇O2max), leucine
oxidation has been shown to increase by ~2-3 fold above resting rates during (Bowtell et al., 1998;
Lamont, McCullough, & Kalhan, 2001; Mazzulla et al., 2017; McKenzie et al., 2000). These
exercise-induced increases in BCAA particularly, leucine oxidation result in the need for AA
losses to be replaced ostensibly through post-exercise dietary protein ingestion to facilitate
recovery.
1.4.4 The effect of CHO availability on AA oxidation during exercise
Exercise-induced AA oxidation increases in response to situations in which endogenous
CHO sources including plasma glucose and muscle glycogen become depleted (e.g. during
extended periods of high intensity CHO depleting exercise) (Kasperek & Snider, 1987;
Wagenmakers et al., 1991). During prolonged exercise conditions when CHO availability is low,
and liver glycogenolysis cannot provide sufficient glucose to support energy demands, the
deamination and irreversible oxidation of AA particularly BCAA increases (Van Hall et al., 1996;
Riddell et al., 2003; Howarth et al., 2010).

17
Bowtell et al. (2000) examined the effect of glucose supplementation during prolonged
exercise (2 h running at 60% V̇O2max) on leucine oxidation in individuals consuming a high (1.8
g⸱kg-1⸱d-1) versus low (0.7 g⸱kg-1⸱d-1) protein diet. In support of the previously discussed role of
CHO availability on AA oxidation, glucose supplementation before and during endurance exercise
suppressed both leucine oxidation (by 20%) and BCOAD activity during exercise compared to
exercise without glucose feeding (Bowtell et al., 2000). In line with this, Howarth et al. (2010)
examined the effect of glycogen availability (high vs low) on muscle and whole-body protein
turnover during exercise and post-exercise recovery. Upon beginning exercise there was an ~150
mmol/g dry mass difference in glycogen content between the experimental conditions. They
showed that muscle phenylalanine NB was negative in the high and low CHO availability
conditions; however, it was significantly lower throughout the duration of exercise in the low-
CHO condition. Further, whole-body leucine oxidation was elevated and leucine NB was more
negative during exercise in the low-CHO but not high-CHO condition when compared to resting
values (Howarth et al., 2010). Similarly, Lemon & Mullin, (1980) examined the effect of muscle
glycogen content (CHO loaded vs depleted) on protein catabolism during cycling exercise (1 h at
65% V̇O2max). They showed that exercise serum urea nitrogen, which reflects AA oxidation, was
elevated significantly in the CHO-depleted condition during and post-exercise. In contrast, serum
urea N in the CHO-loaded condition was not altered during exercise and was increased minimally
post-exercise. Thus, CHO availability from pre-exercise feeding and endogenous muscle glycogen
stores modulate the extent of AA oxidation during endurance exercise.
Elevations in AA mobilization and oxidation during exercise in a glycogen-depleted condition
have been suggested to influence post-exercise protein requirements. Gillen et al. (2019) used the
indicator amino acid oxidation method (IAAO) to examine the effect of high and low CHO

18
availability during endurance exercise on whole-body protein kinetics and dietary protein
requirements. This study implemented a CHO-depleting exercise protocol (10 x 5 min run intervals
at 10-km race pace) the evening before the experimental trial with (high-CHO) or without (low-
CHO) the provision post-exercise CHO to aid in the resynthesis of muscle glycogen. A similar
protocol has been used previously by (Morton et al., 2009) which was reported to enhance aerobic
signalling and lower glycogen content of the vastus lateralis (by 35%) and gastrocnemius (by 45%)
muscles. A single suboptimal protein intake of 0.93 g⸱kg-1⸱d-1 was given and oxidation of an
indicator AA (phenylalanine) which reflects dietary protein requirement was examined. Gillen et
al. (2019) showed that the 10-km training bout in the low CHO availability state resulted in
elevated oxidation of the indicator AA compared to a non-glycogen-depleted, high CHO
availability condition in the post-exercise period. Further, there was an ~15% greater estimated
daily protein requirement in the low-CHO condition, which would be consistent with an exercise-
induced increase in AA oxidation during conditions in which CHO availability is low.
Similarly, Howarth et al. (2010) examined the effect of glycogen availability (low vs high) on
skeletal muscle protein turnover during exercise (2 h of two-legged kicking exercise at 45%
V̇O2peak) and post-exercise recovery. Resting muscle glycogen content was estimated to be ~300
mmol∙ kg dry mass-1 in the low-CHO condition and ~420 mmol∙ kg dry mass-1 in the high-CHO
condition. They showed an ~43% greater rate of leucine oxidation and a more negative
phenylalanine net balance in the low-CHO group during exercise. Importantly, phenylalanine Ra
was elevated within the first 10 min of exercise in the low-CHO condition suggesting that muscle
glycogen content does not necessarily need to be depleted severely but rather, simply reduced, to
see immediate metabolic differences during exercise. Thus, beginning exercise with low CHO
availability (e.g. glycogen-depleted) or performing exercise of sufficient duration and intensity to

19
cause reduced CHO stores can enhance exercise-induced AA oxidative loss and protein
degradation (Beelen et al., 2011; Blomstrand & Saltin, 1999; Gillen et al., 2019; Howarth et al.,
2010; Lemon & Mullin, 1980; Riddell et al., 2003; Tarnopolsky, 2004; Van Hall et al., 1996;
Wagenmakers et al., 1991). Consequently, post-exercise protein requirements to maximize whole-
body protein synthesis are elevated when endurance exercising with low CHO availability (Gillen
et al., 2019).
1.4.5 The effect of exercise intensity and duration on AA oxidation and post-exercise protein requirements
There is a linear relationship between exercise intensity (%V̇O2max) and AA oxidation
during exercise (Millward et al., 1994; Rennie et al., 1981; Tarnopolsky, 2004). Specifically, high
intensity exercise is associated with both elevated total energy demands and greater muscle
glycogen depletion both of which have implications for exercise induced AA oxidative loss.
Lamont et al. (2001) assessed the relationship between whole-body leucine oxidation and oxygen
consumption during steady-state exercise and observed that 60 min of moderate intensity (50%
V̇O2max) cycling exercise resulted in an elevation in leucine oxidation and a suppression in NOLD
during exercise. A modest relationship between leucine oxidation and steady-state O2 consumption
was identified such that approximately half the variance of whole-body leucine oxidation can be
explained by the elevated energy demands of exercise (Lamont et al., 2001). Thus, the elevated
total energy expenditure associated with high-intensity endurance exercise is responsible partially
for the elevated AA oxidation during high-intensity exercise.
As previously described (Section 1.4), the relative utilization of fat and CHO during
exercise is dependent on exercise intensity (Cermak & Van Loon, 2013; Hargreaves, 2000; Romijn
et al., 1993; Van Loon et al., 2001). During high exercise intensities (>65%V̇O2max), the
mobilization and utilization of FFA declines and CHO in the form of circulating glucose and

20
endogenous muscle glycogen (total content 400-600g) are the main sources of fuel (Moore et al.,
2014; Romijn et al., 1993; Van Loon et al., 2001). Ultimately, high-intensity glycogen depleting
exercise increases the reliance on alternate fuel sources including AA (AA oxidation) (Cermak &
Van Loon, 2013; Millward et al., 1994; Tarnopolsky, 2004). Thus, exercise intensity modulates
AA oxidation through elevations in total energy expenditure (Lamont et al., 2001) as well as
depletion of endogenous CHO availability (Bowtell et al. 2000) during exercise. In support of the
relationship between exercise intensity and AA oxidation, early work by Eriksson, Broberg,
Björkman, & Wahren, (1985) compared low intensity endurance exercise (55% V̇O2max) and high
intensity endurance exercise (80% V̇O2max) and showed greater BCAA oxidation and alanine
production (an indicator of AA oxidation) in the high-intensity condition. Further, Lemon, Nagle,
Mullin, & Benevenga, (1982) examined whole-body leucine oxidation in rats at rest and during 1
h exercise at 40% and 80% V̇O2max. They showed that exercise increased leucine oxidation above
resting conditions in an intensity-dependent manner, with greater leucine oxidation in the high
(80% V̇O2max) compared to low (40% V̇O2max) condition. Gillen, (2017) showed that indicator
AA oxidation was not different between a high (~74% V̇O2peak) and low (~50% V̇O2peak)
exercise intensity, reflecting no differences in post-exercise protein requirements between
conditions (Gillen, 2017). Interestingly, they showed that the rate of whole-body protein turnover
tended to be elevated in the high-intensity condition (Gillen, 2017). The lack of between condition
differences (high vs low) may however have been a result of the total exercise times which was
<60 min. Exercise duration is also an important factor that affects substrate utilization/availability.
During moderate to high exercise intensity (70-80% V̇O2max) lasting >90 min, substantial
glycogen depletion occurs and content can become critically low (~25mmol/kg wet mass)
(Gonzalez, Fuchs, Betts, & Van Loon, 2016; Hawley, Schabort, Noakes, & Dennis, 1997). Beyond

21
90 min of moderate-intense exercise, muscle glycogenolysis declines as a function of decreased
glycogen content and the overall contribution of CHO oxidation to support energy needs is lower
(Hargreaves, 2000). This is often reflected in a decreased respiratory exchange ratio (RER) over
time during sustained exercise and relatively greater FFA and AA oxidation for fuel (Blomstrand
& Saltin, 1999; Coyle et al., 1986; Hargreaves, 2000; Murray & Rosenbloom, 2018). Haralambie
& Berg, (1976) showed that serum urea content increased linearly with exercise over 70 minutes,
reflecting elevated AA oxidation during exercise >70 minutes in duration.
It was previously shown that endurance exercise performed at low-moderate intensity (50-
55% V̇O2max) alters whole-body protein kinetics during exercise but has little impact on protein
or AA balance in the 24-h post-exercise period. This may be a result of the relatively lesser CHO
depletion that occurs during low-moderate intensity exercise (Hawley & Hopkins, 1995; Van Loon
et al., 2001). El-Khoury et al. (1997) examined the effect of 90 min of moderate intensity (~46%
V̇O2max) endurance exercise during energy balance on whole-body leucine kinetics during
exercise and in the 24-h post-exercise period. They showed that moderate intensity exercise
increased leucine oxidation during exercise in both a fasted (~50%) and fed (~30%) condition,
however, this did not translate to changes in 24-h leucine oxidation or nitrogen retention. Forslund
et al. (1999) examined the effect of a similar moderate intensity exercise protocol (90 min at 45-
50% V̇O2max) and a ‘high’ (2.5 g⸱kg-1⸱d-1) versus ‘normal’ (1.0 g⸱kg-1⸱d-1) protein intake on
macronutrient utilization during a 24-h period. They showed that exercise significantly increased
protein oxidation and that protein contributed to a higher (22% vs 10%) and CHO to a lower (33%
vs 58%) proportion of total 24-h energy turnover in a high compared to normal protein diet.
Further, high dietary protein intake was associated with a positive protein balance during the 24-h
period. These studies examined only low-moderate exercise intensities (~50% V̇O2max) and as

22
such, these findings may not reflect 24-h protein balance following a high-intensity endurance
exercise bout. Interestingly, Devlin, Brodsky, Scrimgeour, Fuller, & Bier, (1990) showed that
following 3-h of high-intensity cycling exercise (75% V̇O2max), whole-body protein degradation
returned to pre-exercise baseline values within 2-h of exercise. Further, leucine oxidation was
shown to decrease in the post-exercise recovery period perhaps as a means to offset the exercise-
induced rise in AA (leucine) oxidation that occurs during the exercise bout (Rennie et al., 1981).
This study suggests that 24-h protein balance would ultimately be conserved after endurance
exercise. Thus, it may be inaccurate to extrapolate on measures of AA oxidation and protein
kinetics during exercise and over a short post-exercise time frame. These studies highlight the
importance of examining whole-body protein kinetics over an extended period of time in order to
accurately characterize the effects of exercise-induced AA oxidation on whole-body protein
kinetics in the post-exercise period. In agreement with this, a recent study by Bandagen et al.
(2019) (discussed further in Section 1.5.2) examined protein requirements of male endurance
athletes 24-h after their last training bout in order to minimize the acute effects of exercise on study
outcomes.
1.5 Estimating Dietary Protein Requirements for Endurance Athletes
Adequate provision of post-exercise dietary protein supports the repair/remodelling
process, increases whole-body protein synthesis and supports a net positive protein balance (Burd
et al., 2009). Importantly for endurance athletes, post-exercise dietary protein also supplies AA to
the free AA pool to replenish exercise-induced oxidative losses. Moreover, it is suggested to
support recovery, exercise adaptations (e.g. gene expression changes associated with cellular
energy pathways) and aerobic exercise performance (D’Antona et al., 2010; Di Donato et al., 2014;
Ferguson-Stegall et al., 2011; Hawley, Tipton, & Millard-Stafford, 2006; Rennie et al., 2000;

23
Rodriguez et al., 2007; Rowlands et al., 2011). To date, most protein-requirement studies have
used the nitrogen balance (NBAL) method, although recently, a number of studies have used stable
isotope technology through the indicator amino acid oxidation (IAAO) method. Both approaches
are discussed below.
1.5.1 Methods of estimating protein requirements
Nitrogen Balance
The nitrogen balance (NBAL) technique is a method of estimating protein requirements by
means of quantifying all protein that enters the body and all nitrogen that is excreted as a result of
protein breakdown (Tarnopolsky, 2004). NBAL is determined through the measurement of total
nitrogen intake (proteins are ~16% nitrogen) and nitrogen excretion in urine, feces, sweat and other
sources. NBAL is positive when the body is in a positive net protein balance (anabolism) while
NBAL is negative during net negative protein balance (catabolism). Protein requirement
estimations are determined through feeding various doses of protein (~3 intakes per subject) for at
least 7-10 days each to ensure urea pool adaptation to the test intake followed by estimating
nitrogen balance (Pencharz and Ball 2003; Rand et al., 2003). Linear regression analysis is used
to estimate the protein intake that will result in a “zero” NBAL which is the estimated average
requirement (EAR) with one standard deviation being the safe upper intake (Elango et al., 2008).
Tarnopolsky, MacDougall, & Atkinson, (1988) used NBAL to examine protein
requirements in elite-level endurance athletes, elite body builders and sedentary individuals
(control). NBAL data indicated that endurance athletes required 1.37 g⸱kg-1⸱day-1 of protein to
achieve nitrogen balance, which was ~1.7 times more protein than their sedentary counterparts and
also more than the elite body builders (Tarnopolsky et al., 1988). In a group of male endurance
athletes, Meredith and collegues (1989) used the NBAL method and examined whole-body protein

24
turnover and dietary protein requirements. Protein was consumed at three doses (0.6, 0.9 and 1.2
g⸱kg-1⸱d-1) for 10 d. It was determined that a protein intake of 1.2 g⸱kg-1⸱d-1 was required for male
endurance trained athletes. However, the NBAL method is limited in its ability to examine the
individual dynamic processes that influence protein turnover (i.e. rates of protein synthesis,
breakdown and oxidation) (Tarnopolsky, 2004). Additionally, the NBAL method has been
suggested to underestimate an individual’s true protein needs due to: i) inherent methodological
challenges associated with quantifying all routes of nitrogen loss, and ii) the application of linear
regression analysis to a nonlinear physiological response (Humayun, Elango, Ball, & Pencharz,
2007; Kato et al., 2016).
IAAO Method
The IAAO method is a non-invasive method to measure whole-body protein metabolism
through measuring changes in AA oxidation using an orally-administered stable isotope tracer (e.g.
L-[1-13C]phenylalanine), also referred to as the indicator AA (IAA) (Pencharz and Ball 2003;
Humayun et al. 2007; Elango et al., 2008). The IAAO method is based on the basic principle that
when one indispensable AA is deficient for protein synthesis, all other AA (including the
isotopically labelled IAA) will be oxidized (Pencharz and Ball 2003). With increased provision of
the limiting AA, IAA oxidation is reduced (Elango et al., 2008). This method is based on the
fundamental concept that AA cannot be stored in the body but rather have two metabolic fates; i)
incorporation into body protein, or; ii) oxidation (Pencharz and Ball 2003; Elango et al., 2008).
Therefore, when oxidation of the IAA is reduced, by increased provision of the limiting AA, it
reflects greater IAA (and by proxy all other AA) incorporation into body proteins (protein
synthesis).

25
The traditional IAAO method requires individuals to perform multiple metabolic trials in
which they are fed graded intakes of AA in the form of free AA in the composition of egg protein
(except for phenylalanine, tyrosine and the IAA; L-[1-13C]phenylalanine). A modified IAAO
method can be used to examine rates of whole-body protein synthesis and AA oxidation between
different exercise and nutrition conditions. This method involves the provision of a single
suboptimal protein dose for the specific population and measuring the difference in rate of IAA
oxidation (L-[1-13C]phenylalanine) between experimental conditions which is reflective of
differences in protein requirement to maximize whole body protein synthesis (Elango et al., 2008;
Gillen et al., 2019; Pencharz & Ball, 2003).
1.5.2 Current protein intake recommendations for endurance athletes
The IAAO method has been used in a number of studies to estimate protein requirements
in endurance-trained populations (Bandegan, Courtney-Martin, Rafii, Pencharz, & Lemon, 2019;
Kato et al., 2016). Kato et al. (2016) estimated the average protein requirement and recommended
protein intake in male endurance athletes using the IAAO method following a 20-km run at a self-
selected pace. They determined an EAR of 1.65 g⸱kg-1⸱d-1 and a safe intake (upper 95% CI) of
1.83 g⸱kg-1⸱day-1 of protein for this population to maximize protein synthesis (Kato et al., 2016).
Another study used the IAAO method in endurance-trained men over a 24-h period post-exercise
in order to minimize the acute effect of exercise on protein metabolism (Bandegan et al., 2019).
They estimated an EAR of 2.1 g⸱kg-1⸱d-1 and recommendation of 2.6 g⸱kg-1⸱d-1 which, was above
the estimate reported by Kato et al. (2016). Discrepancies seen between these two studies may be
a result of differences in habitual protein intake of the participants examined in each study.
Specifically, Tinline-Goodfellow et al. (2020) showed that a high habitual protein intake increases
the capacity for AA oxidation and may bias protein requirements determined by IAAO

26
methodology. Furthermore, and a 2-d dietary control was shown to be insufficient to ameliorate
the influence of a high protein intake on IAAO derived protein requirements (Tinline-Goodfellow
et al. 2020). Kato et al. (2018) also used the IAAO technique to examine the primary rate-limiting
amino acids following a prolonged 20-km run. Oxidation of ingested [1-13C]phenylalanine was
measured in four conditions; i) an optimal protein intake (1.75 g⸱kg-1⸱d-1) (based on (Kato et al.,
2016), ii) a deficient intake (0.8 g⸱kg-1⸱d-1), iii) a deficient intake with added BCAA, iv) a deficient
intake with added EAA or v) a deficient intake with added NEAA. They showed that breath
enrichment of F13CO2 and phenylalanine oxidation (PheOx) was attenuated in the sufficient protein
intake conditions and also in the deficient protein conditions that were enriched with BCAA, EAA
and NEAA. F13CO2 in the deficient intake + BCAA was 32% lower than the deficient conditions
respectively. The BCAA may be the primary limiting AA in the diets of endurance athletes
following bouts of prolonged endurance exercise and drive the elevated protein requirement. Thus,
the BCAA represent important nutrients to support post-exercise recovery (Kato et al., 2016;
Kimball & Jefferson, 2006). In line with previous NBAL studies, these studies using the IAAO
method indicate that protein needs of endurance athletes are in excess of the current RDA (0.8
g⸱kg-1⸱d-1) by 40-100% (Tarnopolsky et al., 2004; Rodriguez et al., 2007). Additionally, these
recommendations are greater than previous recommendations for endurance athletes of 1.2-1.4
g⸱kg-1⸱d-1 based on NBAL (Tarnopolsky et al., 2004).
1.5.3 Habitual protein intake of endurance athletes
Studies have also examined the habitual dietary protein intake of endurance athletes
(Burke et al., 2003; Gillen et al., 2017; Heikura, Stellingwerff, & Burke, 2018). Gillen et al.
(2017) examined daily dietary protein intake over a 2-4 week period in a large cohort of strength-
, endurance- and team-sport male and female athletes. Overall, the athletes consumed >1.2 g⸱kg-

27
1⸱d-1, while the males consumed approximately 1.5 g⸱kg-1⸱d-1, which was just lower than the
EAR of 1.65 g⸱kg-1⸱d-1 reported by Kato et al. (2016). Burke et al. (2003) reported eating patterns
in Australian Olympic athletes of a range of disciplines and found an average protein intake of
1.9 ± 0.5 g ⸱kg-1⸱d-1 among Australian Olympic endurance athletes participating in running,
cycling and swimming. Despite athletes habitually consuming protein close to (Gillen et al.,
2017) or above (Burke et al., 2003) the recommendation, these studies indicate that the
distribution of protein intake throughout the day and following training sessions was suboptimal
to maximize protein synthesis (Burke et al., 2003; Gillen et al., 2017; Mamerow et al., 2014).
Specifically, 58% of athletes’ protein intakes at breakfast, and 36% of intakes at lunch, were
below 20g, an intake previously shown to maximize PS (Moore et al., 2009) and below the
recent recommendation of 30g to maximize myofibrillar protein synthesis rates following a bout
of endurance exercise (Churchward-Venne et al., 2020).
1.6 Research Gaps and Conclusion
For trained endurance athletes, proper nutrition that is tailored to the unique physical
demands of their sport is an essential element of a successful training program (Heikura et al.,
2018). Understanding the effects of exercise intensity and duration on the relative contribution of
different substrates to support the energy demands of exercise is important to consider for trained
endurance athletes. Exercise intensity throughout a training season, and even over a single week
of training, varies substantially, from high intensity sessions to low intensity ‘recovery’ sessions.
Thus, substrate utilization will differ over an athlete’s training sessions. Accordingly, nutrition
strategies could be tailored to optimally fit athletes’ needs depending on the exercise demands (i.e.
exercise intensity and duration) and associated substrate utilization. Indeed, Heikura et al., (2018)
showed that most athletes focus on pre- and post-exercise nutrition (protein and CHO intake and

28
timing) around important training sessions and periods of the year. Moreover, nutrition strategies
are adopted by athletes with the primary objective of improving physique, exercise adaptations
and exercise performance (Heikura et al., 2018).
The present review summarizes our current understanding of energy metabolism during
exercise and the important influence of factors including CHO availability as well as exercise
intensity and duration on exercise-induced AA oxidation. Previously, it has been shown that
endurance-training athletes have elevated protein needs to facilitate post-exercise recovery and
muscle remodelling and to support exercise adaptations (Kato et al., 2016; Tarnopolsky, 2004).
Despite the well-documented relationship between exercise intensity and substrate utilization (Van
Loon et al., 2001; Romijn et al., 1993), the effect of exercise intensity during prolonged endurance
exercise on post-exercise protein requirements has not been determined. The present study
examined the effect of HIGH vs LOW relative exercise intensity during a prolonged 20-km run on
post-exercise protein requirements to maximize whole body protein synthesis. These findings may
inform periodized nutrition strategies for endurance training athletes throughout their training
season.

29
Chapter 2. Study Proposal
2.1 Introduction
Previous work out of our lab determined the impact of exercise intensity on substrate
utilization and AA oxidation during shorter duration endurance exercise. Male endurance athletes
completed a 10-km run at a high intensity (HIGH; 88-93% HR max) and low intensity (LOW; 70-
75% HR max). There was significantly greater rate of CHO oxidation and an elevated respiratory
exchange ratio (RER) in the high compared to low intensity condition during exercise. This
translated to an elevated rate of protein turnover following high intensity exercise which may
reflect an elevated need for protein remodelling post-exercise. However, oxidation of the indicator
AA in the post-exercise period was not different between the high and low intensity condition
which suggested that protein requirements do not appear to be elevated following a high intensity
CHO depleting exercise bout.
It remains unclear whether the relative intensity of prolonged endurance exercise
influences protein requirements. The 10-km exercise bout required <60 min to complete which
may not have elevated metabolic demands sufficiently to cause severe depletion of endogenous
muscle glycogen and/or elevations in AA oxidation during exercise. We speculated that a
prolonged duration of exercise would result in greater CHO depletion and AA oxidation during
exercise. This would translate to greater indicator AA oxidation in the post exercise period-in a
high compared to low intensity condition which reflects elevated dietary protein requirements
following high intensity exercise.
The present study will use the minimally invasive indicator amino acid oxidation (IAAO)
method to assess the effect of relative exercise intensity (HIGH; 88-93% HR max vs. LOW; 70-
75% HR max) during a prolonged 20-km run on phenylalanine oxidation (i.e. indicator AA) which

30
provides insight into whole body protein metabolism. This research will assess the effect of relative
intensity of a prolonged exercise on whole body protein metabolism and post exercise protein
requirements to support recovery and maximize protein synthesis in male endurance athletes.
2.2 Hypothesis
We hypothesize that performing an endurance exercise bout consisting of a 20-km run at a
high-intensity (88-93% HR max) compared to a low-intensity (70-75% HR max) will result in
greater excretion of our indicator amino acid in the breath which reflects, greater dietary protein
requirement following high- compared to low-intensity endurance exercise.
Secondly, we hypothesize that whole-body protein turnover will be elevated following high
compared to low intensity endurance exercise and that net protein balance will be less positive
following high intensity exercise. These between condition difference would reflect the greater
metabolic demands of high intensity exercise specifically, elevated total CHO oxidation (and
presumably muscle glycogen depletion) and consequently, elevated AA oxidation during exercise.
2.3 Research Phases
The present study has 3 research phases consisting of; i) Introduction and Consent (Session
1) ii) Fitness Assessment and Anthropometric measurement gathering (Session 2) and iii)
Experimental Trials: Two trials consisting of a 20-km run and an 8-hour metabolic trial (Session
3 and 4). Total time commitment for participation in the study was approximately 24 h (Session 1:
1 hours, Session 2: 3 hours, Session 3 & 4: 2 x 10 hours).
Phase 1 – Introduction and Consent (1 hour)
The first phase of data collection took place at the Goldring Centre for High Performance
Sport and lasted approximately 1 h. During this phase, participants received a verbal explanation

31
of the study and any risks and benefits of study participation by the investigator/co-investigator.
This was followed by a demonstration of equipment being used in the study and upon request, the
provision of a sample beverage similar to the hourly drinks provided during the metabolic trial
days. The potential participant had the chance to ask any questions and read over the consent form
(Appendix A) in private before giving their informed consent. Participants were required to
complete additional forms including; i) a CSEP ‘Get Active’ Questionnaire (Appendix B) to
ensure the participants were healthy and suitable to participate in the exercise and ii) a training
log/additional information questionnaire (Appendix C) to confirm that participants met the
inclusion criteria and did not meet any of the exclusion criteria for the study. Following all form
signing and completion, anthropometric measures were taken (height and mass) and second visits
(Phase 2) were scheduled.
Phase 2 - Fitness Assessment and Anthropometric Measurement collection (3 h)
Three days prior to coming into the lab for the second phase of research (Session 2),
participants completed a diet and training log and wore an accelerometer to track their daily
activity. In the morning of Session 2, participants entered the Goldring Centre for High
Performance Sport following a period of fasting (7 h minimum) and completed; i) body
composition analysis (BODPOD), ii) resting energy expenditure determination (Vmax), iii) a
fitness assessment (V̇O2peak test) and iv) a 10-km familiarization run. Successful completion of
each part of phase 2 confirmed the suitability of participates to participate in the study.
Phase 3 – Experimental trial: 20-km exercise and metabolic trial (10 h/session)
In phase 3, participants completed two experimental trials (Session 3 and 4) with exercise
(20-km run) done at a high (88-93% HR max) and low (70-75% HR max) relative intensity. These
two experimental trails were completed in a randomized order. Phase 3 (each experimental trial)

32
consisted of two out of lab days (d 1 and 2) where diet and exercise was controlled and an
accelerometer was worn to assess habitual activity levels. Day 3 took place in the lab in which
participants entered the lab fasted (7 h minimum) and completed; i) a 20-km treadmill run (at a
HIGH or LOW relative intensity) followed by, ii) an 8-hour metabolic trial (IAAO method).
Further information on each element of phase 3 is presented in Chapter 3 (Methodology).

33
Chapter 3. Methodology
3.1 Introduction
The following chapter will provide a detailed description of the elements of the study design
(Table 2.) and methodology in the present study. Each research phase (introduced in Chapter 2)
will be discussed in detail.
Table 1. Overall study protocol (grey bars indicate activities done outside of the lab setting).
Session 1: Introduction • Study explanation and consent form signing • Screening questions
(out of lab) • 3-day control diet, and accelerometer data collection (before session 2)
Session 2: Fitness Assessment and Familiarization
• Anthropometric measurements (BODPOD) • Resting Energy Expenditure determination (REE) • V̇O2peak test • 10-km familiarization run
(out of lab) Session 3 & 4: Day 1 & 2 • 2-day control diet and exercise (d 1: 10km, d 2: 5km at self-selected pace)
Session 3 & 4: Exercise and Metabolic trial (Day 3)
• Exercise session: 20-km run (HIGH or LOW) • Metabolic trial: 8 h drinks, breath and urine sample
collection
3.2 Participants and Study Design
A sample size of n=7 male endurance athletes was used for the present study. This is
consistent with the sample size from the study by Gillen, (2017) which served as the basis for the
study. Moreover, previous studies from our lab implementing different exercise and nutritional
manipulations to athlete populations (Gillen et al., 2019; Kato et al., 2016) have successfully
detected differences in breath 13CO2 enrichment (F13CO2) and phenylalanine oxidation (PheOx)
using the IAAO technique with n =7. The present study implemented a similar sample which we
believe will achieve sufficient statistical power to detect difference in the primary outcome
variable in this study (breath 13CO2 enrichment). Following completion of data collection and
analysis for all participants, post-hoc power analysis will be performed to confirm that we have
sufficient power to detect changes in our primary outcome variable from the intervention.

34
Participants were recruited via posters distributed on the University of Toronto campus and
at local running stores/clubs and through personal communication. Training status of each
participant was confirmed through the completion of a log of the previous month of training,
personal best times and a V̇O2peak test completed in Session 2 (Table 2; Session 2). Eligible
participants were required to; i) run at least 50km/week and/or perform at least 5 h of other aerobic
training modalities (i.e. swimming and cycling) weekly, ii) have a V̇O2peak categorized as “very
good” based on a study by Shvartz & Reibold, (1990) (≥57 ml⸱kg-1⸱min-1 (18-24 y), ≥54 ml⸱kg-
1⸱min-1 (25-29 y), ≥52 ml⸱kg-1⸱min-1 (30-34 y), ≥49 ml⸱kg-1⸱min-1 (35-39 y)) (as determined in
Session 2), iii) successfully complete a 10-km familiarization run (Table 2; Session 2), iv) no use
of tobacco, recreational drugs or banned substances (e.g. growth hormone, testosterone,
marijuana). Participant characteristics are presented in Table 3.
This study was approved by the University of Toronto Health Sciences Research Ethics
Board and was conducted in accordance with the Declaration of Helsinki.
Table 2. Participant characteristics 1 Characteristic Value Age (y) 26 ± 3 Body mass (kg) 76.1 ± 16.1 Fat free mass (kg)2 69.1± 13.8 Height (cm) 181 ± 8 V̇O2 peak (ml⸱kg-1⸱min-1)3 62.2 ± 6.2 Heart rate max (bpm)3 189 ± 5 REE (kJ)4 7568 ± 1518 Habitual dietary protein intake (g⸱kg-1⸱day-1) 1.8 ± 0.6 Habitual dietary CHO intake (g⸱kg-1⸱day-1) 4.8 ± 2.0
1Values are mean ±SD n=7 participants 2Fat-free mass measured using the BODPOD (COSMED USA Inc.) 3V̇O2 peak and Heart rate (HR)max based on V̇O2max test (Session 2) 4REE, Resting energy expenditure based on Weir equation and gas exchange data at rest (30 min)
3.3 Informed Consent and Familiarization
The study was conducted in the Iovate/Muscletech Metabolism and Sports Science Lab at
the Goldring Centre for High Performance Sport. During the initial visit, participants received a

35
verbal explanation of the study and demonstration of equipment and techniques used followed by
completion of a consent form (Appendix A) and screening questions/training log (Appendix C)
(Table 2; Session 1). Prior to Session 2, an accelerometer was used to estimate participant’s
habitual activity level and ‘free-living’ energy expenditure and a 3-d food record was completed
by participants to determine habitual dietary intake.
3.4 Fitness Assessment and Anthropometric Measurement Collection
On the morning of Session 2, participants entered the lab following an overnight fast (7 h
minimum). The BODPOD (COSMED USA Inc.) was used to determine participants’ baseline
whole body mass, fat mass (FM) and fat-free mass (FFM). Following body composition
assessments, resting energy expenditure (REE) was determined with the Weir equation (1) and by
gas exchange data taken at rest in the fasting condition for 30 min using Vmax SPECTRA 29
(SensorMedics Corporation)(Weir, 1949).
REE (kcal/day)= [1440 (3.94 * V̇O2 absolute) + (1.11 * VCO2 absolute)] (1)
Participants then performed a standard treadmill-based ramp protocol fitness assessment
(exercise duration ~12-15 min) to determine peak oxygen consumption. Prior to beginning the test,
participants received a protein-free breakfast (1.2 g·kg-1 CHO) and were familiarized with the
fitness assessment protocol and equipment. A heart rate monitor (Polar) was worn on the chest and
a mask was worn to measure inhaled and exhaled air during the fitness assessment. Gas exchange
was monitored continuously throughout the fitness assessment using GA-300 gas analyzer
(iWorx). Following completion of the fitness assessment, participants were allowed to recover for
a self-selected amount of time and then complete a 10-km run. During the 10-km run, the first 5-
km was completed at a pace that elicited ~70-75% of maximal heart rate (HRmax), and the second
5-km was completed at a pace that elicited ~88-93% HRmax. This session served as familiarization

36
with the high and low intensity running pace for the subsequent exercise trials and informed
investigators on the speed required to elicit the appropriate %HRmax (Table 2; Session 2).
3.5 Experimental Trials
Control diet and exercise
A 2-day standardized diet and exercise program was followed outside of the laboratory by
participants before the completion of both metabolic trials (Figure 2; D 1 & 2). The standardized
exercise consisted of a 10-km run (self-selected pace) on day 1 and a 5-km run (self-selected pace)
on day 2. Standardized exercise ensured that individuals performed exercise that was similar to
their habitual training in the days leading up to each metabolic trial.
The energy content of the control diet was provided as 1.6 * REE plus the exercise-induced
energy expenditure from the 2-d control exercises (estimated as 1kcal·kg-1·km-1 based on Margaria
et al., 2013) as previously used (Gillen et al., 2019; Kato et al., 2016). The controlled diet consisted
of commercially available pre-packaged foods that were tailored to the dietary restrictions and/or
preferences of each participant. Diets contained 8 g·kg-1·d-1 of CHO on d 1 and 7 g·kg-1·d-1 of CHO
on d 2. Diets on both days contained 1.4 g·kg-1·d-1 of protein, which was selected as it has been
suggested to meet the protein requirements of active males (Rodriguez et al., 2000), This intake is
also within the normal day-to-day intake variability of this endurance trained population (Gillen
et al., 2017) and the 95% CI of our previously determined daily requirement for this population
(Kato et al., 2016).
Exercise session and metabolic trail
The exercise session and metabolic trials (Figure 2; Day 3) were completed in the
laboratory with a minimum of 4 d between each trial which served as a washout period. The
participants entered the lab in the morning after an overnight fast (minimum of 7 h) and no caffeine

37
ingestions for a minimum of 12 h before entering the lab. A pre-exercise breakfast drink containing
0.6 g·kg-1 CHO as a 1:1 ratio of maltodextrin (Polycal®; Nutricia, Amsterdam, Netherlands) and
sports drink powder (Gatorade® Endurance Formula; PepsiCo, Purchase, NY) was consumed ~15-
30 min before exercise by all participants. The study followed a crossover design in which
participants were randomized to the two exercise conditions; 20-km run at i) HIGH and ii) LOW
relative exercise intensity. The low intensity 20-km run was completed at 70-75% HR max and
high intensity 20-km was done at 88-93% HR max. Throughout the exercise trials exercise
intensity and relative fuel use were measured. Heart rate (HR) was monitored continuously with a
heart rate monitor (Polar) and oxygen consumption (V̇O2) was measured for ~10 min at 3 time
points during exercise (at 2-km, 8-km and 18-km) using a GA-300 gas analyzer
(iWorx). Following completion the exercise, participants consumed a post-exercise meal to aid in
the recovery and energy replenishment from the 20-km run. The meal consisted of a beverage
containing 2.4 g·kg-1 CHO as a 1:1 ratio of maltodextrin (Polycal®; Nutricia, Amsterdam,
Netherlands) and sports drink powder (Gatorade® Endurance Formula; PepsiCo, Purchase, NY)
and protein free cookies (~10g).

38
Figure 2. Schematic illustration of exercise and metabolic trial sessions.
3.6 Data Collection and Analysis
Assessment of protein required to maximize whole body protein synthesis
Following the 20-km run and post-exercise recovery meal, a metabolic trial using the
IAAO technique was performed to assess between condition (HIGH vs LOW) differences in post-
exercise phenylalanine metabolism. Participants were required to stay in the lab (Goldring Centre
for High Performance Sport) for the duration of the metabolic trial.
The metabolic trial involved the hourly (8 h) consumption of isoenergetic, beverages
containing 0.93 g·kg-1·d-1 of protein (in a crystalline amino acid form) enriched with stable isotope
[13C]phenylalanine and sufficient CHO and fat to meet the post-exercise energy needs of the
Day 3Day 2Day 1Control Diet and Exercise Exercise and Metabolic Trial
Control exercise:10km run (self-selected pace)
Control diet:8g/kg/d CHO1.4g/kg/d PRO
Control exercise:5km run (self-selected pace)
Control diet:7g/kg/d CHO1.4g/kg/d PRO
Enter lab overnight fast (7h) Pre-exercise
0.6g/kg CHO20km run
HIGH (88-93% HRmax)LOW (70-75% HRmax)
Post-exercise 2.4g/kg CHO and
protein free cookies
8-hour metabolic trial

39
participants. The use of a stable isotope tracer allows for the examination of the kinetics of the
indicator AA, phenylalanine in the body. Amino acids in the dinks were modelled off of the
composition of egg protein with the exception of the indicator amino acid (phenylalanine), which
was given at a constant intake (30.5 mg·kg-1·d-1) and tyrosine which was given in excess (40
mg·kg-1·d-1). Providing tyrosine in excess ensured the two fates of the indicator amino acid in the
body to i) incorporation into body protein (protein synthesis) or ii) oxidation (Zello et al., 1990).
When an essential amino acid is deficient for protein synthesis, all other amino acids
including the indicator amino acid (phenylalanine) will be oxidized. With increasing provision of
the limiting (deficient) AA, there is reduced indicator amino acid oxidation which reflects direction
of amino acids towards synthesis. Thus, a protein intake of 0.93 g·kg-1·d-1 was selected as it is the
EAR determined using the NBAL technique (Meredith et al., 1989) and has been shown to be
insufficient for endurance trained males when using the IAAO technique (Kato et al., 2016).
Moreover, this intake has previously been used in our lab when studying male endurance athletes
(Gillen et al., 2019). This intake allowed for the assessment of between condition differences in
oxidation of the indicator AA, which is assumed to reflect the protein requirement to maximize
whole body protein synthesis between the conditions (HIGH vs LOW exercise intensity).
Participants were occasionally required to provide breath and urine samples (illustrated in
Figure 2. Baseline breath (collected at 45, 30 and 15 min) and urine samples (collected at 45 and
15 min) were collected prior to the ingestion of the 5th hourly drink. In the 5th meal, a priming
dose of NaH13CO3 (0.176mg/kg Cambridge Isotope Laboratories (CIL) Canada, Inc.) and L-
[13C]phenylalanine (1.86mg/kg; CIL Canada, Inc.) was ingested with each subsequent hourly
meals containing 1.20mg/kg L-[13C]phenylalanine (CIL Canada, Inc.) to maintain isotopic plateau
(Kato et al., 2016). Two and a half h following the ingestions of the 5th hourly drink, steady state

40
breath samples (6; every 15 min) and urine samples (3; every 30 min) were collected. Breath
samples were collected in Exetainer tubes (Labco, Ltd) and analyzed for 13CO2 enrichment by
continuous-flow isotope ratio mass spectrometry. Urine samples were collected in sterile urine
specimen cups and frozen at -20°C. In preparation for analysis, urine samples were thawed and
then deproteinized and acidified with methanol and spun down in the centrifuge. Supernatant was
removed and the sample was dried down under N2 gas. Samples were than reconstituted in a buffer
made of formic acid and acetonitrile. L-[1-13C]-phenylalanine enrichment in the prepared urine
samples was measured by liquid chromatography-tandem mass spectrometry (LC-MS/MS).
Steady state CO2 production was measured approximately 30 min after the ingestion of the
5th hourly drink by indirect calorimetry (Vmax SPECTRA 29, SensorMedics Corporation) (Figure
2; VCO2). This time point was based on the pilot work from Kato et al. (2016) which confirmed
stable background isotopic and metabolic steady state is achieved at this time point. Determination
of VCO2 allowed for the conversation of [13C]phenylalanine enrichment in the breath to an
absolute value of excretion (F13CO2) which was calculated using an equation (2) by Matthews et
al. (1980) which was previously used (Gillen et al., 2019; Kato et al., 2016).
𝐹%&𝐶𝑂) =+,-.∗0,-.
12 34∗5%.3%44∗4.7%
8 (2)
where FCO2 is the CO2 production rate (cm3 /min); ECO2 is the 13CO2 enrichment in the
expired air at isotopic steady state (APE); W is the participant’s body mass (kg). Constants 60
(min/h) and 41.6µmol/cm3 (at standard temperature and pressure) convert FCO2 to µmol/h. The
factor of 100 changes APE from a percent to a fraction and the factor of 0.81 accounts for the
fraction 13CO2 released from [1-13C]phenylalanine oxidation but not released from the body
bicarbonate pool into the expired air (Matthews et al., 1980).
Tracer kinetics

41
Isotopic enrichment of 13CO2 in the breath and L-[1-13C]-phenylalanine in the urine were
used to calculate Phenylalanine turnover or flux (PheRa µmol∙ kg-1h-1) (3), phenylalanine oxidation
(PheOx µmol∙ kg-1h-1) (4), whole body protein synthesis (PS µmol∙ kg-1h-1) (5) and breakdown (PB
µmol∙ kg-1h-1) (6) and net balance (NB µmol∙ kg-1h-1) (7) were calculated based on the models by
Matthews et al. (1980).
𝑃ℎ𝑒𝑅𝑎 = 𝑖 2?@?A− 18(3)
𝑃ℎ𝑒𝑂𝑥 =𝐹%&𝐶𝑂) 2%0A− %
0@8 ∗ 100 (4)
𝑃𝑆 = 𝑃ℎ𝑒𝑅𝑎 − 𝑃ℎ𝑒𝑂𝑥 (5)
𝑃𝐵 = 𝑃ℎ𝑒𝑅𝑎 − 𝐼 (6)
𝑁𝐵 = 𝑃𝑆 − 𝑃𝐵 (7)
where 𝑖 is the [1-13C]phenylalanine ingestion rate (7.22 µmol∙ kg-1h-1); 𝐸Kis the
enrichment of the [1-13C]phenylalanine ingested (APE) (99%); 𝐸L, the [1-13C]phenylalanine
enrichment in urine at isotopic plateau (APE); I is the rate of exogenous phenylalanine intake
(15.38 µmol∙ kg-1∙h-1)(Matthews et al., 1980).
3.6.1 Statistical Analysis
Paired sample t-tests (IBM SPSS Statistical Software Version 17.0) were performed to
determine between condition (HIGH vs. LOW) differences in mean F13CO2, PheRa and PheOx,
whole body protein synthesis and protein breakdown and net protein balance. Cohen’s d effect size
(small: 0.2, medium: 0.5, large: 0.8) was determined for mean F13CO2, PheRa and PheOx, whole
body protein synthesis and protein breakdown and net protein balance between the HIGH and
LOW condition. A 2-way repeated measures ANOVA (condition x time) was conducted to assess
during exercise variables including substrate utilization, heart rate and oxygen consumption.
Significance was set at P<0.05.
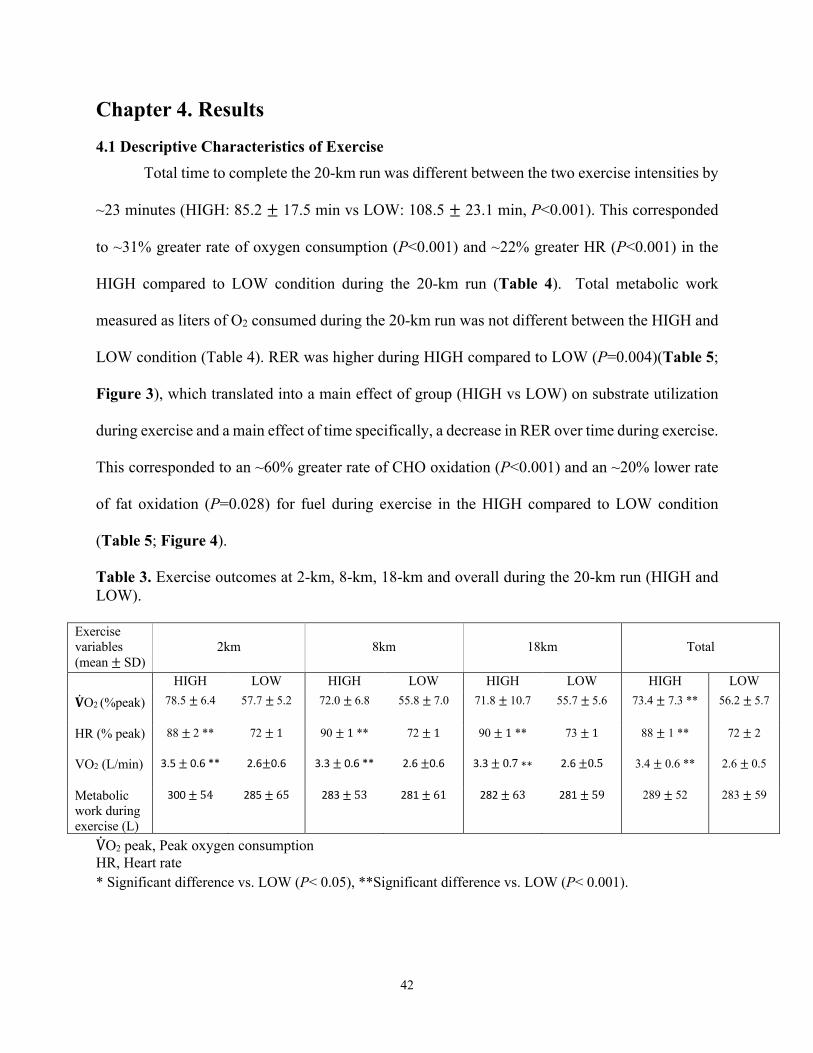
42
Chapter 4. Results
4.1 Descriptive Characteristics of Exercise Total time to complete the 20-km run was different between the two exercise intensities by
~23 minutes (HIGH: 85.2 ± 17.5 min vs LOW: 108.5 ± 23.1 min, P<0.001). This corresponded
to ~31% greater rate of oxygen consumption (P<0.001) and ~22% greater HR (P<0.001) in the
HIGH compared to LOW condition during the 20-km run (Table 4). Total metabolic work
measured as liters of O2 consumed during the 20-km run was not different between the HIGH and
LOW condition (Table 4). RER was higher during HIGH compared to LOW (P=0.004)(Table 5;
Figure 3), which translated into a main effect of group (HIGH vs LOW) on substrate utilization
during exercise and a main effect of time specifically, a decrease in RER over time during exercise.
This corresponded to an ~60% greater rate of CHO oxidation (P<0.001) and an ~20% lower rate
of fat oxidation (P=0.028) for fuel during exercise in the HIGH compared to LOW condition
(Table 5; Figure 4).
Table 3. Exercise outcomes at 2-km, 8-km, 18-km and overall during the 20-km run (HIGH and LOW).
Exercise variables (mean ± SD)
2km
8km
18km
Total
HIGH LOW HIGH LOW HIGH LOW HIGH LOW
�̇�O2 (%peak)
78.5 ±6.4 57.7 ±5.2 72.0 ±6.8 55.8 ±7.0 71.8 ±10.7 55.7 ±5.6 73.4 ± 7.3 ** 56.2 ±5.7
HR (% peak)
88 ±2 ** 72 ±1 90 ±1 ** 72 ±1 90 ±1 ** 73 ±1 88 ± 1 ** 72 ± 2
VO2 (L/min)
3.5 ± 0.6 ** 2.6±0.6 3.3 ± 0.6 ** 2.6 ±0.6 3.3 ±0.7 ∗∗ 2.6 ±0.5 3.4 ± 0.6 ** 2.6 ±0.5
Metabolic work during exercise (L)
300 ±54 285 ±65 283 ±53 281 ±61 282 ±63 281 ±59 289 ± 52 283 ± 59
V̇O2 peak, Peak oxygen consumption HR, Heart rate * Significant difference vs. LOW (P< 0.05), **Significant difference vs. LOW (P< 0.001).

43
Table 4. Substrate utilization during 20-km running performance in LOW and HIGH. Exercise variables (mean ± SD) LOW HIGH
RER 0.88 ±0.03 0.92 ± 0.02 *
Fat oxidation (g/min) 0.5 ± 0.2 0.4 ± 0.2 *
CHO oxidation (g/min) 1.9 ± 0.5 3.1 ± 0.7 **
Total fat oxidation (g) 54.8 ± 20.5 36.1 ± 14.6 *
Total CHO oxidation (g) 204.9 ± 38.9 260.8 ± 50.5 *
RER, Respiratory exchange ratio CHO, Carbohydrate * Significant difference vs LOW (P< 0.05), **Significant difference vs LOW (P< 0.001).
Figure 3. RER at 3 time points (2km, 8km, 18km) during 20-km run in HIGH and LOW. * denotes main effect of condition, letters (A,B, C) denote differences over time (mean ± SD, P<0.05).
2 4 6 8 1 0 1 2 1 4 1 6 1 8
0 .7 5
0 .8 0
0 .8 5
0 .9 0
0 .9 5
1 .0 0
1 .0 5
D is ta n c e (k m )
RE
R
LO W
H IGH *
A
B
C

44
Figure 4: Percent CHO (top) and fat (bottom) oxidation during exercise over time (2km, 8km, 18km). Letters (A, B, C) denote difference over time, * denotes effect of condition (HIGH vs. LOW) (mean ± SD, P<0.05).

45
4.2 Phenylalanine Oxidation and Tracer Kinetics There was no difference in [13C]phenylalanine excretion (F13CO2) between HIGH and
LOW (P=0.91) (Figure 5).
Figure 5. F13CO2 following 20-km run in LOW (grey bars) vs. HIGH (white bars). Black dots represent individual data points (mean ± SD).
Preliminary results of n=6 participants revealed no difference between conditions in
phenylalanine flux (PheRa; HIGH: 52.7 ± 8.7 vs LOW: 42.0 ± 9.8 µmol∙ kg -1∙h-1, P=0.12, d=
0.80, PheRa relative to FFM; HIGH: 58.5 ±8.7 vs. LOW: 47.1 ± 11.4 µmol∙ kg FFM-1∙h-1, P=0.11,
d=0.78). Further, phenylalanine oxidation (PheOx) was not statistically different between
conditions when normalized to body mass (P=0.10) or FFM (P=0.094) despite large effect sizes
of d=0.81 and d=0.84, respectively, in favor of HIGH (Figure 6). Whole-body protein synthesis
(PS; P=0.14, d=0.72) and protein breakdown (PB; P=0.11, d=0.78) were not different between
conditions (Figure 6). Net protein balance (NB) was not statistically different between conditions
when normalized to body mass (P=0.10) or FFM (P=0.09); however, there were large effect sizes
of d=-0.81 and d=-0.84 respectively, in favor of a more positive NB in LOW (Figure 7).

46
Figure 6. PheOx (top), protein synthesis (middle), protein breakdown (bottom) following 20-km run in LOW (grey bars) vs. HIGH (white bars). Black dots represent individual data points (mean ± SD).

47
Figure 7. Net protein balance following 20-km run in LOW (grey bars) vs. HIGH (white bars). Black dots represent individual data points (mean ± SD).

48
Chapter 5. Discussion
5.1 Overview of the Findings
Endurance athletes have dietary protein requirements above the current population RDA
based on NBAL methodology (0.8 g∙ kg-1∙d-1) (Humayun et al., 2007; Rodriguez et al., 2009). The
current protein intake recommendation for endurance trained athletes is 1.2-1.4 g∙ kg-1∙d-1
(Rodriguez et al., 2009). More recently, Kato et al. (2016) estimated a greater dietary protein
requirement and recommendation of 1.65 and 1.83 g∙ kg-1∙d-1 to maximize whole-body protein
synthesis during recovery from exercise in male endurance athletes. These elevated needs are in
part to replenish AA oxidative losses that occur during exercise and maximize whole-body protein
synthesis to support recovery after exercise (Kato et al., 2016; Tarnopolsky, 2004). The present
study used the minimally invasive IAAO technique to examine the effect of a high intensity, CHO-
dependent 20-km run compared to lower intensity 20-km run (HIGH vs LOW) on whole body
protein turnover and to estimate post-exercise protein requirements to maximize whole body
protein synthesis in male endurance athletes. We observed that the rate of CHO utilization during
exercise was significantly greater (~60%) in the high intensity compared to the low intensity
exercise bout. The high intensity condition also increased protein turnover in the post-exercise
period as there was a moderate effect size for PheRa (d=0.78) that nevertheless did not reach
significance (P=0.11), which might have been related to our underpowered preliminary data.
Similar moderate effect sizes were seen for whole-body PS (d=0.72) and PB (d=0.78) post-exercise
although not statistically significant. These results demonstrate that higher-intensity exercise
increases whole-body protein remodelling during the post-exercise recovery period.
We also observed a large effect size (d=0.84) (P=0.094) for indicator AA oxidation
(PheOx) between the high and low intensity condition following exercise. Elevations in PheOx in

49
the high intensity condition reflect AA being limiting and deficient for whole-body PS in the post-
exercise period. In line with these results, there was a large effect size (d=-0.84) for NB with low
intensity being greater than high intensity. Collectively, these data suggest that the post-exercise
protein requirement to maximize whole-body protein synthesis is greater to support a positive net
protein balance following prolonged high-intensity compared to lower-intensity exercise.
5.2 Exercise Outcomes
The exercise stimuli used in the present study were effective in eliciting different exercise
intensities and substrate use. On average, the high and low intensity conditions elicited a heart rate
of ~88% and ~72% HR max, respectively, which corresponded to relative exercise intensities of
~73% and ~56% V̇O2peak, respectively. There was no effect of time on % HRpeak or % V̇O2peak
during exercise, which confirmed that participants were able to sustain the target exercise intensity
throughout the entire duration of exercise. There was a difference in substrate utilization between
the high intensity and low intensity exercise condition in the present study, which is consistent
with previous research (Karlsson et al., 1974; Romijn et al., 1993, 2019; Van Loon et al., 2001).
Specifically, we saw an ~60% greater rate of CHO oxidation during the high intensity exercise
condition that translated into a greater total CHO oxidation during exercise by ~27% or ~57g.
Further, there was a higher RER during high intensity exercise and RER decreased over time
during sustained exercise in both conditions reflecting relatively greater FFA and possibly AA
oxidation for fuel in the later stages of exercise (Blomstrand & Saltin, 1999; Coyle et al., 1986;
Hargreaves, 2000; Murray & Rosenbloom, 2018; Schuetz et al., 1995). Taken together, the
exercise stimuli (HIGH vs. LOW) were completed at significantly different relative exercise
intensities and resulted in diverging patterns of substrate utilization during exericse.

50
It is assumed that RER (i.e. ratio of the volume of CO2 released and O2 consumed at the
mouth) accurately reflects RQ (i.e. ratio of the volume of CO2 release and O2 absorbed at the tissue
level) although, RER does not account for protein in the calculation as it is assumed that protein
oxidation is negliable compared to CHO and lipid oxidation (Patel & Bhardwaj, 2018). In some
sitautions, such as extended periods of glycogen depleting exercise, protein (AA) oxidation
accounts for a considerably larger amount of total energy (5-10%) (Lemon & Nagle, 1981). Protein
oxidation can be estimated from nitrogen excretion in the urine and sweat or by tracer methodoloy
to directly measure AA carbon oxidation or urea production (Lemon & Nagle, 1981) however, this
was not done in the present study. Thus, RER in the present study may overestimate CHO
utilization in late exercise if there is a greater reliance on protein as a substrate (AA oxidation).
Baseline muscle glycogen and the prevision of exogenous pre-exercise CHO also have
important implications for CHO availability and utilization as a function of exercise duration
(Areta & Hopkins, 2018; Hawley et al., 1997). In the present study, the prolonged 20-km exercise
bout took 85±17 min (HIGH) and 108±23 min (LOW) to complete. Near this duration of high
intensity exercise (i.e. >90 minutes) has previously been suggested to result in severe muscle
glycogen depletion (Areta & Hopkins, 2018; Hawley et al., 1997). The prevision of exogenous
pre-exercise CHO (0.6 g∙ kg-1) consumed ~15-30 minutes before the initiation of the exercise bout
in the present study may have limited the time in which individuals were exercising in a low
glycogen state. Pre-exercise CHO ingestion increases glucose uptake into the muscle, replenishes
hepatic glycogen stores and augments muscle glycogenolysis and CHO oxidation (Costill et al.,
1977; Coyle et al., 1997; Jeukendrup & Killer, 2011). It has also been shown to reduce hepatic
glucose reliance during exercise (Costill et al., 1977; Marmy-Conus et al., 1996) and slows the
depletion endogenous CHO (e.g. muscle glycogen) during exercise (Costill et al., 1977; Coyle et

51
al., 1997). Based on the rates of CHO oxidation in the high (3.1 g∙ min-1) and low (1.9 g∙ min-1)
intensity conditions, the pre-exercise exogenous CHO would have fueled exercise for ~13 minutes
in the high intensity and ~22 minutes in the low intensity condition. Thus, while between condition
differences (HIGH vs LOW) in patterns of substrate utilization during exercise were seen, these
differences may have been more pronounced if pre-exercise CHO had not been consumed and/or
if exercise was performed in a fasted condition, both of which are nutrition conditions that athletes
commonly train in (Febbraio et al., 2000).
In the present study, the 2 d control diet consisted of a CHO intake of 8 g∙ kg-1∙d-1 (d 1)
and 7 g∙ kg-1∙d-1 (d 2). This has been used as a dietary lead in for previous studies in of our lab
(Gillen, 2017; Kato et al., 2016, 2018a) and is suggested to meet daily energy demands and be
sufficient to recover muscle glycogen loss from the pre-trial controlled exercise (i.e. 15km over 2
d) (Burke et al., 1995; Costill et al., 1981). Based on the meta-analysis by Areta & Hopkins, (2018)
and the 2-d high CHO intake for endurance trained males in the present study (V̇O2peak ~60 ml∙
kg-1∙min-1), it could be estimated that resting (baseline) glycogen content of participants in the
present study was ~600 mmol∙ kg-1 dry mass upon beginning experimental trial days. Areta &
Hopkins, (2018) showed that the rate of glucosyl unit depletion during exercise at 40% of V̇O2max
was ~1.48 mmol∙ kg dry mass-1∙min-1 and during exercise at 70% V̇O2max was ~2.91mmol ∙ kg dry
mass-1∙min-1. We estimated that in the present study during high intensity exercise, the rate of
glucosyl unit depletion would be slightly greater than the ~2.91 mmol∙ kg dry mass-1∙min-1 seen
during exercise at 70% V̇O2peak by Areta & Hopkins, (2018). During the low intensity condition
we estimate a rate of glucosyl unit depletion of ~2.19 mmol∙ kg dry mass-1 ∙min-1 based on the rates
of depletion at 70% and 40% V̇O2max by Areta & Hopkins, (2018). Thus, we estimate that the
muscle glycogen content would be ~340 mmol∙ kg dry mass-1 at the end of high intensity exercise

52
and ~400mmol∙ kg dry mass-1 at the end of low intensity exercise. Interestingly, the estimation of
muscle glycogen content at the end of high intensity exercise in the present study (~340 mmol∙ kg
dry mass-1) is only slightly higher than the muscle glycogen concentrations at rest and in the early
stages of exercise in a low-CHO condition (~300 mmol∙ kg dry mass-1) in an earlier study by
Howarth et al. (2010). In the Howarth et al. (2010) study, it was shown that compared to a high-
CHO condition (resting muscle glycogen content ~440 mmol∙ kg dry mass-1) a low-CHO condition
(resting muscle glycogen content of ~300 mmol∙ kg dry mass-1) had an ~30% greater rate of leucine
oxidation during exercise. Moreover, leg phenylalanine Ra was elevated within the first 10 min of
exercise in the low-CHO condition suggesting an elevated rate of MPB when CHO availability is
limited during exercise. In line with this, while both conditions had a negative leucine NB, the
low-CHO condition was ~58% more negative during exercise than the high-CHO condition. Thus,
greater CHO utilization during high compared to low intensity exercise in the present study
presumably resulted in a greater reduction in endogenous CHO stores (e.g. muscle glycogen) and
consequently, elevated utilization of AA as a respiratory substrate (AA oxidation) during exercise.
While endogenous CHO stores were likely reduced to a greater extent during high intensity
exercise in the present study, pre-exercise CHO ingestion and high initial muscle glycogen content
may have minimized the between-condition differences in substrate utilization during exercise.
Indeed, high CHO availability augments CHO oxidation during exercise, has a potent inhibitory
effect on fat oxidation and may ultimately reduce exercise-induced AA oxidative loss (Sidossis et
al., 1996; Horowitz et al., 1997; Spriet, 2014). In line with this, Gillen et al. (2019) showed that
individuals performing a prolonged exercise bout in a glycogen depleted state had elevated post-
exercise protein requirements (~15%) to maximize whole-body protein synthesis. This study
highlighted that beginning exercise with reduced resting muscle glycogen content and without the

53
ingestion of pre-exercise CHO may increase AA oxidation and protein catabolism during exercise.
Low endogenous CHO availability (e.g. muscle glycogen) would increase the between-condition
(HIGH vs LOW) differences in indicator AA oxidation and protein requirements in the post-
exercise recovery period. Thus, endurance athletes who commence long runs with limited
endogenous CHO availability (Stellingwerff, 2012) may have to consider the intensity at which
they train at as influencing their post-exercise protein requirements.
5.3 Phenylalanine Oxidation and Tracer Kinetics
To account for the heterogeneity in body composition measures between participants in the
present study (see Chapter 3, Table 3), outcome variables were expressed relative to participant’s
fat free mass (FFM). Due to Covid-19 constraints, preliminary data (n=6) are only available for
the following outcomes; PheRa, PheOx, PS, PB and NB.
Phenylalanine turnover/flux (PheRa)
Preliminary results revealed that phenylalanine turnover/flux (PheRa) was not significantly
different between the high and low relative exercise intensities; however, the moderate effect size
(d=0.78) suggests that we may have been underpowered to detect differences in PheRa and other
protein kinetic outcomes in the preliminary (n=6) results. Indeed, retrospective power analysis
revealed that a sample size of n=7 was needed to achieve statistical power of 0.5 and n=14 was
needed to achieve the desired statistical power (0.8) to detect changes in our primary outcome
variable from the intervention. Nevertheless, our results suggest that the rate of appearance of
phenylalanine in the free AA pool was likely affected by the exercise conditions (HIGH vs LOW
intensity), which indicates that the precursor pool size for PS was influenced by the exercise
conditions in the present study (Koopman et al., 2004). Similar results were seen in the study by
Gillen (2017) who showed that PheRa tended (P=0.08) to be greater in a high intensity compared

54
to low intensity 10-km run condition and had a moderate effect size (d=0.78). Thus, differences in
PheRa (flux) need to be accounted for in calculations of phenylalanine kinetics.
The traditional IAAO method examines excretion 13C in the breath (F13CO2) as a metric of
indicator AA oxidation as F13CO2 is considered to be a robust measure and closely aligned with
rates of phenylalanine hydroxylation (Rafii et al., 2008). There was no difference in 13C excretion
in the breath (F13CO2) between the high intensity and low intensity exercise condition in the
present study. This was consistent with results from Gillen (2017), which were numerically similar
to the present study and showed no difference between high and low exercise intensity on F13CO2
following a 10-km run. Excretion of 13C in the breath (F13CO2) in the present study was ~44%
greater in both the high intensity and low intensity exercise conditions compared to Kato et al.
(2018a) despite similarities in; i) the endurance trained male population examined and ii) the 20-
km exercise stimulus (~72% HRmax). This discrepancy may be due to the use of different
metabolic carts which could impact the VCO2 rates required to calculate F13CO2 (see Chapter 2,
equation 1). Thus, it may be difficult to make comparisons between studies in which different
metabolic carts were used.
Given the influence of the aforementioned factors and the innate interindividual variability,
a crossover study design with the use of a single metabolic cart for all VCO2 measurements is the
most appropriate method of examining differences in 13CO2 breath enrichment. Nevertheless, an
assumption of the traditional IAAO method and specifically for F13CO2 enrichment values is that
the precursor pool size does not change in response to the exercise or dietary intervention. The
moderate effect size seen for phenylalanine flux (PheRa) indicates that the exercise intensity
(HIGH vs LOW) had an effect on precursor pool size (Bross et al., 1998). This may be a result of
an elevated rates of whole-body protein breakdown following high intensity exercise. A possible

55
intensity-dependent increase in whole body protein breakdown is consistent with some, but not all
(Devlin et al., 1990), previous research demonstrating elevated rates of muscle (Harber et al., 2010;
Howarth et al., 2010; Sheffield-Moore et al., 2004; Pikosky et al., 2006) and whole-body PB
(Bowtell et al., 2000; Phillips et al., 1993; Rennie et al., 1981; Sheffield-Moore et al., 2004)
following bouts of endurance exercise. Therefore, as PheOx accounts for differences in precursor
pool size through (i.e. [1-13C]phenylalanine urinary enrichment) it arguably provides a more
physiologically relevant estimate of indicator AA oxidation and a comprehensive picture of protein
kinetics in the present study, as suggested previously (Malowany et al., 2019; Mazzulla et al.,
2020). Thus, the balance of the discussion will focus on the phenylalanine kinetic data.
Phenylalanine Oxidation (PheOx)
Exercise-induced AA oxidation, which is influenced by the intensity and duration of
exercise (Millward et al., 1994; Rennie et al., 1981;Van Loon et al., 2001;), is suggested to be the
primary driver of the catabolic effect of endurance exercise. Additionally, some AA may be used
to increase substrate availability for anaplerosis of the TCA cycle in muscle during exercise
(Gibala et al., 2000). Given the obligatory loss of AA (Bowtell et al., 1998; Knapik et al., 1991;
McKenzie et al., 2000; Mazzulla et al., 2017), dietary protein intake (containing all EAA) is
required to replenish exercise-induced losses and support protein remodelling post-exercise (Kato
et al., 2016, 2018a; Millward et al., 1994). A number of recent studies have also shown that the
NEAA may play a role in replenishing AA when suboptimal EAA intakes are available (Kato et
al., 2018a, 2018b). We showed that oxidation of an indicator amino acid (PheOx) tended (P=0.09)
to be greater (large effect size) in the high compared to low intensity exercise condition in the post-
exercise period, which would suggest one or more AA were relatively limiting for whole-body PS
following high intensity exercise.

56
While we did not measure AA oxidation during exercise in the present study, greater
indicator AA oxidation after exercise in the high compared to low intensity condition likely reflects
the greater CHO utilization (~60%) and CHO depletion (~27% or ~57g CHO) and consequently
elevated rate of AA oxidation during high intensity exercise. (Howarth et al., 2010; Lamont et al.,
1999; Lemon & Mullin, 1980; Millward et al., 1994; Phillips, 1993; Shimomura et al., 2004;
Tarnopolsky, 2004; Van Hall et al., 1996). Elevations in AA oxidation that occurred during
exercise may limit the availability of AA in the post-exercise period to support whole-body PS.
Thus, elevated post-exercise protein requirements are needed to replenish exercise-induced AA
losses (Kato et al., 2016, 2018a; Tarnopolsky, 2004). In line with this, Gillen et al. (2019) showed
that endurance athletes had an ~15% greater post-exercise protein requirement following exercise
performed with low- as compared to high-CHO availability. Moreover, Kato et al. (2018a) showed
that the BCAA, which incidentally may be preferentially oxidized (especially leucine) (Koopman
et al., 2004) during exercise, may be the primary ‘rate-limiting’ AA during recovery from
endurance exercise and largely contributing to the elevated protein requirement in endurance
athlete population. In contrast, El-Khoury et al. (1997) showed that moderate intensity exercise
(90 min at 46% V̇O2max) increased leucine oxidation during exercise but only to ~4-7% of daily
leucine oxidation, which did not have a detectable effect on 24-h leucine homeostasis. Similarly,
Devlin et al. (1990) showed that the elevation in exercise-induced AA degradation and AA
oxidation and the suppression in NOLD are transient and do not extend into the post-exercise
period. Interestingly, they showed that the magnitude of decrease in leucine oxidation after
exercise compared to resting state values was correlated with maximal aerobic capacity. Thus, in
well trained individuals, a single bout of endurance exercise is suggested to have less of an effect

57
on altering AA metabolic pathways on the post-exercise period (Devlin et al., 1990; El-Khoury et
al., 1997; Rennie et al., 1981).
In an unpublished study by Gillen (2017), using the same methodology and study approach
as the present study, PheOx during recovery was not different between a high intensity and low
intensity 10-km run with only a small effect size in favour of greater PheOx in the high intensity
condition (d=0.2). It is possible this difference in indicator AA oxidation (PheOx) between the
present study and Gillen (2017), was a result of the duration of the exercise stimulus. For example,
there was greater total CHO oxidation and likely endogenous CHO depletion during the present
20-km as compared to the previous 10-km run, which presumably would also have led to greater
AA oxidation during exercise (Gonzalez et al., 2016; Hawley et al., 1997; Murray & Rosenbloom,
2018; Romijn et al., 1993; Van Loon et al., 2001). One can also estimate total leucine/protein
oxidation from previously published research assuming a constant rate of leucine oxidation during
high (70% V̇O2peak; ~11mg∙ kg-1∙h-1) Mazzulla et al. (2017) and low (50-60% V̇O2peak; ~ 6.5 mg∙
kg-1∙h-1) (Bowtell et al., 1998; Lamont et al., 2001) intensity exercise and a leucine content of
muscle protein of ~9% (Burd et al., 2013). For example, in a 70kg individual, a 10-km run at a
high intensity (completion time ~45 min) would result in ~0.58g of leucine (~6g of protein)
oxidized while during a low intensity condition (completion time ~60 min) ~0.45g of leucine (~5g
of protein) would be oxidized. If exercise distance is doubled (20-km), a high intensity run
(completion time ~ 85 min) would result in ~1.1g of leucine (~12g of protein) oxidized while a
low intensity run (completion time ~108 min) would result in ~0.8g of leucine (~9g of protein)
oxidized. While these estimates do not appear to account for the differences in post-exercise
PheOX between studies, total protein oxidation may be underestimated in the present study if AA
oxidation was increased during latter parts of the 20-km exercise as muscle glycogen became more

58
limiting. Thus, it is possible that by extending the durations of exercise (completion time ~90 min
or longer), the effect of exercise intensity on post-exercise protein requirements becomes more
pronounced due to critically lower quantities of muscle glycogen (~25mmol/kg wet mass) (Areta
& Hopkins, 2018; Hawley et al., 1997) and consequently, greater AA oxidative loss.
Whole-body protein synthesis (PS)
Whole-body PS has previously been shown by some (Bowtell et al., 1998, 2000; Devlin et
al., 1990; Mazzulla et al., 2017b; Rennie et al., 1981) but not all (Koopman et al., 2004; Phillips
et al., 1993) to be suppressed during endurance exercise. The reduction in whole-body PS that
occurs during exercise could be related to it being an energy consuming process that is down-
regulated during conditions of energy stress (e.g. exercise). This reduction in PS during exercise
increases the availability of AA to be used as a respiratory substrate through AA oxidation, which
has been shown the be elevated during endurance exercise by 2-3 fold (Bowtell et al., 1998, 2000;
Knapik et al., 1991; Mazzulla et al., 2017; Phillips et al., 1993; Rennie et al., 1981; Van Hall et
al., 1996). Early in the post-exercise (Mazzulla et al., 2017) period, rates of whole-body PS have
been shown to return to values near or above pre-exercise baseline values in both the fed (Rennie
et al., 1981) and fasted state (Bowtell et al., 1998; Devlin et al., 1990).
In the present study, whole-body PS was determined as the algebraic difference between
phenylalanine flux (PheRa) and PheOx. Whole-body PS was not statistically different between the
high and low intensity exercise conditions in the post-exercise period but there was a moderate
effect size (d=0.72). The present study revealed that exercise intensity influences the post-exercise
stimulation of whole-body PS with greater stimulation from a high compared to low intensity
conditions. Similar results were seen by Gillen (2017), who showed that rates of whole-body PS
trended to be greater following a high compared to low intensity 10-km run and there was a large

59
effect size (d=0.88). To our knowledge, few other studies have examined the effect of endurance
exercise intensity on whole-body PS in the post-exercise period.
At the skeletal muscle level, endurance exercise has been shown to elevate rates of MPS (i.e.
mixed-muscle, myofibrillar and mitochondrial) (Carraro et al., 1990; Di Donato et al., 2014;
Harber et al., 2010; Mascher et al., 2011; Sheffield-Moore et al., 2004; Wilkinson et al., 2008).
The MPS response to endurance exercise is intensity dependent in nature. Di Donato et al. (2014)
found that high compare to low intensity endurance exercise resulted in a more sustained elevation
in both myofibrillar and mitochondrial PS post-exercise (Di Donato et al., 2014). Moreover, the
myofibrillar subfraction has been shown to be nutritionally sensitive following acute endurance
exercise exhibiting a dose-response relationship this is maximally simulated at a 30 g protein
intake (Churchward-Venne et al., 2020). Interestingly, weight bearing running endurance exercise
as done in the present study was shown to be more damaging and and, therefore, augment the
remodelling and repair of skeletal muscle proteins in the post-exercise period compared to cycling
exercise (Kato et al., 2016; Koller et al., 1998). These results support an effect of exercise intensity
which can also be influenced by the exercise modality.
Whole-body protein breakdown (PB)
It has previously been suggested by some that endurance exercise has little effect on whole-
body PB during exercise or in the post-exercise recovery period (Bowtell et al., 1998; Devlin et
al., 1990; Knapik et al., 1991). In contrast, others have suggested that whole-body PB is elevated
during and after a bout of prolonged endurance exercise (Lemon, 1991; Rennie et al., 1981). At
the level of the muscle, Rennie et al. (1981) also showed that MPB, measured by 3-methylhistidine
production, decreased during exercise but returned to pre-exercise rates in the post-exercise

60
recovery period which is in contrast to Carraro et al., (1990) and Harber et al., (2010) who showed
elevated rates of MPB following endurance exercise.
In the present study, whole-body PB, which is the algebraic difference between
phenylalanine flux (PheRa) and the rate of exogenous phenylalanine intake (15.38 µmol∙ kg-1∙d-1),
had a moderate effect size (d=0.78) in favour of the high intensity condition despite differences
not reaching statistical significance threshold. These results were in line with the large effect size
(d=0.80) in favour of greater whole-body PB during a high compared to low intensity 10-km run
seen by Gillen (2017). It has previously been shown by Di Donato et al., (2014) that the intensity
of exercise influences MPS. However, endurance exercise is generally not associated with
hypertrophy (Wilkinson et al., 2008) which suggests any intensity-dependent stimulation of PS
would be counterbalanced by an reciprocal increase in MPB. Thus, there appears to be a
relationship between endurance exercise intensity and AA degradation in the post-exercise period
that could have influenced whole-body results from the present study.
In contrast to the present study, Howarth et al. (2010) showed that phenylalanine Ra
returned to near baseline values in the post-exercise period and there were no differences in
phenylalanine Ra between a low-CHO and high-CHO exercise condition (Howarth et al., 2010).
Differences in post-exercise phenylalanine Ra between the present study and the study by Howarth
et al. (2010) may have been a result of the different exercise stimuli, the present study arguably
having a more intense and whole-body exercise stimulus (Koller et al., 1998). Mazzulla et al.
(2017) and others (Hawley et al., 2006; Koopman et al., 2004) have shown that ingestion of a
mixed meal containing protein and CHO after exercise causes an insulin-induced attenuation in
the rate of proteolysis (Castellino et al., 1987; Flakoll et al., 1989). In the present study, we

61
provided post-exercise CHO (2.4 g∙ kg-1) and protein (0.93 g∙ kg-1∙d-1 or 0.11 g∙ kg-1∙h-1) which
may have exerted a suppressing effect on whole-body PB.
Net Protein Balance (NB)
While endurance exercise was shown to stimulate whole-body PS in an intensity dependant
manor, elevated AA oxidation during exercise ostensibly limits the availability of AA, particularly
EAA, to support PS in the post-exercise period (Bowtell et al., 2000; Hagg et al., 1982; Harber et
al., 2010). In light of the elevated AA loss, whole-body PB increases to mobilizes AA to support
PS. Given the elevations in whole-body PB, NB which is the algebraic difference between whole-
body PS and PB is constrained following high intensity exercise. In the present study, we provided
a single protein intake that has previously been shown to be suboptimal to maximize whole-body
PS and whole-body NB in male endurance athletes (Kato et al., 2016; Mazzulla et al., 2018).
Although NB was positive in both conditions it tended to be less positive in the high intensity
compared to low intensity conditions and there was a large effect size (d=-0.84). Rennie et al.
(1981) showed that following endurance exercise (3.5 hours at 50% V̇O2max) in the fed state, there
is a rise in both whole-body PS and PB but with change in the former being greater, which led to
a positive NB (Rennie et al., 1981). In line with this, Mazzulla et al. (2017) showed that protein
ingestion enhanced NB in both a resting condition and following endurance exercise that was in
part a result of an insulin-induced suppression in PB. They showed that whole-body NB was
positive in the postprandial period in the resting and exercise conditions; however, when
accounting for the exercise-induced oxidative losses, NB was negative following endurance
exercise. In line with these results, in the present study, the less positive whole-body NB in the
high intensity exercise condition likely reflects an elevated rate of whole-body protein breakdown

62
and elevated exercise-induced AA oxidative loss (Millward et al., 1994; Rennie et al., 1981) that
translated into amino acid limitations that constrain whole-body PS.
In the present study, we provided post-exercise CHO (2.4 g∙ kg-1) and protein (0.93 g∙ kg-1∙d-1
or 0.11 g∙ kg-1∙h-1). The dietary protein intake provided in the present study was suboptimal for
this population of male endurance athletes (Kato et al., 2016); however, we assume the between
condition (HIGH vs LOW) difference in protein kinetic outcomes would be consistent over all
suboptimal intakes. This intake presumably supported a positive NB following both high and low
intensity exercise and served to replenish exercise-induced AA loss and supply the substrates to
build new body proteins (Kim et al., 2015; Mazzulla et al., 2017). Kim et al. (2015) showed that
the anabolic response to a meal containing protein is not limited to the maximal stimulation of
protein synthesis as there is simultaneous protein feeding-induced reduction in whole-body PB
that supports a more positive NB (Beelen et al., 2010). Further, post-exercise CHO feeding causes
an insulin-induced suppression of whole-body PB and supports a more positive NB (Beelen et al.,
2010; Biolo et al., 1999; Dangin et al., 2001; Flakoll et al., 1989; Koopman et al., 2004;
Levenhagen et al., 2001, 2002; Mariotti et al., 2000; Mazzulla et al., 2017). Thus, post-exercise
CHO and protein feeding in the present study may have attenuated whole-body PB and increased
post-exercise whole-body PS contributing to a positive NB following exercise (Børsheim et al.,
2004; Miller et al., 2003).
5.4 Estimating Protein Requirements
The IAAO method measures protein metabolism at the whole-body level which provides
limited insight into the exercise induced changes of specific tissues. However, whole-body
measures may more accurately capture the effects of endurance exercise as it exerts a more
systemic effect compared to resistance exercise which is inherently anabolic and exerts a ‘muscle

63
centric’ effect (Moore, 2020). The traditional IAAO method uses mixed model bi-phasic analysis
of graded protein intakes to generate a breakpoint which is representative of the estimated average
protein requirement (EAR) for the population of interest (Elango et al., 2008). We have recently
explored a modified 3-point IAAO method in which two suboptimal and one optimal protein intake
are given. This allows for the development of a curve and determination of the breakpoint (EAR).
With the two suboptimal doses, the equation (slope) of the line is determined for each participant.
Given our knowledge of the bi-phasic nature of the standard curve for IAAO studies, we believe
phenylalanine oxidation (y-value) of the excess dose will be the same as the y-value at the
breakpoint. Using this y-value and the slope of the line, we can then solve for the unknown x-value
at breakpoint (protein intake) for each participant. The average x-value at the breakpoint is
representative of the protein requirement (EAR) for this population. This modified 3-point IAAO
method has been used in the re-analysis of data from the study by Kato et al. (2016) which revealed
an EAR of 1.59 g∙ kg-1∙d-1 which was similar to the originally determined breakpoint (EAR) of
1.65 g∙ kg-1∙d-1 in male endurance athletes with the traditional IAAO model.
In the present study, we used a single suboptimal protein intake and measured oxidation of
the indicator AA at that single intake (0.93 g∙ kg-1∙d-1) between the two exercise conditions (HIGH
vs. LOW). This approach has used in a number of studies in endurance athletes in the past (Gillen,
2017; Gillen et al., 2019). One important assumption of this method is that the slope relationship
of the experimental conditions (e.g. HIGH vs LOW) is consistent and that differences in
requirement would be reflected as a right or leftward shift on the graph (Figure 8, graph A) without
deviations up or downward (Figure 8, graph B) or in the slope of the lines before the estimated
breakpoint (Figure 8, graph C). Assuming that our data fits a similar relationship to that of Kato
et al. (2016), we estimate an ~21% difference in protein requirement to maximize whole-body

64
protein synthesis between high and low intensity exercise. Kato et al. (2016) had participants
perform a 20-km run at a self-selected pace which elicited an exercise (~70% V̇O2peak) which
aligned closely with the exercise intensity in the high intensity condition of the present study. Kato
et al. (2016) estimated a dietary protein requirement (EAR) of 1.65 g∙ kg-1∙d-1 and recommendation
of 1.83 g∙ kg-1∙d-1 which would likely also reflect the estimate for the high intensity condition in
the present study. Based on the ~21% lower dietary protein requirement following low intensity
exercise seen in the present study, a dietary protein requirement (EAR) of 1.32 g∙ kg-1∙d-1 and
recommendation of 1.47 g∙ kg-1∙d-1 would be needed to maximize whole-body protein synthesis
following low intensity endurance exercise. In line with this reduced dietary protein requirement
following low intensity exercise, El-Khoury et al. (1997) showed that low intensity endurance
exercise (90 min at 46% V̇O2max) did not cause significant deterioration in leucine homeostasis at
a protein intake of only 1 g∙ kg-1∙d-1. Taken together, results indicate that lower intensity endurance
exercise has less of an effect on dietary protein requirements.
Adequate dietary protein is needed to support a positive NB which has previously been
shown to aid in the recovery from physiological stress in endurance athletes (Koopman et al.,
2004) as well as support performance improvements during training (Williamson et al., 2019).
Williamson et al. (2019) showed a dose-response relationship between protein intake and NB with
only a high intake (1.83 g∙ kg-1∙d-1) supporting a positive NB. Interestingly, there was a trend
toward an interaction between performance of a 5-km time trial and dietary protein intake with
high having a moderate effect over low (effect size= 0.56). A number of studies (Burke et al.,
2003; Gillen et al., 2017) examining eating patterns of endurance athletes have shown that athletes’
dietary protein intake tend to be consistent with guidelines. However, the timing and distribution
of dietary protein, especially around the post-exercise period is suboptimal to support recovery.

65
Results from the present study are important for these athletes who are not meeting daily dietary
protein requirements and/or are consuming a protein in suboptimal distributions. Endurance
athletes should consider how their dietary protein needs are influenced by the intensity and
duration of exercise as adequate nutrient intake and recovery ultimately determine exercise
performance.
Figure 8. Assumptions of the IAAO model (single intake method). Differences in requirement are reflected as a right or leftward shift on the graph (A), without deviations up or downward (B) or in the slope of the lines before the estimated breakpoint (C). Blue and grey lines represent two different hypothetical experimental conditions.
5.5 Methodological Limitations
The standard IAAO metabolic trial involves the ingestion of 8 hourly drinks containing
protein (0.11 g∙ kg-1∙h-1). It has been shown that a single bolus of ~20g of protein following
resistance exercise provides a maximal anabolic stimulus in the early (5 h) post-exercise period
(Areta et al., 2013; Moore et al., 2012). Areta et al. (2013) examined how the distribution of
protein feeding during recovery from resistance/strength exercise affects the muscle’s anabolic
response. They showed that a ‘pulse’ feeding style (8 x10g protein every 1.5 h) was inferior to
both an intermediate intake (4 x 20g every 3 h) and a bolus feeding style (2 x 40 g every 6 h) to

66
maximize MPS after exercise. The intermediate feeding style was optimal to maximize MPS after
exercise which is consistent with previous studies (Moore et al., 2009). Thus, the feeding pattern
in the present study, which mimic a pulse style feeding pattern, was likely suboptimal and may
have attenuated the anabolic response at the muscle (Areta et al., 2013) and whole-body level
(Moore et al., 2012). This could have resulted in an underestimation of between condition
differences in phenylalanine kinetic outcomes in the present study. Further, this feeding approach
may not be physiologically relevant given that athletes typically consume a single post-exercise
recovery meal and tend to consume their total daily protein in 3-4 bolus doses that are often skewed
to greater intakes at dinner and lower often suboptimal intakes at breakfast (Burke et al., 2003;
Gillen et al., 2017). Further, in the present study, protein was given in the form of crystalline AA
modelled off the composition of egg protein. This is a high quality protein with respect to its EAA
composition and is rapidly digested, both of these factors supporting anabolism (Moore, 2019).
Thus, the estimates of protein requirements following high intensity and low intensity exercise
derived from the present study may not accurately reflect protein intake needs of low-quality
protein sources with suboptimal AA composition and slower digestion rates (e.g. planted-based
sources) (Tang et al., 2009). As such, when consuming lower-quality, EAA deficient protein
sources, a greater absolute amount is needed to compensate for the suboptimal AA profile of these
proteins (Reidy et al., 2013). Additionally, in the present study, dietary protein was provided in
the form of a beverage however, given that whole foods protein sources have been suggested to
exert a more potent anabolic effect, are more nutrient-dense and represent a more typical source
of protein in athletes’ diets, conducting research with the inclusion of whole foods is warranted
and may strengthen the ecologically valid of future protein requirement studies (Burd et al., 2019;
Moore, 2019).

67
In the present study we did not measure muscle glycogen utilization or AA oxidation during
the 20-km exercise bout. While we can make inferences based on the rate of CHO utilization and
estimated muscle glycogen content upon beginning exercise, a pre-post exercise muscle biopsy
would have provided informative objective measures and valuable insight into the substrate
utilization, rate of glycogen depletion during exercise between the two conditions (Bergström &
Hultman, 1967; Ekblom, 2017; Murray & Rosenbloom, 2018). Further, measuring AA oxidation
during exercise through a stable isotope tracer infusion, similar to Mazzulla et al., (2017), would
have provided valuable insight into whole-body protein turnover during exercise. This would be
important to confirm if exercise-induced AA oxidative loss was a primary driver of the elevated
post-exercise protein needs in this population (Bowtell et al., 2000; Kato et al., 2018a; McKenzie
et al., 2000; Tarnopolsky, 2004).
5.6 Research Implications and Practical Recommendations
In a recent review by Jeukendrup, (2017), periodized nutrition is defined as the strategic
combination of exercise training and nutrition, or nutrition alone with the primary goal of
achieving adaptations that support exercise performance. Given the importance of CHO as a
primary substrate for endurance exercise (Hawley & Hopkins, 1995), endurance athlete frequently
manipulate CHO availability through training with low or high CHO availability to achieve
superior exercise performance (Coggan & Coyle, 1987) and molecular adaptations to exercise
(Jeukendrup, 2014; Psilander et al., 2013).
The present study suggests that dietary protein requirements should also be considered
within periodized nutrition strategies for endurance athletes to meet the changing nutritional needs
of athletes over their training season and optimally support post-exercise recovery. Stellingwerff,
(2012) examined the characteristics of several periodized training approaches for elite male

68
endurance athletes. Total training load was calculated by a training impulse/load calculation
(TRIMP= min x RPE). On average, athletes participated in ~13 training sessions/week. Weekly
TRIMP peaked at week 9 and was the lowest at week 16. Training sessions were classified into 3
zones based on the intensity of exercise; Zone 1-easy to somewhat hard, Zone 2- at lactate
threshold and Zone 3-very hard to maximal intensity. They found that 74% of training was done
in Zone 1, 11% in Zone 2 and 15% in Zone 3 (Stellingwerff, 2012). Based on the training schedules
described by Stellingwerff et al., (2012), athletes should be especially mindful of consuming
adequate post-exercise dietary protein on peak TRIMP weeks (week 9) and following hard to near
maximal intensity training sessions (Zone 3). If we assume that the protein requirement estimate
by Kato et al., (2016) was reflective of the requirement in the high intensity condition in the present
study, there is an ~21% lower estimated intake following low intensity exercise (EAR: 1.32 g∙ kg-
1∙d-1 and recommended intake of 1.47 g∙ kg-1∙d-1). Importantly, Stellingwerff, (2012) showed that
low intensity training (Zone 1) accounts for 74% of total training by athletes which suggests that
dietary protein intakes below the recommendation by Kato et al., (2016) may be sufficient for large
parts of athlete’s training schedules.
Gillen et al., (2017) showed endurance athletes habitually consume ~1.5 g protein∙ kg-1∙d-
1, which is within the recommended range for this population. However, the dietary protein source
and patterns of distribution/timing may be suboptimal to support recovery and remodelling after
exercise. Specifically, endurance athletes were shown to consume approximately 40% of their
protein from EAA deficient plant-based sources that may have lower anabolic properties (Tang,
Moore, Kujbida, Tarnopolsky, & Phillips, 2009). Thus, higher absolute protein intakes may be
needed when consuming low quality EAA deficient sources (Reidy et al., 2013). Moreover,
Gorissen et al., (2018) suggests that combining different plant-based proteins and/or blending

69
animal and plant-based sources can be done to improve dietary protein quality. Further, patterns
of protein intake distribution are skewed towards the dinner meal which accounted for an excessive
dose of ~38g while the intake at breakfast was ~19g. For endurance athletes who have unique
recovery needs and performance goals, particular emphasis on these factors is warranted.
Consuming an evenly spaced distribution of 4-5 intakes of high quality protein would be optimal
to support whole-body anabolism (Areta et al., 2013; Moore et al., 2009).
The present study and previous research from our lab suggest that post-exercise protein
requirements may differ based on a number of constitutional factors including the characteristics
of exercise (e.g. duration and intensity) as well as individuals’ nutrition status, specifically, CHO
availability upon beginning exercise. Previous research by Gillen et al., (2019) suggests that
protein requirements are ~15% greater following exercise in a low-CHO, glycogen depleted state.
We showed performing prolonged bouts of exercise at a high intensity results in an ~21% greater
post-exercise protein requirement comparted to a low-intensity exercise condition. Thus,
performing high-intensity endurance exercise in a low-CHO state may further elevate post-
exercise dietary protein requirements. This is particularly important because low-CHO availability
training is a common strategy frequently utilized by endurance athletes (Burke et al., 2011;
Jeukendrup, 2017). To this point, Stellingwerff, (2012) showed that endurance athletes participate
in low-CHO training bouts ~2.5± 2.3 times/week meaning that over an ~13 session training week,
some athletes may be participating in 36% of training session in a low-CHO availability condition.
Given the previously described relationship between CHO availability during exercise and post-
exercise protein requirements (Gillen et al., 2019), athletes frequently participating in low-CHO
training should be particularly mindful of post-exercise protein ingestion following these sessions.

70
Findings from the present study contribute to the body of knowledge of the combined and
individual effects of nutritional status/CHO availability and exercise intensity and duration on
post-exercise protein requirements in endurance athletes. These findings will inform the
development of nutrition guidelines for endurance athlete populations. Few studies have examined
the implications of dietary protein intake over a period of chronic endurance training on recovery,
injury prevention, exercise adaptations and performance measures. This would be an interesting
and informative area of future direction and aid in the development of comprehensive nutrition
guidelines for endurance athletes. Furthermore, previous research has indicated that sex-
differences in substrate utilization during endurance exercise (Carter et al., 2019; Devries et al.,
2006, 2016; Romijn et al., 2000) and in protein metabolism exist (Phillips et al., 1993; Tarnopolsky
et al., 1990; Tarnopolsky, 2004). Specifically, women use relatively more fat and less CHO as a
respiratory substrate during exercise compared to men (Devries, 2016; Tarnopolsky et al., 1990).
This phenomenon has been attributed to greater concentrations of circulating estrogen in women,
especially during the late-follicular and mid-luteal phase of the female menstrual cycle (Oosthuyse
& Bosch, 2010; Zderic et al., 2017). Based on these findings, the relative difference in substrate
utilization between a high and low intensity exercise bout in women may be less pronounced than
in men. Given these physiological sex-differences, exercise science research in females and
empirical evidence-based nutrition recommendations for female athletes are lacking. Thus, it
would be prudent to include female athletes in future research in this area.
5.7 Summary and Conclusion
It has previously been shown that endurance athletes have unique nutrition needs to
maximize exercise adaptations and support post-exercise recovery (Gillen et al., 2019; Kato et al.,
2016; Mazzulla et al., 2017; Moore et al., 2014; Phillips & Van Loon, 2011; Tarnopolsky, 2004).

71
Throughout an athlete’s training program, the intensity of exercise and total training
duration/volume is modified based on the temporal proximity to major competitions. As such,
nutritional requirements change over an athlete’s training season (from base-training to pre-
competition) (Fiskerstrand & Seiler, 2004).
In the present study, we examined the effect of high intensity, CHO dependant exercise
compared to low intensity exercise on oxidation of an indicator AA which is reflective of post-
exercise protein requirements. The two exercise conditions simulate vastly different types of
training in an athlete’s schedule with the low intensity run simulating a recovery or early base-
season run and the high intensity run simulating a training phase in preparation for major
competition with near maximal effort. We showed a large effect size and found that oxidation of
an indicator amino acid (PheOx) tended to be greater following high intensity compared to low
intensity exercise. This suggested that one or some AA are relatively limiting for whole-body PS
following high intensity endurance exercise. In line with these results, there was a large magnitude
of effect and NB tended to be less positive following high intensity exercise. While protein kinetic
outcomes did not reach statistical significance, the large magnitude of effect size(s) suggest that
we may have been underpowered to detect differences in our outcome variables with the
preliminary data set (n=6) and a type II error occurred. Taken together, these results suggest that
post-exercise protein requirements to maximize whole-body protein synthesis may be greater
following a prolonged high compared to low intensity endurance exercise session.

72
References Abou Sawan, S., Vliet, S. Van, Parel, J.T., Beals, J.W., Mazzulla, M., West, D.W.D., Philp, A., Li, Z.,
Paluska, S.A., Burd, N.A., and Moore, D.R. 2018. Translocation and protein complex co-
localization of mTOR is associated with postprandial myofibrillar protein synthesis at rest and after
endurance exercise. 6: 1–14. doi:10.14814/phy2.13628.
Anne, L.B. 2007. Low Energy Availability in the Marathon and other Endurance Sports. Sport. Med.
37(4–5): 348–352.
Anthony, J.C., Anthony, T.G., Kimball, S.R., and Jefferson, L.S. 2001. Signaling Pathways Involved in
Translational Control of Protein Synthesis in Skeletal Muscle by Leucine. J. Nutr. 131(3): 856S-
860S. doi:10.1093/jn/131.3.856s.
Areta, J.L., Burke, L.M., Ross, M.L., Camera, D.M., West, D.W.D., Broad, E.M., Jeacocke, N.A., Moore,
D.R., Stellingwerff, T., Phillips, S.M., Hawley, J.A., and Coffey, V.G. 2013a. Timing and
distribution of protein ingestion during prolonged recovery from resistance exercise alters
myofibrillar protein synthesis. J. Physiol. 591(9): 2319–2331. doi:10.1113/jphysiol.2012.244897.
Areta, J.L., and Hopkins, W.G. 2018a. Skeletal Muscle Glycogen Content at Rest and During Endurance
Exercise in Humans: A Meta-Analysis. Sport. Med. 48(9): 2091–2102. doi:10.1007/s40279-018-
0941-1.
Areta, J.L., and Hopkins, W.G. 2018b. Skeletal Muscle Glycogen Content at Rest and During Endurance
Exercise in Humans: A Meta-Analysis. Sport. Med. 48(9): 2091–2102. doi:10.1007/s40279-018-
0941-1.
Areta, L., Burke, L.M., Ross, M.L., Camera, D.M., West, D.W.D., Broad, E.M., Jeacocke, N.A., Moore,
D.R., Stellingwerff, T., Phillips, S.M., Hawley, J.A., and Coffey, V.G. 2013b. Timing and
distribution of protein ingestion during prolonged recovery from resistance exercise alters
myofibrillar protein synthesis. 9: 2319–2331. doi:10.1113/jphysiol.2012.244897.
Bandegan, A., Courtney-Martin, G., Rafii, M., Pencharz, P.B., and Lemon, P.W.R. 2019. Indicator amino
acid oxidation protein requirement estimate in endurance-trained men 24 h postexercise exceeds

73
both the EAR and current athlete guidelines. Am. J. Physiol. - Endocrinol. Metab. 316(5): E741–
E748. doi:10.1152/ajpendo.00174.2018.
Barth, E., Albuszies, G., Baumgart, K., Matejovic, M., Wachter, U., Vogt, J., Radermacher, P., and
Calzia, E. 2007. Glucose metabolism and catecholamines. Crit. Care Med. 35(9 SUPPL.).
doi:10.1097/01.CCM.0000278047.06965.20.
Beelen, M., Burke, L.M., Gibala, M.J., and Van Loon, L.J.C. 2010. Nutritional strategies to promote
postexercise recovery. Int. J. Sport Nutr. Exerc. Metab. 20(6): 515–532.
doi:10.1123/ijsnem.20.6.515.
Beelen, M., Zorenc, A., Pennings, B., Senden, J.M., Kuipers, H., and van Loon, L.J.C. 2011. Impact of
protein coingestion on muscle protein synthesis during continuous endurance type exercise. Am. J.
Physiol. - Endocrinol. Metab. 300(6). doi:10.1152/ajpendo.00446.2010.
Berg, J.M., Tymoczko, J.L., and Stryer, L. 2002. The first step in amino acid degradation is the removal
of nitrogen. Biochemistry. Freeman WH.
Bergström, J., and Hultman, E. 1967. A study of the glycogen metabolism during exercise in man. Scand.
J. Clin. Lab. Invest. 19(3): 218–228. doi:10.3109/00365516709090629.
Bijker, K.E., de Groot, G., and Hollander, A.P. 2002. Differences in leg muscle activity during running
and cycling in humans. Eur. J. Appl. Physiol. 87(6): 556–561. doi:10.1007/s00421-002-0663-8.
Biolo, G., Williams, B.D., Fleming, R.Y.D., and Wolfe, R.R. 1999. Insulin action on muscle protein
kinetics and amino acid transport during recovery after resistance exercise. Diabetes 48(5): 949–
957. doi:10.2337/diabetes.48.5.949.
Blomstrand, E., and Saltin, B. 1999. Effect of muscle glycogen on glucose, lactate and amino acid
metabolism during exercise and recovery in human subjects. J. Physiol. 514(1): 293–302.
doi:10.1111/j.1469-7793.1999.293af.x.
Børsheim, E., Cree, M.G., Tipton, K.D., Elliott, T.A., Aarsland, A., and Wolfe, R.R. 2004. Effect of
carbohydrate intake on net muscle protein synthesis during recovery from resistance exercise. J.
Appl. Physiol. 96(2): 674–678. doi:10.1152/japplphysiol.00333.2003.

74
Bowtell, J.L., Leese, G.P., Smith, K., Watt, P.W., Nevill, A., Rooyackers, O., Wagenmakers, A.J.M., and
Rennie, M.J. 1998. Modulation of whole body protein metabolism, during and after exercise, by
variation of dietary protein. J. Appl. Physiol. 85(5): 1744–1752. doi:10.1152/jappl.1998.85.5.1744.
Bowtell, J.L., Leese, G.P., Smith, K., Watt, P.W., Nevill, A., Rooyackers, O., Wagenmakers, A.J.M., and
Rennie, M.J. 2000. Effect of oral glucose on leucine turnover in human subjects at rest and during
exercise at two levels of dietary protein. J. Physiol. 525(1): 271–281. doi:10.1111/j.1469-
7793.2000.00271.x.
Breen, L., Philp, A., Witard, O.C., Jackman, S.R., Selby, A., Smith, K., Baar, K., and Tipton, K.D. 2011.
The influence of carbohydrate-protein co-ingestion following endurance exercise on myofibrillar
and mitochondrial protein synthesis. J. Physiol. 589(16): 4011–4025.
doi:10.1113/jphysiol.2011.211888.
Brooks, G.A. 1998. Mammalian fuel utilization during sustained exercise. 120: 89–107.
Bross, R., Ball, R.O., and Pencharz, P.B. 1998. Development of a minimally invasive protocol for the
determination of phenylalanine and lysine kinetics in humans during the fed state. J. Nutr. 128(11):
1913–1919. doi:10.1093/jn/128.11.1913.
Burd, N.A., Beals, J.W., Martinez, I.G., Salvador, A.F., and Skinner, S.K. 2019. Food-First Approach to
Enhance the Regulation of Post-exercise Skeletal Muscle Protein Synthesis and Remodeling. Sport.
Med. 49(s1): 59–68. Springer International Publishing. doi:10.1007/s40279-018-1009-y.
Burd, N.A., Hamer, H.M., Pennings, B., Pellikaan, W.F., Senden, J.M.G., Gijsen, A.P., and van Loon,
L.J.C. 2013. Substantial Differences between Organ and Muscle Specific Tracer Incorporation Rates
in a Lactating Dairy Cow. PLoS One 8(6). doi:10.1371/journal.pone.0068109.
Burd, N.A., Tang, J.E., Moore, D.R., and Phillips, S.M. 2009. Exercise training and protein metabolism:
influences of contraction, protein intake, and sex-based differences. J. Appl. Physiol. 106(5): 1692–
1701. doi:10.1152/japplphysiol.91351.2008.
Burke, L.M. 2010. Fueling strategies to optimize performance: Training high or training low? Scand. J.
Med. Sci. Sport. 20(SUPPL. 2): 48–58. doi:10.1111/j.1600-0838.2010.01185.x.

75
Burke, L.M., Collier, G.R., Beasley, S.K., Davis, P.G., Fricker, P.A., Heeley, P., Walder, K., and
Hargreaves, M. 1995. Effect of coingestion of fat and protein with carbohydrate feedings on muscle
glycogen storage. J. Appl. Physiol. 78(6): 2187–2192. doi:10.1152/jappl.1995.78.6.2187.
Burke, L.M., Hawley, J.A., Wong, S.H.S., and Jeukendrup, A.E. 2011. Carbohydrates for training and
competition. J. Sports Sci. 29(SUPPL. 1). doi:10.1080/02640414.2011.585473.
Burke, L.M., Kiens, B., and Ivy, J.L. 2004. Carbohydrates and fat for training and recovery. J. Sports Sci.
22(1): 15–30. doi:10.1080/0264041031000140527.
Burke, L.M., Van Loon, L.J.C., and Hawley, J.A. 2017. Postexercise muscle glycogen resynthesis in
humans. J. Appl. Physiol. 122(5): 1055–1067. doi:10.1152/japplphysiol.00860.2016.
Burke, L.M., Slater, G., Broad, E.M., Haukka, J., Modulon, S., and Hopkins, W.G. 2003. Eating Patterns
and Meal Frequency of Elite Australian Athletes. Int. J. Sport Nutr. Exerc. Metab. 13(4): 521–538.
doi:10.1123/ijsnem.13.4.521.
Campbell, P.J., Carlson, M.G., Hill, J.O., and Nurjhan, N. 1992. Regulation of free fatty acid metabolism
by insulin in humans: role of lipolysis and reesterification. Am. J. Physiol. 263(6): E1063-9. United
States. doi:10.1152/ajpendo.2006.263.6.E1063.
Carraro, F., Stuart, C.A., Hartl, W.H., Rosenblatt, J., and Wolfe, R.R. 1990. Effect of exercise and
recovery on muscle protein synthesis in human subjects. Am. J. Physiol. - Endocrinol. Metab. 259(4
22-4): 470–476. doi:10.1152/ajpendo.1990.259.4.e470.
Carter, S.L., Rennie, C., and Tarnopolsky, M.A. 2019. Substrate utilization during endurance exercise in
men and women after endurance training. 5: 898–907.
Castellino, P., Luzi, L., Simonson, D.C., Haymond, M., and DeFronzo, R.A. 1987. Effect of insulin and
plasma amino acid concentrations on leucine metabolism in man. Role of substrate availability on
estimates of whole body protein synthesis. J. Clin. Invest. 80(6): 1784–1793.
doi:10.1172/JCI113272.
Cermak, N.M., and Van Loon, L.J.C. 2013. The use of carbohydrates during exercise as an ergogenic aid.
Sport. Med. 43(11): 1139–1155. doi:10.1007/s40279-013-0079-0.

76
Churchward-Venne, T.A., Pinckaers, P.J.M., Smeets, J.S.J., Betz, M.W., Senden, J.M., Goessens, J.P.B.,
Gijsen, A.P., Rollo, I., Verdijk, L.B., and van Loon, L.J.C. 2020. Dose-response effects of dietary
protein on muscle protein synthesis during recovery from endurance exercise in young men: a
double-blind randomized trial. Am. J. Clin. Nutr.: 1–15. Oxford University Press.
doi:10.1093/ajcn/nqaa073.
Coggan, A.R., and Coyle, E.F. 1987. Reversal of fatigue during prolonged exercise by carbohydrate
infusion or ingestion. J. Appl. Physiol. 63(6): 2388–2395. doi:10.1152/jappl.1987.63.6.2388.
Costill, D.L., Coyle, E., Dalsky, G., Evans, W., Fink, W., and Hoopes, D. 1977. Effects of elevated
plasma FFA and insulin on muscle glycogen usage during exercise. J. Appl. Physiol. Respir.
Environ. Exerc. Physiol. 43(4): 695–699. doi:10.1152/jappl.1977.43.4.695.
Costill, D.L., Sherman, W.M., Fink, W.J., Maresh, C., Witten, M., and Miller, J.M. 1981. The role of
dietary carbohydrates in muscle glycogen resynthesis after strenuous running. Am. J. Clin. Nutr.
34(9): 1831–1836. doi:10.1093/ajcn/34.9.1831.
Coyle, E.F., Coggan, A.R., Hemmert, M.K., and Ivy, J.L. 1986. Muscle glycogen utilization during
prolonged strenous exercise when fed carbohydrate. J. Appl. Physiol. 61(1): 165–172.
doi:10.1152/jappl.1986.61.1.165.
Coyle, E.F., Jeukendrup, A.E., Wagenmakers, A.J.M., and Saris, W.H.M. 1997. Fatty acid oxidation is
directly regulated by carbohydrate metabolism during exercise. Am. J. Physiol. - Endocrinol. Metab.
273(2 36-2). doi:10.1152/ajpendo.1997.273.2.e268.
D’Antona, G., Ragni, M., Cardile, A., Tedesco, L., Dossena, M., Bruttini, F., Caliaro, F., Corsetti, G.,
Bottinelli, R., Carruba, M.O., Valerio, A., and Nisoli, E. 2010. Branched-chain amino acid
supplementation promotes survival and supports cardiac and skeletal muscle mitochondrial
biogenesis in middle-aged mice. Cell Metab. 12(4): 362–372. Elsevier Inc.
doi:10.1016/j.cmet.2010.08.016.
Dangin, M., Boirie, Y., Garcia-Rodenas, C., Gachon, P., Fauquant, J., Callier, P., Ballèvre, O., and
Beaufrère, B. 2001. The digestion rate of protein is an independent regulating factor of postprandial

77
protein retention. Am. J. Physiol. - Endocrinol. Metab. 280(2 43-2).
doi:10.1152/ajpendo.2001.280.2.e340.
Devlin, J.T., Brodsky, I., Scrimgeour, A., Fuller, S., and Bier, D.M. 1990. Amino acid metabolism after
intense exercise. Am. J. Physiol. - Endocrinol. Metab. 258(2 21-2).
doi:10.1152/ajpendo.1990.258.2.e249.
Devries, M.C. 2016. Sex-based differences in endurance exercise muscle metabolism: Impact on exercise
and nutritional strategies to optimize health and performance in women. Exp. Physiol. 101(2): 243–
249. doi:10.1113/EP085369.
Devries, M.C., Hamadeh, M.J., Phillips, S.M., and Tarnopolsky, M.A. 2006. Menstrual cycle phase and
sex influence muscle glycogen utilization and glucose turnover during moderate-intensity endurance
exercise. Am. J. Physiol. Integr. Comp. Physiol. 291(4): R1120–R1128.
doi:10.1152/ajpregu.00700.2005.
Di Donato, D.M., West, D.W.D., Churchward-Venne, T.A., Breen, L., Baker, S.K., and Phillips, S.M.
2014. Influence of aerobic exercise intensity on myofibrillar and mitochondrial protein synthesis in
young men during early and late postexercise recovery. Am. J. Physiol. - Endocrinol. Metab. 306(9):
1025–1032. doi:10.1152/ajpendo.00487.2013.
Dyck, D.J., Putman, C.T., Heigenhauser, G.J.F., Hultman, E., and Spriet, L.L. 1993. Regulation of fat-
carbohydrate interaction in skeletal muscle during intense aerobic cycling. Am. J. Physiol. -
Endocrinol. Metab. 265(6 28-6). doi:10.1152/ajpendo.1993.265.6.e852.
Ekblom, B. 2017. The muscle biopsy technique. Historical and methodological considerations. Scand. J.
Med. Sci. Sport. 27(5): 458–461. doi:10.1111/sms.12808.
El-Khoury, A.E., Forslund, A., Olsson, R., Branth, S., Sjödin, A., Andersson, A., Atkinson, A., Selvaraj,
A., Hambraeus, L., and Young, V.R. 1997. Moderate exercise at energy balance does not affect 24-h
leucine oxidation or nitrogen retention in healthy men. Am. J. Physiol. - Endocrinol. Metab. 273(2
36-2). doi:10.1152/ajpendo.1997.273.2.e394.
Elango, R., Ball, R.O., and Pencharz, P.B. 2008. Indicator Amino Acid Oxidation : Concept. : 243–246.

78
Eriksson, L.S., Broberg, S., Björkman, O., and Wahren, J. 1985. Ammonia metabolism during exercise in
man. Clin. Physiol. 5(4): 325–336. doi:10.1111/j.1475-097X.1985.tb00753.x.
Febbraio, M.A., Chiu, A., Angus, D.J., Arkinstall, M.J., and Hawley, J.A. 2000. Effects of carbohydrate
ingestion before and during exercise on glucose kinetics and performance. J. Appl. Physiol. 89(6):
2220–2226. doi:10.1152/jappl.2000.89.6.2220.
Ferguson-Stegall, L., McCleave, E., Ding, Z., Doerner, P.G., Liu, Y., Wang, B., Healy, M., Kleinert, M.,
Dessard, B., Lassiter, D.G., Kammer, L., and Ivy, J.L. 2011. Aerobic exercise training adaptations
are increased by postexercise carbohydrate-protein supplementation. J. Nutr. Metab. 2011.
doi:10.1155/2011/623182.
Fiskerstrand, Å., and Seiler, K.S. 2004. Training and performance characteristics among Norwegian
International Rowers 1970-2001. Scand. J. Med. Sci. Sport. 14(5): 303–310. doi:10.1111/j.1600-
0838.2003.00370.x.
Flakoll, P.J., Brown, L., Hill, J., Abumrad, N., Paul, J., Kulaylat, M., Frexes-, M., Hourani, H., and
Brown, L.L. 1989. Amino acids augment insulin ’s of whole body proteolysis. Am. J. Physiol. -
Endocrinol. Metab. 89.
Forslund, A.H., El-Khoury, A.E., Olsson, R.M., Sjödin, A.M., Hambraeus, L., and Young, V.R. 1999.
Effect of protein intake and physical activity on 24-h pattern and rate of macronutrient utilization.
Am. J. Physiol. - Endocrinol. Metab. 276(5 39-5). doi:10.1152/ajpendo.1999.276.5.e964.
Gelfand, R.A., and Barrett, E.J. 1987. Effect of physiologic hyperinsulinemia on skeletal muscle protein
synthesis and breakdown in man. J. Clin. Invest. 80(1): 1–6. doi:10.1172/JCI113033.
Gibala, M.J., Young, M.E., and Taegtmeyer, H. 2000. Anaplerosis of the citric acid cycle: Role in energy
metabolism of heart and skeletal muscle. Acta Physiol. Scand. 168(4): 657–665. doi:10.1046/j.1365-
201X.2000.00717.x.
Gillen, J.B., Trommelen, J., Wardenaar, F.C., Brinkmans, N.Y.J., Versteegen, J.J., Jonvik, K.L., Kapp,
C., Vries, J. De, Borne, J.J.G.C. Van Den, Gibala, M.J., and Loon, L.J.C. Van. 2017. Dietary Protein
Intake and Distribution Patterns of Well-Trained Dutch Athletes. : 105–114.

79
Gillen, J.B., West, D.W., Williamson, E.P., Fung, H.J.W., and Moore, D.R. 2019a. Low-Carbohydrate
Training Increases Protein Requirements of Endurance Athletes. Med. Sci. Sport. Exerc. (May): 1.
doi:10.1249/mss.0000000000002036.
Gillen, J.B., West, D.W.D., Williamson, E.P., Fung, H.J.W., and Moore, D.R. 2019b. Low-Carbohydrate
Training Increases Protein Requirements of Endurance Athletes. Med. Sci. Sports Exerc. 51(11):
2294–2301. doi:10.1249/MSS.0000000000002036.
Gollnick, P.D., Pernow, B., Essen, B., Jansson, E., and Saltin, B. 1981. Availability of glycogen and
plasma FFA for substrate utilization in leg muscle of man during exercise. Clin. Physiol. 1(1): 27–
42. doi:10.1111/j.1475-097X.1981.tb00872.x.
Gonzalez, J.T., Fuchs, C.J., Betts, J.A., and van Loon, L.J.C. 2016. Liver glycogen metabolism during
and after prolonged endurance-type exercise. Am. J. Physiol. - Endocrinol. Metab. 311(3): E543–
E553. doi:10.1152/ajpendo.00232.2016.
Gorissen, S.H.M., Crombag, J.J.R., Senden, J.M.G., Waterval, W.A.H., and Bierau, J. 2018. Protein
content and amino acid composition of commercially available plant ‑ based protein isolates. Amino
Acids 50(12): 1685–1695. Springer Vienna. doi:10.1007/s00726-018-2640-5.
Hagg, S.A., Morse, E.L., and Adibi, S.A. 1982. Effect of exercise on rates of oxidation, turnover, and
plasma clearance of leucine in human subjects. Am. J. Physiol. - Endocrinol. Metab. 5(6): 407–410.
doi:10.1152/ajpendo.1982.242.6.e407.
Van Hall, G., MacLean, D.A., Saltin, B., and Wagenmakers, A.J.M. 1996. Mechanisms of activation of
muscle branched-chain α-keto acid dehydrogenase during exercise in man. J. Physiol. 494(3): 899–
905. doi:10.1113/jphysiol.1996.sp021542.
Hansen, A.K., Fischer, C.P., Plomgaard, P., Andersen, J.L., Saltin, B., and Pedersen, B.K. 2005. Skeletal
muscle adaptation: training twice every second day vs. training once daily. J. Appl. Physiol. 98(1):
93–99. United States. doi:10.1152/japplphysiol.00163.2004.
Haralambie, G., and Berg, A. 1976. Serum urea and amino nitrogen changes with exercise duration. Eur.
J. Appl. Physiol. Occup. Physiol. 36(1): 39–48. doi:10.1007/BF00421632.

80
Harber, M.P., Konopka, A.R., Jemiolo, B., Trappe, S.W., Trappe, T.A., and Reidy, P.T. 2010. Muscle
protein synthesis and gene expression during recovery from aerobic exercise in the fasted and fed
states. Am. J. Physiol. - Regul. Integr. Comp. Physiol. 299(5): 1254–1262.
doi:10.1152/ajpregu.00348.2010.
Hargreaves, M. 2000. Proceedings of the Australian Physiological and Pharmacological Society
Symposium Muscle : From Metabolism to Molecules SKELETAL MUSCLE METABOLISM
DURING EXERCISE IN HUMANS. Clin. Exp. Pharmachology Physiol. 27(April 1999): 225–228.
Hargreaves, M. 2015. Exercise, muscle, and CHO metabolism. Scand. J. Med. Sci. Sport. 25: 29–33.
doi:10.1111/sms.12607.
Hawley, J.A., and Hopkins, W.G. 1995. Aerobic Glycolytic and Aerobic Lipolytic Power Systems: A
New Paradigm with Implications for Endurance and Ultraendurance Events. doi:10.2165/00007256-
199519040-00002.
Hawley, J.A., and Morton, J.P. 2014. Ramping up the signal: Promoting endurance training adaptation in
skeletal muscle by nutritional manipulation. Clin. Exp. Pharmacol. Physiol. 41(8): 608–613.
doi:10.1111/1440-1681.12246.
Hawley, J.A., Schabort, E.J., Noakes, T.D., and Dennis, S.C. 1997. Carbohydrate-loading and exercise
performance. Sport. Med. 24(2): 73–81. doi:10.2165/00007256-199724020-00001.
Hawley, J.A., Tipton, K.D., and Millard-Stafford, M.L. 2006. Promoting training adaptations through
nutritional interventions. J. Sports Sci. 24(7): 709–721. doi:10.1080/02640410500482727.
Heikura, I.A., Stellingwerff, T., and Burke, L.M. 2018. Self-Reported Periodization of Nutrition in Elite
Female and Male Runners and Race Walkers. Front. Physiol. 9(December): 1–16.
doi:10.3389/fphys.2018.01732.
Hermansen, L., Hultman, E., and Saltin, B. 1967. Muscle Glycogen during Prolonged Severe Exercise.
Acta Physiol. Scand. 71(2–3): 129–139. doi:10.1111/j.1748-1716.1967.tb03719.x.
Holloszy, J.O., and Coyle, E.F. 2016. Adaptations of skeletal muscle to endurance exercise and their
metabolic consequences. J. Appl. Physiol. 121(6): 831–838.

81
Horowitz, J.F., Mora-Rodriguez, R., Byerley, L.O., and Coyle, E.F. 1997. Lipolytic suppression
following carbohydrate ingestion limits fat oxidation during exercise. Am. J. Physiol. - Endocrinol.
Metab. 273(4 36-4): 768–775. doi:10.1152/ajpendo.1997.273.4.e768.
Howarth, K.R., Moreau, N.A., Phillips, S.M., and Gibala, M.J. 2009. Coingestion of protein with
carbohydrate during recovery from endurance exercise stimulates skeletal muscle protein synthesis
in humans. J. Appl. Physiol. 106(4): 1394–1402. doi:10.1152/japplphysiol.90333.2008.
Howarth, K.R., Phillips, S.M., MacDonald, M.J., Richards, D., Moreau, N.A., and Gibala, M.J. 2010.
Effect of glycogen availability on human skeletal muscle protein turnover during exercise and
recovery. J. Appl. Physiol. 109(2): 431–438. doi:10.1152/japplphysiol.00108.2009.
Hulston, C.J., Venables, M.C., Mann, C.H., Martin, C., Philp, A., Baar, K., and Jeukendrup, A.E. 2010.
Training with low muscle glycogen enhances fat metabolism in well-trained cyclists. Med. Sci.
Sports Exerc. 42(11): 2046–2055. doi:10.1249/MSS.0b013e3181dd5070.
Humayun, M.A., Elango, R., Ball, R.O., and Pencharz, P.B. 2007. Reevaluation of the protein
requirement in young men with the indicator amino acid oxidation technique 1 Ϫ 3. : 995–1002.
Jeukendrup, A.E. 2011. Nutrition for endurance sports: Marathon, triathlon, and road cycling. J. Sports
Sci. 29(SUPPL. 1). doi:10.1080/02640414.2011.610348.
Jeukendrup, A.E. 2017. Periodized Nutrition for Athletes. Sport. Med. 47(s1): 51–63. Springer
International Publishing. doi:10.1007/s40279-017-0694-2.
Jeukendrup, A.E., and Killer, S.C. 2011. The myths surrounding pre-exercise carbohydrate feeding. Ann.
Nutr. Metab. 57(SUPPL. 2): 18–25. doi:10.1159/000322698.
Jeukendrup, A.E., Wagenmakers, A.J.M., Stegen, J.H.C.H., Gijsen, A.P., Brouns, F., and Saris, W.H.M.
1999. Carbohydrate ingestion can completely suppress endogenous glucose production during
exercise. Am. J. Physiol. - Endocrinol. Metab. 276(4 39-4). doi:10.1152/ajpendo.1999.276.4.e672.
Jungas, R.L., Halperin, M.L., and Brosnan, J.T. 1992. Quantitative analysis of amino acid oxidation and
related gluconeogenesis in humans. Physiol. Rev. 72(2): 419–448.
doi:10.1152/physrev.1992.72.2.419.

82
Karlsson, J., Nordesjö, L. ‐O, and Saltin, B. 1974. Muscle Glycogen Utilization during Exercise after
Physical Training. Acta Physiol. Scand. 90(1): 210–217. doi:10.1111/j.1748-1716.1974.tb05579.x.
Kasperek, G.J., and Snider, R.D. 1987a. Effect of exercise intensity and starvation on activation of
branched-chain keto acid dehydrogenase by exercise. Am. J. Physiol. - Endocrinol. Metab. 252(1):
33–37. doi:10.1152/ajpendo.1987.252.1.e33.
Kasperek, G.J., and Snider, R.D. 1987b. Effect of exercise intensity and starvation on activation of
branched-chain keto acid dehydrogenase by exercise. Am. J. Physiol. - Endocrinol. Metab. 252(1):
33–37.
Kato, H., Suzuki, K., Bannai, M., and Moore, D.R. 2016a. Protein requirements are elevated in endurance
athletes after exercise as determined by the indicator amino acid oxidation method. PLoS One 11(6):
1–15. doi:10.1371/journal.pone.0157406.
Kato, H., Suzuki, K., Bannai, M., and Moore, D.R. 2016b. Protein requirements are elevated in endurance
athletes after exercise as determined by the indicator amino acid oxidation method. PLoS One 11(6):
1–15. doi:10.1371/journal.pone.0157406.
Kato, H., Suzuki, K., Bannai, M., and Moore, D.R. 2018a. Branched-chain amino acids are the primary
limiting amino acids in the diets of endurance-trained men after a bout of prolonged exercise. J.
Nutr. 148(6): 925–931. doi:10.1093/jn/nxy048.
Kato, H., Suzuki, K., Bannai, M., and Moore, D.R. 2018b. Branched-chain amino acids are the primary
limiting amino acids in the diets of endurance-trained men after a bout of prolonged exercise. J.
Nutr. 148(6): 925–931. doi:10.1093/jn/nxy048.
Kato, H., Volterman, K.A., West, D.W.D., Suzuki, K., and Moore, D.R. 2018c. Nutritionally non-
essential amino acids are dispensable for whole-body protein synthesis after exercise in endurance
athletes with an adequate essential amino acid intake. Amino Acids 50(12): 1679–1684. Springer
Vienna. doi:10.1007/s00726-018-2639-y.
Kiens, B., Roemen, T.H.M., and Van Der Vusse, G.J. 1999. Muscular long-chain fatty acid content
during graded exercise in humans. Am. J. Physiol. - Endocrinol. Metab. 276(2 39-2): 352–357.

83
doi:10.1152/ajpendo.1999.276.2.e352.
Kim, I.Y., Schutzler, S., Schrader, A., Spencer, H.J., Azhar, G., Ferrando, A.A., and Wolfe, R.R. 2015.
The anabolic response to a meal containing different amounts of protein is not limited by the
maximal stimulation of protein synthesis in healthy young adults. Am. J. Physiol. - Endocrinol.
Metab. 310(1): E73–E80. doi:10.1152/ajpendo.00365.2015.
Kimball, S.R., and Jefferson, L.S. 2006. Branched-Chain Amino Acids : Metabolism , Physiological
Function , and Application Signaling Pathways and Molecular Mechanisms through which
Branched-Chain Amino Acids Mediate Translational Control of. (6): 227–231.
Klein, S., Coyle, E.F., and Wolfe, R.R. 1994. Fat metabolism during low-intensity exercise in endurance-
trained and untrained men. Am. J. Physiol. 267(6 Pt 1): E934-40. United States.
doi:10.1152/ajpendo.1994.267.6.E934.
Knapik, J., Meredith, C., Jones, B., Fielding, R., Young, V., and Evans, W. 1991. Leucine metabolism
during fasting and exercise. J. Appl. Physiol. 70(1): 43–47. doi:10.1152/jappl.1991.70.1.43.
Koller, A., Mair, J., Schobersberger, W., Wohlfarter, T., Haid, C., Mayr, M., Villiger, B., Frey, W., and
Puschendorf, B. 1998. Effects of prolonged strenuous endurance exercise on plasma myosin heavy
chain fragments and other muscular proteins. Cycling vs running. J. Sports Med. Phys. Fitness
38(1): 10–17. Italy.
Koopman, R., Pannemans, D.L.E., Jeukendrup, A.E., Gijsen, A.P., Senden, J.M.G., Halliday, D., Saris,
W.H.M., Van Loon, L.J.C., and Wagenmakers, A.J.M. 2004. Combined ingestion of protein and
carbohydrate improves protein balance during ultra-endurance exercise. Am. J. Physiol. -
Endocrinol. Metab. 287(4 50-4): 712–720. doi:10.1152/ajpendo.00543.2003.
Lamont, L.S., McCullough, A.J., and Kalhan, S.C. 1999. Comparison of leucine kinetics in endurance-
trained and sedentary humans. J. Appl. Physiol. 86(1): 320–325. doi:10.1152/jappl.1999.86.1.320.
Lamont, L.S., McCullough, A.J., and Kalhan, S.C. 2001. Relationship between leucine oxidation and
oxygen consumption during steady-state exercise. Med. Sci. Sports Exerc. 33(2): 237–241.
doi:10.1097/00005768-200102000-00011.

84
Lemon, P.W., and Mullin, J.P. 1980. Effect of initial muscle glycogen levels on protein catabolism during
exercise. J. Appl. Physiol. 48(4): 624–629. United States. doi:10.1152/jappl.1980.48.4.624.
Lemon, P.W., and Nagle, F.J. 1981. Effects of exercise on protein and amino acid metabolism. Med. Sci.
Sports Exerc. 13(3): 141—149. Available from http://europepmc.org/abstract/MED/6789029.
Lemon, P.W.R. 1991. Effect of exercise on protein requirements. J. Sports Sci. 9: 53–70.
doi:10.1080/02640419108729866.
Lemon, P.W.R., Nagle, F.J., Mullin, J.P., and Benevenga, N.J. 1982. In vivo leucine oxidation at rest and
during two intensities of exercise. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 53(4): 947–954.
doi:10.1152/jappl.1982.53.4.947.
Levenhagen, D.K., Carr, C., Carlson, M.G., Maron, D.J., Borel, M.J., and Flakoll, P.J. 2002. Postexercise
protein intake enhances whole-body and leg protein accretion in humans. Med. Sci. Sports Exerc.
34(5): 828–837. doi:10.1097/00005768-200205000-00016.
Levenhagen, D.K., Gresham, J.D., Carlson, M.G., Maron, D.J., Borel, M.J., and Flakoll, P.J. 2001.
Postexercise nutrient intake timing in humans is critical to recovery of leg glucose and protein
homeostasis. Am. J. Physiol. - Endocrinol. Metab. 280(6 43-6): 982–993.
doi:10.1152/ajpendo.2001.280.6.e982.
Loftin, M., Sothern, M., Warren, B., and Udall, J. 2004. Comparison of VO2 peak during treadmill and
cycle ergometry in severely overweight youth. J. Sport. Sci. Med. 3(4): 254–260.
Van Loon, L.J.C., Greenhaff, P.L., Constantin-Teodosiu, D., Saris, W.H.M., and Wagenmakers, A.J.M.
2001. The effects of increasing exercise intensity on muscle fuel utilisation in humans. J. Physiol.
536(1): 295–304. doi:10.1111/j.1469-7793.2001.00295.x.
Loon, L.J.C. Van, Greenhaff, P.L., Saris, W.H.M., and Wagenmakers, A.J.M. 2001. Tjp0536-0295. : 10.
doi:10.1111/J.1469-7793.2001.00295.X.
van Loon, L.J.C., Kruijshoop, M., Verhagen, H., Saris, W.H.M., and Wagenmakers, A.J.M. 2000.
Ingestion of Protein Hydrolysate and Amino Acid–Carbohydrate Mixtures Increases Postexercise
Plasma Insulin Responses in Men. J. Nutr. 130(10): 2508–2513. doi:10.1093/jn/130.10.2508.

85
Malowany, J.M., West, D.W.D., Williamson, E., Volterman, K.A., Abou Sawan, S., Mazzulla, M., and
Moore, D.R. 2019. Protein to Maximize Whole-Body Anabolism in Resistance-trained Females
after Exercise. Med. Sci. Sports Exerc. 51(4): 798–804. doi:10.1249/MSS.0000000000001832.
Mamerow, M.M., Mettler, J.A., English, K.L., Casperson, S.L., Arentson-Lantz, E., Sheffield-Moore, M.,
Layman, D.K., and Paddon-Jones, D. 2014. Dietary Protein Distribution Positively Influences 24-h
Muscle Protein Synthesis in Healthy Adults. J. Nutr. 144(6): 876–880. doi:10.3945/jn.113.185280.
Mariotti, F., Mahe, S., Luengo, C., Benamouzig, R., and Tome, D. 2000. Postprandial modulation of
dietary and whole-body nitrogen utilization by carbohydrates in humans. Am. J. Clin. Nutr. 72(4):
954–962. doi:10.1093/ajcn/72.4.954.
Marmy-Conus, N., Fabris, S., Proietto, J., and Hargreaves, M. 1996. Preexercise glucose ingestion and
glucose kinetics during exercise. J. Appl. Physiol. 81(2): 853–857. doi:10.1152/jappl.1996.81.2.853.
Mascher, H., Ekblom, B., Rooyackers, O., and Blomstrand, E. 2011. Enhanced rates of muscle protein
synthesis and elevated mTOR signalling following endurance exercise in human subjects. Acta
Physiol. 202(2): 175–184. doi:10.1111/j.1748-1716.2011.02274.x.
Matthews, D.E., Motil, K.J., Rohrbaugh, D.K., Burke, J.F., Young, V.R., and Bier, D.M. 1980.
Measurement of leucine metabolism in man from a primed, continuous infusion of L-[1-3C]leucine.
Am. J. Physiol. 238(5). doi:10.1152/ajpendo.1980.238.5.e473.
Mazzulla, M., Parel, J.T., Beals, J.W., Van Vliet, S., Abou Sawan, S., West, D.W.D., Paluska, S.A.,
Ulanov, A. V., Moore, D.R., and Burd, N.A. 2017a. Endurance Exercise Attenuates Postprandial
Whole-Body Leucine Balance in Trained Men. Med. Sci. Sports Exerc. 49(12): 2585–2592.
doi:10.1249/MSS.0000000000001394.
Mazzulla, M., Parel, J.T., Beals, J.W., Van Vliet, S., Abou Sawan, S., West, D.W.D., Paluska, S.A.,
Ulanov, A. V., Moore, D.R., and Burd, N.A. 2017b. Endurance Exercise Attenuates Postprandial
Whole-Body Leucine Balance in Trained Men. Med. Sci. Sports Exerc. 49(12): 2585–2592.
doi:10.1249/MSS.0000000000001394.
Mazzulla, M., Sawan, S.A., Williamson, E., Hannaian, S.J., Volterman, K.A., West, D.W.D., and Moore,

86
D.R. 2020. Protein Intake to Maximize Whole-Body Anabolism during Postexercise Recovery in
Resistance-Trained Men with High Habitual Intakes is Severalfold Greater than the Current
Recommended Dietary Allowance. J. Nutr. 150(3): 505–511. Oxford University Press.
doi:10.1093/jn/nxz249.
Mazzulla, M., Volterman, K.A., Packer, J.E., Wooding, D.J., Brooks, J.C., Kato, H., and Moore, D.R.
2018. Whole-body net protein balance plateaus in response to increasing protein intakes during post-
exercise recovery in adults and adolescents. Nutr. Metab. (Lond). 15(1): 62. Nutrition &
Metabolism. doi:10.1186/s12986-018-0301-z.
McKenzie, S., Phillips, S.M., Carter, S.L., Lowther, S., Gibala, M.J., and Tarnopolsky, M.A. 2000.
Endurance exercise training attenuates leucine oxidation and BCOAD activation during exercise in
humans. Am. J. Physiol. - Endocrinol. Metab. 278(4 41-4): 580–587.
doi:10.1152/ajpendo.2000.278.4.e580.
Meredith, C.N., Zackin, M.J., Frontera, W.R., and Evans, W.J. 1989. Dietary protein requirements and
body protein metabolism in endurance-trained men. J. Appl. Physiol. 66(6): 2850–2856.
doi:10.1152/jappl.1989.66.6.2850.
Miller, S.L., Tipton, K.D., Chinkes, D.L., Wolf, S.E., and Wolfe, R.R. 2003. Independent and combined
effects of amino acids and glucose after resistance exercise. Med. Sci. Sports Exerc. 35(3): 449–455.
doi:10.1249/01.MSS.0000053910.63105.45.
Millet, G.P., Vleck, V.E., and Bentley, D.J. 2009. Physiological differences between cycling and running:
Lessons from triathletes. Sport. Med. 39(3): 179–206. doi:10.2165/00007256-200939030-00002.
Millward, D.J., Bowtell, J.L., Pacy, P., and Rennie, M.J. 1994. Physical activity, protein metabolism and
protein requirements. Proc. Nutr. Soc. 53(1): 223–240. doi:10.1079/pns19940024.
Moore, D.R. 2019. Maximizing Post-exercise Anabolism: The Case for Relative Protein Intakes. Front.
Nutr. 6(September): 1–13. doi:10.3389/fnut.2019.00147.
Moore, D.R. 2020. One size doesn’t fit all: postexercise protein requirements for the endurance athlete.
Am. J. Clin. Nutr. 112(2): 249–250. Oxford University Press. doi:10.1093/ajcn/nqaa144.

87
Moore, D.R., Areta, J., Coffey, V.G., Stellingwerff, T., Phillips, S.M., Burke, L.M., Cléroux, M., Godin,
J.P., and Hawley, J.A. 2012. Daytime pattern of post-exercise protein intake affects whole-body
protein turnover in resistance-trained males. Nutr. Metab. 9: 1–5. doi:10.1186/1743-7075-9-91.
Moore, D.R., Camera, D.M., Areta, J.L., and Hawley, J.A. 2014. Beyond muscle hypertrophy: Why
dietary protein is important for endurance athletes. Appl. Physiol. Nutr. Metab. 39(9): 987–997.
doi:10.1139/apnm-2013-0591.
Moore, D.R., Robinson, M.J., Fry, J.L., Tang, J.E., Glover, E.I., Wilkinson, S.B., Prior, T., Tarnopolsky,
M.A., and Phillips, S.M. 2009. Ingested protein dose response of muscle and albumin protein
synthesis after resistance exercise in young men 1 – 3. : 161–168.
doi:10.3945/ajcn.2008.26401.INTRODUCTION.
Morton, J.P., Croft, L., Bartlett, J.D., MacLaren, D.P.M., Reilly, T., Evans, L., McArdle, A., and Drust,
B. 2009. Reduced carbohydrate availability does not modulate training-induced heat shock protein
adaptations but does upregulate oxidative enzyme activity in human skeletal muscle. J. Appl.
Physiol. 106(5): 1513–1521. doi:10.1152/japplphysiol.00003.2009.
Murray, B., and Rosenbloom, C. 2018a. Fundamentals of glycogen metabolism for coaches and athletes.
Nutr. Rev. 76(4): 243–259. doi:10.1093/NUTRIT/NUY001.
Murray, B., and Rosenbloom, C. 2018b. Fundamentals of glycogen metabolism for coaches and athletes.
Nutr. Rev. 76(4): 243–259. doi:10.1093/NUTRIT/NUY001.
Nair, K.S., Halliday, D., and Griggs, R.C. 1988. Leucine incorporation into mixed skeletal muscle protein
in humans. Am. J. Physiol. - Endocrinol. Metab. 254(2). doi:10.1152/ajpendo.1988.254.2.e208.
Oosthuyse, T., and Bosch, A.N. 2010. The Effect of the Menstrual Cycle on Exercise Metabolism. 40(3):
207–227.
Ott, M., Amunts, A., and Brown, A. 2016. Organization and Regulation of Mitochondrial Protein
Synthesis. Annu. Rev. Biochem. 85(1): 77–101. doi:10.1146/annurev-biochem-060815-014334.
Patel, H., and Bhardwaj, A. 2018. Physiology, respiratory quotient. StatPearls Publishing.
Pencharz, P.B., and Ball, R.O. 2003. D Ifferent a Pproaches To D Efine I Ndividual a Mino a Cid R

88
Equirements . Annu. Rev. Nutr. 23(1): 101–116. doi:10.1146/annurev.nutr.23.011702.073247.
Phillips, S.M., Atkinson, S.A., Tarnopolsky, M.A., and MacDougall, J.D. 1993a. Gender differences in
leucine kinetics and nitrogen balance in endurance athletes. J. Appl. Physiol. 75(5): 2134–2141.
doi:10.1152/jappl.1993.75.5.2134.
Phillips, S.M., Atkinson, S.A., Tarnopolsky, M.A., and MacDougall, J.D. 1993b. Gender differences in
leucine kinetics and nitrogen balance in endurance athletes. J. Appl. Physiol. 75(5): 2134–2141.
doi:10.1152/jappl.1993.75.5.2134.
Phillips, S.M., Hartman, J.W., and Wilkinson, S.B. 2005. Dietary Protein to Support Anabolism with
Resistance Exercise in Young Men. J. Am. Coll. Nutr. 24(2): 134S-139S.
doi:10.1080/07315724.2005.10719454.
Physiol, J.A., and Margaria, R. 2013. Energy cost of running Energy cost of running1. : 367–370.
Pikosky, M.A., Gaine, P.C., Martin, W.F., Grabarz, K.C., Ferrando, A.A., Wolfe, R.R., and Rodriguez,
N.R. 2006. Aerobic Exercise Training Increases Skeletal Muscle Protein Turnover in Healthy Adults
at Rest. J. Nutr. 136(2): 379–383. doi:10.1093/jn/136.2.379.
Psilander, N., Frank, P., Flockhart, M., and Sahlin, K. 2013. Exercise with low glycogen increases PGC-
1α gene expression in human skeletal muscle. Eur. J. Appl. Physiol. 113(4): 951–963.
doi:10.1007/s00421-012-2504-8.
Rafii, M., McKenzie, J.M., Roberts, S.A., Steiner, G., Ball, R.O., and Pencharz, P.B. 2008. In vivo
regulation of phenylalanine hydroxylation to tyrosine, studied using enrichment in apoB-100. Am. J.
Physiol. - Endocrinol. Metab. 294(2): 475–479. doi:10.1152/ajpendo.00604.2007.
Rand, W.M., Pellett, P.L., and Young, V.R. 2003. Meta-analysis of nitrogen balance studies for
estimating protein requirements in healthy adults. Am. J. Clin. Nutr. 77(1): 109–127.
doi:10.1093/ajcn/77.1.109.
Reidy, P.T., Walker, D.K., Dickinson, J.M., Gundermann, D.M., Drummond, M.J., Timmerman, K.L.,
Fry, C.S., Borack, M.S., Cope, M.B., Mukherjea, R., Jennings, K., Volpi, E., and Rasmussen, B.B.
2013. Protein blend ingestion following resistance exercise promotes human muscle protein

89
synthesis. J. Nutr. 143(4): 410–416. doi:10.3945/jn.112.168021.
Rennie, M.J., Edwards, R.H.T., Krywawych, S., Davies, C.T., Halliday, D., Waterlow, J.C., and
Millward, D.J. 1981. Effect of exercise on protein turnover in man. Clin. Sci. 61(5): 627–639.
doi:10.1042/cs0610627.
Riddell, M.C., Partington, S.L., Stupka, N., Armstrong, D., Rennie, C., and Tarnopolsky, M.A. 2003.
Substrate Utilization during Exercise Performed with and Without Glucose Ingestion in Female and
Male Endurance-Trained Athletes. Int. J. Sport Nutr. Exerc. Metab. 13(4): 407–421.
doi:10.1123/ijsnem.13.4.407.
Rodriguez, N.R., Vislocky, L.M., and Gaine, C. (n.d.). Dietary protein , endurance exercise , and human
skeletal-muscle protein turnover. : 40–45.
Romijn, J.A., Coyle, E.F., Sidossis, L.S., Gastaldelli, A., Horowitz, J.F., Endert, E., and Wolfe, R.R.
1993. Regulation of endogenous fat and carbohydrate metabolism in relation to exercise intensity
and duration. Am. J. Physiol. - Endocrinol. Metab. 265(3 28-3): 380–391.
doi:10.1152/ajpendo.1993.265.3.e380.
Romijn, J.A., Coyle, E.F., Sidossis, L.S., Rosenblatt, J., and Wolfe, R.R. 2000. Substrate metabolism
during different exercise intensities in endurance- trained women. J. Appl. Physiol. 88(5): 1707–
1714.
Romijn, J.A., Gastaldelli, A., Horowitz, J.F., Endert, E., and Wolfe, R.R. 2019. Regulation in relation of
endogenous fat and carbohydrate to exercise intensity and duration metabolism. : 380–391.
Rowlands, D.S., Thomson, J.S., Timmons, B.W., Raymond, F., Fuerholz, A., Mansourian, R., Zwahlen,
M.C., Métairon, S., Glover, E., Stellingwerff, T., Kussmann, M., and Tarnopolsky, M.A. 2011.
Transcriptome and translational signaling following endurance exercise in trained skeletal muscle:
Impact of dietary protein. Physiol. Genomics 43(17): 1004–1020.
doi:10.1152/physiolgenomics.00073.2011.
Sawan, S.A., Vliet, S. Van, Parel, J.T., Beals, J.W., Mazzulla, M., West, D.W.D., Philp, A., Li, Z.,
Paluska, S.A., Burd, N.A., and Moore, D.R. 2018. Translocation and protein complex co-

90
localization of mTOR is associated with postprandial myofibrillar protein synthesis at rest and after
endurance exercise. 6: 1–14. doi:10.14814/phy2.13628.
Schuetz, W., Traeger, K., Anhaeupl, T., Schanda, S., Rager, C., Vogt, J., and Georgieff, M. 1995.
Adjustment of metabolism, catecholamines and β-adrenoceptors to 90 min of cycle ergometry. Eur.
J. Appl. Physiol. Occup. Physiol. 70(1): 81–87. doi:10.1007/BF00601813.
Sheffield-Moore, M., Yeckel, C.W., Volpi, E., Wolf, S.E., Morio, B., Chinkes, D.L., Paddon-Jones, D.,
and Wolfe, R.R. 2004a. Postexercise protein metabolism in older and younger men following
moderate-intensity aerobic exercise. Am. J. Physiol. - Endocrinol. Metab. 287(3 50-3).
doi:10.1152/ajpendo.00334.2003.
Sheffield-Moore, M., Yeckel, C.W., Volpi, E., Wolf, S.E., Morio, B., Chinkes, D.L., Paddon-Jones, D.,
and Wolfe, R.R. 2004b. Postexercise protein metabolism in older and younger men following
moderate-intensity aerobic exercise. Am. J. Physiol. - Endocrinol. Metab. 287(3 50-3).
doi:10.1152/ajpendo.00334.2003.
Shepley, B., MacDougall, J.D., Cipriano, N., Sutton, J.R., Tarnopolsky, M.A., and Coates, G. 1992.
Physiological effects of tapering in highly trained athletes. J. Appl. Physiol. 72(2): 706–711.
doi:10.1152/jappl.1992.72.2.706.
Shimomura, Y., Murakami, T., Nakai, N., Nagasaki, M., and Harris, R.A. 2004. Exercise Promotes
BCAA Catabolism: Effects of BCAA Supplementation on Skeletal Muscle during Exercise. J. Nutr.
134(6): 1583S-1587S. doi:10.1093/jn/134.6.1583s.
Sidossis, L.S., Stuart, C.A., Shulman, G.I., Lopaschuk, G.D., and Wolfe, R.R. 1996. Glucose plus insulin
regulate fat oxidation by controlling the rate of fatty acid entry into the mitochondria. J. Clin. Invest.
98(10): 2244–2250. doi:10.1172/JCI119034.
Spriet, L.L. 2014. New insights into the interaction of carbohydrate and fat metabolism during exercise.
Sport. Med. 44(SUPPL.1): 87–96. doi:10.1007/s40279-014-0154-1.
Stellingwerff, T. 2012. Case study: Nutrition and training periodization in three elite marathon runners.
Int. J. Sport Nutr. Exerc. Metab. 22(5): 392–400. doi:10.1123/ijsnem.22.5.392.

91
Stöggl, T.L., and Sperlich, B. 2015. The training intensity distribution among well-trained and elite
endurance athletes. Front. Physiol. 6(OCT): 295. doi:10.3389/fphys.2015.00295.
Tang, J.E., Moore, D.R., Kujbida, G.W., Tarnopolsky, M.A., and Phillips, S.M. 2009. Ingestion of whey
hydrolysate, casein, or soy protein isolate: effects on mixed muscle protein synthesis at rest and
following resistance exercise in young men. J. Appl. Physiol. 107(3): 987–992. United States.
doi:10.1152/japplphysiol.00076.2009.
Tang, J.E., Moore, D.R., Kujbida, G.W., Tarnopolsky, M.A., and Phillips, S.M. 2019. Regulation of
Protein Metabolism in Exercise and Recovery Ingestion of whey hydrolysate , casein , or soy protein
isolate : effects on mixed muscle protein synthesis at rest and following resistance exercise in young
men. : 987–992. doi:10.1152/japplphysiol.00076.2009.
Tarnopolsky, L.J., MacDougall, J.D., Atkinson, S.A., Tarnopolsky, M.A., and Sutton, J.R. 1990. Gender
differences in substrate for endurance exercise. J. Appl. Physiol. 68(1): 302–308.
Tarnopolsky, M. 2004a. Protein requirements for endurance athletes. Nutrition 20(7–8): 662–668.
doi:10.1016/j.nut.2004.04.008.
Tarnopolsky, M. 2004b. Protein requirements for endurance athletes. Nutrition 20(7–8): 662–668.
doi:10.1016/j.nut.2004.04.008.
Tarnopolsky, M. 2004c. Protein requirements for endurance athletes. Eur. J. Sport Sci. 4(1): 1–15.
doi:10.1080/17461390400074102.
Tarnopolsky, M.A., MacDougall, J.D., and Atkinson, S.A. 1988. Influence of protein intake and training
status on nitrogen balance and lean body mass. J. Appl. Physiol. 64(1): 187–193. United States.
doi:10.1152/jappl.1988.64.1.187.
Taylor, R., and Shulman, G.I. 2011. Regulation of Hepatic Glucose Uptake. Compr. Physiol.: 286–294.
doi:10.1002/cphy.cp070226.
Teslaa, T., and Teitell, M.A. 2014. Techniques to monitor glycolysis. Methods Enzymol. 542: 91–114.
doi:10.1016/B978-0-12-416618-9.00005-4.
Tiidus, P.M., Tupling, A.R., and Houston, M.E. 2012. Biochemistry Primer for Exercise Science. Human

92
Kinetics, Incorporated. Available from https://books.google.ca/books?id=EPB6DwAAQBAJ.
Tinline-Goodfellow, C.T., West, D.W.D., Malowany, J.M., Gillen, J.B., and Moore, D.R. 2020. An Acute
Reduction in Habitual Protein Intake Attenuates Post Exercise Anabolism and May Bias Oxidation-
Derived Protein Requirements in Resistance Trained Men. Front. Nutr. 7(April): 1–9.
doi:10.3389/fnut.2020.00055.
Tipton, K.D., Ferrando, A.A., Phillips, S.M., Doyle, D., and Wolfe, R.R. 1999. Postexercise net protein
synthesis in human muscle from orally administered amino acids. Am. J. Physiol. - Endocrinol.
Metab. 276(4 39-4): 628–634. doi:10.1152/ajpendo.1999.276.4.e628.
Wagenmakers 1991.pdf. (n.d.).
Wagenmakers, A.J.M. 1999. Tracers to investigate protein and amino acid metabolism in human subjects.
Proc. Nutr. Soc. 58(4): 987–1000. doi:10.1017/S0029665199001305.
Wagenmakers, A.J.M., Beckers, E.J., Brouns, F., Kuipers, H., Soeters, P.B., Van Der Vusse, G.J., and
Saris, W.H.M. 1991a. Carbohydrate supplementation, glycogen depletion, and amino acid
metabolism during exercise. Am. J. Physiol. - Endocrinol. Metab. 260(6 23/6).
doi:10.1152/ajpendo.1991.260.6.e883.
Wagenmakers, A.J.M., Beckers, E.J., Brouns, F., Kuipers, H., Soeters, P.B., Van Der Vusse, G.J., and
Saris, W.H.M. 1991b. Carbohydrate supplementation, glycogen depletion, and amino acid
metabolism during exercise. Am. J. Physiol. - Endocrinol. Metab. 260(6 23/6).
Wagenmakers, A.J.M., Brookes, J.H., Coakley, J.H., Reilly, T., and Edwards, R.H.T. 1989. Applied
Physiology. : 159–167.
Watt, M.J., and Spriet, L.L. 2004. Regulation and role of hormone-sensitive lipase activity in human
skeletal muscle. Proc. Nutr. Soc. 63(2): 315–322. doi:10.1079/pns2004360.
Weir, J.B. d. V. 1949. New methods for calculating metabolic rate with special reference to protein
metabolism. J. Physiol. 109(1–2): 1–9. doi:10.1113/jphysiol.1949.sp004363.
Wilkinson, S.B., Phillips, S.M., Atherton, P.J., Patel, R., Yarasheski, K.E., Tarnopolsky, M.A., and
Rennie, M.J. 2008. Differential effects of resistance and endurance exercise in the fed state on

93
signalling molecule phosphorylation and protein synthesis in human muscle. 15: 3701–3717.
doi:10.1113/jphysiol.2008.153916.
Wilkinson, S.B., Tarnopolsky, M.A., Macdonald, M.J., Macdonald, J.R., Armstrong, D., and Phillips,
S.M. 2007. Consumption of fluid skim milk promotes greater muscle protein accretion after
resistance exercise than does consumption of an isonitrogenous and isoenergetic soy-protein
beverage 1 – 3. : 9–12.
Williamson, E., Kato, H., Volterman, K.A., Suzuki, K., and Moore, D.R. 2019a. The Effect of Dietary
Protein on Protein Metabolism and Performance in Endurance-trained Males. Med. Sci. Sports
Exerc. 51(2): 352–360. doi:10.1249/MSS.0000000000001791.
Williamson, E., Kato, H., Volterman, K.A., Suzuki, K., and Moore, D.R. 2019b. The Effect of Dietary
Protein on Protein Metabolism and Performance in Endurance-trained Males. Med. Sci. Sports
Exerc. 51(2): 352–360. doi:10.1249/MSS.0000000000001791.
Wolfe, R.R. 2006. The underappreciated role of muscle in health and disease. Am. J. Clin. Nutr. 84(3):
475–482. doi:10.1093/ajcn/84.3.475.
Zderic, T.W., Coggan, A.R., and Ruby, B.C. 2017. Glucose kinetics and substrate oxidation during
exercise in the follicular and luteal phases. J. Appl. Physiol. 90(2): 447–453.
doi:10.1152/jappl.2001.90.2.447.
Zello, G.A., Pencharz, P.B., and Ball, R.O. 1990. Phenylalanine flux, oxidation, and conversion to
tyrosine in humans studied with L-[1-13C]phenylalanine. Am. J. Physiol. - Endocrinol. Metab.
259(6 22-6). doi:10.1152/ajpendo.1990.259.6.e835.

94
Appendices

95
Appendix
A. Consent Form

96
Participant Consent Form
Title of Research Project: Effect of exercise intensity on estimates of dietary protein requirements in male endurance athletes.
Investigators
Daniel Moore, Assistant Professor, Faculty of Kinesiology and Physical Education, University of Toronto Contact information: 416-946-4088; [email protected] Jenna Gillen, Assistant Professor, Faculty of Kinesiology and Physical Education, University of Toronto Contact information: 416-978-3244; [email protected] Carolyn Adams MSc student, Faculty of Kinesiology and Physical Education, University of Toronto Contact information: 519-868-3222; [email protected] Purpose of the Research Project: Protein is composed of small sub-units called amino acids and is an essential nutrient that humans require to build muscle mass and support body functioning. Ingestion of protein is especially important for endurance athletes due to the elevated physical demands of training. Greater physical demands associated with exercise training increase protein needs to support post-exercise recovery. However, it is currently unknown if the intensity at which prolonged endurance exercise is performed impacts how much protein athletes require. The indicator amino acid oxidation (IAAO) method is a non-invasive technique that we will use to measure how the body uses amino acids (the sub-units of protein). Using this technique, we can determine the proportion of ingested amino acids that are used to build new proteins (e.g., muscle proteins) and what proportion of ingested amino acids are used as an energy source. This will allow us to determine whether exercise intensity influences the amount of protein needed to support post-exercise recovery. Ultimately, understanding how exercise intensity influences protein requirements will aid in our ability to develop nutrition strategies that support optimal post-exercise recovery in endurance athletes.
Description of the Research Project: Should you choose to participate, the study will involve 4 visits in which you will have to come to the Iovate/Muscletech lab. The total time commitment for study participants is approximately 24 hours (Session 1: 1 hour, Session 2: 3 hours, Session 3 & 4: 2 x 10hours). (Figure 1.). In addition to initial verbal explanation and this written explanation provided at the beginning of session 1, the beginning of each subsequent session will begin with a brief verbal ‘check-in’. At this point, the investigator/co-investigator will verbally explaining the schedule of events for that day and asking how you are feeling and if you have any questions. Should you have any concerns with your ability to complete any session (for example due to feeling unwell or injured), the investigator/co-investigator will discuss the possibility of rescheduling the session for a different date.

97
Figure 1. Schematic illustration of overall study protocol.
Description of 4 Sessions: Session 1: Introduction (~1 hour) Study participants will be given a complete verbal introduction and explanation of the details of the study (objectives, benefits, associated risk of participation) by the investigator/co-investigator and then then they can decide if they wish to participate and sign this informed consent form. If you agree to participate in the study, session 1 will serve as an introduction to the study. You will be provided with i) an accelerometer (used to measure your normal daily activity and energy expenditure), ii) a survey sheet to record information regarding your habitual training and iii) a diet log (with instructions) to record 3 full days of dietary intake (food and drink). For 3 days prior to returning to the Iovate/Muscletech lab at the Goldring Centre for High Performance Sport at the University of Toronto for Session 2, you will be required to wear the accelerometer and complete the diet log and survey sheet. Session 2: Fitness Assessment and Familiarization (~3 hours) You will be required to report to the lab following an overnight fast (minimum 7h in length). We will begin Session 2 by examining your body composition (amount of fat and fat-free mass) using the BodPod. This is a non-invasive technique commonly used to assess individual’s body composition that does not use cause radiation. The BodPod uses air displacement to assess body composition (whole body fat mass and fat-free mass). It is currently considered the ‘practical gold standard’ in body composition measurements. Following the BodPod measurements, you will be required to breath into a mouth piece for approximately 30 minutes to assess your resting energy expenditure (REE). Following this, you will be given a carbohydrate energy drink before beginning a treadmill-based fitness test (approximately 12 minutes in length) to assess your VO2max/VO2peak. You will be

98
required to wear a facemask throughout the duration of the test so that we can measure your oxygen consumption and carbon dioxide production. There may be slight annoyance or discomfort associated with exercising while wearing the mask. You will also be required to wear a heart rate monitor around your chest to measure your heart rate during the fitness test. The fitness test will begin with comfortable, low intensity exercise and will progressively increase in intensity in a linear fashion until you have reached fatigue (you can no longer continue). After the fitness test, you will be given a rest period prior to beginning the familiarization trial on the treadmill. This period will serve to get you accustomed to the demands of the test for subsequent sessions (Session 3 and 4). The exercise familiarization trial will involve running at a pre-determined pace for 10-km. The first 5-km will be performed at a speed corresponding to 70-75% of maximal heart rate (HRmax). The last 5-km will be run at a speed that corresponds to an effort of 88-93% HRmax. Successful completion of the fitness assessment and the 10-km familiarization run with high and low-intensity components will confirm your eligibility for participation in session 3 and 4. Session 3 & 4: Metabolic trial (~10 hours) (*minimum of 4-day washout between trials) You will complete two metabolic trials. Each of the two metabolic trials will consist of 3-day sessions. Day 1 and 2 of each session will be done outside of the laboratory during the participants own time and the third day of the trial will be done in the laboratory located at the Goldring Centre for High Performance Sport. There will be a minimum of 4 days between each trial. The 2 metabolic trails will be done in a random order and consist of:
• Low-intensity run: 10 and 5 km run for the preceding 2 days prior to the metabolic trial on day 3. Day 3 will consist of a 20-km run at 70-75% HR max.
• High-intensity run: 10 and 5km run for the preceding 2 days prior to the metabolic trial on
day 3. Day 3 will consist of a 20-km run at 88-93% HR max. Session 3 and 4 (day 1 and 2): Participants will wear the accelerometer and receive a controlled diet for the 2 days prior to each metabolic trial which will be match your daily energy expenditure. The controlled diet will consist of commercially available pre-packaged foods and will be tailored to any dietary restrictions and/or preferences that you have. You must abstain from consumption of caffeine and alcohol 12 hours prior to each metabolic trial. You will be required to perform two exercise sessions outside of the lab. The exercise will consist of a 10-km run at a self-selected pace (day 1) and a 5-km run at a self-selected pace (day 2). Session 3 and 4 (day 3): On the final day of session 3 and 4, you will return to the laboratory for the exercise test and metabolic trial after fasting for a minimum of 7 hours prior. Any uneaten food should also be returned to the lab and given to the investigator/co-investigators. Additionally, you will be asked to confirm your compliance with the study protocol and inform the investigator/co-investigators of any deviation from the protocol. You will be given a pre-exercise drink. Following consumption of the pre-exercise beverage, you will begin to perform the 20-km exercise test at; i) high intensity (88-93%HR max) and ii) low intensity (70-75% HR max). During each exercise trial exercise intensity will be monitored by measuring heart rate (HR) continuously during exercise using a small heart rate monitor (around

99
the chest) as well as oxygen consumption (collected with a facemask) which will be measured for 7 minutes at 3 time points during exercise, at 2-km, at 8-km and at 18-km. Following completion of the exercise test, you will consume a post-exercise meal to aid in the recovery and energy replenishment from the 20-km run consisting of a drink and protein free cookies. After the post-exercise recovery meal, the metabolic trial will begin (Figure 2). The metabolic trial will involve the hourly (8 hours) consumption of beverages with protein free cookies which will provide all of your energy needs. Throughout the 8-hour period, you are free to bring school work and entertainment (i.e. books, computer, games) to use. You will occasionally be required to breath into a gas collection chamber and provide urine samples for analysis. Following consumption of the fourth hourly beverage, 3 breath samples (15-minute intervals) and 2 urine samples (30-minute intervals) will be taken for analysis. 2.5 hours after the 5th beverage consumption, breath (6) and urine (3) samples will be taken at 30-minute intervals. Following consumption of all drinks and the collection of all breath and urine samples, the trial will be done.
Figure 2. Schematic of Session 3 and 4: day 3 (20-km exercise test and post-exercise metabolic trial).
Potential Harms, Discomforts or Inconveniences:
• The present research project will involve an initial running treadmill tests to fatigue which serves as a fitness assessment to ensure each participant’s appropriateness in participating in the study. In doing this, we will establish your maximal oxygen uptake (VO2max) or peak oxygen uptake (VO2peak). There may be some physical discomfort associated with running till fatigue however, this should not be greater than the physical demands of your typical training.
Day 3Day 2Day 1Control Diet and Exercise Exercise and Metabolic Trial
Control exercise:10km run (self-selected pace)
Control diet:8g/kg/d CHO1.4g/kg/d PRO
Control exercise:5km run (self-selected pace)
Control diet:7g/kg/d CHO1.4g/kg/d PRO
Enter lab overnight fast (7h) Pre-exercise
0.6g/kg CHO20km run
HIGH (88-93% HRmax)LOW (70-75% HRmax)
Post-exercise 2.4g/kg CHO and
protein free cookies
8-hour metabolic trial

100
• There is minor risk of falling as you become fatigued during running. Handrails are provided on the treadmill and you will be monitored closely by an investigator during the whole test.
• With the treadmill run demands, there is a risk of developing an overuse injury like plantar fasciitis, shin splints, Achilles tendonitis, Iliotibial band (ITB) friction syndrome, and runner’s knee. You are free to stop the trial at any time and if you experience any discomfort or pain. This may be the beginning of an overuse injury and we strongly encourage you to stop exercising at any time to prevent injury worsening and see a sports medicine doctor or physiotherapist if you feel this is necessary.
• You may also feel discomfort or fatigue during completion of the 20-km run and during the recovery period. We do not expect this to be any more severe than the physical discomfort associated with your average weekly training. After the run, you can rest in the lab while you recover, and you will be monitored by the investigator/co-investigator throughout the recovery.
• There is a very small risk (i.e., less than 1 in 10000 chance) of a metabolic event (i.e., acute cardiovascular incident) occurring while you are performing the VO2 max test or the 20-km treadmill runs. This risk will be no greater than the risk when you exercise during a hard training session or race. In the event that medical attention is needed, the Goldring Centre for High Performance Sport at the University of Toronto has emergency first response staff on-call. The investigator and co-investigators are First Aid and CPR certified.
• Some people find treadmill running to be very boring. You will be permitted to run with a personal music device or portable movie player (if you choose) to help pass the time.
• If you are uncomfortable or get anxious in small spaces, you may experience some discomfort in the BodPod. We will try to minimize the time spent in the BodPod machine. In addition, there is a window in the BodPod for you to see out while the analysis is happening. The BodPod test can be terminated at any time by pushing a button located inside the machine which we will identify to you prior to beginning the BodPod test.
• There may be some discomfort or inconvenience associated with consumption of the controlled diet prior to each trial. We will attempt to make the diets as similar to your typical diet as possible (as indicated to us by the completion of the diet log).
• There may also be some discomfort or dislike of the taste of the hourly beverages consumed during the metabolic trial. To make the beverages more palatable, we will also be providing protein-free cookies to be consumed with the drinks.
• The Iovate/Muscletech laboratory has shared laboratory space with other laboratories in the faculty of Kinesiology and Physical Education. Other investigators and participants may be present during your visits. If you are uncomfortable performing any of the study requirements in the presence of other investigators or participants, we can schedule your visits for occasions that there are no other bookings in the lab.
Potential Benefits: • You will be given personal testing results if you choose [e.g. fitness level (VO2max) and
resting energy expenditure]. • Your participation in this study will contribute to our understanding of protein requirements
for endurance-trained athlete who have unique performance demands and recovery needs. • You will be financially compensated for your time. Confidentiality:

101
All of your results and personal information from study participants will be kept confidential unless required by the law (For example, the law could make us give information about you if you have been abused, if you have an illness that could spread to others, if you or someone else talks about suicide or if the court orders us to give them any documents related to this study). At the beginning of the study, all participants will be de-identified by coding with a study identification number. The document connecting the study participant to the identification number will be kept in a locked cabinet in the office of the primary investigator. Study data will be input to a desktop computer in the Goldring Centre for High Performance Sport (room 425). The desktop computer is password protected in a substructure of the U of T server that requires a University of Toronto account and password to gain access and is restricted to only the study investigators/co-investigators. The study files will also be encrypted. The password for the encrypted files will only be made available to the investigator and co-investigators. Reimbursement: The proposed compensation is meant to adequately reimburse the participants for any costs incurred (e.g. parking and a small post-study meal) and to provide a token gift of appreciation for their effort. Compensation will be provided as a total of $230 for complete study participation. Should the participant choose to withdraw from the study for personal reasons, their reimbursement will be pro-rated at $30 per fitness test (Session 2) and $100 per metabolic trial at the lab (Sessions 3 & 4). The investigator/co-investigators is also able to withdraw your participation from the study at any time should you be considered ineligible (you do not meet the inclusion criteria). In this event, your reimbursement will be prorated as described above.
Compliance rules
• You will report to the Iovate/Muscletech Metabolism and Sports Science Lab (I/M Lab) at the Goldring Centre for High Performance Sport, having worn accelerometer for 2 days and having completed a 3-day dietary record prior to Session 2.
• For 2 days before the metabolic trial, you must abstain from consuming any food except for the
maintenance-diet (provided to you by the investigator/co-investigators), or water, coffee and tea without added sugar and milk.
• You will be required to perform standard exercise on the 2 days before the metabolic trials, which
will involve a 10-km run on the first day and a 5-km run on the second day (self-selected running pace).
• You must abstain from consuming caffeine (12 hours) and alcohol (12 hours) where applicable before each metabolic trial day of session 3 and 4.
• It is recommended that you sleep at least 7 hours prior to trial days. • You will report to the Goldring Centre for High Performance Sport at University of Toronto,
having worn the accelerometer for 2 days prior to the Sessions 3 & 4. Participation:

102
• Your participation in the study is entirely voluntary. The investigator/co-investigators will verbally explain the voluntary nature of study. Termination of your participation can be done at any time without need for a reason or explanation. Withdrawal from the study can be done in person or via e-mail to the investigator or co-investigators.
• Withdrawal from the study can be done at any time. Should you choose to withdraw from the study, this can be done in person or by email or phone call to the investigator or co-investigators.
• All data collected prior to participant withdrawal from the study will remain anonymous and not personally identifiable however, will be included in analysis according to; Guidance for Sponsors, Clinical Investigators, and IRBs Data Retention When Subjects Withdraw from FDA-Regulated Clinical Trials (see:https://www.fda.gov/downloads/RegulatoryInformation/Guidances/UCM126489.pdf).
• The research study you are participating in may be reviewed for quality assurance to make sure that the required laws and guidelines are followed. If chosen, (a) representative(s) of the Human Research Ethics Program (HREP) may access study-related data and/or consent materials as part of the review. All information accessed by the HREP will be upheld to the same level of confidentiality that has been stated by the research team Conflict of Interest: We have no conflicts of interest to declare. Declaration of Helsinki This study will be conducted in accordance with the Declaration of Helsinki, which is a set of ethical principles regarding human study developed for the medical community by the World Medical Association.
Consent: By signing this form, I agree that: 1) You have explained this study to me and answered all of my questions. 2) You have explained all possible harms and benefits of this study. 3) I understand that I have the right to refuse to participate in the study or withdraw from study
participation at any time without penalty (any data analyzed during my participation will remain in the study database).
4) I am free now, and in the future, to ask questions to the investigator/co-investigators about the study. 5) I understand that no personal information about myself will be given to anyone or be published without my permission. 6) I have also been provided the study timeline and been given demonstrations of all the measures that will be used in the study. 7) I agree, or consent, that I _________________________________ may take part in this study. __________________________ Printed Name of Participant Participant’s signature & date ___________________________ Printed Name of person who explained consent Signature of Person who explained consent & date

103
If you have questions about your rights as a subject in a study or injuries during a study, please contact either of the investigators or the ethics review board at [email protected] or 416 946 3273. The research study you are participating in may be reviewed for quality assurance to make sure that the required laws and guidelines are followed. If chosen, (a) representative(s) of the Human Research Ethics Program (HREP) may access study-related data and/or consent materials as part of the review. All information accessed by the HREP will be upheld to the same level of confidentiality that has been stated by the research team.

104
Appendix
B. CSEP ‘Get Active’ Questionnaire

105
© Canadian Society for Exercise Physiology, 2017. All rights reserved.
YES NO
For almost everyone, the benefits of physical activity far outweigh any risks. For some individuals, specific advice from a Qualified Exercise Professional (QEP – has post-secondary education in exercise sciences and an advanced certification in the area – see csep.ca/certifications) or health care provider is advisable. This questionnaire is intended for all ages – to help move you along the path to becoming more physically active.
Physical activity improves your physical and mental health. Even small amounts of physical activity are good, and more is better.
YES to any question: go to Reference Document – ADVICE ON WHAT TO DO IF YOU HAVE A YES RESPONSE
The following questions will help to ensure that you have a safe physical activity experience. Please answer YES or NO to each question before you become more physically active. If you are unsure about any question, answer YES.
1 Have you experienced ANY of the following (A to F) within the past six months?
A A diagnosis of/treatment for heart disease or stroke, or pain/discomfort/pressure in your chest during activities of daily living or during physical activity?
B A diagnosis of/treatment for high blood pressure (BP), or a resting BP of 160/90 mmHg or higher?
C Dizziness or lightheadedness during physical activity?
D Shortness of breath at rest?
E Loss of consciousness/fainting for any reason?
F Concussion?
2 Do you currently have pain or swelling in any part of your body (such as from an injury, acute flare-up of arthritis, or back pain) that affects your ability to be physically active?
3 Has a health care provider told you that you should avoid or modify certain types of physical activity?
4 Do you have any other medical or physical condition (such as diabetes, cancer, osteoporosis, asthma, spinal cord injury) that may affect your ability to be physically active?
NO to all questions: go to Page 2 – ASSESS YOUR CURRENT PHYSICAL ACTIVITY
P R E PA R E T O B E C O M E M O R E A C T I V E
PAGE 1 OF 2
Get Active QuestionnaireCANADIAN SOCIETY FOR EXERCISE PHYSIOLOGY – PHYSICAL ACTIVITY TRAINING FOR HEALTH (CSEP-PATH®)
I am completing this questionnaire for myself.
I am completing this questionnaire for my child/dependent as parent/guardian.

106

107
Appendix
C. Training Log and Screening/Information Questionnaire

108
Screening Questionnaire
Investigators Daniel Moore, Assistant Professor, Faculty of Kinesiology and Physical Education, University of Toronto. Contact Information: 416-946-4088, dr.moore@utoronotca Jenna Gillen, Assistant Professor, Faculty of Kinesiology and Physical Education, University of Toronto. Contact Information: 416-978-3244, [email protected] Carolyn Adams, MSc student, Faculty of Kinesiology and Physical Education, University of Toronto. Contact Information: 519-868-3222, [email protected] Subject Information Subject ID:______________________________ Date:_________________________________ Age (years):______ Height (cm):______ Weight (kg):______

109
Training log of previous 1 month: Weekly training volume estimations (time(h) and/or distance (km). Please specify the type of training (i.e. cycling, running, swimming, resistance training). Mon Tues Wed Thurs Fri Sat Sun
Week 1
Week 2
Week 3
Week 4
Additionally comments or important information to be considered from the previous month of training:_____________________________________________________________________________________________________________________________________________________ __________________________________________________________________________________________________________________________________________________________ ______________________________________________________________________________ Are you currently in season/training for upcoming competition in the next 1-2 months? (Y / N) If yes, please briefly describe any changes (i.e. tapering) in your training schedule. ___________________________________________________________________________ ______________________________________________________________________________ ______________________________________________________________________________ ______________________________________________________________________________ Recent Records (in past year): Date: _____________ Distance: ______________ Time: _________________ Date: _____________ Distance: ______________ Time: _________________ Date: _____________ Distance: ______________ Time: _________________ Date: _____________ Distance: ______________ Time: _________________ Drug/Tobacco Use: Regular Tobacco use Y / N Recreational drug use Y / N Anabolic drug use (i.e. testosterone, growth hormone) Y / N Regular Alcohol consumption (greater than 8 drinks per week) Y / N




















