
Peptide YY Regulates Bone Turnover in Rodents
10
Peptide YY Regulates Bone Turnover in Rodents KATHERINE E. WORTLEY,* KAREN GARCIA,* HARUKA OKAMOTO,* KAREN THABET,* KEITH D. ANDERSON,* VICTOR SHEN, ‡ JIM P. HERMAN, § DAVID VALENZUELA,* GEORGE D. YANCOPOULOS,* MATTHIAS H. TSCHÖP, § ANDREW MURPHY,* and MARK W. SLEEMAN* *Regeneron Pharmaceuticals Inc, Tarrytown, New York; ‡ MDS Pharma Services, Bothell, Washington; and § Department of Psychiatry, University of Cincinnati, Cincinnati, Ohio Background & Aims: Peptide YY (PYY) and pancre- atic polypeptide (PPY) are members of the neuropep- tide Y peptide family. The neuropeptide Y receptor signaling pathway has been implicated in a number of physiologic processes, including the regulation of energy balance and bone mass. To investigate the contribution of endogenous PYY and PPY to these processes, we generated both Pyy- and Ppy-deficient mice. Methods: Pyy / and Ppy / mice and their re- spective wild-type littermates were studied from 8 weeks to 9 months of age. Food intake, metabolic parameters, and locomotor activity were monitored using indirect calorimetry. Body composition and bone parameters were analyzed using dual energy x-ray absorptiometry, histomorphometry, and vertebral compression testing. Results: Studies in these mice showed an osteopenic phenotype specific to the Pyy-deficient line, which in- cluded a reduction in trabecular bone mass and a func- tional deficit in bone strength. Furthermore, female Pyy / mice showed a greater sensitivity to ovariectomy- induced bone loss compared with wild-type littermates. No food intake or metabolic phenotype was apparent in male or female Pyy / mice on standard chow. However, female Pyy / mice on a high-fat diet showed a greater propensity to gain body weight and adiposity. No met- abolic or osteopenic phenotype was observed in Ppy- deficient mice. Conclusions: These results indicate that endogenous PYY plays a critical role in regu- lating bone mass. In comparison, its role in regu- lating body weight is minor and is confined to situations of high-fat feeding. P eptide YY (PYY) and pancreatic polypeptide (PPY) are syntenic genes on mouse chromosome 11 that share structural similarity to neuropeptide Y (NPY). 1,2 The neu- ronal and hormonal functions of PYY and PPY have not been fully characterized and have been hampered by their cleavage into multiple bioactive peptides, all of which circulate in the blood stream and bind with varying affinity to a family of G protein– coupled receptors, Y1– Y6. NPY is one of the most potent orexigenic peptides identified and is widely expressed in the brain. 3 In con- trast, PYY and PPY are mainly expressed in peripheral tissues; endocrine L cells have been identified as the major source of circulating PYY, but studies have also documented expression in alpha cells of pancreatic islets and subnuclei of the brainstem. 4–6 PYY secretion is in- creased following food intake in both humans and ro- dents, and it binds with similar affinity to all Y receptors. 7 Full-length PYY is cleaved by dipeptidyl peptidase IV to PYY3-36. This peptide constitutes the majority of total PYY in the postprandial state and is reported to be a more selective ligand for the Y2 receptor subtype, al- though binding and activation of the Y1 and Y5 receptor subtypes are still observed. 8,9 PPY is primarily expressed in pancreatic islets with some expression in the gut. 5 Similar to PYY, PPY secretion increases postprandially but shows selectivity for the Y4 receptor. 10 Whereas the physiologic effects of NPY are well docu- mented, those of PPY are less clear but reports have been published that include inhibition of pancreatic secretion, gallbladder activity, intestinal motility, and a reduction in food intake following peripheral infusion. 10,11 The physiologic function of PYY, particularly in rodents, is perhaps the most controversial. Initial reports high- lighted strong and consistent decreases in food intake and body weight following short-term and long-term administration of this peptide. 12 Many of these effects were, however, not reproducible by other laboratories 13,14 or attributed to a conditioned taste aversion response following short-term administration. 14 In such circum- stances, genetic deletion studies are often useful in gen- erating decisive data in regard to the biological function of a gene. Unfortunately, the analysis of the various Pyy null mice by different investigators has only added to the confusion. Initial reports documented no changes in food intake and body weight in mice that simultaneously lacked Pyy and Ppy. 15 Analysis of an independent line of mice that specifically lack Pyy showed a sex-specific adult- onset mild obese phenotype without any disruption in food intake. 16 In yet a third line of Pyy null mice, a more Abbreviations used in this paper: BMC, bone mineral content; BMD, bone mineral density; NPY, neuropeptide Y; PPY, pancreatic polypep- tide; PYY, peptide YY; WT, wild-type. © 2007 by the AGA Institute 0016-5085/07/$32.00 doi:10.1053/j.gastro.2007.08.024 BASIC– ALIMENTARY TRACT GASTROENTEROLOGY 2007;133:1534 –1543
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Peptide YY Regulates Bone Turnover in Rodents

P
KVA
*C
BatsoocpmstacwhRpctPiNmfpadtlls
PsrbccaYit
BA
SIC–
ALIM
ENTA
RY
TRA
CT
GASTROENTEROLOGY 2007;133:1534–1543
eptide YY Regulates Bone Turnover in Rodents
ATHERINE E. WORTLEY,* KAREN GARCIA,* HARUKA OKAMOTO,* KAREN THABET,* KEITH D. ANDERSON,*ICTOR SHEN,‡ JIM P. HERMAN,§ DAVID VALENZUELA,* GEORGE D. YANCOPOULOS,* MATTHIAS H. TSCHÖP,§
NDREW MURPHY,* and MARK W. SLEEMAN*
Regeneron Pharmaceuticals Inc, Tarrytown, New York; ‡MDS Pharma Services, Bothell, Washington; and §Department of Psychiatry, University of Cincinnati,
incinnati, OhiotmdacdFPPmtsiSb
mpgipplaawofseoncflmof
bt
ackground & Aims: Peptide YY (PYY) and pancre-tic polypeptide (PPY) are members of the neuropep-ide Y peptide family. The neuropeptide Y receptorignaling pathway has been implicated in a numberf physiologic processes, including the regulationf energy balance and bone mass. To investigate theontribution of endogenous PYY and PPY to theserocesses, we generated both Pyy- and Ppy-deficientice. Methods: Pyy�/� and Ppy�/� mice and their re-
pective wild-type littermates were studied from 8 weekso 9 months of age. Food intake, metabolic parameters,nd locomotor activity were monitored using indirectalorimetry. Body composition and bone parametersere analyzed using dual energy x-ray absorptiometry,istomorphometry, and vertebral compression testing.esults: Studies in these mice showed an osteopenichenotype specific to the Pyy-deficient line, which in-luded a reduction in trabecular bone mass and a func-ional deficit in bone strength. Furthermore, femaleyy�/� mice showed a greater sensitivity to ovariectomy-
nduced bone loss compared with wild-type littermates.o food intake or metabolic phenotype was apparent inale or female Pyy�/� mice on standard chow. However,
emale Pyy�/� mice on a high-fat diet showed a greaterropensity to gain body weight and adiposity. No met-bolic or osteopenic phenotype was observed in Ppy-eficient mice. Conclusions: These results indicatehat endogenous PYY plays a critical role in regu-ating bone mass. In comparison, its role in regu-ating body weight is minor and is confined toituations of high-fat feeding.
eptide YY (PYY) and pancreatic polypeptide (PPY) aresyntenic genes on mouse chromosome 11 that share
tructural similarity to neuropeptide Y (NPY).1,2 The neu-onal and hormonal functions of PYY and PPY have noteen fully characterized and have been hampered by theirleavage into multiple bioactive peptides, all of whichirculate in the blood stream and bind with varyingffinity to a family of G protein– coupled receptors, Y1–6. NPY is one of the most potent orexigenic peptides
dentified and is widely expressed in the brain.3 In con-
rast, PYY and PPY are mainly expressed in peripheralissues; endocrine L cells have been identified as theajor source of circulating PYY, but studies have also
ocumented expression in alpha cells of pancreatic isletsnd subnuclei of the brainstem.4 – 6 PYY secretion is in-reased following food intake in both humans and ro-ents, and it binds with similar affinity to all Y receptors.7
ull-length PYY is cleaved by dipeptidyl peptidase IV toYY3-36. This peptide constitutes the majority of totalYY in the postprandial state and is reported to be aore selective ligand for the Y2 receptor subtype, al-
hough binding and activation of the Y1 and Y5 receptorubtypes are still observed.8,9 PPY is primarily expressedn pancreatic islets with some expression in the gut.5
imilar to PYY, PPY secretion increases postprandiallyut shows selectivity for the Y4 receptor.10
Whereas the physiologic effects of NPY are well docu-ented, those of PPY are less clear but reports have been
ublished that include inhibition of pancreatic secretion,allbladder activity, intestinal motility, and a reductionn food intake following peripheral infusion.10,11 Thehysiologic function of PYY, particularly in rodents, iserhaps the most controversial. Initial reports high-
ighted strong and consistent decreases in food intakend body weight following short-term and long-termdministration of this peptide.12 Many of these effectsere, however, not reproducible by other laboratories13,14
r attributed to a conditioned taste aversion responseollowing short-term administration.14 In such circum-tances, genetic deletion studies are often useful in gen-rating decisive data in regard to the biological functionf a gene. Unfortunately, the analysis of the various Pyyull mice by different investigators has only added to theonfusion. Initial reports documented no changes inood intake and body weight in mice that simultaneouslyacked Pyy and Ppy.15 Analysis of an independent line of
ice that specifically lack Pyy showed a sex-specific adult-nset mild obese phenotype without any disruption inood intake.16 In yet a third line of Pyy null mice, a more
Abbreviations used in this paper: BMC, bone mineral content; BMD,one mineral density; NPY, neuropeptide Y; PPY, pancreatic polypep-ide; PYY, peptide YY; WT, wild-type.
© 2007 by the AGA Institute0016-5085/07/$32.00
doi:10.1053/j.gastro.2007.08.024

rm
dtn
h(ocRt(9s
smaLfsalodbh
soafss
iairo4mc
eIfaoaspcaE�m3t
svemrtcnGm
oIekscotp0cfe
f(Mtt
i
BA
SIC–
ALI
MEN
TARY
TRA
CT
November 2007 PYY AND BONE 1535
ecent study reported morbid early-onset obesity andarked hyperphagia.17
In the present study, we generated both Pyy- and Ppy-eficient mice to investigate the physiologic function ofhese peptides and surprisingly found an osteopenic phe-otype specific to the Pyy-deficient line.
Materials and MethodsGeneration of Pyy�/� Mice and Ppy�/� Miceand Experimental ProceduresMice were generated using the previously described
igh-throughput homologous recombination technology18
Supplementary Information; see supplemental materialnline at www.gastrojournal.org). All procedures wereonducted in compliance with protocols approved by theegeneron Institutional Animal Care and Use Commit-
ee. Animals had free access to either standard chow5020; Purina, St Louis, MO) or a high-fat diet (45% fat,3075; Harlan Teklad, Madison, WI) unless otherwisepecified.
Body Composition, Indirect Calorimetry, andFood IntakeBody composition, including bone mineral den-
ity (BMD) and bone mineral content (BMC), was deter-ined before and after a high-fat diet for each individual
nimal using dual emission x-ray absorption (PIXImus;unar, Madison, WI) and the percentage lean muscle andat mass calculated. Metabolic measurements were as-essed both on standard chow (at 8 –10 weeks of age) andfter exposure to a high-fat diet using an Oxymax (Co-umbus Instruments International Corp, Columbus, OH)pen circuit indirect calorimetry system as previouslyescribed.19 Food intake on a standard diet was assessedy automated measurements in metabolic cages or on aigh-fat diet by cage top weights.
BiochemistryAll biochemical analyses were performed on serum
amples. Serum samples were collected either from retro-rbital bleeds from mice fasted for 4 hours and brieflynesthetized with isoflurane or from trunk blood collectedrom decapitated mice fasted overnight. For details on as-ays used, refer to the Supplementary Information (seeupplemental material online at www.gastrojournal.org).
Expression Analysis�-galactosidase staining and immunohistochem-
stry were performed on adult mice that had been deeplynesthetized (240 mg/kg ketamine, 48 mg/kg xylazine,ntramuscularly) and exsanguinated with ice-cold hepa-inized saline. Tissue was fixed by transcardial perfusionf 2% paraformaldehyde (for �-galactosidase staining) or% paraformaldehyde (for immunohistochemistry) in 0.1ol/L phosphate buffer, postfixed for 2 hours, and then
ryoprotected for at least 24 hours in 2 changes of buff- a
red 30% sucrose at 4°C with agitation before sectioning.mmunohistochemical staining of Pyy and Ppy was per-ormed on slide-mounted sections of pancreas, ileum,nd colon (16 �m) and on free-floating coronal sectionsf brain (40 �m) as described19,20 using a rabbit serumgainst porcine/rat Pyy (1:50,000; Bachem, King of Prus-ia, PA) or a guinea pig serum against rat pancreaticolypeptide (1:5000; Linco, St. Charles, MO). Fluores-ently conjugated secondary antisera (Alexa 488 goatnti-rabbit or goat anti– guinea pig; Molecular Probes,ugene, OR) were used at a dilution of 1:200. To visualize-galactosidase, sections were incubated in buffered 1g/mL X-Gal (Molecular Probes) for 12–24 hours at
7°C as described previously.21 The sections were coun-erstained with eosin.
Histology and Histomorphometry of BoneTissueFor histologic analyses, L3–L5 vertebrae were dis-
ected and collected into 10% buffered formalin. Lumbarertebrae were dehydrated through a series of ascendingthanol solution, cleared with xylene, infiltrated withethylmethacrylate, and embedded in methylmethac-
ylate/catalyst. Sagittal sections, through the middle ofhe vertebral body, were obtained using a rotary mi-rotome (model RM2165; Leica Microsystems Inc, Ban-ockburn, IL). One 4-�m section was stained witholdner’s trichrome stain, and one 8-�m section wasounted unstained.Bone histomorphometry was performed using Oste-
Measure image analysis software program (OsteoMetrics,nc, Atlanta, GA) interfaced with an Eclipse E400 light/pifluorescent microscope and video subsystem (Nikon, To-yo, Japan). Histomorphometric evaluations of the stainedlides were performed in a blinded manner, and the nomen-lature and calculations used for all parameters were basedn standardized terms and formulae.22 For measurement ofhe lumbar vertebral body, the region of interest was com-osed of an area of marrow cavity that was approximately.2 mm from the 2 growth plates and the ventral and dorsalortex. Evaluation of structural features was performedrom an image projected using a 10� objective. Six fields onach slide were evaluated.
Compression Test of Lumbar Vertebral BodyCompression testing of L2 vertebrae was per-
ormed using an Instron Mechanical Testing Machinemodel 4465 retrofitted to 5500; Instron Corp, Canton,
A). For more details on the methods used, refer tohe Supplementary Information (see supplemental ma-erial online at www.gastrojournal.org).
Data and Statistical AnalysesData are expressed as mean � SEM, and compar-
sons were performed using a t test or repeated-measures
nalysis of variance where appropriate using the program
St
btPlBPasfiPPaichiacbMafi
egp(smwiwnnict
b2mscbom
wACscls(s
eet5d((NBPPd0v0B.ed0
FgtgwPs
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1536 WORTLEY ET AL GASTROENTEROLOGY Vol. 133, No. 5
tatView (Abacus Concepts, Berkeley, CA). P values lesshan .05 were considered significant.
ResultsGeneration of Pyy�/� and Ppy�/� Null MicePyy�/� and Ppy�/� knockout mice were generated
y constructing and utilizing BAC-based targeting vec-ors that contained a precise deletion of the entire Pyy orpy coding region from the ATG, with insertion of the
acZ reporter gene (Figure 1). Correct targeting of theAC vector and loss of the wild-type/native Pyy or nativepy allele in embryonic stem cells was determined usingquantitative loss-of-native-allele assay as previously de-
cribed.18 Loss of peptide expression was further con-rmed by immunocytochemistry in the Pyy�/� andpy�/� mice. Figure 2 shows strong immunostaining ofyy in wild-type control littermate (WT) mice, which isbsent in Pyy�/� mice. In contrast, Pyy immunostainingn Ppy�/� mice was comparable to that seen in littermateontrol (WT) mice, indicating that the deletion of Ppyad no impact on the expression of Pyy. Likewise, strong
mmunostaining of Ppy was seen in WT mice, which wasbsent in Ppy�/� mice but present in Pyy�/� mice at levelsomparable to WT mice (Figure 2). We observed a normalirth ratio of Pyy�/� and Ppy�/� mice as predicted byendelian genetics, and all mice appeared grossly normal
nd reached normal development milestones during therst 8 weeks of age (data not shown).Consistent with previous reports of endogenous Pyy
xpression, �-galactosidase staining for the lacZ reporterene in Pyy�/� mice showed expression in islet cells of theancreas and in endocrine cells of the ileum and colon
Figure 3). Sparse staining for the lacZ reporter gene waseen in sections of the jejunum and duodenum (Supple-
entary Figure 1; see supplemental material online atww.gastrojournal.org). Cellular expression was also seen
n the brain near the midline in the reticular hindbrainithin the gigantocellular reticular nucleus near to, butot including, the raphe obscurus and raphe magnusuclei (Supplementary Figure 1). �-galactosidase staining
n Ppy�/� mice showed a pattern of expression that wasonfined to the pancreas, ileum, and colon, similar tohat seen in Pyy�/� mice (Figure 3).
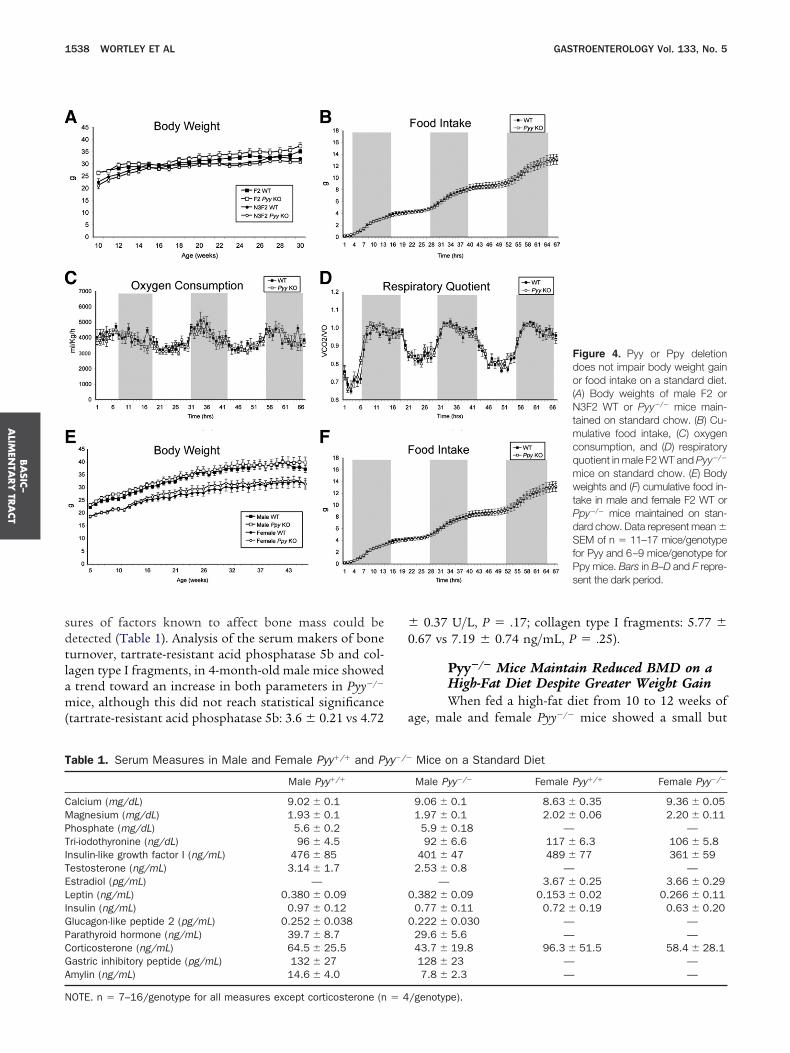
Pyy or Ppy Deletion Does Not Affect FoodIntake or Body Weight in Mice on aStandard DietAnalyses in F2 generation mice with a 75% C57Bl6
ackground revealed no differences in body weight or4-hour food intake (Figure 4A and B) between Pyy�/�
ice and WT littermates when fed a standard diet. Con-istent with these findings, no disturbances in oxygenonsumption, substrate utilization (Figure 4C and D),ody composition, locomotor activity (data not shown),r expression of hypothalamic neuropeptides (Supple-
entary Table 1; see supplemental material online at 0ww.gastrojournal.org) were observed in the Pyy�/� mice.nalysis of mice backcrossed a further generation to57Bl6 mice (N3F2 � 94% C57Bl6 background) also
howed no difference in body weight in male Pyy�/� miceompared with their WT littermates (Figure 4A). Simi-arly, Ppy-deficient mice with a 75% C57Bl6 backgroundhowed no difference in body weight or food intakeFigure 4E and F) or metabolic parameters (data nothown) when compared with their WT littermates.
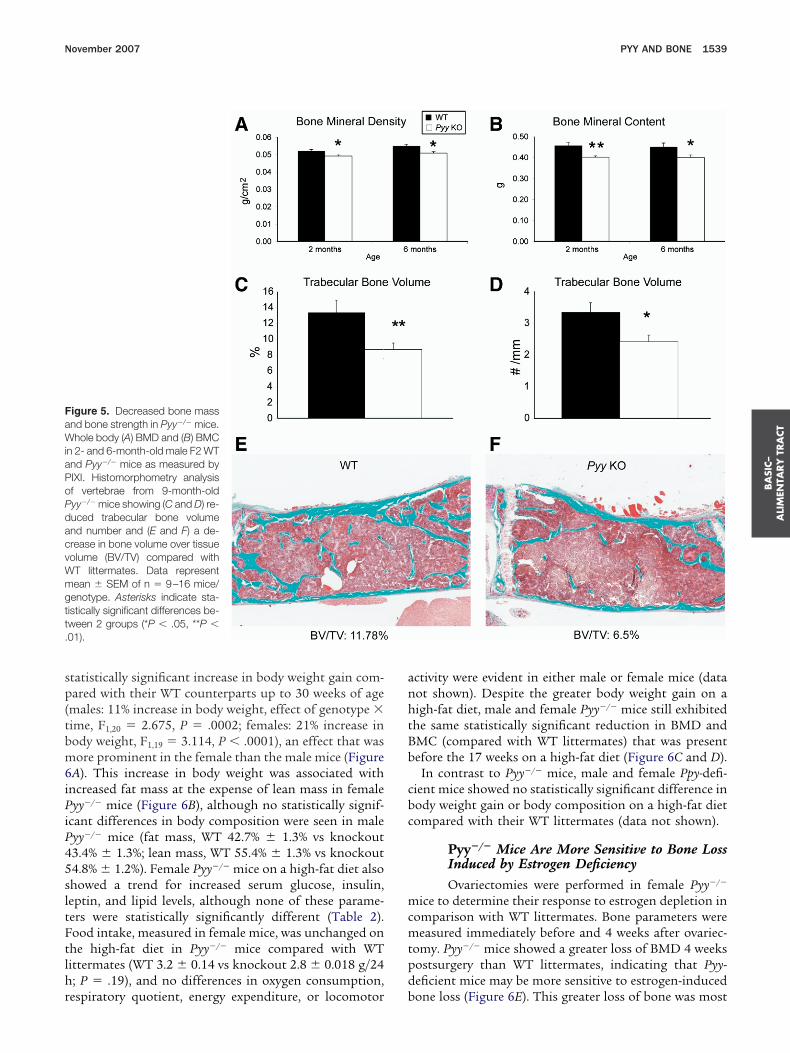
Pyy But Not Ppy Deficiency Causes BoneLossWhereas body composition analysis by dual-en-
rgy x-ray absorptiometry (PIXImus) showed no differ-nce in lean or fat mass in Pyy�/� mice on a standard diet,otal body BMD and BMC were consistently decreased by%–10% in comparison with WT littermates. Initially weetected these differences in F2 generation Pyy�/� mice
75% C57Bl6 background) at 2 and 6 months of ageFigure 5A and B). Further analysis after backcrossing in
3F2 mice showed a similar reduction in total BMD andMC (BMD: 0.0532 � 0.0005 vs 0.0495 � 0.0004 g/cm2,� .0001; BMC: 0.494 � 0.0097 vs 0.443 � 0.0109 g,� .0018), and on refined measurement a loss was
etected in both the vertebrae (BMD: 0.0543 � 0.0011 vs.0495 � 0.0013 g/cm2, P � .01; BMC: 0.0369 � 0.0011s 0.0329 � 0.0011 g, P � .016) and whole femur (BMD:.0672 � 0.0009 vs 0.0633 � 0.0008 g/cm2, P � .005;MC: 0.0319 � 0.0103 vs 0.0283 � 0.00078 g, P �
0089). No gross abnormalities were observed in the skel-tal architecture and no loss of BMD or BMC could beetected in Ppy null mice (BMD: 0.0484 � 0.0022 vs.0477 � 0.0022 g/cm2, P � .83; BMC: 0.428 � 0.0202 vs
igure 1. Generation of Pyy�/� and Ppy�/� mice. (A) Schematic dia-ram of the murine wild-type Pyy alleles and the targeting vectors usedo generate a null Pyy allele by precise substitution of the lacZ reporterene and neo selectable marker. (B) Schematic diagram of the murineild-type Ppy alleles and the targeting vectors used to generate a nullpy allele by precise substitution of the lacZ reporter gene and neoelectable marker.
.419 � 0.0205 g, P � .75).

vsaWt
r1iNsh
tom p
FieitaPmmwPlcivL
mg
BA
SIC–
ALI
MEN
TARY
TRA
CT
November 2007 PYY AND BONE 1537
A blinded histomorphometry analysis of the lumbarertebral bodies taken from male Pyy�/� mice revealed aignificant 35% reduction in trabecular bone volume and
28% reduction in trabecular number compared withT littermates (Figure 5C–F). A subsequent compression
est on the L5 vertebrae of 9-month-old Pyy�/� mice
Figure 2. Immunostaining of Pyy (top panels) and Ppy (bot
igure 3. �-galactosidase stain-ng reveals specific reporter genexpression (LacZ reporter, in blue)
n pancreatic islet cells and en-eroendocrine cells of the ileumnd colon. (Top panels) A femaleyy�/� (right), a 14-week-oldale Pyy�/� (middle), and a fe-ale Pyy�/� mouse (left) (8–16eeks old). (Bottom panels) Malepy�/� mice (16 weeks old). Yel-
ow arrows indicate lacZ-positiveells (bar � 100 �m). D, duct; IL,
slet of Langerhans; IV, intestinalilli; EL, endothelial lining; L, lumen;P, lamina propria; MM, muscularisucosae; MG, mucosal gland; GC,
oblet cell.evealed a 33% decrease in maximal load (20.45 � 2.77 vs3.62 � 1.56 N; P � .023) and a trend toward a decrease
n ultimate strength (10.64 � 1.60 vs 7.48 � 0.92/mm2; P � .062). This reduction in bone mass and
trength did not appear to be secondary to metabolic orormonal changes because no difference in serum mea-
anels) in pancreatic islets of WT, Pyy�/�, and Ppy�/� mice.
sdtlam(
�0
a
T
CMPTITELIGPCGA
N
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1538 WORTLEY ET AL GASTROENTEROLOGY Vol. 133, No. 5
ures of factors known to affect bone mass could beetected (Table 1). Analysis of the serum makers of boneurnover, tartrate-resistant acid phosphatase 5b and col-agen type I fragments, in 4-month-old male mice showed
trend toward an increase in both parameters in Pyy�/�
ice, although this did not reach statistical significancetartrate-resistant acid phosphatase 5b: 3.6 � 0.21 vs 4.72
able 1. Serum Measures in Male and Female Pyy�/� and P
Male Pyy�/�
alcium (mg/dL) 9.02 � 0.1agnesium (mg/dL) 1.93 � 0.1hosphate (mg/dL) 5.6 � 0.2ri-iodothyronine (ng/dL) 96 � 4.5nsulin-like growth factor I (ng/mL) 476 � 85estosterone (ng/mL) 3.14 � 1.7stradiol (pg/mL) —eptin (ng/mL) 0.380 � 0.09nsulin (ng/mL) 0.97 � 0.12lucagon-like peptide 2 (pg/mL) 0.252 � 0.038arathyroid hormone (ng/mL) 39.7 � 8.7orticosterone (ng/mL) 64.5 � 25.5astric inhibitory peptide (pg/mL) 132 � 27mylin (ng/mL) 14.6 � 4.0
OTE. n � 7–16/genotype for all measures except corticosterone (n � 4
0.37 U/L, P � .17; collagen type I fragments: 5.77 �.67 vs 7.19 � 0.74 ng/mL, P � .25).
Pyy�/� Mice Maintain Reduced BMD on aHigh-Fat Diet Despite Greater Weight GainWhen fed a high-fat diet from 10 to 12 weeks of
ge, male and female Pyy�/� mice showed a small but
Figure 4. Pyy or Ppy deletiondoes not impair body weight gainor food intake on a standard diet.(A) Body weights of male F2 orN3F2 WT or Pyy�/� mice main-tained on standard chow. (B) Cu-mulative food intake, (C) oxygenconsumption, and (D) respiratoryquotient in male F2 WT and Pyy�/�
mice on standard chow. (E) Bodyweights and (F) cumulative food in-take in male and female F2 WT orPpy�/� mice maintained on stan-dard chow. Data represent mean �SEM of n � 11–17 mice/genotypefor Pyy and 6–9 mice/genotype forPpy mice. Bars in B–D and F repre-sent the dark period.
Mice on a Standard Diet
Male Pyy�/� Female Pyy�/� Female Pyy�/�
9.06 � 0.1 8.63 � 0.35 9.36 � 0.051.97 � 0.1 2.02 � 0.06 2.20 � 0.115.9 � 0.18 — —92 � 6.6 117 � 6.3 106 � 5.8
401 � 47 489 � 77 361 � 592.53 � 0.8 — —
— 3.67 � 0.25 3.66 � 0.29.382 � 0.09 0.153 � 0.02 0.266 � 0.110.77 � 0.11 0.72 � 0.19 0.63 � 0.20.222 � 0.030 — —29.6 � 5.6 — —43.7 � 19.8 96.3 � 51.5 58.4 � 28.1128 � 23 — —7.8 � 2.3 — —
yy�/�
0
0
/genotype).
sp(tbm6iPiP45sltFtlhr
anhtBb
cbc
mcmtpd
FaWiaPoPdacvWmgtt.
BA
SIC–
ALI
MEN
TARY
TRA
CT
November 2007 PYY AND BONE 1539
tatistically significant increase in body weight gain com-ared with their WT counterparts up to 30 weeks of age
males: 11% increase in body weight, effect of genotype �ime, F1,20 � 2.675, P � .0002; females: 21% increase inody weight, F1,19 � 3.114, P � .0001), an effect that wasore prominent in the female than the male mice (Figure
A). This increase in body weight was associated withncreased fat mass at the expense of lean mass in femaleyy�/� mice (Figure 6B), although no statistically signif-
cant differences in body composition were seen in maleyy�/� mice (fat mass, WT 42.7% � 1.3% vs knockout3.4% � 1.3%; lean mass, WT 55.4% � 1.3% vs knockout4.8% � 1.2%). Female Pyy�/� mice on a high-fat diet alsohowed a trend for increased serum glucose, insulin,eptin, and lipid levels, although none of these parame-ers were statistically significantly different (Table 2).ood intake, measured in female mice, was unchanged onhe high-fat diet in Pyy�/� mice compared with WTittermates (WT 3.2 � 0.14 vs knockout 2.8 � 0.018 g/24; P � .19), and no differences in oxygen consumption,
igure 5. Decreased bone massnd bone strength in Pyy�/� mice.hole body (A) BMD and (B) BMC
n 2- and 6-month-old male F2 WTnd Pyy�/� mice as measured byIXI. Histomorphometry analysisf vertebrae from 9-month-oldyy�/� mice showing (C and D) re-uced trabecular bone volumend number and (E and F) a de-rease in bone volume over tissueolume (BV/TV) compared withT littermates. Data representean � SEM of n � 9–16 mice/enotype. Asterisks indicate sta-istically significant differences be-ween 2 groups (*P � .05, **P �01).
espiratory quotient, energy expenditure, or locomotor b
ctivity were evident in either male or female mice (dataot shown). Despite the greater body weight gain on aigh-fat diet, male and female Pyy�/� mice still exhibitedhe same statistically significant reduction in BMD andMC (compared with WT littermates) that was presentefore the 17 weeks on a high-fat diet (Figure 6C and D).
In contrast to Pyy�/� mice, male and female Ppy-defi-ient mice showed no statistically significant difference inody weight gain or body composition on a high-fat dietompared with their WT littermates (data not shown).
Pyy�/� Mice Are More Sensitive to Bone LossInduced by Estrogen DeficiencyOvariectomies were performed in female Pyy�/�
ice to determine their response to estrogen depletion inomparison with WT littermates. Bone parameters wereeasured immediately before and 4 weeks after ovariec-
omy. Pyy�/� mice showed a greater loss of BMD 4 weeksostsurgery than WT littermates, indicating that Pyy-eficient mice may be more sensitive to estrogen-induced
one loss (Figure 6E). This greater loss of bone was most
nhgvbc
mlPbcNr
cvitflte
nDmedtadrabiw
T
GITNCL
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1540 WORTLEY ET AL GASTROENTEROLOGY Vol. 133, No. 5
oticeable in the lumbar vertebrae, a site with a relativelyigh proportion of trabecular bone. Both genotypesained a similar amount of body weight 4 weeks posto-ariectomy (Figure 6F), indicating that the greater loss ofone in the Pyy�/� mice was not secondary to any greaterhange in body weight.
DiscussionOur evaluation of Pyy�/� mice indicates that the
ost important physiologic role of endogenous Pyy mayie in the control of bone remodeling. Here we show thatyy�/� mice exhibit a substantial reduction in trabecularone mass, resulting in decreased bone strength. This isonsistent with a previous report suggesting that thePY receptor signaling pathway may be implicated in the
egulation of bone turnover.23
Bone tissue remodeling is a continuously ongoing pro-ess. Bone-resorbing osteoclasts constantly remove smallolumes of bone, which are then replaced by bone-form-ng osteoblasts.24 These 2 processes are tightly coupled sohat the rate of bone resorption is equal to that of boneormation, resulting in a constant bone volume in adultife until menopausal and age-related bone loss occur dueo uncoupling of these 2 processes.24 A disturbance in
ither bone resorption or formation causes a change in Net bone turnover, leading to changes in bone volume.eletion of the Pyy gene led to reduced BMD due to aarked decrease in trabecular bone volume, which was
vident as early as 8 weeks of age and is maintaineduring adulthood. Although serum markers of boneurnover were not statistically increased in Pyy�/� micend a dynamic measure of bone resorption is inherentlyifficult to obtain, the data strongly suggest that boneesorption is exceeding bone formation, which leads ton osteoporotic phenotype. An analogous increase inone turnover is seen in models of estrogen deficiency
nduced by ovariectomy in rodents or in postmenopausalomen.25,26
able 2. Serum Measurements in 16-Hour Fasted FemalePyy�/� and Pyy�/� Mice on a High-Fat Diet
Pyy�/� Pyy�/�
lucose (mg/dL) 172 � 8.1 182 � 9.5nsulin (ng/mL) 2.67 � 0.48 4.65 � 1.10riglycerides (mg/dL) 141 � 18 167 � 18onesterified fatty acids (mEq/L) 1.86 � 0.21 2.32 � 0.15holesterol (mg/dL) 142 � 12 155 � 9eptin (ng/mL) 13.4 � 1.8 16.3 � 1.1
Figure 6. Increased sensitivity tohigh-fat diet–induced weight gainand estrogen-induced bone lossin Pyy�/� mice. Exposure to ahigh-fat diet caused (A) greaterweight gain in Pyy�/� mice relativeto WT littermates and (B) inducedgreater adiposity in female miceafter 17 weeks on the diet. (C andD) Male and female Pyy�/� micemaintained reduced BMD andBMC on a high-fat diet despitegreater weight gain. Data repre-sent mean � SEM of n � 8–18mice/genotype. (E) Ovariectomycaused a greater loss of BMD inPyy�/� female mice fed a standarddiet compared with WT litter-mates, whereas (F) body weightremained similar between the 2genotypes. Total BMD and BMDin the lumbar vertebrae and femurwere measured before and 4weeks after ovariectomy. The per-centage loss of BMD relative to theinitial value is shown. Body weightdata represents weights at 4weeks postsurgery. Data repre-sent mean � SEM of n � 4–6mice/genotype. Asterisks indicatestatistically significant differencesbetween 2 groups (*P � .05, **P �.01).
OTE. n � 7–8/genotype.

cfiiwWtaDttubmphatwmsalTutrupu
cgtochmabtsicilatTmr
oDa
cifecmemmfmmaw
prlfaespaWpiSIfaoat
rtatstmtmrgtetogsiT
BA
SIC–
ALI
MEN
TARY
TRA
CT
November 2007 PYY AND BONE 1541
Previous studies have identified hypothalamic Y2 re-eptors as having a major role in the regulation of boneormation. Mice deficient in Y2 receptors (Y2�/�) showncreased osteoblast activity and increased bone mineral-zation and formation rate,27 suggesting that this path-ay has an important tonic inhibitory influence on bone.hereas full-length PYY binds with comparable affinity
o all Y receptors,7 PYY3-36 is known to have the highestffinity for the Y2 receptor as based on detailed studies byumont et al.9,28 It would therefore appear counterintui-
ive that Pyy�/� mice have reduced bone mass in light ofhe fact that Y2�/� mice have a 2-fold increase in trabec-lar bone volume. Nevertheless, one notable differenceetween the Pyy�/� and Y2�/� mice is that the latterodel exhibits increased hypothalamic Npy expression,27
resumably due to the absence of the presynaptic autoin-ibitory Y2 receptor population. Elevated Npy expression iscommon feature in leptin-deficient ob/ob mice as well as
he global and hypothalamus-specific Y2�/� mice, all ofhich show an increase in osteoblast activity.29 The Pyy�/�
ice generated for this study, like other deletions,27
howed no difference in Npy expression in the hypothal-mus, indicating that a change in Npy signaling is un-ikely to be mediating the bone phenotype in these mice.hus, the mechanism through which circulating Pyy reg-lates bone mass is unclear. It is possible that Pyy actshrough a population of postsynaptic Y2 receptors toegulate bone mass or possibly through a novel, as yetnidentified, Y receptor in the brain that influences sym-athetic nervous system activity, which is known to reg-late bone mass.30
The role of PYY in maintaining bone homeostasis isonsistent with emerging evidence that a number ofastrointestinal hormones are important in not only in-egrating nutrient intake but also influencing bone turn-ver.31,32 Glucagon-like peptide 2 is also secreted from Lells and decreases bone resorption, whereas gastric in-ibitory peptide, ghrelin, and amylin stimulate bone for-ation.31–33 Thus, hormones in the gastrointestinal tract
nd pancreas are able to regulate the acute interactionetween food intake and bone turnover. It is possiblehat Pyy regulates bone mass indirectly by regulating theecretion of other hormones that affect bone. However,n our Pyy-deficient mice, we did not detect any clearhange in a number of hormones known to be involvedn the regulation of bone mass. Although circulatingevels of corticosterone, testosterone, and amylin showed
trend to be reduced in the Pyy-deficient mice, none ofhese markers showed a statistically significant reduction.hus, the reduction in bone volume in the Pyy-deficientice does not appear to be associated with a consistent
eduction in any of these hormones.As outlined previously, the role of Pyy in the regulation
f food intake and body weight has been contentious.eletion of the Pyy gene in this mouse model did not
ffect food intake or body weight in mice fed a standard i
how diet. When fed a high-fat diet, Pyy�/� mice exhib-ted greater weight gain, which was more prominent inemale mice and associated with greater adiposity. Theseffects were mild and were not linked to any detectablehanges in 24-hour food intake, locomotor activity, oretabolic parameters such as substrate utilization or
nergy expenditure. These small changes in body weightay reflect the subtle nature of the effects of PYY3-36 onetabolic parameters.13 The increase in adiposity in the
emale Pyy-deficient mice may be easier to detect than inale mice because female mice of this background re-ain comparatively lean on a high-fat diet. Thus, the
bsence of Pyy in female mice may allow them to gaineight more efficiently when exposed to a diet rich in fat.Three other groups have recently characterized inde-
endently generated Pyy�/� mouse models. In the firsteport to appear, Schonhoff et al15 described a mouse lineacking both Pyy and Ppy that shows no impairment ofood intake, body weight, or energy expenditure when fedstandard diet.15 This is significant given that Batterham
t al recently reported that infusion of PPY increasesatiety and decreases food intake in a manner and with aotency almost identical to PYY.11,34 Schonhoff et al used
mixed background of C57BL/6 and FVB/N mice.hereas male C57BL/6 mice are considered obesity
rone, FVB mice are obesity resistant. Similarly, the micen the present study are a mix of C57BL/6 and 129S6/vEvTac, where the latter strain is also obesity resistant.n agreement with the findings of Schonhoff et al, weound no difference in body weight in the Pyy�/� mice onstandard diet. The subtle difference in body weight gainnly became apparent when exposed to the high-fat diet,nd it is possible that rate of weight gain was limited byhe presence of the 129 background.
A specific Pyy null mouse line in which Ppy expressionemains intact was recently reported by Boey et al.16 Inheir studies, they report that female Pyy�/� mice displayn adult-onset obese phenotype on a standard diet. Al-hough this in contrast to our findings, we do noteimilarities in that the increased body weight induced byhe absence of Pyy is more prominent in female than in
ale mice and in the absence of any increase in hypo-halamic Npy expression or food intake in adult Pyy�/�
ice. Although body composition is analyzed in theireport, unfortunately no information on bone mass isiven. Variables that may account for the divergence inhe body weight findings include the mild hyperinsulin-mia observed in both male and female mice in additiono the elevated insulin-like growth factor I and testoster-ne levels seen in the male Pyy�/� mice. As the investi-ators point out in their study, the hyperinsulinemiaeen in these mice is most likely due to the removal of thenhibitory action of PYY on Y1 receptors in the pancreas.he fact that no changes in circulating insulin were seen
n Pyy�/� mice in the present study suggests that inhib-

ig
meacPtsTmitahtftPo
crcitm
pddetbrpsTa
cj
1
1
1
1
1
1
1
1
1
1
2
2
2
22
2
2
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1542 WORTLEY ET AL GASTROENTEROLOGY Vol. 133, No. 5
tory tone on the pancreas by this hormone can vary withenetic background.
More recently, Batterham et al17 described Pyy nullice that exhibit marked obesity and hyperphagia as
arly as 5 weeks of age and on a standard diet. Oncegain, an analysis of bone mass was not reported and weannot draw any comparisons to our findings. Theyy�/� mouse generated by Batterham et al is similar tohe N3F2 Pyy�/� mouse line described in the presenttudy in that the genetic background is �90% C57Bl/6.hus, the different body weight phenotypes in these 2ouse models are difficult to reconcile. However, the
ncreased food intake, body weight gain, and adiposity inhe Pyy�/� mice described by Batterham et al are presum-bly all a direct result of the increased expression ofypothalamic Npy and Agrp. In our Pyy�/� model, hypo-halamic expression of these neuropeptides was unaf-ected by the absence of circulating Pyy, consistent withhe lack of a metabolic phenotype. Why the absence ofyy impacts on hypothalamic neuropeptide expression inne mouse model but not the other is unclear.
Ppy shows selectivity for the NPY Y4 receptor10 and, inontrast to Pyy, this hormone does not appear to play aole in bone homeostasis or energy balance. The lack ofhange in BMD in these mice is consistent with findingsn transgenic mice that over express the Ppy gene or lackhe Y4 gene, both of which show no change in bone
ass.35
In summary, our studies show that neither Pyy nor Ppylay a critical role in regulating food intake. However, theeletion of Pyy does have a mild effect on adiposityuring long-term exposure to a high-fat diet, and thisffect is most prominent in female mice. More impor-antly, however, Pyy deletion causes a marked increase inone turnover and most likely plays a critical role inegulating bone resorption. Pyy cells in the gut are ideallylaced to sense nutrient intake and to integrate theseignals with the circadian rhythm of bone turnover.hus, Pyy mimetics could prove to be a novel therapeuticpproach to treating bone diseases.
Supplementary data
Supplementary data associated with this articlean be found, in the online version, at doi:10.1053/.gastro.2007.08.024.
References
1. Cerda-Reverter JM, Larhammar D. Neuropeptide Y family of pep-tides: structure, anatomical expression, function, and molecularevolution. Biochem Cell Biol 2000;78:371–392.
2. Conlon JM. The origin and evolution of peptide YY (PYY) andpancreatic polypeptide (PP). Peptides 2002;23:269–278.
3. Danger JM, Tonon MC, Jenks BG, et al. Localization in the centralnervous system and neuroendocrine functions. Fundam Clin Phar-
macol 1990;4:307–340.4. Ekblad E, Sundler F. Distribution of pancreatic polypeptide andpeptide YY. Peptides 2002;23:251–261.
5. Ekman R, Wahlestedt C, Bottcher G, et al. Peptide YY-like immu-noreactivity in the central nervous system of the rat. Regul Pept1986;16:157–168.
6. Pieribone VA, Brodin L, Friberg K, et al. Differential expression ofmRNAs for neuropeptide Y-related peptides in rat nervous tis-sues: possible evolutionary conservation. J Neurosci 1992;12:3361–3371.
7. Ballantyne GH. Peptide YY(1-36) and peptide YY(3-36): Part I.Distribution, release and actions. Obes Surg 2006;16:651–658.
8. Grandt D, Teyssen S, Schimiczek M, et al. Novel generation ofhormone receptor specificity by amino terminal processing ofpeptide YY. Biochem Biophys Res Commun 1992;186:1299–1306.
9. Dumont Y, Cadieux A, Pheng LH, et al. Peptide YY derivatives asselective neuropeptide Y/peptide YY Y1 and Y2 agonists de-voided of activity for the Y3 receptor sub-type. Brain Res MolBrain Res 1994 26:320–324.
0. Murphy KG, Dhillo WS, Bloom SR. Gut peptides in the regulationof food intake and energy homeostasis. Endocrinol Rev 200627:719–727.
1. Batterham RL, Le Roux CW, Cohen MA, et al. Pancreatic polypep-tide reduces appetite and food intake in humans. J Clin Endocri-nol Metab 2003;88:3989–3992.
2. Batterham RL, Cowley MA, Small CJ, et al. Gut hormone PYY(3-36)physiologically inhibits food intake. Nature 2002;418:650–654.
3. Tschop M, Castaneda TR, Joost HG, et al. Physiology: does guthormone PYY3-36 decrease food intake in rodents? Nature 2004;430:1.
4. Halatchev IG, Cone RD. Peripheral administration of PYY(3-36)produces conditioned taste aversion in mice. Cell Metab 2005.1:159–68.
5. Schonhoff S, Baggio L, Ratineau C, et al. Energy homeostasis andgastrointestinal endocrine differentiation do not require the anorec-tic hormone peptide YY. Mol Cell Biol 2005;25:4189–4199.
6. Boey D, Lin S, Karl T, et al. Peptide YY ablation in mice leads tothe development of hyperinsulinaemia and obesity. Diabetologia2006;49:1360–1370.
7. Batterham RL, Heffron H, Kapoor S, et al. Critical role for peptideYY in protein-mediated satiation and body-weight regulation. CellMetab 2006;4:223–233.
8. Valenzuela DM, Murphy AJ, Frendewey D, et al. High-throughputengineering of the mouse genome coupled with high-resolutionexpression analysis. Nat Biotechnol 2003;21:652–659.
9. Sleeman MW, Garcia K, Liu R, et al. Ciliary neurotrophic factorimproves diabetic parameters and hepatic steatosis and in-creases basal metabolic rate in db/db mice. Proc Natl Acad SciU S A 2003;100:14297–14302.
0. Peterson WM, Wang Q, Tzekova R, et al. Ciliary neurotrophicfactor and stress stimuli activate the Jak-STAT pathway in retinalneurons and glia. J Neurosci 2000;20:4081–4090.
1. Suri C, Jones PF, Patan S, et al. Requisite role of angiopoietin-1,a ligand for the TIE2 receptor, during embryonic angiogenesis.Cell 1996;87:1171–1180.
2. Parfitt AM, Drezner MK, Glorieux FH, et al. Bone histomorphom-etry: standardization of nomenclature, symbols, and units. Re-port of the ASBMR Histomorphometry Nomenclature Committee.J Bone Miner Res 1987;2:595–610.
3. Allison SJ, Herzog H. NPY and bone. EXS 2006;95:171–182.4. Karsenty G. The complexities of skeletal biology. Nature 2003;
423:316–318.5. Vaananen HK, Harkonen PL. Estrogen and bone metabolism.
Maturitas 1996;23(Suppl):S65–S69.6. Weitzmann MN, Pacifici R. Estrogen deficiency and bone loss: an
inflammatory tale. J Clin Invest 2006;116:1186–1194.

2
2
2
3
3
3
3
3
3
Ss
PM
poTdn
November 2007 PYY AND BONE 1543
7. Baldock PA, Sainsbury A, Couzens M, et al. Hypothalamic Y2 receptorsregulate bone formation. J Clin Invest 2002;109:915–921.
8. Dumont Y, Fournier A, St-Pierre S, et al. Characterization ofneuropeptide Y binding sites in rat brain membrane preparationsusing [125I][Leu31,Pro34]peptide YY and [125I]peptide YY3-36as selective Y1 and Y2 radioligands. J Pharmacol Exp Ther 1995;272:673–680.
9. Baldock PA, Sainsbury A, Allison S, et al. Hypothalamic control ofbone formation: distinct actions of leptin and y2 receptor path-ways. J Bone Miner Res 2005;20:1851–1857.
0. Takeda S, Elefteriou F, Levasseur R, et al. Leptin regulates boneformation via the sympathetic nervous system. Cell 2002;111:305–317.
1. Henriksen DB, Alexandersen P, Bjarnason NH, et al. Role ofgastrointestinal hormones in postprandial reduction of bone re-sorption. J Bone Miner Res 2003;18:2180–2189.
2. Clowes JA, Khosla S, Eastell R. Potential role of pancreatic andenteric hormones in regulating bone turnover. J Bone Miner Res2005;20:1497–1506.
3. Svensson J, Lall S, Dickson SL, et al. Effects of growth hormone
and its secretagogues on bone. Endocrine 2001;14:63–66. o4. Batterham RL, Cohen MA, Ellis SM, et al. Inhibition of food intakein obese subjects by peptide YY3-36. N Engl J Med 2003;349:941–948.
5. Sainsbury A, Baldock PA, Schwarzer C, et al. Synergistic effectsof Y2 and Y4 receptors on adiposity and bone mass revealed indouble knockout mice. Mol Cell Biol 2003;23:5225–5233.
Received March 17, 2007. Accepted July 12, 2007.Address requests for reprints to: Mark W. Sleeman, PhD, 777 Old
aw Mill River Road, Tarrytown, New York 10591. e-mail: [email protected]; fax: (914) 347-5045.The following authors disclose that they own stock in Regeneron
harmacueticals: K.E.W., K.G., H.O., K.T., K.D.A., D.V., G.D.Y., A.M., and.W.S. The remaining authors have no conflict of interest.The authors thank all at Regeneron Pharmaceuticals for their sup-
ort and assistance, especially the Velocigene core for the preparationf targeting vectors, blastocyst injections, and genotyping of mice;om Dechiara, William Poueymirou, and Melissa Meola for the coor-inated breeding of knockout mice; Victor Howard, Jane Murray, Jen-ifer Griffiths, and Hilary Cox for technical help; and Dr Lori Morton and
thers for comments and revisions of the manuscript.BA
SIC–
ALI
MEN
TARY
TRA
CT




















