
Interleukin (IL)-3/granulocyte macrophage-colony stimulating factor/IL5 receptor alpha and beta...
12
Interleukin (IL)-3/granulocyte macrophage-colony stimulating factor/IL-5 receptor alpha and beta chains are preferentially expressed in acute myeloid leukaemias with mutated FMS-related tyrosine kinase 3 receptor Acute myeloid leukaemia (AML) is a clonal disorder of haematopoiesis characterized by the accumulation of large numbers of myeloid blast cells that fail to differentiate into functional blood cells (Fialkow et al, 1987). Although most patients with newly diagnosed AML achieve a complete remission with conventional chemotherapy, the majority relapse and die with chemotherapy-resistant disease (Rowe et al, 1994; Bennet et al, 1997). New therapeutic approaches are therefore needed in this disease. Several lines of evidence suggest that multiple molecular mechanisms are involved in the pathogenesis of AMLs (Hahn & Weinberg, 2002), such as chromosome translocations, muta- tions and activated cell signalling pathways that may be responsible for the proliferation and/or apoptosis escape of leukaemic blasts. In recent years, it has been reported that constitutive extracellular signal-regulated kinase (ERK)1/2 activation is frequently found in solid tumours as well as in haematological diseases, particularly AML (Milella et al, 2001; Ricciardi et al, 2005). In this context, a key role is played by mutations or alterations in the level of expression of membrane receptors involved in the control of the proliferation of haemopoietic cells, such as FMS-related tyrosine kinase 3 (FLT3), cellular-receptor tyrosine kinase (c-kit) and interleu- kin-3 receptor (IL-3R). IL-3R is a heterodimeric membrane receptor (Chang et al, 2003). The a subunit is essential for ligand binding and confers ligand specificity to the receptor. The Roberta Riccioni, 1 Daniela Diverio, 2 Viviana Riti, 2 Sonia Buffolino, 2 Gualtiero Mariani, 1 Alessandra Boe, 1 Michele Cedrone, 3 Tiziana Ottone, 4 Robin Foa ` 2 and Ugo Testa 1 1 Department of Haematology, Oncology and Molecular Medicine, Istituto Superiore di Sanita `, 2 Department of Cellular Biotechnology and Haematology, University ‘‘La Sapienza’’, 3 Haematology Section Ospedale San Giovanni, and 4 Department of Biopathology, Universita ` ‘‘Tor Vergata’’, Rome, Italy Received 15 July 2008; accepted for publication 16 September 2008 Correspondence: Dr Ugo Testa, Department of Haematology, Oncology and Molecular Medicine, Istituto Superiore di Sanita `, Viale Regina Elena 299, Rome 00161, Italy. E-mail: [email protected] Summary The common beta chain subunit (b c ), also known as CDw131, shared by the interleukin-3 (IL-3), granulocytic macrophage colony-stimulating factor (GM-CSF) and IL-5 receptors, is required for high-affinity ligand binding and signal transduction. The present study explored the expression of CDw131 in 105 de novo cases of acute myeloid leukaemia (AML). The levels of CDw131 expression were used to identify two AML subgroups characterized by low (75/ 105) and high (30/105) expression of this receptor chain. It was observed that (i) the level of CDw131 expression strictly correlated with the level of CD116 (GM-CSFa receptor chain) and CD123 (IL-3Ra chain); (ii) AMLs with high CDw131 expression were characterized by low CD34 expression and usually high CD11b, CD14 expression; (iii) AMLs with high CDw131 expression frequently co-expressed receptors for angiogenic growth factors (vascular endothelial growth factor R2, Tie-2); (iv) AMLs with high CDw131 expression were more cycling than those with low CDw131 expression; (v) AMLs with high CDw131 frequently displayed Feline Murine Sarcoma (FMS-related) tyrosine kinase 3 (FLT3) internal tandem duplication and constitutively activated Signal Transducer and Activator of Transcription-5 (STAT5). In conclusion, the analysis of the level of CDw131 expression enabled the identification of a subset of AMLs characterized by a high cycling status, the expression of myelo-monocytic markers, mutated FLT3 and the co-expression of receptors for angiogenic growth factors. These findings are of value for the development of new therapeutic strategies for the treatment of these AMLs. Keywords: leukaemia, growth factor receptors, interleukin-3, granulo- monocytic colony stimulating factor, mutations. research paper First published online 25 November 2008 ª 2008 The Authors doi:10.1111/j.1365-2141.2008.07491.x Journal Compilation ª 2008 Blackwell Publishing Ltd, British Journal of Haematology, 144, 376–387
Transcript of Interleukin (IL)-3/granulocyte macrophage-colony stimulating factor/IL5 receptor alpha and beta...

Interleukin (IL)-3/granulocyte macrophage-colony stimulatingfactor/IL-5 receptor alpha and beta chains are preferentiallyexpressed in acute myeloid leukaemias with mutatedFMS-related tyrosine kinase 3 receptor
Acute myeloid leukaemia (AML) is a clonal disorder of
haematopoiesis characterized by the accumulation of large
numbers of myeloid blast cells that fail to differentiate into
functional blood cells (Fialkow et al, 1987). Although most
patients with newly diagnosed AML achieve a complete
remission with conventional chemotherapy, the majority
relapse and die with chemotherapy-resistant disease (Rowe
et al, 1994; Bennet et al, 1997). New therapeutic approaches
are therefore needed in this disease.
Several lines of evidence suggest that multiple molecular
mechanisms are involved in the pathogenesis of AMLs (Hahn &
Weinberg, 2002), such as chromosome translocations, muta-
tions and activated cell signalling pathways that may be
responsible for the proliferation and/or apoptosis escape of
leukaemic blasts. In recent years, it has been reported that
constitutive extracellular signal-regulated kinase (ERK)1/2
activation is frequently found in solid tumours as well as in
haematological diseases, particularly AML (Milella et al, 2001;
Ricciardi et al, 2005). In this context, a key role is played by
mutations or alterations in the level of expression of membrane
receptors involved in the control of the proliferation of
haemopoietic cells, such as FMS-related tyrosine kinase 3
(FLT3), cellular-receptor tyrosine kinase (c-kit) and interleu-
kin-3 receptor (IL-3R). IL-3R is a heterodimeric membrane
receptor (Chang et al, 2003). The a subunit is essential for ligand
binding and confers ligand specificity to the receptor. The
Roberta Riccioni,1 Daniela Diverio,2
Viviana Riti, 2 Sonia Buffolino,2
Gualtiero Mariani,1 Alessandra Boe,1
Michele Cedrone,3 Tiziana Ottone,4
Robin Foa2 and Ugo Testa1
1Department of Haematology, Oncology and
Molecular Medicine, Istituto Superiore di Sanita,2Department of Cellular Biotechnology and
Haematology, University ‘‘La Sapienza’’,3Haematology Section Ospedale San Giovanni,
and 4Department of Biopathology, Universita
‘‘Tor Vergata’’, Rome, Italy
Received 15 July 2008; accepted for publication
16 September 2008
Correspondence: Dr Ugo Testa, Department of
Haematology, Oncology and Molecular
Medicine, Istituto Superiore di Sanita, Viale
Regina Elena 299, Rome 00161, Italy.
E-mail: [email protected]
Summary
The common beta chain subunit (bc), also known as CDw131, shared by the
interleukin-3 (IL-3), granulocytic macrophage colony-stimulating factor
(GM-CSF) and IL-5 receptors, is required for high-affinity ligand binding
and signal transduction. The present study explored the expression of CDw131
in 105 de novo cases of acute myeloid leukaemia (AML). The levels of CDw131
expression were used to identify two AML subgroups characterized by low (75/
105) and high (30/105) expression of this receptor chain. It was observed that
(i) the level of CDw131 expression strictly correlated with the level of CD116
(GM-CSFa receptor chain) and CD123 (IL-3Ra chain); (ii) AMLs with high
CDw131 expression were characterized by low CD34 expression and usually
high CD11b, CD14 expression; (iii) AMLs with high CDw131 expression
frequently co-expressed receptors for angiogenic growth factors (vascular
endothelial growth factor R2, Tie-2); (iv) AMLs with high CDw131 expression
were more cycling than those with low CDw131 expression; (v) AMLs with
high CDw131 frequently displayed Feline Murine Sarcoma (FMS-related)
tyrosine kinase 3 (FLT3) internal tandem duplication and constitutively
activated Signal Transducer and Activator of Transcription-5 (STAT5). In
conclusion, the analysis of the level of CDw131 expression enabled the
identification of a subset of AMLs characterized by a high cycling status, the
expression of myelo-monocytic markers, mutated FLT3 and the co-expression
of receptors for angiogenic growth factors. These findings are of value for the
development of new therapeutic strategies for the treatment of these AMLs.
Keywords: leukaemia, growth factor receptors, interleukin-3, granulo-
monocytic colony stimulating factor, mutations.
research paper
First published online 25 November 2008 ª 2008 The Authorsdoi:10.1111/j.1365-2141.2008.07491.x Journal Compilation ª 2008 Blackwell Publishing Ltd, British Journal of Haematology, 144, 376–387

common beta chain (bc) subunit, also known as CDw131, which
is shared by the granulocyte macrophage-colony stimulating
factor (GM-CSF), IL-3 and IL-5 receptors, is required for high-
affinity ligand binding and signal transduction. IL-3R is
expressed on leukaemic blasts from the large majority of
patients with AML (Jordan et al, 2000; Testa et al, 2002). In
particular, the a subunit (also known as CD123) is often
expressed at much higher levels in AML than in normal
haematopoietic progenitor and precursor cells and it is also
detected on subpopulations of AML blasts enriched for malig-
nant progenitors, leukaemic stem cells that engraft non-obese
diabetic/severe combined immunodeficient mice (Shultz et al,
1995; Bonnett & Dick, 1997; Jordan et al, 2000). Interestingly,
recent microarray studies confirmed that IL3Ra is amongst the
seven genes that are more overexpressed in AMLs (Stirewalt
et al, 2008). Thus, IL-3R may be an appropriate target for
cytotoxic drugs (i.e., recombinant IL-3 fused with diphtheria
toxin) designed to selectively kill AML cells, whilst sparing their
normal haematopoietic cell counterparts (Testa et al, 2005;
Yalcintepe et al, 2006). IL-3R and GM-CSFR exert their
biological activity through interaction with cell surface receptors
that consist of two subunits, the alpha subunit specific to each
and the common beta chain (b chain or bc) (Miyajima et al,
1993; Bagley et al, 1997). The interaction of the a chain with the
b chain leads to the formation of a high-affinity receptor
complex, able to bind the respective ligand in the range of its
physiological concentrations and to transduce proliferative,
antiapoptotic and differentiative signals (Gearing et al, 1989;
Hayashida et al, 1990; Kitamura et al, 1991). The b chain
expressed alone in the absence of a specific a chain confers little
binding affinity for either IL-3 or GM-CSF (Tavernier et al,
1991; Hara & Miyajima, 1992). The bc chain is not only required
for high-affinity binding of ligands, but is also crucial for the
signalling of downstream pathways through its intracytoplas-
matic tail. In common with other receptors, the membrane
proximal region of bc appears to be both essential and sufficient
for proliferation (Sakamaki et al, 1992). This region is known to
bind the family of Janus Protein Kinase-2 (JAK2), and the
activation of JAK2 by tyrosine phosphorylation correlates
closely with the induction of mitogenesis (Quelle et al, 1994).
It is also essential for the induction of MYC, PIM1 and CIS.
Membrane-distal domains that are responsible for the major
tyrosine phosphorylation of proteins, induction of FOS and JUN
transcription, activation of Ras pathway, and prevention of
apoptosis (Sakamaki et al, 1992; Sato et al, 1993; Kinoshita et al,
1995) have been identified. In addition, the membrane-distal
region has been associated with the induction of differentiation
and negative regulation of receptor signals (Itoh et al, 1996).
Accumulated evidence suggest a role for bc in modulating signal
transduction of other haematopoietic cytokines, including GM-
CSF, erythropoietin (Epo) and stem cell factor (SCF) (Walker
et al, 1985). In addition to phosphorylation and physical
interaction, IL-3R and GM-CSFR are known to co-operate with
EpoR in erythropoiesis (Goodman et al, 1985; Sieff et al, 1989),
and common signal transduction pathways involving STAT5 are
used by these receptors (Pallard et al, 1995). STAT5 is activated
by a variety of ligands in different tissues. In primary leukaemic
cells a constitutive activation of the JAK-STAT pathway has been
demonstrated and the essential role of this pathway for the
malignant transformation of haematopoietic cells has been
suggested (Lin et al, 2000). In addition, bc has been implicated
in signalling by SCF via its receptor, c-kit, and SCF is able to
induce serine/threonine phosphorylation of bc (Liu et al, 1994),
even though there are no evidences about the physiological
significance of this receptor cross-talk in primary haemopoietic
cells (Scott et al, 2000).
The link between endothelial and haematopoietic systems is
well known because they share a common stem cell, the
haemangioblast. However, the role of vascular endothelial
growth factor (VEGF) receptors in myeloid leukaemias is
unclear. Two-thirds of AML patients show VEGF and VEGF
receptor 2 (VEGFR-2) expression, with concomitant increased
marrow microvascular density (Padro et al, 2002). Prolifera-
tion and survival of leukaemic blasts may be sustained by a
paracrine loop. Myeloid blasts release VEGF, which stimulates
adjacent marrow endothelial cells to secrete myeloid growth
factors (Bellamy et al, 1999; Dias et al, 2002). Furthermore, we
have recently shown that Tie-2, the receptor for angiopoietin-1
and -2, is expressed on monocytic and myelomonocytic AMLs
(Riccioni et al, 2007). On the other hand, AML blasts release
angiopoietins that may sustain leukaemic survival through an
autocrine/paracrine mechanism (Wakabayashi et al, 2004).
The current study examined the expression of bc in AML. In
this context, we were particularly interested in exploring
whether bc expression on AMLs might be associated with
specific cellular phenotypes. The level of bc expression was
measured by fluorescent-activated cell sorting (FACS) analysis
in 105 AML samples, which were divided into a group with
low and in a group with high bc expression. AMLs with high bc
expression were found to share myelomonocytic markers.
Furthermore, high bc expression was frequently associated
with the expression of VEGF-R2 and Tie-2. Finally, AMLs with
high bc expression very frequently displayed FLT3-internal
tandem duplication (ITD) mutations.
Materials and methods
Cells
Fresh leukaemic blasts from 105 patients with AML, obtained
after informed consent, were isolated from either bone marrow
or peripheral blood by Ficoll-Hypaque density gradient
centrifugation and immediately processed. All patients were
diagnosed at the Divisions of Haematology of the University
‘La Sapienza’, of the University ‘Tor Vergata’ and of the
Hospital ‘San Giovanni’, Rome, Italy. Leukaemias were
classified by morphological and cytochemical criteria
according to the French–American–British (FAB) classification
(2% corresponded to M0, 19% to M1, 24% to M2, 9% to M3,
26% to M4 and 20% to M5).
Expression of IL-3/GM-CSF/IL-5 in AML
ª 2008 The AuthorsJournal Compilation ª 2008 Blackwell Publishing Ltd, British Journal of Haematology, 144, 376–387 377

The following criteria were fulfiled in all cases diagnosed as
FAB M5 (acute monocytic leukaemia): (i) 70–80% of the
leukaemic cells were morphologically of the monocytic lineage,
including monoblasts, promonocytes and monocytes; (ii) the
leukaemic population contained a population, a percentage of
CD14+ cells >40%; (iii) a minor granulocytic component
(<20%); (iv) in FAB M5a cases the percentage of monoblasts is
predominant (>60–70%), whereas (v) in FAB M5b cases the
percentage of promonocytes and monocytes was predominant
(>70%); 6); finally, the large majority of the leukaemic
population showed intense non-specific esterase activity,
inhibited by sodium fluoride.
All analysed samples contained infiltration by more than
70% leukaemic blasts. Approval for these studies was obtained
from the Institutional Review Board of the Istituto Superiore
di Sanita, Rome, Italy. Informed consent was obtained in
accordance with the Declaration of Helsinki.
Immunophenotypic analysis of leukaemic cells
Analysis of cell surface antigens was performed by flow
cytometry using a FACScan Flow Cytometer (Becton Dickinson,
Bedford, MA, USA). The following antibodies directed to
membrane antigens were used for standard immunophenotypic
analysis of AML blasts: anti-CD3, -CD7, -CD11a, CD11b, -
CD11c, -CD13, -CD14, -CD15, -CD18, -CD19, -CD33, -CD34, -
CD36, -CD38, -CD41, -CD45, -CD61, -CD64, -CD71, -CD90, -
CD116, -CD117, -CD235 and -HLA-DR (all from Pharmingen/
Becton Dickinson). In addition, the following monoclonal
antibodies (mAbs) were used to characterize AML blasts: anti-
CDw123 (Pharmingen/Becton Dickinson) -CD116, CDw131
(Santa Cruz Biotechnology, Santa Cruz, CA, USA), anti-VEGF-
R1, -VEGF-R2, VEGF-R3, -Tie-2, -M-CSFR (Macrophage
Colony Stimulating Factor), -c-met, -IGF-1R (Insulin-like
Growth Factor-1 Receptor) all purchased from R & D Systems,
Minneapolis, MN, USA) -CD133 (Milteny, Bergisch Gladbach,
Germany), anti-Flt3 mAb purchased from Serotec (Oxford,
UK). Cells were labelled and analysed as previously reported
(Testa et al, 2002).
For quantitative evaluation of CDw123, CD116, CDw131
and CD135 expression, leukaemic cells were incubated with
specific antibodies and analysed by flow cytometry. Cell
fluorescence emission was evaluated, maintaining a fixed
photomultiplier tube (PMT) voltage to allow a quantitative
comparison between various samples. Fluorescence data were
evaluated in terms of mean fluorescence intensity (MFI)
calculated as the ratio between MFI observed in cells incubated
with anti-CDw123, CD116, CDw131, CD135 and the MFI
observed for the cells incubated with control IgG.
RNA extraction and RT-PCR analysis
Total RNA was extracted from leukaemic blasts using the
method of Chomczynsky and Sacchi (1987). One microgram
of total RNA was retro-transcribed using random hexamers
primers as published by Biomed Concerted Action Protocol
(van Dongen et al, 1999). To evaluate the expression of FLT3,
2 ll of reverse transcriptase was amplified within the linear
range by 35 PCR cycles: denaturation at 94�C for 30 s,
annealing at 56�C for 45 s, extension at 72�C for 30 s, followed
by 10 min at 72�C for final extension, in a mixture of 50 ll
volume containing 1· PCR buffer, 30 pmol of each specific
primer, 0Æ2 nmol/l dNTPs mix, 1Æ5 mmol/l MgCl2 and 1Æ25 U
Taq gold DNA polymerase. With the aim of simultaneously
analysing both ITD and D835, a multiplex PCR strategy was
adopted using the following four oligonucleotide primers: R5:
5¢-TGTCGAGCAGTACTCTAAACA-3¢ and R6: 5¢-ATCCTA-
GTACCTTCCCAAACTC-3¢, which amplify the entire trans-
membrane domain and the Juxta Membrane (JM) domain of
the gene; and 17F: 5¢-CCGCCAGGAACGTGCTTG-3¢ and
17R: 5¢-GCAGCCTCACATTGCCCC-3¢, which explore the
region containing exon 17 where the D835 mutation is
located. At the end of the PCR, 1Æ5 ll of these amplification
products were digested using the EcoRV restriction endonu-
clease and electrophoresed onto a 3% agarose gel containing
ethidium bromide. The quality of cDNA was assessed by
amplification of Hydroxymethylbilane Synthase (HMBS) using
the same conditions and cycling parameters and furthermore
RNA contamination was excluded using appropriate negative
controls.
Cell cycle analysis
Cell cycle analysis was carried out on nuclei stained with
propidium iodide (PI) using the Cycle TESTTM PLUS DNA
reagent of Becton-Dickinson (San Jose, CA, USA). Briefly,
100000 cells were first washed in buffer solution containing
sodium citrate, sucrose, dimethyl sulphoxide, and then resus-
pended in: 250 ll solution A to digest cell membrane and
cytoskeletons, 200 ll solution B containing trypsin inhibitor
and ribonuclease A to digest the RNA, 200 ll solution C
containing PI and spermine to stabilize nuclear chromatin.
The PI stoichiometrically binds to the DNA at a final
concentration of at least 125 lg/ml. Cells were then analysed
by flow cytometry using a FACScan equipped with software for
cell cycle analysis (Vindelov et al, 1983a,b).
Western blot analysis
To prepare total extracts, the cells were washed twice with cold
phosphate-buffered saline and lysed on ice for 30 min with 1%
Nonidet P40 lysis buffer (20 mmol/l Tris–HCl pH 8Æ0,
137 mmol/l NaCl, 10% glycerol, 2 mmol/l EDTA) in the
presence of 1 mmol/l phenylmethylsulfonylfluoride, 1 mmol/l
dithiothreitol, 1 mmol/l sodium orthovanadate, 2 lg/ml leu-
peptin and 2 lg/ml aprotinin. Cell debris was removed by
centrifugation at 3250 g for 10 min at 4�C, and protein
concentration of supernatants was determined using the Bio-
Rad protein assay (Richmond, CA, USA, http://www.biorad.
com). Aliquots of cells extracts containing 40 lg of total
R. Riccioni et al
ª 2008 The Authors378 Journal Compilation ª 2008 Blackwell Publishing Ltd, British Journal of Haematology, 144, 376–387

protein were resolved by 10% sodium dodecyl sulphate–
polyacrylamide gel electrophoresis under reducing and dena-
turing conditions and transferred onto Hybond-C extra
nitrocellulose membrane (Amersham Pharmacia Biotech,
Buckinghamshire, UK). Filters were blocked for 1 h at room
temperature in 5% non-fat dry milk dissolved in Tris-buffered
saline-Tween (TBS-T; 10 mmol/l Tris–HCl pH 8Æ0, 150 mmol/
l NaCl, 0,2% Tween 20) followed by incubation with primary
antibodies. After washing in TBS-T buffer, the filters were
incubated for 1 h at room temperature in 5% non-fat dry milk
dissolved in TBS-T containing a 1:4000 dilution of corre-
sponding peroxidase-conjugated secondary antibodies. Pro-
teins were visualized with the enhanced chemiluminescence
technique according to the manufacturer’s instructions (Super
Signal West Pico; Pierce, Rockford, IL, USA, http://
www.piercenet.com). Anti-ERK1/2 mAb was purchased from
Cell Signaling (Danvers, MA, USA), anti-pSTAT5 mAb was
purchased from R&D System Inc (Minneapolis, MN, USA),
anti-pERK mAb was purchased from Immunological Science
(Rome, Italy), anti-STAT5 mAb was purchased from Upstate
Biotechnology (Lake Placid, NY, USA, http://www.upstate.
com), anti-actin mAb was purchased from Oncogene Research
Products (Cambridge, MA, USA, http://www.oncogene.com)
and used as loading control.
Cell sorting
Leukaemic blasts stained with phycoerythrin-labelled anti IL-
3Rbc antibody were sorted according to fluorescence intensity
into IL-3Rbcdim and IL-3Rbc
bright fractions using a FACS
Vantage (Becton Dickinson).
Statistical analysis
The statistical significance of various parameters was evaluated
by the Student’s t-test. P values <0Æ05 were considered
significant.
Results
CDw131 expression in AML
The expression of CDw131, CD123 and CD116 on leukae-
mic blasts derived from 105 AML patients was investigated
by flow cytometry using specific mAbs directly conju-
gated with PE. In the majority of patients all the three
membrane receptor chains were detectable (representative
examples are shown in Fig 1). MFI analysis showed a
considerable variation in the label intensity for each of
Fig 1. Flow cytometric analysis of CDw131 (b-chain), CD123 (IL3R-a), CD116 (GM-CSFRa), Tie-2 and KDR in acute myeloid leukaemias (AMLs).
Flow cytometry histograms of three representative AMLs. Top middle and bottom panels correspond to AML cases displaying high, intermediate and
low CDw131 expression respectively. In each histogram the grey area corresponds to the fluorescence observed for cells incubated with control
irrelevant immunoglobulins, whilst the white area indicates the fluorescence observed for cells incubated with the indicated monoclonal antibodies.
IL, interleukin; GM-CSFR, granulocyte macrophage colony-stimulating factor; KDR, kinase domain receptor.
Expression of IL-3/GM-CSF/IL-5 in AML
ª 2008 The AuthorsJournal Compilation ª 2008 Blackwell Publishing Ltd, British Journal of Haematology, 144, 376–387 379
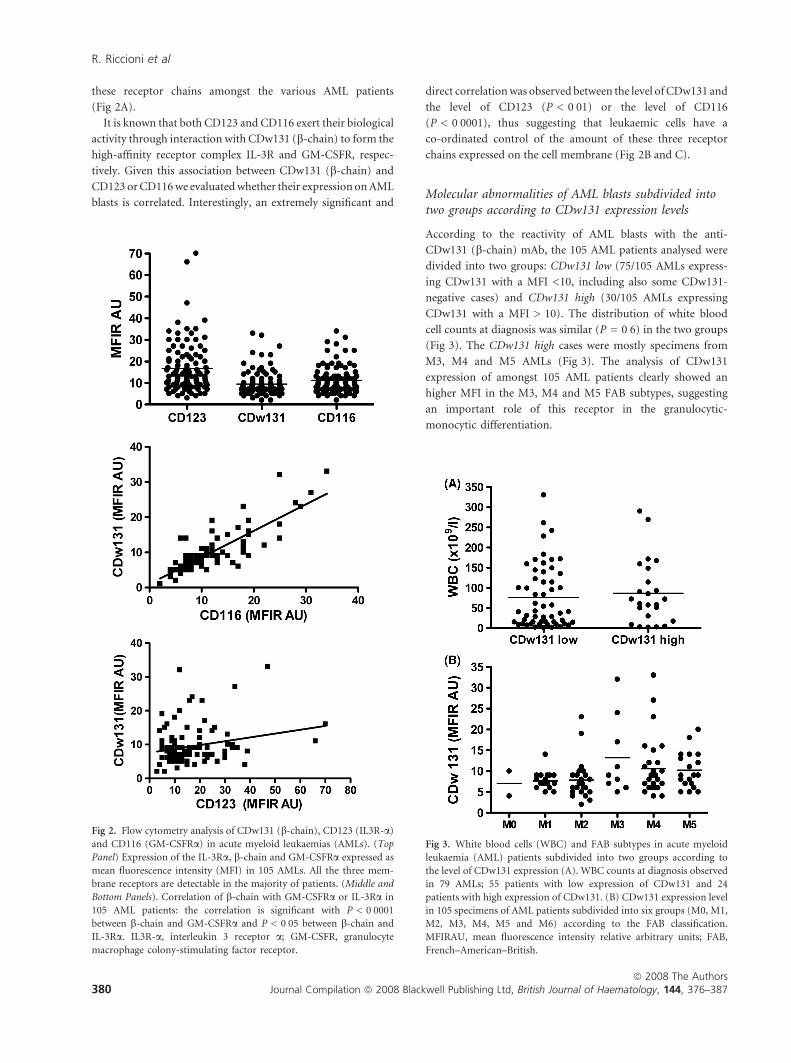
these receptor chains amongst the various AML patients
(Fig 2A).
It is known that both CD123 and CD116 exert their biological
activity through interaction with CDw131 (b-chain) to form the
high-affinity receptor complex IL-3R and GM-CSFR, respec-
tively. Given this association between CDw131 (b-chain) and
CD123 or CD116 we evaluated whether their expression on AML
blasts is correlated. Interestingly, an extremely significant and
direct correlation was observed between the level of CDw131 and
the level of CD123 (P < 0Æ01) or the level of CD116
(P < 0Æ0001), thus suggesting that leukaemic cells have a
co-ordinated control of the amount of these three receptor
chains expressed on the cell membrane (Fig 2B and C).
Molecular abnormalities of AML blasts subdivided intotwo groups according to CDw131 expression levels
According to the reactivity of AML blasts with the anti-
CDw131 (b-chain) mAb, the 105 AML patients analysed were
divided into two groups: CDw131 low (75/105 AMLs express-
ing CDw131 with a MFI <10, including also some CDw131-
negative cases) and CDw131 high (30/105 AMLs expressing
CDw131 with a MFI > 10). The distribution of white blood
cell counts at diagnosis was similar (P = 0Æ6) in the two groups
(Fig 3). The CDw131 high cases were mostly specimens from
M3, M4 and M5 AMLs (Fig 3). The analysis of CDw131
expression of amongst 105 AML patients clearly showed an
higher MFI in the M3, M4 and M5 FAB subtypes, suggesting
an important role of this receptor in the granulocytic-
monocytic differentiation.
Fig 2. Flow cytometry analysis of CDw131 (b-chain), CD123 (IL3R-a)
and CD116 (GM-CSFRa) in acute myeloid leukaemias (AMLs). (Top
Panel) Expression of the IL-3Ra, b-chain and GM-CSFRa expressed as
mean fluorescence intensity (MFI) in 105 AMLs. All the three mem-
brane receptors are detectable in the majority of patients. (Middle and
Bottom Panels). Correlation of b-chain with GM-CSFRa or IL-3Ra in
105 AML patients: the correlation is significant with P < 0Æ0001
between b-chain and GM-CSFRa and P < 0Æ05 between b-chain and
IL-3Ra. IL3R-a, interleukin 3 receptor a; GM-CSFR, granulocyte
macrophage colony-stimulating factor receptor.
Fig 3. White blood cells (WBC) and FAB subtypes in acute myeloid
leukaemia (AML) patients subdivided into two groups according to
the level of CDw131 expression (A). WBC counts at diagnosis observed
in 79 AMLs; 55 patients with low expression of CDw131 and 24
patients with high expression of CDw131. (B) CDw131 expression level
in 105 specimens of AML patients subdivided into six groups (M0, M1,
M2, M3, M4, M5 and M6) according to the FAB classification.
MFIRAU, mean fluorescence intensity relative arbitrary units; FAB,
French–American–British.
R. Riccioni et al
ª 2008 The Authors380 Journal Compilation ª 2008 Blackwell Publishing Ltd, British Journal of Haematology, 144, 376–387

The World Health Organization (WHO) classification for
tumours of haematopoietic tissues has linked recurrent
chromosomal abnormalities and other molecular abnormali-
ties to morphology, immunophenotype and other biological
features of AMLs. We have therefore analysed our patients for
the presence of some recurrent chromosomal rearrange-
ments associated with AMLs (Table I). The analysis was
separately extended to the two different groups with low
and high expression of CDw131 according to most fre-
quent AML chromosome translocation t(15;17) (q22:q21),
t (8;21) (q22:q22), inv16(p13:q22), t (6;9) (p23:q34), t(9;22)
(q34:q11) and t(11:q23; var). The frequency of these alterations
in the present AML series is shown in Table I, no significant
difference was observed between the two groups; however,
AMLs with high CDw131 expression were more positive for
the presence of PML/RARA and CBFB transcripts.
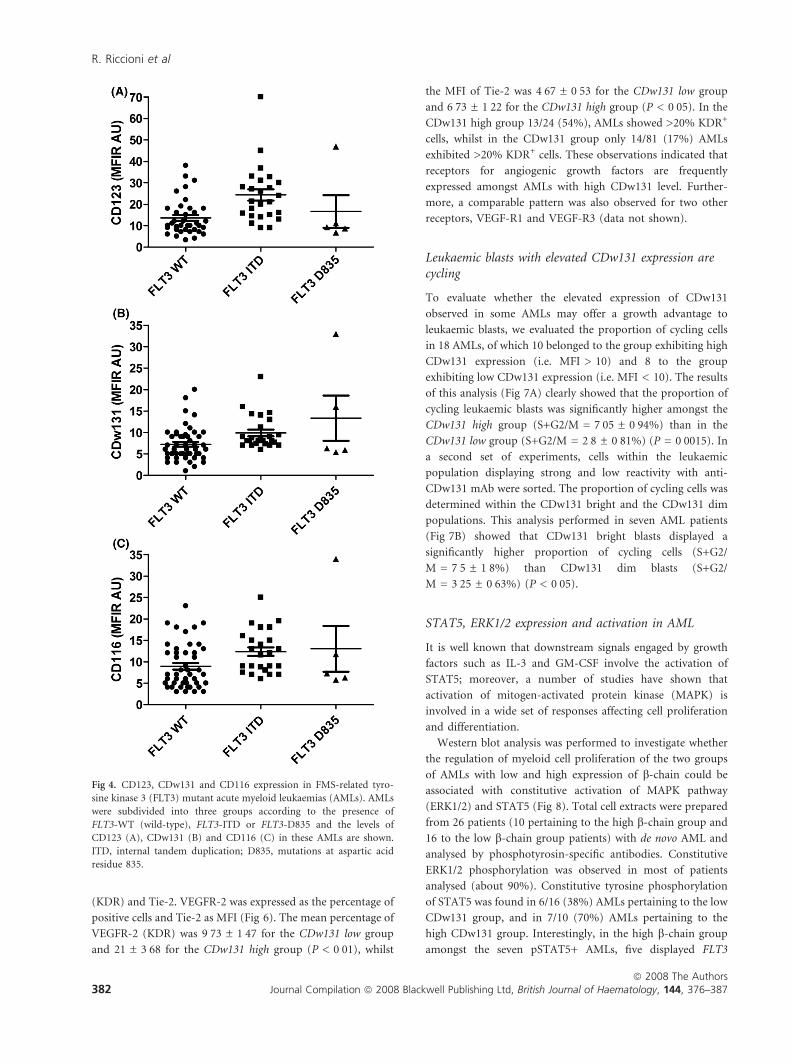
Incidence of FLT3-ITD and D835 mutations
Another important molecular abnormality observed in AMLs
is represented by the mutations of FLT3. To investigate
whether FLT3 mutations were associated with CDw131
expression, the distribution/ frequency of FLT3-ITD and
mutations at aspartic acid residue 835 (Flt3-D835) were
compared amongst 75 patients with de novo AML divided into
CDw131 low and CDw131 high: 57 AMLs low versus 18 AML
high. 20/57 (35%) of CDw131 low AMLs displayed FLT3-ITD/
D835 abnormalities, compared with 11/18 (61%) of CDw131
high AMLs. We further subdivided AMLs into three sub-
groups, FLT3 wild type (WT; 45 cases) FLT3-ITD (25 cases)
and FLT3 D835 (5 cases), and analysed CD123, CD116 and
CDw131 levels in these three groups. The results of this
analysis (Fig 4) showed that: (i) CD123 and CDw131 expres-
sion levels were clearly higher in FLT3-ITD than in FLT3-WT
(wild-type) AML (P £ 0Æ01 for CDw131 and <0Æ001 for
CD123); (ii) CD116 was expressed at higher levels in
FLT3-ITD than in FLT3-WT AML (P £ 0Æ05).
Analysis of immunophenotype
The distribution of the CDw131-positive AMLs along the
various FAB subtypes prompted a careful immunophenotypic
analysis of these AMLs. Expression of surface markers on
leukaemic cells from patients divided in the two CDw131 low
and CDw131 high groups is shown in Fig 5.
CD34, CD133 and CD117 (c-kit) present on haematopoietic
stem/progenitor cells were expressed as the percentage of
positive cells and were detected in most AMLs analysed, with
different patterns. An inverse relationship was observed
between CDw131 high expression and the CD34 marker. The
mean percentage of CD34+ cells in the CDw131 low group was
41 ± 3Æ98 (P < 0Æ01) compared with 24, 15 ± 6, 10 in the
CDw131 high group. CD133 and CD117 markers showed no
differences in the two groups: the mean percentage of CD133
and CD117 was 22Æ87 ± 3Æ45, 51Æ03 ± 3Æ63 for the CDw131 low
group and 27Æ22 ± 5Æ82, 56Æ54 ± 5Æ96 for the CDw131 high
group. CD135 level, expressed as MFI, was higher in the
CDw131 high group than in the CDw131 group (Fig 5).
Differences were observed for the expression of membrane
granulo-monocytic markers CD11b and CD14. These markers
were expressed in the two groups, but differences were
observed in terms of percentage of positive cells for each
marker: the mean percentage of CD11b and CD14 was
28Æ8 ± 2Æ83 and 15Æ13 ± 2Æ24 for CDw131 low group compared
with 48Æ3 ± 5Æ42 and 27Æ77 ± 5Æ14 for CDw131 high group.
The ensemble of these analyses provided several interesting
findings: (i) the majority of the cases in the CDw131 high
group showed a low proportion of CD34+ cells, whilst the
majority of CDw131 low group showed a high proportion of
CD34+ cells (Fig 4); (ii) a significant proportion of AMLs
pertaining to the CDw131 high group exhibited a high
percentage of CD11b and CD14 positive cells, whilst the
frequency of these cells was usually low amongst the CDw131
low group (Fig 5); (iii) FLT3 expression was constantly
elevated amongst the two groups with a higher MFI in the
CDw131 high group than in the CDw131 low group (Fig 5).
The higher CDw131 expression, associated with myelomono-
cytic markers, indicated that bc is involved in the myeloid
differentiation machinery.
Expression of VEGFR-2 (KDR)/Tie-2 angiogenic receptors
We explored also the expression of the receptors for the
angiogenic growth factors, VEGFR-2 Kinase Domain Receptor
Table I. Distribution of chromosomal and
molecular abnormalities amongst 59 AMLs with
CDw131 high and low expression. Chromosome
translocations Fusion gene
Frequency of
alteration CDw131
AML low (42 samples)
Frequency of alteration
CDw131 AML high
(17 samples)
t (15:17)(q22;q21) PML/RARA 6Æ6% 13Æ3%
t (8:21)(q22;q22) RUNX1/RUNX1T1 2Æ4% 0%
inv16 (p13;q22) CBFB/MYH11 19% 11Æ8%
t (6:9)(p23;q34) DEK/NUP214 2Æ4% 0%
t (9:22)(q34;q11) BCR/ABL1 0% 0%
t (11 q23:var.) MLL/var 2Æ4% 0%
AML, acute myeloid leukaemia.
Expression of IL-3/GM-CSF/IL-5 in AML
ª 2008 The AuthorsJournal Compilation ª 2008 Blackwell Publishing Ltd, British Journal of Haematology, 144, 376–387 381
(KDR) and Tie-2. VEGFR-2 was expressed as the percentage of
positive cells and Tie-2 as MFI (Fig 6). The mean percentage of
VEGFR-2 (KDR) was 9Æ73 ± 1Æ47 for the CDw131 low group
and 21 ± 3Æ68 for the CDw131 high group (P < 0Æ01), whilst
the MFI of Tie-2 was 4Æ67 ± 0Æ53 for the CDw131 low group
and 6Æ73 ± 1Æ22 for the CDw131 high group (P < 0Æ05). In the
CDw131 high group 13/24 (54%), AMLs showed >20% KDR+
cells, whilst in the CDw131 group only 14/81 (17%) AMLs
exhibited >20% KDR+ cells. These observations indicated that
receptors for angiogenic growth factors are frequently
expressed amongst AMLs with high CDw131 level. Further-
more, a comparable pattern was also observed for two other
receptors, VEGF-R1 and VEGF-R3 (data not shown).
Leukaemic blasts with elevated CDw131 expression arecycling
To evaluate whether the elevated expression of CDw131
observed in some AMLs may offer a growth advantage to
leukaemic blasts, we evaluated the proportion of cycling cells
in 18 AMLs, of which 10 belonged to the group exhibiting high
CDw131 expression (i.e. MFI > 10) and 8 to the group
exhibiting low CDw131 expression (i.e. MFI < 10). The results
of this analysis (Fig 7A) clearly showed that the proportion of
cycling leukaemic blasts was significantly higher amongst the
CDw131 high group (S+G2/M = 7Æ05 ± 0Æ94%) than in the
CDw131 low group (S+G2/M = 2Æ8 ± 0Æ81%) (P = 0Æ0015). In
a second set of experiments, cells within the leukaemic
population displaying strong and low reactivity with anti-
CDw131 mAb were sorted. The proportion of cycling cells was
determined within the CDw131 bright and the CDw131 dim
populations. This analysis performed in seven AML patients
(Fig 7B) showed that CDw131 bright blasts displayed a
significantly higher proportion of cycling cells (S+G2/
M = 7Æ5 ± 1Æ8%) than CDw131 dim blasts (S+G2/
M = 3Æ25 ± 0Æ63%) (P < 0Æ05).
STAT5, ERK1/2 expression and activation in AML
It is well known that downstream signals engaged by growth
factors such as IL-3 and GM-CSF involve the activation of
STAT5; moreover, a number of studies have shown that
activation of mitogen-activated protein kinase (MAPK) is
involved in a wide set of responses affecting cell proliferation
and differentiation.
Western blot analysis was performed to investigate whether
the regulation of myeloid cell proliferation of the two groups
of AMLs with low and high expression of b-chain could be
associated with constitutive activation of MAPK pathway
(ERK1/2) and STAT5 (Fig 8). Total cell extracts were prepared
from 26 patients (10 pertaining to the high b-chain group and
16 to the low b-chain group patients) with de novo AML and
analysed by phosphotyrosin-specific antibodies. Constitutive
ERK1/2 phosphorylation was observed in most of patients
analysed (about 90%). Constitutive tyrosine phosphorylation
of STAT5 was found in 6/16 (38%) AMLs pertaining to the low
CDw131 group, and in 7/10 (70%) AMLs pertaining to the
high CDw131 group. Interestingly, in the high b-chain group
amongst the seven pSTAT5+ AMLs, five displayed FLT3
Fig 4. CD123, CDw131 and CD116 expression in FMS-related tyro-
sine kinase 3 (FLT3) mutant acute myeloid leukaemias (AMLs). AMLs
were subdivided into three groups according to the presence of
FLT3-WT (wild-type), FLT3-ITD or FLT3-D835 and the levels of
CD123 (A), CDw131 (B) and CD116 (C) in these AMLs are shown.
ITD, internal tandem duplication; D835, mutations at aspartic acid
residue 835.
R. Riccioni et al
ª 2008 The Authors382 Journal Compilation ª 2008 Blackwell Publishing Ltd, British Journal of Haematology, 144, 376–387

Fig 5. Analysis of immunophenotypic features of acute myeloid leukaemias (AMLs) subdivided into two groups (CD131 low and CDw131 high)
according to the level of CDw131 expression. Membrane antigen expression was explored by flow cytometry, and results are expressed as the
percentage of positive cells for CD34, CD133, CD117, CD11b and CD14 and as mean fluorescence intensity (MFI) for FMS-related tyrosine kinase 3
(FLT-3). Differences between the two groups, CDw131 low and CDw131 high, were observed for the expression of CD34 (P < 0Æ01) myelo-monocytic
markers CD11b (P < 0Æ01) and CD14 (P < 0Æ01).
Fig 6. Expression of Tie-2 and vascular endothelial growth factor
receptor (VEGFR-2) (KDR) in acute myeloid leukaemias (AMLs)
subdivided into the two groups, low and high, according to the level of
CDw131 (b-chain). Flow cytometric analysis of Tie-2 and VEGFR-2
(KDR) in AMLs is subdivided into CDw131 low and CDw131 high
groups. The AMLs with high expression of CDw131 showed a higher
mean fluorescence intensity (MFI) of Tie-2 and a higher percentage of
VEGFR-2-positive cells than AMLs with low CDw131 expression.
Fig 7. Cell cycle status of acute myeloid leukaemia (AML) blasts: total
and sorted populations. (A) Evaluation of the proportion of cycling
blasts in 18 AMLs (total population) divided into the two groups with
low (8 AMLs) and high (10 AMLs) expression of CDw131. The per-
centage of cycling cells was higher in the CDw131 high group with
respect to the CDw131 low group. (B) Percentage of S-phase cells at
level of CDw131 bright and CDw131 dim subpopulation in seven
AMLs. The bright and dim populations were isolated by cell sorting.
The bright populations were more cycling than the corresponding dim
populations in all seven patients analysed.
Expression of IL-3/GM-CSF/IL-5 in AML
ª 2008 The AuthorsJournal Compilation ª 2008 Blackwell Publishing Ltd, British Journal of Haematology, 144, 376–387 383

mutations (four FLT3/ITD and one FLT3 D835), whilst in the
low b-chain group the seven pSTAT5+ AMLs, only one
displayed FLT3 mutations.
Discussion
Acute myeloid leukaemia is a heterogeneous group of
neoplastic disorders with great variability in clinical course
and response to therapy as well as in the genetic and molecular
basis of the pathology. Major advances in the understanding of
leukaemogenesis have been achieved by the characterization
and the study of acquired cytogenetic abnormalities; further-
more, gene mutations also constitute key events in AML
pathogenesis. In this context, mutations or enhanced expres-
sion of some membrane receptors contribute to confer a
proliferative advantage to AML blasts. A significant proportion
(about 40%) of AMLs displays mutations of FLT3 or c-kit
receptors, usually associated with poor cure rates due to
increased relapse risk (Small, 2006; Renneville et al, 2008). In
addition, FLT3 is overexpressed in some AML patients in the
absence of any mutation: the overexpressed FLT3 receptors are
auto-phosphorylated and constitutively signal-like mutated
receptors (Ozeki et al, 2004).
We recently reported that the IL-3Ra chain was over-
expressed in about 45% of AML patients and this overexpres-
sion was associated with increased proliferation of leukaemic
blasts and poor prognosis (Testa et al, 2002). The present
study explored the level of CDw131 expression in AMLs, with
the particular aim of evaluating whether this parameter may be
related to peculiar features and to the autonomous growth of
leukaemic blasts. Our observations indicated that CDw131 is
expressed by the large majority of AMLs and about 23% of
AML samples express this receptor chain at significantly high
levels. One observation suggests that elevated CDw131 expres-
sion may have a role in the proliferation of leukaemic cells. In
fact, we observed that within each leukaemic sample, the
fraction of cycling blasts was preferentially observed in the
fraction of cells expressing the highest CDw131 levels.
Interestingly, a significant and direct correlation between the
level of CDw131 and the level of CD123 or the level of CD116
was observed, thus suggesting that leukaemic cells have a
co-ordinated control of the amount of these receptor chains
expressed on the cell membrane. The comparative analysis of
the immunophenotypic features of CDw131 high and CDw131
low AMLs showed that: (i) CD34 was usually scarcely expressed
in CDw131 high AMLs; (ii) the majority of CDw131 high AMLs
consistently expressed both CD11b and CD14. These observa-
tions indicated that an elevated CDw131 expression on
leukaemic blasts is associated with more differentiated AMLs,
mainly pertaining to M4 and M5 subtypes.
The analysis of FLT3 mutations in AMLs highly expressing
CDw131 showed that about 61% of these AMLs displayed
FLT3 mutations (mainly FLT3-ITD) compared with a fre-
quency of about 35% of FLT3 mutants observed in AMLs
expressing low CDw131 levels. On the other hand, we observed
also that FLT3-ITD+ AMLs displayed an elevated CD123 and
CDw131 expression. These observations suggest that elevated
expression of CDw131 and CD123 is associated with FLT3-
ITD and that these two genes may represent molecular targets
for FLT3-ITD. Interestingly, in a previous study aiming to
evaluate the nuclear morphology of AMLs with mutated FLT3,
Kussick et al (2004) reported preliminary observations sug-
gesting that AMLs with FLT3-ITD strongly expressed CD123.
The mechanism through which FLT3-ITD may stimulate
CDw131 and CD123 expression remains to be determined.
The observation that AMLs with FLT3-ITD strongly express
CD123 may also have some implications for the development
of new therapies for the treatment of this category of AML
patients, who notoriously have a poor prognosis. Our obser-
vations suggest that these AMLs are potential candidates for
experimental treatment with diphtheria toxin-IL3 fusion
protein (Frankel et al, 2008).
Constitutive activation of STAT5 has been demonstrated in
>50% of AML cases (Birkenkamp et al, 2001; Spiekermann
et al, 2003) and may be attributed to activating mutations in
upstream kinases such as FLT3 (Hayakawa et al, 2000;
Fig 8. Analysis of constitutive extracellular signal-regulated kinase
(ERK1/2) and STAT5 phosphorylation in acute myeloid leukaemias
(AMLs) subdivided according to the level of CDw131 expression. Sixteen
AML samples were evaluated by western blotting for phosphor-STAT5
(pSTAT5), total STAT5 (STAT5 TOT), phosphor-ERK (pERK), total
ERK (ERK-TOT) and actin content. Samples corresponding to AMLs
with high CDw131 expression are indicated by an asterisk.
R. Riccioni et al
ª 2008 The Authors384 Journal Compilation ª 2008 Blackwell Publishing Ltd, British Journal of Haematology, 144, 376–387

Spiekermann et al, 2003; Yao et al, 2005; Rocnik et al, 2006),
Kinase Inducing Tyrosine (KIT) (Growney et al, 2005) or
JAK2 (Walters et al, 2006) or alternatively be due to autocrine
growth factor production (Birkenkamp et al, 2001). Finally, a
recent study showed that in AMLs with constitutive STAT5
activation, STAT5 is required for leukaemic cell proliferation
and for maintenance of leukaemic stem/progenitor cells
(Schepers et al, 2007). In the present study we reported the
frequent constitutive STAT5 activation in high CDw131 AMLs
(70%) compared with a lower frequency observed in low
CDw131 AMLs (38%). The high frequency of constitutive
STAT5 activation observed in high CDw131 AMLs was related
to the high frequency of FLT3-ITD observed in these AMLs.
We previously reported the frequent constitutive STAT5
activation in AMLs overexpressing IL-3Ra (Testa et al,
2002). This finding may be now easily explained by the
present observations showing that FLT3-ITD+ AMLs fre-
quently overexpress IL-3Ra.
An additional interesting finding of the present study
was the observation that CDw131 high AMLs frequently
co-expressed membrane endothelial markers such as Tie-2,
VEGF-R1 and VEGF-R2. These findings indicate that these
AMLs may represent a subtype of AMLs, as recently described
(Riccioni et al, 2007) and corresponding to myeloid cells
expressing endothelial markers. The normal counterpart of
these cells represent a population of haematopoietic proangio-
genic myeloid cells required for tumour vessel formation
(Yang et al, 2004; De Palma et al, 2005; Ahn & Brown, 2008).
References
Ahn, G. & Brown, J.M. (2008) Matrix metalloproteinase-9 is required
for tumor vasculogenesis but not for angiogenesis: role of bone
marrow-derived myelomonocytic cells. Cancer Cell, 13, 193–205.
Bagley, C.J., Woodcock, J.M., Stomski, F.C. & Lopez, A.F. (1997) The
structural and functional basis of cytokine receptor activation: les-
sons from the common b subunit of the granulocyte-macrophage
colony-stimulating factor, interlukin-3 (IL-3) and IL-5 receptor.
Blood, 89, 1471–1482.
Bellamy, W.T., Richter, L., Frutiger, Y. & Grogan, T.M. (1999)
Expression of vascular endothelial growth factor and its receptors in
hematopoietic malignancies. Cancer Research, 59, 728–733.
Bennet, J.M., Young, M.L., Andersen, J.W., Cassileth, P.A., Tallman,
M.S., Paietta, E., Wiernik, P.H. & Rowe, J.M. (1997) Long-term
survival in acute myeloid leukemia: the Eastern Cooperative
Oncology Group experience. Cancer, 80, 2205–2209.
Birkenkamp, K.U., Geugien, M., Lemmink, H.H., Kruijer, W. & Vel-
lenga, E. (2001) Regulation of constitutive STAT5 phosphorylation
in acute myeloid leukemia blasts. Leukemia, 15, 1923–1931.
Bonnett, D. & Dick, J.E. (1997) Human acute myeloid leukemia is
organized as a hierarchy that originates from a primitive hemato-
poietic cell. Nature Medicine, 3, 730–736.
Chang, F., Steelman, L.S., Lee, J.T., Shelton, J.G., Navolanic, P.M.,
Blalock, W.L., Franklin, R.A. & McCubrey, J.A. (2003) Signal
transduction mediated by the RAS/Raf/MEK/ERK pathway from
cytokine receptors to transcription factors: potential targeting for
therapeutic intervention. Leukemia, 17, 1263–1293.
Chomczynsky, P. & Sacchi, N. (1987) Single step method of RNA
isolation by acid guanidium thiocyanate-phenol chloroform
extraction. Analytical Biochemistry, 162, 156–159.
De Palma, M., Venneri, M.A., Galli, R., Sergi Sergi, L., Politi, L.S.,
Sampaolesi, M. & Naldini, L. (2005) Tie2 identifies a hematopoietic
lineage of proangiogenic monocytes required for tumor vessel for-
mation and a mesenchymal population of pericyte progenitors.
Cancer Cell, 8, 211–226.
Dias, S., Choy, M., Alitalo, K. & Rafii, S. (2002) Vascular endothelial
growth factor (VEGF)-C signaling through FLT-4 (VEGFR-3)
mediates leukemic cell proliferation, survival, and resistance to
chemotheraphy. Blood, 99, 2179–2186.
van Dongen, J.J., Macintyre, E.A., Gabert, J.A., Delabesse, E., Rossi, V.,
Saglio, G., Gottardi, E., Rambaldi, A., Dotti, G., Griesinger, F.,
Parreira, A., Gameiro, P., Diaz, M.G., Malec, M., Langerak, A.W.,
San Miguel, J.F. & Biondi, A. (1999) Standardized RT-PCR analysis
of fusion gene transcripts from chromosome aberrations in acute
leukemia for detection of minimal residual disease. Report of the
BIOMED-1 concerted action: investigation of minimal residual
disease in acute leukemia. Leukemia, 13, 1901–1928.
Fialkow, P.J., Singer, J.W., Raskind, W.H., Adamson, J.W., Jacobson,
R.J., Bernstein, I.D., Dow, L.W., Naifeld, V. & Veith, R. (1987)
Clonal development, stem-cell differentiation, and clinical remis-
sions in acute nonlymphocytic leukemia. The New England Journal
of Medicine, 317, 468–473.
Frankel, A., Liu, J.S., Rizzieri, D. & Hogge, D. (2008) Phase I clinical
study of diphteria toxin-interleukin-3 fusion protein in patients with
acute myeloid leukemia and myelodysplasia. Leukaemia & Lym-
phoma, 49, 543–553.
Gearing, D.P., King, J.A., Gough, N.M. & Nicola, N.A. (1989)
Expression cloning of a receptor for human granulocyte-macro-
phage colony stimulating factor. The EMBO Journal, 12, 3667–
3676.
Goodman, J.W., Hall, E.A., Miller, K.L. & Shinpock, S.G. (1985)
Interleukin 3 promotes erythroid burst formation in ‘‘serum-free’’
cultures without detectable erythropoietin. Proceedings of the
National Academy of Sciences of the United States of America, 82,
3291–3295.
Growney, J.D., Clark, J.J. & Adelsperger, J. (2005) Activation muta-
tions of human c-KIT resistant to imatinib mesylate are sensitive to
the tyrosine kinase PKC412. Blood, 106, 721–724.
Hahn, W.C. & Weinberg, R.A. (2002) Rules for making human tumor
cells. The New England Journal of Medicine, 347, 1593–1603.
Hara, T. & Miyajima, A. (1992) Two distinct functional high-affinity
receptors for mouse interleukin-3 (IL-3). The EMBO Journal, 11,
1875–1884.
Hayakawa, F., Towatari, M. & Kiyoi, H. (2000) Tandem-duplicated
Flt3 constitutively activates STAT5 and MAPK and introduces
autonomous cell growth in IL-3-dependent cell lines. Oncogene, 19,
624–631.
Hayashida, K., Kitamura, T., Gorman, D.M., Arai, K., Yokota, T. &
Miyajima, A. (1990) Molecular cloning of a second subunit of the
receptor for human granulocyte-macrophage colony-stimulating
factor (GM-CSF): reconstitution of a high affinity GM-CSF recep-
tor. Proceedings of the National Academy of Sciences of the United
States of America, 87, 9655–9659.
Itoh, T., Liu, R., Arai, K. & Watanabe, S. (1996) Roles of tyrosine
phosphorylation of the receptor b subunit in human GM-CSF signal
transduction. Blood, 88(Suppl. 1), 446a.
Expression of IL-3/GM-CSF/IL-5 in AML
ª 2008 The AuthorsJournal Compilation ª 2008 Blackwell Publishing Ltd, British Journal of Haematology, 144, 376–387 385

Jordan, C.T., Upchurch, D., Szilvassy, S.J., Guzman, M.L., Howard,
D.S., Pettigrew, A.L., Meyerrose, T., Rossi, R., Grimes, B., Rizzieri,
D.A., Luger, S.M. & Phillips, G.L. (2000) The interleukin-3 receptor
a chain is a unique marker for human acute myelogenous leukemia
stem cells. Leukemia, 14, 1777–1784.
Kinoshita, T., Yokota, T., Arai, K. & Miyajima, A. (1995) Suppression
of apoptotic death in hematopoietic cells by signaling through the
IL-3/GM-CSF receptors. The EMBO Journal, 14, 266–275.
Kitamura, T., Hayashida, K., Sakamaki, K., Yokota, T., Arai, K. &
Miyajima, A. (1991) Reconstitution of functional receptors for human
granulocyte/macrophage colony stimulating factor (GM-CSF) :
evidence that the protein encoded by the AIC2B cDNA is a subunit of
the murine GM-CSF receptor. Proceedings of the National Academy of
Sciences of the United States of America, 88, 5082–5086.
Kussick, S., Stirewalt, D.L., Yi, H.S., Sheets, K.M., Pogosowa-Agadja-
nyan, E., Braswell, S., Norwood, T.H., Radich, J.P. & Wood, B.L.
(2004) A distinctive nuclear morphology in acute myeloid leukemia
is strongly associated with loss of HLA-DR expression and FLT3
internal tandem duplication. Leukemia, 18, 1591–1598.
Lin, T.S., Mahajan, S. & Frank, D.A. (2000) STAT signaling in the
phatogenesis and treatment of leukemias. Oncogene, 19, 2496–2504.
Liu, L., Cutler, R.L., Mui, A.L. & Krystal, G. (1994) Steel factor stim-
ulates the serine/threonine phosphorylation of the interleukin-3
receptor. The Journal of Biological Chemistry, 269, 16774–16779.
Milella, M., Kornblau, S.M., Estrov, Z., Carter, B.Z., Lapillonne, H.,
Harris, D., Konopleva, M., Zhao, S., Estey, E. & Andreeff, M. (2001)
Therapeutic targeting of the MEK/MAPK signaling transduction
module in acute myeloid leukemia. The Journal of Clinical Investi-
gation, 108, 851–859.
Miyajima, A., Mui, A.l., Orogochi, T. & Sakamaki, K. (1993) Receptors
for granulocyte-macrophage colony-stimulating factor, interleukin-
3, and interleukin-5. Blood, 82, 1960–1974.
Ozeki, K., Kiyoi, H., Hirose, Y., Iwai, M., Ninomiya, M., Kodera, Y.,
Miyawaki, S., Kuriyama, K., Shimazaki, C., Akiyama, H., Nishimura,
M., Motoji, T., Shinagawa, K., Takeshita, A., Ueda, R., Ohno, R.,
Emi, N. & Naoe, T. (2004) Biologic and clinical significance of the
FLT3 transcript level in acute myeloid leukemia. Blood, 103, 1901–
1908.
Padro, T., Biker, R., Ruiz, S., Steins, M., Retzlaff, S., Burger, H.,
Buchner, T., Kessler, T., Herrera, F., Kienast, J., Muller-Tidow, C.,
Serve, H., Berdel, W.E. & Mesters, R.M. (2002) Overexpression of
vascular endothelial growth factor (VEGF) and its cellular receptor
KDR (VEGFR-2) in the bone marrow of patients with acute myeloid
leukemia. Leukemia, 16, 1302–1310.
Pallard, C., Gouilleux, F., Charon, M., Groner, B., Gisselbrecht, S. &
Dusanter-Fourt, I. (1995) Interleukin-3, erythropoietin, and pro-
lactin activate a STAT5-like factor in lymphoid cells. The Journal of
Biological Chemistry, 270, 15942–15945.
Quelle, F.W., Sato, N., Witthuhn, B.A., Inhorn, R.C., Eder, M.,
Miyajima, A., Griffin, J.D. & Ihle, J.N. (1994) JAK2 associates with
the bc chain of the receptor for granulocyte-macrophage colony-
stimulating factor, and its activation requires the membrane-prox-
imal region. Molecular and Cellular Biology, 14, 4335–4341.
Renneville, A., Roumier, C., Biggio, V., Nibourel, O., Boissel, N.,
Fenaux, P. & Preudhomme, C. (2008) Cooperating gene mutations
in acute myeloid leukemia: a review of the literature. Leukemia, 22,
915–931.
Ricciardi, M.R., McQueen, T., Chism, D., Milella, M., Estey, E.,
Kaldjian, E., Sebolt-Leopold, J., Konopleva, M. & Andreef, M.
(2005) Quantitative single cell determination of ERK phosphoryla-
tion and regulation in relapsed and refractory primary acute myeloid
leukemia. Leukemia, 19, 1543–1549.
Riccioni, R., Diverio, D., Mariani, G., Buffolino, S., Riti, V., Saulle, E.,
Petrucci, E., Cedrone, M., Lo-Coco, F., Foa, R., Peschle, C. & Testa,
U. (2007) Expression of Tie-2 and other receptors for endothelial
growth factors in acute myeloid leukemias is associated with
monocytic features of leukemic blasts. Stem Cells, 25, 1862–1871.
Rocnik, J.L., Okabe, R., Yu, J.C., Lee, B.H., Giese, N., Schenkein, D.P.
& Gilliland, D.G. (2006) Roles of tyrosine 589 and 591 in STAT5
activation and transformation mediated by FLT3-ITD. Blood, 108,
1339–1345.
Rowe, J.M., Andersen, J., Cassileth, P.A., Oken, M.M., Bennet, J.M. &
Wiernik, P.H. (1994) Clinical trials of adults with acute myelo-
genous leukemia: experience of the Eastern Cooperative Oncology
Group. In: Acute Leukemias IV. Prognostic factors and treatment
strategies (by W. Hiddemann, T. Buchner, B. Wormann &
M. Keating), pp. 541–546. Springer-Verlag, Germany.
Sakamaki, K., Miyajima, I., Kitamura, T. & Miyajima, A. (1992)
Critical cytoplasmic domains of the common b subunit of the
human GM-CSF, IL-3R and IL-5 receptors for the growth signal
transduction and tyrosine phosphorylation. The EMBO Journal, 11,
3541–3549.
Sato, N., Sakamaki, K., Terada, N., Arai, K. & Miyajima, A. (1993)
Signal transduction by the high-affinity GM-CSF receptor: two
distinct cytoplasmic regions of the common b subunit responsible
for different signaling. The EMBO Journal, 12, 4181–4189.
Schepers, H., Van Gosliga, D., Wierenga, A.T., Eggen, B., Schuringa,
J.J. & Vellenga, E. (2007) STAT5 is required for long-term mainte-
nance of normal and leukemic human stem/progenitor cells. Blood,
110, 2880–2888.
Scott, C.L., Robb, L., Papaevangeliou, B., Mansfield, R., Nicola, N.A. &
Begley, C.G. (2000) Reassessment of interactions between hemato-
poietic receptors using common beta-chain and interleukin-3-
specific receptor beta-chain-null cells: no evidence of functional
interactions with receptors for erythropoietin, granulocyte
colony-stimulating factor, or stem cell factor. Blood, 96, 1588–
1590.
Shultz, L.D., Schweitzer, P.A., Christianson, S.W., Gott, B., Schweitzer,
I.B., Tennent, B., McKenna, S., Mobraaten, L., Rajan, T.V., Greiner,
D.L. & Leiter, E.H. (1995) Multiple defects in innate and adaptive
immunologic function in NOD/LtSz-scid mice. Journal of Immu-
nology, 154, 180–191.
Sieff, C.A., Ekern, S.C., Nathan, D.G. & Anderson, J.W. (1989)
Combinations of recombinant colony-stimulating factors are
required for optimal hematopoietic differentiation in serum-
deprived culture. Blood, 73, 688–693.
Small, D. (2006) FLT3 mutations: biology and treatment. Hematology/
the Education Program of the American Society of Hematology.
American Society of Hematology. Education Program, 2006, 178–184.
Spiekermann, K., Bagrintseva, K., Schwab, R., Schmieja, K. & Hidde-
mann, W. (2003) Overexpression and constitutive activation of
FLT3 induces STAT5 activation in primary acute myeloid leukemia
blast cells. Clinical Cancer Research, 9, 2140–2150.
Stirewalt, D.L., Meshinchi, S., Kopecky, K.J., Fan, W., Pogosova-
Agadjanyan, E.L., Engel, J.H., Cronk, M.R., Dorcy, K.S., McQuary,
A.R., Hockenbery, D., Wood, B., Heimfeld, S. & Radich, J.P. (2008)
Identification of genes with abnormal expression changes in acute
myeloid leukemia. Genes, Chromosomes and Cancer, 47, 8–20.
R. Riccioni et al
ª 2008 The Authors386 Journal Compilation ª 2008 Blackwell Publishing Ltd, British Journal of Haematology, 144, 376–387

Tavernier, J., Devos, R., Cornelis, S., Tuypens, T., Van der Heyden, J.,
Fiers, W. & Plaetinck, G. (1991) A human high-affinity interleukin-5
receptor (IL-5R) is composed of an IL-5 specific alpha chain and a
beta chain shared with the receptor for GM-CSF. Cell, 66, 1175–
1184.
Testa, U., Riccioni, R., Militi, S., Coccia, E., Stellacci, E., Samoggia, P.,
Latagliata, R., Mariani, G., Rossini, A., Battistini, A., Lo-Coco, F. &
Peschle, C. (2002) Elevated expression of IL-3Ra in acute myelo-
genous leukemia is associated with enhanced blast proliferation,
increased cellularity, and poor prognosis. Blood, 100, 2980–2988.
Testa, U., Riccioni, R., Biffoni, M., Diverio, D., Lo-Coco, F., Foa, R.,
Peschle, C. & Frankel, A.E. (2005) Diphteria toxin fused to variant
human interleukin-3 induces cytotoxicity of blasts from patients
with acute myeloid leukemia according to the level of interleukin-3
receptor expression. Blood, 106, 2527–2529.
Vindelov, L.L., Christensen, I.J., Jensen, G. & Nissen, N.I. (1983a)
Limits of detection of nuclear DNA abnormalities by flow cytometric
DNA analysis. Results obtained by a set of methods for sample-
storage, staining and internal standardization. Cytometry, 3, 332–339.
Vindelov, L.L., Christensen, I.J. & Nissen, N.I. (1983b) A detergent-
trypsin method for the preparation of nuclei for flow cytometric
DNA analysis. Cytometry, 3, 323–327.
Wakabayashi, M., Miwa, H., Shikami, M., Hiramatsu, A., Ikai, T.,
Tajima, E., Yamamoto, H., Miura, K., Satoh, A., Itoh, M., Imamura,
A., Mihara, H., Katoh, Y. & Nitta, M. (2004) Autocrine pathway
of angiopoietin-Tie2 system in AML cells: association with
phosphatidyl-inositol 3 kinase. The Hematology Journal, 5, 353–
360.
Walker, F., Nicola, N.A., Metcalf, D. & Burgess, A.W. (1985) Hierar-
chical down-modulation of hemopoietic growth factor receptors.
Cell, 43, 269–276.
Walters, D.K., Goss, V.L. & Stroffegen, E.P. (2006) Phosphoproteomic
analysis of AML cell lines identifies leukemic oncogenes. Leukemia
Research, 30, 1097–1104.
Yalcintepe, L., Frankel, A.E. & Hogge, D.E. (2006) Expression of
interleukin-3 receptor subunits on defined subpopulations of acute
myeloid leukaemia blasts predicts the cytotoxicity of diphtheria
toxin interleukin-3 fusion protein against malignant progenitors
that engraft in immunodeficient mice. Blood, 108, 3530–3537.
Yang, L., DeBusk, L.M., Fukuda, K., Fingleton, B., Green-Jarvis, B.,
Shyr, Y., Matrisian, L.M., Carbone, D.P. & Lin, P.C. (2004)
Expansion of myeloid immune suppressor Gr+CD11b+ cells in
tumor-bearing host directly promotes tumor angiogenesis. Cancer
Cell, 6, 409–421.
Yao, Q., Nishiuchi, R., Kitamura, T. & Kersey, J.H. (2005) Human
leukemias with mutated FLT3 kinase are synergistically sensitive to
FLT3 and Hsp90 inhibitors: the key role of the STAT5 signal
transduction pathway. Leukemia, 19, 1605–1612.
Expression of IL-3/GM-CSF/IL-5 in AML
ª 2008 The AuthorsJournal Compilation ª 2008 Blackwell Publishing Ltd, British Journal of Haematology, 144, 376–387 387




















